
Retortamonadida (with Notes on Carpediemonas-Like ... · Retortamonadida (with Notes on...
32
Retortamonadida (with Notes on Carpediemonas-Like Organisms and Caviomonadidae) Jaroslav Kulda, Eva Nohýnková, and Ivan Čepička Contents Summary Classification ........................................................................... 2 Introduction ....................................................................................... 3 General Characteristics ........................................................................ 3 Occurrence .................................................................................... 3 Literature ...................................................................................... 4 History of Knowledge ........................................................................ 4 Practical Importance .......................................................................... 5 Habitat and Ecology .............................................................................. 6 Recognition and Characterization ................................................................ 6 The Trophozoite ............................................................................... 6 The Cyst Stage ................................................................................ 12 Reproduction .................................................................................. 12 Maintenance and Cultivation ..................................................................... 18 Evolutionary History .......................................................................... 19 Coda: Carpediemonas-Like Organisms and Caviomonadidae .................................. 20 Introduction ................................................................................... 20 Light Microscopy ............................................................................. 21 Flagellar Apparatus ........................................................................... 21 Flagellar Transformation ...................................................................... 25 Nucleus and Cytoplasmic Organelles ......................................................... 25 Ultrastructure of Cysts ........................................................................ 26 Taxonomy and Evolutionary History ......................................................... 26 Morphology, Ultrastructure, and Ecology of Caviomonadidae .............................. 27 References ........................................................................................ 28 J. Kulda (*) • I. Čepička Faculty of Science, Charles University, Prague 2, Czech Republic e-mail: [email protected]; [email protected] E. Nohýnková First Faculty of Medicine, Charles University, Prague 2, Czech Republic e-mail: [email protected] # Springer International Publishing AG 2016 J.M. Archibald et al. (eds.), Handbook of the Protists, DOI 10.1007/978-3-319-32669-6_3-1 1
Transcript of Retortamonadida (with Notes on Carpediemonas-Like ... · Retortamonadida (with Notes on...

Retortamonadida (with Notes onCarpediemonas-Like Organisms andCaviomonadidae)
Jaroslav Kulda, Eva Nohýnková, and Ivan Čepička
ContentsSummary Classification . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
General Characteristics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3Occurrence . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3Literature . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4History of Knowledge . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4Practical Importance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
Habitat and Ecology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6Recognition and Characterization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
The Trophozoite . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6The Cyst Stage . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12Reproduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
Maintenance and Cultivation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18Evolutionary History . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
Coda: Carpediemonas-Like Organisms and Caviomonadidae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20Light Microscopy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21Flagellar Apparatus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21Flagellar Transformation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25Nucleus and Cytoplasmic Organelles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25Ultrastructure of Cysts . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26Taxonomy and Evolutionary History . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26Morphology, Ultrastructure, and Ecology of Caviomonadidae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
J. Kulda (*) • I. ČepičkaFaculty of Science, Charles University, Prague 2, Czech Republice-mail: [email protected]; [email protected]
E. NohýnkováFirst Faculty of Medicine, Charles University, Prague 2, Czech Republice-mail: [email protected]
# Springer International Publishing AG 2016J.M. Archibald et al. (eds.), Handbook of the Protists,DOI 10.1007/978-3-319-32669-6_3-1
1

AbstractRetortamonadida (retortamonads) is a group of bacterivorous metamonadsbelonging to Fornicata, currently represented by the single familyRetortamonadidae, with two genera, Retortamonas and Chilomastix, and about60 species. They are adapted to low-oxygen environments and live predomi-nantly as endocommensals in intestines of animal hosts, including humans. Twospecies were reported to be potential pathogens causing diarrhea in humans(C. mesnili) or unadapted avian hosts (C. gallinarum). One species(C. cuspidata) is free-living in hypoxic water sediments. Retortamonads aretypical excavates with a single karyomastigont possessing four basal bodies andtwo or four flagella. One flagellum is recurrent, has two or three lateral vanes, andis associated with a ventral feeding groove. Double-membrane-bounded organ-elles without cristae, assumed to be mitochondrial derivatives, were found inChilomastix. Retortamonads reproduce by binary division and produce a resistantcyst stage. The cysts of endobiotic species are discharged with feces and serve tospread the infection. Recent phylogenetic analysis and ultrastructural observa-tions indicate that Retortamonas species from insects are close relatives ofChilomastix, while species from vertebrates appear to be relatives ofDiplomonads and should be excluded from this genus. Carpediemonas-likeorganisms (CLOs) comprise a few species of small, free-living Fornicata. Theyare typical excavates and are biflagellated, though usually possess three or fourbasal bodies in the mastigont. CLOs form a paraphyletic grade in phylogenetictrees, having retortamonads, diplomonads, and Caviomonadidae (i.e., the rest ofFornicata) nested within them. Caviomonadidae is a group of morphologicallyreduced uniflagellates that were thought to belong to Diplomonadida untilrecently. Instead, they are closely related to the CLO genera Hicanonectes andAduncisulcus. Caviomonadidae includes three endobiotic species and oneundescribed free-living, marine isolate.
KeywordsBacterivore • Carpediemonas-like organisms • Caviomonadidae • Excavata •Flagellate • Protozoa • Retortamonadida
Summary Classification
● Fornicata●● Carpediemonas●● Ergobibamus●● Aduncisulcus●● Hicanonectes●● Caviomonadidae●●● Caviomonas●●● Iotanema
2 J. Kulda et al.

●● Retortamonadida●●● Chilomastix●●● Retortamonas *●● Kipferlia●● Dysnectes●● Diplomonadida **
*Likely polyphyletic with its present species composition – see text.**See chapter ▶Diplomonadida.
Introduction
General Characteristics
Retortamonads are small (5–20 μm) bacterivorous protists with two or four flagella.They possess a large ventral feeding groove elaborated into a conspicuouscytostome-cytopharynx complex supported by microtubules and other cytoskeletalcomponents (Kulda and Nohýnková 1978). One of the flagella, equipped with vanes,is directed posteriorly and runs through or is confined to the cytostomal cavity.Motion of this flagellum generates the feeding current. Retortamonads lack classicalmitochondria, peroxisome-like organelles, and a morphologically developed Golgiapparatus, but possess double-membrane-bounded organelles that resemblehydrogenosomes in their morphology (Hampl and Simpson 2008). The life cycleof retortamonads includes two stages: the motile trophozoite stage, which multipliesby longitudinal binary fission, and the resting cyst stage, which is protected by athick cyst wall (Kulda and Nohýnková 1978). Because no biochemical or pertinentgenomic data are available, the physiology and cell biology of retortamonads arelargely unknown. No sexual processes have been observed. At present only onefamily, Retortamonadidae, and two genera, Retortamonas and Chilomastix, arerecognized, with about 60 species described (Kulda and Nohýnková 1978). Molec-ular phylogenetic analyses show that Retortamonadida is not monophyletic with itscurrent composition (Cepicka et al. 2008; Takishita et al. 2012).
Occurrence
Retortamonads typically exist as endocommensals in the digestive tracts of verte-brate and invertebrate animals; two species Retortamonas intestinalis andChilomastix mesnili are found in the human large intestine (Kulda and Nohýnková1978). Chilomastix cuspidata is a free-living species of this group that inhabitshypoxic water sediments (Bernard et al. 2000).
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 3

Literature
General information on retortamonads can be found in protozoology books such asDoflein and Reichenow (1952) and Levine (1973). The most complete data can befound in Grassé (1952) and Kulda and Nohýnková (1978). Brugerolle and Mignot(1990) and Brugerolle and Lee (2000) published comprehensive chapters on theseorganisms. Numerous earlier studies provide information on the morphology, tax-onomy, and cell division of different retortamonad species based on light micros-copy. Of these, publications by Bělař (1921), Boeck and Tanabe (1926), Bishop(1931, 1935), Wenrich (1932), Geinman (1935), Nie (1948, 1950), Kirby andHonigberg (1950), Moskowitz (1951), and McDowell (1953) deserve particularattention. The ultrastructure of Retortamonas and Chilomastix was examined byBrugerolle (1973, 1977) and Bernard et al. (1997). Molecular phylogenetic analysesbased on sequences of the small subunit ribosomal RNA (SSU rRNA) gene werepresented by Silberman et al. (2002) and Cepicka et al. (2008), and a multigenephylogenetic analysis was presented by Takishita et al. (2012).
History of Knowledge
Grassi (1879) described the first species of Retortamonas in the intestine of the molecricket Gryllotalpa. The generic names Embadomonas and Waskia were usedthereafter for several species described in insects and vertebrates (e.g., Brug 1922;Hegner and Schumaker 1928; see also Wenrich 1932). Later, the three names weretreated as subjective synonyms, and only a single genus of biflagellateretortamonads, Retortamonas, was recognized (Wenrich 1932). Alexeieffestablished the quadriflagellate genus Chilomastix in 1912. Synonyms ofChilomastix include Macrostoma, Fanapepea, Tetrachilomastix, Cyathomastix,and Difamus. Both Alexeieff (1912) and Mackinnon (1915) observed the morpho-logical similarity between Retortamonas and Chilomastix, but Alexeieff (1917) wasthe first to unite these genera into a single family, designated Embadomonadidae.Wenrich (1932) confirmed the relationship, restored the name Retortamonas for thegenus possessing two flagella, and established the family Retortamonadidae. Thisclassification, however, was not generally accepted, because it conflicted with thetraditional grouping of flagellates in Protomonadida and Polymastigida (Blochman1895), based on the number of flagella. Accordingly, Retortamonas (possessing twoflagella) and Chilomastix (four flagella) were usually assigned to different orders ofMastigophora. Grassé (1952) dismissed this rigid concept and proposed a morenatural grouping based on comparative morphology. He introduced several neworders of flagellates, including Retortamonadida, which comprised Wenrich’s familyand again brought together Chilomastix and Retortamonas. Subsequent electronmicroscopic observations (Brugerolle 1973, 1977) confirmed this relationship.
During the period 1926–1938, investigators published valuable observations onthe morphology of members of the Retortamonadida based on light microscopy. Theintroduction of the protargol staining method in the early1950s (see Nie 1950 for a
4 J. Kulda et al.

workable protocol) allowed for a better resolution of karyomastigont structures, thuscontributing to the definition of taxonomically sound morphological characters. Thework of investigators from the Wenrich and Kirby schools during this periodrepresents the best light microscopic morphology studies of the retortamonads andother intestinal flagellates. Electron microscopic studies by Brugerolle (1973, 1977,1991) and Bernard et al. (1997) revealed the subcellular organization of bothChilomastix and Retortamonas and provided useful morphological data for phylo-genetic considerations of retortamonads in the context of other “excavate taxa”(Simpson 2003). Early molecular phylogenetic analyses based on the SSU rRNAgene and involving several strains of Retortamonas from vertebrates (Silbermanet al. 2002) showed a close relationship between retortamonads and diplomonads.When Chilomastix SSU rRNA gene sequences were included in analyses (Cepickaet al. 2008; Takishita et al. 2012), the monophyly of the Retortamonadida, thoughexpected on the basis of ultrastructural observations (Brugerolle 1973, 1977), wasnot supported. As mentioned by Cepicka et al. (2008), these discrepancies may resultfrom different taxon sampling, as all Retortamonas strains subjected to molecularphylogenetic analyses were isolated from vertebrates, whereas ultrastructural studieswere performed on species from insects only. It is therefore likely that the genusRetortamonas is polyphyletic, which should lead to substantial taxonomic revisionof the group in the future.
Practical Importance
Most members of the Retortamonadida are assumed to be harmless commensals;however, two species, Chilomastix mesnili in humans and Chilomastix gallinarum inpoultry, have been reported as potential pathogens.
The medical importance of Chilomastix mesnili is marginal. The parasite isdistributed worldwide and shows a low incidence in standard surveys (0.2–1%;e.g., Waikagul et al. 2002; Jacobsen et al. 2007; Nasiri et al. 2009). No markeddifferences in incidence have been observed in surveys performed in differentgeographic areas; however, an increased prevalence of 11–40% has been reportedin specific groups of people suffering from diarrhea, including a closed communityof institutionalized children (Červa and Větrovská 1958), military troops deployedon overseas missions (Oyofo et al. 1997), and selected groups of diarrheic patients(Felsenfeld and Young 1946). Although the association of Chilomastix infectionwith diarrhea does not necessarily reflect a causal relationship in all reported cases,the pathogenic potential of this species cannot be ruled out (Westphal 1939; forfurther information, see Kulda and Nohýnková (2006)). No specific treatment isrecommended for asymptomatic infections; successful treatment of symptomaticpatients by carbasone (Červa and Větrovská 1958) or metronidazole (Barnham1977) has been reported. Because the infection is acquired by swallowing cystsreleased from the feces of an infected person, good sanitation and personal hygieneare the most effective preventive measures.
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 5

Chilomastix gallinarum, a common commensal in the ceca of chicken, turkeys,pheasants, and ducks, is potentially pathogenic to unadapted avian hosts, such asquail. Davis et al. (1964) described an outbreak of acute diarrhea and mortality inyoung pen-raised quail involving 35,000 birds. Chilomastix, which was abundant inthe lower digestive tract of the affected birds, was identified as the etiologic agent,and its pathogenicity in quail was experimentally confirmed.
Habitat and Ecology
A common feature of the retortamonads is their adaptation to low-oxygen environ-ments. Most retortamonad species live in the intestines of various animals; species ofboth Retortamonas and Chilomastix can be found in the human intestine. It seemsthat the typical association of retortamonads with particular hosts is commensalism(Kulda and Nohýnková 1978). Transmission among susceptible hosts occurs viaingestion of resistant cysts discharged into the environment from the feces ofinfected individuals. Most retortamonad species are most likely host specific, but afew of them may exhibit a wider host spectrum, including either related animals ormembers of a common biotope.
Species of the genus Retortamonas live in the intestines of both vertebrates andinvertebrates. The typical habitat is the midgut or hindgut of insects, such as molecrickets, cockroaches, water-dwelling larvae of crane flies and beetles, and sometermites (Grassé 1952; Brugerolle 2006). Their primary habitat in vertebrates is inthe cecum and colon of mammals and the cloaca of amphibians and reptiles (Kuldaand Nohýnková 1978). The ostrich Struthio camelus is the only avian host ofRetortamonas reported thus far (Martínez-Díaz et al. 2001). Retortamonasintestinalis is an infrequent nonpathogenic inhabitant of human cecum, with lowprevalence (up to 2%) even in populations with poor hygiene standards. Species ofChilomastix have been recorded in numerous vertebrates, including humans (Kuldaand Nohýnková 1978), and in a few invertebrates (the horseleech Haemopissanguisuga and some termites). The localization of these species in vertebrates issimilar to that of Retortamonas. The free-living species Chilomastix cuspidata hasbeen reported in hypoxic sediments of marine, brackish, and freshwaters inAustralia, the United States, Northern Ireland, and Denmark (see Bernard et al.1997 for pertinent references).
Recognition and Characterization
The Trophozoite
Light MicroscopyTrophozoites of both retortamonad genera are pyriform or carrot-shaped cells thatare rounded anteriorly and tapered posteriorly, with a posterior spike in somespecies. In Chilomastix, the cell is ventrally flattened, dorsally convex, and
6 J. Kulda et al.

sometimes twisted in its posterior portion. The most conspicuous feature is thecytostome, a large pouch-like oral aperture on the ventral side (Fig. 1a, c). Thecytostomal cavity extends into a tubelike cytopharynx through which food particlesenter the interior of the cell and are endocytosed. After protargol or hematoxylinstaining, two fibers supporting the lips that surround the cytostome become evident.The right fiber is more prominent and longer than the left. The former is curvedposteriorly and forms a hook along the cytopharynx.
The spherical nucleus is located near the anterior margin of the cell, dorsally, andto the left of the cytostome. The flagella and both cytostomal fibrils originate in abasal body complex above the arched anterior margin of the cytostome, close to the
Fig. 1 Diagrammatic representation of Chilomastix and Retortamonas trophozoites and cysts,based on light microscopic observations. (a) Chilomastix trophozoite. Ventral view. The cell bearsfour flagella, three anterior (aFl), and one recurrent (rFl) that is located in the cytostomal pouch(Ct). There are two cytostomal fibers, the right (rCF) and the left one (lCF) outlining the margins ofthe cytostome on each side. The more prominent right cytostomal fiber is hooked posteriorly. Allflagella and both fibers originate in the basal body complex situated close to the lower right marginof the nucleus (N ). The cytoplasm contains numerous digestive vacuoles (dV) with ingestedbacteria. (b) The pear-shaped Chilomastix cyst is enveloped by a thick cyst wall (CW) and containsa single nucleus (N ), both cytostomal fibers (RCF, LCF), and internalized flagella (Fl).(c) Retortamonas trophozoite. The cell bears two flagella inserted near the nucleus (N ). One isdirected anteriorly (aFl); the other is recurrent (rFl), passing through the cytostomal pouch (Ct) andextending outside by its distal portion. Digestive vacuoles (dV) containing bacteria are present in thecytoplasm. (d) The cyst of Retortamonas showing a cyst wall (CW), nucleus (N ), internalizedflagella (Fl), and cytostomal fibers (CF)
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 7

nucleus. In both genera, one flagellum is directed backward into the cytostome, andthe others emerge anteriorly as free flagella. There are three anterior flagellain Chilomastix (Fig. 1a), but only one in Retortamonas (Fig. 1c). The recurrentflagellum (also known as the cytostomal flagellum) is usually short and entirelylocated in the cytostomal cavity in Chilomastix (Fig. 1a), but the recurrent flagellumof Retortamonas is proportionately longer, extending through the cytostomal grooveand emerging outside with a free terminal portion (Fig. 1c). The beating of anteriorflagella moves the cell, and undulations of the recurrent flagellum propel food intothe cytopharynx. Detailed descriptions of some species were published by Wenrich(1932) and Nie (1948).
Ultrastructure
CytoskeletonAlthough the two genera of retortamonads differ in the number of flagella, electronmicroscopy reveals that the kinetids are tetrakont in both, possessing four basalbodies (kinetosomes) arranged in two orthogonal pairs. Whereas all basal bodies inChilomastix bear flagellar axonemes (Fig. 3c), one basal body in each pair is barrenin Retortamonas (Figs. 2 and 5a). All flagella have the typical 9 + 2 organizationof axonemal microtubules. The cytostomal (recurrent) flagellum is modified by twoor three lateral vanes stiffened by paraxonemal lamellae (Figs. 2, 3a, e, and 5c, d).Electron microscopy provides unequivocal evidence that the recurrent/cytostomalflagellum of retortamonads does not form an undulating membrane attached to oneof the cytostomal lips, thus correcting earlier interpretations based on light micros-copy (Boeck and Tanabe 1926; Nie 1948).
The basal bodies also give rise to several different fibrillar roots, most of whichintegrate into the complex cytoskeleton of the cytostome. The most prominent is themicrotubular root that descends from the basal body of the recurrent flagellum(root 2, according to the nomenclature recommended by Yubuki et al. 2013), amajor support of the right cytostomal lip. In the kinetosomal area, this root originatesas a hooked row of microtubules that is rolled into an incomplete cylinder on its leftside (Fig. 2). The cavity of this “gutter” (Fig. 5b) opens posteriorly into thecytostomal groove (Fig. 5b, d). The curved band of the gutter microtubules separatesfrom the main row to form the inner subroot (the hooked band), which extends intothe cytopharynx as its microtubular support (Figs. 2 and 3b). The right portion of theroot gradually expands into a flat ribbon through the addition of microtubules andserves as a major support of the right wall of the cytostomal groove (Figs. 2 and 3b).The second microtubular root originating at the recurrent basal body descends to theleft wall of the groove, strengthening the left cytostomal lip (root 1, according toYubuki et al. 2013; Figs. 2 and 5d).
Another prominent structure originating at the kinetosomes is the striated(paracrystalline) lamella (composite fiber), which is adjacent to the inner faceof the microtubular root of the right cytostomal wall and extends posteriorly intothe cytopharynx (Figs. 2, 3a, b, e, f, and 4). Both lips of the cytostome areconnected at the top of the groove by an arched microfibrillar bundle, the
8 J. Kulda et al.
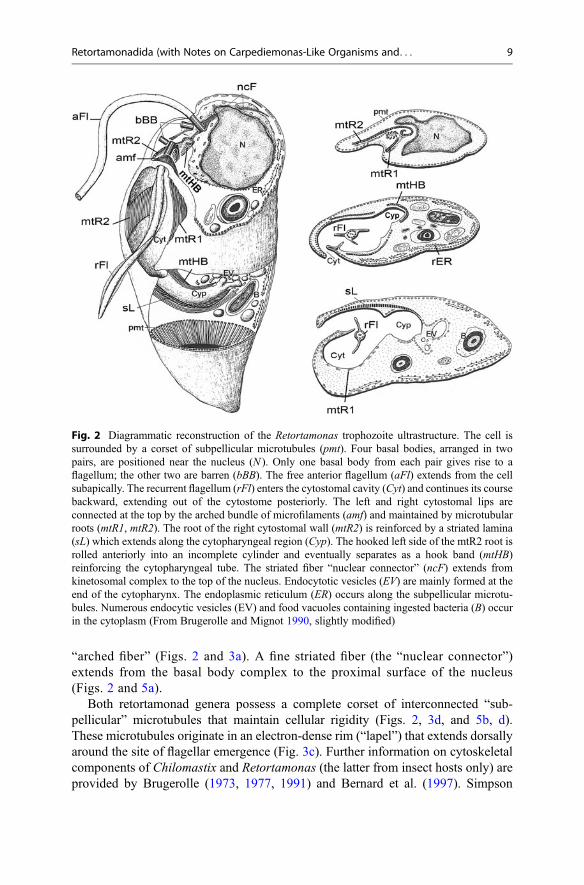
“arched fiber” (Figs. 2 and 3a). A fine striated fiber (the “nuclear connector”)extends from the basal body complex to the proximal surface of the nucleus(Figs. 2 and 5a).
Both retortamonad genera possess a complete corset of interconnected “sub-pellicular” microtubules that maintain cellular rigidity (Figs. 2, 3d, and 5b, d).These microtubules originate in an electron-dense rim (“lapel”) that extends dorsallyaround the site of flagellar emergence (Fig. 3c). Further information on cytoskeletalcomponents of Chilomastix and Retortamonas (the latter from insect hosts only) areprovided by Brugerolle (1973, 1977, 1991) and Bernard et al. (1997). Simpson
Fig. 2 Diagrammatic reconstruction of the Retortamonas trophozoite ultrastructure. The cell issurrounded by a corset of subpellicular microtubules (pmt). Four basal bodies, arranged in twopairs, are positioned near the nucleus (N ). Only one basal body from each pair gives rise to aflagellum; the other two are barren (bBB). The free anterior flagellum (aFl) extends from the cellsubapically. The recurrent flagellum (rFl) enters the cytostomal cavity (Cyt) and continues its coursebackward, extending out of the cytostome posteriorly. The left and right cytostomal lips areconnected at the top by the arched bundle of microfilaments (amf) and maintained by microtubularroots (mtR1, mtR2). The root of the right cytostomal wall (mtR2) is reinforced by a striated lamina(sL) which extends along the cytopharyngeal region (Cyp). The hooked left side of the mtR2 root isrolled anteriorly into an incomplete cylinder and eventually separates as a hook band (mtHB)reinforcing the cytopharyngeal tube. The striated fiber “nuclear connector” (ncF) extends fromkinetosomal complex to the top of the nucleus. Endocytotic vesicles (EV) are mainly formed at theend of the cytopharynx. The endoplasmic reticulum (ER) occurs along the subpellicular microtu-bules. Numerous endocytic vesicles (EV) and food vacuoles containing ingested bacteria (B) occurin the cytoplasm (From Brugerolle and Mignot 1990, slightly modified)
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 9

Fig. 3 The ultrastructure of trophozoites of Chilomastix caulleryi from a toad. (a) Longitudinalsection through the cytostomal cavity (Cyt) showing the recurrent flagellum (rFl) with lateral vanes,the microtubular roots (mtR1,mtR2) bordering the left and right cytostomal lips, and an arched bundleof microfilaments (amf) connecting both lips at the top of the cytostome (“arched fiber”). The striatedlamina (sL) adjacent to the mtR2 is curved posteriorly around the cytopharyngeal region (Cyp). One
10 J. Kulda et al.

(2003) published a proposal of universal terminology for flagellar roots and fibers inexcavates, which was revised by Yubuki et al. (2013).
Weerakoon et al. (1999) examined the localization of centrin in Chilomastixcuspidata cells using conventional and confocal immunofluorescent microscopy.The authors found that in addition to its localization in the basal body area, thecentrin signal colocalized with the tubulin signal along a microtubular root thatpasses through the cytostomal pouch. The major site of centrin association was mostlikely the hook band subroot of the cytostomal skeleton. Centrin is a calcium-binding protein that is a known component of the pericentriolar complex, but it isalso involved in contractility and a variety of signal functions. Weerakoon et al.(1999) speculated that root-associated centrin in the Chilomastix cytostome might beinvolved in signaling related to the capture and endocytosis of food particles in thecytopharynx.
Nucleus and Cytoplasmic OrganellesThe nucleus is always located in the most anterior part of the cell, close to the basalbodies (Figs. 1a, c, 2, and 3a). Its shape is almost spherical, with some depressions inthe area facing the basal bodies and the intracytoplasmic portion of the recurrentflagellum axoneme. Condensed material visualized by transmission electron micros-copy in the posterior part of the interphase nuclei of Retortamonas and Chilomastix(Brugerolle 1973, 1977) most likely represents the nucleolus and apparently corre-sponds to the membrane-associated chromatic plaques that give a negative result inFeulgen reactions, as described by Nie (1948).
The cytoplasm of the retortamonads contains numerous pinocytotic vesicles andlarge digestive vacuoles with ingested bacteria. Endocytosis occurs in the terminalparts of the cytopharynx, where the cytoskeletal armature is discontinuous. Therough endoplasmic reticulum is present, but it is usually not abundant. A single ordouble layer of reticulum cisternae is typically located at the cell periphery and
�
Fig. 3 (continued) anterior flagellum (aFl), the subpellicular microtubules (pmt), and the nucleus(N ) are also visible. (b) Transverse section through the cytopharyngeal region where the endocy-tosis occurs. Note the naked cytoplasmic membrane between the cytoskeletal structures of thecytopharynx, numerous pinocytic vesicles in the neighboring cytoplasm, and an engulfed bacterium(B) in the lumen of the cytopharynx. The derivatives of the hook band microtubules (mtHB) can beseen at both sides of the cytopharyngeal tube. Also shown is the striated lamina (sL) and a part of thecytostomal compartment (Cyt) with the recurrent flagellum (rFl). (c) The mastigont of Chilomastix,with two pairs of basal bodies and associated fibers. Basal bodies 2, 3, and 4 underlie the anteriorflagella; the recurrent one (1) give rise to recurrent/cytostomal flagellum. The arrow points to thelapel, the nucleating site of subpellicular microtubules. (d) Transverse section through the layer ofsubpellicular microtubules (pmt) interconnected by side arms. (e) Transverse section through thestriated lamina (sL) of the right wall of the cytostomal pouch underlain with rough endoplasmicreticulum (rER) and through the recurrent flagellum (rFl) with two lateral vanes. (f) Tangentialsection of the striated lamina showing its paracrystalline structure. (g) Chilomastix aulastomi cystfrom the horseleech is surrounded by a thick cyst wall (Cw) and containing the nucleus (N ),internalized flagella (Fl), and cytostomal fibers (Cyt) (From Brugerolle and Mignot 1990)
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 11

around the nucleus (Figs. 2 and 3a, e). No Golgi apparatus is apparent (at least in theform of organized stacks of dictyosomes), and peroxisome organelles are absent.Brugerolle (1973) observed in Chilomastix a circular reticulum of smooth cisternaebeneath the nucleus, but the function of this structure is unknown. Retortamonads donot possess typical mitochondria; however, double-membrane-bounded organelleswithout cristae that resemble hydrogenosomes have been demonstrated inChilomastix cuspidata (Hampl and Simpson 2008) and other Chilomastix species(Fig. 4). Additional cytoplasmic components of retortamonads include free ribo-somes and inclusions of multigranular glycogen.
The Cyst Stage
The cyst is the resting stage that is resistant to unfavorable environments and servesto transmit infection. Retortamonad cysts are ovoid, pyriform, or lemon-shaped andexhibit a thick cell wall made of filamentous material (Figs. 1b, d and 3g). There isno operculum or pore in the cyst wall. The interior of the cyst contains basal bodieswith internalized flagella, the retained cytoskeletal armature of the cytostome andcytopharynx, glycogen granules, and a single nucleus (Fig. 3 (12)). Brugerolle(1973) observed precystic stages of Chilomastix with flagella withdrawn into thecytoplasm, partly disorganized “subpellicular” microtubules, and numerous secre-tory vesicles trafficking cyst wall material to the cell surface. Retortamonads do notdivide inside the cyst. A previous description of within-cyst division by Kofoid andSwezy (1920) has not been confirmed by subsequent investigators (Geiman 1935).
Reproduction
Trophozoites of retortamonads reproduce by binary fission, but the details of theirmitosis and cell division are not well known. Somewhat conflicting observations byearly authors and electron microscopic evidence (Brugerolle 1973, 1977) suggestthat the process is similar to that observed in diplomonads (Brugerolle 1974).Mitosis proceeds inside the nucleus, which is enveloped by the persisting nuclearmembrane, with the aid of intranuclear microtubules, but the mitotic spindle nucle-ates outside the nucleus as a “hemispindle” (i.e., the process is a type of semi-openmitosis). The two pairs of basal bodies (kinetosomes) separate during prophase andlocalize to the nuclear poles (Boeck and Tanabe 1926). The spindle microtubulesthen nucleate in the perikinetosomal area of the polar basal bodies (see Fig. 7 inBrugerolle 1973). The formation of a “paradesmose” (the extranuclear spindlecharacteristic of mitosis in Parabasalia), reported by some investigators (Bishop1934, 1935; Geiman 1935), does not in fact occur in retortamonads.
The chromosomes condense in early prophase; six have been detected inChilomastix intestinalis by the Feulgen reaction (Nie 1948). The cytostome of theparent cell disintegrates by the end of the prophase, and the de novo assembly of thedaughter organelles is completed during telophase (Nie 1948). The daughter
12 J. Kulda et al.

mastigonts are fully reorganized during the final phase of cytokinesis. The parentflagella are probably retained and redistributed to the daughter mastigonts (Bishop1931, 1934). Further studies employing advanced methodology are necessary togain better insight into mitosis and cell division in retortamonads.
TaxonomyGrassé (1952) established the order Retortamonadida and placed it in the superorderMetamonadina, which at the time encompassed oxymonads, retortamonads, andseveral orders of parabasalids. Metamonads were later transferred to Archezoa(Cavalier-Smith 1983, 1998; see below). Alternatively, Simpson and Patterson(1999) proposed an “excavate hypothesis” grouping flagellates equipped with aventral groove associated with the recurrent flagellum and a characteristic cytoskel-eton. After several revisions of the metamonad grouping, Cavalier-Smith (2003)returned to something similar to Grassé’s concept, which he modified by includingdiplomonads and the free-living anaerobe Carpediemonas. Cavalier-Smith (2003)also abandoned Archezoa as a taxon and placed the Metamonada as a phylum of theExcavata infrakingdom. Meanwhile, Simpson (2003) proposed Fornicata as a taxonto include a phylogenetically coherent subset of metamonads, consisting ofretortamonads, diplomonads, and Carpediemonas (and later including the subse-quently characterized Carpediemonas-like organisms; see below and Kolisko et al.
Fig. 4 Hydrogenosome-like organelles of Chilomastix caulleryi. Transmission electron micro-graph of section through area close to the right cytostomal lip shows the elongate hydrogenosome-like organelles (H ). Also shown is the striated cytostomal lamina (sL) adjacent to microtubules ofthe mtR2 root. As apparent at higher magnification (inset), the organelles are limited by two closelyadjacent membranes (arrow) and lack cristae
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 13

Fig. 5 The ultrastructure of Retortamonas from crane fly larvae. (a) Basal body arrangement inRetortamonas. The basal bodies, localized close to the nucleus (N ), are grouped in two pairs: theanterior pair (2, 4) and the posterior pair (3, 1). Basal body 2 gives rise to the anterior flagellum(aFl); the recurrent flagellum (rFl) originates from basal body 1. The basal bodies 3 and 4 arebarren. The origin of the nuclear connector fiber (nc) can be seen at the base of the basal body 2.(b) Transverse section through the upper part of the cytostomal pocket, showing the opening of thegutter (gt) into the cytostomal cavity to form the cytopharyngeal tube. The density attached distallyto mtR1 microtubules is an anchoring point for microfilaments of the “arched fiber” (amF). The cellmembrane of Retortamonas is supported by a corset of subpellicular microtubules (pmt).(c) Transverse section through the central part of the cytostomal pocket showing the recurrent
14 J. Kulda et al.

2010). In the revised classification of eukaryotes proposed by the InternationalSociety of Protistologists (Adl et al. 2012), Retortamonadida, together withDiplomonadida and Carpediemonas-like organisms, are placed in the Fornicata(Simpson 2003), a subgroup of Metamonada (Cavalier-Smith 1987), as emendedby Cavalier-Smith (2003), within the supergroup Excavata (Cavalier-Smith 2002),as emended by Simpson (2003).
Order: Retortamonadida Grassé 1952
Retortamonadida are Fornicata with a single karyomastigont possessing four basalbodies and two or four flagella. One flagellum is recurrent, has two or three lateralvanes, and is associated with the ventral cytostomal pouch. In the phylogenetic treeof Fornicata (Fig. 6), Retortamonadida is split into two different clades, indicatingthat Retortamonadidae is currently polyphyletic (see ▶Evolutionary History).
Family: Retortamonadidae, Wenrich 1932
Characteristics as for order Retortamonadida
Genus: Retortamonas Grassi 1879
Objective synonym: Plagiomonas Grassi 1881Subjective synonyms: Embadomonas Mackinnon 1911; Waskia Wenyon and
O’Connor 1917.Members of the genus possess one anterior and one recurrent flagellum. The
recurrent flagellum is directed into the cytostomal pocket, from which a free distalportion of the flagellum extends. Distinctive morphological characters among spe-cies include body shape and size, the size of the posterior spike, the relative length ofthe flagella, the length of the free portion of the cytostomal flagellum, and the size ofthe cytostome. All known species are intestinal endobionts of insects andvertebrates.
Type species:Retortamonas gryllotalpae Grassi 1879 [Synonym: Plagiomonas gryllotalpae
Grassi 1881], from the intestine of the mole cricket. The species was redescribedby Wenrich (1932).
�
Fig. 5 (continued) flagellum (rFl) with three vanes and the microtubular roots supporting the right(mtR2) and the left (mtRl) wall of the cytostome (Cyt). Digestive vacuoles (dV), some with ingestedbacteria, are present in the cytoplasm. (d) Transverse section through the posterior part of thecytostome (Cyt) and the cytopharynx (Cyp) showing the recurrent flagellum (rFl) with two vanessectioned, the microtubules of the left cytostomal wall (mtR1), the striated lamina (sL) whichextends to the cytopharyngeal tube, and the cytopharyngeal derivatives of the hooked bandmicrotubules (mtHB). A digestive vacuole (dV) and an endocytosed bacterium (B) are shown inthe cytoplasm. Note the corset of subpellicular microtubules (pmt) (Pictures by courtesy ofG. Brugerolle)
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 15

More than 20 species of Retortamonas have been described. The retortamonadsfrom crane fly larvae, Retortamonas agilis (Mackinnon 1911) and Retortamonasalexeieffi (Mackinnon 1912), were subjected to relatively detailed electron micro-scopic studies (Brugerolle 1977) and thus represent the model species of the genus,though Brugerolle (2006) also briefly described the ultrastructure of R. hodotermitis.Other Retortamonas species were found in larvae of beetles (R. caudacus,R. phylophagae), in roaches (R. blattae), termites (R. termitis, R. hodotermitis),water bugs (R. belostomae), amphibians (R. dobelli), reptiles (R. boae,R. saurarum, R. testudae), and various mammals including humans (e.g.,R. intestinalis, R. bradypi, R. cuniculi, R. mitrula, R. ovis, R. caviae,
Fig. 6 Phylogenetic tree of Fornicata based on SSU rDNA sequences. The tree was constructed bythe maximum likelihood method in RAxML (GTRGAMMAI model). The values at the branchesrepresent statistical support in bootstrap values (RAxML)/posterior probability (MrBayes); supportvalues below 50/0.95 are not shown. The putative position of Retortamonas from insects is shownin the tree
16 J. Kulda et al.

R. ruminantium). The ostrich Struthio camelus is only avian host of retortamonasreported (MartinezDiaz et al. 2001). For an almost complete list of Retortamonasspecies, see Ansari (1955).
Genus Chilomastix Alexeieff 1912
Objective synonym: Macrostoma Alexeieff 1909Subjective synonyms: Fanapepea Prowazek 1911; Cyathomastix Provazek and
Werner 1915; Difamus Gäbel 1914; Tetrachilomastix da Fonseca 1915.Members of this genus are endobiotic or free-living Retortamonadidae with three
anterior and one recurrent flagellum. The recurrent flagellum is located in thecytostomal pocket and does not extend from it. Distinctive features among speciesinclude body size and shape; the relative length of the posterior spike; the relativelength of the anterior flagella; the size, shape, and position of the cytostome; and, insome species, the supposed host specificity. Members of the genus are intestinalendobionts of many classes of vertebrates (including humans) and some inverte-brates. One species is free-living.
Type species:Chilomastix caulleryi (Alexeieff 1909) [Macrostoma caulleryi Alexeieff 1909]This species is commonly found in the cloaca of amphibians. The original generic
name Macrostoma was rejected because it is a junior homonym of MacrostomaRisso 1826.
Over 30 species of Chilomastix have been described. Light microscopic studies ofgeneral importance were published by Boeck and Tanabe (1926), Geiman (1935),Bishop (1935), Nie (1948, 1950), McDowell (1953), and Russel Gabel (1954). Theultrastructure of the genus was studied by Brugerolle (1973) and Bernardet al. (1997).
Examples of Chilomastix species:Chilomastix mesnili (Wenyon 1910) [Synonyms: Fanapepea intestinalis Pro-
wazek 1911, Cyathomastix hominis Prowazek and Werner 1914, Difamus tunensisGäbell 1914, Tetrachilomastix bengalensis Chatterjee 1923] occurs in the cecum andcolon of humans, monkeys, and pigs. The susceptibility of monkeys and pigs tohuman isolates was confirmed experimentally (Kessel 1924, 1928). The potentialpathogenicity of this species in humans is discussed above.
Other Chilomastix species with mammalian hosts were described from horses,goats, cattle, hares, and rabbits. The major rodent species, with relatively wide hostspectra, are Chilomastix bettencourti da Fonseca 1915 and Chilomastix intestinalisKuczynski 1914. The former is commonly found in laboratory and wild rats, as wellas mice, hamsters, voles, and gerbils. The latter occurs in guinea pigs, hamsters, andrabbits. Another frequent inhabitant of guinea pig cecum is Chilomastix wenrichiNie 1948.
Chilomastix gallinarum Martin and Robertson 1911 is a common avian speciespresent in the ceca of chicken, ducks, turkeys, pheasants, quails, and partridges.Boeck and Tanabe (1926) and McDowell (1953) published morphological studies ofthis species. Its pathogenic potential is discussed above. Three Chilomastix species
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 17

were recorded in marine fishes (Lavier 1936), and C. bursa Moskowitz 1951 in theintestines of frogs.
The type species of the genus C. caulleryi (Alexeieff 1909) is found in the cloacaof anuran and urodelan amphibians. This large species can apparently infect thehorseleech Haemopis sanguisuga in shared biotopes; however the autochthonousspecies of the horseleech, C. aulastomi Bělař 1921, is also identifiable bymorphology.
The host list, with pertinent references and further information on Chilomastixspecies from vertebrates, is available from Kulda and Nohýnková (1978). Speciesliving in invertebrates such as termites and sea urchins are listed by Grassé (1952).
The free-living species Chilomastix cuspidata (Larsen and Patterson 1990),originally assigned to the genus Percolomonas, is widely distributed in anoxicmarine, brackish, and freshwater habitats (Bernard et al. 1997, 2000).
Maintenance and Cultivation
Various species of the Retortamonadida from vertebrates, including humans, can beisolated and maintained in xenic cultures (i.e., cultures containing a concomitantmixture of unidentified bacteria). None of the retortamonad isolates have beengrown axenically, and no attempts to obtain monoxenic cultures (or any other typeof gnotobiotic cultures) have been reported. Some isolates of Retortamonas andChilomastix spontaneously form cysts in culture.
The crude biphasic media originally developed for the cultivation of intestinalamoebae, such as Locke-egg-serum medium (Boeck and Drbohlav 1925) or serum-Ringer-egg medium (Dobell and Laidlaw 1926), are still the best choices for theisolation of primary cultures of endobiotic species. Cultures established in biphasicmedia can usually be transferred and maintained in simple liquid media, such asLoeffler’s serum-Ringer medium (Wenrich 1947) or TYSGM medium (Diamond1982). The optimal pH for the cultivation of retortamonads is 7.2–7.4, and therecommended temperatures for mammalian and avian species are 35–37 �C and38–40 �C, respectively. Under these conditions, cultures should be transferred at 2-to 3-day intervals. Isolates from amphibians, reptiles, and some invertebrates grow atroom temperature (18–25 �C), and transfers at 1- or 2-week intervals arerecommended.
The free-living species Chilomastix cuspidata can be maintained in seawater-Cerophyl medium (ATCC Medium 1525, American Type Culture Collection, www.attc.org). The medium consists of a 1:1 mixture of filter-sterilized artificial seawaterand an autoclaved solution prepared by boiling rye grass Cerophyl or its equivalent.
In our experience, isolates of retortamonads can be cryopreserved in the presenceof 5% dimethyl sulfoxide using a standard procedure and stored indefinitely in liquidnitrogen.
Reports on the cultivation of retortamonads from humans and animals werepublished by Hogue (1921), Boeck (1921), Boeck and Tanabe (1926), Collier andBoeck (1926), Bishop (1934), and Dobell (1935); brief procedures are also available
18 J. Kulda et al.

in publications of Silberman et al. (2002), Cepicka et al. (2008), and Takishitaet al. (2012).
Chilomastix cuspidata and two Retortamonas cultures can be obtained from theAmerican Type Culture Collection; several isolates of Retortamonas andChilomastix from mammals, amphibians, and reptiles are available in the culturecollection of the Department of Parasitology, Charles University in Prague.
Evolutionary History
Retortamonads were once thought to belong to the “Archezoa,” a group of putativelyprimitive eukaryotes that diverged before the acquisition of mitochondria by anancestor of all living eukaryotes with this organelle (Cavalier-Smith 1983, 1987).Famously, the “archezoa hypothesis” has declined in popularity in the twenty-firstcentury, as organelles interpreted as mitochondrial derivatives (e.g.,hydrogenosomes and mitosomes) have been found in most putative archezoans(see Lindmark and Müller 1973; Simpson and Roger 2004; Tovar et al. 1999,2003; Tachezy and Šmíd 2007). Double-membrane-bounded organelles resemblinghydrogenosomes have been found also in Chilomastix cuspidata (Hampl andSimpson 2008) and some other Chilomastix species (Fig. 4 (13)). However, asidefrom gross morphology, no data are currently available on the biogenesis andfunction of these organelles in retortamonads.
The retortamonads are currently classified within the taxon Fornicata (Simpson2003). According to phylogenetic analyses based on the SSU rRNA gene and aconcatenate of several protein-coding genes (Takishita et al. 2012; Cepicka et al.2008), the genera Retortamonas and Chilomastix form two unrelated clades.Retortamonas, represented by species from vertebrates, appears to be the closestrelatives of diplomonads, while Chilomastix branched within the paraphyletic gradeof Carpediemonas-like organisms. Cepicka et al. (2008) suggested thatRetortamonas might be polyphyletic as well, because species from insects possessa complete microtubular corset, similar to that of Chilomastix, while the species fromvertebrates seem to lack it (Cepicka et al. 2008). Accordingly, sequence data fromRetortamonas species from insects, presented by Smejkalová et al. at InternationalCongress of Protistology in Vancouver, Canada (2013), indicate that insectretortamonads are closely related to Chilomastix, thus confirming the polyphyly ofthe genus Retortamonas (see also Fig. 6 (18)). These findings indicate that taxo-nomic revision of Retortamonadida can be expected soon.
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 19

Coda: Carpediemonas-Like Organisms and Caviomonadidae
Introduction
Carpediemonas-like organisms (CLOs) are a small assemblage of poorly knownexcavates. CLO cells are usually biflagellate, with one anterior and one recurrentflagellum, and possess a longitudinal ventral groove. All CLOs are anaerobic andalmost all are free-living, occurring in marine hypoxic sediments. They seem to berelatively common in such environments and have been isolated worldwide. Variousmedia for anaerobic free-living marine protists were shown to be suitable for free-living CLOs (see Kolisko et al. 2010). It was shown only recently that the endobioticgenus Caviomonas, which had been previously thought to be a highly reduceddiplomonad, belongs to CLOs, being closely related to the endobiotic genusIotanema and free-living, marine isolate PCS (Yubuki et al. 2016). Becausecaviomonads substantially differ from other CLOs by possessing a single flagellumand a highly reduced flagellar apparatus, they will mainly be treated in a separatesection (see below). Together with diplomonads and retortamonads, the CLOs formthe clade Fornicata. Recent molecular phylogenetic analyses showed that CLOs areparaphyletic.
The first known CLO was Kipferlia bialata, described by Ruinen (1938) asCryptobia bialata. This species was, however, neglected for more than 50 yearsand was not recorded again before the end of the twentieth century (Fenchel et al.1995). Two species of Caviomonas were described in 1950 and 1970, respectively(Nie 1950; Navarathnam 1970). Carpediemonas membranifera, the best knownCLO, was described by Larsen and Patterson (1990) as Percolomonasmembranifera, but was later transferred by Ekebom et al. (1996) to their newlycreated genus Carpediemonas. An ultrastructural study of C. membranifera bySimpson and Patterson (1999) was crucial for the formulation of the so-calledexcavate hypothesis. The morphological similarity of Carpediemonas membraniferaand Cryptobia bialata was noted by Lee and Patterson (2000), who transferred thelatter species to the genus Carpediemonas. The remaining species of free-livingCLOs, Dysnectes brevis, Hicanonectes teleskopos, Ergobibamus cyprinoides, andAduncisulcus paluster, were all described recently, as was the caviomonad Iotanemaspirale (Yubuki et al. 2007, 2016; Park et al. 2009, 2010). At the same time, it wasshown by SSU rRNA gene phylogenies that Carpediemonas membranifera andC. bialata are not closely to each other, and the genus Kipferlia was therefore createdfor C. bialata (Kolisko et al. 2010); the distant relationship between Carpediemonasand Kipferlia was corroborated by a multigene phylogenetic analysis of Fornicata(Takishita et al. 2012) and an ultrastructural study of Kipferlia bialata (Yubuki et al.2013).
20 J. Kulda et al.

Light Microscopy
CLOs are small flagellates (5–18 μm) with two heterodynamic flagella and aconspicuous ventral groove (but see below for Caviomonadidae). The trophozoitesare naked and possess a single, anteriorly located nucleus. The flagella are insertedslightly subapically, on the ventral side of the cell, close to the nucleus. Oneflagellum (anterior flagellum, F2) is directed anteriorly and is approximately thesame length as the cell. The other flagellum (recurrent, R, F1) is directed posteriorly,runs through the ventral groove, and continues behind the cell. This flagellum isusually substantially longer than the cell; in Dysnectes, however, the recurrentflagellum is shorter than the cell body. The recurrent flagellum beats in the ventralgroove and creates a current that draws bacteria into the posterior part of the groovewhere they are ingested. The longitudinal groove occupies almost the whole ventralside of the cell.
Slow movement with little rotation is typical of Carpediemonas andErgobibamus; occasional and very slow swimming is characteristic of Dysnectes,which usually lies on the substratum and rocks back and forth by moving the anteriorflagellum. Kipferlia also swims slowly and often adheres to the substratum (Yubukiet al. 2013), whereas Hicanonectes rotates rapidly while moving (Park et al. 2009).
Flagellar Apparatus
In general, the organization of the flagellar apparatus of CLOs is similar to that ofretortamonads (but see below for Caviomonadidae). Most CLOs possess two flagella(members of Caviomonadidae possess a single flagellum; see below), the recurrentflagellum (F1, R) and the anterior flagellum (F2) (Fig. 8a–e). Transmission electronmicroscopy shows that they differ in the number of basal bodies; however; inaddition to the two basal bodies bearing flagellar axonemes (i.e., basal bodies1 and 2), there are barren basal bodies: one in Carpediemonas and Caviomonas;two in Kipferlia (Fig. 8b), Ergobibamus, Aduncisulcus, and Hicanonectes; and threein Iotanema. Ultrastructurally, the basal bodies and the flagellar axonemes have atypical eukaryotic organization. As in retortamonads, the anterior flagellum of CLOsis smooth with no accessory structures, whereas the recurrent flagellum has one tothree longitudinal vanes, each supported by a paraxonemal lamella with differentultrastructures and unknown biochemical composition (though these have been lostin Caviomonadidae). The ventral vane is always the broadest and is the only vanepresent in Kipferlia (Fig. 8c).
The two flagella-bearing basal bodies are arranged almost perpendicularly or at aslightly obtuse angle (Fig. 8a). The basal body of the anterior flagellum (basal body2) is associated with the anterior root which consists of a single (Kipferlia,Dysnectes) (Yubuki et al. 2007, 2013) or several microtubules (e.g., two inCarpediemonas, five in Aduncisulcus, six in Ergobibamus, and nine inHicanonectes; Simpson and Patterson 1999; Park et al. 2009, 2010; Yubuki et al.2016). The anterior root (also known as root 3; see Yubuki et al. 2013) originates
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 21

Fig. 7 Light microscopic photographs of free-living Carpediemonas-like organisms. (a), (b)Carpediemonas membranifera. (c), (d) Kipferlia bialata. (e) Dysnectes brevis. (f) Dysnectessp. SIVOTA. (g) Hicanonectes teleskopos. (h) Ergobibamus cyprinoides. (i), (j) Carpediemonas-like organism NC. (k), (l) Aduncisulcus sp. PCE. (m), (n) Aduncisulcus paluster. Scale bar is 5 μmfor all figures (From Kolisko et al. 2010, with permission of the Society for Applied Microbiologyand Blackwell Publishing Ltd.)
22 J. Kulda et al.

Fig. 8 The ultrastructure of Kipferlia bialata. (a) A longitudinal section through the anterior part ofthe cell showing perpendicularly arranged basal bodies of the recurrent flagellum (1) and anteriorflagellum (2) and a dense fiber (dF) originating dorsally to basal body 2 and extending along themicrotubular root 3 (mtR3). The recurrent flagellum (Fl1), microtubules of the left root (mtR1), andthe B fiber can also be seen. (b) A transverse section through the anterior part of the cell showing thebasal body of the recurrent flagellum (1), the barren basal bodies 3 and 4, and associated structures ofthe mastigont including the left and the right microtubular roots (mtR1 andmtR2), and the A, B,C, andI fibers. The I fiber (arrowhead) reinforces the ventral face of mtR2. The B fiber stretches from mtR1
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 23

from the anterior side of basal body 2 and curves to run posteriorly down the left sideof the cell (except in Hicanonectes, where the root travels anteriorly). InAduncisulcus, Carpediemonas, Ergobibamus, and Hicanonectes, the anterior rootis closely associated with microtubules of a dorsal fan, which supports the dorsal cellmembrane, much like the subpellicular microtubules in Retortamonas andChilomastix. The extent of the fan differs across CLO genera and is entirely absentin Kipferlia and Dysnectes (Yubuki et al. 2007, 2013).
The basal body of the recurrent flagellum (basal body 1 or R) is associated withcytoskeletal structures that support the ventral groove, namely, the left microtubularroot (R1 or LR), the right microtubular root (R2 or RR), the singlet microtubularroot, and several non-microtubular fibers (A, B, C, and I fibers) (Fig. 8b).
The right root originates from the right (or right dorsal) side of the basal body 1 asa single concavely curved row of 15–18 microtubules (Fig. 8b) that split into twobranches near the distal end of the basal body 1: a narrow inner branch (iR2 or IRR)and a more conspicuous outer branch (oR2 or ORR) (Fig. 8c–e). Both branchesextend posteriorly. In most CLOs, the outer branch expands through additionalmicrotubules and forms a ribbon supporting the right wall of the groove; the innerbranch is associated with the floor of the groove. In Kipferlia, the only CLO with adeep gutter within the ventral groove, the outer and inner root branches support theright and left walls of the gutter, respectively (Fig. 8c–e).
The right root is accompanied by two fibers: the short A fiber (on the dorsal side)and the layered I fiber closely associated with its ventral side through fine fibrousconnections between the fiber and the root microtubules (Fig. 8b). Posterior to theroot splitting, the I fiber continues with the outer branch only and usually terminateshalfway down the groove, whereas the root microtubules extend farther. Kipferliaand Hicanonectes cells have a prominent, posteriorly located cytopharynx, and thisis supported by several microtubules ultimately derived from the right root.
The left root originates from the left side of the basal body 1. It is formed from asingle row of closely adjacent microtubules, which extend posteriorly to support theleft margin of the ventral groove. The left root is associated with two fibers. TheC fiber is found on the dorsal side of the left root (Fig. 8b). The fiber is short,
�
Fig. 8 (continued) to mtR2 (and the I fiber), around the ventral side of the recurrent basal body (1).(c) Transverse section through the anterior part of the cell showing the ventral groove (G) borderedby the hood (H ) and the gutter (gt) on the groove floor. The mtR1 root and individual microtubules(arrowheads) support the left wall of the groove. Note two microtubular bands (oR2 and iR2)resulting from splitting of the right root. The I fiber reinforces the oR2 band. The recurrent flagellumwith a prominent vane (arrow), the nucleus (N ), and the B fiber supporting the hood are also visible.(d) Transverse section through the anterior part of the gutter and its associated structures: themicrotubular bands oR2 and iR2, microtubules (arrowheads) beneath the gutter membrane, and thefibers I and A. The left root supports the left wall of the apical part of the ventral groove covered bythe hood. The axoneme of the recurrent flagellum ( fl1Ax) is visible shortly before the flagellumdescends into the ventral groove. (e) A detailed view of the gutter region (more posteriorly than in36) to show organization of cytoskeleton that supports the wall of the gutter. oR2 and iR2, fiveindividual microtubules (arrowheads), and the I fiber are shown (Pictures by courtesy of N. Yubuki)
24 J. Kulda et al.

terminating at the level of the groove opening, except in Ergobibamus in which the Cfiber runs farther along the left root. The B fiber connects the left and right roots. Thefiber traverses the ventral side of the basal body of the recurrent flagellum (Fig. 8b).The left region is always short and located ventral to the left root. The right region,beneath the cell membrane, descends the right side of the groove with the outerbranch of the right root. In Kipferlia, the B fiber is the main support for the hood, amembranous lip covering the anterior region of the cell above the anterior flagellumand extending down the left and right margins of the ventral groove (Fig. 8c; Yubukiet al. 2013).
Flagellar Transformation
During the cell division of Kipferlia bialata, the parental anterior flagellum(F2) becomes the recurrent flagellum (F1) in one daughter cell, while the parentalrecurrent flagellum (F1) remains recurrent in the other daughter cell. In bothdaughter cells, the anterior flagellum is formed de novo (Yubuki et al. 2013).Thus, the flagella in K. bialata are inherited in a semiconservative way, and therecurrent flagellum is older than the anterior flagellum. Kipferlia thus represents thefirst “typical excavate” in which flagellar transformation has been thoroughlydocumented (see Nohýnková et al. (2006) for flagellar transformation in the related,but atypical Giardia intestinalis).
Nucleus and Cytoplasmic Organelles
The nucleus of the CLOs is always located at the anterior part of the cell, in closeassociation with the basal bodies. Its shape is mostly spherical except for the ovoidnucleus of Ergobibamus. The nucleus lacks a central nucleolus. Instead, electron-dense material forms a large mass located either subcentrally (Carpediemonas) or onone side of the nucleus (Hicanonectes and Aduncisulcus); alternatively this massmay form several small aggregates beneath the nuclear envelope, as in Ergobibamus(Simpson and Patterson 1999, Park et al. 2009, 2010).
Typical mitochondria are absent; however, double-membrane-bounded organ-elles that resemble hydrogenosomes are present in all CLO genera, at an abundanceof several per cell. The limiting membranes of these organelles are very closelyadjacent and do not form cristae. In most genera, these organelles are rounded andare 300–500 nm in diameter, though in Carpediemonas, they are elongated ordumbbell-shaped (Simpson and Patterson 1999; Park et al. 2009, 2010; Yubukiet al. 2007, 2013, 2016).
No discrete Golgi apparatus was observed in most CLOs. The exception isCarpediemonas membranifera, in which a single Golgi dictyosome with three tofour cisternae is placed left of the ventral groove and ventral to the nucleus (Simpsonand Patterson 1999).
Food vacuoles are usually located in the posterior two-thirds of the cell.
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 25

Ultrastructure of Cysts
The cyst stage of Hicanonectes telescopos and Iotanema spirale was observed byTEM (Park et al. 2009; Yubuki et al. 2016). The rounded cyst of Hicanonectes iscovered by a relatively thin, single-layered cyst wall, which is separated by emptyspace from the cytoplasm. The nucleus, hydrogenosome-like organelles, and inter-nalized flagellar axonemes are located within the cytoplasm. Only a single cyst ofIotanema spirale was documented, and almost no details were observed (Yubukiet al. 2016).
Taxonomy and Evolutionary History
The CLOs are members of the Fornicata. To date, six species of free-living CLOshave been described (see below for Caviomonadidae), each currently belonging to aseparate genus: Aduncisulcus paluster, Carpediemonas membranifera, Dysnectesbrevis, Ergobibamus cyprinoides, Hicanonectes teleskopos, and Kipferlia bialata(Ekebom et al. 1996; Kolisko et al. 2010; Larsen and Patterson 1990; Park et al.2009, 2010; Ruinen 1938; Yubuki et al. 2007, 2016). This assignment of knownCLOs to described genera is now replacing a more informal division into cladeslabeled CL1-CL6 (Kolisko et al. 2010).
Cavalier-Smith recently (2013) divided CLOs into three families: Carpedie-monadidae Cavalier-Smith 2003 (Carpediemonas, Hicanonectes, andErgobibamus), Kipferliidae Cavalier-Smith 2013 (Kipferlia), and DysnectidaeCavalier-Smith 2013 (Dysnectes). However, the family Carpediemonadidae sensuCavalier-Smith 2013 is paraphyletic. Aduncisulcus has not been accommodated inany family (Yubuki et al. 2016).
Almost all CLOs were unknown before the end of the twentieth century, andconsequently their phylogenetic affinities were barely considered. Simpson andPatterson (1999) recognized the so-called excavate taxa, to which they assignedCarpediemonas membranifera (among others), based on the presence of the ventralgroove and the associated cytoskeleton. The specific relationship of CLOs,diplomonads, and retortamonads was revealed in 2002, when sequence data forCarpediemonas membranifera became available (Simpson et al. 2002); the threegroups were subsequently united in the taxon Fornicata (Simpson 2003). It was latershown by analyses of the SSU rRNA gene that the CLOs are diverse and likelyrepresent several distinct lineages (Kolisko et al. 2010; Park et al. 2009; Yubuki et al.2007). Notably, Dysnectes brevis appeared to be the closest relative of theDiplomonadida + vertebrate Retortamonas clade (except for the report of Yubukiet al. 2007 in which this species branched more basally than Carpediemonasmembranifera), but the SSU rRNA gene showed little power to resolve interrelation-ships within the Fornicata. A multigene analysis (Takishita et al. 2012) confirmedthat the CLOs are paraphyletic, with diplomonads and retortamonads nested withinthem. Briefly, Fornicata split into four clades without resolved interrelationships:(1) Carpediemonas, (2) Ergobibamus, (3) Hicanonectes and Aduncisulcus (and
26 J. Kulda et al.

Caviomonadidae, see below), and (4) Chilomastix, Kipferlia, Dysnectes,Retortamonas, and Diplomonadida. Within the latter Dysnectes was closely relatedto the Retortamonas + Diplomonadida lineage.
Recently it was established that the endobiotic genus Caviomonas, which hadpreviously been considered an ▶Enteromonad Diplomonad, actually descendedindependently from CLOs (see below).
Morphology, Ultrastructure, and Ecology of Caviomonadidae
Family Caviomonadidae, originally created by Cavalier-Smith (2013), currentlyconsists of genera Caviomonas and Iotanema and an undescribed isolate PCS.Members of Caviomonadidae are small uniflagellate organisms with a single nucleusand no clear ventral groove (Fig. 9a–d). Preliminary electron microscopy ofCaviomonas and Iotanema shows a simple flagellar apparatus, with two or fourbasal bodies, respectively (Brugerolle and Regnault 2001; Yubuki et al. 2016). Onebasal body (B2) bears the single flagellum, which was identified as F2 (i.e., homol-ogous to the anterior flagellum of CLOs and other excavates). One (Caviomonas) orthree (Iotanema) basal bodies are barren, including B1, which bears the recurrentflagellum F1 in other fornicates. The cytoskeleton associated with the basal bodies issimple and consists of a single microtubular fiber (called the nuclear fiber) and adorsal fan of microtubules. The nuclear fiber is accompanied by a multilayered fiberand a connecting fiber (both are non-microtubular; the latter was observed only inIotanema). It has been putatively homologized with root R1 of typical excavates,though more data are needed (see Yubuki et al. 2016). Mitochondria have not yetbeen detected in Caviomonadidae.
Genus Caviomonas comprises two endobiotic species, C. mobilis andC. frugivori, found in the large intestine of rodents (Nie 1950; Navarathnam1970); they are considered harmless commensals. The only species of Iotanema,
Fig. 9 Light microscopic photographs of Caviomonadidae. (a), (b) Free-living strain PCS. (c), (d)Iotanema spirale. Scale bar is 5 μm for all figures ((a), (b) from Kolisko et al. 2010, with permissionof the Society for Applied Microbiology and Blackwell Publishing Ltd.; (c), (d) by courtesy ofN. Yubuki)
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 27

I. spirale, was obtained from feces of a gecko (Yubuki et al. 2016). By contrast, theorganism “PCS” is free-living and was isolated from marine anoxic sediments(Kolisko et al. 2010). Iotanema spirale was successfully cultured in serum-Ringer-egg medium (Dobell and Laidlaw 1926); see Kolisko et al. (2010) for suitableculture medium for PCS.
Phylogenies of the SSU rRNA gene of Iotanema and PCS show a close relation-ship with the CLO Hicanonectes (Yubuki et al. 2016); sequence data of Caviomonasare still unavailable. It was concluded that Caviomonadidae are descended fromCLOs, losing most of the typical excavate features and, in the case of Caviomonas(and almost certainly Iotanema), becoming endobionts. Therefore, they represent thethird origin of the endobiotic lifestyle in Fornicata, independent of bothretortamonads sensu stricto and the diplomonads + “vertebrate Retortamonas”lineage.
Acknowledgments The authors would like to thank Guy Brugerolle for the kind permission to usethe micrographs featured in Fig. 3a–g and Naoji Yubuki for the kind permission to use themicrographs featured in Figs. 8a–e and 9c, d. This work was supported by the Czech ScienceFoundation (project GA14-14105S).
References
Adl, S. M., Simpson, A. G. B., Lane, C. L., et al. (2012). The revised classification of eukaryotes.Journal of Eukaryotic Microbiology, 59, 429–493.
Alexeieff, A. (1912). Sur quelques noms de genres des flagellés qui doivent disparaitre de lanomenclature pour cause de synonymie ou pour cause autre raison. Diagnoses de quelquesgenres récemment étudiés. Zoologischer Anzeiger, 39, 674–680.
Alexeieff, A. (1917). Mitochondries et corps parabasal chez les Flagellés. Comptes Rendus desSeances de la Societe de Biologie et de Filiales, 80, 358–361.
Ansari, M. A. R. (1955). The genus Retortamonas Grassi (Mastigophora, Retortamonadidae).Biologia Lahore, 1, 40–69.
Barnham, M. (1977). Is Chilomastix harmless? The Lancet, 2, 1077–1078.Bernard, C., Simpson, A. G. B., & Patterson, D. J. (1997). An ultrastructural study of a free-living
retortamonad Chilomastix cuspidata (Larsen & Patterson, 1990) n. comb. (Retortamonadida,Protista). European Journal of Protistology, 33, 254–265.
Bernard, C., Simpson, A. G. B., & Patterson, D. J. (2000). Some free-living flagellates (Protista)from anoxic habitats. Ophelia, 52, 113–142.
Bělař, K. (1921). Protozoenstüdien III. Archiv für Protistenkunde, 43, 431–462.Bishop, A. (1931). A description of Embadomonas n. spp. from Blatta orientalis, Rana temporaria,
Bufo vulgaris, Salamandra maculosa; with a note upon the “cyst” of Trichomonasbatrachorum. Parasitology, 23, 286–300.
Bishop, A. (1934). Observations upon Embadomonas intestinalis in culture. Parasitology, 26,17–25.
Bishop, A. (1935). Observations upon Chilomastix from Bufo vulgaris, with notes on Chilomastixaulastomi. Parasitology, 27, 507–518.
Boeck, W. C. (1921). Chilomastix mesnili and a method for its culture. Journal of ExperimentalMedicine, 33, 147–175.
Boeck, W. C., & Drbohlav, J. (1925). The cultivation of Entamoeba histolytica. American Journalof Hygiene, 5, 371–407.
28 J. Kulda et al.

Boeck, W. C., & Tanabe, M. (1926). Chilomastix gallinarum,morphology, division and cultivation.American Journal of Hygiene, 6, 319–336.
Brug, S. L. (1922). Quelques observations sur les protozoaires parasites intestinaux de l’homme etdes animaux. Bulletin de la Societe de Pathologie Exotique, 15, 132–139.
Brugerolle, G. (1973). Ètude ultrastructurale du trophozoite et du kyste chez le genre ChilomastixAlexeieff, 1910 (Zoomastigophorea, Retortamonadida Grassé, 1952). Journal of Protozoology,20, 574–585.
Brugerolle, G. (1974). Contribution à l’étude cytologique et phylétique des Diplozoaires (Zoomasti-gophorea, Diplozoa, Dangeard 1910). III. Ètude ultrastructurale du genre Hexamita (Dujardin1838). Protistologica, 10, 83–90.
Brugerolle, G. (1977). Ultrastructure du genre Retortamonas Grassi 1879 (Zoomastigophorea,Retortamonadida, Wenrich 1932). Protistologica, 13, 233–240.
Brugerolle, G. (1991). Flagellar and cytoskeletal systems in amitochondrial flagellates:Archamoeba, Metamonada and Parabasala. Protoplasma, 164, 70–90.
Brugerolle, G. (2006). The symbiotic fauna of the African termite Hodotermes mossambicusidentification of four flagellate species of the genera Spironympha, Trichomonoides andRetortamonas. Parasitology Research, 98, 257–263.
Brugerolle, G., & Lee, J. J. (2000). Order Retortamonadida, Grassé. In J. J. Lee, G. F. Leedale, &P. Bradbury (Eds.), The illustrated guide to the protozoa (Vol. 2, 2nd ed., pp. 1250–1253).Lawrence: The Society of Protozoologists/Allen Press.
Brugerolle, G., & Mignot, J. P. (1990). Phylum Zoomastigina, Class Retortamonadida. InL. Margulis, J. O. Corliss, M. Melkonian, & D. J. Chapman (Eds.), Handbook of Protoctista(1st ed., pp. 259–265). Boston: Jones and Bartlett.
Brugerolle, G., & Regnault, J. P. (2001). Ultrastructure of the enteromonad flagellate Caviomonasmobilis. Parasitology Research, 87, 662–665.
Cavalier-Smith, T. (1983). A 6-kinkdom classification and a united phylogeny. In H. E. A. Schen-g& W. S. Schwemmler (Eds.), Endocytobiology II. Intracellular Space as Oligogenetic Ecosys-tem. Berlin: Walter de Gruiter.
Cavalier-Smith, T. (1987). Eukaryotes with no mitochondria. Nature, 326, 332–333.Cavalier-Smith, T. (1998). A revised six-kingdom system of life. Biological Reviews of the
Cambridge Philosophical Society, 73, 203–266.Cavalier-Smith, T. (2003). The excavate protozoan phyla Metamonada Grassé emend.
(Anaeromonadea, Parabasalia, Carpediemonas, Eopharyngia) and Loukozoa emend. (Jakobea,Malavimonas): their evolutionary affinities and new higher taxa. International Journal ofSystematic and Evolutionary Microbiology, 53, 1741–1758.
Cavalier-Smith, T. (2013). Early evolution of eukaryote feeding modes, cell structural diversity, andclassification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa. European Journal ofProtistology, 49, 115–178.
Cepicka, I., Kostka, M., Uzlíková, M., Kulda, J., & Flegr, J. (2008). Non-monophyly ofRetortamonadida and high genetic diversity of the genus Chilomastix suggested by analysisof SSU rDNA. Molecular Phylogenetics and Evolution, 48, 770–775.
Collier, J., & Boeck, W. C. (1926). The morphology and cultivation of Embadomonas cuniculin. sp. Journal of Parasitology, 12, 131–140.
Červa, L., & Větrovská, G. (1958). Towards the question of pathogenic character of the flagellateChilomastix mesnili. Czechoslovac Epidemiology Microbiology and Immunology, 7, 125–185.
Davis, D. E., Schwartz, L. D., & Jordan, H. E. (1964). A case report: Chilomastix sp. infection inpen-raised quail. Avian Diseases, 8, 465–470.
Diamond, L. S. (1982). A new liquid medium for xenic cultivation of Entamoeba histolytica andother lumen dwelling protozoa. Journal of Parasitology, 68, 958–959.
Dobell, C. (1935). Researches on the intestinal protozoa of monkeys and man VII. On theEnteromonas of macaques and Embadomonas intestinalis. Parasitology, 27, 564–592.
Dobell, C., & Laidlaw, P. P. (1926). On the cultivation of Entamoeba histolytica and some otherentozoic amoebae. Parasitology, 18, 283–318.
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 29

Doflein, F., & Reichenow, E. (1952). Lehrbuch der Protozoenkunde, Spezielle Naturgeschichte derProtozoen (Vol. 2). Jena: Fischer Verlag.
Ekebom, J., Patterson, D. J., & Vors, N. (1996). Heterotrophic flagellates from coral reef sediments(Great Barrier Reef, Australia). Archiv für Protistenkunde, 146, 251–272.
Felsenfeld, O., & Young, V. M. (1946). The correlation of intestinal protozoa and enteric microor-ganisms of known and doubtful pathogenicity. The American Journal of Digestive Diseases, 13,233–234.
Fenchel, T., Bernard, C., Esteban, G., Finlay, B. J., Hansen, P. J., & Iversen, N. (1995). Microbialdiversity and activity in a Danish fjord with anoxic deep water. Ophelia, 43, 45–100.
Geiman, W. M. (1935). Cytological studies of the Chilomastix (Protozoa, Flagellata) of man andother mammals. Journal of Morphology, 57, 429–459.
Grassé, P. P. (1952). Ordre des Retortamonadines nov. In Traité de Zoologie. Phylogénie. Pro-tozoaires: Généralités. Flagellés (Vol. 1, pp. 824–835). Paris: Masson and Cie.
Grassi, B. (1879). Dei protozoi parassiti e specialmente di quelli che sono nell’uomo. GazzettaMedica Italiana Lombardia, 39, 445–448.
Hampl, V., & Simpson, A. G. B. (2008). Possible mitochondria-related organelles in poorly studied“amitochondriate” eukaryotes. In J. Tachezy (Ed.), Hydrogenosomes and Mitosomes: theMitochondria of Anaerobic Eukaryotes (pp. 265–282). Berlin/Heidelberg: Springer.
Hegner, R. W., & Schumaker, E. (1928). Some intestinal amoebae and flagellates from thechimpanzee, three-toed sheep, and guinea-pig. Journal of Parasitology, 15, 31–37.
Hogue, M. J. (1921).Waskia intestinalis: its cultivation and cyst formation. Journal of the AmericanMedical Association, 77, 112–113.
Jacobsen, K. H., Ribeiro, P. S., Quist, B. K., & Rydbeck, B. V. (2007). Prevalence of intestinalparasites in young Quichua children in the highlands of rural Ecuador. Journal of Health,Population and Nutrition, 25, 399–405.
Kessel, J. F. (1924). The experimental transfer of certain intestinal protozoa from man to monkeys.Proceedings of the Society for Experimental Biology and Medicine, 22, 206–208.
Kessel, J. F. (1928). Intestinal protozoa of the domestic pig. American Journal of TropicalMedicine, 8, 481–497.
Kirby, H., & Honigberg, B. M. (1950). Intestinal flagellates from a wallaroo, Macropus robustusGould. University of California Publications in Zoology, 55, 35–66.
Kofoid, C. A., & Swezy, O. (1920). On the morphology and mitosis of Chilomastix mesnili(Wenyon), a common flagellate of the human intestine. University of California Publicationsin Zoology, 20, 117–144.
Kolisko, M., Silberman, J. D., Cepicka, I., Yubuki, N., Takishita, K., Yabuki, A., Leander, B. S.,Inouye, I., Inagaki, Y., Roger, A. J., & Simpson, A. G. B. (2010). Awide diversity of previouslyundetected free-living relatives of diplomonads isolated from marine saline habitats. Environ-mental Microbiology, 12, 2700–2710.
Kulda, J., & Nohýnková, E. (1978). Flagellates of the human intestine and of intestines of otherspecies. In J. P. Kreier (Ed.), Parasitic Protozoa (Vol. 2, pp. 1–138). New York: Academic Press.
Kulda, J., Nohýnková, E.: Dientamoeba fragilis and other intestinal flagellates. In: Topley &Wilson’s microbiology and microbial infections 10th ed. Parasitology (Cox, F.E.G., Wakelin,D., Gillespie, S. H., Despommier, D.D., eds.), pp. 266–274. London: Hodder Arnold (2006).
Larsen, J., & Patterson, D. J. (1990). Some flagellates (Protista) from tropical marine sediments.Journal of Natural History, 24, 801–937.
Lavier, G. (1936). Sur quelques flagellés intestinaux de poissons marins. Annales de ParasitologieHumaine et Comparee, 14, 278–289.
Lee, W. J., & Patterson, D. J. (2000). Heterotrophic flagellates (Protista) from marine sediments ofBotany Bay, Australia. Journal of Natural History, 34, 483–662.
Levine, N. D. (1973). Protozoan parasites of domestic animals and of man (2nd ed.). Minneapolis:Burgess.
30 J. Kulda et al.

Lindmark, D. G., & Müller, M. (1973). Hydrogenosome, a cytoplasmic organelle of the anaerobicflagellate Tritrichomonas foetus, and its role in pyruvate metabolism. Journal of BiologicalChemistry, 248, 7724–7728.
Mackinnon, D. L. (1915). Studies on parasitic protozoa III. (a) Notes on the flagellatesEmbadomonas. (b) The multiplication cysts of a trichomastigine. Quarterly Journal of Micro-scopical Science, 61, 105–118.
Martínez-Díaz, R., Castro, T. A., Herrera, S., & Ponce, F. (2001). First report of the genusRetortamonas (Sarcomastigophora: Retortamonadidae) in birds. Memórias do InstitutoOswaldo Cruz, 96, 96–963.
McDowell, S. (1953). A morphological and taxonomy study of the caecal Protozoa of the commonfowl, Gallus gallus L. Journal of Morphology, 92, 337–400.
Moskowitz, N. (1951). Observations on some intestinal flagellates from reptilian host (Squamata).Journal of Morphology, 89, 257–321.
Nasiri, V., Esmailnia, K., Karim, G., Nasir, M., & Akhavan, O. (2009). Intestinal parasitic infectionsamong inhabitants of Karaj City, Tehran province, Iran 2006-2008. Korean Journal of Parasi-tology, 47, 265–268.
Navarathnam, E. S. (1970). Intestinal flagellates of the common Indian rat Rattus rattus frugivo-rous. Acta Protozoologica, 8, 155–165.
Nie, D. (1948). The structure and division of Chilomastix intestinalis Kuczynski, with notes onsimilar forms in man and other vertebrates. Journal of Morphology, 82, 287–318.
Nie, D. (1950). Morphology and taxonomy of the intestinal protozoa of the guinea-pig, Caviaporcella. Journal of Morphology, 86, 381–493.
Nohýnková, E., Tůmová, P., & Kulda, J. (2006). Cell division of Giardia intestinalis: flagellardevelopmental cycle involves transformation and exchange of flagella between mastigonts of adiplomonad cell. Eukaryotic Cell, 5, 753–761.
Oyofo, B. A., Peruski, I. F., Ismail, T. F., el-Etr, S. H., Churilla, A. M., Wasfy, M. O., Petruccelli,B. F., & Gabriel, M. E. (1997). Enteropathogens associated with diarrhea among militarypersonel during Operation Bright Star 96, in Alexandria, Egypt. Military Medicine, 162,396–400.
Park, J. S., Kolisko, M., & Simpson, A. G. B. (2010). Cell morphology and formal description ofErgobibamus cyprinoides n.g., n.sp., another Carpediemonas-like relative of diplomonads.Journal of Eukaryotic Microbiology, 57, 520–528.
Park, J. S., Kolisko, M., Heiss, A. A., & Simpson, A. G. B. (2009). Light microscopic observations,ultrastructure, and molecular phylogeny of Hicanonectes teleskopos n.g., n.sp., a deep-branching relative of diplomonads. Journal of Eukaryotic Microbiology, 56, 373–384.
Ruinen, J. (1938). Notizen über Salzflagellaten II. Über die Verbreitung der Salzflagellaten. Archivfür Protistenkunde, 90, 210–258.
Russel Gabel, J. (1954). The morphology and taxonomy of the intestinal protozoa of the Americanwoodchuck Marmota monax Linnaeus. Journal of Morphology, 94, 473–449.
Silberman, J. D., Simpson, A. G. B., Kulda, J., Cepicka, I., Hampl, V., Johnson, P. J., & Roger, A. J.(2002). Retortamonad flagellates are closely related to diplomonads – Implications for themitochondrial function in eukaryote evolution.Molecular Biology and Evolution, 19, 777–786.
Simpson, A. G. B. (2003). Cytoskeletal organization, phylogenetic affinities and systematics in thecontentious taxon Excavata (Eukaryota). International Journal of Systematic and EvolutionaryMicrobiology, 53, 1759–1777.
Simpson, A. G. B., & Patterson, D. J. (1999). The ultrastructure od Carpediemonas membranifera(Eukaryota) with reference to the “Excavate hypothesis”. European Journal of Protistology, 35,353–370.
Simpson, A. G. B., & Roger, A. J. (2004). Excavata and origin of amitochondriate eukaryotes. InR. P. Hirt & D. S. Horner (Eds.), Organelles, genomes, and eukaryote phylogeny: an evolu-tionary synthesis in the age of genomics (pp. 27–53). Boca Raton: CRC Press.
Simpson, A. G. B., Roger, A. J., Silberman, J. D., Leipe, D. D., Edgcomb, V. P., Jermiin, L. S.,Patterson, D. J., & Sogin, M. L. (2002). Evolutionary history of early “early-diverging”
Retortamonadida (with Notes on Carpediemonas-Like Organisms and. . . 31

eukaryotes: the excavate taxon Carpediemonas is a close relative od Giardia. MolecularBiology and Evolution, 19, 1782–1791.
Tachezy, J., & Šmíd, O. (2007). Mitosomes in parasitic protists. In J. Tachezy (Ed.),Hydrogenosomes and mitosomes: mitochondria of anaerobic eukaryotes. Berlin/Heidelberg:Springer-Verlag.
Takishita, K., Kolísko, M., Komatzuzaki, H., Yabuki, A., Inagaki, N., Cepicka, I., Smejkalová, P.,Silberman, J. D., Hashimoto, T., Roger, A. J., & Simpson, A. G. B. (2012). Multigenephylogenies of diverse Carpediemonas-like organisms identify the closest relatives of‘amitochondriate‘ diplomonads and retortamonads. Protist, 163, 344–355.
Tovar, J., Fischer, A., & Clark, C. G. (1999). The mitosome, a novel organelle related to mitochon-dria in the amitochondrial parasite Entamoeba. Molecular Microbiology, 32, 1013–1021.
Tovar, J., León-Avila, G., Sanchéz, L. B., Sutak, R., Tachezy, J., van der Giezen, M., Hernández,M., Müller, M., & Lucocq, J. M. (2003). Mitochondrial remnant organelles of Giardia functionin iron-sulphur protein maturation. Nature, 426, 172–176.
Waikagul, J., Krudsood, S., Radomyos, P., Radomyos, B., Chalemrut, K., Jonsuksuntigul, P.,Kojima, S., Looareesuwan, S., & Thaineau, W. (2002). A cross-sectional study of intestinalparasitic infections among schoolchildren in Nan Province, Northern Thailand. Southeast AsianJournal of Tropical Medicine and Public Health, 33, 218–223.
Weerakon, N. D., Harper, J. D. I., Simpson, A. G. B., & Patterson, D. J. (1999). Centrin in thegroove: immunolocalization of centrin and microtubules in the putatively primitive protistChilomastix cuspidata (Retortamonadida). Protoplasma, 210, 75–84.
Wenrich, D. H. (1932). The relation of the protozoan flagellate Retortamonas gryllotalpae (Grassi,1879) Stiles, 1902, to the species of the genus EmbadomonasMackinnon, 1911. Transactions ofthe American Microscopical Society, 51, 225–237.
Wenrich, D. H. (1947). Culture experiments on intestinal flagellates III. Species from amphibiansand reptiles. Parasitology, 33, 62–70.
Westphal, A. (1939). Beziehungen zwischen Infektionsstärke and “Krankheitsbild” bei Infektionenmit Chilomastix mesnili und anderen Dickdarm-flagellaten. Zeitschrift für Hygiene undInfektionskrankheiten, 122, 146–158.
Yubuki, N., Inagaki, Y., Nakayama, T., & Inouye, I. (2007). Ultrastructure and ribosomal RNAphylogeny of the free-living heterotrophic flagellate Dysnectes brevis n. gen., n. sp., a newmember of the Fornicata. Journal of Eukaryotic Microbiology, 54, 191–200.
Yubuki, N., Simpson, A. G. B., & Leander, B. S. (2013). Comprehensive ultrastructure of Kipferliabialata provides evidence for character evolution within the Fornicata (Excavata). Protist, 164,423–439.
Yubuki, N., Huang, S. S. C., & Leander, B. S. (2016). Comparative ultrastructure of fornicateexcavates, including a novel free-living relative of diplomonads: Aduncisulcus paluster gen. etsp. nov. Protist, 167, 584–596.
Yubuki, N., Zadrobílková, E., Čepička, I. (2016). Ultrastructure and molecular phylogeny ofIotanema spirale gen. nov. et sp. nov., a new lineage of endobiotic Fornicata with strikinglysimplified ultrastructure. Journal of Eukaryotic Microbiology. doi:10.1111/jeu.12376.
32 J. Kulda et al.


















