
Role Gene Bacteriophage T4 Repair UV-Irradiated Alkylatedjvi.asm.org/content/16/1/5.full.pdf ·...
12
JOURNAL OF VIROLOGY, July 1975, p. 5-16 Copyright 0 1975 American Society for Microbiology Vol. 16, No. 1 Printed in U.SA. Role of Gene 59 of Bacteriophage T4 in Repair of UV-Irradiated and Alkylated DNA In Vivo REEN WU,I JEN-LEIH WU, AND YUN-CHI YEH* Department of Biochemistry, School of Medicine, University of Arkansas, Little Rock, Arkansas 72201 Received for publication 15 October 1974 Nonsense mutants in gene 59 (amC5, amHL628) were used to study the role of this gene in the repair of UV-damaged and alkylated DNA of bacteriophage T4 in vivo. The higher sensitivity to UV irradiation and alkylation of gene 59 mutants after exposure to these agents was established by a comparison of the survival fractions with wild type. Zonal centrifugal analysis of both parental and nascent mutant intracellular DNA molecules after UV irradiation showed that immedi- ately after exposure the size of single-stranded DNA fragments was the same as the wild-type intracellular DNA. However, the capability of rejoining fragmented ntracellular DNA was greatly reduced in the mutant. In contrast, the wild- type-infected cells under the same condition resumed DNA replication and repaired its DNA to normal size. Methyl methanesulfonate induced more randomly fragmented intracellular DNA, when compared to UV irradiation. The rate of rejoining under these conditions as judged from their sedimentation profiles was also greatly reduced in mutant-infected cells. Further evidence is presented that UV repair is not a simple consequence of arrested DNA replication, which is a phenotype of the mutant when infected in a nonpermissive host, Escherichia coli B (suj), but rather that the DNA repair function of gene 59 is independent of the replication function. These and other data presented indicate that a product(s) of gene 59 is essential for both repair of UV lesions and repair of alkylation damage of DNA in vivo. It is suggested that gene 59 may have two functions during viral development: DNA replication and replication repair of DNA molecules. Several lines of evidence suggest that the mechanisms for DNA replication, repair, and genetic recombination are closely related (9, 16, 22, 26, 39). However, due to the complexity of each mechanism, the nature and role of many proteins involved in these processes have not been elucidated in prokaryotic as well as eukaryotic cells. Excision repair and postreplication repair mechanisms for DNA lesions have been de- scribed in Escherichia coli (6, 10, 18, 28, 30, 39). Repair mechanisms similar to these two mech- anisms have been reported in phage T4 (2, 5, 15, 17, 23, 25, 32, 36). The first process is excision repair which is initiated by a thymine dimer- specific endonuclease coded for by the v gene (14, 15, 31, 37, 43). This process is followed by excision of the dimer, reinsertion of new nucleo- tides, and rejoining of newly formed fragments to the parental strand. E. coli DNA polymerase 1 (gene polA) and T4 genes 1, 30, 42, 45, 56 are 'Present address: Department of Biology, University of California, San Diego, LaJolla, Calif. 92037. 5 thought to be involved in excision repair. From the viewpoint of radiobiological behavior, the v mutant is similar to uvr mutant in E. coli which is sensitive to UV irradiation, but not sensitive to alkylating agents and X ray (25). The second repair process, replication repair, is mediated by x, y, and other early genes of phage T4 which are required for normal phage replication. T4 DNA polymerase (gene 43), but not E. coli DNA polymerase I, is involved in this process (36). In this repair process, T4 x and y are somewhat similar to E. coli recA, -B, and -C which are sensitive to UV irradiation, alkylat- ing agents, and X ray (25). Gene 59 is an essential gene for phage DNA replication (12, 41; R. Wu and Y. C. Yeh, Bacteriol. Proc., p. 167, 1970). Mutants of gene 59, T4amC5 and T4amHL628, have a pheno- type of arrested DNA synthesis after a short interval of infection in a nonpermissive host, E. coli B (41). In wild-type-infected cells the DNA is initially associated with cell membrane frac- tions (1,600S). In contrast, we found mutant on July 18, 2018 by guest http://jvi.asm.org/ Downloaded from
Transcript of Role Gene Bacteriophage T4 Repair UV-Irradiated Alkylatedjvi.asm.org/content/16/1/5.full.pdf ·...

JOURNAL OF VIROLOGY, July 1975, p. 5-16Copyright 0 1975 American Society for Microbiology
Vol. 16, No. 1Printed in U.SA.
Role of Gene 59 of Bacteriophage T4 in Repair of UV-Irradiatedand Alkylated DNA In Vivo
REEN WU,I JEN-LEIH WU, AND YUN-CHI YEH*
Department of Biochemistry, School of Medicine, University of Arkansas, Little Rock, Arkansas 72201
Received for publication 15 October 1974
Nonsense mutants in gene 59 (amC5, amHL628) were used to study the role ofthis gene in the repair of UV-damaged and alkylated DNA of bacteriophage T4 invivo. The higher sensitivity to UV irradiation and alkylation of gene 59 mutantsafter exposure to these agents was established by a comparison of the survivalfractions with wild type. Zonal centrifugal analysis of both parental and nascentmutant intracellular DNA molecules after UV irradiation showed that immedi-ately after exposure the size of single-stranded DNA fragments was the same asthe wild-type intracellular DNA. However, the capability of rejoining fragmentedntracellular DNA was greatly reduced in the mutant. In contrast, the wild-type-infected cells under the same condition resumed DNA replication andrepaired its DNA to normal size. Methyl methanesulfonate induced morerandomly fragmented intracellular DNA, when compared to UV irradiation. Therate of rejoining under these conditions as judged from their sedimentationprofiles was also greatly reduced in mutant-infected cells. Further evidence ispresented that UV repair is not a simple consequence of arrested DNAreplication, which is a phenotype of the mutant when infected in a nonpermissivehost, Escherichia coli B (suj), but rather that the DNA repair function of gene 59is independent of the replication function. These and other data presentedindicate that a product(s) of gene 59 is essential for both repair of UV lesions andrepair of alkylation damage of DNA in vivo. It is suggested that gene 59 may havetwo functions during viral development: DNA replication and replication repairof DNA molecules.
Several lines of evidence suggest that themechanisms for DNA replication, repair, andgenetic recombination are closely related (9, 16,22, 26, 39). However, due to the complexity ofeach mechanism, the nature and role of manyproteins involved in these processes have notbeen elucidated in prokaryotic as well aseukaryotic cells.
Excision repair and postreplication repairmechanisms for DNA lesions have been de-scribed in Escherichia coli (6, 10, 18, 28, 30, 39).Repair mechanisms similar to these two mech-anisms have been reported in phage T4 (2, 5, 15,17, 23, 25, 32, 36). The first process is excisionrepair which is initiated by a thymine dimer-specific endonuclease coded for by the v gene(14, 15, 31, 37, 43). This process is followed byexcision of the dimer, reinsertion of new nucleo-tides, and rejoining of newly formed fragmentsto the parental strand. E. coli DNA polymerase1 (gene polA) and T4 genes 1, 30, 42, 45, 56 are
'Present address: Department of Biology, University ofCalifornia, San Diego, LaJolla, Calif. 92037.
5
thought to be involved in excision repair. Fromthe viewpoint of radiobiological behavior, the vmutant is similar to uvr mutant in E. coli whichis sensitive to UV irradiation, but not sensitiveto alkylating agents and X ray (25). The secondrepair process, replication repair, is mediatedby x, y, and other early genes of phage T4 whichare required for normal phage replication. T4DNA polymerase (gene 43), but not E. coliDNA polymerase I, is involved in this process(36). In this repair process, T4 x and y aresomewhat similar to E. coli recA, -B, and -Cwhich are sensitive to UV irradiation, alkylat-ing agents, and X ray (25).Gene 59 is an essential gene for phage DNA
replication (12, 41; R. Wu and Y. C. Yeh,Bacteriol. Proc., p. 167, 1970). Mutants of gene59, T4amC5 and T4amHL628, have a pheno-type of arrested DNA synthesis after a shortinterval of infection in a nonpermissive host, E.coli B (41). In wild-type-infected cells the DNAis initially associated with cell membrane frac-tions (1,600S). In contrast, we found mutant
on July 18, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

6 WU, WU, AND YEH
DNA was released from the cell membrane 2 to3 min after the initiation of DNA replicationand gradually accumulated (42). Furthermore,abnormality in concatemer formation as well assuppression of arrested DNA synthesis by chlor-amphenicol or by the introduction of exergenicmutations were observed (41, 42; R. Wu and Y.C. Yeh, Cold Spring Harbor Phage Meet., p. 59,1970). However, the role of this gene product inrepair of DNA lesions induced by UV irradiationand alkylation is still unknown. In this paper,we shall present biochemical as well as biologi-cal evidence which show that gene 59 is essen-tial for the replication repair of damages causedby UV and alkylating agents in vivo. There hasbeen a preliminary account of some of theseexperiments (Wu and Yeh, Bacteriol. Proc., p.167, 1970).
MATERIALS AND METHODSBacteria and bacteriophage. Escherichia coli K
strain CR63 was used as a permissive host for prepar-ing stocks of T4 amber mutants. E. coli B strainTr201, a low thymine-requiring mutant obtained fromG. R. Greenberg, was used as a nonpermissive host foramber mutants (B. Dale, Ph.D. thesis, Univ. ofMichigan, Ann Arbor, 1968). E. coli BB and E. coliB021 were also used as nonpermissive hosts. Twoamber mutants in gene 59 of the T4 bacteriophage,T4amC5 and T4amHL628, were isolated and kindlysupplied by R. Edgar. Both amber mutants werepurified by crossing twice with wild-type T4D.
Media. The following medium was used in theisotope-labeling experiments and for growing bacteria(38). It was made by mixing 0.1 ml of 20% glucose, 0.2ml of 5% casein hydrolysates, and 10 ml of 1x saltsolution. Salt solution (50x) was prepared by dissolv-ing 10 g of MgSO4-7H2O, 100 g of citric acid-water,500 g of K2HPO4, and 175 g of NaNH4HPO4-4H20 in670 ml of water. This mixture was named lx CT-medium. The medium containing thymine or thymi-dine (5 pg/ml) was named 1x CT+ medium.
Chemical reagents. All radioisotopes were pur-chased from the New England Nuclear Corp.Methyl methanesulfonate (MMS) was obtained fromthe Sigma Company. Other reagents and chemicalswere standard grade products.
Preparation of the parental labeled DNA phage.A culture of E. coli was infected at a multiplicity of0.1 phage per bacterium in the presence of [methyl-3Hlthymine (50 ,Ci/ml, 2.1 Ci/mmol). After 5 h ofincubation, a few drops of chloroform was added andaeration was continued for an additional 10 min. Thislysate was purified by the standard method (1). Inmost cases, the isotope-labeled phage was used withina week.Growth and infection of cells. An overnight cul-
ture of E. coli B Tr201 was diluted 50 times with freshlx CT+ medium. The bacteria were grown at 37 Cuntil the optical density reached 0.8 at 590 nm on aZeiss spectrophotometer, corresponding to a viable
count of about 5 x 108 cells/ml. These cells werecentrifuged and resuspended in an equal volume of 1 xCT- medium in the presence of 20 Ag of D,L-trypto-phan per ml. Phage was added at a multiplicity of fiveparticles per bacterium. Three minutes after infec-tion, [14C]thymine (1 ,Ci/ml, 42 mCi/mmol) or[methyl-3H]thymine (10 sCi/ml, 0.42 Ci/mmol) wasadded.Measurement of kinetics of DNA synthesis. The
radioactivity in acid-insoluble fractions was deter-mined by pipetting 0.1 ml of a sample onto a 25-mmdisk of Whatman no. 3 filter paper. The disks werewashed for 10 min in three successive 200-ml volumesof 5% trichloroacetic acid followed by two successiverinses in 200 ml of 95% ethanol. This procedure is aslight modification of the procedure as described byBollum (4). The filters were then dried and placed invials containing 5 ml each of toluene phosphor [4 g of2,5-diphenyloxazole and 50 mg of 1,4-bis-2-(5-phenyl-oxazolyl) benzene per liter of toluene] and counted ina Nuclear Chicago liquid scintillation counter.UV irradiation. At various times after infection, a
2-ml sample of the infected culture was put on a petridish (4.25 cm in diameter) with shaking and wasirradiated at room temperature in a dark room.Irradiation was carried out using a 15-W germicidallamp (General Electric) at a distance of 10.5 inches.Under this condition, the UV dosage was 24 ergs/mm2per s as measured by a Black-Ray Ultra-violet Meter(model J-225, UV Products, Inc., San Gabriel, Calif.).Other conditions used are specified in the appropriatelegends.MMS treatment. The purified phage at a concen-
tration of 5 x 1010 PFU/ml which had been suspendedin 50 mM Tris-hydrochloride buffer (pH 8.0) wastreated with different concentrations of MMS. After20 min at 37 C, the phage suspension was diluted with9 volumes of cold 1x CT- medium and kept at 0 C,before infection. Other conditions used are describedin the appropriate legends.Lysing procedure. For alkaline sucrose gradient
centrifugation, the cells were suspended in a mixtureof 50 mM Tris, 50 mM NaCl, 50 mM EDTA, andlysozyme (100 jig/ml), pH 8.0, and chilled for 10 min.Triton X-100 was added to a concentration of 1%, and10 min later the DNA was denatured by addition ofNaOH to 0.4 N (24).
Sucrose gradients and centrifugation. Linear 5 to20% sucrose gradients were layered over 1 ml ofsaturated sucrose to improve recovery. Alkaline su-crose solutions were prepared in 0.05 M NaPO4 buffer(pH 12.2) in the presence of 0.015 M EDTA. Centrifu-gation was performed at 30,000 rpm for 60 min in aSW40 rotor of a Beckman model L2-65B preparativeultracentrifuge. The tubes were pierced and eightdrops per fraction were collected onto Whatman no. 3filter papers. After drying, the radioactivity of theacid-insoluble fractions was assayed.
Calculation. Molecular weight of DNA was calcu-lated according to the equation of Studier (35). Thenumber of nicks in DNA was estimated using themethod of Litwin et al. (21). T4 standard DNA wastaken as 123 x 106 for double-stranded DNA (19).
J. VIROL.
on July 18, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

REPAIR OF UV-IRRADIATED AND ALKYLATED DNA 7
RESULTSEffect on mutant progeny formation. (i)
Extracellular irradiation by UV light. Theeffect on progeny formation of mutant phage byUV irradiation before infection was examined.Purified mutant or wild-type phage particleswere suspended in phosphate-buffered saline ata concentration of 109 PFU/ml. The irradiatedphage T4amC5 were plated on E. coli B021.Survival fraction was calculated for the differ-ent time intervals of UV irradiation. The plat-ing efficiency of T4amC5 and T4amHL628 on
E. coli B was 0.5% as compared to 100% on E.coli CR63 before the irradiation (41). Our re-
sults clearly showed that both amber mutantsof gene 59 (amC5, amHL628) have higher sensi-tivity to UV irradiation when compared to thewild-type control (Fig. 1). It was shown thatwild-type phage particles require 864 ergs/mm2for 0.1% survival. The 0.1% survival dosages forT4amC5 and T4amHL628 were 576 and 502ergs/mm2, respectively. The ratio of 0.1% sur-
vival dosage between the mutants and wild typewas 0.66 and 0.58 for T4amC5 and T4amHL628,respectively. These values are close to otherUV-sensitive genes, x, y, and gene 32, accordingto the data reported by Harm (17) and Baldy(2). However, it was more sensitive than y
mutant which is also a known UV-sensitivemutant (J. R. Wu, unpublished data).
(ii) Intracellular irradiation by UV light.Studies on the effect of UV irradiation on
progeny formation showed that T4amHL628was also highly sensitive to UV (Table 1). Theculture of E. coli B was infected withT4amHL628 or T4am+, followed by irradiation9 min later. The progeny phage formed underthese conditions was determined by plating on
E. coli CR63, a permissive host, 20 and 180 minafter infection. The yield of phage was com-
pared to a nonirradiated control. The phageyield was about 10% of the control after 180 minin wild-type-infected cells, but it was less than0.03% of the control in mutant-infected cells.
Effect ofUV irradiation on kinetics ofDNAsynthesis. The initial rate of viral DNA synthe-sis of amber nonsense mutants of gene 59(amC5, amHL628) in a nonpermissive host, E.coli B, is indistinguishable from the wild-type-infected cells (41, 42). However, the mutantDNA synthesis is arrested completely after itsinitiation as judged from both pulse and contin-uous labeling by thymine (41). Figure 2 showsthe effects of UV irradiation on kinetics of viralDNA synthesis in T4amC5 and wild-type-infected cells. UV light was administered 2 min
z0
U. 6
CD)
.a3 T4D
1-4 |- T4jM[LL62 810
T\4S=C5
10 20 30 40 50
UV DOSE (SEC)FIG. 1. Survival of extracellular phage particles as
a function of UV dosage. After two cycles of low- andhigh-speed centrifugation, phage was suspended inphosphate-buffered saline (K2HPO4, 1.57 glliter;KH,P04, 0.41 glliter; NaCI, 7.36 glliter) and ad-justed to a titer of 109 PFU/ml. Two-millilitersamples were irradiated as described for the infectedculture in Materials and Methods. At various timesafter exposure, the irradiated phage stocks wereplated immediately in a nonpermissive host, E. coliB021. The surviving fraction was plotted against thetime of exposure to UV light. Survival fraction =(PFU [+UV] in E. coli B021)/(PFU [-UV] in E. coliB021).
after the viral DNA synthesis had been initi-ated. DNA synthesis in mutant-infected cellswas stopped immediately after an exposure ofUV light (600 ergs/mm2). The same result wasobtained when irradiation was administered 9min after infection. Under the same conditions,the wild-type-infected cells resumed the normalrate of synthesis after 1.5 min.We have described previously that arrested
VOL. 16, 1975
on July 18, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
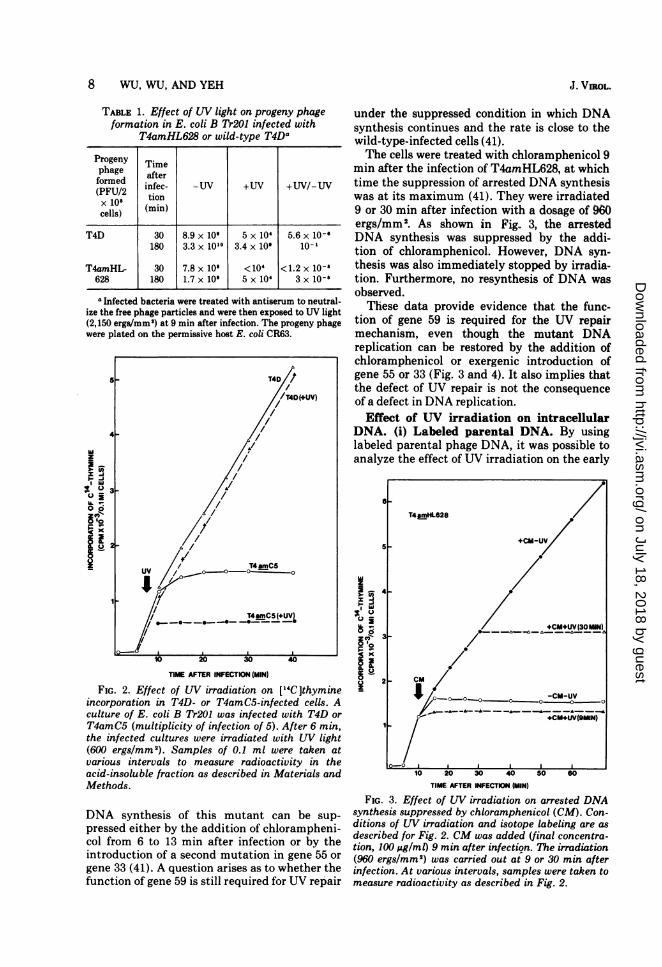
8 WU, WU, AND YEH
TABLE 1. Effect of UV light on progeny phageformation in E. coli B Tr201 infected with
T4amHL628 or wild-type T4Da
Progeny Timephage after
(forme infec- -UV +UV +UV/-UV(PU2 tionx 10', (min)cells)
T4D 30 8.9 x 109 5 x 10' 5.6 x 10-'180 3.3 x 10"° 3.4 x 10' 10-
T4amHL- 30 7.8 x 108 < 104 <1.2 x 10-'628 180 1.7 x 10' 5 x 104 3 x 10-'
aInfected bacteria were treated with antiserum to neutral-ize the free phage particles and were then exposed to UV light(2,150 ergs/mm') at 9 min after infection. The progeny phagewere plated on the permissive host E. coli CR63.
5 T4D/
1/T4D(+UV)
//~~~~~~
4A-
Kj 3
2-
FIG 2EfetoWiradito, nl4lhmn
incorporation ~ ~ ~ TinTD rT4 m5ifcte els
UV3
T4am5(mltipicit of nfetio o ).Ate mn
10- ~ ~
FIG.2inEfect oftue wer irradiation ont Wlhyight
inopoain nT4-o T T45jnfcte els
T4amC(muliplicty ofinfetionmof5(UfteV6J in
FtG.2.ece Effeturof UVr irradiationwion[LThyminet
(600 ergslmm2). Samples of 0.1 ml were taken atvarious intervals to measure radioactivity in theacid-insoluble fraction as described in Materials andMethods.
DNA synthesis of this mutant can be sup-pressed either by the addition of chlorampheni-col from 6 to 13 min after infection or by theintroduction of a second mutation in gene 55 or
gene 33 (41). A question arises as to whether thefunction of gene 59 is still required for UV repair
under the suppressed condition in which DNAsynthesis continues and the rate is close to thewild-type-infected cells (41).The cells were treated with chloramphenicol 9
min after the infection of T4amHL628, at whichtime the suppression of arrested DNA synthesiswas at its maximum (41). They were irradiated9 or 30 min after infection with a dosage of 960
ergs/mm2. As shown in Fig.. 3, the arrestedDNA synthesis was suppressed by the addi-tion of chloramphenicol. However, DNA syn-thesis was also immediately stopped by irradia-tion. Furthermore, no resynthesis of DNA wasobserved.These data provide evidence that the func-
tion of gene 59 is required for the UV repairmechanism, even though the mutant DNAreplication can be restored by the addition ofchloramphenicol or exergenic introduction ofgene 55 or 33 (Fig. 3 and 4). It also implies thatthe defect of UV repair is not the consequenceof a defect in DNA replication.Effect of UV irradiation on intracellular
DNA. (i) Labeled parental DNA. By usinglabeled parental phage DNA, it was possible toanalyze the effect of UV irradiation on the early
_a
x>ze
6 ;
x
a_2
10 20 30 40
TIME AFTER INFECTION (MIN)
FIG. 3. Effect of UV irradiation on arrested DNAsynthesis suppressed by chloramphenicol (CM). Con-ditions of UV irradiation and isotope labeling are asdescribed for Fig. 2. CM was added (final concentra-tion, 100 Ag/ml) 9 min after infection. The irradiation(960 ergs/mm2) was carried out at 9 or 30 min afterinfection. At various intervals, samples were taken tomeasure radioactivity as described in Fig. 2.
T4mHL628
+CM-UV
+CM+UV(30WMN)
/
I° 0 0°0~~O o...-Cm-UV
_A__ -&- - _-_.---&-- _-_
+CM.NJV(9tMN)
I I tZ
J. VIROL.
on July 18, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

VO1,15REPAIR OF UV-IRRADIATED AND ALKYLATED DNA
z
-6.WjE
i X8
FIG. 4incorporT4amBIiCi/mI,infectiorat 15 mimeasureA, T4ar(+UV);(+UV).
replicasedimedenatualkalinIrradiation atcorresp
Pare]fourthP, eqgradierthe disstandaimutantUV-daiual incshowncontrasvarioussame nrepairunder t
(ii)]IDNA['H Ith3
cold thymine per ml immediately after irradia-tion. More than 60% of the [3H ]thymine-labeled DNA in both mutant and wild-typephage-infected cells was fragmented to a molec-
/. A ular weight of less than 12 million (D,/D1 =0.53) in alkaline sucrose gradients (Fig. 6). The
oOZ degraded DNA strands in the wild-type-in-fected cells probably undergo repair by theaction of enzymes such as DNA polymerases
- uv /oU and nucleases and then are finally joined bypolynucleotide ligase. After 60 min, the sedi-mentation of UV-damaged DNA coincided with
/// ^ the reference T4 DNA. However, the mutant-infected cells could not repair the UV-damagedDNA (Fig. 6).Sedimentation profiles of zonal centrifuga-
tion of both parental and newly synthesizedDNA gave consistent data showing that mutantDNA was hydrolyzed immediately from one-fourth to one-sixth of the original molecular
lo 20 30 40 weight and that the rejoining of DNA did notTMr AFTER IW(ECT4N(M) occur. Our preliminary analysis of thymine
dimers indicated that the dimers were excised4. Effect of LW irradiation on [HH]thymine and excision of UV-damaged DNA are notmation in T4amC5amBL292 (gene 59-55)- or defective for the mutant. Rather, the mutant isL292 (gene 55)-infected cells. ['HWthymine (10 defective fore polynuleothe ortantps0.42 Ci/mmol) was added at 3 min after defective in reJoiningofpolynucleotides orstepss followed by UV irradiation (960 ergs/mm2) after the excision of thymine dimers.n. At various intervals, a sample was taken to Sensitivity of T4amC5 (gene 59) to alkylat-radioactivity as described in Fig. 2. Symbols: ing agent. Our simple assumption for the repairmC5amBL292 (-UL); A, T4amC5amBL292 mechanisms of damage caused by UV and0, T4amBL292 (-UV); 0, T4amBL292 alkylating agents is that not all mechanisms
acting on UV damages will repair damagescaused by alkylating agents and that each
tive intermediates. Figure 5 shows the mechanism involves several gene products (36).,ntation analysis of [3H ]thymine-labeled To determine whether the function of gene 59 isred parental phage DNA in 5 to 20% also involved in repair of damage caused byte sucrose gradients after UV irradiation. the alkylating agent, the following experimentLtion was carried out 6.5 min after infec- was performed. Purified T4amC5 or wild-typea dosage of 360 ergs/mm2 (one lethal hit phage particles were treated with 20 mM MMSponds to a dosage of 144 ergs/mm2). at 1010 PFU/ml suspended in 50 mM Tris-ntal phage DNA was fragmented to one- hydrochloride buffer, pH 8.0. At various timeof its molecular weight (18 x 106, D/D, intervals after the treatment, the phage wasjuals the distance from the top of the plated on E. coli B. The kinetics of survivingit of the intracellular DNA and Dl equals fractions as a function of duration of exposure to,tance from the top of the gradient of the MMS is shown in Fig. 7. The survival of 1% forrd phage DNA molecules] = 0.608) in the mutant required an exposure time of only 19t- and wild-type-infected cells. Repair of min, as compared with 50 min for that of themaged DNA was evidenced by the grad- wild-type phage. This result shows thatrease in size of single-stranded DNA, as T4amC5 (gene 59) at the cellular level exhibitsin wild-type-infected cells (Fig. 5). In much greater susceptibility to inactivation
at, the mutant parental DNA, isolated at caused by a monofunctional alkylating agent. In3 times after irradiation, retained the addition, we have also plated MMS-treatednolecular weight (Fig. 5). Evidently, no T4amC5 on E. coli CR63, a strain carryingoccurred on the mutant parental DNA suppressor (SuI) for nonsense mutation. Essen-this condition. tially, no difference in the amount of survivingNewly synthesized viral DNA. The fractions after treatment was observed betweenwas labeled from 3 to 18 min with T4amC5 and wild type in this strain (Fig. 7).ymine and then chased with 1.5 mg of The increased sensitivity of gene 59 mutant to
9VOL. 16, 1975
on July 18, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

10 WU, WU, AND YEH
T4D
10 20 30BOTTOM
I
IC.)lf'U
10 20 304I BOTTOM
FRACTION NUMBERFIG. 5. Effect of UV irradiation on sedimentation profiles of parental phage DNA ([3H]thymine labeled)
after infection. The LTV irradiation (360 ergs/mm2) was carried out at 6.5 min after infection. DNA was ex-tracted and followed by centrifugation in 5 to 20%o alkaline sucrose gradient as described in Materials andMethods. Samples were taken at 7 (0), 20 (0), and 30 min (U). 0, "4C-labeled reference T4 DNA.
alkylation was apparently suppressed by E. coliCR63. This result supports the hypothesis thatthe defect in repair of alkylated DNA is due tononsense mutation in gene 59.
Effect of alkylating agent on mutant DNAreplication. We have studied the effect ofMMS on kinetics of DNA replication. Thisexperiment was feasible because DNA replica-tion has been shown to be normal up to 2 to 3min after it is initiated under a nonpermissivecondition (41). T4amC5 (gene 59) or T4am+ wasincubated with different concentrations ofMMS at 37 C for 20 min. A culture of E. coliB Tr201 was infected at a multiplicity of infec-tion of 5 with the MMS-treated phages. Asshown in Fig. 8, incorporation of [8H ]thyminein T4amC5-infected cells, namely, replicationof mutant DNA, was affected drastically afterMMS treatment. Total amounts of DNA syn-thesized were reduced, possibly because thegrowing point for DNA replication may bestopped without further repair at or near
the alkylated lesions. At a concentration of 2mM MMS, the effect of MMS on DNAsynthesis was evident in mutant-infected cells.When the concentration of MMS was increasedto 0.25 M, the amount of DNA synthesizedwas less than 20% of controls without MMStreatment. In contrast, the rate of DNA syn-thesis of wild-type phage which had beentreated with 0.25 M MMS became normalafter a lag period of 20 min. These resultsagain suggest that T4amC5 (gene 59)-infectedcells were defective in the repair function ofsingle-stranded DNA breaks which are pre-sumably the main result of alkylation.Repair of intracellular DNA after MMS
treatment. An analysis of intracellular DNA inalkaline sucrose gradients should reflect thedegree of breakage induced by the alkylatingagent and rejoining of DNA single strands.We performed the following experiments de-signed to give more direct evidence of theeffect of MMS at the molecular level. A culture
II I
1
Xc
0C.)cc
I-
'U0mall'U
J. VIROL.
on July 18, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
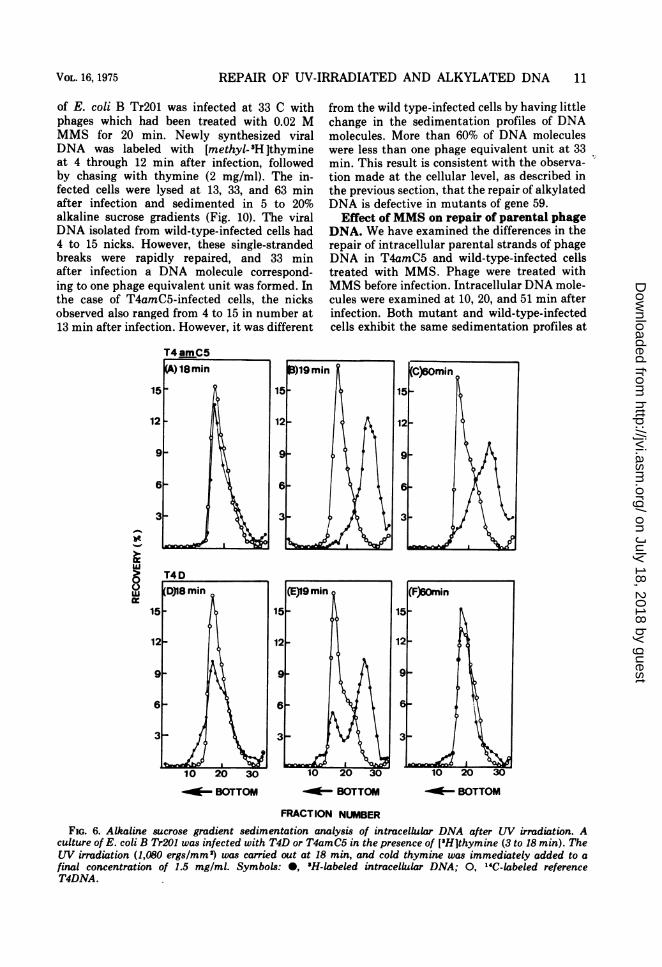
REPAIR OF UV-IRRADIATED AND ALKYLATED DNA 11
of E. coli B Tr201 was infected at 33 C withphages which had been treated with 0.02 MMMS for 20 min. Newly synthesized viralDNA was labeled with [methyl-PH Ithymineat 4 through 12 min after infection, followedby chasing with thymine (2 mg/ml). The in-fected cells were lysed at 13, 33, and 63 minafter infection and sedimented in 5 to 20%alkaline sucrose gradients (Fig. 10). The viralDNA isolated from wild-type-infected cells had4 to 15 nicks. However, these single-strandedbreaks were rapidly repaired, and 33 minafter infection a DNA molecule correspond-ing to one phage equivalent unit was formed. Inthe case of T4amC5-infected cells, the nicksobserved also ranged from 4 to 15 in number at13 min after infection. However, it was different
-
uJ
from the wild type-infected cells by having littlechange in the sedimentation profiles of DNAmolecules. More than 60% of DNA moleculeswere less than one phage equivalent unit at 33min. This result is consistent with the observa-tion made at the cellular level, as described inthe previous section, that the repair of alkylatedDNA is defective in mutants of gene 59.
Effect of MMS on repair of parental phageDNA. We have examined the differences in therepair of intracellular parental strands of phageDNA in T4amC5 and wild-type-infected cellstreated with MMS. Phage were treated withMMS before infection. Intracellular DNA mole-cules were examined at 10, 20, and 51 min afterinfection. Both mutant and wild-type-infectedcells exhibit the same sedimentation profiles at
- BOTTOM - BOTTOM - BOTTOM
FRACTION NUMBERFIG. 6. Alkaline sucrose gradient sedimentation analysis of intracellular DNA after LW irradiation. A
culture of E. coli B Tr201 was infected with T4D or T4amC5 in the presence of ['H]thymine (3 to 18 min). TheLW irradiation (1,080 ergs/mm2) was carried out at 18 min, and cold thymine was immediately added to afinal concentration of 1.5 mg/mI. Symbols: 0, 'H-labeled intracellular DNA; 0, "IC-labeled referenceT4DNA.
VOL. 16, 1975
on July 18, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

12 WU, WU, AND YEH
z0
U.
0
z
'I
cc
10 20 30 40 50 60
TIME(MIN)FIG. 7. Sensitivity of T4amC5 or T4D phage parti-
cles to MMS treatment. Phage T4 was incubated at37 C at a concentration of 1010 PFU/ml in 50 mMTris-hydrochloride buffer (pH 8.0) containing 20 mMMMS. At various times after incubation, the MMS-treated phages were plated on E. coli BB or E. coliCR63. Survival fraction = (PFU [+MMS] in E. coliBB)/(PFU [-MMS] in E. coli BB).
11 min after infection. The mutant-infectedcells failed to repair the MMS-damaged paren-tal DNA, whereas the wild-type parental DNAmolecules were gradually repaired to a normalsize (63S) at 51 min after infection (Fig. 11). Weinterpret these results as meaning that repair ofalkylated lesions in both newly synthesized andparental DNA is greatly reduced by a mutationin gene 59.
DISCUSSIONIt is known that phage T4 controls two
processes for repairing its UV-damaged DNA(23, 36). The excision repair process is thoughtto consist of four steps: (i) incision by v geneproduct, a thymine dimer-specific endonucle-ase; (ii) excision of dimer as part of an oligonu-cleotide by DNA polymerase I; (iii) resynthesisof new complementary strands by DNA polym-
erase I; (iv) rejoining of newly synthesizedfragments to the parental strand by polynucleo-tide ligase (gene 30). T4 genes 1 (hydroxymeth-yl-dCMP kinase), 42 (dCMP hydroxymethyl-ase), 45 (unknown function), and 56 (dCTPpyrophosphatase) are also shown to be involvedin this process (36). The mechanism for thereplication repair process is still unknown.However, this process reportedly involves genesx, y, 43 (DNA polymerase), 32 (DNA unwindingprotein), 30, 1, 41, (unknown function), 42, 44(unknown function), 45, and 56 (36). Genes xand y mutants are sensitive to UV, X-ray, andalkylating agents (25). Among these genes, gene30 is required for the repair of UV-damaged (2,36) and alkylated DNA molecules (3, 27). Gene43 is required for the repair of alkylated DNAmolecules (27); however, the requirement forUV repair is still uncertain (2, 29).The results of this study indicate that gene 59
3
za1- ;2x
J
aaxI X.
z
TIME AFTER INFECTION (MIN)
FIG. 8. Effect of MMS treatment on [methyl-'HJthymine incorporation in T4amC5-infected cells.A suspension of T4amC5 was incubated with differentconcentrations of MMS in 50mM Tris-hydrochloridebuffer, pH 8.0, at 37 C for 20 min. A culture of E. coliB Tr201 was infected with MMS-treated T4amC5(multiplicity of infection of 5) with aeration at 37Cfor 5 min. The infected cells were centrifuged andresuspended in I x CT- medium containing [methyl-3H]thymine (10 /uCi/ml, 0.42 Ci/mmol). Samples of0.1 ml were withdrawn at various intervals for count-ing as described in Materials and Methods. Symbols:0, without MMS; 0, 2 x 10-'M MMS; 0, 10- 2 M
MMS;A, 5 x 10-2MMMS; V, 2.5 x 10- MMMS.
T4 am CSRESIJSPENSION
/ _v v7- 9
J. VIROL.
on July 18, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

VREPAIROF UV-IRRADIATED AND ALKYLATED DNA
is involved in the replication repair process. Ourconclusion is based on the following results: (i)T4amC5 (gene 59) is sensitive to both UVirradiation and MMS treatment; (ii) the resultsof alkaline sucrose gradient sedimentation ofintracellular mutant DNA, immediately afterexposure to UV, showed that DNA moleculeswere fragmented to the same size as that of wildtype (Fig. 5.and 6). These results indicate thatincision of the UV-irradiated DNA, presumablycatalyzed by a v gene product, is normal in themutant-infected cells; (iii) gene 59 is involved innormal phage DNA replication (41, 42; Wu andYeh, Bacteriol. Proc., p. 167, 1970). It has beenreported that genes 1, 30, 42, 45, and 56 areinvolved in both excision and replication repairprocesses (36). From our previous and presentdata we cannot exclude the possibility that gene59 is also involved in both excision and replica-tion repair processes.
Genetically, gene 59 is continguous to gene32. Their functions are required for replicationrepair and DNA replication, but their roles inthe replication repair mechanism probably isdifferent. It has been shown that a gene 32product is required in stoichiometric amountsfor phage growth (33) and plays a unique role inthe repair mechanism (40). It has been postu-lated that the possible mode of action of un-winding protein (gene 32) in UV repair is thatthis protein protects the excised area (gaps)from single-stranded nucleases, which, in ef-fect, preserves the structure which can berepaired by proper DNA polymerases (40; B.Alberts, personal communication).We have examined whether or not the defect
in repair of UV-induced lesions is a simpleconsequence of arrest in DNA replication. Asshown in Fig. 3 and 4, even though the arrestedDNA synthesis is restored to the normal by theaddition of chloramphenicol or by the introduc-tion of exogenic mutation, the repair of UV-induced lesions is still defective. We have alsoisolated a late mutant, darl, which specificallyrestores the arrested DNA synthesis of gene 59mutant. Phage DNA synthesis is restored to thenormal level in darl-amC5 double mutant-infected cells but UV sensitivity caused by gene59 mutation is not restored (J. R. Wu and Y. C.Yeh, Cold Spring Harbor DNA ReplicationMeet., p. 66, 1974). In short, darl suppresses thedefect in DNA replication but does not suppressthe defect in UV repair of amC5. We concludefrom these results that the defect in repair ofUV-damaged DNA is not a simple consequenceof arrested DNA replication, but rather implies
14 /
12-
10 1
z
XL)wj -j 8
0I. 7 A
TIE AFTERINFECTIU(MNINO)
FIG. 9. Effect of MMS treatment on [methyl-'H]thymine incorporation in T4D-infected cells. Thecondition was identical to that described in Fig. 8.Symbols: 0, without MMS; 0, 10-2 M MMS; A,5 x 10-2MMMS;V, 2.5 x 10- 1MMMS.
that gene 59 plays an independent role in repairsynthesis. Therefore, the function of gene 59 isrequired for DNA replication as well as repair ofUV lesions.
Considerable information is available aboutthe repair process of UV damage. However, lessis known about the repair process of chemicallydamaged DNA. In this study, MMS, a mono-functional alkylating agent, was used to investi-gate the involvement of gene 59 in repair ofalkylated DNA. It has been reported that alkyl-ation occurs at the purine bases, particularly atN-7 of guanine and N-3 of adenine (7, 8, 11, 20).Depurination followed because of the unstablealkylated purines. The rate of N-3 alkylade-nine loss is about six times the rate of loss ofN-7 alkylguanine. The amount of N-3 alk-yladenine is approximately one-tenth of the N-7alkylguanine; therefore, depurination occursapproximately equally on guanine and adenine.These apurinic acid sites are converted to sin-gle-stranded breaks by a ,B-elimination process(20, 34) and by enzymatic actions (13). Alkaline
T4D
VOL. 16, 1975 13
on July 18, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
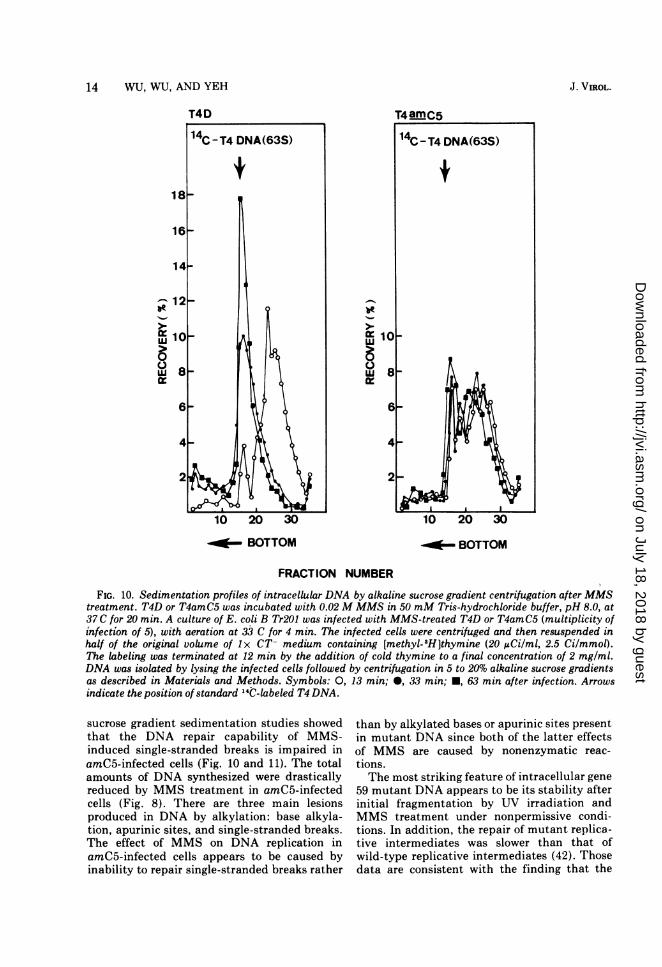
14 WU, WU, AND YEH
I-
I-,
C.)w
T4D
I--
wC.)r
...ilf- BOTTOM ~'- BOTTOM
FRACTION NUMBER
FIG. 10. Sedimentation profiles of intracellular DNA by alkaline sucrose gradient centrifugation after MMStreatment. T4D or T4amC5 was incubated with 0.02 M MMS in 50 mM Tris-hydrochloride buffer, pH 8.0, at37 C for 20 min. A culture of E. coli B Tr201 was infected with MMS-treated T4D or T4amC5 (multiplicity ofinfection of 5), with aeration at 33 C for 4 min. The infected cells were centrifuged and then resuspended inhalf of the original volume of 1x CT- medium containing [methyl-3H]thymine (20 AiCi/ml, 2.5 Ci/mmol).The labeling was terminated at 12 min by the addition of cold thymine to a final concentration of 2 mg/ml.DNA was isolated by lysing the infected cells followed by centrifugation in 5 to 20% alkaline sucrose gradientsas described in Materials and Methods. Symbols: 0, 13 min; 0, 33 min; *, 63 min after infection. Arrowsindicate the position of standard 14C-labeled T4 DNA.
sucrose gradient sedimentation studies showedthat the DNA repair capability of MMS-induced single-stranded breaks is impaired inamC5-infected cells (Fig. 10 and 11). The totalamounts of DNA synthesized were drasticallyreduced by MMS treatment in amC5-infectedcells (Fig. 8). There are three main lesionsproduced in DNA by alkylation: base alkyla-tion, apurinic sites, and single-stranded breaks.The effect of MMS on DNA replication inamC5-infected cells appears to be caused byinability to repair single-stranded breaks rather
than by alkylated bases or apurinic sites presentin mutant DNA since both of the latter effectsof MMS are caused by nonenzymatic reac-tions.The most striking feature of intracellular gene
59 mutant DNA appears to be its stability afterinitial fragmentation by UV irradiation andMMS treatment under nonpermissive condi-tions. In addition, the repair of mutant replica-tive intermediates was slower than that ofwild-type replicative intermediates (42). Thosedata are consistent with the finding that the
J. VIROL.
on July 18, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
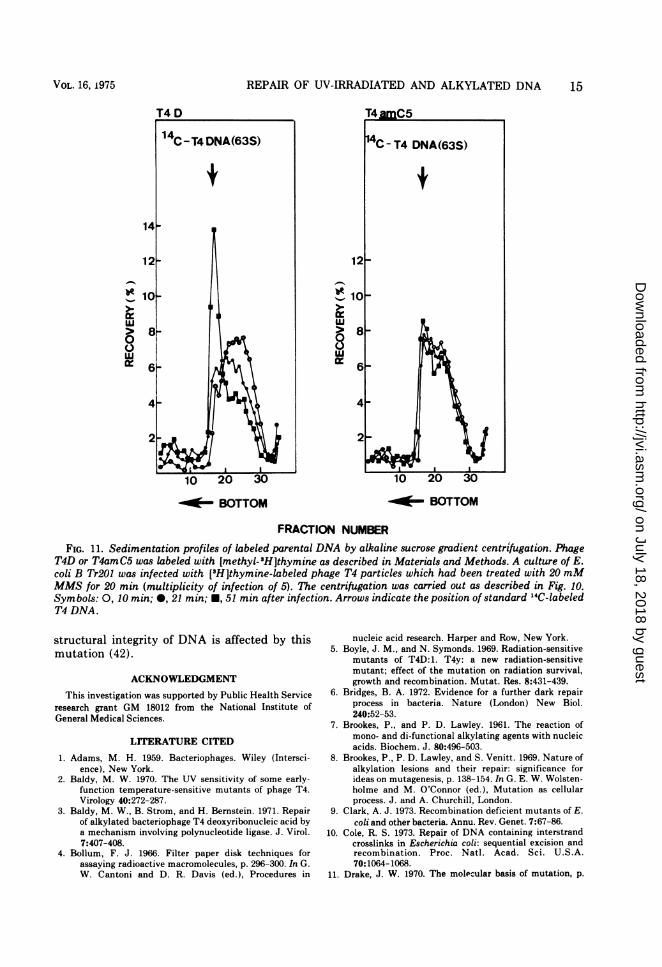
REPAIR OF UV-IRRADIATED AND ALKYLATED DNA
1-
uIII
00L:
'-I
II
8cz
-OE- BOTTOM .~u~ BOTTOM
FRACTION NUMBER
FIG. 11. Sedimentation profiles of labeled parental DNA by alkaline sucrose gradient centrifugation. PhageT4D or T4amC5 was labeled with [methyl-3H]thymine as described in Materials and Methods. A culture of E.coli B Tr201 was infected with [3H]thymine-labeled phage T4 particles which had been treated with 20 mMMMS for 20 min (multiplicity of infection of 5). The centrifugation was carried out as described in Fig. 10.Symbols: 0, 10 min; 0, 21 min; *, 51 min after infection. Arrows indicate the position of standard "4C-labeledT4 DNA.
structural integrity of DNA is affected by thismutation (42).
ACKNOWLEDGMENT
This investigation was supported by Public Health Serviceresearch grant GM 18012 from the National Institute ofGeneral Medical Sciences.
LITERATURE CITED
1. Adams, M. H. 1959. Bacteriophages. Wiley (Intersci-ence), New York.
2. Baldy, M. W. 1970. The UV sensitivity of some early-function temperature-sensitive mutants of phage T4.Virology 40:272-287.
3. Baldy, M. W., B. Strom, and H. Bernstein. 1971. Repairof alkylated bacteriophage T4 deoxyribonucleic acid bya mechanism involving polynucleotide ligase. J. Virol.7:407-408.
4. Bollum, F. J. 1966. Filter paper disk techniques forassaying radioactive macromolecules, p. 296-300. In G.W. Cantoni and D. R. Davis (ed.), Procedures in
nucleic acid research. Harper and Row, New York.5. Boyle, J. M., and N. Symonds. 1969. Radiation-sensitive
mutants of T4D:1. T4y: a new radiation-sensitivemutant; effect of the mutation on radiation survival,growth and recombination. Mutat. Res. 8:431-439.
6. Bridges, B. A. 1972. Evidence for a further dark repairprocess in bacteria. Nature (London) New Biol.240:52-53.
7. Brookes, P., and P. D. Lawley. 1961. The reaction ofmono- and di-functional alkylating agents with nucleicacids. Biochem. J. 80:496-503.
8. Brookes, P., P. D. Lawley, and S. Venitt. 1969. Nature ofalkylation lesions and their repair: significance forideas on mutagenesis, p. 138-154. In G. E. W. Wolsten-holme and M. O'Connor (ed.), Mutation as cellularprocess. J. and A. Churchill, London.
9. Clark, A. J. 1973. Recombination deficient mutants of E.coli and other bacteria. Annu. Rev. Genet. 7:67-86.
10. Cole, R. S. 1973. Repair of DNA containing interstrandcrosslinks in Escherichia coli: sequential excision andrecombination. Proc. Natl. Acad. Sci. U.S.A.70:1064-1068.
11. Drake, J. W. 1970. The molecular basis of mutation, p.
VOL. 16, 1975 15
on July 18, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

16 WU, WU, AND YEH J
146-159. Holden-Day, San Francisco.12. Epstein, R. H., A. Bolle, C. M. Steinberg, E. Kellen-
berger, E. Boy de la Tour, R. Chevalley, R. S. Edgar,M. Susman, G. H. Denhardt, and A. Lielausis. 1963.Physiological studies of conditional lethal mutants ofbacteriophage T4D. Cold Spring Harbor Symp. Quant.Biol. 28:375-394.
13. Friedberg, E. C., and D. Goldthwait. 1969. EndonucleaseII of E. coli. L. Isolation and purification. Proc. Natl.Acad. Sci. U.S.A. 62:934-940.
14. Friedberg, E. C., and J. J. King. 1969. Endonucleolyticcleavage of UV-irradiated DNA controlled by the V+gene in phage T4. Biochem. Biophys. Res. Commun.37:646-651.
15. Friedberg, E. C., and J. J. King. 1971. Dark repair ofultraviolet-irradiated deoxyribonucleic acid by bacte-riophage T4: purification and characterization of a
dimer-specific phage-induced endonuclease. J. Bacte-riol. 106:500-507.
16. Hanawalt, P. C. 1971. Repair replication of damagedDNA in vivo, p. 528-547. In 23rd Symp. Fundam.Cancer Res. The Williams and Wilkins Co., Baltimore.
17. Harm, W. 1963. Mutants of phage T4 with increasedsensitivity to ultraviolet. Virology 19:66-71.
18. Kelly, R. B., M. R. Atkinson, J. A. Huberman, and A.Kornberg. 1969. Excision of thymine dimers and othermismatched sequences by DNA polymerase of Esche-richia coli. Nature (London) 224:495-501.
19. Kim, J. S., and N. Davidson. 1974. Electron microscopeheteroduplex study of sequence relations of T2, T4 andT6 bacteriophage DNAs. Virology 57:93-111.
20. Lawley, P. D. 1966. Effects of some chemical mutagensand carcinogens on nucleic acids. Prog. Nucleic AcidRes. Mol. Biol. 5:89-131.
21. Litwin, S., E. Shahn, and A. W. Kozinski. 1969. Interpre-tation of sucrose gradient sedimentation pattern ofdeoxyribonucleic acid fragments resulting from ran-
dom breaks. J. Virol. 4:24-30.22. Lohman, P. H. M., and D. Bootsma. 1974. Repair
mechanisms in mammalian cells. Mutat. Res.23:147-150.
23. Maynard-Smith, S., and N. Symonds. 1973. Involvementof bacteriophage T4 genes in radiation repair. J. Mol.Biol. 74:33-44.
24. Miller, R. C., Jr., A. W. Kozinski, and S. Litwin. 1970.Molecular recombination in T4 bacteriophage deoxyri-bonucleic acid. III. Formation of long single strandsduring recombination. J. Virol. 5:368-380.
25. Mortelmans, K., and E. C. Friedberg. 1972. Deoxyribonu-cleic acid repair in bacteriophage T4: observations on
the roles of the X and V genes and of host factors. J.Virol. 10:730-736.
26. Radding, C. M. 1973. Molecular mechanisms in geneticrecombination. Annu. Rev. Genet. 7:87-111.
27. Ray, U., L. Bartenstein, and J. W. Drake. 1972. Inactiva-
tion of bacteriophage T4 by ethyl methanesulfonate:influence of host and viral genotypes. J. Virol.9:440-447.
28. Rupp, W. D., and P. Howard-Flanders. 1968. Discon-tinuities in the DNA synthesized in an excision-defec-tive strain of Escherichia coli following ultravioletirradiation. J. Mol. Biol. 31:291-304.
29. Schnitzlein, C. F., I. Albrecht, and J. W. Drake. 1974. Isbacteriophage T4 DNA polymerase involved in therepair of ultraviolet damage? Virology 59:580-583.
30. Setlow, R. B. 1968. The photochemistry, photobiology,and repair of polynucleotides. Prog. Nucleic Acid Res.Mol. Biol. 8:257-295.
31. Setlow, R. B., and W. L. Carrier. 1966. Pyrimidine dimersin ultraviolet-irradiated DNA's. J. Mol. Biol.17:237-254.
32. Smith, S. M., and N. Symonds. 1973. The unexpectedlocation of a gene conferring abnormal radiation sensi-tivitv on phage T4. Nature (London) 241:395-396.
33. Snustad, D. P. 1968. Dominance interactions in Esche-richia coli cells mixedly infected with bacteriophageT4D wild-type and amber mutants and their possibleimplications as to type of gene-product function: cata-lytic vs. stoichiometric. Virology 35:550-563.
34. Strauss, B., M. Coyle, and M. Robbins. 1968. Alkylationdamage and its repair. Cold Spring Harbor Symp.Quant. Biol. 33:277-287.
35. Studier, F. W. 1965. Sedimentation studies on the sizeand shape of DNA. J. Mol. Biol. 11:373-390.
36. Symonds, N., H. Heindl, and P. White. 1973. Radiationsensitive mutants of phage T4. A comparative study.Mol. Gen. Genet. 120:253-259.
37. Taketo, A., S. Yasuda, and M. Sekiguchi. 1972. Initialstep of excision repair in Escherichia coli: replace-ment of defective function of uvr mutants by T4endonuclease v. J. Mol. Biol. 70:1-14.
38. Vogel, H. J., and D. M. Bonner. 1956. Acetylornithinaseof Escherichia coli: partial purification and someproperties. J. Biol. Chem. 218:97-106.
39. Witkin, E. M. 1969. Ultraviolet-induced mutation andDNA repair. Annu. Rev. Genet. 3:525-552.
40. Wu, J. R., and Y. C. Yeh. 1973. Requirement of afunctional gene 32 product of bacteriophage T4 in UVrepair. J. Virol. 12:758-765.
41. Wu, R., F. Y. Ma, and Y. C. Yeh. 1972. Suppression ofDNA-arrested synthesis in mutants defective in gene59 of bacteriophage T4. Virology 47:147-156.
42. Wu, R. and Y. C. Yeh. 1974. DNA arrested mutants ofgene 59 of bacteriophage T4. II. Replicative intermedi-ates. Virology 59:108-122.
43. Yasuda, A., and M. Sekiguchi. 1970. Mechanism of repairof DNA in bacteriophage. II. Inability of ultraviolet-sensitive strains of bacteriophage in inducing an en-zyme activity to excise pyrimidine dimers. J. Mol. Biol.47:243-255.
J. VIROL.
on July 18, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from


















