Responses of neurons in the region of human thalamic principal somatic sensory nucleus to mechanical...
15
Responses of Neurons in the Region of Human Thalamic Principal Somatic Sensory Nucleus to Mechanical and Thermal Stimuli Graded Into the Painful Range J.-I. LEE,* P.M. DOUGHERTY, D. ANTEZANA, AND F.A. LENZ Departments of Neurosurgery and Neuroscience, Johns Hopkins Hospital, Baltimore, Maryland 21287-7713 ABSTRACT The role of the region of the principal somatic sensory nucleus of the human thalamus (ventral caudal – Vc) in signaling painful sensations is unclear. We have now studied the response of cells (n 5 57) in this region to both thermal and mechanical stimuli graded into the painful range during surgeries (n 5 24) for treatment of movement disorders. Fifteen cells had a graded response to mechanical stimuli extending into the painful range and, thus, were classified in the wide dynamic range (WDR) category. The mean stimulus-response function of cells in the WDR class, normalized to baseline, showed a fourfold mean increase in firing rate above baseline across the mechanical series of stimuli. Seven of these cells responded to heat stimuli (WDR-H) and two responded to cold stimuli (WDR-C). Twenty-five cells were in a class (multiple receptive – MR) that showed a response to both brush and compressive stimuli, although the responses were not graded into the painful range. Three of these cells (MR-H) had a response to heat stimuli and five cells responded to cold stimuli (MR-C). Nine cells responded to brushing without a response to the compressive stimuli (low threshold – LT). Cells responsive to painful mechanical and thermal stimuli were located throughout the thalamic region where cells responded to nonpainful cutaneous stimulation. These results show that cells in the region of the human thalamic principal somatic sensory nucleus respond to mechanical and thermal stimuli extending into the painful range. J. Comp. Neurol. 410:541–555, 1999. r 1999 Wiley-Liss, Inc. Indexing terms: pain; ventral posterior nucleus; posterior nucleus; human thalamus; thermoreception; mechanoreception In the monkey, several lines of evidence implicate the region of the thalamic principal somatic sensory nucleus (ventral posterior nucleus - VP) in pain signaling path- ways (Price and Dubner, 1977; Willis, 1985). Anatomic studies demonstrate that the spinothalamic tract (STT) terminates in the region of VP (Boivie, 1979; Berkley, 1980; Jones, 1985; Willis, 1985; Apkarian and Hodge, 1989; Ralston and Ralston, 1992, 1994; Craig et al., 1994). Physiologic studies demonstrate that cells in VP, and posterior inferior to VP, respond to noxious stimuli in anesthetized monkeys (Perl and Whitlock, 1961; Kenshalo et al., 1980; Casey and Morrow, 1983, 1987; Chung et al., 1986a,b; Bushnell and Duncan, 1987; Morrow and Casey, 1992; Bushnell et al., 1993; Apkarian and Shi, 1994) and unanesthetized monkeys (Casey, 1966; Bushnell and Dun- can, 1987; Bushnell et al., 1993). Blockade of the activity of these cells by thalamic injection of local anesthetic signifi- cantly interferes with the monkey’s ability to discriminate temperature in both the innocuous and noxious range (Duncan et al., 1993). These studies establish that cells in the region of the monkey principal somatic sensory nucleus are involved in discrimination of cutaneous thermal stimuli into the noxious range. Anatomic studies in patients after cordotomy demon- strate that nuclei where the STT terminates in humans Grant sponsor: National Institutes of Health; Grant number: P01 NS32386-Proj. 1. *Correspondence to: F. A. Lenz, Department of Neurosurgery, Meyer Building 7–113, Johns Hopkins Hospital, 600 North Wolfe Street, Balti- more, MD 21287-7713. E-mail: [email protected] Received 30 July 1998; Revised 25 January 1999; Accepted 4 March 1999 THE JOURNAL OF COMPARATIVE NEUROLOGY 410:541–555 (1999) r 1999 WILEY-LISS, INC.
Transcript of Responses of neurons in the region of human thalamic principal somatic sensory nucleus to mechanical...

Responses of Neurons in the Region ofHuman Thalamic Principal Somatic
Sensory Nucleus to Mechanicaland Thermal Stimuli Graded
Into the Painful Range
J.-I. LEE,* P.M. DOUGHERTY, D. ANTEZANA, AND F.A. LENZ
Departments of Neurosurgery and Neuroscience, Johns Hopkins Hospital,Baltimore, Maryland 21287-7713
ABSTRACTThe role of the region of the principal somatic sensory nucleus of the human thalamus
(ventral caudal – Vc) in signaling painful sensations is unclear. We have now studied theresponse of cells (n 5 57) in this region to both thermal and mechanical stimuli graded into thepainful range during surgeries (n 5 24) for treatment of movement disorders. Fifteen cells hada graded response to mechanical stimuli extending into the painful range and, thus, wereclassified in the wide dynamic range (WDR) category. The mean stimulus-response function ofcells in the WDR class, normalized to baseline, showed a fourfold mean increase in firing rateabove baseline across the mechanical series of stimuli. Seven of these cells responded to heatstimuli (WDR-H) and two responded to cold stimuli (WDR-C). Twenty-five cells were in a class(multiple receptive – MR) that showed a response to both brush and compressive stimuli,although the responses were not graded into the painful range. Three of these cells (MR-H)had a response to heat stimuli and five cells responded to cold stimuli (MR-C). Nine cellsresponded to brushing without a response to the compressive stimuli (low threshold – LT).Cells responsive to painful mechanical and thermal stimuli were located throughout thethalamic region where cells responded to nonpainful cutaneous stimulation. These resultsshow that cells in the region of the human thalamic principal somatic sensory nucleus respondto mechanical and thermal stimuli extending into the painful range. J. Comp. Neurol.410:541–555, 1999. r 1999 Wiley-Liss, Inc.
Indexing terms: pain; ventral posterior nucleus; posterior nucleus; human thalamus;
thermoreception; mechanoreception
In the monkey, several lines of evidence implicate theregion of the thalamic principal somatic sensory nucleus(ventral posterior nucleus - VP) in pain signaling path-ways (Price and Dubner, 1977; Willis, 1985). Anatomicstudies demonstrate that the spinothalamic tract (STT)terminates in the region of VP (Boivie, 1979; Berkley, 1980;Jones, 1985; Willis, 1985; Apkarian and Hodge, 1989;Ralston and Ralston, 1992, 1994; Craig et al., 1994).Physiologic studies demonstrate that cells in VP, andposterior inferior to VP, respond to noxious stimuli inanesthetized monkeys (Perl and Whitlock, 1961; Kenshaloet al., 1980; Casey and Morrow, 1983, 1987; Chung et al.,1986a,b; Bushnell and Duncan, 1987; Morrow and Casey,1992; Bushnell et al., 1993; Apkarian and Shi, 1994) andunanesthetized monkeys (Casey, 1966; Bushnell and Dun-can, 1987; Bushnell et al., 1993). Blockade of the activity of
these cells by thalamic injection of local anesthetic signifi-cantly interferes with the monkey’s ability to discriminatetemperature in both the innocuous and noxious range(Duncan et al., 1993). These studies establish that cells inthe region of the monkey principal somatic sensory nucleusare involved in discrimination of cutaneous thermal stimuliinto the noxious range.
Anatomic studies in patients after cordotomy demon-strate that nuclei where the STT terminates in humans
Grant sponsor: National Institutes of Health; Grant number: P01NS32386-Proj. 1.
*Correspondence to: F. A. Lenz, Department of Neurosurgery, MeyerBuilding 7–113, Johns Hopkins Hospital, 600 North Wolfe Street, Balti-more, MD 21287-7713. E-mail: [email protected]
Received 30 July 1998; Revised 25 January 1999; Accepted 4 March 1999
THE JOURNAL OF COMPARATIVE NEUROLOGY 410:541–555 (1999)
r 1999 WILEY-LISS, INC.

are similar to those in monkeys (Walker, 1943; Bowsher,1957; Mehler et al., 1960; Mehler, 1962, 1966). Our physi-ologic studies have demonstrated that cells in the region ofthe human principal somatic sensory nucleus (Ventralcaudal – Vc) respond to painful mechanical stimuli (Lenzet al., 1994b) and painful heat stimuli (Lenz et al., 1993b).The degree of convergence of thermal and mechanicalmodalities graded into the painful range has not previ-ously been studied. Cells responding to both types ofstimuli may explain both the sensation of pain, i.e.,hyperalgesia and allodynia, evoked by normally nonpain-ful stimuli (Fruhstorfer and Lindblom, 1984), and thealleviation of pain by thermal stimuli (Lehmann and deLateur, 1994). We now report responses of human thalamiccells to painful and nonpainful thermal and mechanicalstimuli in patients undergoing thalamic procedures for thetreatment of movement disorders.
MATERIALS AND METHODS
Thalamic exploration was performed as a stereotacticprocedure with use of the Leksell frame. First, the three-dimensional frame coordinates of the anterior and poste-rior commissures were measured by CT scan, MRI scan, orboth. A computer program then shrank or expanded thesagittal sections of the Schaltenbrand and Bailey Atlas(Schaltenbrand and Bailey, 1959) to conform to the pa-tient’s anterior commissure-posterior commissure (ACPC)line (Hawrylyshyn et al., 1976). Physiologic corroborationof anatomic loci was then performed under local anesthetic(i.e., patient fully conscious) by using single unit recordingand microstimulation with a high impedance platinum-iridium microelectrode (Lenz et al., 1988a). EMG activity
was routinely monitored in four muscles on the contralat-eral arm (Lenz et al., 1988b). Initial microelectrode trajec-tories targeted Vc as located on the computer-generatedmap described above (Hawrylyshyn et al., 1976). Themicroelectrode was switched to the output of an opticallyisolated, constant current stimulator (BSI-2 and 8PG-2,BAK Electronics, Rockville, MD). Microstimulation atcurrent levels of up to 40 µA was carried out at 0.5-mmintervals along the trajectory.
Preoperative testing
Physiologic definition of the anterior and inferior borderof region where cells respond to innocuous somatic sensoryinput is required to carry out thalamic stereotactic proce-dures for treatment of movement disorders. The presentstudies were carried out during trajectories through thisregion by adding about 1 hour to the operative time. Theprotocol of these studies was reviewed and approvedyearly by the Hopkins Joint Committee for Clinical Inves-tigation. All patients signed an informed consent form.
Each patient had a complete history, physical examina-tion, and MRI scan before the procedure. Initial somaticsensory testing included determination of mechanical andheat thresholds and description of these stimuli with useof a questionnaire of ideal type pain descriptors (Torgersonet al., 1988; Lenz et al., 1994a, 1998a) and verbal descrip-tor visual analog scale (VAS) rating the intensity andunpleasantness of the pain evoked by these stimuli (Gracelyet al., 1978, 1979). These 10-point scales had verbaldescriptors of the intensity of pain appropriately spacedfrom zero (‘‘no pain’’) to 10 (‘‘the most intense pain imagin-able’’). Somatic sensory testing included the followingmechanical stimuli: brushing the skin with a camel-hairbrush, and separate application of a large, medium, orsmall arterial clip to a fold of skin (Chung et al., 1986a;Guilbaud et al., 1987; Surmeier et al., 1988). Mechanicalstimulation also included a nonpenetrating towel clip(110–121, Jarit Surgical Instruments, New York, NY) withtwo parallel 4 3 5 mm serrated surfaces that could beapproximated in 10 reproducible steps by using a ratchet.Approximation of the surfaces of the towel clip has beendescribed in terms of the number of steps taken on theratchet (e.g., F3 indicates three steps). Thermal stimuliwere applied by using a Peltier device (LTS-3, 1-inch head,Thermal Devices, Golden Valley, MN) to deliver a gradedseries of thermal stimuli from a baseline of 33°C.
Study of thalamic activity
The output of the microelectrode preamplifier was fur-ther amplified and displayed on an oscilloscope. As asearch stimulus for neurons that might not be spontane-ously active, manual stroking, tapping, plus painful andnonpainful pinching were carried out on the contralateralface and hand and in the projected field for microstimula-tion at adjacent sites. It was anticipated that this searchtechnique might identify cells without spontaneous activ-ity but no such cells were found. Quantitative testing wascarried out on cells that responded to the search stimuli.Toward the end of each operation, when time was limited,cells responding to pinch were often studied, whereasthose responding to stroking were not. Thus, there was abias to study cells that responded to pinching, eitherpainful or nonpainful. A small number of cells that re-sponded to tapping (n 5 4) were also studied; these cellswere found to be unresponsive to stimuli used in this
Abbreviations
ACPC line anterior commissure-posterior commissure lineBR brushcore region the part of region of Vc where cells respond to Innocuous
cutaneous stimuli stimulus produced by applying threesteps on the ratchet of the nonpenetrating towel
F3 clip/forceps (F4 for four steps, etc.)HT high threshold cellsLC large clipLT low threshold cellsMC medium clipMR multiple receptive cellsPF projected field, the part of the body where a sensation is
evoked by electrical stimulation of the thalamusPo monkey and human thalamic posterior nucleuspostero-
inferiorregion
region of Vc posterior and inferior to the core region (seecore region above)
RF receptive fieldSC small clipSEM standard error of meanSTT spinothalamic tractTMIS threshold electrical microstimulationVAS visual analog scale (of pain intensity)Vc human thalamic ventral caudal nucleus corresponding to
monkey VPVcpor ventral caudal portae, posterior subnucleus of Vc that corre-
sponds to monkey pulvinar oralis nucleusVcpc ventral caudal parvocellular nucleus, inferior subnucleus of
Vc that corresponds to monkey ventral posterior inferiornucleus, VPI
Vim human thalamic nucleus ventralis intermedius (cerebellarrelay nucleus)
VP ventral posterior nucleusVPI ventral posterior inferior nucleusWDR wide dynamic range cells
542 J.-I. LEE ET AL.

study. Another four cells that responded to manual pinch-ing or stroking did not respond significantly to the mechani-cal stimuli used in this study.
When a cell was isolated, spontaneous activity was firstrecorded for a period of 30 to 60 seconds. The center andboundaries of the RF were then defined by manual exami-nation with innocuous stimuli. All stimuli described dur-ing preoperative somatic sensory testing were appliedintraoperatively. The timing of mechanical stimuli wasindicated by a voltage signal from a footpedal, incorporat-ing a switch to regulate the output of a constant voltagesource. The footpedal was depressed when the clip hadbeen attached to the skin and released when the clip wasreleased. The timing of thermal stimuli was indicated bythe output of the thermode on the Peltier (thermal stimuli).The site in the RF where the stimulus was applied variedfrom stimulus to stimulus except for thermal stimuli.Stimuli were applied as close as possible to the center ofthe RF, but placed so as to minimize the overlap betweenthe areas stimulated by the different modalities.
Data collection and analysis
The microelectrode signal, the footpedal signal, thethermode signal from the Peltier, and a voice signaldescribing the type of somatic sensory stimulation wererecorded on magnetic tape (Model 4000, Vetter Corp.,Rebersberg, PA). Postoperatively, the spike train wasdigitized and stored on a Pentium-II PC by use of a spikesorting/data acquisition package (Discovery System, Brain-wave Systems, Thornton, CO), which determined times ofoccurrence of discriminated action potentials. Offline, neu-rons were discriminated by the criterion of spike shape asassessed by cluster analysis. The shape of the cluster wasnot altered throughout recordings on an individual cell.Shapes of action potentials discriminated by this methodwere similar at the beginning and end of cellular record-ings (Figs. 2, 4, 6). The program was configured to producea postspike interval of 1.5 msec, during which the programwould not discriminate a second spike. The period duringwhich a second spike could not be discriminated wasjudged to be appropriate given the maximal firing rates ofcells in the region of Vc (Poggio and Mountcastle, 1963;Lenz et al., 1994c, 1998e). The foot pedal signal, thethermode output from the Peltier system, and EMG sig-nals were digitized at 200 Hz.
Cellular classification was based on responses to me-chanical stimuli as characterized by mean firing rateduring stimulation as in the monkey literature (Price andDubner, 1977; Willis, 1985; Chung et al., 1986a, 1986b;Bushnell and Duncan, 1987; Casey and Morrow, 1987) andas illustrated in the results. LT (low threshold) cells weredefined by a significant response to brush but not tocompressive stimuli. The MR (multiple receptive) class ofcells was defined by a significant response to both brushand compressive stimuli, although responses were notgraded into the painful range. The wide dynamic range(WDR) class of cells was defined by a response that wasgraded into the painful range. Cells with a significantresponse to painful but not nonpainful stimuli were classi-fied as high threshold (HT) cells.
RESULTS
The present report describes results in a population of57 cells recorded along 30 trajectories through the tha-
lamic region where cells respond to innocuous stimulationin 24 patients (left side in 15). Patients had movementdisorders including diagnoses of essential tremor (15 pa-tients), intention tremor (2 patients), parkinsonian tremor(5 patients), and dystonia (2 patients). All patients de-scribed the mechanical and thermal stimuli in similarterms to those used by normal subjects (see above andLenz et al., 1993a). The results of preoperative sensorytesting were compared with those in 14 normal subjects todetermine whether the perception of the mechanical andthermal stimuli in this study was the same for thesesubjects as for the patients studied here. All patientsdescribed all cold stimuli as cold, hot stimuli as hot, andmechanical stimuli as mechanical (see next paragraph), asdid the normal subjects. The VAS ratings of high intensitystimuli (forceps F4, F5, F6, medium clip, small clip,45–48°C) for our patients were within two standard devia-tions of the mean for normal subjects. All cells studiedwere in the region where cells responded to innocuoussomatic sensory stimuli (see Fig. 8).
The results of preoperative somatic sensory testing (Fig.1) establish that the series of thermal and mechanicalstimuli included the painful range. On average, the pa-tients described the brush as a natural, nonpainful, sur-face, touch sensation with the sensation moving across theskin (Lenz et al., 1993a). The large arterial clip wasdescribed as an unnatural, nonpainful, pressure sensationinvolving both surface and deep structures; it was painfulin about one-third of patients, on both face and hand. Themedium arterial clip was described as an unnatural,painful, squeezing sensation involving both surface anddeep structures. The small arterial clip was described asan unnatural, painful, squeezing sensation involving bothsurface and deep structures. The nonpenetrating towelclip was usually painful even at the smallest number ofsteps on the ratchet used to approximate the teeth of theclamp (F3); the pain increased monotonically with increas-ing numbers of steps (F4, F5, F6). The Peltier probe at33°C was described as a natural, surface pressure sensa-tion. The Peltier device at cold temperatures was describedas cold sensation involving both surface and deep struc-tures. At 12°C, it was described as nonpainful. The Peltierhead at 48°C was described as a painful, hot sensationinvolving both surface and deep structures. Therefore,both the mechanical and thermal series of stimuli spannedintensities extending into the painful range.
The WDR cells showed a graded response to mechanicalstimuli extending into the painful range. The activity ofthe WDR cell shown in Figure 2 showed a response tostimuli across the thermal series (Fig. 2D) and the mechani-cal series (Fig. 2B, C). The graded response to mechanicalstimuli was seen across the range from nonpainful topainful (Fig. 2B) and within the painful range (Fig. 2C).The increase in firing rate spanned the range of stimuliinto the painful range in the mechanical and thermalseries (Fig. 1). The responses in each series were comparedstatistically with each other and with the baseline firingrate. The response to any stimulus was defined as thefiring rate over the interval during which the footswitchwas depressed (mechanical series) or the temperature wasdifferent from baseline (thermal series). Comparison offiring rates/100 msec bin (P , 0.05, repeated measuresANOVA, STATISTICA 5.1, StatSoft, Tulsa, OK) foundsignificant differences between firing rates evoked by themechanical stimuli and the baseline firing rate. The
NOCICEPTIVE CONVERGENCE IN HUMAN THALAMIC VP 543

increase between BR and LC, LC and MC, and MC and SCwere significant (Student-Newman-Keuls test, P , 0.05).Comparison of firing rates by thermal stimuli and thebackground (firing rate with the Peltier at skin tempera-ture) found that differences between these firing rateswere significant (repeated measures ANOVA, P , 0.05).The increases between 42°C and both 45° and 48°C weresignificant (Student-Newman-Keuls, P , 0.05). The meanfiring rates were not significantly different between 45°Cand 48°C, although the peak firing rate (over a 1-secondinterval) was higher at 48°C than at 45°C (t-test, P , 0.05).Similar statistical analysis identified a total of 15 cells inthe WDR class (see Table 1). Six of these cells had noresponse to thermal stimuli (termed WDR cells), whereasseven cells had a significant response to heat stimuli(repeated measures ANOVA, P , 0.05) (WDR-H), and twocells responded to cold stimuli (WDR-C).
Electrical microstimulation at the recording site wascarried out for 10 cells in the WDR class. Stimulation atthe site of a WDR cell with an RF on the pad of the distalphalanx of the thumb (36022) evoked a painful, deep, hot,vibration sensation in the forehead at 5 µA. A natural,nonpainful, surface and deep touch sensation was evokedin the RF by stimulation at the site of 1 WDR cell (61093-15 µA). For the remainder, nonpainful, tingling or electriccurrent (9 of 10 sites), deep (3 of 10 sites), surface (3 of 10sites), and both surface and deep sensations (3 of 10 sites)were evoked in a PF that overlapped the RF (8 of 10 sites)at currents of 5 (7 of 10 sites) to 25 µA.
Figure 3A shows the individual stimulus-response func-tions normalized to the baseline firing rate for the WDRclass of cells. Averaged values (mean and SEM) across allcells are shown in Figure 3B. An increase in firing rate isseen across the mechanical series of stimuli for both theindividual and averaged data (P , 0.05, repeated mea-sures ANOVA). The BR response was significantly greaterthan baseline for 10 WDR cells. Among the other five cells,there was a significant response to the LC, which wasnonpainful in the RF of three cells and no significantresponse to the LC in two cells (HT cells-described below).The three cells above exhibited a graded response into thepainful range and were classified as WDR cells, althoughthey did not respond to brush. Figure 3C shows theresponse of the seven WDR-H cells to thermal stimuli;each of these cells had a significant increase in firing ratewith temperature (repeated measures ANOVA, P , 0.05).
Two cells in the WDR class did not respond to brush orlarge clip but had a significant response to the mediumclip, which was painful. These cells might be classified asHT (labeled HT in Fig. 3A). One of these cells had asignificant response to cold stimuli and the other had noresponse to thermal stimuli. Electrical microstimulation
Fig. 1. Sensations evoked by somatic stimuli applied to the face(open bars) or hand (filled bars). A: Percentage of patients for whicheach mechanical and thermal stimulus evoked pain on the face andhand. Stimuli as indicated along the horizontal axis. The number ofsteps on the ratchet of the nonpenetrating towel clip used to generatethe compression stimulus is indicated after the letter F, i.e., F3indicates three steps. B: Mean plus 1 SEM of the VAS for application ofthe mechanical and thermal stimuli to the face and hand. Thenumbers along the right half of the horizontal axis indicate tempera-ture (°C) and ST is skin temperature. For abbreviations, see list.
Fig. 2. Activity of a cell (061093) in Vc responding to painfulmechanical and thermal stimuli. A: Location of the cell (arrow)relative to the positions of trajectories, nuclear boundaries, and otherrecorded cells. The ACPC line is indicated by the horizontal line andthe trajectories are shown by the oblique lines (left, anterior; up,dorsal). Nuclear location was approximated from the position of theACPC line. Lateral location of the cell (in millimeters) is indicatedabove each map. Trajectories have been shifted along the ACPC lineuntil the most posterior cell with a cutaneous RF is aligned with theposterior border of Vc. Because cells responding to innocuous sensorystimuli may be located posterior to Vc (Apkarian and Shi, 1994), thismap represents a first approximation of nuclear location and dimen-sions. The locations of cells are indicated by ticks to the right of eachtrajectory. Cells with cutaneous RFs are indicated by long ticks, thosewithout definable RFs by short ticks. Filled circles attached to the longticks indicate that somatic sensory testing was carried out. The scaleis as indicated. The shape of action potentials recorded at thebeginning of the recording on this cell during application of the brush(upper) and at the end of the recording, during a 12°C stimulus(lower). Action potential discrimination was triggered from up-goingstroke of the action potential by using a voltage threshold of 30 µV. TheRF and PF for the natural, surface and deep, nonpainful, tinglingsensation evoked by TMIS at the recording site (threshold – 15 µA) arealso shown. B: Response to the brush, LC, MC, and SC. C: Theresponse of the neuron to progressive increase in pressure appliedwith the nonpenetrating towel clip, indicated by the number of steps.D: Responses to heat stimuli at 42°, 45°, and 48°C. E: Responses tocold stimuli at 12°, 18°, and 24°C. The upper trace in each panel is afootswitch signal indicating the onset and duration of the stimulus inB and C and the thermode signal in D and E. The scales for the axes forall histograms (bin width 100 msec) are indicated in each panel. Forabbreviations, see list.
544 J.-I. LEE ET AL.

Figure 2
NOCICEPTIVE CONVERGENCE IN HUMAN THALAMIC VP 545

at both sites produced a nonpainful tingling sensation in aPF, including the RF for the cell.
Multiple receptive (MR) cells showed a response to bothbrush and compressive stimuli but responses were notgraded into the painful range. Some cells in the MR classalso responded to painful thermal stimuli (MR-H cells, Fig.4). The cell in Figure 4 showed an increase in firing rate tostimuli across the thermal series (Fig. 4D) but not themechanical series (Fig. 4B, C). The responses in eachseries were compared statistically with each other andwith the baseline firing rate.
The firing rates/100-msec bin at baseline and evoked bymechanical stimuli were significantly different (repeatedmeasures ANOVA, P , 0.05). The differences betweenbaseline and BR, LC, MC, and SC were significant (Student-Newman-Keuls test, P , 0.05); the differences between BRand each of LC, MC, and SC were also significant (Student-Newman-Keuls test, P , 0.05). Differences between thethree compressive stimuli were not significant, the defin-ing characteristic of the MR cell type. There were signifi-cant responses to stimulation at each temperature in-crease tested. An ANOVA used to compare thermal stimuliand the background (Peltier at skin temperature) showedsignificant differences between these firing rates (repeatedmeasures ANOVA, P , 0.05). Post hoc testing demon-strated that differences between baseline and each of 42°,45°, and 48°C were significant and that the differencesbetween 42° and 45° and 48°C were significant (Student-Newman-Keuls test, P , 0.05). Similar statistical analysiswas carried out to identify a total of 25 cells included in theMR class (Table 1). Seventeen cells in the MR class had noresponse to thermal stimuli (termed MR cells). Three cellshad a response to thermal stimuli (MR-H, see Fig. 5) andfive cells responded to cold stimuli (MR-C), a cell typedescribed previously (Lenz and Dougherty, 1998a).
Electrical microstimulation was carried out at the record-ing site of 15 MR cells. Stimulation at one site where anMR cell (61063) was recorded produced a natural, nonpain-ful pressure, and cool sensation in the tongue (10 µA),whereas the RF was on the lower lip. For the remainder, anonpainful, tingling or electric current (15 of 15 sites),pressure (1 of 15 sites), deep (4 of 15 sites), surface (2 of 15sites), and both surface and deep sensation (6 of 15 sites)was evoked at currents of 5 (5 of 15 sites) to 25 µA, in a PFthat overlapped the RF (12 of 15 sites).
The stimulus-response function of cells in the MR class,normalized to the baseline firing rate, is shown in Figure 5(individual responses, Fig. 5A; average response, Fig. 5B).The mechanical responses were significantly greater thanbaseline for each cell. For seven cells, LC evoked thelargest response; this stimulus was nonpainful in the RFfor six of these cells. LC was reported as painful in the RF
of one of these cells (362002, filled diamond in Fig. 5A).This cell had a maximal response to large clip but re-sponses to mechanical stimuli were not graded with thestimulus intensity. Brush evoked the largest response for15 cells. One cell (04902) had no response to mechanicalcompression by arterial clips but responded to applicationof a nonpenetrating towel clip and so was classified as MR(indicated by asterisk in Fig. 5A). For the other three cells,the largest response was to either the medium or smallclip. Responses of these cells to compressive stimuli werenot significantly graded with intensity so that the cellswere classified as MR not WDR.
The activity of a low threshold (LT) cell is shown inFigure 6. There was an increase in firing rate of this cell tothe brush stimulus. A comparison of firing rates/100-msecbin between the responses to mechanical stimuli andbaseline found significant differences between spontane-ous firing rates and those evoked with different mechani-cal stimuli (repeated measures ANOVA, P , 0.05). Posthoc testing showed significance only for brush (Student-Newman-Keuls test, P , 0.05) but not the other mechani-cal and thermal stimuli. Therefore, to a significant degree,this cell encoded nonpainful mechanical stimuli.
Similar analysis identified nine LT cells, none of whichhad a significant response to thermal or compressivestimuli (Table 1). The stimulus-response function of LTcells normalized to the baseline firing rate is shown inFigure 7 (individual responses, Fig. 7A; averaged re-sponse, Fig. 7B). The BR responses were significantlygreater than baseline for all cells, whereas the compres-sive stimuli were not significantly greater than baseline.Stimulation at the recording site was carried out for sevenLT cells. Tingling or electric current (six of seven sites),deep (one of seven sites), surface (three of seven sites) wasevoked in a PF that overlapped the RF (five of seven sites)at currents of 5 (three of seven sites) to 30 µA.
Figure 8 shows the spatial distribution of the cellsdescribed above. WDR, MR, and LT cells were widelydistributed throughout the region of Vc. MR-H and WDR-Hcells are concentrated in the posteriorly and inferiorly.MR-C cells (Lenz and Dougherty, 1998a) and perhapsWDR-C are located dorsally and medially. However, dis-criminant function analysis (STATISTICA 5.1, StatSoft,Tulsa, OK) did not show significant clustering of stereotac-tic coordinates by cell type.
DISCUSSION
The region of the human thalamic principal somaticsensory nucleus (Vc) is implicated in pain and temperaturesignaling pathways by anatomic studies (Walker, 1943;Bowsher, 1957; Mehler et al., 1960; Mehler, 1962, 1966),which indicate that the STT terminates in this region. Wehave now studied the activity of 57 cells in this region ofawake patients at the time of stereotactic surgery. Fifteencells studied had a graded response to mechanical stimuliextending into the painful range (WDR class of cells). Themajority of cells in the WDR class responded to thermalstimuli, either cold (WDR-C, two cells) or heat stimuli(WDR-H, seven cells) but not both. Twenty-five cellsstudied (MR - multiple receptive class of cells) respondedto both brushing and compressive stimuli, although theresponses were not graded into the painful range. Threecells in the MR class (MR-H) responded to heat stimuli and
TABLE 1. Summary of Numbers of Cells in Different Classes Respondingto Mechanical and Thermal Stimuli1
Class of cell
Stimuli
TotalNonpainfulmechanical
Painfulmechanical Cold (2C) Heat (2H)
WDR 13 cells 15 2 (WDR-C) 7 (WDR-H) 15MR 25 22 5 (MR-C) 3 (MR-H) 25LT 9 0 0 0 9Other 0 0 0 0 8
1Name of cell type (e.g., WDR-C) within cell class (e.g., WDR) is indicated, whenappropriate. WDR, wide dynamic range; MR, multiple receptive; LT, low threshold.
546 J.-I. LEE ET AL.

five cells (MR-C) responded to cold stimuli (Lenz andDougherty, 1998a). Nine cells studied responded to brush-ing without a response to compressive or thermal stimuli(LT cells). Cells with responses to heat and painful mechani-cal stimuli were distributed throughout the region wherecells responded to innocuous mechanical stimuli. Theseresults demonstrate that cells in the region of the humanthalamic principal somatic sensory nucleus respond tomechanical and thermal stimuli extending into the painfulrange.
Methodologic considerations
In these studies, it was not feasible to use noxious orthermal search stimuli because of time constraints andpatient discomfort. Noxious search stimuli (Kenshalo etal., 1980; Chung et al., 1986a; Apkarian and Shi, 1994)have been used in studies of anesthetized monkeys andthermal search stimuli have been in studies of anesthe-tized (Burton et al., 1970) and nonanesthetized monkeys
(Bushnell et al., 1993). The use of such search stimuli mayaccount for the large number of WDR cells in these reports(Burton et al., 1970; Chung et al., 1986a; Bushnell et al.,1993). Like most of these monkey studies, we preferen-tially studied cells responding to manual pinching. Theabsence of cells responding exclusively to thermal stimuli(Poulos and Benjamin, 1968; Burton et al., 1970; Chung etal., 1986a) in the present study may be due to the searchstimuli used.
Brief stimuli spanning the range of the VAS (0–10, seeMaterials and Methods section and Fig. 1) were used withpreoperative training and intraoperative testing in thepresent study. This approach is unlike studies of awakemonkeys in which intense noxious stimuli were not used(Casey, 1966; Bushnell and Duncan, 1987; Bushnell et al.,1993). The intense stimuli used in the present study mayexplain the large proportion of WDR cells in the region ofVc in this study. The lack of such cells in another humanstudy (Tasker et al., 1997) may be due to the stimuli used,
Fig. 3. Stimulus response functions for cells in the WDR class.A: Firing rate normalized to the baseline firing rate during stimula-tion by brushing, large clip, medium clip, and small clip for differentcells, as indicated by different symbols. The cell in Figure 2 isindicated by the open triangle pointing up. HT indicates high thresh-old cells. B: Mean firing rate 6 SEM plotted for each mechanical
stimulus. C: Thermal response pattern for WDR-H cells (WDR cellsshowing significant response to heat stimuli) normalized to the firingrate with application of the Peltier device at skin temperature.Symbols in C refer to the same cells as the corresponding symbols in A.The heavy line indicates the average response for these cells. Forabbreviations, see list.
NOCICEPTIVE CONVERGENCE IN HUMAN THALAMIC VP 547
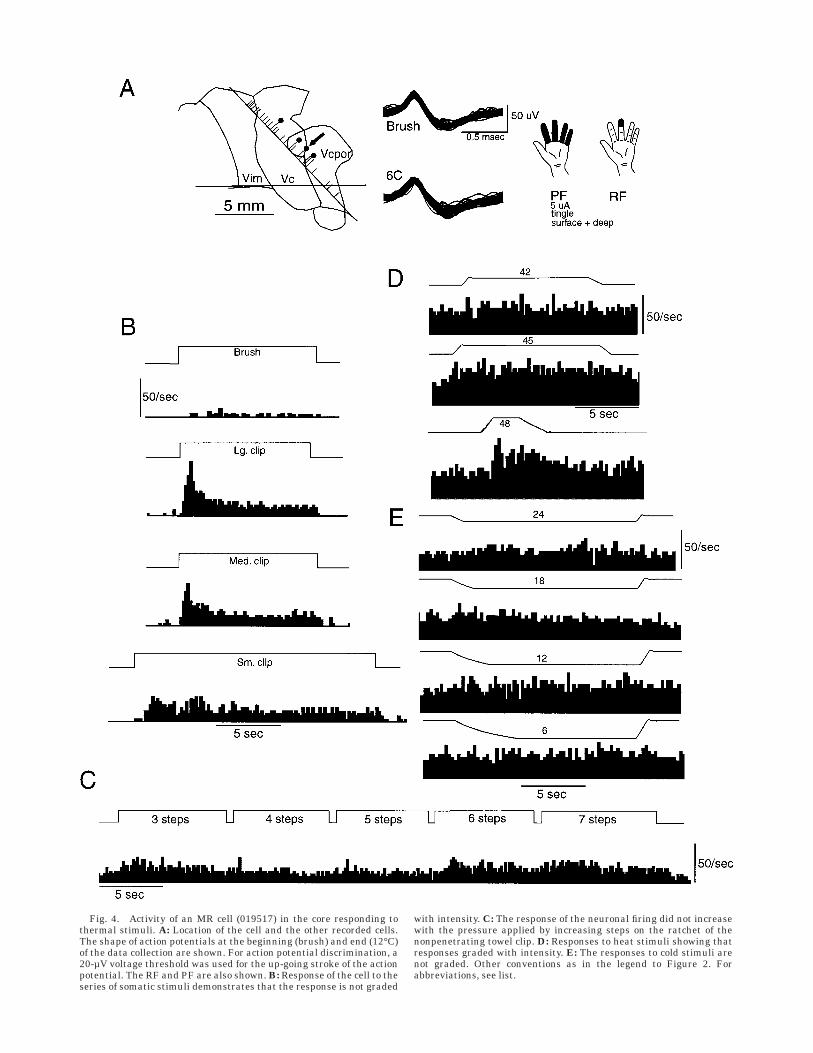
Fig. 4. Activity of an MR cell (019517) in the core responding tothermal stimuli. A: Location of the cell and the other recorded cells.The shape of action potentials at the beginning (brush) and end (12°C)of the data collection are shown. For action potential discrimination, a20-µV voltage threshold was used for the up-going stroke of the actionpotential. The RF and PF are also shown. B: Response of the cell to theseries of somatic stimuli demonstrates that the response is not graded
with intensity. C: The response of the neuronal firing did not increasewith the pressure applied by increasing steps on the ratchet of thenonpenetrating towel clip. D: Responses to heat stimuli showing thatresponses graded with intensity. E: The responses to cold stimuli arenot graded. Other conventions as in the legend to Figure 2. Forabbreviations, see list.

although the methods for that study have not been pub-lished.
In human neurophysiologic studies the location of record-ing sites cannot be determined with the precision expectedin monkey studies. We have mapped anterior-posteriorlocation relative to the ACPC line, a radiologic landmark,and the posterior border of the region where the majorityof cells respond to innocuous somatic sensory stimulation,a physiologic landmark (Fig. 8). In monkeys, cells anatomi-cally verified to be in VPI (Dykes et al., 1981; Apkarian andShi, 1994), pulvinar (Casey, 1966), and posterior nucleus(Po) (Perl and Whitlock, 1961; Apkarian and Shi, 1994)were often responsive to innocuous cutaneous stimuli.These nuclei in monkeys may correspond to the regionwhere cells responded to nonpainful cutaneous stimula-tion in the present report. The maps in Figures 2A, 4A, and
6A show the position of the most posterior cell respondingto innocuous sensory stimuli aligned with the posteriorborder of Vc. Therefore, these maps should be consideredas first approximations to the locations of cells and dimen-sions of nuclei. Because a small number of cells can berecorded in any one operation, the data are pooled frommultiple patients (n 5 24). The relatively large number ofpatients studied adds another source of variability to datarecorded in humans.
Our classification includes MR cells with responses tobrush and compressive stimuli that were not graded intothe painful range. MR cells respond to compressive stimuli,like WDR cells, and would have been included in the WDRclass in some other studies (Chung et al., 1986a,b; Bush-nell et al., 1993). Some of the compressive stimuli used inthis series activate nociceptors even though they do not
Fig. 5. Stimulus response functions for the MR class of cells. A: Thefiring rate normalized to the baseline firing rate during brushing,large clip, medium clip, and small clip for different cells, as indicatedby different symbols. The cell in Figure 4 is indicated by the filledcircle. B: Mean and SEM for the normalized response to each stimulusfor these cells. C: Firing rate of individual MR-H cells (MR cells
showing significant response to heat stimuli) normalized to thebaseline rate during application of the heat stimuli. Symbols in C referto the same cells as the corresponding symbols in A. In the patient withthe cell indicated by the open circle, the 45°C stimulus evoked amaximal VAS score. Therefore, higher temperatures were not tested.For abbreviations, see list.
NOCICEPTIVE CONVERGENCE IN HUMAN THALAMIC VP 549

Fig. 6. A–E: Activity of an LT cell (04905) in the core. Conventionsas for Figure 2. Action potential discrimination was performed withvoltage threshold of 15 µV for the up-going stroke of the actionpotential and with a relatively wide window for spike height. Theresponse of the cell to the series of somatic stimuli demonstrates a
response to brushing but no significant response to any other stimulus(B,C) except for the onset of the compressive stimuli. Responses toheat stimuli (D, 45° and 51°C) and cold stimuli (E, 12°, 18°, and 24°C)were not graded. For abbreviations, see list.

cause pain (Adriaensen et al., 1984). Input from nocicep-tors might be transmitted to the thalamus through thedorsal column pathway because cells with similar proper-ties have been reported in the dorsal column nuclei ofanesthetized primates (Ferrington et al., 1988). Similarresponses have been recorded in the cells of origin of theSTT (Willis et al., 1973). Thus, inputs from nociceptorsmay be transmitted to the thalamus through the dorsalcolumn or the STT.
Comparison of the present resultswith studies in monkeys
The present results are consistent with the resultsreported in awake and anesthetized monkeys. In a study ofresponses of cells in VPM of awake monkeys to gradedmechanical stimuli (Bushnell and Duncan, 1987), 10% ofcells (9 of 89) are classified as WDR. These cells wereclustered in ventral VPM. Another study reports 22 ther-mal responsive cells from a population of hundreds re-corded in alert, trained, cynomolgus monkeys (Bushnell etal., 1993). Eighteen percent (4 of 22) of these cells re-sponded to noxious heat only. No such cells were found inthe present analysis, perhaps because of the search stimuliused. Among cells responding to thermal stimuli thosethat also displayed a WDR response pattern composed27% of cells (6 of 22) in that series and 53% of cells (9 of 17)in the present series.
The human region of Vc where cells respond to innocu-ous mechanical stimuli (Fig. 8) may correspond to monkeyVP, VPI, pulvinar, and Po because cells in these nuclei alsorespond to innocuous cutaneous stimuli (Perl and Whit-lock, 1961; Dykes et al., 1981; Apkarian and Shi, 1994). Ina recent study, 40 cells in these nuclei responded tonoxious mechanical stimuli; of these, 23 cells also re-sponded to noxious heat and 9 responded to noxious cold(Apkarian and Shi, 1994). These cells were located in VPIand Po more commonly than in VP, consistent with earlierstudies (Casey, 1966; Perl and Whitlock, 1961). Studies inawake squirrel monkeys have found that 8% (3 of 36) to12% (9 of 76) of cells in VP responded to noxious mechani-cal stimuli (Casey, 1966; Casey and Morrow, 1983). Thesecells were widely distributed throughout VP. In anotherstudy, a smaller number of WDR and HT cells were foundthroughout VP in anesthetized rhesus monkeys (73 cells/thousands of cells) (Kenshalo et al., 1980). Overall, mon-key studies suggest that cells responsive to noxious stimuliare located in nuclei where cells respond to innocuousstimuli - VP, VPI, pulvinar, and Po (Perl and Whitlock,1961; Casey, 1966; Casey and Morrow, 1983; Apkarian andShi, 1994). Although we do not know the anatomic locationof recorded neurons, the present results demonstrate thatcells responsive to painful mechanical stimuli are locatedin nuclei where cells respond to nonpainful cutaneousstimuli.
Fig. 7. Stimulus response functions for LT cells. The cell illustrated in Figure 6 is indicated by theopen square. Conventions as for Figure 3. For abbreviations, see list.
NOCICEPTIVE CONVERGENCE IN HUMAN THALAMIC VP 551

Pathways transmitting nociceptive inputto the region of Vc
Studies of patients at autopsy after lesions of the STTshow that the human STT ascends to the thalamus medialto the medial geniculate (Walker, 1943; Bowsher, 1957;Mehler et al., 1960; Mehler, 1962, 1966) before terminat-ing in the magnocellular medial geniculate (Mehler, 1962,1969), limitans, and Vc portae nuclei, posterior to Vc(Mehler, 1966). More anteriorly the STT makes its mostdense termination as irregular clusters in Vc (Walker,1943; Bowsher, 1957; Mehler et al., 1960; Mehler, 1962,1966). The STT terminations are concentrated in posteriorinferior Vc and in dorsal Vc parvocellularis (Mehler, 1966).
Similarly, in monkeys, STT terminals occur as denseclusters in VPL (Mehler et al., 1960; Boivie, 1979; Berkley,1980; Mantyh, 1983; Apkarian and Hodge, 1989). A moreuniform, less-dense termination is found in ventral poste-rior inferior - VPI (Apkarian and Hodge, 1989) correspond-ing to human Vc parvocellularis (Hirai and Jones, 1989)and in the posterior nuclear group including posteriornucleus (Boivie, 1979; Berkley, 1980; Burton and Craig,1983; Mantyh, 1983; Ralston and Ralston, 1992), pulvinaroralis (Apkarian and Hodge, 1989), limitans (Apkarianand Hodge, 1989; Mantyh, 1983), magnocellular medialgeniculate (Berkley, 1980), suprageniculate nuclei (Berk-ley, 1980; Mantyh, 1983; Apkarian and Hodge, 1989;
Fig. 8. Locations of different cell types. Cellular location is shownrelative to the most posterior cell with an RF (vertical line) and theACPC (horizontal) line for planes 14 mm or less from the midline (B)
and more than 14 mm from the midline (A). WDR-H and -C; MR-H and-C indicate WDR and MR cells with significant response to hear or coldstimuli. For abbreviations, see list.
552 J.-I. LEE ET AL.

Ralston and Ralston, 1992), and in the posterior division ofthe ventral medial nucleus (Craig et al., 1994). Thus, thereis ample evidence of inputs from the STT to the region ofVc that could explain the occurrence of cellular responsesto noxious and thermal stimuli.
Function of cells responding to painfulmechanical and thermal stimuli
Microstimulation studies suggest that there is partition-ing of thermal/pain sensations at different locations in theregion of Vc. Stimulation sites where thermal/pain sensa-tions are evoked are located near the posterior border ofthe core and within the posterior inferior region (Lenz etal., 1993a). Microstimulation in the posteroinferior regionevokes thermal sensations or pain often referred to largeRFs and subcutaneous structures. Other reports identifysites where pain but not thermal sensations are evokedposterior and inferior to the core (Hassler and Reichert,1959; Halliday and Logue, 1972; Dostrovsky et al., 1991).These stimulation results demonstrate that the posteriorpart of the core area and the posteroinferior area areinvolved in signaling thermal sensations and pain. Thus,the location where cells responded to heat stimuli (Fig. 8)apparently overlapped with the location where stimula-tion evoked pain. In a previous study, pain was evokedsignificantly more commonly by stimulation at sites wherecells responding to noxious thermal stimuli were recorded(Lenz et al., 1993b).
The graded response of WDR-H cells to both mechanicaland thermal stimuli strongly suggests that these cellsencode pain for these two modalities. However, except fortwo cells that might be classified as HT cells, cells in theWDR class always responded to nonpainful stimuli. Thefunction of such cells is less clear than that of cells thatrespond to noxious stimuli alone (Craig et al., 1994).Microstimulation-evoked sensations provide one indicatorof the modality signaled by the population of cells at aparticular location (Ranck, 1975). In the present results,the sensations evoked by microstimulation at the siteswhere these cells were recorded did not correlate with thesensations produced by the stimuli that evoked cellularactivity.
The mechanism by which cells with convergent inputs(Casey, 1966; Kenshalo et al., 1980; Casey and Morrow,1983; Bushnell and Duncan, 1987; Bushnell et al., 1993;Apkarian and Shi, 1994) might signal specific modalities ofsensation is uncertain. One such mechanism may involvegating by the combined activity of multiple neuronalchannels (LeBars and Chitour, 1983). Thus, application ofnoxious mechanical stimuli activates cells in the principalsomatic sensory nucleus of the thalamus, medial thala-mus, and posterior group (Lenz and Dougherty, 1997).Inputs from these nuclei may explain pain-related activa-tion of the contralateral anterior cingulate gyrus (Jones etal., 1991; Talbot et al., 1991; Casey et al., 1994; Lenz et al.,1998c), ipsilateral cingulate gyrus (Vogt et al., 1996; Lenzet al., 1998c), primary and secondary somatic sensoryareas, insular cortex (Jones et al., 1991; Casey et al., 1994;Lenz et al., 1998b), and supplementary motor area (Talbotet al., 1991; Casey et al., 1994). Thus, the perception ofpain may involve simultaneous activation of multiplecortical structures through populations of thalamic cellswith convergent input, including responses to painfulstimuli.
The principal sensory nucleus receives substantial inputfrom the medial lemniscus, which sends a dense projectionto Vc and signals innocuous mechanical sensations (Berk-ley, 1980; Jones, 1985). It is possible that the sensationevoked by stimulation in the region of Vc is dominated bythe activation of medial lemniscal fibers, which are morenumerous in Vc than STT fibers (Jones, 1985). STT inputmay allow cells in Vc to signal pain and temperaturesensations or may provide them with the potential tosignal these sensations. This actual or potential functionmay be activated or transformed in different circum-stances, like injuries to the nervous system. For example,the sensation of pain is evoked by stimulation in the coreregion of patients with central pain after strokes morecommonly than in patients operated on for the treatmentof movement disorders (Davis et al., 1996). The increase inthe number of sites where pain is evoked is equal to thedecrease in sites where heat and cool sensations areevoked (Lenz et al., 1998a). This reciprocal relation be-tween pain and thermal sensation could be explained by atransformation in the function of cells that respond topainful mechanical and thermal sensations.
ACKNOWLEDGMENTS
We thank L. Rowland for expert technical assistance.F.A.L. received support from the National Institutes ofHealth.
LITERATURE CITED
Adriaensen H, Gybels J, Handwerker HO, Van Hees J. 1984. Nociceptordischarges and sensations due to prolonged noxious mechanical stimu-lation. Hum Neurobiol 3:53–58.
Apkarian AV, Hodge CJ. 1989. Primate spinothalamic pathways: III.Thalamic terminations of the dorsolateral and ventral spinothalamicpathways. J Comp Neurol 288:493–511.
Apkarian AV, Shi T. 1994. Squirrel monkey lateral thalamus I Somaticnociresponsive neurons and their relation to spinothalamic terminals. JNeurosci 14:6779–6795.
Berkley KJ. 1980. Spatial relationships between the terminations ofsomatic sensory and motor pathways in the rostral brainstem of catsand monkeys. I. Ascending somatic sensory inputs to lateral diencepha-lon. J Comp Neurol 193:283–317.
Boivie J. 1979. An anatomic reinvestigation of the termination of thespinothalamic tract in the monkey. J Comp Neurol 168:343–370.
Bowsher D. 1957. Termination of the central pain pathway in man: theconscious appreciation of pain. Brain 80:606–620.
Burton H, Craig AD Jr. 1983. Spinothalamic projections in cat, raccoon andmonkey: a study based on anterograde transport of horseradish peroxi-dase. In: Macchi G, editor. Somatosensory integration in the thalamus.Amsterdam: Elsevier. p 17–41.
Burton H, Forbes DJ, Benjamin RM. 1970. Thalamic neurons responsive totemperature changes of glabrous hand and foot skin in squirrel monkey.Brain Res 24:179–190.
Bushnell MC, Duncan GH. 1987. Mechanical response properties ofventroposterior medial thalamic neurons in the alert monkey. ExpBrain Res 67:603–614.
Bushnell MC, Duncan GH, Tremblay N. 1993. Thalamic VPM nucleus inthe behaving monkey. I. Multimodal and discriminative properties ofthermosensitive neurons. J Neurophysiol 69:739–752.
Casey KL. 1966. Unit analysis of nociceptive mechanisms in the thalamusof the awake squirrel monkey. J Neurophysiol 29:727–750.
Casey KL, Morrow TJ. 1983. Ventral posterior thalamic neurons differen-tially responsive to noxious stimulation of the awake monkey. Science221:675–677.
Casey KL, Morrow TJ. 1987. Nociceptive neurons in the ventral posteriorthalamus of the awake squirrel monkey: observations on identification,modulation, and drug effects. In: Besson JM, Guilbaud G, PeschanskiM, editors. Thalamus and pain. New York, Amsterdam, Oxford: ElsevierScience Publishers BV. p 211–257.
NOCICEPTIVE CONVERGENCE IN HUMAN THALAMIC VP 553

Casey KL, Minoshima S, Berger KL, Koeppe RA, Morrow TJ, Frey KA.1994. Positron emission tomographic analysis of cerebral structuresactivated specifically by repetitive noxious heat stimuli. J Neurophysiol71:802–807.
Chung JM, Lee KH, Surmeier DJ, Sorkin LS, Kim I, Willis WD. 1986a.Response characteristics of neurons in the ventral posterior lateralnucleus of the monkey thalamus. J Neurophysiol 56:370–390.
Chung JM, Surmeier DJ, Lee KH, Sorkin LS, Honda N, Tsong Y, Willis WD.1986b. Classification of primate spinothalamic and somatosensorythalamic neurons based on cluster analysis. J Neurophysiol 56:308–327.
Craig AD, Bushnell MC, Zhang ET, Blomqvist A. 1994. A specific thalamicnucleus for pain and temperature sensation in macaques and humans.Nature 372:770–773.
Davis KD, Kiss ZHT, Tasker RR, Dostrovsky JO. 1996. Thalamic stimula-tion-evoked sensations in chronic pain patients and nonpain (move-ment disorder) patients. J Neurophysiol 75:1026–1037.
Dostrovsky JO, Wells FEB, Tasker RR. 1991. Pain evoked by stimulation inhuman thalamus. In: Sjigenaga Y, editor. International symposium onprocessing nociceptive information. Amsterdam: Elsevier .
Duncan GH, Bushnell MC, Oliveras JL, Bastrash N, Tremblay N. 1993.Thalamic VPM nucleus in the behaving monkey. III. Effects of revers-ible inactivation by lidocaine on thermal and mechanical discrimina-tion. J Neurophysiol 70:2086–2096.
Dykes RW, Sur M, Merzenich MM, Kaas JH, Nelson RJ. 1981. Regionalsegregation of neurons responding to quickly adapting, slowly adapt-ing, deep and pacinian receptors within thalamic ventroposteriorlateral and ventroposterior inferior nuclei in the squirrel monkey.Neuroscience 6:1687–1692.
Ferrington DG, Downie JW, Willis WD Jr. 1988. Primate nucleus gracilisneurons: responses to innocuous and noxious stimuli. J Neurophysiol59:886–907.
Fruhstorfer H, Lindblom U. 1984. Sensibility abnormalities in neuralgicpatients studied by thermal and tactile pulse stimulation. In: von EulerC, editor. Somatosensory mechanisms. London: MacMillian. p 353–361.
Gracely RH, McGrath P, Dubner R. 1978. Validity and sensitivity of ratioscales of sensory and affective verbal pain descriptors: manipulation ofaffect by Diazepam. Pain 5:19–29.
Gracely RH, Dubner R, McGrath PA. 1979. Narcotic analgesia: fentanylreduces the intensity but not the unpleasantness of painful tooth pulpstimulation. Science 203:1261–1263.
Guilbaud G, Neil A, Benoist JM, Kayser V, Gautron M. 1987. Thresholdsand encoding of neuronal responses to mechanical stimuli in theventro-basal thalamus during carrageenin-induced hyperalgesic inflam-mation in the rat. Exp Brain Res 68:311–318.
Halliday AM, Logue V. 1972. Painful sensations evoked by electricalstimulation in the thalamus. In: Somjen GG, editor. Neurophysiologystudied in man. Amsterdam: Excerpta Medica. p 221–230.
Hassler R, Reichert T. 1959. Klinische und anatomische Befunde beistereotaktischen Schmerzoperationen im Thalamus. Arch PsychiatrNervenkr 200:93–122.
Hawrylyshyn P, Rowe IH, Tasker RR, Organ LW. 1976. A computer systemfor stereotaxic neurosurgery. Comput Biol Med 6:87–97.
Hirai T, Jones EG. 1989. A new parcellation of the human thalamus on thebasis of histochemical staining. Brain Res Rev 14:1–34.
Jones EG. 1985. The thalamus. New York: Plenum.Jones AKP, Brown WD, Friston KJ, Qi LY, Frackowiak RSJ. 1991. Cortical
and subcortical localization of response to pain in man using positronemission tomography. Proc R Soc Lond Biol 244:39–44.
Kenshalo DR, Giesler GJ, Leonard RB, Willis WD. 1980. Responses ofneurons in primate ventral posterior lateral nucleus to noxious stimuli.J Neurophysiol 43:1594–1614.
LeBars D, Chitour D. 1983. Do convergent neurons in the spinal dorsalhorn discriminate nociceptive from non-nociceptive information? Pain17:1–19.
Lehmann JF, de Lateur BJ. 1994. Ultrasound, shortwave, microwave, laser,superficial heat and cold in the treatment of pain. In: Wall PD, MelzackR, editors. Textbook of pain. New York: Churchill Livingstone. p1237–1250.
Lenz FA, Dougherty PM. 1997. Pain processing in the human thalamus. In:Steriade M, Jones EG, McCormick DA, editors. Thalamus: Vol II.Oxford: Elsevier. p 617–651.
Lenz FA, Dougherty PM. 1998. Cells in the human principal thalamicsensory nucleus (Ventralis Caudalis - Vc) respond to innocuous mechani-cal and cool stimuli. J Neurophysiol 79:2227–2230.
Lenz FA, Dostrovsky JO, Kwan HC, Tasker RR, Yamashiro K, Murphy JT.1988a. Methods for microstimulation and recording of single neuronsand evoked potentials in the human central nervous system. J Neuro-surg 68:630–634.
Lenz FA, Tasker RR, Kwan HC, Schnider S, Kwong R, Murayama Y,Dostrovsky JO, Murphy JT. 1988b. Single unit analysis of the humanventral thalamic nuclear group: correlation of thalamic ‘‘tremor cells’’with the 3–6 Hz component of parkinsonian tremor. J Neurosci8:754–764.
Lenz, FA, M Seike, YC Lin, FH Baker, RT Richardson, and RH Gracely.1993a. Thermal and pain sensations evoked by microstimulation in thearea of the human ventrocaudal nucleus. Vc. J Neurophysiol 70:200–212.
Lenz, FA, M Seike, YC Lin, FH Baker, LH Rowland, RH Gracely, and RTRichardson. 1993b. Neurons in the area of human thalamic nucleusventralis caudalis respond to painful heat stimuli Brain Res 623:235–240.
Lenz FA, Gracely RH, Hope EJ, Baker FH, Rowland LH, Dougherty PM,Richardson RT. 1994a. The sensation of angina can be evoked bystimulation of the human thalamus. Pain 59:119–125.
Lenz FA, Gracely RH, Rowland LH, Dougherty PM. 1994b. A population ofcells in the human principal sensory nucleus respond to painfulmechanical stimuli. Neurosci Lett 180:46–50.
Lenz FA, Kwan HC, Martin R, Tasker R, Richardson RT, Dostrovsky JO.1994c. Characteristics of somatotopic organization and spontaneousneuronal activity in the region of the thalamic principal sensorynucleus in patients with spinal cord transection. J Neurophysiol72:1570–1587.
Lenz FA, Gracely RH, Baker FH, Richardson RT, Dougherty PM. 1998a.Reorganization of sensory modalities evoked by stimulation in theregion of the principal sensory nucleus (ventral caudal - Vc) in patientswith pain secondary to neural injury. J Comp Neurol 399:125–138.
Lenz FA, Rios MR, Chau D, Krauss G, Zirh TA, Lesser RP. 1998b. Painfulstimuli evoke potentials recorded over the parasylvian cortex in hu-mans. J Neurophysiol 80:2077–2088.
Lenz FA, Rios MR, Zirh TA, Krauss G, Lesser RP. 1998c. Painful stimulievoke potentials recorded over the human anterior cingulate gyrus. JNeurophysiol 79:2231–2234.
Lenz FA, Zirh AT, Garonzik IM, Dougherty PM. 1998d. Neuronal activity inthe region of the principal sensory nucleus of human thalamus (ventra-lis caudalis) in patients with pain following amputations. Neuroscience86:1065–1081.
Mantyh PW. 1983. The spinothalamic tract in primate: a re-examinationusing wheat germ agglutinin conjugated with horseradish peroxidase.Neuroscience 9:847–862.
Mehler WR. 1962. The anatomy of the so-called ‘‘pain tract’’ in man: ananalysis of the course and distribution of the ascending fibers of thefasciculus anterolateralis. In: French JD, Porter RW, editors. Basicresearch in paraplegia. Springfield: Thomas. p 26–55.
Mehler WR. 1966. The posterior thalamic region in man. Confin Neurol27:18–29.
Mehler WR. 1969. Some neurological species differences: a posteriori. AnnNY Acad Sci 167:424–468.
Mehler WR, Feferman ME, Nauta WHJ. 1960. Ascending axon degenera-tion following anterolateral cordotomy. An experimental study in themonkey. Brain 83:718–750.
Morrow TJ, Casey KL. 1992. State-related modulation of thalamic somato-sensory responses in the awake monkey. J Neurophysiol 67:305–317.
Perl ER, Whitlock DG. 1961. Somatic stimuli exciting spinothalamicprojections to thalamic neurons in cat and monkey. Exp Neurol3:256–296.
Poggio GF, Mountcastle VB. 1963. The functional properties of ventrobasalthalamic neurons studied in ananesthetized monkeys. J Neurophysiol26:775–806.
Poulos DA Benjamin RM. 1968. Response of thalamic neurons to thermalstimulation of the tongue. J Neurophysiol 31:28–43.
Price DD, Dubner R. 1977. Neurons that subserve the sensory-discrimina-tive aspects of pain. Pain 3:307–338.
Ralston HJ, Ralston DD. 1992. The primate dorsal spinothalamic tract:evidence for a specific termination in the posterior nuclei [Po/SG] of thethalamus. Pain 48:107–118.
Ralston HJ, Ralston DD. 1994. Medial lemniscal and spinal projections tothe Macaque thalamus: an electron microscopic study of differing
554 J.-I. LEE ET AL.

GABAergic circuitry serving thalamic somatosensory mechanisms. JNeurosci 14:2485–2502.
Ranck JB. 1975. Which elements are excited in electrical stimulation ofmammalian central nervous system: a review. Brain Res 98:417–440.
Schaltenbrand G, Bailey P. 1959. Introduction to stereotaxis with an atlasof the human brain. Stuttgart: Thieme.
Surmeier DG, Honda CN, Willis WD. 1988. Natural groupings of primatespinothalamic neurons based on cutaneous stimulation. J Neurophysiol59:833–860.
Talbot JD, Marrett S, Evans AC, Meyer E, Bushnell MC, Duncan GH.1991.Multiple representations of pain in human cerebral cortex. Sci-ence 251:1355–1358.
Tasker RR, Davis KD, Hutchinson WD, Dostrovsky JO. 1997. Subcorticaland thalamic mapping in functional neurosurgery. In Gildenberg PL,
Tasker RR, editors. Stereotactic and functional neurosurgery. NewYork: McGraw-Hill. p 883–923.
Torgerson WS, BenDebba M, Mason KJ. 1988. Varieties of pain. In: DubnerR, Gebhart GF, Bond MR, editors. Proceedings of the 5th WorldCongress on Pain. Amsterdam: Elsevier. p 368–374.
Vogt BA, Derbyshire S, Jones AKP. 1996. Pain processing in four regions ofhuman cingulate cortex localized with co-registered PET and MRI. EurJ Neurosci 8:1461–1473.
Walker AE. 1943. Central representation of pain. Res Publ Assoc Res NervMent Dis 23:63–85.
Willis WD. 1985. The pain system. Basel: Karger.Willis WD, Trevino DL, Coulter JD, Maunz RA. 1973. Responses of primate
spinothalamic tract neurons to natural stimulation of hindlimb. JNeurophysiol 37:358–372.
NOCICEPTIVE CONVERGENCE IN HUMAN THALAMIC VP 555