
Rifomycin - Semantic Scholar · after the extraction, purification, and crystallization of this...
11
Rifomycin XIV. Production of Rifomycin B P. MARGALITH' AND H. PAGANI Research Laboratories Lepetit S.p.A., Milan, Italy Received for publication October 14, 1960 ABSTRAkCT MARGALITH, P. (Lepetit S.p.A., Milan, Italy), AND H. PAGANI. Rifomycin. XIV. Production of rifomycin B. Appl. Microbiol. 9:325-334. 1961.-The production of a fraction of the rifomycin complex (rifomycin B) by Streptomyces mediterranei has been described. Both qualitative and quantitative aspects of the fermentation are determined by suitable agitation-aeration param- eters. In addition to conventional nutrient ingredi- ents, the presence of some of the barbituric acid derivatives was found to be essential for rifomycin B production. An attempt was made to explain the mode of action of these compounds. In the foregoing papers of this series the biological characteristics of Streptomyces mediterranei and the production of rifomycin complex (a mixture of at least five different antibiotic principles) were described (Margalith and Beretta, 1960; Margalith and Pagani, 1961). It has been pointed out that for various reasons the production of the complex met with great difficulty both on laboratory and pilot plant scale. Attempts to isolate strains of S. mediterranei that would produce more abundantly one of the fractions of the complex were relatively successful, insofar as the prevalent production of fractions C and D could be promoted. In this paper the fermentation of S. mediter- ranei for the sole production of fraction B of the com- plex (rifomycin B) will be described. MATERIALS AND METHODS Materials. All synthetic compounds employed in this research were of commercial origin. Fermentation. Essentially the same procedure for inoculum and vegetative growth were employed as for the production of rifomycin complex (Margalith and Pagani, 1961). The main modifications were in the composition of the media. Mycelial growth. Samples of 10-ml mash were cen- trifuged at 3,000 rev/min for 10 min and the volume of mycelium was determined. Growth is expressed as percentage of mycelial volume (pmv). 1 Present address: Department of Microbiology, Israel Insti- tute of Technology, Technion, Haifa, Israel. Antibiotic activity. Unless stated otherwise, rifomycin B assays were carried out spectrophotometrically since the presence of small amounts of other fractions of the complex with different antibiotic activities made the microbiological assay difficult in interpretation. The spectrophotometric method for the assay of rifomycin B in fermentation broths has been described by Gallo, Sensi, and Radaelli (1960). Furthermore, uniformity in rifomycin production was established by observing the extinction curve of the ethyl acetate extract when oxidized by shaking with a potassium ferricyanide solution. The appearance of a shoulder peak at 460 my was considered as indication for the presence of frac- tions C and D (Sensi, Greco, and Ballotta, 1960). Use of chemically defined stimulating substances. Strain selection of the rifomycin complex-producing streptomyces resulted in the isolation of a strain that produced fractions C and D in prevalent amounts. This made us believe that on further screening a strain that would synthetize predominantly one of the other components of the complex would be eventually iso- TABLE 1. Composition of media and rifomycin B yields Nutrient ingredients (g/l,OOO ml) ASS/6* ASS/7 ASS/8 Glucose 50 70 70 Soya bean meal 5 15 5 Phaseolus beanst (Varese 20 type) (NH4)2SO4 7 7 7 MgSO4*7H20 1 1 1 KH2PO4 3 2 2 CaCO3 9 6 6 Diethylbarbiturate 2 2 2 CuSO 4 * 5H20 0.0033 0.0033 0.0033 FeSO4-7H20 0.010 0.010 0.010 ZnSO4*7H20 0.050 0.050 0.050 MnSO4-4H20 0.004 0.004 0.004 CoCl*26H20 - 0.002 0.002 (NH4)2MoO 4 0.001 0.001 Avg rifomycin B potency 200-250 400-450 650-800 obtained (ug/ml) pH was adjusted to 7.0 with 40% NaOH. Sterilization 30 min at 121 C. * Medium ASS/4 like ASS/6 but without barbital. t Phaseolus beans were cooked for 30 min before addition to medium. 325 on October 15, 2017 by guest http://aem.asm.org/ Downloaded from on October 15, 2017 by guest http://aem.asm.org/ Downloaded from on October 15, 2017 by guest http://aem.asm.org/ Downloaded from
Transcript of Rifomycin - Semantic Scholar · after the extraction, purification, and crystallization of this...

RifomycinXIV. Production of Rifomycin B
P. MARGALITH' AND H. PAGANI
Research Laboratories Lepetit S.p.A., Milan, Italy
Received for publication October 14, 1960
ABSTRAkCT
MARGALITH, P. (Lepetit S.p.A., Milan, Italy), ANDH. PAGANI. Rifomycin. XIV. Production of rifomycinB. Appl. Microbiol. 9:325-334. 1961.-The productionof a fraction of the rifomycin complex (rifomycin B)by Streptomyces mediterranei has been described. Bothqualitative and quantitative aspects of the fermentationare determined by suitable agitation-aeration param-eters. In addition to conventional nutrient ingredi-ents, the presence of some of the barbituric acidderivatives was found to be essential for rifomycin Bproduction. An attempt was made to explain the modeof action of these compounds.
In the foregoing papers of this series the biologicalcharacteristics of Streptomyces mediterranei and theproduction of rifomycin complex (a mixture of at leastfive different antibiotic principles) were described(Margalith and Beretta, 1960; Margalith and Pagani,1961). It has been pointed out that for various reasonsthe production of the complex met with great difficultyboth on laboratory and pilot plant scale.Attempts to isolate strains of S. mediterranei that
would produce more abundantly one of the fractions ofthe complex were relatively successful, insofar as theprevalent production of fractions C and D could bepromoted. In this paper the fermentation of S. mediter-ranei for the sole production of fraction B of the com-plex (rifomycin B) will be described.
MATERIALS AND METHODS
Materials. All synthetic compounds employed inthis research were of commercial origin.
Fermentation. Essentially the same procedure forinoculum and vegetative growth were employed as forthe production of rifomycin complex (Margalith andPagani, 1961). The main modifications were in thecomposition of the media.
Mycelial growth. Samples of 10-ml mash were cen-trifuged at 3,000 rev/min for 10 min and the volumeof mycelium was determined. Growth is expressed aspercentage of mycelial volume (pmv).
1 Present address: Department of Microbiology, Israel Insti-tute of Technology, Technion, Haifa, Israel.
Antibiotic activity. Unless stated otherwise, rifomycinB assays were carried out spectrophotometrically sincethe presence of small amounts of other fractions of thecomplex with different antibiotic activities made themicrobiological assay difficult in interpretation. Thespectrophotometric method for the assay of rifomycinB in fermentation broths has been described by Gallo,Sensi, and Radaelli (1960). Furthermore, uniformityin rifomycin production was established by observingthe extinction curve of the ethyl acetate extract whenoxidized by shaking with a potassium ferricyanidesolution. The appearance of a shoulder peak at 460 mywas considered as indication for the presence of frac-tions C and D (Sensi, Greco, and Ballotta, 1960).
Use of chemically defined stimulating substances.Strain selection of the rifomycin complex-producingstreptomyces resulted in the isolation of a strain thatproduced fractions C and D in prevalent amounts. Thismade us believe that on further screening a strain thatwould synthetize predominantly one of the othercomponents of the complex would be eventually iso-
TABLE 1. Composition of media and rifomycin B yields
Nutrient ingredients (g/l,OOO ml) ASS/6* ASS/7 ASS/8
Glucose 50 70 70Soya bean meal 5 15 5Phaseolus beanst (Varese 20
type)(NH4)2SO4 7 7 7MgSO4*7H20 1 1 1KH2PO4 3 2 2CaCO3 9 6 6Diethylbarbiturate 2 2 2CuSO 4 *5H20 0.0033 0.0033 0.0033FeSO4-7H20 0.010 0.010 0.010ZnSO4*7H20 0.050 0.050 0.050MnSO4-4H20 0.004 0.004 0.004CoCl*26H20 - 0.002 0.002(NH4)2MoO 4 0.001 0.001
Avg rifomycin B potency 200-250 400-450 650-800obtained (ug/ml)
pH was adjusted to 7.0 with 40% NaOH. Sterilization 30min at 121 C.
* Medium ASS/4 like ASS/6 but without barbital.t Phaseolus beans were cooked for 30 min before addition
to medium.
325
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from
on O
ctober 15, 2017 by guesthttp://aem
.asm.org/
Dow
nloaded from
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

P. MIARGALITH AN-D H. PAGANI[
lated. Our interest in fraction B became more inquisiteafter the extraction, purification, and crystallization ofthis component was achieved (Sensi et al., 1960). How-ever, the screening of 3,000 isolates after variousmutagenic treatments revealed only rifomycin com-plex-producing strainis similar to the parent culture.Therefore, further studies were carried out to find asubstance or precursor which wotuld change the relativecomposition of the antibiotic complex. Chemicallydefined substances as well as complex organic materialsfrom plant and animal soturces were tested in thefermentation medium. Only in one case a strikingdifference in the spectrum (between wavelengths375 to 480 m,) of the antibiotic fractions could beascertained. In this case, S. mediterranei was fermentedusing medium ASS/6 in the final stage (see Table 1).This medium was different from that employed for theproduction of rifomycin complex, principally in thepresence of sodium diethylbarbiturate (barbital). The
characteristic spectrophotometric curve of the ethylacetate extract is given in Fig. 1.
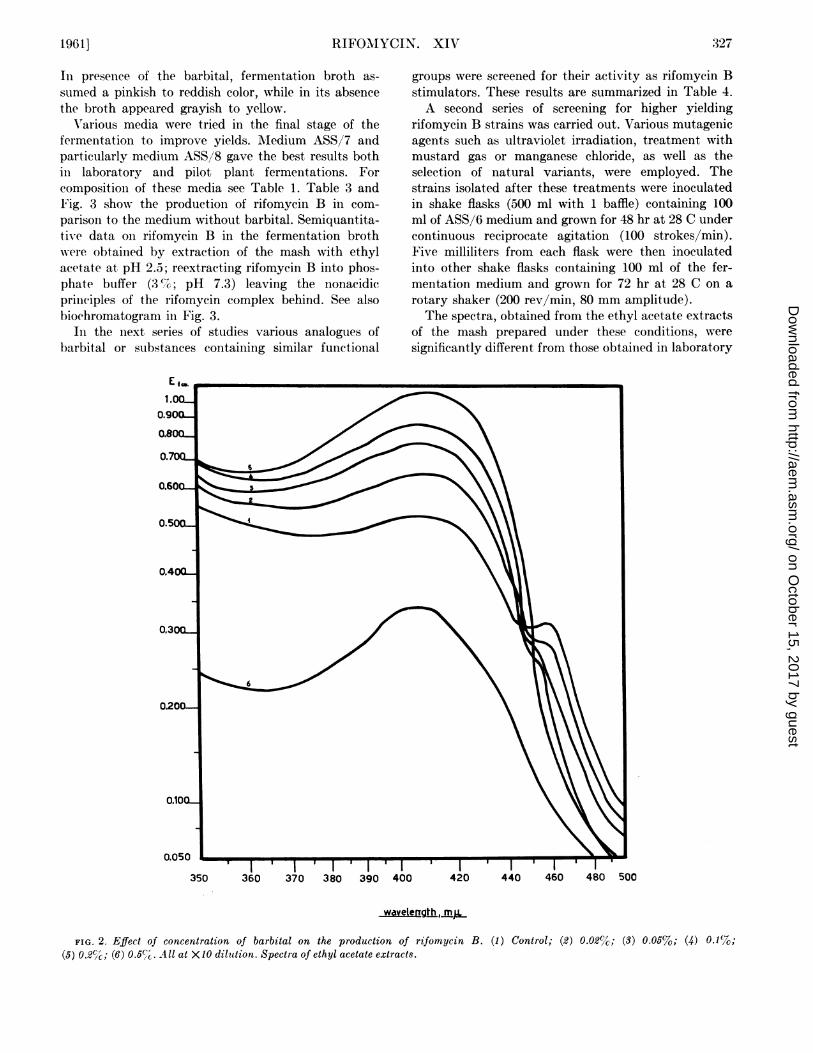
It is evident from Fig. 2 and Table 2 that, in thepresence of barbital, most if not all the antibioticproduced by S. mediterranei is rifomycin B. It isparticularly interesting to note that the microbiologicalactivity of the fermentation broth in the presence ofbarbital is inversely proportional to the concentrationof the barbiturate. The absolute amount of rifomycinB produced in the mash increases up to a concentrationof 0.2 %. This fact can be explained by the lowermicrobiological activity of rifomycin B with respect tothe other fractions of the complex as tested in theroutine agar plate technique.
Fermentations of S. mediterranei, in medium Conl-taining soya bean meal, ammonium sulfate, glucose,and 0.2/7% barbital, produce 200 to 300 ,ug/ml of rifo-mycin B. In the absence of barbital, rifomycin complexwas synthesized with negligible amounts of fraction B
420 440 460 480 500
wavelength ,IFIG. 1. Spectrumrl of rifonmycin B in ethyl acetate as obtained by fermentation in presence of barbital. (1) No barbital; X2dilu.tiott.
(2) NYo barbital oxidized formn; X2 diluition. (3) ITVith barbital; X5 diluition. (4) WTith barbital, oxidized forrm; X5 dilution.
[VOL. 9326 on O
ctober 15, 2017 by guesthttp://aem
.asm.org/
Dow
nloaded from
RIFONIYCIN. XIV
In presence of the barbital, fermentation broth as-sumed a pinkish to reddish color, while in its absencethe broth appeared grayish to yellow.
Vrarious media were tried in the final stage of thefermentation to improve yields. Medium ASS/7 andparticularly medium ASS/8 gave the best results bothin laboratory anid pilot plant fermentations. Forcomposition of these media see Table 1. Table 3 andFig. 3 show the production of rifomycin B in com-parison to the medium without barbital. Semiquantita-tive data on rifomycin B in the fermentation brothwere obtained by extraction of the mash with ethylacetate at pH 2.5; reextracting rifomycin B into phos-phate btuffer (3 c,; pH 7.3) leaving the nonacidicprinciples of the rifomycin complex behind. See alsobiochromatogram in Fig. 3.
In the next series of studies various analogues ofbarbital or substances containing similar functional
groups were screened for their activity as rifomycin Bstimulators. These results are summarized in Table 4.A second series of screening for higher yielding
rifomycin B strains was carried out. Various mutagenicagents such as ultraviolet irradiation, treatment withmustard gas or manganese chloride, as well as theselection of natural variants, were employed. Thestrains isolated after these treatments were inoculatedin shake flasks (500 ml with 1 baffle) containing 100ml of ASS/6 medium and grown for 48 hr at 28 C undercontinuous reciprocate agitation (100 strokes/min).Five milliliters from each flask were then inoculatedinto other shake flasks containing 100 ml of the fer-mentation medium and grown for 72 hr at 28 C on arotary shaker (200 rev/min, 80 mm amplitude).The spectra, obtained from the ethyl acetate extracts
of the mash prepared under these conditions, weresignificantly different from those obtained in laboratory
EI..
0.050 I I I II IIII I
350 360 370 380 390 400 420 440 460 480 500
wavelergtgh,-m,>
FIG. 2. Effect of concentration of barbital on the production of rifomycin B. (1) Control; (2) 0.02%c; (3) 0.056; (4) 0.1%7;(5) 0.2%X-; (6) 0.5%G. All at X10 dilution. Spectra of ethyl acetate extracts.
1961] 327
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

P. MARGALITH AND H. PAGANI
fermentors. Antibiotic titers were most variable. Theextinction curves had the tendency of becoming more
and more pointed and in some of the cases a very
characteristic peak with maximal extinction at 410 m,t(see Fig. 4) was observed. To investigate the possi-bility that these differences were due to the differentbehavior of mutated strains, isolates that produced a
pronounced pointed curve were repeatedly fermentedin shake flasks. Of the five isolates fermented, only one
isolate showed the same spectrum as observed in thefermentors. Not being able to explain this behavior on
genetical grounds, the variability in the spectrum was
therefore suspected to be due to uncontrollable differ-ences in the conditions of shake flask fermentations. Inan attempt to confirm this hypothesis, experimentswere set up under different conditions, affecting agita-tion and aeration, to determine their influence on thenature of the spectrum. The results of these experi-ments are summarized in Table 5.
It is evident from Table 5 that the production ofrifomycin B is extremely sensitive to the agitation-aeration conditions of the fermenting unit. Not onlythe type of spectrum obtained, but also the amount ofrifomycin B produced, as judged from the extinction at410 to 420 m,u, are pronouncedly affected by aeration.The fact that under reduced aeration conditions thecharacteristically pointed curve was obtained inducedus to attempt a fermentation on similar lines to isolatethe substance responsible for this characteristic spec-
trum. A 20-liter glass fermentor with 10 liters of ASS/6medium was set up, reducing the agitation to 300rev/min (instead of 800 rev/min in the usual runs) andan aeration of 0.8 v:v:min. After 48 hr the charac-teristically pointed curve, identical with the spectrafrom shake flasks, was obtained. The fermentation mashwas extracted in the same manner as for rifomycin B.
TABLE 2. Effect of Na-diethylbarbiturate on the formation ofrifomycin B
Microbio- Maximal% Barbital in logical extinction at Remarksmedium activitv wavelengths
(Ag/mlj* (mn)
Control 240 460; 420 Rifomycin complex0.01 190 460; 420 Rifomycin complex0.03 165 460; 420 Rifomycin complex0.05 140 460; 420 Rifomycin complex0.10 105 460; 420 Rifomycin complex0.20 70 ; 420 Rifomycin only0.30 50 ; 420 Rifomycin only0.50 50 -; 420 Rifomycin only
* Activity as compared to a sample of fractions C-D stan-dard preparation.
Fermentations were run in 10-liter glass fermentors with 4liters of ASS/4 medium (see Table 1); 800 rev/min; 1.5 v: v:min
aeration.Sampling and determination of antibiotic activity were per-
formed after 48 hr.
The crude product revealed the same pointed spectrum.However, when submitted to the microbiologicalassay, no antibacterial activity could be demonstrated.Therefore, fermentations with reduced aeration had tobe abandoned. The effect of aeration on rifomycin Bproduction is illustrated in Table 6.
Further screening for higher rifomycin B-yieldingmutants was carried out by fermenting strains in shakeflasks with reduced volume of medium (35 ml/500 mlflask), thus assuring better and more constant yields ofthe control flasks. Under these conditions, only normalspectra of rifomycin B were obtained. Data concerningthe metabolism S. mediterranei in three fermentationmedia are reported in Fig. 5.
DISCUSSIONAs reported in the Materials and Methods section,
when certain substances of the general formulaR CO-NH\/6 1\C5 2C0/\4 3/
R CO NH
are present in the culture media, a shift toward theproduction of fraction B of rifomycins takes place.This is most clearly achieved in the presence of sodiumdiethylbarbiturate, when practically all the antibioticproduced is rifomycin B.The complete disappearance of fraction C and D
and the simultaneous increase of fraction B during the
TABLE 3. Rifomycin B production in ASS/8 and controlmedium
Control 0.2% Barbital
Total ethyl Phosphate Total ethyl PhosphateAge acetate extract; buffer (pH 7.3); acetate extract; buffer (pH 7.3);
maximal maximal maximal maximalextinction at: extinction at extinction at extinction at
410 mp 460 mpu* 425 mpt 460 miA 410 my 460 mu 425 mju 460 mA
hr
36 1.87 1.70 0.96 - 2.75 - 3.05 -
42 2.40 1.90 1.55 - 4.00 - 4.80 -
48 3.40 2.50 2.45 - 4.70 - 6.40 -
54 4.00 2.30 2.60 - 5.30 - 7.00 -60 3.20 3.40 6.00 - 8.30 -72 4.60 3.30 - 6.20 - 7.2096 4.80 3.30 - 7.00 - 0.20 -
120 4.70 3.90 - 7.00 - 7.80 -
* Maximum corresponding to fractions C and D of thecomplex.
t Maximal extinction mostly due to back ground absorp-tion; spectrophotometric curve not characteristic of rifomycinB.
I Extinction coefficient of rifomycin B in phosphate bufferpH 7.3 is somewhat higher than in ethyl acetate; whereasmaximum is shifted from 410 to 425 m,u.
Remarks: Note disappearance of fractions C and D fromcontrol medium after 54 hr due probably to demolition onprolonged fermentation.
[VOL. 9328
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

191]RIFIOYIYCIN\-. XIV2r
fermentation canl be seen from the biochromatogramin Fig. 3. Interestingly enough another fraction (F)with an RF of about 0.9 appeared in small amounts.It is likely that this spot is due to the "activation" ofrifomycin B (Sensi, Timbal, and Maffii, 1960).As canl be seeni from Table 4, none of the moieties of
diethylbarbituric acid, such as urea, malonamide,diethylmalonic acid, or even barbituric acid, behavesin a similar manner. However, the substitution inposition 5 of the barbiturate is not limited to thediethyl analogtue. 5-Ethyl-5-methylbarbiturate haspractically the same effect. 5-Phenyl-5-ethylbarbitu-rate, although greatly stimulating the biosynthesis ofrifomycin B, does not prevent completely some forma-tion of fractionis C and D. a-Ethyl-5-isopropylbar-biturate, although promoting the biosynthesis ofrifomycini B, should be regarded as a poor stimulatorinsofar as only low titers could be obtained. On theother hand, 5, 5-dimethylbarbiturate promotes theformation of all fractions of the complex, also yieldinggood titers of fractionis C and D. Pyrimidines andpurines were found to be ineffective. It is interestingto note that rather recently studies on the use ofbarbiturates in the fermentation media, for the pro-duiction of streptomycin or other antibiotics, have beenicarried otut (Ferguson, Huang, and Davisson, 1957;
Herold, 1959). Ferguson et al. (1957), in fermentingS. griseus in a synthetic medium in the presence of vari-ous barbiturates (10 to 30 X 10-s M), observed a sharpincrease in streptomycin production of about 3 to 4..5times the control activity. Barbiturates were consideredto be one of the synthetic "activity factors" suggestedby Waksman, Schatz, and Reilly (1946) as being neces-sary for mycelial metabolism so as to reach the samelevels of antibiotic production as obtained in completeorganic media. Since streptomycin fermentation pre-sents a distinct biphasic pattern in growth and pro-duction, and since the presence of barbiturates dturingthe growth phase was obligatory, the authors ex-clude the possibility that these substances serve asprecursors or coenzymes but attribute the activity tothe induction of the antibiotic producingenzyme system.Further, Ferguson et al. (1957) state that, for optimalactivity, there are two requirements: (i) the barbituricacid nucleus and (ii) the disubstitution of alkyl groups inposition 5 of C2, C2 or C2 and C3 as optimal range.However, both diethylmalondiamide and ,3-ethyl-3-methylglutarimide gave similar, yet somewhat loweractivity. No activity was found with 5-phenyl-5-ethylbarbiturate. -No correlation betweeni hypnoticaction of the barbiturates and streptomycin prodtuctionof these compounds could be demonstrated.
* Sodinnttl diethylbarbiturate (barbital)FIG. 3. Biochromzatogram. of rifonmycin produiction in ASS/8 and control mediulm
32919)611
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

TABLE 4. Effect of barbiturates and analogues on rifomycin B production*
Ethyl acetate extract;Optimal maximal extinction at: Phosphate
Compound Formula concn buffer; maximal Remarksused extinction at(g/liter) 405-420 460 nu 425 m;&
Control
Urea
Malonic acid
Malonamide
Diethylmalonicacid
Barbituric acid
Dimethylbarbituricacid
Ethylmethylbarbi-turic acid
Diethylbarbituricacid
Phenylethylbarbi-turic acid
Ethylisopropylbar-bituric acid
Ethylisoamylbarbi-turic acid
Dipropylbarbituricacid
NH2
CO
NH2
COOH
CH2
COOH
CONH2
CH2
CONH2
C2H5 COOH
C
C2H5 COOH
CH-NH/\
CH2 CO\/
CO-NH
CH3 CO-NH
\C/ \CO\ c/
CH3 CO-NH
C2H5 CO-NH
\C/ \COCH3 CO-NH
C2H5 CO-NH
\C/ \CO\ c/
C2H5 CO-NH
C6H5 CO-NH
\C/ \CO/\ /
C2H5 CO-NH
C2H5 CO-NH
\C/ \CO(CH3)2CH CO-NH
C2H5 CO-NH
\C/ \CO/\ /
(CH3)2CH(CH2)2 CO-NH
C3H7 CO-NH
\CO \COC3H7 CO-NH
0.8
2.0
2.0
2.0
4.0
2.0
2.0
2.0
0.5
2.0
2.0
1.0
1.95
1.30
No
1.70
1.78
1.00
maxima
1.55
No maxima
3.2
2.60
3.07
3.20
2.60
0.51
1.15
Shoulder
1.97
Shoulder
0.18
0.93
No maxima
No maxima
No maxima
No maxima
No maxima
2.82
3.32
2.15
0.53
No maxima
No maxima
Rifomycin B
Abundant formationof fractions C andD
Rifomycin B
Rifomycin B
Predominantly rifo-mycin B
Rifomycin B
330
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

RIFOMYCIN. XIV
TABLE 4. Continued
Ethyl acetate extract;Optimal maximal extinction at: Phosphate buffer;
Compound Formula concn maximal extinc- Remarksused tion at 425 mp&(g/liter) 405-420 460m
n y
Diallylbarbituric CH2: CHCH2 CO-NH 2.0 0.56 0.32 0.49 (at 400acid C CmO m)
CH2:CHCH2 CO-NH
5-Nitrobarbituric CO-NH 1.0 2.15 1.70 No maximaacid N02-CH CO
CO-NH
5-Aminobarbituric CO-NH 2.0 1.15 0.85 No maximaacid/
NH2-CH CO
CO-NH
Thiobarbituric acid CO-NH 2.0 2.12 1.40 No maxima
CH2 Cs
CO-NH
Diethylamino- (C2H5)2NCH2CH20H 4.0 1.55 1.35 No maximaethanol
* All test were carried out on pooled samples of 5 X 35 ml.rotary shaker for 72 hr at 28 C.
The effect of barbiturates on rifomycin B productionis a different case. The fermentation of rifomycin Bdoes not show a distinct biphasic pattern; most rifomy-cin B formation is taking place during the growth of themycelium or immediately thereafter (see Fig. 5). Bar-bital is necessary for the suppression of the productionof fractions C and D during the fermentation in a com-plex, organic media.
In an attempt to explain the effect of barbital on thefermentation of rifomycin B we come into twohypotheses:A) Barbital can be a precursor of the antibiotic in a
strict sense, i.e., barbital may constitute part of therifomycin B molecule.B) Barbital can play the role of an activator or
inhibitor of certain enzymic systems. By shifting thebiosynthetic pathways (metabolic shunt) from theformation of the whole rifomycin complex to the bio-synthesis of rifomycin B only, all the antibiotic potentialof S. mediterranei is exploited for the production of thedesired fraction.The facts accounted for in the experimental part
favor to some extent the latter hypothesis. The pre-adaptation of S. mediterranei by growing the vegetativemycelium in the presence of a high concentration ofbarbital (up to 0.5 % w/v) did not result in the induc-tion of the enzymic system responsible for the soleproduction of rifomycin B. Further, mycelium sampled
ASS/8 medium in shake flasks of 500 ml. Growth took place on a
at different time intervals from a fermentor containingASS/8 medium, washed three times in buffer toeliminate both nutrient ingredients as well as barbital,suspended in equal amounts of phosphate buffer(M/15, pH 6.8), and shaken for 24 hr at 28 C wouldagain produce the whole rifomycin complex (withmaxima at 420 and 460 m,u). This appears to excludethe possibility of an adaptive nature of the system.When adding barbital to the fermentor after growthhas taken place for 48 hr no shifting of the spectrum(rifomycin B production) would occur. This might bedue to the absence of the stimulating compound in thecritical period of enzyme formation preceding that ofrifomycin B production, which under normal conditionscontinues up to 72 to 96 hr. This would imply thatbarbital induces a shift in the synthesis of the enzymesystem, being without effect after the antibiotic-formingsystem has already been established. However, these re-sults stand in contrast with those obtained from pre-viously mentioned experiments. Therefore, it is possiblethat the presence of barbital is obligatory during all theperiod of rifomycin B production, thus suppressing theformation of fractions C and D. The question whetherbarbital is metabolized during rifomycin B productionis still open, since attempts to follow the consumptionof barbiturate failed due to analytical difficulties. Thereappears to be an optimal concentration of barbital forrifomycin B production (Fig. 2). It can be seen that
1961] 331
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

wavelength, w,
FIG. 4. Spectra obtained from. Streptomyces nmediterraniei shake flask fermentation (ethyl acetate extract); 100 ml mnedimn (- ); 35ndl medium; X5 dilution (--).
TABLE 5. Effect of physical tariants on rifomycin B production*
Vol of MaximalType of vessel medium eaxtin41i4 Remarks
1-baffled, 500-ml shake 25 4.30 Rifomycin Bflask 30 2.4 Rifomycin B
75 0.52 "Pointed spec-trum"t
100 0.25 No maxima200 0.56 No maxima
3-baffled, 500-mil shake 100 0.50 Rifomvcin Bflask
Test ttubes, 185 by 16 10 0.68 Slightly "pointedmm |spectrum"
* Five per cent inoculum of 24-hr growth in seed medium.Fermentation in ASS/7 medium for 72 hr at 28 C. on a rotaryshaker (200 rev/min), except for test tubes which wereagitated on a reciprocal shaker. Broth from small samples(<75 ml) were pooled before extraction.
t See text for explanation.
TABLE 6. Effect of aeration on rifomycin B production*
MycelialAir flow (v/v) Time (hr) pH growth (% E410-420m,volume)
0.5 48 6.0 30 3.6072 6.0 25 4.2596 6.0 20 4.55
1.0 48 6.0 30 3.7072 6.0 30 4.5096 6.0 28 4.65
1.5 48 6.0 30 3.6572 6.0 30 5.3096 6.0 30 5.80
2.0 48 6.0 30 4.0572 6.0 30 7.096 6.0 30 8.5
2.5 48 6.0 30 4.8072 6.0 35 7.6096 6.0 40 9.40
* Fermentation medium ASS/8; agitation 800 rev/min.
332
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

RIFOMYCIN. XIV
over 0.2 % no better results could be obtained. At0.5 % a negative effect can be observed.
These results are in agreement with those of Fergusonet al. (1957). At higher concentrations, the spectro-photometric curve appears to resemble more the oneobtained under unfavorable conditions of aeration. Itis interesting to note that the optimal concentration isindependent of the medium employed, it being identicalunder the conditions that permit the production of200 to 300 ,g/ml or 600 to 800 ,ug/ml. This is anotherindication for the existence of a regulative mechanismof the enzyme system.No doubt the addition of barbiturates, per se, has a
certain toxic effect upon S. mediterranei. Mycelialgrowth in fermentation media without these compoundsis about 20 to 25 % more abundant in comparison tothe barbiturate-containing media. The presence of0.2 % barbital reveals a certain inhibition in colonialgrowth in surface cultures. No significant differencescould be noted in the metabolism of carbohydrates,nitrogen, or phosphorus as compared to the controlmedium. The effect of barbiturates in suppressing theformation of fractions C and D appears to be that of amild inhibitor. Coming back to the observations madeby Ferguson et al. (1957) it is possible to explain theformation of rifomycin B by a so-called "dual effect"of barbiturates. As in streptomycin production, all5, 5-disubstituted barbiturates (with certain restrictions
QILN--totalmg/ml
2.5
as to the nature of the derivative), including dimethyl-barbiturate and possibly also the phenylethyl analogue,stimulate the total biosynthesis of the antibiotic frac-tions. However, only a limited number of analogues,with at least one ethyl group in position 5, will promotethe production of rifomycin B. Phenyl ethyl barbiturateoccupies an intermediate position, i.e., as generalstimulator and to a certain extent that of a rifomycinB promoter. This can be considered as another indica-tion for the "dual effect" of certain barbiturates onrifomycin B production.
It has been shown that rifomycin B production is notdue to the action of an inductive enzyme system butrather to a "metabolic shunt." In agreement with thedata of Ferguson et al. (1957), barbiturates have ageneral stimulating effect on the various antibioticprinciples. Of these, however, only 5-methyl-5-ethyl-and 5,5-diethylbarbiturate promote the production ofonly one fraction, i.e., rifomycin B, while simultane-ously suppressing the other antibiotic components.
LITERATURE CITED
FERGUSON, J. H., H. T. HUANG, AND J. W. DAVISSON. 1957.Stimulation of streptomycin production by a series ofsynthetic organic compounds. Appl. Microbiol. 5:339-343.
GALLO, G. G., P. SENSI, AND P. RADAELLI. 1960. RifomicinaVII. Analisi spettrofotometrica della rifomicina B.Farmaco (Pavia), Ed. pract. 15(5):283-291.
E 420 mun 8.0
7.0
6.0
5.0
4.0
3.0
2.0
1.0
Time, Hours
FIG. 5. Metabolism of Streptomyces mediterranei in three fermentation media
19611 333
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

J. KRAMER AND A. KIRSHBAUM
HEROLD, M. 1959. Conditions particularly affecting the engi-neering aspects of the biosynthesis of antibiotics. Mainlecture. Technological section. Antibiotic symposium,Prague, May 1959.
MARGALITH, P., AND G. BERETTA. 1960. Rifomycin. XI. Taxo-nomic study on Streptomyces mediterranei nov. sp. Myco-pathol. et Mycol. Appl., 8(4):321-330.
MARGALITH, P., AND H. PAGANI. 1961. Rifomycin. XIII.Fermentation and production of rifomycin complex. Appl.Microbiol. 9:320-324.
SENSI, P., A. M. GRECO, AND R. BALLOTTA. 1960. Rifomycin.I. Isolation and properties of rifomycin B and rifomycin-complex. Antibiotics Ann. 1959/60:262-270.
SENSI, P., M. T. TIMBAL, AND G. MAFFII. 1960. Rifomycin.IX. Two new antibiotics of rifomycin family: RifomycinS and rifomycin SV. Preliminary report. Explrientia16(9) :412-414.
WAKSMAN, S. A., A. SCHATZ, AND H. C. REILLY. 1946. Metab-olism and the chemical nature of Streptomyces griseus.J. Bacteriol. 51:753-759.
Effect of Paper on the Performance Assay in the Control ofAntibiotic Sensitivity Discs
JULIAN KRAMER AND AMIEL KIRSHBAUM
Department of Health, Education, and Welfare, Food and Drug Administration, Washington, D. C.
Received for publication October 19, 1960
ABSTRACT
KRAMER, JULIAN (Food and Drug Administration,Washington, D. C.), AND AMIEL KIRSHBAUM. Effect ofpaper on the performance assay in the control of anti-biotic sensitivity discs. Appl. Microbiol. 9:334-336.1961.-Antibiotic discs were prepared, using severalseveral batches of papers meeting Food and DrugAdministration specifications. The analysis of 1,152zones of inhibition produced showed no performancedifferences among these batches. Other discs were
prepared using papers of different grades. These pro-
duced large differences in performance. It is obvious,therefore, that the use of a specified disc paper is neces-
sary for standardizing the performances of the productsof various manufacturers and that reproducible resultscan be attained with the grade of paper specified.
The characteristics of the paper used are an
important consideration in the manufacture of sen-
sitivity discs. Because of the use of paper as matrix inthis product, these characteristics rank highly as
potential sources of variation in the performance ofdiscs. The various manufacturers have selected papers
suitable to the requirements for their final products.The specifications of these papers vary somewhat as
to weight, thickness, anid absorbability of water. Forthis very reason, paper must play a major role in thestandardization of discs. The proposed regulations ofthe Food and Drug Administration (FDA) for theassay of discs (Anonymous, 1960) specify the paper tobe used for the preparation of the control discs. Thispresent work was undertaken to answer questions
raised about the reproducibility of results that can beobtained with different commercial batches of paperof the specified grade. In this publication "differentbatches" refer to paper of the same specifications butmanufactured at different times, whereas "differentgrades" refer to papers with different specifications.The Food and Drug Administration specifies that
the blank discs used in the control of commercial discs"be 14 inch in diameter, of paper weighing approxi-mately 30 mg per square centimeter that will absorbapproximately 3 times its weight of distilled water."
Ostrander and Griffith (1959) used an exquisitelysensitive indicator organism in a study of the effectsof 16 different papers used to make penicillin discs.With this sensitive organism, two units of penicillinis in the upper range of concentrations where furtherincreases of concentration produce no further response.This study indicated that unless some other agents,such as certain dyes, were present, the paper used madeno difference in the performance of the disc.
In our study the organisms used were those desig-nated in the FDA proposed regulations.' Each of theseorganisms was originally selected because its sensitivityis such that an essentially linear response is obtainedto the concentrations of antibiotics used. The study wasmade in two parts so that these questions might beanswered: (i) Is there any difference in performanceamong discs made with different commercial batchesof paper meeting Food and Drug specifications, and(ii) are there performance differences among discsmade with papers that do not meet these specifications?
I The organism used for polymyxin was Bordetella bron-chiseptica.
::334 [VOL. 9
on October 15, 2017 by guest
http://aem.asm
.org/D
ownloaded from

INFRARED SPECTROPHOTONMETRY OF S. A URE'US
dissimilarities brought about by the canceling of somebands and reversal of others is lost, even though enoughof the balancing effect to allow greater scale expansion ismaintained. Another difficulty is that the double-beamdifferential analysis cannot detect a real difference whosemagnitude is similar to that of naturally occurring sample-to-sample variability. Strains with high natural variabilitycannot be analyzed with good reproducibility.
Factors such as warping of the Nernst glower, differencesin batches of the culture medium, and individual tech-niques become more important, as sources of significantvariation when the ordinate scale of the spectrum isexpanded. These factors are not subject to control bythe described method and make it even more difficult toreproduce differential spectra repeatedly.
In general, the method described would probably servebetter in association with physiological and biochemicalstuidies than as a means of differentiating strains inepidemiological investigations. A satisfactory interpreta-tion of the dissimilarities exposed by this method is notpossible without further experimentation. The significanceof the 'band at 11.6 to 11.7 , in the spectra is of particularinterest since it does make possible one broad separationof strains. The glycogen spectrum studied by Levine et al.(1953a) in enteric organisms produced bands in this regionand may offer a clue to the factors responsible for theband. Its nature and possible correlation with othercharacteristics of the organism are subjects now underconisideration.
LITERATURE CITED
KENNER, B. A., J. W. RIDDI.E, S. W. ROCKWOOD, AND R.H. BORDNER. 1958. Bacterial identification by infrared spee-
trophotometry. II. Effect of instrumental and environmentalvariables. J. Bacteriol. 75:16-20.
KULL, F. C., AND MI. R. GRIMMI. 1956. Differentiation of resistantmutants by infrared analysis. J. Bacteriol. 71:342-347.
LEVI, L., B. H. MATHESON, AND F. S. THATCHER. 1956. Detectionof staphylococcus enterotoxin by infrared spectrophotometry.Science 123:64-65.
LEVINE, S., H. J. R. STEVENSON, AND R. H. BORDNER, 1953a.Identification of glycogen in whole bacterial cells by infraredspectrophotometry. Science 118:141-142.
LEVINE, S., H. J. R. STEVENSON, L. A. CHAMBER, AND B.A. KENNER. 1953b. Infrared spectrophotometry of the entericbacteria. J. Bacteriol. 65:10-15.
LEVINE. S., H. J. R. STEVENSON, E. C. TABOR, R. H. BORDNER,AND L. A. CHAMBER. 1953c. Glycogen of enteric bacteria. J.Bacteriol. 66:664-670.
O'CONNOR, R. R., E. R. MCCALL, AND E. F. DUPRE. 1957. Differen-tiation of microorganisms by means of the infrared spectra oftheir acetone extracts. J. Bacteriol. 73:303-313.
PERKIN-ELMER CORPORATION. 1957. Instruction manual for theordinate scale expander. Norwalk, Conin.
RANDALL, H. M., D. W. SMITH, A. COLM, AND W. J. NUNGESTER.1951. Correlation of biologic properties of strains of myco-bacterium with their infrared spectrums. I. Reproducibility ofextracts of M. tuberculosis as determined by infrared spec-troscopy. Am. Rev. Tuberc. 63:372-380.
RANDALL, H. M., D. W. SMITH, AND W. J. NUNGESTER. 1952. Cor-relation of biologic properties of strains of mycobacteriumwith their infrared spectrums. II. The differentiation of twostrains. H37Rv and H37Ra of M. tuberculosis by means oftheir infrared spectrums. Am. Rev. Tuberc. 65:477-480.
RIDDLE, J. W., P. W. KABLER, B. A. KENNER, R. H. BORDNER,S. W. ROCKWOOD, AND H. J. R. STEVENSON. 1956. Bacterialidentification by infrared spectrophotometry. J. Bacteriol.72:593-603.
STEVENSON, H. J. R., AND 0. E. A. BOLDUAN. 1952. Infrared spec-trophotometry as a means for identification of bacteria. Sci-ence 116:111-113.
ERRATUMIn the paper "Rifomycin. XIV. Production of Rifomycin B" by P. Margalith
and H. Pagani, Appl. Microbiol. 9:325-334 (1961), an error was made in the set-ing of TABLE 4. Effect of barbiturates and analogues on rifomycin B production. Theentry in the Remarks column for the Control material should read Rifomycincomplex, not Rifomycin B.
179


















