
RESEARCH ARTICLE Open Access Drug ... - BMC Microbiology
15
RESEARCH ARTICLE Open Access Drug-resistant and hospital-associated Enterococcus faecium from wastewater, riverine estuary and anthropogenically impacted marine catchment basin Ewa Sadowy 1 and Aneta Luczkiewicz 2* Abstract Background: Enterococci, ubiquitous colonizers of humans and other animals, play an increasingly important role in health-care associated infections (HAIs). It is believed that the recent evolution of two clinically relevant species, Enterococcus faecalis and Enterococcus faecium occurred in a big part in a hospital environment, leading to formation of high-risk enterococcal clonal complexes (HiRECCs), which combine multidrug resistance with increased pathogenicity and epidemicity. The aim of this study was to establish the species composition in wastewater, its marine recipient as well as a river estuary and to investigate the antimicrobial susceptibility of collected isolates. Molecular methods were additionally applied to test the presence of HiRRECC-related E. faecium. Results: Two wastewater treatment plants (WWTPs), their marine outfalls and Vistula river that influence significantly the quality of waters in Gulf of Gdansk were sampled to investigate the presence of Enterococcus spp. Four-hundred-twenty-eight isolates were obtained, including E. faecium (244 isolates, 57.0%), E. hirae (113 isolates, 26.4%) and E. faecalis (63 isolates, 14.7%); other species (E. gallinarum/casseliflavus, E. durans and E. avium) accounted for 1.9%. Antimicrobial susceptibility testing revealed the presence of isolates resistant to erythromycin, tetracycline, amipicillin, fluoroquinolones and aminoglycosides (high-level resistance), especially among E. faecium, where such isolates were usually characterized by multilocus sequence types associated with nosocomial lineages 17, 18 and 78 of this species representing HiRECC, formerly called CC17. These isolates not only carried several resistance determinants but were also enriched in genes encoding pathogenicity factors (Esp, pili) and genes associated with mobile genetic elements (MGE), a feature also typical for nosocomial HiRECC. Conclusions: Our data show that WWTPs constitute an important source of enterococcal strains carrying antimicrobial resistance determinants, often associated with the presence of MGE, for the recipient water environment, thus increasing a pool of such genes for other organisms. The presence of HiRECCs in wastewaters and marine/river environment may indicate that adaptations gained in hospitals may be also beneficial for survival of such clones in other settings. There is an obvious need to monitor the release and spread of such strains in order to elucidate better ways to curb their dissemination. Keywords: WWTP, Marine waters, Enterococcus, Resistance, HiRECC, Plasmid, Replicon type, Toxin-antitoxin system, Esp, Pili * Correspondence: [email protected] 2 Gdansk University of Technology, ul. G. Narutowicza 11/12, Gdansk 80-952, Poland Full list of author information is available at the end of the article © 2014 Sadowy and Luczkiewicz; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. Sadowy and Luczkiewicz BMC Microbiology 2014, 14:66 http://www.biomedcentral.com/1471-2180/14/66
Transcript of RESEARCH ARTICLE Open Access Drug ... - BMC Microbiology

Sadowy and Luczkiewicz BMC Microbiology 2014 1466httpwwwbiomedcentralcom1471-21801466
RESEARCH ARTICLE Open Access
Drug-resistant and hospital-associatedEnterococcus faecium from wastewater riverineestuary and anthropogenically impacted marinecatchment basinEwa Sadowy1 and Aneta Luczkiewicz2
Abstract
Background Enterococci ubiquitous colonizers of humans and other animals play an increasingly important rolein health-care associated infections (HAIs) It is believed that the recent evolution of two clinically relevant speciesEnterococcus faecalis and Enterococcus faecium occurred in a big part in a hospital environment leading toformation of high-risk enterococcal clonal complexes (HiRECCs) which combine multidrug resistance with increasedpathogenicity and epidemicity The aim of this study was to establish the species composition in wastewater itsmarine recipient as well as a river estuary and to investigate the antimicrobial susceptibility of collected isolatesMolecular methods were additionally applied to test the presence of HiRRECC-related E faecium
Results Two wastewater treatment plants (WWTPs) their marine outfalls and Vistula river that influencesignificantly the quality of waters in Gulf of Gdansk were sampled to investigate the presence of Enterococcus sppFour-hundred-twenty-eight isolates were obtained including E faecium (244 isolates 570) E hirae (113 isolates264) and E faecalis (63 isolates 147) other species (E gallinarumcasseliflavus E durans and E avium) accountedfor 19 Antimicrobial susceptibility testing revealed the presence of isolates resistant to erythromycin tetracyclineamipicillin fluoroquinolones and aminoglycosides (high-level resistance) especially among E faecium where suchisolates were usually characterized by multilocus sequence types associated with nosocomial lineages 17 18 and78 of this species representing HiRECC formerly called CC17 These isolates not only carried several resistancedeterminants but were also enriched in genes encoding pathogenicity factors (Esp pili) and genes associated withmobile genetic elements (MGE) a feature also typical for nosocomial HiRECC
Conclusions Our data show that WWTPs constitute an important source of enterococcal strains carryingantimicrobial resistance determinants often associated with the presence of MGE for the recipient waterenvironment thus increasing a pool of such genes for other organisms The presence of HiRECCs in wastewatersand marineriver environment may indicate that adaptations gained in hospitals may be also beneficial for survivalof such clones in other settings There is an obvious need to monitor the release and spread of such strains inorder to elucidate better ways to curb their dissemination
Keywords WWTP Marine waters Enterococcus Resistance HiRECC Plasmid Replicon type Toxin-antitoxin systemEsp Pili
Correspondence ansobpggdapl2Gdansk University of Technology ul G Narutowicza 1112 Gdansk 80-952PolandFull list of author information is available at the end of the article
copy 2014 Sadowy and Luczkiewicz licensee BioMed Central Ltd This is an Open Access article distributed under the terms ofthe Creative Commons Attribution License (httpcreativecommonsorglicensesby20) which permits unrestricted usedistribution and reproduction in any medium provided the original work is properly credited The Creative Commons PublicDomain Dedication waiver (httpcreativecommonsorgpublicdomainzero10) applies to the data made available in thisarticle unless otherwise stated
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 2 of 15httpwwwbiomedcentralcom1471-21801466
BackgroundEnterococci are commensal bacteria of intestinal tractof mammals and birds but these bacteria can also befound in the gut of reptiles and insects and in differentenvironmental compartments like soil and plants infood (both as starters and contamination) as well as inaquatic ecosystems both distributed with treated waste-water and lacking such anthropogenic impact [1] In thehuman intestine the Enterococcus faecalis and Entero-coccus faecium are most abundant and these twospecies are also most commonly isolated from humanenterococcal infections however other species such asEnterococcus avium Enterococcus casseliflavus Entero-coccus durans Enterococcus gallinarum and Entero-coccus raffinosus have also been detected [23] RecentlyE faecalis and E faecium play more and more import-ant role in hospital-acquired infections (HAIs) such asbacteremia endocarditis infections of urinary tract andpost-surgery wounds [4] and nowadays represent a sec-ond etiologic agent of hospital bloodstream infectionsin Europe [5] In the recent years the relative propor-tion of E faecium to E faecalis is increasing in the USand Europe [6-8] which is likely due to the spread of aparticular hospital-adapted polyclonal high-risk entero-coccal complex (HiRECC) of E faecium initially describedas clonal complex 17 CC17 [8] This nosocomial subpop-ulation consists of three main lineages named from thecentral sequence type (ST) defined by multilocus se-quence typing (MLST) as lineages 17 18 and 78 [9] Theacquisition of resistance to antimicrobials of several clas-ses constitutes an important feature of hospital-associatedE faecium what not only renders them resilient to ther-apy but also creates a reservoir of antimicrobial resistancegenes often associated with mobile genetic elements(MGE) such as conjugative transposons and plasmids[10] The most important from the clinical point of viewexamples of acquired resistance among enterococci in-clude resistance to (amino)penicillins fluoroquinolonesglycopeptides (vancomycin-resistant enterococci VRE)and high-level resistance to aminoglycosides (HLAR)[3] Resistance to ampicillin associated with the changesin penicillin binding protein 5 (PBP5) and resistance toquinolones determined by point mutations in genes en-coding bacterial gyrase and topoisomerase IV are char-acteristic phenotypic features of nosocomial E faecium[1112] Such strains also commonly carry resistancegenes to other classes of antimicrobials such as tetracy-clines tet(M) tet(L) tet(S) and tet(O) macrolides erm(B) and aminoglycosides [13] The HLAR phenotype isusually determined by the aac(6prime)-Ie-aph(2Prime) gene en-coding a so-called bi-functional enzyme responsible forresistance to all aminoglycosides except for streptomycinhigh-level resistance to this compound is typically speci-fied by ant(6prime)-Ia [3] Hospital-associated E faecium is
also enriched in several virulence factors such as en-terococcal surface protein (Esp) and MSCRAMM pro-teins [14-16]Several studies have addressed the spread of HiRECCs
especially VRE within as well as among health-care facil-ities [17] Much less is known however about the releaseof drug-resistant enterococci and HiRECCs into the envir-onment and their subsequent survival Since it is sus-pected that human-associated bacteria can be regarded asvectors of gene transmission into environmental popula-tions [18] and positive selection of bacteria resistant toantimicrobial agents was observed in wastewater processes[1920] the potential role of effluents from wastewatertreatment plant (WWTP) in dissemination of clinicallyrelevant bacteria and genes needs evaluation It is particu-larly important in ecosystems subjected to strong an-thropogenic impact such as the Gulf of Gdansk and itsshallow western part the Puck Bay Significant pollutionload is discharged to this coastal area through the numer-ous local rivers and marine outfalls Due to limited waterexchange in the Gulf of Gdansk safe wastewater disposalis essential to prevent the environmental degradation andto preserve the public health Epidemiological studiesreported a direct relationship between the risk of gastro-enteritis among swimmers and density of enterococci insurface waters [21] Additionally resistant and multiresis-tant enterococci were detected in effluent of WWTPs[182022] suggesting their survival capacity in treatmentprocesses However no microbiological standards havebeen set on WWTP effluents in the majority of Europeancountries and it is required instead to monitor the qualityof recreational water [23] The US Environmental Protec-tion Agency [21] as well as the European Union legislation[23] recommended detection of fecal enterococci togetherwith Escherichia coli as indicators of fecal contaminationof bathing waterAim of the current study was to investigate the pres-
ence and species composition of enterococci in treatedwastewater river estuary water and their recipienthighly anthropogenically impacted Gulf of Gdansk Forthis purpose enterococci were isolated from wastewaterof two local WWTPs Gdansk-Wschod (influent - W-INFeffluent - W-EFF and bioreactor - W-BR) and Gdynia-Debogorze (effluent - D-EFF) as well as from their marineoutfalls (W-MOut and D-MOut respectively) Add-itionally the Vistula River mouth was sampled due toecological importance of this river flows for the Gulfof Gdansk area (Figure 1) Identification and furthercharacterization of drug-resistant enterococci especiallythe most prevalent E faecium including its resistancevirulence and MGE genes was in the special focus of thestudy due to the problem posed by such clones for safeeconomical and reliable way of wastewater disposal incostal ecosystems
54ordm22rsquo588rsquorsquoN 18ordm58rsquo048rsquorsquoE
54ordm37rsquo084rsquorsquoN 18ordm33rsquo288rsquorsquoE
54ordm37rsquo084rsquorsquoN 18ordm33rsquo288rsquorsquoE
Figure 1 Sampling sites location WWTP Gdansk-Wschod ndash influent (W-INF) bioreactor (W-BR) effluent (W-EFF) and marine outfall(W-MOut) WWTP Gdynia-Debogorze effluent (D-EFF) and marine outfall (D-MOut) Vistula River mouth (VR) Both WWTPs treat mainlymunicipal wastewater industrial and not disinfected hospital wastewater consist approximately 10 and 02 of daily inflow respectively TheWWTP Gdansk-Wschod works in a modified University of Cape Town system Qav = 96 000 m3d serves 570 000 people in operation since 2001The WWTP Gdynia-Debogorze works in Bardenpho system including the Barnard modification Qav = 55 000 m3d serves 350 000 people inoperation since 2011 The Vistula River is the second biggest river (1 048 km long) in the catchmentrsquos area of the Baltic Sea the average flow atthe mouth to the Baltic Sea ca 1050 m3sminus1
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 3 of 15httpwwwbiomedcentralcom1471-21801466
ResultsSpecies composition and antimicrobial susceptibility ofenterococcal isolatesIn this study the number of enterococci in WWTP efflu-ents (W-EFF and D-EFF) remained high for both WWTPyielding up to 61x105 CFU per 100 mL (Table 1) althoughthe WWTP Gdansk-Wschod showed over 99 efficiencyin the removal of enterococci In marine outfalls of the twoWWTPs (W-MOut D-MOut) as well as at the Vistula
Table 1 Presence (CFU 100 mLminus1) of enterococci number ()composition at particular sites
Gdansk-W
W-INF W-BR
Presence CFU 100 mLminus1 (07ndash23) times 107 na (
Number () of recovered isolates 33 (77) 55 (128)
Species compositionnumber ()
E faecium 19 (576) 36 (655)
E faecalis 5 (152) 8 (145)
E hirae 8 (242) 11 (200)
Ecasseliflavusgallinarum 1 (30) nd
E durans nd nd
E avium nd nd
na ndash not applicable nd ndash not detectedAbbreviations of sampling pointsrsquo locations and their description provided in Figure
River mouth enterococci were detected at low densitiesie below 100 CFU per 100 mL Altogether 428 isolateswere collected including 243 isolates from wastewater 116isolates from marine outfalls and 69 from Vistula Rivermouth (Table 1) These 428 isolates were mainly identifiedas E faecium (altogether 244 isolates 57) Enterococcushirae (113 isolates 264) and E faecalis (63 isolates147) with an occasional occurrence of E gallinarumcasseliflavus E durans and E avium (4 3 and 1 isolates
of enterococcal isolates recovered and species
WWTP Vistulariver Totalschod Gdynia-Debogorze
W-EFF W-MOut D-EFF D-MOut VR
01ndash61) times 105 3 ndash 60 (08ndash31) times 105 2 ndash 80 1 ndash 30 -
82 (192) 45 (105) 73 (171) 71 (166) 69 (161) 428 (100)
53 (646) 29 (644) 41 (562) 40 (563) 26 (377) 244 (57)
19 (232) 4 (89) 19 (260) 3 (43) 5 (72) 63 (147)
7 (85) 11 (244) 12 (164) 27 (380) 37 (536) 113 (264)
2 (25) nd 1 (14) nd nd 4 (10)
1 (12) 1 (23) nd 1 (14) nd 3 (07)
nd nd nd nd 1 (15) 1 (02)
1
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 4 of 15httpwwwbiomedcentralcom1471-21801466
respectively altogether 19) The most common speciesin all sampled sites was E faecium (562 ndash 655) withthe exception of Vistula River mouth where E hirae waspredominant (536 of isolates) followed by E faecalis(377)In the next step antimicrobial susceptibility was tested
for E faecium and E faecalis isolates (Table 2) It shouldbe noted that at some points a low number of E faecalisisolates was recovered (fewer than 10 isolates) and thuscertain resistance rates obtained for E feacalis should betreated with caution Resistance to erythromycin was de-tected with the highest prevalence among both specieshowever rates of resistance to this compound variedover a broad range Erythromycin-resistant E faeciumcomprised 368 of isolates in raw wastewater (W-INF)453 in treated wastewater (W-EFF) and 586 inmarine outflow (W-MOut) of WWTP Gdansk-WschodSimilar tendency was seen for E faecalis isolates whereresistance rates in the sampling points listed above wereequal to 200 684 and 100 respectively For WWTPGdynia-Debogorze effluent (D-EFF) and marine outflow(D-MOut) erythromycin resistance rates for E faeciumvaried between 682 and 550 respectively while for Efaecalis were equal 684 and 667 respectively In thecase of fluoroquinolones resistance to ciprofloxacin among
Table 2 Antimicrobial resistance of E faecium and E faecalis
Num
Resistant to a single antim
GM ST AM CIP LVX
E faecium
W= INF (n = 19) 1 (53) 3 (158) 1 (53) 6 (316) 1 (53
W-BR (n = 36) 1 (28) 2 (56) 1 (28) 12 (332) 3 (83
W-EFF (n = 53) 3 (57) 2 (38) 4 (75) 23 (434) 10 (18
W-MOut (n = 29) 0 6 (207) 6 (207) 13 (448) 4 (138
D-EFF (n = 41) 1 (24) 6 (146) 5 (122) 13 (317) 2 (49
D-MOut (n = 40) 1 (25) 2 (5) 2 (5) 14 (350) 3 (75
VR (n = 26) 0 1 (38) 0 8 (308) 1 (38
All (n = 244) 7 (29) 22 (90) 19 (78) 90 (369) 24 (98
E faecalis
W-INF (n = 5) 0 1 (200) 0 1 (200) 0
W-BR (n = 8) 2 (250) 2 (250) 1 (125) 2 (250) 2 (250
W-EFF (n = 19) 0 1 (53) 1 (53) 3 (158) 3 (158
W-MOut (n = 4) 0 0 0 0 0
D-EFF (n = 19) 2 (105) 5 (263) 0 1 (53) 0
D-MOut (n = 3) 1 (333) 1 (333) 0 0 0
VR (n = 5) 0 0 0 2 (400) 0
All (n = 63) 5 (79) 10 (159) 2 (32) 9 (143) 5 (79
GM gentamicin ST streptomycin AM ampicillin CIP ciprofloxacin LVX levofloxaciall tested antimicrobial agents MDR multiple drug resistance na not applicableAbbreviations of sampling pointsrsquo locations and their description provided in Figure
E faecium varied between 308 in Vistula River mouth to448 in W-MOut while resistance to levofloxacin did notexceed 20 Among E faecalis resistance to ciprofloxacinand levofloxacin was in general lower then reported for Efaecium in the corresponding sampling points (up to250) except Vistula River mouth The high-level strepto-mycin resistance (HLSR) was prevalent and reached 207for E faecium isolates from marine outflow of WWTPGdansk-Wschod (W-MOut) Also for E faecalis HLSRphenotype was detected among up to 16 of all tested iso-lates Resistance to glycopeptides linezolid and daptomycinwas not detected among all tested enterococci
Clonal structure of E faecium isolates and distribution ofvirulence resistance and MGE genesSixty-nine isolates of E faecium (283 of all isolates ofthis species) were further subjected to MLST and ana-lysis of the presence of resistance virulence and MGEgenes These isolates presented various phenotypes of re-sistance to antimicrobial compounds as described belowand were obtained mostly from effluents and marine out-falls of both WWTPs (Table 3) MLST discerned 54 differ-ent STs (Figure 2) including 23 new ones (STs 632ndash654)The comparative eBURST analysis against the wholeMLST E faecium database (as of the 5th April 2013)
ber () of isolates
icrobial agent Sensitive MDRphenotypeTE SYN E S
n = 244
) 5 (263) 6 (316) 7 (368) 6 (316) 4 (211)
) 7 (194) 19 (528) 16 (444) 10 (278) 8 (222)
9) 10 (189) 18 (340) 24 (453) 12 (226) 14 (264)
) 8 (276) 14 (483) 17 (586) 5 (174) 9 (310)
) 10 (244) 12 (292) 28 (682) 4 (98) 8 (195)
) 2 (50) 21 (525) 22 (550) 7 (175) 3 (75)
) 2 (77) 15 (577) 11 (424) 3 (115) 3 (115)
) 44 (180) 105 (430) 125 (512) 49 (201) 47 (193)
n = 63
1 (200) na 1 (200) 4 (800) 0
) 2 (250) na 7 (875) 1 (125) 2 (250)
) 4 (211) na 13 (684) 0 3 (158)
0 na 4 (100) 0 0
9 (474) na 13 (684) 4 (211) 5 (263)
2 (667) na 2 (667) 0 1 (333)
0 na 1 (200) 2 (400) 0
) 18 (286) na 41 (651) 11 (175) 11 (175)
n TE tetracycline SYN quinupristin-dalfopristin E erythromycin S sensitive to
1
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium
LineageCCST
MT Isolationsite
Virulence genes Resistance determinants Transposongenes
Plasmid rep genes Plasmid TAS
espintA
fms5 fms17 fms19 fms21 pbp5 GyrA ParC aac(6prime)-Ie-aph(2Prime)
ant(6prime)-Ia
tet(M)
tet(L)
tet(S)
intTn916 tndX rep1pIP501
rep2pRE25
rep17pRUM
rep18pEF418
reppLG1
reppMG1
axe-txe
ω-ε-ζ
relBE
Lineage 17
17 (2) 1 8 W-EFF 2 2 2 2 2 pbp5-30
S84R R61GS80R
1 1 2 2 1 2 2 1 1
386 7 W-MOut 1 1 1 1 1 1 1 1 1 1 1 1 1
Lineage 18
18 (3) 1(3)
D-EFFW-MOut
3 3 3 1 pbp5-20
S84Y R61GS80I
1 2 3 3 1 2 1 3
262 10 W-EFF 1 1 1 pbp5-25
S84Y R61G 1 1 1 1 1 1 1
574 402 VR 1 1 1 1
Lineage 78
78 (4) 159(4)
W-EFFW-MOut
4 4 4 4 4 pbp5-28
S84RY
R61GS80R
2 1 2 2 1 4 4 2 4 4 3
266 (2) 139(2)
W-INFD-EFF
2 1 2 2 pbp5-24
1 1 2 1 1 2 2 2
323 12 D-MOut 1 1 1 1 pbp5-26
wt R61GS80I
1 1 1 1 1 1 1 1
564 302 D-EFF 1 1 1 1 pbp5-29
S84I R61GS80R
1 1 1 1 1 1 1
653 302 W-EFF 1 nd S84I R61GS80R
1 1 1
CC5
29 (2) W-EFFW-MOut
2 1 1 2 1
66 W-MOut 1 1 1 1 1 1 1
97 W-EFF 1 wt R61GS80R
1 1
123 W-EFF 1 1 wt R61G 1 1 1
148 W-MOut 1 pbp5-30
wt R61GS80R
1 1 1 1 1
168 W-MOut 1 1 pbp5-31
1 1 1 1 1 1 1 1 1
505 W-EFF 1 wt R61G 1 1
588 D-MOut 1 pbp5-30
1 1 1 1 1 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
5of
15httpw
wwbiom
edcentralcom1471-21801466
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium (Continued)
635 D-EFF 1 1 1 1
636 W-EFF 1 wt R61G 1
637 W-EFF 1 1 1 wt R61G 1 1
638 W-EFF 1 wt R61GS80I
1
642 W-EFF 1 1 1 1 1
645 D-EFF 1 1 1 1 1
646 W-EFF 1 1 1 1 1
649 W-BR 1 1 1 pbp5-32
wt R61G 1
650 (3) D-EFF 3 3 pbp5-27
2 2 2 1 1 3
CC 9
8 D-EFF 1 1 1 1 1 1 1 1 1 1 1
640 W-EFF 1 1 1 1 1
CC 22
21 W-EFF 1 1 1 1 1 1 1 1
22 W-EFF 1 1 1 1 1 1
25 W-EFF 1 1 pbp5-32
wt R61G 1 1 1
32 (3) W-EFF 3 3 1 wt R61G 2 1
92 W-EFF 1 1 1 1 1
441 W-EFF 1 1 1 1 1 1 1 1
533 W-EFF 1 1 1
633 W-EFF 1 1 1 1 1
644 D-EFF 1 1 1 1 1
CC27
652 W-EFF 1 1 1 1
CC46
47 VR 1 1 1 1
69 D-EFF 1 1 1 1 1 1
CC94
94 W-MOut 1 1 1 1
361 (4) D-EFFD-MOut
3 4 4 4 pbp5-29
1 1 1 4 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
6of
15httpw
wwbiom
edcentralcom1471-21801466
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium (Continued)
CC296
296 W-EFF 1 1
CC639
639 W-EFF 1 1 1 1 1
CC648
634 W-EFF 1 1 1 1 1
648 W-EFF 1 1 1 1 1
Doubletons
632 W-MOut 1 1 1
Singletons
264 W-INF 1 1 1 1 1 1 1
641 D-EFF 1 1 1 1
643 W-EFF 1 1 1 1 1 1
647 W-MOut 1 1 1 1 1
651 D-EFF 1 1 1 1 1 1
654 VR 1 1 1 1 1 1
All (69) 8 57 49 29 46 6 17 20 17 1 9 4 39 27 14 7 55 2 12 10 5
CC clonal complex ST sequence type MT MLVA-type nd not determined due to lack of amplification of the pbp5 gene wt wild-type STs associated with MDR phenotype in boldAbbreviations of sampling pointsrsquo locations and their description provided in Figure 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
7of
15httpw
wwbiom
edcentralcom1471-21801466
Figure 2 MLST-based similarity tree of E faecium isolates constructed using the START software and the UPGMA clustering algorithm
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 8 of 15httpwwwbiomedcentralcom1471-21801466
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 9 of 15httpwwwbiomedcentralcom1471-21801466
grouped the 38 STs specific for 50 isolates into the majorlsquosuper-complex 17rsquo characteristic for this species (data notshown) Further detailed analysis of the localization ofparticular STs within this structure revealed that 11 STscharacteristic for 17 isolates belonged to the nosocomialcomplex of E faecium representing all three majorlineages 17 18 and 78 of a former CC17 as defined byWillems et al [9] (Table 3) Seventeen STs (20 isolates)nine STs (11 isolates) and two STs with single isolates eachbelonged to CC5 CC22 and CC9 respectively as definedby Freitas et al [24] Of the remaining 16 STs nine wereincluded into six small CCs (CCs 27 46 94 296 639648) unlinked to the major lsquosuper-complexrsquo ST632formed a lsquodoubletonrsquo (ie was linked by a single-locus vari-ant link to only one ST from the database) and six STsrepresented by single isolates were singletons ie werenot related to any ST in the database at the time of ana-lysis Thirty isolates (435) in the investigated groupshowed the MDR phenotype which was significantly asso-ciated (p = 000006) with isolates from the nosocomialcomplex (Table 3 and Figure 3) but found also in CC5CC22 and CC94 this phenotype did not occur amongsmall CCs or singletons Seventeen isolates repre-senting the nosocomial complex were additionallystudied by multilocus VNTR (variable number tan-dem repeats) analysis (MLVA) to better elucidate therelationships among these isolates Altogether nineMLVA-types (MTs) were found (Table 3) Importantlythe same MLVA-type MT159 associated with ST78was characteristic for isolates derived from samplesfrom W-EFF and W-MOut suggesting survival of thisparticular clone
0
10
20
30
40
50
60
70
80
90
100
IS16
espintA
fms5
fms1
7
fms1
9
fms2
1
MDR
AM-R
CIP-R
HLG
R
p
ositi
ve
isol
ates
Figure 3 Distribution () of IS16 resistance phenotypes and virulenccolumns isolates with STs typical for nosocomial E faecium grey columnsgroups are indicated by an asterisk MDR multi-drug resistance AM-R ampgentamicin resistance HLSR high-level streptomycin resistance
IS16 was present in all 17 isolates included by MLST tothree lineages of nosocomial E faecium and this markerwas limited to this group (Table 3 and Figure 3) TheespEfm gene occurred always together with intA among allisolates of lineage 17 and in some isolates of lineage 78The fms19 pilin gene detected in 29 isolates was over-represented among STs typical for nosocomial E faeciumwhile other pilin genes such as fms5 fms17 and fms21detected in 57 49 and 46 isolates respectively occurredamong various CCs and singleton STs Ampicillin resist-ance significantly associated with isolates of nosocomialcomplex (10 of 17 isolates 588 p = 0001) occurred alsoin CC5 (6 isolates) CC22 and CC94 (1 isolate each) Se-quencing of the 3prime part of the pbp5 gene revealed its 11nucleotide alleles corresponding to 11 variants of the C-terminal part of PBP5 Six alleles of the gene were new(pbp5-24 to pbp5-29) two alleles (pbp5-20 pbp5-22)were described previously among Polish hospital VREmisolates [13] and three alleles were found in other stud-ies pbp5-30 occurred in a Portuguese animal strain ofE faecium [25] and pbp5-31 and pbp5-32 in clinical iso-lates TCGE70411_2 and 164306 also from Portugal (de-posited as JN208888 and JN208885 in the GenBankrespectively) For a single ampicillin-resistant isolate ofST653 (lineage 78) pbp5 amplification was not achieveddespite repeated attempts The majority (11 of 17 iso-lates 647) of isolates belonging to three lineages ofnosocomial E faecium showed resistance to ciprofloxa-cin determined by mutations R61G and S80IR in ParCand S84IRY in GyrA Ciprofloxacin resistance wasalso significantly over-represented among isolates ofSTs typical for nosocomial E faecium (p = 00006) Nine
HLS
R
tetM tetL
rep1
-pIP50
1
rep2
-pRE25
rep1
7-pR
UM
rep1
8-pE
F418
rep-pL
G1
axe-txe
relBE
e resistance and plasmid genes among E faecium isolates Blackother E faecium isolates Significant differences (p lt 005) between twoicillin resistance CIP-R ciprofloxacin resistance HLGR high-level
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 10 of 15httpwwwbiomedcentralcom1471-21801466
isolates resistant to ciprofloxacin were found also inCCs 5 9 and 22 These isolates possessed mutations inParC mentioned above but had the wild-type QRDR inGyrA HLGR occurred in five isolates of lineages 18 and78 and in single isolates of CC5 and CC22 this pheno-type was determined by the presence of aac(6prime)-Ie-aph(2Prime) with an exception of an isolate from CC5 whichwas negative for this gene High-level resistance tostreptomycin (HLSR) was widely distributed amongstudied isolates and determined by the ant(6prime)-Ia geneTetracycline resistance was found also in various STsthe major determinant of this phenotype was tet(M) (20isolates) followed by tet(L) (17 isolates) These twogenes occurred together in 16 isolates A single isolateof ST633 belonging to CC22 harbored tet(S) The tet(O) gene was absent in the studied group For a singletetracycline-resistant isolate of ST17 none of the fourtetracycline determinants studied was detected Nineisolates carried int of Tn916 and four the tndX gene ofTn5397 These transposon-specific genes were not char-acteristic for any lineage or complex (Table 3) Plasmid-replication genes such as rep2pRE25 rep17pRUM andrep18 pEF418 (26 14 and 7 isolates respectively) andgenes specifying toxin-antitoxin systems (TAS) axe-txeand ω-ε-ζ (12 and 10 isolates respectively) were signifi-cantly associated with the group of isolates of nosocomialE faecium Two other rep genes (rep1pIP501 reppLG1) werewidely distributed among all studied E faecium isolatesand the reppMG1 gene occurred in two isolates (in a singleisolate of ST533 from CC22 and in a singleton ST647) Allω-ε-ζ-positive isolates carried rep2pRE25 and nine isolatesshowed a concomitant presence of axe-txe and rep17pRUMThe relBE genes were found solely among five non-nosocomial E faecium (a difference in distribution with-out statistical significance p = 043) among various STs
DiscussionIn this study Enterococcus spp was used to evaluate theimpact posed by treated wastewater of two local WWTPson their receiver the costal waters of Gulf of GdanskTotal counts of enterococci observed for raw (W-INF)and treated (W-EFF D-EFF) wastewater are consistentwith our previous study showing similar over 99 reduc-tion capacity of applied treatment processes It is alsoreported in the literature that wastewater treatment pro-cesses based on activated sludge although they focus onremoval of nutrients and organic matter simultaneouslyreduce faecal indicators with high efficiency reaching upto 999 [1822] Due to the high number of enterococciin raw wastewater they are nevertheless released from aWWTP in a number up to 61x105 CFU per 100 mLThus marine water sampled in the area directly impactedby treated wastewater (W-MOut and D-MOut) showed ahigher number of enterococci than the one observed in
Vistula River mouth however lower by four orders ofmagnitude in comparison to corresponding treated waste-water (W-EFF and D-EFF) It should be noted that in mar-ine outfalls (W-MOut D-MOut) as well as in VistulaRiver mouth enterococci were detected in number lowerthan 100 CFU per 100 mL ie the value set out by NewBathing Water Directive 20067EC for the costal water ofldquoexcellent qualityrdquo [23]It is suggested that enterococcal species composition in
faecal-impacted environment depends from the source ofcontamination In this study two species most commonlycolonizing humans E faecalis and E faecium comprisedtogether from 73 to 88 of enterococcal isolates intested wastewater samples (W-INF W-BR W-EFF and D-EFF) 73 and 61 in marine outflows (W-MOut and D-MOut respectively) and 45 in the Vistula River mouthThe predominant species in wastewater and marine waterimpacted by treated wastewater was E faecium (from 56to 65) which stayed at a similar level during treatmentprocesses as determined also in our previous study [20]Species distribution in wastewater samples is not wellunderstood and affecting factors may include differencesin diet climate season and methods of detection E feaca-lis was found to be the most prevalent species in wastewa-ter samples in Sweden [26] E faecium in Spain UKCanada France and Switzerland [26-29] while E hiraepredominated in Portugal and United States [2230] Al-though E faecalis is typically the most abundant colonizerof humans in the community (eg [2631]) E faecium isgenerally most often associated with human microbialcontamination [2729] E faecalis was reported fromagriculture-impacted waters [32] and E hirae likelyoriginates mainly from cattle and pig faecal contamin-ation [26] which explains high relative proportion ofthese two species in Vistula River mouth Apart from thevariability of contamination sources proportion of par-ticular species may differ due to the differences of abilityto sustain environmental stresses In this study enterococ-cal composition might have been influenced by wastewa-ter treatment processes and marine water characteristicsThe effect of wastewater treatment and marine environ-
ment was seen on phenotypes of antimicrobial resistanceThe positive selection of bacteria with resistance patternshas been already suggested and noted in wastewater pro-cesses [192022] In this study the resistance rates to ampi-cillin ciprofloxacin levofloxacin and erythromycin notedfor E faecium from raw wastewater (W-INF) were lowerthan these observed for this species in the correspondingtreated wastewater (W-EFF) however not statistically sig-nificant (p gt 005) Moreover resistance rates to ampicillinciprofloxacin tetracycline and erythromycin observed inmarine outflow of WWTP Gdansk-Wschod (W-MOut)were higher than in treated wastewater of this plantSuch phenomenon was not observed for treated
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 11 of 15httpwwwbiomedcentralcom1471-21801466
wastewater of WWTP Gdynia-Debogorze (D-EFF) andits marine outfall (D-MOut) Resistance rates among Efaecium originating from D-MOut were in generalcomparable to these detected in the Vistula Rivermouth It should be noted that treated wastewater fromWWTP Gdansk-Wschod has been discharged via mar-ine outfall (W-MOut) for last 12 years while marineoutfall of WWTP Gdynia-Debogorze (D-MOut) hasbeen operating for last two years The observed differ-ences in resistance rate corresponding to the local im-pact caused by long and short-term operated marineoutfalls (W-MOut and D-MOut) need further attentionSignificant rates of antimicrobial resistance observed
for isolates of E faecalis from wastewater and marineriver waters impacted by wastewater are in agreementwith a recent study performed in six European countriesshowing that in Poland the rates of resistance for humanisolates of this species were usually among the highestboth in the community and hospitals [33] To our know-ledge no corresponding data are available for E faeciumcolonization in the community in Poland but a studyfocused on hospital VRE isolates showed their ubiquitousMDR phenotype and a high load of resistance genes [13]In our study no vancomycin resistance were detectedlikely due to relatively low (below 10) prevalence of VREin Polish hospitals [5] although such isolates are isolatedfrom the environment in other countries eg [34]This study shows that hospital clones of E faecium are
important carriers of resistance genes in wastewatersMost likely in a significant part they originated fromhospital wastewater treated together with communalwastewater In our estimation hospital wastewater con-stitutes approximately 02 of both WWTPsrsquo daily in-flow The release of hospital-associated clones of Efaecium to the environment was observed also by others[24283536] In our study isolates belonging to themajor nosocomial HiRECC (formerly named CC17)constituted altogether 246 of all isolates of E faeciumsubjected to typing and were present at all samplingsites These isolates apart from their inclusion into theHiRECC by MLST demonstrated several other pheno-typic and genotypic features typical for nosocomial Efaecium ie they were resistant to ciprofloxacin andampicillin in vast majority showed MDR phenotypeand carried several resistance determinants associatedboth with mutations of chromosomal genes (gyrA parCpbp5) and in the case of aminoglycoside and tetracyclineresistance with gene acquisition Plasmids likely played animportant role in the later process as some plasmid genessuch as rep2pRE25 rep17pRUM and rep18 pEF418 and TASgenes axe-txe and ω-ε-ζ were significantly more abundantin isolates associated with HiRECC than among theremaining isolates As yet the knowledge of distributionof plasmid-associated genes in populations of E faecium
especially in the context of their clonal compositionremains limited A study on 93 isolates of differentgeographical origins and from various sites showed anover-representation of rep17pRUM and reppMG1 amongnosocomial E faecium while in contrast to our observa-tions rep18 pEF418 was not detected [37] Moreover wedid not observe reppMG1 among HiRECC of E faeciumThese differences may be due to a high prevalence of VRE(58 isolates) among 93 isolates studied by Rosvoll et alwhat may explain differences in the plasmid content Thesame study also reported the co-localization of ω-ε-ζ withrep2pRE25 and axe-txe with rep17pRUM on the same plas-mids in concordance with our findings showing a fre-quent joint occurrence of these pairs of genes Anotherstudy performed on 99 invasive isolates from Norwaymostly of HLGR phenotype showed a significantly higherprevalence of rep2pRE25 and rep17pRUM among the majornosocomial HiRECC [38] This study as well some others[2439] also demonstrated a common presence of reppLG1among E faecium from hospital settings and in animalsOur findings indicate that this type of replicon is evenmore ubiquitous in E faecium than previously reportedAll rep genes mentioned above are typically associatedwith plasmids carrying various antimicrobial resistancegenes [3940] GenBank accession number AF408195 Inaddition to genes determining antimicrobial resistanceand genes associated with MGE isolates of nosocomialHiRECC analyzed in our study carried such molecularmarkers as IS16 and the esp gene together with the intAgene characteristic for ICEEfm [4142] Another patho-genicity factor the fms19 gene of the pilin gene cluster4 (PGC4) showed increased prevalence among theseisolates Similar observation was made for a representa-tive collection of 433 E faecium isolates from varioussources [43]In a study that followed enterococci in a continuum
from hospital and retirement home wastewaters througha WWTP to a river a gradual decrease in the proportionof E faecium isolates representing hospital-associatedSTs was observed likely due to the ldquodilutionrdquo of HiRECCrepresentatives by more diverse and less drug-resistantstrains originating from the community [28] Because inour study only selected representatives of obtained iso-lates were investigated it is not possible to determineprecise tendencies in their frequency from the WWTPinfluent to effluent and water however resistant entero-cocci both HiRECC and non-HiRECC are clearly ableto survive the treatment process what results in their re-lease into marine and river waters Moreover using anadditional typing method MLVA we were able to findisolates with the same characteristics in the effluent ofWWTP and its marine outfall suggesting ability of suchclones to survive in the environment for at least sometime It seems plausible that hospital resistant clones
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 12 of 15httpwwwbiomedcentralcom1471-21801466
accumulate several determinants that may promote theirsurvival not only in the hospital settings but also in theenvironment Such features would include resistance todisinfectants and ultraviolet light additional metabolicpathways and ability to form biofilms Moreover determi-nants of some of the mentioned adaptation factors arefound co-resident in plasmids harboring resistance genessuggesting co-selection For example pLG1-type plasmidsof E faecium carry not only resistance determinantsagainst aminoglycosides glycopeptides and macrolidesbut also pili genes and carbon uptake-utilization genes[3839] Pheromone-responsive plasmids of E faecalisapart from resistance to aminoglycosides glycopeptidespenicillins and macrolides may encode bacteriocins add-itional pili and UV-resistance determinants [40] Thusnosocomial HiRECCs released by WWTPs are equippedin whole sets of genes with various adaptive benefits andoften located within MGE It was also demonstrated thatMGEs can be transferred in a WWTP [44] further under-ling the importance of threat posed by such organismsThere is a selection pressure for survival of resistantstrains and their tranconjugants due to release of antibi-otics to the environment especially in the hospital efflu-ents [45] and even sub-inhibitory concentrations may beeffective in selection for resistance [46]
ConclusionsThis study describes the species composition resistanceprofiles and clonal relationships of enterococci isolatedfrom WWTPs located in Gdansk their marine outfallsto the Gulf of Gdansk and from Vistula River It was ob-served that these WWTPs release enterococcal strainsresistant to various antimicrobial compounds to the re-cipient water environment Molecular analyses revealedthe presence of clones associated with nosocomial HiR-ECCs and in the case of E faecium isolates of lineages17 18 and 78 of former CC17 abundant in resistance de-terminants MGE genes and pathogenicity factors Thepresence of HiRECCs in wastewaters and marineriverenvironment highlights the need for further detailedanalyses to better understand the survival and spread ofdrug-resistant strains in water ecosystems and to eluci-date the ways to curb such dissemination
MethodsAcquisition of samples and isolation of enterococciOnce a month in April June August and October in 2011of the flow proportioned composited samples of wastewa-ter were taken from two local WWTPs Gdansk-Wschodfrom influent (W-INF) effluent (W-EFF) and wastewa-ter from activated sludge bioreactor (W-BR) and fromGdynia-Debogorze effluent (D-EFF) The two WWTPstreat sewage from approximately 920 000 inhabitants(for details see Figure 1) In the same period of time and
with the same frequency marine outfalls of those WWTPs(W-MOut and D-MOut respectively) as well as VistulaRiver mouth were also sampled for isolation of entero-cocci Marine samples were taken at the depth of approxi-mately one meter below the surface once a month inApril June August and October of 2011 Location ofsampling points and their characteristics are provided inFigure 1 Enterococci were detected using the membranefiltration method To this end appropriate dilutions ofanalyzed samples were filtered in duplicate through045 μm cellulose-acetate filters (EMD Millipore Cor-poration Billerica MA USA) which were then placedon Enterococcus selective agar (Merck DarmstadtGermany) and incubated at 37degC for 48 h (ISO 7899ndash22000) Dark red or maroon colonies were consideredpresumptive enterococci For further investigationsrepresentative isolates were taken from membranes pre-senting from 20 to 50 typical colonies and stored in nu-trient broth supplemented with 15 glycerol at minus80degC
Species identification DNA isolation susceptibility testingand statistical analysisThe species identification and drug susceptibility of pre-sumptive enterococci were determined by the PhoenixtradeAutomated Microbiology System (BD New Jersey USA)according to the manufacturerrsquos instructions Since it isnot possible to distinguish E casseliflavus and E galli-narum using this system the classification E casselifla-vusgallinarum was used The susceptibility tests basedon microdilution were carried out for 11 antimicrobialagents gentamicin (GM) streptomycin (ST) ampicillin(AM) daptomycin (DAP) vancomycin (VA) teicoplanin(TEC) linezolid (LZD) fluoroquinolones (ciprofloxacin -CIP levofloxacin - LVX) tetracycline (TE) erythromycin(E) and quinupristin-dalfopristin (SYN) (E faecium only)Obtained minimal inhibitory concentrations (MIC) wereevaluated according to the guidelines of Clinical and La-boratory Standards Institute [47] Isolates were defined asmultidrug-resistant (MDR) when they showed resistanceto three or more compounds tested [48] Bacterial DNAwas purified using the Genomic DNA Prep Plus kitfollowing the manufacturerrsquos instructions (AampA Biotech-nology Gdynia Poland) Differences in distributions wereevaluated using the χ2 test with the p values lt 005 con-sidered significant
MLST and MLVA of E faeciumMLST for E faecium was performed by sequencing ofseven house-keeping genes as described by Homan et al[49] The allele numbers and STs were assigned with theuse of Internet database httpefaeciummlstnet (5th July2013 date last accessed) Novel alleles and allelic profileswere submitted to the database MLST data were analyzedwith Sequence Type Analysis and Recombinational Tests
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 13 of 15httpwwwbiomedcentralcom1471-21801466
(START) software [50] and the eBURST analysis [51] wasused to elucidate the relationships of STs with knownCCs using the comparative eBURST option available athttpeburstmlstnetv3enter_datacomparative (5th July2013 date last accessed) Further inclusion of E faeciumSTs into specific CCs and lineages was performed fol-lowing other studies [924] MLVA was performed as de-scribed by others [52]
Detection and analysis of genes associated with virulenceantimicrobial resistance and MGE in E faeciumThe full list of primers used for gene detection and se-quencing is provided in Additional file 1 Table S1 PCR-based detection of IS16 virulence-associated genes (fms21fms17 fms5 and fms19 representing each of four pilingene clusters in E faecium and the espEfm gene) genes de-termining resistance to tetracycline tet(M) tet(O) tet(L)tet(S) HLGR (aac(6prime)-Ie-aph(2Prime) HLSR (ant(6prime)-Ia) andgenes specific for MGE such as intA of ICEEfm1 int ofTn916 transposon tndX of Tn5397 transposon entero-coccal plasmid replication genes rep1pIP501 rep2pRE25rep17pRUM rep18pEF418 reppMG1 reppLG1 and TAS genesaxe-txe ω-ε-ζ relBE was performed as described in otherstudies [134153-59] following the conditions describedherein Previously characterized clinical isolates of Strepto-coccus agalactiae E faecium and E faecalis harbouringpilin genes IS16 espEfm intA of ICEEfm1 int of Tn916tndX of Tn5397 tet(M) tet(O) tet(L) tet(S) aac(6prime)-Ie-aph(2Prime) ant(6prime)-Ia rep1pIP501 rep2pRE25 ω-ε-ζ [136061]served as positive controls In cases when positive controlswere not available (rep18pEF418 reppMG1 reppLG1 axe-txerelBE) a few randomly selected PCR products wereverified by sequencing and served further as controlsAmpicillin-resistance determinant the pbp5 gene was ana-lysed by sequencing of the part encoding C-terminaltranspeptidase domain in PBP5 [62] New sequence vari-ants pbp5-24 to pbp5-29 alleles were submitted to theGenBank (acc numbers KC594860-KC594865 respect-ively) The quinolone-determining regions (QRDR) in thegyrA and parC genes were sequenced and analysed asdescribed by others [63]
Additional file
Additional file 1 Table S1 Primers used in the study for PCR andsequencing
Competing interestThe authors declare that they have no competing interest
Authorsrsquo contributionsES participated in the study design performed experimental work andparticipated in the manuscript preparation AL designed the studyperformed experimental work and participated in the manuscriptpreparation Both authors read and approved the final manuscript
AcknowledgementsWe thank Rob Willems and Janetta Top for assigning new alleles new STsand MTs This publication made use of the Enterococcus faecium MLSTwebsite (httpefaeciummlstnet) which is hosted at Imperial College of theUniversity of Oxford and funded by the Wellcome Trust This work wassupported by project N N305 461139 from the Narodowe Centrum Nauki(NCN)
Author details1National Medicines Institute ul Chelmska 3034 Warsaw 00-725 Poland2Gdansk University of Technology ul G Narutowicza 1112 Gdansk 80-952Poland
Received 5 September 2013 Accepted 27 February 2014Published 14 March 2014
References1 Aarestrup FM Butaye P Witte W Nonhuman reservoirs of Enterococci In
American Society for Microbiology Edited by Gilmore MS Clewell DB CourvalinP Dunny GM Murray BE Rice LB Washington DC The Enterococcipathogenesis molecular biology and antibiotic resistance 2002
2 Tendolkar PM Baghdayan AS Shankar N Pathogenic enterococci newdevelopments in the 21st century Cell Mol Life Sci 2003 602622ndash2636
3 Sood S Malhotra M Das BK Kapil A Enterococcal infections ampantimicrobial resistance Indian J Med Res 2008 128111ndash121
4 Sydnor ER Perl TM Hospital epidemiology and infection control inacute-care settings Clin Microbiol Rev 2011 24141ndash173
5 European Centre for Disease Prevention and Control Annual EpidemiologicalReport Reporting on 2010 surveillance data and 2011 epidemic intelligencedata Stockholm ECDC 20122013
6 Hidron AI Edwards JR Patel J Horan TC Sievert DM Pollock DA Fridkin SKNational Healthcare Safety Network Team Participating National HealthcareSafety Network Facilities NHSN annual update antimicrobial-resistantpathogens associated with healthcare-associated infections annualsummary of data reported to the National Healthcare Safety Network atthe Centers for Disease Control and Prevention 2006ndash2007 Infect ControlHosp Epidemiol 2008 29996ndash1011
7 De Kraker ME Jarlier V Monen JC Heuer OE van de Sande N GrundmannH The changing epidemiology of bacteraemias in Europe trends fromthe European Antimicrobial Resistance Surveillance System Clin MicrobiolInfect 2012 doi1011111469-0691
8 Leavis HL Bonten MJ Willems RJ Identification of high-risk enterococcalclonal complexes global dispersion and antibiotic resistance Curr OpinMicrobiol 2006 9454ndash460
9 Willems RJL Top J van Schaik W Leavis H Bonten M Sireacuten J Hanage WPCorander J Restricted gene flow among hospital subpopulations ofEnterococcus faecium mBio 2012 3e00151ndash12
10 Hegstad K Mikalsen T Coque TM Werner G Sundsfjord A Mobile geneticelements and their contribution to the emergence of antimicrobialresistant Enterococcus faecalis and Enterococcus faecium Clin MicrobiolInfect 2010 16541ndash554
11 Shepard BD Gilmore MS Antibiotic-resistant enterococci themechanisms and dynamics of drug introduction and resistance MicrobesInfect 2002 4215ndash224
12 Bonten MJ Willems R Weinstein RA Vancomycin-resistant enterococciwhy are they here and where do they come from Lancet Infect Dis 20011314ndash325
13 Sadowy E Sieńko A Gawryszewska I Bojarska A Malinowska K HryniewiczW High abundance and diversity of antimicrobial resistancedeterminants among early vancomycin-resistant Enterococcus faecium inPoland Eur J Clin Microbiol Infect Dis 2013 321193ndash1203
14 Willems RJ Homan W Top J van Santen-Verheuvel M Tribe D Manzioros XGaillard C Vandenbroucke-Grauls CM Mascini EM van Kregten E vanEmbden JD Bonten MJ Variant esp gene as a marker of a distinct geneticlineage of vancomycin-resistant Enterococcus faecium spreading inhospitals Lancet 2001 357853ndash855
15 Sillanpaumlauml J Nallapareddy SR Prakash VP Qin X Houmloumlk M Weinstock GMMurray BE Identification and phenotypic characterization of a secondcollagen adhesin Scm and genome-based identification and analysis of13 other predicted MSCRAMMs including four distinct pilus loci inEnterococcus faecium Microbiology 2008 1543199ndash3211
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 14 of 15httpwwwbiomedcentralcom1471-21801466
16 Galloway-Pentildea JR Nallapareddy SR Arias CA Eliopoulos GM Murray BEAnalysis of clonality and antibiotic resistance among early clinicalisolates of Enterococcus faecium in the United States J Infect Dis 20092001566ndash1573
17 Top J Willems R Bonten M Emergence of CC17 Enterococcus faeciumfrom commensal to hospital-adapted pathogen FEMS Immunol MedMicrobiol 2008 52297ndash308
18 Martins da Costa P Vaz-Pires P Bernardo F Antimicrobial resistance inEnterococcus spp isolated in inflow effluent and sludge from municipalsewage water treatment plants Water Res 2006 401735ndash1740
19 Reinthaler FF Posch J Feierl G Wust G Haas D Ruckenbauer G Mascher FMarth E Antibiotic resistance of Escherichia coli in sewage and sludgeWater Res 2003 371685ndash1690
20 Luczkiewicz A Fudala-Ksiazek S Jankowska K Olanczuk-Neyman KAntimicrobial resistance of fecal indicators in municipal wastewatertreatment plant Water Res 2010 445089ndash5097
21 Dulfour AP Ballentine RK Ambient water quality criteria for bacteria-1986 EPA4405-84-002 Washington DC US Environmental Protection Agency 1986
22 Ferreira da Silva M Tiago I Veriacutessimo A Boaventura RA Nunes OC Manaia CMAntimicrobial resistance patterns in Enterobacteriaceae isolated from anurban wastewater treatment plant FEMS Microbiol Ecol 2007 60166ndash176
23 DIRECTIVE 20067EC of the European Parliament and of the Council 15February 2006 concerning the management of bathing water quality andrepealing Directive 76160EEC httpeur-lexeuropaeuLexUriServLexUriServdouri=OJL200606400370051ENPDF
24 Freitas AR Novais C Ruiz-Garbajosa P Coque TM Peixe L Dispersion ofmultidrug-resistant Enterococcus faecium isolates belonging to majorclonal complexes in different Portuguese settings Appl Environ Microbiol2009 754904ndash4908
25 Poeta P Costa D Igrejas G Saacuteenz Y Zarazaga M Rodrigues J Torres CPolymorphisms of the pbp5 gene and correlation with ampicillinresistance in Enterococcus faecium isolates of animal origin J MedMicrobiol 2007 56236ndash240
26 Kuumlhn I Iversen A Burman LG Olsson-Liljequist B Franklin A Finn MAarestrup F Seyfarth AM Blanch AR Vilanova X Taylor H Caplin J MorenoMA Dominguez L Herrero IA Moumlllby R Comparison of enterococcalpopulations in animals humans and the environment ndash a Europeanstudy Int J Food Microbiol 2003 88133ndash145
27 Lanthier M Scott A Lapen DR Zhang Y Topp E Frequency of virulencegenes and antibiotic resistances in Enterococcus spp isolates fromwastewater and feces of domesticated mammals and birds and wildlifeCan J Microbiol 2010 56715ndash729
28 Leclercq R Oberleacute K Galopin S Cattoir V Budzinski H Petit F Changes inenterococcal populations and related antibiotic resistance along amedical center-wastewater treatment plant-river continuum Appl EnvironMicrobiol 2013 792428ndash2434
29 Thevenon F Regier N Benagli C Tonolla M Adatte T Wildi W Poteacute JCharacterization of fecal indicator bacteria in sediments cores from thelargest freshwater lake of Western Europe (Lake Geneva Switzerland)Ecotoxicol Environ Safety 2012 7850ndash56
30 Bonilla TD Nowosielski K Esiobu N McCorquodale DS Rogerson ASpecies assemblages of Enterococcus indicate potential sources offecal bacteria at a south Florida recreational beach Mar Pollut Bull2006 52807ndash810
31 Wang JT Chang SC Wang HY Chen PC Shiau YR Lauderdale TL TSARHospitals High rates of multidrug resistance in Enterococcus faecalis andE faecium isolated from inpatients and outpatients in TaiwanDiagn Microbiol Infect Dis 2013 75406ndash411
32 Lanthier M Scott A Zhang Y Cloutier M Durie D Henderson VC WilkesG Lapen DR Topp E Distribution of selected virulence genes andantibiotic resistance in Enterococcus species isolated from the SouthNation River drainage basin Ontario Canada J Appl Microbiol 2011110407ndash421
33 Kuch A Willems RJ Werner G Coque TM Hammerum AM Sundsfjord AKlare I Ruiz-Garbajosa P Simonsen GS van Luit-Asbroek M Hryniewicz WSadowy E Insight into antimicrobial susceptibility and population struc-ture of contemporary human Enterococcus faecalis isolates fromEurope J Antimicrob Chemother 2012 67551ndash558
34 Novais C Coque TM Ferreira H Sousa JC Peixe L Environmentalcontamination with vancomycin-resistant enterococci from hospitalsewage in Portugal Appl Environ Microbiol 2005 713364ndash3368
35 Iversen A Kuumlhn I Rahman M Franklin A Burman LG Olsson-Liljequist BTorell E Moumlllby R Evidence for transmission between humans and theenvironment of a nosocomial strain of Enterococcus faeciumEnviron Microbiol 2004 655ndash59
36 Rathnayake I Hargreaves M Huygens F SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterwayAustralia and associated antibiotic resistance gene profilesBMC Microbiol 2011 11201
37 Rosvoll TC Pedersen T Sletvold H Johnsen PJ Sollid JE Simonsen GSJensen LB Nielsen KM Sundsfjord A PCR-based plasmid typing inEnterococcus faecium strains reveals widely distributed pRE25-pRUM- pIP501- and pHT-related replicons associated with glycopeptideresistance and stabilizing toxin-antitoxin systems FEMS Immunol MedMicrobiol 2010 58254ndash268
38 Rosvoll TC Lindstad BL Lunde TM Hegstad K Aasnaes B Hammerum AMLester CH Simonsen GS Sundsfjord A Pedersen T Increased high-levelgentamicin resistance in invasive Enterococcus faecium is associated withaac(6prime)Ie-aph(2Prime)Ia-encoding transferable megaplasmids hosted by majorhospital-adapted lineages FEMS Immunol Med Microbiol 2012 66166ndash176
39 Laverde Gomez JA van Schaik W Freitas AR Coque TM Weaver KE FranciaMV Witte W Werner G A multiresistance megaplasmid pLG1 bearing ahylEfm genomic island in hospital Enterococcus faecium isolates Int J MedMicrobiol 2011 301165ndash175
40 Coque TM Evolutionary biology of pathogenic enterococci InEvolutionary Biology of Bacterial and Fungal Pathogens Coque TM Edited byBaquero F Nombela C Cassell GH Gutierrez JA Washington DC ASMPress501ndash523
41 Werner G Fleige C Geringer U van Schaik W Klare I Witte W IS elementIS16 as a molecular screening tool to identify hospital-associated strainsof Enterococcus faecium BMC Infect Dis 2011 1180
42 Top J Sinnige JC Majoor EA Bonten MJ Willems RJ van Schaik W Therecombinase IntA is required for excision of esp-containing ICEEfm1 inEnterococcus faecium J Bacteriol 2011 1931003ndash1006
43 Sillanpaumlauml J Prakash VP Nallapareddy SR Murray BE Distribution of genesencoding MSCRAMMs and Pili in clinical and natural populations ofEnterococcus faecium J Clin Microbiol 2009 47896ndash901
44 Marcinek H Wirth R Muscholl-Silberhorn A Gauer M Enterococcus faecalisgene transfer under natural conditions in municipal sewage watertreatment plants Appl Environ Microbiol 1998 64626ndash632
45 Kuumlmmerer K Significance of antibiotics in the environment J AntimicrobChemother 2003 525ndash7
46 Liu A Fong A Becket E Yuan J Tamae C Medrano L Maiz M Wahba C LeeC Lee K Tran KP Yang H Hoffman RM Salih A Miller JHP Selectiveadvantage of resistant strains at trace levels of antibiotics a simple andultrasensitive color test for detection of antibiotics and genotoxicagents Antimicrob Agents Chemother 2011 551204ndash1210
47 Clinical and Laboratory Standards Institute Performance standards forantimicrobial susceptibility testing 21st informational supplement Wayne CLSI2011 PA M100-S20
48 Magiorakos AP Srinivasan A Carey RB Carmeli Y Falagas ME Giske CGHarbarth S Hindler JF Kahlmeter G Olsson-Liljequist B Paterson DL Rice LBStelling J Struelens MJ Vatopoulos A Weber JT Monnet DLMultidrug-resistant extensively drug-resistant and pandrug-resistantbacteria an international expert proposal for interim standarddefinitions for acquired resistance Clin Microbiol Infect 2012 18268ndash281
49 Homan WL Tribe D Poznanski S Li M Hogg G Spalburg E Van Embden JDWillems RJ Multilocus sequence typing scheme for Enterococcus faeciumJ Clin Microbiol 2002 401963ndash1971
50 Jolley KA Feil EJ Chan MS Maiden MC Sequence type analysis andrecombinational tests (START) Bioinformatics 2001 171230ndash1231
51 Feil EJ Li BC Aanensen DM Hanage WP Spratt BG eBURST inferringpatterns of evolutionary descent among clusters of related bacterialgenotypes from multilocus sequence typing data J Bacteriol 20041861518ndash1530
52 Top J Banga1 NMI Hayes R Willems RJ Bonten MJM Hayden MKComparison of multiple-locus variable-number tandem repeat analysisand pulsed-field gel electrophoresis in a setting of polyclonal endemicityof vancomycin-resistant Enterococcus faecium Clin Microbiol Infect 200814363ndash369
53 Vankerckhoven V Van Autgaerden T Vael C Lammens C Chapelle S RossiR Jabes D Goossens H Development of a multiplex PCR for the
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-

Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 2 of 15httpwwwbiomedcentralcom1471-21801466
BackgroundEnterococci are commensal bacteria of intestinal tractof mammals and birds but these bacteria can also befound in the gut of reptiles and insects and in differentenvironmental compartments like soil and plants infood (both as starters and contamination) as well as inaquatic ecosystems both distributed with treated waste-water and lacking such anthropogenic impact [1] In thehuman intestine the Enterococcus faecalis and Entero-coccus faecium are most abundant and these twospecies are also most commonly isolated from humanenterococcal infections however other species such asEnterococcus avium Enterococcus casseliflavus Entero-coccus durans Enterococcus gallinarum and Entero-coccus raffinosus have also been detected [23] RecentlyE faecalis and E faecium play more and more import-ant role in hospital-acquired infections (HAIs) such asbacteremia endocarditis infections of urinary tract andpost-surgery wounds [4] and nowadays represent a sec-ond etiologic agent of hospital bloodstream infectionsin Europe [5] In the recent years the relative propor-tion of E faecium to E faecalis is increasing in the USand Europe [6-8] which is likely due to the spread of aparticular hospital-adapted polyclonal high-risk entero-coccal complex (HiRECC) of E faecium initially describedas clonal complex 17 CC17 [8] This nosocomial subpop-ulation consists of three main lineages named from thecentral sequence type (ST) defined by multilocus se-quence typing (MLST) as lineages 17 18 and 78 [9] Theacquisition of resistance to antimicrobials of several clas-ses constitutes an important feature of hospital-associatedE faecium what not only renders them resilient to ther-apy but also creates a reservoir of antimicrobial resistancegenes often associated with mobile genetic elements(MGE) such as conjugative transposons and plasmids[10] The most important from the clinical point of viewexamples of acquired resistance among enterococci in-clude resistance to (amino)penicillins fluoroquinolonesglycopeptides (vancomycin-resistant enterococci VRE)and high-level resistance to aminoglycosides (HLAR)[3] Resistance to ampicillin associated with the changesin penicillin binding protein 5 (PBP5) and resistance toquinolones determined by point mutations in genes en-coding bacterial gyrase and topoisomerase IV are char-acteristic phenotypic features of nosocomial E faecium[1112] Such strains also commonly carry resistancegenes to other classes of antimicrobials such as tetracy-clines tet(M) tet(L) tet(S) and tet(O) macrolides erm(B) and aminoglycosides [13] The HLAR phenotype isusually determined by the aac(6prime)-Ie-aph(2Prime) gene en-coding a so-called bi-functional enzyme responsible forresistance to all aminoglycosides except for streptomycinhigh-level resistance to this compound is typically speci-fied by ant(6prime)-Ia [3] Hospital-associated E faecium is
also enriched in several virulence factors such as en-terococcal surface protein (Esp) and MSCRAMM pro-teins [14-16]Several studies have addressed the spread of HiRECCs
especially VRE within as well as among health-care facil-ities [17] Much less is known however about the releaseof drug-resistant enterococci and HiRECCs into the envir-onment and their subsequent survival Since it is sus-pected that human-associated bacteria can be regarded asvectors of gene transmission into environmental popula-tions [18] and positive selection of bacteria resistant toantimicrobial agents was observed in wastewater processes[1920] the potential role of effluents from wastewatertreatment plant (WWTP) in dissemination of clinicallyrelevant bacteria and genes needs evaluation It is particu-larly important in ecosystems subjected to strong an-thropogenic impact such as the Gulf of Gdansk and itsshallow western part the Puck Bay Significant pollutionload is discharged to this coastal area through the numer-ous local rivers and marine outfalls Due to limited waterexchange in the Gulf of Gdansk safe wastewater disposalis essential to prevent the environmental degradation andto preserve the public health Epidemiological studiesreported a direct relationship between the risk of gastro-enteritis among swimmers and density of enterococci insurface waters [21] Additionally resistant and multiresis-tant enterococci were detected in effluent of WWTPs[182022] suggesting their survival capacity in treatmentprocesses However no microbiological standards havebeen set on WWTP effluents in the majority of Europeancountries and it is required instead to monitor the qualityof recreational water [23] The US Environmental Protec-tion Agency [21] as well as the European Union legislation[23] recommended detection of fecal enterococci togetherwith Escherichia coli as indicators of fecal contaminationof bathing waterAim of the current study was to investigate the pres-
ence and species composition of enterococci in treatedwastewater river estuary water and their recipienthighly anthropogenically impacted Gulf of Gdansk Forthis purpose enterococci were isolated from wastewaterof two local WWTPs Gdansk-Wschod (influent - W-INFeffluent - W-EFF and bioreactor - W-BR) and Gdynia-Debogorze (effluent - D-EFF) as well as from their marineoutfalls (W-MOut and D-MOut respectively) Add-itionally the Vistula River mouth was sampled due toecological importance of this river flows for the Gulfof Gdansk area (Figure 1) Identification and furthercharacterization of drug-resistant enterococci especiallythe most prevalent E faecium including its resistancevirulence and MGE genes was in the special focus of thestudy due to the problem posed by such clones for safeeconomical and reliable way of wastewater disposal incostal ecosystems
54ordm22rsquo588rsquorsquoN 18ordm58rsquo048rsquorsquoE
54ordm37rsquo084rsquorsquoN 18ordm33rsquo288rsquorsquoE
54ordm37rsquo084rsquorsquoN 18ordm33rsquo288rsquorsquoE
Figure 1 Sampling sites location WWTP Gdansk-Wschod ndash influent (W-INF) bioreactor (W-BR) effluent (W-EFF) and marine outfall(W-MOut) WWTP Gdynia-Debogorze effluent (D-EFF) and marine outfall (D-MOut) Vistula River mouth (VR) Both WWTPs treat mainlymunicipal wastewater industrial and not disinfected hospital wastewater consist approximately 10 and 02 of daily inflow respectively TheWWTP Gdansk-Wschod works in a modified University of Cape Town system Qav = 96 000 m3d serves 570 000 people in operation since 2001The WWTP Gdynia-Debogorze works in Bardenpho system including the Barnard modification Qav = 55 000 m3d serves 350 000 people inoperation since 2011 The Vistula River is the second biggest river (1 048 km long) in the catchmentrsquos area of the Baltic Sea the average flow atthe mouth to the Baltic Sea ca 1050 m3sminus1
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 3 of 15httpwwwbiomedcentralcom1471-21801466
ResultsSpecies composition and antimicrobial susceptibility ofenterococcal isolatesIn this study the number of enterococci in WWTP efflu-ents (W-EFF and D-EFF) remained high for both WWTPyielding up to 61x105 CFU per 100 mL (Table 1) althoughthe WWTP Gdansk-Wschod showed over 99 efficiencyin the removal of enterococci In marine outfalls of the twoWWTPs (W-MOut D-MOut) as well as at the Vistula
Table 1 Presence (CFU 100 mLminus1) of enterococci number ()composition at particular sites
Gdansk-W
W-INF W-BR
Presence CFU 100 mLminus1 (07ndash23) times 107 na (
Number () of recovered isolates 33 (77) 55 (128)
Species compositionnumber ()
E faecium 19 (576) 36 (655)
E faecalis 5 (152) 8 (145)
E hirae 8 (242) 11 (200)
Ecasseliflavusgallinarum 1 (30) nd
E durans nd nd
E avium nd nd
na ndash not applicable nd ndash not detectedAbbreviations of sampling pointsrsquo locations and their description provided in Figure
River mouth enterococci were detected at low densitiesie below 100 CFU per 100 mL Altogether 428 isolateswere collected including 243 isolates from wastewater 116isolates from marine outfalls and 69 from Vistula Rivermouth (Table 1) These 428 isolates were mainly identifiedas E faecium (altogether 244 isolates 57) Enterococcushirae (113 isolates 264) and E faecalis (63 isolates147) with an occasional occurrence of E gallinarumcasseliflavus E durans and E avium (4 3 and 1 isolates
of enterococcal isolates recovered and species
WWTP Vistulariver Totalschod Gdynia-Debogorze
W-EFF W-MOut D-EFF D-MOut VR
01ndash61) times 105 3 ndash 60 (08ndash31) times 105 2 ndash 80 1 ndash 30 -
82 (192) 45 (105) 73 (171) 71 (166) 69 (161) 428 (100)
53 (646) 29 (644) 41 (562) 40 (563) 26 (377) 244 (57)
19 (232) 4 (89) 19 (260) 3 (43) 5 (72) 63 (147)
7 (85) 11 (244) 12 (164) 27 (380) 37 (536) 113 (264)
2 (25) nd 1 (14) nd nd 4 (10)
1 (12) 1 (23) nd 1 (14) nd 3 (07)
nd nd nd nd 1 (15) 1 (02)
1
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 4 of 15httpwwwbiomedcentralcom1471-21801466
respectively altogether 19) The most common speciesin all sampled sites was E faecium (562 ndash 655) withthe exception of Vistula River mouth where E hirae waspredominant (536 of isolates) followed by E faecalis(377)In the next step antimicrobial susceptibility was tested
for E faecium and E faecalis isolates (Table 2) It shouldbe noted that at some points a low number of E faecalisisolates was recovered (fewer than 10 isolates) and thuscertain resistance rates obtained for E feacalis should betreated with caution Resistance to erythromycin was de-tected with the highest prevalence among both specieshowever rates of resistance to this compound variedover a broad range Erythromycin-resistant E faeciumcomprised 368 of isolates in raw wastewater (W-INF)453 in treated wastewater (W-EFF) and 586 inmarine outflow (W-MOut) of WWTP Gdansk-WschodSimilar tendency was seen for E faecalis isolates whereresistance rates in the sampling points listed above wereequal to 200 684 and 100 respectively For WWTPGdynia-Debogorze effluent (D-EFF) and marine outflow(D-MOut) erythromycin resistance rates for E faeciumvaried between 682 and 550 respectively while for Efaecalis were equal 684 and 667 respectively In thecase of fluoroquinolones resistance to ciprofloxacin among
Table 2 Antimicrobial resistance of E faecium and E faecalis
Num
Resistant to a single antim
GM ST AM CIP LVX
E faecium
W= INF (n = 19) 1 (53) 3 (158) 1 (53) 6 (316) 1 (53
W-BR (n = 36) 1 (28) 2 (56) 1 (28) 12 (332) 3 (83
W-EFF (n = 53) 3 (57) 2 (38) 4 (75) 23 (434) 10 (18
W-MOut (n = 29) 0 6 (207) 6 (207) 13 (448) 4 (138
D-EFF (n = 41) 1 (24) 6 (146) 5 (122) 13 (317) 2 (49
D-MOut (n = 40) 1 (25) 2 (5) 2 (5) 14 (350) 3 (75
VR (n = 26) 0 1 (38) 0 8 (308) 1 (38
All (n = 244) 7 (29) 22 (90) 19 (78) 90 (369) 24 (98
E faecalis
W-INF (n = 5) 0 1 (200) 0 1 (200) 0
W-BR (n = 8) 2 (250) 2 (250) 1 (125) 2 (250) 2 (250
W-EFF (n = 19) 0 1 (53) 1 (53) 3 (158) 3 (158
W-MOut (n = 4) 0 0 0 0 0
D-EFF (n = 19) 2 (105) 5 (263) 0 1 (53) 0
D-MOut (n = 3) 1 (333) 1 (333) 0 0 0
VR (n = 5) 0 0 0 2 (400) 0
All (n = 63) 5 (79) 10 (159) 2 (32) 9 (143) 5 (79
GM gentamicin ST streptomycin AM ampicillin CIP ciprofloxacin LVX levofloxaciall tested antimicrobial agents MDR multiple drug resistance na not applicableAbbreviations of sampling pointsrsquo locations and their description provided in Figure
E faecium varied between 308 in Vistula River mouth to448 in W-MOut while resistance to levofloxacin did notexceed 20 Among E faecalis resistance to ciprofloxacinand levofloxacin was in general lower then reported for Efaecium in the corresponding sampling points (up to250) except Vistula River mouth The high-level strepto-mycin resistance (HLSR) was prevalent and reached 207for E faecium isolates from marine outflow of WWTPGdansk-Wschod (W-MOut) Also for E faecalis HLSRphenotype was detected among up to 16 of all tested iso-lates Resistance to glycopeptides linezolid and daptomycinwas not detected among all tested enterococci
Clonal structure of E faecium isolates and distribution ofvirulence resistance and MGE genesSixty-nine isolates of E faecium (283 of all isolates ofthis species) were further subjected to MLST and ana-lysis of the presence of resistance virulence and MGEgenes These isolates presented various phenotypes of re-sistance to antimicrobial compounds as described belowand were obtained mostly from effluents and marine out-falls of both WWTPs (Table 3) MLST discerned 54 differ-ent STs (Figure 2) including 23 new ones (STs 632ndash654)The comparative eBURST analysis against the wholeMLST E faecium database (as of the 5th April 2013)
ber () of isolates
icrobial agent Sensitive MDRphenotypeTE SYN E S
n = 244
) 5 (263) 6 (316) 7 (368) 6 (316) 4 (211)
) 7 (194) 19 (528) 16 (444) 10 (278) 8 (222)
9) 10 (189) 18 (340) 24 (453) 12 (226) 14 (264)
) 8 (276) 14 (483) 17 (586) 5 (174) 9 (310)
) 10 (244) 12 (292) 28 (682) 4 (98) 8 (195)
) 2 (50) 21 (525) 22 (550) 7 (175) 3 (75)
) 2 (77) 15 (577) 11 (424) 3 (115) 3 (115)
) 44 (180) 105 (430) 125 (512) 49 (201) 47 (193)
n = 63
1 (200) na 1 (200) 4 (800) 0
) 2 (250) na 7 (875) 1 (125) 2 (250)
) 4 (211) na 13 (684) 0 3 (158)
0 na 4 (100) 0 0
9 (474) na 13 (684) 4 (211) 5 (263)
2 (667) na 2 (667) 0 1 (333)
0 na 1 (200) 2 (400) 0
) 18 (286) na 41 (651) 11 (175) 11 (175)
n TE tetracycline SYN quinupristin-dalfopristin E erythromycin S sensitive to
1
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium
LineageCCST
MT Isolationsite
Virulence genes Resistance determinants Transposongenes
Plasmid rep genes Plasmid TAS
espintA
fms5 fms17 fms19 fms21 pbp5 GyrA ParC aac(6prime)-Ie-aph(2Prime)
ant(6prime)-Ia
tet(M)
tet(L)
tet(S)
intTn916 tndX rep1pIP501
rep2pRE25
rep17pRUM
rep18pEF418
reppLG1
reppMG1
axe-txe
ω-ε-ζ
relBE
Lineage 17
17 (2) 1 8 W-EFF 2 2 2 2 2 pbp5-30
S84R R61GS80R
1 1 2 2 1 2 2 1 1
386 7 W-MOut 1 1 1 1 1 1 1 1 1 1 1 1 1
Lineage 18
18 (3) 1(3)
D-EFFW-MOut
3 3 3 1 pbp5-20
S84Y R61GS80I
1 2 3 3 1 2 1 3
262 10 W-EFF 1 1 1 pbp5-25
S84Y R61G 1 1 1 1 1 1 1
574 402 VR 1 1 1 1
Lineage 78
78 (4) 159(4)
W-EFFW-MOut
4 4 4 4 4 pbp5-28
S84RY
R61GS80R
2 1 2 2 1 4 4 2 4 4 3
266 (2) 139(2)
W-INFD-EFF
2 1 2 2 pbp5-24
1 1 2 1 1 2 2 2
323 12 D-MOut 1 1 1 1 pbp5-26
wt R61GS80I
1 1 1 1 1 1 1 1
564 302 D-EFF 1 1 1 1 pbp5-29
S84I R61GS80R
1 1 1 1 1 1 1
653 302 W-EFF 1 nd S84I R61GS80R
1 1 1
CC5
29 (2) W-EFFW-MOut
2 1 1 2 1
66 W-MOut 1 1 1 1 1 1 1
97 W-EFF 1 wt R61GS80R
1 1
123 W-EFF 1 1 wt R61G 1 1 1
148 W-MOut 1 pbp5-30
wt R61GS80R
1 1 1 1 1
168 W-MOut 1 1 pbp5-31
1 1 1 1 1 1 1 1 1
505 W-EFF 1 wt R61G 1 1
588 D-MOut 1 pbp5-30
1 1 1 1 1 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
5of
15httpw
wwbiom
edcentralcom1471-21801466
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium (Continued)
635 D-EFF 1 1 1 1
636 W-EFF 1 wt R61G 1
637 W-EFF 1 1 1 wt R61G 1 1
638 W-EFF 1 wt R61GS80I
1
642 W-EFF 1 1 1 1 1
645 D-EFF 1 1 1 1 1
646 W-EFF 1 1 1 1 1
649 W-BR 1 1 1 pbp5-32
wt R61G 1
650 (3) D-EFF 3 3 pbp5-27
2 2 2 1 1 3
CC 9
8 D-EFF 1 1 1 1 1 1 1 1 1 1 1
640 W-EFF 1 1 1 1 1
CC 22
21 W-EFF 1 1 1 1 1 1 1 1
22 W-EFF 1 1 1 1 1 1
25 W-EFF 1 1 pbp5-32
wt R61G 1 1 1
32 (3) W-EFF 3 3 1 wt R61G 2 1
92 W-EFF 1 1 1 1 1
441 W-EFF 1 1 1 1 1 1 1 1
533 W-EFF 1 1 1
633 W-EFF 1 1 1 1 1
644 D-EFF 1 1 1 1 1
CC27
652 W-EFF 1 1 1 1
CC46
47 VR 1 1 1 1
69 D-EFF 1 1 1 1 1 1
CC94
94 W-MOut 1 1 1 1
361 (4) D-EFFD-MOut
3 4 4 4 pbp5-29
1 1 1 4 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
6of
15httpw
wwbiom
edcentralcom1471-21801466
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium (Continued)
CC296
296 W-EFF 1 1
CC639
639 W-EFF 1 1 1 1 1
CC648
634 W-EFF 1 1 1 1 1
648 W-EFF 1 1 1 1 1
Doubletons
632 W-MOut 1 1 1
Singletons
264 W-INF 1 1 1 1 1 1 1
641 D-EFF 1 1 1 1
643 W-EFF 1 1 1 1 1 1
647 W-MOut 1 1 1 1 1
651 D-EFF 1 1 1 1 1 1
654 VR 1 1 1 1 1 1
All (69) 8 57 49 29 46 6 17 20 17 1 9 4 39 27 14 7 55 2 12 10 5
CC clonal complex ST sequence type MT MLVA-type nd not determined due to lack of amplification of the pbp5 gene wt wild-type STs associated with MDR phenotype in boldAbbreviations of sampling pointsrsquo locations and their description provided in Figure 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
7of
15httpw
wwbiom
edcentralcom1471-21801466
Figure 2 MLST-based similarity tree of E faecium isolates constructed using the START software and the UPGMA clustering algorithm
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 8 of 15httpwwwbiomedcentralcom1471-21801466
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 9 of 15httpwwwbiomedcentralcom1471-21801466
grouped the 38 STs specific for 50 isolates into the majorlsquosuper-complex 17rsquo characteristic for this species (data notshown) Further detailed analysis of the localization ofparticular STs within this structure revealed that 11 STscharacteristic for 17 isolates belonged to the nosocomialcomplex of E faecium representing all three majorlineages 17 18 and 78 of a former CC17 as defined byWillems et al [9] (Table 3) Seventeen STs (20 isolates)nine STs (11 isolates) and two STs with single isolates eachbelonged to CC5 CC22 and CC9 respectively as definedby Freitas et al [24] Of the remaining 16 STs nine wereincluded into six small CCs (CCs 27 46 94 296 639648) unlinked to the major lsquosuper-complexrsquo ST632formed a lsquodoubletonrsquo (ie was linked by a single-locus vari-ant link to only one ST from the database) and six STsrepresented by single isolates were singletons ie werenot related to any ST in the database at the time of ana-lysis Thirty isolates (435) in the investigated groupshowed the MDR phenotype which was significantly asso-ciated (p = 000006) with isolates from the nosocomialcomplex (Table 3 and Figure 3) but found also in CC5CC22 and CC94 this phenotype did not occur amongsmall CCs or singletons Seventeen isolates repre-senting the nosocomial complex were additionallystudied by multilocus VNTR (variable number tan-dem repeats) analysis (MLVA) to better elucidate therelationships among these isolates Altogether nineMLVA-types (MTs) were found (Table 3) Importantlythe same MLVA-type MT159 associated with ST78was characteristic for isolates derived from samplesfrom W-EFF and W-MOut suggesting survival of thisparticular clone
0
10
20
30
40
50
60
70
80
90
100
IS16
espintA
fms5
fms1
7
fms1
9
fms2
1
MDR
AM-R
CIP-R
HLG
R
p
ositi
ve
isol
ates
Figure 3 Distribution () of IS16 resistance phenotypes and virulenccolumns isolates with STs typical for nosocomial E faecium grey columnsgroups are indicated by an asterisk MDR multi-drug resistance AM-R ampgentamicin resistance HLSR high-level streptomycin resistance
IS16 was present in all 17 isolates included by MLST tothree lineages of nosocomial E faecium and this markerwas limited to this group (Table 3 and Figure 3) TheespEfm gene occurred always together with intA among allisolates of lineage 17 and in some isolates of lineage 78The fms19 pilin gene detected in 29 isolates was over-represented among STs typical for nosocomial E faeciumwhile other pilin genes such as fms5 fms17 and fms21detected in 57 49 and 46 isolates respectively occurredamong various CCs and singleton STs Ampicillin resist-ance significantly associated with isolates of nosocomialcomplex (10 of 17 isolates 588 p = 0001) occurred alsoin CC5 (6 isolates) CC22 and CC94 (1 isolate each) Se-quencing of the 3prime part of the pbp5 gene revealed its 11nucleotide alleles corresponding to 11 variants of the C-terminal part of PBP5 Six alleles of the gene were new(pbp5-24 to pbp5-29) two alleles (pbp5-20 pbp5-22)were described previously among Polish hospital VREmisolates [13] and three alleles were found in other stud-ies pbp5-30 occurred in a Portuguese animal strain ofE faecium [25] and pbp5-31 and pbp5-32 in clinical iso-lates TCGE70411_2 and 164306 also from Portugal (de-posited as JN208888 and JN208885 in the GenBankrespectively) For a single ampicillin-resistant isolate ofST653 (lineage 78) pbp5 amplification was not achieveddespite repeated attempts The majority (11 of 17 iso-lates 647) of isolates belonging to three lineages ofnosocomial E faecium showed resistance to ciprofloxa-cin determined by mutations R61G and S80IR in ParCand S84IRY in GyrA Ciprofloxacin resistance wasalso significantly over-represented among isolates ofSTs typical for nosocomial E faecium (p = 00006) Nine
HLS
R
tetM tetL
rep1
-pIP50
1
rep2
-pRE25
rep1
7-pR
UM
rep1
8-pE
F418
rep-pL
G1
axe-txe
relBE
e resistance and plasmid genes among E faecium isolates Blackother E faecium isolates Significant differences (p lt 005) between twoicillin resistance CIP-R ciprofloxacin resistance HLGR high-level
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 10 of 15httpwwwbiomedcentralcom1471-21801466
isolates resistant to ciprofloxacin were found also inCCs 5 9 and 22 These isolates possessed mutations inParC mentioned above but had the wild-type QRDR inGyrA HLGR occurred in five isolates of lineages 18 and78 and in single isolates of CC5 and CC22 this pheno-type was determined by the presence of aac(6prime)-Ie-aph(2Prime) with an exception of an isolate from CC5 whichwas negative for this gene High-level resistance tostreptomycin (HLSR) was widely distributed amongstudied isolates and determined by the ant(6prime)-Ia geneTetracycline resistance was found also in various STsthe major determinant of this phenotype was tet(M) (20isolates) followed by tet(L) (17 isolates) These twogenes occurred together in 16 isolates A single isolateof ST633 belonging to CC22 harbored tet(S) The tet(O) gene was absent in the studied group For a singletetracycline-resistant isolate of ST17 none of the fourtetracycline determinants studied was detected Nineisolates carried int of Tn916 and four the tndX gene ofTn5397 These transposon-specific genes were not char-acteristic for any lineage or complex (Table 3) Plasmid-replication genes such as rep2pRE25 rep17pRUM andrep18 pEF418 (26 14 and 7 isolates respectively) andgenes specifying toxin-antitoxin systems (TAS) axe-txeand ω-ε-ζ (12 and 10 isolates respectively) were signifi-cantly associated with the group of isolates of nosocomialE faecium Two other rep genes (rep1pIP501 reppLG1) werewidely distributed among all studied E faecium isolatesand the reppMG1 gene occurred in two isolates (in a singleisolate of ST533 from CC22 and in a singleton ST647) Allω-ε-ζ-positive isolates carried rep2pRE25 and nine isolatesshowed a concomitant presence of axe-txe and rep17pRUMThe relBE genes were found solely among five non-nosocomial E faecium (a difference in distribution with-out statistical significance p = 043) among various STs
DiscussionIn this study Enterococcus spp was used to evaluate theimpact posed by treated wastewater of two local WWTPson their receiver the costal waters of Gulf of GdanskTotal counts of enterococci observed for raw (W-INF)and treated (W-EFF D-EFF) wastewater are consistentwith our previous study showing similar over 99 reduc-tion capacity of applied treatment processes It is alsoreported in the literature that wastewater treatment pro-cesses based on activated sludge although they focus onremoval of nutrients and organic matter simultaneouslyreduce faecal indicators with high efficiency reaching upto 999 [1822] Due to the high number of enterococciin raw wastewater they are nevertheless released from aWWTP in a number up to 61x105 CFU per 100 mLThus marine water sampled in the area directly impactedby treated wastewater (W-MOut and D-MOut) showed ahigher number of enterococci than the one observed in
Vistula River mouth however lower by four orders ofmagnitude in comparison to corresponding treated waste-water (W-EFF and D-EFF) It should be noted that in mar-ine outfalls (W-MOut D-MOut) as well as in VistulaRiver mouth enterococci were detected in number lowerthan 100 CFU per 100 mL ie the value set out by NewBathing Water Directive 20067EC for the costal water ofldquoexcellent qualityrdquo [23]It is suggested that enterococcal species composition in
faecal-impacted environment depends from the source ofcontamination In this study two species most commonlycolonizing humans E faecalis and E faecium comprisedtogether from 73 to 88 of enterococcal isolates intested wastewater samples (W-INF W-BR W-EFF and D-EFF) 73 and 61 in marine outflows (W-MOut and D-MOut respectively) and 45 in the Vistula River mouthThe predominant species in wastewater and marine waterimpacted by treated wastewater was E faecium (from 56to 65) which stayed at a similar level during treatmentprocesses as determined also in our previous study [20]Species distribution in wastewater samples is not wellunderstood and affecting factors may include differencesin diet climate season and methods of detection E feaca-lis was found to be the most prevalent species in wastewa-ter samples in Sweden [26] E faecium in Spain UKCanada France and Switzerland [26-29] while E hiraepredominated in Portugal and United States [2230] Al-though E faecalis is typically the most abundant colonizerof humans in the community (eg [2631]) E faecium isgenerally most often associated with human microbialcontamination [2729] E faecalis was reported fromagriculture-impacted waters [32] and E hirae likelyoriginates mainly from cattle and pig faecal contamin-ation [26] which explains high relative proportion ofthese two species in Vistula River mouth Apart from thevariability of contamination sources proportion of par-ticular species may differ due to the differences of abilityto sustain environmental stresses In this study enterococ-cal composition might have been influenced by wastewa-ter treatment processes and marine water characteristicsThe effect of wastewater treatment and marine environ-
ment was seen on phenotypes of antimicrobial resistanceThe positive selection of bacteria with resistance patternshas been already suggested and noted in wastewater pro-cesses [192022] In this study the resistance rates to ampi-cillin ciprofloxacin levofloxacin and erythromycin notedfor E faecium from raw wastewater (W-INF) were lowerthan these observed for this species in the correspondingtreated wastewater (W-EFF) however not statistically sig-nificant (p gt 005) Moreover resistance rates to ampicillinciprofloxacin tetracycline and erythromycin observed inmarine outflow of WWTP Gdansk-Wschod (W-MOut)were higher than in treated wastewater of this plantSuch phenomenon was not observed for treated
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 11 of 15httpwwwbiomedcentralcom1471-21801466
wastewater of WWTP Gdynia-Debogorze (D-EFF) andits marine outfall (D-MOut) Resistance rates among Efaecium originating from D-MOut were in generalcomparable to these detected in the Vistula Rivermouth It should be noted that treated wastewater fromWWTP Gdansk-Wschod has been discharged via mar-ine outfall (W-MOut) for last 12 years while marineoutfall of WWTP Gdynia-Debogorze (D-MOut) hasbeen operating for last two years The observed differ-ences in resistance rate corresponding to the local im-pact caused by long and short-term operated marineoutfalls (W-MOut and D-MOut) need further attentionSignificant rates of antimicrobial resistance observed
for isolates of E faecalis from wastewater and marineriver waters impacted by wastewater are in agreementwith a recent study performed in six European countriesshowing that in Poland the rates of resistance for humanisolates of this species were usually among the highestboth in the community and hospitals [33] To our know-ledge no corresponding data are available for E faeciumcolonization in the community in Poland but a studyfocused on hospital VRE isolates showed their ubiquitousMDR phenotype and a high load of resistance genes [13]In our study no vancomycin resistance were detectedlikely due to relatively low (below 10) prevalence of VREin Polish hospitals [5] although such isolates are isolatedfrom the environment in other countries eg [34]This study shows that hospital clones of E faecium are
important carriers of resistance genes in wastewatersMost likely in a significant part they originated fromhospital wastewater treated together with communalwastewater In our estimation hospital wastewater con-stitutes approximately 02 of both WWTPsrsquo daily in-flow The release of hospital-associated clones of Efaecium to the environment was observed also by others[24283536] In our study isolates belonging to themajor nosocomial HiRECC (formerly named CC17)constituted altogether 246 of all isolates of E faeciumsubjected to typing and were present at all samplingsites These isolates apart from their inclusion into theHiRECC by MLST demonstrated several other pheno-typic and genotypic features typical for nosocomial Efaecium ie they were resistant to ciprofloxacin andampicillin in vast majority showed MDR phenotypeand carried several resistance determinants associatedboth with mutations of chromosomal genes (gyrA parCpbp5) and in the case of aminoglycoside and tetracyclineresistance with gene acquisition Plasmids likely played animportant role in the later process as some plasmid genessuch as rep2pRE25 rep17pRUM and rep18 pEF418 and TASgenes axe-txe and ω-ε-ζ were significantly more abundantin isolates associated with HiRECC than among theremaining isolates As yet the knowledge of distributionof plasmid-associated genes in populations of E faecium
especially in the context of their clonal compositionremains limited A study on 93 isolates of differentgeographical origins and from various sites showed anover-representation of rep17pRUM and reppMG1 amongnosocomial E faecium while in contrast to our observa-tions rep18 pEF418 was not detected [37] Moreover wedid not observe reppMG1 among HiRECC of E faeciumThese differences may be due to a high prevalence of VRE(58 isolates) among 93 isolates studied by Rosvoll et alwhat may explain differences in the plasmid content Thesame study also reported the co-localization of ω-ε-ζ withrep2pRE25 and axe-txe with rep17pRUM on the same plas-mids in concordance with our findings showing a fre-quent joint occurrence of these pairs of genes Anotherstudy performed on 99 invasive isolates from Norwaymostly of HLGR phenotype showed a significantly higherprevalence of rep2pRE25 and rep17pRUM among the majornosocomial HiRECC [38] This study as well some others[2439] also demonstrated a common presence of reppLG1among E faecium from hospital settings and in animalsOur findings indicate that this type of replicon is evenmore ubiquitous in E faecium than previously reportedAll rep genes mentioned above are typically associatedwith plasmids carrying various antimicrobial resistancegenes [3940] GenBank accession number AF408195 Inaddition to genes determining antimicrobial resistanceand genes associated with MGE isolates of nosocomialHiRECC analyzed in our study carried such molecularmarkers as IS16 and the esp gene together with the intAgene characteristic for ICEEfm [4142] Another patho-genicity factor the fms19 gene of the pilin gene cluster4 (PGC4) showed increased prevalence among theseisolates Similar observation was made for a representa-tive collection of 433 E faecium isolates from varioussources [43]In a study that followed enterococci in a continuum
from hospital and retirement home wastewaters througha WWTP to a river a gradual decrease in the proportionof E faecium isolates representing hospital-associatedSTs was observed likely due to the ldquodilutionrdquo of HiRECCrepresentatives by more diverse and less drug-resistantstrains originating from the community [28] Because inour study only selected representatives of obtained iso-lates were investigated it is not possible to determineprecise tendencies in their frequency from the WWTPinfluent to effluent and water however resistant entero-cocci both HiRECC and non-HiRECC are clearly ableto survive the treatment process what results in their re-lease into marine and river waters Moreover using anadditional typing method MLVA we were able to findisolates with the same characteristics in the effluent ofWWTP and its marine outfall suggesting ability of suchclones to survive in the environment for at least sometime It seems plausible that hospital resistant clones
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 12 of 15httpwwwbiomedcentralcom1471-21801466
accumulate several determinants that may promote theirsurvival not only in the hospital settings but also in theenvironment Such features would include resistance todisinfectants and ultraviolet light additional metabolicpathways and ability to form biofilms Moreover determi-nants of some of the mentioned adaptation factors arefound co-resident in plasmids harboring resistance genessuggesting co-selection For example pLG1-type plasmidsof E faecium carry not only resistance determinantsagainst aminoglycosides glycopeptides and macrolidesbut also pili genes and carbon uptake-utilization genes[3839] Pheromone-responsive plasmids of E faecalisapart from resistance to aminoglycosides glycopeptidespenicillins and macrolides may encode bacteriocins add-itional pili and UV-resistance determinants [40] Thusnosocomial HiRECCs released by WWTPs are equippedin whole sets of genes with various adaptive benefits andoften located within MGE It was also demonstrated thatMGEs can be transferred in a WWTP [44] further under-ling the importance of threat posed by such organismsThere is a selection pressure for survival of resistantstrains and their tranconjugants due to release of antibi-otics to the environment especially in the hospital efflu-ents [45] and even sub-inhibitory concentrations may beeffective in selection for resistance [46]
ConclusionsThis study describes the species composition resistanceprofiles and clonal relationships of enterococci isolatedfrom WWTPs located in Gdansk their marine outfallsto the Gulf of Gdansk and from Vistula River It was ob-served that these WWTPs release enterococcal strainsresistant to various antimicrobial compounds to the re-cipient water environment Molecular analyses revealedthe presence of clones associated with nosocomial HiR-ECCs and in the case of E faecium isolates of lineages17 18 and 78 of former CC17 abundant in resistance de-terminants MGE genes and pathogenicity factors Thepresence of HiRECCs in wastewaters and marineriverenvironment highlights the need for further detailedanalyses to better understand the survival and spread ofdrug-resistant strains in water ecosystems and to eluci-date the ways to curb such dissemination
MethodsAcquisition of samples and isolation of enterococciOnce a month in April June August and October in 2011of the flow proportioned composited samples of wastewa-ter were taken from two local WWTPs Gdansk-Wschodfrom influent (W-INF) effluent (W-EFF) and wastewa-ter from activated sludge bioreactor (W-BR) and fromGdynia-Debogorze effluent (D-EFF) The two WWTPstreat sewage from approximately 920 000 inhabitants(for details see Figure 1) In the same period of time and
with the same frequency marine outfalls of those WWTPs(W-MOut and D-MOut respectively) as well as VistulaRiver mouth were also sampled for isolation of entero-cocci Marine samples were taken at the depth of approxi-mately one meter below the surface once a month inApril June August and October of 2011 Location ofsampling points and their characteristics are provided inFigure 1 Enterococci were detected using the membranefiltration method To this end appropriate dilutions ofanalyzed samples were filtered in duplicate through045 μm cellulose-acetate filters (EMD Millipore Cor-poration Billerica MA USA) which were then placedon Enterococcus selective agar (Merck DarmstadtGermany) and incubated at 37degC for 48 h (ISO 7899ndash22000) Dark red or maroon colonies were consideredpresumptive enterococci For further investigationsrepresentative isolates were taken from membranes pre-senting from 20 to 50 typical colonies and stored in nu-trient broth supplemented with 15 glycerol at minus80degC
Species identification DNA isolation susceptibility testingand statistical analysisThe species identification and drug susceptibility of pre-sumptive enterococci were determined by the PhoenixtradeAutomated Microbiology System (BD New Jersey USA)according to the manufacturerrsquos instructions Since it isnot possible to distinguish E casseliflavus and E galli-narum using this system the classification E casselifla-vusgallinarum was used The susceptibility tests basedon microdilution were carried out for 11 antimicrobialagents gentamicin (GM) streptomycin (ST) ampicillin(AM) daptomycin (DAP) vancomycin (VA) teicoplanin(TEC) linezolid (LZD) fluoroquinolones (ciprofloxacin -CIP levofloxacin - LVX) tetracycline (TE) erythromycin(E) and quinupristin-dalfopristin (SYN) (E faecium only)Obtained minimal inhibitory concentrations (MIC) wereevaluated according to the guidelines of Clinical and La-boratory Standards Institute [47] Isolates were defined asmultidrug-resistant (MDR) when they showed resistanceto three or more compounds tested [48] Bacterial DNAwas purified using the Genomic DNA Prep Plus kitfollowing the manufacturerrsquos instructions (AampA Biotech-nology Gdynia Poland) Differences in distributions wereevaluated using the χ2 test with the p values lt 005 con-sidered significant
MLST and MLVA of E faeciumMLST for E faecium was performed by sequencing ofseven house-keeping genes as described by Homan et al[49] The allele numbers and STs were assigned with theuse of Internet database httpefaeciummlstnet (5th July2013 date last accessed) Novel alleles and allelic profileswere submitted to the database MLST data were analyzedwith Sequence Type Analysis and Recombinational Tests
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 13 of 15httpwwwbiomedcentralcom1471-21801466
(START) software [50] and the eBURST analysis [51] wasused to elucidate the relationships of STs with knownCCs using the comparative eBURST option available athttpeburstmlstnetv3enter_datacomparative (5th July2013 date last accessed) Further inclusion of E faeciumSTs into specific CCs and lineages was performed fol-lowing other studies [924] MLVA was performed as de-scribed by others [52]
Detection and analysis of genes associated with virulenceantimicrobial resistance and MGE in E faeciumThe full list of primers used for gene detection and se-quencing is provided in Additional file 1 Table S1 PCR-based detection of IS16 virulence-associated genes (fms21fms17 fms5 and fms19 representing each of four pilingene clusters in E faecium and the espEfm gene) genes de-termining resistance to tetracycline tet(M) tet(O) tet(L)tet(S) HLGR (aac(6prime)-Ie-aph(2Prime) HLSR (ant(6prime)-Ia) andgenes specific for MGE such as intA of ICEEfm1 int ofTn916 transposon tndX of Tn5397 transposon entero-coccal plasmid replication genes rep1pIP501 rep2pRE25rep17pRUM rep18pEF418 reppMG1 reppLG1 and TAS genesaxe-txe ω-ε-ζ relBE was performed as described in otherstudies [134153-59] following the conditions describedherein Previously characterized clinical isolates of Strepto-coccus agalactiae E faecium and E faecalis harbouringpilin genes IS16 espEfm intA of ICEEfm1 int of Tn916tndX of Tn5397 tet(M) tet(O) tet(L) tet(S) aac(6prime)-Ie-aph(2Prime) ant(6prime)-Ia rep1pIP501 rep2pRE25 ω-ε-ζ [136061]served as positive controls In cases when positive controlswere not available (rep18pEF418 reppMG1 reppLG1 axe-txerelBE) a few randomly selected PCR products wereverified by sequencing and served further as controlsAmpicillin-resistance determinant the pbp5 gene was ana-lysed by sequencing of the part encoding C-terminaltranspeptidase domain in PBP5 [62] New sequence vari-ants pbp5-24 to pbp5-29 alleles were submitted to theGenBank (acc numbers KC594860-KC594865 respect-ively) The quinolone-determining regions (QRDR) in thegyrA and parC genes were sequenced and analysed asdescribed by others [63]
Additional file
Additional file 1 Table S1 Primers used in the study for PCR andsequencing
Competing interestThe authors declare that they have no competing interest
Authorsrsquo contributionsES participated in the study design performed experimental work andparticipated in the manuscript preparation AL designed the studyperformed experimental work and participated in the manuscriptpreparation Both authors read and approved the final manuscript
AcknowledgementsWe thank Rob Willems and Janetta Top for assigning new alleles new STsand MTs This publication made use of the Enterococcus faecium MLSTwebsite (httpefaeciummlstnet) which is hosted at Imperial College of theUniversity of Oxford and funded by the Wellcome Trust This work wassupported by project N N305 461139 from the Narodowe Centrum Nauki(NCN)
Author details1National Medicines Institute ul Chelmska 3034 Warsaw 00-725 Poland2Gdansk University of Technology ul G Narutowicza 1112 Gdansk 80-952Poland
Received 5 September 2013 Accepted 27 February 2014Published 14 March 2014
References1 Aarestrup FM Butaye P Witte W Nonhuman reservoirs of Enterococci In
American Society for Microbiology Edited by Gilmore MS Clewell DB CourvalinP Dunny GM Murray BE Rice LB Washington DC The Enterococcipathogenesis molecular biology and antibiotic resistance 2002
2 Tendolkar PM Baghdayan AS Shankar N Pathogenic enterococci newdevelopments in the 21st century Cell Mol Life Sci 2003 602622ndash2636
3 Sood S Malhotra M Das BK Kapil A Enterococcal infections ampantimicrobial resistance Indian J Med Res 2008 128111ndash121
4 Sydnor ER Perl TM Hospital epidemiology and infection control inacute-care settings Clin Microbiol Rev 2011 24141ndash173
5 European Centre for Disease Prevention and Control Annual EpidemiologicalReport Reporting on 2010 surveillance data and 2011 epidemic intelligencedata Stockholm ECDC 20122013
6 Hidron AI Edwards JR Patel J Horan TC Sievert DM Pollock DA Fridkin SKNational Healthcare Safety Network Team Participating National HealthcareSafety Network Facilities NHSN annual update antimicrobial-resistantpathogens associated with healthcare-associated infections annualsummary of data reported to the National Healthcare Safety Network atthe Centers for Disease Control and Prevention 2006ndash2007 Infect ControlHosp Epidemiol 2008 29996ndash1011
7 De Kraker ME Jarlier V Monen JC Heuer OE van de Sande N GrundmannH The changing epidemiology of bacteraemias in Europe trends fromthe European Antimicrobial Resistance Surveillance System Clin MicrobiolInfect 2012 doi1011111469-0691
8 Leavis HL Bonten MJ Willems RJ Identification of high-risk enterococcalclonal complexes global dispersion and antibiotic resistance Curr OpinMicrobiol 2006 9454ndash460
9 Willems RJL Top J van Schaik W Leavis H Bonten M Sireacuten J Hanage WPCorander J Restricted gene flow among hospital subpopulations ofEnterococcus faecium mBio 2012 3e00151ndash12
10 Hegstad K Mikalsen T Coque TM Werner G Sundsfjord A Mobile geneticelements and their contribution to the emergence of antimicrobialresistant Enterococcus faecalis and Enterococcus faecium Clin MicrobiolInfect 2010 16541ndash554
11 Shepard BD Gilmore MS Antibiotic-resistant enterococci themechanisms and dynamics of drug introduction and resistance MicrobesInfect 2002 4215ndash224
12 Bonten MJ Willems R Weinstein RA Vancomycin-resistant enterococciwhy are they here and where do they come from Lancet Infect Dis 20011314ndash325
13 Sadowy E Sieńko A Gawryszewska I Bojarska A Malinowska K HryniewiczW High abundance and diversity of antimicrobial resistancedeterminants among early vancomycin-resistant Enterococcus faecium inPoland Eur J Clin Microbiol Infect Dis 2013 321193ndash1203
14 Willems RJ Homan W Top J van Santen-Verheuvel M Tribe D Manzioros XGaillard C Vandenbroucke-Grauls CM Mascini EM van Kregten E vanEmbden JD Bonten MJ Variant esp gene as a marker of a distinct geneticlineage of vancomycin-resistant Enterococcus faecium spreading inhospitals Lancet 2001 357853ndash855
15 Sillanpaumlauml J Nallapareddy SR Prakash VP Qin X Houmloumlk M Weinstock GMMurray BE Identification and phenotypic characterization of a secondcollagen adhesin Scm and genome-based identification and analysis of13 other predicted MSCRAMMs including four distinct pilus loci inEnterococcus faecium Microbiology 2008 1543199ndash3211
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 14 of 15httpwwwbiomedcentralcom1471-21801466
16 Galloway-Pentildea JR Nallapareddy SR Arias CA Eliopoulos GM Murray BEAnalysis of clonality and antibiotic resistance among early clinicalisolates of Enterococcus faecium in the United States J Infect Dis 20092001566ndash1573
17 Top J Willems R Bonten M Emergence of CC17 Enterococcus faeciumfrom commensal to hospital-adapted pathogen FEMS Immunol MedMicrobiol 2008 52297ndash308
18 Martins da Costa P Vaz-Pires P Bernardo F Antimicrobial resistance inEnterococcus spp isolated in inflow effluent and sludge from municipalsewage water treatment plants Water Res 2006 401735ndash1740
19 Reinthaler FF Posch J Feierl G Wust G Haas D Ruckenbauer G Mascher FMarth E Antibiotic resistance of Escherichia coli in sewage and sludgeWater Res 2003 371685ndash1690
20 Luczkiewicz A Fudala-Ksiazek S Jankowska K Olanczuk-Neyman KAntimicrobial resistance of fecal indicators in municipal wastewatertreatment plant Water Res 2010 445089ndash5097
21 Dulfour AP Ballentine RK Ambient water quality criteria for bacteria-1986 EPA4405-84-002 Washington DC US Environmental Protection Agency 1986
22 Ferreira da Silva M Tiago I Veriacutessimo A Boaventura RA Nunes OC Manaia CMAntimicrobial resistance patterns in Enterobacteriaceae isolated from anurban wastewater treatment plant FEMS Microbiol Ecol 2007 60166ndash176
23 DIRECTIVE 20067EC of the European Parliament and of the Council 15February 2006 concerning the management of bathing water quality andrepealing Directive 76160EEC httpeur-lexeuropaeuLexUriServLexUriServdouri=OJL200606400370051ENPDF
24 Freitas AR Novais C Ruiz-Garbajosa P Coque TM Peixe L Dispersion ofmultidrug-resistant Enterococcus faecium isolates belonging to majorclonal complexes in different Portuguese settings Appl Environ Microbiol2009 754904ndash4908
25 Poeta P Costa D Igrejas G Saacuteenz Y Zarazaga M Rodrigues J Torres CPolymorphisms of the pbp5 gene and correlation with ampicillinresistance in Enterococcus faecium isolates of animal origin J MedMicrobiol 2007 56236ndash240
26 Kuumlhn I Iversen A Burman LG Olsson-Liljequist B Franklin A Finn MAarestrup F Seyfarth AM Blanch AR Vilanova X Taylor H Caplin J MorenoMA Dominguez L Herrero IA Moumlllby R Comparison of enterococcalpopulations in animals humans and the environment ndash a Europeanstudy Int J Food Microbiol 2003 88133ndash145
27 Lanthier M Scott A Lapen DR Zhang Y Topp E Frequency of virulencegenes and antibiotic resistances in Enterococcus spp isolates fromwastewater and feces of domesticated mammals and birds and wildlifeCan J Microbiol 2010 56715ndash729
28 Leclercq R Oberleacute K Galopin S Cattoir V Budzinski H Petit F Changes inenterococcal populations and related antibiotic resistance along amedical center-wastewater treatment plant-river continuum Appl EnvironMicrobiol 2013 792428ndash2434
29 Thevenon F Regier N Benagli C Tonolla M Adatte T Wildi W Poteacute JCharacterization of fecal indicator bacteria in sediments cores from thelargest freshwater lake of Western Europe (Lake Geneva Switzerland)Ecotoxicol Environ Safety 2012 7850ndash56
30 Bonilla TD Nowosielski K Esiobu N McCorquodale DS Rogerson ASpecies assemblages of Enterococcus indicate potential sources offecal bacteria at a south Florida recreational beach Mar Pollut Bull2006 52807ndash810
31 Wang JT Chang SC Wang HY Chen PC Shiau YR Lauderdale TL TSARHospitals High rates of multidrug resistance in Enterococcus faecalis andE faecium isolated from inpatients and outpatients in TaiwanDiagn Microbiol Infect Dis 2013 75406ndash411
32 Lanthier M Scott A Zhang Y Cloutier M Durie D Henderson VC WilkesG Lapen DR Topp E Distribution of selected virulence genes andantibiotic resistance in Enterococcus species isolated from the SouthNation River drainage basin Ontario Canada J Appl Microbiol 2011110407ndash421
33 Kuch A Willems RJ Werner G Coque TM Hammerum AM Sundsfjord AKlare I Ruiz-Garbajosa P Simonsen GS van Luit-Asbroek M Hryniewicz WSadowy E Insight into antimicrobial susceptibility and population struc-ture of contemporary human Enterococcus faecalis isolates fromEurope J Antimicrob Chemother 2012 67551ndash558
34 Novais C Coque TM Ferreira H Sousa JC Peixe L Environmentalcontamination with vancomycin-resistant enterococci from hospitalsewage in Portugal Appl Environ Microbiol 2005 713364ndash3368
35 Iversen A Kuumlhn I Rahman M Franklin A Burman LG Olsson-Liljequist BTorell E Moumlllby R Evidence for transmission between humans and theenvironment of a nosocomial strain of Enterococcus faeciumEnviron Microbiol 2004 655ndash59
36 Rathnayake I Hargreaves M Huygens F SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterwayAustralia and associated antibiotic resistance gene profilesBMC Microbiol 2011 11201
37 Rosvoll TC Pedersen T Sletvold H Johnsen PJ Sollid JE Simonsen GSJensen LB Nielsen KM Sundsfjord A PCR-based plasmid typing inEnterococcus faecium strains reveals widely distributed pRE25-pRUM- pIP501- and pHT-related replicons associated with glycopeptideresistance and stabilizing toxin-antitoxin systems FEMS Immunol MedMicrobiol 2010 58254ndash268
38 Rosvoll TC Lindstad BL Lunde TM Hegstad K Aasnaes B Hammerum AMLester CH Simonsen GS Sundsfjord A Pedersen T Increased high-levelgentamicin resistance in invasive Enterococcus faecium is associated withaac(6prime)Ie-aph(2Prime)Ia-encoding transferable megaplasmids hosted by majorhospital-adapted lineages FEMS Immunol Med Microbiol 2012 66166ndash176
39 Laverde Gomez JA van Schaik W Freitas AR Coque TM Weaver KE FranciaMV Witte W Werner G A multiresistance megaplasmid pLG1 bearing ahylEfm genomic island in hospital Enterococcus faecium isolates Int J MedMicrobiol 2011 301165ndash175
40 Coque TM Evolutionary biology of pathogenic enterococci InEvolutionary Biology of Bacterial and Fungal Pathogens Coque TM Edited byBaquero F Nombela C Cassell GH Gutierrez JA Washington DC ASMPress501ndash523
41 Werner G Fleige C Geringer U van Schaik W Klare I Witte W IS elementIS16 as a molecular screening tool to identify hospital-associated strainsof Enterococcus faecium BMC Infect Dis 2011 1180
42 Top J Sinnige JC Majoor EA Bonten MJ Willems RJ van Schaik W Therecombinase IntA is required for excision of esp-containing ICEEfm1 inEnterococcus faecium J Bacteriol 2011 1931003ndash1006
43 Sillanpaumlauml J Prakash VP Nallapareddy SR Murray BE Distribution of genesencoding MSCRAMMs and Pili in clinical and natural populations ofEnterococcus faecium J Clin Microbiol 2009 47896ndash901
44 Marcinek H Wirth R Muscholl-Silberhorn A Gauer M Enterococcus faecalisgene transfer under natural conditions in municipal sewage watertreatment plants Appl Environ Microbiol 1998 64626ndash632
45 Kuumlmmerer K Significance of antibiotics in the environment J AntimicrobChemother 2003 525ndash7
46 Liu A Fong A Becket E Yuan J Tamae C Medrano L Maiz M Wahba C LeeC Lee K Tran KP Yang H Hoffman RM Salih A Miller JHP Selectiveadvantage of resistant strains at trace levels of antibiotics a simple andultrasensitive color test for detection of antibiotics and genotoxicagents Antimicrob Agents Chemother 2011 551204ndash1210
47 Clinical and Laboratory Standards Institute Performance standards forantimicrobial susceptibility testing 21st informational supplement Wayne CLSI2011 PA M100-S20
48 Magiorakos AP Srinivasan A Carey RB Carmeli Y Falagas ME Giske CGHarbarth S Hindler JF Kahlmeter G Olsson-Liljequist B Paterson DL Rice LBStelling J Struelens MJ Vatopoulos A Weber JT Monnet DLMultidrug-resistant extensively drug-resistant and pandrug-resistantbacteria an international expert proposal for interim standarddefinitions for acquired resistance Clin Microbiol Infect 2012 18268ndash281
49 Homan WL Tribe D Poznanski S Li M Hogg G Spalburg E Van Embden JDWillems RJ Multilocus sequence typing scheme for Enterococcus faeciumJ Clin Microbiol 2002 401963ndash1971
50 Jolley KA Feil EJ Chan MS Maiden MC Sequence type analysis andrecombinational tests (START) Bioinformatics 2001 171230ndash1231
51 Feil EJ Li BC Aanensen DM Hanage WP Spratt BG eBURST inferringpatterns of evolutionary descent among clusters of related bacterialgenotypes from multilocus sequence typing data J Bacteriol 20041861518ndash1530
52 Top J Banga1 NMI Hayes R Willems RJ Bonten MJM Hayden MKComparison of multiple-locus variable-number tandem repeat analysisand pulsed-field gel electrophoresis in a setting of polyclonal endemicityof vancomycin-resistant Enterococcus faecium Clin Microbiol Infect 200814363ndash369
53 Vankerckhoven V Van Autgaerden T Vael C Lammens C Chapelle S RossiR Jabes D Goossens H Development of a multiplex PCR for the
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-
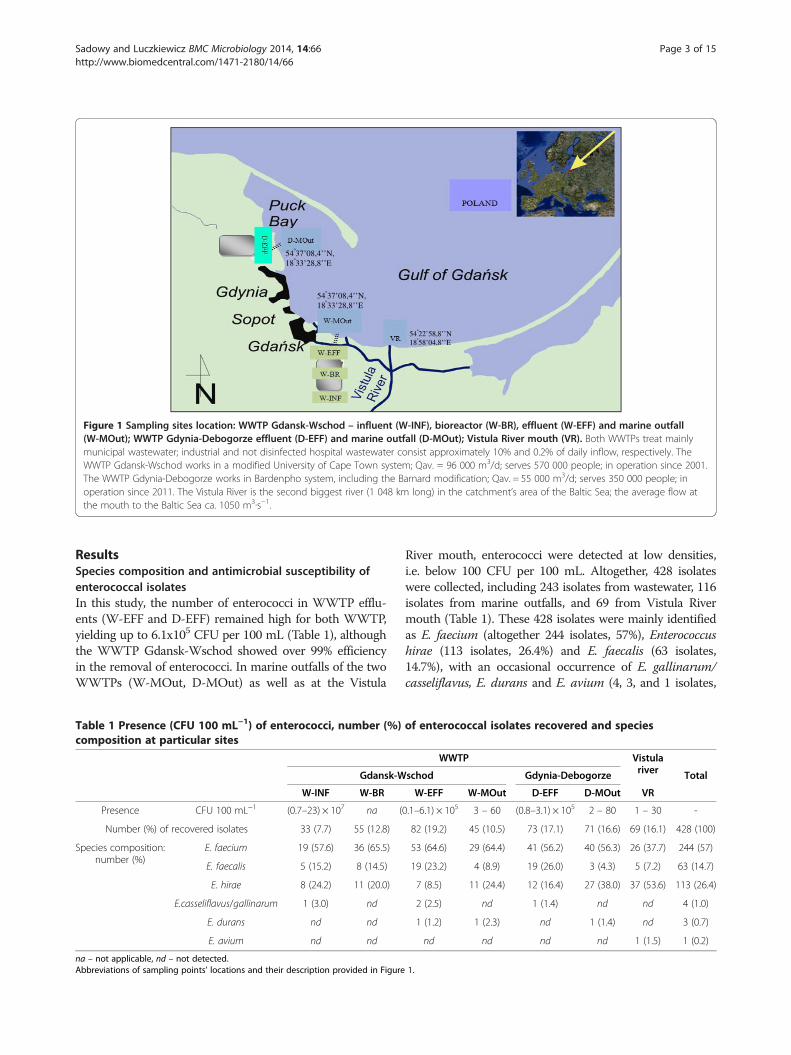
54ordm22rsquo588rsquorsquoN 18ordm58rsquo048rsquorsquoE
54ordm37rsquo084rsquorsquoN 18ordm33rsquo288rsquorsquoE
54ordm37rsquo084rsquorsquoN 18ordm33rsquo288rsquorsquoE
Figure 1 Sampling sites location WWTP Gdansk-Wschod ndash influent (W-INF) bioreactor (W-BR) effluent (W-EFF) and marine outfall(W-MOut) WWTP Gdynia-Debogorze effluent (D-EFF) and marine outfall (D-MOut) Vistula River mouth (VR) Both WWTPs treat mainlymunicipal wastewater industrial and not disinfected hospital wastewater consist approximately 10 and 02 of daily inflow respectively TheWWTP Gdansk-Wschod works in a modified University of Cape Town system Qav = 96 000 m3d serves 570 000 people in operation since 2001The WWTP Gdynia-Debogorze works in Bardenpho system including the Barnard modification Qav = 55 000 m3d serves 350 000 people inoperation since 2011 The Vistula River is the second biggest river (1 048 km long) in the catchmentrsquos area of the Baltic Sea the average flow atthe mouth to the Baltic Sea ca 1050 m3sminus1
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 3 of 15httpwwwbiomedcentralcom1471-21801466
ResultsSpecies composition and antimicrobial susceptibility ofenterococcal isolatesIn this study the number of enterococci in WWTP efflu-ents (W-EFF and D-EFF) remained high for both WWTPyielding up to 61x105 CFU per 100 mL (Table 1) althoughthe WWTP Gdansk-Wschod showed over 99 efficiencyin the removal of enterococci In marine outfalls of the twoWWTPs (W-MOut D-MOut) as well as at the Vistula
Table 1 Presence (CFU 100 mLminus1) of enterococci number ()composition at particular sites
Gdansk-W
W-INF W-BR
Presence CFU 100 mLminus1 (07ndash23) times 107 na (
Number () of recovered isolates 33 (77) 55 (128)
Species compositionnumber ()
E faecium 19 (576) 36 (655)
E faecalis 5 (152) 8 (145)
E hirae 8 (242) 11 (200)
Ecasseliflavusgallinarum 1 (30) nd
E durans nd nd
E avium nd nd
na ndash not applicable nd ndash not detectedAbbreviations of sampling pointsrsquo locations and their description provided in Figure
River mouth enterococci were detected at low densitiesie below 100 CFU per 100 mL Altogether 428 isolateswere collected including 243 isolates from wastewater 116isolates from marine outfalls and 69 from Vistula Rivermouth (Table 1) These 428 isolates were mainly identifiedas E faecium (altogether 244 isolates 57) Enterococcushirae (113 isolates 264) and E faecalis (63 isolates147) with an occasional occurrence of E gallinarumcasseliflavus E durans and E avium (4 3 and 1 isolates
of enterococcal isolates recovered and species
WWTP Vistulariver Totalschod Gdynia-Debogorze
W-EFF W-MOut D-EFF D-MOut VR
01ndash61) times 105 3 ndash 60 (08ndash31) times 105 2 ndash 80 1 ndash 30 -
82 (192) 45 (105) 73 (171) 71 (166) 69 (161) 428 (100)
53 (646) 29 (644) 41 (562) 40 (563) 26 (377) 244 (57)
19 (232) 4 (89) 19 (260) 3 (43) 5 (72) 63 (147)
7 (85) 11 (244) 12 (164) 27 (380) 37 (536) 113 (264)
2 (25) nd 1 (14) nd nd 4 (10)
1 (12) 1 (23) nd 1 (14) nd 3 (07)
nd nd nd nd 1 (15) 1 (02)
1
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 4 of 15httpwwwbiomedcentralcom1471-21801466
respectively altogether 19) The most common speciesin all sampled sites was E faecium (562 ndash 655) withthe exception of Vistula River mouth where E hirae waspredominant (536 of isolates) followed by E faecalis(377)In the next step antimicrobial susceptibility was tested
for E faecium and E faecalis isolates (Table 2) It shouldbe noted that at some points a low number of E faecalisisolates was recovered (fewer than 10 isolates) and thuscertain resistance rates obtained for E feacalis should betreated with caution Resistance to erythromycin was de-tected with the highest prevalence among both specieshowever rates of resistance to this compound variedover a broad range Erythromycin-resistant E faeciumcomprised 368 of isolates in raw wastewater (W-INF)453 in treated wastewater (W-EFF) and 586 inmarine outflow (W-MOut) of WWTP Gdansk-WschodSimilar tendency was seen for E faecalis isolates whereresistance rates in the sampling points listed above wereequal to 200 684 and 100 respectively For WWTPGdynia-Debogorze effluent (D-EFF) and marine outflow(D-MOut) erythromycin resistance rates for E faeciumvaried between 682 and 550 respectively while for Efaecalis were equal 684 and 667 respectively In thecase of fluoroquinolones resistance to ciprofloxacin among
Table 2 Antimicrobial resistance of E faecium and E faecalis
Num
Resistant to a single antim
GM ST AM CIP LVX
E faecium
W= INF (n = 19) 1 (53) 3 (158) 1 (53) 6 (316) 1 (53
W-BR (n = 36) 1 (28) 2 (56) 1 (28) 12 (332) 3 (83
W-EFF (n = 53) 3 (57) 2 (38) 4 (75) 23 (434) 10 (18
W-MOut (n = 29) 0 6 (207) 6 (207) 13 (448) 4 (138
D-EFF (n = 41) 1 (24) 6 (146) 5 (122) 13 (317) 2 (49
D-MOut (n = 40) 1 (25) 2 (5) 2 (5) 14 (350) 3 (75
VR (n = 26) 0 1 (38) 0 8 (308) 1 (38
All (n = 244) 7 (29) 22 (90) 19 (78) 90 (369) 24 (98
E faecalis
W-INF (n = 5) 0 1 (200) 0 1 (200) 0
W-BR (n = 8) 2 (250) 2 (250) 1 (125) 2 (250) 2 (250
W-EFF (n = 19) 0 1 (53) 1 (53) 3 (158) 3 (158
W-MOut (n = 4) 0 0 0 0 0
D-EFF (n = 19) 2 (105) 5 (263) 0 1 (53) 0
D-MOut (n = 3) 1 (333) 1 (333) 0 0 0
VR (n = 5) 0 0 0 2 (400) 0
All (n = 63) 5 (79) 10 (159) 2 (32) 9 (143) 5 (79
GM gentamicin ST streptomycin AM ampicillin CIP ciprofloxacin LVX levofloxaciall tested antimicrobial agents MDR multiple drug resistance na not applicableAbbreviations of sampling pointsrsquo locations and their description provided in Figure
E faecium varied between 308 in Vistula River mouth to448 in W-MOut while resistance to levofloxacin did notexceed 20 Among E faecalis resistance to ciprofloxacinand levofloxacin was in general lower then reported for Efaecium in the corresponding sampling points (up to250) except Vistula River mouth The high-level strepto-mycin resistance (HLSR) was prevalent and reached 207for E faecium isolates from marine outflow of WWTPGdansk-Wschod (W-MOut) Also for E faecalis HLSRphenotype was detected among up to 16 of all tested iso-lates Resistance to glycopeptides linezolid and daptomycinwas not detected among all tested enterococci
Clonal structure of E faecium isolates and distribution ofvirulence resistance and MGE genesSixty-nine isolates of E faecium (283 of all isolates ofthis species) were further subjected to MLST and ana-lysis of the presence of resistance virulence and MGEgenes These isolates presented various phenotypes of re-sistance to antimicrobial compounds as described belowand were obtained mostly from effluents and marine out-falls of both WWTPs (Table 3) MLST discerned 54 differ-ent STs (Figure 2) including 23 new ones (STs 632ndash654)The comparative eBURST analysis against the wholeMLST E faecium database (as of the 5th April 2013)
ber () of isolates
icrobial agent Sensitive MDRphenotypeTE SYN E S
n = 244
) 5 (263) 6 (316) 7 (368) 6 (316) 4 (211)
) 7 (194) 19 (528) 16 (444) 10 (278) 8 (222)
9) 10 (189) 18 (340) 24 (453) 12 (226) 14 (264)
) 8 (276) 14 (483) 17 (586) 5 (174) 9 (310)
) 10 (244) 12 (292) 28 (682) 4 (98) 8 (195)
) 2 (50) 21 (525) 22 (550) 7 (175) 3 (75)
) 2 (77) 15 (577) 11 (424) 3 (115) 3 (115)
) 44 (180) 105 (430) 125 (512) 49 (201) 47 (193)
n = 63
1 (200) na 1 (200) 4 (800) 0
) 2 (250) na 7 (875) 1 (125) 2 (250)
) 4 (211) na 13 (684) 0 3 (158)
0 na 4 (100) 0 0
9 (474) na 13 (684) 4 (211) 5 (263)
2 (667) na 2 (667) 0 1 (333)
0 na 1 (200) 2 (400) 0
) 18 (286) na 41 (651) 11 (175) 11 (175)
n TE tetracycline SYN quinupristin-dalfopristin E erythromycin S sensitive to
1
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium
LineageCCST
MT Isolationsite
Virulence genes Resistance determinants Transposongenes
Plasmid rep genes Plasmid TAS
espintA
fms5 fms17 fms19 fms21 pbp5 GyrA ParC aac(6prime)-Ie-aph(2Prime)
ant(6prime)-Ia
tet(M)
tet(L)
tet(S)
intTn916 tndX rep1pIP501
rep2pRE25
rep17pRUM
rep18pEF418
reppLG1
reppMG1
axe-txe
ω-ε-ζ
relBE
Lineage 17
17 (2) 1 8 W-EFF 2 2 2 2 2 pbp5-30
S84R R61GS80R
1 1 2 2 1 2 2 1 1
386 7 W-MOut 1 1 1 1 1 1 1 1 1 1 1 1 1
Lineage 18
18 (3) 1(3)
D-EFFW-MOut
3 3 3 1 pbp5-20
S84Y R61GS80I
1 2 3 3 1 2 1 3
262 10 W-EFF 1 1 1 pbp5-25
S84Y R61G 1 1 1 1 1 1 1
574 402 VR 1 1 1 1
Lineage 78
78 (4) 159(4)
W-EFFW-MOut
4 4 4 4 4 pbp5-28
S84RY
R61GS80R
2 1 2 2 1 4 4 2 4 4 3
266 (2) 139(2)
W-INFD-EFF
2 1 2 2 pbp5-24
1 1 2 1 1 2 2 2
323 12 D-MOut 1 1 1 1 pbp5-26
wt R61GS80I
1 1 1 1 1 1 1 1
564 302 D-EFF 1 1 1 1 pbp5-29
S84I R61GS80R
1 1 1 1 1 1 1
653 302 W-EFF 1 nd S84I R61GS80R
1 1 1
CC5
29 (2) W-EFFW-MOut
2 1 1 2 1
66 W-MOut 1 1 1 1 1 1 1
97 W-EFF 1 wt R61GS80R
1 1
123 W-EFF 1 1 wt R61G 1 1 1
148 W-MOut 1 pbp5-30
wt R61GS80R
1 1 1 1 1
168 W-MOut 1 1 pbp5-31
1 1 1 1 1 1 1 1 1
505 W-EFF 1 wt R61G 1 1
588 D-MOut 1 pbp5-30
1 1 1 1 1 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
5of
15httpw
wwbiom
edcentralcom1471-21801466
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium (Continued)
635 D-EFF 1 1 1 1
636 W-EFF 1 wt R61G 1
637 W-EFF 1 1 1 wt R61G 1 1
638 W-EFF 1 wt R61GS80I
1
642 W-EFF 1 1 1 1 1
645 D-EFF 1 1 1 1 1
646 W-EFF 1 1 1 1 1
649 W-BR 1 1 1 pbp5-32
wt R61G 1
650 (3) D-EFF 3 3 pbp5-27
2 2 2 1 1 3
CC 9
8 D-EFF 1 1 1 1 1 1 1 1 1 1 1
640 W-EFF 1 1 1 1 1
CC 22
21 W-EFF 1 1 1 1 1 1 1 1
22 W-EFF 1 1 1 1 1 1
25 W-EFF 1 1 pbp5-32
wt R61G 1 1 1
32 (3) W-EFF 3 3 1 wt R61G 2 1
92 W-EFF 1 1 1 1 1
441 W-EFF 1 1 1 1 1 1 1 1
533 W-EFF 1 1 1
633 W-EFF 1 1 1 1 1
644 D-EFF 1 1 1 1 1
CC27
652 W-EFF 1 1 1 1
CC46
47 VR 1 1 1 1
69 D-EFF 1 1 1 1 1 1
CC94
94 W-MOut 1 1 1 1
361 (4) D-EFFD-MOut
3 4 4 4 pbp5-29
1 1 1 4 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
6of
15httpw
wwbiom
edcentralcom1471-21801466
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium (Continued)
CC296
296 W-EFF 1 1
CC639
639 W-EFF 1 1 1 1 1
CC648
634 W-EFF 1 1 1 1 1
648 W-EFF 1 1 1 1 1
Doubletons
632 W-MOut 1 1 1
Singletons
264 W-INF 1 1 1 1 1 1 1
641 D-EFF 1 1 1 1
643 W-EFF 1 1 1 1 1 1
647 W-MOut 1 1 1 1 1
651 D-EFF 1 1 1 1 1 1
654 VR 1 1 1 1 1 1
All (69) 8 57 49 29 46 6 17 20 17 1 9 4 39 27 14 7 55 2 12 10 5
CC clonal complex ST sequence type MT MLVA-type nd not determined due to lack of amplification of the pbp5 gene wt wild-type STs associated with MDR phenotype in boldAbbreviations of sampling pointsrsquo locations and their description provided in Figure 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
7of
15httpw
wwbiom
edcentralcom1471-21801466
Figure 2 MLST-based similarity tree of E faecium isolates constructed using the START software and the UPGMA clustering algorithm
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 8 of 15httpwwwbiomedcentralcom1471-21801466
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 9 of 15httpwwwbiomedcentralcom1471-21801466
grouped the 38 STs specific for 50 isolates into the majorlsquosuper-complex 17rsquo characteristic for this species (data notshown) Further detailed analysis of the localization ofparticular STs within this structure revealed that 11 STscharacteristic for 17 isolates belonged to the nosocomialcomplex of E faecium representing all three majorlineages 17 18 and 78 of a former CC17 as defined byWillems et al [9] (Table 3) Seventeen STs (20 isolates)nine STs (11 isolates) and two STs with single isolates eachbelonged to CC5 CC22 and CC9 respectively as definedby Freitas et al [24] Of the remaining 16 STs nine wereincluded into six small CCs (CCs 27 46 94 296 639648) unlinked to the major lsquosuper-complexrsquo ST632formed a lsquodoubletonrsquo (ie was linked by a single-locus vari-ant link to only one ST from the database) and six STsrepresented by single isolates were singletons ie werenot related to any ST in the database at the time of ana-lysis Thirty isolates (435) in the investigated groupshowed the MDR phenotype which was significantly asso-ciated (p = 000006) with isolates from the nosocomialcomplex (Table 3 and Figure 3) but found also in CC5CC22 and CC94 this phenotype did not occur amongsmall CCs or singletons Seventeen isolates repre-senting the nosocomial complex were additionallystudied by multilocus VNTR (variable number tan-dem repeats) analysis (MLVA) to better elucidate therelationships among these isolates Altogether nineMLVA-types (MTs) were found (Table 3) Importantlythe same MLVA-type MT159 associated with ST78was characteristic for isolates derived from samplesfrom W-EFF and W-MOut suggesting survival of thisparticular clone
0
10
20
30
40
50
60
70
80
90
100
IS16
espintA
fms5
fms1
7
fms1
9
fms2
1
MDR
AM-R
CIP-R
HLG
R
p
ositi
ve
isol
ates
Figure 3 Distribution () of IS16 resistance phenotypes and virulenccolumns isolates with STs typical for nosocomial E faecium grey columnsgroups are indicated by an asterisk MDR multi-drug resistance AM-R ampgentamicin resistance HLSR high-level streptomycin resistance
IS16 was present in all 17 isolates included by MLST tothree lineages of nosocomial E faecium and this markerwas limited to this group (Table 3 and Figure 3) TheespEfm gene occurred always together with intA among allisolates of lineage 17 and in some isolates of lineage 78The fms19 pilin gene detected in 29 isolates was over-represented among STs typical for nosocomial E faeciumwhile other pilin genes such as fms5 fms17 and fms21detected in 57 49 and 46 isolates respectively occurredamong various CCs and singleton STs Ampicillin resist-ance significantly associated with isolates of nosocomialcomplex (10 of 17 isolates 588 p = 0001) occurred alsoin CC5 (6 isolates) CC22 and CC94 (1 isolate each) Se-quencing of the 3prime part of the pbp5 gene revealed its 11nucleotide alleles corresponding to 11 variants of the C-terminal part of PBP5 Six alleles of the gene were new(pbp5-24 to pbp5-29) two alleles (pbp5-20 pbp5-22)were described previously among Polish hospital VREmisolates [13] and three alleles were found in other stud-ies pbp5-30 occurred in a Portuguese animal strain ofE faecium [25] and pbp5-31 and pbp5-32 in clinical iso-lates TCGE70411_2 and 164306 also from Portugal (de-posited as JN208888 and JN208885 in the GenBankrespectively) For a single ampicillin-resistant isolate ofST653 (lineage 78) pbp5 amplification was not achieveddespite repeated attempts The majority (11 of 17 iso-lates 647) of isolates belonging to three lineages ofnosocomial E faecium showed resistance to ciprofloxa-cin determined by mutations R61G and S80IR in ParCand S84IRY in GyrA Ciprofloxacin resistance wasalso significantly over-represented among isolates ofSTs typical for nosocomial E faecium (p = 00006) Nine
HLS
R
tetM tetL
rep1
-pIP50
1
rep2
-pRE25
rep1
7-pR
UM
rep1
8-pE
F418
rep-pL
G1
axe-txe
relBE
e resistance and plasmid genes among E faecium isolates Blackother E faecium isolates Significant differences (p lt 005) between twoicillin resistance CIP-R ciprofloxacin resistance HLGR high-level
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 10 of 15httpwwwbiomedcentralcom1471-21801466
isolates resistant to ciprofloxacin were found also inCCs 5 9 and 22 These isolates possessed mutations inParC mentioned above but had the wild-type QRDR inGyrA HLGR occurred in five isolates of lineages 18 and78 and in single isolates of CC5 and CC22 this pheno-type was determined by the presence of aac(6prime)-Ie-aph(2Prime) with an exception of an isolate from CC5 whichwas negative for this gene High-level resistance tostreptomycin (HLSR) was widely distributed amongstudied isolates and determined by the ant(6prime)-Ia geneTetracycline resistance was found also in various STsthe major determinant of this phenotype was tet(M) (20isolates) followed by tet(L) (17 isolates) These twogenes occurred together in 16 isolates A single isolateof ST633 belonging to CC22 harbored tet(S) The tet(O) gene was absent in the studied group For a singletetracycline-resistant isolate of ST17 none of the fourtetracycline determinants studied was detected Nineisolates carried int of Tn916 and four the tndX gene ofTn5397 These transposon-specific genes were not char-acteristic for any lineage or complex (Table 3) Plasmid-replication genes such as rep2pRE25 rep17pRUM andrep18 pEF418 (26 14 and 7 isolates respectively) andgenes specifying toxin-antitoxin systems (TAS) axe-txeand ω-ε-ζ (12 and 10 isolates respectively) were signifi-cantly associated with the group of isolates of nosocomialE faecium Two other rep genes (rep1pIP501 reppLG1) werewidely distributed among all studied E faecium isolatesand the reppMG1 gene occurred in two isolates (in a singleisolate of ST533 from CC22 and in a singleton ST647) Allω-ε-ζ-positive isolates carried rep2pRE25 and nine isolatesshowed a concomitant presence of axe-txe and rep17pRUMThe relBE genes were found solely among five non-nosocomial E faecium (a difference in distribution with-out statistical significance p = 043) among various STs
DiscussionIn this study Enterococcus spp was used to evaluate theimpact posed by treated wastewater of two local WWTPson their receiver the costal waters of Gulf of GdanskTotal counts of enterococci observed for raw (W-INF)and treated (W-EFF D-EFF) wastewater are consistentwith our previous study showing similar over 99 reduc-tion capacity of applied treatment processes It is alsoreported in the literature that wastewater treatment pro-cesses based on activated sludge although they focus onremoval of nutrients and organic matter simultaneouslyreduce faecal indicators with high efficiency reaching upto 999 [1822] Due to the high number of enterococciin raw wastewater they are nevertheless released from aWWTP in a number up to 61x105 CFU per 100 mLThus marine water sampled in the area directly impactedby treated wastewater (W-MOut and D-MOut) showed ahigher number of enterococci than the one observed in
Vistula River mouth however lower by four orders ofmagnitude in comparison to corresponding treated waste-water (W-EFF and D-EFF) It should be noted that in mar-ine outfalls (W-MOut D-MOut) as well as in VistulaRiver mouth enterococci were detected in number lowerthan 100 CFU per 100 mL ie the value set out by NewBathing Water Directive 20067EC for the costal water ofldquoexcellent qualityrdquo [23]It is suggested that enterococcal species composition in
faecal-impacted environment depends from the source ofcontamination In this study two species most commonlycolonizing humans E faecalis and E faecium comprisedtogether from 73 to 88 of enterococcal isolates intested wastewater samples (W-INF W-BR W-EFF and D-EFF) 73 and 61 in marine outflows (W-MOut and D-MOut respectively) and 45 in the Vistula River mouthThe predominant species in wastewater and marine waterimpacted by treated wastewater was E faecium (from 56to 65) which stayed at a similar level during treatmentprocesses as determined also in our previous study [20]Species distribution in wastewater samples is not wellunderstood and affecting factors may include differencesin diet climate season and methods of detection E feaca-lis was found to be the most prevalent species in wastewa-ter samples in Sweden [26] E faecium in Spain UKCanada France and Switzerland [26-29] while E hiraepredominated in Portugal and United States [2230] Al-though E faecalis is typically the most abundant colonizerof humans in the community (eg [2631]) E faecium isgenerally most often associated with human microbialcontamination [2729] E faecalis was reported fromagriculture-impacted waters [32] and E hirae likelyoriginates mainly from cattle and pig faecal contamin-ation [26] which explains high relative proportion ofthese two species in Vistula River mouth Apart from thevariability of contamination sources proportion of par-ticular species may differ due to the differences of abilityto sustain environmental stresses In this study enterococ-cal composition might have been influenced by wastewa-ter treatment processes and marine water characteristicsThe effect of wastewater treatment and marine environ-
ment was seen on phenotypes of antimicrobial resistanceThe positive selection of bacteria with resistance patternshas been already suggested and noted in wastewater pro-cesses [192022] In this study the resistance rates to ampi-cillin ciprofloxacin levofloxacin and erythromycin notedfor E faecium from raw wastewater (W-INF) were lowerthan these observed for this species in the correspondingtreated wastewater (W-EFF) however not statistically sig-nificant (p gt 005) Moreover resistance rates to ampicillinciprofloxacin tetracycline and erythromycin observed inmarine outflow of WWTP Gdansk-Wschod (W-MOut)were higher than in treated wastewater of this plantSuch phenomenon was not observed for treated
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 11 of 15httpwwwbiomedcentralcom1471-21801466
wastewater of WWTP Gdynia-Debogorze (D-EFF) andits marine outfall (D-MOut) Resistance rates among Efaecium originating from D-MOut were in generalcomparable to these detected in the Vistula Rivermouth It should be noted that treated wastewater fromWWTP Gdansk-Wschod has been discharged via mar-ine outfall (W-MOut) for last 12 years while marineoutfall of WWTP Gdynia-Debogorze (D-MOut) hasbeen operating for last two years The observed differ-ences in resistance rate corresponding to the local im-pact caused by long and short-term operated marineoutfalls (W-MOut and D-MOut) need further attentionSignificant rates of antimicrobial resistance observed
for isolates of E faecalis from wastewater and marineriver waters impacted by wastewater are in agreementwith a recent study performed in six European countriesshowing that in Poland the rates of resistance for humanisolates of this species were usually among the highestboth in the community and hospitals [33] To our know-ledge no corresponding data are available for E faeciumcolonization in the community in Poland but a studyfocused on hospital VRE isolates showed their ubiquitousMDR phenotype and a high load of resistance genes [13]In our study no vancomycin resistance were detectedlikely due to relatively low (below 10) prevalence of VREin Polish hospitals [5] although such isolates are isolatedfrom the environment in other countries eg [34]This study shows that hospital clones of E faecium are
important carriers of resistance genes in wastewatersMost likely in a significant part they originated fromhospital wastewater treated together with communalwastewater In our estimation hospital wastewater con-stitutes approximately 02 of both WWTPsrsquo daily in-flow The release of hospital-associated clones of Efaecium to the environment was observed also by others[24283536] In our study isolates belonging to themajor nosocomial HiRECC (formerly named CC17)constituted altogether 246 of all isolates of E faeciumsubjected to typing and were present at all samplingsites These isolates apart from their inclusion into theHiRECC by MLST demonstrated several other pheno-typic and genotypic features typical for nosocomial Efaecium ie they were resistant to ciprofloxacin andampicillin in vast majority showed MDR phenotypeand carried several resistance determinants associatedboth with mutations of chromosomal genes (gyrA parCpbp5) and in the case of aminoglycoside and tetracyclineresistance with gene acquisition Plasmids likely played animportant role in the later process as some plasmid genessuch as rep2pRE25 rep17pRUM and rep18 pEF418 and TASgenes axe-txe and ω-ε-ζ were significantly more abundantin isolates associated with HiRECC than among theremaining isolates As yet the knowledge of distributionof plasmid-associated genes in populations of E faecium
especially in the context of their clonal compositionremains limited A study on 93 isolates of differentgeographical origins and from various sites showed anover-representation of rep17pRUM and reppMG1 amongnosocomial E faecium while in contrast to our observa-tions rep18 pEF418 was not detected [37] Moreover wedid not observe reppMG1 among HiRECC of E faeciumThese differences may be due to a high prevalence of VRE(58 isolates) among 93 isolates studied by Rosvoll et alwhat may explain differences in the plasmid content Thesame study also reported the co-localization of ω-ε-ζ withrep2pRE25 and axe-txe with rep17pRUM on the same plas-mids in concordance with our findings showing a fre-quent joint occurrence of these pairs of genes Anotherstudy performed on 99 invasive isolates from Norwaymostly of HLGR phenotype showed a significantly higherprevalence of rep2pRE25 and rep17pRUM among the majornosocomial HiRECC [38] This study as well some others[2439] also demonstrated a common presence of reppLG1among E faecium from hospital settings and in animalsOur findings indicate that this type of replicon is evenmore ubiquitous in E faecium than previously reportedAll rep genes mentioned above are typically associatedwith plasmids carrying various antimicrobial resistancegenes [3940] GenBank accession number AF408195 Inaddition to genes determining antimicrobial resistanceand genes associated with MGE isolates of nosocomialHiRECC analyzed in our study carried such molecularmarkers as IS16 and the esp gene together with the intAgene characteristic for ICEEfm [4142] Another patho-genicity factor the fms19 gene of the pilin gene cluster4 (PGC4) showed increased prevalence among theseisolates Similar observation was made for a representa-tive collection of 433 E faecium isolates from varioussources [43]In a study that followed enterococci in a continuum
from hospital and retirement home wastewaters througha WWTP to a river a gradual decrease in the proportionof E faecium isolates representing hospital-associatedSTs was observed likely due to the ldquodilutionrdquo of HiRECCrepresentatives by more diverse and less drug-resistantstrains originating from the community [28] Because inour study only selected representatives of obtained iso-lates were investigated it is not possible to determineprecise tendencies in their frequency from the WWTPinfluent to effluent and water however resistant entero-cocci both HiRECC and non-HiRECC are clearly ableto survive the treatment process what results in their re-lease into marine and river waters Moreover using anadditional typing method MLVA we were able to findisolates with the same characteristics in the effluent ofWWTP and its marine outfall suggesting ability of suchclones to survive in the environment for at least sometime It seems plausible that hospital resistant clones
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 12 of 15httpwwwbiomedcentralcom1471-21801466
accumulate several determinants that may promote theirsurvival not only in the hospital settings but also in theenvironment Such features would include resistance todisinfectants and ultraviolet light additional metabolicpathways and ability to form biofilms Moreover determi-nants of some of the mentioned adaptation factors arefound co-resident in plasmids harboring resistance genessuggesting co-selection For example pLG1-type plasmidsof E faecium carry not only resistance determinantsagainst aminoglycosides glycopeptides and macrolidesbut also pili genes and carbon uptake-utilization genes[3839] Pheromone-responsive plasmids of E faecalisapart from resistance to aminoglycosides glycopeptidespenicillins and macrolides may encode bacteriocins add-itional pili and UV-resistance determinants [40] Thusnosocomial HiRECCs released by WWTPs are equippedin whole sets of genes with various adaptive benefits andoften located within MGE It was also demonstrated thatMGEs can be transferred in a WWTP [44] further under-ling the importance of threat posed by such organismsThere is a selection pressure for survival of resistantstrains and their tranconjugants due to release of antibi-otics to the environment especially in the hospital efflu-ents [45] and even sub-inhibitory concentrations may beeffective in selection for resistance [46]
ConclusionsThis study describes the species composition resistanceprofiles and clonal relationships of enterococci isolatedfrom WWTPs located in Gdansk their marine outfallsto the Gulf of Gdansk and from Vistula River It was ob-served that these WWTPs release enterococcal strainsresistant to various antimicrobial compounds to the re-cipient water environment Molecular analyses revealedthe presence of clones associated with nosocomial HiR-ECCs and in the case of E faecium isolates of lineages17 18 and 78 of former CC17 abundant in resistance de-terminants MGE genes and pathogenicity factors Thepresence of HiRECCs in wastewaters and marineriverenvironment highlights the need for further detailedanalyses to better understand the survival and spread ofdrug-resistant strains in water ecosystems and to eluci-date the ways to curb such dissemination
MethodsAcquisition of samples and isolation of enterococciOnce a month in April June August and October in 2011of the flow proportioned composited samples of wastewa-ter were taken from two local WWTPs Gdansk-Wschodfrom influent (W-INF) effluent (W-EFF) and wastewa-ter from activated sludge bioreactor (W-BR) and fromGdynia-Debogorze effluent (D-EFF) The two WWTPstreat sewage from approximately 920 000 inhabitants(for details see Figure 1) In the same period of time and
with the same frequency marine outfalls of those WWTPs(W-MOut and D-MOut respectively) as well as VistulaRiver mouth were also sampled for isolation of entero-cocci Marine samples were taken at the depth of approxi-mately one meter below the surface once a month inApril June August and October of 2011 Location ofsampling points and their characteristics are provided inFigure 1 Enterococci were detected using the membranefiltration method To this end appropriate dilutions ofanalyzed samples were filtered in duplicate through045 μm cellulose-acetate filters (EMD Millipore Cor-poration Billerica MA USA) which were then placedon Enterococcus selective agar (Merck DarmstadtGermany) and incubated at 37degC for 48 h (ISO 7899ndash22000) Dark red or maroon colonies were consideredpresumptive enterococci For further investigationsrepresentative isolates were taken from membranes pre-senting from 20 to 50 typical colonies and stored in nu-trient broth supplemented with 15 glycerol at minus80degC
Species identification DNA isolation susceptibility testingand statistical analysisThe species identification and drug susceptibility of pre-sumptive enterococci were determined by the PhoenixtradeAutomated Microbiology System (BD New Jersey USA)according to the manufacturerrsquos instructions Since it isnot possible to distinguish E casseliflavus and E galli-narum using this system the classification E casselifla-vusgallinarum was used The susceptibility tests basedon microdilution were carried out for 11 antimicrobialagents gentamicin (GM) streptomycin (ST) ampicillin(AM) daptomycin (DAP) vancomycin (VA) teicoplanin(TEC) linezolid (LZD) fluoroquinolones (ciprofloxacin -CIP levofloxacin - LVX) tetracycline (TE) erythromycin(E) and quinupristin-dalfopristin (SYN) (E faecium only)Obtained minimal inhibitory concentrations (MIC) wereevaluated according to the guidelines of Clinical and La-boratory Standards Institute [47] Isolates were defined asmultidrug-resistant (MDR) when they showed resistanceto three or more compounds tested [48] Bacterial DNAwas purified using the Genomic DNA Prep Plus kitfollowing the manufacturerrsquos instructions (AampA Biotech-nology Gdynia Poland) Differences in distributions wereevaluated using the χ2 test with the p values lt 005 con-sidered significant
MLST and MLVA of E faeciumMLST for E faecium was performed by sequencing ofseven house-keeping genes as described by Homan et al[49] The allele numbers and STs were assigned with theuse of Internet database httpefaeciummlstnet (5th July2013 date last accessed) Novel alleles and allelic profileswere submitted to the database MLST data were analyzedwith Sequence Type Analysis and Recombinational Tests
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 13 of 15httpwwwbiomedcentralcom1471-21801466
(START) software [50] and the eBURST analysis [51] wasused to elucidate the relationships of STs with knownCCs using the comparative eBURST option available athttpeburstmlstnetv3enter_datacomparative (5th July2013 date last accessed) Further inclusion of E faeciumSTs into specific CCs and lineages was performed fol-lowing other studies [924] MLVA was performed as de-scribed by others [52]
Detection and analysis of genes associated with virulenceantimicrobial resistance and MGE in E faeciumThe full list of primers used for gene detection and se-quencing is provided in Additional file 1 Table S1 PCR-based detection of IS16 virulence-associated genes (fms21fms17 fms5 and fms19 representing each of four pilingene clusters in E faecium and the espEfm gene) genes de-termining resistance to tetracycline tet(M) tet(O) tet(L)tet(S) HLGR (aac(6prime)-Ie-aph(2Prime) HLSR (ant(6prime)-Ia) andgenes specific for MGE such as intA of ICEEfm1 int ofTn916 transposon tndX of Tn5397 transposon entero-coccal plasmid replication genes rep1pIP501 rep2pRE25rep17pRUM rep18pEF418 reppMG1 reppLG1 and TAS genesaxe-txe ω-ε-ζ relBE was performed as described in otherstudies [134153-59] following the conditions describedherein Previously characterized clinical isolates of Strepto-coccus agalactiae E faecium and E faecalis harbouringpilin genes IS16 espEfm intA of ICEEfm1 int of Tn916tndX of Tn5397 tet(M) tet(O) tet(L) tet(S) aac(6prime)-Ie-aph(2Prime) ant(6prime)-Ia rep1pIP501 rep2pRE25 ω-ε-ζ [136061]served as positive controls In cases when positive controlswere not available (rep18pEF418 reppMG1 reppLG1 axe-txerelBE) a few randomly selected PCR products wereverified by sequencing and served further as controlsAmpicillin-resistance determinant the pbp5 gene was ana-lysed by sequencing of the part encoding C-terminaltranspeptidase domain in PBP5 [62] New sequence vari-ants pbp5-24 to pbp5-29 alleles were submitted to theGenBank (acc numbers KC594860-KC594865 respect-ively) The quinolone-determining regions (QRDR) in thegyrA and parC genes were sequenced and analysed asdescribed by others [63]
Additional file
Additional file 1 Table S1 Primers used in the study for PCR andsequencing
Competing interestThe authors declare that they have no competing interest
Authorsrsquo contributionsES participated in the study design performed experimental work andparticipated in the manuscript preparation AL designed the studyperformed experimental work and participated in the manuscriptpreparation Both authors read and approved the final manuscript
AcknowledgementsWe thank Rob Willems and Janetta Top for assigning new alleles new STsand MTs This publication made use of the Enterococcus faecium MLSTwebsite (httpefaeciummlstnet) which is hosted at Imperial College of theUniversity of Oxford and funded by the Wellcome Trust This work wassupported by project N N305 461139 from the Narodowe Centrum Nauki(NCN)
Author details1National Medicines Institute ul Chelmska 3034 Warsaw 00-725 Poland2Gdansk University of Technology ul G Narutowicza 1112 Gdansk 80-952Poland
Received 5 September 2013 Accepted 27 February 2014Published 14 March 2014
References1 Aarestrup FM Butaye P Witte W Nonhuman reservoirs of Enterococci In
American Society for Microbiology Edited by Gilmore MS Clewell DB CourvalinP Dunny GM Murray BE Rice LB Washington DC The Enterococcipathogenesis molecular biology and antibiotic resistance 2002
2 Tendolkar PM Baghdayan AS Shankar N Pathogenic enterococci newdevelopments in the 21st century Cell Mol Life Sci 2003 602622ndash2636
3 Sood S Malhotra M Das BK Kapil A Enterococcal infections ampantimicrobial resistance Indian J Med Res 2008 128111ndash121
4 Sydnor ER Perl TM Hospital epidemiology and infection control inacute-care settings Clin Microbiol Rev 2011 24141ndash173
5 European Centre for Disease Prevention and Control Annual EpidemiologicalReport Reporting on 2010 surveillance data and 2011 epidemic intelligencedata Stockholm ECDC 20122013
6 Hidron AI Edwards JR Patel J Horan TC Sievert DM Pollock DA Fridkin SKNational Healthcare Safety Network Team Participating National HealthcareSafety Network Facilities NHSN annual update antimicrobial-resistantpathogens associated with healthcare-associated infections annualsummary of data reported to the National Healthcare Safety Network atthe Centers for Disease Control and Prevention 2006ndash2007 Infect ControlHosp Epidemiol 2008 29996ndash1011
7 De Kraker ME Jarlier V Monen JC Heuer OE van de Sande N GrundmannH The changing epidemiology of bacteraemias in Europe trends fromthe European Antimicrobial Resistance Surveillance System Clin MicrobiolInfect 2012 doi1011111469-0691
8 Leavis HL Bonten MJ Willems RJ Identification of high-risk enterococcalclonal complexes global dispersion and antibiotic resistance Curr OpinMicrobiol 2006 9454ndash460
9 Willems RJL Top J van Schaik W Leavis H Bonten M Sireacuten J Hanage WPCorander J Restricted gene flow among hospital subpopulations ofEnterococcus faecium mBio 2012 3e00151ndash12
10 Hegstad K Mikalsen T Coque TM Werner G Sundsfjord A Mobile geneticelements and their contribution to the emergence of antimicrobialresistant Enterococcus faecalis and Enterococcus faecium Clin MicrobiolInfect 2010 16541ndash554
11 Shepard BD Gilmore MS Antibiotic-resistant enterococci themechanisms and dynamics of drug introduction and resistance MicrobesInfect 2002 4215ndash224
12 Bonten MJ Willems R Weinstein RA Vancomycin-resistant enterococciwhy are they here and where do they come from Lancet Infect Dis 20011314ndash325
13 Sadowy E Sieńko A Gawryszewska I Bojarska A Malinowska K HryniewiczW High abundance and diversity of antimicrobial resistancedeterminants among early vancomycin-resistant Enterococcus faecium inPoland Eur J Clin Microbiol Infect Dis 2013 321193ndash1203
14 Willems RJ Homan W Top J van Santen-Verheuvel M Tribe D Manzioros XGaillard C Vandenbroucke-Grauls CM Mascini EM van Kregten E vanEmbden JD Bonten MJ Variant esp gene as a marker of a distinct geneticlineage of vancomycin-resistant Enterococcus faecium spreading inhospitals Lancet 2001 357853ndash855
15 Sillanpaumlauml J Nallapareddy SR Prakash VP Qin X Houmloumlk M Weinstock GMMurray BE Identification and phenotypic characterization of a secondcollagen adhesin Scm and genome-based identification and analysis of13 other predicted MSCRAMMs including four distinct pilus loci inEnterococcus faecium Microbiology 2008 1543199ndash3211
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 14 of 15httpwwwbiomedcentralcom1471-21801466
16 Galloway-Pentildea JR Nallapareddy SR Arias CA Eliopoulos GM Murray BEAnalysis of clonality and antibiotic resistance among early clinicalisolates of Enterococcus faecium in the United States J Infect Dis 20092001566ndash1573
17 Top J Willems R Bonten M Emergence of CC17 Enterococcus faeciumfrom commensal to hospital-adapted pathogen FEMS Immunol MedMicrobiol 2008 52297ndash308
18 Martins da Costa P Vaz-Pires P Bernardo F Antimicrobial resistance inEnterococcus spp isolated in inflow effluent and sludge from municipalsewage water treatment plants Water Res 2006 401735ndash1740
19 Reinthaler FF Posch J Feierl G Wust G Haas D Ruckenbauer G Mascher FMarth E Antibiotic resistance of Escherichia coli in sewage and sludgeWater Res 2003 371685ndash1690
20 Luczkiewicz A Fudala-Ksiazek S Jankowska K Olanczuk-Neyman KAntimicrobial resistance of fecal indicators in municipal wastewatertreatment plant Water Res 2010 445089ndash5097
21 Dulfour AP Ballentine RK Ambient water quality criteria for bacteria-1986 EPA4405-84-002 Washington DC US Environmental Protection Agency 1986
22 Ferreira da Silva M Tiago I Veriacutessimo A Boaventura RA Nunes OC Manaia CMAntimicrobial resistance patterns in Enterobacteriaceae isolated from anurban wastewater treatment plant FEMS Microbiol Ecol 2007 60166ndash176
23 DIRECTIVE 20067EC of the European Parliament and of the Council 15February 2006 concerning the management of bathing water quality andrepealing Directive 76160EEC httpeur-lexeuropaeuLexUriServLexUriServdouri=OJL200606400370051ENPDF
24 Freitas AR Novais C Ruiz-Garbajosa P Coque TM Peixe L Dispersion ofmultidrug-resistant Enterococcus faecium isolates belonging to majorclonal complexes in different Portuguese settings Appl Environ Microbiol2009 754904ndash4908
25 Poeta P Costa D Igrejas G Saacuteenz Y Zarazaga M Rodrigues J Torres CPolymorphisms of the pbp5 gene and correlation with ampicillinresistance in Enterococcus faecium isolates of animal origin J MedMicrobiol 2007 56236ndash240
26 Kuumlhn I Iversen A Burman LG Olsson-Liljequist B Franklin A Finn MAarestrup F Seyfarth AM Blanch AR Vilanova X Taylor H Caplin J MorenoMA Dominguez L Herrero IA Moumlllby R Comparison of enterococcalpopulations in animals humans and the environment ndash a Europeanstudy Int J Food Microbiol 2003 88133ndash145
27 Lanthier M Scott A Lapen DR Zhang Y Topp E Frequency of virulencegenes and antibiotic resistances in Enterococcus spp isolates fromwastewater and feces of domesticated mammals and birds and wildlifeCan J Microbiol 2010 56715ndash729
28 Leclercq R Oberleacute K Galopin S Cattoir V Budzinski H Petit F Changes inenterococcal populations and related antibiotic resistance along amedical center-wastewater treatment plant-river continuum Appl EnvironMicrobiol 2013 792428ndash2434
29 Thevenon F Regier N Benagli C Tonolla M Adatte T Wildi W Poteacute JCharacterization of fecal indicator bacteria in sediments cores from thelargest freshwater lake of Western Europe (Lake Geneva Switzerland)Ecotoxicol Environ Safety 2012 7850ndash56
30 Bonilla TD Nowosielski K Esiobu N McCorquodale DS Rogerson ASpecies assemblages of Enterococcus indicate potential sources offecal bacteria at a south Florida recreational beach Mar Pollut Bull2006 52807ndash810
31 Wang JT Chang SC Wang HY Chen PC Shiau YR Lauderdale TL TSARHospitals High rates of multidrug resistance in Enterococcus faecalis andE faecium isolated from inpatients and outpatients in TaiwanDiagn Microbiol Infect Dis 2013 75406ndash411
32 Lanthier M Scott A Zhang Y Cloutier M Durie D Henderson VC WilkesG Lapen DR Topp E Distribution of selected virulence genes andantibiotic resistance in Enterococcus species isolated from the SouthNation River drainage basin Ontario Canada J Appl Microbiol 2011110407ndash421
33 Kuch A Willems RJ Werner G Coque TM Hammerum AM Sundsfjord AKlare I Ruiz-Garbajosa P Simonsen GS van Luit-Asbroek M Hryniewicz WSadowy E Insight into antimicrobial susceptibility and population struc-ture of contemporary human Enterococcus faecalis isolates fromEurope J Antimicrob Chemother 2012 67551ndash558
34 Novais C Coque TM Ferreira H Sousa JC Peixe L Environmentalcontamination with vancomycin-resistant enterococci from hospitalsewage in Portugal Appl Environ Microbiol 2005 713364ndash3368
35 Iversen A Kuumlhn I Rahman M Franklin A Burman LG Olsson-Liljequist BTorell E Moumlllby R Evidence for transmission between humans and theenvironment of a nosocomial strain of Enterococcus faeciumEnviron Microbiol 2004 655ndash59
36 Rathnayake I Hargreaves M Huygens F SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterwayAustralia and associated antibiotic resistance gene profilesBMC Microbiol 2011 11201
37 Rosvoll TC Pedersen T Sletvold H Johnsen PJ Sollid JE Simonsen GSJensen LB Nielsen KM Sundsfjord A PCR-based plasmid typing inEnterococcus faecium strains reveals widely distributed pRE25-pRUM- pIP501- and pHT-related replicons associated with glycopeptideresistance and stabilizing toxin-antitoxin systems FEMS Immunol MedMicrobiol 2010 58254ndash268
38 Rosvoll TC Lindstad BL Lunde TM Hegstad K Aasnaes B Hammerum AMLester CH Simonsen GS Sundsfjord A Pedersen T Increased high-levelgentamicin resistance in invasive Enterococcus faecium is associated withaac(6prime)Ie-aph(2Prime)Ia-encoding transferable megaplasmids hosted by majorhospital-adapted lineages FEMS Immunol Med Microbiol 2012 66166ndash176
39 Laverde Gomez JA van Schaik W Freitas AR Coque TM Weaver KE FranciaMV Witte W Werner G A multiresistance megaplasmid pLG1 bearing ahylEfm genomic island in hospital Enterococcus faecium isolates Int J MedMicrobiol 2011 301165ndash175
40 Coque TM Evolutionary biology of pathogenic enterococci InEvolutionary Biology of Bacterial and Fungal Pathogens Coque TM Edited byBaquero F Nombela C Cassell GH Gutierrez JA Washington DC ASMPress501ndash523
41 Werner G Fleige C Geringer U van Schaik W Klare I Witte W IS elementIS16 as a molecular screening tool to identify hospital-associated strainsof Enterococcus faecium BMC Infect Dis 2011 1180
42 Top J Sinnige JC Majoor EA Bonten MJ Willems RJ van Schaik W Therecombinase IntA is required for excision of esp-containing ICEEfm1 inEnterococcus faecium J Bacteriol 2011 1931003ndash1006
43 Sillanpaumlauml J Prakash VP Nallapareddy SR Murray BE Distribution of genesencoding MSCRAMMs and Pili in clinical and natural populations ofEnterococcus faecium J Clin Microbiol 2009 47896ndash901
44 Marcinek H Wirth R Muscholl-Silberhorn A Gauer M Enterococcus faecalisgene transfer under natural conditions in municipal sewage watertreatment plants Appl Environ Microbiol 1998 64626ndash632
45 Kuumlmmerer K Significance of antibiotics in the environment J AntimicrobChemother 2003 525ndash7
46 Liu A Fong A Becket E Yuan J Tamae C Medrano L Maiz M Wahba C LeeC Lee K Tran KP Yang H Hoffman RM Salih A Miller JHP Selectiveadvantage of resistant strains at trace levels of antibiotics a simple andultrasensitive color test for detection of antibiotics and genotoxicagents Antimicrob Agents Chemother 2011 551204ndash1210
47 Clinical and Laboratory Standards Institute Performance standards forantimicrobial susceptibility testing 21st informational supplement Wayne CLSI2011 PA M100-S20
48 Magiorakos AP Srinivasan A Carey RB Carmeli Y Falagas ME Giske CGHarbarth S Hindler JF Kahlmeter G Olsson-Liljequist B Paterson DL Rice LBStelling J Struelens MJ Vatopoulos A Weber JT Monnet DLMultidrug-resistant extensively drug-resistant and pandrug-resistantbacteria an international expert proposal for interim standarddefinitions for acquired resistance Clin Microbiol Infect 2012 18268ndash281
49 Homan WL Tribe D Poznanski S Li M Hogg G Spalburg E Van Embden JDWillems RJ Multilocus sequence typing scheme for Enterococcus faeciumJ Clin Microbiol 2002 401963ndash1971
50 Jolley KA Feil EJ Chan MS Maiden MC Sequence type analysis andrecombinational tests (START) Bioinformatics 2001 171230ndash1231
51 Feil EJ Li BC Aanensen DM Hanage WP Spratt BG eBURST inferringpatterns of evolutionary descent among clusters of related bacterialgenotypes from multilocus sequence typing data J Bacteriol 20041861518ndash1530
52 Top J Banga1 NMI Hayes R Willems RJ Bonten MJM Hayden MKComparison of multiple-locus variable-number tandem repeat analysisand pulsed-field gel electrophoresis in a setting of polyclonal endemicityof vancomycin-resistant Enterococcus faecium Clin Microbiol Infect 200814363ndash369
53 Vankerckhoven V Van Autgaerden T Vael C Lammens C Chapelle S RossiR Jabes D Goossens H Development of a multiplex PCR for the
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-

Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 4 of 15httpwwwbiomedcentralcom1471-21801466
respectively altogether 19) The most common speciesin all sampled sites was E faecium (562 ndash 655) withthe exception of Vistula River mouth where E hirae waspredominant (536 of isolates) followed by E faecalis(377)In the next step antimicrobial susceptibility was tested
for E faecium and E faecalis isolates (Table 2) It shouldbe noted that at some points a low number of E faecalisisolates was recovered (fewer than 10 isolates) and thuscertain resistance rates obtained for E feacalis should betreated with caution Resistance to erythromycin was de-tected with the highest prevalence among both specieshowever rates of resistance to this compound variedover a broad range Erythromycin-resistant E faeciumcomprised 368 of isolates in raw wastewater (W-INF)453 in treated wastewater (W-EFF) and 586 inmarine outflow (W-MOut) of WWTP Gdansk-WschodSimilar tendency was seen for E faecalis isolates whereresistance rates in the sampling points listed above wereequal to 200 684 and 100 respectively For WWTPGdynia-Debogorze effluent (D-EFF) and marine outflow(D-MOut) erythromycin resistance rates for E faeciumvaried between 682 and 550 respectively while for Efaecalis were equal 684 and 667 respectively In thecase of fluoroquinolones resistance to ciprofloxacin among
Table 2 Antimicrobial resistance of E faecium and E faecalis
Num
Resistant to a single antim
GM ST AM CIP LVX
E faecium
W= INF (n = 19) 1 (53) 3 (158) 1 (53) 6 (316) 1 (53
W-BR (n = 36) 1 (28) 2 (56) 1 (28) 12 (332) 3 (83
W-EFF (n = 53) 3 (57) 2 (38) 4 (75) 23 (434) 10 (18
W-MOut (n = 29) 0 6 (207) 6 (207) 13 (448) 4 (138
D-EFF (n = 41) 1 (24) 6 (146) 5 (122) 13 (317) 2 (49
D-MOut (n = 40) 1 (25) 2 (5) 2 (5) 14 (350) 3 (75
VR (n = 26) 0 1 (38) 0 8 (308) 1 (38
All (n = 244) 7 (29) 22 (90) 19 (78) 90 (369) 24 (98
E faecalis
W-INF (n = 5) 0 1 (200) 0 1 (200) 0
W-BR (n = 8) 2 (250) 2 (250) 1 (125) 2 (250) 2 (250
W-EFF (n = 19) 0 1 (53) 1 (53) 3 (158) 3 (158
W-MOut (n = 4) 0 0 0 0 0
D-EFF (n = 19) 2 (105) 5 (263) 0 1 (53) 0
D-MOut (n = 3) 1 (333) 1 (333) 0 0 0
VR (n = 5) 0 0 0 2 (400) 0
All (n = 63) 5 (79) 10 (159) 2 (32) 9 (143) 5 (79
GM gentamicin ST streptomycin AM ampicillin CIP ciprofloxacin LVX levofloxaciall tested antimicrobial agents MDR multiple drug resistance na not applicableAbbreviations of sampling pointsrsquo locations and their description provided in Figure
E faecium varied between 308 in Vistula River mouth to448 in W-MOut while resistance to levofloxacin did notexceed 20 Among E faecalis resistance to ciprofloxacinand levofloxacin was in general lower then reported for Efaecium in the corresponding sampling points (up to250) except Vistula River mouth The high-level strepto-mycin resistance (HLSR) was prevalent and reached 207for E faecium isolates from marine outflow of WWTPGdansk-Wschod (W-MOut) Also for E faecalis HLSRphenotype was detected among up to 16 of all tested iso-lates Resistance to glycopeptides linezolid and daptomycinwas not detected among all tested enterococci
Clonal structure of E faecium isolates and distribution ofvirulence resistance and MGE genesSixty-nine isolates of E faecium (283 of all isolates ofthis species) were further subjected to MLST and ana-lysis of the presence of resistance virulence and MGEgenes These isolates presented various phenotypes of re-sistance to antimicrobial compounds as described belowand were obtained mostly from effluents and marine out-falls of both WWTPs (Table 3) MLST discerned 54 differ-ent STs (Figure 2) including 23 new ones (STs 632ndash654)The comparative eBURST analysis against the wholeMLST E faecium database (as of the 5th April 2013)
ber () of isolates
icrobial agent Sensitive MDRphenotypeTE SYN E S
n = 244
) 5 (263) 6 (316) 7 (368) 6 (316) 4 (211)
) 7 (194) 19 (528) 16 (444) 10 (278) 8 (222)
9) 10 (189) 18 (340) 24 (453) 12 (226) 14 (264)
) 8 (276) 14 (483) 17 (586) 5 (174) 9 (310)
) 10 (244) 12 (292) 28 (682) 4 (98) 8 (195)
) 2 (50) 21 (525) 22 (550) 7 (175) 3 (75)
) 2 (77) 15 (577) 11 (424) 3 (115) 3 (115)
) 44 (180) 105 (430) 125 (512) 49 (201) 47 (193)
n = 63
1 (200) na 1 (200) 4 (800) 0
) 2 (250) na 7 (875) 1 (125) 2 (250)
) 4 (211) na 13 (684) 0 3 (158)
0 na 4 (100) 0 0
9 (474) na 13 (684) 4 (211) 5 (263)
2 (667) na 2 (667) 0 1 (333)
0 na 1 (200) 2 (400) 0
) 18 (286) na 41 (651) 11 (175) 11 (175)
n TE tetracycline SYN quinupristin-dalfopristin E erythromycin S sensitive to
1
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium
LineageCCST
MT Isolationsite
Virulence genes Resistance determinants Transposongenes
Plasmid rep genes Plasmid TAS
espintA
fms5 fms17 fms19 fms21 pbp5 GyrA ParC aac(6prime)-Ie-aph(2Prime)
ant(6prime)-Ia
tet(M)
tet(L)
tet(S)
intTn916 tndX rep1pIP501
rep2pRE25
rep17pRUM
rep18pEF418
reppLG1
reppMG1
axe-txe
ω-ε-ζ
relBE
Lineage 17
17 (2) 1 8 W-EFF 2 2 2 2 2 pbp5-30
S84R R61GS80R
1 1 2 2 1 2 2 1 1
386 7 W-MOut 1 1 1 1 1 1 1 1 1 1 1 1 1
Lineage 18
18 (3) 1(3)
D-EFFW-MOut
3 3 3 1 pbp5-20
S84Y R61GS80I
1 2 3 3 1 2 1 3
262 10 W-EFF 1 1 1 pbp5-25
S84Y R61G 1 1 1 1 1 1 1
574 402 VR 1 1 1 1
Lineage 78
78 (4) 159(4)
W-EFFW-MOut
4 4 4 4 4 pbp5-28
S84RY
R61GS80R
2 1 2 2 1 4 4 2 4 4 3
266 (2) 139(2)
W-INFD-EFF
2 1 2 2 pbp5-24
1 1 2 1 1 2 2 2
323 12 D-MOut 1 1 1 1 pbp5-26
wt R61GS80I
1 1 1 1 1 1 1 1
564 302 D-EFF 1 1 1 1 pbp5-29
S84I R61GS80R
1 1 1 1 1 1 1
653 302 W-EFF 1 nd S84I R61GS80R
1 1 1
CC5
29 (2) W-EFFW-MOut
2 1 1 2 1
66 W-MOut 1 1 1 1 1 1 1
97 W-EFF 1 wt R61GS80R
1 1
123 W-EFF 1 1 wt R61G 1 1 1
148 W-MOut 1 pbp5-30
wt R61GS80R
1 1 1 1 1
168 W-MOut 1 1 pbp5-31
1 1 1 1 1 1 1 1 1
505 W-EFF 1 wt R61G 1 1
588 D-MOut 1 pbp5-30
1 1 1 1 1 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
5of
15httpw
wwbiom
edcentralcom1471-21801466
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium (Continued)
635 D-EFF 1 1 1 1
636 W-EFF 1 wt R61G 1
637 W-EFF 1 1 1 wt R61G 1 1
638 W-EFF 1 wt R61GS80I
1
642 W-EFF 1 1 1 1 1
645 D-EFF 1 1 1 1 1
646 W-EFF 1 1 1 1 1
649 W-BR 1 1 1 pbp5-32
wt R61G 1
650 (3) D-EFF 3 3 pbp5-27
2 2 2 1 1 3
CC 9
8 D-EFF 1 1 1 1 1 1 1 1 1 1 1
640 W-EFF 1 1 1 1 1
CC 22
21 W-EFF 1 1 1 1 1 1 1 1
22 W-EFF 1 1 1 1 1 1
25 W-EFF 1 1 pbp5-32
wt R61G 1 1 1
32 (3) W-EFF 3 3 1 wt R61G 2 1
92 W-EFF 1 1 1 1 1
441 W-EFF 1 1 1 1 1 1 1 1
533 W-EFF 1 1 1
633 W-EFF 1 1 1 1 1
644 D-EFF 1 1 1 1 1
CC27
652 W-EFF 1 1 1 1
CC46
47 VR 1 1 1 1
69 D-EFF 1 1 1 1 1 1
CC94
94 W-MOut 1 1 1 1
361 (4) D-EFFD-MOut
3 4 4 4 pbp5-29
1 1 1 4 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
6of
15httpw
wwbiom
edcentralcom1471-21801466
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium (Continued)
CC296
296 W-EFF 1 1
CC639
639 W-EFF 1 1 1 1 1
CC648
634 W-EFF 1 1 1 1 1
648 W-EFF 1 1 1 1 1
Doubletons
632 W-MOut 1 1 1
Singletons
264 W-INF 1 1 1 1 1 1 1
641 D-EFF 1 1 1 1
643 W-EFF 1 1 1 1 1 1
647 W-MOut 1 1 1 1 1
651 D-EFF 1 1 1 1 1 1
654 VR 1 1 1 1 1 1
All (69) 8 57 49 29 46 6 17 20 17 1 9 4 39 27 14 7 55 2 12 10 5
CC clonal complex ST sequence type MT MLVA-type nd not determined due to lack of amplification of the pbp5 gene wt wild-type STs associated with MDR phenotype in boldAbbreviations of sampling pointsrsquo locations and their description provided in Figure 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
7of
15httpw
wwbiom
edcentralcom1471-21801466
Figure 2 MLST-based similarity tree of E faecium isolates constructed using the START software and the UPGMA clustering algorithm
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 8 of 15httpwwwbiomedcentralcom1471-21801466
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 9 of 15httpwwwbiomedcentralcom1471-21801466
grouped the 38 STs specific for 50 isolates into the majorlsquosuper-complex 17rsquo characteristic for this species (data notshown) Further detailed analysis of the localization ofparticular STs within this structure revealed that 11 STscharacteristic for 17 isolates belonged to the nosocomialcomplex of E faecium representing all three majorlineages 17 18 and 78 of a former CC17 as defined byWillems et al [9] (Table 3) Seventeen STs (20 isolates)nine STs (11 isolates) and two STs with single isolates eachbelonged to CC5 CC22 and CC9 respectively as definedby Freitas et al [24] Of the remaining 16 STs nine wereincluded into six small CCs (CCs 27 46 94 296 639648) unlinked to the major lsquosuper-complexrsquo ST632formed a lsquodoubletonrsquo (ie was linked by a single-locus vari-ant link to only one ST from the database) and six STsrepresented by single isolates were singletons ie werenot related to any ST in the database at the time of ana-lysis Thirty isolates (435) in the investigated groupshowed the MDR phenotype which was significantly asso-ciated (p = 000006) with isolates from the nosocomialcomplex (Table 3 and Figure 3) but found also in CC5CC22 and CC94 this phenotype did not occur amongsmall CCs or singletons Seventeen isolates repre-senting the nosocomial complex were additionallystudied by multilocus VNTR (variable number tan-dem repeats) analysis (MLVA) to better elucidate therelationships among these isolates Altogether nineMLVA-types (MTs) were found (Table 3) Importantlythe same MLVA-type MT159 associated with ST78was characteristic for isolates derived from samplesfrom W-EFF and W-MOut suggesting survival of thisparticular clone
0
10
20
30
40
50
60
70
80
90
100
IS16
espintA
fms5
fms1
7
fms1
9
fms2
1
MDR
AM-R
CIP-R
HLG
R
p
ositi
ve
isol
ates
Figure 3 Distribution () of IS16 resistance phenotypes and virulenccolumns isolates with STs typical for nosocomial E faecium grey columnsgroups are indicated by an asterisk MDR multi-drug resistance AM-R ampgentamicin resistance HLSR high-level streptomycin resistance
IS16 was present in all 17 isolates included by MLST tothree lineages of nosocomial E faecium and this markerwas limited to this group (Table 3 and Figure 3) TheespEfm gene occurred always together with intA among allisolates of lineage 17 and in some isolates of lineage 78The fms19 pilin gene detected in 29 isolates was over-represented among STs typical for nosocomial E faeciumwhile other pilin genes such as fms5 fms17 and fms21detected in 57 49 and 46 isolates respectively occurredamong various CCs and singleton STs Ampicillin resist-ance significantly associated with isolates of nosocomialcomplex (10 of 17 isolates 588 p = 0001) occurred alsoin CC5 (6 isolates) CC22 and CC94 (1 isolate each) Se-quencing of the 3prime part of the pbp5 gene revealed its 11nucleotide alleles corresponding to 11 variants of the C-terminal part of PBP5 Six alleles of the gene were new(pbp5-24 to pbp5-29) two alleles (pbp5-20 pbp5-22)were described previously among Polish hospital VREmisolates [13] and three alleles were found in other stud-ies pbp5-30 occurred in a Portuguese animal strain ofE faecium [25] and pbp5-31 and pbp5-32 in clinical iso-lates TCGE70411_2 and 164306 also from Portugal (de-posited as JN208888 and JN208885 in the GenBankrespectively) For a single ampicillin-resistant isolate ofST653 (lineage 78) pbp5 amplification was not achieveddespite repeated attempts The majority (11 of 17 iso-lates 647) of isolates belonging to three lineages ofnosocomial E faecium showed resistance to ciprofloxa-cin determined by mutations R61G and S80IR in ParCand S84IRY in GyrA Ciprofloxacin resistance wasalso significantly over-represented among isolates ofSTs typical for nosocomial E faecium (p = 00006) Nine
HLS
R
tetM tetL
rep1
-pIP50
1
rep2
-pRE25
rep1
7-pR
UM
rep1
8-pE
F418
rep-pL
G1
axe-txe
relBE
e resistance and plasmid genes among E faecium isolates Blackother E faecium isolates Significant differences (p lt 005) between twoicillin resistance CIP-R ciprofloxacin resistance HLGR high-level
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 10 of 15httpwwwbiomedcentralcom1471-21801466
isolates resistant to ciprofloxacin were found also inCCs 5 9 and 22 These isolates possessed mutations inParC mentioned above but had the wild-type QRDR inGyrA HLGR occurred in five isolates of lineages 18 and78 and in single isolates of CC5 and CC22 this pheno-type was determined by the presence of aac(6prime)-Ie-aph(2Prime) with an exception of an isolate from CC5 whichwas negative for this gene High-level resistance tostreptomycin (HLSR) was widely distributed amongstudied isolates and determined by the ant(6prime)-Ia geneTetracycline resistance was found also in various STsthe major determinant of this phenotype was tet(M) (20isolates) followed by tet(L) (17 isolates) These twogenes occurred together in 16 isolates A single isolateof ST633 belonging to CC22 harbored tet(S) The tet(O) gene was absent in the studied group For a singletetracycline-resistant isolate of ST17 none of the fourtetracycline determinants studied was detected Nineisolates carried int of Tn916 and four the tndX gene ofTn5397 These transposon-specific genes were not char-acteristic for any lineage or complex (Table 3) Plasmid-replication genes such as rep2pRE25 rep17pRUM andrep18 pEF418 (26 14 and 7 isolates respectively) andgenes specifying toxin-antitoxin systems (TAS) axe-txeand ω-ε-ζ (12 and 10 isolates respectively) were signifi-cantly associated with the group of isolates of nosocomialE faecium Two other rep genes (rep1pIP501 reppLG1) werewidely distributed among all studied E faecium isolatesand the reppMG1 gene occurred in two isolates (in a singleisolate of ST533 from CC22 and in a singleton ST647) Allω-ε-ζ-positive isolates carried rep2pRE25 and nine isolatesshowed a concomitant presence of axe-txe and rep17pRUMThe relBE genes were found solely among five non-nosocomial E faecium (a difference in distribution with-out statistical significance p = 043) among various STs
DiscussionIn this study Enterococcus spp was used to evaluate theimpact posed by treated wastewater of two local WWTPson their receiver the costal waters of Gulf of GdanskTotal counts of enterococci observed for raw (W-INF)and treated (W-EFF D-EFF) wastewater are consistentwith our previous study showing similar over 99 reduc-tion capacity of applied treatment processes It is alsoreported in the literature that wastewater treatment pro-cesses based on activated sludge although they focus onremoval of nutrients and organic matter simultaneouslyreduce faecal indicators with high efficiency reaching upto 999 [1822] Due to the high number of enterococciin raw wastewater they are nevertheless released from aWWTP in a number up to 61x105 CFU per 100 mLThus marine water sampled in the area directly impactedby treated wastewater (W-MOut and D-MOut) showed ahigher number of enterococci than the one observed in
Vistula River mouth however lower by four orders ofmagnitude in comparison to corresponding treated waste-water (W-EFF and D-EFF) It should be noted that in mar-ine outfalls (W-MOut D-MOut) as well as in VistulaRiver mouth enterococci were detected in number lowerthan 100 CFU per 100 mL ie the value set out by NewBathing Water Directive 20067EC for the costal water ofldquoexcellent qualityrdquo [23]It is suggested that enterococcal species composition in
faecal-impacted environment depends from the source ofcontamination In this study two species most commonlycolonizing humans E faecalis and E faecium comprisedtogether from 73 to 88 of enterococcal isolates intested wastewater samples (W-INF W-BR W-EFF and D-EFF) 73 and 61 in marine outflows (W-MOut and D-MOut respectively) and 45 in the Vistula River mouthThe predominant species in wastewater and marine waterimpacted by treated wastewater was E faecium (from 56to 65) which stayed at a similar level during treatmentprocesses as determined also in our previous study [20]Species distribution in wastewater samples is not wellunderstood and affecting factors may include differencesin diet climate season and methods of detection E feaca-lis was found to be the most prevalent species in wastewa-ter samples in Sweden [26] E faecium in Spain UKCanada France and Switzerland [26-29] while E hiraepredominated in Portugal and United States [2230] Al-though E faecalis is typically the most abundant colonizerof humans in the community (eg [2631]) E faecium isgenerally most often associated with human microbialcontamination [2729] E faecalis was reported fromagriculture-impacted waters [32] and E hirae likelyoriginates mainly from cattle and pig faecal contamin-ation [26] which explains high relative proportion ofthese two species in Vistula River mouth Apart from thevariability of contamination sources proportion of par-ticular species may differ due to the differences of abilityto sustain environmental stresses In this study enterococ-cal composition might have been influenced by wastewa-ter treatment processes and marine water characteristicsThe effect of wastewater treatment and marine environ-
ment was seen on phenotypes of antimicrobial resistanceThe positive selection of bacteria with resistance patternshas been already suggested and noted in wastewater pro-cesses [192022] In this study the resistance rates to ampi-cillin ciprofloxacin levofloxacin and erythromycin notedfor E faecium from raw wastewater (W-INF) were lowerthan these observed for this species in the correspondingtreated wastewater (W-EFF) however not statistically sig-nificant (p gt 005) Moreover resistance rates to ampicillinciprofloxacin tetracycline and erythromycin observed inmarine outflow of WWTP Gdansk-Wschod (W-MOut)were higher than in treated wastewater of this plantSuch phenomenon was not observed for treated
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 11 of 15httpwwwbiomedcentralcom1471-21801466
wastewater of WWTP Gdynia-Debogorze (D-EFF) andits marine outfall (D-MOut) Resistance rates among Efaecium originating from D-MOut were in generalcomparable to these detected in the Vistula Rivermouth It should be noted that treated wastewater fromWWTP Gdansk-Wschod has been discharged via mar-ine outfall (W-MOut) for last 12 years while marineoutfall of WWTP Gdynia-Debogorze (D-MOut) hasbeen operating for last two years The observed differ-ences in resistance rate corresponding to the local im-pact caused by long and short-term operated marineoutfalls (W-MOut and D-MOut) need further attentionSignificant rates of antimicrobial resistance observed
for isolates of E faecalis from wastewater and marineriver waters impacted by wastewater are in agreementwith a recent study performed in six European countriesshowing that in Poland the rates of resistance for humanisolates of this species were usually among the highestboth in the community and hospitals [33] To our know-ledge no corresponding data are available for E faeciumcolonization in the community in Poland but a studyfocused on hospital VRE isolates showed their ubiquitousMDR phenotype and a high load of resistance genes [13]In our study no vancomycin resistance were detectedlikely due to relatively low (below 10) prevalence of VREin Polish hospitals [5] although such isolates are isolatedfrom the environment in other countries eg [34]This study shows that hospital clones of E faecium are
important carriers of resistance genes in wastewatersMost likely in a significant part they originated fromhospital wastewater treated together with communalwastewater In our estimation hospital wastewater con-stitutes approximately 02 of both WWTPsrsquo daily in-flow The release of hospital-associated clones of Efaecium to the environment was observed also by others[24283536] In our study isolates belonging to themajor nosocomial HiRECC (formerly named CC17)constituted altogether 246 of all isolates of E faeciumsubjected to typing and were present at all samplingsites These isolates apart from their inclusion into theHiRECC by MLST demonstrated several other pheno-typic and genotypic features typical for nosocomial Efaecium ie they were resistant to ciprofloxacin andampicillin in vast majority showed MDR phenotypeand carried several resistance determinants associatedboth with mutations of chromosomal genes (gyrA parCpbp5) and in the case of aminoglycoside and tetracyclineresistance with gene acquisition Plasmids likely played animportant role in the later process as some plasmid genessuch as rep2pRE25 rep17pRUM and rep18 pEF418 and TASgenes axe-txe and ω-ε-ζ were significantly more abundantin isolates associated with HiRECC than among theremaining isolates As yet the knowledge of distributionof plasmid-associated genes in populations of E faecium
especially in the context of their clonal compositionremains limited A study on 93 isolates of differentgeographical origins and from various sites showed anover-representation of rep17pRUM and reppMG1 amongnosocomial E faecium while in contrast to our observa-tions rep18 pEF418 was not detected [37] Moreover wedid not observe reppMG1 among HiRECC of E faeciumThese differences may be due to a high prevalence of VRE(58 isolates) among 93 isolates studied by Rosvoll et alwhat may explain differences in the plasmid content Thesame study also reported the co-localization of ω-ε-ζ withrep2pRE25 and axe-txe with rep17pRUM on the same plas-mids in concordance with our findings showing a fre-quent joint occurrence of these pairs of genes Anotherstudy performed on 99 invasive isolates from Norwaymostly of HLGR phenotype showed a significantly higherprevalence of rep2pRE25 and rep17pRUM among the majornosocomial HiRECC [38] This study as well some others[2439] also demonstrated a common presence of reppLG1among E faecium from hospital settings and in animalsOur findings indicate that this type of replicon is evenmore ubiquitous in E faecium than previously reportedAll rep genes mentioned above are typically associatedwith plasmids carrying various antimicrobial resistancegenes [3940] GenBank accession number AF408195 Inaddition to genes determining antimicrobial resistanceand genes associated with MGE isolates of nosocomialHiRECC analyzed in our study carried such molecularmarkers as IS16 and the esp gene together with the intAgene characteristic for ICEEfm [4142] Another patho-genicity factor the fms19 gene of the pilin gene cluster4 (PGC4) showed increased prevalence among theseisolates Similar observation was made for a representa-tive collection of 433 E faecium isolates from varioussources [43]In a study that followed enterococci in a continuum
from hospital and retirement home wastewaters througha WWTP to a river a gradual decrease in the proportionof E faecium isolates representing hospital-associatedSTs was observed likely due to the ldquodilutionrdquo of HiRECCrepresentatives by more diverse and less drug-resistantstrains originating from the community [28] Because inour study only selected representatives of obtained iso-lates were investigated it is not possible to determineprecise tendencies in their frequency from the WWTPinfluent to effluent and water however resistant entero-cocci both HiRECC and non-HiRECC are clearly ableto survive the treatment process what results in their re-lease into marine and river waters Moreover using anadditional typing method MLVA we were able to findisolates with the same characteristics in the effluent ofWWTP and its marine outfall suggesting ability of suchclones to survive in the environment for at least sometime It seems plausible that hospital resistant clones
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 12 of 15httpwwwbiomedcentralcom1471-21801466
accumulate several determinants that may promote theirsurvival not only in the hospital settings but also in theenvironment Such features would include resistance todisinfectants and ultraviolet light additional metabolicpathways and ability to form biofilms Moreover determi-nants of some of the mentioned adaptation factors arefound co-resident in plasmids harboring resistance genessuggesting co-selection For example pLG1-type plasmidsof E faecium carry not only resistance determinantsagainst aminoglycosides glycopeptides and macrolidesbut also pili genes and carbon uptake-utilization genes[3839] Pheromone-responsive plasmids of E faecalisapart from resistance to aminoglycosides glycopeptidespenicillins and macrolides may encode bacteriocins add-itional pili and UV-resistance determinants [40] Thusnosocomial HiRECCs released by WWTPs are equippedin whole sets of genes with various adaptive benefits andoften located within MGE It was also demonstrated thatMGEs can be transferred in a WWTP [44] further under-ling the importance of threat posed by such organismsThere is a selection pressure for survival of resistantstrains and their tranconjugants due to release of antibi-otics to the environment especially in the hospital efflu-ents [45] and even sub-inhibitory concentrations may beeffective in selection for resistance [46]
ConclusionsThis study describes the species composition resistanceprofiles and clonal relationships of enterococci isolatedfrom WWTPs located in Gdansk their marine outfallsto the Gulf of Gdansk and from Vistula River It was ob-served that these WWTPs release enterococcal strainsresistant to various antimicrobial compounds to the re-cipient water environment Molecular analyses revealedthe presence of clones associated with nosocomial HiR-ECCs and in the case of E faecium isolates of lineages17 18 and 78 of former CC17 abundant in resistance de-terminants MGE genes and pathogenicity factors Thepresence of HiRECCs in wastewaters and marineriverenvironment highlights the need for further detailedanalyses to better understand the survival and spread ofdrug-resistant strains in water ecosystems and to eluci-date the ways to curb such dissemination
MethodsAcquisition of samples and isolation of enterococciOnce a month in April June August and October in 2011of the flow proportioned composited samples of wastewa-ter were taken from two local WWTPs Gdansk-Wschodfrom influent (W-INF) effluent (W-EFF) and wastewa-ter from activated sludge bioreactor (W-BR) and fromGdynia-Debogorze effluent (D-EFF) The two WWTPstreat sewage from approximately 920 000 inhabitants(for details see Figure 1) In the same period of time and
with the same frequency marine outfalls of those WWTPs(W-MOut and D-MOut respectively) as well as VistulaRiver mouth were also sampled for isolation of entero-cocci Marine samples were taken at the depth of approxi-mately one meter below the surface once a month inApril June August and October of 2011 Location ofsampling points and their characteristics are provided inFigure 1 Enterococci were detected using the membranefiltration method To this end appropriate dilutions ofanalyzed samples were filtered in duplicate through045 μm cellulose-acetate filters (EMD Millipore Cor-poration Billerica MA USA) which were then placedon Enterococcus selective agar (Merck DarmstadtGermany) and incubated at 37degC for 48 h (ISO 7899ndash22000) Dark red or maroon colonies were consideredpresumptive enterococci For further investigationsrepresentative isolates were taken from membranes pre-senting from 20 to 50 typical colonies and stored in nu-trient broth supplemented with 15 glycerol at minus80degC
Species identification DNA isolation susceptibility testingand statistical analysisThe species identification and drug susceptibility of pre-sumptive enterococci were determined by the PhoenixtradeAutomated Microbiology System (BD New Jersey USA)according to the manufacturerrsquos instructions Since it isnot possible to distinguish E casseliflavus and E galli-narum using this system the classification E casselifla-vusgallinarum was used The susceptibility tests basedon microdilution were carried out for 11 antimicrobialagents gentamicin (GM) streptomycin (ST) ampicillin(AM) daptomycin (DAP) vancomycin (VA) teicoplanin(TEC) linezolid (LZD) fluoroquinolones (ciprofloxacin -CIP levofloxacin - LVX) tetracycline (TE) erythromycin(E) and quinupristin-dalfopristin (SYN) (E faecium only)Obtained minimal inhibitory concentrations (MIC) wereevaluated according to the guidelines of Clinical and La-boratory Standards Institute [47] Isolates were defined asmultidrug-resistant (MDR) when they showed resistanceto three or more compounds tested [48] Bacterial DNAwas purified using the Genomic DNA Prep Plus kitfollowing the manufacturerrsquos instructions (AampA Biotech-nology Gdynia Poland) Differences in distributions wereevaluated using the χ2 test with the p values lt 005 con-sidered significant
MLST and MLVA of E faeciumMLST for E faecium was performed by sequencing ofseven house-keeping genes as described by Homan et al[49] The allele numbers and STs were assigned with theuse of Internet database httpefaeciummlstnet (5th July2013 date last accessed) Novel alleles and allelic profileswere submitted to the database MLST data were analyzedwith Sequence Type Analysis and Recombinational Tests
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 13 of 15httpwwwbiomedcentralcom1471-21801466
(START) software [50] and the eBURST analysis [51] wasused to elucidate the relationships of STs with knownCCs using the comparative eBURST option available athttpeburstmlstnetv3enter_datacomparative (5th July2013 date last accessed) Further inclusion of E faeciumSTs into specific CCs and lineages was performed fol-lowing other studies [924] MLVA was performed as de-scribed by others [52]
Detection and analysis of genes associated with virulenceantimicrobial resistance and MGE in E faeciumThe full list of primers used for gene detection and se-quencing is provided in Additional file 1 Table S1 PCR-based detection of IS16 virulence-associated genes (fms21fms17 fms5 and fms19 representing each of four pilingene clusters in E faecium and the espEfm gene) genes de-termining resistance to tetracycline tet(M) tet(O) tet(L)tet(S) HLGR (aac(6prime)-Ie-aph(2Prime) HLSR (ant(6prime)-Ia) andgenes specific for MGE such as intA of ICEEfm1 int ofTn916 transposon tndX of Tn5397 transposon entero-coccal plasmid replication genes rep1pIP501 rep2pRE25rep17pRUM rep18pEF418 reppMG1 reppLG1 and TAS genesaxe-txe ω-ε-ζ relBE was performed as described in otherstudies [134153-59] following the conditions describedherein Previously characterized clinical isolates of Strepto-coccus agalactiae E faecium and E faecalis harbouringpilin genes IS16 espEfm intA of ICEEfm1 int of Tn916tndX of Tn5397 tet(M) tet(O) tet(L) tet(S) aac(6prime)-Ie-aph(2Prime) ant(6prime)-Ia rep1pIP501 rep2pRE25 ω-ε-ζ [136061]served as positive controls In cases when positive controlswere not available (rep18pEF418 reppMG1 reppLG1 axe-txerelBE) a few randomly selected PCR products wereverified by sequencing and served further as controlsAmpicillin-resistance determinant the pbp5 gene was ana-lysed by sequencing of the part encoding C-terminaltranspeptidase domain in PBP5 [62] New sequence vari-ants pbp5-24 to pbp5-29 alleles were submitted to theGenBank (acc numbers KC594860-KC594865 respect-ively) The quinolone-determining regions (QRDR) in thegyrA and parC genes were sequenced and analysed asdescribed by others [63]
Additional file
Additional file 1 Table S1 Primers used in the study for PCR andsequencing
Competing interestThe authors declare that they have no competing interest
Authorsrsquo contributionsES participated in the study design performed experimental work andparticipated in the manuscript preparation AL designed the studyperformed experimental work and participated in the manuscriptpreparation Both authors read and approved the final manuscript
AcknowledgementsWe thank Rob Willems and Janetta Top for assigning new alleles new STsand MTs This publication made use of the Enterococcus faecium MLSTwebsite (httpefaeciummlstnet) which is hosted at Imperial College of theUniversity of Oxford and funded by the Wellcome Trust This work wassupported by project N N305 461139 from the Narodowe Centrum Nauki(NCN)
Author details1National Medicines Institute ul Chelmska 3034 Warsaw 00-725 Poland2Gdansk University of Technology ul G Narutowicza 1112 Gdansk 80-952Poland
Received 5 September 2013 Accepted 27 February 2014Published 14 March 2014
References1 Aarestrup FM Butaye P Witte W Nonhuman reservoirs of Enterococci In
American Society for Microbiology Edited by Gilmore MS Clewell DB CourvalinP Dunny GM Murray BE Rice LB Washington DC The Enterococcipathogenesis molecular biology and antibiotic resistance 2002
2 Tendolkar PM Baghdayan AS Shankar N Pathogenic enterococci newdevelopments in the 21st century Cell Mol Life Sci 2003 602622ndash2636
3 Sood S Malhotra M Das BK Kapil A Enterococcal infections ampantimicrobial resistance Indian J Med Res 2008 128111ndash121
4 Sydnor ER Perl TM Hospital epidemiology and infection control inacute-care settings Clin Microbiol Rev 2011 24141ndash173
5 European Centre for Disease Prevention and Control Annual EpidemiologicalReport Reporting on 2010 surveillance data and 2011 epidemic intelligencedata Stockholm ECDC 20122013
6 Hidron AI Edwards JR Patel J Horan TC Sievert DM Pollock DA Fridkin SKNational Healthcare Safety Network Team Participating National HealthcareSafety Network Facilities NHSN annual update antimicrobial-resistantpathogens associated with healthcare-associated infections annualsummary of data reported to the National Healthcare Safety Network atthe Centers for Disease Control and Prevention 2006ndash2007 Infect ControlHosp Epidemiol 2008 29996ndash1011
7 De Kraker ME Jarlier V Monen JC Heuer OE van de Sande N GrundmannH The changing epidemiology of bacteraemias in Europe trends fromthe European Antimicrobial Resistance Surveillance System Clin MicrobiolInfect 2012 doi1011111469-0691
8 Leavis HL Bonten MJ Willems RJ Identification of high-risk enterococcalclonal complexes global dispersion and antibiotic resistance Curr OpinMicrobiol 2006 9454ndash460
9 Willems RJL Top J van Schaik W Leavis H Bonten M Sireacuten J Hanage WPCorander J Restricted gene flow among hospital subpopulations ofEnterococcus faecium mBio 2012 3e00151ndash12
10 Hegstad K Mikalsen T Coque TM Werner G Sundsfjord A Mobile geneticelements and their contribution to the emergence of antimicrobialresistant Enterococcus faecalis and Enterococcus faecium Clin MicrobiolInfect 2010 16541ndash554
11 Shepard BD Gilmore MS Antibiotic-resistant enterococci themechanisms and dynamics of drug introduction and resistance MicrobesInfect 2002 4215ndash224
12 Bonten MJ Willems R Weinstein RA Vancomycin-resistant enterococciwhy are they here and where do they come from Lancet Infect Dis 20011314ndash325
13 Sadowy E Sieńko A Gawryszewska I Bojarska A Malinowska K HryniewiczW High abundance and diversity of antimicrobial resistancedeterminants among early vancomycin-resistant Enterococcus faecium inPoland Eur J Clin Microbiol Infect Dis 2013 321193ndash1203
14 Willems RJ Homan W Top J van Santen-Verheuvel M Tribe D Manzioros XGaillard C Vandenbroucke-Grauls CM Mascini EM van Kregten E vanEmbden JD Bonten MJ Variant esp gene as a marker of a distinct geneticlineage of vancomycin-resistant Enterococcus faecium spreading inhospitals Lancet 2001 357853ndash855
15 Sillanpaumlauml J Nallapareddy SR Prakash VP Qin X Houmloumlk M Weinstock GMMurray BE Identification and phenotypic characterization of a secondcollagen adhesin Scm and genome-based identification and analysis of13 other predicted MSCRAMMs including four distinct pilus loci inEnterococcus faecium Microbiology 2008 1543199ndash3211
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 14 of 15httpwwwbiomedcentralcom1471-21801466
16 Galloway-Pentildea JR Nallapareddy SR Arias CA Eliopoulos GM Murray BEAnalysis of clonality and antibiotic resistance among early clinicalisolates of Enterococcus faecium in the United States J Infect Dis 20092001566ndash1573
17 Top J Willems R Bonten M Emergence of CC17 Enterococcus faeciumfrom commensal to hospital-adapted pathogen FEMS Immunol MedMicrobiol 2008 52297ndash308
18 Martins da Costa P Vaz-Pires P Bernardo F Antimicrobial resistance inEnterococcus spp isolated in inflow effluent and sludge from municipalsewage water treatment plants Water Res 2006 401735ndash1740
19 Reinthaler FF Posch J Feierl G Wust G Haas D Ruckenbauer G Mascher FMarth E Antibiotic resistance of Escherichia coli in sewage and sludgeWater Res 2003 371685ndash1690
20 Luczkiewicz A Fudala-Ksiazek S Jankowska K Olanczuk-Neyman KAntimicrobial resistance of fecal indicators in municipal wastewatertreatment plant Water Res 2010 445089ndash5097
21 Dulfour AP Ballentine RK Ambient water quality criteria for bacteria-1986 EPA4405-84-002 Washington DC US Environmental Protection Agency 1986
22 Ferreira da Silva M Tiago I Veriacutessimo A Boaventura RA Nunes OC Manaia CMAntimicrobial resistance patterns in Enterobacteriaceae isolated from anurban wastewater treatment plant FEMS Microbiol Ecol 2007 60166ndash176
23 DIRECTIVE 20067EC of the European Parliament and of the Council 15February 2006 concerning the management of bathing water quality andrepealing Directive 76160EEC httpeur-lexeuropaeuLexUriServLexUriServdouri=OJL200606400370051ENPDF
24 Freitas AR Novais C Ruiz-Garbajosa P Coque TM Peixe L Dispersion ofmultidrug-resistant Enterococcus faecium isolates belonging to majorclonal complexes in different Portuguese settings Appl Environ Microbiol2009 754904ndash4908
25 Poeta P Costa D Igrejas G Saacuteenz Y Zarazaga M Rodrigues J Torres CPolymorphisms of the pbp5 gene and correlation with ampicillinresistance in Enterococcus faecium isolates of animal origin J MedMicrobiol 2007 56236ndash240
26 Kuumlhn I Iversen A Burman LG Olsson-Liljequist B Franklin A Finn MAarestrup F Seyfarth AM Blanch AR Vilanova X Taylor H Caplin J MorenoMA Dominguez L Herrero IA Moumlllby R Comparison of enterococcalpopulations in animals humans and the environment ndash a Europeanstudy Int J Food Microbiol 2003 88133ndash145
27 Lanthier M Scott A Lapen DR Zhang Y Topp E Frequency of virulencegenes and antibiotic resistances in Enterococcus spp isolates fromwastewater and feces of domesticated mammals and birds and wildlifeCan J Microbiol 2010 56715ndash729
28 Leclercq R Oberleacute K Galopin S Cattoir V Budzinski H Petit F Changes inenterococcal populations and related antibiotic resistance along amedical center-wastewater treatment plant-river continuum Appl EnvironMicrobiol 2013 792428ndash2434
29 Thevenon F Regier N Benagli C Tonolla M Adatte T Wildi W Poteacute JCharacterization of fecal indicator bacteria in sediments cores from thelargest freshwater lake of Western Europe (Lake Geneva Switzerland)Ecotoxicol Environ Safety 2012 7850ndash56
30 Bonilla TD Nowosielski K Esiobu N McCorquodale DS Rogerson ASpecies assemblages of Enterococcus indicate potential sources offecal bacteria at a south Florida recreational beach Mar Pollut Bull2006 52807ndash810
31 Wang JT Chang SC Wang HY Chen PC Shiau YR Lauderdale TL TSARHospitals High rates of multidrug resistance in Enterococcus faecalis andE faecium isolated from inpatients and outpatients in TaiwanDiagn Microbiol Infect Dis 2013 75406ndash411
32 Lanthier M Scott A Zhang Y Cloutier M Durie D Henderson VC WilkesG Lapen DR Topp E Distribution of selected virulence genes andantibiotic resistance in Enterococcus species isolated from the SouthNation River drainage basin Ontario Canada J Appl Microbiol 2011110407ndash421
33 Kuch A Willems RJ Werner G Coque TM Hammerum AM Sundsfjord AKlare I Ruiz-Garbajosa P Simonsen GS van Luit-Asbroek M Hryniewicz WSadowy E Insight into antimicrobial susceptibility and population struc-ture of contemporary human Enterococcus faecalis isolates fromEurope J Antimicrob Chemother 2012 67551ndash558
34 Novais C Coque TM Ferreira H Sousa JC Peixe L Environmentalcontamination with vancomycin-resistant enterococci from hospitalsewage in Portugal Appl Environ Microbiol 2005 713364ndash3368
35 Iversen A Kuumlhn I Rahman M Franklin A Burman LG Olsson-Liljequist BTorell E Moumlllby R Evidence for transmission between humans and theenvironment of a nosocomial strain of Enterococcus faeciumEnviron Microbiol 2004 655ndash59
36 Rathnayake I Hargreaves M Huygens F SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterwayAustralia and associated antibiotic resistance gene profilesBMC Microbiol 2011 11201
37 Rosvoll TC Pedersen T Sletvold H Johnsen PJ Sollid JE Simonsen GSJensen LB Nielsen KM Sundsfjord A PCR-based plasmid typing inEnterococcus faecium strains reveals widely distributed pRE25-pRUM- pIP501- and pHT-related replicons associated with glycopeptideresistance and stabilizing toxin-antitoxin systems FEMS Immunol MedMicrobiol 2010 58254ndash268
38 Rosvoll TC Lindstad BL Lunde TM Hegstad K Aasnaes B Hammerum AMLester CH Simonsen GS Sundsfjord A Pedersen T Increased high-levelgentamicin resistance in invasive Enterococcus faecium is associated withaac(6prime)Ie-aph(2Prime)Ia-encoding transferable megaplasmids hosted by majorhospital-adapted lineages FEMS Immunol Med Microbiol 2012 66166ndash176
39 Laverde Gomez JA van Schaik W Freitas AR Coque TM Weaver KE FranciaMV Witte W Werner G A multiresistance megaplasmid pLG1 bearing ahylEfm genomic island in hospital Enterococcus faecium isolates Int J MedMicrobiol 2011 301165ndash175
40 Coque TM Evolutionary biology of pathogenic enterococci InEvolutionary Biology of Bacterial and Fungal Pathogens Coque TM Edited byBaquero F Nombela C Cassell GH Gutierrez JA Washington DC ASMPress501ndash523
41 Werner G Fleige C Geringer U van Schaik W Klare I Witte W IS elementIS16 as a molecular screening tool to identify hospital-associated strainsof Enterococcus faecium BMC Infect Dis 2011 1180
42 Top J Sinnige JC Majoor EA Bonten MJ Willems RJ van Schaik W Therecombinase IntA is required for excision of esp-containing ICEEfm1 inEnterococcus faecium J Bacteriol 2011 1931003ndash1006
43 Sillanpaumlauml J Prakash VP Nallapareddy SR Murray BE Distribution of genesencoding MSCRAMMs and Pili in clinical and natural populations ofEnterococcus faecium J Clin Microbiol 2009 47896ndash901
44 Marcinek H Wirth R Muscholl-Silberhorn A Gauer M Enterococcus faecalisgene transfer under natural conditions in municipal sewage watertreatment plants Appl Environ Microbiol 1998 64626ndash632
45 Kuumlmmerer K Significance of antibiotics in the environment J AntimicrobChemother 2003 525ndash7
46 Liu A Fong A Becket E Yuan J Tamae C Medrano L Maiz M Wahba C LeeC Lee K Tran KP Yang H Hoffman RM Salih A Miller JHP Selectiveadvantage of resistant strains at trace levels of antibiotics a simple andultrasensitive color test for detection of antibiotics and genotoxicagents Antimicrob Agents Chemother 2011 551204ndash1210
47 Clinical and Laboratory Standards Institute Performance standards forantimicrobial susceptibility testing 21st informational supplement Wayne CLSI2011 PA M100-S20
48 Magiorakos AP Srinivasan A Carey RB Carmeli Y Falagas ME Giske CGHarbarth S Hindler JF Kahlmeter G Olsson-Liljequist B Paterson DL Rice LBStelling J Struelens MJ Vatopoulos A Weber JT Monnet DLMultidrug-resistant extensively drug-resistant and pandrug-resistantbacteria an international expert proposal for interim standarddefinitions for acquired resistance Clin Microbiol Infect 2012 18268ndash281
49 Homan WL Tribe D Poznanski S Li M Hogg G Spalburg E Van Embden JDWillems RJ Multilocus sequence typing scheme for Enterococcus faeciumJ Clin Microbiol 2002 401963ndash1971
50 Jolley KA Feil EJ Chan MS Maiden MC Sequence type analysis andrecombinational tests (START) Bioinformatics 2001 171230ndash1231
51 Feil EJ Li BC Aanensen DM Hanage WP Spratt BG eBURST inferringpatterns of evolutionary descent among clusters of related bacterialgenotypes from multilocus sequence typing data J Bacteriol 20041861518ndash1530
52 Top J Banga1 NMI Hayes R Willems RJ Bonten MJM Hayden MKComparison of multiple-locus variable-number tandem repeat analysisand pulsed-field gel electrophoresis in a setting of polyclonal endemicityof vancomycin-resistant Enterococcus faecium Clin Microbiol Infect 200814363ndash369
53 Vankerckhoven V Van Autgaerden T Vael C Lammens C Chapelle S RossiR Jabes D Goossens H Development of a multiplex PCR for the
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-
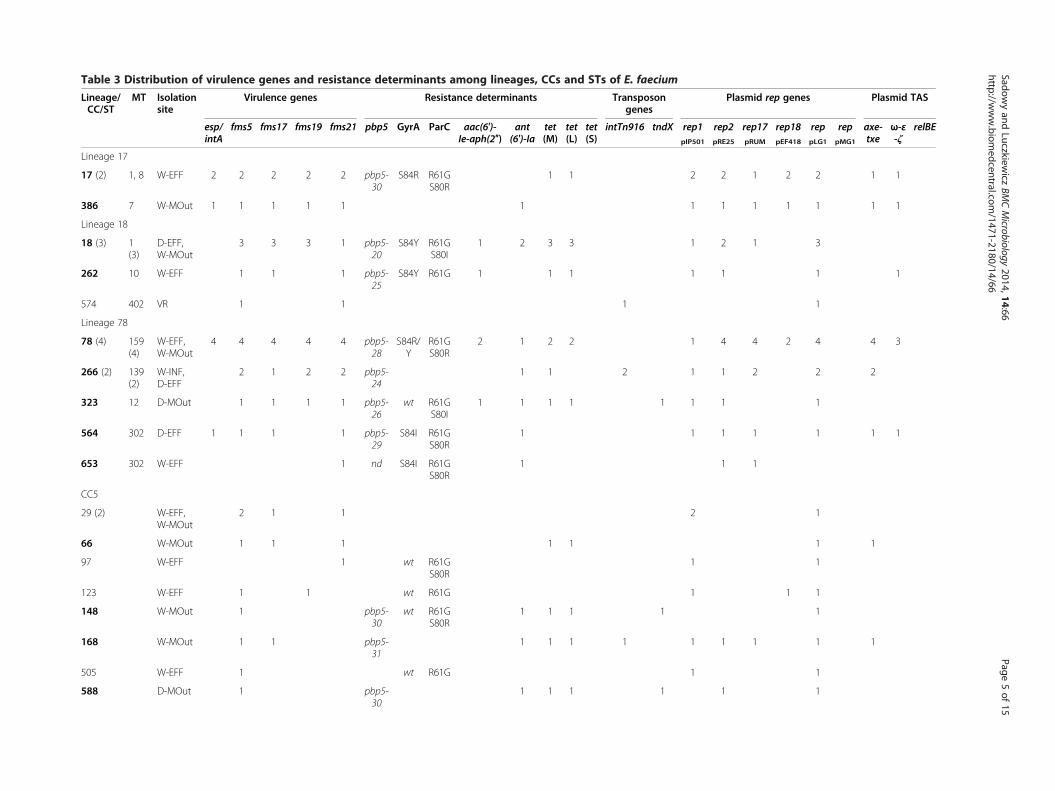
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium
LineageCCST
MT Isolationsite
Virulence genes Resistance determinants Transposongenes
Plasmid rep genes Plasmid TAS
espintA
fms5 fms17 fms19 fms21 pbp5 GyrA ParC aac(6prime)-Ie-aph(2Prime)
ant(6prime)-Ia
tet(M)
tet(L)
tet(S)
intTn916 tndX rep1pIP501
rep2pRE25
rep17pRUM
rep18pEF418
reppLG1
reppMG1
axe-txe
ω-ε-ζ
relBE
Lineage 17
17 (2) 1 8 W-EFF 2 2 2 2 2 pbp5-30
S84R R61GS80R
1 1 2 2 1 2 2 1 1
386 7 W-MOut 1 1 1 1 1 1 1 1 1 1 1 1 1
Lineage 18
18 (3) 1(3)
D-EFFW-MOut
3 3 3 1 pbp5-20
S84Y R61GS80I
1 2 3 3 1 2 1 3
262 10 W-EFF 1 1 1 pbp5-25
S84Y R61G 1 1 1 1 1 1 1
574 402 VR 1 1 1 1
Lineage 78
78 (4) 159(4)
W-EFFW-MOut
4 4 4 4 4 pbp5-28
S84RY
R61GS80R
2 1 2 2 1 4 4 2 4 4 3
266 (2) 139(2)
W-INFD-EFF
2 1 2 2 pbp5-24
1 1 2 1 1 2 2 2
323 12 D-MOut 1 1 1 1 pbp5-26
wt R61GS80I
1 1 1 1 1 1 1 1
564 302 D-EFF 1 1 1 1 pbp5-29
S84I R61GS80R
1 1 1 1 1 1 1
653 302 W-EFF 1 nd S84I R61GS80R
1 1 1
CC5
29 (2) W-EFFW-MOut
2 1 1 2 1
66 W-MOut 1 1 1 1 1 1 1
97 W-EFF 1 wt R61GS80R
1 1
123 W-EFF 1 1 wt R61G 1 1 1
148 W-MOut 1 pbp5-30
wt R61GS80R
1 1 1 1 1
168 W-MOut 1 1 pbp5-31
1 1 1 1 1 1 1 1 1
505 W-EFF 1 wt R61G 1 1
588 D-MOut 1 pbp5-30
1 1 1 1 1 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
5of
15httpw
wwbiom
edcentralcom1471-21801466
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium (Continued)
635 D-EFF 1 1 1 1
636 W-EFF 1 wt R61G 1
637 W-EFF 1 1 1 wt R61G 1 1
638 W-EFF 1 wt R61GS80I
1
642 W-EFF 1 1 1 1 1
645 D-EFF 1 1 1 1 1
646 W-EFF 1 1 1 1 1
649 W-BR 1 1 1 pbp5-32
wt R61G 1
650 (3) D-EFF 3 3 pbp5-27
2 2 2 1 1 3
CC 9
8 D-EFF 1 1 1 1 1 1 1 1 1 1 1
640 W-EFF 1 1 1 1 1
CC 22
21 W-EFF 1 1 1 1 1 1 1 1
22 W-EFF 1 1 1 1 1 1
25 W-EFF 1 1 pbp5-32
wt R61G 1 1 1
32 (3) W-EFF 3 3 1 wt R61G 2 1
92 W-EFF 1 1 1 1 1
441 W-EFF 1 1 1 1 1 1 1 1
533 W-EFF 1 1 1
633 W-EFF 1 1 1 1 1
644 D-EFF 1 1 1 1 1
CC27
652 W-EFF 1 1 1 1
CC46
47 VR 1 1 1 1
69 D-EFF 1 1 1 1 1 1
CC94
94 W-MOut 1 1 1 1
361 (4) D-EFFD-MOut
3 4 4 4 pbp5-29
1 1 1 4 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
6of
15httpw
wwbiom
edcentralcom1471-21801466
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium (Continued)
CC296
296 W-EFF 1 1
CC639
639 W-EFF 1 1 1 1 1
CC648
634 W-EFF 1 1 1 1 1
648 W-EFF 1 1 1 1 1
Doubletons
632 W-MOut 1 1 1
Singletons
264 W-INF 1 1 1 1 1 1 1
641 D-EFF 1 1 1 1
643 W-EFF 1 1 1 1 1 1
647 W-MOut 1 1 1 1 1
651 D-EFF 1 1 1 1 1 1
654 VR 1 1 1 1 1 1
All (69) 8 57 49 29 46 6 17 20 17 1 9 4 39 27 14 7 55 2 12 10 5
CC clonal complex ST sequence type MT MLVA-type nd not determined due to lack of amplification of the pbp5 gene wt wild-type STs associated with MDR phenotype in boldAbbreviations of sampling pointsrsquo locations and their description provided in Figure 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
7of
15httpw
wwbiom
edcentralcom1471-21801466
Figure 2 MLST-based similarity tree of E faecium isolates constructed using the START software and the UPGMA clustering algorithm
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 8 of 15httpwwwbiomedcentralcom1471-21801466
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 9 of 15httpwwwbiomedcentralcom1471-21801466
grouped the 38 STs specific for 50 isolates into the majorlsquosuper-complex 17rsquo characteristic for this species (data notshown) Further detailed analysis of the localization ofparticular STs within this structure revealed that 11 STscharacteristic for 17 isolates belonged to the nosocomialcomplex of E faecium representing all three majorlineages 17 18 and 78 of a former CC17 as defined byWillems et al [9] (Table 3) Seventeen STs (20 isolates)nine STs (11 isolates) and two STs with single isolates eachbelonged to CC5 CC22 and CC9 respectively as definedby Freitas et al [24] Of the remaining 16 STs nine wereincluded into six small CCs (CCs 27 46 94 296 639648) unlinked to the major lsquosuper-complexrsquo ST632formed a lsquodoubletonrsquo (ie was linked by a single-locus vari-ant link to only one ST from the database) and six STsrepresented by single isolates were singletons ie werenot related to any ST in the database at the time of ana-lysis Thirty isolates (435) in the investigated groupshowed the MDR phenotype which was significantly asso-ciated (p = 000006) with isolates from the nosocomialcomplex (Table 3 and Figure 3) but found also in CC5CC22 and CC94 this phenotype did not occur amongsmall CCs or singletons Seventeen isolates repre-senting the nosocomial complex were additionallystudied by multilocus VNTR (variable number tan-dem repeats) analysis (MLVA) to better elucidate therelationships among these isolates Altogether nineMLVA-types (MTs) were found (Table 3) Importantlythe same MLVA-type MT159 associated with ST78was characteristic for isolates derived from samplesfrom W-EFF and W-MOut suggesting survival of thisparticular clone
0
10
20
30
40
50
60
70
80
90
100
IS16
espintA
fms5
fms1
7
fms1
9
fms2
1
MDR
AM-R
CIP-R
HLG
R
p
ositi
ve
isol
ates
Figure 3 Distribution () of IS16 resistance phenotypes and virulenccolumns isolates with STs typical for nosocomial E faecium grey columnsgroups are indicated by an asterisk MDR multi-drug resistance AM-R ampgentamicin resistance HLSR high-level streptomycin resistance
IS16 was present in all 17 isolates included by MLST tothree lineages of nosocomial E faecium and this markerwas limited to this group (Table 3 and Figure 3) TheespEfm gene occurred always together with intA among allisolates of lineage 17 and in some isolates of lineage 78The fms19 pilin gene detected in 29 isolates was over-represented among STs typical for nosocomial E faeciumwhile other pilin genes such as fms5 fms17 and fms21detected in 57 49 and 46 isolates respectively occurredamong various CCs and singleton STs Ampicillin resist-ance significantly associated with isolates of nosocomialcomplex (10 of 17 isolates 588 p = 0001) occurred alsoin CC5 (6 isolates) CC22 and CC94 (1 isolate each) Se-quencing of the 3prime part of the pbp5 gene revealed its 11nucleotide alleles corresponding to 11 variants of the C-terminal part of PBP5 Six alleles of the gene were new(pbp5-24 to pbp5-29) two alleles (pbp5-20 pbp5-22)were described previously among Polish hospital VREmisolates [13] and three alleles were found in other stud-ies pbp5-30 occurred in a Portuguese animal strain ofE faecium [25] and pbp5-31 and pbp5-32 in clinical iso-lates TCGE70411_2 and 164306 also from Portugal (de-posited as JN208888 and JN208885 in the GenBankrespectively) For a single ampicillin-resistant isolate ofST653 (lineage 78) pbp5 amplification was not achieveddespite repeated attempts The majority (11 of 17 iso-lates 647) of isolates belonging to three lineages ofnosocomial E faecium showed resistance to ciprofloxa-cin determined by mutations R61G and S80IR in ParCand S84IRY in GyrA Ciprofloxacin resistance wasalso significantly over-represented among isolates ofSTs typical for nosocomial E faecium (p = 00006) Nine
HLS
R
tetM tetL
rep1
-pIP50
1
rep2
-pRE25
rep1
7-pR
UM
rep1
8-pE
F418
rep-pL
G1
axe-txe
relBE
e resistance and plasmid genes among E faecium isolates Blackother E faecium isolates Significant differences (p lt 005) between twoicillin resistance CIP-R ciprofloxacin resistance HLGR high-level
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 10 of 15httpwwwbiomedcentralcom1471-21801466
isolates resistant to ciprofloxacin were found also inCCs 5 9 and 22 These isolates possessed mutations inParC mentioned above but had the wild-type QRDR inGyrA HLGR occurred in five isolates of lineages 18 and78 and in single isolates of CC5 and CC22 this pheno-type was determined by the presence of aac(6prime)-Ie-aph(2Prime) with an exception of an isolate from CC5 whichwas negative for this gene High-level resistance tostreptomycin (HLSR) was widely distributed amongstudied isolates and determined by the ant(6prime)-Ia geneTetracycline resistance was found also in various STsthe major determinant of this phenotype was tet(M) (20isolates) followed by tet(L) (17 isolates) These twogenes occurred together in 16 isolates A single isolateof ST633 belonging to CC22 harbored tet(S) The tet(O) gene was absent in the studied group For a singletetracycline-resistant isolate of ST17 none of the fourtetracycline determinants studied was detected Nineisolates carried int of Tn916 and four the tndX gene ofTn5397 These transposon-specific genes were not char-acteristic for any lineage or complex (Table 3) Plasmid-replication genes such as rep2pRE25 rep17pRUM andrep18 pEF418 (26 14 and 7 isolates respectively) andgenes specifying toxin-antitoxin systems (TAS) axe-txeand ω-ε-ζ (12 and 10 isolates respectively) were signifi-cantly associated with the group of isolates of nosocomialE faecium Two other rep genes (rep1pIP501 reppLG1) werewidely distributed among all studied E faecium isolatesand the reppMG1 gene occurred in two isolates (in a singleisolate of ST533 from CC22 and in a singleton ST647) Allω-ε-ζ-positive isolates carried rep2pRE25 and nine isolatesshowed a concomitant presence of axe-txe and rep17pRUMThe relBE genes were found solely among five non-nosocomial E faecium (a difference in distribution with-out statistical significance p = 043) among various STs
DiscussionIn this study Enterococcus spp was used to evaluate theimpact posed by treated wastewater of two local WWTPson their receiver the costal waters of Gulf of GdanskTotal counts of enterococci observed for raw (W-INF)and treated (W-EFF D-EFF) wastewater are consistentwith our previous study showing similar over 99 reduc-tion capacity of applied treatment processes It is alsoreported in the literature that wastewater treatment pro-cesses based on activated sludge although they focus onremoval of nutrients and organic matter simultaneouslyreduce faecal indicators with high efficiency reaching upto 999 [1822] Due to the high number of enterococciin raw wastewater they are nevertheless released from aWWTP in a number up to 61x105 CFU per 100 mLThus marine water sampled in the area directly impactedby treated wastewater (W-MOut and D-MOut) showed ahigher number of enterococci than the one observed in
Vistula River mouth however lower by four orders ofmagnitude in comparison to corresponding treated waste-water (W-EFF and D-EFF) It should be noted that in mar-ine outfalls (W-MOut D-MOut) as well as in VistulaRiver mouth enterococci were detected in number lowerthan 100 CFU per 100 mL ie the value set out by NewBathing Water Directive 20067EC for the costal water ofldquoexcellent qualityrdquo [23]It is suggested that enterococcal species composition in
faecal-impacted environment depends from the source ofcontamination In this study two species most commonlycolonizing humans E faecalis and E faecium comprisedtogether from 73 to 88 of enterococcal isolates intested wastewater samples (W-INF W-BR W-EFF and D-EFF) 73 and 61 in marine outflows (W-MOut and D-MOut respectively) and 45 in the Vistula River mouthThe predominant species in wastewater and marine waterimpacted by treated wastewater was E faecium (from 56to 65) which stayed at a similar level during treatmentprocesses as determined also in our previous study [20]Species distribution in wastewater samples is not wellunderstood and affecting factors may include differencesin diet climate season and methods of detection E feaca-lis was found to be the most prevalent species in wastewa-ter samples in Sweden [26] E faecium in Spain UKCanada France and Switzerland [26-29] while E hiraepredominated in Portugal and United States [2230] Al-though E faecalis is typically the most abundant colonizerof humans in the community (eg [2631]) E faecium isgenerally most often associated with human microbialcontamination [2729] E faecalis was reported fromagriculture-impacted waters [32] and E hirae likelyoriginates mainly from cattle and pig faecal contamin-ation [26] which explains high relative proportion ofthese two species in Vistula River mouth Apart from thevariability of contamination sources proportion of par-ticular species may differ due to the differences of abilityto sustain environmental stresses In this study enterococ-cal composition might have been influenced by wastewa-ter treatment processes and marine water characteristicsThe effect of wastewater treatment and marine environ-
ment was seen on phenotypes of antimicrobial resistanceThe positive selection of bacteria with resistance patternshas been already suggested and noted in wastewater pro-cesses [192022] In this study the resistance rates to ampi-cillin ciprofloxacin levofloxacin and erythromycin notedfor E faecium from raw wastewater (W-INF) were lowerthan these observed for this species in the correspondingtreated wastewater (W-EFF) however not statistically sig-nificant (p gt 005) Moreover resistance rates to ampicillinciprofloxacin tetracycline and erythromycin observed inmarine outflow of WWTP Gdansk-Wschod (W-MOut)were higher than in treated wastewater of this plantSuch phenomenon was not observed for treated
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 11 of 15httpwwwbiomedcentralcom1471-21801466
wastewater of WWTP Gdynia-Debogorze (D-EFF) andits marine outfall (D-MOut) Resistance rates among Efaecium originating from D-MOut were in generalcomparable to these detected in the Vistula Rivermouth It should be noted that treated wastewater fromWWTP Gdansk-Wschod has been discharged via mar-ine outfall (W-MOut) for last 12 years while marineoutfall of WWTP Gdynia-Debogorze (D-MOut) hasbeen operating for last two years The observed differ-ences in resistance rate corresponding to the local im-pact caused by long and short-term operated marineoutfalls (W-MOut and D-MOut) need further attentionSignificant rates of antimicrobial resistance observed
for isolates of E faecalis from wastewater and marineriver waters impacted by wastewater are in agreementwith a recent study performed in six European countriesshowing that in Poland the rates of resistance for humanisolates of this species were usually among the highestboth in the community and hospitals [33] To our know-ledge no corresponding data are available for E faeciumcolonization in the community in Poland but a studyfocused on hospital VRE isolates showed their ubiquitousMDR phenotype and a high load of resistance genes [13]In our study no vancomycin resistance were detectedlikely due to relatively low (below 10) prevalence of VREin Polish hospitals [5] although such isolates are isolatedfrom the environment in other countries eg [34]This study shows that hospital clones of E faecium are
important carriers of resistance genes in wastewatersMost likely in a significant part they originated fromhospital wastewater treated together with communalwastewater In our estimation hospital wastewater con-stitutes approximately 02 of both WWTPsrsquo daily in-flow The release of hospital-associated clones of Efaecium to the environment was observed also by others[24283536] In our study isolates belonging to themajor nosocomial HiRECC (formerly named CC17)constituted altogether 246 of all isolates of E faeciumsubjected to typing and were present at all samplingsites These isolates apart from their inclusion into theHiRECC by MLST demonstrated several other pheno-typic and genotypic features typical for nosocomial Efaecium ie they were resistant to ciprofloxacin andampicillin in vast majority showed MDR phenotypeand carried several resistance determinants associatedboth with mutations of chromosomal genes (gyrA parCpbp5) and in the case of aminoglycoside and tetracyclineresistance with gene acquisition Plasmids likely played animportant role in the later process as some plasmid genessuch as rep2pRE25 rep17pRUM and rep18 pEF418 and TASgenes axe-txe and ω-ε-ζ were significantly more abundantin isolates associated with HiRECC than among theremaining isolates As yet the knowledge of distributionof plasmid-associated genes in populations of E faecium
especially in the context of their clonal compositionremains limited A study on 93 isolates of differentgeographical origins and from various sites showed anover-representation of rep17pRUM and reppMG1 amongnosocomial E faecium while in contrast to our observa-tions rep18 pEF418 was not detected [37] Moreover wedid not observe reppMG1 among HiRECC of E faeciumThese differences may be due to a high prevalence of VRE(58 isolates) among 93 isolates studied by Rosvoll et alwhat may explain differences in the plasmid content Thesame study also reported the co-localization of ω-ε-ζ withrep2pRE25 and axe-txe with rep17pRUM on the same plas-mids in concordance with our findings showing a fre-quent joint occurrence of these pairs of genes Anotherstudy performed on 99 invasive isolates from Norwaymostly of HLGR phenotype showed a significantly higherprevalence of rep2pRE25 and rep17pRUM among the majornosocomial HiRECC [38] This study as well some others[2439] also demonstrated a common presence of reppLG1among E faecium from hospital settings and in animalsOur findings indicate that this type of replicon is evenmore ubiquitous in E faecium than previously reportedAll rep genes mentioned above are typically associatedwith plasmids carrying various antimicrobial resistancegenes [3940] GenBank accession number AF408195 Inaddition to genes determining antimicrobial resistanceand genes associated with MGE isolates of nosocomialHiRECC analyzed in our study carried such molecularmarkers as IS16 and the esp gene together with the intAgene characteristic for ICEEfm [4142] Another patho-genicity factor the fms19 gene of the pilin gene cluster4 (PGC4) showed increased prevalence among theseisolates Similar observation was made for a representa-tive collection of 433 E faecium isolates from varioussources [43]In a study that followed enterococci in a continuum
from hospital and retirement home wastewaters througha WWTP to a river a gradual decrease in the proportionof E faecium isolates representing hospital-associatedSTs was observed likely due to the ldquodilutionrdquo of HiRECCrepresentatives by more diverse and less drug-resistantstrains originating from the community [28] Because inour study only selected representatives of obtained iso-lates were investigated it is not possible to determineprecise tendencies in their frequency from the WWTPinfluent to effluent and water however resistant entero-cocci both HiRECC and non-HiRECC are clearly ableto survive the treatment process what results in their re-lease into marine and river waters Moreover using anadditional typing method MLVA we were able to findisolates with the same characteristics in the effluent ofWWTP and its marine outfall suggesting ability of suchclones to survive in the environment for at least sometime It seems plausible that hospital resistant clones
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 12 of 15httpwwwbiomedcentralcom1471-21801466
accumulate several determinants that may promote theirsurvival not only in the hospital settings but also in theenvironment Such features would include resistance todisinfectants and ultraviolet light additional metabolicpathways and ability to form biofilms Moreover determi-nants of some of the mentioned adaptation factors arefound co-resident in plasmids harboring resistance genessuggesting co-selection For example pLG1-type plasmidsof E faecium carry not only resistance determinantsagainst aminoglycosides glycopeptides and macrolidesbut also pili genes and carbon uptake-utilization genes[3839] Pheromone-responsive plasmids of E faecalisapart from resistance to aminoglycosides glycopeptidespenicillins and macrolides may encode bacteriocins add-itional pili and UV-resistance determinants [40] Thusnosocomial HiRECCs released by WWTPs are equippedin whole sets of genes with various adaptive benefits andoften located within MGE It was also demonstrated thatMGEs can be transferred in a WWTP [44] further under-ling the importance of threat posed by such organismsThere is a selection pressure for survival of resistantstrains and their tranconjugants due to release of antibi-otics to the environment especially in the hospital efflu-ents [45] and even sub-inhibitory concentrations may beeffective in selection for resistance [46]
ConclusionsThis study describes the species composition resistanceprofiles and clonal relationships of enterococci isolatedfrom WWTPs located in Gdansk their marine outfallsto the Gulf of Gdansk and from Vistula River It was ob-served that these WWTPs release enterococcal strainsresistant to various antimicrobial compounds to the re-cipient water environment Molecular analyses revealedthe presence of clones associated with nosocomial HiR-ECCs and in the case of E faecium isolates of lineages17 18 and 78 of former CC17 abundant in resistance de-terminants MGE genes and pathogenicity factors Thepresence of HiRECCs in wastewaters and marineriverenvironment highlights the need for further detailedanalyses to better understand the survival and spread ofdrug-resistant strains in water ecosystems and to eluci-date the ways to curb such dissemination
MethodsAcquisition of samples and isolation of enterococciOnce a month in April June August and October in 2011of the flow proportioned composited samples of wastewa-ter were taken from two local WWTPs Gdansk-Wschodfrom influent (W-INF) effluent (W-EFF) and wastewa-ter from activated sludge bioreactor (W-BR) and fromGdynia-Debogorze effluent (D-EFF) The two WWTPstreat sewage from approximately 920 000 inhabitants(for details see Figure 1) In the same period of time and
with the same frequency marine outfalls of those WWTPs(W-MOut and D-MOut respectively) as well as VistulaRiver mouth were also sampled for isolation of entero-cocci Marine samples were taken at the depth of approxi-mately one meter below the surface once a month inApril June August and October of 2011 Location ofsampling points and their characteristics are provided inFigure 1 Enterococci were detected using the membranefiltration method To this end appropriate dilutions ofanalyzed samples were filtered in duplicate through045 μm cellulose-acetate filters (EMD Millipore Cor-poration Billerica MA USA) which were then placedon Enterococcus selective agar (Merck DarmstadtGermany) and incubated at 37degC for 48 h (ISO 7899ndash22000) Dark red or maroon colonies were consideredpresumptive enterococci For further investigationsrepresentative isolates were taken from membranes pre-senting from 20 to 50 typical colonies and stored in nu-trient broth supplemented with 15 glycerol at minus80degC
Species identification DNA isolation susceptibility testingand statistical analysisThe species identification and drug susceptibility of pre-sumptive enterococci were determined by the PhoenixtradeAutomated Microbiology System (BD New Jersey USA)according to the manufacturerrsquos instructions Since it isnot possible to distinguish E casseliflavus and E galli-narum using this system the classification E casselifla-vusgallinarum was used The susceptibility tests basedon microdilution were carried out for 11 antimicrobialagents gentamicin (GM) streptomycin (ST) ampicillin(AM) daptomycin (DAP) vancomycin (VA) teicoplanin(TEC) linezolid (LZD) fluoroquinolones (ciprofloxacin -CIP levofloxacin - LVX) tetracycline (TE) erythromycin(E) and quinupristin-dalfopristin (SYN) (E faecium only)Obtained minimal inhibitory concentrations (MIC) wereevaluated according to the guidelines of Clinical and La-boratory Standards Institute [47] Isolates were defined asmultidrug-resistant (MDR) when they showed resistanceto three or more compounds tested [48] Bacterial DNAwas purified using the Genomic DNA Prep Plus kitfollowing the manufacturerrsquos instructions (AampA Biotech-nology Gdynia Poland) Differences in distributions wereevaluated using the χ2 test with the p values lt 005 con-sidered significant
MLST and MLVA of E faeciumMLST for E faecium was performed by sequencing ofseven house-keeping genes as described by Homan et al[49] The allele numbers and STs were assigned with theuse of Internet database httpefaeciummlstnet (5th July2013 date last accessed) Novel alleles and allelic profileswere submitted to the database MLST data were analyzedwith Sequence Type Analysis and Recombinational Tests
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 13 of 15httpwwwbiomedcentralcom1471-21801466
(START) software [50] and the eBURST analysis [51] wasused to elucidate the relationships of STs with knownCCs using the comparative eBURST option available athttpeburstmlstnetv3enter_datacomparative (5th July2013 date last accessed) Further inclusion of E faeciumSTs into specific CCs and lineages was performed fol-lowing other studies [924] MLVA was performed as de-scribed by others [52]
Detection and analysis of genes associated with virulenceantimicrobial resistance and MGE in E faeciumThe full list of primers used for gene detection and se-quencing is provided in Additional file 1 Table S1 PCR-based detection of IS16 virulence-associated genes (fms21fms17 fms5 and fms19 representing each of four pilingene clusters in E faecium and the espEfm gene) genes de-termining resistance to tetracycline tet(M) tet(O) tet(L)tet(S) HLGR (aac(6prime)-Ie-aph(2Prime) HLSR (ant(6prime)-Ia) andgenes specific for MGE such as intA of ICEEfm1 int ofTn916 transposon tndX of Tn5397 transposon entero-coccal plasmid replication genes rep1pIP501 rep2pRE25rep17pRUM rep18pEF418 reppMG1 reppLG1 and TAS genesaxe-txe ω-ε-ζ relBE was performed as described in otherstudies [134153-59] following the conditions describedherein Previously characterized clinical isolates of Strepto-coccus agalactiae E faecium and E faecalis harbouringpilin genes IS16 espEfm intA of ICEEfm1 int of Tn916tndX of Tn5397 tet(M) tet(O) tet(L) tet(S) aac(6prime)-Ie-aph(2Prime) ant(6prime)-Ia rep1pIP501 rep2pRE25 ω-ε-ζ [136061]served as positive controls In cases when positive controlswere not available (rep18pEF418 reppMG1 reppLG1 axe-txerelBE) a few randomly selected PCR products wereverified by sequencing and served further as controlsAmpicillin-resistance determinant the pbp5 gene was ana-lysed by sequencing of the part encoding C-terminaltranspeptidase domain in PBP5 [62] New sequence vari-ants pbp5-24 to pbp5-29 alleles were submitted to theGenBank (acc numbers KC594860-KC594865 respect-ively) The quinolone-determining regions (QRDR) in thegyrA and parC genes were sequenced and analysed asdescribed by others [63]
Additional file
Additional file 1 Table S1 Primers used in the study for PCR andsequencing
Competing interestThe authors declare that they have no competing interest
Authorsrsquo contributionsES participated in the study design performed experimental work andparticipated in the manuscript preparation AL designed the studyperformed experimental work and participated in the manuscriptpreparation Both authors read and approved the final manuscript
AcknowledgementsWe thank Rob Willems and Janetta Top for assigning new alleles new STsand MTs This publication made use of the Enterococcus faecium MLSTwebsite (httpefaeciummlstnet) which is hosted at Imperial College of theUniversity of Oxford and funded by the Wellcome Trust This work wassupported by project N N305 461139 from the Narodowe Centrum Nauki(NCN)
Author details1National Medicines Institute ul Chelmska 3034 Warsaw 00-725 Poland2Gdansk University of Technology ul G Narutowicza 1112 Gdansk 80-952Poland
Received 5 September 2013 Accepted 27 February 2014Published 14 March 2014
References1 Aarestrup FM Butaye P Witte W Nonhuman reservoirs of Enterococci In
American Society for Microbiology Edited by Gilmore MS Clewell DB CourvalinP Dunny GM Murray BE Rice LB Washington DC The Enterococcipathogenesis molecular biology and antibiotic resistance 2002
2 Tendolkar PM Baghdayan AS Shankar N Pathogenic enterococci newdevelopments in the 21st century Cell Mol Life Sci 2003 602622ndash2636
3 Sood S Malhotra M Das BK Kapil A Enterococcal infections ampantimicrobial resistance Indian J Med Res 2008 128111ndash121
4 Sydnor ER Perl TM Hospital epidemiology and infection control inacute-care settings Clin Microbiol Rev 2011 24141ndash173
5 European Centre for Disease Prevention and Control Annual EpidemiologicalReport Reporting on 2010 surveillance data and 2011 epidemic intelligencedata Stockholm ECDC 20122013
6 Hidron AI Edwards JR Patel J Horan TC Sievert DM Pollock DA Fridkin SKNational Healthcare Safety Network Team Participating National HealthcareSafety Network Facilities NHSN annual update antimicrobial-resistantpathogens associated with healthcare-associated infections annualsummary of data reported to the National Healthcare Safety Network atthe Centers for Disease Control and Prevention 2006ndash2007 Infect ControlHosp Epidemiol 2008 29996ndash1011
7 De Kraker ME Jarlier V Monen JC Heuer OE van de Sande N GrundmannH The changing epidemiology of bacteraemias in Europe trends fromthe European Antimicrobial Resistance Surveillance System Clin MicrobiolInfect 2012 doi1011111469-0691
8 Leavis HL Bonten MJ Willems RJ Identification of high-risk enterococcalclonal complexes global dispersion and antibiotic resistance Curr OpinMicrobiol 2006 9454ndash460
9 Willems RJL Top J van Schaik W Leavis H Bonten M Sireacuten J Hanage WPCorander J Restricted gene flow among hospital subpopulations ofEnterococcus faecium mBio 2012 3e00151ndash12
10 Hegstad K Mikalsen T Coque TM Werner G Sundsfjord A Mobile geneticelements and their contribution to the emergence of antimicrobialresistant Enterococcus faecalis and Enterococcus faecium Clin MicrobiolInfect 2010 16541ndash554
11 Shepard BD Gilmore MS Antibiotic-resistant enterococci themechanisms and dynamics of drug introduction and resistance MicrobesInfect 2002 4215ndash224
12 Bonten MJ Willems R Weinstein RA Vancomycin-resistant enterococciwhy are they here and where do they come from Lancet Infect Dis 20011314ndash325
13 Sadowy E Sieńko A Gawryszewska I Bojarska A Malinowska K HryniewiczW High abundance and diversity of antimicrobial resistancedeterminants among early vancomycin-resistant Enterococcus faecium inPoland Eur J Clin Microbiol Infect Dis 2013 321193ndash1203
14 Willems RJ Homan W Top J van Santen-Verheuvel M Tribe D Manzioros XGaillard C Vandenbroucke-Grauls CM Mascini EM van Kregten E vanEmbden JD Bonten MJ Variant esp gene as a marker of a distinct geneticlineage of vancomycin-resistant Enterococcus faecium spreading inhospitals Lancet 2001 357853ndash855
15 Sillanpaumlauml J Nallapareddy SR Prakash VP Qin X Houmloumlk M Weinstock GMMurray BE Identification and phenotypic characterization of a secondcollagen adhesin Scm and genome-based identification and analysis of13 other predicted MSCRAMMs including four distinct pilus loci inEnterococcus faecium Microbiology 2008 1543199ndash3211
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 14 of 15httpwwwbiomedcentralcom1471-21801466
16 Galloway-Pentildea JR Nallapareddy SR Arias CA Eliopoulos GM Murray BEAnalysis of clonality and antibiotic resistance among early clinicalisolates of Enterococcus faecium in the United States J Infect Dis 20092001566ndash1573
17 Top J Willems R Bonten M Emergence of CC17 Enterococcus faeciumfrom commensal to hospital-adapted pathogen FEMS Immunol MedMicrobiol 2008 52297ndash308
18 Martins da Costa P Vaz-Pires P Bernardo F Antimicrobial resistance inEnterococcus spp isolated in inflow effluent and sludge from municipalsewage water treatment plants Water Res 2006 401735ndash1740
19 Reinthaler FF Posch J Feierl G Wust G Haas D Ruckenbauer G Mascher FMarth E Antibiotic resistance of Escherichia coli in sewage and sludgeWater Res 2003 371685ndash1690
20 Luczkiewicz A Fudala-Ksiazek S Jankowska K Olanczuk-Neyman KAntimicrobial resistance of fecal indicators in municipal wastewatertreatment plant Water Res 2010 445089ndash5097
21 Dulfour AP Ballentine RK Ambient water quality criteria for bacteria-1986 EPA4405-84-002 Washington DC US Environmental Protection Agency 1986
22 Ferreira da Silva M Tiago I Veriacutessimo A Boaventura RA Nunes OC Manaia CMAntimicrobial resistance patterns in Enterobacteriaceae isolated from anurban wastewater treatment plant FEMS Microbiol Ecol 2007 60166ndash176
23 DIRECTIVE 20067EC of the European Parliament and of the Council 15February 2006 concerning the management of bathing water quality andrepealing Directive 76160EEC httpeur-lexeuropaeuLexUriServLexUriServdouri=OJL200606400370051ENPDF
24 Freitas AR Novais C Ruiz-Garbajosa P Coque TM Peixe L Dispersion ofmultidrug-resistant Enterococcus faecium isolates belonging to majorclonal complexes in different Portuguese settings Appl Environ Microbiol2009 754904ndash4908
25 Poeta P Costa D Igrejas G Saacuteenz Y Zarazaga M Rodrigues J Torres CPolymorphisms of the pbp5 gene and correlation with ampicillinresistance in Enterococcus faecium isolates of animal origin J MedMicrobiol 2007 56236ndash240
26 Kuumlhn I Iversen A Burman LG Olsson-Liljequist B Franklin A Finn MAarestrup F Seyfarth AM Blanch AR Vilanova X Taylor H Caplin J MorenoMA Dominguez L Herrero IA Moumlllby R Comparison of enterococcalpopulations in animals humans and the environment ndash a Europeanstudy Int J Food Microbiol 2003 88133ndash145
27 Lanthier M Scott A Lapen DR Zhang Y Topp E Frequency of virulencegenes and antibiotic resistances in Enterococcus spp isolates fromwastewater and feces of domesticated mammals and birds and wildlifeCan J Microbiol 2010 56715ndash729
28 Leclercq R Oberleacute K Galopin S Cattoir V Budzinski H Petit F Changes inenterococcal populations and related antibiotic resistance along amedical center-wastewater treatment plant-river continuum Appl EnvironMicrobiol 2013 792428ndash2434
29 Thevenon F Regier N Benagli C Tonolla M Adatte T Wildi W Poteacute JCharacterization of fecal indicator bacteria in sediments cores from thelargest freshwater lake of Western Europe (Lake Geneva Switzerland)Ecotoxicol Environ Safety 2012 7850ndash56
30 Bonilla TD Nowosielski K Esiobu N McCorquodale DS Rogerson ASpecies assemblages of Enterococcus indicate potential sources offecal bacteria at a south Florida recreational beach Mar Pollut Bull2006 52807ndash810
31 Wang JT Chang SC Wang HY Chen PC Shiau YR Lauderdale TL TSARHospitals High rates of multidrug resistance in Enterococcus faecalis andE faecium isolated from inpatients and outpatients in TaiwanDiagn Microbiol Infect Dis 2013 75406ndash411
32 Lanthier M Scott A Zhang Y Cloutier M Durie D Henderson VC WilkesG Lapen DR Topp E Distribution of selected virulence genes andantibiotic resistance in Enterococcus species isolated from the SouthNation River drainage basin Ontario Canada J Appl Microbiol 2011110407ndash421
33 Kuch A Willems RJ Werner G Coque TM Hammerum AM Sundsfjord AKlare I Ruiz-Garbajosa P Simonsen GS van Luit-Asbroek M Hryniewicz WSadowy E Insight into antimicrobial susceptibility and population struc-ture of contemporary human Enterococcus faecalis isolates fromEurope J Antimicrob Chemother 2012 67551ndash558
34 Novais C Coque TM Ferreira H Sousa JC Peixe L Environmentalcontamination with vancomycin-resistant enterococci from hospitalsewage in Portugal Appl Environ Microbiol 2005 713364ndash3368
35 Iversen A Kuumlhn I Rahman M Franklin A Burman LG Olsson-Liljequist BTorell E Moumlllby R Evidence for transmission between humans and theenvironment of a nosocomial strain of Enterococcus faeciumEnviron Microbiol 2004 655ndash59
36 Rathnayake I Hargreaves M Huygens F SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterwayAustralia and associated antibiotic resistance gene profilesBMC Microbiol 2011 11201
37 Rosvoll TC Pedersen T Sletvold H Johnsen PJ Sollid JE Simonsen GSJensen LB Nielsen KM Sundsfjord A PCR-based plasmid typing inEnterococcus faecium strains reveals widely distributed pRE25-pRUM- pIP501- and pHT-related replicons associated with glycopeptideresistance and stabilizing toxin-antitoxin systems FEMS Immunol MedMicrobiol 2010 58254ndash268
38 Rosvoll TC Lindstad BL Lunde TM Hegstad K Aasnaes B Hammerum AMLester CH Simonsen GS Sundsfjord A Pedersen T Increased high-levelgentamicin resistance in invasive Enterococcus faecium is associated withaac(6prime)Ie-aph(2Prime)Ia-encoding transferable megaplasmids hosted by majorhospital-adapted lineages FEMS Immunol Med Microbiol 2012 66166ndash176
39 Laverde Gomez JA van Schaik W Freitas AR Coque TM Weaver KE FranciaMV Witte W Werner G A multiresistance megaplasmid pLG1 bearing ahylEfm genomic island in hospital Enterococcus faecium isolates Int J MedMicrobiol 2011 301165ndash175
40 Coque TM Evolutionary biology of pathogenic enterococci InEvolutionary Biology of Bacterial and Fungal Pathogens Coque TM Edited byBaquero F Nombela C Cassell GH Gutierrez JA Washington DC ASMPress501ndash523
41 Werner G Fleige C Geringer U van Schaik W Klare I Witte W IS elementIS16 as a molecular screening tool to identify hospital-associated strainsof Enterococcus faecium BMC Infect Dis 2011 1180
42 Top J Sinnige JC Majoor EA Bonten MJ Willems RJ van Schaik W Therecombinase IntA is required for excision of esp-containing ICEEfm1 inEnterococcus faecium J Bacteriol 2011 1931003ndash1006
43 Sillanpaumlauml J Prakash VP Nallapareddy SR Murray BE Distribution of genesencoding MSCRAMMs and Pili in clinical and natural populations ofEnterococcus faecium J Clin Microbiol 2009 47896ndash901
44 Marcinek H Wirth R Muscholl-Silberhorn A Gauer M Enterococcus faecalisgene transfer under natural conditions in municipal sewage watertreatment plants Appl Environ Microbiol 1998 64626ndash632
45 Kuumlmmerer K Significance of antibiotics in the environment J AntimicrobChemother 2003 525ndash7
46 Liu A Fong A Becket E Yuan J Tamae C Medrano L Maiz M Wahba C LeeC Lee K Tran KP Yang H Hoffman RM Salih A Miller JHP Selectiveadvantage of resistant strains at trace levels of antibiotics a simple andultrasensitive color test for detection of antibiotics and genotoxicagents Antimicrob Agents Chemother 2011 551204ndash1210
47 Clinical and Laboratory Standards Institute Performance standards forantimicrobial susceptibility testing 21st informational supplement Wayne CLSI2011 PA M100-S20
48 Magiorakos AP Srinivasan A Carey RB Carmeli Y Falagas ME Giske CGHarbarth S Hindler JF Kahlmeter G Olsson-Liljequist B Paterson DL Rice LBStelling J Struelens MJ Vatopoulos A Weber JT Monnet DLMultidrug-resistant extensively drug-resistant and pandrug-resistantbacteria an international expert proposal for interim standarddefinitions for acquired resistance Clin Microbiol Infect 2012 18268ndash281
49 Homan WL Tribe D Poznanski S Li M Hogg G Spalburg E Van Embden JDWillems RJ Multilocus sequence typing scheme for Enterococcus faeciumJ Clin Microbiol 2002 401963ndash1971
50 Jolley KA Feil EJ Chan MS Maiden MC Sequence type analysis andrecombinational tests (START) Bioinformatics 2001 171230ndash1231
51 Feil EJ Li BC Aanensen DM Hanage WP Spratt BG eBURST inferringpatterns of evolutionary descent among clusters of related bacterialgenotypes from multilocus sequence typing data J Bacteriol 20041861518ndash1530
52 Top J Banga1 NMI Hayes R Willems RJ Bonten MJM Hayden MKComparison of multiple-locus variable-number tandem repeat analysisand pulsed-field gel electrophoresis in a setting of polyclonal endemicityof vancomycin-resistant Enterococcus faecium Clin Microbiol Infect 200814363ndash369
53 Vankerckhoven V Van Autgaerden T Vael C Lammens C Chapelle S RossiR Jabes D Goossens H Development of a multiplex PCR for the
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-
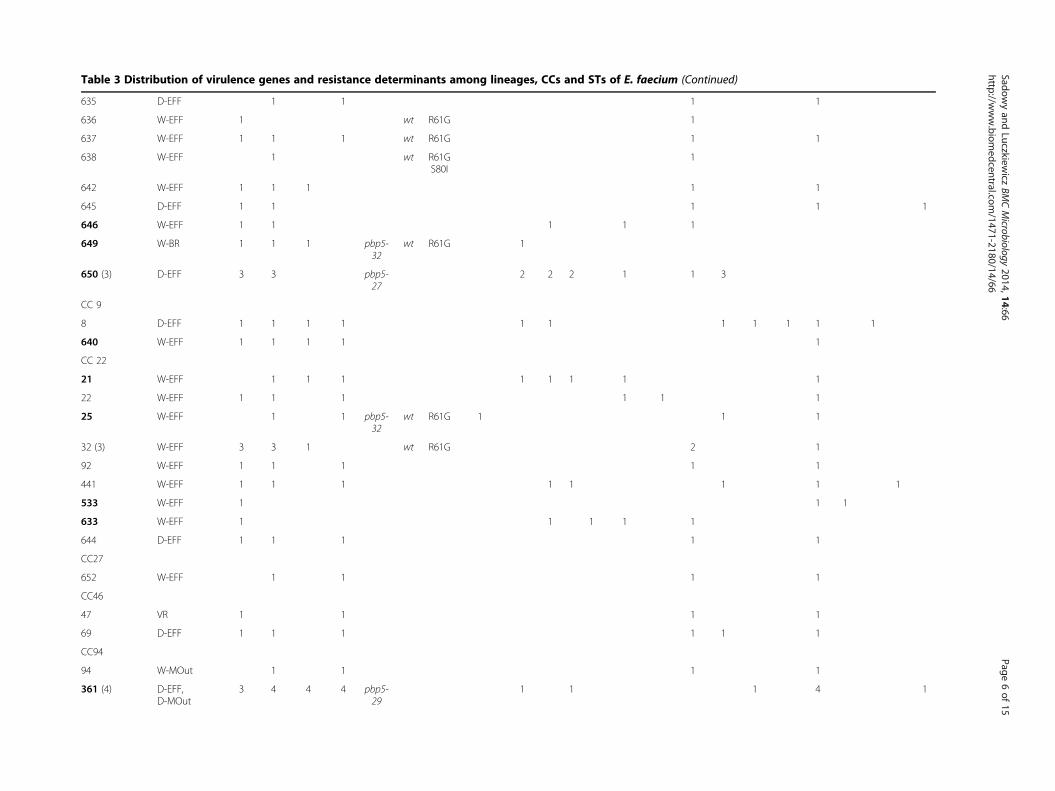
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium (Continued)
635 D-EFF 1 1 1 1
636 W-EFF 1 wt R61G 1
637 W-EFF 1 1 1 wt R61G 1 1
638 W-EFF 1 wt R61GS80I
1
642 W-EFF 1 1 1 1 1
645 D-EFF 1 1 1 1 1
646 W-EFF 1 1 1 1 1
649 W-BR 1 1 1 pbp5-32
wt R61G 1
650 (3) D-EFF 3 3 pbp5-27
2 2 2 1 1 3
CC 9
8 D-EFF 1 1 1 1 1 1 1 1 1 1 1
640 W-EFF 1 1 1 1 1
CC 22
21 W-EFF 1 1 1 1 1 1 1 1
22 W-EFF 1 1 1 1 1 1
25 W-EFF 1 1 pbp5-32
wt R61G 1 1 1
32 (3) W-EFF 3 3 1 wt R61G 2 1
92 W-EFF 1 1 1 1 1
441 W-EFF 1 1 1 1 1 1 1 1
533 W-EFF 1 1 1
633 W-EFF 1 1 1 1 1
644 D-EFF 1 1 1 1 1
CC27
652 W-EFF 1 1 1 1
CC46
47 VR 1 1 1 1
69 D-EFF 1 1 1 1 1 1
CC94
94 W-MOut 1 1 1 1
361 (4) D-EFFD-MOut
3 4 4 4 pbp5-29
1 1 1 4 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
6of
15httpw
wwbiom
edcentralcom1471-21801466
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium (Continued)
CC296
296 W-EFF 1 1
CC639
639 W-EFF 1 1 1 1 1
CC648
634 W-EFF 1 1 1 1 1
648 W-EFF 1 1 1 1 1
Doubletons
632 W-MOut 1 1 1
Singletons
264 W-INF 1 1 1 1 1 1 1
641 D-EFF 1 1 1 1
643 W-EFF 1 1 1 1 1 1
647 W-MOut 1 1 1 1 1
651 D-EFF 1 1 1 1 1 1
654 VR 1 1 1 1 1 1
All (69) 8 57 49 29 46 6 17 20 17 1 9 4 39 27 14 7 55 2 12 10 5
CC clonal complex ST sequence type MT MLVA-type nd not determined due to lack of amplification of the pbp5 gene wt wild-type STs associated with MDR phenotype in boldAbbreviations of sampling pointsrsquo locations and their description provided in Figure 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
7of
15httpw
wwbiom
edcentralcom1471-21801466
Figure 2 MLST-based similarity tree of E faecium isolates constructed using the START software and the UPGMA clustering algorithm
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 8 of 15httpwwwbiomedcentralcom1471-21801466
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 9 of 15httpwwwbiomedcentralcom1471-21801466
grouped the 38 STs specific for 50 isolates into the majorlsquosuper-complex 17rsquo characteristic for this species (data notshown) Further detailed analysis of the localization ofparticular STs within this structure revealed that 11 STscharacteristic for 17 isolates belonged to the nosocomialcomplex of E faecium representing all three majorlineages 17 18 and 78 of a former CC17 as defined byWillems et al [9] (Table 3) Seventeen STs (20 isolates)nine STs (11 isolates) and two STs with single isolates eachbelonged to CC5 CC22 and CC9 respectively as definedby Freitas et al [24] Of the remaining 16 STs nine wereincluded into six small CCs (CCs 27 46 94 296 639648) unlinked to the major lsquosuper-complexrsquo ST632formed a lsquodoubletonrsquo (ie was linked by a single-locus vari-ant link to only one ST from the database) and six STsrepresented by single isolates were singletons ie werenot related to any ST in the database at the time of ana-lysis Thirty isolates (435) in the investigated groupshowed the MDR phenotype which was significantly asso-ciated (p = 000006) with isolates from the nosocomialcomplex (Table 3 and Figure 3) but found also in CC5CC22 and CC94 this phenotype did not occur amongsmall CCs or singletons Seventeen isolates repre-senting the nosocomial complex were additionallystudied by multilocus VNTR (variable number tan-dem repeats) analysis (MLVA) to better elucidate therelationships among these isolates Altogether nineMLVA-types (MTs) were found (Table 3) Importantlythe same MLVA-type MT159 associated with ST78was characteristic for isolates derived from samplesfrom W-EFF and W-MOut suggesting survival of thisparticular clone
0
10
20
30
40
50
60
70
80
90
100
IS16
espintA
fms5
fms1
7
fms1
9
fms2
1
MDR
AM-R
CIP-R
HLG
R
p
ositi
ve
isol
ates
Figure 3 Distribution () of IS16 resistance phenotypes and virulenccolumns isolates with STs typical for nosocomial E faecium grey columnsgroups are indicated by an asterisk MDR multi-drug resistance AM-R ampgentamicin resistance HLSR high-level streptomycin resistance
IS16 was present in all 17 isolates included by MLST tothree lineages of nosocomial E faecium and this markerwas limited to this group (Table 3 and Figure 3) TheespEfm gene occurred always together with intA among allisolates of lineage 17 and in some isolates of lineage 78The fms19 pilin gene detected in 29 isolates was over-represented among STs typical for nosocomial E faeciumwhile other pilin genes such as fms5 fms17 and fms21detected in 57 49 and 46 isolates respectively occurredamong various CCs and singleton STs Ampicillin resist-ance significantly associated with isolates of nosocomialcomplex (10 of 17 isolates 588 p = 0001) occurred alsoin CC5 (6 isolates) CC22 and CC94 (1 isolate each) Se-quencing of the 3prime part of the pbp5 gene revealed its 11nucleotide alleles corresponding to 11 variants of the C-terminal part of PBP5 Six alleles of the gene were new(pbp5-24 to pbp5-29) two alleles (pbp5-20 pbp5-22)were described previously among Polish hospital VREmisolates [13] and three alleles were found in other stud-ies pbp5-30 occurred in a Portuguese animal strain ofE faecium [25] and pbp5-31 and pbp5-32 in clinical iso-lates TCGE70411_2 and 164306 also from Portugal (de-posited as JN208888 and JN208885 in the GenBankrespectively) For a single ampicillin-resistant isolate ofST653 (lineage 78) pbp5 amplification was not achieveddespite repeated attempts The majority (11 of 17 iso-lates 647) of isolates belonging to three lineages ofnosocomial E faecium showed resistance to ciprofloxa-cin determined by mutations R61G and S80IR in ParCand S84IRY in GyrA Ciprofloxacin resistance wasalso significantly over-represented among isolates ofSTs typical for nosocomial E faecium (p = 00006) Nine
HLS
R
tetM tetL
rep1
-pIP50
1
rep2
-pRE25
rep1
7-pR
UM
rep1
8-pE
F418
rep-pL
G1
axe-txe
relBE
e resistance and plasmid genes among E faecium isolates Blackother E faecium isolates Significant differences (p lt 005) between twoicillin resistance CIP-R ciprofloxacin resistance HLGR high-level
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 10 of 15httpwwwbiomedcentralcom1471-21801466
isolates resistant to ciprofloxacin were found also inCCs 5 9 and 22 These isolates possessed mutations inParC mentioned above but had the wild-type QRDR inGyrA HLGR occurred in five isolates of lineages 18 and78 and in single isolates of CC5 and CC22 this pheno-type was determined by the presence of aac(6prime)-Ie-aph(2Prime) with an exception of an isolate from CC5 whichwas negative for this gene High-level resistance tostreptomycin (HLSR) was widely distributed amongstudied isolates and determined by the ant(6prime)-Ia geneTetracycline resistance was found also in various STsthe major determinant of this phenotype was tet(M) (20isolates) followed by tet(L) (17 isolates) These twogenes occurred together in 16 isolates A single isolateof ST633 belonging to CC22 harbored tet(S) The tet(O) gene was absent in the studied group For a singletetracycline-resistant isolate of ST17 none of the fourtetracycline determinants studied was detected Nineisolates carried int of Tn916 and four the tndX gene ofTn5397 These transposon-specific genes were not char-acteristic for any lineage or complex (Table 3) Plasmid-replication genes such as rep2pRE25 rep17pRUM andrep18 pEF418 (26 14 and 7 isolates respectively) andgenes specifying toxin-antitoxin systems (TAS) axe-txeand ω-ε-ζ (12 and 10 isolates respectively) were signifi-cantly associated with the group of isolates of nosocomialE faecium Two other rep genes (rep1pIP501 reppLG1) werewidely distributed among all studied E faecium isolatesand the reppMG1 gene occurred in two isolates (in a singleisolate of ST533 from CC22 and in a singleton ST647) Allω-ε-ζ-positive isolates carried rep2pRE25 and nine isolatesshowed a concomitant presence of axe-txe and rep17pRUMThe relBE genes were found solely among five non-nosocomial E faecium (a difference in distribution with-out statistical significance p = 043) among various STs
DiscussionIn this study Enterococcus spp was used to evaluate theimpact posed by treated wastewater of two local WWTPson their receiver the costal waters of Gulf of GdanskTotal counts of enterococci observed for raw (W-INF)and treated (W-EFF D-EFF) wastewater are consistentwith our previous study showing similar over 99 reduc-tion capacity of applied treatment processes It is alsoreported in the literature that wastewater treatment pro-cesses based on activated sludge although they focus onremoval of nutrients and organic matter simultaneouslyreduce faecal indicators with high efficiency reaching upto 999 [1822] Due to the high number of enterococciin raw wastewater they are nevertheless released from aWWTP in a number up to 61x105 CFU per 100 mLThus marine water sampled in the area directly impactedby treated wastewater (W-MOut and D-MOut) showed ahigher number of enterococci than the one observed in
Vistula River mouth however lower by four orders ofmagnitude in comparison to corresponding treated waste-water (W-EFF and D-EFF) It should be noted that in mar-ine outfalls (W-MOut D-MOut) as well as in VistulaRiver mouth enterococci were detected in number lowerthan 100 CFU per 100 mL ie the value set out by NewBathing Water Directive 20067EC for the costal water ofldquoexcellent qualityrdquo [23]It is suggested that enterococcal species composition in
faecal-impacted environment depends from the source ofcontamination In this study two species most commonlycolonizing humans E faecalis and E faecium comprisedtogether from 73 to 88 of enterococcal isolates intested wastewater samples (W-INF W-BR W-EFF and D-EFF) 73 and 61 in marine outflows (W-MOut and D-MOut respectively) and 45 in the Vistula River mouthThe predominant species in wastewater and marine waterimpacted by treated wastewater was E faecium (from 56to 65) which stayed at a similar level during treatmentprocesses as determined also in our previous study [20]Species distribution in wastewater samples is not wellunderstood and affecting factors may include differencesin diet climate season and methods of detection E feaca-lis was found to be the most prevalent species in wastewa-ter samples in Sweden [26] E faecium in Spain UKCanada France and Switzerland [26-29] while E hiraepredominated in Portugal and United States [2230] Al-though E faecalis is typically the most abundant colonizerof humans in the community (eg [2631]) E faecium isgenerally most often associated with human microbialcontamination [2729] E faecalis was reported fromagriculture-impacted waters [32] and E hirae likelyoriginates mainly from cattle and pig faecal contamin-ation [26] which explains high relative proportion ofthese two species in Vistula River mouth Apart from thevariability of contamination sources proportion of par-ticular species may differ due to the differences of abilityto sustain environmental stresses In this study enterococ-cal composition might have been influenced by wastewa-ter treatment processes and marine water characteristicsThe effect of wastewater treatment and marine environ-
ment was seen on phenotypes of antimicrobial resistanceThe positive selection of bacteria with resistance patternshas been already suggested and noted in wastewater pro-cesses [192022] In this study the resistance rates to ampi-cillin ciprofloxacin levofloxacin and erythromycin notedfor E faecium from raw wastewater (W-INF) were lowerthan these observed for this species in the correspondingtreated wastewater (W-EFF) however not statistically sig-nificant (p gt 005) Moreover resistance rates to ampicillinciprofloxacin tetracycline and erythromycin observed inmarine outflow of WWTP Gdansk-Wschod (W-MOut)were higher than in treated wastewater of this plantSuch phenomenon was not observed for treated
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 11 of 15httpwwwbiomedcentralcom1471-21801466
wastewater of WWTP Gdynia-Debogorze (D-EFF) andits marine outfall (D-MOut) Resistance rates among Efaecium originating from D-MOut were in generalcomparable to these detected in the Vistula Rivermouth It should be noted that treated wastewater fromWWTP Gdansk-Wschod has been discharged via mar-ine outfall (W-MOut) for last 12 years while marineoutfall of WWTP Gdynia-Debogorze (D-MOut) hasbeen operating for last two years The observed differ-ences in resistance rate corresponding to the local im-pact caused by long and short-term operated marineoutfalls (W-MOut and D-MOut) need further attentionSignificant rates of antimicrobial resistance observed
for isolates of E faecalis from wastewater and marineriver waters impacted by wastewater are in agreementwith a recent study performed in six European countriesshowing that in Poland the rates of resistance for humanisolates of this species were usually among the highestboth in the community and hospitals [33] To our know-ledge no corresponding data are available for E faeciumcolonization in the community in Poland but a studyfocused on hospital VRE isolates showed their ubiquitousMDR phenotype and a high load of resistance genes [13]In our study no vancomycin resistance were detectedlikely due to relatively low (below 10) prevalence of VREin Polish hospitals [5] although such isolates are isolatedfrom the environment in other countries eg [34]This study shows that hospital clones of E faecium are
important carriers of resistance genes in wastewatersMost likely in a significant part they originated fromhospital wastewater treated together with communalwastewater In our estimation hospital wastewater con-stitutes approximately 02 of both WWTPsrsquo daily in-flow The release of hospital-associated clones of Efaecium to the environment was observed also by others[24283536] In our study isolates belonging to themajor nosocomial HiRECC (formerly named CC17)constituted altogether 246 of all isolates of E faeciumsubjected to typing and were present at all samplingsites These isolates apart from their inclusion into theHiRECC by MLST demonstrated several other pheno-typic and genotypic features typical for nosocomial Efaecium ie they were resistant to ciprofloxacin andampicillin in vast majority showed MDR phenotypeand carried several resistance determinants associatedboth with mutations of chromosomal genes (gyrA parCpbp5) and in the case of aminoglycoside and tetracyclineresistance with gene acquisition Plasmids likely played animportant role in the later process as some plasmid genessuch as rep2pRE25 rep17pRUM and rep18 pEF418 and TASgenes axe-txe and ω-ε-ζ were significantly more abundantin isolates associated with HiRECC than among theremaining isolates As yet the knowledge of distributionof plasmid-associated genes in populations of E faecium
especially in the context of their clonal compositionremains limited A study on 93 isolates of differentgeographical origins and from various sites showed anover-representation of rep17pRUM and reppMG1 amongnosocomial E faecium while in contrast to our observa-tions rep18 pEF418 was not detected [37] Moreover wedid not observe reppMG1 among HiRECC of E faeciumThese differences may be due to a high prevalence of VRE(58 isolates) among 93 isolates studied by Rosvoll et alwhat may explain differences in the plasmid content Thesame study also reported the co-localization of ω-ε-ζ withrep2pRE25 and axe-txe with rep17pRUM on the same plas-mids in concordance with our findings showing a fre-quent joint occurrence of these pairs of genes Anotherstudy performed on 99 invasive isolates from Norwaymostly of HLGR phenotype showed a significantly higherprevalence of rep2pRE25 and rep17pRUM among the majornosocomial HiRECC [38] This study as well some others[2439] also demonstrated a common presence of reppLG1among E faecium from hospital settings and in animalsOur findings indicate that this type of replicon is evenmore ubiquitous in E faecium than previously reportedAll rep genes mentioned above are typically associatedwith plasmids carrying various antimicrobial resistancegenes [3940] GenBank accession number AF408195 Inaddition to genes determining antimicrobial resistanceand genes associated with MGE isolates of nosocomialHiRECC analyzed in our study carried such molecularmarkers as IS16 and the esp gene together with the intAgene characteristic for ICEEfm [4142] Another patho-genicity factor the fms19 gene of the pilin gene cluster4 (PGC4) showed increased prevalence among theseisolates Similar observation was made for a representa-tive collection of 433 E faecium isolates from varioussources [43]In a study that followed enterococci in a continuum
from hospital and retirement home wastewaters througha WWTP to a river a gradual decrease in the proportionof E faecium isolates representing hospital-associatedSTs was observed likely due to the ldquodilutionrdquo of HiRECCrepresentatives by more diverse and less drug-resistantstrains originating from the community [28] Because inour study only selected representatives of obtained iso-lates were investigated it is not possible to determineprecise tendencies in their frequency from the WWTPinfluent to effluent and water however resistant entero-cocci both HiRECC and non-HiRECC are clearly ableto survive the treatment process what results in their re-lease into marine and river waters Moreover using anadditional typing method MLVA we were able to findisolates with the same characteristics in the effluent ofWWTP and its marine outfall suggesting ability of suchclones to survive in the environment for at least sometime It seems plausible that hospital resistant clones
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 12 of 15httpwwwbiomedcentralcom1471-21801466
accumulate several determinants that may promote theirsurvival not only in the hospital settings but also in theenvironment Such features would include resistance todisinfectants and ultraviolet light additional metabolicpathways and ability to form biofilms Moreover determi-nants of some of the mentioned adaptation factors arefound co-resident in plasmids harboring resistance genessuggesting co-selection For example pLG1-type plasmidsof E faecium carry not only resistance determinantsagainst aminoglycosides glycopeptides and macrolidesbut also pili genes and carbon uptake-utilization genes[3839] Pheromone-responsive plasmids of E faecalisapart from resistance to aminoglycosides glycopeptidespenicillins and macrolides may encode bacteriocins add-itional pili and UV-resistance determinants [40] Thusnosocomial HiRECCs released by WWTPs are equippedin whole sets of genes with various adaptive benefits andoften located within MGE It was also demonstrated thatMGEs can be transferred in a WWTP [44] further under-ling the importance of threat posed by such organismsThere is a selection pressure for survival of resistantstrains and their tranconjugants due to release of antibi-otics to the environment especially in the hospital efflu-ents [45] and even sub-inhibitory concentrations may beeffective in selection for resistance [46]
ConclusionsThis study describes the species composition resistanceprofiles and clonal relationships of enterococci isolatedfrom WWTPs located in Gdansk their marine outfallsto the Gulf of Gdansk and from Vistula River It was ob-served that these WWTPs release enterococcal strainsresistant to various antimicrobial compounds to the re-cipient water environment Molecular analyses revealedthe presence of clones associated with nosocomial HiR-ECCs and in the case of E faecium isolates of lineages17 18 and 78 of former CC17 abundant in resistance de-terminants MGE genes and pathogenicity factors Thepresence of HiRECCs in wastewaters and marineriverenvironment highlights the need for further detailedanalyses to better understand the survival and spread ofdrug-resistant strains in water ecosystems and to eluci-date the ways to curb such dissemination
MethodsAcquisition of samples and isolation of enterococciOnce a month in April June August and October in 2011of the flow proportioned composited samples of wastewa-ter were taken from two local WWTPs Gdansk-Wschodfrom influent (W-INF) effluent (W-EFF) and wastewa-ter from activated sludge bioreactor (W-BR) and fromGdynia-Debogorze effluent (D-EFF) The two WWTPstreat sewage from approximately 920 000 inhabitants(for details see Figure 1) In the same period of time and
with the same frequency marine outfalls of those WWTPs(W-MOut and D-MOut respectively) as well as VistulaRiver mouth were also sampled for isolation of entero-cocci Marine samples were taken at the depth of approxi-mately one meter below the surface once a month inApril June August and October of 2011 Location ofsampling points and their characteristics are provided inFigure 1 Enterococci were detected using the membranefiltration method To this end appropriate dilutions ofanalyzed samples were filtered in duplicate through045 μm cellulose-acetate filters (EMD Millipore Cor-poration Billerica MA USA) which were then placedon Enterococcus selective agar (Merck DarmstadtGermany) and incubated at 37degC for 48 h (ISO 7899ndash22000) Dark red or maroon colonies were consideredpresumptive enterococci For further investigationsrepresentative isolates were taken from membranes pre-senting from 20 to 50 typical colonies and stored in nu-trient broth supplemented with 15 glycerol at minus80degC
Species identification DNA isolation susceptibility testingand statistical analysisThe species identification and drug susceptibility of pre-sumptive enterococci were determined by the PhoenixtradeAutomated Microbiology System (BD New Jersey USA)according to the manufacturerrsquos instructions Since it isnot possible to distinguish E casseliflavus and E galli-narum using this system the classification E casselifla-vusgallinarum was used The susceptibility tests basedon microdilution were carried out for 11 antimicrobialagents gentamicin (GM) streptomycin (ST) ampicillin(AM) daptomycin (DAP) vancomycin (VA) teicoplanin(TEC) linezolid (LZD) fluoroquinolones (ciprofloxacin -CIP levofloxacin - LVX) tetracycline (TE) erythromycin(E) and quinupristin-dalfopristin (SYN) (E faecium only)Obtained minimal inhibitory concentrations (MIC) wereevaluated according to the guidelines of Clinical and La-boratory Standards Institute [47] Isolates were defined asmultidrug-resistant (MDR) when they showed resistanceto three or more compounds tested [48] Bacterial DNAwas purified using the Genomic DNA Prep Plus kitfollowing the manufacturerrsquos instructions (AampA Biotech-nology Gdynia Poland) Differences in distributions wereevaluated using the χ2 test with the p values lt 005 con-sidered significant
MLST and MLVA of E faeciumMLST for E faecium was performed by sequencing ofseven house-keeping genes as described by Homan et al[49] The allele numbers and STs were assigned with theuse of Internet database httpefaeciummlstnet (5th July2013 date last accessed) Novel alleles and allelic profileswere submitted to the database MLST data were analyzedwith Sequence Type Analysis and Recombinational Tests
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 13 of 15httpwwwbiomedcentralcom1471-21801466
(START) software [50] and the eBURST analysis [51] wasused to elucidate the relationships of STs with knownCCs using the comparative eBURST option available athttpeburstmlstnetv3enter_datacomparative (5th July2013 date last accessed) Further inclusion of E faeciumSTs into specific CCs and lineages was performed fol-lowing other studies [924] MLVA was performed as de-scribed by others [52]
Detection and analysis of genes associated with virulenceantimicrobial resistance and MGE in E faeciumThe full list of primers used for gene detection and se-quencing is provided in Additional file 1 Table S1 PCR-based detection of IS16 virulence-associated genes (fms21fms17 fms5 and fms19 representing each of four pilingene clusters in E faecium and the espEfm gene) genes de-termining resistance to tetracycline tet(M) tet(O) tet(L)tet(S) HLGR (aac(6prime)-Ie-aph(2Prime) HLSR (ant(6prime)-Ia) andgenes specific for MGE such as intA of ICEEfm1 int ofTn916 transposon tndX of Tn5397 transposon entero-coccal plasmid replication genes rep1pIP501 rep2pRE25rep17pRUM rep18pEF418 reppMG1 reppLG1 and TAS genesaxe-txe ω-ε-ζ relBE was performed as described in otherstudies [134153-59] following the conditions describedherein Previously characterized clinical isolates of Strepto-coccus agalactiae E faecium and E faecalis harbouringpilin genes IS16 espEfm intA of ICEEfm1 int of Tn916tndX of Tn5397 tet(M) tet(O) tet(L) tet(S) aac(6prime)-Ie-aph(2Prime) ant(6prime)-Ia rep1pIP501 rep2pRE25 ω-ε-ζ [136061]served as positive controls In cases when positive controlswere not available (rep18pEF418 reppMG1 reppLG1 axe-txerelBE) a few randomly selected PCR products wereverified by sequencing and served further as controlsAmpicillin-resistance determinant the pbp5 gene was ana-lysed by sequencing of the part encoding C-terminaltranspeptidase domain in PBP5 [62] New sequence vari-ants pbp5-24 to pbp5-29 alleles were submitted to theGenBank (acc numbers KC594860-KC594865 respect-ively) The quinolone-determining regions (QRDR) in thegyrA and parC genes were sequenced and analysed asdescribed by others [63]
Additional file
Additional file 1 Table S1 Primers used in the study for PCR andsequencing
Competing interestThe authors declare that they have no competing interest
Authorsrsquo contributionsES participated in the study design performed experimental work andparticipated in the manuscript preparation AL designed the studyperformed experimental work and participated in the manuscriptpreparation Both authors read and approved the final manuscript
AcknowledgementsWe thank Rob Willems and Janetta Top for assigning new alleles new STsand MTs This publication made use of the Enterococcus faecium MLSTwebsite (httpefaeciummlstnet) which is hosted at Imperial College of theUniversity of Oxford and funded by the Wellcome Trust This work wassupported by project N N305 461139 from the Narodowe Centrum Nauki(NCN)
Author details1National Medicines Institute ul Chelmska 3034 Warsaw 00-725 Poland2Gdansk University of Technology ul G Narutowicza 1112 Gdansk 80-952Poland
Received 5 September 2013 Accepted 27 February 2014Published 14 March 2014
References1 Aarestrup FM Butaye P Witte W Nonhuman reservoirs of Enterococci In
American Society for Microbiology Edited by Gilmore MS Clewell DB CourvalinP Dunny GM Murray BE Rice LB Washington DC The Enterococcipathogenesis molecular biology and antibiotic resistance 2002
2 Tendolkar PM Baghdayan AS Shankar N Pathogenic enterococci newdevelopments in the 21st century Cell Mol Life Sci 2003 602622ndash2636
3 Sood S Malhotra M Das BK Kapil A Enterococcal infections ampantimicrobial resistance Indian J Med Res 2008 128111ndash121
4 Sydnor ER Perl TM Hospital epidemiology and infection control inacute-care settings Clin Microbiol Rev 2011 24141ndash173
5 European Centre for Disease Prevention and Control Annual EpidemiologicalReport Reporting on 2010 surveillance data and 2011 epidemic intelligencedata Stockholm ECDC 20122013
6 Hidron AI Edwards JR Patel J Horan TC Sievert DM Pollock DA Fridkin SKNational Healthcare Safety Network Team Participating National HealthcareSafety Network Facilities NHSN annual update antimicrobial-resistantpathogens associated with healthcare-associated infections annualsummary of data reported to the National Healthcare Safety Network atthe Centers for Disease Control and Prevention 2006ndash2007 Infect ControlHosp Epidemiol 2008 29996ndash1011
7 De Kraker ME Jarlier V Monen JC Heuer OE van de Sande N GrundmannH The changing epidemiology of bacteraemias in Europe trends fromthe European Antimicrobial Resistance Surveillance System Clin MicrobiolInfect 2012 doi1011111469-0691
8 Leavis HL Bonten MJ Willems RJ Identification of high-risk enterococcalclonal complexes global dispersion and antibiotic resistance Curr OpinMicrobiol 2006 9454ndash460
9 Willems RJL Top J van Schaik W Leavis H Bonten M Sireacuten J Hanage WPCorander J Restricted gene flow among hospital subpopulations ofEnterococcus faecium mBio 2012 3e00151ndash12
10 Hegstad K Mikalsen T Coque TM Werner G Sundsfjord A Mobile geneticelements and their contribution to the emergence of antimicrobialresistant Enterococcus faecalis and Enterococcus faecium Clin MicrobiolInfect 2010 16541ndash554
11 Shepard BD Gilmore MS Antibiotic-resistant enterococci themechanisms and dynamics of drug introduction and resistance MicrobesInfect 2002 4215ndash224
12 Bonten MJ Willems R Weinstein RA Vancomycin-resistant enterococciwhy are they here and where do they come from Lancet Infect Dis 20011314ndash325
13 Sadowy E Sieńko A Gawryszewska I Bojarska A Malinowska K HryniewiczW High abundance and diversity of antimicrobial resistancedeterminants among early vancomycin-resistant Enterococcus faecium inPoland Eur J Clin Microbiol Infect Dis 2013 321193ndash1203
14 Willems RJ Homan W Top J van Santen-Verheuvel M Tribe D Manzioros XGaillard C Vandenbroucke-Grauls CM Mascini EM van Kregten E vanEmbden JD Bonten MJ Variant esp gene as a marker of a distinct geneticlineage of vancomycin-resistant Enterococcus faecium spreading inhospitals Lancet 2001 357853ndash855
15 Sillanpaumlauml J Nallapareddy SR Prakash VP Qin X Houmloumlk M Weinstock GMMurray BE Identification and phenotypic characterization of a secondcollagen adhesin Scm and genome-based identification and analysis of13 other predicted MSCRAMMs including four distinct pilus loci inEnterococcus faecium Microbiology 2008 1543199ndash3211
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 14 of 15httpwwwbiomedcentralcom1471-21801466
16 Galloway-Pentildea JR Nallapareddy SR Arias CA Eliopoulos GM Murray BEAnalysis of clonality and antibiotic resistance among early clinicalisolates of Enterococcus faecium in the United States J Infect Dis 20092001566ndash1573
17 Top J Willems R Bonten M Emergence of CC17 Enterococcus faeciumfrom commensal to hospital-adapted pathogen FEMS Immunol MedMicrobiol 2008 52297ndash308
18 Martins da Costa P Vaz-Pires P Bernardo F Antimicrobial resistance inEnterococcus spp isolated in inflow effluent and sludge from municipalsewage water treatment plants Water Res 2006 401735ndash1740
19 Reinthaler FF Posch J Feierl G Wust G Haas D Ruckenbauer G Mascher FMarth E Antibiotic resistance of Escherichia coli in sewage and sludgeWater Res 2003 371685ndash1690
20 Luczkiewicz A Fudala-Ksiazek S Jankowska K Olanczuk-Neyman KAntimicrobial resistance of fecal indicators in municipal wastewatertreatment plant Water Res 2010 445089ndash5097
21 Dulfour AP Ballentine RK Ambient water quality criteria for bacteria-1986 EPA4405-84-002 Washington DC US Environmental Protection Agency 1986
22 Ferreira da Silva M Tiago I Veriacutessimo A Boaventura RA Nunes OC Manaia CMAntimicrobial resistance patterns in Enterobacteriaceae isolated from anurban wastewater treatment plant FEMS Microbiol Ecol 2007 60166ndash176
23 DIRECTIVE 20067EC of the European Parliament and of the Council 15February 2006 concerning the management of bathing water quality andrepealing Directive 76160EEC httpeur-lexeuropaeuLexUriServLexUriServdouri=OJL200606400370051ENPDF
24 Freitas AR Novais C Ruiz-Garbajosa P Coque TM Peixe L Dispersion ofmultidrug-resistant Enterococcus faecium isolates belonging to majorclonal complexes in different Portuguese settings Appl Environ Microbiol2009 754904ndash4908
25 Poeta P Costa D Igrejas G Saacuteenz Y Zarazaga M Rodrigues J Torres CPolymorphisms of the pbp5 gene and correlation with ampicillinresistance in Enterococcus faecium isolates of animal origin J MedMicrobiol 2007 56236ndash240
26 Kuumlhn I Iversen A Burman LG Olsson-Liljequist B Franklin A Finn MAarestrup F Seyfarth AM Blanch AR Vilanova X Taylor H Caplin J MorenoMA Dominguez L Herrero IA Moumlllby R Comparison of enterococcalpopulations in animals humans and the environment ndash a Europeanstudy Int J Food Microbiol 2003 88133ndash145
27 Lanthier M Scott A Lapen DR Zhang Y Topp E Frequency of virulencegenes and antibiotic resistances in Enterococcus spp isolates fromwastewater and feces of domesticated mammals and birds and wildlifeCan J Microbiol 2010 56715ndash729
28 Leclercq R Oberleacute K Galopin S Cattoir V Budzinski H Petit F Changes inenterococcal populations and related antibiotic resistance along amedical center-wastewater treatment plant-river continuum Appl EnvironMicrobiol 2013 792428ndash2434
29 Thevenon F Regier N Benagli C Tonolla M Adatte T Wildi W Poteacute JCharacterization of fecal indicator bacteria in sediments cores from thelargest freshwater lake of Western Europe (Lake Geneva Switzerland)Ecotoxicol Environ Safety 2012 7850ndash56
30 Bonilla TD Nowosielski K Esiobu N McCorquodale DS Rogerson ASpecies assemblages of Enterococcus indicate potential sources offecal bacteria at a south Florida recreational beach Mar Pollut Bull2006 52807ndash810
31 Wang JT Chang SC Wang HY Chen PC Shiau YR Lauderdale TL TSARHospitals High rates of multidrug resistance in Enterococcus faecalis andE faecium isolated from inpatients and outpatients in TaiwanDiagn Microbiol Infect Dis 2013 75406ndash411
32 Lanthier M Scott A Zhang Y Cloutier M Durie D Henderson VC WilkesG Lapen DR Topp E Distribution of selected virulence genes andantibiotic resistance in Enterococcus species isolated from the SouthNation River drainage basin Ontario Canada J Appl Microbiol 2011110407ndash421
33 Kuch A Willems RJ Werner G Coque TM Hammerum AM Sundsfjord AKlare I Ruiz-Garbajosa P Simonsen GS van Luit-Asbroek M Hryniewicz WSadowy E Insight into antimicrobial susceptibility and population struc-ture of contemporary human Enterococcus faecalis isolates fromEurope J Antimicrob Chemother 2012 67551ndash558
34 Novais C Coque TM Ferreira H Sousa JC Peixe L Environmentalcontamination with vancomycin-resistant enterococci from hospitalsewage in Portugal Appl Environ Microbiol 2005 713364ndash3368
35 Iversen A Kuumlhn I Rahman M Franklin A Burman LG Olsson-Liljequist BTorell E Moumlllby R Evidence for transmission between humans and theenvironment of a nosocomial strain of Enterococcus faeciumEnviron Microbiol 2004 655ndash59
36 Rathnayake I Hargreaves M Huygens F SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterwayAustralia and associated antibiotic resistance gene profilesBMC Microbiol 2011 11201
37 Rosvoll TC Pedersen T Sletvold H Johnsen PJ Sollid JE Simonsen GSJensen LB Nielsen KM Sundsfjord A PCR-based plasmid typing inEnterococcus faecium strains reveals widely distributed pRE25-pRUM- pIP501- and pHT-related replicons associated with glycopeptideresistance and stabilizing toxin-antitoxin systems FEMS Immunol MedMicrobiol 2010 58254ndash268
38 Rosvoll TC Lindstad BL Lunde TM Hegstad K Aasnaes B Hammerum AMLester CH Simonsen GS Sundsfjord A Pedersen T Increased high-levelgentamicin resistance in invasive Enterococcus faecium is associated withaac(6prime)Ie-aph(2Prime)Ia-encoding transferable megaplasmids hosted by majorhospital-adapted lineages FEMS Immunol Med Microbiol 2012 66166ndash176
39 Laverde Gomez JA van Schaik W Freitas AR Coque TM Weaver KE FranciaMV Witte W Werner G A multiresistance megaplasmid pLG1 bearing ahylEfm genomic island in hospital Enterococcus faecium isolates Int J MedMicrobiol 2011 301165ndash175
40 Coque TM Evolutionary biology of pathogenic enterococci InEvolutionary Biology of Bacterial and Fungal Pathogens Coque TM Edited byBaquero F Nombela C Cassell GH Gutierrez JA Washington DC ASMPress501ndash523
41 Werner G Fleige C Geringer U van Schaik W Klare I Witte W IS elementIS16 as a molecular screening tool to identify hospital-associated strainsof Enterococcus faecium BMC Infect Dis 2011 1180
42 Top J Sinnige JC Majoor EA Bonten MJ Willems RJ van Schaik W Therecombinase IntA is required for excision of esp-containing ICEEfm1 inEnterococcus faecium J Bacteriol 2011 1931003ndash1006
43 Sillanpaumlauml J Prakash VP Nallapareddy SR Murray BE Distribution of genesencoding MSCRAMMs and Pili in clinical and natural populations ofEnterococcus faecium J Clin Microbiol 2009 47896ndash901
44 Marcinek H Wirth R Muscholl-Silberhorn A Gauer M Enterococcus faecalisgene transfer under natural conditions in municipal sewage watertreatment plants Appl Environ Microbiol 1998 64626ndash632
45 Kuumlmmerer K Significance of antibiotics in the environment J AntimicrobChemother 2003 525ndash7
46 Liu A Fong A Becket E Yuan J Tamae C Medrano L Maiz M Wahba C LeeC Lee K Tran KP Yang H Hoffman RM Salih A Miller JHP Selectiveadvantage of resistant strains at trace levels of antibiotics a simple andultrasensitive color test for detection of antibiotics and genotoxicagents Antimicrob Agents Chemother 2011 551204ndash1210
47 Clinical and Laboratory Standards Institute Performance standards forantimicrobial susceptibility testing 21st informational supplement Wayne CLSI2011 PA M100-S20
48 Magiorakos AP Srinivasan A Carey RB Carmeli Y Falagas ME Giske CGHarbarth S Hindler JF Kahlmeter G Olsson-Liljequist B Paterson DL Rice LBStelling J Struelens MJ Vatopoulos A Weber JT Monnet DLMultidrug-resistant extensively drug-resistant and pandrug-resistantbacteria an international expert proposal for interim standarddefinitions for acquired resistance Clin Microbiol Infect 2012 18268ndash281
49 Homan WL Tribe D Poznanski S Li M Hogg G Spalburg E Van Embden JDWillems RJ Multilocus sequence typing scheme for Enterococcus faeciumJ Clin Microbiol 2002 401963ndash1971
50 Jolley KA Feil EJ Chan MS Maiden MC Sequence type analysis andrecombinational tests (START) Bioinformatics 2001 171230ndash1231
51 Feil EJ Li BC Aanensen DM Hanage WP Spratt BG eBURST inferringpatterns of evolutionary descent among clusters of related bacterialgenotypes from multilocus sequence typing data J Bacteriol 20041861518ndash1530
52 Top J Banga1 NMI Hayes R Willems RJ Bonten MJM Hayden MKComparison of multiple-locus variable-number tandem repeat analysisand pulsed-field gel electrophoresis in a setting of polyclonal endemicityof vancomycin-resistant Enterococcus faecium Clin Microbiol Infect 200814363ndash369
53 Vankerckhoven V Van Autgaerden T Vael C Lammens C Chapelle S RossiR Jabes D Goossens H Development of a multiplex PCR for the
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-
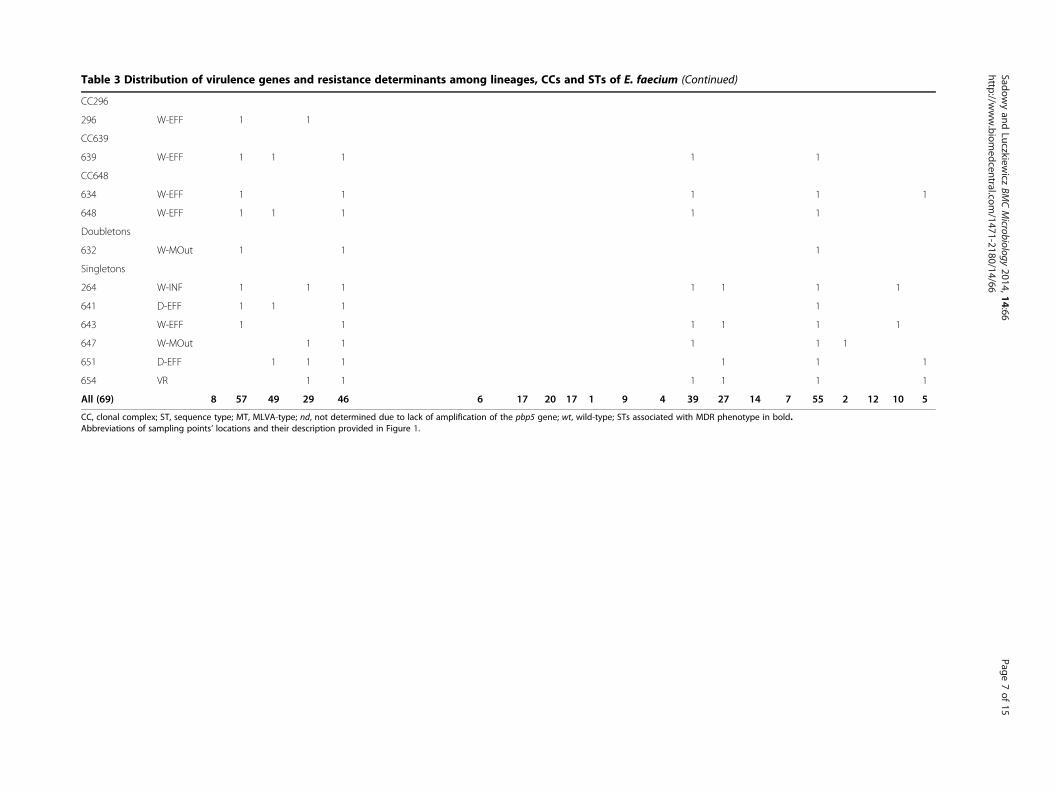
Table 3 Distribution of virulence genes and resistance determinants among lineages CCs and STs of E faecium (Continued)
CC296
296 W-EFF 1 1
CC639
639 W-EFF 1 1 1 1 1
CC648
634 W-EFF 1 1 1 1 1
648 W-EFF 1 1 1 1 1
Doubletons
632 W-MOut 1 1 1
Singletons
264 W-INF 1 1 1 1 1 1 1
641 D-EFF 1 1 1 1
643 W-EFF 1 1 1 1 1 1
647 W-MOut 1 1 1 1 1
651 D-EFF 1 1 1 1 1 1
654 VR 1 1 1 1 1 1
All (69) 8 57 49 29 46 6 17 20 17 1 9 4 39 27 14 7 55 2 12 10 5
CC clonal complex ST sequence type MT MLVA-type nd not determined due to lack of amplification of the pbp5 gene wt wild-type STs associated with MDR phenotype in boldAbbreviations of sampling pointsrsquo locations and their description provided in Figure 1
Sadowyand
Luczkiewicz
BMCMicrobiology
20141466Page
7of
15httpw
wwbiom
edcentralcom1471-21801466
Figure 2 MLST-based similarity tree of E faecium isolates constructed using the START software and the UPGMA clustering algorithm
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 8 of 15httpwwwbiomedcentralcom1471-21801466
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 9 of 15httpwwwbiomedcentralcom1471-21801466
grouped the 38 STs specific for 50 isolates into the majorlsquosuper-complex 17rsquo characteristic for this species (data notshown) Further detailed analysis of the localization ofparticular STs within this structure revealed that 11 STscharacteristic for 17 isolates belonged to the nosocomialcomplex of E faecium representing all three majorlineages 17 18 and 78 of a former CC17 as defined byWillems et al [9] (Table 3) Seventeen STs (20 isolates)nine STs (11 isolates) and two STs with single isolates eachbelonged to CC5 CC22 and CC9 respectively as definedby Freitas et al [24] Of the remaining 16 STs nine wereincluded into six small CCs (CCs 27 46 94 296 639648) unlinked to the major lsquosuper-complexrsquo ST632formed a lsquodoubletonrsquo (ie was linked by a single-locus vari-ant link to only one ST from the database) and six STsrepresented by single isolates were singletons ie werenot related to any ST in the database at the time of ana-lysis Thirty isolates (435) in the investigated groupshowed the MDR phenotype which was significantly asso-ciated (p = 000006) with isolates from the nosocomialcomplex (Table 3 and Figure 3) but found also in CC5CC22 and CC94 this phenotype did not occur amongsmall CCs or singletons Seventeen isolates repre-senting the nosocomial complex were additionallystudied by multilocus VNTR (variable number tan-dem repeats) analysis (MLVA) to better elucidate therelationships among these isolates Altogether nineMLVA-types (MTs) were found (Table 3) Importantlythe same MLVA-type MT159 associated with ST78was characteristic for isolates derived from samplesfrom W-EFF and W-MOut suggesting survival of thisparticular clone
0
10
20
30
40
50
60
70
80
90
100
IS16
espintA
fms5
fms1
7
fms1
9
fms2
1
MDR
AM-R
CIP-R
HLG
R
p
ositi
ve
isol
ates
Figure 3 Distribution () of IS16 resistance phenotypes and virulenccolumns isolates with STs typical for nosocomial E faecium grey columnsgroups are indicated by an asterisk MDR multi-drug resistance AM-R ampgentamicin resistance HLSR high-level streptomycin resistance
IS16 was present in all 17 isolates included by MLST tothree lineages of nosocomial E faecium and this markerwas limited to this group (Table 3 and Figure 3) TheespEfm gene occurred always together with intA among allisolates of lineage 17 and in some isolates of lineage 78The fms19 pilin gene detected in 29 isolates was over-represented among STs typical for nosocomial E faeciumwhile other pilin genes such as fms5 fms17 and fms21detected in 57 49 and 46 isolates respectively occurredamong various CCs and singleton STs Ampicillin resist-ance significantly associated with isolates of nosocomialcomplex (10 of 17 isolates 588 p = 0001) occurred alsoin CC5 (6 isolates) CC22 and CC94 (1 isolate each) Se-quencing of the 3prime part of the pbp5 gene revealed its 11nucleotide alleles corresponding to 11 variants of the C-terminal part of PBP5 Six alleles of the gene were new(pbp5-24 to pbp5-29) two alleles (pbp5-20 pbp5-22)were described previously among Polish hospital VREmisolates [13] and three alleles were found in other stud-ies pbp5-30 occurred in a Portuguese animal strain ofE faecium [25] and pbp5-31 and pbp5-32 in clinical iso-lates TCGE70411_2 and 164306 also from Portugal (de-posited as JN208888 and JN208885 in the GenBankrespectively) For a single ampicillin-resistant isolate ofST653 (lineage 78) pbp5 amplification was not achieveddespite repeated attempts The majority (11 of 17 iso-lates 647) of isolates belonging to three lineages ofnosocomial E faecium showed resistance to ciprofloxa-cin determined by mutations R61G and S80IR in ParCand S84IRY in GyrA Ciprofloxacin resistance wasalso significantly over-represented among isolates ofSTs typical for nosocomial E faecium (p = 00006) Nine
HLS
R
tetM tetL
rep1
-pIP50
1
rep2
-pRE25
rep1
7-pR
UM
rep1
8-pE
F418
rep-pL
G1
axe-txe
relBE
e resistance and plasmid genes among E faecium isolates Blackother E faecium isolates Significant differences (p lt 005) between twoicillin resistance CIP-R ciprofloxacin resistance HLGR high-level
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 10 of 15httpwwwbiomedcentralcom1471-21801466
isolates resistant to ciprofloxacin were found also inCCs 5 9 and 22 These isolates possessed mutations inParC mentioned above but had the wild-type QRDR inGyrA HLGR occurred in five isolates of lineages 18 and78 and in single isolates of CC5 and CC22 this pheno-type was determined by the presence of aac(6prime)-Ie-aph(2Prime) with an exception of an isolate from CC5 whichwas negative for this gene High-level resistance tostreptomycin (HLSR) was widely distributed amongstudied isolates and determined by the ant(6prime)-Ia geneTetracycline resistance was found also in various STsthe major determinant of this phenotype was tet(M) (20isolates) followed by tet(L) (17 isolates) These twogenes occurred together in 16 isolates A single isolateof ST633 belonging to CC22 harbored tet(S) The tet(O) gene was absent in the studied group For a singletetracycline-resistant isolate of ST17 none of the fourtetracycline determinants studied was detected Nineisolates carried int of Tn916 and four the tndX gene ofTn5397 These transposon-specific genes were not char-acteristic for any lineage or complex (Table 3) Plasmid-replication genes such as rep2pRE25 rep17pRUM andrep18 pEF418 (26 14 and 7 isolates respectively) andgenes specifying toxin-antitoxin systems (TAS) axe-txeand ω-ε-ζ (12 and 10 isolates respectively) were signifi-cantly associated with the group of isolates of nosocomialE faecium Two other rep genes (rep1pIP501 reppLG1) werewidely distributed among all studied E faecium isolatesand the reppMG1 gene occurred in two isolates (in a singleisolate of ST533 from CC22 and in a singleton ST647) Allω-ε-ζ-positive isolates carried rep2pRE25 and nine isolatesshowed a concomitant presence of axe-txe and rep17pRUMThe relBE genes were found solely among five non-nosocomial E faecium (a difference in distribution with-out statistical significance p = 043) among various STs
DiscussionIn this study Enterococcus spp was used to evaluate theimpact posed by treated wastewater of two local WWTPson their receiver the costal waters of Gulf of GdanskTotal counts of enterococci observed for raw (W-INF)and treated (W-EFF D-EFF) wastewater are consistentwith our previous study showing similar over 99 reduc-tion capacity of applied treatment processes It is alsoreported in the literature that wastewater treatment pro-cesses based on activated sludge although they focus onremoval of nutrients and organic matter simultaneouslyreduce faecal indicators with high efficiency reaching upto 999 [1822] Due to the high number of enterococciin raw wastewater they are nevertheless released from aWWTP in a number up to 61x105 CFU per 100 mLThus marine water sampled in the area directly impactedby treated wastewater (W-MOut and D-MOut) showed ahigher number of enterococci than the one observed in
Vistula River mouth however lower by four orders ofmagnitude in comparison to corresponding treated waste-water (W-EFF and D-EFF) It should be noted that in mar-ine outfalls (W-MOut D-MOut) as well as in VistulaRiver mouth enterococci were detected in number lowerthan 100 CFU per 100 mL ie the value set out by NewBathing Water Directive 20067EC for the costal water ofldquoexcellent qualityrdquo [23]It is suggested that enterococcal species composition in
faecal-impacted environment depends from the source ofcontamination In this study two species most commonlycolonizing humans E faecalis and E faecium comprisedtogether from 73 to 88 of enterococcal isolates intested wastewater samples (W-INF W-BR W-EFF and D-EFF) 73 and 61 in marine outflows (W-MOut and D-MOut respectively) and 45 in the Vistula River mouthThe predominant species in wastewater and marine waterimpacted by treated wastewater was E faecium (from 56to 65) which stayed at a similar level during treatmentprocesses as determined also in our previous study [20]Species distribution in wastewater samples is not wellunderstood and affecting factors may include differencesin diet climate season and methods of detection E feaca-lis was found to be the most prevalent species in wastewa-ter samples in Sweden [26] E faecium in Spain UKCanada France and Switzerland [26-29] while E hiraepredominated in Portugal and United States [2230] Al-though E faecalis is typically the most abundant colonizerof humans in the community (eg [2631]) E faecium isgenerally most often associated with human microbialcontamination [2729] E faecalis was reported fromagriculture-impacted waters [32] and E hirae likelyoriginates mainly from cattle and pig faecal contamin-ation [26] which explains high relative proportion ofthese two species in Vistula River mouth Apart from thevariability of contamination sources proportion of par-ticular species may differ due to the differences of abilityto sustain environmental stresses In this study enterococ-cal composition might have been influenced by wastewa-ter treatment processes and marine water characteristicsThe effect of wastewater treatment and marine environ-
ment was seen on phenotypes of antimicrobial resistanceThe positive selection of bacteria with resistance patternshas been already suggested and noted in wastewater pro-cesses [192022] In this study the resistance rates to ampi-cillin ciprofloxacin levofloxacin and erythromycin notedfor E faecium from raw wastewater (W-INF) were lowerthan these observed for this species in the correspondingtreated wastewater (W-EFF) however not statistically sig-nificant (p gt 005) Moreover resistance rates to ampicillinciprofloxacin tetracycline and erythromycin observed inmarine outflow of WWTP Gdansk-Wschod (W-MOut)were higher than in treated wastewater of this plantSuch phenomenon was not observed for treated
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 11 of 15httpwwwbiomedcentralcom1471-21801466
wastewater of WWTP Gdynia-Debogorze (D-EFF) andits marine outfall (D-MOut) Resistance rates among Efaecium originating from D-MOut were in generalcomparable to these detected in the Vistula Rivermouth It should be noted that treated wastewater fromWWTP Gdansk-Wschod has been discharged via mar-ine outfall (W-MOut) for last 12 years while marineoutfall of WWTP Gdynia-Debogorze (D-MOut) hasbeen operating for last two years The observed differ-ences in resistance rate corresponding to the local im-pact caused by long and short-term operated marineoutfalls (W-MOut and D-MOut) need further attentionSignificant rates of antimicrobial resistance observed
for isolates of E faecalis from wastewater and marineriver waters impacted by wastewater are in agreementwith a recent study performed in six European countriesshowing that in Poland the rates of resistance for humanisolates of this species were usually among the highestboth in the community and hospitals [33] To our know-ledge no corresponding data are available for E faeciumcolonization in the community in Poland but a studyfocused on hospital VRE isolates showed their ubiquitousMDR phenotype and a high load of resistance genes [13]In our study no vancomycin resistance were detectedlikely due to relatively low (below 10) prevalence of VREin Polish hospitals [5] although such isolates are isolatedfrom the environment in other countries eg [34]This study shows that hospital clones of E faecium are
important carriers of resistance genes in wastewatersMost likely in a significant part they originated fromhospital wastewater treated together with communalwastewater In our estimation hospital wastewater con-stitutes approximately 02 of both WWTPsrsquo daily in-flow The release of hospital-associated clones of Efaecium to the environment was observed also by others[24283536] In our study isolates belonging to themajor nosocomial HiRECC (formerly named CC17)constituted altogether 246 of all isolates of E faeciumsubjected to typing and were present at all samplingsites These isolates apart from their inclusion into theHiRECC by MLST demonstrated several other pheno-typic and genotypic features typical for nosocomial Efaecium ie they were resistant to ciprofloxacin andampicillin in vast majority showed MDR phenotypeand carried several resistance determinants associatedboth with mutations of chromosomal genes (gyrA parCpbp5) and in the case of aminoglycoside and tetracyclineresistance with gene acquisition Plasmids likely played animportant role in the later process as some plasmid genessuch as rep2pRE25 rep17pRUM and rep18 pEF418 and TASgenes axe-txe and ω-ε-ζ were significantly more abundantin isolates associated with HiRECC than among theremaining isolates As yet the knowledge of distributionof plasmid-associated genes in populations of E faecium
especially in the context of their clonal compositionremains limited A study on 93 isolates of differentgeographical origins and from various sites showed anover-representation of rep17pRUM and reppMG1 amongnosocomial E faecium while in contrast to our observa-tions rep18 pEF418 was not detected [37] Moreover wedid not observe reppMG1 among HiRECC of E faeciumThese differences may be due to a high prevalence of VRE(58 isolates) among 93 isolates studied by Rosvoll et alwhat may explain differences in the plasmid content Thesame study also reported the co-localization of ω-ε-ζ withrep2pRE25 and axe-txe with rep17pRUM on the same plas-mids in concordance with our findings showing a fre-quent joint occurrence of these pairs of genes Anotherstudy performed on 99 invasive isolates from Norwaymostly of HLGR phenotype showed a significantly higherprevalence of rep2pRE25 and rep17pRUM among the majornosocomial HiRECC [38] This study as well some others[2439] also demonstrated a common presence of reppLG1among E faecium from hospital settings and in animalsOur findings indicate that this type of replicon is evenmore ubiquitous in E faecium than previously reportedAll rep genes mentioned above are typically associatedwith plasmids carrying various antimicrobial resistancegenes [3940] GenBank accession number AF408195 Inaddition to genes determining antimicrobial resistanceand genes associated with MGE isolates of nosocomialHiRECC analyzed in our study carried such molecularmarkers as IS16 and the esp gene together with the intAgene characteristic for ICEEfm [4142] Another patho-genicity factor the fms19 gene of the pilin gene cluster4 (PGC4) showed increased prevalence among theseisolates Similar observation was made for a representa-tive collection of 433 E faecium isolates from varioussources [43]In a study that followed enterococci in a continuum
from hospital and retirement home wastewaters througha WWTP to a river a gradual decrease in the proportionof E faecium isolates representing hospital-associatedSTs was observed likely due to the ldquodilutionrdquo of HiRECCrepresentatives by more diverse and less drug-resistantstrains originating from the community [28] Because inour study only selected representatives of obtained iso-lates were investigated it is not possible to determineprecise tendencies in their frequency from the WWTPinfluent to effluent and water however resistant entero-cocci both HiRECC and non-HiRECC are clearly ableto survive the treatment process what results in their re-lease into marine and river waters Moreover using anadditional typing method MLVA we were able to findisolates with the same characteristics in the effluent ofWWTP and its marine outfall suggesting ability of suchclones to survive in the environment for at least sometime It seems plausible that hospital resistant clones
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 12 of 15httpwwwbiomedcentralcom1471-21801466
accumulate several determinants that may promote theirsurvival not only in the hospital settings but also in theenvironment Such features would include resistance todisinfectants and ultraviolet light additional metabolicpathways and ability to form biofilms Moreover determi-nants of some of the mentioned adaptation factors arefound co-resident in plasmids harboring resistance genessuggesting co-selection For example pLG1-type plasmidsof E faecium carry not only resistance determinantsagainst aminoglycosides glycopeptides and macrolidesbut also pili genes and carbon uptake-utilization genes[3839] Pheromone-responsive plasmids of E faecalisapart from resistance to aminoglycosides glycopeptidespenicillins and macrolides may encode bacteriocins add-itional pili and UV-resistance determinants [40] Thusnosocomial HiRECCs released by WWTPs are equippedin whole sets of genes with various adaptive benefits andoften located within MGE It was also demonstrated thatMGEs can be transferred in a WWTP [44] further under-ling the importance of threat posed by such organismsThere is a selection pressure for survival of resistantstrains and their tranconjugants due to release of antibi-otics to the environment especially in the hospital efflu-ents [45] and even sub-inhibitory concentrations may beeffective in selection for resistance [46]
ConclusionsThis study describes the species composition resistanceprofiles and clonal relationships of enterococci isolatedfrom WWTPs located in Gdansk their marine outfallsto the Gulf of Gdansk and from Vistula River It was ob-served that these WWTPs release enterococcal strainsresistant to various antimicrobial compounds to the re-cipient water environment Molecular analyses revealedthe presence of clones associated with nosocomial HiR-ECCs and in the case of E faecium isolates of lineages17 18 and 78 of former CC17 abundant in resistance de-terminants MGE genes and pathogenicity factors Thepresence of HiRECCs in wastewaters and marineriverenvironment highlights the need for further detailedanalyses to better understand the survival and spread ofdrug-resistant strains in water ecosystems and to eluci-date the ways to curb such dissemination
MethodsAcquisition of samples and isolation of enterococciOnce a month in April June August and October in 2011of the flow proportioned composited samples of wastewa-ter were taken from two local WWTPs Gdansk-Wschodfrom influent (W-INF) effluent (W-EFF) and wastewa-ter from activated sludge bioreactor (W-BR) and fromGdynia-Debogorze effluent (D-EFF) The two WWTPstreat sewage from approximately 920 000 inhabitants(for details see Figure 1) In the same period of time and
with the same frequency marine outfalls of those WWTPs(W-MOut and D-MOut respectively) as well as VistulaRiver mouth were also sampled for isolation of entero-cocci Marine samples were taken at the depth of approxi-mately one meter below the surface once a month inApril June August and October of 2011 Location ofsampling points and their characteristics are provided inFigure 1 Enterococci were detected using the membranefiltration method To this end appropriate dilutions ofanalyzed samples were filtered in duplicate through045 μm cellulose-acetate filters (EMD Millipore Cor-poration Billerica MA USA) which were then placedon Enterococcus selective agar (Merck DarmstadtGermany) and incubated at 37degC for 48 h (ISO 7899ndash22000) Dark red or maroon colonies were consideredpresumptive enterococci For further investigationsrepresentative isolates were taken from membranes pre-senting from 20 to 50 typical colonies and stored in nu-trient broth supplemented with 15 glycerol at minus80degC
Species identification DNA isolation susceptibility testingand statistical analysisThe species identification and drug susceptibility of pre-sumptive enterococci were determined by the PhoenixtradeAutomated Microbiology System (BD New Jersey USA)according to the manufacturerrsquos instructions Since it isnot possible to distinguish E casseliflavus and E galli-narum using this system the classification E casselifla-vusgallinarum was used The susceptibility tests basedon microdilution were carried out for 11 antimicrobialagents gentamicin (GM) streptomycin (ST) ampicillin(AM) daptomycin (DAP) vancomycin (VA) teicoplanin(TEC) linezolid (LZD) fluoroquinolones (ciprofloxacin -CIP levofloxacin - LVX) tetracycline (TE) erythromycin(E) and quinupristin-dalfopristin (SYN) (E faecium only)Obtained minimal inhibitory concentrations (MIC) wereevaluated according to the guidelines of Clinical and La-boratory Standards Institute [47] Isolates were defined asmultidrug-resistant (MDR) when they showed resistanceto three or more compounds tested [48] Bacterial DNAwas purified using the Genomic DNA Prep Plus kitfollowing the manufacturerrsquos instructions (AampA Biotech-nology Gdynia Poland) Differences in distributions wereevaluated using the χ2 test with the p values lt 005 con-sidered significant
MLST and MLVA of E faeciumMLST for E faecium was performed by sequencing ofseven house-keeping genes as described by Homan et al[49] The allele numbers and STs were assigned with theuse of Internet database httpefaeciummlstnet (5th July2013 date last accessed) Novel alleles and allelic profileswere submitted to the database MLST data were analyzedwith Sequence Type Analysis and Recombinational Tests
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 13 of 15httpwwwbiomedcentralcom1471-21801466
(START) software [50] and the eBURST analysis [51] wasused to elucidate the relationships of STs with knownCCs using the comparative eBURST option available athttpeburstmlstnetv3enter_datacomparative (5th July2013 date last accessed) Further inclusion of E faeciumSTs into specific CCs and lineages was performed fol-lowing other studies [924] MLVA was performed as de-scribed by others [52]
Detection and analysis of genes associated with virulenceantimicrobial resistance and MGE in E faeciumThe full list of primers used for gene detection and se-quencing is provided in Additional file 1 Table S1 PCR-based detection of IS16 virulence-associated genes (fms21fms17 fms5 and fms19 representing each of four pilingene clusters in E faecium and the espEfm gene) genes de-termining resistance to tetracycline tet(M) tet(O) tet(L)tet(S) HLGR (aac(6prime)-Ie-aph(2Prime) HLSR (ant(6prime)-Ia) andgenes specific for MGE such as intA of ICEEfm1 int ofTn916 transposon tndX of Tn5397 transposon entero-coccal plasmid replication genes rep1pIP501 rep2pRE25rep17pRUM rep18pEF418 reppMG1 reppLG1 and TAS genesaxe-txe ω-ε-ζ relBE was performed as described in otherstudies [134153-59] following the conditions describedherein Previously characterized clinical isolates of Strepto-coccus agalactiae E faecium and E faecalis harbouringpilin genes IS16 espEfm intA of ICEEfm1 int of Tn916tndX of Tn5397 tet(M) tet(O) tet(L) tet(S) aac(6prime)-Ie-aph(2Prime) ant(6prime)-Ia rep1pIP501 rep2pRE25 ω-ε-ζ [136061]served as positive controls In cases when positive controlswere not available (rep18pEF418 reppMG1 reppLG1 axe-txerelBE) a few randomly selected PCR products wereverified by sequencing and served further as controlsAmpicillin-resistance determinant the pbp5 gene was ana-lysed by sequencing of the part encoding C-terminaltranspeptidase domain in PBP5 [62] New sequence vari-ants pbp5-24 to pbp5-29 alleles were submitted to theGenBank (acc numbers KC594860-KC594865 respect-ively) The quinolone-determining regions (QRDR) in thegyrA and parC genes were sequenced and analysed asdescribed by others [63]
Additional file
Additional file 1 Table S1 Primers used in the study for PCR andsequencing
Competing interestThe authors declare that they have no competing interest
Authorsrsquo contributionsES participated in the study design performed experimental work andparticipated in the manuscript preparation AL designed the studyperformed experimental work and participated in the manuscriptpreparation Both authors read and approved the final manuscript
AcknowledgementsWe thank Rob Willems and Janetta Top for assigning new alleles new STsand MTs This publication made use of the Enterococcus faecium MLSTwebsite (httpefaeciummlstnet) which is hosted at Imperial College of theUniversity of Oxford and funded by the Wellcome Trust This work wassupported by project N N305 461139 from the Narodowe Centrum Nauki(NCN)
Author details1National Medicines Institute ul Chelmska 3034 Warsaw 00-725 Poland2Gdansk University of Technology ul G Narutowicza 1112 Gdansk 80-952Poland
Received 5 September 2013 Accepted 27 February 2014Published 14 March 2014
References1 Aarestrup FM Butaye P Witte W Nonhuman reservoirs of Enterococci In
American Society for Microbiology Edited by Gilmore MS Clewell DB CourvalinP Dunny GM Murray BE Rice LB Washington DC The Enterococcipathogenesis molecular biology and antibiotic resistance 2002
2 Tendolkar PM Baghdayan AS Shankar N Pathogenic enterococci newdevelopments in the 21st century Cell Mol Life Sci 2003 602622ndash2636
3 Sood S Malhotra M Das BK Kapil A Enterococcal infections ampantimicrobial resistance Indian J Med Res 2008 128111ndash121
4 Sydnor ER Perl TM Hospital epidemiology and infection control inacute-care settings Clin Microbiol Rev 2011 24141ndash173
5 European Centre for Disease Prevention and Control Annual EpidemiologicalReport Reporting on 2010 surveillance data and 2011 epidemic intelligencedata Stockholm ECDC 20122013
6 Hidron AI Edwards JR Patel J Horan TC Sievert DM Pollock DA Fridkin SKNational Healthcare Safety Network Team Participating National HealthcareSafety Network Facilities NHSN annual update antimicrobial-resistantpathogens associated with healthcare-associated infections annualsummary of data reported to the National Healthcare Safety Network atthe Centers for Disease Control and Prevention 2006ndash2007 Infect ControlHosp Epidemiol 2008 29996ndash1011
7 De Kraker ME Jarlier V Monen JC Heuer OE van de Sande N GrundmannH The changing epidemiology of bacteraemias in Europe trends fromthe European Antimicrobial Resistance Surveillance System Clin MicrobiolInfect 2012 doi1011111469-0691
8 Leavis HL Bonten MJ Willems RJ Identification of high-risk enterococcalclonal complexes global dispersion and antibiotic resistance Curr OpinMicrobiol 2006 9454ndash460
9 Willems RJL Top J van Schaik W Leavis H Bonten M Sireacuten J Hanage WPCorander J Restricted gene flow among hospital subpopulations ofEnterococcus faecium mBio 2012 3e00151ndash12
10 Hegstad K Mikalsen T Coque TM Werner G Sundsfjord A Mobile geneticelements and their contribution to the emergence of antimicrobialresistant Enterococcus faecalis and Enterococcus faecium Clin MicrobiolInfect 2010 16541ndash554
11 Shepard BD Gilmore MS Antibiotic-resistant enterococci themechanisms and dynamics of drug introduction and resistance MicrobesInfect 2002 4215ndash224
12 Bonten MJ Willems R Weinstein RA Vancomycin-resistant enterococciwhy are they here and where do they come from Lancet Infect Dis 20011314ndash325
13 Sadowy E Sieńko A Gawryszewska I Bojarska A Malinowska K HryniewiczW High abundance and diversity of antimicrobial resistancedeterminants among early vancomycin-resistant Enterococcus faecium inPoland Eur J Clin Microbiol Infect Dis 2013 321193ndash1203
14 Willems RJ Homan W Top J van Santen-Verheuvel M Tribe D Manzioros XGaillard C Vandenbroucke-Grauls CM Mascini EM van Kregten E vanEmbden JD Bonten MJ Variant esp gene as a marker of a distinct geneticlineage of vancomycin-resistant Enterococcus faecium spreading inhospitals Lancet 2001 357853ndash855
15 Sillanpaumlauml J Nallapareddy SR Prakash VP Qin X Houmloumlk M Weinstock GMMurray BE Identification and phenotypic characterization of a secondcollagen adhesin Scm and genome-based identification and analysis of13 other predicted MSCRAMMs including four distinct pilus loci inEnterococcus faecium Microbiology 2008 1543199ndash3211
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 14 of 15httpwwwbiomedcentralcom1471-21801466
16 Galloway-Pentildea JR Nallapareddy SR Arias CA Eliopoulos GM Murray BEAnalysis of clonality and antibiotic resistance among early clinicalisolates of Enterococcus faecium in the United States J Infect Dis 20092001566ndash1573
17 Top J Willems R Bonten M Emergence of CC17 Enterococcus faeciumfrom commensal to hospital-adapted pathogen FEMS Immunol MedMicrobiol 2008 52297ndash308
18 Martins da Costa P Vaz-Pires P Bernardo F Antimicrobial resistance inEnterococcus spp isolated in inflow effluent and sludge from municipalsewage water treatment plants Water Res 2006 401735ndash1740
19 Reinthaler FF Posch J Feierl G Wust G Haas D Ruckenbauer G Mascher FMarth E Antibiotic resistance of Escherichia coli in sewage and sludgeWater Res 2003 371685ndash1690
20 Luczkiewicz A Fudala-Ksiazek S Jankowska K Olanczuk-Neyman KAntimicrobial resistance of fecal indicators in municipal wastewatertreatment plant Water Res 2010 445089ndash5097
21 Dulfour AP Ballentine RK Ambient water quality criteria for bacteria-1986 EPA4405-84-002 Washington DC US Environmental Protection Agency 1986
22 Ferreira da Silva M Tiago I Veriacutessimo A Boaventura RA Nunes OC Manaia CMAntimicrobial resistance patterns in Enterobacteriaceae isolated from anurban wastewater treatment plant FEMS Microbiol Ecol 2007 60166ndash176
23 DIRECTIVE 20067EC of the European Parliament and of the Council 15February 2006 concerning the management of bathing water quality andrepealing Directive 76160EEC httpeur-lexeuropaeuLexUriServLexUriServdouri=OJL200606400370051ENPDF
24 Freitas AR Novais C Ruiz-Garbajosa P Coque TM Peixe L Dispersion ofmultidrug-resistant Enterococcus faecium isolates belonging to majorclonal complexes in different Portuguese settings Appl Environ Microbiol2009 754904ndash4908
25 Poeta P Costa D Igrejas G Saacuteenz Y Zarazaga M Rodrigues J Torres CPolymorphisms of the pbp5 gene and correlation with ampicillinresistance in Enterococcus faecium isolates of animal origin J MedMicrobiol 2007 56236ndash240
26 Kuumlhn I Iversen A Burman LG Olsson-Liljequist B Franklin A Finn MAarestrup F Seyfarth AM Blanch AR Vilanova X Taylor H Caplin J MorenoMA Dominguez L Herrero IA Moumlllby R Comparison of enterococcalpopulations in animals humans and the environment ndash a Europeanstudy Int J Food Microbiol 2003 88133ndash145
27 Lanthier M Scott A Lapen DR Zhang Y Topp E Frequency of virulencegenes and antibiotic resistances in Enterococcus spp isolates fromwastewater and feces of domesticated mammals and birds and wildlifeCan J Microbiol 2010 56715ndash729
28 Leclercq R Oberleacute K Galopin S Cattoir V Budzinski H Petit F Changes inenterococcal populations and related antibiotic resistance along amedical center-wastewater treatment plant-river continuum Appl EnvironMicrobiol 2013 792428ndash2434
29 Thevenon F Regier N Benagli C Tonolla M Adatte T Wildi W Poteacute JCharacterization of fecal indicator bacteria in sediments cores from thelargest freshwater lake of Western Europe (Lake Geneva Switzerland)Ecotoxicol Environ Safety 2012 7850ndash56
30 Bonilla TD Nowosielski K Esiobu N McCorquodale DS Rogerson ASpecies assemblages of Enterococcus indicate potential sources offecal bacteria at a south Florida recreational beach Mar Pollut Bull2006 52807ndash810
31 Wang JT Chang SC Wang HY Chen PC Shiau YR Lauderdale TL TSARHospitals High rates of multidrug resistance in Enterococcus faecalis andE faecium isolated from inpatients and outpatients in TaiwanDiagn Microbiol Infect Dis 2013 75406ndash411
32 Lanthier M Scott A Zhang Y Cloutier M Durie D Henderson VC WilkesG Lapen DR Topp E Distribution of selected virulence genes andantibiotic resistance in Enterococcus species isolated from the SouthNation River drainage basin Ontario Canada J Appl Microbiol 2011110407ndash421
33 Kuch A Willems RJ Werner G Coque TM Hammerum AM Sundsfjord AKlare I Ruiz-Garbajosa P Simonsen GS van Luit-Asbroek M Hryniewicz WSadowy E Insight into antimicrobial susceptibility and population struc-ture of contemporary human Enterococcus faecalis isolates fromEurope J Antimicrob Chemother 2012 67551ndash558
34 Novais C Coque TM Ferreira H Sousa JC Peixe L Environmentalcontamination with vancomycin-resistant enterococci from hospitalsewage in Portugal Appl Environ Microbiol 2005 713364ndash3368
35 Iversen A Kuumlhn I Rahman M Franklin A Burman LG Olsson-Liljequist BTorell E Moumlllby R Evidence for transmission between humans and theenvironment of a nosocomial strain of Enterococcus faeciumEnviron Microbiol 2004 655ndash59
36 Rathnayake I Hargreaves M Huygens F SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterwayAustralia and associated antibiotic resistance gene profilesBMC Microbiol 2011 11201
37 Rosvoll TC Pedersen T Sletvold H Johnsen PJ Sollid JE Simonsen GSJensen LB Nielsen KM Sundsfjord A PCR-based plasmid typing inEnterococcus faecium strains reveals widely distributed pRE25-pRUM- pIP501- and pHT-related replicons associated with glycopeptideresistance and stabilizing toxin-antitoxin systems FEMS Immunol MedMicrobiol 2010 58254ndash268
38 Rosvoll TC Lindstad BL Lunde TM Hegstad K Aasnaes B Hammerum AMLester CH Simonsen GS Sundsfjord A Pedersen T Increased high-levelgentamicin resistance in invasive Enterococcus faecium is associated withaac(6prime)Ie-aph(2Prime)Ia-encoding transferable megaplasmids hosted by majorhospital-adapted lineages FEMS Immunol Med Microbiol 2012 66166ndash176
39 Laverde Gomez JA van Schaik W Freitas AR Coque TM Weaver KE FranciaMV Witte W Werner G A multiresistance megaplasmid pLG1 bearing ahylEfm genomic island in hospital Enterococcus faecium isolates Int J MedMicrobiol 2011 301165ndash175
40 Coque TM Evolutionary biology of pathogenic enterococci InEvolutionary Biology of Bacterial and Fungal Pathogens Coque TM Edited byBaquero F Nombela C Cassell GH Gutierrez JA Washington DC ASMPress501ndash523
41 Werner G Fleige C Geringer U van Schaik W Klare I Witte W IS elementIS16 as a molecular screening tool to identify hospital-associated strainsof Enterococcus faecium BMC Infect Dis 2011 1180
42 Top J Sinnige JC Majoor EA Bonten MJ Willems RJ van Schaik W Therecombinase IntA is required for excision of esp-containing ICEEfm1 inEnterococcus faecium J Bacteriol 2011 1931003ndash1006
43 Sillanpaumlauml J Prakash VP Nallapareddy SR Murray BE Distribution of genesencoding MSCRAMMs and Pili in clinical and natural populations ofEnterococcus faecium J Clin Microbiol 2009 47896ndash901
44 Marcinek H Wirth R Muscholl-Silberhorn A Gauer M Enterococcus faecalisgene transfer under natural conditions in municipal sewage watertreatment plants Appl Environ Microbiol 1998 64626ndash632
45 Kuumlmmerer K Significance of antibiotics in the environment J AntimicrobChemother 2003 525ndash7
46 Liu A Fong A Becket E Yuan J Tamae C Medrano L Maiz M Wahba C LeeC Lee K Tran KP Yang H Hoffman RM Salih A Miller JHP Selectiveadvantage of resistant strains at trace levels of antibiotics a simple andultrasensitive color test for detection of antibiotics and genotoxicagents Antimicrob Agents Chemother 2011 551204ndash1210
47 Clinical and Laboratory Standards Institute Performance standards forantimicrobial susceptibility testing 21st informational supplement Wayne CLSI2011 PA M100-S20
48 Magiorakos AP Srinivasan A Carey RB Carmeli Y Falagas ME Giske CGHarbarth S Hindler JF Kahlmeter G Olsson-Liljequist B Paterson DL Rice LBStelling J Struelens MJ Vatopoulos A Weber JT Monnet DLMultidrug-resistant extensively drug-resistant and pandrug-resistantbacteria an international expert proposal for interim standarddefinitions for acquired resistance Clin Microbiol Infect 2012 18268ndash281
49 Homan WL Tribe D Poznanski S Li M Hogg G Spalburg E Van Embden JDWillems RJ Multilocus sequence typing scheme for Enterococcus faeciumJ Clin Microbiol 2002 401963ndash1971
50 Jolley KA Feil EJ Chan MS Maiden MC Sequence type analysis andrecombinational tests (START) Bioinformatics 2001 171230ndash1231
51 Feil EJ Li BC Aanensen DM Hanage WP Spratt BG eBURST inferringpatterns of evolutionary descent among clusters of related bacterialgenotypes from multilocus sequence typing data J Bacteriol 20041861518ndash1530
52 Top J Banga1 NMI Hayes R Willems RJ Bonten MJM Hayden MKComparison of multiple-locus variable-number tandem repeat analysisand pulsed-field gel electrophoresis in a setting of polyclonal endemicityof vancomycin-resistant Enterococcus faecium Clin Microbiol Infect 200814363ndash369
53 Vankerckhoven V Van Autgaerden T Vael C Lammens C Chapelle S RossiR Jabes D Goossens H Development of a multiplex PCR for the
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-
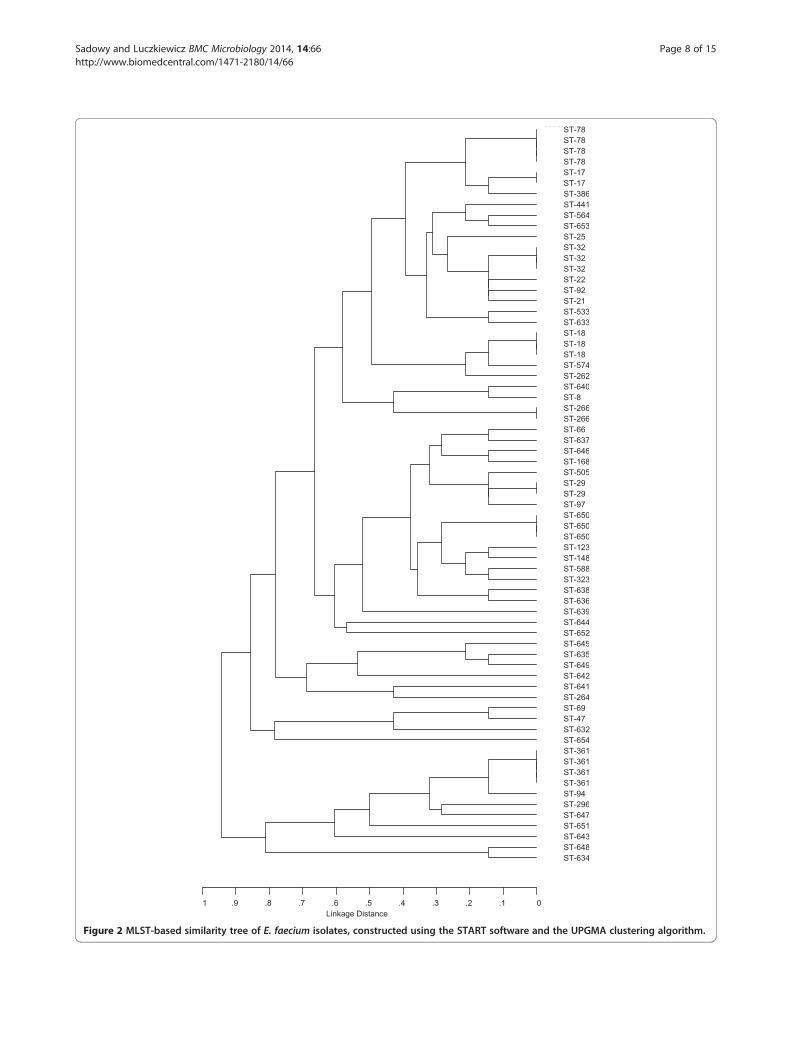
Figure 2 MLST-based similarity tree of E faecium isolates constructed using the START software and the UPGMA clustering algorithm
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 8 of 15httpwwwbiomedcentralcom1471-21801466
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 9 of 15httpwwwbiomedcentralcom1471-21801466
grouped the 38 STs specific for 50 isolates into the majorlsquosuper-complex 17rsquo characteristic for this species (data notshown) Further detailed analysis of the localization ofparticular STs within this structure revealed that 11 STscharacteristic for 17 isolates belonged to the nosocomialcomplex of E faecium representing all three majorlineages 17 18 and 78 of a former CC17 as defined byWillems et al [9] (Table 3) Seventeen STs (20 isolates)nine STs (11 isolates) and two STs with single isolates eachbelonged to CC5 CC22 and CC9 respectively as definedby Freitas et al [24] Of the remaining 16 STs nine wereincluded into six small CCs (CCs 27 46 94 296 639648) unlinked to the major lsquosuper-complexrsquo ST632formed a lsquodoubletonrsquo (ie was linked by a single-locus vari-ant link to only one ST from the database) and six STsrepresented by single isolates were singletons ie werenot related to any ST in the database at the time of ana-lysis Thirty isolates (435) in the investigated groupshowed the MDR phenotype which was significantly asso-ciated (p = 000006) with isolates from the nosocomialcomplex (Table 3 and Figure 3) but found also in CC5CC22 and CC94 this phenotype did not occur amongsmall CCs or singletons Seventeen isolates repre-senting the nosocomial complex were additionallystudied by multilocus VNTR (variable number tan-dem repeats) analysis (MLVA) to better elucidate therelationships among these isolates Altogether nineMLVA-types (MTs) were found (Table 3) Importantlythe same MLVA-type MT159 associated with ST78was characteristic for isolates derived from samplesfrom W-EFF and W-MOut suggesting survival of thisparticular clone
0
10
20
30
40
50
60
70
80
90
100
IS16
espintA
fms5
fms1
7
fms1
9
fms2
1
MDR
AM-R
CIP-R
HLG
R
p
ositi
ve
isol
ates
Figure 3 Distribution () of IS16 resistance phenotypes and virulenccolumns isolates with STs typical for nosocomial E faecium grey columnsgroups are indicated by an asterisk MDR multi-drug resistance AM-R ampgentamicin resistance HLSR high-level streptomycin resistance
IS16 was present in all 17 isolates included by MLST tothree lineages of nosocomial E faecium and this markerwas limited to this group (Table 3 and Figure 3) TheespEfm gene occurred always together with intA among allisolates of lineage 17 and in some isolates of lineage 78The fms19 pilin gene detected in 29 isolates was over-represented among STs typical for nosocomial E faeciumwhile other pilin genes such as fms5 fms17 and fms21detected in 57 49 and 46 isolates respectively occurredamong various CCs and singleton STs Ampicillin resist-ance significantly associated with isolates of nosocomialcomplex (10 of 17 isolates 588 p = 0001) occurred alsoin CC5 (6 isolates) CC22 and CC94 (1 isolate each) Se-quencing of the 3prime part of the pbp5 gene revealed its 11nucleotide alleles corresponding to 11 variants of the C-terminal part of PBP5 Six alleles of the gene were new(pbp5-24 to pbp5-29) two alleles (pbp5-20 pbp5-22)were described previously among Polish hospital VREmisolates [13] and three alleles were found in other stud-ies pbp5-30 occurred in a Portuguese animal strain ofE faecium [25] and pbp5-31 and pbp5-32 in clinical iso-lates TCGE70411_2 and 164306 also from Portugal (de-posited as JN208888 and JN208885 in the GenBankrespectively) For a single ampicillin-resistant isolate ofST653 (lineage 78) pbp5 amplification was not achieveddespite repeated attempts The majority (11 of 17 iso-lates 647) of isolates belonging to three lineages ofnosocomial E faecium showed resistance to ciprofloxa-cin determined by mutations R61G and S80IR in ParCand S84IRY in GyrA Ciprofloxacin resistance wasalso significantly over-represented among isolates ofSTs typical for nosocomial E faecium (p = 00006) Nine
HLS
R
tetM tetL
rep1
-pIP50
1
rep2
-pRE25
rep1
7-pR
UM
rep1
8-pE
F418
rep-pL
G1
axe-txe
relBE
e resistance and plasmid genes among E faecium isolates Blackother E faecium isolates Significant differences (p lt 005) between twoicillin resistance CIP-R ciprofloxacin resistance HLGR high-level
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 10 of 15httpwwwbiomedcentralcom1471-21801466
isolates resistant to ciprofloxacin were found also inCCs 5 9 and 22 These isolates possessed mutations inParC mentioned above but had the wild-type QRDR inGyrA HLGR occurred in five isolates of lineages 18 and78 and in single isolates of CC5 and CC22 this pheno-type was determined by the presence of aac(6prime)-Ie-aph(2Prime) with an exception of an isolate from CC5 whichwas negative for this gene High-level resistance tostreptomycin (HLSR) was widely distributed amongstudied isolates and determined by the ant(6prime)-Ia geneTetracycline resistance was found also in various STsthe major determinant of this phenotype was tet(M) (20isolates) followed by tet(L) (17 isolates) These twogenes occurred together in 16 isolates A single isolateof ST633 belonging to CC22 harbored tet(S) The tet(O) gene was absent in the studied group For a singletetracycline-resistant isolate of ST17 none of the fourtetracycline determinants studied was detected Nineisolates carried int of Tn916 and four the tndX gene ofTn5397 These transposon-specific genes were not char-acteristic for any lineage or complex (Table 3) Plasmid-replication genes such as rep2pRE25 rep17pRUM andrep18 pEF418 (26 14 and 7 isolates respectively) andgenes specifying toxin-antitoxin systems (TAS) axe-txeand ω-ε-ζ (12 and 10 isolates respectively) were signifi-cantly associated with the group of isolates of nosocomialE faecium Two other rep genes (rep1pIP501 reppLG1) werewidely distributed among all studied E faecium isolatesand the reppMG1 gene occurred in two isolates (in a singleisolate of ST533 from CC22 and in a singleton ST647) Allω-ε-ζ-positive isolates carried rep2pRE25 and nine isolatesshowed a concomitant presence of axe-txe and rep17pRUMThe relBE genes were found solely among five non-nosocomial E faecium (a difference in distribution with-out statistical significance p = 043) among various STs
DiscussionIn this study Enterococcus spp was used to evaluate theimpact posed by treated wastewater of two local WWTPson their receiver the costal waters of Gulf of GdanskTotal counts of enterococci observed for raw (W-INF)and treated (W-EFF D-EFF) wastewater are consistentwith our previous study showing similar over 99 reduc-tion capacity of applied treatment processes It is alsoreported in the literature that wastewater treatment pro-cesses based on activated sludge although they focus onremoval of nutrients and organic matter simultaneouslyreduce faecal indicators with high efficiency reaching upto 999 [1822] Due to the high number of enterococciin raw wastewater they are nevertheless released from aWWTP in a number up to 61x105 CFU per 100 mLThus marine water sampled in the area directly impactedby treated wastewater (W-MOut and D-MOut) showed ahigher number of enterococci than the one observed in
Vistula River mouth however lower by four orders ofmagnitude in comparison to corresponding treated waste-water (W-EFF and D-EFF) It should be noted that in mar-ine outfalls (W-MOut D-MOut) as well as in VistulaRiver mouth enterococci were detected in number lowerthan 100 CFU per 100 mL ie the value set out by NewBathing Water Directive 20067EC for the costal water ofldquoexcellent qualityrdquo [23]It is suggested that enterococcal species composition in
faecal-impacted environment depends from the source ofcontamination In this study two species most commonlycolonizing humans E faecalis and E faecium comprisedtogether from 73 to 88 of enterococcal isolates intested wastewater samples (W-INF W-BR W-EFF and D-EFF) 73 and 61 in marine outflows (W-MOut and D-MOut respectively) and 45 in the Vistula River mouthThe predominant species in wastewater and marine waterimpacted by treated wastewater was E faecium (from 56to 65) which stayed at a similar level during treatmentprocesses as determined also in our previous study [20]Species distribution in wastewater samples is not wellunderstood and affecting factors may include differencesin diet climate season and methods of detection E feaca-lis was found to be the most prevalent species in wastewa-ter samples in Sweden [26] E faecium in Spain UKCanada France and Switzerland [26-29] while E hiraepredominated in Portugal and United States [2230] Al-though E faecalis is typically the most abundant colonizerof humans in the community (eg [2631]) E faecium isgenerally most often associated with human microbialcontamination [2729] E faecalis was reported fromagriculture-impacted waters [32] and E hirae likelyoriginates mainly from cattle and pig faecal contamin-ation [26] which explains high relative proportion ofthese two species in Vistula River mouth Apart from thevariability of contamination sources proportion of par-ticular species may differ due to the differences of abilityto sustain environmental stresses In this study enterococ-cal composition might have been influenced by wastewa-ter treatment processes and marine water characteristicsThe effect of wastewater treatment and marine environ-
ment was seen on phenotypes of antimicrobial resistanceThe positive selection of bacteria with resistance patternshas been already suggested and noted in wastewater pro-cesses [192022] In this study the resistance rates to ampi-cillin ciprofloxacin levofloxacin and erythromycin notedfor E faecium from raw wastewater (W-INF) were lowerthan these observed for this species in the correspondingtreated wastewater (W-EFF) however not statistically sig-nificant (p gt 005) Moreover resistance rates to ampicillinciprofloxacin tetracycline and erythromycin observed inmarine outflow of WWTP Gdansk-Wschod (W-MOut)were higher than in treated wastewater of this plantSuch phenomenon was not observed for treated
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 11 of 15httpwwwbiomedcentralcom1471-21801466
wastewater of WWTP Gdynia-Debogorze (D-EFF) andits marine outfall (D-MOut) Resistance rates among Efaecium originating from D-MOut were in generalcomparable to these detected in the Vistula Rivermouth It should be noted that treated wastewater fromWWTP Gdansk-Wschod has been discharged via mar-ine outfall (W-MOut) for last 12 years while marineoutfall of WWTP Gdynia-Debogorze (D-MOut) hasbeen operating for last two years The observed differ-ences in resistance rate corresponding to the local im-pact caused by long and short-term operated marineoutfalls (W-MOut and D-MOut) need further attentionSignificant rates of antimicrobial resistance observed
for isolates of E faecalis from wastewater and marineriver waters impacted by wastewater are in agreementwith a recent study performed in six European countriesshowing that in Poland the rates of resistance for humanisolates of this species were usually among the highestboth in the community and hospitals [33] To our know-ledge no corresponding data are available for E faeciumcolonization in the community in Poland but a studyfocused on hospital VRE isolates showed their ubiquitousMDR phenotype and a high load of resistance genes [13]In our study no vancomycin resistance were detectedlikely due to relatively low (below 10) prevalence of VREin Polish hospitals [5] although such isolates are isolatedfrom the environment in other countries eg [34]This study shows that hospital clones of E faecium are
important carriers of resistance genes in wastewatersMost likely in a significant part they originated fromhospital wastewater treated together with communalwastewater In our estimation hospital wastewater con-stitutes approximately 02 of both WWTPsrsquo daily in-flow The release of hospital-associated clones of Efaecium to the environment was observed also by others[24283536] In our study isolates belonging to themajor nosocomial HiRECC (formerly named CC17)constituted altogether 246 of all isolates of E faeciumsubjected to typing and were present at all samplingsites These isolates apart from their inclusion into theHiRECC by MLST demonstrated several other pheno-typic and genotypic features typical for nosocomial Efaecium ie they were resistant to ciprofloxacin andampicillin in vast majority showed MDR phenotypeand carried several resistance determinants associatedboth with mutations of chromosomal genes (gyrA parCpbp5) and in the case of aminoglycoside and tetracyclineresistance with gene acquisition Plasmids likely played animportant role in the later process as some plasmid genessuch as rep2pRE25 rep17pRUM and rep18 pEF418 and TASgenes axe-txe and ω-ε-ζ were significantly more abundantin isolates associated with HiRECC than among theremaining isolates As yet the knowledge of distributionof plasmid-associated genes in populations of E faecium
especially in the context of their clonal compositionremains limited A study on 93 isolates of differentgeographical origins and from various sites showed anover-representation of rep17pRUM and reppMG1 amongnosocomial E faecium while in contrast to our observa-tions rep18 pEF418 was not detected [37] Moreover wedid not observe reppMG1 among HiRECC of E faeciumThese differences may be due to a high prevalence of VRE(58 isolates) among 93 isolates studied by Rosvoll et alwhat may explain differences in the plasmid content Thesame study also reported the co-localization of ω-ε-ζ withrep2pRE25 and axe-txe with rep17pRUM on the same plas-mids in concordance with our findings showing a fre-quent joint occurrence of these pairs of genes Anotherstudy performed on 99 invasive isolates from Norwaymostly of HLGR phenotype showed a significantly higherprevalence of rep2pRE25 and rep17pRUM among the majornosocomial HiRECC [38] This study as well some others[2439] also demonstrated a common presence of reppLG1among E faecium from hospital settings and in animalsOur findings indicate that this type of replicon is evenmore ubiquitous in E faecium than previously reportedAll rep genes mentioned above are typically associatedwith plasmids carrying various antimicrobial resistancegenes [3940] GenBank accession number AF408195 Inaddition to genes determining antimicrobial resistanceand genes associated with MGE isolates of nosocomialHiRECC analyzed in our study carried such molecularmarkers as IS16 and the esp gene together with the intAgene characteristic for ICEEfm [4142] Another patho-genicity factor the fms19 gene of the pilin gene cluster4 (PGC4) showed increased prevalence among theseisolates Similar observation was made for a representa-tive collection of 433 E faecium isolates from varioussources [43]In a study that followed enterococci in a continuum
from hospital and retirement home wastewaters througha WWTP to a river a gradual decrease in the proportionof E faecium isolates representing hospital-associatedSTs was observed likely due to the ldquodilutionrdquo of HiRECCrepresentatives by more diverse and less drug-resistantstrains originating from the community [28] Because inour study only selected representatives of obtained iso-lates were investigated it is not possible to determineprecise tendencies in their frequency from the WWTPinfluent to effluent and water however resistant entero-cocci both HiRECC and non-HiRECC are clearly ableto survive the treatment process what results in their re-lease into marine and river waters Moreover using anadditional typing method MLVA we were able to findisolates with the same characteristics in the effluent ofWWTP and its marine outfall suggesting ability of suchclones to survive in the environment for at least sometime It seems plausible that hospital resistant clones
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 12 of 15httpwwwbiomedcentralcom1471-21801466
accumulate several determinants that may promote theirsurvival not only in the hospital settings but also in theenvironment Such features would include resistance todisinfectants and ultraviolet light additional metabolicpathways and ability to form biofilms Moreover determi-nants of some of the mentioned adaptation factors arefound co-resident in plasmids harboring resistance genessuggesting co-selection For example pLG1-type plasmidsof E faecium carry not only resistance determinantsagainst aminoglycosides glycopeptides and macrolidesbut also pili genes and carbon uptake-utilization genes[3839] Pheromone-responsive plasmids of E faecalisapart from resistance to aminoglycosides glycopeptidespenicillins and macrolides may encode bacteriocins add-itional pili and UV-resistance determinants [40] Thusnosocomial HiRECCs released by WWTPs are equippedin whole sets of genes with various adaptive benefits andoften located within MGE It was also demonstrated thatMGEs can be transferred in a WWTP [44] further under-ling the importance of threat posed by such organismsThere is a selection pressure for survival of resistantstrains and their tranconjugants due to release of antibi-otics to the environment especially in the hospital efflu-ents [45] and even sub-inhibitory concentrations may beeffective in selection for resistance [46]
ConclusionsThis study describes the species composition resistanceprofiles and clonal relationships of enterococci isolatedfrom WWTPs located in Gdansk their marine outfallsto the Gulf of Gdansk and from Vistula River It was ob-served that these WWTPs release enterococcal strainsresistant to various antimicrobial compounds to the re-cipient water environment Molecular analyses revealedthe presence of clones associated with nosocomial HiR-ECCs and in the case of E faecium isolates of lineages17 18 and 78 of former CC17 abundant in resistance de-terminants MGE genes and pathogenicity factors Thepresence of HiRECCs in wastewaters and marineriverenvironment highlights the need for further detailedanalyses to better understand the survival and spread ofdrug-resistant strains in water ecosystems and to eluci-date the ways to curb such dissemination
MethodsAcquisition of samples and isolation of enterococciOnce a month in April June August and October in 2011of the flow proportioned composited samples of wastewa-ter were taken from two local WWTPs Gdansk-Wschodfrom influent (W-INF) effluent (W-EFF) and wastewa-ter from activated sludge bioreactor (W-BR) and fromGdynia-Debogorze effluent (D-EFF) The two WWTPstreat sewage from approximately 920 000 inhabitants(for details see Figure 1) In the same period of time and
with the same frequency marine outfalls of those WWTPs(W-MOut and D-MOut respectively) as well as VistulaRiver mouth were also sampled for isolation of entero-cocci Marine samples were taken at the depth of approxi-mately one meter below the surface once a month inApril June August and October of 2011 Location ofsampling points and their characteristics are provided inFigure 1 Enterococci were detected using the membranefiltration method To this end appropriate dilutions ofanalyzed samples were filtered in duplicate through045 μm cellulose-acetate filters (EMD Millipore Cor-poration Billerica MA USA) which were then placedon Enterococcus selective agar (Merck DarmstadtGermany) and incubated at 37degC for 48 h (ISO 7899ndash22000) Dark red or maroon colonies were consideredpresumptive enterococci For further investigationsrepresentative isolates were taken from membranes pre-senting from 20 to 50 typical colonies and stored in nu-trient broth supplemented with 15 glycerol at minus80degC
Species identification DNA isolation susceptibility testingand statistical analysisThe species identification and drug susceptibility of pre-sumptive enterococci were determined by the PhoenixtradeAutomated Microbiology System (BD New Jersey USA)according to the manufacturerrsquos instructions Since it isnot possible to distinguish E casseliflavus and E galli-narum using this system the classification E casselifla-vusgallinarum was used The susceptibility tests basedon microdilution were carried out for 11 antimicrobialagents gentamicin (GM) streptomycin (ST) ampicillin(AM) daptomycin (DAP) vancomycin (VA) teicoplanin(TEC) linezolid (LZD) fluoroquinolones (ciprofloxacin -CIP levofloxacin - LVX) tetracycline (TE) erythromycin(E) and quinupristin-dalfopristin (SYN) (E faecium only)Obtained minimal inhibitory concentrations (MIC) wereevaluated according to the guidelines of Clinical and La-boratory Standards Institute [47] Isolates were defined asmultidrug-resistant (MDR) when they showed resistanceto three or more compounds tested [48] Bacterial DNAwas purified using the Genomic DNA Prep Plus kitfollowing the manufacturerrsquos instructions (AampA Biotech-nology Gdynia Poland) Differences in distributions wereevaluated using the χ2 test with the p values lt 005 con-sidered significant
MLST and MLVA of E faeciumMLST for E faecium was performed by sequencing ofseven house-keeping genes as described by Homan et al[49] The allele numbers and STs were assigned with theuse of Internet database httpefaeciummlstnet (5th July2013 date last accessed) Novel alleles and allelic profileswere submitted to the database MLST data were analyzedwith Sequence Type Analysis and Recombinational Tests
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 13 of 15httpwwwbiomedcentralcom1471-21801466
(START) software [50] and the eBURST analysis [51] wasused to elucidate the relationships of STs with knownCCs using the comparative eBURST option available athttpeburstmlstnetv3enter_datacomparative (5th July2013 date last accessed) Further inclusion of E faeciumSTs into specific CCs and lineages was performed fol-lowing other studies [924] MLVA was performed as de-scribed by others [52]
Detection and analysis of genes associated with virulenceantimicrobial resistance and MGE in E faeciumThe full list of primers used for gene detection and se-quencing is provided in Additional file 1 Table S1 PCR-based detection of IS16 virulence-associated genes (fms21fms17 fms5 and fms19 representing each of four pilingene clusters in E faecium and the espEfm gene) genes de-termining resistance to tetracycline tet(M) tet(O) tet(L)tet(S) HLGR (aac(6prime)-Ie-aph(2Prime) HLSR (ant(6prime)-Ia) andgenes specific for MGE such as intA of ICEEfm1 int ofTn916 transposon tndX of Tn5397 transposon entero-coccal plasmid replication genes rep1pIP501 rep2pRE25rep17pRUM rep18pEF418 reppMG1 reppLG1 and TAS genesaxe-txe ω-ε-ζ relBE was performed as described in otherstudies [134153-59] following the conditions describedherein Previously characterized clinical isolates of Strepto-coccus agalactiae E faecium and E faecalis harbouringpilin genes IS16 espEfm intA of ICEEfm1 int of Tn916tndX of Tn5397 tet(M) tet(O) tet(L) tet(S) aac(6prime)-Ie-aph(2Prime) ant(6prime)-Ia rep1pIP501 rep2pRE25 ω-ε-ζ [136061]served as positive controls In cases when positive controlswere not available (rep18pEF418 reppMG1 reppLG1 axe-txerelBE) a few randomly selected PCR products wereverified by sequencing and served further as controlsAmpicillin-resistance determinant the pbp5 gene was ana-lysed by sequencing of the part encoding C-terminaltranspeptidase domain in PBP5 [62] New sequence vari-ants pbp5-24 to pbp5-29 alleles were submitted to theGenBank (acc numbers KC594860-KC594865 respect-ively) The quinolone-determining regions (QRDR) in thegyrA and parC genes were sequenced and analysed asdescribed by others [63]
Additional file
Additional file 1 Table S1 Primers used in the study for PCR andsequencing
Competing interestThe authors declare that they have no competing interest
Authorsrsquo contributionsES participated in the study design performed experimental work andparticipated in the manuscript preparation AL designed the studyperformed experimental work and participated in the manuscriptpreparation Both authors read and approved the final manuscript
AcknowledgementsWe thank Rob Willems and Janetta Top for assigning new alleles new STsand MTs This publication made use of the Enterococcus faecium MLSTwebsite (httpefaeciummlstnet) which is hosted at Imperial College of theUniversity of Oxford and funded by the Wellcome Trust This work wassupported by project N N305 461139 from the Narodowe Centrum Nauki(NCN)
Author details1National Medicines Institute ul Chelmska 3034 Warsaw 00-725 Poland2Gdansk University of Technology ul G Narutowicza 1112 Gdansk 80-952Poland
Received 5 September 2013 Accepted 27 February 2014Published 14 March 2014
References1 Aarestrup FM Butaye P Witte W Nonhuman reservoirs of Enterococci In
American Society for Microbiology Edited by Gilmore MS Clewell DB CourvalinP Dunny GM Murray BE Rice LB Washington DC The Enterococcipathogenesis molecular biology and antibiotic resistance 2002
2 Tendolkar PM Baghdayan AS Shankar N Pathogenic enterococci newdevelopments in the 21st century Cell Mol Life Sci 2003 602622ndash2636
3 Sood S Malhotra M Das BK Kapil A Enterococcal infections ampantimicrobial resistance Indian J Med Res 2008 128111ndash121
4 Sydnor ER Perl TM Hospital epidemiology and infection control inacute-care settings Clin Microbiol Rev 2011 24141ndash173
5 European Centre for Disease Prevention and Control Annual EpidemiologicalReport Reporting on 2010 surveillance data and 2011 epidemic intelligencedata Stockholm ECDC 20122013
6 Hidron AI Edwards JR Patel J Horan TC Sievert DM Pollock DA Fridkin SKNational Healthcare Safety Network Team Participating National HealthcareSafety Network Facilities NHSN annual update antimicrobial-resistantpathogens associated with healthcare-associated infections annualsummary of data reported to the National Healthcare Safety Network atthe Centers for Disease Control and Prevention 2006ndash2007 Infect ControlHosp Epidemiol 2008 29996ndash1011
7 De Kraker ME Jarlier V Monen JC Heuer OE van de Sande N GrundmannH The changing epidemiology of bacteraemias in Europe trends fromthe European Antimicrobial Resistance Surveillance System Clin MicrobiolInfect 2012 doi1011111469-0691
8 Leavis HL Bonten MJ Willems RJ Identification of high-risk enterococcalclonal complexes global dispersion and antibiotic resistance Curr OpinMicrobiol 2006 9454ndash460
9 Willems RJL Top J van Schaik W Leavis H Bonten M Sireacuten J Hanage WPCorander J Restricted gene flow among hospital subpopulations ofEnterococcus faecium mBio 2012 3e00151ndash12
10 Hegstad K Mikalsen T Coque TM Werner G Sundsfjord A Mobile geneticelements and their contribution to the emergence of antimicrobialresistant Enterococcus faecalis and Enterococcus faecium Clin MicrobiolInfect 2010 16541ndash554
11 Shepard BD Gilmore MS Antibiotic-resistant enterococci themechanisms and dynamics of drug introduction and resistance MicrobesInfect 2002 4215ndash224
12 Bonten MJ Willems R Weinstein RA Vancomycin-resistant enterococciwhy are they here and where do they come from Lancet Infect Dis 20011314ndash325
13 Sadowy E Sieńko A Gawryszewska I Bojarska A Malinowska K HryniewiczW High abundance and diversity of antimicrobial resistancedeterminants among early vancomycin-resistant Enterococcus faecium inPoland Eur J Clin Microbiol Infect Dis 2013 321193ndash1203
14 Willems RJ Homan W Top J van Santen-Verheuvel M Tribe D Manzioros XGaillard C Vandenbroucke-Grauls CM Mascini EM van Kregten E vanEmbden JD Bonten MJ Variant esp gene as a marker of a distinct geneticlineage of vancomycin-resistant Enterococcus faecium spreading inhospitals Lancet 2001 357853ndash855
15 Sillanpaumlauml J Nallapareddy SR Prakash VP Qin X Houmloumlk M Weinstock GMMurray BE Identification and phenotypic characterization of a secondcollagen adhesin Scm and genome-based identification and analysis of13 other predicted MSCRAMMs including four distinct pilus loci inEnterococcus faecium Microbiology 2008 1543199ndash3211
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 14 of 15httpwwwbiomedcentralcom1471-21801466
16 Galloway-Pentildea JR Nallapareddy SR Arias CA Eliopoulos GM Murray BEAnalysis of clonality and antibiotic resistance among early clinicalisolates of Enterococcus faecium in the United States J Infect Dis 20092001566ndash1573
17 Top J Willems R Bonten M Emergence of CC17 Enterococcus faeciumfrom commensal to hospital-adapted pathogen FEMS Immunol MedMicrobiol 2008 52297ndash308
18 Martins da Costa P Vaz-Pires P Bernardo F Antimicrobial resistance inEnterococcus spp isolated in inflow effluent and sludge from municipalsewage water treatment plants Water Res 2006 401735ndash1740
19 Reinthaler FF Posch J Feierl G Wust G Haas D Ruckenbauer G Mascher FMarth E Antibiotic resistance of Escherichia coli in sewage and sludgeWater Res 2003 371685ndash1690
20 Luczkiewicz A Fudala-Ksiazek S Jankowska K Olanczuk-Neyman KAntimicrobial resistance of fecal indicators in municipal wastewatertreatment plant Water Res 2010 445089ndash5097
21 Dulfour AP Ballentine RK Ambient water quality criteria for bacteria-1986 EPA4405-84-002 Washington DC US Environmental Protection Agency 1986
22 Ferreira da Silva M Tiago I Veriacutessimo A Boaventura RA Nunes OC Manaia CMAntimicrobial resistance patterns in Enterobacteriaceae isolated from anurban wastewater treatment plant FEMS Microbiol Ecol 2007 60166ndash176
23 DIRECTIVE 20067EC of the European Parliament and of the Council 15February 2006 concerning the management of bathing water quality andrepealing Directive 76160EEC httpeur-lexeuropaeuLexUriServLexUriServdouri=OJL200606400370051ENPDF
24 Freitas AR Novais C Ruiz-Garbajosa P Coque TM Peixe L Dispersion ofmultidrug-resistant Enterococcus faecium isolates belonging to majorclonal complexes in different Portuguese settings Appl Environ Microbiol2009 754904ndash4908
25 Poeta P Costa D Igrejas G Saacuteenz Y Zarazaga M Rodrigues J Torres CPolymorphisms of the pbp5 gene and correlation with ampicillinresistance in Enterococcus faecium isolates of animal origin J MedMicrobiol 2007 56236ndash240
26 Kuumlhn I Iversen A Burman LG Olsson-Liljequist B Franklin A Finn MAarestrup F Seyfarth AM Blanch AR Vilanova X Taylor H Caplin J MorenoMA Dominguez L Herrero IA Moumlllby R Comparison of enterococcalpopulations in animals humans and the environment ndash a Europeanstudy Int J Food Microbiol 2003 88133ndash145
27 Lanthier M Scott A Lapen DR Zhang Y Topp E Frequency of virulencegenes and antibiotic resistances in Enterococcus spp isolates fromwastewater and feces of domesticated mammals and birds and wildlifeCan J Microbiol 2010 56715ndash729
28 Leclercq R Oberleacute K Galopin S Cattoir V Budzinski H Petit F Changes inenterococcal populations and related antibiotic resistance along amedical center-wastewater treatment plant-river continuum Appl EnvironMicrobiol 2013 792428ndash2434
29 Thevenon F Regier N Benagli C Tonolla M Adatte T Wildi W Poteacute JCharacterization of fecal indicator bacteria in sediments cores from thelargest freshwater lake of Western Europe (Lake Geneva Switzerland)Ecotoxicol Environ Safety 2012 7850ndash56
30 Bonilla TD Nowosielski K Esiobu N McCorquodale DS Rogerson ASpecies assemblages of Enterococcus indicate potential sources offecal bacteria at a south Florida recreational beach Mar Pollut Bull2006 52807ndash810
31 Wang JT Chang SC Wang HY Chen PC Shiau YR Lauderdale TL TSARHospitals High rates of multidrug resistance in Enterococcus faecalis andE faecium isolated from inpatients and outpatients in TaiwanDiagn Microbiol Infect Dis 2013 75406ndash411
32 Lanthier M Scott A Zhang Y Cloutier M Durie D Henderson VC WilkesG Lapen DR Topp E Distribution of selected virulence genes andantibiotic resistance in Enterococcus species isolated from the SouthNation River drainage basin Ontario Canada J Appl Microbiol 2011110407ndash421
33 Kuch A Willems RJ Werner G Coque TM Hammerum AM Sundsfjord AKlare I Ruiz-Garbajosa P Simonsen GS van Luit-Asbroek M Hryniewicz WSadowy E Insight into antimicrobial susceptibility and population struc-ture of contemporary human Enterococcus faecalis isolates fromEurope J Antimicrob Chemother 2012 67551ndash558
34 Novais C Coque TM Ferreira H Sousa JC Peixe L Environmentalcontamination with vancomycin-resistant enterococci from hospitalsewage in Portugal Appl Environ Microbiol 2005 713364ndash3368
35 Iversen A Kuumlhn I Rahman M Franklin A Burman LG Olsson-Liljequist BTorell E Moumlllby R Evidence for transmission between humans and theenvironment of a nosocomial strain of Enterococcus faeciumEnviron Microbiol 2004 655ndash59
36 Rathnayake I Hargreaves M Huygens F SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterwayAustralia and associated antibiotic resistance gene profilesBMC Microbiol 2011 11201
37 Rosvoll TC Pedersen T Sletvold H Johnsen PJ Sollid JE Simonsen GSJensen LB Nielsen KM Sundsfjord A PCR-based plasmid typing inEnterococcus faecium strains reveals widely distributed pRE25-pRUM- pIP501- and pHT-related replicons associated with glycopeptideresistance and stabilizing toxin-antitoxin systems FEMS Immunol MedMicrobiol 2010 58254ndash268
38 Rosvoll TC Lindstad BL Lunde TM Hegstad K Aasnaes B Hammerum AMLester CH Simonsen GS Sundsfjord A Pedersen T Increased high-levelgentamicin resistance in invasive Enterococcus faecium is associated withaac(6prime)Ie-aph(2Prime)Ia-encoding transferable megaplasmids hosted by majorhospital-adapted lineages FEMS Immunol Med Microbiol 2012 66166ndash176
39 Laverde Gomez JA van Schaik W Freitas AR Coque TM Weaver KE FranciaMV Witte W Werner G A multiresistance megaplasmid pLG1 bearing ahylEfm genomic island in hospital Enterococcus faecium isolates Int J MedMicrobiol 2011 301165ndash175
40 Coque TM Evolutionary biology of pathogenic enterococci InEvolutionary Biology of Bacterial and Fungal Pathogens Coque TM Edited byBaquero F Nombela C Cassell GH Gutierrez JA Washington DC ASMPress501ndash523
41 Werner G Fleige C Geringer U van Schaik W Klare I Witte W IS elementIS16 as a molecular screening tool to identify hospital-associated strainsof Enterococcus faecium BMC Infect Dis 2011 1180
42 Top J Sinnige JC Majoor EA Bonten MJ Willems RJ van Schaik W Therecombinase IntA is required for excision of esp-containing ICEEfm1 inEnterococcus faecium J Bacteriol 2011 1931003ndash1006
43 Sillanpaumlauml J Prakash VP Nallapareddy SR Murray BE Distribution of genesencoding MSCRAMMs and Pili in clinical and natural populations ofEnterococcus faecium J Clin Microbiol 2009 47896ndash901
44 Marcinek H Wirth R Muscholl-Silberhorn A Gauer M Enterococcus faecalisgene transfer under natural conditions in municipal sewage watertreatment plants Appl Environ Microbiol 1998 64626ndash632
45 Kuumlmmerer K Significance of antibiotics in the environment J AntimicrobChemother 2003 525ndash7
46 Liu A Fong A Becket E Yuan J Tamae C Medrano L Maiz M Wahba C LeeC Lee K Tran KP Yang H Hoffman RM Salih A Miller JHP Selectiveadvantage of resistant strains at trace levels of antibiotics a simple andultrasensitive color test for detection of antibiotics and genotoxicagents Antimicrob Agents Chemother 2011 551204ndash1210
47 Clinical and Laboratory Standards Institute Performance standards forantimicrobial susceptibility testing 21st informational supplement Wayne CLSI2011 PA M100-S20
48 Magiorakos AP Srinivasan A Carey RB Carmeli Y Falagas ME Giske CGHarbarth S Hindler JF Kahlmeter G Olsson-Liljequist B Paterson DL Rice LBStelling J Struelens MJ Vatopoulos A Weber JT Monnet DLMultidrug-resistant extensively drug-resistant and pandrug-resistantbacteria an international expert proposal for interim standarddefinitions for acquired resistance Clin Microbiol Infect 2012 18268ndash281
49 Homan WL Tribe D Poznanski S Li M Hogg G Spalburg E Van Embden JDWillems RJ Multilocus sequence typing scheme for Enterococcus faeciumJ Clin Microbiol 2002 401963ndash1971
50 Jolley KA Feil EJ Chan MS Maiden MC Sequence type analysis andrecombinational tests (START) Bioinformatics 2001 171230ndash1231
51 Feil EJ Li BC Aanensen DM Hanage WP Spratt BG eBURST inferringpatterns of evolutionary descent among clusters of related bacterialgenotypes from multilocus sequence typing data J Bacteriol 20041861518ndash1530
52 Top J Banga1 NMI Hayes R Willems RJ Bonten MJM Hayden MKComparison of multiple-locus variable-number tandem repeat analysisand pulsed-field gel electrophoresis in a setting of polyclonal endemicityof vancomycin-resistant Enterococcus faecium Clin Microbiol Infect 200814363ndash369
53 Vankerckhoven V Van Autgaerden T Vael C Lammens C Chapelle S RossiR Jabes D Goossens H Development of a multiplex PCR for the
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-
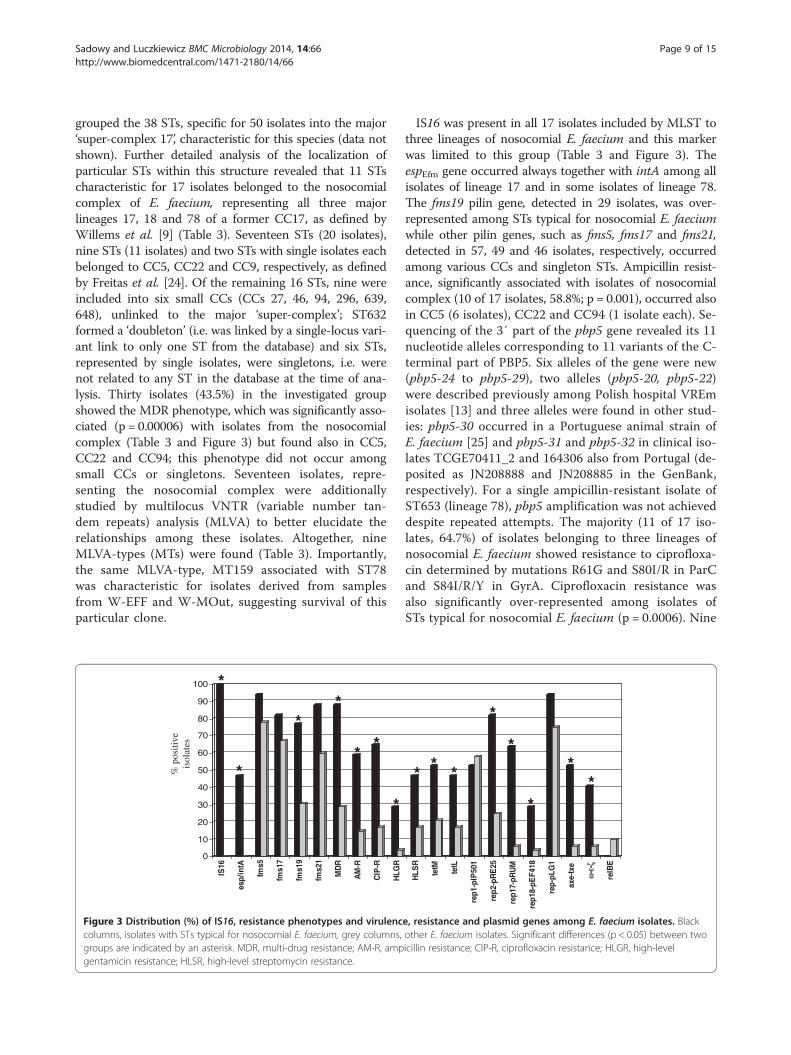
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 9 of 15httpwwwbiomedcentralcom1471-21801466
grouped the 38 STs specific for 50 isolates into the majorlsquosuper-complex 17rsquo characteristic for this species (data notshown) Further detailed analysis of the localization ofparticular STs within this structure revealed that 11 STscharacteristic for 17 isolates belonged to the nosocomialcomplex of E faecium representing all three majorlineages 17 18 and 78 of a former CC17 as defined byWillems et al [9] (Table 3) Seventeen STs (20 isolates)nine STs (11 isolates) and two STs with single isolates eachbelonged to CC5 CC22 and CC9 respectively as definedby Freitas et al [24] Of the remaining 16 STs nine wereincluded into six small CCs (CCs 27 46 94 296 639648) unlinked to the major lsquosuper-complexrsquo ST632formed a lsquodoubletonrsquo (ie was linked by a single-locus vari-ant link to only one ST from the database) and six STsrepresented by single isolates were singletons ie werenot related to any ST in the database at the time of ana-lysis Thirty isolates (435) in the investigated groupshowed the MDR phenotype which was significantly asso-ciated (p = 000006) with isolates from the nosocomialcomplex (Table 3 and Figure 3) but found also in CC5CC22 and CC94 this phenotype did not occur amongsmall CCs or singletons Seventeen isolates repre-senting the nosocomial complex were additionallystudied by multilocus VNTR (variable number tan-dem repeats) analysis (MLVA) to better elucidate therelationships among these isolates Altogether nineMLVA-types (MTs) were found (Table 3) Importantlythe same MLVA-type MT159 associated with ST78was characteristic for isolates derived from samplesfrom W-EFF and W-MOut suggesting survival of thisparticular clone
0
10
20
30
40
50
60
70
80
90
100
IS16
espintA
fms5
fms1
7
fms1
9
fms2
1
MDR
AM-R
CIP-R
HLG
R
p
ositi
ve
isol
ates
Figure 3 Distribution () of IS16 resistance phenotypes and virulenccolumns isolates with STs typical for nosocomial E faecium grey columnsgroups are indicated by an asterisk MDR multi-drug resistance AM-R ampgentamicin resistance HLSR high-level streptomycin resistance
IS16 was present in all 17 isolates included by MLST tothree lineages of nosocomial E faecium and this markerwas limited to this group (Table 3 and Figure 3) TheespEfm gene occurred always together with intA among allisolates of lineage 17 and in some isolates of lineage 78The fms19 pilin gene detected in 29 isolates was over-represented among STs typical for nosocomial E faeciumwhile other pilin genes such as fms5 fms17 and fms21detected in 57 49 and 46 isolates respectively occurredamong various CCs and singleton STs Ampicillin resist-ance significantly associated with isolates of nosocomialcomplex (10 of 17 isolates 588 p = 0001) occurred alsoin CC5 (6 isolates) CC22 and CC94 (1 isolate each) Se-quencing of the 3prime part of the pbp5 gene revealed its 11nucleotide alleles corresponding to 11 variants of the C-terminal part of PBP5 Six alleles of the gene were new(pbp5-24 to pbp5-29) two alleles (pbp5-20 pbp5-22)were described previously among Polish hospital VREmisolates [13] and three alleles were found in other stud-ies pbp5-30 occurred in a Portuguese animal strain ofE faecium [25] and pbp5-31 and pbp5-32 in clinical iso-lates TCGE70411_2 and 164306 also from Portugal (de-posited as JN208888 and JN208885 in the GenBankrespectively) For a single ampicillin-resistant isolate ofST653 (lineage 78) pbp5 amplification was not achieveddespite repeated attempts The majority (11 of 17 iso-lates 647) of isolates belonging to three lineages ofnosocomial E faecium showed resistance to ciprofloxa-cin determined by mutations R61G and S80IR in ParCand S84IRY in GyrA Ciprofloxacin resistance wasalso significantly over-represented among isolates ofSTs typical for nosocomial E faecium (p = 00006) Nine
HLS
R
tetM tetL
rep1
-pIP50
1
rep2
-pRE25
rep1
7-pR
UM
rep1
8-pE
F418
rep-pL
G1
axe-txe
relBE
e resistance and plasmid genes among E faecium isolates Blackother E faecium isolates Significant differences (p lt 005) between twoicillin resistance CIP-R ciprofloxacin resistance HLGR high-level
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 10 of 15httpwwwbiomedcentralcom1471-21801466
isolates resistant to ciprofloxacin were found also inCCs 5 9 and 22 These isolates possessed mutations inParC mentioned above but had the wild-type QRDR inGyrA HLGR occurred in five isolates of lineages 18 and78 and in single isolates of CC5 and CC22 this pheno-type was determined by the presence of aac(6prime)-Ie-aph(2Prime) with an exception of an isolate from CC5 whichwas negative for this gene High-level resistance tostreptomycin (HLSR) was widely distributed amongstudied isolates and determined by the ant(6prime)-Ia geneTetracycline resistance was found also in various STsthe major determinant of this phenotype was tet(M) (20isolates) followed by tet(L) (17 isolates) These twogenes occurred together in 16 isolates A single isolateof ST633 belonging to CC22 harbored tet(S) The tet(O) gene was absent in the studied group For a singletetracycline-resistant isolate of ST17 none of the fourtetracycline determinants studied was detected Nineisolates carried int of Tn916 and four the tndX gene ofTn5397 These transposon-specific genes were not char-acteristic for any lineage or complex (Table 3) Plasmid-replication genes such as rep2pRE25 rep17pRUM andrep18 pEF418 (26 14 and 7 isolates respectively) andgenes specifying toxin-antitoxin systems (TAS) axe-txeand ω-ε-ζ (12 and 10 isolates respectively) were signifi-cantly associated with the group of isolates of nosocomialE faecium Two other rep genes (rep1pIP501 reppLG1) werewidely distributed among all studied E faecium isolatesand the reppMG1 gene occurred in two isolates (in a singleisolate of ST533 from CC22 and in a singleton ST647) Allω-ε-ζ-positive isolates carried rep2pRE25 and nine isolatesshowed a concomitant presence of axe-txe and rep17pRUMThe relBE genes were found solely among five non-nosocomial E faecium (a difference in distribution with-out statistical significance p = 043) among various STs
DiscussionIn this study Enterococcus spp was used to evaluate theimpact posed by treated wastewater of two local WWTPson their receiver the costal waters of Gulf of GdanskTotal counts of enterococci observed for raw (W-INF)and treated (W-EFF D-EFF) wastewater are consistentwith our previous study showing similar over 99 reduc-tion capacity of applied treatment processes It is alsoreported in the literature that wastewater treatment pro-cesses based on activated sludge although they focus onremoval of nutrients and organic matter simultaneouslyreduce faecal indicators with high efficiency reaching upto 999 [1822] Due to the high number of enterococciin raw wastewater they are nevertheless released from aWWTP in a number up to 61x105 CFU per 100 mLThus marine water sampled in the area directly impactedby treated wastewater (W-MOut and D-MOut) showed ahigher number of enterococci than the one observed in
Vistula River mouth however lower by four orders ofmagnitude in comparison to corresponding treated waste-water (W-EFF and D-EFF) It should be noted that in mar-ine outfalls (W-MOut D-MOut) as well as in VistulaRiver mouth enterococci were detected in number lowerthan 100 CFU per 100 mL ie the value set out by NewBathing Water Directive 20067EC for the costal water ofldquoexcellent qualityrdquo [23]It is suggested that enterococcal species composition in
faecal-impacted environment depends from the source ofcontamination In this study two species most commonlycolonizing humans E faecalis and E faecium comprisedtogether from 73 to 88 of enterococcal isolates intested wastewater samples (W-INF W-BR W-EFF and D-EFF) 73 and 61 in marine outflows (W-MOut and D-MOut respectively) and 45 in the Vistula River mouthThe predominant species in wastewater and marine waterimpacted by treated wastewater was E faecium (from 56to 65) which stayed at a similar level during treatmentprocesses as determined also in our previous study [20]Species distribution in wastewater samples is not wellunderstood and affecting factors may include differencesin diet climate season and methods of detection E feaca-lis was found to be the most prevalent species in wastewa-ter samples in Sweden [26] E faecium in Spain UKCanada France and Switzerland [26-29] while E hiraepredominated in Portugal and United States [2230] Al-though E faecalis is typically the most abundant colonizerof humans in the community (eg [2631]) E faecium isgenerally most often associated with human microbialcontamination [2729] E faecalis was reported fromagriculture-impacted waters [32] and E hirae likelyoriginates mainly from cattle and pig faecal contamin-ation [26] which explains high relative proportion ofthese two species in Vistula River mouth Apart from thevariability of contamination sources proportion of par-ticular species may differ due to the differences of abilityto sustain environmental stresses In this study enterococ-cal composition might have been influenced by wastewa-ter treatment processes and marine water characteristicsThe effect of wastewater treatment and marine environ-
ment was seen on phenotypes of antimicrobial resistanceThe positive selection of bacteria with resistance patternshas been already suggested and noted in wastewater pro-cesses [192022] In this study the resistance rates to ampi-cillin ciprofloxacin levofloxacin and erythromycin notedfor E faecium from raw wastewater (W-INF) were lowerthan these observed for this species in the correspondingtreated wastewater (W-EFF) however not statistically sig-nificant (p gt 005) Moreover resistance rates to ampicillinciprofloxacin tetracycline and erythromycin observed inmarine outflow of WWTP Gdansk-Wschod (W-MOut)were higher than in treated wastewater of this plantSuch phenomenon was not observed for treated
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 11 of 15httpwwwbiomedcentralcom1471-21801466
wastewater of WWTP Gdynia-Debogorze (D-EFF) andits marine outfall (D-MOut) Resistance rates among Efaecium originating from D-MOut were in generalcomparable to these detected in the Vistula Rivermouth It should be noted that treated wastewater fromWWTP Gdansk-Wschod has been discharged via mar-ine outfall (W-MOut) for last 12 years while marineoutfall of WWTP Gdynia-Debogorze (D-MOut) hasbeen operating for last two years The observed differ-ences in resistance rate corresponding to the local im-pact caused by long and short-term operated marineoutfalls (W-MOut and D-MOut) need further attentionSignificant rates of antimicrobial resistance observed
for isolates of E faecalis from wastewater and marineriver waters impacted by wastewater are in agreementwith a recent study performed in six European countriesshowing that in Poland the rates of resistance for humanisolates of this species were usually among the highestboth in the community and hospitals [33] To our know-ledge no corresponding data are available for E faeciumcolonization in the community in Poland but a studyfocused on hospital VRE isolates showed their ubiquitousMDR phenotype and a high load of resistance genes [13]In our study no vancomycin resistance were detectedlikely due to relatively low (below 10) prevalence of VREin Polish hospitals [5] although such isolates are isolatedfrom the environment in other countries eg [34]This study shows that hospital clones of E faecium are
important carriers of resistance genes in wastewatersMost likely in a significant part they originated fromhospital wastewater treated together with communalwastewater In our estimation hospital wastewater con-stitutes approximately 02 of both WWTPsrsquo daily in-flow The release of hospital-associated clones of Efaecium to the environment was observed also by others[24283536] In our study isolates belonging to themajor nosocomial HiRECC (formerly named CC17)constituted altogether 246 of all isolates of E faeciumsubjected to typing and were present at all samplingsites These isolates apart from their inclusion into theHiRECC by MLST demonstrated several other pheno-typic and genotypic features typical for nosocomial Efaecium ie they were resistant to ciprofloxacin andampicillin in vast majority showed MDR phenotypeand carried several resistance determinants associatedboth with mutations of chromosomal genes (gyrA parCpbp5) and in the case of aminoglycoside and tetracyclineresistance with gene acquisition Plasmids likely played animportant role in the later process as some plasmid genessuch as rep2pRE25 rep17pRUM and rep18 pEF418 and TASgenes axe-txe and ω-ε-ζ were significantly more abundantin isolates associated with HiRECC than among theremaining isolates As yet the knowledge of distributionof plasmid-associated genes in populations of E faecium
especially in the context of their clonal compositionremains limited A study on 93 isolates of differentgeographical origins and from various sites showed anover-representation of rep17pRUM and reppMG1 amongnosocomial E faecium while in contrast to our observa-tions rep18 pEF418 was not detected [37] Moreover wedid not observe reppMG1 among HiRECC of E faeciumThese differences may be due to a high prevalence of VRE(58 isolates) among 93 isolates studied by Rosvoll et alwhat may explain differences in the plasmid content Thesame study also reported the co-localization of ω-ε-ζ withrep2pRE25 and axe-txe with rep17pRUM on the same plas-mids in concordance with our findings showing a fre-quent joint occurrence of these pairs of genes Anotherstudy performed on 99 invasive isolates from Norwaymostly of HLGR phenotype showed a significantly higherprevalence of rep2pRE25 and rep17pRUM among the majornosocomial HiRECC [38] This study as well some others[2439] also demonstrated a common presence of reppLG1among E faecium from hospital settings and in animalsOur findings indicate that this type of replicon is evenmore ubiquitous in E faecium than previously reportedAll rep genes mentioned above are typically associatedwith plasmids carrying various antimicrobial resistancegenes [3940] GenBank accession number AF408195 Inaddition to genes determining antimicrobial resistanceand genes associated with MGE isolates of nosocomialHiRECC analyzed in our study carried such molecularmarkers as IS16 and the esp gene together with the intAgene characteristic for ICEEfm [4142] Another patho-genicity factor the fms19 gene of the pilin gene cluster4 (PGC4) showed increased prevalence among theseisolates Similar observation was made for a representa-tive collection of 433 E faecium isolates from varioussources [43]In a study that followed enterococci in a continuum
from hospital and retirement home wastewaters througha WWTP to a river a gradual decrease in the proportionof E faecium isolates representing hospital-associatedSTs was observed likely due to the ldquodilutionrdquo of HiRECCrepresentatives by more diverse and less drug-resistantstrains originating from the community [28] Because inour study only selected representatives of obtained iso-lates were investigated it is not possible to determineprecise tendencies in their frequency from the WWTPinfluent to effluent and water however resistant entero-cocci both HiRECC and non-HiRECC are clearly ableto survive the treatment process what results in their re-lease into marine and river waters Moreover using anadditional typing method MLVA we were able to findisolates with the same characteristics in the effluent ofWWTP and its marine outfall suggesting ability of suchclones to survive in the environment for at least sometime It seems plausible that hospital resistant clones
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 12 of 15httpwwwbiomedcentralcom1471-21801466
accumulate several determinants that may promote theirsurvival not only in the hospital settings but also in theenvironment Such features would include resistance todisinfectants and ultraviolet light additional metabolicpathways and ability to form biofilms Moreover determi-nants of some of the mentioned adaptation factors arefound co-resident in plasmids harboring resistance genessuggesting co-selection For example pLG1-type plasmidsof E faecium carry not only resistance determinantsagainst aminoglycosides glycopeptides and macrolidesbut also pili genes and carbon uptake-utilization genes[3839] Pheromone-responsive plasmids of E faecalisapart from resistance to aminoglycosides glycopeptidespenicillins and macrolides may encode bacteriocins add-itional pili and UV-resistance determinants [40] Thusnosocomial HiRECCs released by WWTPs are equippedin whole sets of genes with various adaptive benefits andoften located within MGE It was also demonstrated thatMGEs can be transferred in a WWTP [44] further under-ling the importance of threat posed by such organismsThere is a selection pressure for survival of resistantstrains and their tranconjugants due to release of antibi-otics to the environment especially in the hospital efflu-ents [45] and even sub-inhibitory concentrations may beeffective in selection for resistance [46]
ConclusionsThis study describes the species composition resistanceprofiles and clonal relationships of enterococci isolatedfrom WWTPs located in Gdansk their marine outfallsto the Gulf of Gdansk and from Vistula River It was ob-served that these WWTPs release enterococcal strainsresistant to various antimicrobial compounds to the re-cipient water environment Molecular analyses revealedthe presence of clones associated with nosocomial HiR-ECCs and in the case of E faecium isolates of lineages17 18 and 78 of former CC17 abundant in resistance de-terminants MGE genes and pathogenicity factors Thepresence of HiRECCs in wastewaters and marineriverenvironment highlights the need for further detailedanalyses to better understand the survival and spread ofdrug-resistant strains in water ecosystems and to eluci-date the ways to curb such dissemination
MethodsAcquisition of samples and isolation of enterococciOnce a month in April June August and October in 2011of the flow proportioned composited samples of wastewa-ter were taken from two local WWTPs Gdansk-Wschodfrom influent (W-INF) effluent (W-EFF) and wastewa-ter from activated sludge bioreactor (W-BR) and fromGdynia-Debogorze effluent (D-EFF) The two WWTPstreat sewage from approximately 920 000 inhabitants(for details see Figure 1) In the same period of time and
with the same frequency marine outfalls of those WWTPs(W-MOut and D-MOut respectively) as well as VistulaRiver mouth were also sampled for isolation of entero-cocci Marine samples were taken at the depth of approxi-mately one meter below the surface once a month inApril June August and October of 2011 Location ofsampling points and their characteristics are provided inFigure 1 Enterococci were detected using the membranefiltration method To this end appropriate dilutions ofanalyzed samples were filtered in duplicate through045 μm cellulose-acetate filters (EMD Millipore Cor-poration Billerica MA USA) which were then placedon Enterococcus selective agar (Merck DarmstadtGermany) and incubated at 37degC for 48 h (ISO 7899ndash22000) Dark red or maroon colonies were consideredpresumptive enterococci For further investigationsrepresentative isolates were taken from membranes pre-senting from 20 to 50 typical colonies and stored in nu-trient broth supplemented with 15 glycerol at minus80degC
Species identification DNA isolation susceptibility testingand statistical analysisThe species identification and drug susceptibility of pre-sumptive enterococci were determined by the PhoenixtradeAutomated Microbiology System (BD New Jersey USA)according to the manufacturerrsquos instructions Since it isnot possible to distinguish E casseliflavus and E galli-narum using this system the classification E casselifla-vusgallinarum was used The susceptibility tests basedon microdilution were carried out for 11 antimicrobialagents gentamicin (GM) streptomycin (ST) ampicillin(AM) daptomycin (DAP) vancomycin (VA) teicoplanin(TEC) linezolid (LZD) fluoroquinolones (ciprofloxacin -CIP levofloxacin - LVX) tetracycline (TE) erythromycin(E) and quinupristin-dalfopristin (SYN) (E faecium only)Obtained minimal inhibitory concentrations (MIC) wereevaluated according to the guidelines of Clinical and La-boratory Standards Institute [47] Isolates were defined asmultidrug-resistant (MDR) when they showed resistanceto three or more compounds tested [48] Bacterial DNAwas purified using the Genomic DNA Prep Plus kitfollowing the manufacturerrsquos instructions (AampA Biotech-nology Gdynia Poland) Differences in distributions wereevaluated using the χ2 test with the p values lt 005 con-sidered significant
MLST and MLVA of E faeciumMLST for E faecium was performed by sequencing ofseven house-keeping genes as described by Homan et al[49] The allele numbers and STs were assigned with theuse of Internet database httpefaeciummlstnet (5th July2013 date last accessed) Novel alleles and allelic profileswere submitted to the database MLST data were analyzedwith Sequence Type Analysis and Recombinational Tests
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 13 of 15httpwwwbiomedcentralcom1471-21801466
(START) software [50] and the eBURST analysis [51] wasused to elucidate the relationships of STs with knownCCs using the comparative eBURST option available athttpeburstmlstnetv3enter_datacomparative (5th July2013 date last accessed) Further inclusion of E faeciumSTs into specific CCs and lineages was performed fol-lowing other studies [924] MLVA was performed as de-scribed by others [52]
Detection and analysis of genes associated with virulenceantimicrobial resistance and MGE in E faeciumThe full list of primers used for gene detection and se-quencing is provided in Additional file 1 Table S1 PCR-based detection of IS16 virulence-associated genes (fms21fms17 fms5 and fms19 representing each of four pilingene clusters in E faecium and the espEfm gene) genes de-termining resistance to tetracycline tet(M) tet(O) tet(L)tet(S) HLGR (aac(6prime)-Ie-aph(2Prime) HLSR (ant(6prime)-Ia) andgenes specific for MGE such as intA of ICEEfm1 int ofTn916 transposon tndX of Tn5397 transposon entero-coccal plasmid replication genes rep1pIP501 rep2pRE25rep17pRUM rep18pEF418 reppMG1 reppLG1 and TAS genesaxe-txe ω-ε-ζ relBE was performed as described in otherstudies [134153-59] following the conditions describedherein Previously characterized clinical isolates of Strepto-coccus agalactiae E faecium and E faecalis harbouringpilin genes IS16 espEfm intA of ICEEfm1 int of Tn916tndX of Tn5397 tet(M) tet(O) tet(L) tet(S) aac(6prime)-Ie-aph(2Prime) ant(6prime)-Ia rep1pIP501 rep2pRE25 ω-ε-ζ [136061]served as positive controls In cases when positive controlswere not available (rep18pEF418 reppMG1 reppLG1 axe-txerelBE) a few randomly selected PCR products wereverified by sequencing and served further as controlsAmpicillin-resistance determinant the pbp5 gene was ana-lysed by sequencing of the part encoding C-terminaltranspeptidase domain in PBP5 [62] New sequence vari-ants pbp5-24 to pbp5-29 alleles were submitted to theGenBank (acc numbers KC594860-KC594865 respect-ively) The quinolone-determining regions (QRDR) in thegyrA and parC genes were sequenced and analysed asdescribed by others [63]
Additional file
Additional file 1 Table S1 Primers used in the study for PCR andsequencing
Competing interestThe authors declare that they have no competing interest
Authorsrsquo contributionsES participated in the study design performed experimental work andparticipated in the manuscript preparation AL designed the studyperformed experimental work and participated in the manuscriptpreparation Both authors read and approved the final manuscript
AcknowledgementsWe thank Rob Willems and Janetta Top for assigning new alleles new STsand MTs This publication made use of the Enterococcus faecium MLSTwebsite (httpefaeciummlstnet) which is hosted at Imperial College of theUniversity of Oxford and funded by the Wellcome Trust This work wassupported by project N N305 461139 from the Narodowe Centrum Nauki(NCN)
Author details1National Medicines Institute ul Chelmska 3034 Warsaw 00-725 Poland2Gdansk University of Technology ul G Narutowicza 1112 Gdansk 80-952Poland
Received 5 September 2013 Accepted 27 February 2014Published 14 March 2014
References1 Aarestrup FM Butaye P Witte W Nonhuman reservoirs of Enterococci In
American Society for Microbiology Edited by Gilmore MS Clewell DB CourvalinP Dunny GM Murray BE Rice LB Washington DC The Enterococcipathogenesis molecular biology and antibiotic resistance 2002
2 Tendolkar PM Baghdayan AS Shankar N Pathogenic enterococci newdevelopments in the 21st century Cell Mol Life Sci 2003 602622ndash2636
3 Sood S Malhotra M Das BK Kapil A Enterococcal infections ampantimicrobial resistance Indian J Med Res 2008 128111ndash121
4 Sydnor ER Perl TM Hospital epidemiology and infection control inacute-care settings Clin Microbiol Rev 2011 24141ndash173
5 European Centre for Disease Prevention and Control Annual EpidemiologicalReport Reporting on 2010 surveillance data and 2011 epidemic intelligencedata Stockholm ECDC 20122013
6 Hidron AI Edwards JR Patel J Horan TC Sievert DM Pollock DA Fridkin SKNational Healthcare Safety Network Team Participating National HealthcareSafety Network Facilities NHSN annual update antimicrobial-resistantpathogens associated with healthcare-associated infections annualsummary of data reported to the National Healthcare Safety Network atthe Centers for Disease Control and Prevention 2006ndash2007 Infect ControlHosp Epidemiol 2008 29996ndash1011
7 De Kraker ME Jarlier V Monen JC Heuer OE van de Sande N GrundmannH The changing epidemiology of bacteraemias in Europe trends fromthe European Antimicrobial Resistance Surveillance System Clin MicrobiolInfect 2012 doi1011111469-0691
8 Leavis HL Bonten MJ Willems RJ Identification of high-risk enterococcalclonal complexes global dispersion and antibiotic resistance Curr OpinMicrobiol 2006 9454ndash460
9 Willems RJL Top J van Schaik W Leavis H Bonten M Sireacuten J Hanage WPCorander J Restricted gene flow among hospital subpopulations ofEnterococcus faecium mBio 2012 3e00151ndash12
10 Hegstad K Mikalsen T Coque TM Werner G Sundsfjord A Mobile geneticelements and their contribution to the emergence of antimicrobialresistant Enterococcus faecalis and Enterococcus faecium Clin MicrobiolInfect 2010 16541ndash554
11 Shepard BD Gilmore MS Antibiotic-resistant enterococci themechanisms and dynamics of drug introduction and resistance MicrobesInfect 2002 4215ndash224
12 Bonten MJ Willems R Weinstein RA Vancomycin-resistant enterococciwhy are they here and where do they come from Lancet Infect Dis 20011314ndash325
13 Sadowy E Sieńko A Gawryszewska I Bojarska A Malinowska K HryniewiczW High abundance and diversity of antimicrobial resistancedeterminants among early vancomycin-resistant Enterococcus faecium inPoland Eur J Clin Microbiol Infect Dis 2013 321193ndash1203
14 Willems RJ Homan W Top J van Santen-Verheuvel M Tribe D Manzioros XGaillard C Vandenbroucke-Grauls CM Mascini EM van Kregten E vanEmbden JD Bonten MJ Variant esp gene as a marker of a distinct geneticlineage of vancomycin-resistant Enterococcus faecium spreading inhospitals Lancet 2001 357853ndash855
15 Sillanpaumlauml J Nallapareddy SR Prakash VP Qin X Houmloumlk M Weinstock GMMurray BE Identification and phenotypic characterization of a secondcollagen adhesin Scm and genome-based identification and analysis of13 other predicted MSCRAMMs including four distinct pilus loci inEnterococcus faecium Microbiology 2008 1543199ndash3211
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 14 of 15httpwwwbiomedcentralcom1471-21801466
16 Galloway-Pentildea JR Nallapareddy SR Arias CA Eliopoulos GM Murray BEAnalysis of clonality and antibiotic resistance among early clinicalisolates of Enterococcus faecium in the United States J Infect Dis 20092001566ndash1573
17 Top J Willems R Bonten M Emergence of CC17 Enterococcus faeciumfrom commensal to hospital-adapted pathogen FEMS Immunol MedMicrobiol 2008 52297ndash308
18 Martins da Costa P Vaz-Pires P Bernardo F Antimicrobial resistance inEnterococcus spp isolated in inflow effluent and sludge from municipalsewage water treatment plants Water Res 2006 401735ndash1740
19 Reinthaler FF Posch J Feierl G Wust G Haas D Ruckenbauer G Mascher FMarth E Antibiotic resistance of Escherichia coli in sewage and sludgeWater Res 2003 371685ndash1690
20 Luczkiewicz A Fudala-Ksiazek S Jankowska K Olanczuk-Neyman KAntimicrobial resistance of fecal indicators in municipal wastewatertreatment plant Water Res 2010 445089ndash5097
21 Dulfour AP Ballentine RK Ambient water quality criteria for bacteria-1986 EPA4405-84-002 Washington DC US Environmental Protection Agency 1986
22 Ferreira da Silva M Tiago I Veriacutessimo A Boaventura RA Nunes OC Manaia CMAntimicrobial resistance patterns in Enterobacteriaceae isolated from anurban wastewater treatment plant FEMS Microbiol Ecol 2007 60166ndash176
23 DIRECTIVE 20067EC of the European Parliament and of the Council 15February 2006 concerning the management of bathing water quality andrepealing Directive 76160EEC httpeur-lexeuropaeuLexUriServLexUriServdouri=OJL200606400370051ENPDF
24 Freitas AR Novais C Ruiz-Garbajosa P Coque TM Peixe L Dispersion ofmultidrug-resistant Enterococcus faecium isolates belonging to majorclonal complexes in different Portuguese settings Appl Environ Microbiol2009 754904ndash4908
25 Poeta P Costa D Igrejas G Saacuteenz Y Zarazaga M Rodrigues J Torres CPolymorphisms of the pbp5 gene and correlation with ampicillinresistance in Enterococcus faecium isolates of animal origin J MedMicrobiol 2007 56236ndash240
26 Kuumlhn I Iversen A Burman LG Olsson-Liljequist B Franklin A Finn MAarestrup F Seyfarth AM Blanch AR Vilanova X Taylor H Caplin J MorenoMA Dominguez L Herrero IA Moumlllby R Comparison of enterococcalpopulations in animals humans and the environment ndash a Europeanstudy Int J Food Microbiol 2003 88133ndash145
27 Lanthier M Scott A Lapen DR Zhang Y Topp E Frequency of virulencegenes and antibiotic resistances in Enterococcus spp isolates fromwastewater and feces of domesticated mammals and birds and wildlifeCan J Microbiol 2010 56715ndash729
28 Leclercq R Oberleacute K Galopin S Cattoir V Budzinski H Petit F Changes inenterococcal populations and related antibiotic resistance along amedical center-wastewater treatment plant-river continuum Appl EnvironMicrobiol 2013 792428ndash2434
29 Thevenon F Regier N Benagli C Tonolla M Adatte T Wildi W Poteacute JCharacterization of fecal indicator bacteria in sediments cores from thelargest freshwater lake of Western Europe (Lake Geneva Switzerland)Ecotoxicol Environ Safety 2012 7850ndash56
30 Bonilla TD Nowosielski K Esiobu N McCorquodale DS Rogerson ASpecies assemblages of Enterococcus indicate potential sources offecal bacteria at a south Florida recreational beach Mar Pollut Bull2006 52807ndash810
31 Wang JT Chang SC Wang HY Chen PC Shiau YR Lauderdale TL TSARHospitals High rates of multidrug resistance in Enterococcus faecalis andE faecium isolated from inpatients and outpatients in TaiwanDiagn Microbiol Infect Dis 2013 75406ndash411
32 Lanthier M Scott A Zhang Y Cloutier M Durie D Henderson VC WilkesG Lapen DR Topp E Distribution of selected virulence genes andantibiotic resistance in Enterococcus species isolated from the SouthNation River drainage basin Ontario Canada J Appl Microbiol 2011110407ndash421
33 Kuch A Willems RJ Werner G Coque TM Hammerum AM Sundsfjord AKlare I Ruiz-Garbajosa P Simonsen GS van Luit-Asbroek M Hryniewicz WSadowy E Insight into antimicrobial susceptibility and population struc-ture of contemporary human Enterococcus faecalis isolates fromEurope J Antimicrob Chemother 2012 67551ndash558
34 Novais C Coque TM Ferreira H Sousa JC Peixe L Environmentalcontamination with vancomycin-resistant enterococci from hospitalsewage in Portugal Appl Environ Microbiol 2005 713364ndash3368
35 Iversen A Kuumlhn I Rahman M Franklin A Burman LG Olsson-Liljequist BTorell E Moumlllby R Evidence for transmission between humans and theenvironment of a nosocomial strain of Enterococcus faeciumEnviron Microbiol 2004 655ndash59
36 Rathnayake I Hargreaves M Huygens F SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterwayAustralia and associated antibiotic resistance gene profilesBMC Microbiol 2011 11201
37 Rosvoll TC Pedersen T Sletvold H Johnsen PJ Sollid JE Simonsen GSJensen LB Nielsen KM Sundsfjord A PCR-based plasmid typing inEnterococcus faecium strains reveals widely distributed pRE25-pRUM- pIP501- and pHT-related replicons associated with glycopeptideresistance and stabilizing toxin-antitoxin systems FEMS Immunol MedMicrobiol 2010 58254ndash268
38 Rosvoll TC Lindstad BL Lunde TM Hegstad K Aasnaes B Hammerum AMLester CH Simonsen GS Sundsfjord A Pedersen T Increased high-levelgentamicin resistance in invasive Enterococcus faecium is associated withaac(6prime)Ie-aph(2Prime)Ia-encoding transferable megaplasmids hosted by majorhospital-adapted lineages FEMS Immunol Med Microbiol 2012 66166ndash176
39 Laverde Gomez JA van Schaik W Freitas AR Coque TM Weaver KE FranciaMV Witte W Werner G A multiresistance megaplasmid pLG1 bearing ahylEfm genomic island in hospital Enterococcus faecium isolates Int J MedMicrobiol 2011 301165ndash175
40 Coque TM Evolutionary biology of pathogenic enterococci InEvolutionary Biology of Bacterial and Fungal Pathogens Coque TM Edited byBaquero F Nombela C Cassell GH Gutierrez JA Washington DC ASMPress501ndash523
41 Werner G Fleige C Geringer U van Schaik W Klare I Witte W IS elementIS16 as a molecular screening tool to identify hospital-associated strainsof Enterococcus faecium BMC Infect Dis 2011 1180
42 Top J Sinnige JC Majoor EA Bonten MJ Willems RJ van Schaik W Therecombinase IntA is required for excision of esp-containing ICEEfm1 inEnterococcus faecium J Bacteriol 2011 1931003ndash1006
43 Sillanpaumlauml J Prakash VP Nallapareddy SR Murray BE Distribution of genesencoding MSCRAMMs and Pili in clinical and natural populations ofEnterococcus faecium J Clin Microbiol 2009 47896ndash901
44 Marcinek H Wirth R Muscholl-Silberhorn A Gauer M Enterococcus faecalisgene transfer under natural conditions in municipal sewage watertreatment plants Appl Environ Microbiol 1998 64626ndash632
45 Kuumlmmerer K Significance of antibiotics in the environment J AntimicrobChemother 2003 525ndash7
46 Liu A Fong A Becket E Yuan J Tamae C Medrano L Maiz M Wahba C LeeC Lee K Tran KP Yang H Hoffman RM Salih A Miller JHP Selectiveadvantage of resistant strains at trace levels of antibiotics a simple andultrasensitive color test for detection of antibiotics and genotoxicagents Antimicrob Agents Chemother 2011 551204ndash1210
47 Clinical and Laboratory Standards Institute Performance standards forantimicrobial susceptibility testing 21st informational supplement Wayne CLSI2011 PA M100-S20
48 Magiorakos AP Srinivasan A Carey RB Carmeli Y Falagas ME Giske CGHarbarth S Hindler JF Kahlmeter G Olsson-Liljequist B Paterson DL Rice LBStelling J Struelens MJ Vatopoulos A Weber JT Monnet DLMultidrug-resistant extensively drug-resistant and pandrug-resistantbacteria an international expert proposal for interim standarddefinitions for acquired resistance Clin Microbiol Infect 2012 18268ndash281
49 Homan WL Tribe D Poznanski S Li M Hogg G Spalburg E Van Embden JDWillems RJ Multilocus sequence typing scheme for Enterococcus faeciumJ Clin Microbiol 2002 401963ndash1971
50 Jolley KA Feil EJ Chan MS Maiden MC Sequence type analysis andrecombinational tests (START) Bioinformatics 2001 171230ndash1231
51 Feil EJ Li BC Aanensen DM Hanage WP Spratt BG eBURST inferringpatterns of evolutionary descent among clusters of related bacterialgenotypes from multilocus sequence typing data J Bacteriol 20041861518ndash1530
52 Top J Banga1 NMI Hayes R Willems RJ Bonten MJM Hayden MKComparison of multiple-locus variable-number tandem repeat analysisand pulsed-field gel electrophoresis in a setting of polyclonal endemicityof vancomycin-resistant Enterococcus faecium Clin Microbiol Infect 200814363ndash369
53 Vankerckhoven V Van Autgaerden T Vael C Lammens C Chapelle S RossiR Jabes D Goossens H Development of a multiplex PCR for the
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-

Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 10 of 15httpwwwbiomedcentralcom1471-21801466
isolates resistant to ciprofloxacin were found also inCCs 5 9 and 22 These isolates possessed mutations inParC mentioned above but had the wild-type QRDR inGyrA HLGR occurred in five isolates of lineages 18 and78 and in single isolates of CC5 and CC22 this pheno-type was determined by the presence of aac(6prime)-Ie-aph(2Prime) with an exception of an isolate from CC5 whichwas negative for this gene High-level resistance tostreptomycin (HLSR) was widely distributed amongstudied isolates and determined by the ant(6prime)-Ia geneTetracycline resistance was found also in various STsthe major determinant of this phenotype was tet(M) (20isolates) followed by tet(L) (17 isolates) These twogenes occurred together in 16 isolates A single isolateof ST633 belonging to CC22 harbored tet(S) The tet(O) gene was absent in the studied group For a singletetracycline-resistant isolate of ST17 none of the fourtetracycline determinants studied was detected Nineisolates carried int of Tn916 and four the tndX gene ofTn5397 These transposon-specific genes were not char-acteristic for any lineage or complex (Table 3) Plasmid-replication genes such as rep2pRE25 rep17pRUM andrep18 pEF418 (26 14 and 7 isolates respectively) andgenes specifying toxin-antitoxin systems (TAS) axe-txeand ω-ε-ζ (12 and 10 isolates respectively) were signifi-cantly associated with the group of isolates of nosocomialE faecium Two other rep genes (rep1pIP501 reppLG1) werewidely distributed among all studied E faecium isolatesand the reppMG1 gene occurred in two isolates (in a singleisolate of ST533 from CC22 and in a singleton ST647) Allω-ε-ζ-positive isolates carried rep2pRE25 and nine isolatesshowed a concomitant presence of axe-txe and rep17pRUMThe relBE genes were found solely among five non-nosocomial E faecium (a difference in distribution with-out statistical significance p = 043) among various STs
DiscussionIn this study Enterococcus spp was used to evaluate theimpact posed by treated wastewater of two local WWTPson their receiver the costal waters of Gulf of GdanskTotal counts of enterococci observed for raw (W-INF)and treated (W-EFF D-EFF) wastewater are consistentwith our previous study showing similar over 99 reduc-tion capacity of applied treatment processes It is alsoreported in the literature that wastewater treatment pro-cesses based on activated sludge although they focus onremoval of nutrients and organic matter simultaneouslyreduce faecal indicators with high efficiency reaching upto 999 [1822] Due to the high number of enterococciin raw wastewater they are nevertheless released from aWWTP in a number up to 61x105 CFU per 100 mLThus marine water sampled in the area directly impactedby treated wastewater (W-MOut and D-MOut) showed ahigher number of enterococci than the one observed in
Vistula River mouth however lower by four orders ofmagnitude in comparison to corresponding treated waste-water (W-EFF and D-EFF) It should be noted that in mar-ine outfalls (W-MOut D-MOut) as well as in VistulaRiver mouth enterococci were detected in number lowerthan 100 CFU per 100 mL ie the value set out by NewBathing Water Directive 20067EC for the costal water ofldquoexcellent qualityrdquo [23]It is suggested that enterococcal species composition in
faecal-impacted environment depends from the source ofcontamination In this study two species most commonlycolonizing humans E faecalis and E faecium comprisedtogether from 73 to 88 of enterococcal isolates intested wastewater samples (W-INF W-BR W-EFF and D-EFF) 73 and 61 in marine outflows (W-MOut and D-MOut respectively) and 45 in the Vistula River mouthThe predominant species in wastewater and marine waterimpacted by treated wastewater was E faecium (from 56to 65) which stayed at a similar level during treatmentprocesses as determined also in our previous study [20]Species distribution in wastewater samples is not wellunderstood and affecting factors may include differencesin diet climate season and methods of detection E feaca-lis was found to be the most prevalent species in wastewa-ter samples in Sweden [26] E faecium in Spain UKCanada France and Switzerland [26-29] while E hiraepredominated in Portugal and United States [2230] Al-though E faecalis is typically the most abundant colonizerof humans in the community (eg [2631]) E faecium isgenerally most often associated with human microbialcontamination [2729] E faecalis was reported fromagriculture-impacted waters [32] and E hirae likelyoriginates mainly from cattle and pig faecal contamin-ation [26] which explains high relative proportion ofthese two species in Vistula River mouth Apart from thevariability of contamination sources proportion of par-ticular species may differ due to the differences of abilityto sustain environmental stresses In this study enterococ-cal composition might have been influenced by wastewa-ter treatment processes and marine water characteristicsThe effect of wastewater treatment and marine environ-
ment was seen on phenotypes of antimicrobial resistanceThe positive selection of bacteria with resistance patternshas been already suggested and noted in wastewater pro-cesses [192022] In this study the resistance rates to ampi-cillin ciprofloxacin levofloxacin and erythromycin notedfor E faecium from raw wastewater (W-INF) were lowerthan these observed for this species in the correspondingtreated wastewater (W-EFF) however not statistically sig-nificant (p gt 005) Moreover resistance rates to ampicillinciprofloxacin tetracycline and erythromycin observed inmarine outflow of WWTP Gdansk-Wschod (W-MOut)were higher than in treated wastewater of this plantSuch phenomenon was not observed for treated
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 11 of 15httpwwwbiomedcentralcom1471-21801466
wastewater of WWTP Gdynia-Debogorze (D-EFF) andits marine outfall (D-MOut) Resistance rates among Efaecium originating from D-MOut were in generalcomparable to these detected in the Vistula Rivermouth It should be noted that treated wastewater fromWWTP Gdansk-Wschod has been discharged via mar-ine outfall (W-MOut) for last 12 years while marineoutfall of WWTP Gdynia-Debogorze (D-MOut) hasbeen operating for last two years The observed differ-ences in resistance rate corresponding to the local im-pact caused by long and short-term operated marineoutfalls (W-MOut and D-MOut) need further attentionSignificant rates of antimicrobial resistance observed
for isolates of E faecalis from wastewater and marineriver waters impacted by wastewater are in agreementwith a recent study performed in six European countriesshowing that in Poland the rates of resistance for humanisolates of this species were usually among the highestboth in the community and hospitals [33] To our know-ledge no corresponding data are available for E faeciumcolonization in the community in Poland but a studyfocused on hospital VRE isolates showed their ubiquitousMDR phenotype and a high load of resistance genes [13]In our study no vancomycin resistance were detectedlikely due to relatively low (below 10) prevalence of VREin Polish hospitals [5] although such isolates are isolatedfrom the environment in other countries eg [34]This study shows that hospital clones of E faecium are
important carriers of resistance genes in wastewatersMost likely in a significant part they originated fromhospital wastewater treated together with communalwastewater In our estimation hospital wastewater con-stitutes approximately 02 of both WWTPsrsquo daily in-flow The release of hospital-associated clones of Efaecium to the environment was observed also by others[24283536] In our study isolates belonging to themajor nosocomial HiRECC (formerly named CC17)constituted altogether 246 of all isolates of E faeciumsubjected to typing and were present at all samplingsites These isolates apart from their inclusion into theHiRECC by MLST demonstrated several other pheno-typic and genotypic features typical for nosocomial Efaecium ie they were resistant to ciprofloxacin andampicillin in vast majority showed MDR phenotypeand carried several resistance determinants associatedboth with mutations of chromosomal genes (gyrA parCpbp5) and in the case of aminoglycoside and tetracyclineresistance with gene acquisition Plasmids likely played animportant role in the later process as some plasmid genessuch as rep2pRE25 rep17pRUM and rep18 pEF418 and TASgenes axe-txe and ω-ε-ζ were significantly more abundantin isolates associated with HiRECC than among theremaining isolates As yet the knowledge of distributionof plasmid-associated genes in populations of E faecium
especially in the context of their clonal compositionremains limited A study on 93 isolates of differentgeographical origins and from various sites showed anover-representation of rep17pRUM and reppMG1 amongnosocomial E faecium while in contrast to our observa-tions rep18 pEF418 was not detected [37] Moreover wedid not observe reppMG1 among HiRECC of E faeciumThese differences may be due to a high prevalence of VRE(58 isolates) among 93 isolates studied by Rosvoll et alwhat may explain differences in the plasmid content Thesame study also reported the co-localization of ω-ε-ζ withrep2pRE25 and axe-txe with rep17pRUM on the same plas-mids in concordance with our findings showing a fre-quent joint occurrence of these pairs of genes Anotherstudy performed on 99 invasive isolates from Norwaymostly of HLGR phenotype showed a significantly higherprevalence of rep2pRE25 and rep17pRUM among the majornosocomial HiRECC [38] This study as well some others[2439] also demonstrated a common presence of reppLG1among E faecium from hospital settings and in animalsOur findings indicate that this type of replicon is evenmore ubiquitous in E faecium than previously reportedAll rep genes mentioned above are typically associatedwith plasmids carrying various antimicrobial resistancegenes [3940] GenBank accession number AF408195 Inaddition to genes determining antimicrobial resistanceand genes associated with MGE isolates of nosocomialHiRECC analyzed in our study carried such molecularmarkers as IS16 and the esp gene together with the intAgene characteristic for ICEEfm [4142] Another patho-genicity factor the fms19 gene of the pilin gene cluster4 (PGC4) showed increased prevalence among theseisolates Similar observation was made for a representa-tive collection of 433 E faecium isolates from varioussources [43]In a study that followed enterococci in a continuum
from hospital and retirement home wastewaters througha WWTP to a river a gradual decrease in the proportionof E faecium isolates representing hospital-associatedSTs was observed likely due to the ldquodilutionrdquo of HiRECCrepresentatives by more diverse and less drug-resistantstrains originating from the community [28] Because inour study only selected representatives of obtained iso-lates were investigated it is not possible to determineprecise tendencies in their frequency from the WWTPinfluent to effluent and water however resistant entero-cocci both HiRECC and non-HiRECC are clearly ableto survive the treatment process what results in their re-lease into marine and river waters Moreover using anadditional typing method MLVA we were able to findisolates with the same characteristics in the effluent ofWWTP and its marine outfall suggesting ability of suchclones to survive in the environment for at least sometime It seems plausible that hospital resistant clones
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 12 of 15httpwwwbiomedcentralcom1471-21801466
accumulate several determinants that may promote theirsurvival not only in the hospital settings but also in theenvironment Such features would include resistance todisinfectants and ultraviolet light additional metabolicpathways and ability to form biofilms Moreover determi-nants of some of the mentioned adaptation factors arefound co-resident in plasmids harboring resistance genessuggesting co-selection For example pLG1-type plasmidsof E faecium carry not only resistance determinantsagainst aminoglycosides glycopeptides and macrolidesbut also pili genes and carbon uptake-utilization genes[3839] Pheromone-responsive plasmids of E faecalisapart from resistance to aminoglycosides glycopeptidespenicillins and macrolides may encode bacteriocins add-itional pili and UV-resistance determinants [40] Thusnosocomial HiRECCs released by WWTPs are equippedin whole sets of genes with various adaptive benefits andoften located within MGE It was also demonstrated thatMGEs can be transferred in a WWTP [44] further under-ling the importance of threat posed by such organismsThere is a selection pressure for survival of resistantstrains and their tranconjugants due to release of antibi-otics to the environment especially in the hospital efflu-ents [45] and even sub-inhibitory concentrations may beeffective in selection for resistance [46]
ConclusionsThis study describes the species composition resistanceprofiles and clonal relationships of enterococci isolatedfrom WWTPs located in Gdansk their marine outfallsto the Gulf of Gdansk and from Vistula River It was ob-served that these WWTPs release enterococcal strainsresistant to various antimicrobial compounds to the re-cipient water environment Molecular analyses revealedthe presence of clones associated with nosocomial HiR-ECCs and in the case of E faecium isolates of lineages17 18 and 78 of former CC17 abundant in resistance de-terminants MGE genes and pathogenicity factors Thepresence of HiRECCs in wastewaters and marineriverenvironment highlights the need for further detailedanalyses to better understand the survival and spread ofdrug-resistant strains in water ecosystems and to eluci-date the ways to curb such dissemination
MethodsAcquisition of samples and isolation of enterococciOnce a month in April June August and October in 2011of the flow proportioned composited samples of wastewa-ter were taken from two local WWTPs Gdansk-Wschodfrom influent (W-INF) effluent (W-EFF) and wastewa-ter from activated sludge bioreactor (W-BR) and fromGdynia-Debogorze effluent (D-EFF) The two WWTPstreat sewage from approximately 920 000 inhabitants(for details see Figure 1) In the same period of time and
with the same frequency marine outfalls of those WWTPs(W-MOut and D-MOut respectively) as well as VistulaRiver mouth were also sampled for isolation of entero-cocci Marine samples were taken at the depth of approxi-mately one meter below the surface once a month inApril June August and October of 2011 Location ofsampling points and their characteristics are provided inFigure 1 Enterococci were detected using the membranefiltration method To this end appropriate dilutions ofanalyzed samples were filtered in duplicate through045 μm cellulose-acetate filters (EMD Millipore Cor-poration Billerica MA USA) which were then placedon Enterococcus selective agar (Merck DarmstadtGermany) and incubated at 37degC for 48 h (ISO 7899ndash22000) Dark red or maroon colonies were consideredpresumptive enterococci For further investigationsrepresentative isolates were taken from membranes pre-senting from 20 to 50 typical colonies and stored in nu-trient broth supplemented with 15 glycerol at minus80degC
Species identification DNA isolation susceptibility testingand statistical analysisThe species identification and drug susceptibility of pre-sumptive enterococci were determined by the PhoenixtradeAutomated Microbiology System (BD New Jersey USA)according to the manufacturerrsquos instructions Since it isnot possible to distinguish E casseliflavus and E galli-narum using this system the classification E casselifla-vusgallinarum was used The susceptibility tests basedon microdilution were carried out for 11 antimicrobialagents gentamicin (GM) streptomycin (ST) ampicillin(AM) daptomycin (DAP) vancomycin (VA) teicoplanin(TEC) linezolid (LZD) fluoroquinolones (ciprofloxacin -CIP levofloxacin - LVX) tetracycline (TE) erythromycin(E) and quinupristin-dalfopristin (SYN) (E faecium only)Obtained minimal inhibitory concentrations (MIC) wereevaluated according to the guidelines of Clinical and La-boratory Standards Institute [47] Isolates were defined asmultidrug-resistant (MDR) when they showed resistanceto three or more compounds tested [48] Bacterial DNAwas purified using the Genomic DNA Prep Plus kitfollowing the manufacturerrsquos instructions (AampA Biotech-nology Gdynia Poland) Differences in distributions wereevaluated using the χ2 test with the p values lt 005 con-sidered significant
MLST and MLVA of E faeciumMLST for E faecium was performed by sequencing ofseven house-keeping genes as described by Homan et al[49] The allele numbers and STs were assigned with theuse of Internet database httpefaeciummlstnet (5th July2013 date last accessed) Novel alleles and allelic profileswere submitted to the database MLST data were analyzedwith Sequence Type Analysis and Recombinational Tests
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 13 of 15httpwwwbiomedcentralcom1471-21801466
(START) software [50] and the eBURST analysis [51] wasused to elucidate the relationships of STs with knownCCs using the comparative eBURST option available athttpeburstmlstnetv3enter_datacomparative (5th July2013 date last accessed) Further inclusion of E faeciumSTs into specific CCs and lineages was performed fol-lowing other studies [924] MLVA was performed as de-scribed by others [52]
Detection and analysis of genes associated with virulenceantimicrobial resistance and MGE in E faeciumThe full list of primers used for gene detection and se-quencing is provided in Additional file 1 Table S1 PCR-based detection of IS16 virulence-associated genes (fms21fms17 fms5 and fms19 representing each of four pilingene clusters in E faecium and the espEfm gene) genes de-termining resistance to tetracycline tet(M) tet(O) tet(L)tet(S) HLGR (aac(6prime)-Ie-aph(2Prime) HLSR (ant(6prime)-Ia) andgenes specific for MGE such as intA of ICEEfm1 int ofTn916 transposon tndX of Tn5397 transposon entero-coccal plasmid replication genes rep1pIP501 rep2pRE25rep17pRUM rep18pEF418 reppMG1 reppLG1 and TAS genesaxe-txe ω-ε-ζ relBE was performed as described in otherstudies [134153-59] following the conditions describedherein Previously characterized clinical isolates of Strepto-coccus agalactiae E faecium and E faecalis harbouringpilin genes IS16 espEfm intA of ICEEfm1 int of Tn916tndX of Tn5397 tet(M) tet(O) tet(L) tet(S) aac(6prime)-Ie-aph(2Prime) ant(6prime)-Ia rep1pIP501 rep2pRE25 ω-ε-ζ [136061]served as positive controls In cases when positive controlswere not available (rep18pEF418 reppMG1 reppLG1 axe-txerelBE) a few randomly selected PCR products wereverified by sequencing and served further as controlsAmpicillin-resistance determinant the pbp5 gene was ana-lysed by sequencing of the part encoding C-terminaltranspeptidase domain in PBP5 [62] New sequence vari-ants pbp5-24 to pbp5-29 alleles were submitted to theGenBank (acc numbers KC594860-KC594865 respect-ively) The quinolone-determining regions (QRDR) in thegyrA and parC genes were sequenced and analysed asdescribed by others [63]
Additional file
Additional file 1 Table S1 Primers used in the study for PCR andsequencing
Competing interestThe authors declare that they have no competing interest
Authorsrsquo contributionsES participated in the study design performed experimental work andparticipated in the manuscript preparation AL designed the studyperformed experimental work and participated in the manuscriptpreparation Both authors read and approved the final manuscript
AcknowledgementsWe thank Rob Willems and Janetta Top for assigning new alleles new STsand MTs This publication made use of the Enterococcus faecium MLSTwebsite (httpefaeciummlstnet) which is hosted at Imperial College of theUniversity of Oxford and funded by the Wellcome Trust This work wassupported by project N N305 461139 from the Narodowe Centrum Nauki(NCN)
Author details1National Medicines Institute ul Chelmska 3034 Warsaw 00-725 Poland2Gdansk University of Technology ul G Narutowicza 1112 Gdansk 80-952Poland
Received 5 September 2013 Accepted 27 February 2014Published 14 March 2014
References1 Aarestrup FM Butaye P Witte W Nonhuman reservoirs of Enterococci In
American Society for Microbiology Edited by Gilmore MS Clewell DB CourvalinP Dunny GM Murray BE Rice LB Washington DC The Enterococcipathogenesis molecular biology and antibiotic resistance 2002
2 Tendolkar PM Baghdayan AS Shankar N Pathogenic enterococci newdevelopments in the 21st century Cell Mol Life Sci 2003 602622ndash2636
3 Sood S Malhotra M Das BK Kapil A Enterococcal infections ampantimicrobial resistance Indian J Med Res 2008 128111ndash121
4 Sydnor ER Perl TM Hospital epidemiology and infection control inacute-care settings Clin Microbiol Rev 2011 24141ndash173
5 European Centre for Disease Prevention and Control Annual EpidemiologicalReport Reporting on 2010 surveillance data and 2011 epidemic intelligencedata Stockholm ECDC 20122013
6 Hidron AI Edwards JR Patel J Horan TC Sievert DM Pollock DA Fridkin SKNational Healthcare Safety Network Team Participating National HealthcareSafety Network Facilities NHSN annual update antimicrobial-resistantpathogens associated with healthcare-associated infections annualsummary of data reported to the National Healthcare Safety Network atthe Centers for Disease Control and Prevention 2006ndash2007 Infect ControlHosp Epidemiol 2008 29996ndash1011
7 De Kraker ME Jarlier V Monen JC Heuer OE van de Sande N GrundmannH The changing epidemiology of bacteraemias in Europe trends fromthe European Antimicrobial Resistance Surveillance System Clin MicrobiolInfect 2012 doi1011111469-0691
8 Leavis HL Bonten MJ Willems RJ Identification of high-risk enterococcalclonal complexes global dispersion and antibiotic resistance Curr OpinMicrobiol 2006 9454ndash460
9 Willems RJL Top J van Schaik W Leavis H Bonten M Sireacuten J Hanage WPCorander J Restricted gene flow among hospital subpopulations ofEnterococcus faecium mBio 2012 3e00151ndash12
10 Hegstad K Mikalsen T Coque TM Werner G Sundsfjord A Mobile geneticelements and their contribution to the emergence of antimicrobialresistant Enterococcus faecalis and Enterococcus faecium Clin MicrobiolInfect 2010 16541ndash554
11 Shepard BD Gilmore MS Antibiotic-resistant enterococci themechanisms and dynamics of drug introduction and resistance MicrobesInfect 2002 4215ndash224
12 Bonten MJ Willems R Weinstein RA Vancomycin-resistant enterococciwhy are they here and where do they come from Lancet Infect Dis 20011314ndash325
13 Sadowy E Sieńko A Gawryszewska I Bojarska A Malinowska K HryniewiczW High abundance and diversity of antimicrobial resistancedeterminants among early vancomycin-resistant Enterococcus faecium inPoland Eur J Clin Microbiol Infect Dis 2013 321193ndash1203
14 Willems RJ Homan W Top J van Santen-Verheuvel M Tribe D Manzioros XGaillard C Vandenbroucke-Grauls CM Mascini EM van Kregten E vanEmbden JD Bonten MJ Variant esp gene as a marker of a distinct geneticlineage of vancomycin-resistant Enterococcus faecium spreading inhospitals Lancet 2001 357853ndash855
15 Sillanpaumlauml J Nallapareddy SR Prakash VP Qin X Houmloumlk M Weinstock GMMurray BE Identification and phenotypic characterization of a secondcollagen adhesin Scm and genome-based identification and analysis of13 other predicted MSCRAMMs including four distinct pilus loci inEnterococcus faecium Microbiology 2008 1543199ndash3211
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 14 of 15httpwwwbiomedcentralcom1471-21801466
16 Galloway-Pentildea JR Nallapareddy SR Arias CA Eliopoulos GM Murray BEAnalysis of clonality and antibiotic resistance among early clinicalisolates of Enterococcus faecium in the United States J Infect Dis 20092001566ndash1573
17 Top J Willems R Bonten M Emergence of CC17 Enterococcus faeciumfrom commensal to hospital-adapted pathogen FEMS Immunol MedMicrobiol 2008 52297ndash308
18 Martins da Costa P Vaz-Pires P Bernardo F Antimicrobial resistance inEnterococcus spp isolated in inflow effluent and sludge from municipalsewage water treatment plants Water Res 2006 401735ndash1740
19 Reinthaler FF Posch J Feierl G Wust G Haas D Ruckenbauer G Mascher FMarth E Antibiotic resistance of Escherichia coli in sewage and sludgeWater Res 2003 371685ndash1690
20 Luczkiewicz A Fudala-Ksiazek S Jankowska K Olanczuk-Neyman KAntimicrobial resistance of fecal indicators in municipal wastewatertreatment plant Water Res 2010 445089ndash5097
21 Dulfour AP Ballentine RK Ambient water quality criteria for bacteria-1986 EPA4405-84-002 Washington DC US Environmental Protection Agency 1986
22 Ferreira da Silva M Tiago I Veriacutessimo A Boaventura RA Nunes OC Manaia CMAntimicrobial resistance patterns in Enterobacteriaceae isolated from anurban wastewater treatment plant FEMS Microbiol Ecol 2007 60166ndash176
23 DIRECTIVE 20067EC of the European Parliament and of the Council 15February 2006 concerning the management of bathing water quality andrepealing Directive 76160EEC httpeur-lexeuropaeuLexUriServLexUriServdouri=OJL200606400370051ENPDF
24 Freitas AR Novais C Ruiz-Garbajosa P Coque TM Peixe L Dispersion ofmultidrug-resistant Enterococcus faecium isolates belonging to majorclonal complexes in different Portuguese settings Appl Environ Microbiol2009 754904ndash4908
25 Poeta P Costa D Igrejas G Saacuteenz Y Zarazaga M Rodrigues J Torres CPolymorphisms of the pbp5 gene and correlation with ampicillinresistance in Enterococcus faecium isolates of animal origin J MedMicrobiol 2007 56236ndash240
26 Kuumlhn I Iversen A Burman LG Olsson-Liljequist B Franklin A Finn MAarestrup F Seyfarth AM Blanch AR Vilanova X Taylor H Caplin J MorenoMA Dominguez L Herrero IA Moumlllby R Comparison of enterococcalpopulations in animals humans and the environment ndash a Europeanstudy Int J Food Microbiol 2003 88133ndash145
27 Lanthier M Scott A Lapen DR Zhang Y Topp E Frequency of virulencegenes and antibiotic resistances in Enterococcus spp isolates fromwastewater and feces of domesticated mammals and birds and wildlifeCan J Microbiol 2010 56715ndash729
28 Leclercq R Oberleacute K Galopin S Cattoir V Budzinski H Petit F Changes inenterococcal populations and related antibiotic resistance along amedical center-wastewater treatment plant-river continuum Appl EnvironMicrobiol 2013 792428ndash2434
29 Thevenon F Regier N Benagli C Tonolla M Adatte T Wildi W Poteacute JCharacterization of fecal indicator bacteria in sediments cores from thelargest freshwater lake of Western Europe (Lake Geneva Switzerland)Ecotoxicol Environ Safety 2012 7850ndash56
30 Bonilla TD Nowosielski K Esiobu N McCorquodale DS Rogerson ASpecies assemblages of Enterococcus indicate potential sources offecal bacteria at a south Florida recreational beach Mar Pollut Bull2006 52807ndash810
31 Wang JT Chang SC Wang HY Chen PC Shiau YR Lauderdale TL TSARHospitals High rates of multidrug resistance in Enterococcus faecalis andE faecium isolated from inpatients and outpatients in TaiwanDiagn Microbiol Infect Dis 2013 75406ndash411
32 Lanthier M Scott A Zhang Y Cloutier M Durie D Henderson VC WilkesG Lapen DR Topp E Distribution of selected virulence genes andantibiotic resistance in Enterococcus species isolated from the SouthNation River drainage basin Ontario Canada J Appl Microbiol 2011110407ndash421
33 Kuch A Willems RJ Werner G Coque TM Hammerum AM Sundsfjord AKlare I Ruiz-Garbajosa P Simonsen GS van Luit-Asbroek M Hryniewicz WSadowy E Insight into antimicrobial susceptibility and population struc-ture of contemporary human Enterococcus faecalis isolates fromEurope J Antimicrob Chemother 2012 67551ndash558
34 Novais C Coque TM Ferreira H Sousa JC Peixe L Environmentalcontamination with vancomycin-resistant enterococci from hospitalsewage in Portugal Appl Environ Microbiol 2005 713364ndash3368
35 Iversen A Kuumlhn I Rahman M Franklin A Burman LG Olsson-Liljequist BTorell E Moumlllby R Evidence for transmission between humans and theenvironment of a nosocomial strain of Enterococcus faeciumEnviron Microbiol 2004 655ndash59
36 Rathnayake I Hargreaves M Huygens F SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterwayAustralia and associated antibiotic resistance gene profilesBMC Microbiol 2011 11201
37 Rosvoll TC Pedersen T Sletvold H Johnsen PJ Sollid JE Simonsen GSJensen LB Nielsen KM Sundsfjord A PCR-based plasmid typing inEnterococcus faecium strains reveals widely distributed pRE25-pRUM- pIP501- and pHT-related replicons associated with glycopeptideresistance and stabilizing toxin-antitoxin systems FEMS Immunol MedMicrobiol 2010 58254ndash268
38 Rosvoll TC Lindstad BL Lunde TM Hegstad K Aasnaes B Hammerum AMLester CH Simonsen GS Sundsfjord A Pedersen T Increased high-levelgentamicin resistance in invasive Enterococcus faecium is associated withaac(6prime)Ie-aph(2Prime)Ia-encoding transferable megaplasmids hosted by majorhospital-adapted lineages FEMS Immunol Med Microbiol 2012 66166ndash176
39 Laverde Gomez JA van Schaik W Freitas AR Coque TM Weaver KE FranciaMV Witte W Werner G A multiresistance megaplasmid pLG1 bearing ahylEfm genomic island in hospital Enterococcus faecium isolates Int J MedMicrobiol 2011 301165ndash175
40 Coque TM Evolutionary biology of pathogenic enterococci InEvolutionary Biology of Bacterial and Fungal Pathogens Coque TM Edited byBaquero F Nombela C Cassell GH Gutierrez JA Washington DC ASMPress501ndash523
41 Werner G Fleige C Geringer U van Schaik W Klare I Witte W IS elementIS16 as a molecular screening tool to identify hospital-associated strainsof Enterococcus faecium BMC Infect Dis 2011 1180
42 Top J Sinnige JC Majoor EA Bonten MJ Willems RJ van Schaik W Therecombinase IntA is required for excision of esp-containing ICEEfm1 inEnterococcus faecium J Bacteriol 2011 1931003ndash1006
43 Sillanpaumlauml J Prakash VP Nallapareddy SR Murray BE Distribution of genesencoding MSCRAMMs and Pili in clinical and natural populations ofEnterococcus faecium J Clin Microbiol 2009 47896ndash901
44 Marcinek H Wirth R Muscholl-Silberhorn A Gauer M Enterococcus faecalisgene transfer under natural conditions in municipal sewage watertreatment plants Appl Environ Microbiol 1998 64626ndash632
45 Kuumlmmerer K Significance of antibiotics in the environment J AntimicrobChemother 2003 525ndash7
46 Liu A Fong A Becket E Yuan J Tamae C Medrano L Maiz M Wahba C LeeC Lee K Tran KP Yang H Hoffman RM Salih A Miller JHP Selectiveadvantage of resistant strains at trace levels of antibiotics a simple andultrasensitive color test for detection of antibiotics and genotoxicagents Antimicrob Agents Chemother 2011 551204ndash1210
47 Clinical and Laboratory Standards Institute Performance standards forantimicrobial susceptibility testing 21st informational supplement Wayne CLSI2011 PA M100-S20
48 Magiorakos AP Srinivasan A Carey RB Carmeli Y Falagas ME Giske CGHarbarth S Hindler JF Kahlmeter G Olsson-Liljequist B Paterson DL Rice LBStelling J Struelens MJ Vatopoulos A Weber JT Monnet DLMultidrug-resistant extensively drug-resistant and pandrug-resistantbacteria an international expert proposal for interim standarddefinitions for acquired resistance Clin Microbiol Infect 2012 18268ndash281
49 Homan WL Tribe D Poznanski S Li M Hogg G Spalburg E Van Embden JDWillems RJ Multilocus sequence typing scheme for Enterococcus faeciumJ Clin Microbiol 2002 401963ndash1971
50 Jolley KA Feil EJ Chan MS Maiden MC Sequence type analysis andrecombinational tests (START) Bioinformatics 2001 171230ndash1231
51 Feil EJ Li BC Aanensen DM Hanage WP Spratt BG eBURST inferringpatterns of evolutionary descent among clusters of related bacterialgenotypes from multilocus sequence typing data J Bacteriol 20041861518ndash1530
52 Top J Banga1 NMI Hayes R Willems RJ Bonten MJM Hayden MKComparison of multiple-locus variable-number tandem repeat analysisand pulsed-field gel electrophoresis in a setting of polyclonal endemicityof vancomycin-resistant Enterococcus faecium Clin Microbiol Infect 200814363ndash369
53 Vankerckhoven V Van Autgaerden T Vael C Lammens C Chapelle S RossiR Jabes D Goossens H Development of a multiplex PCR for the
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-

Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 11 of 15httpwwwbiomedcentralcom1471-21801466
wastewater of WWTP Gdynia-Debogorze (D-EFF) andits marine outfall (D-MOut) Resistance rates among Efaecium originating from D-MOut were in generalcomparable to these detected in the Vistula Rivermouth It should be noted that treated wastewater fromWWTP Gdansk-Wschod has been discharged via mar-ine outfall (W-MOut) for last 12 years while marineoutfall of WWTP Gdynia-Debogorze (D-MOut) hasbeen operating for last two years The observed differ-ences in resistance rate corresponding to the local im-pact caused by long and short-term operated marineoutfalls (W-MOut and D-MOut) need further attentionSignificant rates of antimicrobial resistance observed
for isolates of E faecalis from wastewater and marineriver waters impacted by wastewater are in agreementwith a recent study performed in six European countriesshowing that in Poland the rates of resistance for humanisolates of this species were usually among the highestboth in the community and hospitals [33] To our know-ledge no corresponding data are available for E faeciumcolonization in the community in Poland but a studyfocused on hospital VRE isolates showed their ubiquitousMDR phenotype and a high load of resistance genes [13]In our study no vancomycin resistance were detectedlikely due to relatively low (below 10) prevalence of VREin Polish hospitals [5] although such isolates are isolatedfrom the environment in other countries eg [34]This study shows that hospital clones of E faecium are
important carriers of resistance genes in wastewatersMost likely in a significant part they originated fromhospital wastewater treated together with communalwastewater In our estimation hospital wastewater con-stitutes approximately 02 of both WWTPsrsquo daily in-flow The release of hospital-associated clones of Efaecium to the environment was observed also by others[24283536] In our study isolates belonging to themajor nosocomial HiRECC (formerly named CC17)constituted altogether 246 of all isolates of E faeciumsubjected to typing and were present at all samplingsites These isolates apart from their inclusion into theHiRECC by MLST demonstrated several other pheno-typic and genotypic features typical for nosocomial Efaecium ie they were resistant to ciprofloxacin andampicillin in vast majority showed MDR phenotypeand carried several resistance determinants associatedboth with mutations of chromosomal genes (gyrA parCpbp5) and in the case of aminoglycoside and tetracyclineresistance with gene acquisition Plasmids likely played animportant role in the later process as some plasmid genessuch as rep2pRE25 rep17pRUM and rep18 pEF418 and TASgenes axe-txe and ω-ε-ζ were significantly more abundantin isolates associated with HiRECC than among theremaining isolates As yet the knowledge of distributionof plasmid-associated genes in populations of E faecium
especially in the context of their clonal compositionremains limited A study on 93 isolates of differentgeographical origins and from various sites showed anover-representation of rep17pRUM and reppMG1 amongnosocomial E faecium while in contrast to our observa-tions rep18 pEF418 was not detected [37] Moreover wedid not observe reppMG1 among HiRECC of E faeciumThese differences may be due to a high prevalence of VRE(58 isolates) among 93 isolates studied by Rosvoll et alwhat may explain differences in the plasmid content Thesame study also reported the co-localization of ω-ε-ζ withrep2pRE25 and axe-txe with rep17pRUM on the same plas-mids in concordance with our findings showing a fre-quent joint occurrence of these pairs of genes Anotherstudy performed on 99 invasive isolates from Norwaymostly of HLGR phenotype showed a significantly higherprevalence of rep2pRE25 and rep17pRUM among the majornosocomial HiRECC [38] This study as well some others[2439] also demonstrated a common presence of reppLG1among E faecium from hospital settings and in animalsOur findings indicate that this type of replicon is evenmore ubiquitous in E faecium than previously reportedAll rep genes mentioned above are typically associatedwith plasmids carrying various antimicrobial resistancegenes [3940] GenBank accession number AF408195 Inaddition to genes determining antimicrobial resistanceand genes associated with MGE isolates of nosocomialHiRECC analyzed in our study carried such molecularmarkers as IS16 and the esp gene together with the intAgene characteristic for ICEEfm [4142] Another patho-genicity factor the fms19 gene of the pilin gene cluster4 (PGC4) showed increased prevalence among theseisolates Similar observation was made for a representa-tive collection of 433 E faecium isolates from varioussources [43]In a study that followed enterococci in a continuum
from hospital and retirement home wastewaters througha WWTP to a river a gradual decrease in the proportionof E faecium isolates representing hospital-associatedSTs was observed likely due to the ldquodilutionrdquo of HiRECCrepresentatives by more diverse and less drug-resistantstrains originating from the community [28] Because inour study only selected representatives of obtained iso-lates were investigated it is not possible to determineprecise tendencies in their frequency from the WWTPinfluent to effluent and water however resistant entero-cocci both HiRECC and non-HiRECC are clearly ableto survive the treatment process what results in their re-lease into marine and river waters Moreover using anadditional typing method MLVA we were able to findisolates with the same characteristics in the effluent ofWWTP and its marine outfall suggesting ability of suchclones to survive in the environment for at least sometime It seems plausible that hospital resistant clones
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 12 of 15httpwwwbiomedcentralcom1471-21801466
accumulate several determinants that may promote theirsurvival not only in the hospital settings but also in theenvironment Such features would include resistance todisinfectants and ultraviolet light additional metabolicpathways and ability to form biofilms Moreover determi-nants of some of the mentioned adaptation factors arefound co-resident in plasmids harboring resistance genessuggesting co-selection For example pLG1-type plasmidsof E faecium carry not only resistance determinantsagainst aminoglycosides glycopeptides and macrolidesbut also pili genes and carbon uptake-utilization genes[3839] Pheromone-responsive plasmids of E faecalisapart from resistance to aminoglycosides glycopeptidespenicillins and macrolides may encode bacteriocins add-itional pili and UV-resistance determinants [40] Thusnosocomial HiRECCs released by WWTPs are equippedin whole sets of genes with various adaptive benefits andoften located within MGE It was also demonstrated thatMGEs can be transferred in a WWTP [44] further under-ling the importance of threat posed by such organismsThere is a selection pressure for survival of resistantstrains and their tranconjugants due to release of antibi-otics to the environment especially in the hospital efflu-ents [45] and even sub-inhibitory concentrations may beeffective in selection for resistance [46]
ConclusionsThis study describes the species composition resistanceprofiles and clonal relationships of enterococci isolatedfrom WWTPs located in Gdansk their marine outfallsto the Gulf of Gdansk and from Vistula River It was ob-served that these WWTPs release enterococcal strainsresistant to various antimicrobial compounds to the re-cipient water environment Molecular analyses revealedthe presence of clones associated with nosocomial HiR-ECCs and in the case of E faecium isolates of lineages17 18 and 78 of former CC17 abundant in resistance de-terminants MGE genes and pathogenicity factors Thepresence of HiRECCs in wastewaters and marineriverenvironment highlights the need for further detailedanalyses to better understand the survival and spread ofdrug-resistant strains in water ecosystems and to eluci-date the ways to curb such dissemination
MethodsAcquisition of samples and isolation of enterococciOnce a month in April June August and October in 2011of the flow proportioned composited samples of wastewa-ter were taken from two local WWTPs Gdansk-Wschodfrom influent (W-INF) effluent (W-EFF) and wastewa-ter from activated sludge bioreactor (W-BR) and fromGdynia-Debogorze effluent (D-EFF) The two WWTPstreat sewage from approximately 920 000 inhabitants(for details see Figure 1) In the same period of time and
with the same frequency marine outfalls of those WWTPs(W-MOut and D-MOut respectively) as well as VistulaRiver mouth were also sampled for isolation of entero-cocci Marine samples were taken at the depth of approxi-mately one meter below the surface once a month inApril June August and October of 2011 Location ofsampling points and their characteristics are provided inFigure 1 Enterococci were detected using the membranefiltration method To this end appropriate dilutions ofanalyzed samples were filtered in duplicate through045 μm cellulose-acetate filters (EMD Millipore Cor-poration Billerica MA USA) which were then placedon Enterococcus selective agar (Merck DarmstadtGermany) and incubated at 37degC for 48 h (ISO 7899ndash22000) Dark red or maroon colonies were consideredpresumptive enterococci For further investigationsrepresentative isolates were taken from membranes pre-senting from 20 to 50 typical colonies and stored in nu-trient broth supplemented with 15 glycerol at minus80degC
Species identification DNA isolation susceptibility testingand statistical analysisThe species identification and drug susceptibility of pre-sumptive enterococci were determined by the PhoenixtradeAutomated Microbiology System (BD New Jersey USA)according to the manufacturerrsquos instructions Since it isnot possible to distinguish E casseliflavus and E galli-narum using this system the classification E casselifla-vusgallinarum was used The susceptibility tests basedon microdilution were carried out for 11 antimicrobialagents gentamicin (GM) streptomycin (ST) ampicillin(AM) daptomycin (DAP) vancomycin (VA) teicoplanin(TEC) linezolid (LZD) fluoroquinolones (ciprofloxacin -CIP levofloxacin - LVX) tetracycline (TE) erythromycin(E) and quinupristin-dalfopristin (SYN) (E faecium only)Obtained minimal inhibitory concentrations (MIC) wereevaluated according to the guidelines of Clinical and La-boratory Standards Institute [47] Isolates were defined asmultidrug-resistant (MDR) when they showed resistanceto three or more compounds tested [48] Bacterial DNAwas purified using the Genomic DNA Prep Plus kitfollowing the manufacturerrsquos instructions (AampA Biotech-nology Gdynia Poland) Differences in distributions wereevaluated using the χ2 test with the p values lt 005 con-sidered significant
MLST and MLVA of E faeciumMLST for E faecium was performed by sequencing ofseven house-keeping genes as described by Homan et al[49] The allele numbers and STs were assigned with theuse of Internet database httpefaeciummlstnet (5th July2013 date last accessed) Novel alleles and allelic profileswere submitted to the database MLST data were analyzedwith Sequence Type Analysis and Recombinational Tests
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 13 of 15httpwwwbiomedcentralcom1471-21801466
(START) software [50] and the eBURST analysis [51] wasused to elucidate the relationships of STs with knownCCs using the comparative eBURST option available athttpeburstmlstnetv3enter_datacomparative (5th July2013 date last accessed) Further inclusion of E faeciumSTs into specific CCs and lineages was performed fol-lowing other studies [924] MLVA was performed as de-scribed by others [52]
Detection and analysis of genes associated with virulenceantimicrobial resistance and MGE in E faeciumThe full list of primers used for gene detection and se-quencing is provided in Additional file 1 Table S1 PCR-based detection of IS16 virulence-associated genes (fms21fms17 fms5 and fms19 representing each of four pilingene clusters in E faecium and the espEfm gene) genes de-termining resistance to tetracycline tet(M) tet(O) tet(L)tet(S) HLGR (aac(6prime)-Ie-aph(2Prime) HLSR (ant(6prime)-Ia) andgenes specific for MGE such as intA of ICEEfm1 int ofTn916 transposon tndX of Tn5397 transposon entero-coccal plasmid replication genes rep1pIP501 rep2pRE25rep17pRUM rep18pEF418 reppMG1 reppLG1 and TAS genesaxe-txe ω-ε-ζ relBE was performed as described in otherstudies [134153-59] following the conditions describedherein Previously characterized clinical isolates of Strepto-coccus agalactiae E faecium and E faecalis harbouringpilin genes IS16 espEfm intA of ICEEfm1 int of Tn916tndX of Tn5397 tet(M) tet(O) tet(L) tet(S) aac(6prime)-Ie-aph(2Prime) ant(6prime)-Ia rep1pIP501 rep2pRE25 ω-ε-ζ [136061]served as positive controls In cases when positive controlswere not available (rep18pEF418 reppMG1 reppLG1 axe-txerelBE) a few randomly selected PCR products wereverified by sequencing and served further as controlsAmpicillin-resistance determinant the pbp5 gene was ana-lysed by sequencing of the part encoding C-terminaltranspeptidase domain in PBP5 [62] New sequence vari-ants pbp5-24 to pbp5-29 alleles were submitted to theGenBank (acc numbers KC594860-KC594865 respect-ively) The quinolone-determining regions (QRDR) in thegyrA and parC genes were sequenced and analysed asdescribed by others [63]
Additional file
Additional file 1 Table S1 Primers used in the study for PCR andsequencing
Competing interestThe authors declare that they have no competing interest
Authorsrsquo contributionsES participated in the study design performed experimental work andparticipated in the manuscript preparation AL designed the studyperformed experimental work and participated in the manuscriptpreparation Both authors read and approved the final manuscript
AcknowledgementsWe thank Rob Willems and Janetta Top for assigning new alleles new STsand MTs This publication made use of the Enterococcus faecium MLSTwebsite (httpefaeciummlstnet) which is hosted at Imperial College of theUniversity of Oxford and funded by the Wellcome Trust This work wassupported by project N N305 461139 from the Narodowe Centrum Nauki(NCN)
Author details1National Medicines Institute ul Chelmska 3034 Warsaw 00-725 Poland2Gdansk University of Technology ul G Narutowicza 1112 Gdansk 80-952Poland
Received 5 September 2013 Accepted 27 February 2014Published 14 March 2014
References1 Aarestrup FM Butaye P Witte W Nonhuman reservoirs of Enterococci In
American Society for Microbiology Edited by Gilmore MS Clewell DB CourvalinP Dunny GM Murray BE Rice LB Washington DC The Enterococcipathogenesis molecular biology and antibiotic resistance 2002
2 Tendolkar PM Baghdayan AS Shankar N Pathogenic enterococci newdevelopments in the 21st century Cell Mol Life Sci 2003 602622ndash2636
3 Sood S Malhotra M Das BK Kapil A Enterococcal infections ampantimicrobial resistance Indian J Med Res 2008 128111ndash121
4 Sydnor ER Perl TM Hospital epidemiology and infection control inacute-care settings Clin Microbiol Rev 2011 24141ndash173
5 European Centre for Disease Prevention and Control Annual EpidemiologicalReport Reporting on 2010 surveillance data and 2011 epidemic intelligencedata Stockholm ECDC 20122013
6 Hidron AI Edwards JR Patel J Horan TC Sievert DM Pollock DA Fridkin SKNational Healthcare Safety Network Team Participating National HealthcareSafety Network Facilities NHSN annual update antimicrobial-resistantpathogens associated with healthcare-associated infections annualsummary of data reported to the National Healthcare Safety Network atthe Centers for Disease Control and Prevention 2006ndash2007 Infect ControlHosp Epidemiol 2008 29996ndash1011
7 De Kraker ME Jarlier V Monen JC Heuer OE van de Sande N GrundmannH The changing epidemiology of bacteraemias in Europe trends fromthe European Antimicrobial Resistance Surveillance System Clin MicrobiolInfect 2012 doi1011111469-0691
8 Leavis HL Bonten MJ Willems RJ Identification of high-risk enterococcalclonal complexes global dispersion and antibiotic resistance Curr OpinMicrobiol 2006 9454ndash460
9 Willems RJL Top J van Schaik W Leavis H Bonten M Sireacuten J Hanage WPCorander J Restricted gene flow among hospital subpopulations ofEnterococcus faecium mBio 2012 3e00151ndash12
10 Hegstad K Mikalsen T Coque TM Werner G Sundsfjord A Mobile geneticelements and their contribution to the emergence of antimicrobialresistant Enterococcus faecalis and Enterococcus faecium Clin MicrobiolInfect 2010 16541ndash554
11 Shepard BD Gilmore MS Antibiotic-resistant enterococci themechanisms and dynamics of drug introduction and resistance MicrobesInfect 2002 4215ndash224
12 Bonten MJ Willems R Weinstein RA Vancomycin-resistant enterococciwhy are they here and where do they come from Lancet Infect Dis 20011314ndash325
13 Sadowy E Sieńko A Gawryszewska I Bojarska A Malinowska K HryniewiczW High abundance and diversity of antimicrobial resistancedeterminants among early vancomycin-resistant Enterococcus faecium inPoland Eur J Clin Microbiol Infect Dis 2013 321193ndash1203
14 Willems RJ Homan W Top J van Santen-Verheuvel M Tribe D Manzioros XGaillard C Vandenbroucke-Grauls CM Mascini EM van Kregten E vanEmbden JD Bonten MJ Variant esp gene as a marker of a distinct geneticlineage of vancomycin-resistant Enterococcus faecium spreading inhospitals Lancet 2001 357853ndash855
15 Sillanpaumlauml J Nallapareddy SR Prakash VP Qin X Houmloumlk M Weinstock GMMurray BE Identification and phenotypic characterization of a secondcollagen adhesin Scm and genome-based identification and analysis of13 other predicted MSCRAMMs including four distinct pilus loci inEnterococcus faecium Microbiology 2008 1543199ndash3211
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 14 of 15httpwwwbiomedcentralcom1471-21801466
16 Galloway-Pentildea JR Nallapareddy SR Arias CA Eliopoulos GM Murray BEAnalysis of clonality and antibiotic resistance among early clinicalisolates of Enterococcus faecium in the United States J Infect Dis 20092001566ndash1573
17 Top J Willems R Bonten M Emergence of CC17 Enterococcus faeciumfrom commensal to hospital-adapted pathogen FEMS Immunol MedMicrobiol 2008 52297ndash308
18 Martins da Costa P Vaz-Pires P Bernardo F Antimicrobial resistance inEnterococcus spp isolated in inflow effluent and sludge from municipalsewage water treatment plants Water Res 2006 401735ndash1740
19 Reinthaler FF Posch J Feierl G Wust G Haas D Ruckenbauer G Mascher FMarth E Antibiotic resistance of Escherichia coli in sewage and sludgeWater Res 2003 371685ndash1690
20 Luczkiewicz A Fudala-Ksiazek S Jankowska K Olanczuk-Neyman KAntimicrobial resistance of fecal indicators in municipal wastewatertreatment plant Water Res 2010 445089ndash5097
21 Dulfour AP Ballentine RK Ambient water quality criteria for bacteria-1986 EPA4405-84-002 Washington DC US Environmental Protection Agency 1986
22 Ferreira da Silva M Tiago I Veriacutessimo A Boaventura RA Nunes OC Manaia CMAntimicrobial resistance patterns in Enterobacteriaceae isolated from anurban wastewater treatment plant FEMS Microbiol Ecol 2007 60166ndash176
23 DIRECTIVE 20067EC of the European Parliament and of the Council 15February 2006 concerning the management of bathing water quality andrepealing Directive 76160EEC httpeur-lexeuropaeuLexUriServLexUriServdouri=OJL200606400370051ENPDF
24 Freitas AR Novais C Ruiz-Garbajosa P Coque TM Peixe L Dispersion ofmultidrug-resistant Enterococcus faecium isolates belonging to majorclonal complexes in different Portuguese settings Appl Environ Microbiol2009 754904ndash4908
25 Poeta P Costa D Igrejas G Saacuteenz Y Zarazaga M Rodrigues J Torres CPolymorphisms of the pbp5 gene and correlation with ampicillinresistance in Enterococcus faecium isolates of animal origin J MedMicrobiol 2007 56236ndash240
26 Kuumlhn I Iversen A Burman LG Olsson-Liljequist B Franklin A Finn MAarestrup F Seyfarth AM Blanch AR Vilanova X Taylor H Caplin J MorenoMA Dominguez L Herrero IA Moumlllby R Comparison of enterococcalpopulations in animals humans and the environment ndash a Europeanstudy Int J Food Microbiol 2003 88133ndash145
27 Lanthier M Scott A Lapen DR Zhang Y Topp E Frequency of virulencegenes and antibiotic resistances in Enterococcus spp isolates fromwastewater and feces of domesticated mammals and birds and wildlifeCan J Microbiol 2010 56715ndash729
28 Leclercq R Oberleacute K Galopin S Cattoir V Budzinski H Petit F Changes inenterococcal populations and related antibiotic resistance along amedical center-wastewater treatment plant-river continuum Appl EnvironMicrobiol 2013 792428ndash2434
29 Thevenon F Regier N Benagli C Tonolla M Adatte T Wildi W Poteacute JCharacterization of fecal indicator bacteria in sediments cores from thelargest freshwater lake of Western Europe (Lake Geneva Switzerland)Ecotoxicol Environ Safety 2012 7850ndash56
30 Bonilla TD Nowosielski K Esiobu N McCorquodale DS Rogerson ASpecies assemblages of Enterococcus indicate potential sources offecal bacteria at a south Florida recreational beach Mar Pollut Bull2006 52807ndash810
31 Wang JT Chang SC Wang HY Chen PC Shiau YR Lauderdale TL TSARHospitals High rates of multidrug resistance in Enterococcus faecalis andE faecium isolated from inpatients and outpatients in TaiwanDiagn Microbiol Infect Dis 2013 75406ndash411
32 Lanthier M Scott A Zhang Y Cloutier M Durie D Henderson VC WilkesG Lapen DR Topp E Distribution of selected virulence genes andantibiotic resistance in Enterococcus species isolated from the SouthNation River drainage basin Ontario Canada J Appl Microbiol 2011110407ndash421
33 Kuch A Willems RJ Werner G Coque TM Hammerum AM Sundsfjord AKlare I Ruiz-Garbajosa P Simonsen GS van Luit-Asbroek M Hryniewicz WSadowy E Insight into antimicrobial susceptibility and population struc-ture of contemporary human Enterococcus faecalis isolates fromEurope J Antimicrob Chemother 2012 67551ndash558
34 Novais C Coque TM Ferreira H Sousa JC Peixe L Environmentalcontamination with vancomycin-resistant enterococci from hospitalsewage in Portugal Appl Environ Microbiol 2005 713364ndash3368
35 Iversen A Kuumlhn I Rahman M Franklin A Burman LG Olsson-Liljequist BTorell E Moumlllby R Evidence for transmission between humans and theenvironment of a nosocomial strain of Enterococcus faeciumEnviron Microbiol 2004 655ndash59
36 Rathnayake I Hargreaves M Huygens F SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterwayAustralia and associated antibiotic resistance gene profilesBMC Microbiol 2011 11201
37 Rosvoll TC Pedersen T Sletvold H Johnsen PJ Sollid JE Simonsen GSJensen LB Nielsen KM Sundsfjord A PCR-based plasmid typing inEnterococcus faecium strains reveals widely distributed pRE25-pRUM- pIP501- and pHT-related replicons associated with glycopeptideresistance and stabilizing toxin-antitoxin systems FEMS Immunol MedMicrobiol 2010 58254ndash268
38 Rosvoll TC Lindstad BL Lunde TM Hegstad K Aasnaes B Hammerum AMLester CH Simonsen GS Sundsfjord A Pedersen T Increased high-levelgentamicin resistance in invasive Enterococcus faecium is associated withaac(6prime)Ie-aph(2Prime)Ia-encoding transferable megaplasmids hosted by majorhospital-adapted lineages FEMS Immunol Med Microbiol 2012 66166ndash176
39 Laverde Gomez JA van Schaik W Freitas AR Coque TM Weaver KE FranciaMV Witte W Werner G A multiresistance megaplasmid pLG1 bearing ahylEfm genomic island in hospital Enterococcus faecium isolates Int J MedMicrobiol 2011 301165ndash175
40 Coque TM Evolutionary biology of pathogenic enterococci InEvolutionary Biology of Bacterial and Fungal Pathogens Coque TM Edited byBaquero F Nombela C Cassell GH Gutierrez JA Washington DC ASMPress501ndash523
41 Werner G Fleige C Geringer U van Schaik W Klare I Witte W IS elementIS16 as a molecular screening tool to identify hospital-associated strainsof Enterococcus faecium BMC Infect Dis 2011 1180
42 Top J Sinnige JC Majoor EA Bonten MJ Willems RJ van Schaik W Therecombinase IntA is required for excision of esp-containing ICEEfm1 inEnterococcus faecium J Bacteriol 2011 1931003ndash1006
43 Sillanpaumlauml J Prakash VP Nallapareddy SR Murray BE Distribution of genesencoding MSCRAMMs and Pili in clinical and natural populations ofEnterococcus faecium J Clin Microbiol 2009 47896ndash901
44 Marcinek H Wirth R Muscholl-Silberhorn A Gauer M Enterococcus faecalisgene transfer under natural conditions in municipal sewage watertreatment plants Appl Environ Microbiol 1998 64626ndash632
45 Kuumlmmerer K Significance of antibiotics in the environment J AntimicrobChemother 2003 525ndash7
46 Liu A Fong A Becket E Yuan J Tamae C Medrano L Maiz M Wahba C LeeC Lee K Tran KP Yang H Hoffman RM Salih A Miller JHP Selectiveadvantage of resistant strains at trace levels of antibiotics a simple andultrasensitive color test for detection of antibiotics and genotoxicagents Antimicrob Agents Chemother 2011 551204ndash1210
47 Clinical and Laboratory Standards Institute Performance standards forantimicrobial susceptibility testing 21st informational supplement Wayne CLSI2011 PA M100-S20
48 Magiorakos AP Srinivasan A Carey RB Carmeli Y Falagas ME Giske CGHarbarth S Hindler JF Kahlmeter G Olsson-Liljequist B Paterson DL Rice LBStelling J Struelens MJ Vatopoulos A Weber JT Monnet DLMultidrug-resistant extensively drug-resistant and pandrug-resistantbacteria an international expert proposal for interim standarddefinitions for acquired resistance Clin Microbiol Infect 2012 18268ndash281
49 Homan WL Tribe D Poznanski S Li M Hogg G Spalburg E Van Embden JDWillems RJ Multilocus sequence typing scheme for Enterococcus faeciumJ Clin Microbiol 2002 401963ndash1971
50 Jolley KA Feil EJ Chan MS Maiden MC Sequence type analysis andrecombinational tests (START) Bioinformatics 2001 171230ndash1231
51 Feil EJ Li BC Aanensen DM Hanage WP Spratt BG eBURST inferringpatterns of evolutionary descent among clusters of related bacterialgenotypes from multilocus sequence typing data J Bacteriol 20041861518ndash1530
52 Top J Banga1 NMI Hayes R Willems RJ Bonten MJM Hayden MKComparison of multiple-locus variable-number tandem repeat analysisand pulsed-field gel electrophoresis in a setting of polyclonal endemicityof vancomycin-resistant Enterococcus faecium Clin Microbiol Infect 200814363ndash369
53 Vankerckhoven V Van Autgaerden T Vael C Lammens C Chapelle S RossiR Jabes D Goossens H Development of a multiplex PCR for the
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-

Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 12 of 15httpwwwbiomedcentralcom1471-21801466
accumulate several determinants that may promote theirsurvival not only in the hospital settings but also in theenvironment Such features would include resistance todisinfectants and ultraviolet light additional metabolicpathways and ability to form biofilms Moreover determi-nants of some of the mentioned adaptation factors arefound co-resident in plasmids harboring resistance genessuggesting co-selection For example pLG1-type plasmidsof E faecium carry not only resistance determinantsagainst aminoglycosides glycopeptides and macrolidesbut also pili genes and carbon uptake-utilization genes[3839] Pheromone-responsive plasmids of E faecalisapart from resistance to aminoglycosides glycopeptidespenicillins and macrolides may encode bacteriocins add-itional pili and UV-resistance determinants [40] Thusnosocomial HiRECCs released by WWTPs are equippedin whole sets of genes with various adaptive benefits andoften located within MGE It was also demonstrated thatMGEs can be transferred in a WWTP [44] further under-ling the importance of threat posed by such organismsThere is a selection pressure for survival of resistantstrains and their tranconjugants due to release of antibi-otics to the environment especially in the hospital efflu-ents [45] and even sub-inhibitory concentrations may beeffective in selection for resistance [46]
ConclusionsThis study describes the species composition resistanceprofiles and clonal relationships of enterococci isolatedfrom WWTPs located in Gdansk their marine outfallsto the Gulf of Gdansk and from Vistula River It was ob-served that these WWTPs release enterococcal strainsresistant to various antimicrobial compounds to the re-cipient water environment Molecular analyses revealedthe presence of clones associated with nosocomial HiR-ECCs and in the case of E faecium isolates of lineages17 18 and 78 of former CC17 abundant in resistance de-terminants MGE genes and pathogenicity factors Thepresence of HiRECCs in wastewaters and marineriverenvironment highlights the need for further detailedanalyses to better understand the survival and spread ofdrug-resistant strains in water ecosystems and to eluci-date the ways to curb such dissemination
MethodsAcquisition of samples and isolation of enterococciOnce a month in April June August and October in 2011of the flow proportioned composited samples of wastewa-ter were taken from two local WWTPs Gdansk-Wschodfrom influent (W-INF) effluent (W-EFF) and wastewa-ter from activated sludge bioreactor (W-BR) and fromGdynia-Debogorze effluent (D-EFF) The two WWTPstreat sewage from approximately 920 000 inhabitants(for details see Figure 1) In the same period of time and
with the same frequency marine outfalls of those WWTPs(W-MOut and D-MOut respectively) as well as VistulaRiver mouth were also sampled for isolation of entero-cocci Marine samples were taken at the depth of approxi-mately one meter below the surface once a month inApril June August and October of 2011 Location ofsampling points and their characteristics are provided inFigure 1 Enterococci were detected using the membranefiltration method To this end appropriate dilutions ofanalyzed samples were filtered in duplicate through045 μm cellulose-acetate filters (EMD Millipore Cor-poration Billerica MA USA) which were then placedon Enterococcus selective agar (Merck DarmstadtGermany) and incubated at 37degC for 48 h (ISO 7899ndash22000) Dark red or maroon colonies were consideredpresumptive enterococci For further investigationsrepresentative isolates were taken from membranes pre-senting from 20 to 50 typical colonies and stored in nu-trient broth supplemented with 15 glycerol at minus80degC
Species identification DNA isolation susceptibility testingand statistical analysisThe species identification and drug susceptibility of pre-sumptive enterococci were determined by the PhoenixtradeAutomated Microbiology System (BD New Jersey USA)according to the manufacturerrsquos instructions Since it isnot possible to distinguish E casseliflavus and E galli-narum using this system the classification E casselifla-vusgallinarum was used The susceptibility tests basedon microdilution were carried out for 11 antimicrobialagents gentamicin (GM) streptomycin (ST) ampicillin(AM) daptomycin (DAP) vancomycin (VA) teicoplanin(TEC) linezolid (LZD) fluoroquinolones (ciprofloxacin -CIP levofloxacin - LVX) tetracycline (TE) erythromycin(E) and quinupristin-dalfopristin (SYN) (E faecium only)Obtained minimal inhibitory concentrations (MIC) wereevaluated according to the guidelines of Clinical and La-boratory Standards Institute [47] Isolates were defined asmultidrug-resistant (MDR) when they showed resistanceto three or more compounds tested [48] Bacterial DNAwas purified using the Genomic DNA Prep Plus kitfollowing the manufacturerrsquos instructions (AampA Biotech-nology Gdynia Poland) Differences in distributions wereevaluated using the χ2 test with the p values lt 005 con-sidered significant
MLST and MLVA of E faeciumMLST for E faecium was performed by sequencing ofseven house-keeping genes as described by Homan et al[49] The allele numbers and STs were assigned with theuse of Internet database httpefaeciummlstnet (5th July2013 date last accessed) Novel alleles and allelic profileswere submitted to the database MLST data were analyzedwith Sequence Type Analysis and Recombinational Tests
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 13 of 15httpwwwbiomedcentralcom1471-21801466
(START) software [50] and the eBURST analysis [51] wasused to elucidate the relationships of STs with knownCCs using the comparative eBURST option available athttpeburstmlstnetv3enter_datacomparative (5th July2013 date last accessed) Further inclusion of E faeciumSTs into specific CCs and lineages was performed fol-lowing other studies [924] MLVA was performed as de-scribed by others [52]
Detection and analysis of genes associated with virulenceantimicrobial resistance and MGE in E faeciumThe full list of primers used for gene detection and se-quencing is provided in Additional file 1 Table S1 PCR-based detection of IS16 virulence-associated genes (fms21fms17 fms5 and fms19 representing each of four pilingene clusters in E faecium and the espEfm gene) genes de-termining resistance to tetracycline tet(M) tet(O) tet(L)tet(S) HLGR (aac(6prime)-Ie-aph(2Prime) HLSR (ant(6prime)-Ia) andgenes specific for MGE such as intA of ICEEfm1 int ofTn916 transposon tndX of Tn5397 transposon entero-coccal plasmid replication genes rep1pIP501 rep2pRE25rep17pRUM rep18pEF418 reppMG1 reppLG1 and TAS genesaxe-txe ω-ε-ζ relBE was performed as described in otherstudies [134153-59] following the conditions describedherein Previously characterized clinical isolates of Strepto-coccus agalactiae E faecium and E faecalis harbouringpilin genes IS16 espEfm intA of ICEEfm1 int of Tn916tndX of Tn5397 tet(M) tet(O) tet(L) tet(S) aac(6prime)-Ie-aph(2Prime) ant(6prime)-Ia rep1pIP501 rep2pRE25 ω-ε-ζ [136061]served as positive controls In cases when positive controlswere not available (rep18pEF418 reppMG1 reppLG1 axe-txerelBE) a few randomly selected PCR products wereverified by sequencing and served further as controlsAmpicillin-resistance determinant the pbp5 gene was ana-lysed by sequencing of the part encoding C-terminaltranspeptidase domain in PBP5 [62] New sequence vari-ants pbp5-24 to pbp5-29 alleles were submitted to theGenBank (acc numbers KC594860-KC594865 respect-ively) The quinolone-determining regions (QRDR) in thegyrA and parC genes were sequenced and analysed asdescribed by others [63]
Additional file
Additional file 1 Table S1 Primers used in the study for PCR andsequencing
Competing interestThe authors declare that they have no competing interest
Authorsrsquo contributionsES participated in the study design performed experimental work andparticipated in the manuscript preparation AL designed the studyperformed experimental work and participated in the manuscriptpreparation Both authors read and approved the final manuscript
AcknowledgementsWe thank Rob Willems and Janetta Top for assigning new alleles new STsand MTs This publication made use of the Enterococcus faecium MLSTwebsite (httpefaeciummlstnet) which is hosted at Imperial College of theUniversity of Oxford and funded by the Wellcome Trust This work wassupported by project N N305 461139 from the Narodowe Centrum Nauki(NCN)
Author details1National Medicines Institute ul Chelmska 3034 Warsaw 00-725 Poland2Gdansk University of Technology ul G Narutowicza 1112 Gdansk 80-952Poland
Received 5 September 2013 Accepted 27 February 2014Published 14 March 2014
References1 Aarestrup FM Butaye P Witte W Nonhuman reservoirs of Enterococci In
American Society for Microbiology Edited by Gilmore MS Clewell DB CourvalinP Dunny GM Murray BE Rice LB Washington DC The Enterococcipathogenesis molecular biology and antibiotic resistance 2002
2 Tendolkar PM Baghdayan AS Shankar N Pathogenic enterococci newdevelopments in the 21st century Cell Mol Life Sci 2003 602622ndash2636
3 Sood S Malhotra M Das BK Kapil A Enterococcal infections ampantimicrobial resistance Indian J Med Res 2008 128111ndash121
4 Sydnor ER Perl TM Hospital epidemiology and infection control inacute-care settings Clin Microbiol Rev 2011 24141ndash173
5 European Centre for Disease Prevention and Control Annual EpidemiologicalReport Reporting on 2010 surveillance data and 2011 epidemic intelligencedata Stockholm ECDC 20122013
6 Hidron AI Edwards JR Patel J Horan TC Sievert DM Pollock DA Fridkin SKNational Healthcare Safety Network Team Participating National HealthcareSafety Network Facilities NHSN annual update antimicrobial-resistantpathogens associated with healthcare-associated infections annualsummary of data reported to the National Healthcare Safety Network atthe Centers for Disease Control and Prevention 2006ndash2007 Infect ControlHosp Epidemiol 2008 29996ndash1011
7 De Kraker ME Jarlier V Monen JC Heuer OE van de Sande N GrundmannH The changing epidemiology of bacteraemias in Europe trends fromthe European Antimicrobial Resistance Surveillance System Clin MicrobiolInfect 2012 doi1011111469-0691
8 Leavis HL Bonten MJ Willems RJ Identification of high-risk enterococcalclonal complexes global dispersion and antibiotic resistance Curr OpinMicrobiol 2006 9454ndash460
9 Willems RJL Top J van Schaik W Leavis H Bonten M Sireacuten J Hanage WPCorander J Restricted gene flow among hospital subpopulations ofEnterococcus faecium mBio 2012 3e00151ndash12
10 Hegstad K Mikalsen T Coque TM Werner G Sundsfjord A Mobile geneticelements and their contribution to the emergence of antimicrobialresistant Enterococcus faecalis and Enterococcus faecium Clin MicrobiolInfect 2010 16541ndash554
11 Shepard BD Gilmore MS Antibiotic-resistant enterococci themechanisms and dynamics of drug introduction and resistance MicrobesInfect 2002 4215ndash224
12 Bonten MJ Willems R Weinstein RA Vancomycin-resistant enterococciwhy are they here and where do they come from Lancet Infect Dis 20011314ndash325
13 Sadowy E Sieńko A Gawryszewska I Bojarska A Malinowska K HryniewiczW High abundance and diversity of antimicrobial resistancedeterminants among early vancomycin-resistant Enterococcus faecium inPoland Eur J Clin Microbiol Infect Dis 2013 321193ndash1203
14 Willems RJ Homan W Top J van Santen-Verheuvel M Tribe D Manzioros XGaillard C Vandenbroucke-Grauls CM Mascini EM van Kregten E vanEmbden JD Bonten MJ Variant esp gene as a marker of a distinct geneticlineage of vancomycin-resistant Enterococcus faecium spreading inhospitals Lancet 2001 357853ndash855
15 Sillanpaumlauml J Nallapareddy SR Prakash VP Qin X Houmloumlk M Weinstock GMMurray BE Identification and phenotypic characterization of a secondcollagen adhesin Scm and genome-based identification and analysis of13 other predicted MSCRAMMs including four distinct pilus loci inEnterococcus faecium Microbiology 2008 1543199ndash3211
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 14 of 15httpwwwbiomedcentralcom1471-21801466
16 Galloway-Pentildea JR Nallapareddy SR Arias CA Eliopoulos GM Murray BEAnalysis of clonality and antibiotic resistance among early clinicalisolates of Enterococcus faecium in the United States J Infect Dis 20092001566ndash1573
17 Top J Willems R Bonten M Emergence of CC17 Enterococcus faeciumfrom commensal to hospital-adapted pathogen FEMS Immunol MedMicrobiol 2008 52297ndash308
18 Martins da Costa P Vaz-Pires P Bernardo F Antimicrobial resistance inEnterococcus spp isolated in inflow effluent and sludge from municipalsewage water treatment plants Water Res 2006 401735ndash1740
19 Reinthaler FF Posch J Feierl G Wust G Haas D Ruckenbauer G Mascher FMarth E Antibiotic resistance of Escherichia coli in sewage and sludgeWater Res 2003 371685ndash1690
20 Luczkiewicz A Fudala-Ksiazek S Jankowska K Olanczuk-Neyman KAntimicrobial resistance of fecal indicators in municipal wastewatertreatment plant Water Res 2010 445089ndash5097
21 Dulfour AP Ballentine RK Ambient water quality criteria for bacteria-1986 EPA4405-84-002 Washington DC US Environmental Protection Agency 1986
22 Ferreira da Silva M Tiago I Veriacutessimo A Boaventura RA Nunes OC Manaia CMAntimicrobial resistance patterns in Enterobacteriaceae isolated from anurban wastewater treatment plant FEMS Microbiol Ecol 2007 60166ndash176
23 DIRECTIVE 20067EC of the European Parliament and of the Council 15February 2006 concerning the management of bathing water quality andrepealing Directive 76160EEC httpeur-lexeuropaeuLexUriServLexUriServdouri=OJL200606400370051ENPDF
24 Freitas AR Novais C Ruiz-Garbajosa P Coque TM Peixe L Dispersion ofmultidrug-resistant Enterococcus faecium isolates belonging to majorclonal complexes in different Portuguese settings Appl Environ Microbiol2009 754904ndash4908
25 Poeta P Costa D Igrejas G Saacuteenz Y Zarazaga M Rodrigues J Torres CPolymorphisms of the pbp5 gene and correlation with ampicillinresistance in Enterococcus faecium isolates of animal origin J MedMicrobiol 2007 56236ndash240
26 Kuumlhn I Iversen A Burman LG Olsson-Liljequist B Franklin A Finn MAarestrup F Seyfarth AM Blanch AR Vilanova X Taylor H Caplin J MorenoMA Dominguez L Herrero IA Moumlllby R Comparison of enterococcalpopulations in animals humans and the environment ndash a Europeanstudy Int J Food Microbiol 2003 88133ndash145
27 Lanthier M Scott A Lapen DR Zhang Y Topp E Frequency of virulencegenes and antibiotic resistances in Enterococcus spp isolates fromwastewater and feces of domesticated mammals and birds and wildlifeCan J Microbiol 2010 56715ndash729
28 Leclercq R Oberleacute K Galopin S Cattoir V Budzinski H Petit F Changes inenterococcal populations and related antibiotic resistance along amedical center-wastewater treatment plant-river continuum Appl EnvironMicrobiol 2013 792428ndash2434
29 Thevenon F Regier N Benagli C Tonolla M Adatte T Wildi W Poteacute JCharacterization of fecal indicator bacteria in sediments cores from thelargest freshwater lake of Western Europe (Lake Geneva Switzerland)Ecotoxicol Environ Safety 2012 7850ndash56
30 Bonilla TD Nowosielski K Esiobu N McCorquodale DS Rogerson ASpecies assemblages of Enterococcus indicate potential sources offecal bacteria at a south Florida recreational beach Mar Pollut Bull2006 52807ndash810
31 Wang JT Chang SC Wang HY Chen PC Shiau YR Lauderdale TL TSARHospitals High rates of multidrug resistance in Enterococcus faecalis andE faecium isolated from inpatients and outpatients in TaiwanDiagn Microbiol Infect Dis 2013 75406ndash411
32 Lanthier M Scott A Zhang Y Cloutier M Durie D Henderson VC WilkesG Lapen DR Topp E Distribution of selected virulence genes andantibiotic resistance in Enterococcus species isolated from the SouthNation River drainage basin Ontario Canada J Appl Microbiol 2011110407ndash421
33 Kuch A Willems RJ Werner G Coque TM Hammerum AM Sundsfjord AKlare I Ruiz-Garbajosa P Simonsen GS van Luit-Asbroek M Hryniewicz WSadowy E Insight into antimicrobial susceptibility and population struc-ture of contemporary human Enterococcus faecalis isolates fromEurope J Antimicrob Chemother 2012 67551ndash558
34 Novais C Coque TM Ferreira H Sousa JC Peixe L Environmentalcontamination with vancomycin-resistant enterococci from hospitalsewage in Portugal Appl Environ Microbiol 2005 713364ndash3368
35 Iversen A Kuumlhn I Rahman M Franklin A Burman LG Olsson-Liljequist BTorell E Moumlllby R Evidence for transmission between humans and theenvironment of a nosocomial strain of Enterococcus faeciumEnviron Microbiol 2004 655ndash59
36 Rathnayake I Hargreaves M Huygens F SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterwayAustralia and associated antibiotic resistance gene profilesBMC Microbiol 2011 11201
37 Rosvoll TC Pedersen T Sletvold H Johnsen PJ Sollid JE Simonsen GSJensen LB Nielsen KM Sundsfjord A PCR-based plasmid typing inEnterococcus faecium strains reveals widely distributed pRE25-pRUM- pIP501- and pHT-related replicons associated with glycopeptideresistance and stabilizing toxin-antitoxin systems FEMS Immunol MedMicrobiol 2010 58254ndash268
38 Rosvoll TC Lindstad BL Lunde TM Hegstad K Aasnaes B Hammerum AMLester CH Simonsen GS Sundsfjord A Pedersen T Increased high-levelgentamicin resistance in invasive Enterococcus faecium is associated withaac(6prime)Ie-aph(2Prime)Ia-encoding transferable megaplasmids hosted by majorhospital-adapted lineages FEMS Immunol Med Microbiol 2012 66166ndash176
39 Laverde Gomez JA van Schaik W Freitas AR Coque TM Weaver KE FranciaMV Witte W Werner G A multiresistance megaplasmid pLG1 bearing ahylEfm genomic island in hospital Enterococcus faecium isolates Int J MedMicrobiol 2011 301165ndash175
40 Coque TM Evolutionary biology of pathogenic enterococci InEvolutionary Biology of Bacterial and Fungal Pathogens Coque TM Edited byBaquero F Nombela C Cassell GH Gutierrez JA Washington DC ASMPress501ndash523
41 Werner G Fleige C Geringer U van Schaik W Klare I Witte W IS elementIS16 as a molecular screening tool to identify hospital-associated strainsof Enterococcus faecium BMC Infect Dis 2011 1180
42 Top J Sinnige JC Majoor EA Bonten MJ Willems RJ van Schaik W Therecombinase IntA is required for excision of esp-containing ICEEfm1 inEnterococcus faecium J Bacteriol 2011 1931003ndash1006
43 Sillanpaumlauml J Prakash VP Nallapareddy SR Murray BE Distribution of genesencoding MSCRAMMs and Pili in clinical and natural populations ofEnterococcus faecium J Clin Microbiol 2009 47896ndash901
44 Marcinek H Wirth R Muscholl-Silberhorn A Gauer M Enterococcus faecalisgene transfer under natural conditions in municipal sewage watertreatment plants Appl Environ Microbiol 1998 64626ndash632
45 Kuumlmmerer K Significance of antibiotics in the environment J AntimicrobChemother 2003 525ndash7
46 Liu A Fong A Becket E Yuan J Tamae C Medrano L Maiz M Wahba C LeeC Lee K Tran KP Yang H Hoffman RM Salih A Miller JHP Selectiveadvantage of resistant strains at trace levels of antibiotics a simple andultrasensitive color test for detection of antibiotics and genotoxicagents Antimicrob Agents Chemother 2011 551204ndash1210
47 Clinical and Laboratory Standards Institute Performance standards forantimicrobial susceptibility testing 21st informational supplement Wayne CLSI2011 PA M100-S20
48 Magiorakos AP Srinivasan A Carey RB Carmeli Y Falagas ME Giske CGHarbarth S Hindler JF Kahlmeter G Olsson-Liljequist B Paterson DL Rice LBStelling J Struelens MJ Vatopoulos A Weber JT Monnet DLMultidrug-resistant extensively drug-resistant and pandrug-resistantbacteria an international expert proposal for interim standarddefinitions for acquired resistance Clin Microbiol Infect 2012 18268ndash281
49 Homan WL Tribe D Poznanski S Li M Hogg G Spalburg E Van Embden JDWillems RJ Multilocus sequence typing scheme for Enterococcus faeciumJ Clin Microbiol 2002 401963ndash1971
50 Jolley KA Feil EJ Chan MS Maiden MC Sequence type analysis andrecombinational tests (START) Bioinformatics 2001 171230ndash1231
51 Feil EJ Li BC Aanensen DM Hanage WP Spratt BG eBURST inferringpatterns of evolutionary descent among clusters of related bacterialgenotypes from multilocus sequence typing data J Bacteriol 20041861518ndash1530
52 Top J Banga1 NMI Hayes R Willems RJ Bonten MJM Hayden MKComparison of multiple-locus variable-number tandem repeat analysisand pulsed-field gel electrophoresis in a setting of polyclonal endemicityof vancomycin-resistant Enterococcus faecium Clin Microbiol Infect 200814363ndash369
53 Vankerckhoven V Van Autgaerden T Vael C Lammens C Chapelle S RossiR Jabes D Goossens H Development of a multiplex PCR for the
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-

Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 13 of 15httpwwwbiomedcentralcom1471-21801466
(START) software [50] and the eBURST analysis [51] wasused to elucidate the relationships of STs with knownCCs using the comparative eBURST option available athttpeburstmlstnetv3enter_datacomparative (5th July2013 date last accessed) Further inclusion of E faeciumSTs into specific CCs and lineages was performed fol-lowing other studies [924] MLVA was performed as de-scribed by others [52]
Detection and analysis of genes associated with virulenceantimicrobial resistance and MGE in E faeciumThe full list of primers used for gene detection and se-quencing is provided in Additional file 1 Table S1 PCR-based detection of IS16 virulence-associated genes (fms21fms17 fms5 and fms19 representing each of four pilingene clusters in E faecium and the espEfm gene) genes de-termining resistance to tetracycline tet(M) tet(O) tet(L)tet(S) HLGR (aac(6prime)-Ie-aph(2Prime) HLSR (ant(6prime)-Ia) andgenes specific for MGE such as intA of ICEEfm1 int ofTn916 transposon tndX of Tn5397 transposon entero-coccal plasmid replication genes rep1pIP501 rep2pRE25rep17pRUM rep18pEF418 reppMG1 reppLG1 and TAS genesaxe-txe ω-ε-ζ relBE was performed as described in otherstudies [134153-59] following the conditions describedherein Previously characterized clinical isolates of Strepto-coccus agalactiae E faecium and E faecalis harbouringpilin genes IS16 espEfm intA of ICEEfm1 int of Tn916tndX of Tn5397 tet(M) tet(O) tet(L) tet(S) aac(6prime)-Ie-aph(2Prime) ant(6prime)-Ia rep1pIP501 rep2pRE25 ω-ε-ζ [136061]served as positive controls In cases when positive controlswere not available (rep18pEF418 reppMG1 reppLG1 axe-txerelBE) a few randomly selected PCR products wereverified by sequencing and served further as controlsAmpicillin-resistance determinant the pbp5 gene was ana-lysed by sequencing of the part encoding C-terminaltranspeptidase domain in PBP5 [62] New sequence vari-ants pbp5-24 to pbp5-29 alleles were submitted to theGenBank (acc numbers KC594860-KC594865 respect-ively) The quinolone-determining regions (QRDR) in thegyrA and parC genes were sequenced and analysed asdescribed by others [63]
Additional file
Additional file 1 Table S1 Primers used in the study for PCR andsequencing
Competing interestThe authors declare that they have no competing interest
Authorsrsquo contributionsES participated in the study design performed experimental work andparticipated in the manuscript preparation AL designed the studyperformed experimental work and participated in the manuscriptpreparation Both authors read and approved the final manuscript
AcknowledgementsWe thank Rob Willems and Janetta Top for assigning new alleles new STsand MTs This publication made use of the Enterococcus faecium MLSTwebsite (httpefaeciummlstnet) which is hosted at Imperial College of theUniversity of Oxford and funded by the Wellcome Trust This work wassupported by project N N305 461139 from the Narodowe Centrum Nauki(NCN)
Author details1National Medicines Institute ul Chelmska 3034 Warsaw 00-725 Poland2Gdansk University of Technology ul G Narutowicza 1112 Gdansk 80-952Poland
Received 5 September 2013 Accepted 27 February 2014Published 14 March 2014
References1 Aarestrup FM Butaye P Witte W Nonhuman reservoirs of Enterococci In
American Society for Microbiology Edited by Gilmore MS Clewell DB CourvalinP Dunny GM Murray BE Rice LB Washington DC The Enterococcipathogenesis molecular biology and antibiotic resistance 2002
2 Tendolkar PM Baghdayan AS Shankar N Pathogenic enterococci newdevelopments in the 21st century Cell Mol Life Sci 2003 602622ndash2636
3 Sood S Malhotra M Das BK Kapil A Enterococcal infections ampantimicrobial resistance Indian J Med Res 2008 128111ndash121
4 Sydnor ER Perl TM Hospital epidemiology and infection control inacute-care settings Clin Microbiol Rev 2011 24141ndash173
5 European Centre for Disease Prevention and Control Annual EpidemiologicalReport Reporting on 2010 surveillance data and 2011 epidemic intelligencedata Stockholm ECDC 20122013
6 Hidron AI Edwards JR Patel J Horan TC Sievert DM Pollock DA Fridkin SKNational Healthcare Safety Network Team Participating National HealthcareSafety Network Facilities NHSN annual update antimicrobial-resistantpathogens associated with healthcare-associated infections annualsummary of data reported to the National Healthcare Safety Network atthe Centers for Disease Control and Prevention 2006ndash2007 Infect ControlHosp Epidemiol 2008 29996ndash1011
7 De Kraker ME Jarlier V Monen JC Heuer OE van de Sande N GrundmannH The changing epidemiology of bacteraemias in Europe trends fromthe European Antimicrobial Resistance Surveillance System Clin MicrobiolInfect 2012 doi1011111469-0691
8 Leavis HL Bonten MJ Willems RJ Identification of high-risk enterococcalclonal complexes global dispersion and antibiotic resistance Curr OpinMicrobiol 2006 9454ndash460
9 Willems RJL Top J van Schaik W Leavis H Bonten M Sireacuten J Hanage WPCorander J Restricted gene flow among hospital subpopulations ofEnterococcus faecium mBio 2012 3e00151ndash12
10 Hegstad K Mikalsen T Coque TM Werner G Sundsfjord A Mobile geneticelements and their contribution to the emergence of antimicrobialresistant Enterococcus faecalis and Enterococcus faecium Clin MicrobiolInfect 2010 16541ndash554
11 Shepard BD Gilmore MS Antibiotic-resistant enterococci themechanisms and dynamics of drug introduction and resistance MicrobesInfect 2002 4215ndash224
12 Bonten MJ Willems R Weinstein RA Vancomycin-resistant enterococciwhy are they here and where do they come from Lancet Infect Dis 20011314ndash325
13 Sadowy E Sieńko A Gawryszewska I Bojarska A Malinowska K HryniewiczW High abundance and diversity of antimicrobial resistancedeterminants among early vancomycin-resistant Enterococcus faecium inPoland Eur J Clin Microbiol Infect Dis 2013 321193ndash1203
14 Willems RJ Homan W Top J van Santen-Verheuvel M Tribe D Manzioros XGaillard C Vandenbroucke-Grauls CM Mascini EM van Kregten E vanEmbden JD Bonten MJ Variant esp gene as a marker of a distinct geneticlineage of vancomycin-resistant Enterococcus faecium spreading inhospitals Lancet 2001 357853ndash855
15 Sillanpaumlauml J Nallapareddy SR Prakash VP Qin X Houmloumlk M Weinstock GMMurray BE Identification and phenotypic characterization of a secondcollagen adhesin Scm and genome-based identification and analysis of13 other predicted MSCRAMMs including four distinct pilus loci inEnterococcus faecium Microbiology 2008 1543199ndash3211
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 14 of 15httpwwwbiomedcentralcom1471-21801466
16 Galloway-Pentildea JR Nallapareddy SR Arias CA Eliopoulos GM Murray BEAnalysis of clonality and antibiotic resistance among early clinicalisolates of Enterococcus faecium in the United States J Infect Dis 20092001566ndash1573
17 Top J Willems R Bonten M Emergence of CC17 Enterococcus faeciumfrom commensal to hospital-adapted pathogen FEMS Immunol MedMicrobiol 2008 52297ndash308
18 Martins da Costa P Vaz-Pires P Bernardo F Antimicrobial resistance inEnterococcus spp isolated in inflow effluent and sludge from municipalsewage water treatment plants Water Res 2006 401735ndash1740
19 Reinthaler FF Posch J Feierl G Wust G Haas D Ruckenbauer G Mascher FMarth E Antibiotic resistance of Escherichia coli in sewage and sludgeWater Res 2003 371685ndash1690
20 Luczkiewicz A Fudala-Ksiazek S Jankowska K Olanczuk-Neyman KAntimicrobial resistance of fecal indicators in municipal wastewatertreatment plant Water Res 2010 445089ndash5097
21 Dulfour AP Ballentine RK Ambient water quality criteria for bacteria-1986 EPA4405-84-002 Washington DC US Environmental Protection Agency 1986
22 Ferreira da Silva M Tiago I Veriacutessimo A Boaventura RA Nunes OC Manaia CMAntimicrobial resistance patterns in Enterobacteriaceae isolated from anurban wastewater treatment plant FEMS Microbiol Ecol 2007 60166ndash176
23 DIRECTIVE 20067EC of the European Parliament and of the Council 15February 2006 concerning the management of bathing water quality andrepealing Directive 76160EEC httpeur-lexeuropaeuLexUriServLexUriServdouri=OJL200606400370051ENPDF
24 Freitas AR Novais C Ruiz-Garbajosa P Coque TM Peixe L Dispersion ofmultidrug-resistant Enterococcus faecium isolates belonging to majorclonal complexes in different Portuguese settings Appl Environ Microbiol2009 754904ndash4908
25 Poeta P Costa D Igrejas G Saacuteenz Y Zarazaga M Rodrigues J Torres CPolymorphisms of the pbp5 gene and correlation with ampicillinresistance in Enterococcus faecium isolates of animal origin J MedMicrobiol 2007 56236ndash240
26 Kuumlhn I Iversen A Burman LG Olsson-Liljequist B Franklin A Finn MAarestrup F Seyfarth AM Blanch AR Vilanova X Taylor H Caplin J MorenoMA Dominguez L Herrero IA Moumlllby R Comparison of enterococcalpopulations in animals humans and the environment ndash a Europeanstudy Int J Food Microbiol 2003 88133ndash145
27 Lanthier M Scott A Lapen DR Zhang Y Topp E Frequency of virulencegenes and antibiotic resistances in Enterococcus spp isolates fromwastewater and feces of domesticated mammals and birds and wildlifeCan J Microbiol 2010 56715ndash729
28 Leclercq R Oberleacute K Galopin S Cattoir V Budzinski H Petit F Changes inenterococcal populations and related antibiotic resistance along amedical center-wastewater treatment plant-river continuum Appl EnvironMicrobiol 2013 792428ndash2434
29 Thevenon F Regier N Benagli C Tonolla M Adatte T Wildi W Poteacute JCharacterization of fecal indicator bacteria in sediments cores from thelargest freshwater lake of Western Europe (Lake Geneva Switzerland)Ecotoxicol Environ Safety 2012 7850ndash56
30 Bonilla TD Nowosielski K Esiobu N McCorquodale DS Rogerson ASpecies assemblages of Enterococcus indicate potential sources offecal bacteria at a south Florida recreational beach Mar Pollut Bull2006 52807ndash810
31 Wang JT Chang SC Wang HY Chen PC Shiau YR Lauderdale TL TSARHospitals High rates of multidrug resistance in Enterococcus faecalis andE faecium isolated from inpatients and outpatients in TaiwanDiagn Microbiol Infect Dis 2013 75406ndash411
32 Lanthier M Scott A Zhang Y Cloutier M Durie D Henderson VC WilkesG Lapen DR Topp E Distribution of selected virulence genes andantibiotic resistance in Enterococcus species isolated from the SouthNation River drainage basin Ontario Canada J Appl Microbiol 2011110407ndash421
33 Kuch A Willems RJ Werner G Coque TM Hammerum AM Sundsfjord AKlare I Ruiz-Garbajosa P Simonsen GS van Luit-Asbroek M Hryniewicz WSadowy E Insight into antimicrobial susceptibility and population struc-ture of contemporary human Enterococcus faecalis isolates fromEurope J Antimicrob Chemother 2012 67551ndash558
34 Novais C Coque TM Ferreira H Sousa JC Peixe L Environmentalcontamination with vancomycin-resistant enterococci from hospitalsewage in Portugal Appl Environ Microbiol 2005 713364ndash3368
35 Iversen A Kuumlhn I Rahman M Franklin A Burman LG Olsson-Liljequist BTorell E Moumlllby R Evidence for transmission between humans and theenvironment of a nosocomial strain of Enterococcus faeciumEnviron Microbiol 2004 655ndash59
36 Rathnayake I Hargreaves M Huygens F SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterwayAustralia and associated antibiotic resistance gene profilesBMC Microbiol 2011 11201
37 Rosvoll TC Pedersen T Sletvold H Johnsen PJ Sollid JE Simonsen GSJensen LB Nielsen KM Sundsfjord A PCR-based plasmid typing inEnterococcus faecium strains reveals widely distributed pRE25-pRUM- pIP501- and pHT-related replicons associated with glycopeptideresistance and stabilizing toxin-antitoxin systems FEMS Immunol MedMicrobiol 2010 58254ndash268
38 Rosvoll TC Lindstad BL Lunde TM Hegstad K Aasnaes B Hammerum AMLester CH Simonsen GS Sundsfjord A Pedersen T Increased high-levelgentamicin resistance in invasive Enterococcus faecium is associated withaac(6prime)Ie-aph(2Prime)Ia-encoding transferable megaplasmids hosted by majorhospital-adapted lineages FEMS Immunol Med Microbiol 2012 66166ndash176
39 Laverde Gomez JA van Schaik W Freitas AR Coque TM Weaver KE FranciaMV Witte W Werner G A multiresistance megaplasmid pLG1 bearing ahylEfm genomic island in hospital Enterococcus faecium isolates Int J MedMicrobiol 2011 301165ndash175
40 Coque TM Evolutionary biology of pathogenic enterococci InEvolutionary Biology of Bacterial and Fungal Pathogens Coque TM Edited byBaquero F Nombela C Cassell GH Gutierrez JA Washington DC ASMPress501ndash523
41 Werner G Fleige C Geringer U van Schaik W Klare I Witte W IS elementIS16 as a molecular screening tool to identify hospital-associated strainsof Enterococcus faecium BMC Infect Dis 2011 1180
42 Top J Sinnige JC Majoor EA Bonten MJ Willems RJ van Schaik W Therecombinase IntA is required for excision of esp-containing ICEEfm1 inEnterococcus faecium J Bacteriol 2011 1931003ndash1006
43 Sillanpaumlauml J Prakash VP Nallapareddy SR Murray BE Distribution of genesencoding MSCRAMMs and Pili in clinical and natural populations ofEnterococcus faecium J Clin Microbiol 2009 47896ndash901
44 Marcinek H Wirth R Muscholl-Silberhorn A Gauer M Enterococcus faecalisgene transfer under natural conditions in municipal sewage watertreatment plants Appl Environ Microbiol 1998 64626ndash632
45 Kuumlmmerer K Significance of antibiotics in the environment J AntimicrobChemother 2003 525ndash7
46 Liu A Fong A Becket E Yuan J Tamae C Medrano L Maiz M Wahba C LeeC Lee K Tran KP Yang H Hoffman RM Salih A Miller JHP Selectiveadvantage of resistant strains at trace levels of antibiotics a simple andultrasensitive color test for detection of antibiotics and genotoxicagents Antimicrob Agents Chemother 2011 551204ndash1210
47 Clinical and Laboratory Standards Institute Performance standards forantimicrobial susceptibility testing 21st informational supplement Wayne CLSI2011 PA M100-S20
48 Magiorakos AP Srinivasan A Carey RB Carmeli Y Falagas ME Giske CGHarbarth S Hindler JF Kahlmeter G Olsson-Liljequist B Paterson DL Rice LBStelling J Struelens MJ Vatopoulos A Weber JT Monnet DLMultidrug-resistant extensively drug-resistant and pandrug-resistantbacteria an international expert proposal for interim standarddefinitions for acquired resistance Clin Microbiol Infect 2012 18268ndash281
49 Homan WL Tribe D Poznanski S Li M Hogg G Spalburg E Van Embden JDWillems RJ Multilocus sequence typing scheme for Enterococcus faeciumJ Clin Microbiol 2002 401963ndash1971
50 Jolley KA Feil EJ Chan MS Maiden MC Sequence type analysis andrecombinational tests (START) Bioinformatics 2001 171230ndash1231
51 Feil EJ Li BC Aanensen DM Hanage WP Spratt BG eBURST inferringpatterns of evolutionary descent among clusters of related bacterialgenotypes from multilocus sequence typing data J Bacteriol 20041861518ndash1530
52 Top J Banga1 NMI Hayes R Willems RJ Bonten MJM Hayden MKComparison of multiple-locus variable-number tandem repeat analysisand pulsed-field gel electrophoresis in a setting of polyclonal endemicityof vancomycin-resistant Enterococcus faecium Clin Microbiol Infect 200814363ndash369
53 Vankerckhoven V Van Autgaerden T Vael C Lammens C Chapelle S RossiR Jabes D Goossens H Development of a multiplex PCR for the
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-

Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 14 of 15httpwwwbiomedcentralcom1471-21801466
16 Galloway-Pentildea JR Nallapareddy SR Arias CA Eliopoulos GM Murray BEAnalysis of clonality and antibiotic resistance among early clinicalisolates of Enterococcus faecium in the United States J Infect Dis 20092001566ndash1573
17 Top J Willems R Bonten M Emergence of CC17 Enterococcus faeciumfrom commensal to hospital-adapted pathogen FEMS Immunol MedMicrobiol 2008 52297ndash308
18 Martins da Costa P Vaz-Pires P Bernardo F Antimicrobial resistance inEnterococcus spp isolated in inflow effluent and sludge from municipalsewage water treatment plants Water Res 2006 401735ndash1740
19 Reinthaler FF Posch J Feierl G Wust G Haas D Ruckenbauer G Mascher FMarth E Antibiotic resistance of Escherichia coli in sewage and sludgeWater Res 2003 371685ndash1690
20 Luczkiewicz A Fudala-Ksiazek S Jankowska K Olanczuk-Neyman KAntimicrobial resistance of fecal indicators in municipal wastewatertreatment plant Water Res 2010 445089ndash5097
21 Dulfour AP Ballentine RK Ambient water quality criteria for bacteria-1986 EPA4405-84-002 Washington DC US Environmental Protection Agency 1986
22 Ferreira da Silva M Tiago I Veriacutessimo A Boaventura RA Nunes OC Manaia CMAntimicrobial resistance patterns in Enterobacteriaceae isolated from anurban wastewater treatment plant FEMS Microbiol Ecol 2007 60166ndash176
23 DIRECTIVE 20067EC of the European Parliament and of the Council 15February 2006 concerning the management of bathing water quality andrepealing Directive 76160EEC httpeur-lexeuropaeuLexUriServLexUriServdouri=OJL200606400370051ENPDF
24 Freitas AR Novais C Ruiz-Garbajosa P Coque TM Peixe L Dispersion ofmultidrug-resistant Enterococcus faecium isolates belonging to majorclonal complexes in different Portuguese settings Appl Environ Microbiol2009 754904ndash4908
25 Poeta P Costa D Igrejas G Saacuteenz Y Zarazaga M Rodrigues J Torres CPolymorphisms of the pbp5 gene and correlation with ampicillinresistance in Enterococcus faecium isolates of animal origin J MedMicrobiol 2007 56236ndash240
26 Kuumlhn I Iversen A Burman LG Olsson-Liljequist B Franklin A Finn MAarestrup F Seyfarth AM Blanch AR Vilanova X Taylor H Caplin J MorenoMA Dominguez L Herrero IA Moumlllby R Comparison of enterococcalpopulations in animals humans and the environment ndash a Europeanstudy Int J Food Microbiol 2003 88133ndash145
27 Lanthier M Scott A Lapen DR Zhang Y Topp E Frequency of virulencegenes and antibiotic resistances in Enterococcus spp isolates fromwastewater and feces of domesticated mammals and birds and wildlifeCan J Microbiol 2010 56715ndash729
28 Leclercq R Oberleacute K Galopin S Cattoir V Budzinski H Petit F Changes inenterococcal populations and related antibiotic resistance along amedical center-wastewater treatment plant-river continuum Appl EnvironMicrobiol 2013 792428ndash2434
29 Thevenon F Regier N Benagli C Tonolla M Adatte T Wildi W Poteacute JCharacterization of fecal indicator bacteria in sediments cores from thelargest freshwater lake of Western Europe (Lake Geneva Switzerland)Ecotoxicol Environ Safety 2012 7850ndash56
30 Bonilla TD Nowosielski K Esiobu N McCorquodale DS Rogerson ASpecies assemblages of Enterococcus indicate potential sources offecal bacteria at a south Florida recreational beach Mar Pollut Bull2006 52807ndash810
31 Wang JT Chang SC Wang HY Chen PC Shiau YR Lauderdale TL TSARHospitals High rates of multidrug resistance in Enterococcus faecalis andE faecium isolated from inpatients and outpatients in TaiwanDiagn Microbiol Infect Dis 2013 75406ndash411
32 Lanthier M Scott A Zhang Y Cloutier M Durie D Henderson VC WilkesG Lapen DR Topp E Distribution of selected virulence genes andantibiotic resistance in Enterococcus species isolated from the SouthNation River drainage basin Ontario Canada J Appl Microbiol 2011110407ndash421
33 Kuch A Willems RJ Werner G Coque TM Hammerum AM Sundsfjord AKlare I Ruiz-Garbajosa P Simonsen GS van Luit-Asbroek M Hryniewicz WSadowy E Insight into antimicrobial susceptibility and population struc-ture of contemporary human Enterococcus faecalis isolates fromEurope J Antimicrob Chemother 2012 67551ndash558
34 Novais C Coque TM Ferreira H Sousa JC Peixe L Environmentalcontamination with vancomycin-resistant enterococci from hospitalsewage in Portugal Appl Environ Microbiol 2005 713364ndash3368
35 Iversen A Kuumlhn I Rahman M Franklin A Burman LG Olsson-Liljequist BTorell E Moumlllby R Evidence for transmission between humans and theenvironment of a nosocomial strain of Enterococcus faeciumEnviron Microbiol 2004 655ndash59
36 Rathnayake I Hargreaves M Huygens F SNP diversity of Enterococcusfaecalis and Enterococcus faecium in a South East Queensland waterwayAustralia and associated antibiotic resistance gene profilesBMC Microbiol 2011 11201
37 Rosvoll TC Pedersen T Sletvold H Johnsen PJ Sollid JE Simonsen GSJensen LB Nielsen KM Sundsfjord A PCR-based plasmid typing inEnterococcus faecium strains reveals widely distributed pRE25-pRUM- pIP501- and pHT-related replicons associated with glycopeptideresistance and stabilizing toxin-antitoxin systems FEMS Immunol MedMicrobiol 2010 58254ndash268
38 Rosvoll TC Lindstad BL Lunde TM Hegstad K Aasnaes B Hammerum AMLester CH Simonsen GS Sundsfjord A Pedersen T Increased high-levelgentamicin resistance in invasive Enterococcus faecium is associated withaac(6prime)Ie-aph(2Prime)Ia-encoding transferable megaplasmids hosted by majorhospital-adapted lineages FEMS Immunol Med Microbiol 2012 66166ndash176
39 Laverde Gomez JA van Schaik W Freitas AR Coque TM Weaver KE FranciaMV Witte W Werner G A multiresistance megaplasmid pLG1 bearing ahylEfm genomic island in hospital Enterococcus faecium isolates Int J MedMicrobiol 2011 301165ndash175
40 Coque TM Evolutionary biology of pathogenic enterococci InEvolutionary Biology of Bacterial and Fungal Pathogens Coque TM Edited byBaquero F Nombela C Cassell GH Gutierrez JA Washington DC ASMPress501ndash523
41 Werner G Fleige C Geringer U van Schaik W Klare I Witte W IS elementIS16 as a molecular screening tool to identify hospital-associated strainsof Enterococcus faecium BMC Infect Dis 2011 1180
42 Top J Sinnige JC Majoor EA Bonten MJ Willems RJ van Schaik W Therecombinase IntA is required for excision of esp-containing ICEEfm1 inEnterococcus faecium J Bacteriol 2011 1931003ndash1006
43 Sillanpaumlauml J Prakash VP Nallapareddy SR Murray BE Distribution of genesencoding MSCRAMMs and Pili in clinical and natural populations ofEnterococcus faecium J Clin Microbiol 2009 47896ndash901
44 Marcinek H Wirth R Muscholl-Silberhorn A Gauer M Enterococcus faecalisgene transfer under natural conditions in municipal sewage watertreatment plants Appl Environ Microbiol 1998 64626ndash632
45 Kuumlmmerer K Significance of antibiotics in the environment J AntimicrobChemother 2003 525ndash7
46 Liu A Fong A Becket E Yuan J Tamae C Medrano L Maiz M Wahba C LeeC Lee K Tran KP Yang H Hoffman RM Salih A Miller JHP Selectiveadvantage of resistant strains at trace levels of antibiotics a simple andultrasensitive color test for detection of antibiotics and genotoxicagents Antimicrob Agents Chemother 2011 551204ndash1210
47 Clinical and Laboratory Standards Institute Performance standards forantimicrobial susceptibility testing 21st informational supplement Wayne CLSI2011 PA M100-S20
48 Magiorakos AP Srinivasan A Carey RB Carmeli Y Falagas ME Giske CGHarbarth S Hindler JF Kahlmeter G Olsson-Liljequist B Paterson DL Rice LBStelling J Struelens MJ Vatopoulos A Weber JT Monnet DLMultidrug-resistant extensively drug-resistant and pandrug-resistantbacteria an international expert proposal for interim standarddefinitions for acquired resistance Clin Microbiol Infect 2012 18268ndash281
49 Homan WL Tribe D Poznanski S Li M Hogg G Spalburg E Van Embden JDWillems RJ Multilocus sequence typing scheme for Enterococcus faeciumJ Clin Microbiol 2002 401963ndash1971
50 Jolley KA Feil EJ Chan MS Maiden MC Sequence type analysis andrecombinational tests (START) Bioinformatics 2001 171230ndash1231
51 Feil EJ Li BC Aanensen DM Hanage WP Spratt BG eBURST inferringpatterns of evolutionary descent among clusters of related bacterialgenotypes from multilocus sequence typing data J Bacteriol 20041861518ndash1530
52 Top J Banga1 NMI Hayes R Willems RJ Bonten MJM Hayden MKComparison of multiple-locus variable-number tandem repeat analysisand pulsed-field gel electrophoresis in a setting of polyclonal endemicityof vancomycin-resistant Enterococcus faecium Clin Microbiol Infect 200814363ndash369
53 Vankerckhoven V Van Autgaerden T Vael C Lammens C Chapelle S RossiR Jabes D Goossens H Development of a multiplex PCR for the
Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-

Sadowy and Luczkiewicz BMC Microbiology 2014 1466 Page 15 of 15httpwwwbiomedcentralcom1471-21801466
detection of asa1 gelE cylA esp and hyl genes in enterococci andsurvey for virulence determinants among European hospital isolates ofEnterococcus faecium J Clin Microbiol 2004 424473ndash4479
54 Doherty N Trzcinski K Pickerill P Zawadzki P Dowson CG Genetic diversity ofthe tet(M) gene in tetracycline-resistant clonal lineages of Streptococcuspneumoniae Antimicrob Agents Chemother 2000 442979ndash2984
55 Trzcinski K Cooper BS Hryniewicz W Dowson CG Expression of resistanceto tetracyclines in strains of methicillin-resistant Staphylococcus aureusJ Antimicrob Chemother 2000 45763ndash770
56 Leelaporn A Yodkamol K Waywa D Pattanachaiwit S A novel structure ofTn4001-truncated element type V in clinical enterococcal isolates andmultiplex PCR for detecting aminoglycoside resistance genes Int JAntimicrob Agents 2008 31250ndash254
57 Agersoslash Y Pedersen AG Aarestrup FM Identification of Tn5397-like andTn916-like transposons and diversity of the tetracycline resistance genetet(M) in enterococci from humans pigs and poultry J AntimicrobChemother 2006 57832ndash839
58 Jensen LB Garcia Migura L Valenzuela AJ Loslashhr M Hasman H Aarestrup FMA classification system for plasmids from enterococci and othergram-positive bacteria J Microbiol Methods 2010 8025ndash43
59 Moritz EM Hergenrother PJ Toxin-antitoxin systems are ubiquitous andplasmid-encoded in vancomycin-resistant enterococci Proc Natl Acad SciU S A 2007 104311ndash316
60 Wardal E Gawryszewska I Hryniewicz W Sadowy E Abundance anddiversity of plasmid-associated genes among clinical isolates ofEnterococcus faecalis Plasmid 2013 70329ndash342
61 Sadowy E Matynia B Hryniewicz W Population structure virulence factorsand resistance determinants of invasive non-invasive and colonizingStreptococcus agalactiae in Poland J Antimicrob Chemother 2010651907ndash1914
62 Hsieh SE Hsu LL Hsu WH Chen CY Chen HJ Liao CT Importance ofamino acid alterations and expression of penicillin-binding protein 5 toampicillin resistance of Enterococcus faecium in Taiwan Int J AntimicrobAgents 2006 28514ndash519
63 el Amin NA Jalal S Wretlind B Alterations in GyrA and ParC associatedwith fluoroquinolone resistance in Enterococcus faecium AntimicrobAgents Chemother 1999 43947ndash949
doi1011861471-2180-14-66Cite this article as Sadowy and Luczkiewicz Drug-resistant andhospital-associated Enterococcus faecium from wastewater riverineestuary and anthropogenically impacted marine catchment basin BMCMicrobiology 2014 1466
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Species composition and antimicrobial susceptibility of enterococcal isolates
- Clonal structure of E faecium isolates and distribution of virulence resistance and MGE genes
-
- Discussion
- Conclusions
- Methods
-
- Acquisition of samples and isolation of enterococci
- Species identification DNA isolation susceptibility testing and statistical analysis
- MLST and MLVA of E faecium
- Detection and analysis of genes associated with virulence antimicrobial resistance and MGE in E faecium
-
- Additional file
- Competing interest
- Authorsrsquo contributions
- Acknowledgements
- Author details
- References
-


















