
Research Article MiR-183 Regulates ITGB1P Expression...
11
Research Article MiR-183 Regulates ITGB1P Expression and Promotes Invasion of Endometrial Stromal Cells Jie Chen, 1 Lin Gu, 1 Jie Ni, 1 Ping Hu, 1 Kai Hu, 1 and Ying-Li Shi 2 1 State Key Laboratory of Reproductive Medicine, Department of Gynecology, Nanjing Maternity and Child Health Care Hospital Affiliated to Nanjing Medical University, 123 Tianfei Road, Nanjing, Jiangsu 210029, China 2 Shanghai Ji Ai Genetics & IVF Institute, Obstetrics & Gynecology Hospital, Fudan University, 588 Fangxie Road, Shanghai 200011, China Correspondence should be addressed to Ying-Li Shi; [email protected] Received 14 January 2015; Accepted 24 February 2015 Academic Editor: Shi-Wen Jiang Copyright © 2015 Jie Chen et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. We applied in the previous study miRNA microarray screening analysis to identify several differentially expressed miRNAs, including miR-183 in normal, eutopic, and ectopic endometrium. Knockdown of miR-183 expression induced the invasiveness and inhibition of apoptosis in endometrial stromal cells. e current study aims to identify the miR-183 targets with relevance to cell functions in endometrial stromal cells, to verify the interaction of miR-183 with its target genes, and to confirm the role of miR-183 in the process of endometriosis. Using microarray analysis, we identified 27 differentially expressed genes (19 were upregulated and 8 downregulated), from which we selected 4 downregulated genes (ITGB1, AMIGO2, VAV3, and PSEN2) based on GO databases for functional analysis and significant pathway analysis. Western blotting analyses showed that integrin 1 (ITGB1), but not AMIGO2, was affected by miR-183 overexpression, whereas no protein expression of VAV3 and PSEN2 was detected. Luciferase reporter assay verified that ITGB1 is a target gene of miR-183. Moreover, we found that ITGB1 is overexpressed in the endometrium of endometriosis patients. Furthermore, overexpression of ITGB1 rescued the repressive effects of miR-183 on the invasiveness of endometrial stromal cells. ese findings, together with the fact that ITGB1 is a critical factor for cell adhesion and invasiveness, suggest that miR-183 may be involved in the development of endometriosis by regulating ITGB1 in endometrial stromal cells. 1. Introduction Affecting 6–10% of women at reproductive age, endometrio- sis is a common chronic gynecological disorder associated with infertility, severe pelvic pain, and menstrual disorders [1]. Despite intensive research efforts, the pathogenic mecha- nism of endometriosis remains poorly understood. So far, it is still a highly underdiagnosed disease, and the current surgical and hormonal treatment of endometriosis is not effective and recurrence is oſten observed. MicroRNAs (miRNAs) are small, noncoding regulatory RNAs that regulate the stability and translation of mRNAs by inhibiting ribosome functions, deadenylating the poly(A) tail, and degrading target mRNA [2]. Aberrant miRNA expression has been linked to a variety of human diseases such as gynecological diseases, cancers, inflammatory dis- eases, and cardiovascular disorders [3]. A single microRNA molecule is able to modulate multiple target genes; thus it is involved in multiple cell functions including proliferation, invasiveness, and differentiation [4]. In 2007, Pan et al. detected deregulated expression of miRNAs in paired ectopic and eutopic endometrial tissues using microarray analysis. From then on, many studies have con- firmed the aberrant expression patterns of miRNAs in ectopic as well as eutopic endometrial tissues from endometri- otic patients. MiR-520, miR-199a, miR-23a, miR-23b, miR- 145, miR-196, miR-10, miR-20a, let-7, miR-126, and miR- 135 are among the most studied miRNAs in endometriosis. Adammek et al. reported that miR-145 inhibited endometri- otic cell proliferation, invasiveness, and stemness by tar- geting pluripotency factors, cytoskeletal elements, and pro- tease inhibitors [5]. MiR-196b was implicated in HOXA10 expression and in the proliferation of endometriotic stromal cells [6]. Loss of eutopic endometrial miR-451 expression is Hindawi Publishing Corporation BioMed Research International Volume 2015, Article ID 340218, 10 pages http://dx.doi.org/10.1155/2015/340218
Transcript of Research Article MiR-183 Regulates ITGB1P Expression...

Research ArticleMiR-183 Regulates ITGB1P Expression and Promotes Invasionof Endometrial Stromal Cells
Jie Chen1 Lin Gu1 Jie Ni1 Ping Hu1 Kai Hu1 and Ying-Li Shi2
1State Key Laboratory of Reproductive Medicine Department of Gynecology Nanjing Maternity and Child Health Care HospitalAffiliated to Nanjing Medical University 123 Tianfei Road Nanjing Jiangsu 210029 China2Shanghai Ji Ai Genetics amp IVF Institute Obstetrics amp Gynecology Hospital Fudan University 588 Fangxie RoadShanghai 200011 China
Correspondence should be addressed to Ying-Li Shi ylshi2005163com
Received 14 January 2015 Accepted 24 February 2015
Academic Editor Shi-Wen Jiang
Copyright copy 2015 Jie Chen et alThis is an open access article distributed under the Creative Commons Attribution License whichpermits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
We applied in the previous study miRNA microarray screening analysis to identify several differentially expressed miRNAsincluding miR-183 in normal eutopic and ectopic endometrium Knockdown of miR-183 expression induced the invasiveness andinhibition of apoptosis in endometrial stromal cells The current study aims to identify the miR-183 targets with relevance to cellfunctions in endometrial stromal cells to verify the interaction of miR-183 with its target genes and to confirm the role of miR-183in the process of endometriosis Using microarray analysis we identified 27 differentially expressed genes (19 were upregulated and8 downregulated) fromwhichwe selected 4 downregulated genes (ITGB1 AMIGO2VAV3 and PSEN2) based onGOdatabases forfunctional analysis and significant pathway analysis Western blotting analyses showed that integrin 1205731 (ITGB1) but not AMIGO2was affected by miR-183 overexpression whereas no protein expression of VAV3 and PSEN2 was detected Luciferase reporterassay verified that ITGB1 is a target gene of miR-183 Moreover we found that ITGB1 is overexpressed in the endometrium ofendometriosis patients Furthermore overexpression of ITGB1 rescued the repressive effects of miR-183 on the invasiveness ofendometrial stromal cells These findings together with the fact that ITGB1 is a critical factor for cell adhesion and invasivenesssuggest that miR-183 may be involved in the development of endometriosis by regulating ITGB1 in endometrial stromal cells
1 Introduction
Affecting 6ndash10 of women at reproductive age endometrio-sis is a common chronic gynecological disorder associatedwith infertility severe pelvic pain and menstrual disorders[1] Despite intensive research efforts the pathogenic mecha-nismof endometriosis remains poorly understood So far it isstill a highly underdiagnosed disease and the current surgicaland hormonal treatment of endometriosis is not effective andrecurrence is often observed
MicroRNAs (miRNAs) are small noncoding regulatoryRNAs that regulate the stability and translation of mRNAsby inhibiting ribosome functions deadenylating the poly(A)tail and degrading target mRNA [2] Aberrant miRNAexpression has been linked to a variety of human diseasessuch as gynecological diseases cancers inflammatory dis-eases and cardiovascular disorders [3]
A singlemicroRNAmolecule is able tomodulatemultipletarget genes thus it is involved in multiple cell functionsincluding proliferation invasiveness and differentiation [4]In 2007 Pan et al detected deregulated expression ofmiRNAsin paired ectopic and eutopic endometrial tissues usingmicroarray analysis From then on many studies have con-firmed the aberrant expression patterns ofmiRNAs in ectopicas well as eutopic endometrial tissues from endometri-otic patients MiR-520 miR-199a miR-23a miR-23b miR-145 miR-196 miR-10 miR-20a let-7 miR-126 and miR-135 are among the most studied miRNAs in endometriosisAdammek et al reported that miR-145 inhibited endometri-otic cell proliferation invasiveness and stemness by tar-geting pluripotency factors cytoskeletal elements and pro-tease inhibitors [5] MiR-196b was implicated in HOXA10expression and in the proliferation of endometriotic stromalcells [6] Loss of eutopic endometrial miR-451 expression is
Hindawi Publishing CorporationBioMed Research InternationalVolume 2015 Article ID 340218 10 pageshttpdxdoiorg1011552015340218
2 BioMed Research International
associated with decreased establishment of endometriosis ina mouse model [7] These data landed strong support foran important role of miRNAs in the development of endo-metriosis through regulating cell proliferation cell apoptosiscell migration cell invasiveness and estrogenic signaling
In a previous study we identified several differentially ex-pressed miRNAs among normal eutopic and ectopic endo-metrium MiR-183 was found to be the most downregulatedmiRNA in both the ectopic and eutopic tissues comparedto normal endometrium Functional studies indicated thatmiR-183may contribute to endometrial stromal cell apoptosisand impose a negative regulatory impact on cell invasivenessbut it has no effect on endometrial stromal cell proliferationThe study suggested that aberrant miR-183 expressionmay beinvolved in the development and progression of endometrio-sis [8]
In the present study we aim to identify the functionaltargets of miR-183 using an overexpression model Weexamine if miR-183 exerts its effects by directly binding toand inhibiting the ITGB1 promoter activity In addition weinvestigate ifmiR-183 could regulate stromal cell invasivenessThe findings from these experiments will help us to betterunderstand the miR-183-mediated molecular mechanism inendometric stromal cells
2 Materials and Methods
21 Tissue Acquisition and Cell Culture All the eutopic andnormal endometrial tissues were obtained at the NanjingMaternity and Child Health Care Hospital Affiliated to Nan-jing Medical University by uterine curettage from patientswith or without endometriosis None of the patients hadreceived preoperative hormonal therapy and all samples werehistologically confirmed by pathologists All samples werecollected from the proliferative phase of the menstrual cycleThe phase of the menstrual cycle was determined based onhistologic evaluation of the endometrium with the assistanceof pathologists The average age of the patients was 305 plusmn52 Patients consented to tissue donation prior to surgeryThe research project and consent form were approved by thehospital ethics committee Each sample was divided into twoparts one part was snap-frozen and stored at minus20∘C and laterused for mRNA and protein extraction the other part wasrinsed thoroughly with cold PBS and directly subjected todigestion and stromal cell isolation
The endometrial stromal cells (ESC) fromnormal womenwithout endometriosis were used as control Tissue digestionand stromal cell isolation and culturewere performed accord-ing to the method described in previous publication [9]
22 MiR-183-Lentivirus Construction and Transduction Thelentivirus gene transfer vector carrying precursor of hsa-miR-183 (Genbank accession number MIMAT0000261) andencoding green fluorescence protein (GFP) were constructedby Genechem Co Ltd Shanghai China and confirmed byDNA sequencing The primers of RNA were 51015840-GAGGAT-CCCCGGGTACCAAGGGAGTGGGCAGGCTA and 51015840-ATAAGCTTGATATCGTCCCTGCACCCTTGGAAGCA
The recombinant lentivirus formiR-183 overexpression (miR-183-lentivirus) and the control lentivirus (GFP-lentivirus)were prepared and titered to 50119864 + 8TUmL (transfectionunit per mL)
The sequence of inhibitor of hsa-miR-183-5p was TATG-GCACTGGTAGAATTCACT The recombinant lentivirus ofmiR-183-5p inhibitor (In-miR-183 lentivirus) and the controllentivirus (GFP-lentivirus) were prepared and titered to40119864 + 8TUmL (transfection unit per mL)
ESC from women without endometriosis were plated in6-well plates (5 times 104 cellswell) overnight The lentiviruseswere diluted in 02mL complete medium containing poly-brene (8mgmL) and added to the cells for 12 h of incubationat 37∘C followed by incubation in 03mL of freshly preparedpolybrene-DMEM for 24 h The media were replaced withfresh DMEM and the cells were cultured for 3 days Thelentivirus transduction efficiency of ESC was determined bythe detection of GFP signals under a fluorescencemicroscopeat 72 h after transduction The miR-183 expression in stablytransduced ESC was measured by real-time PCR The ESCtransfected with miR-183-lentivirus In-miR-183-lentivirusand GFP-lentivirus were kept for further analysis
23 RNA Extraction and Microarray For the microarrayanalyses groups were divided into the ESC with miR-183 overexpression and the control ones Total RNA wasextracted using TRIzol (Invitrogen) according to the man-ufacturerrsquos instructions Gene expression profiling was con-ducted using PrimeView Human Gene Expression Array(Affymetrix) The array contains 530000 probes coveringmore than 36000 transcripts and variants which representmore than 20000 genes mapped through RefSeq or viaUniGene annotation All subsequent technical proceduresand quality controls were performed by Genechem Co LtdShanghai China The arrays were scanned using a GeneChipScanner 3000 (Affymetrix Inc CA USA) Raw data wereextracted from the scanned images and analyzed usingGeneSpring GX software version 115 (Agilent TechnologiesCA USA)
The data were normalized using the PLIER default proto-col Significant differentially expressed genes were analyzedusing an unpaired 119905-test Hierarchical cluster analysis wasused to assess correlations among samples for each identifiedgene set with Euclidean distance and average linkage statisti-cal methods
24MicroarrayDataAnalyses Differentially expressed geneswere uploaded to the Ingenuity database (Ingenuity Sys-tems Redwood City CA USA) for pathway and functionalanalyses Genes were further annotated and classified basedon the Gene Ontology (GO) consortium annotations fromthe GO Bostaurus database using GOEAST (Gene OntologyEnrichment Analysis Software Toolkit)
25Western Blotting ESC proteins were extracted with RIPAbuffer [(50mM Tris-HCl pH 80 150mM NaCl 1 (vv)NP-40 05 (vo) sodium deoxycholate and 01 (wo)
BioMed Research International 3
sodium dodecyl sulphate (SDS)] supplemented with a pro-tease inhibitor cocktail (Sigma) After incubating on ice for30min cell debris was removed by centrifugation at 12000 gfor 10min Cell lysates were resolved in 12 SDS-PAGEelectrophoresis and transferred to the PVDFmembrane (Bio-Rad Hercules CA) Nonspecific binding was blocked byincubating the membranes in Tris-buffered saline containing5 nonfat milk (TBST 50mmolL TrisndashHCl 150mmolLNaCl and 01 Tween-20) for 1 h at room temperatureMembranes were incubated overnight at 4∘C with AMIGO2antibody (1 400 RD) integrin 1205731 antibody (1 1000 Cell Sig-naling) Presenilin 2 antibody (1 500 ABCAM) and VAV3antibody (1 500 ABCAM) After three washes with TBSTthe membranes were incubated for 1 h at room temperaturewith the horseradish peroxidase-labeled secondary antibodyThe signals were visualized with chemiluminescence andquantified with Quantity-One (Bio-Rad America)
26 Luciferase Assay The mutant construct of ITGB131015840UTR was obtained by introducing the mutation intothe 7 nucleotides (GUGCCAUU) of the seed region formiR-183 The mutant construct of AMIGO2 31015840UTR wasobtained by introducing twomutations into the 7 nucleotides(GUGCCAUA) (UGCCAUA) of the seed region for miR-183 The miR-183 target sequences in the coding region ofITGB1AMIGO2 were amplified by PCR and cloned intoGV143 that contained a firefly luciferase reporter gene Wild-type ITBG1AMIGO2 31015840UTR or mutant ITBG1AMIGO231015840UTR and the empty 31015840UTR vector were cotransfected intoESC Cell transfection using Lipofectamine 2000 and nor-malization for transfection efficiency was performed accord-ing to the recommendation of manufacturer (Invitrogen)Luciferase activity was measured 24 h after transfection usingthe Dual-Luciferase Reporter Assay System (Promega CorpMadison WI)
27 Quantitative RT-PCR Endometrial tissue was mincedwith a scalpel blade Total RNA was isolated using TRIzol(Takara Otsu Shiga Japan) and cDNA was synthesizedusing the SYBR Prime Script RT-PCR kit (Takara) on theABI Prism 7500 Sequence Detection System according tomanufacturerrsquos instructions The housekeeping gene 120573-actinwas used for normalization The primers used were 51015840-TCG-TCACGTTCCGGTTATTC (sense) and 51015840-CTTTTACTT-ACGGTTTACCC (antisense) for ITGB1 51015840-CCTCGCCTT-TGCCGATCCG (sense) and 51015840-GCCGGAGCCGTTGTC-GACG (antisense) for 120573-actin The cycling conditions were94∘C for 1min 35 cycles of 94∘C for 30 sec 66∘C for 30 secand 68∘C for 3min followed by extension at 68∘C for3min Quantitative PCR was carried out using SYBR-GreenJumpStart Taq ReadyMix (Sigma) and the 7300 Real-TimePCRDetection System (ABI)The results were analyzed usingthe comparative threshold cycle (CT) method
28 Rescue Experiment To further validate direct targetingof ITGB1 by miR-183 functional rescue experiment was per-formed by cotransfection with miR-183 mimic and plasmidconstructs expressing ITGB1 in ESC using Lipofectamine
2000 (Invitrogen) as described aboveThe expression plasmidpcDNA-ITGB1 encoding human ITGB1was purchased fromGenechem The presence of complete ITGB1 coding regionswas confirmed by DNA sequencing
29 Invasion (Matrigel) Chamber Assay ESC (25 times 104) wereseeded on a transwell insert coated with extracellular matrix(ECM) (8mm pore size 24-well format Corning Costar) in2 FBS medium Complete medium (10 FBS) was addedto the lower chamber To determine the amount of invasioncells were incubated for 24 h and then removed from theupper chamber using a cotton swab The invaded cells onthe underside of the insert were fixed with methanol stainedwith crystal violet for 2min and rinsed with phosphatebuffered saline (PBS) The undersides of the membrane werephotographed to compare the number of invaded cells perinsert The transmigrated cells were counted using a lightmicroscope Invasive cells were scored by counting 10 randomhigh-power fields per filter
210 Statistical Analyses Data represents mean plusmn SEM fromat least 3 independent experiments Difference betweenexperimental and control groupswas determined by Studentrsquos119905-test while one-way ANOVA analysis was employed tocompare three or more groups 119875 lt 005 was considered asstatistical significance
3 Results
31 Gene Expression Profiling following miR-183 Overexpres-sion In order to screen target genes in response to miR-183we used microarrays representing more than 20000 genesmapped through RefSeq or via UniGene annotation Westudied gene expression alterations (up- or downregulation)at 24 h after transfection The changes of gene expressionin miR-183-overexpressing endometrial stromal cells wereanalyzed Differential expression was found in 27 genes at119875 value lt 005 with folds of change ge15 Of these 19 wereupregulated and 8 downregulated (ITGB1 AMIGO2 VAV3PSEN2 LHFPL2 HS2ST1 AHSA2 and UQCRB) Resultsof hierarchical cluster analyses of these genes are shownin Figure 1 and supplementary 1 in Supplementary Materialavailable online at httpdxdoiorg1011552015340218
32 Functional Analysis with GO Databases By using theGene Ontology (GO) database we systematically extractedand analyzed the information of threeGOcategories ldquobiolog-ical processrdquo ldquomolecular functionrdquo and ldquocellular componentrdquoIt was revealed that the identified genes were involvedin hemophilic cell adhesion (ITGB1 AMIGO2) cell-celladhesion (ITGB1 AMIGO2) cell migration (ITGB1MYH9)positive regulation of catalytic activity (PSEN2 SHC1) andproteolysis (MYH9 PSEN2) (Table 1)
33 Significant Pathway Analysis Significant pathway analy-sis revealed that the gene expression alterations in endome-trial stromal cells were involved in pathways of PTEN(ITGB1 SHC1) TFF (ITGB1 SHC1) ECM (ITGB1 SHC1)
4 BioMed Research International
Table 1 List of genes with fold of change ge15 (119875 lt 005) and their biological functions
Gene Gene set name 119875
ITGB1 AMIGO2 Hemophilic cell adhesion 197119864 minus 03
ITGB1 AMIGO2 Cell to cell adhesion 502119864 minus 02
ITGB1 MYH9 Cell migration 610119864 minus 02
PSEN2 SHC1 Positive regulation of catalytic activity 152119864 minus 01
MYH9 PSEN2 Proteolysis 191119864 minus 01
UQCRBAMIGO2HS2ST1AHSA2VAV3LHFPL2ITGB1PSEN2PICALMHES4SHC1HYOU1PPIFHPCAL1TUBBCBSMZT1CDT1GNB2PPP2R5DPTPRHCHMP4BVPS35PTPRAMYH9C6orf150HDGF
minus1 minus05 0 05 1 15Row Z-score
X461_1 X461_3 X461_2 X462_3 X462_1 X462_2
Figure 1 Hierarchical clustering of differentially expressed genes in miR-183-overexpressing endometrial stromal cells versus control cellsGene expression profiling was conducted using PrimeViewHumanGene ExpressionArray Raw data were extracted from the scanned imagesand analyzed with GeneSpring GX software version 115 The data was normalized using the iterative PLIER default protocol Differentiallyexpressed genes were analyzed with the unpaired Student 119905-test Statistically different genes (119875 lt 005) with a greater than 15-fold increaseor decrease were recorded Red color indicates low expression green color indicates relatively high expression
ERK (ITGB1 SHC1) integrin (ITGB1 SHC1) pathogenicEscherichia coli infection (ITGB1 TUBB) chemokine sig-naling pathway (SHC1 VAV3 and GNB2) focal adhe-sion (ITGB1 SHC1 and VAV3) regulation of cytoskele-ton (ITGB1 VAV3 and MYH9) leukocyte transendothelialmigration (ITGB1 VAV3) natural killer cell-mediated cyto-toxicity (SHC1 VAV3) and Alzheimerrsquos disease (UQCRBPSEN2) (Table 2)
34 Confirmation of Microarray Data by Western BlottingBecause of the inhibitory property of miRNA on target geneswe chose from the list of 8 downregulated genes (ITGB1AMIGO2 VAV3 PSEN2 LHFPL2 HS2ST1 AHSA2 andUQCRB) in miR-183-overexpressing cells Biological func-tion analysis using GO databases revealed that ITGB1 and
AMIGO2 were involved in cell adhesion andor cell migra-tionThese two geneswere selected for further study PubMedreports showed VAV3 and PSEN2 were both involved in cellinvasion [10 11] and these two genes were included as studytargets as well
Western blotting (Figure 2) was used to confirm theiralterations on protein levels Using the initial sample sets andthe criteria of ge15-fold of change integrin 1205731 yielded consis-tent results by the two technologies Compared to the controlgroup the levels of integrin 1205731 showed a significant decreasein the miR-183-overexpression group whereas the miR-183-downexpressing group (119875 lt 005) showed a sharp increaseAlthough endometrial stromal cells express AMIGO2 pro-tein the amount was largely unchanged in the miR-183 up-or downexpression groups suggesting it was not regulatedby miR-183 On the other hand no protein expression of
BioMed Research International 5
Table 2 List of genes with fold of change ge15 (119875 lt 005) and the pathways involved
Gene Gene set name Pathways 119875
ITGB1 SHC1 BIOCARTA PTEN PATHWAY PTEN dependent cell cycle arrest andapoptosis 308119864 minus 03
ITGB1 SHC1 BIOCARTA TFF PATHWAY Trefoil factors initiate mucosal healing 419119864 minus 03
ITGB1 SHC1 BIOCARTA ECM PATHWAY Erk and PI-3 kinase are necessary forcollagen binding in corneal epithelia 546119864 minus 03
ITGB1 SHC1 BIOCARTA ERK PATHWAY Erk1Erk2 Mapk signaling pathway 739119864 minus 03
ITGB1 SHC1 BIOCARTA INTEGRIN PATHWAY Integrin signaling pathway 134119864 minus 02
ITGB1 TUBB KEGG PATHOGENIC ESCHERICHIA COLI INFECFECTION Pathogenic Escherichiacoli infection 306119864 minus 02
SHC1 VAV3GNB2 KEGG CHEMOKINE SIGNALING PATHWAY Chemokine signaling pathway 575119864 minus 02
ITGB1 SHC1VAV3 KEGG FOCAL ADHESION Focal adhesion 659119864 minus 02
ITGB1 VAV3MYH9 KEGG REGULATION OF ACTIN CYTOSKELETON Regulation of actin cytoskeleton 782119864 minus 02
ITGB1 VAV3 KEGG LEUKOCYTE TRANSENDOTHELIAL MIGRATRATION Leukocyte transendothelial migration 105119864 minus 01
002040608
11214
U937
CON
NC-
low
dos
e
MiR
-dow
n-lo
w d
ose
MiR
-up-
low
dos
e
NC-
high
dos
e
MiR
-dow
n-hi
gh d
ose
MiR
-up-
high
dos
e
lowastlowast
lowastlowast
Integrin 1205731
GAPDH
1 2 3 4 5 6 7 8
Rela
tive i
nteg
rin1205731
prot
ein
leve
ls
(a)
0
02
04
06
08
1
12
Relat
ive A
MIG
O2
U937
CON
NC-
low
dos
e
MiR
-dow
n-lo
w d
ose
MiR
-up-
low
dos
e
NC-
high
dos
e
MiR
-dow
n-hi
gh d
ose
MiR
-up-
high
dos
e
Integrin 1205731
GAPDH
1 2 3 4 5 6 7 8pr
otei
n le
vels
(b)
Figure 2 Confirmation of microarray data withWestern blotting analysesThe endometrial stromal cells transfected withmiR-183-lentivirus(low dose or high dose) In-miR-183-lentivirus (low dose or high dose) and GFP-lentivirus (NC normal control) were used to examine theexpression alterations of integrin 1205731 (a) and AMIGO2 (b) 120573-actin served as protein loading control Compared with the control groupintegrin 1205731 expression in the miR-183-overexpression group was significantly decreased whereas it increased sharply in cells with low miR-183 expression AMIGO2 was largely not regulated by miR-183 lowast119875 lt 005 when compared to the negative control Error bars represent plusmnSEM
Presenilin 2 and VAV3 was detected in endometrial stromalcells (data not shown) and these two genes were not furtherstudied
35 MiR-183 Directly Targets ITBG1 but Not AMIGO2 Toinvestigate if ITBG1 and AMIGO2 are the direct targets of
miR-183 we performed luciferase assay to determinewhethermiR-183 binds to the 31015840UTR of ITBG1AMIGO2 mRNA(Figure 3(a)) Our results showed that reporter plasmidcontaining the miR-183 targeting sequence of ITGB1 mRNAdisplayed a significantly decreased luciferase activity in cellstransfected with miR-183 (119875 lt 005) whereas luciferase
6 BioMed Research International
3998400UTR5998400UTRAMIGO2
Has-miR-183
Mutant binding site
Coding region
5998400-UUCAGUUCCUCAAUGUGCCAUA-3998400
3998400-UCACUUAAGAUGGUCACGGUAU-5998400
5998400-UUCAGUUCCUCAAUCAUAACGC-3998400
174ndash181 bp
3998400UTR5998400UTRAMIGO2
Mutant binding site
Coding region
5998400-UCAUCAUAAUUUGUUGCCAUAA-3998400
3998400-CACUUAAGAUGGUCACGGUAU-5998400
5998400-UCAUCAUAAUUUGUAUAACGC-3998400
691ndash697 bp
3998400UTR5998400UTR
ITGB1
Has-miR-183
Mutant binding site
Coding region
5998400-GCUUUAAAACCUGUGUGCCAUU-3998400
3998400-UCACUUAAGAUGGUCACGGUAU-5998400
5998400-GCUUUAAAACCUGUUGUGACGU-3998400
695ndash701 bp
ITGB1 3998400UTR
998400Mutant 3 UTR
AMIGO2 3998400UTR
Mutant 3998400UTR
Has-miR-183
AMIGO2 3998400UTR
Mutant 3998400UTR
(a)
0
02
04
06
08
1
12
14
16
ITGB1-NC Wild-type Mutant type
Relat
ive l
ucife
rase
activ
ity (I
TGB1
)
MiR-183-NCMiR-183
0
02
04
06
08
1
12
14
16
AMIGO2-NC Wild-type Mutant type
Relat
ive L
ucife
rase
activ
ity (A
MIG
O2)
MiR-183-NCMiR-183
3998400UTR 3998400UTR 3998400UTR 3998400UTR
lowast
(b)
Figure 3 ITGB1 but not AMIGO2 is the direct target of miR-183 (a)The binding site of miR-183 in the 31015840UTR of ITGB1 (top) and AMIGO2(bottom) 31015840UTR along with the mutant construct in the predicted binding region (b) MiR-183 but not AMIGO2 directly targets ITBG1Endometrial stromal cells were cotransfected with wild-type reporter containing the ITBG1AMIGO2 31015840UTR or mutant ITBG1AMIGO231015840UTR plus miR-183 or negative control using Invitrogen Lipofectamine 2000 reagents Luminescence was measured after 24 hours oftransfection Reporter plasmid expressing ITGB1 mRNA that contains sequences potentially targeted by miR-183 displayed a significantlydecreased luciferase activity in cells transfected with miR-183 (119875 lt 005) whereas luciferase activity of reporter plasmid containing mutantsequence of ITGB1 was not changed following forced expression of miR-183 lowast119875 lt 005 when compared to the negative control Error barsrepresent plusmn SEM
BioMed Research International 7
0
02
04
06
08
1
12
14
Normal EM Eutopic EM
ITG
B1 ex
pres
sion
(nor
mal
ized
)
lowast
Figure 4 Validation of ITGB1 expression in eutopic endometriumwith endometriosis (119899 = 19) versus normal group (119899 = 18) byqPCR The results were analyzed using the comparative thresholdcycle (CT) method Gene expression data is presented as the foldof change relative to the levels of reference gene lowast119875 lt 005 whencompared to the normal group Error bars represent plusmn SEM
activity of reporter plasmid containing mutant sequence ofITGB1 was not changed following forced expression of miR-183 (Figure 3(b)) We found no significant change whencompared to the luciferase activity between cells transfectedwith wild-type or mutated AMIGO2 31015840UTR (119875 gt 005Figure 3(b) Thus these data suggested that miR-183 nega-tively regulates the expression of ITBG1 by directly target-ing their 31015840UTR whereas miR-183 does not directly targetAMIGO2 31015840UTR
36 ITGB1 Is Overexpressed in Endometrium from Endomet-riosis Patients Real-time RT-PCR results indicated thatITGB was significantly increased in the eutopic endometrialtissues from patients with endometriosis (119899 = 19) comparedwith endometrium from normal women (119899 = 18) (119875 lt 005)(Figure 4)
37 Overexpression of ITGB1 Rescued the Repressive Effectsof miR-183 on Endometrial Stromal Cells To ascertain thatmiR-183 regulates the function of endometrial stromal cellsthrough its interaction with ITGB1 a rescue experiment wasperformed Overexpression of ITGB1 partially rescued therepressive effects of miR-183 leading to elevated invasionabilities in the cells (Figure 5) This data indicated that miR-183 targets ITGB1 which in turn led to negative regulation onthe invasive activity of endometrial stromal cells
4 Discussion
In the previous study we performed miRNA microarrayscreening and identified several differentially expressed miR-NAs in the normal eutopic and ectopic endometriumAmong these miRNA species miR-183 was found to be
downregulated in the ectopic and eutopic tissues Functionalanalysis indicated that miR-183 promoted endometrial stro-mal cell apoptosis and had a negative regulatory impact onthe invasive ability of cells although it had no effect on cellproliferation [8] These findings suggested that the down-regulation of miR-183 expression might be involved in thepathogenesis of endometriosis The molecular mechanismshowever remain to be characterized
MiR-183 is a member of a miRNA family (miR-183 miR-182 and miR-96) that clusters within a 2ndash4 kb region atchromosome 7q32These miRNAs are known to regulate celldifferentiation apoptosis motility adhesion and invasion[12] Moreover it has been reported that miR-183 is upregu-lated in colorectal cancer prostate cancer and hepatocellularcarcinomas and downregulated in ovarian cancer breastcancer stem cells and osteosarcomas pointing to varied andcell type-dependent function of miR-183 Several target genesof miR-183 including ITGB1 Taok1 Ezrin EGR1 PDCD4and LRP6 have been identified and validated [13ndash19]
To systematically investigate the potential mechanismby which miR-183 may contribute to the development ofendometriosis we performed microarray analysis to identifythe target genes with relevance to cell functions in miR-183-overexpressing endometrial stromal cells Eight genes werefound to be downregulated in endometrial stromal cells withmiR-183 overexpression Among them the GO databasespathway analyses and PubMed reports pointed to importantassociation of 4 genes (ITGB1 AMIGO2 VAV3 and PSEN2)with cell adhesion cell migration and cell invasivenessWestern blotting results showed that endometrial stromalcell secreted integrin 1205731 and AMIGO2 but not VAV3 andPresenilin 2 The protein expression of integrin 1205731 appearedto be a direct regulation target of miR-183 which has beenproved in human trabecular meshwork cells human diploidfibroblasts and hela cells [19] AMIGO2 was not directlyregulated by miR-183
According to Sampsonrsquos theory endometriosis is thoughtto be initiated via retrograde endometrium into peri-toneal cavity during menstruation However while this phe-nomenon occurs in approximately 75ndash90 of women farfewer suffer the disease suggesting that additional factorsmust contribute to the process Invasion to ectopic locations isthe key step for the initiation of endometriosis during whichadhesion molecules play an important role The integrinfamily includes glycoproteins that formdimeric structure andmediate cell attachment to the ECM components [20] It hasbeen reported that 1205731 integrin-depleted cells had reducedinvasive capabilities and 1205731 integrin signaling was involvedin cancer cell invasion [21] Integrin 1205731 is constitutivelyexpressed in endometrial stromal cells and endometrioticlesions [22] Increased integrin 1205731 protein expression inendometrial stromal cells was observed in endometrioticcompared to normal endometrial tissues [23] It was reportedthat downregulation of prostaglandin E2 receptors EP2 andEP4 led to inhibited adhesion of human endometrioticepithelial and stromal cells Importantly this inhibitioncould be mediated through suppression of integrin-mediatedmechanisms [24] This finding provided a strong evidence
8 BioMed Research International
(a)
0
05
1
15
2
25
NC MiR-183 MiR-183ITGB1
Inva
sive r
ate (
OO
D57
0)
lowast
(b)
Figure 5 Overexpression of ITGB1 rescued the repressive effects of miR-183 on endometrial stromal cells leading to elevated invasiveabilities in transwell assays Endometrial stromal cells were infected with miR-183-lentivirus miR-183ITGB1-lentivirus and GFP-lentivirusin upper wells After 24 hours the number of cells that invaded through Matrigel was counted in at least 10 fields per well (a) Representativephotographs show that miR-183 inhibited the invasiveness of endometrial stromal cells whereas ITGB1 partially rescued the repressive effectsof miR-183 (b) Cell counting results indicate that ITGB1 overexpression rescued the repressive effects of miR-183 lowast119875 lt 005 when comparedto the negative control Error bars represent plusmn SEM
for the significance of integrins in the adhesion function ofendometrial stromal cells
AMIGO2 (ALI1 DEGA) was first reported to be prefer-entially expressed in the central nervous system Sequenceanalyses revealed that the protein contains seven leucine-richrepeats one IgC2-like loop and a transmembrane domainand displays homology to Kek and Trk families [25ndash27] Inthe gastric adenocarcinoma cell line AMIGO2 was reportedto be involved in cell adhesion extracellular matrix andbasement membrane formation [28] AMIGO2 was alsoimplicated in oral carcinogenesis by preventing the arrest ofcell cycle progression at the G1 phase through inhibiting theexpression of cyclin-dependent kinase inhibitors Howeverlittle is known about the role of AMIGO2 in the developmentof endometriosis
To investigate whether miR-183 directly targets ITGB1and AMIGO2 dual luciferase reporter assays were carriedout We found that miR-183 negatively regulated the expres-sion of ITBG1 by directly targeting its 31015840UTR but mightnot directly regulate the expression of AMIGO2 We further
detected differential expression of ITGB1 in endometriumwith or without endometriosis by quantitative RT-PCR Thehigher expression of integrin 1205731 in endometrium tissues inendometriosis was confirmed which is consistent with theresult for higher integrin 1205731 expression in ectopic stromalcells than normal endometrial stromal cells Previous studieshave shown that integrin 1205731 mediates cell adherence toextracellular matrix [29] Indeed the rescue experimentindicated that overexpression of ITGB1 blocked the repressiveeffects of miR-183 on endometrial stromal cell invasivenesswhich provided a critical evidence supporting that the lowerexpression of miR-183 may promote ITGB1 expression andcontribute to the development of endometriosis
In conclusion altered miR-183 expression may causederegulation of its target gene ITBG1 affecting the celladhesiveness and invasiveness of endometrial stromal cellswhich in turn lead to pathogenesis of endometriosis Thesefindings revealed a novel pathologic role of miR-183 forendometriosis It would be of great interest to investigate ifthese alterations may serve as biomarkers for the prediction
BioMed Research International 9
andor treatment of endometriosisThe regulation ofmiR-183and the exact mechanism by which miR-183 regulates ITBG1expression remain to be characterized
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This studywas supported by grants from theNational NaturalScience Foundation of China (nos 81100405 and 81471438)the Key Program ofNanjingMedical Science and TechnologyDevelopment Foundation Nanjing Department of Health(no QRX11109)
References
[1] D R Nyholt S-K Low C A Anderson et al ldquoGenome-wide association meta-analysis identifies new endometriosisrisk locirdquo Nature Genetics vol 44 no 12 pp 1355ndash1359 2012
[2] W Filipowicz S N Bhattacharyya and N Sonenberg ldquoMecha-nisms of post-transcriptional regulation by microRNAs are theanswers in sightrdquoNature Reviews Genetics vol 9 no 2 pp 102ndash114 2008
[3] X Santamaria and H Taylor ldquoMicroRNA and gynecologicalreproductive diseasesrdquo Fertility and Sterility vol 101 no 6 pp1545ndash1551 2014
[4] Y Shimono M Zabala R W Cho et al ldquoDownregulation ofmiRNA-200c links breast cancer stem cells with normal stemcellsrdquo Cell vol 138 no 3 pp 592ndash603 2009
[5] M Adammek B Greve N Kassens et al ldquoMicroRNA miR-145 inhibits proliferation invasiveness and stem cell phenotypeof an in vitro endometriosis model by targeting multiplecytoskeletal elements and pluripotency factorsrdquo Fertility andSterility vol 99 no 5 pp 1346e5ndash1355e5 2013
[6] W Abe K Nasu C Nakada Y Kawano M Moriyama andH Narahara ldquoMiR-196b targets c-myc and Bcl-2 expressioninhibits proliferation and induces apoptosis in endometrioticstromal cellsrdquo Human Reproduction vol 28 no 3 pp 750ndash7612013
[7] W B Nothnick A Graham J Holbert et al ldquomiR-451deficiency is associated with altered endometrial fibrinogenalpha chain expression and reduced endometriotic implantestablishment in an experimental mouse modelrdquo PLoS ONEvol 9 no 6 Article ID e100336 2014
[8] X-Y Shi L Gu J Chen X-R Guo and Y-L Shi ldquoDownregu-lation of miR-183 inhibits apoptosis and enhances the invasivepotential of endometrial stromal cells in endometriosisrdquo Inter-national Journal of Molecular Medicine vol 33 no 1 pp 59ndash672014
[9] Y-L Shi X-Z Luo X-Y Zhu K-Q Hua Y Zhu and D-JLi ldquoEffects of combined 17120573-estradiol with TCDD on secretionof chemokine IL-8 and expression of its receptor CXCR1 inendometriotic focus-associated cells in co-culturerdquo HumanReproduction vol 21 no 4 pp 870ndash879 2006
[10] B Salhia N L Tran A Chan et al ldquoThe guanine nucleotideexchange factors Trio Ect2 and Vav3 mediate the invasivebehavior of glioblastomardquo American Journal of Pathology vol173 no 6 pp 1828ndash1838 2008
[11] B Liu L Wang L-L Shen et al ldquoRNAi-mediated inhibitionof presenilin 2 inhibits glioma cell growth and invasion andis involved in the regulation of Nrg1ErbB signalingrdquo Neuro-Oncology vol 14 no 8 pp 994ndash1006 2012
[12] S D Weeraratne V Amani N Teider et al ldquoPleiotropiceffects of miR-183sim96sim182 converge to regulate cell survivalproliferation and migration in medulloblastomardquo Acta Neu-ropathologica vol 123 no 4 pp 539ndash552 2012
[13] M Patel Q Cai D Ding R Salvi Z Hu and B H Hu ldquoThemiR-183Taok1 target pair is implicated in cochlear responsesto acoustic traumardquo PLoS ONE vol 8 no 3 Article ID e584712013
[14] J Zhu Y Feng Z Ke et al ldquoDown-regulation of miR-183promotes migration and invasion of osteosarcoma by targetingEzrinrdquo The American Journal of Pathology vol 180 no 6 pp2440ndash2451 2012
[15] A J Lowery N Miller R M Dwyer and M J Kerin ldquoDysreg-ulated miR-183 inhibits migration in breast cancer cellsrdquo BMCCancer vol 10 article 502 2010
[16] A L Sarver H Li and S Subramanian ldquoMicroRNA miR-183functions as an oncogene by targeting the transcription factorEGR1 and promoting tumor cell migrationrdquo Cancer Researchvol 70 no 23 pp 9570ndash9580 2010
[17] J Li H Fu C Xu et al ldquoMiR-183 inhibits TGF-beta1-inducedapoptosis by downregulation of PDCD4 expression in humanhepatocellular carcinoma cellsrdquoBMCCancer vol 10 article 3542010
[18] C Chen H Xiang Y-L Peng J Peng and S-W JiangldquoMature miR-183 negatively regulated by transcription factorGATA3 promotes 3T3-L1 adipogenesis through inhibition ofthe canonical Wnt120573-catenin signaling pathway by targetingLRP6rdquo Cellular Signalling vol 26 no 6 pp 1155ndash1165 2014
[19] G Li C Luna J Qiu D L Epstein and P Gonzalez ldquoTargetingof integrin beta1 and kinesin 2alpha by microRNA 183rdquo TheJournal of Biological Chemistry vol 285 no 8 pp 5461ndash54712010
[20] A M Schooley N M Andrews H Zhao and C L Addisonldquo1205731 integrin is required for anchorage-independent growth andinvasion of tumor cells in a context dependent mannerrdquo CancerLetters vol 316 no 2 pp 157ndash167 2012
[21] Y Zhang YGongDHu et al ldquoNuclear SIPA1 activates integrin1205731 promoter and promotes invasion of breast cancer cellsrdquoOncogene vol 34 no 11 pp 1451ndash1462 2015
[22] Y B Aznaurova M B Zhumataev T K Roberts A M Aliperand A A Zhavoronkov ldquoMolecular aspects of developmentand regulation of endometriosisrdquo Reproductive Biology andEndocrinology vol 12 article 50 2014
[23] M Adachi K Nasu A Tsuno A Yuge Y Kawano and HNarahara ldquoAttachment to extracellular matrices is enhancedin human endometriotic stromal cells a possible mechanismunderlying the pathogenesis of endometriosisrdquo European Jour-nal of Obstetrics Gynecology and Reproductive Biology vol 155no 1 pp 85ndash88 2011
[24] J Lee S K Banu R C Burghardt A Starzinski-Powitz andJ A Arosh ldquoSelective inhibition of prostaglandin E2 receptorsEP2 and EP4 inhibits adhesion of human endometriotic epithe-lial and stromal cells through suppression of integrin-mediatedmechanismsrdquo Biology of Reproduction vol 88 no 3 article 772013
[25] A Laeremans J NysW Luyten R DrsquoHoogeM Paulussen andL Arckens ldquoAMIGO2 mRNA expression in hippocampal CA2
10 BioMed Research International
and CA3ardquo Brain Structure and Function vol 218 no 1 pp 123ndash130 2013
[26] T Ono N Sekino-Suzuki Y Kikkawa H Yonekawa andS Kawashima ldquoAlivin 1 a novel neuronal activity-dependentgene inhibits apoptosis and promotes survival of cerebellargranule neuronsrdquo Journal of Neuroscience vol 23 no 13 pp5887ndash5896 2003
[27] J Kuja-Panula M Kiiltomaki T Yamashiro A Rouhiainenand H Rauvala ldquoAMIGO a transmembrane protein implicatedin axon tract development defines a novel protein family withleucine-rich repeatsrdquoThe Journal of Cell Biology vol 160 no 6pp 963ndash973 2003
[28] K E Rabenau J M OrsquoToole R Bassi et al ldquoDEGAAMIGO-2 a leucine-rich repeat family member differentially expressedin human gastric adenocarcinoma effects on ploidy chromo-somal stability cell adhesionmigration and tumorigenicityrdquoOncogene vol 23 no 29 pp 5056ndash5067 2004
[29] J A Garcia-Velasco and A Arici ldquoInterleukin-8 expression inendometrial stromal cells is regulated by integrin-dependentcell adhesionrdquo Molecular Human Reproduction vol 5 no 12pp 1135ndash1140 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 BioMed Research International
associated with decreased establishment of endometriosis ina mouse model [7] These data landed strong support foran important role of miRNAs in the development of endo-metriosis through regulating cell proliferation cell apoptosiscell migration cell invasiveness and estrogenic signaling
In a previous study we identified several differentially ex-pressed miRNAs among normal eutopic and ectopic endo-metrium MiR-183 was found to be the most downregulatedmiRNA in both the ectopic and eutopic tissues comparedto normal endometrium Functional studies indicated thatmiR-183may contribute to endometrial stromal cell apoptosisand impose a negative regulatory impact on cell invasivenessbut it has no effect on endometrial stromal cell proliferationThe study suggested that aberrant miR-183 expressionmay beinvolved in the development and progression of endometrio-sis [8]
In the present study we aim to identify the functionaltargets of miR-183 using an overexpression model Weexamine if miR-183 exerts its effects by directly binding toand inhibiting the ITGB1 promoter activity In addition weinvestigate ifmiR-183 could regulate stromal cell invasivenessThe findings from these experiments will help us to betterunderstand the miR-183-mediated molecular mechanism inendometric stromal cells
2 Materials and Methods
21 Tissue Acquisition and Cell Culture All the eutopic andnormal endometrial tissues were obtained at the NanjingMaternity and Child Health Care Hospital Affiliated to Nan-jing Medical University by uterine curettage from patientswith or without endometriosis None of the patients hadreceived preoperative hormonal therapy and all samples werehistologically confirmed by pathologists All samples werecollected from the proliferative phase of the menstrual cycleThe phase of the menstrual cycle was determined based onhistologic evaluation of the endometrium with the assistanceof pathologists The average age of the patients was 305 plusmn52 Patients consented to tissue donation prior to surgeryThe research project and consent form were approved by thehospital ethics committee Each sample was divided into twoparts one part was snap-frozen and stored at minus20∘C and laterused for mRNA and protein extraction the other part wasrinsed thoroughly with cold PBS and directly subjected todigestion and stromal cell isolation
The endometrial stromal cells (ESC) fromnormal womenwithout endometriosis were used as control Tissue digestionand stromal cell isolation and culturewere performed accord-ing to the method described in previous publication [9]
22 MiR-183-Lentivirus Construction and Transduction Thelentivirus gene transfer vector carrying precursor of hsa-miR-183 (Genbank accession number MIMAT0000261) andencoding green fluorescence protein (GFP) were constructedby Genechem Co Ltd Shanghai China and confirmed byDNA sequencing The primers of RNA were 51015840-GAGGAT-CCCCGGGTACCAAGGGAGTGGGCAGGCTA and 51015840-ATAAGCTTGATATCGTCCCTGCACCCTTGGAAGCA
The recombinant lentivirus formiR-183 overexpression (miR-183-lentivirus) and the control lentivirus (GFP-lentivirus)were prepared and titered to 50119864 + 8TUmL (transfectionunit per mL)
The sequence of inhibitor of hsa-miR-183-5p was TATG-GCACTGGTAGAATTCACT The recombinant lentivirus ofmiR-183-5p inhibitor (In-miR-183 lentivirus) and the controllentivirus (GFP-lentivirus) were prepared and titered to40119864 + 8TUmL (transfection unit per mL)
ESC from women without endometriosis were plated in6-well plates (5 times 104 cellswell) overnight The lentiviruseswere diluted in 02mL complete medium containing poly-brene (8mgmL) and added to the cells for 12 h of incubationat 37∘C followed by incubation in 03mL of freshly preparedpolybrene-DMEM for 24 h The media were replaced withfresh DMEM and the cells were cultured for 3 days Thelentivirus transduction efficiency of ESC was determined bythe detection of GFP signals under a fluorescencemicroscopeat 72 h after transduction The miR-183 expression in stablytransduced ESC was measured by real-time PCR The ESCtransfected with miR-183-lentivirus In-miR-183-lentivirusand GFP-lentivirus were kept for further analysis
23 RNA Extraction and Microarray For the microarrayanalyses groups were divided into the ESC with miR-183 overexpression and the control ones Total RNA wasextracted using TRIzol (Invitrogen) according to the man-ufacturerrsquos instructions Gene expression profiling was con-ducted using PrimeView Human Gene Expression Array(Affymetrix) The array contains 530000 probes coveringmore than 36000 transcripts and variants which representmore than 20000 genes mapped through RefSeq or viaUniGene annotation All subsequent technical proceduresand quality controls were performed by Genechem Co LtdShanghai China The arrays were scanned using a GeneChipScanner 3000 (Affymetrix Inc CA USA) Raw data wereextracted from the scanned images and analyzed usingGeneSpring GX software version 115 (Agilent TechnologiesCA USA)
The data were normalized using the PLIER default proto-col Significant differentially expressed genes were analyzedusing an unpaired 119905-test Hierarchical cluster analysis wasused to assess correlations among samples for each identifiedgene set with Euclidean distance and average linkage statisti-cal methods
24MicroarrayDataAnalyses Differentially expressed geneswere uploaded to the Ingenuity database (Ingenuity Sys-tems Redwood City CA USA) for pathway and functionalanalyses Genes were further annotated and classified basedon the Gene Ontology (GO) consortium annotations fromthe GO Bostaurus database using GOEAST (Gene OntologyEnrichment Analysis Software Toolkit)
25Western Blotting ESC proteins were extracted with RIPAbuffer [(50mM Tris-HCl pH 80 150mM NaCl 1 (vv)NP-40 05 (vo) sodium deoxycholate and 01 (wo)
BioMed Research International 3
sodium dodecyl sulphate (SDS)] supplemented with a pro-tease inhibitor cocktail (Sigma) After incubating on ice for30min cell debris was removed by centrifugation at 12000 gfor 10min Cell lysates were resolved in 12 SDS-PAGEelectrophoresis and transferred to the PVDFmembrane (Bio-Rad Hercules CA) Nonspecific binding was blocked byincubating the membranes in Tris-buffered saline containing5 nonfat milk (TBST 50mmolL TrisndashHCl 150mmolLNaCl and 01 Tween-20) for 1 h at room temperatureMembranes were incubated overnight at 4∘C with AMIGO2antibody (1 400 RD) integrin 1205731 antibody (1 1000 Cell Sig-naling) Presenilin 2 antibody (1 500 ABCAM) and VAV3antibody (1 500 ABCAM) After three washes with TBSTthe membranes were incubated for 1 h at room temperaturewith the horseradish peroxidase-labeled secondary antibodyThe signals were visualized with chemiluminescence andquantified with Quantity-One (Bio-Rad America)
26 Luciferase Assay The mutant construct of ITGB131015840UTR was obtained by introducing the mutation intothe 7 nucleotides (GUGCCAUU) of the seed region formiR-183 The mutant construct of AMIGO2 31015840UTR wasobtained by introducing twomutations into the 7 nucleotides(GUGCCAUA) (UGCCAUA) of the seed region for miR-183 The miR-183 target sequences in the coding region ofITGB1AMIGO2 were amplified by PCR and cloned intoGV143 that contained a firefly luciferase reporter gene Wild-type ITBG1AMIGO2 31015840UTR or mutant ITBG1AMIGO231015840UTR and the empty 31015840UTR vector were cotransfected intoESC Cell transfection using Lipofectamine 2000 and nor-malization for transfection efficiency was performed accord-ing to the recommendation of manufacturer (Invitrogen)Luciferase activity was measured 24 h after transfection usingthe Dual-Luciferase Reporter Assay System (Promega CorpMadison WI)
27 Quantitative RT-PCR Endometrial tissue was mincedwith a scalpel blade Total RNA was isolated using TRIzol(Takara Otsu Shiga Japan) and cDNA was synthesizedusing the SYBR Prime Script RT-PCR kit (Takara) on theABI Prism 7500 Sequence Detection System according tomanufacturerrsquos instructions The housekeeping gene 120573-actinwas used for normalization The primers used were 51015840-TCG-TCACGTTCCGGTTATTC (sense) and 51015840-CTTTTACTT-ACGGTTTACCC (antisense) for ITGB1 51015840-CCTCGCCTT-TGCCGATCCG (sense) and 51015840-GCCGGAGCCGTTGTC-GACG (antisense) for 120573-actin The cycling conditions were94∘C for 1min 35 cycles of 94∘C for 30 sec 66∘C for 30 secand 68∘C for 3min followed by extension at 68∘C for3min Quantitative PCR was carried out using SYBR-GreenJumpStart Taq ReadyMix (Sigma) and the 7300 Real-TimePCRDetection System (ABI)The results were analyzed usingthe comparative threshold cycle (CT) method
28 Rescue Experiment To further validate direct targetingof ITGB1 by miR-183 functional rescue experiment was per-formed by cotransfection with miR-183 mimic and plasmidconstructs expressing ITGB1 in ESC using Lipofectamine
2000 (Invitrogen) as described aboveThe expression plasmidpcDNA-ITGB1 encoding human ITGB1was purchased fromGenechem The presence of complete ITGB1 coding regionswas confirmed by DNA sequencing
29 Invasion (Matrigel) Chamber Assay ESC (25 times 104) wereseeded on a transwell insert coated with extracellular matrix(ECM) (8mm pore size 24-well format Corning Costar) in2 FBS medium Complete medium (10 FBS) was addedto the lower chamber To determine the amount of invasioncells were incubated for 24 h and then removed from theupper chamber using a cotton swab The invaded cells onthe underside of the insert were fixed with methanol stainedwith crystal violet for 2min and rinsed with phosphatebuffered saline (PBS) The undersides of the membrane werephotographed to compare the number of invaded cells perinsert The transmigrated cells were counted using a lightmicroscope Invasive cells were scored by counting 10 randomhigh-power fields per filter
210 Statistical Analyses Data represents mean plusmn SEM fromat least 3 independent experiments Difference betweenexperimental and control groupswas determined by Studentrsquos119905-test while one-way ANOVA analysis was employed tocompare three or more groups 119875 lt 005 was considered asstatistical significance
3 Results
31 Gene Expression Profiling following miR-183 Overexpres-sion In order to screen target genes in response to miR-183we used microarrays representing more than 20000 genesmapped through RefSeq or via UniGene annotation Westudied gene expression alterations (up- or downregulation)at 24 h after transfection The changes of gene expressionin miR-183-overexpressing endometrial stromal cells wereanalyzed Differential expression was found in 27 genes at119875 value lt 005 with folds of change ge15 Of these 19 wereupregulated and 8 downregulated (ITGB1 AMIGO2 VAV3PSEN2 LHFPL2 HS2ST1 AHSA2 and UQCRB) Resultsof hierarchical cluster analyses of these genes are shownin Figure 1 and supplementary 1 in Supplementary Materialavailable online at httpdxdoiorg1011552015340218
32 Functional Analysis with GO Databases By using theGene Ontology (GO) database we systematically extractedand analyzed the information of threeGOcategories ldquobiolog-ical processrdquo ldquomolecular functionrdquo and ldquocellular componentrdquoIt was revealed that the identified genes were involvedin hemophilic cell adhesion (ITGB1 AMIGO2) cell-celladhesion (ITGB1 AMIGO2) cell migration (ITGB1MYH9)positive regulation of catalytic activity (PSEN2 SHC1) andproteolysis (MYH9 PSEN2) (Table 1)
33 Significant Pathway Analysis Significant pathway analy-sis revealed that the gene expression alterations in endome-trial stromal cells were involved in pathways of PTEN(ITGB1 SHC1) TFF (ITGB1 SHC1) ECM (ITGB1 SHC1)
4 BioMed Research International
Table 1 List of genes with fold of change ge15 (119875 lt 005) and their biological functions
Gene Gene set name 119875
ITGB1 AMIGO2 Hemophilic cell adhesion 197119864 minus 03
ITGB1 AMIGO2 Cell to cell adhesion 502119864 minus 02
ITGB1 MYH9 Cell migration 610119864 minus 02
PSEN2 SHC1 Positive regulation of catalytic activity 152119864 minus 01
MYH9 PSEN2 Proteolysis 191119864 minus 01
UQCRBAMIGO2HS2ST1AHSA2VAV3LHFPL2ITGB1PSEN2PICALMHES4SHC1HYOU1PPIFHPCAL1TUBBCBSMZT1CDT1GNB2PPP2R5DPTPRHCHMP4BVPS35PTPRAMYH9C6orf150HDGF
minus1 minus05 0 05 1 15Row Z-score
X461_1 X461_3 X461_2 X462_3 X462_1 X462_2
Figure 1 Hierarchical clustering of differentially expressed genes in miR-183-overexpressing endometrial stromal cells versus control cellsGene expression profiling was conducted using PrimeViewHumanGene ExpressionArray Raw data were extracted from the scanned imagesand analyzed with GeneSpring GX software version 115 The data was normalized using the iterative PLIER default protocol Differentiallyexpressed genes were analyzed with the unpaired Student 119905-test Statistically different genes (119875 lt 005) with a greater than 15-fold increaseor decrease were recorded Red color indicates low expression green color indicates relatively high expression
ERK (ITGB1 SHC1) integrin (ITGB1 SHC1) pathogenicEscherichia coli infection (ITGB1 TUBB) chemokine sig-naling pathway (SHC1 VAV3 and GNB2) focal adhe-sion (ITGB1 SHC1 and VAV3) regulation of cytoskele-ton (ITGB1 VAV3 and MYH9) leukocyte transendothelialmigration (ITGB1 VAV3) natural killer cell-mediated cyto-toxicity (SHC1 VAV3) and Alzheimerrsquos disease (UQCRBPSEN2) (Table 2)
34 Confirmation of Microarray Data by Western BlottingBecause of the inhibitory property of miRNA on target geneswe chose from the list of 8 downregulated genes (ITGB1AMIGO2 VAV3 PSEN2 LHFPL2 HS2ST1 AHSA2 andUQCRB) in miR-183-overexpressing cells Biological func-tion analysis using GO databases revealed that ITGB1 and
AMIGO2 were involved in cell adhesion andor cell migra-tionThese two geneswere selected for further study PubMedreports showed VAV3 and PSEN2 were both involved in cellinvasion [10 11] and these two genes were included as studytargets as well
Western blotting (Figure 2) was used to confirm theiralterations on protein levels Using the initial sample sets andthe criteria of ge15-fold of change integrin 1205731 yielded consis-tent results by the two technologies Compared to the controlgroup the levels of integrin 1205731 showed a significant decreasein the miR-183-overexpression group whereas the miR-183-downexpressing group (119875 lt 005) showed a sharp increaseAlthough endometrial stromal cells express AMIGO2 pro-tein the amount was largely unchanged in the miR-183 up-or downexpression groups suggesting it was not regulatedby miR-183 On the other hand no protein expression of
BioMed Research International 5
Table 2 List of genes with fold of change ge15 (119875 lt 005) and the pathways involved
Gene Gene set name Pathways 119875
ITGB1 SHC1 BIOCARTA PTEN PATHWAY PTEN dependent cell cycle arrest andapoptosis 308119864 minus 03
ITGB1 SHC1 BIOCARTA TFF PATHWAY Trefoil factors initiate mucosal healing 419119864 minus 03
ITGB1 SHC1 BIOCARTA ECM PATHWAY Erk and PI-3 kinase are necessary forcollagen binding in corneal epithelia 546119864 minus 03
ITGB1 SHC1 BIOCARTA ERK PATHWAY Erk1Erk2 Mapk signaling pathway 739119864 minus 03
ITGB1 SHC1 BIOCARTA INTEGRIN PATHWAY Integrin signaling pathway 134119864 minus 02
ITGB1 TUBB KEGG PATHOGENIC ESCHERICHIA COLI INFECFECTION Pathogenic Escherichiacoli infection 306119864 minus 02
SHC1 VAV3GNB2 KEGG CHEMOKINE SIGNALING PATHWAY Chemokine signaling pathway 575119864 minus 02
ITGB1 SHC1VAV3 KEGG FOCAL ADHESION Focal adhesion 659119864 minus 02
ITGB1 VAV3MYH9 KEGG REGULATION OF ACTIN CYTOSKELETON Regulation of actin cytoskeleton 782119864 minus 02
ITGB1 VAV3 KEGG LEUKOCYTE TRANSENDOTHELIAL MIGRATRATION Leukocyte transendothelial migration 105119864 minus 01
002040608
11214
U937
CON
NC-
low
dos
e
MiR
-dow
n-lo
w d
ose
MiR
-up-
low
dos
e
NC-
high
dos
e
MiR
-dow
n-hi
gh d
ose
MiR
-up-
high
dos
e
lowastlowast
lowastlowast
Integrin 1205731
GAPDH
1 2 3 4 5 6 7 8
Rela
tive i
nteg
rin1205731
prot
ein
leve
ls
(a)
0
02
04
06
08
1
12
Relat
ive A
MIG
O2
U937
CON
NC-
low
dos
e
MiR
-dow
n-lo
w d
ose
MiR
-up-
low
dos
e
NC-
high
dos
e
MiR
-dow
n-hi
gh d
ose
MiR
-up-
high
dos
e
Integrin 1205731
GAPDH
1 2 3 4 5 6 7 8pr
otei
n le
vels
(b)
Figure 2 Confirmation of microarray data withWestern blotting analysesThe endometrial stromal cells transfected withmiR-183-lentivirus(low dose or high dose) In-miR-183-lentivirus (low dose or high dose) and GFP-lentivirus (NC normal control) were used to examine theexpression alterations of integrin 1205731 (a) and AMIGO2 (b) 120573-actin served as protein loading control Compared with the control groupintegrin 1205731 expression in the miR-183-overexpression group was significantly decreased whereas it increased sharply in cells with low miR-183 expression AMIGO2 was largely not regulated by miR-183 lowast119875 lt 005 when compared to the negative control Error bars represent plusmnSEM
Presenilin 2 and VAV3 was detected in endometrial stromalcells (data not shown) and these two genes were not furtherstudied
35 MiR-183 Directly Targets ITBG1 but Not AMIGO2 Toinvestigate if ITBG1 and AMIGO2 are the direct targets of
miR-183 we performed luciferase assay to determinewhethermiR-183 binds to the 31015840UTR of ITBG1AMIGO2 mRNA(Figure 3(a)) Our results showed that reporter plasmidcontaining the miR-183 targeting sequence of ITGB1 mRNAdisplayed a significantly decreased luciferase activity in cellstransfected with miR-183 (119875 lt 005) whereas luciferase
6 BioMed Research International
3998400UTR5998400UTRAMIGO2
Has-miR-183
Mutant binding site
Coding region
5998400-UUCAGUUCCUCAAUGUGCCAUA-3998400
3998400-UCACUUAAGAUGGUCACGGUAU-5998400
5998400-UUCAGUUCCUCAAUCAUAACGC-3998400
174ndash181 bp
3998400UTR5998400UTRAMIGO2
Mutant binding site
Coding region
5998400-UCAUCAUAAUUUGUUGCCAUAA-3998400
3998400-CACUUAAGAUGGUCACGGUAU-5998400
5998400-UCAUCAUAAUUUGUAUAACGC-3998400
691ndash697 bp
3998400UTR5998400UTR
ITGB1
Has-miR-183
Mutant binding site
Coding region
5998400-GCUUUAAAACCUGUGUGCCAUU-3998400
3998400-UCACUUAAGAUGGUCACGGUAU-5998400
5998400-GCUUUAAAACCUGUUGUGACGU-3998400
695ndash701 bp
ITGB1 3998400UTR
998400Mutant 3 UTR
AMIGO2 3998400UTR
Mutant 3998400UTR
Has-miR-183
AMIGO2 3998400UTR
Mutant 3998400UTR
(a)
0
02
04
06
08
1
12
14
16
ITGB1-NC Wild-type Mutant type
Relat
ive l
ucife
rase
activ
ity (I
TGB1
)
MiR-183-NCMiR-183
0
02
04
06
08
1
12
14
16
AMIGO2-NC Wild-type Mutant type
Relat
ive L
ucife
rase
activ
ity (A
MIG
O2)
MiR-183-NCMiR-183
3998400UTR 3998400UTR 3998400UTR 3998400UTR
lowast
(b)
Figure 3 ITGB1 but not AMIGO2 is the direct target of miR-183 (a)The binding site of miR-183 in the 31015840UTR of ITGB1 (top) and AMIGO2(bottom) 31015840UTR along with the mutant construct in the predicted binding region (b) MiR-183 but not AMIGO2 directly targets ITBG1Endometrial stromal cells were cotransfected with wild-type reporter containing the ITBG1AMIGO2 31015840UTR or mutant ITBG1AMIGO231015840UTR plus miR-183 or negative control using Invitrogen Lipofectamine 2000 reagents Luminescence was measured after 24 hours oftransfection Reporter plasmid expressing ITGB1 mRNA that contains sequences potentially targeted by miR-183 displayed a significantlydecreased luciferase activity in cells transfected with miR-183 (119875 lt 005) whereas luciferase activity of reporter plasmid containing mutantsequence of ITGB1 was not changed following forced expression of miR-183 lowast119875 lt 005 when compared to the negative control Error barsrepresent plusmn SEM
BioMed Research International 7
0
02
04
06
08
1
12
14
Normal EM Eutopic EM
ITG
B1 ex
pres
sion
(nor
mal
ized
)
lowast
Figure 4 Validation of ITGB1 expression in eutopic endometriumwith endometriosis (119899 = 19) versus normal group (119899 = 18) byqPCR The results were analyzed using the comparative thresholdcycle (CT) method Gene expression data is presented as the foldof change relative to the levels of reference gene lowast119875 lt 005 whencompared to the normal group Error bars represent plusmn SEM
activity of reporter plasmid containing mutant sequence ofITGB1 was not changed following forced expression of miR-183 (Figure 3(b)) We found no significant change whencompared to the luciferase activity between cells transfectedwith wild-type or mutated AMIGO2 31015840UTR (119875 gt 005Figure 3(b) Thus these data suggested that miR-183 nega-tively regulates the expression of ITBG1 by directly target-ing their 31015840UTR whereas miR-183 does not directly targetAMIGO2 31015840UTR
36 ITGB1 Is Overexpressed in Endometrium from Endomet-riosis Patients Real-time RT-PCR results indicated thatITGB was significantly increased in the eutopic endometrialtissues from patients with endometriosis (119899 = 19) comparedwith endometrium from normal women (119899 = 18) (119875 lt 005)(Figure 4)
37 Overexpression of ITGB1 Rescued the Repressive Effectsof miR-183 on Endometrial Stromal Cells To ascertain thatmiR-183 regulates the function of endometrial stromal cellsthrough its interaction with ITGB1 a rescue experiment wasperformed Overexpression of ITGB1 partially rescued therepressive effects of miR-183 leading to elevated invasionabilities in the cells (Figure 5) This data indicated that miR-183 targets ITGB1 which in turn led to negative regulation onthe invasive activity of endometrial stromal cells
4 Discussion
In the previous study we performed miRNA microarrayscreening and identified several differentially expressed miR-NAs in the normal eutopic and ectopic endometriumAmong these miRNA species miR-183 was found to be
downregulated in the ectopic and eutopic tissues Functionalanalysis indicated that miR-183 promoted endometrial stro-mal cell apoptosis and had a negative regulatory impact onthe invasive ability of cells although it had no effect on cellproliferation [8] These findings suggested that the down-regulation of miR-183 expression might be involved in thepathogenesis of endometriosis The molecular mechanismshowever remain to be characterized
MiR-183 is a member of a miRNA family (miR-183 miR-182 and miR-96) that clusters within a 2ndash4 kb region atchromosome 7q32These miRNAs are known to regulate celldifferentiation apoptosis motility adhesion and invasion[12] Moreover it has been reported that miR-183 is upregu-lated in colorectal cancer prostate cancer and hepatocellularcarcinomas and downregulated in ovarian cancer breastcancer stem cells and osteosarcomas pointing to varied andcell type-dependent function of miR-183 Several target genesof miR-183 including ITGB1 Taok1 Ezrin EGR1 PDCD4and LRP6 have been identified and validated [13ndash19]
To systematically investigate the potential mechanismby which miR-183 may contribute to the development ofendometriosis we performed microarray analysis to identifythe target genes with relevance to cell functions in miR-183-overexpressing endometrial stromal cells Eight genes werefound to be downregulated in endometrial stromal cells withmiR-183 overexpression Among them the GO databasespathway analyses and PubMed reports pointed to importantassociation of 4 genes (ITGB1 AMIGO2 VAV3 and PSEN2)with cell adhesion cell migration and cell invasivenessWestern blotting results showed that endometrial stromalcell secreted integrin 1205731 and AMIGO2 but not VAV3 andPresenilin 2 The protein expression of integrin 1205731 appearedto be a direct regulation target of miR-183 which has beenproved in human trabecular meshwork cells human diploidfibroblasts and hela cells [19] AMIGO2 was not directlyregulated by miR-183
According to Sampsonrsquos theory endometriosis is thoughtto be initiated via retrograde endometrium into peri-toneal cavity during menstruation However while this phe-nomenon occurs in approximately 75ndash90 of women farfewer suffer the disease suggesting that additional factorsmust contribute to the process Invasion to ectopic locations isthe key step for the initiation of endometriosis during whichadhesion molecules play an important role The integrinfamily includes glycoproteins that formdimeric structure andmediate cell attachment to the ECM components [20] It hasbeen reported that 1205731 integrin-depleted cells had reducedinvasive capabilities and 1205731 integrin signaling was involvedin cancer cell invasion [21] Integrin 1205731 is constitutivelyexpressed in endometrial stromal cells and endometrioticlesions [22] Increased integrin 1205731 protein expression inendometrial stromal cells was observed in endometrioticcompared to normal endometrial tissues [23] It was reportedthat downregulation of prostaglandin E2 receptors EP2 andEP4 led to inhibited adhesion of human endometrioticepithelial and stromal cells Importantly this inhibitioncould be mediated through suppression of integrin-mediatedmechanisms [24] This finding provided a strong evidence
8 BioMed Research International
(a)
0
05
1
15
2
25
NC MiR-183 MiR-183ITGB1
Inva
sive r
ate (
OO
D57
0)
lowast
(b)
Figure 5 Overexpression of ITGB1 rescued the repressive effects of miR-183 on endometrial stromal cells leading to elevated invasiveabilities in transwell assays Endometrial stromal cells were infected with miR-183-lentivirus miR-183ITGB1-lentivirus and GFP-lentivirusin upper wells After 24 hours the number of cells that invaded through Matrigel was counted in at least 10 fields per well (a) Representativephotographs show that miR-183 inhibited the invasiveness of endometrial stromal cells whereas ITGB1 partially rescued the repressive effectsof miR-183 (b) Cell counting results indicate that ITGB1 overexpression rescued the repressive effects of miR-183 lowast119875 lt 005 when comparedto the negative control Error bars represent plusmn SEM
for the significance of integrins in the adhesion function ofendometrial stromal cells
AMIGO2 (ALI1 DEGA) was first reported to be prefer-entially expressed in the central nervous system Sequenceanalyses revealed that the protein contains seven leucine-richrepeats one IgC2-like loop and a transmembrane domainand displays homology to Kek and Trk families [25ndash27] Inthe gastric adenocarcinoma cell line AMIGO2 was reportedto be involved in cell adhesion extracellular matrix andbasement membrane formation [28] AMIGO2 was alsoimplicated in oral carcinogenesis by preventing the arrest ofcell cycle progression at the G1 phase through inhibiting theexpression of cyclin-dependent kinase inhibitors Howeverlittle is known about the role of AMIGO2 in the developmentof endometriosis
To investigate whether miR-183 directly targets ITGB1and AMIGO2 dual luciferase reporter assays were carriedout We found that miR-183 negatively regulated the expres-sion of ITBG1 by directly targeting its 31015840UTR but mightnot directly regulate the expression of AMIGO2 We further
detected differential expression of ITGB1 in endometriumwith or without endometriosis by quantitative RT-PCR Thehigher expression of integrin 1205731 in endometrium tissues inendometriosis was confirmed which is consistent with theresult for higher integrin 1205731 expression in ectopic stromalcells than normal endometrial stromal cells Previous studieshave shown that integrin 1205731 mediates cell adherence toextracellular matrix [29] Indeed the rescue experimentindicated that overexpression of ITGB1 blocked the repressiveeffects of miR-183 on endometrial stromal cell invasivenesswhich provided a critical evidence supporting that the lowerexpression of miR-183 may promote ITGB1 expression andcontribute to the development of endometriosis
In conclusion altered miR-183 expression may causederegulation of its target gene ITBG1 affecting the celladhesiveness and invasiveness of endometrial stromal cellswhich in turn lead to pathogenesis of endometriosis Thesefindings revealed a novel pathologic role of miR-183 forendometriosis It would be of great interest to investigate ifthese alterations may serve as biomarkers for the prediction
BioMed Research International 9
andor treatment of endometriosisThe regulation ofmiR-183and the exact mechanism by which miR-183 regulates ITBG1expression remain to be characterized
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This studywas supported by grants from theNational NaturalScience Foundation of China (nos 81100405 and 81471438)the Key Program ofNanjingMedical Science and TechnologyDevelopment Foundation Nanjing Department of Health(no QRX11109)
References
[1] D R Nyholt S-K Low C A Anderson et al ldquoGenome-wide association meta-analysis identifies new endometriosisrisk locirdquo Nature Genetics vol 44 no 12 pp 1355ndash1359 2012
[2] W Filipowicz S N Bhattacharyya and N Sonenberg ldquoMecha-nisms of post-transcriptional regulation by microRNAs are theanswers in sightrdquoNature Reviews Genetics vol 9 no 2 pp 102ndash114 2008
[3] X Santamaria and H Taylor ldquoMicroRNA and gynecologicalreproductive diseasesrdquo Fertility and Sterility vol 101 no 6 pp1545ndash1551 2014
[4] Y Shimono M Zabala R W Cho et al ldquoDownregulation ofmiRNA-200c links breast cancer stem cells with normal stemcellsrdquo Cell vol 138 no 3 pp 592ndash603 2009
[5] M Adammek B Greve N Kassens et al ldquoMicroRNA miR-145 inhibits proliferation invasiveness and stem cell phenotypeof an in vitro endometriosis model by targeting multiplecytoskeletal elements and pluripotency factorsrdquo Fertility andSterility vol 99 no 5 pp 1346e5ndash1355e5 2013
[6] W Abe K Nasu C Nakada Y Kawano M Moriyama andH Narahara ldquoMiR-196b targets c-myc and Bcl-2 expressioninhibits proliferation and induces apoptosis in endometrioticstromal cellsrdquo Human Reproduction vol 28 no 3 pp 750ndash7612013
[7] W B Nothnick A Graham J Holbert et al ldquomiR-451deficiency is associated with altered endometrial fibrinogenalpha chain expression and reduced endometriotic implantestablishment in an experimental mouse modelrdquo PLoS ONEvol 9 no 6 Article ID e100336 2014
[8] X-Y Shi L Gu J Chen X-R Guo and Y-L Shi ldquoDownregu-lation of miR-183 inhibits apoptosis and enhances the invasivepotential of endometrial stromal cells in endometriosisrdquo Inter-national Journal of Molecular Medicine vol 33 no 1 pp 59ndash672014
[9] Y-L Shi X-Z Luo X-Y Zhu K-Q Hua Y Zhu and D-JLi ldquoEffects of combined 17120573-estradiol with TCDD on secretionof chemokine IL-8 and expression of its receptor CXCR1 inendometriotic focus-associated cells in co-culturerdquo HumanReproduction vol 21 no 4 pp 870ndash879 2006
[10] B Salhia N L Tran A Chan et al ldquoThe guanine nucleotideexchange factors Trio Ect2 and Vav3 mediate the invasivebehavior of glioblastomardquo American Journal of Pathology vol173 no 6 pp 1828ndash1838 2008
[11] B Liu L Wang L-L Shen et al ldquoRNAi-mediated inhibitionof presenilin 2 inhibits glioma cell growth and invasion andis involved in the regulation of Nrg1ErbB signalingrdquo Neuro-Oncology vol 14 no 8 pp 994ndash1006 2012
[12] S D Weeraratne V Amani N Teider et al ldquoPleiotropiceffects of miR-183sim96sim182 converge to regulate cell survivalproliferation and migration in medulloblastomardquo Acta Neu-ropathologica vol 123 no 4 pp 539ndash552 2012
[13] M Patel Q Cai D Ding R Salvi Z Hu and B H Hu ldquoThemiR-183Taok1 target pair is implicated in cochlear responsesto acoustic traumardquo PLoS ONE vol 8 no 3 Article ID e584712013
[14] J Zhu Y Feng Z Ke et al ldquoDown-regulation of miR-183promotes migration and invasion of osteosarcoma by targetingEzrinrdquo The American Journal of Pathology vol 180 no 6 pp2440ndash2451 2012
[15] A J Lowery N Miller R M Dwyer and M J Kerin ldquoDysreg-ulated miR-183 inhibits migration in breast cancer cellsrdquo BMCCancer vol 10 article 502 2010
[16] A L Sarver H Li and S Subramanian ldquoMicroRNA miR-183functions as an oncogene by targeting the transcription factorEGR1 and promoting tumor cell migrationrdquo Cancer Researchvol 70 no 23 pp 9570ndash9580 2010
[17] J Li H Fu C Xu et al ldquoMiR-183 inhibits TGF-beta1-inducedapoptosis by downregulation of PDCD4 expression in humanhepatocellular carcinoma cellsrdquoBMCCancer vol 10 article 3542010
[18] C Chen H Xiang Y-L Peng J Peng and S-W JiangldquoMature miR-183 negatively regulated by transcription factorGATA3 promotes 3T3-L1 adipogenesis through inhibition ofthe canonical Wnt120573-catenin signaling pathway by targetingLRP6rdquo Cellular Signalling vol 26 no 6 pp 1155ndash1165 2014
[19] G Li C Luna J Qiu D L Epstein and P Gonzalez ldquoTargetingof integrin beta1 and kinesin 2alpha by microRNA 183rdquo TheJournal of Biological Chemistry vol 285 no 8 pp 5461ndash54712010
[20] A M Schooley N M Andrews H Zhao and C L Addisonldquo1205731 integrin is required for anchorage-independent growth andinvasion of tumor cells in a context dependent mannerrdquo CancerLetters vol 316 no 2 pp 157ndash167 2012
[21] Y Zhang YGongDHu et al ldquoNuclear SIPA1 activates integrin1205731 promoter and promotes invasion of breast cancer cellsrdquoOncogene vol 34 no 11 pp 1451ndash1462 2015
[22] Y B Aznaurova M B Zhumataev T K Roberts A M Aliperand A A Zhavoronkov ldquoMolecular aspects of developmentand regulation of endometriosisrdquo Reproductive Biology andEndocrinology vol 12 article 50 2014
[23] M Adachi K Nasu A Tsuno A Yuge Y Kawano and HNarahara ldquoAttachment to extracellular matrices is enhancedin human endometriotic stromal cells a possible mechanismunderlying the pathogenesis of endometriosisrdquo European Jour-nal of Obstetrics Gynecology and Reproductive Biology vol 155no 1 pp 85ndash88 2011
[24] J Lee S K Banu R C Burghardt A Starzinski-Powitz andJ A Arosh ldquoSelective inhibition of prostaglandin E2 receptorsEP2 and EP4 inhibits adhesion of human endometriotic epithe-lial and stromal cells through suppression of integrin-mediatedmechanismsrdquo Biology of Reproduction vol 88 no 3 article 772013
[25] A Laeremans J NysW Luyten R DrsquoHoogeM Paulussen andL Arckens ldquoAMIGO2 mRNA expression in hippocampal CA2
10 BioMed Research International
and CA3ardquo Brain Structure and Function vol 218 no 1 pp 123ndash130 2013
[26] T Ono N Sekino-Suzuki Y Kikkawa H Yonekawa andS Kawashima ldquoAlivin 1 a novel neuronal activity-dependentgene inhibits apoptosis and promotes survival of cerebellargranule neuronsrdquo Journal of Neuroscience vol 23 no 13 pp5887ndash5896 2003
[27] J Kuja-Panula M Kiiltomaki T Yamashiro A Rouhiainenand H Rauvala ldquoAMIGO a transmembrane protein implicatedin axon tract development defines a novel protein family withleucine-rich repeatsrdquoThe Journal of Cell Biology vol 160 no 6pp 963ndash973 2003
[28] K E Rabenau J M OrsquoToole R Bassi et al ldquoDEGAAMIGO-2 a leucine-rich repeat family member differentially expressedin human gastric adenocarcinoma effects on ploidy chromo-somal stability cell adhesionmigration and tumorigenicityrdquoOncogene vol 23 no 29 pp 5056ndash5067 2004
[29] J A Garcia-Velasco and A Arici ldquoInterleukin-8 expression inendometrial stromal cells is regulated by integrin-dependentcell adhesionrdquo Molecular Human Reproduction vol 5 no 12pp 1135ndash1140 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 3
sodium dodecyl sulphate (SDS)] supplemented with a pro-tease inhibitor cocktail (Sigma) After incubating on ice for30min cell debris was removed by centrifugation at 12000 gfor 10min Cell lysates were resolved in 12 SDS-PAGEelectrophoresis and transferred to the PVDFmembrane (Bio-Rad Hercules CA) Nonspecific binding was blocked byincubating the membranes in Tris-buffered saline containing5 nonfat milk (TBST 50mmolL TrisndashHCl 150mmolLNaCl and 01 Tween-20) for 1 h at room temperatureMembranes were incubated overnight at 4∘C with AMIGO2antibody (1 400 RD) integrin 1205731 antibody (1 1000 Cell Sig-naling) Presenilin 2 antibody (1 500 ABCAM) and VAV3antibody (1 500 ABCAM) After three washes with TBSTthe membranes were incubated for 1 h at room temperaturewith the horseradish peroxidase-labeled secondary antibodyThe signals were visualized with chemiluminescence andquantified with Quantity-One (Bio-Rad America)
26 Luciferase Assay The mutant construct of ITGB131015840UTR was obtained by introducing the mutation intothe 7 nucleotides (GUGCCAUU) of the seed region formiR-183 The mutant construct of AMIGO2 31015840UTR wasobtained by introducing twomutations into the 7 nucleotides(GUGCCAUA) (UGCCAUA) of the seed region for miR-183 The miR-183 target sequences in the coding region ofITGB1AMIGO2 were amplified by PCR and cloned intoGV143 that contained a firefly luciferase reporter gene Wild-type ITBG1AMIGO2 31015840UTR or mutant ITBG1AMIGO231015840UTR and the empty 31015840UTR vector were cotransfected intoESC Cell transfection using Lipofectamine 2000 and nor-malization for transfection efficiency was performed accord-ing to the recommendation of manufacturer (Invitrogen)Luciferase activity was measured 24 h after transfection usingthe Dual-Luciferase Reporter Assay System (Promega CorpMadison WI)
27 Quantitative RT-PCR Endometrial tissue was mincedwith a scalpel blade Total RNA was isolated using TRIzol(Takara Otsu Shiga Japan) and cDNA was synthesizedusing the SYBR Prime Script RT-PCR kit (Takara) on theABI Prism 7500 Sequence Detection System according tomanufacturerrsquos instructions The housekeeping gene 120573-actinwas used for normalization The primers used were 51015840-TCG-TCACGTTCCGGTTATTC (sense) and 51015840-CTTTTACTT-ACGGTTTACCC (antisense) for ITGB1 51015840-CCTCGCCTT-TGCCGATCCG (sense) and 51015840-GCCGGAGCCGTTGTC-GACG (antisense) for 120573-actin The cycling conditions were94∘C for 1min 35 cycles of 94∘C for 30 sec 66∘C for 30 secand 68∘C for 3min followed by extension at 68∘C for3min Quantitative PCR was carried out using SYBR-GreenJumpStart Taq ReadyMix (Sigma) and the 7300 Real-TimePCRDetection System (ABI)The results were analyzed usingthe comparative threshold cycle (CT) method
28 Rescue Experiment To further validate direct targetingof ITGB1 by miR-183 functional rescue experiment was per-formed by cotransfection with miR-183 mimic and plasmidconstructs expressing ITGB1 in ESC using Lipofectamine
2000 (Invitrogen) as described aboveThe expression plasmidpcDNA-ITGB1 encoding human ITGB1was purchased fromGenechem The presence of complete ITGB1 coding regionswas confirmed by DNA sequencing
29 Invasion (Matrigel) Chamber Assay ESC (25 times 104) wereseeded on a transwell insert coated with extracellular matrix(ECM) (8mm pore size 24-well format Corning Costar) in2 FBS medium Complete medium (10 FBS) was addedto the lower chamber To determine the amount of invasioncells were incubated for 24 h and then removed from theupper chamber using a cotton swab The invaded cells onthe underside of the insert were fixed with methanol stainedwith crystal violet for 2min and rinsed with phosphatebuffered saline (PBS) The undersides of the membrane werephotographed to compare the number of invaded cells perinsert The transmigrated cells were counted using a lightmicroscope Invasive cells were scored by counting 10 randomhigh-power fields per filter
210 Statistical Analyses Data represents mean plusmn SEM fromat least 3 independent experiments Difference betweenexperimental and control groupswas determined by Studentrsquos119905-test while one-way ANOVA analysis was employed tocompare three or more groups 119875 lt 005 was considered asstatistical significance
3 Results
31 Gene Expression Profiling following miR-183 Overexpres-sion In order to screen target genes in response to miR-183we used microarrays representing more than 20000 genesmapped through RefSeq or via UniGene annotation Westudied gene expression alterations (up- or downregulation)at 24 h after transfection The changes of gene expressionin miR-183-overexpressing endometrial stromal cells wereanalyzed Differential expression was found in 27 genes at119875 value lt 005 with folds of change ge15 Of these 19 wereupregulated and 8 downregulated (ITGB1 AMIGO2 VAV3PSEN2 LHFPL2 HS2ST1 AHSA2 and UQCRB) Resultsof hierarchical cluster analyses of these genes are shownin Figure 1 and supplementary 1 in Supplementary Materialavailable online at httpdxdoiorg1011552015340218
32 Functional Analysis with GO Databases By using theGene Ontology (GO) database we systematically extractedand analyzed the information of threeGOcategories ldquobiolog-ical processrdquo ldquomolecular functionrdquo and ldquocellular componentrdquoIt was revealed that the identified genes were involvedin hemophilic cell adhesion (ITGB1 AMIGO2) cell-celladhesion (ITGB1 AMIGO2) cell migration (ITGB1MYH9)positive regulation of catalytic activity (PSEN2 SHC1) andproteolysis (MYH9 PSEN2) (Table 1)
33 Significant Pathway Analysis Significant pathway analy-sis revealed that the gene expression alterations in endome-trial stromal cells were involved in pathways of PTEN(ITGB1 SHC1) TFF (ITGB1 SHC1) ECM (ITGB1 SHC1)
4 BioMed Research International
Table 1 List of genes with fold of change ge15 (119875 lt 005) and their biological functions
Gene Gene set name 119875
ITGB1 AMIGO2 Hemophilic cell adhesion 197119864 minus 03
ITGB1 AMIGO2 Cell to cell adhesion 502119864 minus 02
ITGB1 MYH9 Cell migration 610119864 minus 02
PSEN2 SHC1 Positive regulation of catalytic activity 152119864 minus 01
MYH9 PSEN2 Proteolysis 191119864 minus 01
UQCRBAMIGO2HS2ST1AHSA2VAV3LHFPL2ITGB1PSEN2PICALMHES4SHC1HYOU1PPIFHPCAL1TUBBCBSMZT1CDT1GNB2PPP2R5DPTPRHCHMP4BVPS35PTPRAMYH9C6orf150HDGF
minus1 minus05 0 05 1 15Row Z-score
X461_1 X461_3 X461_2 X462_3 X462_1 X462_2
Figure 1 Hierarchical clustering of differentially expressed genes in miR-183-overexpressing endometrial stromal cells versus control cellsGene expression profiling was conducted using PrimeViewHumanGene ExpressionArray Raw data were extracted from the scanned imagesand analyzed with GeneSpring GX software version 115 The data was normalized using the iterative PLIER default protocol Differentiallyexpressed genes were analyzed with the unpaired Student 119905-test Statistically different genes (119875 lt 005) with a greater than 15-fold increaseor decrease were recorded Red color indicates low expression green color indicates relatively high expression
ERK (ITGB1 SHC1) integrin (ITGB1 SHC1) pathogenicEscherichia coli infection (ITGB1 TUBB) chemokine sig-naling pathway (SHC1 VAV3 and GNB2) focal adhe-sion (ITGB1 SHC1 and VAV3) regulation of cytoskele-ton (ITGB1 VAV3 and MYH9) leukocyte transendothelialmigration (ITGB1 VAV3) natural killer cell-mediated cyto-toxicity (SHC1 VAV3) and Alzheimerrsquos disease (UQCRBPSEN2) (Table 2)
34 Confirmation of Microarray Data by Western BlottingBecause of the inhibitory property of miRNA on target geneswe chose from the list of 8 downregulated genes (ITGB1AMIGO2 VAV3 PSEN2 LHFPL2 HS2ST1 AHSA2 andUQCRB) in miR-183-overexpressing cells Biological func-tion analysis using GO databases revealed that ITGB1 and
AMIGO2 were involved in cell adhesion andor cell migra-tionThese two geneswere selected for further study PubMedreports showed VAV3 and PSEN2 were both involved in cellinvasion [10 11] and these two genes were included as studytargets as well
Western blotting (Figure 2) was used to confirm theiralterations on protein levels Using the initial sample sets andthe criteria of ge15-fold of change integrin 1205731 yielded consis-tent results by the two technologies Compared to the controlgroup the levels of integrin 1205731 showed a significant decreasein the miR-183-overexpression group whereas the miR-183-downexpressing group (119875 lt 005) showed a sharp increaseAlthough endometrial stromal cells express AMIGO2 pro-tein the amount was largely unchanged in the miR-183 up-or downexpression groups suggesting it was not regulatedby miR-183 On the other hand no protein expression of
BioMed Research International 5
Table 2 List of genes with fold of change ge15 (119875 lt 005) and the pathways involved
Gene Gene set name Pathways 119875
ITGB1 SHC1 BIOCARTA PTEN PATHWAY PTEN dependent cell cycle arrest andapoptosis 308119864 minus 03
ITGB1 SHC1 BIOCARTA TFF PATHWAY Trefoil factors initiate mucosal healing 419119864 minus 03
ITGB1 SHC1 BIOCARTA ECM PATHWAY Erk and PI-3 kinase are necessary forcollagen binding in corneal epithelia 546119864 minus 03
ITGB1 SHC1 BIOCARTA ERK PATHWAY Erk1Erk2 Mapk signaling pathway 739119864 minus 03
ITGB1 SHC1 BIOCARTA INTEGRIN PATHWAY Integrin signaling pathway 134119864 minus 02
ITGB1 TUBB KEGG PATHOGENIC ESCHERICHIA COLI INFECFECTION Pathogenic Escherichiacoli infection 306119864 minus 02
SHC1 VAV3GNB2 KEGG CHEMOKINE SIGNALING PATHWAY Chemokine signaling pathway 575119864 minus 02
ITGB1 SHC1VAV3 KEGG FOCAL ADHESION Focal adhesion 659119864 minus 02
ITGB1 VAV3MYH9 KEGG REGULATION OF ACTIN CYTOSKELETON Regulation of actin cytoskeleton 782119864 minus 02
ITGB1 VAV3 KEGG LEUKOCYTE TRANSENDOTHELIAL MIGRATRATION Leukocyte transendothelial migration 105119864 minus 01
002040608
11214
U937
CON
NC-
low
dos
e
MiR
-dow
n-lo
w d
ose
MiR
-up-
low
dos
e
NC-
high
dos
e
MiR
-dow
n-hi
gh d
ose
MiR
-up-
high
dos
e
lowastlowast
lowastlowast
Integrin 1205731
GAPDH
1 2 3 4 5 6 7 8
Rela
tive i
nteg
rin1205731
prot
ein
leve
ls
(a)
0
02
04
06
08
1
12
Relat
ive A
MIG
O2
U937
CON
NC-
low
dos
e
MiR
-dow
n-lo
w d
ose
MiR
-up-
low
dos
e
NC-
high
dos
e
MiR
-dow
n-hi
gh d
ose
MiR
-up-
high
dos
e
Integrin 1205731
GAPDH
1 2 3 4 5 6 7 8pr
otei
n le
vels
(b)
Figure 2 Confirmation of microarray data withWestern blotting analysesThe endometrial stromal cells transfected withmiR-183-lentivirus(low dose or high dose) In-miR-183-lentivirus (low dose or high dose) and GFP-lentivirus (NC normal control) were used to examine theexpression alterations of integrin 1205731 (a) and AMIGO2 (b) 120573-actin served as protein loading control Compared with the control groupintegrin 1205731 expression in the miR-183-overexpression group was significantly decreased whereas it increased sharply in cells with low miR-183 expression AMIGO2 was largely not regulated by miR-183 lowast119875 lt 005 when compared to the negative control Error bars represent plusmnSEM
Presenilin 2 and VAV3 was detected in endometrial stromalcells (data not shown) and these two genes were not furtherstudied
35 MiR-183 Directly Targets ITBG1 but Not AMIGO2 Toinvestigate if ITBG1 and AMIGO2 are the direct targets of
miR-183 we performed luciferase assay to determinewhethermiR-183 binds to the 31015840UTR of ITBG1AMIGO2 mRNA(Figure 3(a)) Our results showed that reporter plasmidcontaining the miR-183 targeting sequence of ITGB1 mRNAdisplayed a significantly decreased luciferase activity in cellstransfected with miR-183 (119875 lt 005) whereas luciferase
6 BioMed Research International
3998400UTR5998400UTRAMIGO2
Has-miR-183
Mutant binding site
Coding region
5998400-UUCAGUUCCUCAAUGUGCCAUA-3998400
3998400-UCACUUAAGAUGGUCACGGUAU-5998400
5998400-UUCAGUUCCUCAAUCAUAACGC-3998400
174ndash181 bp
3998400UTR5998400UTRAMIGO2
Mutant binding site
Coding region
5998400-UCAUCAUAAUUUGUUGCCAUAA-3998400
3998400-CACUUAAGAUGGUCACGGUAU-5998400
5998400-UCAUCAUAAUUUGUAUAACGC-3998400
691ndash697 bp
3998400UTR5998400UTR
ITGB1
Has-miR-183
Mutant binding site
Coding region
5998400-GCUUUAAAACCUGUGUGCCAUU-3998400
3998400-UCACUUAAGAUGGUCACGGUAU-5998400
5998400-GCUUUAAAACCUGUUGUGACGU-3998400
695ndash701 bp
ITGB1 3998400UTR
998400Mutant 3 UTR
AMIGO2 3998400UTR
Mutant 3998400UTR
Has-miR-183
AMIGO2 3998400UTR
Mutant 3998400UTR
(a)
0
02
04
06
08
1
12
14
16
ITGB1-NC Wild-type Mutant type
Relat
ive l
ucife
rase
activ
ity (I
TGB1
)
MiR-183-NCMiR-183
0
02
04
06
08
1
12
14
16
AMIGO2-NC Wild-type Mutant type
Relat
ive L
ucife
rase
activ
ity (A
MIG
O2)
MiR-183-NCMiR-183
3998400UTR 3998400UTR 3998400UTR 3998400UTR
lowast
(b)
Figure 3 ITGB1 but not AMIGO2 is the direct target of miR-183 (a)The binding site of miR-183 in the 31015840UTR of ITGB1 (top) and AMIGO2(bottom) 31015840UTR along with the mutant construct in the predicted binding region (b) MiR-183 but not AMIGO2 directly targets ITBG1Endometrial stromal cells were cotransfected with wild-type reporter containing the ITBG1AMIGO2 31015840UTR or mutant ITBG1AMIGO231015840UTR plus miR-183 or negative control using Invitrogen Lipofectamine 2000 reagents Luminescence was measured after 24 hours oftransfection Reporter plasmid expressing ITGB1 mRNA that contains sequences potentially targeted by miR-183 displayed a significantlydecreased luciferase activity in cells transfected with miR-183 (119875 lt 005) whereas luciferase activity of reporter plasmid containing mutantsequence of ITGB1 was not changed following forced expression of miR-183 lowast119875 lt 005 when compared to the negative control Error barsrepresent plusmn SEM
BioMed Research International 7
0
02
04
06
08
1
12
14
Normal EM Eutopic EM
ITG
B1 ex
pres
sion
(nor
mal
ized
)
lowast
Figure 4 Validation of ITGB1 expression in eutopic endometriumwith endometriosis (119899 = 19) versus normal group (119899 = 18) byqPCR The results were analyzed using the comparative thresholdcycle (CT) method Gene expression data is presented as the foldof change relative to the levels of reference gene lowast119875 lt 005 whencompared to the normal group Error bars represent plusmn SEM
activity of reporter plasmid containing mutant sequence ofITGB1 was not changed following forced expression of miR-183 (Figure 3(b)) We found no significant change whencompared to the luciferase activity between cells transfectedwith wild-type or mutated AMIGO2 31015840UTR (119875 gt 005Figure 3(b) Thus these data suggested that miR-183 nega-tively regulates the expression of ITBG1 by directly target-ing their 31015840UTR whereas miR-183 does not directly targetAMIGO2 31015840UTR
36 ITGB1 Is Overexpressed in Endometrium from Endomet-riosis Patients Real-time RT-PCR results indicated thatITGB was significantly increased in the eutopic endometrialtissues from patients with endometriosis (119899 = 19) comparedwith endometrium from normal women (119899 = 18) (119875 lt 005)(Figure 4)
37 Overexpression of ITGB1 Rescued the Repressive Effectsof miR-183 on Endometrial Stromal Cells To ascertain thatmiR-183 regulates the function of endometrial stromal cellsthrough its interaction with ITGB1 a rescue experiment wasperformed Overexpression of ITGB1 partially rescued therepressive effects of miR-183 leading to elevated invasionabilities in the cells (Figure 5) This data indicated that miR-183 targets ITGB1 which in turn led to negative regulation onthe invasive activity of endometrial stromal cells
4 Discussion
In the previous study we performed miRNA microarrayscreening and identified several differentially expressed miR-NAs in the normal eutopic and ectopic endometriumAmong these miRNA species miR-183 was found to be
downregulated in the ectopic and eutopic tissues Functionalanalysis indicated that miR-183 promoted endometrial stro-mal cell apoptosis and had a negative regulatory impact onthe invasive ability of cells although it had no effect on cellproliferation [8] These findings suggested that the down-regulation of miR-183 expression might be involved in thepathogenesis of endometriosis The molecular mechanismshowever remain to be characterized
MiR-183 is a member of a miRNA family (miR-183 miR-182 and miR-96) that clusters within a 2ndash4 kb region atchromosome 7q32These miRNAs are known to regulate celldifferentiation apoptosis motility adhesion and invasion[12] Moreover it has been reported that miR-183 is upregu-lated in colorectal cancer prostate cancer and hepatocellularcarcinomas and downregulated in ovarian cancer breastcancer stem cells and osteosarcomas pointing to varied andcell type-dependent function of miR-183 Several target genesof miR-183 including ITGB1 Taok1 Ezrin EGR1 PDCD4and LRP6 have been identified and validated [13ndash19]
To systematically investigate the potential mechanismby which miR-183 may contribute to the development ofendometriosis we performed microarray analysis to identifythe target genes with relevance to cell functions in miR-183-overexpressing endometrial stromal cells Eight genes werefound to be downregulated in endometrial stromal cells withmiR-183 overexpression Among them the GO databasespathway analyses and PubMed reports pointed to importantassociation of 4 genes (ITGB1 AMIGO2 VAV3 and PSEN2)with cell adhesion cell migration and cell invasivenessWestern blotting results showed that endometrial stromalcell secreted integrin 1205731 and AMIGO2 but not VAV3 andPresenilin 2 The protein expression of integrin 1205731 appearedto be a direct regulation target of miR-183 which has beenproved in human trabecular meshwork cells human diploidfibroblasts and hela cells [19] AMIGO2 was not directlyregulated by miR-183
According to Sampsonrsquos theory endometriosis is thoughtto be initiated via retrograde endometrium into peri-toneal cavity during menstruation However while this phe-nomenon occurs in approximately 75ndash90 of women farfewer suffer the disease suggesting that additional factorsmust contribute to the process Invasion to ectopic locations isthe key step for the initiation of endometriosis during whichadhesion molecules play an important role The integrinfamily includes glycoproteins that formdimeric structure andmediate cell attachment to the ECM components [20] It hasbeen reported that 1205731 integrin-depleted cells had reducedinvasive capabilities and 1205731 integrin signaling was involvedin cancer cell invasion [21] Integrin 1205731 is constitutivelyexpressed in endometrial stromal cells and endometrioticlesions [22] Increased integrin 1205731 protein expression inendometrial stromal cells was observed in endometrioticcompared to normal endometrial tissues [23] It was reportedthat downregulation of prostaglandin E2 receptors EP2 andEP4 led to inhibited adhesion of human endometrioticepithelial and stromal cells Importantly this inhibitioncould be mediated through suppression of integrin-mediatedmechanisms [24] This finding provided a strong evidence
8 BioMed Research International
(a)
0
05
1
15
2
25
NC MiR-183 MiR-183ITGB1
Inva
sive r
ate (
OO
D57
0)
lowast
(b)
Figure 5 Overexpression of ITGB1 rescued the repressive effects of miR-183 on endometrial stromal cells leading to elevated invasiveabilities in transwell assays Endometrial stromal cells were infected with miR-183-lentivirus miR-183ITGB1-lentivirus and GFP-lentivirusin upper wells After 24 hours the number of cells that invaded through Matrigel was counted in at least 10 fields per well (a) Representativephotographs show that miR-183 inhibited the invasiveness of endometrial stromal cells whereas ITGB1 partially rescued the repressive effectsof miR-183 (b) Cell counting results indicate that ITGB1 overexpression rescued the repressive effects of miR-183 lowast119875 lt 005 when comparedto the negative control Error bars represent plusmn SEM
for the significance of integrins in the adhesion function ofendometrial stromal cells
AMIGO2 (ALI1 DEGA) was first reported to be prefer-entially expressed in the central nervous system Sequenceanalyses revealed that the protein contains seven leucine-richrepeats one IgC2-like loop and a transmembrane domainand displays homology to Kek and Trk families [25ndash27] Inthe gastric adenocarcinoma cell line AMIGO2 was reportedto be involved in cell adhesion extracellular matrix andbasement membrane formation [28] AMIGO2 was alsoimplicated in oral carcinogenesis by preventing the arrest ofcell cycle progression at the G1 phase through inhibiting theexpression of cyclin-dependent kinase inhibitors Howeverlittle is known about the role of AMIGO2 in the developmentof endometriosis
To investigate whether miR-183 directly targets ITGB1and AMIGO2 dual luciferase reporter assays were carriedout We found that miR-183 negatively regulated the expres-sion of ITBG1 by directly targeting its 31015840UTR but mightnot directly regulate the expression of AMIGO2 We further
detected differential expression of ITGB1 in endometriumwith or without endometriosis by quantitative RT-PCR Thehigher expression of integrin 1205731 in endometrium tissues inendometriosis was confirmed which is consistent with theresult for higher integrin 1205731 expression in ectopic stromalcells than normal endometrial stromal cells Previous studieshave shown that integrin 1205731 mediates cell adherence toextracellular matrix [29] Indeed the rescue experimentindicated that overexpression of ITGB1 blocked the repressiveeffects of miR-183 on endometrial stromal cell invasivenesswhich provided a critical evidence supporting that the lowerexpression of miR-183 may promote ITGB1 expression andcontribute to the development of endometriosis
In conclusion altered miR-183 expression may causederegulation of its target gene ITBG1 affecting the celladhesiveness and invasiveness of endometrial stromal cellswhich in turn lead to pathogenesis of endometriosis Thesefindings revealed a novel pathologic role of miR-183 forendometriosis It would be of great interest to investigate ifthese alterations may serve as biomarkers for the prediction
BioMed Research International 9
andor treatment of endometriosisThe regulation ofmiR-183and the exact mechanism by which miR-183 regulates ITBG1expression remain to be characterized
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This studywas supported by grants from theNational NaturalScience Foundation of China (nos 81100405 and 81471438)the Key Program ofNanjingMedical Science and TechnologyDevelopment Foundation Nanjing Department of Health(no QRX11109)
References
[1] D R Nyholt S-K Low C A Anderson et al ldquoGenome-wide association meta-analysis identifies new endometriosisrisk locirdquo Nature Genetics vol 44 no 12 pp 1355ndash1359 2012
[2] W Filipowicz S N Bhattacharyya and N Sonenberg ldquoMecha-nisms of post-transcriptional regulation by microRNAs are theanswers in sightrdquoNature Reviews Genetics vol 9 no 2 pp 102ndash114 2008
[3] X Santamaria and H Taylor ldquoMicroRNA and gynecologicalreproductive diseasesrdquo Fertility and Sterility vol 101 no 6 pp1545ndash1551 2014
[4] Y Shimono M Zabala R W Cho et al ldquoDownregulation ofmiRNA-200c links breast cancer stem cells with normal stemcellsrdquo Cell vol 138 no 3 pp 592ndash603 2009
[5] M Adammek B Greve N Kassens et al ldquoMicroRNA miR-145 inhibits proliferation invasiveness and stem cell phenotypeof an in vitro endometriosis model by targeting multiplecytoskeletal elements and pluripotency factorsrdquo Fertility andSterility vol 99 no 5 pp 1346e5ndash1355e5 2013
[6] W Abe K Nasu C Nakada Y Kawano M Moriyama andH Narahara ldquoMiR-196b targets c-myc and Bcl-2 expressioninhibits proliferation and induces apoptosis in endometrioticstromal cellsrdquo Human Reproduction vol 28 no 3 pp 750ndash7612013
[7] W B Nothnick A Graham J Holbert et al ldquomiR-451deficiency is associated with altered endometrial fibrinogenalpha chain expression and reduced endometriotic implantestablishment in an experimental mouse modelrdquo PLoS ONEvol 9 no 6 Article ID e100336 2014
[8] X-Y Shi L Gu J Chen X-R Guo and Y-L Shi ldquoDownregu-lation of miR-183 inhibits apoptosis and enhances the invasivepotential of endometrial stromal cells in endometriosisrdquo Inter-national Journal of Molecular Medicine vol 33 no 1 pp 59ndash672014
[9] Y-L Shi X-Z Luo X-Y Zhu K-Q Hua Y Zhu and D-JLi ldquoEffects of combined 17120573-estradiol with TCDD on secretionof chemokine IL-8 and expression of its receptor CXCR1 inendometriotic focus-associated cells in co-culturerdquo HumanReproduction vol 21 no 4 pp 870ndash879 2006
[10] B Salhia N L Tran A Chan et al ldquoThe guanine nucleotideexchange factors Trio Ect2 and Vav3 mediate the invasivebehavior of glioblastomardquo American Journal of Pathology vol173 no 6 pp 1828ndash1838 2008
[11] B Liu L Wang L-L Shen et al ldquoRNAi-mediated inhibitionof presenilin 2 inhibits glioma cell growth and invasion andis involved in the regulation of Nrg1ErbB signalingrdquo Neuro-Oncology vol 14 no 8 pp 994ndash1006 2012
[12] S D Weeraratne V Amani N Teider et al ldquoPleiotropiceffects of miR-183sim96sim182 converge to regulate cell survivalproliferation and migration in medulloblastomardquo Acta Neu-ropathologica vol 123 no 4 pp 539ndash552 2012
[13] M Patel Q Cai D Ding R Salvi Z Hu and B H Hu ldquoThemiR-183Taok1 target pair is implicated in cochlear responsesto acoustic traumardquo PLoS ONE vol 8 no 3 Article ID e584712013
[14] J Zhu Y Feng Z Ke et al ldquoDown-regulation of miR-183promotes migration and invasion of osteosarcoma by targetingEzrinrdquo The American Journal of Pathology vol 180 no 6 pp2440ndash2451 2012
[15] A J Lowery N Miller R M Dwyer and M J Kerin ldquoDysreg-ulated miR-183 inhibits migration in breast cancer cellsrdquo BMCCancer vol 10 article 502 2010
[16] A L Sarver H Li and S Subramanian ldquoMicroRNA miR-183functions as an oncogene by targeting the transcription factorEGR1 and promoting tumor cell migrationrdquo Cancer Researchvol 70 no 23 pp 9570ndash9580 2010
[17] J Li H Fu C Xu et al ldquoMiR-183 inhibits TGF-beta1-inducedapoptosis by downregulation of PDCD4 expression in humanhepatocellular carcinoma cellsrdquoBMCCancer vol 10 article 3542010
[18] C Chen H Xiang Y-L Peng J Peng and S-W JiangldquoMature miR-183 negatively regulated by transcription factorGATA3 promotes 3T3-L1 adipogenesis through inhibition ofthe canonical Wnt120573-catenin signaling pathway by targetingLRP6rdquo Cellular Signalling vol 26 no 6 pp 1155ndash1165 2014
[19] G Li C Luna J Qiu D L Epstein and P Gonzalez ldquoTargetingof integrin beta1 and kinesin 2alpha by microRNA 183rdquo TheJournal of Biological Chemistry vol 285 no 8 pp 5461ndash54712010
[20] A M Schooley N M Andrews H Zhao and C L Addisonldquo1205731 integrin is required for anchorage-independent growth andinvasion of tumor cells in a context dependent mannerrdquo CancerLetters vol 316 no 2 pp 157ndash167 2012
[21] Y Zhang YGongDHu et al ldquoNuclear SIPA1 activates integrin1205731 promoter and promotes invasion of breast cancer cellsrdquoOncogene vol 34 no 11 pp 1451ndash1462 2015
[22] Y B Aznaurova M B Zhumataev T K Roberts A M Aliperand A A Zhavoronkov ldquoMolecular aspects of developmentand regulation of endometriosisrdquo Reproductive Biology andEndocrinology vol 12 article 50 2014
[23] M Adachi K Nasu A Tsuno A Yuge Y Kawano and HNarahara ldquoAttachment to extracellular matrices is enhancedin human endometriotic stromal cells a possible mechanismunderlying the pathogenesis of endometriosisrdquo European Jour-nal of Obstetrics Gynecology and Reproductive Biology vol 155no 1 pp 85ndash88 2011
[24] J Lee S K Banu R C Burghardt A Starzinski-Powitz andJ A Arosh ldquoSelective inhibition of prostaglandin E2 receptorsEP2 and EP4 inhibits adhesion of human endometriotic epithe-lial and stromal cells through suppression of integrin-mediatedmechanismsrdquo Biology of Reproduction vol 88 no 3 article 772013
[25] A Laeremans J NysW Luyten R DrsquoHoogeM Paulussen andL Arckens ldquoAMIGO2 mRNA expression in hippocampal CA2
10 BioMed Research International
and CA3ardquo Brain Structure and Function vol 218 no 1 pp 123ndash130 2013
[26] T Ono N Sekino-Suzuki Y Kikkawa H Yonekawa andS Kawashima ldquoAlivin 1 a novel neuronal activity-dependentgene inhibits apoptosis and promotes survival of cerebellargranule neuronsrdquo Journal of Neuroscience vol 23 no 13 pp5887ndash5896 2003
[27] J Kuja-Panula M Kiiltomaki T Yamashiro A Rouhiainenand H Rauvala ldquoAMIGO a transmembrane protein implicatedin axon tract development defines a novel protein family withleucine-rich repeatsrdquoThe Journal of Cell Biology vol 160 no 6pp 963ndash973 2003
[28] K E Rabenau J M OrsquoToole R Bassi et al ldquoDEGAAMIGO-2 a leucine-rich repeat family member differentially expressedin human gastric adenocarcinoma effects on ploidy chromo-somal stability cell adhesionmigration and tumorigenicityrdquoOncogene vol 23 no 29 pp 5056ndash5067 2004
[29] J A Garcia-Velasco and A Arici ldquoInterleukin-8 expression inendometrial stromal cells is regulated by integrin-dependentcell adhesionrdquo Molecular Human Reproduction vol 5 no 12pp 1135ndash1140 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
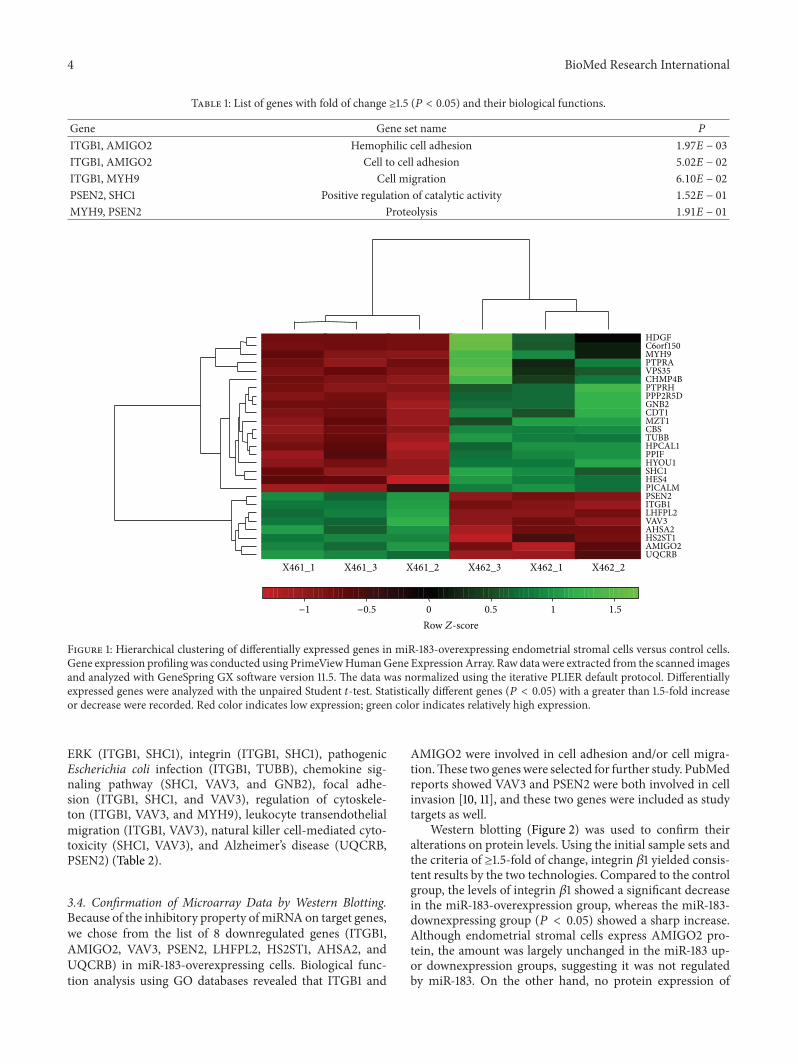
4 BioMed Research International
Table 1 List of genes with fold of change ge15 (119875 lt 005) and their biological functions
Gene Gene set name 119875
ITGB1 AMIGO2 Hemophilic cell adhesion 197119864 minus 03
ITGB1 AMIGO2 Cell to cell adhesion 502119864 minus 02
ITGB1 MYH9 Cell migration 610119864 minus 02
PSEN2 SHC1 Positive regulation of catalytic activity 152119864 minus 01
MYH9 PSEN2 Proteolysis 191119864 minus 01
UQCRBAMIGO2HS2ST1AHSA2VAV3LHFPL2ITGB1PSEN2PICALMHES4SHC1HYOU1PPIFHPCAL1TUBBCBSMZT1CDT1GNB2PPP2R5DPTPRHCHMP4BVPS35PTPRAMYH9C6orf150HDGF
minus1 minus05 0 05 1 15Row Z-score
X461_1 X461_3 X461_2 X462_3 X462_1 X462_2
Figure 1 Hierarchical clustering of differentially expressed genes in miR-183-overexpressing endometrial stromal cells versus control cellsGene expression profiling was conducted using PrimeViewHumanGene ExpressionArray Raw data were extracted from the scanned imagesand analyzed with GeneSpring GX software version 115 The data was normalized using the iterative PLIER default protocol Differentiallyexpressed genes were analyzed with the unpaired Student 119905-test Statistically different genes (119875 lt 005) with a greater than 15-fold increaseor decrease were recorded Red color indicates low expression green color indicates relatively high expression
ERK (ITGB1 SHC1) integrin (ITGB1 SHC1) pathogenicEscherichia coli infection (ITGB1 TUBB) chemokine sig-naling pathway (SHC1 VAV3 and GNB2) focal adhe-sion (ITGB1 SHC1 and VAV3) regulation of cytoskele-ton (ITGB1 VAV3 and MYH9) leukocyte transendothelialmigration (ITGB1 VAV3) natural killer cell-mediated cyto-toxicity (SHC1 VAV3) and Alzheimerrsquos disease (UQCRBPSEN2) (Table 2)
34 Confirmation of Microarray Data by Western BlottingBecause of the inhibitory property of miRNA on target geneswe chose from the list of 8 downregulated genes (ITGB1AMIGO2 VAV3 PSEN2 LHFPL2 HS2ST1 AHSA2 andUQCRB) in miR-183-overexpressing cells Biological func-tion analysis using GO databases revealed that ITGB1 and
AMIGO2 were involved in cell adhesion andor cell migra-tionThese two geneswere selected for further study PubMedreports showed VAV3 and PSEN2 were both involved in cellinvasion [10 11] and these two genes were included as studytargets as well
Western blotting (Figure 2) was used to confirm theiralterations on protein levels Using the initial sample sets andthe criteria of ge15-fold of change integrin 1205731 yielded consis-tent results by the two technologies Compared to the controlgroup the levels of integrin 1205731 showed a significant decreasein the miR-183-overexpression group whereas the miR-183-downexpressing group (119875 lt 005) showed a sharp increaseAlthough endometrial stromal cells express AMIGO2 pro-tein the amount was largely unchanged in the miR-183 up-or downexpression groups suggesting it was not regulatedby miR-183 On the other hand no protein expression of
BioMed Research International 5
Table 2 List of genes with fold of change ge15 (119875 lt 005) and the pathways involved
Gene Gene set name Pathways 119875
ITGB1 SHC1 BIOCARTA PTEN PATHWAY PTEN dependent cell cycle arrest andapoptosis 308119864 minus 03
ITGB1 SHC1 BIOCARTA TFF PATHWAY Trefoil factors initiate mucosal healing 419119864 minus 03
ITGB1 SHC1 BIOCARTA ECM PATHWAY Erk and PI-3 kinase are necessary forcollagen binding in corneal epithelia 546119864 minus 03
ITGB1 SHC1 BIOCARTA ERK PATHWAY Erk1Erk2 Mapk signaling pathway 739119864 minus 03
ITGB1 SHC1 BIOCARTA INTEGRIN PATHWAY Integrin signaling pathway 134119864 minus 02
ITGB1 TUBB KEGG PATHOGENIC ESCHERICHIA COLI INFECFECTION Pathogenic Escherichiacoli infection 306119864 minus 02
SHC1 VAV3GNB2 KEGG CHEMOKINE SIGNALING PATHWAY Chemokine signaling pathway 575119864 minus 02
ITGB1 SHC1VAV3 KEGG FOCAL ADHESION Focal adhesion 659119864 minus 02
ITGB1 VAV3MYH9 KEGG REGULATION OF ACTIN CYTOSKELETON Regulation of actin cytoskeleton 782119864 minus 02
ITGB1 VAV3 KEGG LEUKOCYTE TRANSENDOTHELIAL MIGRATRATION Leukocyte transendothelial migration 105119864 minus 01
002040608
11214
U937
CON
NC-
low
dos
e
MiR
-dow
n-lo
w d
ose
MiR
-up-
low
dos
e
NC-
high
dos
e
MiR
-dow
n-hi
gh d
ose
MiR
-up-
high
dos
e
lowastlowast
lowastlowast
Integrin 1205731
GAPDH
1 2 3 4 5 6 7 8
Rela
tive i
nteg
rin1205731
prot
ein
leve
ls
(a)
0
02
04
06
08
1
12
Relat
ive A
MIG
O2
U937
CON
NC-
low
dos
e
MiR
-dow
n-lo
w d
ose
MiR
-up-
low
dos
e
NC-
high
dos
e
MiR
-dow
n-hi
gh d
ose
MiR
-up-
high
dos
e
Integrin 1205731
GAPDH
1 2 3 4 5 6 7 8pr
otei
n le
vels
(b)
Figure 2 Confirmation of microarray data withWestern blotting analysesThe endometrial stromal cells transfected withmiR-183-lentivirus(low dose or high dose) In-miR-183-lentivirus (low dose or high dose) and GFP-lentivirus (NC normal control) were used to examine theexpression alterations of integrin 1205731 (a) and AMIGO2 (b) 120573-actin served as protein loading control Compared with the control groupintegrin 1205731 expression in the miR-183-overexpression group was significantly decreased whereas it increased sharply in cells with low miR-183 expression AMIGO2 was largely not regulated by miR-183 lowast119875 lt 005 when compared to the negative control Error bars represent plusmnSEM
Presenilin 2 and VAV3 was detected in endometrial stromalcells (data not shown) and these two genes were not furtherstudied
35 MiR-183 Directly Targets ITBG1 but Not AMIGO2 Toinvestigate if ITBG1 and AMIGO2 are the direct targets of
miR-183 we performed luciferase assay to determinewhethermiR-183 binds to the 31015840UTR of ITBG1AMIGO2 mRNA(Figure 3(a)) Our results showed that reporter plasmidcontaining the miR-183 targeting sequence of ITGB1 mRNAdisplayed a significantly decreased luciferase activity in cellstransfected with miR-183 (119875 lt 005) whereas luciferase
6 BioMed Research International
3998400UTR5998400UTRAMIGO2
Has-miR-183
Mutant binding site
Coding region
5998400-UUCAGUUCCUCAAUGUGCCAUA-3998400
3998400-UCACUUAAGAUGGUCACGGUAU-5998400
5998400-UUCAGUUCCUCAAUCAUAACGC-3998400
174ndash181 bp
3998400UTR5998400UTRAMIGO2
Mutant binding site
Coding region
5998400-UCAUCAUAAUUUGUUGCCAUAA-3998400
3998400-CACUUAAGAUGGUCACGGUAU-5998400
5998400-UCAUCAUAAUUUGUAUAACGC-3998400
691ndash697 bp
3998400UTR5998400UTR
ITGB1
Has-miR-183
Mutant binding site
Coding region
5998400-GCUUUAAAACCUGUGUGCCAUU-3998400
3998400-UCACUUAAGAUGGUCACGGUAU-5998400
5998400-GCUUUAAAACCUGUUGUGACGU-3998400
695ndash701 bp
ITGB1 3998400UTR
998400Mutant 3 UTR
AMIGO2 3998400UTR
Mutant 3998400UTR
Has-miR-183
AMIGO2 3998400UTR
Mutant 3998400UTR
(a)
0
02
04
06
08
1
12
14
16
ITGB1-NC Wild-type Mutant type
Relat
ive l
ucife
rase
activ
ity (I
TGB1
)
MiR-183-NCMiR-183
0
02
04
06
08
1
12
14
16
AMIGO2-NC Wild-type Mutant type
Relat
ive L
ucife
rase
activ
ity (A
MIG
O2)
MiR-183-NCMiR-183
3998400UTR 3998400UTR 3998400UTR 3998400UTR
lowast
(b)
Figure 3 ITGB1 but not AMIGO2 is the direct target of miR-183 (a)The binding site of miR-183 in the 31015840UTR of ITGB1 (top) and AMIGO2(bottom) 31015840UTR along with the mutant construct in the predicted binding region (b) MiR-183 but not AMIGO2 directly targets ITBG1Endometrial stromal cells were cotransfected with wild-type reporter containing the ITBG1AMIGO2 31015840UTR or mutant ITBG1AMIGO231015840UTR plus miR-183 or negative control using Invitrogen Lipofectamine 2000 reagents Luminescence was measured after 24 hours oftransfection Reporter plasmid expressing ITGB1 mRNA that contains sequences potentially targeted by miR-183 displayed a significantlydecreased luciferase activity in cells transfected with miR-183 (119875 lt 005) whereas luciferase activity of reporter plasmid containing mutantsequence of ITGB1 was not changed following forced expression of miR-183 lowast119875 lt 005 when compared to the negative control Error barsrepresent plusmn SEM
BioMed Research International 7
0
02
04
06
08
1
12
14
Normal EM Eutopic EM
ITG
B1 ex
pres
sion
(nor
mal
ized
)
lowast
Figure 4 Validation of ITGB1 expression in eutopic endometriumwith endometriosis (119899 = 19) versus normal group (119899 = 18) byqPCR The results were analyzed using the comparative thresholdcycle (CT) method Gene expression data is presented as the foldof change relative to the levels of reference gene lowast119875 lt 005 whencompared to the normal group Error bars represent plusmn SEM
activity of reporter plasmid containing mutant sequence ofITGB1 was not changed following forced expression of miR-183 (Figure 3(b)) We found no significant change whencompared to the luciferase activity between cells transfectedwith wild-type or mutated AMIGO2 31015840UTR (119875 gt 005Figure 3(b) Thus these data suggested that miR-183 nega-tively regulates the expression of ITBG1 by directly target-ing their 31015840UTR whereas miR-183 does not directly targetAMIGO2 31015840UTR
36 ITGB1 Is Overexpressed in Endometrium from Endomet-riosis Patients Real-time RT-PCR results indicated thatITGB was significantly increased in the eutopic endometrialtissues from patients with endometriosis (119899 = 19) comparedwith endometrium from normal women (119899 = 18) (119875 lt 005)(Figure 4)
37 Overexpression of ITGB1 Rescued the Repressive Effectsof miR-183 on Endometrial Stromal Cells To ascertain thatmiR-183 regulates the function of endometrial stromal cellsthrough its interaction with ITGB1 a rescue experiment wasperformed Overexpression of ITGB1 partially rescued therepressive effects of miR-183 leading to elevated invasionabilities in the cells (Figure 5) This data indicated that miR-183 targets ITGB1 which in turn led to negative regulation onthe invasive activity of endometrial stromal cells
4 Discussion
In the previous study we performed miRNA microarrayscreening and identified several differentially expressed miR-NAs in the normal eutopic and ectopic endometriumAmong these miRNA species miR-183 was found to be
downregulated in the ectopic and eutopic tissues Functionalanalysis indicated that miR-183 promoted endometrial stro-mal cell apoptosis and had a negative regulatory impact onthe invasive ability of cells although it had no effect on cellproliferation [8] These findings suggested that the down-regulation of miR-183 expression might be involved in thepathogenesis of endometriosis The molecular mechanismshowever remain to be characterized
MiR-183 is a member of a miRNA family (miR-183 miR-182 and miR-96) that clusters within a 2ndash4 kb region atchromosome 7q32These miRNAs are known to regulate celldifferentiation apoptosis motility adhesion and invasion[12] Moreover it has been reported that miR-183 is upregu-lated in colorectal cancer prostate cancer and hepatocellularcarcinomas and downregulated in ovarian cancer breastcancer stem cells and osteosarcomas pointing to varied andcell type-dependent function of miR-183 Several target genesof miR-183 including ITGB1 Taok1 Ezrin EGR1 PDCD4and LRP6 have been identified and validated [13ndash19]
To systematically investigate the potential mechanismby which miR-183 may contribute to the development ofendometriosis we performed microarray analysis to identifythe target genes with relevance to cell functions in miR-183-overexpressing endometrial stromal cells Eight genes werefound to be downregulated in endometrial stromal cells withmiR-183 overexpression Among them the GO databasespathway analyses and PubMed reports pointed to importantassociation of 4 genes (ITGB1 AMIGO2 VAV3 and PSEN2)with cell adhesion cell migration and cell invasivenessWestern blotting results showed that endometrial stromalcell secreted integrin 1205731 and AMIGO2 but not VAV3 andPresenilin 2 The protein expression of integrin 1205731 appearedto be a direct regulation target of miR-183 which has beenproved in human trabecular meshwork cells human diploidfibroblasts and hela cells [19] AMIGO2 was not directlyregulated by miR-183
According to Sampsonrsquos theory endometriosis is thoughtto be initiated via retrograde endometrium into peri-toneal cavity during menstruation However while this phe-nomenon occurs in approximately 75ndash90 of women farfewer suffer the disease suggesting that additional factorsmust contribute to the process Invasion to ectopic locations isthe key step for the initiation of endometriosis during whichadhesion molecules play an important role The integrinfamily includes glycoproteins that formdimeric structure andmediate cell attachment to the ECM components [20] It hasbeen reported that 1205731 integrin-depleted cells had reducedinvasive capabilities and 1205731 integrin signaling was involvedin cancer cell invasion [21] Integrin 1205731 is constitutivelyexpressed in endometrial stromal cells and endometrioticlesions [22] Increased integrin 1205731 protein expression inendometrial stromal cells was observed in endometrioticcompared to normal endometrial tissues [23] It was reportedthat downregulation of prostaglandin E2 receptors EP2 andEP4 led to inhibited adhesion of human endometrioticepithelial and stromal cells Importantly this inhibitioncould be mediated through suppression of integrin-mediatedmechanisms [24] This finding provided a strong evidence
8 BioMed Research International
(a)
0
05
1
15
2
25
NC MiR-183 MiR-183ITGB1
Inva
sive r
ate (
OO
D57
0)
lowast
(b)
Figure 5 Overexpression of ITGB1 rescued the repressive effects of miR-183 on endometrial stromal cells leading to elevated invasiveabilities in transwell assays Endometrial stromal cells were infected with miR-183-lentivirus miR-183ITGB1-lentivirus and GFP-lentivirusin upper wells After 24 hours the number of cells that invaded through Matrigel was counted in at least 10 fields per well (a) Representativephotographs show that miR-183 inhibited the invasiveness of endometrial stromal cells whereas ITGB1 partially rescued the repressive effectsof miR-183 (b) Cell counting results indicate that ITGB1 overexpression rescued the repressive effects of miR-183 lowast119875 lt 005 when comparedto the negative control Error bars represent plusmn SEM
for the significance of integrins in the adhesion function ofendometrial stromal cells
AMIGO2 (ALI1 DEGA) was first reported to be prefer-entially expressed in the central nervous system Sequenceanalyses revealed that the protein contains seven leucine-richrepeats one IgC2-like loop and a transmembrane domainand displays homology to Kek and Trk families [25ndash27] Inthe gastric adenocarcinoma cell line AMIGO2 was reportedto be involved in cell adhesion extracellular matrix andbasement membrane formation [28] AMIGO2 was alsoimplicated in oral carcinogenesis by preventing the arrest ofcell cycle progression at the G1 phase through inhibiting theexpression of cyclin-dependent kinase inhibitors Howeverlittle is known about the role of AMIGO2 in the developmentof endometriosis
To investigate whether miR-183 directly targets ITGB1and AMIGO2 dual luciferase reporter assays were carriedout We found that miR-183 negatively regulated the expres-sion of ITBG1 by directly targeting its 31015840UTR but mightnot directly regulate the expression of AMIGO2 We further
detected differential expression of ITGB1 in endometriumwith or without endometriosis by quantitative RT-PCR Thehigher expression of integrin 1205731 in endometrium tissues inendometriosis was confirmed which is consistent with theresult for higher integrin 1205731 expression in ectopic stromalcells than normal endometrial stromal cells Previous studieshave shown that integrin 1205731 mediates cell adherence toextracellular matrix [29] Indeed the rescue experimentindicated that overexpression of ITGB1 blocked the repressiveeffects of miR-183 on endometrial stromal cell invasivenesswhich provided a critical evidence supporting that the lowerexpression of miR-183 may promote ITGB1 expression andcontribute to the development of endometriosis
In conclusion altered miR-183 expression may causederegulation of its target gene ITBG1 affecting the celladhesiveness and invasiveness of endometrial stromal cellswhich in turn lead to pathogenesis of endometriosis Thesefindings revealed a novel pathologic role of miR-183 forendometriosis It would be of great interest to investigate ifthese alterations may serve as biomarkers for the prediction
BioMed Research International 9
andor treatment of endometriosisThe regulation ofmiR-183and the exact mechanism by which miR-183 regulates ITBG1expression remain to be characterized
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This studywas supported by grants from theNational NaturalScience Foundation of China (nos 81100405 and 81471438)the Key Program ofNanjingMedical Science and TechnologyDevelopment Foundation Nanjing Department of Health(no QRX11109)
References
[1] D R Nyholt S-K Low C A Anderson et al ldquoGenome-wide association meta-analysis identifies new endometriosisrisk locirdquo Nature Genetics vol 44 no 12 pp 1355ndash1359 2012
[2] W Filipowicz S N Bhattacharyya and N Sonenberg ldquoMecha-nisms of post-transcriptional regulation by microRNAs are theanswers in sightrdquoNature Reviews Genetics vol 9 no 2 pp 102ndash114 2008
[3] X Santamaria and H Taylor ldquoMicroRNA and gynecologicalreproductive diseasesrdquo Fertility and Sterility vol 101 no 6 pp1545ndash1551 2014
[4] Y Shimono M Zabala R W Cho et al ldquoDownregulation ofmiRNA-200c links breast cancer stem cells with normal stemcellsrdquo Cell vol 138 no 3 pp 592ndash603 2009
[5] M Adammek B Greve N Kassens et al ldquoMicroRNA miR-145 inhibits proliferation invasiveness and stem cell phenotypeof an in vitro endometriosis model by targeting multiplecytoskeletal elements and pluripotency factorsrdquo Fertility andSterility vol 99 no 5 pp 1346e5ndash1355e5 2013
[6] W Abe K Nasu C Nakada Y Kawano M Moriyama andH Narahara ldquoMiR-196b targets c-myc and Bcl-2 expressioninhibits proliferation and induces apoptosis in endometrioticstromal cellsrdquo Human Reproduction vol 28 no 3 pp 750ndash7612013
[7] W B Nothnick A Graham J Holbert et al ldquomiR-451deficiency is associated with altered endometrial fibrinogenalpha chain expression and reduced endometriotic implantestablishment in an experimental mouse modelrdquo PLoS ONEvol 9 no 6 Article ID e100336 2014
[8] X-Y Shi L Gu J Chen X-R Guo and Y-L Shi ldquoDownregu-lation of miR-183 inhibits apoptosis and enhances the invasivepotential of endometrial stromal cells in endometriosisrdquo Inter-national Journal of Molecular Medicine vol 33 no 1 pp 59ndash672014
[9] Y-L Shi X-Z Luo X-Y Zhu K-Q Hua Y Zhu and D-JLi ldquoEffects of combined 17120573-estradiol with TCDD on secretionof chemokine IL-8 and expression of its receptor CXCR1 inendometriotic focus-associated cells in co-culturerdquo HumanReproduction vol 21 no 4 pp 870ndash879 2006
[10] B Salhia N L Tran A Chan et al ldquoThe guanine nucleotideexchange factors Trio Ect2 and Vav3 mediate the invasivebehavior of glioblastomardquo American Journal of Pathology vol173 no 6 pp 1828ndash1838 2008
[11] B Liu L Wang L-L Shen et al ldquoRNAi-mediated inhibitionof presenilin 2 inhibits glioma cell growth and invasion andis involved in the regulation of Nrg1ErbB signalingrdquo Neuro-Oncology vol 14 no 8 pp 994ndash1006 2012
[12] S D Weeraratne V Amani N Teider et al ldquoPleiotropiceffects of miR-183sim96sim182 converge to regulate cell survivalproliferation and migration in medulloblastomardquo Acta Neu-ropathologica vol 123 no 4 pp 539ndash552 2012
[13] M Patel Q Cai D Ding R Salvi Z Hu and B H Hu ldquoThemiR-183Taok1 target pair is implicated in cochlear responsesto acoustic traumardquo PLoS ONE vol 8 no 3 Article ID e584712013
[14] J Zhu Y Feng Z Ke et al ldquoDown-regulation of miR-183promotes migration and invasion of osteosarcoma by targetingEzrinrdquo The American Journal of Pathology vol 180 no 6 pp2440ndash2451 2012
[15] A J Lowery N Miller R M Dwyer and M J Kerin ldquoDysreg-ulated miR-183 inhibits migration in breast cancer cellsrdquo BMCCancer vol 10 article 502 2010
[16] A L Sarver H Li and S Subramanian ldquoMicroRNA miR-183functions as an oncogene by targeting the transcription factorEGR1 and promoting tumor cell migrationrdquo Cancer Researchvol 70 no 23 pp 9570ndash9580 2010
[17] J Li H Fu C Xu et al ldquoMiR-183 inhibits TGF-beta1-inducedapoptosis by downregulation of PDCD4 expression in humanhepatocellular carcinoma cellsrdquoBMCCancer vol 10 article 3542010
[18] C Chen H Xiang Y-L Peng J Peng and S-W JiangldquoMature miR-183 negatively regulated by transcription factorGATA3 promotes 3T3-L1 adipogenesis through inhibition ofthe canonical Wnt120573-catenin signaling pathway by targetingLRP6rdquo Cellular Signalling vol 26 no 6 pp 1155ndash1165 2014
[19] G Li C Luna J Qiu D L Epstein and P Gonzalez ldquoTargetingof integrin beta1 and kinesin 2alpha by microRNA 183rdquo TheJournal of Biological Chemistry vol 285 no 8 pp 5461ndash54712010
[20] A M Schooley N M Andrews H Zhao and C L Addisonldquo1205731 integrin is required for anchorage-independent growth andinvasion of tumor cells in a context dependent mannerrdquo CancerLetters vol 316 no 2 pp 157ndash167 2012
[21] Y Zhang YGongDHu et al ldquoNuclear SIPA1 activates integrin1205731 promoter and promotes invasion of breast cancer cellsrdquoOncogene vol 34 no 11 pp 1451ndash1462 2015
[22] Y B Aznaurova M B Zhumataev T K Roberts A M Aliperand A A Zhavoronkov ldquoMolecular aspects of developmentand regulation of endometriosisrdquo Reproductive Biology andEndocrinology vol 12 article 50 2014
[23] M Adachi K Nasu A Tsuno A Yuge Y Kawano and HNarahara ldquoAttachment to extracellular matrices is enhancedin human endometriotic stromal cells a possible mechanismunderlying the pathogenesis of endometriosisrdquo European Jour-nal of Obstetrics Gynecology and Reproductive Biology vol 155no 1 pp 85ndash88 2011
[24] J Lee S K Banu R C Burghardt A Starzinski-Powitz andJ A Arosh ldquoSelective inhibition of prostaglandin E2 receptorsEP2 and EP4 inhibits adhesion of human endometriotic epithe-lial and stromal cells through suppression of integrin-mediatedmechanismsrdquo Biology of Reproduction vol 88 no 3 article 772013
[25] A Laeremans J NysW Luyten R DrsquoHoogeM Paulussen andL Arckens ldquoAMIGO2 mRNA expression in hippocampal CA2
10 BioMed Research International
and CA3ardquo Brain Structure and Function vol 218 no 1 pp 123ndash130 2013
[26] T Ono N Sekino-Suzuki Y Kikkawa H Yonekawa andS Kawashima ldquoAlivin 1 a novel neuronal activity-dependentgene inhibits apoptosis and promotes survival of cerebellargranule neuronsrdquo Journal of Neuroscience vol 23 no 13 pp5887ndash5896 2003
[27] J Kuja-Panula M Kiiltomaki T Yamashiro A Rouhiainenand H Rauvala ldquoAMIGO a transmembrane protein implicatedin axon tract development defines a novel protein family withleucine-rich repeatsrdquoThe Journal of Cell Biology vol 160 no 6pp 963ndash973 2003
[28] K E Rabenau J M OrsquoToole R Bassi et al ldquoDEGAAMIGO-2 a leucine-rich repeat family member differentially expressedin human gastric adenocarcinoma effects on ploidy chromo-somal stability cell adhesionmigration and tumorigenicityrdquoOncogene vol 23 no 29 pp 5056ndash5067 2004
[29] J A Garcia-Velasco and A Arici ldquoInterleukin-8 expression inendometrial stromal cells is regulated by integrin-dependentcell adhesionrdquo Molecular Human Reproduction vol 5 no 12pp 1135ndash1140 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
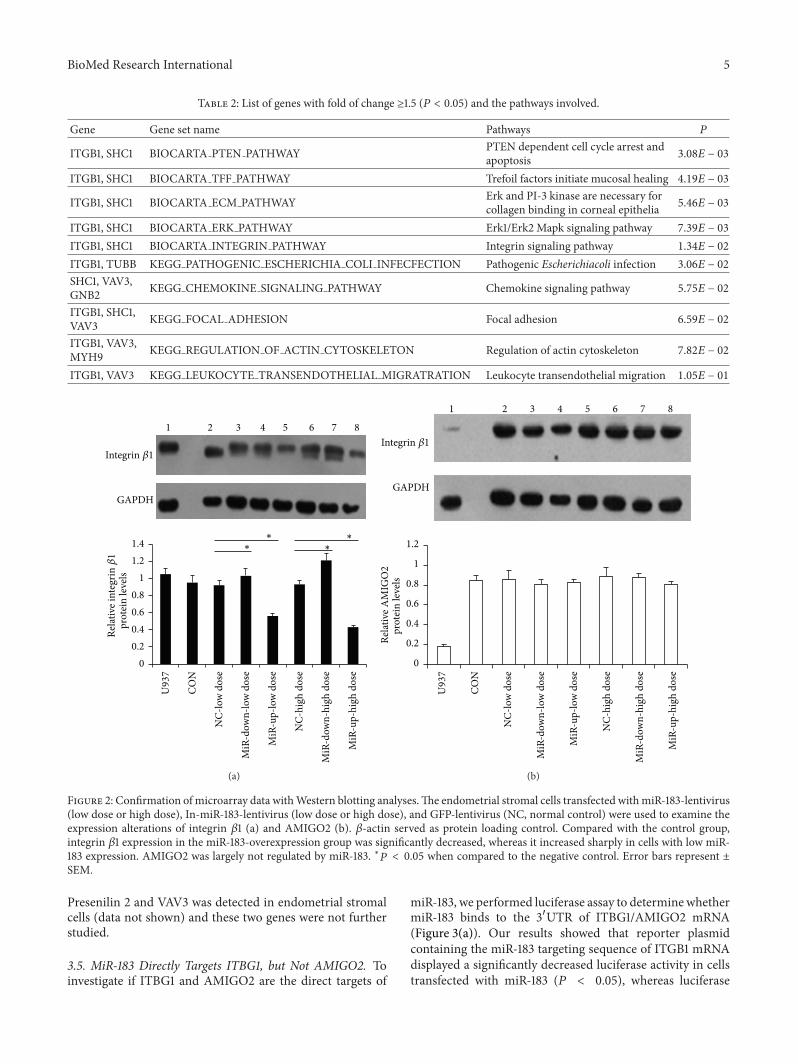
BioMed Research International 5
Table 2 List of genes with fold of change ge15 (119875 lt 005) and the pathways involved
Gene Gene set name Pathways 119875
ITGB1 SHC1 BIOCARTA PTEN PATHWAY PTEN dependent cell cycle arrest andapoptosis 308119864 minus 03
ITGB1 SHC1 BIOCARTA TFF PATHWAY Trefoil factors initiate mucosal healing 419119864 minus 03
ITGB1 SHC1 BIOCARTA ECM PATHWAY Erk and PI-3 kinase are necessary forcollagen binding in corneal epithelia 546119864 minus 03
ITGB1 SHC1 BIOCARTA ERK PATHWAY Erk1Erk2 Mapk signaling pathway 739119864 minus 03
ITGB1 SHC1 BIOCARTA INTEGRIN PATHWAY Integrin signaling pathway 134119864 minus 02
ITGB1 TUBB KEGG PATHOGENIC ESCHERICHIA COLI INFECFECTION Pathogenic Escherichiacoli infection 306119864 minus 02
SHC1 VAV3GNB2 KEGG CHEMOKINE SIGNALING PATHWAY Chemokine signaling pathway 575119864 minus 02
ITGB1 SHC1VAV3 KEGG FOCAL ADHESION Focal adhesion 659119864 minus 02
ITGB1 VAV3MYH9 KEGG REGULATION OF ACTIN CYTOSKELETON Regulation of actin cytoskeleton 782119864 minus 02
ITGB1 VAV3 KEGG LEUKOCYTE TRANSENDOTHELIAL MIGRATRATION Leukocyte transendothelial migration 105119864 minus 01
002040608
11214
U937
CON
NC-
low
dos
e
MiR
-dow
n-lo
w d
ose
MiR
-up-
low
dos
e
NC-
high
dos
e
MiR
-dow
n-hi
gh d
ose
MiR
-up-
high
dos
e
lowastlowast
lowastlowast
Integrin 1205731
GAPDH
1 2 3 4 5 6 7 8
Rela
tive i
nteg
rin1205731
prot
ein
leve
ls
(a)
0
02
04
06
08
1
12
Relat
ive A
MIG
O2
U937
CON
NC-
low
dos
e
MiR
-dow
n-lo
w d
ose
MiR
-up-
low
dos
e
NC-
high
dos
e
MiR
-dow
n-hi
gh d
ose
MiR
-up-
high
dos
e
Integrin 1205731
GAPDH
1 2 3 4 5 6 7 8pr
otei
n le
vels
(b)
Figure 2 Confirmation of microarray data withWestern blotting analysesThe endometrial stromal cells transfected withmiR-183-lentivirus(low dose or high dose) In-miR-183-lentivirus (low dose or high dose) and GFP-lentivirus (NC normal control) were used to examine theexpression alterations of integrin 1205731 (a) and AMIGO2 (b) 120573-actin served as protein loading control Compared with the control groupintegrin 1205731 expression in the miR-183-overexpression group was significantly decreased whereas it increased sharply in cells with low miR-183 expression AMIGO2 was largely not regulated by miR-183 lowast119875 lt 005 when compared to the negative control Error bars represent plusmnSEM
Presenilin 2 and VAV3 was detected in endometrial stromalcells (data not shown) and these two genes were not furtherstudied
35 MiR-183 Directly Targets ITBG1 but Not AMIGO2 Toinvestigate if ITBG1 and AMIGO2 are the direct targets of
miR-183 we performed luciferase assay to determinewhethermiR-183 binds to the 31015840UTR of ITBG1AMIGO2 mRNA(Figure 3(a)) Our results showed that reporter plasmidcontaining the miR-183 targeting sequence of ITGB1 mRNAdisplayed a significantly decreased luciferase activity in cellstransfected with miR-183 (119875 lt 005) whereas luciferase
6 BioMed Research International
3998400UTR5998400UTRAMIGO2
Has-miR-183
Mutant binding site
Coding region
5998400-UUCAGUUCCUCAAUGUGCCAUA-3998400
3998400-UCACUUAAGAUGGUCACGGUAU-5998400
5998400-UUCAGUUCCUCAAUCAUAACGC-3998400
174ndash181 bp
3998400UTR5998400UTRAMIGO2
Mutant binding site
Coding region
5998400-UCAUCAUAAUUUGUUGCCAUAA-3998400
3998400-CACUUAAGAUGGUCACGGUAU-5998400
5998400-UCAUCAUAAUUUGUAUAACGC-3998400
691ndash697 bp
3998400UTR5998400UTR
ITGB1
Has-miR-183
Mutant binding site
Coding region
5998400-GCUUUAAAACCUGUGUGCCAUU-3998400
3998400-UCACUUAAGAUGGUCACGGUAU-5998400
5998400-GCUUUAAAACCUGUUGUGACGU-3998400
695ndash701 bp
ITGB1 3998400UTR
998400Mutant 3 UTR
AMIGO2 3998400UTR
Mutant 3998400UTR
Has-miR-183
AMIGO2 3998400UTR
Mutant 3998400UTR
(a)
0
02
04
06
08
1
12
14
16
ITGB1-NC Wild-type Mutant type
Relat
ive l
ucife
rase
activ
ity (I
TGB1
)
MiR-183-NCMiR-183
0
02
04
06
08
1
12
14
16
AMIGO2-NC Wild-type Mutant type
Relat
ive L
ucife
rase
activ
ity (A
MIG
O2)
MiR-183-NCMiR-183
3998400UTR 3998400UTR 3998400UTR 3998400UTR
lowast
(b)
Figure 3 ITGB1 but not AMIGO2 is the direct target of miR-183 (a)The binding site of miR-183 in the 31015840UTR of ITGB1 (top) and AMIGO2(bottom) 31015840UTR along with the mutant construct in the predicted binding region (b) MiR-183 but not AMIGO2 directly targets ITBG1Endometrial stromal cells were cotransfected with wild-type reporter containing the ITBG1AMIGO2 31015840UTR or mutant ITBG1AMIGO231015840UTR plus miR-183 or negative control using Invitrogen Lipofectamine 2000 reagents Luminescence was measured after 24 hours oftransfection Reporter plasmid expressing ITGB1 mRNA that contains sequences potentially targeted by miR-183 displayed a significantlydecreased luciferase activity in cells transfected with miR-183 (119875 lt 005) whereas luciferase activity of reporter plasmid containing mutantsequence of ITGB1 was not changed following forced expression of miR-183 lowast119875 lt 005 when compared to the negative control Error barsrepresent plusmn SEM
BioMed Research International 7
0
02
04
06
08
1
12
14
Normal EM Eutopic EM
ITG
B1 ex
pres
sion
(nor
mal
ized
)
lowast
Figure 4 Validation of ITGB1 expression in eutopic endometriumwith endometriosis (119899 = 19) versus normal group (119899 = 18) byqPCR The results were analyzed using the comparative thresholdcycle (CT) method Gene expression data is presented as the foldof change relative to the levels of reference gene lowast119875 lt 005 whencompared to the normal group Error bars represent plusmn SEM
activity of reporter plasmid containing mutant sequence ofITGB1 was not changed following forced expression of miR-183 (Figure 3(b)) We found no significant change whencompared to the luciferase activity between cells transfectedwith wild-type or mutated AMIGO2 31015840UTR (119875 gt 005Figure 3(b) Thus these data suggested that miR-183 nega-tively regulates the expression of ITBG1 by directly target-ing their 31015840UTR whereas miR-183 does not directly targetAMIGO2 31015840UTR
36 ITGB1 Is Overexpressed in Endometrium from Endomet-riosis Patients Real-time RT-PCR results indicated thatITGB was significantly increased in the eutopic endometrialtissues from patients with endometriosis (119899 = 19) comparedwith endometrium from normal women (119899 = 18) (119875 lt 005)(Figure 4)
37 Overexpression of ITGB1 Rescued the Repressive Effectsof miR-183 on Endometrial Stromal Cells To ascertain thatmiR-183 regulates the function of endometrial stromal cellsthrough its interaction with ITGB1 a rescue experiment wasperformed Overexpression of ITGB1 partially rescued therepressive effects of miR-183 leading to elevated invasionabilities in the cells (Figure 5) This data indicated that miR-183 targets ITGB1 which in turn led to negative regulation onthe invasive activity of endometrial stromal cells
4 Discussion
In the previous study we performed miRNA microarrayscreening and identified several differentially expressed miR-NAs in the normal eutopic and ectopic endometriumAmong these miRNA species miR-183 was found to be
downregulated in the ectopic and eutopic tissues Functionalanalysis indicated that miR-183 promoted endometrial stro-mal cell apoptosis and had a negative regulatory impact onthe invasive ability of cells although it had no effect on cellproliferation [8] These findings suggested that the down-regulation of miR-183 expression might be involved in thepathogenesis of endometriosis The molecular mechanismshowever remain to be characterized
MiR-183 is a member of a miRNA family (miR-183 miR-182 and miR-96) that clusters within a 2ndash4 kb region atchromosome 7q32These miRNAs are known to regulate celldifferentiation apoptosis motility adhesion and invasion[12] Moreover it has been reported that miR-183 is upregu-lated in colorectal cancer prostate cancer and hepatocellularcarcinomas and downregulated in ovarian cancer breastcancer stem cells and osteosarcomas pointing to varied andcell type-dependent function of miR-183 Several target genesof miR-183 including ITGB1 Taok1 Ezrin EGR1 PDCD4and LRP6 have been identified and validated [13ndash19]
To systematically investigate the potential mechanismby which miR-183 may contribute to the development ofendometriosis we performed microarray analysis to identifythe target genes with relevance to cell functions in miR-183-overexpressing endometrial stromal cells Eight genes werefound to be downregulated in endometrial stromal cells withmiR-183 overexpression Among them the GO databasespathway analyses and PubMed reports pointed to importantassociation of 4 genes (ITGB1 AMIGO2 VAV3 and PSEN2)with cell adhesion cell migration and cell invasivenessWestern blotting results showed that endometrial stromalcell secreted integrin 1205731 and AMIGO2 but not VAV3 andPresenilin 2 The protein expression of integrin 1205731 appearedto be a direct regulation target of miR-183 which has beenproved in human trabecular meshwork cells human diploidfibroblasts and hela cells [19] AMIGO2 was not directlyregulated by miR-183
According to Sampsonrsquos theory endometriosis is thoughtto be initiated via retrograde endometrium into peri-toneal cavity during menstruation However while this phe-nomenon occurs in approximately 75ndash90 of women farfewer suffer the disease suggesting that additional factorsmust contribute to the process Invasion to ectopic locations isthe key step for the initiation of endometriosis during whichadhesion molecules play an important role The integrinfamily includes glycoproteins that formdimeric structure andmediate cell attachment to the ECM components [20] It hasbeen reported that 1205731 integrin-depleted cells had reducedinvasive capabilities and 1205731 integrin signaling was involvedin cancer cell invasion [21] Integrin 1205731 is constitutivelyexpressed in endometrial stromal cells and endometrioticlesions [22] Increased integrin 1205731 protein expression inendometrial stromal cells was observed in endometrioticcompared to normal endometrial tissues [23] It was reportedthat downregulation of prostaglandin E2 receptors EP2 andEP4 led to inhibited adhesion of human endometrioticepithelial and stromal cells Importantly this inhibitioncould be mediated through suppression of integrin-mediatedmechanisms [24] This finding provided a strong evidence
8 BioMed Research International
(a)
0
05
1
15
2
25
NC MiR-183 MiR-183ITGB1
Inva
sive r
ate (
OO
D57
0)
lowast
(b)
Figure 5 Overexpression of ITGB1 rescued the repressive effects of miR-183 on endometrial stromal cells leading to elevated invasiveabilities in transwell assays Endometrial stromal cells were infected with miR-183-lentivirus miR-183ITGB1-lentivirus and GFP-lentivirusin upper wells After 24 hours the number of cells that invaded through Matrigel was counted in at least 10 fields per well (a) Representativephotographs show that miR-183 inhibited the invasiveness of endometrial stromal cells whereas ITGB1 partially rescued the repressive effectsof miR-183 (b) Cell counting results indicate that ITGB1 overexpression rescued the repressive effects of miR-183 lowast119875 lt 005 when comparedto the negative control Error bars represent plusmn SEM
for the significance of integrins in the adhesion function ofendometrial stromal cells
AMIGO2 (ALI1 DEGA) was first reported to be prefer-entially expressed in the central nervous system Sequenceanalyses revealed that the protein contains seven leucine-richrepeats one IgC2-like loop and a transmembrane domainand displays homology to Kek and Trk families [25ndash27] Inthe gastric adenocarcinoma cell line AMIGO2 was reportedto be involved in cell adhesion extracellular matrix andbasement membrane formation [28] AMIGO2 was alsoimplicated in oral carcinogenesis by preventing the arrest ofcell cycle progression at the G1 phase through inhibiting theexpression of cyclin-dependent kinase inhibitors Howeverlittle is known about the role of AMIGO2 in the developmentof endometriosis
To investigate whether miR-183 directly targets ITGB1and AMIGO2 dual luciferase reporter assays were carriedout We found that miR-183 negatively regulated the expres-sion of ITBG1 by directly targeting its 31015840UTR but mightnot directly regulate the expression of AMIGO2 We further
detected differential expression of ITGB1 in endometriumwith or without endometriosis by quantitative RT-PCR Thehigher expression of integrin 1205731 in endometrium tissues inendometriosis was confirmed which is consistent with theresult for higher integrin 1205731 expression in ectopic stromalcells than normal endometrial stromal cells Previous studieshave shown that integrin 1205731 mediates cell adherence toextracellular matrix [29] Indeed the rescue experimentindicated that overexpression of ITGB1 blocked the repressiveeffects of miR-183 on endometrial stromal cell invasivenesswhich provided a critical evidence supporting that the lowerexpression of miR-183 may promote ITGB1 expression andcontribute to the development of endometriosis
In conclusion altered miR-183 expression may causederegulation of its target gene ITBG1 affecting the celladhesiveness and invasiveness of endometrial stromal cellswhich in turn lead to pathogenesis of endometriosis Thesefindings revealed a novel pathologic role of miR-183 forendometriosis It would be of great interest to investigate ifthese alterations may serve as biomarkers for the prediction
BioMed Research International 9
andor treatment of endometriosisThe regulation ofmiR-183and the exact mechanism by which miR-183 regulates ITBG1expression remain to be characterized
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This studywas supported by grants from theNational NaturalScience Foundation of China (nos 81100405 and 81471438)the Key Program ofNanjingMedical Science and TechnologyDevelopment Foundation Nanjing Department of Health(no QRX11109)
References
[1] D R Nyholt S-K Low C A Anderson et al ldquoGenome-wide association meta-analysis identifies new endometriosisrisk locirdquo Nature Genetics vol 44 no 12 pp 1355ndash1359 2012
[2] W Filipowicz S N Bhattacharyya and N Sonenberg ldquoMecha-nisms of post-transcriptional regulation by microRNAs are theanswers in sightrdquoNature Reviews Genetics vol 9 no 2 pp 102ndash114 2008
[3] X Santamaria and H Taylor ldquoMicroRNA and gynecologicalreproductive diseasesrdquo Fertility and Sterility vol 101 no 6 pp1545ndash1551 2014
[4] Y Shimono M Zabala R W Cho et al ldquoDownregulation ofmiRNA-200c links breast cancer stem cells with normal stemcellsrdquo Cell vol 138 no 3 pp 592ndash603 2009
[5] M Adammek B Greve N Kassens et al ldquoMicroRNA miR-145 inhibits proliferation invasiveness and stem cell phenotypeof an in vitro endometriosis model by targeting multiplecytoskeletal elements and pluripotency factorsrdquo Fertility andSterility vol 99 no 5 pp 1346e5ndash1355e5 2013
[6] W Abe K Nasu C Nakada Y Kawano M Moriyama andH Narahara ldquoMiR-196b targets c-myc and Bcl-2 expressioninhibits proliferation and induces apoptosis in endometrioticstromal cellsrdquo Human Reproduction vol 28 no 3 pp 750ndash7612013
[7] W B Nothnick A Graham J Holbert et al ldquomiR-451deficiency is associated with altered endometrial fibrinogenalpha chain expression and reduced endometriotic implantestablishment in an experimental mouse modelrdquo PLoS ONEvol 9 no 6 Article ID e100336 2014
[8] X-Y Shi L Gu J Chen X-R Guo and Y-L Shi ldquoDownregu-lation of miR-183 inhibits apoptosis and enhances the invasivepotential of endometrial stromal cells in endometriosisrdquo Inter-national Journal of Molecular Medicine vol 33 no 1 pp 59ndash672014
[9] Y-L Shi X-Z Luo X-Y Zhu K-Q Hua Y Zhu and D-JLi ldquoEffects of combined 17120573-estradiol with TCDD on secretionof chemokine IL-8 and expression of its receptor CXCR1 inendometriotic focus-associated cells in co-culturerdquo HumanReproduction vol 21 no 4 pp 870ndash879 2006
[10] B Salhia N L Tran A Chan et al ldquoThe guanine nucleotideexchange factors Trio Ect2 and Vav3 mediate the invasivebehavior of glioblastomardquo American Journal of Pathology vol173 no 6 pp 1828ndash1838 2008
[11] B Liu L Wang L-L Shen et al ldquoRNAi-mediated inhibitionof presenilin 2 inhibits glioma cell growth and invasion andis involved in the regulation of Nrg1ErbB signalingrdquo Neuro-Oncology vol 14 no 8 pp 994ndash1006 2012
[12] S D Weeraratne V Amani N Teider et al ldquoPleiotropiceffects of miR-183sim96sim182 converge to regulate cell survivalproliferation and migration in medulloblastomardquo Acta Neu-ropathologica vol 123 no 4 pp 539ndash552 2012
[13] M Patel Q Cai D Ding R Salvi Z Hu and B H Hu ldquoThemiR-183Taok1 target pair is implicated in cochlear responsesto acoustic traumardquo PLoS ONE vol 8 no 3 Article ID e584712013
[14] J Zhu Y Feng Z Ke et al ldquoDown-regulation of miR-183promotes migration and invasion of osteosarcoma by targetingEzrinrdquo The American Journal of Pathology vol 180 no 6 pp2440ndash2451 2012
[15] A J Lowery N Miller R M Dwyer and M J Kerin ldquoDysreg-ulated miR-183 inhibits migration in breast cancer cellsrdquo BMCCancer vol 10 article 502 2010
[16] A L Sarver H Li and S Subramanian ldquoMicroRNA miR-183functions as an oncogene by targeting the transcription factorEGR1 and promoting tumor cell migrationrdquo Cancer Researchvol 70 no 23 pp 9570ndash9580 2010
[17] J Li H Fu C Xu et al ldquoMiR-183 inhibits TGF-beta1-inducedapoptosis by downregulation of PDCD4 expression in humanhepatocellular carcinoma cellsrdquoBMCCancer vol 10 article 3542010
[18] C Chen H Xiang Y-L Peng J Peng and S-W JiangldquoMature miR-183 negatively regulated by transcription factorGATA3 promotes 3T3-L1 adipogenesis through inhibition ofthe canonical Wnt120573-catenin signaling pathway by targetingLRP6rdquo Cellular Signalling vol 26 no 6 pp 1155ndash1165 2014
[19] G Li C Luna J Qiu D L Epstein and P Gonzalez ldquoTargetingof integrin beta1 and kinesin 2alpha by microRNA 183rdquo TheJournal of Biological Chemistry vol 285 no 8 pp 5461ndash54712010
[20] A M Schooley N M Andrews H Zhao and C L Addisonldquo1205731 integrin is required for anchorage-independent growth andinvasion of tumor cells in a context dependent mannerrdquo CancerLetters vol 316 no 2 pp 157ndash167 2012
[21] Y Zhang YGongDHu et al ldquoNuclear SIPA1 activates integrin1205731 promoter and promotes invasion of breast cancer cellsrdquoOncogene vol 34 no 11 pp 1451ndash1462 2015
[22] Y B Aznaurova M B Zhumataev T K Roberts A M Aliperand A A Zhavoronkov ldquoMolecular aspects of developmentand regulation of endometriosisrdquo Reproductive Biology andEndocrinology vol 12 article 50 2014
[23] M Adachi K Nasu A Tsuno A Yuge Y Kawano and HNarahara ldquoAttachment to extracellular matrices is enhancedin human endometriotic stromal cells a possible mechanismunderlying the pathogenesis of endometriosisrdquo European Jour-nal of Obstetrics Gynecology and Reproductive Biology vol 155no 1 pp 85ndash88 2011
[24] J Lee S K Banu R C Burghardt A Starzinski-Powitz andJ A Arosh ldquoSelective inhibition of prostaglandin E2 receptorsEP2 and EP4 inhibits adhesion of human endometriotic epithe-lial and stromal cells through suppression of integrin-mediatedmechanismsrdquo Biology of Reproduction vol 88 no 3 article 772013
[25] A Laeremans J NysW Luyten R DrsquoHoogeM Paulussen andL Arckens ldquoAMIGO2 mRNA expression in hippocampal CA2
10 BioMed Research International
and CA3ardquo Brain Structure and Function vol 218 no 1 pp 123ndash130 2013
[26] T Ono N Sekino-Suzuki Y Kikkawa H Yonekawa andS Kawashima ldquoAlivin 1 a novel neuronal activity-dependentgene inhibits apoptosis and promotes survival of cerebellargranule neuronsrdquo Journal of Neuroscience vol 23 no 13 pp5887ndash5896 2003
[27] J Kuja-Panula M Kiiltomaki T Yamashiro A Rouhiainenand H Rauvala ldquoAMIGO a transmembrane protein implicatedin axon tract development defines a novel protein family withleucine-rich repeatsrdquoThe Journal of Cell Biology vol 160 no 6pp 963ndash973 2003
[28] K E Rabenau J M OrsquoToole R Bassi et al ldquoDEGAAMIGO-2 a leucine-rich repeat family member differentially expressedin human gastric adenocarcinoma effects on ploidy chromo-somal stability cell adhesionmigration and tumorigenicityrdquoOncogene vol 23 no 29 pp 5056ndash5067 2004
[29] J A Garcia-Velasco and A Arici ldquoInterleukin-8 expression inendometrial stromal cells is regulated by integrin-dependentcell adhesionrdquo Molecular Human Reproduction vol 5 no 12pp 1135ndash1140 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
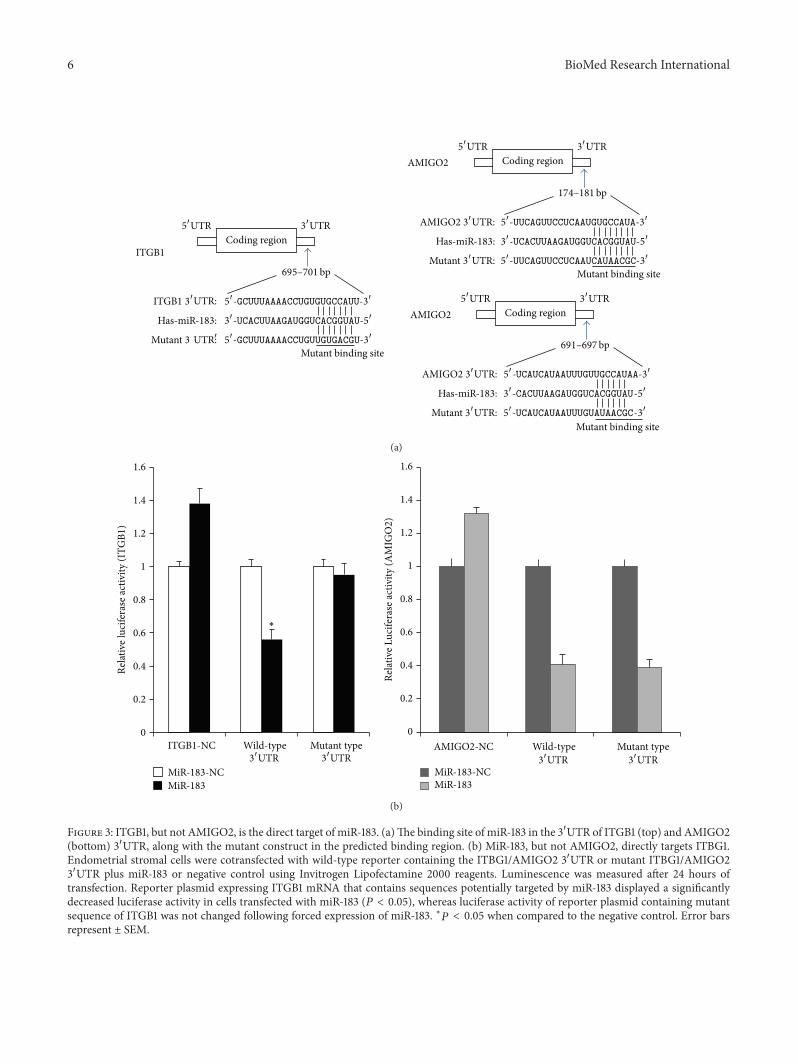
6 BioMed Research International
3998400UTR5998400UTRAMIGO2
Has-miR-183
Mutant binding site
Coding region
5998400-UUCAGUUCCUCAAUGUGCCAUA-3998400
3998400-UCACUUAAGAUGGUCACGGUAU-5998400
5998400-UUCAGUUCCUCAAUCAUAACGC-3998400
174ndash181 bp
3998400UTR5998400UTRAMIGO2
Mutant binding site
Coding region
5998400-UCAUCAUAAUUUGUUGCCAUAA-3998400
3998400-CACUUAAGAUGGUCACGGUAU-5998400
5998400-UCAUCAUAAUUUGUAUAACGC-3998400
691ndash697 bp
3998400UTR5998400UTR
ITGB1
Has-miR-183
Mutant binding site
Coding region
5998400-GCUUUAAAACCUGUGUGCCAUU-3998400
3998400-UCACUUAAGAUGGUCACGGUAU-5998400
5998400-GCUUUAAAACCUGUUGUGACGU-3998400
695ndash701 bp
ITGB1 3998400UTR
998400Mutant 3 UTR
AMIGO2 3998400UTR
Mutant 3998400UTR
Has-miR-183
AMIGO2 3998400UTR
Mutant 3998400UTR
(a)
0
02
04
06
08
1
12
14
16
ITGB1-NC Wild-type Mutant type
Relat
ive l
ucife
rase
activ
ity (I
TGB1
)
MiR-183-NCMiR-183
0
02
04
06
08
1
12
14
16
AMIGO2-NC Wild-type Mutant type
Relat
ive L
ucife
rase
activ
ity (A
MIG
O2)
MiR-183-NCMiR-183
3998400UTR 3998400UTR 3998400UTR 3998400UTR
lowast
(b)
Figure 3 ITGB1 but not AMIGO2 is the direct target of miR-183 (a)The binding site of miR-183 in the 31015840UTR of ITGB1 (top) and AMIGO2(bottom) 31015840UTR along with the mutant construct in the predicted binding region (b) MiR-183 but not AMIGO2 directly targets ITBG1Endometrial stromal cells were cotransfected with wild-type reporter containing the ITBG1AMIGO2 31015840UTR or mutant ITBG1AMIGO231015840UTR plus miR-183 or negative control using Invitrogen Lipofectamine 2000 reagents Luminescence was measured after 24 hours oftransfection Reporter plasmid expressing ITGB1 mRNA that contains sequences potentially targeted by miR-183 displayed a significantlydecreased luciferase activity in cells transfected with miR-183 (119875 lt 005) whereas luciferase activity of reporter plasmid containing mutantsequence of ITGB1 was not changed following forced expression of miR-183 lowast119875 lt 005 when compared to the negative control Error barsrepresent plusmn SEM
BioMed Research International 7
0
02
04
06
08
1
12
14
Normal EM Eutopic EM
ITG
B1 ex
pres
sion
(nor
mal
ized
)
lowast
Figure 4 Validation of ITGB1 expression in eutopic endometriumwith endometriosis (119899 = 19) versus normal group (119899 = 18) byqPCR The results were analyzed using the comparative thresholdcycle (CT) method Gene expression data is presented as the foldof change relative to the levels of reference gene lowast119875 lt 005 whencompared to the normal group Error bars represent plusmn SEM
activity of reporter plasmid containing mutant sequence ofITGB1 was not changed following forced expression of miR-183 (Figure 3(b)) We found no significant change whencompared to the luciferase activity between cells transfectedwith wild-type or mutated AMIGO2 31015840UTR (119875 gt 005Figure 3(b) Thus these data suggested that miR-183 nega-tively regulates the expression of ITBG1 by directly target-ing their 31015840UTR whereas miR-183 does not directly targetAMIGO2 31015840UTR
36 ITGB1 Is Overexpressed in Endometrium from Endomet-riosis Patients Real-time RT-PCR results indicated thatITGB was significantly increased in the eutopic endometrialtissues from patients with endometriosis (119899 = 19) comparedwith endometrium from normal women (119899 = 18) (119875 lt 005)(Figure 4)
37 Overexpression of ITGB1 Rescued the Repressive Effectsof miR-183 on Endometrial Stromal Cells To ascertain thatmiR-183 regulates the function of endometrial stromal cellsthrough its interaction with ITGB1 a rescue experiment wasperformed Overexpression of ITGB1 partially rescued therepressive effects of miR-183 leading to elevated invasionabilities in the cells (Figure 5) This data indicated that miR-183 targets ITGB1 which in turn led to negative regulation onthe invasive activity of endometrial stromal cells
4 Discussion
In the previous study we performed miRNA microarrayscreening and identified several differentially expressed miR-NAs in the normal eutopic and ectopic endometriumAmong these miRNA species miR-183 was found to be
downregulated in the ectopic and eutopic tissues Functionalanalysis indicated that miR-183 promoted endometrial stro-mal cell apoptosis and had a negative regulatory impact onthe invasive ability of cells although it had no effect on cellproliferation [8] These findings suggested that the down-regulation of miR-183 expression might be involved in thepathogenesis of endometriosis The molecular mechanismshowever remain to be characterized
MiR-183 is a member of a miRNA family (miR-183 miR-182 and miR-96) that clusters within a 2ndash4 kb region atchromosome 7q32These miRNAs are known to regulate celldifferentiation apoptosis motility adhesion and invasion[12] Moreover it has been reported that miR-183 is upregu-lated in colorectal cancer prostate cancer and hepatocellularcarcinomas and downregulated in ovarian cancer breastcancer stem cells and osteosarcomas pointing to varied andcell type-dependent function of miR-183 Several target genesof miR-183 including ITGB1 Taok1 Ezrin EGR1 PDCD4and LRP6 have been identified and validated [13ndash19]
To systematically investigate the potential mechanismby which miR-183 may contribute to the development ofendometriosis we performed microarray analysis to identifythe target genes with relevance to cell functions in miR-183-overexpressing endometrial stromal cells Eight genes werefound to be downregulated in endometrial stromal cells withmiR-183 overexpression Among them the GO databasespathway analyses and PubMed reports pointed to importantassociation of 4 genes (ITGB1 AMIGO2 VAV3 and PSEN2)with cell adhesion cell migration and cell invasivenessWestern blotting results showed that endometrial stromalcell secreted integrin 1205731 and AMIGO2 but not VAV3 andPresenilin 2 The protein expression of integrin 1205731 appearedto be a direct regulation target of miR-183 which has beenproved in human trabecular meshwork cells human diploidfibroblasts and hela cells [19] AMIGO2 was not directlyregulated by miR-183
According to Sampsonrsquos theory endometriosis is thoughtto be initiated via retrograde endometrium into peri-toneal cavity during menstruation However while this phe-nomenon occurs in approximately 75ndash90 of women farfewer suffer the disease suggesting that additional factorsmust contribute to the process Invasion to ectopic locations isthe key step for the initiation of endometriosis during whichadhesion molecules play an important role The integrinfamily includes glycoproteins that formdimeric structure andmediate cell attachment to the ECM components [20] It hasbeen reported that 1205731 integrin-depleted cells had reducedinvasive capabilities and 1205731 integrin signaling was involvedin cancer cell invasion [21] Integrin 1205731 is constitutivelyexpressed in endometrial stromal cells and endometrioticlesions [22] Increased integrin 1205731 protein expression inendometrial stromal cells was observed in endometrioticcompared to normal endometrial tissues [23] It was reportedthat downregulation of prostaglandin E2 receptors EP2 andEP4 led to inhibited adhesion of human endometrioticepithelial and stromal cells Importantly this inhibitioncould be mediated through suppression of integrin-mediatedmechanisms [24] This finding provided a strong evidence
8 BioMed Research International
(a)
0
05
1
15
2
25
NC MiR-183 MiR-183ITGB1
Inva
sive r
ate (
OO
D57
0)
lowast
(b)
Figure 5 Overexpression of ITGB1 rescued the repressive effects of miR-183 on endometrial stromal cells leading to elevated invasiveabilities in transwell assays Endometrial stromal cells were infected with miR-183-lentivirus miR-183ITGB1-lentivirus and GFP-lentivirusin upper wells After 24 hours the number of cells that invaded through Matrigel was counted in at least 10 fields per well (a) Representativephotographs show that miR-183 inhibited the invasiveness of endometrial stromal cells whereas ITGB1 partially rescued the repressive effectsof miR-183 (b) Cell counting results indicate that ITGB1 overexpression rescued the repressive effects of miR-183 lowast119875 lt 005 when comparedto the negative control Error bars represent plusmn SEM
for the significance of integrins in the adhesion function ofendometrial stromal cells
AMIGO2 (ALI1 DEGA) was first reported to be prefer-entially expressed in the central nervous system Sequenceanalyses revealed that the protein contains seven leucine-richrepeats one IgC2-like loop and a transmembrane domainand displays homology to Kek and Trk families [25ndash27] Inthe gastric adenocarcinoma cell line AMIGO2 was reportedto be involved in cell adhesion extracellular matrix andbasement membrane formation [28] AMIGO2 was alsoimplicated in oral carcinogenesis by preventing the arrest ofcell cycle progression at the G1 phase through inhibiting theexpression of cyclin-dependent kinase inhibitors Howeverlittle is known about the role of AMIGO2 in the developmentof endometriosis
To investigate whether miR-183 directly targets ITGB1and AMIGO2 dual luciferase reporter assays were carriedout We found that miR-183 negatively regulated the expres-sion of ITBG1 by directly targeting its 31015840UTR but mightnot directly regulate the expression of AMIGO2 We further
detected differential expression of ITGB1 in endometriumwith or without endometriosis by quantitative RT-PCR Thehigher expression of integrin 1205731 in endometrium tissues inendometriosis was confirmed which is consistent with theresult for higher integrin 1205731 expression in ectopic stromalcells than normal endometrial stromal cells Previous studieshave shown that integrin 1205731 mediates cell adherence toextracellular matrix [29] Indeed the rescue experimentindicated that overexpression of ITGB1 blocked the repressiveeffects of miR-183 on endometrial stromal cell invasivenesswhich provided a critical evidence supporting that the lowerexpression of miR-183 may promote ITGB1 expression andcontribute to the development of endometriosis
In conclusion altered miR-183 expression may causederegulation of its target gene ITBG1 affecting the celladhesiveness and invasiveness of endometrial stromal cellswhich in turn lead to pathogenesis of endometriosis Thesefindings revealed a novel pathologic role of miR-183 forendometriosis It would be of great interest to investigate ifthese alterations may serve as biomarkers for the prediction
BioMed Research International 9
andor treatment of endometriosisThe regulation ofmiR-183and the exact mechanism by which miR-183 regulates ITBG1expression remain to be characterized
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This studywas supported by grants from theNational NaturalScience Foundation of China (nos 81100405 and 81471438)the Key Program ofNanjingMedical Science and TechnologyDevelopment Foundation Nanjing Department of Health(no QRX11109)
References
[1] D R Nyholt S-K Low C A Anderson et al ldquoGenome-wide association meta-analysis identifies new endometriosisrisk locirdquo Nature Genetics vol 44 no 12 pp 1355ndash1359 2012
[2] W Filipowicz S N Bhattacharyya and N Sonenberg ldquoMecha-nisms of post-transcriptional regulation by microRNAs are theanswers in sightrdquoNature Reviews Genetics vol 9 no 2 pp 102ndash114 2008
[3] X Santamaria and H Taylor ldquoMicroRNA and gynecologicalreproductive diseasesrdquo Fertility and Sterility vol 101 no 6 pp1545ndash1551 2014
[4] Y Shimono M Zabala R W Cho et al ldquoDownregulation ofmiRNA-200c links breast cancer stem cells with normal stemcellsrdquo Cell vol 138 no 3 pp 592ndash603 2009
[5] M Adammek B Greve N Kassens et al ldquoMicroRNA miR-145 inhibits proliferation invasiveness and stem cell phenotypeof an in vitro endometriosis model by targeting multiplecytoskeletal elements and pluripotency factorsrdquo Fertility andSterility vol 99 no 5 pp 1346e5ndash1355e5 2013
[6] W Abe K Nasu C Nakada Y Kawano M Moriyama andH Narahara ldquoMiR-196b targets c-myc and Bcl-2 expressioninhibits proliferation and induces apoptosis in endometrioticstromal cellsrdquo Human Reproduction vol 28 no 3 pp 750ndash7612013
[7] W B Nothnick A Graham J Holbert et al ldquomiR-451deficiency is associated with altered endometrial fibrinogenalpha chain expression and reduced endometriotic implantestablishment in an experimental mouse modelrdquo PLoS ONEvol 9 no 6 Article ID e100336 2014
[8] X-Y Shi L Gu J Chen X-R Guo and Y-L Shi ldquoDownregu-lation of miR-183 inhibits apoptosis and enhances the invasivepotential of endometrial stromal cells in endometriosisrdquo Inter-national Journal of Molecular Medicine vol 33 no 1 pp 59ndash672014
[9] Y-L Shi X-Z Luo X-Y Zhu K-Q Hua Y Zhu and D-JLi ldquoEffects of combined 17120573-estradiol with TCDD on secretionof chemokine IL-8 and expression of its receptor CXCR1 inendometriotic focus-associated cells in co-culturerdquo HumanReproduction vol 21 no 4 pp 870ndash879 2006
[10] B Salhia N L Tran A Chan et al ldquoThe guanine nucleotideexchange factors Trio Ect2 and Vav3 mediate the invasivebehavior of glioblastomardquo American Journal of Pathology vol173 no 6 pp 1828ndash1838 2008
[11] B Liu L Wang L-L Shen et al ldquoRNAi-mediated inhibitionof presenilin 2 inhibits glioma cell growth and invasion andis involved in the regulation of Nrg1ErbB signalingrdquo Neuro-Oncology vol 14 no 8 pp 994ndash1006 2012
[12] S D Weeraratne V Amani N Teider et al ldquoPleiotropiceffects of miR-183sim96sim182 converge to regulate cell survivalproliferation and migration in medulloblastomardquo Acta Neu-ropathologica vol 123 no 4 pp 539ndash552 2012
[13] M Patel Q Cai D Ding R Salvi Z Hu and B H Hu ldquoThemiR-183Taok1 target pair is implicated in cochlear responsesto acoustic traumardquo PLoS ONE vol 8 no 3 Article ID e584712013
[14] J Zhu Y Feng Z Ke et al ldquoDown-regulation of miR-183promotes migration and invasion of osteosarcoma by targetingEzrinrdquo The American Journal of Pathology vol 180 no 6 pp2440ndash2451 2012
[15] A J Lowery N Miller R M Dwyer and M J Kerin ldquoDysreg-ulated miR-183 inhibits migration in breast cancer cellsrdquo BMCCancer vol 10 article 502 2010
[16] A L Sarver H Li and S Subramanian ldquoMicroRNA miR-183functions as an oncogene by targeting the transcription factorEGR1 and promoting tumor cell migrationrdquo Cancer Researchvol 70 no 23 pp 9570ndash9580 2010
[17] J Li H Fu C Xu et al ldquoMiR-183 inhibits TGF-beta1-inducedapoptosis by downregulation of PDCD4 expression in humanhepatocellular carcinoma cellsrdquoBMCCancer vol 10 article 3542010
[18] C Chen H Xiang Y-L Peng J Peng and S-W JiangldquoMature miR-183 negatively regulated by transcription factorGATA3 promotes 3T3-L1 adipogenesis through inhibition ofthe canonical Wnt120573-catenin signaling pathway by targetingLRP6rdquo Cellular Signalling vol 26 no 6 pp 1155ndash1165 2014
[19] G Li C Luna J Qiu D L Epstein and P Gonzalez ldquoTargetingof integrin beta1 and kinesin 2alpha by microRNA 183rdquo TheJournal of Biological Chemistry vol 285 no 8 pp 5461ndash54712010
[20] A M Schooley N M Andrews H Zhao and C L Addisonldquo1205731 integrin is required for anchorage-independent growth andinvasion of tumor cells in a context dependent mannerrdquo CancerLetters vol 316 no 2 pp 157ndash167 2012
[21] Y Zhang YGongDHu et al ldquoNuclear SIPA1 activates integrin1205731 promoter and promotes invasion of breast cancer cellsrdquoOncogene vol 34 no 11 pp 1451ndash1462 2015
[22] Y B Aznaurova M B Zhumataev T K Roberts A M Aliperand A A Zhavoronkov ldquoMolecular aspects of developmentand regulation of endometriosisrdquo Reproductive Biology andEndocrinology vol 12 article 50 2014
[23] M Adachi K Nasu A Tsuno A Yuge Y Kawano and HNarahara ldquoAttachment to extracellular matrices is enhancedin human endometriotic stromal cells a possible mechanismunderlying the pathogenesis of endometriosisrdquo European Jour-nal of Obstetrics Gynecology and Reproductive Biology vol 155no 1 pp 85ndash88 2011
[24] J Lee S K Banu R C Burghardt A Starzinski-Powitz andJ A Arosh ldquoSelective inhibition of prostaglandin E2 receptorsEP2 and EP4 inhibits adhesion of human endometriotic epithe-lial and stromal cells through suppression of integrin-mediatedmechanismsrdquo Biology of Reproduction vol 88 no 3 article 772013
[25] A Laeremans J NysW Luyten R DrsquoHoogeM Paulussen andL Arckens ldquoAMIGO2 mRNA expression in hippocampal CA2
10 BioMed Research International
and CA3ardquo Brain Structure and Function vol 218 no 1 pp 123ndash130 2013
[26] T Ono N Sekino-Suzuki Y Kikkawa H Yonekawa andS Kawashima ldquoAlivin 1 a novel neuronal activity-dependentgene inhibits apoptosis and promotes survival of cerebellargranule neuronsrdquo Journal of Neuroscience vol 23 no 13 pp5887ndash5896 2003
[27] J Kuja-Panula M Kiiltomaki T Yamashiro A Rouhiainenand H Rauvala ldquoAMIGO a transmembrane protein implicatedin axon tract development defines a novel protein family withleucine-rich repeatsrdquoThe Journal of Cell Biology vol 160 no 6pp 963ndash973 2003
[28] K E Rabenau J M OrsquoToole R Bassi et al ldquoDEGAAMIGO-2 a leucine-rich repeat family member differentially expressedin human gastric adenocarcinoma effects on ploidy chromo-somal stability cell adhesionmigration and tumorigenicityrdquoOncogene vol 23 no 29 pp 5056ndash5067 2004
[29] J A Garcia-Velasco and A Arici ldquoInterleukin-8 expression inendometrial stromal cells is regulated by integrin-dependentcell adhesionrdquo Molecular Human Reproduction vol 5 no 12pp 1135ndash1140 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 7
0
02
04
06
08
1
12
14
Normal EM Eutopic EM
ITG
B1 ex
pres
sion
(nor
mal
ized
)
lowast
Figure 4 Validation of ITGB1 expression in eutopic endometriumwith endometriosis (119899 = 19) versus normal group (119899 = 18) byqPCR The results were analyzed using the comparative thresholdcycle (CT) method Gene expression data is presented as the foldof change relative to the levels of reference gene lowast119875 lt 005 whencompared to the normal group Error bars represent plusmn SEM
activity of reporter plasmid containing mutant sequence ofITGB1 was not changed following forced expression of miR-183 (Figure 3(b)) We found no significant change whencompared to the luciferase activity between cells transfectedwith wild-type or mutated AMIGO2 31015840UTR (119875 gt 005Figure 3(b) Thus these data suggested that miR-183 nega-tively regulates the expression of ITBG1 by directly target-ing their 31015840UTR whereas miR-183 does not directly targetAMIGO2 31015840UTR
36 ITGB1 Is Overexpressed in Endometrium from Endomet-riosis Patients Real-time RT-PCR results indicated thatITGB was significantly increased in the eutopic endometrialtissues from patients with endometriosis (119899 = 19) comparedwith endometrium from normal women (119899 = 18) (119875 lt 005)(Figure 4)
37 Overexpression of ITGB1 Rescued the Repressive Effectsof miR-183 on Endometrial Stromal Cells To ascertain thatmiR-183 regulates the function of endometrial stromal cellsthrough its interaction with ITGB1 a rescue experiment wasperformed Overexpression of ITGB1 partially rescued therepressive effects of miR-183 leading to elevated invasionabilities in the cells (Figure 5) This data indicated that miR-183 targets ITGB1 which in turn led to negative regulation onthe invasive activity of endometrial stromal cells
4 Discussion
In the previous study we performed miRNA microarrayscreening and identified several differentially expressed miR-NAs in the normal eutopic and ectopic endometriumAmong these miRNA species miR-183 was found to be
downregulated in the ectopic and eutopic tissues Functionalanalysis indicated that miR-183 promoted endometrial stro-mal cell apoptosis and had a negative regulatory impact onthe invasive ability of cells although it had no effect on cellproliferation [8] These findings suggested that the down-regulation of miR-183 expression might be involved in thepathogenesis of endometriosis The molecular mechanismshowever remain to be characterized
MiR-183 is a member of a miRNA family (miR-183 miR-182 and miR-96) that clusters within a 2ndash4 kb region atchromosome 7q32These miRNAs are known to regulate celldifferentiation apoptosis motility adhesion and invasion[12] Moreover it has been reported that miR-183 is upregu-lated in colorectal cancer prostate cancer and hepatocellularcarcinomas and downregulated in ovarian cancer breastcancer stem cells and osteosarcomas pointing to varied andcell type-dependent function of miR-183 Several target genesof miR-183 including ITGB1 Taok1 Ezrin EGR1 PDCD4and LRP6 have been identified and validated [13ndash19]
To systematically investigate the potential mechanismby which miR-183 may contribute to the development ofendometriosis we performed microarray analysis to identifythe target genes with relevance to cell functions in miR-183-overexpressing endometrial stromal cells Eight genes werefound to be downregulated in endometrial stromal cells withmiR-183 overexpression Among them the GO databasespathway analyses and PubMed reports pointed to importantassociation of 4 genes (ITGB1 AMIGO2 VAV3 and PSEN2)with cell adhesion cell migration and cell invasivenessWestern blotting results showed that endometrial stromalcell secreted integrin 1205731 and AMIGO2 but not VAV3 andPresenilin 2 The protein expression of integrin 1205731 appearedto be a direct regulation target of miR-183 which has beenproved in human trabecular meshwork cells human diploidfibroblasts and hela cells [19] AMIGO2 was not directlyregulated by miR-183
According to Sampsonrsquos theory endometriosis is thoughtto be initiated via retrograde endometrium into peri-toneal cavity during menstruation However while this phe-nomenon occurs in approximately 75ndash90 of women farfewer suffer the disease suggesting that additional factorsmust contribute to the process Invasion to ectopic locations isthe key step for the initiation of endometriosis during whichadhesion molecules play an important role The integrinfamily includes glycoproteins that formdimeric structure andmediate cell attachment to the ECM components [20] It hasbeen reported that 1205731 integrin-depleted cells had reducedinvasive capabilities and 1205731 integrin signaling was involvedin cancer cell invasion [21] Integrin 1205731 is constitutivelyexpressed in endometrial stromal cells and endometrioticlesions [22] Increased integrin 1205731 protein expression inendometrial stromal cells was observed in endometrioticcompared to normal endometrial tissues [23] It was reportedthat downregulation of prostaglandin E2 receptors EP2 andEP4 led to inhibited adhesion of human endometrioticepithelial and stromal cells Importantly this inhibitioncould be mediated through suppression of integrin-mediatedmechanisms [24] This finding provided a strong evidence
8 BioMed Research International
(a)
0
05
1
15
2
25
NC MiR-183 MiR-183ITGB1
Inva
sive r
ate (
OO
D57
0)
lowast
(b)
Figure 5 Overexpression of ITGB1 rescued the repressive effects of miR-183 on endometrial stromal cells leading to elevated invasiveabilities in transwell assays Endometrial stromal cells were infected with miR-183-lentivirus miR-183ITGB1-lentivirus and GFP-lentivirusin upper wells After 24 hours the number of cells that invaded through Matrigel was counted in at least 10 fields per well (a) Representativephotographs show that miR-183 inhibited the invasiveness of endometrial stromal cells whereas ITGB1 partially rescued the repressive effectsof miR-183 (b) Cell counting results indicate that ITGB1 overexpression rescued the repressive effects of miR-183 lowast119875 lt 005 when comparedto the negative control Error bars represent plusmn SEM
for the significance of integrins in the adhesion function ofendometrial stromal cells
AMIGO2 (ALI1 DEGA) was first reported to be prefer-entially expressed in the central nervous system Sequenceanalyses revealed that the protein contains seven leucine-richrepeats one IgC2-like loop and a transmembrane domainand displays homology to Kek and Trk families [25ndash27] Inthe gastric adenocarcinoma cell line AMIGO2 was reportedto be involved in cell adhesion extracellular matrix andbasement membrane formation [28] AMIGO2 was alsoimplicated in oral carcinogenesis by preventing the arrest ofcell cycle progression at the G1 phase through inhibiting theexpression of cyclin-dependent kinase inhibitors Howeverlittle is known about the role of AMIGO2 in the developmentof endometriosis
To investigate whether miR-183 directly targets ITGB1and AMIGO2 dual luciferase reporter assays were carriedout We found that miR-183 negatively regulated the expres-sion of ITBG1 by directly targeting its 31015840UTR but mightnot directly regulate the expression of AMIGO2 We further
detected differential expression of ITGB1 in endometriumwith or without endometriosis by quantitative RT-PCR Thehigher expression of integrin 1205731 in endometrium tissues inendometriosis was confirmed which is consistent with theresult for higher integrin 1205731 expression in ectopic stromalcells than normal endometrial stromal cells Previous studieshave shown that integrin 1205731 mediates cell adherence toextracellular matrix [29] Indeed the rescue experimentindicated that overexpression of ITGB1 blocked the repressiveeffects of miR-183 on endometrial stromal cell invasivenesswhich provided a critical evidence supporting that the lowerexpression of miR-183 may promote ITGB1 expression andcontribute to the development of endometriosis
In conclusion altered miR-183 expression may causederegulation of its target gene ITBG1 affecting the celladhesiveness and invasiveness of endometrial stromal cellswhich in turn lead to pathogenesis of endometriosis Thesefindings revealed a novel pathologic role of miR-183 forendometriosis It would be of great interest to investigate ifthese alterations may serve as biomarkers for the prediction
BioMed Research International 9
andor treatment of endometriosisThe regulation ofmiR-183and the exact mechanism by which miR-183 regulates ITBG1expression remain to be characterized
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This studywas supported by grants from theNational NaturalScience Foundation of China (nos 81100405 and 81471438)the Key Program ofNanjingMedical Science and TechnologyDevelopment Foundation Nanjing Department of Health(no QRX11109)
References
[1] D R Nyholt S-K Low C A Anderson et al ldquoGenome-wide association meta-analysis identifies new endometriosisrisk locirdquo Nature Genetics vol 44 no 12 pp 1355ndash1359 2012
[2] W Filipowicz S N Bhattacharyya and N Sonenberg ldquoMecha-nisms of post-transcriptional regulation by microRNAs are theanswers in sightrdquoNature Reviews Genetics vol 9 no 2 pp 102ndash114 2008
[3] X Santamaria and H Taylor ldquoMicroRNA and gynecologicalreproductive diseasesrdquo Fertility and Sterility vol 101 no 6 pp1545ndash1551 2014
[4] Y Shimono M Zabala R W Cho et al ldquoDownregulation ofmiRNA-200c links breast cancer stem cells with normal stemcellsrdquo Cell vol 138 no 3 pp 592ndash603 2009
[5] M Adammek B Greve N Kassens et al ldquoMicroRNA miR-145 inhibits proliferation invasiveness and stem cell phenotypeof an in vitro endometriosis model by targeting multiplecytoskeletal elements and pluripotency factorsrdquo Fertility andSterility vol 99 no 5 pp 1346e5ndash1355e5 2013
[6] W Abe K Nasu C Nakada Y Kawano M Moriyama andH Narahara ldquoMiR-196b targets c-myc and Bcl-2 expressioninhibits proliferation and induces apoptosis in endometrioticstromal cellsrdquo Human Reproduction vol 28 no 3 pp 750ndash7612013
[7] W B Nothnick A Graham J Holbert et al ldquomiR-451deficiency is associated with altered endometrial fibrinogenalpha chain expression and reduced endometriotic implantestablishment in an experimental mouse modelrdquo PLoS ONEvol 9 no 6 Article ID e100336 2014
[8] X-Y Shi L Gu J Chen X-R Guo and Y-L Shi ldquoDownregu-lation of miR-183 inhibits apoptosis and enhances the invasivepotential of endometrial stromal cells in endometriosisrdquo Inter-national Journal of Molecular Medicine vol 33 no 1 pp 59ndash672014
[9] Y-L Shi X-Z Luo X-Y Zhu K-Q Hua Y Zhu and D-JLi ldquoEffects of combined 17120573-estradiol with TCDD on secretionof chemokine IL-8 and expression of its receptor CXCR1 inendometriotic focus-associated cells in co-culturerdquo HumanReproduction vol 21 no 4 pp 870ndash879 2006
[10] B Salhia N L Tran A Chan et al ldquoThe guanine nucleotideexchange factors Trio Ect2 and Vav3 mediate the invasivebehavior of glioblastomardquo American Journal of Pathology vol173 no 6 pp 1828ndash1838 2008
[11] B Liu L Wang L-L Shen et al ldquoRNAi-mediated inhibitionof presenilin 2 inhibits glioma cell growth and invasion andis involved in the regulation of Nrg1ErbB signalingrdquo Neuro-Oncology vol 14 no 8 pp 994ndash1006 2012
[12] S D Weeraratne V Amani N Teider et al ldquoPleiotropiceffects of miR-183sim96sim182 converge to regulate cell survivalproliferation and migration in medulloblastomardquo Acta Neu-ropathologica vol 123 no 4 pp 539ndash552 2012
[13] M Patel Q Cai D Ding R Salvi Z Hu and B H Hu ldquoThemiR-183Taok1 target pair is implicated in cochlear responsesto acoustic traumardquo PLoS ONE vol 8 no 3 Article ID e584712013
[14] J Zhu Y Feng Z Ke et al ldquoDown-regulation of miR-183promotes migration and invasion of osteosarcoma by targetingEzrinrdquo The American Journal of Pathology vol 180 no 6 pp2440ndash2451 2012
[15] A J Lowery N Miller R M Dwyer and M J Kerin ldquoDysreg-ulated miR-183 inhibits migration in breast cancer cellsrdquo BMCCancer vol 10 article 502 2010
[16] A L Sarver H Li and S Subramanian ldquoMicroRNA miR-183functions as an oncogene by targeting the transcription factorEGR1 and promoting tumor cell migrationrdquo Cancer Researchvol 70 no 23 pp 9570ndash9580 2010
[17] J Li H Fu C Xu et al ldquoMiR-183 inhibits TGF-beta1-inducedapoptosis by downregulation of PDCD4 expression in humanhepatocellular carcinoma cellsrdquoBMCCancer vol 10 article 3542010
[18] C Chen H Xiang Y-L Peng J Peng and S-W JiangldquoMature miR-183 negatively regulated by transcription factorGATA3 promotes 3T3-L1 adipogenesis through inhibition ofthe canonical Wnt120573-catenin signaling pathway by targetingLRP6rdquo Cellular Signalling vol 26 no 6 pp 1155ndash1165 2014
[19] G Li C Luna J Qiu D L Epstein and P Gonzalez ldquoTargetingof integrin beta1 and kinesin 2alpha by microRNA 183rdquo TheJournal of Biological Chemistry vol 285 no 8 pp 5461ndash54712010
[20] A M Schooley N M Andrews H Zhao and C L Addisonldquo1205731 integrin is required for anchorage-independent growth andinvasion of tumor cells in a context dependent mannerrdquo CancerLetters vol 316 no 2 pp 157ndash167 2012
[21] Y Zhang YGongDHu et al ldquoNuclear SIPA1 activates integrin1205731 promoter and promotes invasion of breast cancer cellsrdquoOncogene vol 34 no 11 pp 1451ndash1462 2015
[22] Y B Aznaurova M B Zhumataev T K Roberts A M Aliperand A A Zhavoronkov ldquoMolecular aspects of developmentand regulation of endometriosisrdquo Reproductive Biology andEndocrinology vol 12 article 50 2014
[23] M Adachi K Nasu A Tsuno A Yuge Y Kawano and HNarahara ldquoAttachment to extracellular matrices is enhancedin human endometriotic stromal cells a possible mechanismunderlying the pathogenesis of endometriosisrdquo European Jour-nal of Obstetrics Gynecology and Reproductive Biology vol 155no 1 pp 85ndash88 2011
[24] J Lee S K Banu R C Burghardt A Starzinski-Powitz andJ A Arosh ldquoSelective inhibition of prostaglandin E2 receptorsEP2 and EP4 inhibits adhesion of human endometriotic epithe-lial and stromal cells through suppression of integrin-mediatedmechanismsrdquo Biology of Reproduction vol 88 no 3 article 772013
[25] A Laeremans J NysW Luyten R DrsquoHoogeM Paulussen andL Arckens ldquoAMIGO2 mRNA expression in hippocampal CA2
10 BioMed Research International
and CA3ardquo Brain Structure and Function vol 218 no 1 pp 123ndash130 2013
[26] T Ono N Sekino-Suzuki Y Kikkawa H Yonekawa andS Kawashima ldquoAlivin 1 a novel neuronal activity-dependentgene inhibits apoptosis and promotes survival of cerebellargranule neuronsrdquo Journal of Neuroscience vol 23 no 13 pp5887ndash5896 2003
[27] J Kuja-Panula M Kiiltomaki T Yamashiro A Rouhiainenand H Rauvala ldquoAMIGO a transmembrane protein implicatedin axon tract development defines a novel protein family withleucine-rich repeatsrdquoThe Journal of Cell Biology vol 160 no 6pp 963ndash973 2003
[28] K E Rabenau J M OrsquoToole R Bassi et al ldquoDEGAAMIGO-2 a leucine-rich repeat family member differentially expressedin human gastric adenocarcinoma effects on ploidy chromo-somal stability cell adhesionmigration and tumorigenicityrdquoOncogene vol 23 no 29 pp 5056ndash5067 2004
[29] J A Garcia-Velasco and A Arici ldquoInterleukin-8 expression inendometrial stromal cells is regulated by integrin-dependentcell adhesionrdquo Molecular Human Reproduction vol 5 no 12pp 1135ndash1140 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 BioMed Research International
(a)
0
05
1
15
2
25
NC MiR-183 MiR-183ITGB1
Inva
sive r
ate (
OO
D57
0)
lowast
(b)
Figure 5 Overexpression of ITGB1 rescued the repressive effects of miR-183 on endometrial stromal cells leading to elevated invasiveabilities in transwell assays Endometrial stromal cells were infected with miR-183-lentivirus miR-183ITGB1-lentivirus and GFP-lentivirusin upper wells After 24 hours the number of cells that invaded through Matrigel was counted in at least 10 fields per well (a) Representativephotographs show that miR-183 inhibited the invasiveness of endometrial stromal cells whereas ITGB1 partially rescued the repressive effectsof miR-183 (b) Cell counting results indicate that ITGB1 overexpression rescued the repressive effects of miR-183 lowast119875 lt 005 when comparedto the negative control Error bars represent plusmn SEM
for the significance of integrins in the adhesion function ofendometrial stromal cells
AMIGO2 (ALI1 DEGA) was first reported to be prefer-entially expressed in the central nervous system Sequenceanalyses revealed that the protein contains seven leucine-richrepeats one IgC2-like loop and a transmembrane domainand displays homology to Kek and Trk families [25ndash27] Inthe gastric adenocarcinoma cell line AMIGO2 was reportedto be involved in cell adhesion extracellular matrix andbasement membrane formation [28] AMIGO2 was alsoimplicated in oral carcinogenesis by preventing the arrest ofcell cycle progression at the G1 phase through inhibiting theexpression of cyclin-dependent kinase inhibitors Howeverlittle is known about the role of AMIGO2 in the developmentof endometriosis
To investigate whether miR-183 directly targets ITGB1and AMIGO2 dual luciferase reporter assays were carriedout We found that miR-183 negatively regulated the expres-sion of ITBG1 by directly targeting its 31015840UTR but mightnot directly regulate the expression of AMIGO2 We further
detected differential expression of ITGB1 in endometriumwith or without endometriosis by quantitative RT-PCR Thehigher expression of integrin 1205731 in endometrium tissues inendometriosis was confirmed which is consistent with theresult for higher integrin 1205731 expression in ectopic stromalcells than normal endometrial stromal cells Previous studieshave shown that integrin 1205731 mediates cell adherence toextracellular matrix [29] Indeed the rescue experimentindicated that overexpression of ITGB1 blocked the repressiveeffects of miR-183 on endometrial stromal cell invasivenesswhich provided a critical evidence supporting that the lowerexpression of miR-183 may promote ITGB1 expression andcontribute to the development of endometriosis
In conclusion altered miR-183 expression may causederegulation of its target gene ITBG1 affecting the celladhesiveness and invasiveness of endometrial stromal cellswhich in turn lead to pathogenesis of endometriosis Thesefindings revealed a novel pathologic role of miR-183 forendometriosis It would be of great interest to investigate ifthese alterations may serve as biomarkers for the prediction
BioMed Research International 9
andor treatment of endometriosisThe regulation ofmiR-183and the exact mechanism by which miR-183 regulates ITBG1expression remain to be characterized
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This studywas supported by grants from theNational NaturalScience Foundation of China (nos 81100405 and 81471438)the Key Program ofNanjingMedical Science and TechnologyDevelopment Foundation Nanjing Department of Health(no QRX11109)
References
[1] D R Nyholt S-K Low C A Anderson et al ldquoGenome-wide association meta-analysis identifies new endometriosisrisk locirdquo Nature Genetics vol 44 no 12 pp 1355ndash1359 2012
[2] W Filipowicz S N Bhattacharyya and N Sonenberg ldquoMecha-nisms of post-transcriptional regulation by microRNAs are theanswers in sightrdquoNature Reviews Genetics vol 9 no 2 pp 102ndash114 2008
[3] X Santamaria and H Taylor ldquoMicroRNA and gynecologicalreproductive diseasesrdquo Fertility and Sterility vol 101 no 6 pp1545ndash1551 2014
[4] Y Shimono M Zabala R W Cho et al ldquoDownregulation ofmiRNA-200c links breast cancer stem cells with normal stemcellsrdquo Cell vol 138 no 3 pp 592ndash603 2009
[5] M Adammek B Greve N Kassens et al ldquoMicroRNA miR-145 inhibits proliferation invasiveness and stem cell phenotypeof an in vitro endometriosis model by targeting multiplecytoskeletal elements and pluripotency factorsrdquo Fertility andSterility vol 99 no 5 pp 1346e5ndash1355e5 2013
[6] W Abe K Nasu C Nakada Y Kawano M Moriyama andH Narahara ldquoMiR-196b targets c-myc and Bcl-2 expressioninhibits proliferation and induces apoptosis in endometrioticstromal cellsrdquo Human Reproduction vol 28 no 3 pp 750ndash7612013
[7] W B Nothnick A Graham J Holbert et al ldquomiR-451deficiency is associated with altered endometrial fibrinogenalpha chain expression and reduced endometriotic implantestablishment in an experimental mouse modelrdquo PLoS ONEvol 9 no 6 Article ID e100336 2014
[8] X-Y Shi L Gu J Chen X-R Guo and Y-L Shi ldquoDownregu-lation of miR-183 inhibits apoptosis and enhances the invasivepotential of endometrial stromal cells in endometriosisrdquo Inter-national Journal of Molecular Medicine vol 33 no 1 pp 59ndash672014
[9] Y-L Shi X-Z Luo X-Y Zhu K-Q Hua Y Zhu and D-JLi ldquoEffects of combined 17120573-estradiol with TCDD on secretionof chemokine IL-8 and expression of its receptor CXCR1 inendometriotic focus-associated cells in co-culturerdquo HumanReproduction vol 21 no 4 pp 870ndash879 2006
[10] B Salhia N L Tran A Chan et al ldquoThe guanine nucleotideexchange factors Trio Ect2 and Vav3 mediate the invasivebehavior of glioblastomardquo American Journal of Pathology vol173 no 6 pp 1828ndash1838 2008
[11] B Liu L Wang L-L Shen et al ldquoRNAi-mediated inhibitionof presenilin 2 inhibits glioma cell growth and invasion andis involved in the regulation of Nrg1ErbB signalingrdquo Neuro-Oncology vol 14 no 8 pp 994ndash1006 2012
[12] S D Weeraratne V Amani N Teider et al ldquoPleiotropiceffects of miR-183sim96sim182 converge to regulate cell survivalproliferation and migration in medulloblastomardquo Acta Neu-ropathologica vol 123 no 4 pp 539ndash552 2012
[13] M Patel Q Cai D Ding R Salvi Z Hu and B H Hu ldquoThemiR-183Taok1 target pair is implicated in cochlear responsesto acoustic traumardquo PLoS ONE vol 8 no 3 Article ID e584712013
[14] J Zhu Y Feng Z Ke et al ldquoDown-regulation of miR-183promotes migration and invasion of osteosarcoma by targetingEzrinrdquo The American Journal of Pathology vol 180 no 6 pp2440ndash2451 2012
[15] A J Lowery N Miller R M Dwyer and M J Kerin ldquoDysreg-ulated miR-183 inhibits migration in breast cancer cellsrdquo BMCCancer vol 10 article 502 2010
[16] A L Sarver H Li and S Subramanian ldquoMicroRNA miR-183functions as an oncogene by targeting the transcription factorEGR1 and promoting tumor cell migrationrdquo Cancer Researchvol 70 no 23 pp 9570ndash9580 2010
[17] J Li H Fu C Xu et al ldquoMiR-183 inhibits TGF-beta1-inducedapoptosis by downregulation of PDCD4 expression in humanhepatocellular carcinoma cellsrdquoBMCCancer vol 10 article 3542010
[18] C Chen H Xiang Y-L Peng J Peng and S-W JiangldquoMature miR-183 negatively regulated by transcription factorGATA3 promotes 3T3-L1 adipogenesis through inhibition ofthe canonical Wnt120573-catenin signaling pathway by targetingLRP6rdquo Cellular Signalling vol 26 no 6 pp 1155ndash1165 2014
[19] G Li C Luna J Qiu D L Epstein and P Gonzalez ldquoTargetingof integrin beta1 and kinesin 2alpha by microRNA 183rdquo TheJournal of Biological Chemistry vol 285 no 8 pp 5461ndash54712010
[20] A M Schooley N M Andrews H Zhao and C L Addisonldquo1205731 integrin is required for anchorage-independent growth andinvasion of tumor cells in a context dependent mannerrdquo CancerLetters vol 316 no 2 pp 157ndash167 2012
[21] Y Zhang YGongDHu et al ldquoNuclear SIPA1 activates integrin1205731 promoter and promotes invasion of breast cancer cellsrdquoOncogene vol 34 no 11 pp 1451ndash1462 2015
[22] Y B Aznaurova M B Zhumataev T K Roberts A M Aliperand A A Zhavoronkov ldquoMolecular aspects of developmentand regulation of endometriosisrdquo Reproductive Biology andEndocrinology vol 12 article 50 2014
[23] M Adachi K Nasu A Tsuno A Yuge Y Kawano and HNarahara ldquoAttachment to extracellular matrices is enhancedin human endometriotic stromal cells a possible mechanismunderlying the pathogenesis of endometriosisrdquo European Jour-nal of Obstetrics Gynecology and Reproductive Biology vol 155no 1 pp 85ndash88 2011
[24] J Lee S K Banu R C Burghardt A Starzinski-Powitz andJ A Arosh ldquoSelective inhibition of prostaglandin E2 receptorsEP2 and EP4 inhibits adhesion of human endometriotic epithe-lial and stromal cells through suppression of integrin-mediatedmechanismsrdquo Biology of Reproduction vol 88 no 3 article 772013
[25] A Laeremans J NysW Luyten R DrsquoHoogeM Paulussen andL Arckens ldquoAMIGO2 mRNA expression in hippocampal CA2
10 BioMed Research International
and CA3ardquo Brain Structure and Function vol 218 no 1 pp 123ndash130 2013
[26] T Ono N Sekino-Suzuki Y Kikkawa H Yonekawa andS Kawashima ldquoAlivin 1 a novel neuronal activity-dependentgene inhibits apoptosis and promotes survival of cerebellargranule neuronsrdquo Journal of Neuroscience vol 23 no 13 pp5887ndash5896 2003
[27] J Kuja-Panula M Kiiltomaki T Yamashiro A Rouhiainenand H Rauvala ldquoAMIGO a transmembrane protein implicatedin axon tract development defines a novel protein family withleucine-rich repeatsrdquoThe Journal of Cell Biology vol 160 no 6pp 963ndash973 2003
[28] K E Rabenau J M OrsquoToole R Bassi et al ldquoDEGAAMIGO-2 a leucine-rich repeat family member differentially expressedin human gastric adenocarcinoma effects on ploidy chromo-somal stability cell adhesionmigration and tumorigenicityrdquoOncogene vol 23 no 29 pp 5056ndash5067 2004
[29] J A Garcia-Velasco and A Arici ldquoInterleukin-8 expression inendometrial stromal cells is regulated by integrin-dependentcell adhesionrdquo Molecular Human Reproduction vol 5 no 12pp 1135ndash1140 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 9
andor treatment of endometriosisThe regulation ofmiR-183and the exact mechanism by which miR-183 regulates ITBG1expression remain to be characterized
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
This studywas supported by grants from theNational NaturalScience Foundation of China (nos 81100405 and 81471438)the Key Program ofNanjingMedical Science and TechnologyDevelopment Foundation Nanjing Department of Health(no QRX11109)
References
[1] D R Nyholt S-K Low C A Anderson et al ldquoGenome-wide association meta-analysis identifies new endometriosisrisk locirdquo Nature Genetics vol 44 no 12 pp 1355ndash1359 2012
[2] W Filipowicz S N Bhattacharyya and N Sonenberg ldquoMecha-nisms of post-transcriptional regulation by microRNAs are theanswers in sightrdquoNature Reviews Genetics vol 9 no 2 pp 102ndash114 2008
[3] X Santamaria and H Taylor ldquoMicroRNA and gynecologicalreproductive diseasesrdquo Fertility and Sterility vol 101 no 6 pp1545ndash1551 2014
[4] Y Shimono M Zabala R W Cho et al ldquoDownregulation ofmiRNA-200c links breast cancer stem cells with normal stemcellsrdquo Cell vol 138 no 3 pp 592ndash603 2009
[5] M Adammek B Greve N Kassens et al ldquoMicroRNA miR-145 inhibits proliferation invasiveness and stem cell phenotypeof an in vitro endometriosis model by targeting multiplecytoskeletal elements and pluripotency factorsrdquo Fertility andSterility vol 99 no 5 pp 1346e5ndash1355e5 2013
[6] W Abe K Nasu C Nakada Y Kawano M Moriyama andH Narahara ldquoMiR-196b targets c-myc and Bcl-2 expressioninhibits proliferation and induces apoptosis in endometrioticstromal cellsrdquo Human Reproduction vol 28 no 3 pp 750ndash7612013
[7] W B Nothnick A Graham J Holbert et al ldquomiR-451deficiency is associated with altered endometrial fibrinogenalpha chain expression and reduced endometriotic implantestablishment in an experimental mouse modelrdquo PLoS ONEvol 9 no 6 Article ID e100336 2014
[8] X-Y Shi L Gu J Chen X-R Guo and Y-L Shi ldquoDownregu-lation of miR-183 inhibits apoptosis and enhances the invasivepotential of endometrial stromal cells in endometriosisrdquo Inter-national Journal of Molecular Medicine vol 33 no 1 pp 59ndash672014
[9] Y-L Shi X-Z Luo X-Y Zhu K-Q Hua Y Zhu and D-JLi ldquoEffects of combined 17120573-estradiol with TCDD on secretionof chemokine IL-8 and expression of its receptor CXCR1 inendometriotic focus-associated cells in co-culturerdquo HumanReproduction vol 21 no 4 pp 870ndash879 2006
[10] B Salhia N L Tran A Chan et al ldquoThe guanine nucleotideexchange factors Trio Ect2 and Vav3 mediate the invasivebehavior of glioblastomardquo American Journal of Pathology vol173 no 6 pp 1828ndash1838 2008
[11] B Liu L Wang L-L Shen et al ldquoRNAi-mediated inhibitionof presenilin 2 inhibits glioma cell growth and invasion andis involved in the regulation of Nrg1ErbB signalingrdquo Neuro-Oncology vol 14 no 8 pp 994ndash1006 2012
[12] S D Weeraratne V Amani N Teider et al ldquoPleiotropiceffects of miR-183sim96sim182 converge to regulate cell survivalproliferation and migration in medulloblastomardquo Acta Neu-ropathologica vol 123 no 4 pp 539ndash552 2012
[13] M Patel Q Cai D Ding R Salvi Z Hu and B H Hu ldquoThemiR-183Taok1 target pair is implicated in cochlear responsesto acoustic traumardquo PLoS ONE vol 8 no 3 Article ID e584712013
[14] J Zhu Y Feng Z Ke et al ldquoDown-regulation of miR-183promotes migration and invasion of osteosarcoma by targetingEzrinrdquo The American Journal of Pathology vol 180 no 6 pp2440ndash2451 2012
[15] A J Lowery N Miller R M Dwyer and M J Kerin ldquoDysreg-ulated miR-183 inhibits migration in breast cancer cellsrdquo BMCCancer vol 10 article 502 2010
[16] A L Sarver H Li and S Subramanian ldquoMicroRNA miR-183functions as an oncogene by targeting the transcription factorEGR1 and promoting tumor cell migrationrdquo Cancer Researchvol 70 no 23 pp 9570ndash9580 2010
[17] J Li H Fu C Xu et al ldquoMiR-183 inhibits TGF-beta1-inducedapoptosis by downregulation of PDCD4 expression in humanhepatocellular carcinoma cellsrdquoBMCCancer vol 10 article 3542010
[18] C Chen H Xiang Y-L Peng J Peng and S-W JiangldquoMature miR-183 negatively regulated by transcription factorGATA3 promotes 3T3-L1 adipogenesis through inhibition ofthe canonical Wnt120573-catenin signaling pathway by targetingLRP6rdquo Cellular Signalling vol 26 no 6 pp 1155ndash1165 2014
[19] G Li C Luna J Qiu D L Epstein and P Gonzalez ldquoTargetingof integrin beta1 and kinesin 2alpha by microRNA 183rdquo TheJournal of Biological Chemistry vol 285 no 8 pp 5461ndash54712010
[20] A M Schooley N M Andrews H Zhao and C L Addisonldquo1205731 integrin is required for anchorage-independent growth andinvasion of tumor cells in a context dependent mannerrdquo CancerLetters vol 316 no 2 pp 157ndash167 2012
[21] Y Zhang YGongDHu et al ldquoNuclear SIPA1 activates integrin1205731 promoter and promotes invasion of breast cancer cellsrdquoOncogene vol 34 no 11 pp 1451ndash1462 2015
[22] Y B Aznaurova M B Zhumataev T K Roberts A M Aliperand A A Zhavoronkov ldquoMolecular aspects of developmentand regulation of endometriosisrdquo Reproductive Biology andEndocrinology vol 12 article 50 2014
[23] M Adachi K Nasu A Tsuno A Yuge Y Kawano and HNarahara ldquoAttachment to extracellular matrices is enhancedin human endometriotic stromal cells a possible mechanismunderlying the pathogenesis of endometriosisrdquo European Jour-nal of Obstetrics Gynecology and Reproductive Biology vol 155no 1 pp 85ndash88 2011
[24] J Lee S K Banu R C Burghardt A Starzinski-Powitz andJ A Arosh ldquoSelective inhibition of prostaglandin E2 receptorsEP2 and EP4 inhibits adhesion of human endometriotic epithe-lial and stromal cells through suppression of integrin-mediatedmechanismsrdquo Biology of Reproduction vol 88 no 3 article 772013
[25] A Laeremans J NysW Luyten R DrsquoHoogeM Paulussen andL Arckens ldquoAMIGO2 mRNA expression in hippocampal CA2
10 BioMed Research International
and CA3ardquo Brain Structure and Function vol 218 no 1 pp 123ndash130 2013
[26] T Ono N Sekino-Suzuki Y Kikkawa H Yonekawa andS Kawashima ldquoAlivin 1 a novel neuronal activity-dependentgene inhibits apoptosis and promotes survival of cerebellargranule neuronsrdquo Journal of Neuroscience vol 23 no 13 pp5887ndash5896 2003
[27] J Kuja-Panula M Kiiltomaki T Yamashiro A Rouhiainenand H Rauvala ldquoAMIGO a transmembrane protein implicatedin axon tract development defines a novel protein family withleucine-rich repeatsrdquoThe Journal of Cell Biology vol 160 no 6pp 963ndash973 2003
[28] K E Rabenau J M OrsquoToole R Bassi et al ldquoDEGAAMIGO-2 a leucine-rich repeat family member differentially expressedin human gastric adenocarcinoma effects on ploidy chromo-somal stability cell adhesionmigration and tumorigenicityrdquoOncogene vol 23 no 29 pp 5056ndash5067 2004
[29] J A Garcia-Velasco and A Arici ldquoInterleukin-8 expression inendometrial stromal cells is regulated by integrin-dependentcell adhesionrdquo Molecular Human Reproduction vol 5 no 12pp 1135ndash1140 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 BioMed Research International
and CA3ardquo Brain Structure and Function vol 218 no 1 pp 123ndash130 2013
[26] T Ono N Sekino-Suzuki Y Kikkawa H Yonekawa andS Kawashima ldquoAlivin 1 a novel neuronal activity-dependentgene inhibits apoptosis and promotes survival of cerebellargranule neuronsrdquo Journal of Neuroscience vol 23 no 13 pp5887ndash5896 2003
[27] J Kuja-Panula M Kiiltomaki T Yamashiro A Rouhiainenand H Rauvala ldquoAMIGO a transmembrane protein implicatedin axon tract development defines a novel protein family withleucine-rich repeatsrdquoThe Journal of Cell Biology vol 160 no 6pp 963ndash973 2003
[28] K E Rabenau J M OrsquoToole R Bassi et al ldquoDEGAAMIGO-2 a leucine-rich repeat family member differentially expressedin human gastric adenocarcinoma effects on ploidy chromo-somal stability cell adhesionmigration and tumorigenicityrdquoOncogene vol 23 no 29 pp 5056ndash5067 2004
[29] J A Garcia-Velasco and A Arici ldquoInterleukin-8 expression inendometrial stromal cells is regulated by integrin-dependentcell adhesionrdquo Molecular Human Reproduction vol 5 no 12pp 1135ndash1140 1999
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom


















