
Retinal determination genes as targets and possible ... · Retinal determination genes as targets...
10
Retinal determination genes as targets and possible effectors of extracellular signals Lucy C. Firth 1 , Nicholas E. Baker ⁎ Department of Genetics, Albert Einstein College of Medicine,1300 Morris Park Avenue, Bronx, NY 10461, USA abstract article info Article history: Received for publication 21 October 2008 Revised 15 December 2008 Accepted 16 December 2008 Available online 25 December 2008 Keywords: Retinal determination Drosophila eye Master regulatory gene Positional information Retinal determination genes are sufficient to specify eyes in ectopic locations, raising the question of how these master regulatory genes define an eye developmental field. Genetic mosaic studies establish that expression of the retinal determination genes eyeless, teashirt, homothorax, eyes absent, sine oculis, and dachshund are each regulated by combinations of Dpp, Hh, N, Wg, and Ras signals in Drosophila. Dpp and Hh control eyeless, teashirt, sine oculis, and dachshund expression, Dpp and Ras control homothorax, and all the signaling pathways affect eyes absent expression. These results suggest that eye-specific development uses retinal determination gene expression to relay positional information to eye target genes, because the distinct, overlapping patterns of retinal determination gene expression reflect the activities of the extracellular signaling pathways. © 2008 Elsevier Inc. All rights reserved. Introduction Pax6/Eyeless was described as a ‘Master Regulatory gene’ when it was found that its ectopic expression could direct development of an ectopic eye (Halder et al., 1995). A small set of other genes have also been found capable of directing ectopic eye development, including another Pax6 homolog twin of eyeless (toy), sine oculis (so), eyes absent (eya) and dacshund (dac) (reviewed in Pappu and Mardon, 2002; Silver and Rebay, 2005). These retinal determination (RD) genes have roles in mammalian eye development, suggesting conserved master-regulatory gene mechanisms (reviewed in Nils- son, 2004; Treisman, 2004). It has been suggested that RD gene products define the eye field by binding to many genes that are expressed in the eye, and interacting with the various transcription factors that are direct effectors of positional signals throughout the body, so that the RD protein/positional information combination defines eye-specific programs of gene expression in response to extracellular signals (Curtiss et al., 2002; Mann and Carroll, 2002). This combinatorial mechanism might be illustrated by an eye-specific enhancer of the hh gene, which is activated by the Ras pathway through a Ras- dependent transcription factor Pnt, in combination with the RD protein So that provides eye specificity (Rogers et al., 2005). The model that RD gene products provide for eye-specific interpretation of positional information could explain why extracellular signals have such an influence on the locations where RD gene expression can form ectopic eyes, something that is harder to understand if RD genes simply activate eye programs irrespective of the cell's position (Chen et al., 1999; Pappu et al., 2003; Bessa and Casares, 2005; Weasner et al., 2006). The notion that RD genes define the eye response to positional signals is complicated by the fact that RD gene expression is itself dynamic in space and time (Bessa et al., 2002; Pappu and Mardon, 2002). Dynamic RD gene expression is partly attributable to regulatory interactions between the RD genes themselves, but these are not sufficient to explain all the spatial and temporal aspects, which also depend on Dpp signaling to cue changes in RD gene expression, as summarized below (Bessa et al., 2002). One explanation could be that there is a distinction between extracellular signaling pathways. As there are several examples of RD gene regulation by Dpp signaling, it could be that Dpp plays a unique role in coordinating RD gene expression. By contrast, Wnt, Hh, Notch, and Ras pathways might provide the positional information that induces specific eye target genes and patterns eye-specific fates within the eye field, according to the status of RD gene expression at the time. To investigate the control of RD gene expression in more detail, we made a systematic study of the contribution of extracellular signaling pathways during the third larval instar, when patterning, specifica- tion, and differentiation of individual retinal cells occurs. We used clonal analysis of a battery of mutations affecting signal reception and transduction to explain the spatial and temporal dynamics of RD gene expression in terms of specific roles for Hh, Wg, Notch and Ras signals, in addition to Dpp. We found that all the extracellular signals regulate RD gene expression. The results suggest that Dpp does not play a unique role, and lead to a new discussion of the relationship between RD genes as master regulators and the role of extracellular signals and positional information. Developmental Biology 327 (2009) 366–375 ⁎ Corresponding author. E-mail addresses: lucy.fi[email protected] (L.C. Firth), [email protected] (N.E. Baker). 1 Present address: Syngenta, Jealott's Hill International Research Centre Bracknell, Berkshire, RG42 6EY, UK. 0012-1606/$ – see front matter © 2008 Elsevier Inc. All rights reserved. doi:10.1016/j.ydbio.2008.12.021 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology
-
Upload
nguyendieu -
Category
Documents
-
view
215 -
download
0
Transcript of Retinal determination genes as targets and possible ... · Retinal determination genes as targets...

Developmental Biology 327 (2009) 366–375
Contents lists available at ScienceDirect
Developmental Biology
j ourna l homepage: www.e lsev ie r.com/deve lopmenta lb io logy
Retinal determination genes as targets and possible effectors of extracellular signals
Lucy C. Firth 1, Nicholas E. Baker ⁎Department of Genetics, Albert Einstein College of Medicine, 1300 Morris Park Avenue, Bronx, NY 10461, USA
⁎ Corresponding author.E-mail addresses: [email protected] (L.C. Firth
(N.E. Baker).1 Present address: Syngenta, Jealott's Hill Internation
Berkshire, RG42 6EY, UK.
0012-1606/$ – see front matter © 2008 Elsevier Inc. Aldoi:10.1016/j.ydbio.2008.12.021
a b s t r a c t
a r t i c l e i n f oArticle history:
Retinal determination gene Received for publication 21 October 2008Revised 15 December 2008Accepted 16 December 2008Available online 25 December 2008Keywords:Retinal determinationDrosophila eyeMaster regulatory genePositional information
s are sufficient to specify eyes in ectopic locations, raising the question of howthese master regulatory genes define an eye developmental field. Genetic mosaic studies establish thatexpression of the retinal determination genes eyeless, teashirt, homothorax, eyes absent, sine oculis, anddachshund are each regulated by combinations of Dpp, Hh, N, Wg, and Ras signals in Drosophila. Dpp andHh control eyeless, teashirt, sine oculis, and dachshund expression, Dpp and Ras control homothorax, and allthe signaling pathways affect eyes absent expression. These results suggest that eye-specific developmentuses retinal determination gene expression to relay positional information to eye target genes, because thedistinct, overlapping patterns of retinal determination gene expression reflect the activities of theextracellular signaling pathways.
© 2008 Elsevier Inc. All rights reserved.
Introduction
Pax6/Eyeless was described as a ‘Master Regulatory gene’ whenit was found that its ectopic expression could direct development ofan ectopic eye (Halder et al., 1995). A small set of other genes havealso been found capable of directing ectopic eye development,including another Pax6 homolog twin of eyeless (toy), sine oculis(so), eyes absent (eya) and dacshund (dac) (reviewed in Pappu andMardon, 2002; Silver and Rebay, 2005). These retinal determination(RD) genes have roles in mammalian eye development, suggestingconserved master-regulatory gene mechanisms (reviewed in Nils-son, 2004; Treisman, 2004).
It has been suggested that RD gene products define the eye fieldby binding to many genes that are expressed in the eye, andinteracting with the various transcription factors that are directeffectors of positional signals throughout the body, so that the RDprotein/positional information combination defines eye-specificprograms of gene expression in response to extracellular signals(Curtiss et al., 2002; Mann and Carroll, 2002). This combinatorialmechanism might be illustrated by an eye-specific enhancer of thehh gene, which is activated by the Ras pathway through a Ras-dependent transcription factor Pnt, in combination with the RDprotein So that provides eye specificity (Rogers et al., 2005). Themodel that RD gene products provide for eye-specific interpretationof positional information could explain why extracellular signalshave such an influence on the locations where RD gene expression
al Research Centre Bracknell,
l rights reserved.
can form ectopic eyes, something that is harder to understand if RDgenes simply activate eye programs irrespective of the cell's position(Chen et al., 1999; Pappu et al., 2003; Bessa and Casares, 2005;Weasner et al., 2006).
The notion that RD genes define the eye response to positionalsignals is complicated by the fact that RD gene expression is itselfdynamic in space and time (Bessa et al., 2002; Pappu and Mardon,2002). Dynamic RD gene expression is partly attributable to regulatoryinteractions between the RD genes themselves, but these are notsufficient to explain all the spatial and temporal aspects, which alsodepend on Dpp signaling to cue changes in RD gene expression, assummarized below (Bessa et al., 2002).
One explanation could be that there is a distinction betweenextracellular signaling pathways. As there are several examples of RDgene regulation by Dpp signaling, it could be that Dpp plays a uniquerole in coordinating RD gene expression. By contrast, Wnt, Hh, Notch,and Ras pathways might provide the positional information thatinduces specific eye target genes and patterns eye-specific fateswithin the eye field, according to the status of RD gene expression atthe time.
To investigate the control of RD gene expression in more detail, wemade a systematic study of the contribution of extracellular signalingpathways during the third larval instar, when patterning, specifica-tion, and differentiation of individual retinal cells occurs. We usedclonal analysis of a battery of mutations affecting signal reception andtransduction to explain the spatial and temporal dynamics of RD geneexpression in terms of specific roles for Hh, Wg, Notch and Ras signals,in addition to Dpp. We found that all the extracellular signals regulateRD gene expression. The results suggest that Dpp does not play aunique role, and lead to a new discussion of the relationship betweenRD genes as master regulators and the role of extracellular signals andpositional information.

367L.C. Firth, N.E. Baker / Developmental Biology 327 (2009) 366–375
Specification and differentiation of individual retinal cells occurwithin a domain of the eye imaginal disc that co-expresses So, Eyaand Dac. This co-expression domain spreads anteriorly across the eyeimaginal disc, progressively replacing expression of three othergenes, Ey, Tsh and Hth (Fig. 1A–E) (Bessa et al., 2002). Ey, Tsh and Hthare all DNA-binding transcription factors that interact directly, andpromote each other's expression (Bessa et al., 2002). The Ey/Tsh/Hthcombination represses expression of Eya and Dac (Bessa et al., 2002).Eya and Dac interact with the DNA-binding protein So, and there isevidence that Dac can also bind DNA (Bonini et al., 1997; Chen et al.,1997; Pignoni et al., 1997; Kim et al., 2002). The combination of So/Eya/Dac promotes expression of each of their own genes, andrepresses expression of Ey/Tsh/Hth genes (Bessa et al., 2002). Cellsin the eye disc may be poised to switch between these two geneexpression combinations by the cell-autonomous antagonismbetween them. However, Hth shuts off earlier than Ey and Tsh do,and So and Eya are expressed earlier in most eye cells than Dac is,suggesting that the notion of a single switch between only twoexpression states is oversimplified.
Fig.1. (A) A cartoon of the third instar eye imaginal disc relating proliferation, G1 arrest, themthe left in this and other figures. Hth is expressed most anteriorly and overlaps with more pobegin expression 5–8 cell diameters anterior to where cell cycle arrest occurs in G1. 6–10 cellpersist during differentiation. Yellow bar shows the initial stripe of atonal expression in respoof Hth (green), Ey (red) and Eya (blue). Arrow indicates the position of the morphogenetic furof ectopic Dpp signaling (actNtkv⁎, Lac Z) on (F) Hth, (G) Ey, (H) Tsh, (I) Eya, and (J) Dac levels.in panel (J) indicate anterior clones where Dac is induced poorly.
The dynamic gene expression patterns correlate with a morpho-genetic furrow that moves anteriorly across the eye disc and marksthe progression of retinal differentiation (Figs. 1A–E) (Bonini et al.,1993; Pichaud and Casares, 2000; Bessa et al., 2002; Kenyon et al.,2003). The morphogenetic furrow moves across the eye disc inresponse to Hh and Dpp signaling, antagonized by Wg signaling, andwith a contribution from N (Ma et al., 1993; Ma and Moses, 1995;Treisman and Rubin, 1995; Greenwood and Struhl, 1999; Curtiss andMlodzik, 2000; Baonza and Freeman, 2001; Fu and Baker, 2003). Hh,Dpp, and N ligands are all expressed in various cells in or near theposterior, differentiating parts of the retina as it expands; Wg isexpressed ahead of the furrow by undifferentiated cells at the lateraland anterior margins. Dpp signaling is already known to contributeto activation of the so, eya, and dac genes, and to acceleraterepression of the hth and ey genes (Curtiss and Mlodzik, 2000; Leeand Treisman, 2001; Bessa et al., 2002). These changes areunaffected or only delayed in cells deficient for Dpp signaltransduction, suggesting that other signals also contribute toregulate RD genes.
orphogenetic furrow (MF) and differentiation to gene expression domains. Anterior is tosterior expression of Ey and Tsh. Ey and Tsh overlap in turn with Eya, So and Dac, whichdiameters posterior to cell cycle arrest, Ey and Tsh disappear, leaving Eya, So and Dac tonse to Dpp and Hh signaling. (B) Wild type eye disc epithelium showing the expressionrow. Individual channels are shown in panels (C) Hth, (D) Ey and (E) Eya. (F–J) The effectFlip-out clones expressing activated Tkv⁎ and β-Galactosidase are labeled green. Arrows

368 L.C. Firth, N.E. Baker / Developmental Biology 327 (2009) 366–375
Materials and methods
Loss of function clones
Clones of cells mutant for genes were obtained by the FLP-mediated mitotic recombination technique with a heat shockinducible FLPase (Golic, 1991; Xu and Rubin, 1993). Homozygousmutant cells were identified through the absence of transgene-encoded arm-β-gal (Vincent et al., 1994). Apart from dac4, clones ofthe genotypes below were generated in a Minute heterozygousbackground (Morata and Ripoll, 1975). Minute heterozygous larvaewere heat shocked at 48–96 h AEL for 1 h at 37 °C.
The following alleles were used:Mad12 FRT40 (Sekelsky et al., 1995); smo3 FRT40 and smoD16 FRT40
(Chen and Struhl, 1998); Su(H) Δ47 FRT40[w+ l(2)35Bg+] (Morel andSchweisguth, 2000); dac4 FRT40 (Chen et al., 1997); FRT42 arr2 (Wehrliet al., 2000); FRT42 mago3 (Boswell et al., 1991); FRT42 shn3(Chase andBaker, 1995); FRT42 shn1 (Arora et al., 1995). Double and triple mutantclones were obtained using the following chromosomes:
smo3 Su(H) Δ47 FRT40 [w+ l(2)35Bg+];smo3 Mad12 FRT40 [w+ l(2)35Bg+];Mad12 Su(H) Δ47 FRT40 [w+ l(2)35Bg+];FRT42 mago3 shn3;FRT42 arr2 shn1;smo3 Mad12 Su(H) Δ47 FRT40 [w+ l(2)35Bg+];FRT42 arr2 shn1.Marked, Minute chromosomes used included:M21 [armLacZ] FRT40;FRT42 [armLacZ] M(2)56F;FRT82 M(3)96C [armLacZ].
Transgenes
UAS-ArmS10 (Pai et al., 1997); UAS-Dac (Chen et al., 1997); UAS-Dac(Shen and Mardon, 1997); UAS-SoC.2 (Pignoni et al., 1997); UAS-RasV12
(Karim and Rubin, 1998); yw; p[actNCD2NGal4]; UAS-LacZ (Pignoniand Zipursky, 1997); UAS-GFPnls.
MARCM analysis
Dac and/or So were overexpressed in smo Mad cells using theMARCM technique (Lee and Luo, 2001). y w hsFLP, UAS-GFP/+; smo3
Mad12 FRT40/TubGal80 FRT40; TubGal4/UAS-Dac, y w hsFLP, UAS-GFP/+;smo3 Mad12 FRT40/TubGal80 FRT40; TubGal4/UAS-SoC2 and y w hsFLP,UAS-GFP/+; smo3 Mad12 FRT40/TubGal80 FRT40; TubGal4/UAS-Dac,UAS-SoC2 were dissected. Wingless signaling was activated inMad and smoMad Su(H) clones in y w hsFLP, UAS-GFP/UAS-arm⁎; Mad10 FRT40/TubGal80 FRT40; TubGal4/+ and y w hsFLP, UAS-GFP/UAS-arm⁎; smo3
Mad12 Su(H)Δ47 FRT40 [w+ l(2)35Bg+]/TubGal80 FRT40; TubGal4/+.Larvae were heat shocked 24–72 h AEL. Mutant cells were positivelymarked and detected with an antibody against GFP or Dac ectopicexpression.
Immunohistochemistry
Labeling of eye discs was performed as described (Firth et al.,2006). Preparations were examined on the BioRad Radiance2000confocal microscope. Images were processed using Adobe Photoshop6.0 and NIH Image J software. The following antibodies were used:rabbit anti-βGalactosidase (Cappel); mouse anti-βGgalactosidase(mAb40-1a); mouse anti-Dac (2–3) (Mardon et al., 1994); mouseanti-Elav (mAb9F8A9) (O'Neill et al., 1994); rabbit anti-Ey (Halder etal., 1998); mouse anti-Eya (10H6) (Bonini et al., 1993); guinea pig anti-Hth (Casares and Mann, 1998); guinea pig anti-So (Mutsuddi et al.,2005); rabbit anti-Tsh (Wu and Cohen, 2000); mouse anti-Wg (4D4)(Brook and Cohen, 1996); rabbit and mouse anti-GFP (Invitrogen).
Results
In this study we examined the cell-autonomous roles ofextracellular molecules by generating clones of cells, located withinthe developing retinal field, that are mutant for downstreamsignaling components (Wassarman et al., 1995; Bray, 2006; Clevers,2006; Affolter and Basler, 2007; Variosalo and Taipale, 2008). Thesestudies aim to identify the extracellular signals to which retinal cellsrespond directly and cell-autonomously, and to distinguish signalsthat act indirectly and non-autonomously. Our study also focuses onthe cells within the eye field that themselves differentiate to retinalfates; the initiation of the morphogenetic furrow at the posterioreye margin is genetically distinct (Lee and Treisman, 2002; Kenyonet al., 2003).
BMP signaling does not explain all transcriptional transitions
Dpp signaling regulates all the RD genes, and has been proposedas the chief source of their spatial patterning (Bessa et al., 2002). Ifthis was so, we would expect that ectopic Dpp signaling would besufficient to repress Ey, Tsh, and Hth, and to activate expression ofEya, So, and Dac. Ectopic expression of an activated Dpp receptor(actNtkv⁎) was used to test this hypothesis. Although ectopic Dppsignaling could repress or activate the expression of some of thesegenes, it was not sufficient for all. Specifically, Tkv⁎ repressed Hth,but Ey was not repressed and repression of Tsh was variable (Figs.1F–H) (Bessa et al., 2002). In addition, whereas Tkv⁎ activated Eyaexpression (Fig. 1I), Dac was induced only in cells just ahead ofthe MF (Fig. 1J), corresponding to a modest acceleration of thenormal program. Therefore, Dpp signaling was not sufficient toaccount for spatial patterning of retinal determination geneexpression, and other spatial information within the eye discmust be important also.
Hh contributes to Ey and Tsh repression, but does not repress Hth
The most obvious suspect to collaborate with Dpp is Hh. Dppand Hh together are required for the onset of differentiation andcell cycle arrest as the morphogenetic furrow progresses (Green-wood and Struhl, 1999; Curtiss and Mlodzik, 2000; Firth andBaker, 2005). The contribution of Hh was first assessed withregard to the Hth, Ey and Tsh genes that are repressed as the MFapproaches. When the Dpp pathway was eliminated, in clones ofcells mutant for Mothers against Dpp (Mad), repression of all threegenes was delayed with respect to wild type cells, consistent withprevious reports (Figs. 2A, D and G) (Lee and Treisman, 2001;Bessa et al., 2002).
Cells mutant for smoothened (smo) were used to examine the roleof Hh signaling. In smo− clones near the MF, repression of Ey and Tshwas delayed (Figs. 2B and E). Repression of Hth occurred normally insmo− clones, however (Fig. 2H). These data show that Hh signalingcontributes to the normal repression of Ey and Tsh.
The combined roles of Hh and Dpp were examined in clones ofcells simultaneously mutant for both Mad and smo. The smo Mad−
clones completely failed to repress Ey or Tsh, and expression of theseproteins was maintained in all eye disc cells (Figs. 2C and F). Bycontrast, Hth repressionwas only delayed, similar to what was seen inMad− clones (Fig. 2I). These findings indicate that ey and tshexpression are repressed by both Dpp and Hh. Each pathwaycontributes to timely repression, and neither is completely sufficientalone. Cells that cannot respond to Hh or Dpp maintain Ey and Tsh,suggesting that these two pathways together account for the spatialregulation of ey and tsh expression in the eye disc. Certainly, no othersignal is present that is sufficient for their repression. By contrast, Hthmust be regulated by Dpp and some other spatial signal or signals,distinct from Hh.

Fig. 2. Combinatorial repression of Ey, Tsh and Hth by Dpp and other signals. The effects of losing of Dpp and Hh signaling on Ey, Tsh and Hth expressionwere assessed by generatingmitotic clones of Mad12, smoD16, and smo3 Mad12 respectively (panels A–I). Hth expression was also examined in additional genotypes (panels J–N). Homozygous clones are outlinedby absence of the β-Galactosidase in the magenta channel (except for panel N). Ey, Tsh or Hth proteins were labeled in the green channel and are shown as maximum projections ofthe z-axis of the disc epithelium. (A) Ey expression persisted in Mad clones but was eventually repressed (eg yellow arrows). (B) Ey expression persisted in smo clones but waseventually repressed (eg yellow arrows). (C). Ey was never repressed in smo Mad clones (eg blue arrow). (D) Tsh expression persisted in Mad clones but was eventually repressed(eg yellow arrow). (E) Tsh expression persisted in smo clones but was eventually repressed (eg yellow arrows). (F) Tsh was never repressed in smo Mad clones (eg blue arrows).(G) Hth expression persisted in Mad clones but was eventually repressed (eg yellow arrows). In addition, the second phase of Hth expression that occurs in the posterior eye beganearly in Mad clones (eg orange arrows). Note that presence or absence of Hth depends on location in the anterior–posterior axis, not size of the clones. (H). Hth expression wasrepressed normally in smo clones (eg yellow arrow). (I–J) Hth expression persisted in smo Mad clones (I) or smo Mad Su(H) clones (J) but was downregulated after a delay (eg yellowarrows). Hth re-expression was accelerated (eg orange arrows). These phenotypes resemble the effect of Mad alone. Hth repression sometimes seems more complete in Mad clones(see panel G), which are expected to maintain higher Ras activity than smo Mad or smo Mad Su(H) clones. (K) Hth expression persisted in shn clones but was eventually repressed(eg yellow arrow). Hth re-expression was accelerated (eg orange arrow). Note that presence or absence of Hth depends on location in the anterior–posterior axis, not size of theclones. (L) Hth was repressed inmago clones (eg yellow arrow), after a slight delay. (M). Hth was never repressed in shn mago clones (blue arrows). (N) actNRasV12, GFP clones labeledfor GFP in red. Differentiating neural cells are labeled with ElaV in blue. Ras activity repressed Hth in the anterior domain (eg blue arrows), evenwhere there were no differentiatingneural cells. Hth was also reduced by Ras activation and ectopic differentiation in the posterior domain (eg yellow arrows).
369L.C. Firth, N.E. Baker / Developmental Biology 327 (2009) 366–375

370 L.C. Firth, N.E. Baker / Developmental Biology 327 (2009) 366–375
Hth repression is controlled by Dpp and Ras signals
The Notch (N) and Ras/MAPK signaling pathways are activated byligand expression as eye development proceeds. We assessed the roleof Notch by generating clones of cells mutant for the transcriptionfactor that acts downstream of N, Suppressor of Hairless (Su(H)). Hthwas normal in Su(H)– or smo Su(H)– clones (data not shown). AlthoughSu(H) has both positive and negative roles in N signaling (Morel andSchweisguth, 2000; Bray, 2006), Su(H) mutant cells cannot beinfluenced by N signaling so spatiotemporal differences between Su(H)mutant cells must be responses to other pathways. When Dpp andN signaling were both affected, in Mad Su(H)– clones, Hth repressionwas delayed to the same extent as when only Dpp signaling wasremoved (data not shown). We also removed Dpp, N and Hh signalingsimultaneously. Hth repression was similarly delayed in smo Mad Su(H)− clones (Fig. 2J). Thus, neither N nor Hh had any detectable role inrepressing Hth, alone or in the absence of Dpp signaling.
We assessed the role of Ras/MAPK signaling in clones of cellsmutant for a component of the Ras/MAPK signal transductionpathway, mago nashi (mago) (J.Y. Roignant and J. Treisman, personalcommunication). The mago gene is conveniently linked to schnurri(shn), which encodes a Mad co-repressor that is required for mosteffects of Dpp signaling (Affolter and Basler, 2007). Hth repressionwasdelayed in shn− clones as it was in Mad– clones, confirming that thiseffect of Dpp required shn (Fig. 2K). Therewas also a small delay in Hthrepression in mago− or egfr− clones (Fig. 2L and data not shown). Inmago shn− clones, Hth was not repressed, and Hth levels weremaintained, throughout the eye disc (Fig. 2M). The data indicate thatHth is repressed by Dpp and Ras/MAPK signaling.
To test whether Ras/MAPK pathway activity is sufficient to repressHth, we examined clones of cells ectopically expressing an activatedform of Ras (actNRasV12). As predicted, expression of activated RasV12
repressed Hth (Fig. 2N). Ectopic Ras/MAPK activity can induce ectopicphotoreceptor differentiation (Freeman, 1996), possibly leading toectopic Dpp expression, but we found that actNRasV12 clonesrepressed Hth even when they lacked ectopic photoreceptors, andin addition that repression appeared cell-autonomous (Fig. 2N).RasV12 also repressed Hth expression in the peripodial epithelium(data not shown).
These findings indicate that hth expression is repressed by bothDpp and Ras signaling. Each pathway contributes to timely repressionat the proper location, and neither is completely sufficient alone. Cellsthat cannot respond to Dpp or Ras maintain Hth, suggesting that thesetwo pathways together account for the spatial regulation of hthexpression in the eye disc. No other signal is present that is sufficientfor hth repression.
Maintaining Hth repression depends on Dpp
Hth repression is transient, and Hth is expressed again about20 h later in undifferentiated cells with basal nuclei that are fated tocontributed to retinal pigment and sensory bristle cells (Pichaudand Casares, 2000). This second stage of hth expression posterior tothe morphogenetic furrow began prematurely in clones of Mad− orshn− mutant cells unable to respond to Dpp (orange arrows Figs. 2Gand K). This suggests that Dpp signaling represses Hth continuously,and that Hth re-expressed posterior to the furrow is due toincreasing distance from the morphogenetic furrow where Dpp isexpressed in a stripe (Blackman et al., 1991). Roles of Dpp signalinganterior to the morphogenetic furrow are well-known, but to ourknowledge initiation and maintenance of Hth repression providesthe first indication that Dpp might diffuse and acts both anteriorand posterior of the Dpp expression stripe in the morphogeneticfurrow. This is intriguing as phosphorylation of the Mad protein ismore apparent anterior to the furrow than in the differentiatinretina (unpublished observations).
Hth re-expression occurred with normal timing in mago– clones,or egfr– clones, indicating that a drop in MAPK signaling is notsufficient for Hth re-expression (Fig. 2L, and data not shown). Bycontrast, elevated Ras signaling, which recruits ectopic photoreceptorcells, also prevented Hth re-expression posterior to the furrow (Fig.2N). It is possible that the role of Ras signaling in photoreceptordifferentiation normally helps restrict Hth expression to undiffer-entiated cells posterior to the furrow.
Dpp and Hh are required for dac and so expression
The RD genes eya, so and dac are turned on as the MF approaches(Bonini et al., 1993; Cheyette et al., 1994; Mardon et al., 1994; Serikakuand O'Tousa, 1994). Because Dpp signaling was only sufficient to turnon Dac close to the furrow, and Dac expression occurs in cells unableto respond to Dpp, albeit after a delay, we examined the contributionof Hh to Dac regulation. As expected, Dac expression was delayed inMad− clones unable to respond to Dpp (Fig. 3A). The timing of Dacexpression was unaffected in smo− clones that are unable to respondto Hh, but Dac levels appeared lower (Fig. 3B). Dac was cell-autonomously absent (or severely reduced) in smo Mad– clones thatcannot respond to Hh or Dpp (Fig. 3C). Thus, Dpp and Hh actredundantly to establish Dac expression.
The regulation of So expression has not been examined previously.The level of So expression was subtly reduced in most internal Mad−
clones that are unable to respond to Dpp (Fig. 3D). Sometimes this wasaccompanied by a short delay (data not shown). Expression wasunaffected in smo− clones that are unable to respond to Hh (Fig. 3E). Bycontrast, Sowas almost absent in smoMad– clones that cannot respondto Hh or Dpp (Fig. 3F). The effect was cell-autonomous. Sometimes alow level of So protein remained. This did not seem to correlate withthe clone boundaries, so it was hard to conclude that such residualexpression indicates a role for another signaling pathway, althoughthis might be the case. Taken together, the data indicate that Dpp andHh act redundantly to establish most So expression.
Multiple signals turn on Eya expression
Similar experiments led to a complex picture for regulation of Eya(Fig. 4; higher magnification images in Supplementary Fig. 1). Dppsignaling is sufficient to turn on eya expression (Fig. 1I). Although Dppsignaling contributes to initiating eya expression promptly at theproper location, Eya is only delayed in the absence of Dpp signaling(Fig. 4A) (Curtiss andMlodzik, 2000). We found that although Eyawasstill expressed in smoMad– cells, therewas a longer delay than inMad–
clones alone (Figs. 4B). This suggested that Hh also contributes to eyaexpression, but that additional cues can activate Eya more posteriorly.
Eya expression was still further delayed in Mad Su(H) ci– cellscompared to Mad ci–, indicating that N signaling contributes to Eyaexpression (Fig. 4C, and data not shown). Eya expression was normalin Su(H)– or Su(H) ci– clones, showing that Eyawas not very dependenton N when Dpp signaling was normal (data not shown). Although cihas both positive and negative roles in Hh signaling (Methot andBasler, 1999; Variosalo and Taipale, 2008), ci mutant cells cannot beinfluenced by Hh so spatiotemporal differences between ci mutantcells must indicate patterning by other pathways.
Eya expression was reduced and slightly delayed in mago– clonesthat lack Ras signaling (Fig. 4D). The absence of Dpp and Ras signalingin mago shn– clones did not delay Eya expression more than in shn–
clones, although Eya levels were also reduced as inmago– clones (Figs.4E and F). Thus, MAPK signaling appears to contibute to the level ofEya expression, Dpp signaling to the timely onset. These data suggestthat in addition to Dpp signaling, Hh, N, and MAPK signaling eachmake contributions to eya expression. It is possible that still furtherspatial cues exist, because none of the genotypes yet examined abolishEya expression.
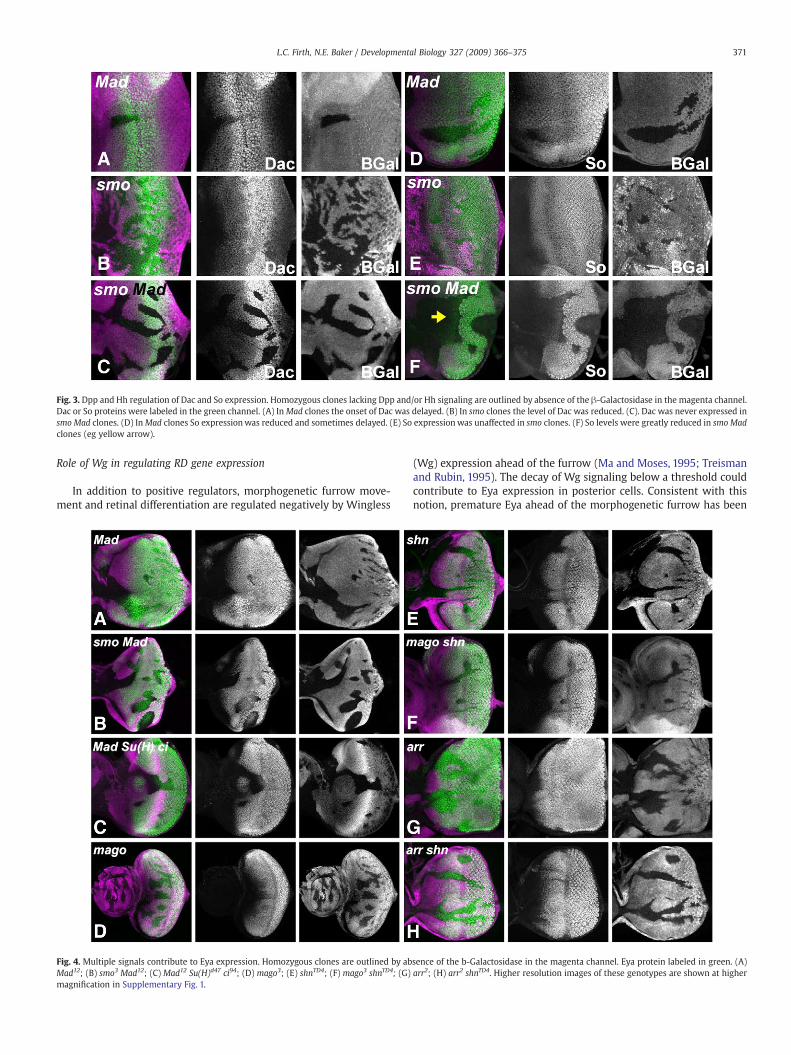
Fig. 3. Dpp and Hh regulation of Dac and So expression. Homozygous clones lacking Dpp and/or Hh signaling are outlined by absence of the β-Galactosidase in the magenta channel.Dac or So proteins were labeled in the green channel. (A) InMad clones the onset of Dac was delayed. (B) In smo clones the level of Dac was reduced. (C). Dac was never expressed insmo Mad clones. (D) InMad clones So expressionwas reduced and sometimes delayed. (E) So expressionwas unaffected in smo clones. (F) So levels were greatly reduced in smo Madclones (eg yellow arrow).
371L.C. Firth, N.E. Baker / Developmental Biology 327 (2009) 366–375
Role of Wg in regulating RD gene expression
In addition to positive regulators, morphogenetic furrow move-ment and retinal differentiation are regulated negatively by Wingless
Fig. 4. Multiple signals contribute to Eya expression. Homozygous clones are outlined by abMad12; (B) smo3 Mad12; (C) Mad12 Su(H)d47 ci94; (D) mago3; (E) shnTD4; (F) mago3 shnTD4; (G)magnification in Supplementary Fig. 1.
(Wg) expression ahead of the furrow (Ma and Moses, 1995; Treismanand Rubin, 1995). The decay of Wg signaling below a threshold couldcontribute to Eya expression in posterior cells. Consistent with thisnotion, premature Eya ahead of the morphogenetic furrow has been
sence of the b-Galactosidase in the magenta channel. Eya protein labeled in green. (A)arr2; (H) arr2 shnTD4. Higher resolution images of these genotypes are shown at higher
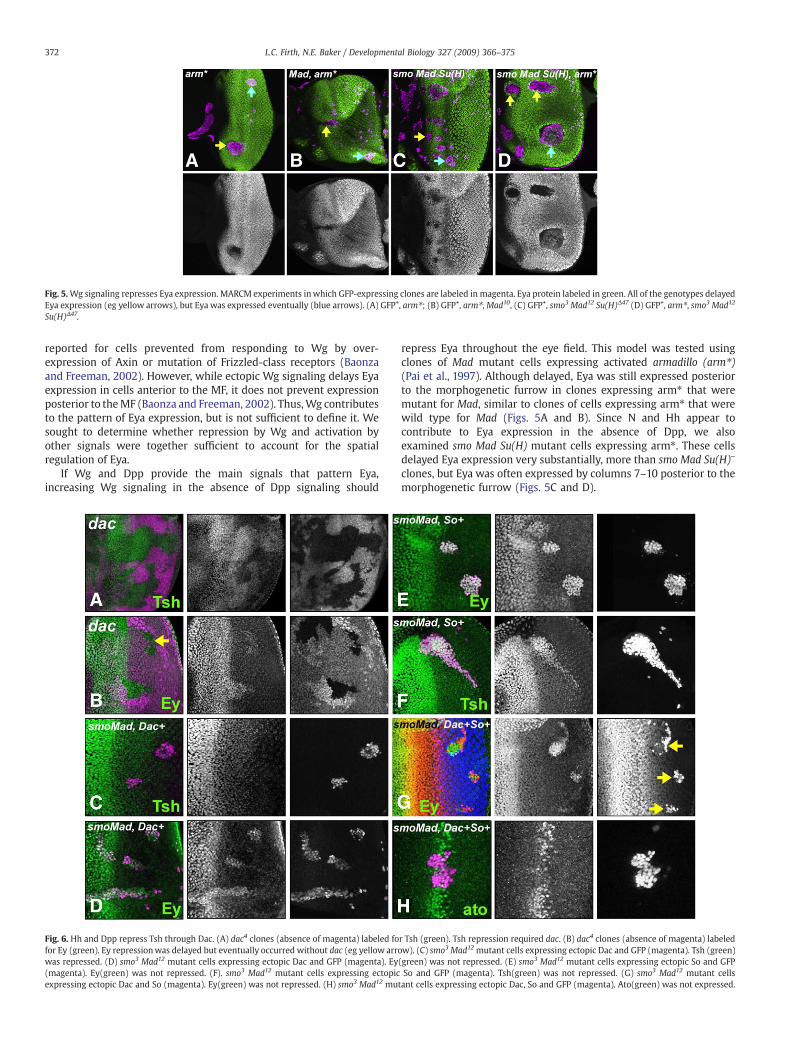
Fig. 5.Wg signaling represses Eya expression. MARCM experiments inwhich GFP-expressing clones are labeled in magenta. Eya protein labeled in green. All of the genotypes delayedEya expression (eg yellow arrows), but Eya was expressed eventually (blue arrows). (A) GFP+, arm⁎; (B) GFP+, arm⁎,Mad10, (C) GFP+, smo3 Mad12 Su(H)Δ47 (D) GFP+, arm⁎, smo3 Mad12
Su(H)Δ47.
372 L.C. Firth, N.E. Baker / Developmental Biology 327 (2009) 366–375
reported for cells prevented from responding to Wg by over-expression of Axin or mutation of Frizzled-class receptors (Baonzaand Freeman, 2002). However, while ectopic Wg signaling delays Eyaexpression in cells anterior to the MF, it does not prevent expressionposterior to theMF (Baonza and Freeman, 2002). Thus,Wg contributesto the pattern of Eya expression, but is not sufficient to define it. Wesought to determine whether repression by Wg and activation byother signals were together sufficient to account for the spatialregulation of Eya.
If Wg and Dpp provide the main signals that pattern Eya,increasing Wg signaling in the absence of Dpp signaling should
Fig. 6. Hh and Dpp repress Tsh through Dac. (A) dac4 clones (absence of magenta) labeled fofor Ey (green). Ey repressionwas delayed but eventually occurred without dac (eg yellow arrowas repressed. (D) smo3 Mad12 mutant cells expressing ectopic Dac and GFP (magenta). Ey(magenta). Ey(green) was not repressed. (F). smo3 Mad12 mutant cells expressing ectopicexpressing ectopic Dac and So (magenta). Ey(green) was not repressed. (H) smo3 Mad12 mu
repress Eya throughout the eye field. This model was tested usingclones of Mad mutant cells expressing activated armadillo (arm⁎)(Pai et al., 1997). Although delayed, Eya was still expressed posteriorto the morphogenetic furrow in clones expressing arm⁎ that weremutant for Mad, similar to clones of cells expressing arm⁎ that werewild type for Mad (Figs. 5A and B). Since N and Hh appear tocontribute to Eya expression in the absence of Dpp, we alsoexamined smo Mad Su(H) mutant cells expressing arm⁎. These cellsdelayed Eya expression very substantially, more than smo Mad Su(H)–
clones, but Eya was often expressed by columns 7–10 posterior to themorphogenetic furrow (Figs. 5C and D).
r Tsh (green). Tsh repression required dac. (B) dac4 clones (absence of magenta) labeledw). (C) smo3 Mad12 mutant cells expressing ectopic Dac and GFP (magenta). Tsh (green)(green) was not repressed. (E) smo3 Mad12 mutant cells expressing ectopic So and GFPSo and GFP (magenta). Tsh(green) was not repressed. (G) smo3 Mad12 mutant cells
tant cells expressing ectopic Dac, So and GFP (magenta). Ato(green) was not expressed.

Fig. 7. Extracellular signals regulate and act through retinal determination genes. Acartoon showing the spatial signals that affect retinal determination gene expression inthe third instar eye disc, as the morphogenetic furrow induces differentiation withincells coexpressing Eya, So, and Dac, and extinguishes Hth, Ey and Tsh expression. Clonalanalysis has defined cell-autonomous effects of extracellular signaling pathways, whichmust be ‘direct’ in that any intermediate steps have to be intracellular. Themorphogenetic furrow is associated with and driven forwards by expression of Dppand Hh; ligands for N and receptor tyrosine kinases are also expressed. Effects ongrowth suggest that EGFR and Ras function anterior to the furrow as well as posterior,although the ligands involved are uncertain. Non-autonomous interactions betweenthese signals, which contribute to their pattern of expression, are not shown here.Morphogenetic furrow movement is antagonized by Wg, which is expressed in theanterior eye disc. The retinal determination genes are color-coded to indicate whichextracellular signals are responsible for their spatial and temporal regulation. Roles ofWg signaling in the initiation of Hth, Ey and Tsh expression at earlier developmentalstages are not shown. Potential indirect regulation of the Ey and Tsh genes is indicatedby grey arrows. Our findings are consistent with repression of Tsh and to some extent Eyby Hh occurring indirectly via the activation of Dac. The data do not exclude additional,parallel regulation by Hh and Dpp directly, however. In addition, any of the regulatoryrelationships shown here may act through unidentified intermediates, so long as thesefunction cell-autonomously. For example, Dac might regulate Tsh or Ey indirectly.
373L.C. Firth, N.E. Baker / Developmental Biology 327 (2009) 366–375
These findings were consistent with the notion that Wgsignaling contributed to patterning Eya expression, in addition tosignals that act positively. However, Wg might not act indepen-dently, but instead antagonize one or more positive signals.Conversely, one or more positive signals might contribute onlyindirectly, by interfering with Wg. These possibilities wereinvestigated for the case of Dpp, using loss of function mutations.As expected, Eya was expressed prematurely in cells mutated forthe Wg receptor Arrow (Arr) (Fig. 4G). If the role of Wg were toprevent Dpp from inducing Eya, we would expect that Wg signalingwould not affect cells unable to respond to Dpp, which would delayEya expression. By contrast, Eya expression in arr shn doublemutant cells began at an intermediate stage, close to that of wildtype cells (Fig. 4H). This suggests that Wg and Dpp contributeindependently to Eya expression, in addition to an underlyingpattern of other spatial information that includes the Hh, MAPKand N pathways, and perhaps other pathways not investigated here,which can turn on Eya almost normally in the absence of both Wgand Dpp signals.
Dpp and Hh repress Tsh, but not Ey, through Dac
Our studies identify extracellular sources of patterning for thehth, Ey, tsh, dac, and eya genes that are direct in the sense that cell-autonomous requirements rule out indirect effects mediated byother cell–cell signaling mechanisms. The cell-autonomousresponses might be multi-layered, however. For example, regulationof ey, tsh, so and dac expression by Hh and Dpp signaling couldindicate direct binding of the Ci, Mad and Brk transcription factorsto each of the Ey, tsh, so and dac genes (Brk is a repressor controlledby Mad and Shn so that Brk targets are under Dpp regulation(Affolter and Basler, 2007)). Alternatively, Hh and Dpp mightdirectly regulate one of these genes, or even some other transcrip-tion factor gene, which then regulates ey, tsh, and dac expressioncell-autonomously.
There is already an extensive literature documenting cross-regulation between eye specification genes (reviewed in Pappu andMardon, 2002). We found evidence that Tsh is partly regulatedthough Dac, which is the more direct target of Dpp and Hh signaling.In dac− clones, Tsh was not repressed posterior to the MF, andrepression of Ey was delayed (Figs. 6A and B). Reduced Ey expressionwas still present in more posterior dac− clones however (arrow in Fig.6B). If Hh and Dpp repress Ey and Tsh by establishing Dac, we wouldpredict that Ey and Tsh would be repressed in smo Mad− clones if Dacexpression was restored. We found that smo Mad− clones thatexpressed dac repressed Tsh but not Ey (Figs. 6C and D). Thisindicated that Dpp and Hh signaling were dispensable for Tshrepression if Dac was present, consistent with Dac expressionmediating the repression of Tsh posterior to the furrow. Expressionof Dac was not sufficient to repress Tsh prematurely ahead of thefurrow, however, and the two proteins are coexpressed in some cellsin normal development. Although Dac was partially required for Eyrepression, ectopic Dac was not sufficient to repress Ey in smo Madmutant cells (data not shown), indicating that there is another targetof Hh and Dpp that is required.
As there is evidence that So might repress Ey (Pignoni et al.,1997), we tested whether Hh and Dpp repressed Ey through So byexpressing So in smo Mad mutant cells. However, So was notsufficient to repress Ey (Fig. 6E). So also did not restore Tshrepression in smo Mad mutant cells (Fig. 6F). Co-over expression ofDac and So had only minor effects on Ey expression in smo Madmutant cells (Fig. 6G).
We also noted that expression of Dac and So failed to restoreeither Ato expression or ommatidial differentiation to smo Mad−
clones, even at stages where smo Mad cells express Eya (Fig. 6H,and data not shown).
Discussion
Retinal determination gene expression is regulated spatially andtemporally by extracellular signals
We found that all the extracellular signaling pathways played rolesin spatial and temporal regulation of RD genes. Although Dpp wasimportant, none of the hth, tsh, ey, eya, dac, and so genes wereregulated exclusively by Dpp. Instead, three distinct combinatorial‘codes’, comprising Dpp+Hh, Dpp+Ras, and Dpp+Hh+Ras+Notch+Wg, each contributed to the expression of particular RD genes. Hthwas repressed redundantly by Dpp and Ras signaling, Tsh, Ey, So andDac were regulated redundantly by Dpp and Hh signals, and Eyaregulation was affected by all five extracellular signaling pathwaysexamined. Therefore, dynamic RD gene expression is a function ofextracellular signaling by all the core signaling pathways, and perhapsby still further spatial cues not examined in our study (Fig. 7).
Two kinds of data show that signals other than Dpp wereimportant in wild type flies, and did not just contribute to robustnessor as back-up pathways that are mainly relevant in mutant genotypes.First, Dpp signaling alonewas not sufficient to regulate all the genes inthe third instar. Specifically, we found that activated Tkv was notsufficient for Ey repression, only partially repressed Tsh, and was notsufficient to turn on Dac throughout the eye field (Figs.1F–J). Secondly,cells that remain able to respond to Dpp signaling, but are deficient inresponding to other signals, showed abnormal patterns of geneexpression that indicated roles for other signals even when Dppsignaling was intact. Specifically, cells defective in Ras signaling

374 L.C. Firth, N.E. Baker / Developmental Biology 327 (2009) 366–375
delayed repression of Hth (Fig. 2L) and had reduced levels of Eya (Fig.4D); cells defective in Hh signaling had reduced Dac levels anddelayed repression of Ey and Tsh (Figs. 2B, E). Therefore, Dpp signalingis important but not sufficient by itself to specify the normal spatio-temporal pattern of retinal determination gene expression.
Retinal determination genes may define eye-specific, indirect targetgenes of extracellular signals
One idea has been that retinal determination proteins define theeye field through eye-specific responses to core extracellularsignaling pathways (Curtiss et al., 2002; Mann and Carroll, 2002).How can RD genes play such a high-level role if RD gene expressionis itself changing dynamically in response to all the signalingpathways, like all the effector genes responsible for eye differentia-tion and morphogenesis? Rather than surrender the idea that RDgenes, like other master regulatory genes, define how a particularorgan interprets positional information, we suggest another possibleexplanation. If dynamic RD gene expression captures enoughfeatures of the extracellular signaling pattern, overlapping expressionpatterns of RD genes can themselves encapsulate the activity ofextracellular signals, and act as a relay of this positional information.In this view, target genes of RD proteins are being regulated by theextracellular positional information, but indirectly, through thedynamic, overlapping combinations of RD gene expression. Thisregulation would be eye-specific. For example, a gene that lacks anybinding site for Ci could respond cell autonomously to Hh signalingthrough regulation by one or more retinal determination genes thatare regulated by Hh, and this Hh-responsiveness would be eyespecific. This model would only be plausible if RD gene expressionreflected the activity of all the major extracellular signaling path-ways, which we find is indeed the case.
Is there any evidence for eye genes that are regulated byextracellular signals indirectly through an RD gene relay? The atonalgene, which encodes the proneural bHLH protein for the founder R8photoreceptor cells of the neural retina, may be an example (Tanaka-Matakatsu and Du, 2008). Hh and Dpp signaling induce atotranscription in a stripe just ahead of the morphogenetic furrow(Jarman et al., 1994; Greenwood and Struhl,1999; Curtiss andMlodzik,2000). This ato expression depends an enhancer located 3′ to thegene, yet the 384 bp that contain this enhancer lack any consensusbinding site for the transcription factors Ci, Mad, or Brk, whichmediate Hh and Dpp signaling (Sun et al., 1998; Zhang et al., 2006)(our unpublished observations). Instead, the enhancer activitydepends on binding sites for Ey and So (Zhang et al., 2006). Ourstudy now shows that the expression of both ey and so depends on Hhand Dpp signaling. Therefore the Hh- and Dpp-dependent transcrip-tion of ato may be explained by Hh- and Dpp-regulation of ey and so.Theremust be another Hh/Dpp-dependent factor in addition to Ey andSo, however, because restoring So (and Dac) expression did not rescueato expression to smo Mad mutant cells (these smo Mad cells alreadyexpress Ey and Eya) (Fig. 6H). We predict that the missing factordepends heavily on Hh signaling, because ato expression is morecritically dependent on smo than on Dpp signaling (Dominguez, 1999;Fu and Baker, 2003).
It will require more than one example for the positionalinformation relay model to prove useful. It will be interesting todetermine whether Dpp regulates ey and tsh through Mad or Brkbinding, or indirectly through Hth and Dac (Fig. 7). The Ras and Notchpathways have many roles in eye-specific cell fates, and it will beinteresting to determine whether some of the target genes involvedare regulated indirectly through Hth and Eya, which respond to Rasand Notch signals (Fig. 7). The ‘positional-information relay’ modeldoes not mean that RD gene products cannot act in combination withextracellular signals on the same promoters as has been proposedbefore. Indeed this is likely at some promoters (Curtiss et al., 2002;
Mann and Carroll, 2002; Rogers et al., 2005; Hayashi et al., 2008) andprobably necessary in activating expression of the RD genesthemselves (Bessa and Casares, 2005; Pappu et al., 2005). We suggest,however, that evolution used RD genes and the relay mechanism toexpand the repertoire of eye-specific gene expression and recruittarget genes that lack binding sites for direct transducers ofextracellular signals.
Acknowledgments
We gratefully thank J. Curtiss, S. Cohen, R. Holmgren J. Kumar, R.Padgett, R. Mann, G. Mardon, F. Pignoni, L. Raftery, I. Rebay and theDrosophila Stock Center at Bloomington, Indiana for the antibodiesand for the genetic strains. The anti-Dac, anti-Elav, anti-Eya, and anti-Wg antibodies were obtained from the Developmental StudiesHybridoma Bank developed under the auspices of the NICHD andmaintained by The University of Iowa, Department of BiologicalSciences, Iowa City, IA 52242. We thank A. Cvekl, C. Desplan, R. Mann,F. Pignoni and J. Treisman for comments on the manuscript, and J.Treisman and J.-Y. Roignant for generously sharing unpublishedreagents and data. Confocal microscopy was performed in theAnalytical Imaging Facility at the Albert Einstein College of Medicine.This work was supported by grant from the National Institutes ofHealth (GM47892).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.ydbio.2008.12.021.
References
Affolter, M., Basler, K., 2007. The Decapentaplegic morphogen gradient: from patternformation to growth regulation. Nat. Rev. Genet. 8, 663–674.
Arora, K., Dai, H., Kazuko, S.G., Jamal, J., O'Connor, M.B., Letsou, A., Warrior, R., 1995.The Drosophila schnurri gene acts in the Dpp/TGF beta signaling pathway andencodes a transcription factor homologous to the human MBP family. Cell 81,781–790.
Baonza, A., Freeman,M., 2001. Notch signalling and the initiation of neural developmentin the Drosophila eye. Development 128, 3889–3898.
Baonza, A., Freeman, M., 2002. Control of Drosophila eye specification by Winglesssignalling. Development 129, 5313–5322.
Bessa, J., Casares, F., 2005. Restricted teashirt expression confers eye-specificresponsiveness to Dpp and Wg signals during eye specification in Drosophila.Development 132, 5011–5020.
Bessa, J., Gebelein, B., Pichaud, F., Casares, F., Mann, R.S., 2002. Combinatorial control ofDrosophila eye development by Eyeless, Homothorax, and Teashirt. Genes Dev. 16,2415–2427.
Blackman, R.K., Sanicola, M., Raftery, L.A., Gillevet, T., Gelbart, W.M., 1991. An extensive3′ cis-regulatory region directs the imaginal disk expression of decapentaplegic, amember of the TGF-β family in Drosophila. Development 111, 657–666.
Bonini, N.M., Leiserson, W.M., Benzer, S., 1993. The eyes absent gene: genetic controlof cell survival and differentiation in the developing Drosophila eye. Cell 72,379–395.
Bonini, N.M., Bui, Q.T., Gray-Board, G.L., Warrick, J.M., 1997. The Drosophila eyes absentgene directs ectopic eye formation in a pathway conserved between flies andvertebrates. Development 124, 4819–4826.
Boswell, R.E., Prout, M.E., Steichen, J.C., 1991. Mutations in a newly identified Drosophilamelanogaster gene, mago nashi, disrupt germ cell formation and result in theformation of mirror-image symmetrical double abdomen embryos. Development113, 373–384.
Bray, S.J., 2006. Notch signalling: a simple pathway becomes complex. Nat. Rev. Mol. CellBiol. 7, 678–689.
Brook, W.J., Cohen, S.M., 1996. Antagonistic interactions between wingless anddecapentaplegic responsible for dorsal–ventral pattern in the Drosophila leg.Science 273, 1373–1377.
Casares, F., Mann, R.S., 1998. Control of antennal versus leg development in Drosophila.Nature 392, 723–726.
Chase, B.A., Baker, B.S., 1995. A genetic analysis of intersex, a gene regulating sexualdifferentiation in Drosophila melanogaster females. Genetics 139, 1649–1661.
Chen, Y., Struhl, G., 1998. In vivo evidence that Patched and Smoothened constitutedistinct binding and transducing components of a Hh receptor complex.Development 125, 4943–4948.
Chen, R., Amoui, M., Zhang, Z., Mardon, G., 1997. Dachshund and eyes absent proteinsform a complex and function synergistically to induce ectopic eye development inDrosophila. Cell 91, 893–903.

375L.C. Firth, N.E. Baker / Developmental Biology 327 (2009) 366–375
Chen, R., Halder, G., Zhang, Z., Mardon, G., 1999. Signaling by the TGF-beta homologdecapentaplegic functions reiteratively within the network of genes controllingretinal cell fate determination in Drosophila. Development 126, 935–943.
Cheyette, B.N., Green, P.J., Martin, K., Garren, H., Hartenstein, V., Zipursky, S.L., 1994. TheDrosophila sine oculis locus encodes a homeodomain-containing protein requiredfor the development of the entire visual system. Neuron. 12, 977–996.
Clevers, H., 2006. Wnt/beta-catenin signaling in development and disease. Cell 127,469–480.
Curtiss, J., Mlodzik, M., 2000. Morphogenetic furrow initiation and progression duringeye development in Drosophila: the roles of decapentaplegic, hedgehog and eyesabsent. Development 127, 1325–1336.
Curtiss, J., Halder, G., Mlodzik, M., 2002. Selector and signalling molecules cooperate inorgan patterning. Nat. Cell Biol. 4, E48–51.
Dominguez, M., 1999. Dual role for Hedgehog in the regulation of the proneural geneatonal during ommatidia development. Development 126, 2345–2353.
Firth, L.C., Baker, N.E., 2005. Extracellular signals responsible for spatially regulatedproliferation in the differentiating Drosophila eye. Dev. Cell. 8, 541–551.
Firth, L.C., Li, W., Zhang, H., Baker, N.E., 2006. Analyses of RAS regulation of eyedevelopment in Drosophila melanogaster. Methods Enzymol. 407, 711–721.
Freeman, M., 1996. Reiterative use of the EGF receptor triggers differentiation of all celltypes in the Drosophila eye. Cell 87, 651–660.
Fu, W., Baker, N.E., 2003. Deciphering synergistic and redundant roles of Hedgehog,Decapentaplegic and Delta that drive the wave of differentiation in Drosophila eyedevelopment. Development 130, 5229–5239.
Golic, K.G., 1991. Site-specific recombination between homologous chromosomes inDrosophila. Science 252, 958–961.
Greenwood, S., Struhl, G., 1999. Progression of the morphogenetic furrow in the Dro-sophila eye: the roles of Hedgehog, Decapentaplegic and the Raf pathway.Development 126, 5795–5808.
Halder, G., Callaerts, P., Gehring, W.J., 1995. Induction of ectopic eyes by targetedexpression of the eyeless gene in Drosophila. Science 267, 1788–1792.
Halder, G., Callaerts, P., Flister, S., Walldorf, U., Kloter, U., Gehring, W.J., 1998. Eyelessinitiates the expression of both sine oculis and eyes absent during Drosophilacompound eye development. Development 125, 2181–2191.
Hayashi, T., Xu, C., Carthew, R.W., 2008. Cell-type-specific transcription of Prospero iscontrolled by combinatorial signaling in the Drosophila eye. Development 135,2787–2796.
Jarman, A.P., Grell, E.H., Ackerman, L., Jan, L.Y., Jan, Y.N., 1994. atonal is the proneuralgene for Drosophila photoreceptors. Nature 369, 398–400.
Karim, F.D., Rubin, G.M., 1998. Ectopic expression of activated Ras1 induces hyperplasticgrowth and increased cell death in Drosophila imaginal discs. Development 125,1–9.
Kenyon, K.L., Ranade, S.S., Curtiss, J., Mlodzik, M., Pignoni, F., 2003. Coordinatingproliferation and tissue specification to promote regional identity in the Drosophilahead. Dev. Cell. 5, 403–414.
Kim, S.S., Zhang, R., Braunstein, S.E., Joachlmlak, A., Cvekl, A., Hegde, R.S., 2002.Structure of the retinal determination protein Dachshund reveals a DNA bindingmotif. Structure 10, 787–795.
Lee, J.D., Treisman, J.E., 2001. The role of Wingless signaling in establishing theanteroposterior and dorsoventral axes of the eye disc. Development 128,1519–1529.
Lee, J.D., Treisman, J.E., 2002. Regulators of the morphogenetic furrow. Results Probl.Cell Differ. 37, 21–33.
Lee, T., Luo, L., 2001. Mosaic analysis with a repressible cell marker (MARCM) for Dro-sophila neural development. Trends Neurosci. 24, 251–254.
Ma, C., Moses, K., 1995. Wingless and patched are negative regulators of themorphogenetic furrow and can affect tissue polarity in the developing Drosophilacompound eye. Development 121, 2279–2289.
Ma, C., Zhou, Y., Beachy, P.A., Moses, K., 1993. The segment gene hedgehog is required forprogression of the morphogenetic furrow in the developing Drosophila eye. Cell 75,927–938.
Mann, R.S., Carroll, S.B., 2002. Molecular mechanisms of selector gene function andevolution. Curr. Opin. Genet. Dev. 12, 592–600.
Mardon, G., Solomon, N.M., Rubin, G.M., 1994. dachshund encodes a nuclear proteinrequired for normal eye and leg development in Drosophila. Development 120,3473–3486.
Methot, N., Basler, K., 1999. Hedgehog controls limb development by regulating theactivities of distinct transcriptional activator and transcriptional repressor forms ofCubitus interruptus. Cell 96, 819–831.
Morata, G., Ripoll, P., 1975. Minutes: mutants of Drosophila autonomously affecting celldivision rate. Dev. Biol. 42, 211–221.
Morel, V., Schweisguth, F., 2000. Repression by suppressor of Hairless and activation byNotch are required to define a single row of single-minded expressing cells in theDrosophila embryo. Genes Dev. 14, 377–388.
Mutsuddi, M., Chaffee, B., Cassidy, J., Silver, S.J., Tootle, T.L., Rebay, I., 2005. Using Dro-sophila to decipher how mutations associated with human branchio-oto-renalsyndrome and optical defects compromise the protein tyrosine phosphatase andtranscriptional functions of eyes absent. Genetics 170, 687–695.
Nilsson, D., 2004. Eye evolution: a question of genetic promiscuity. Curr. Opin.Neurobiol. 14, 407–414.
O'Neill, E.M., Rebay, I., Tjian, R., Rubin, G.M., 1994. The activities of two Ets-relatedtranscription factors required for Drosophila eye development are modulated bythe Ras/MAPK pathway. Cell 78, 137–147.
Pai, L.M., Orsulic, S., Bejsovec, A., Peifer, M., 1997. Negative regulation of Armadillo, aWingless effector in Drosophila. Development 124, 2255–2266.
Pappu, K., Mardon, G., 2002. Retinal specification and determination in Drosophila.Results Probl. Cell Differ. 37, 5–20.
Pappu, K., Chen, R., Middlebrooks, B.W., Woo, C., Heberlein, U., Mardon, G., 2003.Mechanism of Hedgehog signaling during Drosophila eye development. Develop-ment 130, 3053–3062.
Pappu, K.S., Ostrin, E.J., Middlebrooks, B.W., Sili, B.T., Chen, R., Atkins, M.R., Gibbs, R.,Mardon, G., 2005. Dual regulation and redundant function of two eye-specificenhancers of the Drosophila retinal determination gene dachshund. Development132, 2895–2905.
Pichaud, F., Casares, F., 2000. homothorax and iroquois C genes are required for theestablishment of territories within the developing eye disc. Mech. Dev. 96,15–25.
Pignoni, F., Hu, B., Zavitz, K.H., Xiao, J., Garrity, P.A., Zipursky, S.L., 1997. The eye-specification proteins So and Eya form a complex and regulate multiple steps inDrosophila eye development. Cell 91, 881–891.
Pignoni, F., Zipursky, L., 1997. Induction of Drosophila eye development by Decapenta-plegic. Development 124, 271–278.
Rogers, E.M., Brennan, C.A., Mortimer, N.T., Cook, S., Morris, A.R., Moses, K., 2005.Pointed regulates an eye-specific transcriptional enhancer in the Drosophilahedgehog gene, which is required for the movement of the morphogenetic furrow.Development 132, 4833–4843.
Sekelsky, J.J., Newfeld, S.J., Raftery, L.A., Chartoff, E.H., Gelbart, W.M., 1995. Geneticcharacterization and cloning of mothers against dpp, a gene required fordecapentaplegic function in Drosophila melanogaster. Genetics 139, 1347–1358.
Serikaku, M.A., O'Tousa, J.E., 1994. sine oculis is a homeobox gene required for Droso-phila visual system development. Genetics 138, 1137–1150.
Shen, W., Mardon, G., 1997. Ectopic eye development in Drosophila induced by directeddachshund expression. Development 124, 45–52.
Silver, S.J., Rebay, I., 2005. Signaling circuitries in development: insights from the retinaldetermination gene network. Development 132, 3–13.
Sun, Y., Jan, L.Y., Jan, Y.N., 1998. Transcriptional regulation of atonal duringdevelopment of the Drosophila peripheral nervous system. Development 125,3731–3740.
Tanaka-Matakatsu, M., Du, W., 2008. Direct control of the proneural gene atonal byretinal determination factors during Drosophila eye development. Dev. Biol. 313,787–801.
Treisman, J.E., 2004. How to make an eye. Development 131, 3823–3827.Treisman, J.E., Rubin, G.M., 1995. Wingless inhibits morphogenetic furrowmovement in
the Drosophila eye disc. Development 121, 3519–3527.Variosalo, M., Taipale, J., 2008. Hedgehog: functions and mechanisms. Genes and Dev.
22, 2454–2472.Vincent, J., Girdham, C., O'Farrell, P., 1994. A cell-autonomous, ubiquitous marker for the
analysis of Drosophila genetic mosaics. Dev. Biol. 164, 328–331.Wassarman, D.A., Therrien, M., Rubin, G.M., 1995. The Ras signaling pathway in Dro-
sophila. Curr. Opin. Genet. Dev. 5, 44–50.Weasner, B., Salzer, C., Kumar, J.P., 2006. Sine oculis, a member of the SIX family of
transcription factors, directs eye formation. Dev. Biol. 303, 756–771.Wehrli, M., Dougan, S.T., Caldwell, K., O'Keefe, L., Schwartz, S., Vaizel-Ohayon, D.,
Schejter, E.D., Tomlinson, A., DiNardo, S., 2000. arrow encodes an LDL-receptor-related protein essential for Wingless signaling. Nature 407, 527–530.
Wu, J., Cohen, S.M., 2000. Proximal distal axis formation in the Drosophila leg: distinctfunctions of teashirt and homothorax in the proximal leg. Mech. Dev. 94, 47–56.
Xu, T., Rubin, G.M., 1993. Analysis of genetic mosaics in the developing and adult Dro-sophila tissues. Development 117, 1223–1236.
Zhang, T., Ranade, S., Cai, C.Q., Clouser, C., Pignoni, F., 2006. Direct control ofneurogenesis by selector factors in the fly eye: regulation of atonal by Ey and So.Development 133, 4881–4889.


















