
Research Article Lysophosphatidic Acid Inhibits Apoptosis Induced...
13
Research Article Lysophosphatidic Acid Inhibits Apoptosis Induced by Cisplatin in Cervical Cancer Cells Yanxia Sui, 1,2 Ya Yang, 3 Ji Wang, 4 Yi Li, 5 Hongbing Ma, 6 Hui Cai, 7 Xiaoping Liu, 1 Yong Zhang, 8 Shufeng Wang, 9 Zongfang Li, 6 Xiaozhi Zhang, 1 Jiansheng Wang, 9 Rui Liu, 1 Yanli Yan, 3 Chaofan Xue, 3 Xiaowei Shi, 3 Li Tan, 3 and Juan Ren 1 1 Department of Radiotherapy Oncology, First Affiliated Hospital of Xi’an Jiaotong University, Xi’an, Shaanxi 710061, China 2 Pathology Department, First Affiliated Hospital of Xi’an Jiaotong University, Xi’an, Shaanxi 710061, China 3 Medical School, Xi’an Jiaotong University, Xi’an, Shaanxi 710061, China 4 ICU, China Meitan General Hospital, 29 Xi Ba He Nan Li, Chaoyang District, Beijing 100028, China 5 Department of Chemotherapy Oncology, First Affiliated Hospital of Xi’an Jiaotong University, Xi’an, Shaanxi 710061, China 6 Department of Oncology, Second Affiliated Hospital of Xi’an Jiaotong University, Xi’an, Shaanxi 710004, China 7 Department of Anesthesia, Xi’an Jiaotong University, Xi’an, Shaanxi 710061, China 8 Department of Orthopedics, Xi’an Children Hospital, Xi’an, Shaanxi 710000, China 9 Department of Surgery, First Affiliated Hospital of Xi’an Jiaotong University, Xi’an, Shaanxi 710061, China Correspondence should be addressed to Juan Ren; [email protected] Received 19 February 2015; Revised 17 April 2015; Accepted 28 April 2015 Academic Editor: Brian Brost Copyright © 2015 Yanxia Sui et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Cervical cancer is the second most common cause of cancer death in women worldwide. Lysophosphatidic acid (LPA) level has been found significantly increased in the serum of patients with ovarian, cervical, and colon cancers. LPA level in cervical cancer patients is significantly higher than in healthy controls. LPA receptors were found highly expressed in cervical cancer cells, suggesting LPA may play a role in the development of cervical cancer. e aim of this study is to investigate the effect of LPA on the apoptosis induced by cisplatin (DDP) in cervical cancer cell line and the underlying changes in signaling pathways. Our study found that cisplatin induced apoptosis of Hela cell through inhibiting expression of Bcl-2, upregulating the expression of Bax, Fas-L, and the enzyme activity of caspase-3 ( < 0.05); LPA significantly provided protection against the apoptosis induced by cisplatin by inhibiting the above alterations in apoptotic factor caused by cisplatin ( < 0.05). Moreover, PI3K/AKT pathway was found to be important for the LPA antiapoptosis effect, and administration of PI3K/AKT partially reversed the LPA-mediated protection against cisplatin- induced apoptosis ( < 0.05). ese findings have shed new lights on the LPA bioactivity in cervical cancer cells and pointed to a possible sensitization scheme through combined administration of PI3K inhibitor and cisplatin for better treatment of cervical cancer patients, especially those with elevated LPA levels. 1. Introduction Cervical cancer is the second most common type, and the second most cause of deaths, of malignancy, in females world- wide. An estimated 500,000 new cases of cervical cancer are diagnosed, leading to 280,000 deaths, each year worldwide. e highest incidences of cervical cancers occur in devel- oping countries. While surgery and chemoradiotherapy can cure 80%–95% of women with early stage cervical cancer, the recurrence and metastasis events are oſten associated with poor prognosis. In addition to the efforts for more effective prevention, new diagnosis and treatment modalities are urgently needed for better management of this life-threaten- ing disease. High levels of lysophosphatidic acid (LPA) were firstly found in the ascites of ovarian cancers patients [1, 2]. LPA is known as an “ovarian cancer activating factor” to exert a growth factor-like effect through binding to 4 specific G protein-coupled receptors (LPA1-4). e biological activities of LPA in ovarian cancer have been studied for many years. Hindawi Publishing Corporation BioMed Research International Volume 2015, Article ID 598386, 12 pages http://dx.doi.org/10.1155/2015/598386
Transcript of Research Article Lysophosphatidic Acid Inhibits Apoptosis Induced...

Research ArticleLysophosphatidic Acid Inhibits Apoptosis Induced byCisplatin in Cervical Cancer Cells
Yanxia Sui12 Ya Yang3 Ji Wang4 Yi Li5 Hongbing Ma6 Hui Cai7 Xiaoping Liu1
Yong Zhang8 Shufeng Wang9 Zongfang Li6 Xiaozhi Zhang1 Jiansheng Wang9 Rui Liu1
Yanli Yan3 Chaofan Xue3 Xiaowei Shi3 Li Tan3 and Juan Ren1
1Department of Radiotherapy Oncology First Affiliated Hospital of Xirsquoan Jiaotong University Xirsquoan Shaanxi 710061 China2Pathology Department First Affiliated Hospital of Xirsquoan Jiaotong University Xirsquoan Shaanxi 710061 China3Medical School Xirsquoan Jiaotong University Xirsquoan Shaanxi 710061 China4ICU China Meitan General Hospital 29 Xi Ba He Nan Li Chaoyang District Beijing 100028 China5Department of Chemotherapy Oncology First Affiliated Hospital of Xirsquoan Jiaotong University Xirsquoan Shaanxi 710061 China6Department of Oncology Second Affiliated Hospital of Xirsquoan Jiaotong University Xirsquoan Shaanxi 710004 China7Department of Anesthesia Xirsquoan Jiaotong University Xirsquoan Shaanxi 710061 China8Department of Orthopedics Xirsquoan Children Hospital Xirsquoan Shaanxi 710000 China9Department of Surgery First Affiliated Hospital of Xirsquoan Jiaotong University Xirsquoan Shaanxi 710061 China
Correspondence should be addressed to Juan Ren 869491533qqcom
Received 19 February 2015 Revised 17 April 2015 Accepted 28 April 2015
Academic Editor Brian Brost
Copyright copy 2015 Yanxia Sui et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Cervical cancer is the secondmost common cause of cancer death inwomenworldwide Lysophosphatidic acid (LPA) level has beenfound significantly increased in the serum of patients with ovarian cervical and colon cancers LPA level in cervical cancer patientsis significantly higher than in healthy controls LPA receptors were found highly expressed in cervical cancer cells suggesting LPAmay play a role in the development of cervical cancerThe aimof this study is to investigate the effect of LPAon the apoptosis inducedby cisplatin (DDP) in cervical cancer cell line and the underlying changes in signaling pathways Our study found that cisplatininduced apoptosis of Hela cell through inhibiting expression of Bcl-2 upregulating the expression of Bax Fas-L and the enzymeactivity of caspase-3 (119901 lt 005) LPA significantly provided protection against the apoptosis induced by cisplatin by inhibiting theabove alterations in apoptotic factor caused by cisplatin (119901 lt 005) Moreover PI3KAKT pathway was found to be important forthe LPA antiapoptosis effect and administration of PI3KAKT partially reversed the LPA-mediated protection against cisplatin-induced apoptosis (119901 lt 005) These findings have shed new lights on the LPA bioactivity in cervical cancer cells and pointed toa possible sensitization scheme through combined administration of PI3K inhibitor and cisplatin for better treatment of cervicalcancer patients especially those with elevated LPA levels
1 Introduction
Cervical cancer is the second most common type and thesecondmost cause of deaths ofmalignancy in femalesworld-wide An estimated 500000 new cases of cervical cancer arediagnosed leading to 280000 deaths each year worldwideThe highest incidences of cervical cancers occur in devel-oping countries While surgery and chemoradiotherapy cancure 80ndash95 of women with early stage cervical cancer therecurrence and metastasis events are often associated with
poor prognosis In addition to the efforts for more effectiveprevention new diagnosis and treatment modalities areurgently needed for better management of this life-threaten-ing disease
High levels of lysophosphatidic acid (LPA) were firstlyfound in the ascites of ovarian cancers patients [1 2] LPAis known as an ldquoovarian cancer activating factorrdquo to exerta growth factor-like effect through binding to 4 specific Gprotein-coupled receptors (LPA1-4) The biological activitiesof LPA in ovarian cancer have been studied for many years
Hindawi Publishing CorporationBioMed Research InternationalVolume 2015 Article ID 598386 12 pageshttpdxdoiorg1011552015598386
2 BioMed Research International
Increased level of lysophosphatidic acid is also found inpatients with acute myocardial infarction LPA has beenimplicated in the development of the cardiovascular systemassisting in its progression to a fully functional state [3 4] LPAis a bioactive glycerophospholipid generated and releasedby platelets macrophages epithelial cells and tumor cellsLPAmodulates a broad range of cellular responses includingalterations of cell proliferation protection against apoptosismodulation of chemotaxis and transcellular migration [56] thereby affecting the survival of ovarian cancer cellsmacrophages fibroblasts and neonatal cardiacmyocytesThesignificant role of LPA in triggering these cellular responseshas implicated LPA in tumor progression
It has also been reported that LPA is increased in theplasma of cervical cancer patients [2] Xu et al found thatstage I and stage IV cervical cancer patients had significantlyhigher plasma LPA levels than normal controls ElevatedLPA levels were detected in all the 6 cervical cancer patientsexamined [2] In addition therewas an increased ratio of totalLPAlysophosphatidylinositol (LPI) [7] Similarly Shen et alreported that the ratio of unsaturated LPALPI subspecieswassignificantly higher in patients with cervical cancer than inhealthy controls [7] The significantly increased LPA in theplasma of patients of cervical cancer points to its possiblerole(s) for the development of this malignancy Indeed LPAreceptorswere also found to be extensively expressed in cervi-cal cell lines including Hela CaSki and Siha [8ndash11] Previousstudies from this group confirmed a high expression level ofLPA receptors especially the LPA receptor 2 inHela cells [12]providing a basis for using this cell line as a study model toinvestigate the LPA bioactivity and the underlying pathways
Cisplatin (DDP) has been used as the first line chemother-apy drug for adjuvant treatment of cervical cancer patientsCisplatin-induced DNA damage activates multiple signalingpathways leading to cell apoptosis [13ndash15] DNA damagecaused by cisplatin induces the phosphorylation and stabi-lization of p53 [16] p53 promotes cisplatin-induced apop-tosis by antagonizing the antiapoptotic effect of Bcl-xL [17]Phosphatidylinositol 3-kinaseAKT pathway is also involvedin apoptosis regulation Yan et al found that suppression ofPI3KAKT pathway caused apoptosis in the HepG2 humanhepatoma cell line [18] On the other hand Wang et alfound that LPA protects bone marrow-derived mesenchymalstem cells (BMSCs) against hypoxia- or serum deprivation-induced apoptosis [19] LPA rescues H
2O2-induced apoptosis
by activating ERK12 and PI3KAKT pathways in mesenchy-mal stem cells [19] However LPA effect on the apoptosis incervical cancer cells and the potential mechanism remainsunclear
In this study we investigate how LPA-triggered cellresponsesmay affect the cell apoptosis induced by cisplatin ina cervical cancer cell lineWe characterized the effects of LPAon cell apoptotic factors including Bcl-2 Bax and caspase-3in Hela cells treated with cisplatin The influence of LPA onthe upstream pathway PI3KAKT was also determined Thefindings provide insights on the general bioactivity as well asthe chemotherapeutic concerns of LPA produced by cervicalcancer cells
2 Materials and Methods
21 Materials 1-Oleoyl LPA (18 1 LPA) was purchased fromAvanti Polar Lipids (Birmingham AL USA) Inhibitor ofphosphoinositide 3-kinase (PI3K) LY290042 and inhibitorof mitogen-activated protein kinase (MAPK) PD98059were from Cell Signaling (Beverly MA USA) Rho kinaseinhibitor Y-27632 was from Biomol (Beverly MA USA)Hela cells were cultured in Dulbeccorsquos modified Eaglersquosmedium (DMEM) supplemented with 100mLL fetal bovineserum (FBS) streptomycin (100mgL) and penicillin(100KUL) at 37∘C in 50mLL CO
2incubator Cells were
serum starved for 12 hours before LPA treatment
22 Cell Proliferation Assay 3-(45-Dimethylthiazol-2-yl)-25-diphenyltetrazolium bromide (MTT) colorimetry assaywas employed to measure cell proliferation Hela cells (2 times103well) were seeded at 96-well plate After being starvedfor 12 hours cells were fed with DMEM containing LPAsupplemented with 1 gL bovine serum albumin Following24 48 72 and 96 hours of culture 20120583L of MTT solution(5 gL) was added to each well Four hours later the mediumwas removed and 150 120583L of dimethyl sulfoxide (DMSO) wasadded to each well Absorbance was measured at 490 nm ona Microplate Reader (EXL800) Each assay was performed inquintuplicate
23 Annexin V Staining and Flow Cytometry Assay Cellswere starved in serum-free DMEM for 12 hours and treatedwith different concentrations of LPA For kinase inhibitorexperiments LY294002 (50120583molL) PD98059 (10 120583molL)and Y-27632 (10 120583molL) were applied to cells 30minbefore addition of LPA Cells were resuspended in bindingbuffer (10mmolL HEPESNaOH pH74 140mmolL NaCl25mmolL CaCl
2) and stained with 5120583L of annexin-FITC
and 5mgL propidium iodide (PI) Following washing cellswere analyzed with flow cytometry (FACS Calibur cytometerBD Biosciences USA) and CellQuest (BD Biosciences) forquantification of cell apoptosis The experiment was per-formed in triplicate and repeated twice
24 Caspase-3 Activity Assay Activity of caspase-3 wasdetected using the CaspACE Assay System (Promega USA)Briefly cell lysates from treated and untreated control cellswere prepared Cell lysates were centrifuged at 15000timesg for20min at 4∘C and the supernatant was collected The assaywas performed in a total volume of 100 120583L in 96-well platesCell extracts with an equal amount of protein (50ndash100120583g oftotal protein) were mixed with specific colorimetric substrate(DEVD-pNA) for caspase-3 The mixture was incubated at37∘C for 4 h according to the manufacturerrsquos protocol Theabsorbance was measured at a wave length of 405 nm
25 Western Blotting Antibodies against AKT (1 200 dilu-tion times) and phosphorylated-AKT (1 200 dilution times)were purchased from Cell Signaling Transduction (USA)Antibodies against Fas (1 200 dilution times) Bcl-2 (1 200dilution times) and Bax (1 200 dilution times) were pur-chased from Santa Cruz After treatment cells were rinsed
BioMed Research International 3
800
600
400
200
000
0 24 48 72 96
(hours)
Cel
l num
ber (times10
4)
Control011
520500
(a)
800
600
400
200
000
0 24 48 72 96
(hours)
Cel
l num
ber (times10
4)
ControlLPA + PD98059LPA + LY294002
LPA + Y-27632LPA
(b)
Figure 1 LPA effects on cell proliferation of Hela cells (a) Hela cells were treated with different doses and time of LPA MTT essay wasperformed LPA significantly stimulated the proliferation of Hela cells in a dose-dependent and time-dependent manner (b) Effect ofinhibitors on LPA stimulation of cell proliferation Hela cells were treatedwith LPA at 20120583Mplus different inhibitors for different time periodsbefore MTT essay PD98059 (MAPK inhibitor at 10 120583M) PD98059 (MAPK inhibitor at 10 120583M) and Y-27632 (Rho inhibitor at 10 120583M) wereapplied respectively MAPK inhibitor PD98059 significantly blocked the stimulation effect of LPA on cell proliferation in Hela cells PI3Kinhibitor LY294002 partially blocked the LPA effect on cell proliferation
with ice-cold PBS and then lysed in SDS sample buffer Sam-ples were resolved in electrophoresis using 10ndash12 sodiumdodecyl sulfate (SDS) polyacrylamide gels and transferredto PVDF membranes (Bio-Rad Hercules CA) Immunoblotanalyses were carried out using the appropriate antibodiesSpecific proteins were detected with the enhanced chemilu-minescence (ECL) system (Amersham Pharmacia BiotechPiscataway NJ)
26 Statistical Analysis Statistical significance was assessedby one-wayANOVAusing SPSS130 softwareTheBand in theWestern blotting was quantified with software Quantity OneData are presented as the means plusmn standard error 119901 le 005was used as standard for statistical significance
3 Results
31 Effects of LPA on Proliferation of Hela Cell Cells weretreated with LPA at different concentrations including 0120583M01 120583M 1 120583M 5 120583M 20120583M and 50 120583M for different timeperiods including 24 hours 48 hours 72 hours and 96hours It was found that LPA significantly stimulated theproliferation of Hela cells (119901 lt 005) Between 01 120583M and20120583M the higher the LPA concentration is the greater thestimulation effect on cell proliferation was observed Therewas no significant difference between the stimulation effectof 20120583M LPA and 50 120583M LPA on cell proliferation Thestimulation effect of LPA on proliferation of Hela cells wasalso increased along with the time The longer the exposuretime the greater the stimulation effect of LPA (Figure 1(a))
Several inhibitors were employed to determine which sig-naling transduction pathway was involved in the LPA effect
on Hela cell proliferation The inhibitors include LY294002(PI3K inhibitor) PD98059 (MAPKinase inhibitor) and Y-27632 (Rho kinase inhibitor) Cells were treated with 20120583Mof LPA plus different inhibitors It was found that PD98059(MEK1 inhibitor) significantly blocked the stimulation effectof LPA on the proliferation of Hela cell (119901 lt 005) LY294002(PI3K inhibitor) also partially blocked the stimulation effectof LPA on the proliferation of Hela cell (Figure 1(b)) Theresults suggested that RasRaf-MAPK signal pathway may beinvolved in the LPA stimulation of Hela cells proliferation
32 Effect of LPA on Apoptosis Induced by Cisplatin Follow-ing treatmentwith increasing concentrations of cisplatin (50ndash250 120583gmL) for 8 hours many floating cells were observed(Figure 2(a)) indicating the occurrence of extensive celldeaths To confirm the cells died of apoptosis Hela cellstreated with cisplatin were subject to flow cytometry anal-ysis and the assay of caspase-3 activity Similar levels ofapoptosis were detected by flow cytometry andmeasurementof caspase-3 activity The apoptosis rates elevated with theincrease of cisplatin concentrations The apoptosis rate was096 plusmn 007 1365 plusmn 136 1949 plusmn 075 2144 plusmn 1163748 plusmn 165 and 4415 plusmn 133when cisplatin was applied at50 100 150 200 and 250 120583gmL respectively (Figure 2(b))
To examine the LPA effects on cisplatin-induced apopto-sis Hela cells were treated by 200120583gmL cisplatin alone orplus LPA for 4 hours It was found that apoptosis inducedby cisplatin was significantly inhibited by LPA Moreoverthis inhibition displayed a dose-dependency The inhibitionfor cell apoptosis became significant (119901 lt 005) when theconcentration of LPA was ge20120583M (Figure 2(c))
4 BioMed Research International
50120583gmL 100120583gmL
150120583gmL 200120583gmL 250120583gmL
0120583gmL
(a)
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
50120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
100120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
150120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
200120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
250120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
0120583gmL
50
40
30
20
10
0
Apop
tosis
rate
()
2500 50 100 150 200
Cisplatin (120583gmL)
(b)
Figure 2 Continued
BioMed Research International 5
50
40
30
20
10
0
Apop
tosis
rate
()
0 50 100 150 200 250
LPA (120583M)
⋆⋆
⋆
⋆
(c)
60
40
20
0
Caspase-3 activity
pNA
den
sity
(120583M
mL)
Con
trol
Gro
up
Cisp
latin
LPA
+ ci
splat
in
⋆
(d)
Figure 2 LPA effects on apoptosis induced by cisplatin in Hela cell (a) Cell pictures (times20) Hela cells were treated with cisplatin at differentdoses After 8 hours of treatment with cisplatin many floating cells were observed The bigger the dose of cisplatin the more the floatingcells (b) Flow cytometry detection of apoptosis with Annexin VPI staining Hela cells were treated with cisplatin at different doses for 4hours Apoptotic cells were detected and quantified with flow cytometry Increased apoptosis following cisplatin treatment was observed (c)Quantification of LPA effects on apoptosis induced by cisplatin Hela cells were treated with cisplatin (at 200 120583gmL) and LPA (at differentconcentrations) for 4 hours The apoptotic cells were detected by flow cytometry and the apoptosis rate of Hela cells was quantified andshown Cell apoptosis induced by cisplatin was significantly inhibited by LPA (d) Caspase-3 activity Hela cells were treated with 200120583gmLof cisplatin plus LPA at 20 120583M for 4 hours The caspase-3 activity was measured and shown Cisplatin dramatically increased the caspase-3activity but the upregulated activity of caspase-3 was partially reversed by LPA treatment
In caspase-3 activity assay Hela cells were treated with200120583gmL of cisplatin alone or plus 20 120583M of LPA for 4hours It was found that cisplatin treatment significantlyincreased the caspase-3 activity but the upregulation ofcaspase-3 activitywas partially reversed by LPA (Figure 2(d))
33 LPA Effects on the Expression of Apoptosis Proteins Sincecisplatin induced apoptosis in cervical cancer Hela cell wefurther characterized alterations of key proteins involvedin the apoptotic pathway Western blotting showed thatthe expression of Fas-1 was significantly increased by cis-platin treatment in a dose-dependent manner (Figure 3(a))Expression of the antiapoptotic protein Bcl-2 was signif-icantly inhibited by cisplatin Moreover the inhibition ofBcl-2 expression was dependent on the dose of cisplatin(Figure 3(b)) Expression of another important apoptoticprotein Bax was also significantly increased by cisplatintreatment (Figure 3(c)) Since LPA appeared to rescue Helacells from apoptosis induced by cisplatin we examinedthe LPA effect on the change of apoptosis-related proteinsinduced by cisplatin Western blotting showed that theincrease of Fas-1 expression induced by cisplatin was signifi-cantly inhibited by LPA (Figure 4(a)) Moreover LPA treat-ment restored the inhibition of Bcl-2 expression by cisplatin(Figure 4(b)) in comparison to cells treated with cisplatinalone (Figure 2(b)) In addition LPA significantly inhibitedthe increased Bax expression induced by cisplatin Theincrease of Bax expression induced by cisplatin was signifi-cantly inhibited by LPA (Figure 4(c)) These results indicated
that LPA was able to block the apoptotic effects of cisplatinby reversing the changes in apoptotic proteins caused bycisplatin treatment
34 Signaling Transduction Pathways Involved in the LPAInhibition of Apoptosis Induced by Cisplatin Since severalstudies have shown cell apoptosis is associated with theactivation of PI3KAKT RasMAPK andRHOGEFsRHOApathways we applied specific inhibitors specifically targetingthese pathways to elucidate the mechanism underlying theLPA antiapoptotic activity Cells were treated by 200120583gmLcisplatin alone or plus 20120583M LPA and additionally dif-ferent inhibitors including PD98059 (MAPK inhibitor at10 120583M) LY294002 (PI3K inhibitor at 10 120583M) or Y-27632(Rho inhibitor at 10 120583M) It was found that the PI3K inhibitorLY294002 significantly reversed the protection effect of LPAon apoptosis induced by cisplatin (119901 = 0003) while MAPKinhibitor PD98059 and Rho inhibitor Y-27632 had no signif-icant effect (Figures 5(a) and 5(b)) These results suggestedthat the LPA effect on apoptosis induced by cisplatin werelikely mediated by the PI3KAKT signaling pathway but notRasMAPK and RHOGEFsRHOA pathways
35 Effects of LPA on the Phosphorylation of AKT Since AKTinhibitor significantly reversed the LPA protection of cellapoptosis caused by cisplatin we subsequently investigatedif LPA could activate the PI3KAKT pathway by increasingthe phosphorylation of AKT Cells were treated by LPA atdifferent concentrations including 0120583M 01 120583M 1 120583M 5 120583M
6 BioMed Research International
10
8
6
4
2
00 50 100 150 200 250
0 50 100 150 200 250
Fas-1
120573-actinCisplatin (120583gmL)
Cisplatin (120583gmL)
Fas-1 expression
Band
inte
nsity
(a)
0 50 100 150 200 250
Cisplatin (120583gmL)
0 50 100 150 200 250
Bcl-2
120573-actin
Cisplatin (120583gmL)
Bcl-2 expression3
2
1
0
Band
inte
nsity
(b)
5
4
3
2
1
0
0 50 100 150 200 250
0 50 100 150 200 250
Bax
120573-actin
Cisplatin (120583gmL)
Cisplatin (120583gmL)
Bax expression
Band
inte
nsity
(c)
Figure 3Western blotting analysis of apoptotic proteins in cells treated with cisplatin Hela cells were treated with cisplatin at different dosesincluding 050 120583gmL 100 120583gmL 150 120583gmL 200120583gmL and 250 120583gmL for 4 hours 120573-actin was used as a protein loading control Theresultant bandswere subject to densitometry analysis for quantification (a)Western blotting of Fas-LThe expression of Fas-Lwas significantlyinduced by cisplatin in a dose-dependentmanner (b)Western blotting of Bcl-2The expression of Bcl-2 was significantly inhibited by cisplatinin a dose-dependent manner (c) Western blotting of Bax The expression of Bax was significantly induced by cisplatin in a dose-dependentmanner
20120583M and 50120583Mof LPA for 1 hour (Figure 6(a))The resultsshowed that LPA treatment led to an increased phosphoryla-tion of AKT in a dose-dependent manner In addition LPA-mediated activation of AKT was also characterized by thetime-dependentmannerWhen cells were treatedwith 20120583Mof LPA for different time AKT phosphorylation graduallyincreased reaching the peak at the 2-hour time point Therewas a significant difference between control and the LPA-treated groups at 5min 10min 30min 1 hour and 2 hours
(Figure 6(b)) These results indicated that LPA protectioneffect on the apoptosis induced by cisplatin may be relatedto the phosphorylation of AKT
36 Effects of LPA on the Phosphorylation of ERK SinceMAPK inhibitor PD98059 significantly inhibited the pro-liferation stimulated by LPA which suggested ERKMEKMAPK pathway was involved in the LPA functions in theproliferation in Hela cells we investigated whether LPA can
BioMed Research International 7
Fas-1
120573-actin
Fas-1 expression5
4
3
2
1
0
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(a)
Bcl-2
120573-actin
Bcl-2 expression4
3
2
1
0
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(b)
4
3
2
1
0
Bax
120573-actin
Bax expression
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(c)
Figure 4 LPA effects on the expression of apoptotic proteins induced by cisplatin Hela cells were treated with cisplatin (at 200120583gmL) aloneor plus LPA (at 20 120583M) for 4 hours Western blotting was used to determine the expression of apoptosis related proteins 120573-actin was used as aprotein loading control (a) Western blotting of Fas-L The increased Fas-L expression induced by cisplatin was significantly reversed by LPAtreatment (b) Western blotting of Bcl-2 LPA restored the Bcl-2 expression which was inhibited by cisplatin (c) Western blotting of BaxTheupregulation of Bax expression induced by cisplatin was significantly inhibited by LPA
activate the phosphorylation of ERKThe results showed thatLPA significantly activated the phosphorylation of ERK ina dose-dependent manner Cells were treated with LPA atdifferent concentrations including 0120583M 01 120583M 1 120583M 5 120583M20120583M and 50120583M for 1 hour The effect on ERK phosphory-lation of LPA was increased along with the increase of LPAconcentration (Figure 7(a))
LPA also activated ERK in a time-dependent mannerCells were treated by 20 120583M LPA at different time pointsincluding 0min 5min 10min 30min 1 hour 2 hours 3hours and 4 hours ERK phosphorylation reached the peakat 3 hours following LPA treatment (Figure 7(b))
4 Discussion
In the intrinsic or mitochondrial pathway of apoptosis Bcl-2is able to prevent cell apoptosis while a related protein Baxcan migrate to the surface of the mitochondrion where itinhibits the protective effect of Bcl-2 by interrupting themembrane structure causing the release of cytochrome Cand caspase activation In the extrinsic or death receptorpathway of apoptosis Fas and the TNF receptor are integralmembrane proteins receiving apoptotic signals Uponbinding by ligand they transmit signals to the cytoplasmwhich leads to activation of caspase and phagocytosis
8 BioMed Research International
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
Control LPAPI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + PD98069
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + LY294002
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + Y-27632
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
(a)
Con
trol
LPA+
PD98069
LPA+
LY294002
LPA+
Y-27632
LPA
Apop
tosis
rate
0
5
10
15
20
25
30
35
40
45
50
(b)
Figure 5 Flow cytometry detection of cell apoptosis by Annexin VPI staining (a) Hela cells were treated with cisplatin (200120583gmL) andLPA (20 120583M) alone or in combination with different inhibitors for 4 hours Cisplatin and LPA were used in each inhibitors-treated groupInhibitors including PD98059 (MAPK inhibitor at 10 120583M) PD98059 (MAPK inhibitor at 10 120583M) and Y-27632 (Rho inhibitor at 10 120583M)wereapplied respectively (b) The apoptosis rate of Hela cells was quantified Cisplatin-treated group served as control PI3K inhibitor LY294002significantly reversed the LPA effect on apoptosis induced by cisplatin while MAPK inhibitor PD98059 and Rho inhibitor Y-27632 had nosignificant effect
BioMed Research International 9
Fold
chan
ge o
f ban
d in
tens
ity
5
4
3
2
1
00 01 1 5 20 50
LPA (120583M)
0 01 1 5 20 50 LPA (120583M)
P-AKT
AKT
(a)
Fold
chan
ge o
f ban
d in
tens
ity
P-AKT
Total-AKT
4
35
3
25
2
15
1
05
00 5min 10min 30min 1h 2h 3h 4h
Time
0 5min 10min 30min 1h 2h 3h 4h Time
AKT activation
(b)
Figure 6 Western blotting analysis of phosphorylated AKT in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01120583M 1 120583M 5 120583M 20 120583M and 50 120583M for 1 hour Western blotting was conducted to detect the expressionof phosphorylated AKT Total AKT was detected and the results were used as protein input control Densitometry analysis was carried outfor quantification purpose LPA treatment led to increased phosphorylation of AKT in a dose-dependent manner (b) Time-response effectof LPA on the phosphorylation of AKT Hela cells were treated with LPA at 20 120583M for different times including 5min 10min 30min 1 hour2 hours 3 hours and 4 hours Western blotting was used to detect the expression of phosphorylated AKT Compared to the total AKT LPAtreatment activated the phosphorylation of AKT in a time-dependent manner
25
20
15
10
05
00
Fold
chan
ge
0 01 10 50 20 50
LPA (120583M)
0 01 10 05 20 50 LPA (120583M)
P-ERK
ERK
(a)
25
20
15
10
05
00
Fold
chan
ge
0 min5 min10 min30 1h 2h 3h 4hTime
0 min5 min10 min30 1h 2h 3h 4h Time
P-ERK
ERK
(b)
Figure 7 Western blotting analysis of phosphorylated ERK in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01 120583M 1 120583M 5120583M 20 120583M and 50 120583M for 1 hour Western blotting was performed to detect the levels ofphosphorylated ERK Total ERK was used as protein input control The density of bands was quantified and compared LPA activated thephosphorylation of ERK in a dose-dependent manner (b) Hela cells were treated with LPA at 20120583M for different times including 5min10min 30min 1 hour 2 hours 3 hours and 4 hours A time-dependent increase in the ERK phosphorylation was detected
10 BioMed Research International
Some studies found that cell apoptosis induced by cis-platinwasmediated by death receptors signaling pathway [2021] while others claimed that it was mediated by mitochon-drial pathway [22 23] The discrepancy in previous studiesmay be caused by the divergent cell lines methodologiesand conditions used by different groups In this study weobserved that cisplatin significantly induced the apoptosis ofHela cells in a concentration-dependent manner Cisplatintreatment was accompanied by increased Fas-1 and Baxexpression and the increased enzyme activity of caspase-3 Meanwhile cisplatin inhibited Bcl-2 expression Theseobservations suggested that cisplatin-induced apoptosis ofHela cell may be mediated by both intrinsic and extrinsicpathways Indeed extensive cross talks between the two path-ways have been identified [24 25] Simultaneous activationof the two pathways has been observed in many types of cellsfollowing various treatments
Our previous study found that both ovarian cancer cellsand peritoneal mesothelial cells contribute to the elevatedLPA levels in ovarian cancer ascites Under normal cultureconditions ovarian cancer cells and prostate cancer cellsbut not breast cancer cell lines constitutively synthesize andrelease significant amounts of LPA to the culture medium Amajor finding of this study is that LPA was able to rescue theHela cells apoptosis induced by cisplatin The protective roleof LPA against apoptosis was observed in ovarian cancer cells[26]We observed that LPA treatment blocked the alterationsof Bcl-2 Fas-1 and Bax expression that were induced bycisplatin LPA also significantly inhibited the upregulationof the caspase-3 enzyme activity induced by cisplatin Ourobservations of LPA effect on cervical cancer were consistentwith other reports on LPA effects in different kinds of cellsMeng et al found that LPA may protect epithelial ovariancancer from immune cell attack and cytoskeleton disruptingreagents induced apoptosis through multiple pathways andLPA inhibited anti-Fas-induced apoptosis enhanced by actindepolymerization [26 27] Actin depolymerization acceler-ated caspase-8 activation while LPA inhibited the associationand activation of caspase-8 at the DISC in epithelial ovariancancer OVCAR3 cells Thus our observations have raisedan interesting question on the potential role of LPA forthe development of chemoresistance by cervical cancer cellsIn addition could LPA be an effective biomarker for theprediction of chemoresistance in cervical cancer Furtherstudies are required to answer these questions
In this study three specific kinase inhibitors were appliedto determine the signaling pathways involved in the pro-tection effect of LPA on apoptosis It was found that PI3Kinhibitor (LY290042) almost completely blocked the LPAprotection against apoptosis induced by cisplatin Subsequentexperiments confirmed that LPA treatment of Hela cells ledto increased phosphorylation of AKT in a dose- and time-dependent manner This indicated that LPA protected Helacells from apoptosis induced by cisplatin This effect of LPAmay be through the activation of PI3KAKT pathway Thisfinding appears to be consistent with published data showingthat PI3KAKT pathway plays a key role in the regulationof cell apoptosis [28 29] Wan et al observed that PI3Kinhibition augmented staurosporine-induced apoptosis in
the endometrial carcinoma cell line [28] In that experimentstaurosporine reduced the levels of phosphorylation in AKTand Bad In a study by Kim et al 24-bis(p-hydroxyphenyl)-2-butenal (HPB242) induced apoptosis via the inhibition ofPI3KAKT pathway in human cervical cancer cells HPB242treatment decreased phosphatidyl inositol 3-kinase and p-AKT expression levels demonstrating that this survivalpathway may also be inhibited by HPB242 [30] In anotherstudy Cui et al found that MiR-125b inhibited tumor growthand promoted apoptosis of cervical cancer cells by targetingphosphoinositide 3-kinase catalytic subunit delta Overex-pression of MiR-125b in Hela cervical cancer cells inducedapoptosis and downregulated the expression of PIK3CDand Phospho-AKT [31] Kang et al found that thioridazineinduces apoptosis by targeting the PI3KAKTmTOR path-way in cervical cancer cells [32] Thanaketpaisarn et alfound that Artesunate enhanced TRAIL-induced apoptosisin human cervical carcinoma cells through inhibition of thePI3KAKT and NF-120581B signaling pathways [33] Wang et alfound that Stathmin is involved in arsenic trioxide-inducedapoptosis in human cervical cancer cell lines via PI3K linkedsignal pathway As2O3-induced stathmin downregulation ismediated through the phosphatidylinositol-3-kinase (PI3K)signaling pathway PI3K inhibitor effectively attenuated thestathmin downregulation and cell apoptosis upon As2O3-treatment [34] Recurrent and metastatic cervical cancersoften acquire chemoresistance to cisplatin Our findingpointed to a possible sensitization scheme through combinedadministration of PI3K inhibitor and cisplatin
It should be pointed out that while our current studyfocused on the apoptotic pathways andmechanisms the LPAeffects on cell cycle control and cell proliferation should notbe excluded In fact the PI3KAKT pathway is known to playan active role in the control of cell cycle [35] We found thatMEK1 inhibitor (PD98059) significantly inhibited the effectof LPA on the proliferation of Hela cells RasRaf1MEKERKsignal transduction pathway is one of the most importantmembers of the mitogen-activated protein kinase pathwaysregulating cell proliferation [36] MEK1 a MAPK representsa key molecule of the RasRaf1MEKERK signal pathway[37] Several lines of evidence suggest that in cervical cancerthe RasRaf1MEKERK pathway but not the JNK pathwayor the p38 MAPK pathway is the major regulator of cellproliferation [38ndash40]
In summary we demonstrated that cisplatin inducedapoptosis of Hela cell via the downregulation of Bcl-2 andupregulation of Bax Fas-L and increased enzyme activityof caspase-3 LPA was able to block the alterations in theapoptotic factors caused by cisplatin and ultimately reversedthe apoptosis induced by cisplatin Cisplatin-induced cellapoptosis appeared to be partially dependent on PI3KAKTpathway and AKT inhibitor was able to reverse the pro-tective effects of LPA against cisplatin-induced apoptosisLPA stimulated the proliferation of Hela cells throughthe RasRaf1MEKERK pathway These data enriched ourknowledge regarding the LPA activity in cervical cancer cellsIn addition the finding on the positive impact of PI3KAKTinhibitor on the apoptotic effects of cisplatin has provideduseful information for the design of sensitization strategy in
BioMed Research International 11
cervical cancer patients especially those with high levels ofLPA
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Yanxia Sui and Ya Yang contributed equally to this work
Acknowledgments
This study is supported by the following funding sourcesThe National Natural Science Foundations of China (JuanRen 31201060C0709 Juan Ren 30973175H1621 and JuanRen 81172490H1621) Program for New Century ExcellentTalents in University (Juan Ren NCET-12-0440) Scientificand Technological Research Foundation of Shaanxi Province(Juan Ren 2012K13-01-06 Juan Ren 2007K09-09) Scien-tific Research Foundation for the Returned overseas Chi-nese Scholars of State Education Ministry (Juan Ren 0601-18920006) Research Foundation of Health Department ofShaanrsquoxi Province (Juan Ren 2010D41) Qing Nian Jiao ShiGenZongJiHua ofXirsquoan JiaotongUniversity (JuanRen 2012)Program for Changjiang Scholars and Innovative ResearchTeam inUniversity (Zongfang Li PCSIRT1171) andResearchFoundation of Xirsquoan Jiao Tong University of China (JuanRen)
References
[1] J R Erickson Y Hasegawa X Fang et al ldquoLysophosphatidicacid and ovarian cancer a paradigm for tumorogenesis andpatient managementrdquo Prostaglandins and Other Lipid Media-tors vol 64 no 1ndash4 pp 63ndash81 2001
[2] Y Xu Z Z Shen DWWiper et al ldquoLysophosphatidic acid as apotential biomarker for ovarian and other gynecologic cancersrdquoThe Journal of the American Medical Association vol 280 no 8pp 719ndash723 1998
[3] J S Karliner ldquoLysophospholipids and the cardiovascular sys-temrdquoBiochimica et Biophysica ActamdashMolecular and Cell Biologyof Lipids vol 1582 no 1ndash3 pp 216ndash221 2002
[4] C Schulze C Smales L L Rubin and J M StaddonldquoLysophosphatidic acid increases tight junction permeability incultured brain endothelial cellsrdquo Journal of Neurochemistry vol68 no 3 pp 991ndash1000 1997
[5] I Ishii N Fukushima X Ye and J Chun ldquoLysophospholipidreceptors signaling and biologyrdquo Annual Review of Biochem-istry vol 73 pp 321ndash354 2004
[6] T S Panetti ldquoDifferential effects of sphingosine 1-phosphateand lysophosphatidic acid on endothelial cellsrdquo Biochimica etBiophysica ActamdashMolecular and Cell Biology of Lipids vol 1582no 1ndash3 pp 190ndash196 2002
[7] Z Z Shen M Z Wu P Elson et al ldquoFatty acid composition oflysophosphatidic acid and lysophosphatidylinositol in plasmafrom patients with ovarian cancer and other gynecologicaldiseasesrdquo Gynecologic Oncology vol 83 no 1 pp 25ndash30 2001
[8] M M Murph ldquoAgonist-induced endocytosis of lysophospha-tidic acid-coupled LPA1EDG-2 receptors via a dynamin2- andRab5-dependent pathwayrdquo Journal of Cell Science vol 116 no10 pp 1969ndash1980 2003
[9] L M Baudhuin K L Cristina J Lu and Y Xu ldquoAkt activationinduced by lysophosphatidic acid and sphingosine-1-phosphaterequires both mitogen-activated protein kinase kinase andp38 mitogen-activated protein kinase and is cell-line specificrdquoMolecular Pharmacology vol 62 no 3 pp 660ndash671 2002
[10] R-J Chen S-U Chen C-H Chou andM-C Lin ldquoLysophos-phatidic acid receptor 23-mediated IL-8-dependent angiogen-esis in cervical cancer cellsrdquo International Journal of Cancer vol131 no 4 pp 789ndash802 2012
[11] J W Choi S Lim Y-S Oh et al ldquoSubtype-specific role ofphospholipase C-120573 in bradykinin and LPA signaling throughdifferential binding of different PDZ scaffold proteinsrdquo CellularSignalling vol 22 no 7 pp 1153ndash1161 2010
[12] Y Kishi S Okudaira M Tanaka et al ldquoAutotaxin is over-expressed in glioblastoma multiforme and contributes to cellmotility of glioblastoma by converting lysophosphatidylcholineTO lysophosphatidic acidrdquoThe Journal of Biological Chemistryvol 281 no 25 pp 17492ndash17500 2006
[13] A Eastman ldquoThe formation isolation and characterization ofDNA adducts produced by anticancer platinum complexesrdquoPharmacology andTherapeutics vol 34 no 2 pp 155ndash166 1987
[14] K Wozniak and J Błasiak ldquoRecognition and repair of DNA-cisplatin adductsrdquo Acta Biochimica Polonica vol 49 no 3 pp583ndash596 2002
[15] A Basu and S Krishnamurthy ldquoCellular responses to cisplatin-induced DNA damagerdquo Journal of Nucleic Acids vol 2010Article ID 201367 16 pages 2010
[16] A Efeyan and M Serrano ldquop53 guardian of the genome andpoliceman of the oncogenesrdquo Cell Cycle vol 6 no 9 pp 1006ndash1010 2007
[17] O Kutuk E D Arisan T Tezil M C Shoshan and H BasagaldquoCisplatin overcomesBcl-2-mediated resistance to apoptosis viapreferential engagement of Bak critical role of Noxa-mediatedlipid peroxidationrdquo Carcinogenesis vol 30 no 9 pp 1517ndash15272009
[18] C M Yan E Q Chai H Y Cai G Miao and W MaldquoOleuropein induces apoptosis via activation of caspases andsuppression of phosphatidylinositol 3-kinaseprotein kinaseB pathway in HepG2 human hepatoma cell linerdquo MolecularMedicine Reports vol 11 pp 4617ndash4624 2015
[19] X Y Wang X S Fan L Cai S Liu X F Cong and X ChenldquoLysophosphatidic acid rescues bone mesenchymal stem cellsfrom hydrogen peroxide-induced apoptosisrdquo Apoptosis vol 20no 3 pp 273ndash284 2015
[20] A Brozovic G Fritz M Christmann et al ldquoLong-term activa-tion of SAPKJNK p38 kinase and fas-L expression by cisplatinis attenuated in human carcinoma cells that acquired drugresistancerdquo International Journal of Cancer vol 112 no 6 pp974ndash985 2004
[21] Y Q Liu H Xing X B Han et al ldquoApoptosis of HeLa cellsinduced by cisplatin and its mechanismrdquo Journal of HuazhongUniversity of Science and TechnologymdashMedical Science vol 28no 2 pp 197ndash199 2008
[22] K L Jin J-Y Park E J Noh et al ldquoThe effect of combinedtreatment with cisplatin and histone deacetylase inhibitors onHeLa cellsrdquo Journal of Gynecologic Oncology vol 21 no 4 pp262ndash268 2010
12 BioMed Research International
[23] T Sakaeda T Nakamura M Hirai et al ldquoMDR1 up-regulatedby apoptotic stimuli suppresses apoptotic signalingrdquo Pharma-ceutical Research vol 19 no 9 pp 1323ndash1329 2002
[24] K Kandasamy S M Srinivasula E S Alnemri et al ldquoInvolve-ment of proapoptotic molecules Bax and Bak in tumor necro-sis factor-related apoptosis-inducing ligand (TRAIL)-inducedmitochondrial disruption and apoptosis differential regulationof cytochrome c and SmacDIABLO releaserdquo Cancer Researchvol 63 no 7 pp 1712ndash1721 2003
[25] H Toyota N Yanase T Yoshimoto M Moriyama T Sudoand J Mizuguchi ldquoCalpain-induced Bax-cleavage product is amore potent inducer of apoptotic cell death thanwild-type BaxrdquoCancer Letters vol 189 no 2 pp 221ndash230 2003
[26] Y R Meng S J Kang and D A Fishman ldquoLysophosphatidicacid inhibits anti-Fas-mediated apoptosis enhanced by actindepolymerization in epithelial ovarian cancerrdquo FEBS Lettersvol 579 no 5 pp 1311ndash1319 2005
[27] Y Meng S Kang J So S Reierstad and D A FishmanldquoTranslocation of Fas by LPA prevents ovarian cancer cells fromanti-Fas-induced apoptosisrdquo Gynecologic Oncology vol 96 no2 pp 462ndash469 2005
[28] X Wan Y Yokoyama A Shinohara Y Takahashi and TTamaya ldquoPTEN augments staurosporine-induced apoptosis inPTEN-null Ishikawa cells by downregulating PI3KAkt signal-ing pathwayrdquo Cell Death and Differentiation vol 9 no 4 pp414ndash420 2002
[29] J Yu Y Zhang J Mcilroy T Rordorf-Nikolic G A Orr and JM Backer ldquoRegulation of the p85p110 phosphatidylinositol 31015840-kinase stabilization and inhibition of the p110120572 catalytic subunitby the p85 regulatory subunitrdquoMolecular and Cellular Biologyvol 18 no 3 pp 1379ndash1387 1998
[30] M S Kim J H Kim Y Bak et al ldquo24-bis (p-hydroxyphenyl)-2-butenal (HPB242) induces apoptosis via modulating E7expression and inhibition of PI3KAkt pathway in SiHa humancervical cancer cellsrdquo Nutrition and Cancer vol 64 no 8 pp1236ndash1244 2012
[31] F Cui X L Li X Y Zhu et al ldquoMiR-125b inhibits tumorgrowth and promotes apoptosis of cervical cancer cells bytargeting phosphoinositide 3-kinase catalytic subunit deltardquoCellular Physiology and Biochemistry vol 30 no 5 pp 1310ndash1318 2012
[32] S Kang S M Dong B-R Kim et al ldquoThioridazine inducesapoptosis by targeting the PI3KAktmTOR pathway in cervicaland endometrial cancer cellsrdquo Apoptosis vol 17 no 9 pp 989ndash997 2012
[33] O Thanaketpaisarn P Waiwut H Sakurai and I Saiki ldquoArte-sunate enhances TRAIL-induced apoptosis in human cervicalcarcinoma cells through inhibition of the NF-120581B and PI3KAktsignaling pathwaysrdquo International Journal of Oncology vol 39no 1 pp 279ndash285 2011
[34] XWang J-H Ren F Lin et al ldquoStathmin is involved in arsenictrioxide-induced apoptosis in human cervical cancer cell linesvia PI3K linked signal pathwayrdquo Cancer Biology and Therapyvol 10 no 6 pp 632ndash643 2010
[35] C W Lee N H Kim H K Choi et al ldquoLysophosphatidicacid-induced c-fos up-regulation involves cyclic AMP responseelement-binding protein activated by mitogen- and stress-activated protein kinase-1rdquo Journal of Cellular Biochemistry vol104 no 3 pp 785ndash794 2008
[36] DW Hommes M P Peppelenbosch and S J H van DeventerldquoMitogen activated protein (MAP) kinase signal transduction
pathways and novel anti-inflammatory targetsrdquoGut vol 52 no1 pp 144ndash151 2003
[37] M Mukai F Imamura M Ayaki et al ldquoInhibition of tumorinvasion andmetastasis by a novel lysophosphatidic acid (cyclicLPA)rdquo International Journal of Cancer vol 81 no 6 pp 918ndash9221999
[38] E-Y Huang Y-F Chen Y-M Chen et al ldquoA novel radiore-sistant mechanism of galectin-1 mediated by H-Ras-dependentpathways in cervical cancer cellsrdquo Cell Death and Disease vol3 no 1 article e251 2012
[39] W-H Su P-C Chuang E-Y Huang and K D YangldquoRadiation-induced increase in cell migration and metastaticpotential of cervical cancer cells operates via theK-ras pathwayrdquoThe American Journal of Pathology vol 180 no 2 pp 862ndash8712012
[40] H Rincon-Arano R Rosales N Mora A Rodriguez-Castaneda and C Rosales ldquoR-Ras promotes tumor growthof cervical epithelial cellsrdquo Cancer vol 97 no 3 pp 575ndash5852003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 BioMed Research International
Increased level of lysophosphatidic acid is also found inpatients with acute myocardial infarction LPA has beenimplicated in the development of the cardiovascular systemassisting in its progression to a fully functional state [3 4] LPAis a bioactive glycerophospholipid generated and releasedby platelets macrophages epithelial cells and tumor cellsLPAmodulates a broad range of cellular responses includingalterations of cell proliferation protection against apoptosismodulation of chemotaxis and transcellular migration [56] thereby affecting the survival of ovarian cancer cellsmacrophages fibroblasts and neonatal cardiacmyocytesThesignificant role of LPA in triggering these cellular responseshas implicated LPA in tumor progression
It has also been reported that LPA is increased in theplasma of cervical cancer patients [2] Xu et al found thatstage I and stage IV cervical cancer patients had significantlyhigher plasma LPA levels than normal controls ElevatedLPA levels were detected in all the 6 cervical cancer patientsexamined [2] In addition therewas an increased ratio of totalLPAlysophosphatidylinositol (LPI) [7] Similarly Shen et alreported that the ratio of unsaturated LPALPI subspecieswassignificantly higher in patients with cervical cancer than inhealthy controls [7] The significantly increased LPA in theplasma of patients of cervical cancer points to its possiblerole(s) for the development of this malignancy Indeed LPAreceptorswere also found to be extensively expressed in cervi-cal cell lines including Hela CaSki and Siha [8ndash11] Previousstudies from this group confirmed a high expression level ofLPA receptors especially the LPA receptor 2 inHela cells [12]providing a basis for using this cell line as a study model toinvestigate the LPA bioactivity and the underlying pathways
Cisplatin (DDP) has been used as the first line chemother-apy drug for adjuvant treatment of cervical cancer patientsCisplatin-induced DNA damage activates multiple signalingpathways leading to cell apoptosis [13ndash15] DNA damagecaused by cisplatin induces the phosphorylation and stabi-lization of p53 [16] p53 promotes cisplatin-induced apop-tosis by antagonizing the antiapoptotic effect of Bcl-xL [17]Phosphatidylinositol 3-kinaseAKT pathway is also involvedin apoptosis regulation Yan et al found that suppression ofPI3KAKT pathway caused apoptosis in the HepG2 humanhepatoma cell line [18] On the other hand Wang et alfound that LPA protects bone marrow-derived mesenchymalstem cells (BMSCs) against hypoxia- or serum deprivation-induced apoptosis [19] LPA rescues H
2O2-induced apoptosis
by activating ERK12 and PI3KAKT pathways in mesenchy-mal stem cells [19] However LPA effect on the apoptosis incervical cancer cells and the potential mechanism remainsunclear
In this study we investigate how LPA-triggered cellresponsesmay affect the cell apoptosis induced by cisplatin ina cervical cancer cell lineWe characterized the effects of LPAon cell apoptotic factors including Bcl-2 Bax and caspase-3in Hela cells treated with cisplatin The influence of LPA onthe upstream pathway PI3KAKT was also determined Thefindings provide insights on the general bioactivity as well asthe chemotherapeutic concerns of LPA produced by cervicalcancer cells
2 Materials and Methods
21 Materials 1-Oleoyl LPA (18 1 LPA) was purchased fromAvanti Polar Lipids (Birmingham AL USA) Inhibitor ofphosphoinositide 3-kinase (PI3K) LY290042 and inhibitorof mitogen-activated protein kinase (MAPK) PD98059were from Cell Signaling (Beverly MA USA) Rho kinaseinhibitor Y-27632 was from Biomol (Beverly MA USA)Hela cells were cultured in Dulbeccorsquos modified Eaglersquosmedium (DMEM) supplemented with 100mLL fetal bovineserum (FBS) streptomycin (100mgL) and penicillin(100KUL) at 37∘C in 50mLL CO
2incubator Cells were
serum starved for 12 hours before LPA treatment
22 Cell Proliferation Assay 3-(45-Dimethylthiazol-2-yl)-25-diphenyltetrazolium bromide (MTT) colorimetry assaywas employed to measure cell proliferation Hela cells (2 times103well) were seeded at 96-well plate After being starvedfor 12 hours cells were fed with DMEM containing LPAsupplemented with 1 gL bovine serum albumin Following24 48 72 and 96 hours of culture 20120583L of MTT solution(5 gL) was added to each well Four hours later the mediumwas removed and 150 120583L of dimethyl sulfoxide (DMSO) wasadded to each well Absorbance was measured at 490 nm ona Microplate Reader (EXL800) Each assay was performed inquintuplicate
23 Annexin V Staining and Flow Cytometry Assay Cellswere starved in serum-free DMEM for 12 hours and treatedwith different concentrations of LPA For kinase inhibitorexperiments LY294002 (50120583molL) PD98059 (10 120583molL)and Y-27632 (10 120583molL) were applied to cells 30minbefore addition of LPA Cells were resuspended in bindingbuffer (10mmolL HEPESNaOH pH74 140mmolL NaCl25mmolL CaCl
2) and stained with 5120583L of annexin-FITC
and 5mgL propidium iodide (PI) Following washing cellswere analyzed with flow cytometry (FACS Calibur cytometerBD Biosciences USA) and CellQuest (BD Biosciences) forquantification of cell apoptosis The experiment was per-formed in triplicate and repeated twice
24 Caspase-3 Activity Assay Activity of caspase-3 wasdetected using the CaspACE Assay System (Promega USA)Briefly cell lysates from treated and untreated control cellswere prepared Cell lysates were centrifuged at 15000timesg for20min at 4∘C and the supernatant was collected The assaywas performed in a total volume of 100 120583L in 96-well platesCell extracts with an equal amount of protein (50ndash100120583g oftotal protein) were mixed with specific colorimetric substrate(DEVD-pNA) for caspase-3 The mixture was incubated at37∘C for 4 h according to the manufacturerrsquos protocol Theabsorbance was measured at a wave length of 405 nm
25 Western Blotting Antibodies against AKT (1 200 dilu-tion times) and phosphorylated-AKT (1 200 dilution times)were purchased from Cell Signaling Transduction (USA)Antibodies against Fas (1 200 dilution times) Bcl-2 (1 200dilution times) and Bax (1 200 dilution times) were pur-chased from Santa Cruz After treatment cells were rinsed
BioMed Research International 3
800
600
400
200
000
0 24 48 72 96
(hours)
Cel
l num
ber (times10
4)
Control011
520500
(a)
800
600
400
200
000
0 24 48 72 96
(hours)
Cel
l num
ber (times10
4)
ControlLPA + PD98059LPA + LY294002
LPA + Y-27632LPA
(b)
Figure 1 LPA effects on cell proliferation of Hela cells (a) Hela cells were treated with different doses and time of LPA MTT essay wasperformed LPA significantly stimulated the proliferation of Hela cells in a dose-dependent and time-dependent manner (b) Effect ofinhibitors on LPA stimulation of cell proliferation Hela cells were treatedwith LPA at 20120583Mplus different inhibitors for different time periodsbefore MTT essay PD98059 (MAPK inhibitor at 10 120583M) PD98059 (MAPK inhibitor at 10 120583M) and Y-27632 (Rho inhibitor at 10 120583M) wereapplied respectively MAPK inhibitor PD98059 significantly blocked the stimulation effect of LPA on cell proliferation in Hela cells PI3Kinhibitor LY294002 partially blocked the LPA effect on cell proliferation
with ice-cold PBS and then lysed in SDS sample buffer Sam-ples were resolved in electrophoresis using 10ndash12 sodiumdodecyl sulfate (SDS) polyacrylamide gels and transferredto PVDF membranes (Bio-Rad Hercules CA) Immunoblotanalyses were carried out using the appropriate antibodiesSpecific proteins were detected with the enhanced chemilu-minescence (ECL) system (Amersham Pharmacia BiotechPiscataway NJ)
26 Statistical Analysis Statistical significance was assessedby one-wayANOVAusing SPSS130 softwareTheBand in theWestern blotting was quantified with software Quantity OneData are presented as the means plusmn standard error 119901 le 005was used as standard for statistical significance
3 Results
31 Effects of LPA on Proliferation of Hela Cell Cells weretreated with LPA at different concentrations including 0120583M01 120583M 1 120583M 5 120583M 20120583M and 50 120583M for different timeperiods including 24 hours 48 hours 72 hours and 96hours It was found that LPA significantly stimulated theproliferation of Hela cells (119901 lt 005) Between 01 120583M and20120583M the higher the LPA concentration is the greater thestimulation effect on cell proliferation was observed Therewas no significant difference between the stimulation effectof 20120583M LPA and 50 120583M LPA on cell proliferation Thestimulation effect of LPA on proliferation of Hela cells wasalso increased along with the time The longer the exposuretime the greater the stimulation effect of LPA (Figure 1(a))
Several inhibitors were employed to determine which sig-naling transduction pathway was involved in the LPA effect
on Hela cell proliferation The inhibitors include LY294002(PI3K inhibitor) PD98059 (MAPKinase inhibitor) and Y-27632 (Rho kinase inhibitor) Cells were treated with 20120583Mof LPA plus different inhibitors It was found that PD98059(MEK1 inhibitor) significantly blocked the stimulation effectof LPA on the proliferation of Hela cell (119901 lt 005) LY294002(PI3K inhibitor) also partially blocked the stimulation effectof LPA on the proliferation of Hela cell (Figure 1(b)) Theresults suggested that RasRaf-MAPK signal pathway may beinvolved in the LPA stimulation of Hela cells proliferation
32 Effect of LPA on Apoptosis Induced by Cisplatin Follow-ing treatmentwith increasing concentrations of cisplatin (50ndash250 120583gmL) for 8 hours many floating cells were observed(Figure 2(a)) indicating the occurrence of extensive celldeaths To confirm the cells died of apoptosis Hela cellstreated with cisplatin were subject to flow cytometry anal-ysis and the assay of caspase-3 activity Similar levels ofapoptosis were detected by flow cytometry andmeasurementof caspase-3 activity The apoptosis rates elevated with theincrease of cisplatin concentrations The apoptosis rate was096 plusmn 007 1365 plusmn 136 1949 plusmn 075 2144 plusmn 1163748 plusmn 165 and 4415 plusmn 133when cisplatin was applied at50 100 150 200 and 250 120583gmL respectively (Figure 2(b))
To examine the LPA effects on cisplatin-induced apopto-sis Hela cells were treated by 200120583gmL cisplatin alone orplus LPA for 4 hours It was found that apoptosis inducedby cisplatin was significantly inhibited by LPA Moreoverthis inhibition displayed a dose-dependency The inhibitionfor cell apoptosis became significant (119901 lt 005) when theconcentration of LPA was ge20120583M (Figure 2(c))
4 BioMed Research International
50120583gmL 100120583gmL
150120583gmL 200120583gmL 250120583gmL
0120583gmL
(a)
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
50120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
100120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
150120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
200120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
250120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
0120583gmL
50
40
30
20
10
0
Apop
tosis
rate
()
2500 50 100 150 200
Cisplatin (120583gmL)
(b)
Figure 2 Continued
BioMed Research International 5
50
40
30
20
10
0
Apop
tosis
rate
()
0 50 100 150 200 250
LPA (120583M)
⋆⋆
⋆
⋆
(c)
60
40
20
0
Caspase-3 activity
pNA
den
sity
(120583M
mL)
Con
trol
Gro
up
Cisp
latin
LPA
+ ci
splat
in
⋆
(d)
Figure 2 LPA effects on apoptosis induced by cisplatin in Hela cell (a) Cell pictures (times20) Hela cells were treated with cisplatin at differentdoses After 8 hours of treatment with cisplatin many floating cells were observed The bigger the dose of cisplatin the more the floatingcells (b) Flow cytometry detection of apoptosis with Annexin VPI staining Hela cells were treated with cisplatin at different doses for 4hours Apoptotic cells were detected and quantified with flow cytometry Increased apoptosis following cisplatin treatment was observed (c)Quantification of LPA effects on apoptosis induced by cisplatin Hela cells were treated with cisplatin (at 200 120583gmL) and LPA (at differentconcentrations) for 4 hours The apoptotic cells were detected by flow cytometry and the apoptosis rate of Hela cells was quantified andshown Cell apoptosis induced by cisplatin was significantly inhibited by LPA (d) Caspase-3 activity Hela cells were treated with 200120583gmLof cisplatin plus LPA at 20 120583M for 4 hours The caspase-3 activity was measured and shown Cisplatin dramatically increased the caspase-3activity but the upregulated activity of caspase-3 was partially reversed by LPA treatment
In caspase-3 activity assay Hela cells were treated with200120583gmL of cisplatin alone or plus 20 120583M of LPA for 4hours It was found that cisplatin treatment significantlyincreased the caspase-3 activity but the upregulation ofcaspase-3 activitywas partially reversed by LPA (Figure 2(d))
33 LPA Effects on the Expression of Apoptosis Proteins Sincecisplatin induced apoptosis in cervical cancer Hela cell wefurther characterized alterations of key proteins involvedin the apoptotic pathway Western blotting showed thatthe expression of Fas-1 was significantly increased by cis-platin treatment in a dose-dependent manner (Figure 3(a))Expression of the antiapoptotic protein Bcl-2 was signif-icantly inhibited by cisplatin Moreover the inhibition ofBcl-2 expression was dependent on the dose of cisplatin(Figure 3(b)) Expression of another important apoptoticprotein Bax was also significantly increased by cisplatintreatment (Figure 3(c)) Since LPA appeared to rescue Helacells from apoptosis induced by cisplatin we examinedthe LPA effect on the change of apoptosis-related proteinsinduced by cisplatin Western blotting showed that theincrease of Fas-1 expression induced by cisplatin was signifi-cantly inhibited by LPA (Figure 4(a)) Moreover LPA treat-ment restored the inhibition of Bcl-2 expression by cisplatin(Figure 4(b)) in comparison to cells treated with cisplatinalone (Figure 2(b)) In addition LPA significantly inhibitedthe increased Bax expression induced by cisplatin Theincrease of Bax expression induced by cisplatin was signifi-cantly inhibited by LPA (Figure 4(c)) These results indicated
that LPA was able to block the apoptotic effects of cisplatinby reversing the changes in apoptotic proteins caused bycisplatin treatment
34 Signaling Transduction Pathways Involved in the LPAInhibition of Apoptosis Induced by Cisplatin Since severalstudies have shown cell apoptosis is associated with theactivation of PI3KAKT RasMAPK andRHOGEFsRHOApathways we applied specific inhibitors specifically targetingthese pathways to elucidate the mechanism underlying theLPA antiapoptotic activity Cells were treated by 200120583gmLcisplatin alone or plus 20120583M LPA and additionally dif-ferent inhibitors including PD98059 (MAPK inhibitor at10 120583M) LY294002 (PI3K inhibitor at 10 120583M) or Y-27632(Rho inhibitor at 10 120583M) It was found that the PI3K inhibitorLY294002 significantly reversed the protection effect of LPAon apoptosis induced by cisplatin (119901 = 0003) while MAPKinhibitor PD98059 and Rho inhibitor Y-27632 had no signif-icant effect (Figures 5(a) and 5(b)) These results suggestedthat the LPA effect on apoptosis induced by cisplatin werelikely mediated by the PI3KAKT signaling pathway but notRasMAPK and RHOGEFsRHOA pathways
35 Effects of LPA on the Phosphorylation of AKT Since AKTinhibitor significantly reversed the LPA protection of cellapoptosis caused by cisplatin we subsequently investigatedif LPA could activate the PI3KAKT pathway by increasingthe phosphorylation of AKT Cells were treated by LPA atdifferent concentrations including 0120583M 01 120583M 1 120583M 5 120583M
6 BioMed Research International
10
8
6
4
2
00 50 100 150 200 250
0 50 100 150 200 250
Fas-1
120573-actinCisplatin (120583gmL)
Cisplatin (120583gmL)
Fas-1 expression
Band
inte
nsity
(a)
0 50 100 150 200 250
Cisplatin (120583gmL)
0 50 100 150 200 250
Bcl-2
120573-actin
Cisplatin (120583gmL)
Bcl-2 expression3
2
1
0
Band
inte
nsity
(b)
5
4
3
2
1
0
0 50 100 150 200 250
0 50 100 150 200 250
Bax
120573-actin
Cisplatin (120583gmL)
Cisplatin (120583gmL)
Bax expression
Band
inte
nsity
(c)
Figure 3Western blotting analysis of apoptotic proteins in cells treated with cisplatin Hela cells were treated with cisplatin at different dosesincluding 050 120583gmL 100 120583gmL 150 120583gmL 200120583gmL and 250 120583gmL for 4 hours 120573-actin was used as a protein loading control Theresultant bandswere subject to densitometry analysis for quantification (a)Western blotting of Fas-LThe expression of Fas-Lwas significantlyinduced by cisplatin in a dose-dependentmanner (b)Western blotting of Bcl-2The expression of Bcl-2 was significantly inhibited by cisplatinin a dose-dependent manner (c) Western blotting of Bax The expression of Bax was significantly induced by cisplatin in a dose-dependentmanner
20120583M and 50120583Mof LPA for 1 hour (Figure 6(a))The resultsshowed that LPA treatment led to an increased phosphoryla-tion of AKT in a dose-dependent manner In addition LPA-mediated activation of AKT was also characterized by thetime-dependentmannerWhen cells were treatedwith 20120583Mof LPA for different time AKT phosphorylation graduallyincreased reaching the peak at the 2-hour time point Therewas a significant difference between control and the LPA-treated groups at 5min 10min 30min 1 hour and 2 hours
(Figure 6(b)) These results indicated that LPA protectioneffect on the apoptosis induced by cisplatin may be relatedto the phosphorylation of AKT
36 Effects of LPA on the Phosphorylation of ERK SinceMAPK inhibitor PD98059 significantly inhibited the pro-liferation stimulated by LPA which suggested ERKMEKMAPK pathway was involved in the LPA functions in theproliferation in Hela cells we investigated whether LPA can
BioMed Research International 7
Fas-1
120573-actin
Fas-1 expression5
4
3
2
1
0
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(a)
Bcl-2
120573-actin
Bcl-2 expression4
3
2
1
0
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(b)
4
3
2
1
0
Bax
120573-actin
Bax expression
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(c)
Figure 4 LPA effects on the expression of apoptotic proteins induced by cisplatin Hela cells were treated with cisplatin (at 200120583gmL) aloneor plus LPA (at 20 120583M) for 4 hours Western blotting was used to determine the expression of apoptosis related proteins 120573-actin was used as aprotein loading control (a) Western blotting of Fas-L The increased Fas-L expression induced by cisplatin was significantly reversed by LPAtreatment (b) Western blotting of Bcl-2 LPA restored the Bcl-2 expression which was inhibited by cisplatin (c) Western blotting of BaxTheupregulation of Bax expression induced by cisplatin was significantly inhibited by LPA
activate the phosphorylation of ERKThe results showed thatLPA significantly activated the phosphorylation of ERK ina dose-dependent manner Cells were treated with LPA atdifferent concentrations including 0120583M 01 120583M 1 120583M 5 120583M20120583M and 50120583M for 1 hour The effect on ERK phosphory-lation of LPA was increased along with the increase of LPAconcentration (Figure 7(a))
LPA also activated ERK in a time-dependent mannerCells were treated by 20 120583M LPA at different time pointsincluding 0min 5min 10min 30min 1 hour 2 hours 3hours and 4 hours ERK phosphorylation reached the peakat 3 hours following LPA treatment (Figure 7(b))
4 Discussion
In the intrinsic or mitochondrial pathway of apoptosis Bcl-2is able to prevent cell apoptosis while a related protein Baxcan migrate to the surface of the mitochondrion where itinhibits the protective effect of Bcl-2 by interrupting themembrane structure causing the release of cytochrome Cand caspase activation In the extrinsic or death receptorpathway of apoptosis Fas and the TNF receptor are integralmembrane proteins receiving apoptotic signals Uponbinding by ligand they transmit signals to the cytoplasmwhich leads to activation of caspase and phagocytosis
8 BioMed Research International
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
Control LPAPI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + PD98069
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + LY294002
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + Y-27632
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
(a)
Con
trol
LPA+
PD98069
LPA+
LY294002
LPA+
Y-27632
LPA
Apop
tosis
rate
0
5
10
15
20
25
30
35
40
45
50
(b)
Figure 5 Flow cytometry detection of cell apoptosis by Annexin VPI staining (a) Hela cells were treated with cisplatin (200120583gmL) andLPA (20 120583M) alone or in combination with different inhibitors for 4 hours Cisplatin and LPA were used in each inhibitors-treated groupInhibitors including PD98059 (MAPK inhibitor at 10 120583M) PD98059 (MAPK inhibitor at 10 120583M) and Y-27632 (Rho inhibitor at 10 120583M)wereapplied respectively (b) The apoptosis rate of Hela cells was quantified Cisplatin-treated group served as control PI3K inhibitor LY294002significantly reversed the LPA effect on apoptosis induced by cisplatin while MAPK inhibitor PD98059 and Rho inhibitor Y-27632 had nosignificant effect
BioMed Research International 9
Fold
chan
ge o
f ban
d in
tens
ity
5
4
3
2
1
00 01 1 5 20 50
LPA (120583M)
0 01 1 5 20 50 LPA (120583M)
P-AKT
AKT
(a)
Fold
chan
ge o
f ban
d in
tens
ity
P-AKT
Total-AKT
4
35
3
25
2
15
1
05
00 5min 10min 30min 1h 2h 3h 4h
Time
0 5min 10min 30min 1h 2h 3h 4h Time
AKT activation
(b)
Figure 6 Western blotting analysis of phosphorylated AKT in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01120583M 1 120583M 5 120583M 20 120583M and 50 120583M for 1 hour Western blotting was conducted to detect the expressionof phosphorylated AKT Total AKT was detected and the results were used as protein input control Densitometry analysis was carried outfor quantification purpose LPA treatment led to increased phosphorylation of AKT in a dose-dependent manner (b) Time-response effectof LPA on the phosphorylation of AKT Hela cells were treated with LPA at 20 120583M for different times including 5min 10min 30min 1 hour2 hours 3 hours and 4 hours Western blotting was used to detect the expression of phosphorylated AKT Compared to the total AKT LPAtreatment activated the phosphorylation of AKT in a time-dependent manner
25
20
15
10
05
00
Fold
chan
ge
0 01 10 50 20 50
LPA (120583M)
0 01 10 05 20 50 LPA (120583M)
P-ERK
ERK
(a)
25
20
15
10
05
00
Fold
chan
ge
0 min5 min10 min30 1h 2h 3h 4hTime
0 min5 min10 min30 1h 2h 3h 4h Time
P-ERK
ERK
(b)
Figure 7 Western blotting analysis of phosphorylated ERK in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01 120583M 1 120583M 5120583M 20 120583M and 50 120583M for 1 hour Western blotting was performed to detect the levels ofphosphorylated ERK Total ERK was used as protein input control The density of bands was quantified and compared LPA activated thephosphorylation of ERK in a dose-dependent manner (b) Hela cells were treated with LPA at 20120583M for different times including 5min10min 30min 1 hour 2 hours 3 hours and 4 hours A time-dependent increase in the ERK phosphorylation was detected
10 BioMed Research International
Some studies found that cell apoptosis induced by cis-platinwasmediated by death receptors signaling pathway [2021] while others claimed that it was mediated by mitochon-drial pathway [22 23] The discrepancy in previous studiesmay be caused by the divergent cell lines methodologiesand conditions used by different groups In this study weobserved that cisplatin significantly induced the apoptosis ofHela cells in a concentration-dependent manner Cisplatintreatment was accompanied by increased Fas-1 and Baxexpression and the increased enzyme activity of caspase-3 Meanwhile cisplatin inhibited Bcl-2 expression Theseobservations suggested that cisplatin-induced apoptosis ofHela cell may be mediated by both intrinsic and extrinsicpathways Indeed extensive cross talks between the two path-ways have been identified [24 25] Simultaneous activationof the two pathways has been observed in many types of cellsfollowing various treatments
Our previous study found that both ovarian cancer cellsand peritoneal mesothelial cells contribute to the elevatedLPA levels in ovarian cancer ascites Under normal cultureconditions ovarian cancer cells and prostate cancer cellsbut not breast cancer cell lines constitutively synthesize andrelease significant amounts of LPA to the culture medium Amajor finding of this study is that LPA was able to rescue theHela cells apoptosis induced by cisplatin The protective roleof LPA against apoptosis was observed in ovarian cancer cells[26]We observed that LPA treatment blocked the alterationsof Bcl-2 Fas-1 and Bax expression that were induced bycisplatin LPA also significantly inhibited the upregulationof the caspase-3 enzyme activity induced by cisplatin Ourobservations of LPA effect on cervical cancer were consistentwith other reports on LPA effects in different kinds of cellsMeng et al found that LPA may protect epithelial ovariancancer from immune cell attack and cytoskeleton disruptingreagents induced apoptosis through multiple pathways andLPA inhibited anti-Fas-induced apoptosis enhanced by actindepolymerization [26 27] Actin depolymerization acceler-ated caspase-8 activation while LPA inhibited the associationand activation of caspase-8 at the DISC in epithelial ovariancancer OVCAR3 cells Thus our observations have raisedan interesting question on the potential role of LPA forthe development of chemoresistance by cervical cancer cellsIn addition could LPA be an effective biomarker for theprediction of chemoresistance in cervical cancer Furtherstudies are required to answer these questions
In this study three specific kinase inhibitors were appliedto determine the signaling pathways involved in the pro-tection effect of LPA on apoptosis It was found that PI3Kinhibitor (LY290042) almost completely blocked the LPAprotection against apoptosis induced by cisplatin Subsequentexperiments confirmed that LPA treatment of Hela cells ledto increased phosphorylation of AKT in a dose- and time-dependent manner This indicated that LPA protected Helacells from apoptosis induced by cisplatin This effect of LPAmay be through the activation of PI3KAKT pathway Thisfinding appears to be consistent with published data showingthat PI3KAKT pathway plays a key role in the regulationof cell apoptosis [28 29] Wan et al observed that PI3Kinhibition augmented staurosporine-induced apoptosis in
the endometrial carcinoma cell line [28] In that experimentstaurosporine reduced the levels of phosphorylation in AKTand Bad In a study by Kim et al 24-bis(p-hydroxyphenyl)-2-butenal (HPB242) induced apoptosis via the inhibition ofPI3KAKT pathway in human cervical cancer cells HPB242treatment decreased phosphatidyl inositol 3-kinase and p-AKT expression levels demonstrating that this survivalpathway may also be inhibited by HPB242 [30] In anotherstudy Cui et al found that MiR-125b inhibited tumor growthand promoted apoptosis of cervical cancer cells by targetingphosphoinositide 3-kinase catalytic subunit delta Overex-pression of MiR-125b in Hela cervical cancer cells inducedapoptosis and downregulated the expression of PIK3CDand Phospho-AKT [31] Kang et al found that thioridazineinduces apoptosis by targeting the PI3KAKTmTOR path-way in cervical cancer cells [32] Thanaketpaisarn et alfound that Artesunate enhanced TRAIL-induced apoptosisin human cervical carcinoma cells through inhibition of thePI3KAKT and NF-120581B signaling pathways [33] Wang et alfound that Stathmin is involved in arsenic trioxide-inducedapoptosis in human cervical cancer cell lines via PI3K linkedsignal pathway As2O3-induced stathmin downregulation ismediated through the phosphatidylinositol-3-kinase (PI3K)signaling pathway PI3K inhibitor effectively attenuated thestathmin downregulation and cell apoptosis upon As2O3-treatment [34] Recurrent and metastatic cervical cancersoften acquire chemoresistance to cisplatin Our findingpointed to a possible sensitization scheme through combinedadministration of PI3K inhibitor and cisplatin
It should be pointed out that while our current studyfocused on the apoptotic pathways andmechanisms the LPAeffects on cell cycle control and cell proliferation should notbe excluded In fact the PI3KAKT pathway is known to playan active role in the control of cell cycle [35] We found thatMEK1 inhibitor (PD98059) significantly inhibited the effectof LPA on the proliferation of Hela cells RasRaf1MEKERKsignal transduction pathway is one of the most importantmembers of the mitogen-activated protein kinase pathwaysregulating cell proliferation [36] MEK1 a MAPK representsa key molecule of the RasRaf1MEKERK signal pathway[37] Several lines of evidence suggest that in cervical cancerthe RasRaf1MEKERK pathway but not the JNK pathwayor the p38 MAPK pathway is the major regulator of cellproliferation [38ndash40]
In summary we demonstrated that cisplatin inducedapoptosis of Hela cell via the downregulation of Bcl-2 andupregulation of Bax Fas-L and increased enzyme activityof caspase-3 LPA was able to block the alterations in theapoptotic factors caused by cisplatin and ultimately reversedthe apoptosis induced by cisplatin Cisplatin-induced cellapoptosis appeared to be partially dependent on PI3KAKTpathway and AKT inhibitor was able to reverse the pro-tective effects of LPA against cisplatin-induced apoptosisLPA stimulated the proliferation of Hela cells throughthe RasRaf1MEKERK pathway These data enriched ourknowledge regarding the LPA activity in cervical cancer cellsIn addition the finding on the positive impact of PI3KAKTinhibitor on the apoptotic effects of cisplatin has provideduseful information for the design of sensitization strategy in
BioMed Research International 11
cervical cancer patients especially those with high levels ofLPA
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Yanxia Sui and Ya Yang contributed equally to this work
Acknowledgments
This study is supported by the following funding sourcesThe National Natural Science Foundations of China (JuanRen 31201060C0709 Juan Ren 30973175H1621 and JuanRen 81172490H1621) Program for New Century ExcellentTalents in University (Juan Ren NCET-12-0440) Scientificand Technological Research Foundation of Shaanxi Province(Juan Ren 2012K13-01-06 Juan Ren 2007K09-09) Scien-tific Research Foundation for the Returned overseas Chi-nese Scholars of State Education Ministry (Juan Ren 0601-18920006) Research Foundation of Health Department ofShaanrsquoxi Province (Juan Ren 2010D41) Qing Nian Jiao ShiGenZongJiHua ofXirsquoan JiaotongUniversity (JuanRen 2012)Program for Changjiang Scholars and Innovative ResearchTeam inUniversity (Zongfang Li PCSIRT1171) andResearchFoundation of Xirsquoan Jiao Tong University of China (JuanRen)
References
[1] J R Erickson Y Hasegawa X Fang et al ldquoLysophosphatidicacid and ovarian cancer a paradigm for tumorogenesis andpatient managementrdquo Prostaglandins and Other Lipid Media-tors vol 64 no 1ndash4 pp 63ndash81 2001
[2] Y Xu Z Z Shen DWWiper et al ldquoLysophosphatidic acid as apotential biomarker for ovarian and other gynecologic cancersrdquoThe Journal of the American Medical Association vol 280 no 8pp 719ndash723 1998
[3] J S Karliner ldquoLysophospholipids and the cardiovascular sys-temrdquoBiochimica et Biophysica ActamdashMolecular and Cell Biologyof Lipids vol 1582 no 1ndash3 pp 216ndash221 2002
[4] C Schulze C Smales L L Rubin and J M StaddonldquoLysophosphatidic acid increases tight junction permeability incultured brain endothelial cellsrdquo Journal of Neurochemistry vol68 no 3 pp 991ndash1000 1997
[5] I Ishii N Fukushima X Ye and J Chun ldquoLysophospholipidreceptors signaling and biologyrdquo Annual Review of Biochem-istry vol 73 pp 321ndash354 2004
[6] T S Panetti ldquoDifferential effects of sphingosine 1-phosphateand lysophosphatidic acid on endothelial cellsrdquo Biochimica etBiophysica ActamdashMolecular and Cell Biology of Lipids vol 1582no 1ndash3 pp 190ndash196 2002
[7] Z Z Shen M Z Wu P Elson et al ldquoFatty acid composition oflysophosphatidic acid and lysophosphatidylinositol in plasmafrom patients with ovarian cancer and other gynecologicaldiseasesrdquo Gynecologic Oncology vol 83 no 1 pp 25ndash30 2001
[8] M M Murph ldquoAgonist-induced endocytosis of lysophospha-tidic acid-coupled LPA1EDG-2 receptors via a dynamin2- andRab5-dependent pathwayrdquo Journal of Cell Science vol 116 no10 pp 1969ndash1980 2003
[9] L M Baudhuin K L Cristina J Lu and Y Xu ldquoAkt activationinduced by lysophosphatidic acid and sphingosine-1-phosphaterequires both mitogen-activated protein kinase kinase andp38 mitogen-activated protein kinase and is cell-line specificrdquoMolecular Pharmacology vol 62 no 3 pp 660ndash671 2002
[10] R-J Chen S-U Chen C-H Chou andM-C Lin ldquoLysophos-phatidic acid receptor 23-mediated IL-8-dependent angiogen-esis in cervical cancer cellsrdquo International Journal of Cancer vol131 no 4 pp 789ndash802 2012
[11] J W Choi S Lim Y-S Oh et al ldquoSubtype-specific role ofphospholipase C-120573 in bradykinin and LPA signaling throughdifferential binding of different PDZ scaffold proteinsrdquo CellularSignalling vol 22 no 7 pp 1153ndash1161 2010
[12] Y Kishi S Okudaira M Tanaka et al ldquoAutotaxin is over-expressed in glioblastoma multiforme and contributes to cellmotility of glioblastoma by converting lysophosphatidylcholineTO lysophosphatidic acidrdquoThe Journal of Biological Chemistryvol 281 no 25 pp 17492ndash17500 2006
[13] A Eastman ldquoThe formation isolation and characterization ofDNA adducts produced by anticancer platinum complexesrdquoPharmacology andTherapeutics vol 34 no 2 pp 155ndash166 1987
[14] K Wozniak and J Błasiak ldquoRecognition and repair of DNA-cisplatin adductsrdquo Acta Biochimica Polonica vol 49 no 3 pp583ndash596 2002
[15] A Basu and S Krishnamurthy ldquoCellular responses to cisplatin-induced DNA damagerdquo Journal of Nucleic Acids vol 2010Article ID 201367 16 pages 2010
[16] A Efeyan and M Serrano ldquop53 guardian of the genome andpoliceman of the oncogenesrdquo Cell Cycle vol 6 no 9 pp 1006ndash1010 2007
[17] O Kutuk E D Arisan T Tezil M C Shoshan and H BasagaldquoCisplatin overcomesBcl-2-mediated resistance to apoptosis viapreferential engagement of Bak critical role of Noxa-mediatedlipid peroxidationrdquo Carcinogenesis vol 30 no 9 pp 1517ndash15272009
[18] C M Yan E Q Chai H Y Cai G Miao and W MaldquoOleuropein induces apoptosis via activation of caspases andsuppression of phosphatidylinositol 3-kinaseprotein kinaseB pathway in HepG2 human hepatoma cell linerdquo MolecularMedicine Reports vol 11 pp 4617ndash4624 2015
[19] X Y Wang X S Fan L Cai S Liu X F Cong and X ChenldquoLysophosphatidic acid rescues bone mesenchymal stem cellsfrom hydrogen peroxide-induced apoptosisrdquo Apoptosis vol 20no 3 pp 273ndash284 2015
[20] A Brozovic G Fritz M Christmann et al ldquoLong-term activa-tion of SAPKJNK p38 kinase and fas-L expression by cisplatinis attenuated in human carcinoma cells that acquired drugresistancerdquo International Journal of Cancer vol 112 no 6 pp974ndash985 2004
[21] Y Q Liu H Xing X B Han et al ldquoApoptosis of HeLa cellsinduced by cisplatin and its mechanismrdquo Journal of HuazhongUniversity of Science and TechnologymdashMedical Science vol 28no 2 pp 197ndash199 2008
[22] K L Jin J-Y Park E J Noh et al ldquoThe effect of combinedtreatment with cisplatin and histone deacetylase inhibitors onHeLa cellsrdquo Journal of Gynecologic Oncology vol 21 no 4 pp262ndash268 2010
12 BioMed Research International
[23] T Sakaeda T Nakamura M Hirai et al ldquoMDR1 up-regulatedby apoptotic stimuli suppresses apoptotic signalingrdquo Pharma-ceutical Research vol 19 no 9 pp 1323ndash1329 2002
[24] K Kandasamy S M Srinivasula E S Alnemri et al ldquoInvolve-ment of proapoptotic molecules Bax and Bak in tumor necro-sis factor-related apoptosis-inducing ligand (TRAIL)-inducedmitochondrial disruption and apoptosis differential regulationof cytochrome c and SmacDIABLO releaserdquo Cancer Researchvol 63 no 7 pp 1712ndash1721 2003
[25] H Toyota N Yanase T Yoshimoto M Moriyama T Sudoand J Mizuguchi ldquoCalpain-induced Bax-cleavage product is amore potent inducer of apoptotic cell death thanwild-type BaxrdquoCancer Letters vol 189 no 2 pp 221ndash230 2003
[26] Y R Meng S J Kang and D A Fishman ldquoLysophosphatidicacid inhibits anti-Fas-mediated apoptosis enhanced by actindepolymerization in epithelial ovarian cancerrdquo FEBS Lettersvol 579 no 5 pp 1311ndash1319 2005
[27] Y Meng S Kang J So S Reierstad and D A FishmanldquoTranslocation of Fas by LPA prevents ovarian cancer cells fromanti-Fas-induced apoptosisrdquo Gynecologic Oncology vol 96 no2 pp 462ndash469 2005
[28] X Wan Y Yokoyama A Shinohara Y Takahashi and TTamaya ldquoPTEN augments staurosporine-induced apoptosis inPTEN-null Ishikawa cells by downregulating PI3KAkt signal-ing pathwayrdquo Cell Death and Differentiation vol 9 no 4 pp414ndash420 2002
[29] J Yu Y Zhang J Mcilroy T Rordorf-Nikolic G A Orr and JM Backer ldquoRegulation of the p85p110 phosphatidylinositol 31015840-kinase stabilization and inhibition of the p110120572 catalytic subunitby the p85 regulatory subunitrdquoMolecular and Cellular Biologyvol 18 no 3 pp 1379ndash1387 1998
[30] M S Kim J H Kim Y Bak et al ldquo24-bis (p-hydroxyphenyl)-2-butenal (HPB242) induces apoptosis via modulating E7expression and inhibition of PI3KAkt pathway in SiHa humancervical cancer cellsrdquo Nutrition and Cancer vol 64 no 8 pp1236ndash1244 2012
[31] F Cui X L Li X Y Zhu et al ldquoMiR-125b inhibits tumorgrowth and promotes apoptosis of cervical cancer cells bytargeting phosphoinositide 3-kinase catalytic subunit deltardquoCellular Physiology and Biochemistry vol 30 no 5 pp 1310ndash1318 2012
[32] S Kang S M Dong B-R Kim et al ldquoThioridazine inducesapoptosis by targeting the PI3KAktmTOR pathway in cervicaland endometrial cancer cellsrdquo Apoptosis vol 17 no 9 pp 989ndash997 2012
[33] O Thanaketpaisarn P Waiwut H Sakurai and I Saiki ldquoArte-sunate enhances TRAIL-induced apoptosis in human cervicalcarcinoma cells through inhibition of the NF-120581B and PI3KAktsignaling pathwaysrdquo International Journal of Oncology vol 39no 1 pp 279ndash285 2011
[34] XWang J-H Ren F Lin et al ldquoStathmin is involved in arsenictrioxide-induced apoptosis in human cervical cancer cell linesvia PI3K linked signal pathwayrdquo Cancer Biology and Therapyvol 10 no 6 pp 632ndash643 2010
[35] C W Lee N H Kim H K Choi et al ldquoLysophosphatidicacid-induced c-fos up-regulation involves cyclic AMP responseelement-binding protein activated by mitogen- and stress-activated protein kinase-1rdquo Journal of Cellular Biochemistry vol104 no 3 pp 785ndash794 2008
[36] DW Hommes M P Peppelenbosch and S J H van DeventerldquoMitogen activated protein (MAP) kinase signal transduction
pathways and novel anti-inflammatory targetsrdquoGut vol 52 no1 pp 144ndash151 2003
[37] M Mukai F Imamura M Ayaki et al ldquoInhibition of tumorinvasion andmetastasis by a novel lysophosphatidic acid (cyclicLPA)rdquo International Journal of Cancer vol 81 no 6 pp 918ndash9221999
[38] E-Y Huang Y-F Chen Y-M Chen et al ldquoA novel radiore-sistant mechanism of galectin-1 mediated by H-Ras-dependentpathways in cervical cancer cellsrdquo Cell Death and Disease vol3 no 1 article e251 2012
[39] W-H Su P-C Chuang E-Y Huang and K D YangldquoRadiation-induced increase in cell migration and metastaticpotential of cervical cancer cells operates via theK-ras pathwayrdquoThe American Journal of Pathology vol 180 no 2 pp 862ndash8712012
[40] H Rincon-Arano R Rosales N Mora A Rodriguez-Castaneda and C Rosales ldquoR-Ras promotes tumor growthof cervical epithelial cellsrdquo Cancer vol 97 no 3 pp 575ndash5852003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
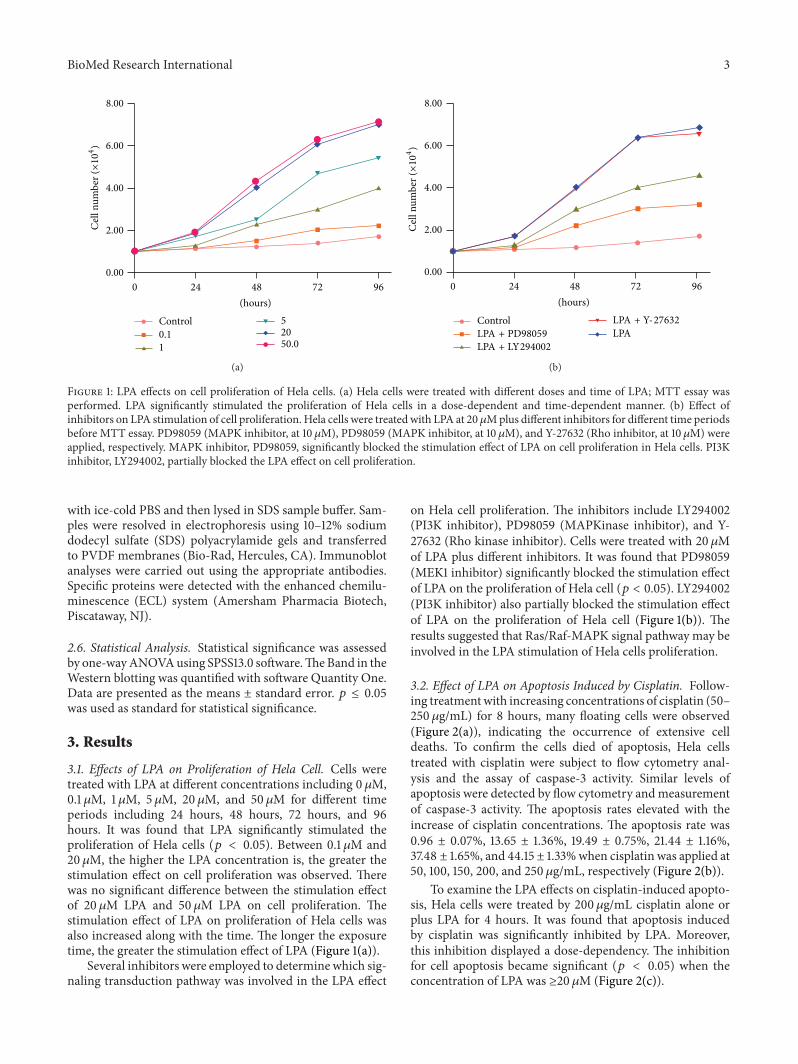
BioMed Research International 3
800
600
400
200
000
0 24 48 72 96
(hours)
Cel
l num
ber (times10
4)
Control011
520500
(a)
800
600
400
200
000
0 24 48 72 96
(hours)
Cel
l num
ber (times10
4)
ControlLPA + PD98059LPA + LY294002
LPA + Y-27632LPA
(b)
Figure 1 LPA effects on cell proliferation of Hela cells (a) Hela cells were treated with different doses and time of LPA MTT essay wasperformed LPA significantly stimulated the proliferation of Hela cells in a dose-dependent and time-dependent manner (b) Effect ofinhibitors on LPA stimulation of cell proliferation Hela cells were treatedwith LPA at 20120583Mplus different inhibitors for different time periodsbefore MTT essay PD98059 (MAPK inhibitor at 10 120583M) PD98059 (MAPK inhibitor at 10 120583M) and Y-27632 (Rho inhibitor at 10 120583M) wereapplied respectively MAPK inhibitor PD98059 significantly blocked the stimulation effect of LPA on cell proliferation in Hela cells PI3Kinhibitor LY294002 partially blocked the LPA effect on cell proliferation
with ice-cold PBS and then lysed in SDS sample buffer Sam-ples were resolved in electrophoresis using 10ndash12 sodiumdodecyl sulfate (SDS) polyacrylamide gels and transferredto PVDF membranes (Bio-Rad Hercules CA) Immunoblotanalyses were carried out using the appropriate antibodiesSpecific proteins were detected with the enhanced chemilu-minescence (ECL) system (Amersham Pharmacia BiotechPiscataway NJ)
26 Statistical Analysis Statistical significance was assessedby one-wayANOVAusing SPSS130 softwareTheBand in theWestern blotting was quantified with software Quantity OneData are presented as the means plusmn standard error 119901 le 005was used as standard for statistical significance
3 Results
31 Effects of LPA on Proliferation of Hela Cell Cells weretreated with LPA at different concentrations including 0120583M01 120583M 1 120583M 5 120583M 20120583M and 50 120583M for different timeperiods including 24 hours 48 hours 72 hours and 96hours It was found that LPA significantly stimulated theproliferation of Hela cells (119901 lt 005) Between 01 120583M and20120583M the higher the LPA concentration is the greater thestimulation effect on cell proliferation was observed Therewas no significant difference between the stimulation effectof 20120583M LPA and 50 120583M LPA on cell proliferation Thestimulation effect of LPA on proliferation of Hela cells wasalso increased along with the time The longer the exposuretime the greater the stimulation effect of LPA (Figure 1(a))
Several inhibitors were employed to determine which sig-naling transduction pathway was involved in the LPA effect
on Hela cell proliferation The inhibitors include LY294002(PI3K inhibitor) PD98059 (MAPKinase inhibitor) and Y-27632 (Rho kinase inhibitor) Cells were treated with 20120583Mof LPA plus different inhibitors It was found that PD98059(MEK1 inhibitor) significantly blocked the stimulation effectof LPA on the proliferation of Hela cell (119901 lt 005) LY294002(PI3K inhibitor) also partially blocked the stimulation effectof LPA on the proliferation of Hela cell (Figure 1(b)) Theresults suggested that RasRaf-MAPK signal pathway may beinvolved in the LPA stimulation of Hela cells proliferation
32 Effect of LPA on Apoptosis Induced by Cisplatin Follow-ing treatmentwith increasing concentrations of cisplatin (50ndash250 120583gmL) for 8 hours many floating cells were observed(Figure 2(a)) indicating the occurrence of extensive celldeaths To confirm the cells died of apoptosis Hela cellstreated with cisplatin were subject to flow cytometry anal-ysis and the assay of caspase-3 activity Similar levels ofapoptosis were detected by flow cytometry andmeasurementof caspase-3 activity The apoptosis rates elevated with theincrease of cisplatin concentrations The apoptosis rate was096 plusmn 007 1365 plusmn 136 1949 plusmn 075 2144 plusmn 1163748 plusmn 165 and 4415 plusmn 133when cisplatin was applied at50 100 150 200 and 250 120583gmL respectively (Figure 2(b))
To examine the LPA effects on cisplatin-induced apopto-sis Hela cells were treated by 200120583gmL cisplatin alone orplus LPA for 4 hours It was found that apoptosis inducedby cisplatin was significantly inhibited by LPA Moreoverthis inhibition displayed a dose-dependency The inhibitionfor cell apoptosis became significant (119901 lt 005) when theconcentration of LPA was ge20120583M (Figure 2(c))
4 BioMed Research International
50120583gmL 100120583gmL
150120583gmL 200120583gmL 250120583gmL
0120583gmL
(a)
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
50120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
100120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
150120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
200120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
250120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
0120583gmL
50
40
30
20
10
0
Apop
tosis
rate
()
2500 50 100 150 200
Cisplatin (120583gmL)
(b)
Figure 2 Continued
BioMed Research International 5
50
40
30
20
10
0
Apop
tosis
rate
()
0 50 100 150 200 250
LPA (120583M)
⋆⋆
⋆
⋆
(c)
60
40
20
0
Caspase-3 activity
pNA
den
sity
(120583M
mL)
Con
trol
Gro
up
Cisp
latin
LPA
+ ci
splat
in
⋆
(d)
Figure 2 LPA effects on apoptosis induced by cisplatin in Hela cell (a) Cell pictures (times20) Hela cells were treated with cisplatin at differentdoses After 8 hours of treatment with cisplatin many floating cells were observed The bigger the dose of cisplatin the more the floatingcells (b) Flow cytometry detection of apoptosis with Annexin VPI staining Hela cells were treated with cisplatin at different doses for 4hours Apoptotic cells were detected and quantified with flow cytometry Increased apoptosis following cisplatin treatment was observed (c)Quantification of LPA effects on apoptosis induced by cisplatin Hela cells were treated with cisplatin (at 200 120583gmL) and LPA (at differentconcentrations) for 4 hours The apoptotic cells were detected by flow cytometry and the apoptosis rate of Hela cells was quantified andshown Cell apoptosis induced by cisplatin was significantly inhibited by LPA (d) Caspase-3 activity Hela cells were treated with 200120583gmLof cisplatin plus LPA at 20 120583M for 4 hours The caspase-3 activity was measured and shown Cisplatin dramatically increased the caspase-3activity but the upregulated activity of caspase-3 was partially reversed by LPA treatment
In caspase-3 activity assay Hela cells were treated with200120583gmL of cisplatin alone or plus 20 120583M of LPA for 4hours It was found that cisplatin treatment significantlyincreased the caspase-3 activity but the upregulation ofcaspase-3 activitywas partially reversed by LPA (Figure 2(d))
33 LPA Effects on the Expression of Apoptosis Proteins Sincecisplatin induced apoptosis in cervical cancer Hela cell wefurther characterized alterations of key proteins involvedin the apoptotic pathway Western blotting showed thatthe expression of Fas-1 was significantly increased by cis-platin treatment in a dose-dependent manner (Figure 3(a))Expression of the antiapoptotic protein Bcl-2 was signif-icantly inhibited by cisplatin Moreover the inhibition ofBcl-2 expression was dependent on the dose of cisplatin(Figure 3(b)) Expression of another important apoptoticprotein Bax was also significantly increased by cisplatintreatment (Figure 3(c)) Since LPA appeared to rescue Helacells from apoptosis induced by cisplatin we examinedthe LPA effect on the change of apoptosis-related proteinsinduced by cisplatin Western blotting showed that theincrease of Fas-1 expression induced by cisplatin was signifi-cantly inhibited by LPA (Figure 4(a)) Moreover LPA treat-ment restored the inhibition of Bcl-2 expression by cisplatin(Figure 4(b)) in comparison to cells treated with cisplatinalone (Figure 2(b)) In addition LPA significantly inhibitedthe increased Bax expression induced by cisplatin Theincrease of Bax expression induced by cisplatin was signifi-cantly inhibited by LPA (Figure 4(c)) These results indicated
that LPA was able to block the apoptotic effects of cisplatinby reversing the changes in apoptotic proteins caused bycisplatin treatment
34 Signaling Transduction Pathways Involved in the LPAInhibition of Apoptosis Induced by Cisplatin Since severalstudies have shown cell apoptosis is associated with theactivation of PI3KAKT RasMAPK andRHOGEFsRHOApathways we applied specific inhibitors specifically targetingthese pathways to elucidate the mechanism underlying theLPA antiapoptotic activity Cells were treated by 200120583gmLcisplatin alone or plus 20120583M LPA and additionally dif-ferent inhibitors including PD98059 (MAPK inhibitor at10 120583M) LY294002 (PI3K inhibitor at 10 120583M) or Y-27632(Rho inhibitor at 10 120583M) It was found that the PI3K inhibitorLY294002 significantly reversed the protection effect of LPAon apoptosis induced by cisplatin (119901 = 0003) while MAPKinhibitor PD98059 and Rho inhibitor Y-27632 had no signif-icant effect (Figures 5(a) and 5(b)) These results suggestedthat the LPA effect on apoptosis induced by cisplatin werelikely mediated by the PI3KAKT signaling pathway but notRasMAPK and RHOGEFsRHOA pathways
35 Effects of LPA on the Phosphorylation of AKT Since AKTinhibitor significantly reversed the LPA protection of cellapoptosis caused by cisplatin we subsequently investigatedif LPA could activate the PI3KAKT pathway by increasingthe phosphorylation of AKT Cells were treated by LPA atdifferent concentrations including 0120583M 01 120583M 1 120583M 5 120583M
6 BioMed Research International
10
8
6
4
2
00 50 100 150 200 250
0 50 100 150 200 250
Fas-1
120573-actinCisplatin (120583gmL)
Cisplatin (120583gmL)
Fas-1 expression
Band
inte
nsity
(a)
0 50 100 150 200 250
Cisplatin (120583gmL)
0 50 100 150 200 250
Bcl-2
120573-actin
Cisplatin (120583gmL)
Bcl-2 expression3
2
1
0
Band
inte
nsity
(b)
5
4
3
2
1
0
0 50 100 150 200 250
0 50 100 150 200 250
Bax
120573-actin
Cisplatin (120583gmL)
Cisplatin (120583gmL)
Bax expression
Band
inte
nsity
(c)
Figure 3Western blotting analysis of apoptotic proteins in cells treated with cisplatin Hela cells were treated with cisplatin at different dosesincluding 050 120583gmL 100 120583gmL 150 120583gmL 200120583gmL and 250 120583gmL for 4 hours 120573-actin was used as a protein loading control Theresultant bandswere subject to densitometry analysis for quantification (a)Western blotting of Fas-LThe expression of Fas-Lwas significantlyinduced by cisplatin in a dose-dependentmanner (b)Western blotting of Bcl-2The expression of Bcl-2 was significantly inhibited by cisplatinin a dose-dependent manner (c) Western blotting of Bax The expression of Bax was significantly induced by cisplatin in a dose-dependentmanner
20120583M and 50120583Mof LPA for 1 hour (Figure 6(a))The resultsshowed that LPA treatment led to an increased phosphoryla-tion of AKT in a dose-dependent manner In addition LPA-mediated activation of AKT was also characterized by thetime-dependentmannerWhen cells were treatedwith 20120583Mof LPA for different time AKT phosphorylation graduallyincreased reaching the peak at the 2-hour time point Therewas a significant difference between control and the LPA-treated groups at 5min 10min 30min 1 hour and 2 hours
(Figure 6(b)) These results indicated that LPA protectioneffect on the apoptosis induced by cisplatin may be relatedto the phosphorylation of AKT
36 Effects of LPA on the Phosphorylation of ERK SinceMAPK inhibitor PD98059 significantly inhibited the pro-liferation stimulated by LPA which suggested ERKMEKMAPK pathway was involved in the LPA functions in theproliferation in Hela cells we investigated whether LPA can
BioMed Research International 7
Fas-1
120573-actin
Fas-1 expression5
4
3
2
1
0
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(a)
Bcl-2
120573-actin
Bcl-2 expression4
3
2
1
0
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(b)
4
3
2
1
0
Bax
120573-actin
Bax expression
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(c)
Figure 4 LPA effects on the expression of apoptotic proteins induced by cisplatin Hela cells were treated with cisplatin (at 200120583gmL) aloneor plus LPA (at 20 120583M) for 4 hours Western blotting was used to determine the expression of apoptosis related proteins 120573-actin was used as aprotein loading control (a) Western blotting of Fas-L The increased Fas-L expression induced by cisplatin was significantly reversed by LPAtreatment (b) Western blotting of Bcl-2 LPA restored the Bcl-2 expression which was inhibited by cisplatin (c) Western blotting of BaxTheupregulation of Bax expression induced by cisplatin was significantly inhibited by LPA
activate the phosphorylation of ERKThe results showed thatLPA significantly activated the phosphorylation of ERK ina dose-dependent manner Cells were treated with LPA atdifferent concentrations including 0120583M 01 120583M 1 120583M 5 120583M20120583M and 50120583M for 1 hour The effect on ERK phosphory-lation of LPA was increased along with the increase of LPAconcentration (Figure 7(a))
LPA also activated ERK in a time-dependent mannerCells were treated by 20 120583M LPA at different time pointsincluding 0min 5min 10min 30min 1 hour 2 hours 3hours and 4 hours ERK phosphorylation reached the peakat 3 hours following LPA treatment (Figure 7(b))
4 Discussion
In the intrinsic or mitochondrial pathway of apoptosis Bcl-2is able to prevent cell apoptosis while a related protein Baxcan migrate to the surface of the mitochondrion where itinhibits the protective effect of Bcl-2 by interrupting themembrane structure causing the release of cytochrome Cand caspase activation In the extrinsic or death receptorpathway of apoptosis Fas and the TNF receptor are integralmembrane proteins receiving apoptotic signals Uponbinding by ligand they transmit signals to the cytoplasmwhich leads to activation of caspase and phagocytosis
8 BioMed Research International
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
Control LPAPI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + PD98069
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + LY294002
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + Y-27632
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
(a)
Con
trol
LPA+
PD98069
LPA+
LY294002
LPA+
Y-27632
LPA
Apop
tosis
rate
0
5
10
15
20
25
30
35
40
45
50
(b)
Figure 5 Flow cytometry detection of cell apoptosis by Annexin VPI staining (a) Hela cells were treated with cisplatin (200120583gmL) andLPA (20 120583M) alone or in combination with different inhibitors for 4 hours Cisplatin and LPA were used in each inhibitors-treated groupInhibitors including PD98059 (MAPK inhibitor at 10 120583M) PD98059 (MAPK inhibitor at 10 120583M) and Y-27632 (Rho inhibitor at 10 120583M)wereapplied respectively (b) The apoptosis rate of Hela cells was quantified Cisplatin-treated group served as control PI3K inhibitor LY294002significantly reversed the LPA effect on apoptosis induced by cisplatin while MAPK inhibitor PD98059 and Rho inhibitor Y-27632 had nosignificant effect
BioMed Research International 9
Fold
chan
ge o
f ban
d in
tens
ity
5
4
3
2
1
00 01 1 5 20 50
LPA (120583M)
0 01 1 5 20 50 LPA (120583M)
P-AKT
AKT
(a)
Fold
chan
ge o
f ban
d in
tens
ity
P-AKT
Total-AKT
4
35
3
25
2
15
1
05
00 5min 10min 30min 1h 2h 3h 4h
Time
0 5min 10min 30min 1h 2h 3h 4h Time
AKT activation
(b)
Figure 6 Western blotting analysis of phosphorylated AKT in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01120583M 1 120583M 5 120583M 20 120583M and 50 120583M for 1 hour Western blotting was conducted to detect the expressionof phosphorylated AKT Total AKT was detected and the results were used as protein input control Densitometry analysis was carried outfor quantification purpose LPA treatment led to increased phosphorylation of AKT in a dose-dependent manner (b) Time-response effectof LPA on the phosphorylation of AKT Hela cells were treated with LPA at 20 120583M for different times including 5min 10min 30min 1 hour2 hours 3 hours and 4 hours Western blotting was used to detect the expression of phosphorylated AKT Compared to the total AKT LPAtreatment activated the phosphorylation of AKT in a time-dependent manner
25
20
15
10
05
00
Fold
chan
ge
0 01 10 50 20 50
LPA (120583M)
0 01 10 05 20 50 LPA (120583M)
P-ERK
ERK
(a)
25
20
15
10
05
00
Fold
chan
ge
0 min5 min10 min30 1h 2h 3h 4hTime
0 min5 min10 min30 1h 2h 3h 4h Time
P-ERK
ERK
(b)
Figure 7 Western blotting analysis of phosphorylated ERK in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01 120583M 1 120583M 5120583M 20 120583M and 50 120583M for 1 hour Western blotting was performed to detect the levels ofphosphorylated ERK Total ERK was used as protein input control The density of bands was quantified and compared LPA activated thephosphorylation of ERK in a dose-dependent manner (b) Hela cells were treated with LPA at 20120583M for different times including 5min10min 30min 1 hour 2 hours 3 hours and 4 hours A time-dependent increase in the ERK phosphorylation was detected
10 BioMed Research International
Some studies found that cell apoptosis induced by cis-platinwasmediated by death receptors signaling pathway [2021] while others claimed that it was mediated by mitochon-drial pathway [22 23] The discrepancy in previous studiesmay be caused by the divergent cell lines methodologiesand conditions used by different groups In this study weobserved that cisplatin significantly induced the apoptosis ofHela cells in a concentration-dependent manner Cisplatintreatment was accompanied by increased Fas-1 and Baxexpression and the increased enzyme activity of caspase-3 Meanwhile cisplatin inhibited Bcl-2 expression Theseobservations suggested that cisplatin-induced apoptosis ofHela cell may be mediated by both intrinsic and extrinsicpathways Indeed extensive cross talks between the two path-ways have been identified [24 25] Simultaneous activationof the two pathways has been observed in many types of cellsfollowing various treatments
Our previous study found that both ovarian cancer cellsand peritoneal mesothelial cells contribute to the elevatedLPA levels in ovarian cancer ascites Under normal cultureconditions ovarian cancer cells and prostate cancer cellsbut not breast cancer cell lines constitutively synthesize andrelease significant amounts of LPA to the culture medium Amajor finding of this study is that LPA was able to rescue theHela cells apoptosis induced by cisplatin The protective roleof LPA against apoptosis was observed in ovarian cancer cells[26]We observed that LPA treatment blocked the alterationsof Bcl-2 Fas-1 and Bax expression that were induced bycisplatin LPA also significantly inhibited the upregulationof the caspase-3 enzyme activity induced by cisplatin Ourobservations of LPA effect on cervical cancer were consistentwith other reports on LPA effects in different kinds of cellsMeng et al found that LPA may protect epithelial ovariancancer from immune cell attack and cytoskeleton disruptingreagents induced apoptosis through multiple pathways andLPA inhibited anti-Fas-induced apoptosis enhanced by actindepolymerization [26 27] Actin depolymerization acceler-ated caspase-8 activation while LPA inhibited the associationand activation of caspase-8 at the DISC in epithelial ovariancancer OVCAR3 cells Thus our observations have raisedan interesting question on the potential role of LPA forthe development of chemoresistance by cervical cancer cellsIn addition could LPA be an effective biomarker for theprediction of chemoresistance in cervical cancer Furtherstudies are required to answer these questions
In this study three specific kinase inhibitors were appliedto determine the signaling pathways involved in the pro-tection effect of LPA on apoptosis It was found that PI3Kinhibitor (LY290042) almost completely blocked the LPAprotection against apoptosis induced by cisplatin Subsequentexperiments confirmed that LPA treatment of Hela cells ledto increased phosphorylation of AKT in a dose- and time-dependent manner This indicated that LPA protected Helacells from apoptosis induced by cisplatin This effect of LPAmay be through the activation of PI3KAKT pathway Thisfinding appears to be consistent with published data showingthat PI3KAKT pathway plays a key role in the regulationof cell apoptosis [28 29] Wan et al observed that PI3Kinhibition augmented staurosporine-induced apoptosis in
the endometrial carcinoma cell line [28] In that experimentstaurosporine reduced the levels of phosphorylation in AKTand Bad In a study by Kim et al 24-bis(p-hydroxyphenyl)-2-butenal (HPB242) induced apoptosis via the inhibition ofPI3KAKT pathway in human cervical cancer cells HPB242treatment decreased phosphatidyl inositol 3-kinase and p-AKT expression levels demonstrating that this survivalpathway may also be inhibited by HPB242 [30] In anotherstudy Cui et al found that MiR-125b inhibited tumor growthand promoted apoptosis of cervical cancer cells by targetingphosphoinositide 3-kinase catalytic subunit delta Overex-pression of MiR-125b in Hela cervical cancer cells inducedapoptosis and downregulated the expression of PIK3CDand Phospho-AKT [31] Kang et al found that thioridazineinduces apoptosis by targeting the PI3KAKTmTOR path-way in cervical cancer cells [32] Thanaketpaisarn et alfound that Artesunate enhanced TRAIL-induced apoptosisin human cervical carcinoma cells through inhibition of thePI3KAKT and NF-120581B signaling pathways [33] Wang et alfound that Stathmin is involved in arsenic trioxide-inducedapoptosis in human cervical cancer cell lines via PI3K linkedsignal pathway As2O3-induced stathmin downregulation ismediated through the phosphatidylinositol-3-kinase (PI3K)signaling pathway PI3K inhibitor effectively attenuated thestathmin downregulation and cell apoptosis upon As2O3-treatment [34] Recurrent and metastatic cervical cancersoften acquire chemoresistance to cisplatin Our findingpointed to a possible sensitization scheme through combinedadministration of PI3K inhibitor and cisplatin
It should be pointed out that while our current studyfocused on the apoptotic pathways andmechanisms the LPAeffects on cell cycle control and cell proliferation should notbe excluded In fact the PI3KAKT pathway is known to playan active role in the control of cell cycle [35] We found thatMEK1 inhibitor (PD98059) significantly inhibited the effectof LPA on the proliferation of Hela cells RasRaf1MEKERKsignal transduction pathway is one of the most importantmembers of the mitogen-activated protein kinase pathwaysregulating cell proliferation [36] MEK1 a MAPK representsa key molecule of the RasRaf1MEKERK signal pathway[37] Several lines of evidence suggest that in cervical cancerthe RasRaf1MEKERK pathway but not the JNK pathwayor the p38 MAPK pathway is the major regulator of cellproliferation [38ndash40]
In summary we demonstrated that cisplatin inducedapoptosis of Hela cell via the downregulation of Bcl-2 andupregulation of Bax Fas-L and increased enzyme activityof caspase-3 LPA was able to block the alterations in theapoptotic factors caused by cisplatin and ultimately reversedthe apoptosis induced by cisplatin Cisplatin-induced cellapoptosis appeared to be partially dependent on PI3KAKTpathway and AKT inhibitor was able to reverse the pro-tective effects of LPA against cisplatin-induced apoptosisLPA stimulated the proliferation of Hela cells throughthe RasRaf1MEKERK pathway These data enriched ourknowledge regarding the LPA activity in cervical cancer cellsIn addition the finding on the positive impact of PI3KAKTinhibitor on the apoptotic effects of cisplatin has provideduseful information for the design of sensitization strategy in
BioMed Research International 11
cervical cancer patients especially those with high levels ofLPA
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Yanxia Sui and Ya Yang contributed equally to this work
Acknowledgments
This study is supported by the following funding sourcesThe National Natural Science Foundations of China (JuanRen 31201060C0709 Juan Ren 30973175H1621 and JuanRen 81172490H1621) Program for New Century ExcellentTalents in University (Juan Ren NCET-12-0440) Scientificand Technological Research Foundation of Shaanxi Province(Juan Ren 2012K13-01-06 Juan Ren 2007K09-09) Scien-tific Research Foundation for the Returned overseas Chi-nese Scholars of State Education Ministry (Juan Ren 0601-18920006) Research Foundation of Health Department ofShaanrsquoxi Province (Juan Ren 2010D41) Qing Nian Jiao ShiGenZongJiHua ofXirsquoan JiaotongUniversity (JuanRen 2012)Program for Changjiang Scholars and Innovative ResearchTeam inUniversity (Zongfang Li PCSIRT1171) andResearchFoundation of Xirsquoan Jiao Tong University of China (JuanRen)
References
[1] J R Erickson Y Hasegawa X Fang et al ldquoLysophosphatidicacid and ovarian cancer a paradigm for tumorogenesis andpatient managementrdquo Prostaglandins and Other Lipid Media-tors vol 64 no 1ndash4 pp 63ndash81 2001
[2] Y Xu Z Z Shen DWWiper et al ldquoLysophosphatidic acid as apotential biomarker for ovarian and other gynecologic cancersrdquoThe Journal of the American Medical Association vol 280 no 8pp 719ndash723 1998
[3] J S Karliner ldquoLysophospholipids and the cardiovascular sys-temrdquoBiochimica et Biophysica ActamdashMolecular and Cell Biologyof Lipids vol 1582 no 1ndash3 pp 216ndash221 2002
[4] C Schulze C Smales L L Rubin and J M StaddonldquoLysophosphatidic acid increases tight junction permeability incultured brain endothelial cellsrdquo Journal of Neurochemistry vol68 no 3 pp 991ndash1000 1997
[5] I Ishii N Fukushima X Ye and J Chun ldquoLysophospholipidreceptors signaling and biologyrdquo Annual Review of Biochem-istry vol 73 pp 321ndash354 2004
[6] T S Panetti ldquoDifferential effects of sphingosine 1-phosphateand lysophosphatidic acid on endothelial cellsrdquo Biochimica etBiophysica ActamdashMolecular and Cell Biology of Lipids vol 1582no 1ndash3 pp 190ndash196 2002
[7] Z Z Shen M Z Wu P Elson et al ldquoFatty acid composition oflysophosphatidic acid and lysophosphatidylinositol in plasmafrom patients with ovarian cancer and other gynecologicaldiseasesrdquo Gynecologic Oncology vol 83 no 1 pp 25ndash30 2001
[8] M M Murph ldquoAgonist-induced endocytosis of lysophospha-tidic acid-coupled LPA1EDG-2 receptors via a dynamin2- andRab5-dependent pathwayrdquo Journal of Cell Science vol 116 no10 pp 1969ndash1980 2003
[9] L M Baudhuin K L Cristina J Lu and Y Xu ldquoAkt activationinduced by lysophosphatidic acid and sphingosine-1-phosphaterequires both mitogen-activated protein kinase kinase andp38 mitogen-activated protein kinase and is cell-line specificrdquoMolecular Pharmacology vol 62 no 3 pp 660ndash671 2002
[10] R-J Chen S-U Chen C-H Chou andM-C Lin ldquoLysophos-phatidic acid receptor 23-mediated IL-8-dependent angiogen-esis in cervical cancer cellsrdquo International Journal of Cancer vol131 no 4 pp 789ndash802 2012
[11] J W Choi S Lim Y-S Oh et al ldquoSubtype-specific role ofphospholipase C-120573 in bradykinin and LPA signaling throughdifferential binding of different PDZ scaffold proteinsrdquo CellularSignalling vol 22 no 7 pp 1153ndash1161 2010
[12] Y Kishi S Okudaira M Tanaka et al ldquoAutotaxin is over-expressed in glioblastoma multiforme and contributes to cellmotility of glioblastoma by converting lysophosphatidylcholineTO lysophosphatidic acidrdquoThe Journal of Biological Chemistryvol 281 no 25 pp 17492ndash17500 2006
[13] A Eastman ldquoThe formation isolation and characterization ofDNA adducts produced by anticancer platinum complexesrdquoPharmacology andTherapeutics vol 34 no 2 pp 155ndash166 1987
[14] K Wozniak and J Błasiak ldquoRecognition and repair of DNA-cisplatin adductsrdquo Acta Biochimica Polonica vol 49 no 3 pp583ndash596 2002
[15] A Basu and S Krishnamurthy ldquoCellular responses to cisplatin-induced DNA damagerdquo Journal of Nucleic Acids vol 2010Article ID 201367 16 pages 2010
[16] A Efeyan and M Serrano ldquop53 guardian of the genome andpoliceman of the oncogenesrdquo Cell Cycle vol 6 no 9 pp 1006ndash1010 2007
[17] O Kutuk E D Arisan T Tezil M C Shoshan and H BasagaldquoCisplatin overcomesBcl-2-mediated resistance to apoptosis viapreferential engagement of Bak critical role of Noxa-mediatedlipid peroxidationrdquo Carcinogenesis vol 30 no 9 pp 1517ndash15272009
[18] C M Yan E Q Chai H Y Cai G Miao and W MaldquoOleuropein induces apoptosis via activation of caspases andsuppression of phosphatidylinositol 3-kinaseprotein kinaseB pathway in HepG2 human hepatoma cell linerdquo MolecularMedicine Reports vol 11 pp 4617ndash4624 2015
[19] X Y Wang X S Fan L Cai S Liu X F Cong and X ChenldquoLysophosphatidic acid rescues bone mesenchymal stem cellsfrom hydrogen peroxide-induced apoptosisrdquo Apoptosis vol 20no 3 pp 273ndash284 2015
[20] A Brozovic G Fritz M Christmann et al ldquoLong-term activa-tion of SAPKJNK p38 kinase and fas-L expression by cisplatinis attenuated in human carcinoma cells that acquired drugresistancerdquo International Journal of Cancer vol 112 no 6 pp974ndash985 2004
[21] Y Q Liu H Xing X B Han et al ldquoApoptosis of HeLa cellsinduced by cisplatin and its mechanismrdquo Journal of HuazhongUniversity of Science and TechnologymdashMedical Science vol 28no 2 pp 197ndash199 2008
[22] K L Jin J-Y Park E J Noh et al ldquoThe effect of combinedtreatment with cisplatin and histone deacetylase inhibitors onHeLa cellsrdquo Journal of Gynecologic Oncology vol 21 no 4 pp262ndash268 2010
12 BioMed Research International
[23] T Sakaeda T Nakamura M Hirai et al ldquoMDR1 up-regulatedby apoptotic stimuli suppresses apoptotic signalingrdquo Pharma-ceutical Research vol 19 no 9 pp 1323ndash1329 2002
[24] K Kandasamy S M Srinivasula E S Alnemri et al ldquoInvolve-ment of proapoptotic molecules Bax and Bak in tumor necro-sis factor-related apoptosis-inducing ligand (TRAIL)-inducedmitochondrial disruption and apoptosis differential regulationof cytochrome c and SmacDIABLO releaserdquo Cancer Researchvol 63 no 7 pp 1712ndash1721 2003
[25] H Toyota N Yanase T Yoshimoto M Moriyama T Sudoand J Mizuguchi ldquoCalpain-induced Bax-cleavage product is amore potent inducer of apoptotic cell death thanwild-type BaxrdquoCancer Letters vol 189 no 2 pp 221ndash230 2003
[26] Y R Meng S J Kang and D A Fishman ldquoLysophosphatidicacid inhibits anti-Fas-mediated apoptosis enhanced by actindepolymerization in epithelial ovarian cancerrdquo FEBS Lettersvol 579 no 5 pp 1311ndash1319 2005
[27] Y Meng S Kang J So S Reierstad and D A FishmanldquoTranslocation of Fas by LPA prevents ovarian cancer cells fromanti-Fas-induced apoptosisrdquo Gynecologic Oncology vol 96 no2 pp 462ndash469 2005
[28] X Wan Y Yokoyama A Shinohara Y Takahashi and TTamaya ldquoPTEN augments staurosporine-induced apoptosis inPTEN-null Ishikawa cells by downregulating PI3KAkt signal-ing pathwayrdquo Cell Death and Differentiation vol 9 no 4 pp414ndash420 2002
[29] J Yu Y Zhang J Mcilroy T Rordorf-Nikolic G A Orr and JM Backer ldquoRegulation of the p85p110 phosphatidylinositol 31015840-kinase stabilization and inhibition of the p110120572 catalytic subunitby the p85 regulatory subunitrdquoMolecular and Cellular Biologyvol 18 no 3 pp 1379ndash1387 1998
[30] M S Kim J H Kim Y Bak et al ldquo24-bis (p-hydroxyphenyl)-2-butenal (HPB242) induces apoptosis via modulating E7expression and inhibition of PI3KAkt pathway in SiHa humancervical cancer cellsrdquo Nutrition and Cancer vol 64 no 8 pp1236ndash1244 2012
[31] F Cui X L Li X Y Zhu et al ldquoMiR-125b inhibits tumorgrowth and promotes apoptosis of cervical cancer cells bytargeting phosphoinositide 3-kinase catalytic subunit deltardquoCellular Physiology and Biochemistry vol 30 no 5 pp 1310ndash1318 2012
[32] S Kang S M Dong B-R Kim et al ldquoThioridazine inducesapoptosis by targeting the PI3KAktmTOR pathway in cervicaland endometrial cancer cellsrdquo Apoptosis vol 17 no 9 pp 989ndash997 2012
[33] O Thanaketpaisarn P Waiwut H Sakurai and I Saiki ldquoArte-sunate enhances TRAIL-induced apoptosis in human cervicalcarcinoma cells through inhibition of the NF-120581B and PI3KAktsignaling pathwaysrdquo International Journal of Oncology vol 39no 1 pp 279ndash285 2011
[34] XWang J-H Ren F Lin et al ldquoStathmin is involved in arsenictrioxide-induced apoptosis in human cervical cancer cell linesvia PI3K linked signal pathwayrdquo Cancer Biology and Therapyvol 10 no 6 pp 632ndash643 2010
[35] C W Lee N H Kim H K Choi et al ldquoLysophosphatidicacid-induced c-fos up-regulation involves cyclic AMP responseelement-binding protein activated by mitogen- and stress-activated protein kinase-1rdquo Journal of Cellular Biochemistry vol104 no 3 pp 785ndash794 2008
[36] DW Hommes M P Peppelenbosch and S J H van DeventerldquoMitogen activated protein (MAP) kinase signal transduction
pathways and novel anti-inflammatory targetsrdquoGut vol 52 no1 pp 144ndash151 2003
[37] M Mukai F Imamura M Ayaki et al ldquoInhibition of tumorinvasion andmetastasis by a novel lysophosphatidic acid (cyclicLPA)rdquo International Journal of Cancer vol 81 no 6 pp 918ndash9221999
[38] E-Y Huang Y-F Chen Y-M Chen et al ldquoA novel radiore-sistant mechanism of galectin-1 mediated by H-Ras-dependentpathways in cervical cancer cellsrdquo Cell Death and Disease vol3 no 1 article e251 2012
[39] W-H Su P-C Chuang E-Y Huang and K D YangldquoRadiation-induced increase in cell migration and metastaticpotential of cervical cancer cells operates via theK-ras pathwayrdquoThe American Journal of Pathology vol 180 no 2 pp 862ndash8712012
[40] H Rincon-Arano R Rosales N Mora A Rodriguez-Castaneda and C Rosales ldquoR-Ras promotes tumor growthof cervical epithelial cellsrdquo Cancer vol 97 no 3 pp 575ndash5852003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
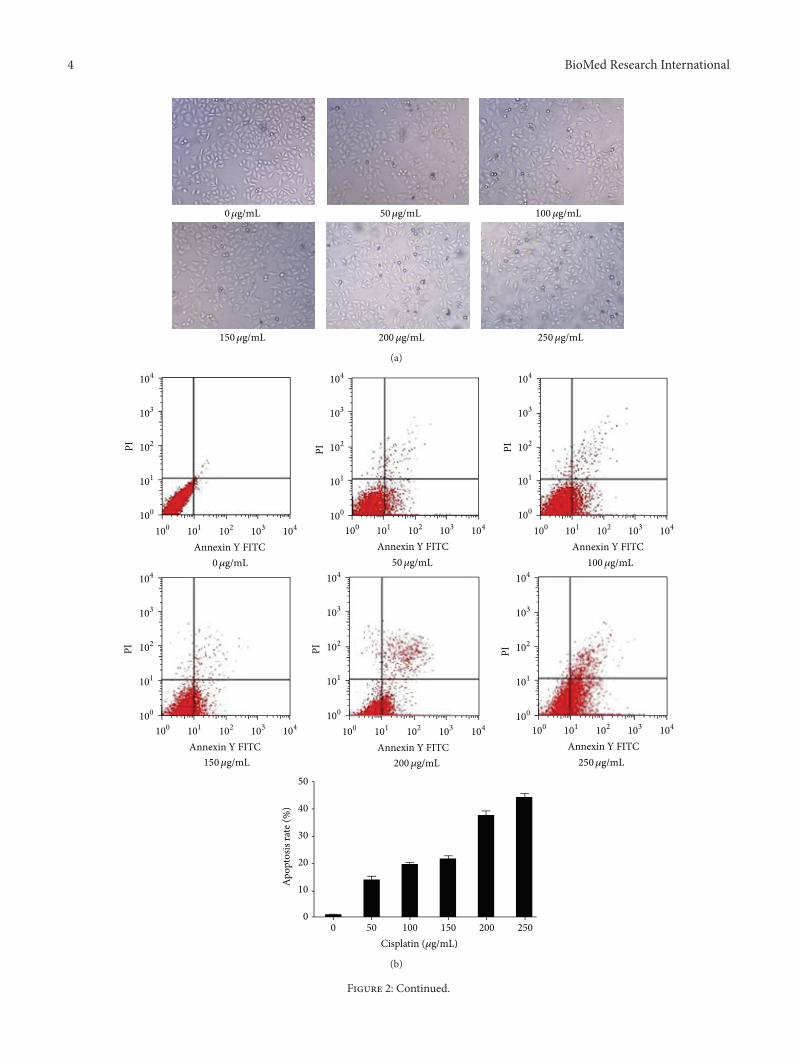
4 BioMed Research International
50120583gmL 100120583gmL
150120583gmL 200120583gmL 250120583gmL
0120583gmL
(a)
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
50120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
100120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
150120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
200120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
250120583gmL
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
0120583gmL
50
40
30
20
10
0
Apop
tosis
rate
()
2500 50 100 150 200
Cisplatin (120583gmL)
(b)
Figure 2 Continued
BioMed Research International 5
50
40
30
20
10
0
Apop
tosis
rate
()
0 50 100 150 200 250
LPA (120583M)
⋆⋆
⋆
⋆
(c)
60
40
20
0
Caspase-3 activity
pNA
den
sity
(120583M
mL)
Con
trol
Gro
up
Cisp
latin
LPA
+ ci
splat
in
⋆
(d)
Figure 2 LPA effects on apoptosis induced by cisplatin in Hela cell (a) Cell pictures (times20) Hela cells were treated with cisplatin at differentdoses After 8 hours of treatment with cisplatin many floating cells were observed The bigger the dose of cisplatin the more the floatingcells (b) Flow cytometry detection of apoptosis with Annexin VPI staining Hela cells were treated with cisplatin at different doses for 4hours Apoptotic cells were detected and quantified with flow cytometry Increased apoptosis following cisplatin treatment was observed (c)Quantification of LPA effects on apoptosis induced by cisplatin Hela cells were treated with cisplatin (at 200 120583gmL) and LPA (at differentconcentrations) for 4 hours The apoptotic cells were detected by flow cytometry and the apoptosis rate of Hela cells was quantified andshown Cell apoptosis induced by cisplatin was significantly inhibited by LPA (d) Caspase-3 activity Hela cells were treated with 200120583gmLof cisplatin plus LPA at 20 120583M for 4 hours The caspase-3 activity was measured and shown Cisplatin dramatically increased the caspase-3activity but the upregulated activity of caspase-3 was partially reversed by LPA treatment
In caspase-3 activity assay Hela cells were treated with200120583gmL of cisplatin alone or plus 20 120583M of LPA for 4hours It was found that cisplatin treatment significantlyincreased the caspase-3 activity but the upregulation ofcaspase-3 activitywas partially reversed by LPA (Figure 2(d))
33 LPA Effects on the Expression of Apoptosis Proteins Sincecisplatin induced apoptosis in cervical cancer Hela cell wefurther characterized alterations of key proteins involvedin the apoptotic pathway Western blotting showed thatthe expression of Fas-1 was significantly increased by cis-platin treatment in a dose-dependent manner (Figure 3(a))Expression of the antiapoptotic protein Bcl-2 was signif-icantly inhibited by cisplatin Moreover the inhibition ofBcl-2 expression was dependent on the dose of cisplatin(Figure 3(b)) Expression of another important apoptoticprotein Bax was also significantly increased by cisplatintreatment (Figure 3(c)) Since LPA appeared to rescue Helacells from apoptosis induced by cisplatin we examinedthe LPA effect on the change of apoptosis-related proteinsinduced by cisplatin Western blotting showed that theincrease of Fas-1 expression induced by cisplatin was signifi-cantly inhibited by LPA (Figure 4(a)) Moreover LPA treat-ment restored the inhibition of Bcl-2 expression by cisplatin(Figure 4(b)) in comparison to cells treated with cisplatinalone (Figure 2(b)) In addition LPA significantly inhibitedthe increased Bax expression induced by cisplatin Theincrease of Bax expression induced by cisplatin was signifi-cantly inhibited by LPA (Figure 4(c)) These results indicated
that LPA was able to block the apoptotic effects of cisplatinby reversing the changes in apoptotic proteins caused bycisplatin treatment
34 Signaling Transduction Pathways Involved in the LPAInhibition of Apoptosis Induced by Cisplatin Since severalstudies have shown cell apoptosis is associated with theactivation of PI3KAKT RasMAPK andRHOGEFsRHOApathways we applied specific inhibitors specifically targetingthese pathways to elucidate the mechanism underlying theLPA antiapoptotic activity Cells were treated by 200120583gmLcisplatin alone or plus 20120583M LPA and additionally dif-ferent inhibitors including PD98059 (MAPK inhibitor at10 120583M) LY294002 (PI3K inhibitor at 10 120583M) or Y-27632(Rho inhibitor at 10 120583M) It was found that the PI3K inhibitorLY294002 significantly reversed the protection effect of LPAon apoptosis induced by cisplatin (119901 = 0003) while MAPKinhibitor PD98059 and Rho inhibitor Y-27632 had no signif-icant effect (Figures 5(a) and 5(b)) These results suggestedthat the LPA effect on apoptosis induced by cisplatin werelikely mediated by the PI3KAKT signaling pathway but notRasMAPK and RHOGEFsRHOA pathways
35 Effects of LPA on the Phosphorylation of AKT Since AKTinhibitor significantly reversed the LPA protection of cellapoptosis caused by cisplatin we subsequently investigatedif LPA could activate the PI3KAKT pathway by increasingthe phosphorylation of AKT Cells were treated by LPA atdifferent concentrations including 0120583M 01 120583M 1 120583M 5 120583M
6 BioMed Research International
10
8
6
4
2
00 50 100 150 200 250
0 50 100 150 200 250
Fas-1
120573-actinCisplatin (120583gmL)
Cisplatin (120583gmL)
Fas-1 expression
Band
inte
nsity
(a)
0 50 100 150 200 250
Cisplatin (120583gmL)
0 50 100 150 200 250
Bcl-2
120573-actin
Cisplatin (120583gmL)
Bcl-2 expression3
2
1
0
Band
inte
nsity
(b)
5
4
3
2
1
0
0 50 100 150 200 250
0 50 100 150 200 250
Bax
120573-actin
Cisplatin (120583gmL)
Cisplatin (120583gmL)
Bax expression
Band
inte
nsity
(c)
Figure 3Western blotting analysis of apoptotic proteins in cells treated with cisplatin Hela cells were treated with cisplatin at different dosesincluding 050 120583gmL 100 120583gmL 150 120583gmL 200120583gmL and 250 120583gmL for 4 hours 120573-actin was used as a protein loading control Theresultant bandswere subject to densitometry analysis for quantification (a)Western blotting of Fas-LThe expression of Fas-Lwas significantlyinduced by cisplatin in a dose-dependentmanner (b)Western blotting of Bcl-2The expression of Bcl-2 was significantly inhibited by cisplatinin a dose-dependent manner (c) Western blotting of Bax The expression of Bax was significantly induced by cisplatin in a dose-dependentmanner
20120583M and 50120583Mof LPA for 1 hour (Figure 6(a))The resultsshowed that LPA treatment led to an increased phosphoryla-tion of AKT in a dose-dependent manner In addition LPA-mediated activation of AKT was also characterized by thetime-dependentmannerWhen cells were treatedwith 20120583Mof LPA for different time AKT phosphorylation graduallyincreased reaching the peak at the 2-hour time point Therewas a significant difference between control and the LPA-treated groups at 5min 10min 30min 1 hour and 2 hours
(Figure 6(b)) These results indicated that LPA protectioneffect on the apoptosis induced by cisplatin may be relatedto the phosphorylation of AKT
36 Effects of LPA on the Phosphorylation of ERK SinceMAPK inhibitor PD98059 significantly inhibited the pro-liferation stimulated by LPA which suggested ERKMEKMAPK pathway was involved in the LPA functions in theproliferation in Hela cells we investigated whether LPA can
BioMed Research International 7
Fas-1
120573-actin
Fas-1 expression5
4
3
2
1
0
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(a)
Bcl-2
120573-actin
Bcl-2 expression4
3
2
1
0
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(b)
4
3
2
1
0
Bax
120573-actin
Bax expression
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(c)
Figure 4 LPA effects on the expression of apoptotic proteins induced by cisplatin Hela cells were treated with cisplatin (at 200120583gmL) aloneor plus LPA (at 20 120583M) for 4 hours Western blotting was used to determine the expression of apoptosis related proteins 120573-actin was used as aprotein loading control (a) Western blotting of Fas-L The increased Fas-L expression induced by cisplatin was significantly reversed by LPAtreatment (b) Western blotting of Bcl-2 LPA restored the Bcl-2 expression which was inhibited by cisplatin (c) Western blotting of BaxTheupregulation of Bax expression induced by cisplatin was significantly inhibited by LPA
activate the phosphorylation of ERKThe results showed thatLPA significantly activated the phosphorylation of ERK ina dose-dependent manner Cells were treated with LPA atdifferent concentrations including 0120583M 01 120583M 1 120583M 5 120583M20120583M and 50120583M for 1 hour The effect on ERK phosphory-lation of LPA was increased along with the increase of LPAconcentration (Figure 7(a))
LPA also activated ERK in a time-dependent mannerCells were treated by 20 120583M LPA at different time pointsincluding 0min 5min 10min 30min 1 hour 2 hours 3hours and 4 hours ERK phosphorylation reached the peakat 3 hours following LPA treatment (Figure 7(b))
4 Discussion
In the intrinsic or mitochondrial pathway of apoptosis Bcl-2is able to prevent cell apoptosis while a related protein Baxcan migrate to the surface of the mitochondrion where itinhibits the protective effect of Bcl-2 by interrupting themembrane structure causing the release of cytochrome Cand caspase activation In the extrinsic or death receptorpathway of apoptosis Fas and the TNF receptor are integralmembrane proteins receiving apoptotic signals Uponbinding by ligand they transmit signals to the cytoplasmwhich leads to activation of caspase and phagocytosis
8 BioMed Research International
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
Control LPAPI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + PD98069
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + LY294002
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + Y-27632
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
(a)
Con
trol
LPA+
PD98069
LPA+
LY294002
LPA+
Y-27632
LPA
Apop
tosis
rate
0
5
10
15
20
25
30
35
40
45
50
(b)
Figure 5 Flow cytometry detection of cell apoptosis by Annexin VPI staining (a) Hela cells were treated with cisplatin (200120583gmL) andLPA (20 120583M) alone or in combination with different inhibitors for 4 hours Cisplatin and LPA were used in each inhibitors-treated groupInhibitors including PD98059 (MAPK inhibitor at 10 120583M) PD98059 (MAPK inhibitor at 10 120583M) and Y-27632 (Rho inhibitor at 10 120583M)wereapplied respectively (b) The apoptosis rate of Hela cells was quantified Cisplatin-treated group served as control PI3K inhibitor LY294002significantly reversed the LPA effect on apoptosis induced by cisplatin while MAPK inhibitor PD98059 and Rho inhibitor Y-27632 had nosignificant effect
BioMed Research International 9
Fold
chan
ge o
f ban
d in
tens
ity
5
4
3
2
1
00 01 1 5 20 50
LPA (120583M)
0 01 1 5 20 50 LPA (120583M)
P-AKT
AKT
(a)
Fold
chan
ge o
f ban
d in
tens
ity
P-AKT
Total-AKT
4
35
3
25
2
15
1
05
00 5min 10min 30min 1h 2h 3h 4h
Time
0 5min 10min 30min 1h 2h 3h 4h Time
AKT activation
(b)
Figure 6 Western blotting analysis of phosphorylated AKT in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01120583M 1 120583M 5 120583M 20 120583M and 50 120583M for 1 hour Western blotting was conducted to detect the expressionof phosphorylated AKT Total AKT was detected and the results were used as protein input control Densitometry analysis was carried outfor quantification purpose LPA treatment led to increased phosphorylation of AKT in a dose-dependent manner (b) Time-response effectof LPA on the phosphorylation of AKT Hela cells were treated with LPA at 20 120583M for different times including 5min 10min 30min 1 hour2 hours 3 hours and 4 hours Western blotting was used to detect the expression of phosphorylated AKT Compared to the total AKT LPAtreatment activated the phosphorylation of AKT in a time-dependent manner
25
20
15
10
05
00
Fold
chan
ge
0 01 10 50 20 50
LPA (120583M)
0 01 10 05 20 50 LPA (120583M)
P-ERK
ERK
(a)
25
20
15
10
05
00
Fold
chan
ge
0 min5 min10 min30 1h 2h 3h 4hTime
0 min5 min10 min30 1h 2h 3h 4h Time
P-ERK
ERK
(b)
Figure 7 Western blotting analysis of phosphorylated ERK in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01 120583M 1 120583M 5120583M 20 120583M and 50 120583M for 1 hour Western blotting was performed to detect the levels ofphosphorylated ERK Total ERK was used as protein input control The density of bands was quantified and compared LPA activated thephosphorylation of ERK in a dose-dependent manner (b) Hela cells were treated with LPA at 20120583M for different times including 5min10min 30min 1 hour 2 hours 3 hours and 4 hours A time-dependent increase in the ERK phosphorylation was detected
10 BioMed Research International
Some studies found that cell apoptosis induced by cis-platinwasmediated by death receptors signaling pathway [2021] while others claimed that it was mediated by mitochon-drial pathway [22 23] The discrepancy in previous studiesmay be caused by the divergent cell lines methodologiesand conditions used by different groups In this study weobserved that cisplatin significantly induced the apoptosis ofHela cells in a concentration-dependent manner Cisplatintreatment was accompanied by increased Fas-1 and Baxexpression and the increased enzyme activity of caspase-3 Meanwhile cisplatin inhibited Bcl-2 expression Theseobservations suggested that cisplatin-induced apoptosis ofHela cell may be mediated by both intrinsic and extrinsicpathways Indeed extensive cross talks between the two path-ways have been identified [24 25] Simultaneous activationof the two pathways has been observed in many types of cellsfollowing various treatments
Our previous study found that both ovarian cancer cellsand peritoneal mesothelial cells contribute to the elevatedLPA levels in ovarian cancer ascites Under normal cultureconditions ovarian cancer cells and prostate cancer cellsbut not breast cancer cell lines constitutively synthesize andrelease significant amounts of LPA to the culture medium Amajor finding of this study is that LPA was able to rescue theHela cells apoptosis induced by cisplatin The protective roleof LPA against apoptosis was observed in ovarian cancer cells[26]We observed that LPA treatment blocked the alterationsof Bcl-2 Fas-1 and Bax expression that were induced bycisplatin LPA also significantly inhibited the upregulationof the caspase-3 enzyme activity induced by cisplatin Ourobservations of LPA effect on cervical cancer were consistentwith other reports on LPA effects in different kinds of cellsMeng et al found that LPA may protect epithelial ovariancancer from immune cell attack and cytoskeleton disruptingreagents induced apoptosis through multiple pathways andLPA inhibited anti-Fas-induced apoptosis enhanced by actindepolymerization [26 27] Actin depolymerization acceler-ated caspase-8 activation while LPA inhibited the associationand activation of caspase-8 at the DISC in epithelial ovariancancer OVCAR3 cells Thus our observations have raisedan interesting question on the potential role of LPA forthe development of chemoresistance by cervical cancer cellsIn addition could LPA be an effective biomarker for theprediction of chemoresistance in cervical cancer Furtherstudies are required to answer these questions
In this study three specific kinase inhibitors were appliedto determine the signaling pathways involved in the pro-tection effect of LPA on apoptosis It was found that PI3Kinhibitor (LY290042) almost completely blocked the LPAprotection against apoptosis induced by cisplatin Subsequentexperiments confirmed that LPA treatment of Hela cells ledto increased phosphorylation of AKT in a dose- and time-dependent manner This indicated that LPA protected Helacells from apoptosis induced by cisplatin This effect of LPAmay be through the activation of PI3KAKT pathway Thisfinding appears to be consistent with published data showingthat PI3KAKT pathway plays a key role in the regulationof cell apoptosis [28 29] Wan et al observed that PI3Kinhibition augmented staurosporine-induced apoptosis in
the endometrial carcinoma cell line [28] In that experimentstaurosporine reduced the levels of phosphorylation in AKTand Bad In a study by Kim et al 24-bis(p-hydroxyphenyl)-2-butenal (HPB242) induced apoptosis via the inhibition ofPI3KAKT pathway in human cervical cancer cells HPB242treatment decreased phosphatidyl inositol 3-kinase and p-AKT expression levels demonstrating that this survivalpathway may also be inhibited by HPB242 [30] In anotherstudy Cui et al found that MiR-125b inhibited tumor growthand promoted apoptosis of cervical cancer cells by targetingphosphoinositide 3-kinase catalytic subunit delta Overex-pression of MiR-125b in Hela cervical cancer cells inducedapoptosis and downregulated the expression of PIK3CDand Phospho-AKT [31] Kang et al found that thioridazineinduces apoptosis by targeting the PI3KAKTmTOR path-way in cervical cancer cells [32] Thanaketpaisarn et alfound that Artesunate enhanced TRAIL-induced apoptosisin human cervical carcinoma cells through inhibition of thePI3KAKT and NF-120581B signaling pathways [33] Wang et alfound that Stathmin is involved in arsenic trioxide-inducedapoptosis in human cervical cancer cell lines via PI3K linkedsignal pathway As2O3-induced stathmin downregulation ismediated through the phosphatidylinositol-3-kinase (PI3K)signaling pathway PI3K inhibitor effectively attenuated thestathmin downregulation and cell apoptosis upon As2O3-treatment [34] Recurrent and metastatic cervical cancersoften acquire chemoresistance to cisplatin Our findingpointed to a possible sensitization scheme through combinedadministration of PI3K inhibitor and cisplatin
It should be pointed out that while our current studyfocused on the apoptotic pathways andmechanisms the LPAeffects on cell cycle control and cell proliferation should notbe excluded In fact the PI3KAKT pathway is known to playan active role in the control of cell cycle [35] We found thatMEK1 inhibitor (PD98059) significantly inhibited the effectof LPA on the proliferation of Hela cells RasRaf1MEKERKsignal transduction pathway is one of the most importantmembers of the mitogen-activated protein kinase pathwaysregulating cell proliferation [36] MEK1 a MAPK representsa key molecule of the RasRaf1MEKERK signal pathway[37] Several lines of evidence suggest that in cervical cancerthe RasRaf1MEKERK pathway but not the JNK pathwayor the p38 MAPK pathway is the major regulator of cellproliferation [38ndash40]
In summary we demonstrated that cisplatin inducedapoptosis of Hela cell via the downregulation of Bcl-2 andupregulation of Bax Fas-L and increased enzyme activityof caspase-3 LPA was able to block the alterations in theapoptotic factors caused by cisplatin and ultimately reversedthe apoptosis induced by cisplatin Cisplatin-induced cellapoptosis appeared to be partially dependent on PI3KAKTpathway and AKT inhibitor was able to reverse the pro-tective effects of LPA against cisplatin-induced apoptosisLPA stimulated the proliferation of Hela cells throughthe RasRaf1MEKERK pathway These data enriched ourknowledge regarding the LPA activity in cervical cancer cellsIn addition the finding on the positive impact of PI3KAKTinhibitor on the apoptotic effects of cisplatin has provideduseful information for the design of sensitization strategy in
BioMed Research International 11
cervical cancer patients especially those with high levels ofLPA
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Yanxia Sui and Ya Yang contributed equally to this work
Acknowledgments
This study is supported by the following funding sourcesThe National Natural Science Foundations of China (JuanRen 31201060C0709 Juan Ren 30973175H1621 and JuanRen 81172490H1621) Program for New Century ExcellentTalents in University (Juan Ren NCET-12-0440) Scientificand Technological Research Foundation of Shaanxi Province(Juan Ren 2012K13-01-06 Juan Ren 2007K09-09) Scien-tific Research Foundation for the Returned overseas Chi-nese Scholars of State Education Ministry (Juan Ren 0601-18920006) Research Foundation of Health Department ofShaanrsquoxi Province (Juan Ren 2010D41) Qing Nian Jiao ShiGenZongJiHua ofXirsquoan JiaotongUniversity (JuanRen 2012)Program for Changjiang Scholars and Innovative ResearchTeam inUniversity (Zongfang Li PCSIRT1171) andResearchFoundation of Xirsquoan Jiao Tong University of China (JuanRen)
References
[1] J R Erickson Y Hasegawa X Fang et al ldquoLysophosphatidicacid and ovarian cancer a paradigm for tumorogenesis andpatient managementrdquo Prostaglandins and Other Lipid Media-tors vol 64 no 1ndash4 pp 63ndash81 2001
[2] Y Xu Z Z Shen DWWiper et al ldquoLysophosphatidic acid as apotential biomarker for ovarian and other gynecologic cancersrdquoThe Journal of the American Medical Association vol 280 no 8pp 719ndash723 1998
[3] J S Karliner ldquoLysophospholipids and the cardiovascular sys-temrdquoBiochimica et Biophysica ActamdashMolecular and Cell Biologyof Lipids vol 1582 no 1ndash3 pp 216ndash221 2002
[4] C Schulze C Smales L L Rubin and J M StaddonldquoLysophosphatidic acid increases tight junction permeability incultured brain endothelial cellsrdquo Journal of Neurochemistry vol68 no 3 pp 991ndash1000 1997
[5] I Ishii N Fukushima X Ye and J Chun ldquoLysophospholipidreceptors signaling and biologyrdquo Annual Review of Biochem-istry vol 73 pp 321ndash354 2004
[6] T S Panetti ldquoDifferential effects of sphingosine 1-phosphateand lysophosphatidic acid on endothelial cellsrdquo Biochimica etBiophysica ActamdashMolecular and Cell Biology of Lipids vol 1582no 1ndash3 pp 190ndash196 2002
[7] Z Z Shen M Z Wu P Elson et al ldquoFatty acid composition oflysophosphatidic acid and lysophosphatidylinositol in plasmafrom patients with ovarian cancer and other gynecologicaldiseasesrdquo Gynecologic Oncology vol 83 no 1 pp 25ndash30 2001
[8] M M Murph ldquoAgonist-induced endocytosis of lysophospha-tidic acid-coupled LPA1EDG-2 receptors via a dynamin2- andRab5-dependent pathwayrdquo Journal of Cell Science vol 116 no10 pp 1969ndash1980 2003
[9] L M Baudhuin K L Cristina J Lu and Y Xu ldquoAkt activationinduced by lysophosphatidic acid and sphingosine-1-phosphaterequires both mitogen-activated protein kinase kinase andp38 mitogen-activated protein kinase and is cell-line specificrdquoMolecular Pharmacology vol 62 no 3 pp 660ndash671 2002
[10] R-J Chen S-U Chen C-H Chou andM-C Lin ldquoLysophos-phatidic acid receptor 23-mediated IL-8-dependent angiogen-esis in cervical cancer cellsrdquo International Journal of Cancer vol131 no 4 pp 789ndash802 2012
[11] J W Choi S Lim Y-S Oh et al ldquoSubtype-specific role ofphospholipase C-120573 in bradykinin and LPA signaling throughdifferential binding of different PDZ scaffold proteinsrdquo CellularSignalling vol 22 no 7 pp 1153ndash1161 2010
[12] Y Kishi S Okudaira M Tanaka et al ldquoAutotaxin is over-expressed in glioblastoma multiforme and contributes to cellmotility of glioblastoma by converting lysophosphatidylcholineTO lysophosphatidic acidrdquoThe Journal of Biological Chemistryvol 281 no 25 pp 17492ndash17500 2006
[13] A Eastman ldquoThe formation isolation and characterization ofDNA adducts produced by anticancer platinum complexesrdquoPharmacology andTherapeutics vol 34 no 2 pp 155ndash166 1987
[14] K Wozniak and J Błasiak ldquoRecognition and repair of DNA-cisplatin adductsrdquo Acta Biochimica Polonica vol 49 no 3 pp583ndash596 2002
[15] A Basu and S Krishnamurthy ldquoCellular responses to cisplatin-induced DNA damagerdquo Journal of Nucleic Acids vol 2010Article ID 201367 16 pages 2010
[16] A Efeyan and M Serrano ldquop53 guardian of the genome andpoliceman of the oncogenesrdquo Cell Cycle vol 6 no 9 pp 1006ndash1010 2007
[17] O Kutuk E D Arisan T Tezil M C Shoshan and H BasagaldquoCisplatin overcomesBcl-2-mediated resistance to apoptosis viapreferential engagement of Bak critical role of Noxa-mediatedlipid peroxidationrdquo Carcinogenesis vol 30 no 9 pp 1517ndash15272009
[18] C M Yan E Q Chai H Y Cai G Miao and W MaldquoOleuropein induces apoptosis via activation of caspases andsuppression of phosphatidylinositol 3-kinaseprotein kinaseB pathway in HepG2 human hepatoma cell linerdquo MolecularMedicine Reports vol 11 pp 4617ndash4624 2015
[19] X Y Wang X S Fan L Cai S Liu X F Cong and X ChenldquoLysophosphatidic acid rescues bone mesenchymal stem cellsfrom hydrogen peroxide-induced apoptosisrdquo Apoptosis vol 20no 3 pp 273ndash284 2015
[20] A Brozovic G Fritz M Christmann et al ldquoLong-term activa-tion of SAPKJNK p38 kinase and fas-L expression by cisplatinis attenuated in human carcinoma cells that acquired drugresistancerdquo International Journal of Cancer vol 112 no 6 pp974ndash985 2004
[21] Y Q Liu H Xing X B Han et al ldquoApoptosis of HeLa cellsinduced by cisplatin and its mechanismrdquo Journal of HuazhongUniversity of Science and TechnologymdashMedical Science vol 28no 2 pp 197ndash199 2008
[22] K L Jin J-Y Park E J Noh et al ldquoThe effect of combinedtreatment with cisplatin and histone deacetylase inhibitors onHeLa cellsrdquo Journal of Gynecologic Oncology vol 21 no 4 pp262ndash268 2010
12 BioMed Research International
[23] T Sakaeda T Nakamura M Hirai et al ldquoMDR1 up-regulatedby apoptotic stimuli suppresses apoptotic signalingrdquo Pharma-ceutical Research vol 19 no 9 pp 1323ndash1329 2002
[24] K Kandasamy S M Srinivasula E S Alnemri et al ldquoInvolve-ment of proapoptotic molecules Bax and Bak in tumor necro-sis factor-related apoptosis-inducing ligand (TRAIL)-inducedmitochondrial disruption and apoptosis differential regulationof cytochrome c and SmacDIABLO releaserdquo Cancer Researchvol 63 no 7 pp 1712ndash1721 2003
[25] H Toyota N Yanase T Yoshimoto M Moriyama T Sudoand J Mizuguchi ldquoCalpain-induced Bax-cleavage product is amore potent inducer of apoptotic cell death thanwild-type BaxrdquoCancer Letters vol 189 no 2 pp 221ndash230 2003
[26] Y R Meng S J Kang and D A Fishman ldquoLysophosphatidicacid inhibits anti-Fas-mediated apoptosis enhanced by actindepolymerization in epithelial ovarian cancerrdquo FEBS Lettersvol 579 no 5 pp 1311ndash1319 2005
[27] Y Meng S Kang J So S Reierstad and D A FishmanldquoTranslocation of Fas by LPA prevents ovarian cancer cells fromanti-Fas-induced apoptosisrdquo Gynecologic Oncology vol 96 no2 pp 462ndash469 2005
[28] X Wan Y Yokoyama A Shinohara Y Takahashi and TTamaya ldquoPTEN augments staurosporine-induced apoptosis inPTEN-null Ishikawa cells by downregulating PI3KAkt signal-ing pathwayrdquo Cell Death and Differentiation vol 9 no 4 pp414ndash420 2002
[29] J Yu Y Zhang J Mcilroy T Rordorf-Nikolic G A Orr and JM Backer ldquoRegulation of the p85p110 phosphatidylinositol 31015840-kinase stabilization and inhibition of the p110120572 catalytic subunitby the p85 regulatory subunitrdquoMolecular and Cellular Biologyvol 18 no 3 pp 1379ndash1387 1998
[30] M S Kim J H Kim Y Bak et al ldquo24-bis (p-hydroxyphenyl)-2-butenal (HPB242) induces apoptosis via modulating E7expression and inhibition of PI3KAkt pathway in SiHa humancervical cancer cellsrdquo Nutrition and Cancer vol 64 no 8 pp1236ndash1244 2012
[31] F Cui X L Li X Y Zhu et al ldquoMiR-125b inhibits tumorgrowth and promotes apoptosis of cervical cancer cells bytargeting phosphoinositide 3-kinase catalytic subunit deltardquoCellular Physiology and Biochemistry vol 30 no 5 pp 1310ndash1318 2012
[32] S Kang S M Dong B-R Kim et al ldquoThioridazine inducesapoptosis by targeting the PI3KAktmTOR pathway in cervicaland endometrial cancer cellsrdquo Apoptosis vol 17 no 9 pp 989ndash997 2012
[33] O Thanaketpaisarn P Waiwut H Sakurai and I Saiki ldquoArte-sunate enhances TRAIL-induced apoptosis in human cervicalcarcinoma cells through inhibition of the NF-120581B and PI3KAktsignaling pathwaysrdquo International Journal of Oncology vol 39no 1 pp 279ndash285 2011
[34] XWang J-H Ren F Lin et al ldquoStathmin is involved in arsenictrioxide-induced apoptosis in human cervical cancer cell linesvia PI3K linked signal pathwayrdquo Cancer Biology and Therapyvol 10 no 6 pp 632ndash643 2010
[35] C W Lee N H Kim H K Choi et al ldquoLysophosphatidicacid-induced c-fos up-regulation involves cyclic AMP responseelement-binding protein activated by mitogen- and stress-activated protein kinase-1rdquo Journal of Cellular Biochemistry vol104 no 3 pp 785ndash794 2008
[36] DW Hommes M P Peppelenbosch and S J H van DeventerldquoMitogen activated protein (MAP) kinase signal transduction
pathways and novel anti-inflammatory targetsrdquoGut vol 52 no1 pp 144ndash151 2003
[37] M Mukai F Imamura M Ayaki et al ldquoInhibition of tumorinvasion andmetastasis by a novel lysophosphatidic acid (cyclicLPA)rdquo International Journal of Cancer vol 81 no 6 pp 918ndash9221999
[38] E-Y Huang Y-F Chen Y-M Chen et al ldquoA novel radiore-sistant mechanism of galectin-1 mediated by H-Ras-dependentpathways in cervical cancer cellsrdquo Cell Death and Disease vol3 no 1 article e251 2012
[39] W-H Su P-C Chuang E-Y Huang and K D YangldquoRadiation-induced increase in cell migration and metastaticpotential of cervical cancer cells operates via theK-ras pathwayrdquoThe American Journal of Pathology vol 180 no 2 pp 862ndash8712012
[40] H Rincon-Arano R Rosales N Mora A Rodriguez-Castaneda and C Rosales ldquoR-Ras promotes tumor growthof cervical epithelial cellsrdquo Cancer vol 97 no 3 pp 575ndash5852003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 5
50
40
30
20
10
0
Apop
tosis
rate
()
0 50 100 150 200 250
LPA (120583M)
⋆⋆
⋆
⋆
(c)
60
40
20
0
Caspase-3 activity
pNA
den
sity
(120583M
mL)
Con
trol
Gro
up
Cisp
latin
LPA
+ ci
splat
in
⋆
(d)
Figure 2 LPA effects on apoptosis induced by cisplatin in Hela cell (a) Cell pictures (times20) Hela cells were treated with cisplatin at differentdoses After 8 hours of treatment with cisplatin many floating cells were observed The bigger the dose of cisplatin the more the floatingcells (b) Flow cytometry detection of apoptosis with Annexin VPI staining Hela cells were treated with cisplatin at different doses for 4hours Apoptotic cells were detected and quantified with flow cytometry Increased apoptosis following cisplatin treatment was observed (c)Quantification of LPA effects on apoptosis induced by cisplatin Hela cells were treated with cisplatin (at 200 120583gmL) and LPA (at differentconcentrations) for 4 hours The apoptotic cells were detected by flow cytometry and the apoptosis rate of Hela cells was quantified andshown Cell apoptosis induced by cisplatin was significantly inhibited by LPA (d) Caspase-3 activity Hela cells were treated with 200120583gmLof cisplatin plus LPA at 20 120583M for 4 hours The caspase-3 activity was measured and shown Cisplatin dramatically increased the caspase-3activity but the upregulated activity of caspase-3 was partially reversed by LPA treatment
In caspase-3 activity assay Hela cells were treated with200120583gmL of cisplatin alone or plus 20 120583M of LPA for 4hours It was found that cisplatin treatment significantlyincreased the caspase-3 activity but the upregulation ofcaspase-3 activitywas partially reversed by LPA (Figure 2(d))
33 LPA Effects on the Expression of Apoptosis Proteins Sincecisplatin induced apoptosis in cervical cancer Hela cell wefurther characterized alterations of key proteins involvedin the apoptotic pathway Western blotting showed thatthe expression of Fas-1 was significantly increased by cis-platin treatment in a dose-dependent manner (Figure 3(a))Expression of the antiapoptotic protein Bcl-2 was signif-icantly inhibited by cisplatin Moreover the inhibition ofBcl-2 expression was dependent on the dose of cisplatin(Figure 3(b)) Expression of another important apoptoticprotein Bax was also significantly increased by cisplatintreatment (Figure 3(c)) Since LPA appeared to rescue Helacells from apoptosis induced by cisplatin we examinedthe LPA effect on the change of apoptosis-related proteinsinduced by cisplatin Western blotting showed that theincrease of Fas-1 expression induced by cisplatin was signifi-cantly inhibited by LPA (Figure 4(a)) Moreover LPA treat-ment restored the inhibition of Bcl-2 expression by cisplatin(Figure 4(b)) in comparison to cells treated with cisplatinalone (Figure 2(b)) In addition LPA significantly inhibitedthe increased Bax expression induced by cisplatin Theincrease of Bax expression induced by cisplatin was signifi-cantly inhibited by LPA (Figure 4(c)) These results indicated
that LPA was able to block the apoptotic effects of cisplatinby reversing the changes in apoptotic proteins caused bycisplatin treatment
34 Signaling Transduction Pathways Involved in the LPAInhibition of Apoptosis Induced by Cisplatin Since severalstudies have shown cell apoptosis is associated with theactivation of PI3KAKT RasMAPK andRHOGEFsRHOApathways we applied specific inhibitors specifically targetingthese pathways to elucidate the mechanism underlying theLPA antiapoptotic activity Cells were treated by 200120583gmLcisplatin alone or plus 20120583M LPA and additionally dif-ferent inhibitors including PD98059 (MAPK inhibitor at10 120583M) LY294002 (PI3K inhibitor at 10 120583M) or Y-27632(Rho inhibitor at 10 120583M) It was found that the PI3K inhibitorLY294002 significantly reversed the protection effect of LPAon apoptosis induced by cisplatin (119901 = 0003) while MAPKinhibitor PD98059 and Rho inhibitor Y-27632 had no signif-icant effect (Figures 5(a) and 5(b)) These results suggestedthat the LPA effect on apoptosis induced by cisplatin werelikely mediated by the PI3KAKT signaling pathway but notRasMAPK and RHOGEFsRHOA pathways
35 Effects of LPA on the Phosphorylation of AKT Since AKTinhibitor significantly reversed the LPA protection of cellapoptosis caused by cisplatin we subsequently investigatedif LPA could activate the PI3KAKT pathway by increasingthe phosphorylation of AKT Cells were treated by LPA atdifferent concentrations including 0120583M 01 120583M 1 120583M 5 120583M
6 BioMed Research International
10
8
6
4
2
00 50 100 150 200 250
0 50 100 150 200 250
Fas-1
120573-actinCisplatin (120583gmL)
Cisplatin (120583gmL)
Fas-1 expression
Band
inte
nsity
(a)
0 50 100 150 200 250
Cisplatin (120583gmL)
0 50 100 150 200 250
Bcl-2
120573-actin
Cisplatin (120583gmL)
Bcl-2 expression3
2
1
0
Band
inte
nsity
(b)
5
4
3
2
1
0
0 50 100 150 200 250
0 50 100 150 200 250
Bax
120573-actin
Cisplatin (120583gmL)
Cisplatin (120583gmL)
Bax expression
Band
inte
nsity
(c)
Figure 3Western blotting analysis of apoptotic proteins in cells treated with cisplatin Hela cells were treated with cisplatin at different dosesincluding 050 120583gmL 100 120583gmL 150 120583gmL 200120583gmL and 250 120583gmL for 4 hours 120573-actin was used as a protein loading control Theresultant bandswere subject to densitometry analysis for quantification (a)Western blotting of Fas-LThe expression of Fas-Lwas significantlyinduced by cisplatin in a dose-dependentmanner (b)Western blotting of Bcl-2The expression of Bcl-2 was significantly inhibited by cisplatinin a dose-dependent manner (c) Western blotting of Bax The expression of Bax was significantly induced by cisplatin in a dose-dependentmanner
20120583M and 50120583Mof LPA for 1 hour (Figure 6(a))The resultsshowed that LPA treatment led to an increased phosphoryla-tion of AKT in a dose-dependent manner In addition LPA-mediated activation of AKT was also characterized by thetime-dependentmannerWhen cells were treatedwith 20120583Mof LPA for different time AKT phosphorylation graduallyincreased reaching the peak at the 2-hour time point Therewas a significant difference between control and the LPA-treated groups at 5min 10min 30min 1 hour and 2 hours
(Figure 6(b)) These results indicated that LPA protectioneffect on the apoptosis induced by cisplatin may be relatedto the phosphorylation of AKT
36 Effects of LPA on the Phosphorylation of ERK SinceMAPK inhibitor PD98059 significantly inhibited the pro-liferation stimulated by LPA which suggested ERKMEKMAPK pathway was involved in the LPA functions in theproliferation in Hela cells we investigated whether LPA can
BioMed Research International 7
Fas-1
120573-actin
Fas-1 expression5
4
3
2
1
0
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(a)
Bcl-2
120573-actin
Bcl-2 expression4
3
2
1
0
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(b)
4
3
2
1
0
Bax
120573-actin
Bax expression
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(c)
Figure 4 LPA effects on the expression of apoptotic proteins induced by cisplatin Hela cells were treated with cisplatin (at 200120583gmL) aloneor plus LPA (at 20 120583M) for 4 hours Western blotting was used to determine the expression of apoptosis related proteins 120573-actin was used as aprotein loading control (a) Western blotting of Fas-L The increased Fas-L expression induced by cisplatin was significantly reversed by LPAtreatment (b) Western blotting of Bcl-2 LPA restored the Bcl-2 expression which was inhibited by cisplatin (c) Western blotting of BaxTheupregulation of Bax expression induced by cisplatin was significantly inhibited by LPA
activate the phosphorylation of ERKThe results showed thatLPA significantly activated the phosphorylation of ERK ina dose-dependent manner Cells were treated with LPA atdifferent concentrations including 0120583M 01 120583M 1 120583M 5 120583M20120583M and 50120583M for 1 hour The effect on ERK phosphory-lation of LPA was increased along with the increase of LPAconcentration (Figure 7(a))
LPA also activated ERK in a time-dependent mannerCells were treated by 20 120583M LPA at different time pointsincluding 0min 5min 10min 30min 1 hour 2 hours 3hours and 4 hours ERK phosphorylation reached the peakat 3 hours following LPA treatment (Figure 7(b))
4 Discussion
In the intrinsic or mitochondrial pathway of apoptosis Bcl-2is able to prevent cell apoptosis while a related protein Baxcan migrate to the surface of the mitochondrion where itinhibits the protective effect of Bcl-2 by interrupting themembrane structure causing the release of cytochrome Cand caspase activation In the extrinsic or death receptorpathway of apoptosis Fas and the TNF receptor are integralmembrane proteins receiving apoptotic signals Uponbinding by ligand they transmit signals to the cytoplasmwhich leads to activation of caspase and phagocytosis
8 BioMed Research International
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
Control LPAPI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + PD98069
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + LY294002
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + Y-27632
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
(a)
Con
trol
LPA+
PD98069
LPA+
LY294002
LPA+
Y-27632
LPA
Apop
tosis
rate
0
5
10
15
20
25
30
35
40
45
50
(b)
Figure 5 Flow cytometry detection of cell apoptosis by Annexin VPI staining (a) Hela cells were treated with cisplatin (200120583gmL) andLPA (20 120583M) alone or in combination with different inhibitors for 4 hours Cisplatin and LPA were used in each inhibitors-treated groupInhibitors including PD98059 (MAPK inhibitor at 10 120583M) PD98059 (MAPK inhibitor at 10 120583M) and Y-27632 (Rho inhibitor at 10 120583M)wereapplied respectively (b) The apoptosis rate of Hela cells was quantified Cisplatin-treated group served as control PI3K inhibitor LY294002significantly reversed the LPA effect on apoptosis induced by cisplatin while MAPK inhibitor PD98059 and Rho inhibitor Y-27632 had nosignificant effect
BioMed Research International 9
Fold
chan
ge o
f ban
d in
tens
ity
5
4
3
2
1
00 01 1 5 20 50
LPA (120583M)
0 01 1 5 20 50 LPA (120583M)
P-AKT
AKT
(a)
Fold
chan
ge o
f ban
d in
tens
ity
P-AKT
Total-AKT
4
35
3
25
2
15
1
05
00 5min 10min 30min 1h 2h 3h 4h
Time
0 5min 10min 30min 1h 2h 3h 4h Time
AKT activation
(b)
Figure 6 Western blotting analysis of phosphorylated AKT in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01120583M 1 120583M 5 120583M 20 120583M and 50 120583M for 1 hour Western blotting was conducted to detect the expressionof phosphorylated AKT Total AKT was detected and the results were used as protein input control Densitometry analysis was carried outfor quantification purpose LPA treatment led to increased phosphorylation of AKT in a dose-dependent manner (b) Time-response effectof LPA on the phosphorylation of AKT Hela cells were treated with LPA at 20 120583M for different times including 5min 10min 30min 1 hour2 hours 3 hours and 4 hours Western blotting was used to detect the expression of phosphorylated AKT Compared to the total AKT LPAtreatment activated the phosphorylation of AKT in a time-dependent manner
25
20
15
10
05
00
Fold
chan
ge
0 01 10 50 20 50
LPA (120583M)
0 01 10 05 20 50 LPA (120583M)
P-ERK
ERK
(a)
25
20
15
10
05
00
Fold
chan
ge
0 min5 min10 min30 1h 2h 3h 4hTime
0 min5 min10 min30 1h 2h 3h 4h Time
P-ERK
ERK
(b)
Figure 7 Western blotting analysis of phosphorylated ERK in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01 120583M 1 120583M 5120583M 20 120583M and 50 120583M for 1 hour Western blotting was performed to detect the levels ofphosphorylated ERK Total ERK was used as protein input control The density of bands was quantified and compared LPA activated thephosphorylation of ERK in a dose-dependent manner (b) Hela cells were treated with LPA at 20120583M for different times including 5min10min 30min 1 hour 2 hours 3 hours and 4 hours A time-dependent increase in the ERK phosphorylation was detected
10 BioMed Research International
Some studies found that cell apoptosis induced by cis-platinwasmediated by death receptors signaling pathway [2021] while others claimed that it was mediated by mitochon-drial pathway [22 23] The discrepancy in previous studiesmay be caused by the divergent cell lines methodologiesand conditions used by different groups In this study weobserved that cisplatin significantly induced the apoptosis ofHela cells in a concentration-dependent manner Cisplatintreatment was accompanied by increased Fas-1 and Baxexpression and the increased enzyme activity of caspase-3 Meanwhile cisplatin inhibited Bcl-2 expression Theseobservations suggested that cisplatin-induced apoptosis ofHela cell may be mediated by both intrinsic and extrinsicpathways Indeed extensive cross talks between the two path-ways have been identified [24 25] Simultaneous activationof the two pathways has been observed in many types of cellsfollowing various treatments
Our previous study found that both ovarian cancer cellsand peritoneal mesothelial cells contribute to the elevatedLPA levels in ovarian cancer ascites Under normal cultureconditions ovarian cancer cells and prostate cancer cellsbut not breast cancer cell lines constitutively synthesize andrelease significant amounts of LPA to the culture medium Amajor finding of this study is that LPA was able to rescue theHela cells apoptosis induced by cisplatin The protective roleof LPA against apoptosis was observed in ovarian cancer cells[26]We observed that LPA treatment blocked the alterationsof Bcl-2 Fas-1 and Bax expression that were induced bycisplatin LPA also significantly inhibited the upregulationof the caspase-3 enzyme activity induced by cisplatin Ourobservations of LPA effect on cervical cancer were consistentwith other reports on LPA effects in different kinds of cellsMeng et al found that LPA may protect epithelial ovariancancer from immune cell attack and cytoskeleton disruptingreagents induced apoptosis through multiple pathways andLPA inhibited anti-Fas-induced apoptosis enhanced by actindepolymerization [26 27] Actin depolymerization acceler-ated caspase-8 activation while LPA inhibited the associationand activation of caspase-8 at the DISC in epithelial ovariancancer OVCAR3 cells Thus our observations have raisedan interesting question on the potential role of LPA forthe development of chemoresistance by cervical cancer cellsIn addition could LPA be an effective biomarker for theprediction of chemoresistance in cervical cancer Furtherstudies are required to answer these questions
In this study three specific kinase inhibitors were appliedto determine the signaling pathways involved in the pro-tection effect of LPA on apoptosis It was found that PI3Kinhibitor (LY290042) almost completely blocked the LPAprotection against apoptosis induced by cisplatin Subsequentexperiments confirmed that LPA treatment of Hela cells ledto increased phosphorylation of AKT in a dose- and time-dependent manner This indicated that LPA protected Helacells from apoptosis induced by cisplatin This effect of LPAmay be through the activation of PI3KAKT pathway Thisfinding appears to be consistent with published data showingthat PI3KAKT pathway plays a key role in the regulationof cell apoptosis [28 29] Wan et al observed that PI3Kinhibition augmented staurosporine-induced apoptosis in
the endometrial carcinoma cell line [28] In that experimentstaurosporine reduced the levels of phosphorylation in AKTand Bad In a study by Kim et al 24-bis(p-hydroxyphenyl)-2-butenal (HPB242) induced apoptosis via the inhibition ofPI3KAKT pathway in human cervical cancer cells HPB242treatment decreased phosphatidyl inositol 3-kinase and p-AKT expression levels demonstrating that this survivalpathway may also be inhibited by HPB242 [30] In anotherstudy Cui et al found that MiR-125b inhibited tumor growthand promoted apoptosis of cervical cancer cells by targetingphosphoinositide 3-kinase catalytic subunit delta Overex-pression of MiR-125b in Hela cervical cancer cells inducedapoptosis and downregulated the expression of PIK3CDand Phospho-AKT [31] Kang et al found that thioridazineinduces apoptosis by targeting the PI3KAKTmTOR path-way in cervical cancer cells [32] Thanaketpaisarn et alfound that Artesunate enhanced TRAIL-induced apoptosisin human cervical carcinoma cells through inhibition of thePI3KAKT and NF-120581B signaling pathways [33] Wang et alfound that Stathmin is involved in arsenic trioxide-inducedapoptosis in human cervical cancer cell lines via PI3K linkedsignal pathway As2O3-induced stathmin downregulation ismediated through the phosphatidylinositol-3-kinase (PI3K)signaling pathway PI3K inhibitor effectively attenuated thestathmin downregulation and cell apoptosis upon As2O3-treatment [34] Recurrent and metastatic cervical cancersoften acquire chemoresistance to cisplatin Our findingpointed to a possible sensitization scheme through combinedadministration of PI3K inhibitor and cisplatin
It should be pointed out that while our current studyfocused on the apoptotic pathways andmechanisms the LPAeffects on cell cycle control and cell proliferation should notbe excluded In fact the PI3KAKT pathway is known to playan active role in the control of cell cycle [35] We found thatMEK1 inhibitor (PD98059) significantly inhibited the effectof LPA on the proliferation of Hela cells RasRaf1MEKERKsignal transduction pathway is one of the most importantmembers of the mitogen-activated protein kinase pathwaysregulating cell proliferation [36] MEK1 a MAPK representsa key molecule of the RasRaf1MEKERK signal pathway[37] Several lines of evidence suggest that in cervical cancerthe RasRaf1MEKERK pathway but not the JNK pathwayor the p38 MAPK pathway is the major regulator of cellproliferation [38ndash40]
In summary we demonstrated that cisplatin inducedapoptosis of Hela cell via the downregulation of Bcl-2 andupregulation of Bax Fas-L and increased enzyme activityof caspase-3 LPA was able to block the alterations in theapoptotic factors caused by cisplatin and ultimately reversedthe apoptosis induced by cisplatin Cisplatin-induced cellapoptosis appeared to be partially dependent on PI3KAKTpathway and AKT inhibitor was able to reverse the pro-tective effects of LPA against cisplatin-induced apoptosisLPA stimulated the proliferation of Hela cells throughthe RasRaf1MEKERK pathway These data enriched ourknowledge regarding the LPA activity in cervical cancer cellsIn addition the finding on the positive impact of PI3KAKTinhibitor on the apoptotic effects of cisplatin has provideduseful information for the design of sensitization strategy in
BioMed Research International 11
cervical cancer patients especially those with high levels ofLPA
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Yanxia Sui and Ya Yang contributed equally to this work
Acknowledgments
This study is supported by the following funding sourcesThe National Natural Science Foundations of China (JuanRen 31201060C0709 Juan Ren 30973175H1621 and JuanRen 81172490H1621) Program for New Century ExcellentTalents in University (Juan Ren NCET-12-0440) Scientificand Technological Research Foundation of Shaanxi Province(Juan Ren 2012K13-01-06 Juan Ren 2007K09-09) Scien-tific Research Foundation for the Returned overseas Chi-nese Scholars of State Education Ministry (Juan Ren 0601-18920006) Research Foundation of Health Department ofShaanrsquoxi Province (Juan Ren 2010D41) Qing Nian Jiao ShiGenZongJiHua ofXirsquoan JiaotongUniversity (JuanRen 2012)Program for Changjiang Scholars and Innovative ResearchTeam inUniversity (Zongfang Li PCSIRT1171) andResearchFoundation of Xirsquoan Jiao Tong University of China (JuanRen)
References
[1] J R Erickson Y Hasegawa X Fang et al ldquoLysophosphatidicacid and ovarian cancer a paradigm for tumorogenesis andpatient managementrdquo Prostaglandins and Other Lipid Media-tors vol 64 no 1ndash4 pp 63ndash81 2001
[2] Y Xu Z Z Shen DWWiper et al ldquoLysophosphatidic acid as apotential biomarker for ovarian and other gynecologic cancersrdquoThe Journal of the American Medical Association vol 280 no 8pp 719ndash723 1998
[3] J S Karliner ldquoLysophospholipids and the cardiovascular sys-temrdquoBiochimica et Biophysica ActamdashMolecular and Cell Biologyof Lipids vol 1582 no 1ndash3 pp 216ndash221 2002
[4] C Schulze C Smales L L Rubin and J M StaddonldquoLysophosphatidic acid increases tight junction permeability incultured brain endothelial cellsrdquo Journal of Neurochemistry vol68 no 3 pp 991ndash1000 1997
[5] I Ishii N Fukushima X Ye and J Chun ldquoLysophospholipidreceptors signaling and biologyrdquo Annual Review of Biochem-istry vol 73 pp 321ndash354 2004
[6] T S Panetti ldquoDifferential effects of sphingosine 1-phosphateand lysophosphatidic acid on endothelial cellsrdquo Biochimica etBiophysica ActamdashMolecular and Cell Biology of Lipids vol 1582no 1ndash3 pp 190ndash196 2002
[7] Z Z Shen M Z Wu P Elson et al ldquoFatty acid composition oflysophosphatidic acid and lysophosphatidylinositol in plasmafrom patients with ovarian cancer and other gynecologicaldiseasesrdquo Gynecologic Oncology vol 83 no 1 pp 25ndash30 2001
[8] M M Murph ldquoAgonist-induced endocytosis of lysophospha-tidic acid-coupled LPA1EDG-2 receptors via a dynamin2- andRab5-dependent pathwayrdquo Journal of Cell Science vol 116 no10 pp 1969ndash1980 2003
[9] L M Baudhuin K L Cristina J Lu and Y Xu ldquoAkt activationinduced by lysophosphatidic acid and sphingosine-1-phosphaterequires both mitogen-activated protein kinase kinase andp38 mitogen-activated protein kinase and is cell-line specificrdquoMolecular Pharmacology vol 62 no 3 pp 660ndash671 2002
[10] R-J Chen S-U Chen C-H Chou andM-C Lin ldquoLysophos-phatidic acid receptor 23-mediated IL-8-dependent angiogen-esis in cervical cancer cellsrdquo International Journal of Cancer vol131 no 4 pp 789ndash802 2012
[11] J W Choi S Lim Y-S Oh et al ldquoSubtype-specific role ofphospholipase C-120573 in bradykinin and LPA signaling throughdifferential binding of different PDZ scaffold proteinsrdquo CellularSignalling vol 22 no 7 pp 1153ndash1161 2010
[12] Y Kishi S Okudaira M Tanaka et al ldquoAutotaxin is over-expressed in glioblastoma multiforme and contributes to cellmotility of glioblastoma by converting lysophosphatidylcholineTO lysophosphatidic acidrdquoThe Journal of Biological Chemistryvol 281 no 25 pp 17492ndash17500 2006
[13] A Eastman ldquoThe formation isolation and characterization ofDNA adducts produced by anticancer platinum complexesrdquoPharmacology andTherapeutics vol 34 no 2 pp 155ndash166 1987
[14] K Wozniak and J Błasiak ldquoRecognition and repair of DNA-cisplatin adductsrdquo Acta Biochimica Polonica vol 49 no 3 pp583ndash596 2002
[15] A Basu and S Krishnamurthy ldquoCellular responses to cisplatin-induced DNA damagerdquo Journal of Nucleic Acids vol 2010Article ID 201367 16 pages 2010
[16] A Efeyan and M Serrano ldquop53 guardian of the genome andpoliceman of the oncogenesrdquo Cell Cycle vol 6 no 9 pp 1006ndash1010 2007
[17] O Kutuk E D Arisan T Tezil M C Shoshan and H BasagaldquoCisplatin overcomesBcl-2-mediated resistance to apoptosis viapreferential engagement of Bak critical role of Noxa-mediatedlipid peroxidationrdquo Carcinogenesis vol 30 no 9 pp 1517ndash15272009
[18] C M Yan E Q Chai H Y Cai G Miao and W MaldquoOleuropein induces apoptosis via activation of caspases andsuppression of phosphatidylinositol 3-kinaseprotein kinaseB pathway in HepG2 human hepatoma cell linerdquo MolecularMedicine Reports vol 11 pp 4617ndash4624 2015
[19] X Y Wang X S Fan L Cai S Liu X F Cong and X ChenldquoLysophosphatidic acid rescues bone mesenchymal stem cellsfrom hydrogen peroxide-induced apoptosisrdquo Apoptosis vol 20no 3 pp 273ndash284 2015
[20] A Brozovic G Fritz M Christmann et al ldquoLong-term activa-tion of SAPKJNK p38 kinase and fas-L expression by cisplatinis attenuated in human carcinoma cells that acquired drugresistancerdquo International Journal of Cancer vol 112 no 6 pp974ndash985 2004
[21] Y Q Liu H Xing X B Han et al ldquoApoptosis of HeLa cellsinduced by cisplatin and its mechanismrdquo Journal of HuazhongUniversity of Science and TechnologymdashMedical Science vol 28no 2 pp 197ndash199 2008
[22] K L Jin J-Y Park E J Noh et al ldquoThe effect of combinedtreatment with cisplatin and histone deacetylase inhibitors onHeLa cellsrdquo Journal of Gynecologic Oncology vol 21 no 4 pp262ndash268 2010
12 BioMed Research International
[23] T Sakaeda T Nakamura M Hirai et al ldquoMDR1 up-regulatedby apoptotic stimuli suppresses apoptotic signalingrdquo Pharma-ceutical Research vol 19 no 9 pp 1323ndash1329 2002
[24] K Kandasamy S M Srinivasula E S Alnemri et al ldquoInvolve-ment of proapoptotic molecules Bax and Bak in tumor necro-sis factor-related apoptosis-inducing ligand (TRAIL)-inducedmitochondrial disruption and apoptosis differential regulationof cytochrome c and SmacDIABLO releaserdquo Cancer Researchvol 63 no 7 pp 1712ndash1721 2003
[25] H Toyota N Yanase T Yoshimoto M Moriyama T Sudoand J Mizuguchi ldquoCalpain-induced Bax-cleavage product is amore potent inducer of apoptotic cell death thanwild-type BaxrdquoCancer Letters vol 189 no 2 pp 221ndash230 2003
[26] Y R Meng S J Kang and D A Fishman ldquoLysophosphatidicacid inhibits anti-Fas-mediated apoptosis enhanced by actindepolymerization in epithelial ovarian cancerrdquo FEBS Lettersvol 579 no 5 pp 1311ndash1319 2005
[27] Y Meng S Kang J So S Reierstad and D A FishmanldquoTranslocation of Fas by LPA prevents ovarian cancer cells fromanti-Fas-induced apoptosisrdquo Gynecologic Oncology vol 96 no2 pp 462ndash469 2005
[28] X Wan Y Yokoyama A Shinohara Y Takahashi and TTamaya ldquoPTEN augments staurosporine-induced apoptosis inPTEN-null Ishikawa cells by downregulating PI3KAkt signal-ing pathwayrdquo Cell Death and Differentiation vol 9 no 4 pp414ndash420 2002
[29] J Yu Y Zhang J Mcilroy T Rordorf-Nikolic G A Orr and JM Backer ldquoRegulation of the p85p110 phosphatidylinositol 31015840-kinase stabilization and inhibition of the p110120572 catalytic subunitby the p85 regulatory subunitrdquoMolecular and Cellular Biologyvol 18 no 3 pp 1379ndash1387 1998
[30] M S Kim J H Kim Y Bak et al ldquo24-bis (p-hydroxyphenyl)-2-butenal (HPB242) induces apoptosis via modulating E7expression and inhibition of PI3KAkt pathway in SiHa humancervical cancer cellsrdquo Nutrition and Cancer vol 64 no 8 pp1236ndash1244 2012
[31] F Cui X L Li X Y Zhu et al ldquoMiR-125b inhibits tumorgrowth and promotes apoptosis of cervical cancer cells bytargeting phosphoinositide 3-kinase catalytic subunit deltardquoCellular Physiology and Biochemistry vol 30 no 5 pp 1310ndash1318 2012
[32] S Kang S M Dong B-R Kim et al ldquoThioridazine inducesapoptosis by targeting the PI3KAktmTOR pathway in cervicaland endometrial cancer cellsrdquo Apoptosis vol 17 no 9 pp 989ndash997 2012
[33] O Thanaketpaisarn P Waiwut H Sakurai and I Saiki ldquoArte-sunate enhances TRAIL-induced apoptosis in human cervicalcarcinoma cells through inhibition of the NF-120581B and PI3KAktsignaling pathwaysrdquo International Journal of Oncology vol 39no 1 pp 279ndash285 2011
[34] XWang J-H Ren F Lin et al ldquoStathmin is involved in arsenictrioxide-induced apoptosis in human cervical cancer cell linesvia PI3K linked signal pathwayrdquo Cancer Biology and Therapyvol 10 no 6 pp 632ndash643 2010
[35] C W Lee N H Kim H K Choi et al ldquoLysophosphatidicacid-induced c-fos up-regulation involves cyclic AMP responseelement-binding protein activated by mitogen- and stress-activated protein kinase-1rdquo Journal of Cellular Biochemistry vol104 no 3 pp 785ndash794 2008
[36] DW Hommes M P Peppelenbosch and S J H van DeventerldquoMitogen activated protein (MAP) kinase signal transduction
pathways and novel anti-inflammatory targetsrdquoGut vol 52 no1 pp 144ndash151 2003
[37] M Mukai F Imamura M Ayaki et al ldquoInhibition of tumorinvasion andmetastasis by a novel lysophosphatidic acid (cyclicLPA)rdquo International Journal of Cancer vol 81 no 6 pp 918ndash9221999
[38] E-Y Huang Y-F Chen Y-M Chen et al ldquoA novel radiore-sistant mechanism of galectin-1 mediated by H-Ras-dependentpathways in cervical cancer cellsrdquo Cell Death and Disease vol3 no 1 article e251 2012
[39] W-H Su P-C Chuang E-Y Huang and K D YangldquoRadiation-induced increase in cell migration and metastaticpotential of cervical cancer cells operates via theK-ras pathwayrdquoThe American Journal of Pathology vol 180 no 2 pp 862ndash8712012
[40] H Rincon-Arano R Rosales N Mora A Rodriguez-Castaneda and C Rosales ldquoR-Ras promotes tumor growthof cervical epithelial cellsrdquo Cancer vol 97 no 3 pp 575ndash5852003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 BioMed Research International
10
8
6
4
2
00 50 100 150 200 250
0 50 100 150 200 250
Fas-1
120573-actinCisplatin (120583gmL)
Cisplatin (120583gmL)
Fas-1 expression
Band
inte
nsity
(a)
0 50 100 150 200 250
Cisplatin (120583gmL)
0 50 100 150 200 250
Bcl-2
120573-actin
Cisplatin (120583gmL)
Bcl-2 expression3
2
1
0
Band
inte
nsity
(b)
5
4
3
2
1
0
0 50 100 150 200 250
0 50 100 150 200 250
Bax
120573-actin
Cisplatin (120583gmL)
Cisplatin (120583gmL)
Bax expression
Band
inte
nsity
(c)
Figure 3Western blotting analysis of apoptotic proteins in cells treated with cisplatin Hela cells were treated with cisplatin at different dosesincluding 050 120583gmL 100 120583gmL 150 120583gmL 200120583gmL and 250 120583gmL for 4 hours 120573-actin was used as a protein loading control Theresultant bandswere subject to densitometry analysis for quantification (a)Western blotting of Fas-LThe expression of Fas-Lwas significantlyinduced by cisplatin in a dose-dependentmanner (b)Western blotting of Bcl-2The expression of Bcl-2 was significantly inhibited by cisplatinin a dose-dependent manner (c) Western blotting of Bax The expression of Bax was significantly induced by cisplatin in a dose-dependentmanner
20120583M and 50120583Mof LPA for 1 hour (Figure 6(a))The resultsshowed that LPA treatment led to an increased phosphoryla-tion of AKT in a dose-dependent manner In addition LPA-mediated activation of AKT was also characterized by thetime-dependentmannerWhen cells were treatedwith 20120583Mof LPA for different time AKT phosphorylation graduallyincreased reaching the peak at the 2-hour time point Therewas a significant difference between control and the LPA-treated groups at 5min 10min 30min 1 hour and 2 hours
(Figure 6(b)) These results indicated that LPA protectioneffect on the apoptosis induced by cisplatin may be relatedto the phosphorylation of AKT
36 Effects of LPA on the Phosphorylation of ERK SinceMAPK inhibitor PD98059 significantly inhibited the pro-liferation stimulated by LPA which suggested ERKMEKMAPK pathway was involved in the LPA functions in theproliferation in Hela cells we investigated whether LPA can
BioMed Research International 7
Fas-1
120573-actin
Fas-1 expression5
4
3
2
1
0
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(a)
Bcl-2
120573-actin
Bcl-2 expression4
3
2
1
0
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(b)
4
3
2
1
0
Bax
120573-actin
Bax expression
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(c)
Figure 4 LPA effects on the expression of apoptotic proteins induced by cisplatin Hela cells were treated with cisplatin (at 200120583gmL) aloneor plus LPA (at 20 120583M) for 4 hours Western blotting was used to determine the expression of apoptosis related proteins 120573-actin was used as aprotein loading control (a) Western blotting of Fas-L The increased Fas-L expression induced by cisplatin was significantly reversed by LPAtreatment (b) Western blotting of Bcl-2 LPA restored the Bcl-2 expression which was inhibited by cisplatin (c) Western blotting of BaxTheupregulation of Bax expression induced by cisplatin was significantly inhibited by LPA
activate the phosphorylation of ERKThe results showed thatLPA significantly activated the phosphorylation of ERK ina dose-dependent manner Cells were treated with LPA atdifferent concentrations including 0120583M 01 120583M 1 120583M 5 120583M20120583M and 50120583M for 1 hour The effect on ERK phosphory-lation of LPA was increased along with the increase of LPAconcentration (Figure 7(a))
LPA also activated ERK in a time-dependent mannerCells were treated by 20 120583M LPA at different time pointsincluding 0min 5min 10min 30min 1 hour 2 hours 3hours and 4 hours ERK phosphorylation reached the peakat 3 hours following LPA treatment (Figure 7(b))
4 Discussion
In the intrinsic or mitochondrial pathway of apoptosis Bcl-2is able to prevent cell apoptosis while a related protein Baxcan migrate to the surface of the mitochondrion where itinhibits the protective effect of Bcl-2 by interrupting themembrane structure causing the release of cytochrome Cand caspase activation In the extrinsic or death receptorpathway of apoptosis Fas and the TNF receptor are integralmembrane proteins receiving apoptotic signals Uponbinding by ligand they transmit signals to the cytoplasmwhich leads to activation of caspase and phagocytosis
8 BioMed Research International
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
Control LPAPI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + PD98069
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + LY294002
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + Y-27632
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
(a)
Con
trol
LPA+
PD98069
LPA+
LY294002
LPA+
Y-27632
LPA
Apop
tosis
rate
0
5
10
15
20
25
30
35
40
45
50
(b)
Figure 5 Flow cytometry detection of cell apoptosis by Annexin VPI staining (a) Hela cells were treated with cisplatin (200120583gmL) andLPA (20 120583M) alone or in combination with different inhibitors for 4 hours Cisplatin and LPA were used in each inhibitors-treated groupInhibitors including PD98059 (MAPK inhibitor at 10 120583M) PD98059 (MAPK inhibitor at 10 120583M) and Y-27632 (Rho inhibitor at 10 120583M)wereapplied respectively (b) The apoptosis rate of Hela cells was quantified Cisplatin-treated group served as control PI3K inhibitor LY294002significantly reversed the LPA effect on apoptosis induced by cisplatin while MAPK inhibitor PD98059 and Rho inhibitor Y-27632 had nosignificant effect
BioMed Research International 9
Fold
chan
ge o
f ban
d in
tens
ity
5
4
3
2
1
00 01 1 5 20 50
LPA (120583M)
0 01 1 5 20 50 LPA (120583M)
P-AKT
AKT
(a)
Fold
chan
ge o
f ban
d in
tens
ity
P-AKT
Total-AKT
4
35
3
25
2
15
1
05
00 5min 10min 30min 1h 2h 3h 4h
Time
0 5min 10min 30min 1h 2h 3h 4h Time
AKT activation
(b)
Figure 6 Western blotting analysis of phosphorylated AKT in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01120583M 1 120583M 5 120583M 20 120583M and 50 120583M for 1 hour Western blotting was conducted to detect the expressionof phosphorylated AKT Total AKT was detected and the results were used as protein input control Densitometry analysis was carried outfor quantification purpose LPA treatment led to increased phosphorylation of AKT in a dose-dependent manner (b) Time-response effectof LPA on the phosphorylation of AKT Hela cells were treated with LPA at 20 120583M for different times including 5min 10min 30min 1 hour2 hours 3 hours and 4 hours Western blotting was used to detect the expression of phosphorylated AKT Compared to the total AKT LPAtreatment activated the phosphorylation of AKT in a time-dependent manner
25
20
15
10
05
00
Fold
chan
ge
0 01 10 50 20 50
LPA (120583M)
0 01 10 05 20 50 LPA (120583M)
P-ERK
ERK
(a)
25
20
15
10
05
00
Fold
chan
ge
0 min5 min10 min30 1h 2h 3h 4hTime
0 min5 min10 min30 1h 2h 3h 4h Time
P-ERK
ERK
(b)
Figure 7 Western blotting analysis of phosphorylated ERK in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01 120583M 1 120583M 5120583M 20 120583M and 50 120583M for 1 hour Western blotting was performed to detect the levels ofphosphorylated ERK Total ERK was used as protein input control The density of bands was quantified and compared LPA activated thephosphorylation of ERK in a dose-dependent manner (b) Hela cells were treated with LPA at 20120583M for different times including 5min10min 30min 1 hour 2 hours 3 hours and 4 hours A time-dependent increase in the ERK phosphorylation was detected
10 BioMed Research International
Some studies found that cell apoptosis induced by cis-platinwasmediated by death receptors signaling pathway [2021] while others claimed that it was mediated by mitochon-drial pathway [22 23] The discrepancy in previous studiesmay be caused by the divergent cell lines methodologiesand conditions used by different groups In this study weobserved that cisplatin significantly induced the apoptosis ofHela cells in a concentration-dependent manner Cisplatintreatment was accompanied by increased Fas-1 and Baxexpression and the increased enzyme activity of caspase-3 Meanwhile cisplatin inhibited Bcl-2 expression Theseobservations suggested that cisplatin-induced apoptosis ofHela cell may be mediated by both intrinsic and extrinsicpathways Indeed extensive cross talks between the two path-ways have been identified [24 25] Simultaneous activationof the two pathways has been observed in many types of cellsfollowing various treatments
Our previous study found that both ovarian cancer cellsand peritoneal mesothelial cells contribute to the elevatedLPA levels in ovarian cancer ascites Under normal cultureconditions ovarian cancer cells and prostate cancer cellsbut not breast cancer cell lines constitutively synthesize andrelease significant amounts of LPA to the culture medium Amajor finding of this study is that LPA was able to rescue theHela cells apoptosis induced by cisplatin The protective roleof LPA against apoptosis was observed in ovarian cancer cells[26]We observed that LPA treatment blocked the alterationsof Bcl-2 Fas-1 and Bax expression that were induced bycisplatin LPA also significantly inhibited the upregulationof the caspase-3 enzyme activity induced by cisplatin Ourobservations of LPA effect on cervical cancer were consistentwith other reports on LPA effects in different kinds of cellsMeng et al found that LPA may protect epithelial ovariancancer from immune cell attack and cytoskeleton disruptingreagents induced apoptosis through multiple pathways andLPA inhibited anti-Fas-induced apoptosis enhanced by actindepolymerization [26 27] Actin depolymerization acceler-ated caspase-8 activation while LPA inhibited the associationand activation of caspase-8 at the DISC in epithelial ovariancancer OVCAR3 cells Thus our observations have raisedan interesting question on the potential role of LPA forthe development of chemoresistance by cervical cancer cellsIn addition could LPA be an effective biomarker for theprediction of chemoresistance in cervical cancer Furtherstudies are required to answer these questions
In this study three specific kinase inhibitors were appliedto determine the signaling pathways involved in the pro-tection effect of LPA on apoptosis It was found that PI3Kinhibitor (LY290042) almost completely blocked the LPAprotection against apoptosis induced by cisplatin Subsequentexperiments confirmed that LPA treatment of Hela cells ledto increased phosphorylation of AKT in a dose- and time-dependent manner This indicated that LPA protected Helacells from apoptosis induced by cisplatin This effect of LPAmay be through the activation of PI3KAKT pathway Thisfinding appears to be consistent with published data showingthat PI3KAKT pathway plays a key role in the regulationof cell apoptosis [28 29] Wan et al observed that PI3Kinhibition augmented staurosporine-induced apoptosis in
the endometrial carcinoma cell line [28] In that experimentstaurosporine reduced the levels of phosphorylation in AKTand Bad In a study by Kim et al 24-bis(p-hydroxyphenyl)-2-butenal (HPB242) induced apoptosis via the inhibition ofPI3KAKT pathway in human cervical cancer cells HPB242treatment decreased phosphatidyl inositol 3-kinase and p-AKT expression levels demonstrating that this survivalpathway may also be inhibited by HPB242 [30] In anotherstudy Cui et al found that MiR-125b inhibited tumor growthand promoted apoptosis of cervical cancer cells by targetingphosphoinositide 3-kinase catalytic subunit delta Overex-pression of MiR-125b in Hela cervical cancer cells inducedapoptosis and downregulated the expression of PIK3CDand Phospho-AKT [31] Kang et al found that thioridazineinduces apoptosis by targeting the PI3KAKTmTOR path-way in cervical cancer cells [32] Thanaketpaisarn et alfound that Artesunate enhanced TRAIL-induced apoptosisin human cervical carcinoma cells through inhibition of thePI3KAKT and NF-120581B signaling pathways [33] Wang et alfound that Stathmin is involved in arsenic trioxide-inducedapoptosis in human cervical cancer cell lines via PI3K linkedsignal pathway As2O3-induced stathmin downregulation ismediated through the phosphatidylinositol-3-kinase (PI3K)signaling pathway PI3K inhibitor effectively attenuated thestathmin downregulation and cell apoptosis upon As2O3-treatment [34] Recurrent and metastatic cervical cancersoften acquire chemoresistance to cisplatin Our findingpointed to a possible sensitization scheme through combinedadministration of PI3K inhibitor and cisplatin
It should be pointed out that while our current studyfocused on the apoptotic pathways andmechanisms the LPAeffects on cell cycle control and cell proliferation should notbe excluded In fact the PI3KAKT pathway is known to playan active role in the control of cell cycle [35] We found thatMEK1 inhibitor (PD98059) significantly inhibited the effectof LPA on the proliferation of Hela cells RasRaf1MEKERKsignal transduction pathway is one of the most importantmembers of the mitogen-activated protein kinase pathwaysregulating cell proliferation [36] MEK1 a MAPK representsa key molecule of the RasRaf1MEKERK signal pathway[37] Several lines of evidence suggest that in cervical cancerthe RasRaf1MEKERK pathway but not the JNK pathwayor the p38 MAPK pathway is the major regulator of cellproliferation [38ndash40]
In summary we demonstrated that cisplatin inducedapoptosis of Hela cell via the downregulation of Bcl-2 andupregulation of Bax Fas-L and increased enzyme activityof caspase-3 LPA was able to block the alterations in theapoptotic factors caused by cisplatin and ultimately reversedthe apoptosis induced by cisplatin Cisplatin-induced cellapoptosis appeared to be partially dependent on PI3KAKTpathway and AKT inhibitor was able to reverse the pro-tective effects of LPA against cisplatin-induced apoptosisLPA stimulated the proliferation of Hela cells throughthe RasRaf1MEKERK pathway These data enriched ourknowledge regarding the LPA activity in cervical cancer cellsIn addition the finding on the positive impact of PI3KAKTinhibitor on the apoptotic effects of cisplatin has provideduseful information for the design of sensitization strategy in
BioMed Research International 11
cervical cancer patients especially those with high levels ofLPA
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Yanxia Sui and Ya Yang contributed equally to this work
Acknowledgments
This study is supported by the following funding sourcesThe National Natural Science Foundations of China (JuanRen 31201060C0709 Juan Ren 30973175H1621 and JuanRen 81172490H1621) Program for New Century ExcellentTalents in University (Juan Ren NCET-12-0440) Scientificand Technological Research Foundation of Shaanxi Province(Juan Ren 2012K13-01-06 Juan Ren 2007K09-09) Scien-tific Research Foundation for the Returned overseas Chi-nese Scholars of State Education Ministry (Juan Ren 0601-18920006) Research Foundation of Health Department ofShaanrsquoxi Province (Juan Ren 2010D41) Qing Nian Jiao ShiGenZongJiHua ofXirsquoan JiaotongUniversity (JuanRen 2012)Program for Changjiang Scholars and Innovative ResearchTeam inUniversity (Zongfang Li PCSIRT1171) andResearchFoundation of Xirsquoan Jiao Tong University of China (JuanRen)
References
[1] J R Erickson Y Hasegawa X Fang et al ldquoLysophosphatidicacid and ovarian cancer a paradigm for tumorogenesis andpatient managementrdquo Prostaglandins and Other Lipid Media-tors vol 64 no 1ndash4 pp 63ndash81 2001
[2] Y Xu Z Z Shen DWWiper et al ldquoLysophosphatidic acid as apotential biomarker for ovarian and other gynecologic cancersrdquoThe Journal of the American Medical Association vol 280 no 8pp 719ndash723 1998
[3] J S Karliner ldquoLysophospholipids and the cardiovascular sys-temrdquoBiochimica et Biophysica ActamdashMolecular and Cell Biologyof Lipids vol 1582 no 1ndash3 pp 216ndash221 2002
[4] C Schulze C Smales L L Rubin and J M StaddonldquoLysophosphatidic acid increases tight junction permeability incultured brain endothelial cellsrdquo Journal of Neurochemistry vol68 no 3 pp 991ndash1000 1997
[5] I Ishii N Fukushima X Ye and J Chun ldquoLysophospholipidreceptors signaling and biologyrdquo Annual Review of Biochem-istry vol 73 pp 321ndash354 2004
[6] T S Panetti ldquoDifferential effects of sphingosine 1-phosphateand lysophosphatidic acid on endothelial cellsrdquo Biochimica etBiophysica ActamdashMolecular and Cell Biology of Lipids vol 1582no 1ndash3 pp 190ndash196 2002
[7] Z Z Shen M Z Wu P Elson et al ldquoFatty acid composition oflysophosphatidic acid and lysophosphatidylinositol in plasmafrom patients with ovarian cancer and other gynecologicaldiseasesrdquo Gynecologic Oncology vol 83 no 1 pp 25ndash30 2001
[8] M M Murph ldquoAgonist-induced endocytosis of lysophospha-tidic acid-coupled LPA1EDG-2 receptors via a dynamin2- andRab5-dependent pathwayrdquo Journal of Cell Science vol 116 no10 pp 1969ndash1980 2003
[9] L M Baudhuin K L Cristina J Lu and Y Xu ldquoAkt activationinduced by lysophosphatidic acid and sphingosine-1-phosphaterequires both mitogen-activated protein kinase kinase andp38 mitogen-activated protein kinase and is cell-line specificrdquoMolecular Pharmacology vol 62 no 3 pp 660ndash671 2002
[10] R-J Chen S-U Chen C-H Chou andM-C Lin ldquoLysophos-phatidic acid receptor 23-mediated IL-8-dependent angiogen-esis in cervical cancer cellsrdquo International Journal of Cancer vol131 no 4 pp 789ndash802 2012
[11] J W Choi S Lim Y-S Oh et al ldquoSubtype-specific role ofphospholipase C-120573 in bradykinin and LPA signaling throughdifferential binding of different PDZ scaffold proteinsrdquo CellularSignalling vol 22 no 7 pp 1153ndash1161 2010
[12] Y Kishi S Okudaira M Tanaka et al ldquoAutotaxin is over-expressed in glioblastoma multiforme and contributes to cellmotility of glioblastoma by converting lysophosphatidylcholineTO lysophosphatidic acidrdquoThe Journal of Biological Chemistryvol 281 no 25 pp 17492ndash17500 2006
[13] A Eastman ldquoThe formation isolation and characterization ofDNA adducts produced by anticancer platinum complexesrdquoPharmacology andTherapeutics vol 34 no 2 pp 155ndash166 1987
[14] K Wozniak and J Błasiak ldquoRecognition and repair of DNA-cisplatin adductsrdquo Acta Biochimica Polonica vol 49 no 3 pp583ndash596 2002
[15] A Basu and S Krishnamurthy ldquoCellular responses to cisplatin-induced DNA damagerdquo Journal of Nucleic Acids vol 2010Article ID 201367 16 pages 2010
[16] A Efeyan and M Serrano ldquop53 guardian of the genome andpoliceman of the oncogenesrdquo Cell Cycle vol 6 no 9 pp 1006ndash1010 2007
[17] O Kutuk E D Arisan T Tezil M C Shoshan and H BasagaldquoCisplatin overcomesBcl-2-mediated resistance to apoptosis viapreferential engagement of Bak critical role of Noxa-mediatedlipid peroxidationrdquo Carcinogenesis vol 30 no 9 pp 1517ndash15272009
[18] C M Yan E Q Chai H Y Cai G Miao and W MaldquoOleuropein induces apoptosis via activation of caspases andsuppression of phosphatidylinositol 3-kinaseprotein kinaseB pathway in HepG2 human hepatoma cell linerdquo MolecularMedicine Reports vol 11 pp 4617ndash4624 2015
[19] X Y Wang X S Fan L Cai S Liu X F Cong and X ChenldquoLysophosphatidic acid rescues bone mesenchymal stem cellsfrom hydrogen peroxide-induced apoptosisrdquo Apoptosis vol 20no 3 pp 273ndash284 2015
[20] A Brozovic G Fritz M Christmann et al ldquoLong-term activa-tion of SAPKJNK p38 kinase and fas-L expression by cisplatinis attenuated in human carcinoma cells that acquired drugresistancerdquo International Journal of Cancer vol 112 no 6 pp974ndash985 2004
[21] Y Q Liu H Xing X B Han et al ldquoApoptosis of HeLa cellsinduced by cisplatin and its mechanismrdquo Journal of HuazhongUniversity of Science and TechnologymdashMedical Science vol 28no 2 pp 197ndash199 2008
[22] K L Jin J-Y Park E J Noh et al ldquoThe effect of combinedtreatment with cisplatin and histone deacetylase inhibitors onHeLa cellsrdquo Journal of Gynecologic Oncology vol 21 no 4 pp262ndash268 2010
12 BioMed Research International
[23] T Sakaeda T Nakamura M Hirai et al ldquoMDR1 up-regulatedby apoptotic stimuli suppresses apoptotic signalingrdquo Pharma-ceutical Research vol 19 no 9 pp 1323ndash1329 2002
[24] K Kandasamy S M Srinivasula E S Alnemri et al ldquoInvolve-ment of proapoptotic molecules Bax and Bak in tumor necro-sis factor-related apoptosis-inducing ligand (TRAIL)-inducedmitochondrial disruption and apoptosis differential regulationof cytochrome c and SmacDIABLO releaserdquo Cancer Researchvol 63 no 7 pp 1712ndash1721 2003
[25] H Toyota N Yanase T Yoshimoto M Moriyama T Sudoand J Mizuguchi ldquoCalpain-induced Bax-cleavage product is amore potent inducer of apoptotic cell death thanwild-type BaxrdquoCancer Letters vol 189 no 2 pp 221ndash230 2003
[26] Y R Meng S J Kang and D A Fishman ldquoLysophosphatidicacid inhibits anti-Fas-mediated apoptosis enhanced by actindepolymerization in epithelial ovarian cancerrdquo FEBS Lettersvol 579 no 5 pp 1311ndash1319 2005
[27] Y Meng S Kang J So S Reierstad and D A FishmanldquoTranslocation of Fas by LPA prevents ovarian cancer cells fromanti-Fas-induced apoptosisrdquo Gynecologic Oncology vol 96 no2 pp 462ndash469 2005
[28] X Wan Y Yokoyama A Shinohara Y Takahashi and TTamaya ldquoPTEN augments staurosporine-induced apoptosis inPTEN-null Ishikawa cells by downregulating PI3KAkt signal-ing pathwayrdquo Cell Death and Differentiation vol 9 no 4 pp414ndash420 2002
[29] J Yu Y Zhang J Mcilroy T Rordorf-Nikolic G A Orr and JM Backer ldquoRegulation of the p85p110 phosphatidylinositol 31015840-kinase stabilization and inhibition of the p110120572 catalytic subunitby the p85 regulatory subunitrdquoMolecular and Cellular Biologyvol 18 no 3 pp 1379ndash1387 1998
[30] M S Kim J H Kim Y Bak et al ldquo24-bis (p-hydroxyphenyl)-2-butenal (HPB242) induces apoptosis via modulating E7expression and inhibition of PI3KAkt pathway in SiHa humancervical cancer cellsrdquo Nutrition and Cancer vol 64 no 8 pp1236ndash1244 2012
[31] F Cui X L Li X Y Zhu et al ldquoMiR-125b inhibits tumorgrowth and promotes apoptosis of cervical cancer cells bytargeting phosphoinositide 3-kinase catalytic subunit deltardquoCellular Physiology and Biochemistry vol 30 no 5 pp 1310ndash1318 2012
[32] S Kang S M Dong B-R Kim et al ldquoThioridazine inducesapoptosis by targeting the PI3KAktmTOR pathway in cervicaland endometrial cancer cellsrdquo Apoptosis vol 17 no 9 pp 989ndash997 2012
[33] O Thanaketpaisarn P Waiwut H Sakurai and I Saiki ldquoArte-sunate enhances TRAIL-induced apoptosis in human cervicalcarcinoma cells through inhibition of the NF-120581B and PI3KAktsignaling pathwaysrdquo International Journal of Oncology vol 39no 1 pp 279ndash285 2011
[34] XWang J-H Ren F Lin et al ldquoStathmin is involved in arsenictrioxide-induced apoptosis in human cervical cancer cell linesvia PI3K linked signal pathwayrdquo Cancer Biology and Therapyvol 10 no 6 pp 632ndash643 2010
[35] C W Lee N H Kim H K Choi et al ldquoLysophosphatidicacid-induced c-fos up-regulation involves cyclic AMP responseelement-binding protein activated by mitogen- and stress-activated protein kinase-1rdquo Journal of Cellular Biochemistry vol104 no 3 pp 785ndash794 2008
[36] DW Hommes M P Peppelenbosch and S J H van DeventerldquoMitogen activated protein (MAP) kinase signal transduction
pathways and novel anti-inflammatory targetsrdquoGut vol 52 no1 pp 144ndash151 2003
[37] M Mukai F Imamura M Ayaki et al ldquoInhibition of tumorinvasion andmetastasis by a novel lysophosphatidic acid (cyclicLPA)rdquo International Journal of Cancer vol 81 no 6 pp 918ndash9221999
[38] E-Y Huang Y-F Chen Y-M Chen et al ldquoA novel radiore-sistant mechanism of galectin-1 mediated by H-Ras-dependentpathways in cervical cancer cellsrdquo Cell Death and Disease vol3 no 1 article e251 2012
[39] W-H Su P-C Chuang E-Y Huang and K D YangldquoRadiation-induced increase in cell migration and metastaticpotential of cervical cancer cells operates via theK-ras pathwayrdquoThe American Journal of Pathology vol 180 no 2 pp 862ndash8712012
[40] H Rincon-Arano R Rosales N Mora A Rodriguez-Castaneda and C Rosales ldquoR-Ras promotes tumor growthof cervical epithelial cellsrdquo Cancer vol 97 no 3 pp 575ndash5852003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
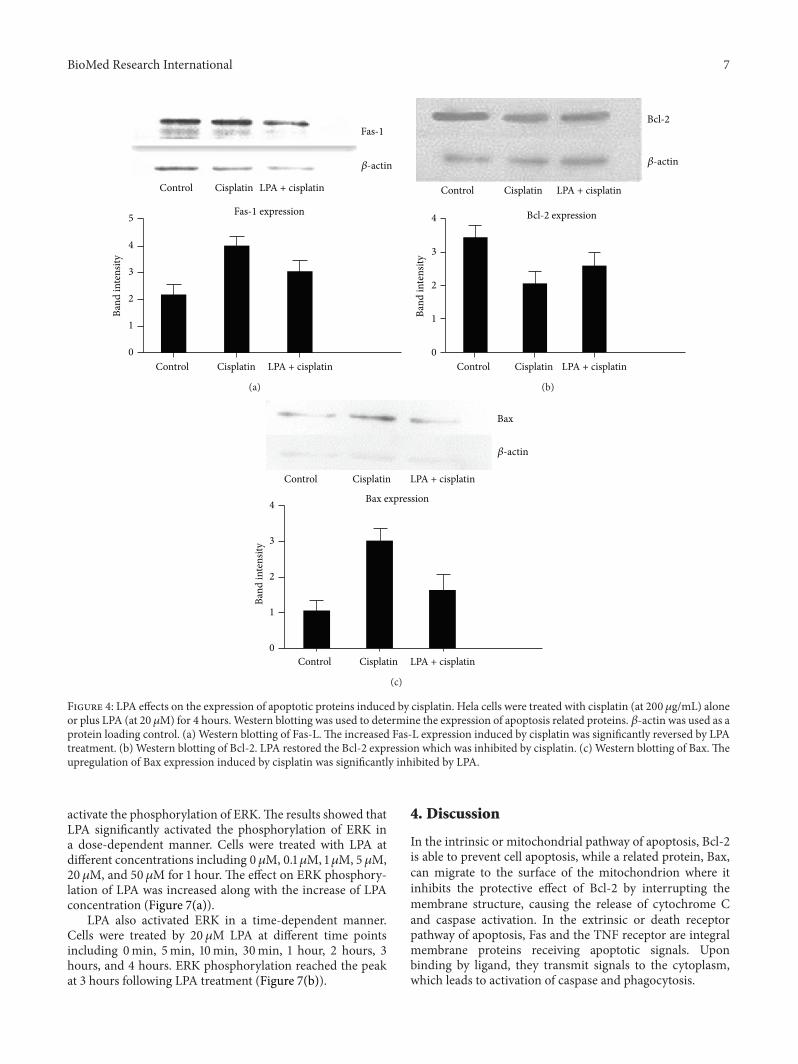
BioMed Research International 7
Fas-1
120573-actin
Fas-1 expression5
4
3
2
1
0
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(a)
Bcl-2
120573-actin
Bcl-2 expression4
3
2
1
0
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(b)
4
3
2
1
0
Bax
120573-actin
Bax expression
Band
inte
nsity
Control Cisplatin LPA + cisplatin
Control Cisplatin LPA + cisplatin
(c)
Figure 4 LPA effects on the expression of apoptotic proteins induced by cisplatin Hela cells were treated with cisplatin (at 200120583gmL) aloneor plus LPA (at 20 120583M) for 4 hours Western blotting was used to determine the expression of apoptosis related proteins 120573-actin was used as aprotein loading control (a) Western blotting of Fas-L The increased Fas-L expression induced by cisplatin was significantly reversed by LPAtreatment (b) Western blotting of Bcl-2 LPA restored the Bcl-2 expression which was inhibited by cisplatin (c) Western blotting of BaxTheupregulation of Bax expression induced by cisplatin was significantly inhibited by LPA
activate the phosphorylation of ERKThe results showed thatLPA significantly activated the phosphorylation of ERK ina dose-dependent manner Cells were treated with LPA atdifferent concentrations including 0120583M 01 120583M 1 120583M 5 120583M20120583M and 50120583M for 1 hour The effect on ERK phosphory-lation of LPA was increased along with the increase of LPAconcentration (Figure 7(a))
LPA also activated ERK in a time-dependent mannerCells were treated by 20 120583M LPA at different time pointsincluding 0min 5min 10min 30min 1 hour 2 hours 3hours and 4 hours ERK phosphorylation reached the peakat 3 hours following LPA treatment (Figure 7(b))
4 Discussion
In the intrinsic or mitochondrial pathway of apoptosis Bcl-2is able to prevent cell apoptosis while a related protein Baxcan migrate to the surface of the mitochondrion where itinhibits the protective effect of Bcl-2 by interrupting themembrane structure causing the release of cytochrome Cand caspase activation In the extrinsic or death receptorpathway of apoptosis Fas and the TNF receptor are integralmembrane proteins receiving apoptotic signals Uponbinding by ligand they transmit signals to the cytoplasmwhich leads to activation of caspase and phagocytosis
8 BioMed Research International
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
Control LPAPI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + PD98069
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + LY294002
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + Y-27632
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
(a)
Con
trol
LPA+
PD98069
LPA+
LY294002
LPA+
Y-27632
LPA
Apop
tosis
rate
0
5
10
15
20
25
30
35
40
45
50
(b)
Figure 5 Flow cytometry detection of cell apoptosis by Annexin VPI staining (a) Hela cells were treated with cisplatin (200120583gmL) andLPA (20 120583M) alone or in combination with different inhibitors for 4 hours Cisplatin and LPA were used in each inhibitors-treated groupInhibitors including PD98059 (MAPK inhibitor at 10 120583M) PD98059 (MAPK inhibitor at 10 120583M) and Y-27632 (Rho inhibitor at 10 120583M)wereapplied respectively (b) The apoptosis rate of Hela cells was quantified Cisplatin-treated group served as control PI3K inhibitor LY294002significantly reversed the LPA effect on apoptosis induced by cisplatin while MAPK inhibitor PD98059 and Rho inhibitor Y-27632 had nosignificant effect
BioMed Research International 9
Fold
chan
ge o
f ban
d in
tens
ity
5
4
3
2
1
00 01 1 5 20 50
LPA (120583M)
0 01 1 5 20 50 LPA (120583M)
P-AKT
AKT
(a)
Fold
chan
ge o
f ban
d in
tens
ity
P-AKT
Total-AKT
4
35
3
25
2
15
1
05
00 5min 10min 30min 1h 2h 3h 4h
Time
0 5min 10min 30min 1h 2h 3h 4h Time
AKT activation
(b)
Figure 6 Western blotting analysis of phosphorylated AKT in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01120583M 1 120583M 5 120583M 20 120583M and 50 120583M for 1 hour Western blotting was conducted to detect the expressionof phosphorylated AKT Total AKT was detected and the results were used as protein input control Densitometry analysis was carried outfor quantification purpose LPA treatment led to increased phosphorylation of AKT in a dose-dependent manner (b) Time-response effectof LPA on the phosphorylation of AKT Hela cells were treated with LPA at 20 120583M for different times including 5min 10min 30min 1 hour2 hours 3 hours and 4 hours Western blotting was used to detect the expression of phosphorylated AKT Compared to the total AKT LPAtreatment activated the phosphorylation of AKT in a time-dependent manner
25
20
15
10
05
00
Fold
chan
ge
0 01 10 50 20 50
LPA (120583M)
0 01 10 05 20 50 LPA (120583M)
P-ERK
ERK
(a)
25
20
15
10
05
00
Fold
chan
ge
0 min5 min10 min30 1h 2h 3h 4hTime
0 min5 min10 min30 1h 2h 3h 4h Time
P-ERK
ERK
(b)
Figure 7 Western blotting analysis of phosphorylated ERK in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01 120583M 1 120583M 5120583M 20 120583M and 50 120583M for 1 hour Western blotting was performed to detect the levels ofphosphorylated ERK Total ERK was used as protein input control The density of bands was quantified and compared LPA activated thephosphorylation of ERK in a dose-dependent manner (b) Hela cells were treated with LPA at 20120583M for different times including 5min10min 30min 1 hour 2 hours 3 hours and 4 hours A time-dependent increase in the ERK phosphorylation was detected
10 BioMed Research International
Some studies found that cell apoptosis induced by cis-platinwasmediated by death receptors signaling pathway [2021] while others claimed that it was mediated by mitochon-drial pathway [22 23] The discrepancy in previous studiesmay be caused by the divergent cell lines methodologiesand conditions used by different groups In this study weobserved that cisplatin significantly induced the apoptosis ofHela cells in a concentration-dependent manner Cisplatintreatment was accompanied by increased Fas-1 and Baxexpression and the increased enzyme activity of caspase-3 Meanwhile cisplatin inhibited Bcl-2 expression Theseobservations suggested that cisplatin-induced apoptosis ofHela cell may be mediated by both intrinsic and extrinsicpathways Indeed extensive cross talks between the two path-ways have been identified [24 25] Simultaneous activationof the two pathways has been observed in many types of cellsfollowing various treatments
Our previous study found that both ovarian cancer cellsand peritoneal mesothelial cells contribute to the elevatedLPA levels in ovarian cancer ascites Under normal cultureconditions ovarian cancer cells and prostate cancer cellsbut not breast cancer cell lines constitutively synthesize andrelease significant amounts of LPA to the culture medium Amajor finding of this study is that LPA was able to rescue theHela cells apoptosis induced by cisplatin The protective roleof LPA against apoptosis was observed in ovarian cancer cells[26]We observed that LPA treatment blocked the alterationsof Bcl-2 Fas-1 and Bax expression that were induced bycisplatin LPA also significantly inhibited the upregulationof the caspase-3 enzyme activity induced by cisplatin Ourobservations of LPA effect on cervical cancer were consistentwith other reports on LPA effects in different kinds of cellsMeng et al found that LPA may protect epithelial ovariancancer from immune cell attack and cytoskeleton disruptingreagents induced apoptosis through multiple pathways andLPA inhibited anti-Fas-induced apoptosis enhanced by actindepolymerization [26 27] Actin depolymerization acceler-ated caspase-8 activation while LPA inhibited the associationand activation of caspase-8 at the DISC in epithelial ovariancancer OVCAR3 cells Thus our observations have raisedan interesting question on the potential role of LPA forthe development of chemoresistance by cervical cancer cellsIn addition could LPA be an effective biomarker for theprediction of chemoresistance in cervical cancer Furtherstudies are required to answer these questions
In this study three specific kinase inhibitors were appliedto determine the signaling pathways involved in the pro-tection effect of LPA on apoptosis It was found that PI3Kinhibitor (LY290042) almost completely blocked the LPAprotection against apoptosis induced by cisplatin Subsequentexperiments confirmed that LPA treatment of Hela cells ledto increased phosphorylation of AKT in a dose- and time-dependent manner This indicated that LPA protected Helacells from apoptosis induced by cisplatin This effect of LPAmay be through the activation of PI3KAKT pathway Thisfinding appears to be consistent with published data showingthat PI3KAKT pathway plays a key role in the regulationof cell apoptosis [28 29] Wan et al observed that PI3Kinhibition augmented staurosporine-induced apoptosis in
the endometrial carcinoma cell line [28] In that experimentstaurosporine reduced the levels of phosphorylation in AKTand Bad In a study by Kim et al 24-bis(p-hydroxyphenyl)-2-butenal (HPB242) induced apoptosis via the inhibition ofPI3KAKT pathway in human cervical cancer cells HPB242treatment decreased phosphatidyl inositol 3-kinase and p-AKT expression levels demonstrating that this survivalpathway may also be inhibited by HPB242 [30] In anotherstudy Cui et al found that MiR-125b inhibited tumor growthand promoted apoptosis of cervical cancer cells by targetingphosphoinositide 3-kinase catalytic subunit delta Overex-pression of MiR-125b in Hela cervical cancer cells inducedapoptosis and downregulated the expression of PIK3CDand Phospho-AKT [31] Kang et al found that thioridazineinduces apoptosis by targeting the PI3KAKTmTOR path-way in cervical cancer cells [32] Thanaketpaisarn et alfound that Artesunate enhanced TRAIL-induced apoptosisin human cervical carcinoma cells through inhibition of thePI3KAKT and NF-120581B signaling pathways [33] Wang et alfound that Stathmin is involved in arsenic trioxide-inducedapoptosis in human cervical cancer cell lines via PI3K linkedsignal pathway As2O3-induced stathmin downregulation ismediated through the phosphatidylinositol-3-kinase (PI3K)signaling pathway PI3K inhibitor effectively attenuated thestathmin downregulation and cell apoptosis upon As2O3-treatment [34] Recurrent and metastatic cervical cancersoften acquire chemoresistance to cisplatin Our findingpointed to a possible sensitization scheme through combinedadministration of PI3K inhibitor and cisplatin
It should be pointed out that while our current studyfocused on the apoptotic pathways andmechanisms the LPAeffects on cell cycle control and cell proliferation should notbe excluded In fact the PI3KAKT pathway is known to playan active role in the control of cell cycle [35] We found thatMEK1 inhibitor (PD98059) significantly inhibited the effectof LPA on the proliferation of Hela cells RasRaf1MEKERKsignal transduction pathway is one of the most importantmembers of the mitogen-activated protein kinase pathwaysregulating cell proliferation [36] MEK1 a MAPK representsa key molecule of the RasRaf1MEKERK signal pathway[37] Several lines of evidence suggest that in cervical cancerthe RasRaf1MEKERK pathway but not the JNK pathwayor the p38 MAPK pathway is the major regulator of cellproliferation [38ndash40]
In summary we demonstrated that cisplatin inducedapoptosis of Hela cell via the downregulation of Bcl-2 andupregulation of Bax Fas-L and increased enzyme activityof caspase-3 LPA was able to block the alterations in theapoptotic factors caused by cisplatin and ultimately reversedthe apoptosis induced by cisplatin Cisplatin-induced cellapoptosis appeared to be partially dependent on PI3KAKTpathway and AKT inhibitor was able to reverse the pro-tective effects of LPA against cisplatin-induced apoptosisLPA stimulated the proliferation of Hela cells throughthe RasRaf1MEKERK pathway These data enriched ourknowledge regarding the LPA activity in cervical cancer cellsIn addition the finding on the positive impact of PI3KAKTinhibitor on the apoptotic effects of cisplatin has provideduseful information for the design of sensitization strategy in
BioMed Research International 11
cervical cancer patients especially those with high levels ofLPA
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Yanxia Sui and Ya Yang contributed equally to this work
Acknowledgments
This study is supported by the following funding sourcesThe National Natural Science Foundations of China (JuanRen 31201060C0709 Juan Ren 30973175H1621 and JuanRen 81172490H1621) Program for New Century ExcellentTalents in University (Juan Ren NCET-12-0440) Scientificand Technological Research Foundation of Shaanxi Province(Juan Ren 2012K13-01-06 Juan Ren 2007K09-09) Scien-tific Research Foundation for the Returned overseas Chi-nese Scholars of State Education Ministry (Juan Ren 0601-18920006) Research Foundation of Health Department ofShaanrsquoxi Province (Juan Ren 2010D41) Qing Nian Jiao ShiGenZongJiHua ofXirsquoan JiaotongUniversity (JuanRen 2012)Program for Changjiang Scholars and Innovative ResearchTeam inUniversity (Zongfang Li PCSIRT1171) andResearchFoundation of Xirsquoan Jiao Tong University of China (JuanRen)
References
[1] J R Erickson Y Hasegawa X Fang et al ldquoLysophosphatidicacid and ovarian cancer a paradigm for tumorogenesis andpatient managementrdquo Prostaglandins and Other Lipid Media-tors vol 64 no 1ndash4 pp 63ndash81 2001
[2] Y Xu Z Z Shen DWWiper et al ldquoLysophosphatidic acid as apotential biomarker for ovarian and other gynecologic cancersrdquoThe Journal of the American Medical Association vol 280 no 8pp 719ndash723 1998
[3] J S Karliner ldquoLysophospholipids and the cardiovascular sys-temrdquoBiochimica et Biophysica ActamdashMolecular and Cell Biologyof Lipids vol 1582 no 1ndash3 pp 216ndash221 2002
[4] C Schulze C Smales L L Rubin and J M StaddonldquoLysophosphatidic acid increases tight junction permeability incultured brain endothelial cellsrdquo Journal of Neurochemistry vol68 no 3 pp 991ndash1000 1997
[5] I Ishii N Fukushima X Ye and J Chun ldquoLysophospholipidreceptors signaling and biologyrdquo Annual Review of Biochem-istry vol 73 pp 321ndash354 2004
[6] T S Panetti ldquoDifferential effects of sphingosine 1-phosphateand lysophosphatidic acid on endothelial cellsrdquo Biochimica etBiophysica ActamdashMolecular and Cell Biology of Lipids vol 1582no 1ndash3 pp 190ndash196 2002
[7] Z Z Shen M Z Wu P Elson et al ldquoFatty acid composition oflysophosphatidic acid and lysophosphatidylinositol in plasmafrom patients with ovarian cancer and other gynecologicaldiseasesrdquo Gynecologic Oncology vol 83 no 1 pp 25ndash30 2001
[8] M M Murph ldquoAgonist-induced endocytosis of lysophospha-tidic acid-coupled LPA1EDG-2 receptors via a dynamin2- andRab5-dependent pathwayrdquo Journal of Cell Science vol 116 no10 pp 1969ndash1980 2003
[9] L M Baudhuin K L Cristina J Lu and Y Xu ldquoAkt activationinduced by lysophosphatidic acid and sphingosine-1-phosphaterequires both mitogen-activated protein kinase kinase andp38 mitogen-activated protein kinase and is cell-line specificrdquoMolecular Pharmacology vol 62 no 3 pp 660ndash671 2002
[10] R-J Chen S-U Chen C-H Chou andM-C Lin ldquoLysophos-phatidic acid receptor 23-mediated IL-8-dependent angiogen-esis in cervical cancer cellsrdquo International Journal of Cancer vol131 no 4 pp 789ndash802 2012
[11] J W Choi S Lim Y-S Oh et al ldquoSubtype-specific role ofphospholipase C-120573 in bradykinin and LPA signaling throughdifferential binding of different PDZ scaffold proteinsrdquo CellularSignalling vol 22 no 7 pp 1153ndash1161 2010
[12] Y Kishi S Okudaira M Tanaka et al ldquoAutotaxin is over-expressed in glioblastoma multiforme and contributes to cellmotility of glioblastoma by converting lysophosphatidylcholineTO lysophosphatidic acidrdquoThe Journal of Biological Chemistryvol 281 no 25 pp 17492ndash17500 2006
[13] A Eastman ldquoThe formation isolation and characterization ofDNA adducts produced by anticancer platinum complexesrdquoPharmacology andTherapeutics vol 34 no 2 pp 155ndash166 1987
[14] K Wozniak and J Błasiak ldquoRecognition and repair of DNA-cisplatin adductsrdquo Acta Biochimica Polonica vol 49 no 3 pp583ndash596 2002
[15] A Basu and S Krishnamurthy ldquoCellular responses to cisplatin-induced DNA damagerdquo Journal of Nucleic Acids vol 2010Article ID 201367 16 pages 2010
[16] A Efeyan and M Serrano ldquop53 guardian of the genome andpoliceman of the oncogenesrdquo Cell Cycle vol 6 no 9 pp 1006ndash1010 2007
[17] O Kutuk E D Arisan T Tezil M C Shoshan and H BasagaldquoCisplatin overcomesBcl-2-mediated resistance to apoptosis viapreferential engagement of Bak critical role of Noxa-mediatedlipid peroxidationrdquo Carcinogenesis vol 30 no 9 pp 1517ndash15272009
[18] C M Yan E Q Chai H Y Cai G Miao and W MaldquoOleuropein induces apoptosis via activation of caspases andsuppression of phosphatidylinositol 3-kinaseprotein kinaseB pathway in HepG2 human hepatoma cell linerdquo MolecularMedicine Reports vol 11 pp 4617ndash4624 2015
[19] X Y Wang X S Fan L Cai S Liu X F Cong and X ChenldquoLysophosphatidic acid rescues bone mesenchymal stem cellsfrom hydrogen peroxide-induced apoptosisrdquo Apoptosis vol 20no 3 pp 273ndash284 2015
[20] A Brozovic G Fritz M Christmann et al ldquoLong-term activa-tion of SAPKJNK p38 kinase and fas-L expression by cisplatinis attenuated in human carcinoma cells that acquired drugresistancerdquo International Journal of Cancer vol 112 no 6 pp974ndash985 2004
[21] Y Q Liu H Xing X B Han et al ldquoApoptosis of HeLa cellsinduced by cisplatin and its mechanismrdquo Journal of HuazhongUniversity of Science and TechnologymdashMedical Science vol 28no 2 pp 197ndash199 2008
[22] K L Jin J-Y Park E J Noh et al ldquoThe effect of combinedtreatment with cisplatin and histone deacetylase inhibitors onHeLa cellsrdquo Journal of Gynecologic Oncology vol 21 no 4 pp262ndash268 2010
12 BioMed Research International
[23] T Sakaeda T Nakamura M Hirai et al ldquoMDR1 up-regulatedby apoptotic stimuli suppresses apoptotic signalingrdquo Pharma-ceutical Research vol 19 no 9 pp 1323ndash1329 2002
[24] K Kandasamy S M Srinivasula E S Alnemri et al ldquoInvolve-ment of proapoptotic molecules Bax and Bak in tumor necro-sis factor-related apoptosis-inducing ligand (TRAIL)-inducedmitochondrial disruption and apoptosis differential regulationof cytochrome c and SmacDIABLO releaserdquo Cancer Researchvol 63 no 7 pp 1712ndash1721 2003
[25] H Toyota N Yanase T Yoshimoto M Moriyama T Sudoand J Mizuguchi ldquoCalpain-induced Bax-cleavage product is amore potent inducer of apoptotic cell death thanwild-type BaxrdquoCancer Letters vol 189 no 2 pp 221ndash230 2003
[26] Y R Meng S J Kang and D A Fishman ldquoLysophosphatidicacid inhibits anti-Fas-mediated apoptosis enhanced by actindepolymerization in epithelial ovarian cancerrdquo FEBS Lettersvol 579 no 5 pp 1311ndash1319 2005
[27] Y Meng S Kang J So S Reierstad and D A FishmanldquoTranslocation of Fas by LPA prevents ovarian cancer cells fromanti-Fas-induced apoptosisrdquo Gynecologic Oncology vol 96 no2 pp 462ndash469 2005
[28] X Wan Y Yokoyama A Shinohara Y Takahashi and TTamaya ldquoPTEN augments staurosporine-induced apoptosis inPTEN-null Ishikawa cells by downregulating PI3KAkt signal-ing pathwayrdquo Cell Death and Differentiation vol 9 no 4 pp414ndash420 2002
[29] J Yu Y Zhang J Mcilroy T Rordorf-Nikolic G A Orr and JM Backer ldquoRegulation of the p85p110 phosphatidylinositol 31015840-kinase stabilization and inhibition of the p110120572 catalytic subunitby the p85 regulatory subunitrdquoMolecular and Cellular Biologyvol 18 no 3 pp 1379ndash1387 1998
[30] M S Kim J H Kim Y Bak et al ldquo24-bis (p-hydroxyphenyl)-2-butenal (HPB242) induces apoptosis via modulating E7expression and inhibition of PI3KAkt pathway in SiHa humancervical cancer cellsrdquo Nutrition and Cancer vol 64 no 8 pp1236ndash1244 2012
[31] F Cui X L Li X Y Zhu et al ldquoMiR-125b inhibits tumorgrowth and promotes apoptosis of cervical cancer cells bytargeting phosphoinositide 3-kinase catalytic subunit deltardquoCellular Physiology and Biochemistry vol 30 no 5 pp 1310ndash1318 2012
[32] S Kang S M Dong B-R Kim et al ldquoThioridazine inducesapoptosis by targeting the PI3KAktmTOR pathway in cervicaland endometrial cancer cellsrdquo Apoptosis vol 17 no 9 pp 989ndash997 2012
[33] O Thanaketpaisarn P Waiwut H Sakurai and I Saiki ldquoArte-sunate enhances TRAIL-induced apoptosis in human cervicalcarcinoma cells through inhibition of the NF-120581B and PI3KAktsignaling pathwaysrdquo International Journal of Oncology vol 39no 1 pp 279ndash285 2011
[34] XWang J-H Ren F Lin et al ldquoStathmin is involved in arsenictrioxide-induced apoptosis in human cervical cancer cell linesvia PI3K linked signal pathwayrdquo Cancer Biology and Therapyvol 10 no 6 pp 632ndash643 2010
[35] C W Lee N H Kim H K Choi et al ldquoLysophosphatidicacid-induced c-fos up-regulation involves cyclic AMP responseelement-binding protein activated by mitogen- and stress-activated protein kinase-1rdquo Journal of Cellular Biochemistry vol104 no 3 pp 785ndash794 2008
[36] DW Hommes M P Peppelenbosch and S J H van DeventerldquoMitogen activated protein (MAP) kinase signal transduction
pathways and novel anti-inflammatory targetsrdquoGut vol 52 no1 pp 144ndash151 2003
[37] M Mukai F Imamura M Ayaki et al ldquoInhibition of tumorinvasion andmetastasis by a novel lysophosphatidic acid (cyclicLPA)rdquo International Journal of Cancer vol 81 no 6 pp 918ndash9221999
[38] E-Y Huang Y-F Chen Y-M Chen et al ldquoA novel radiore-sistant mechanism of galectin-1 mediated by H-Ras-dependentpathways in cervical cancer cellsrdquo Cell Death and Disease vol3 no 1 article e251 2012
[39] W-H Su P-C Chuang E-Y Huang and K D YangldquoRadiation-induced increase in cell migration and metastaticpotential of cervical cancer cells operates via theK-ras pathwayrdquoThe American Journal of Pathology vol 180 no 2 pp 862ndash8712012
[40] H Rincon-Arano R Rosales N Mora A Rodriguez-Castaneda and C Rosales ldquoR-Ras promotes tumor growthof cervical epithelial cellsrdquo Cancer vol 97 no 3 pp 575ndash5852003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 BioMed Research International
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
Control LPAPI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + PD98069
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + LY294002
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
LPA + Y-27632
PI
Annexin Y FITC100 101 102 103 104
100
101
102
103
104
(a)
Con
trol
LPA+
PD98069
LPA+
LY294002
LPA+
Y-27632
LPA
Apop
tosis
rate
0
5
10
15
20
25
30
35
40
45
50
(b)
Figure 5 Flow cytometry detection of cell apoptosis by Annexin VPI staining (a) Hela cells were treated with cisplatin (200120583gmL) andLPA (20 120583M) alone or in combination with different inhibitors for 4 hours Cisplatin and LPA were used in each inhibitors-treated groupInhibitors including PD98059 (MAPK inhibitor at 10 120583M) PD98059 (MAPK inhibitor at 10 120583M) and Y-27632 (Rho inhibitor at 10 120583M)wereapplied respectively (b) The apoptosis rate of Hela cells was quantified Cisplatin-treated group served as control PI3K inhibitor LY294002significantly reversed the LPA effect on apoptosis induced by cisplatin while MAPK inhibitor PD98059 and Rho inhibitor Y-27632 had nosignificant effect
BioMed Research International 9
Fold
chan
ge o
f ban
d in
tens
ity
5
4
3
2
1
00 01 1 5 20 50
LPA (120583M)
0 01 1 5 20 50 LPA (120583M)
P-AKT
AKT
(a)
Fold
chan
ge o
f ban
d in
tens
ity
P-AKT
Total-AKT
4
35
3
25
2
15
1
05
00 5min 10min 30min 1h 2h 3h 4h
Time
0 5min 10min 30min 1h 2h 3h 4h Time
AKT activation
(b)
Figure 6 Western blotting analysis of phosphorylated AKT in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01120583M 1 120583M 5 120583M 20 120583M and 50 120583M for 1 hour Western blotting was conducted to detect the expressionof phosphorylated AKT Total AKT was detected and the results were used as protein input control Densitometry analysis was carried outfor quantification purpose LPA treatment led to increased phosphorylation of AKT in a dose-dependent manner (b) Time-response effectof LPA on the phosphorylation of AKT Hela cells were treated with LPA at 20 120583M for different times including 5min 10min 30min 1 hour2 hours 3 hours and 4 hours Western blotting was used to detect the expression of phosphorylated AKT Compared to the total AKT LPAtreatment activated the phosphorylation of AKT in a time-dependent manner
25
20
15
10
05
00
Fold
chan
ge
0 01 10 50 20 50
LPA (120583M)
0 01 10 05 20 50 LPA (120583M)
P-ERK
ERK
(a)
25
20
15
10
05
00
Fold
chan
ge
0 min5 min10 min30 1h 2h 3h 4hTime
0 min5 min10 min30 1h 2h 3h 4h Time
P-ERK
ERK
(b)
Figure 7 Western blotting analysis of phosphorylated ERK in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01 120583M 1 120583M 5120583M 20 120583M and 50 120583M for 1 hour Western blotting was performed to detect the levels ofphosphorylated ERK Total ERK was used as protein input control The density of bands was quantified and compared LPA activated thephosphorylation of ERK in a dose-dependent manner (b) Hela cells were treated with LPA at 20120583M for different times including 5min10min 30min 1 hour 2 hours 3 hours and 4 hours A time-dependent increase in the ERK phosphorylation was detected
10 BioMed Research International
Some studies found that cell apoptosis induced by cis-platinwasmediated by death receptors signaling pathway [2021] while others claimed that it was mediated by mitochon-drial pathway [22 23] The discrepancy in previous studiesmay be caused by the divergent cell lines methodologiesand conditions used by different groups In this study weobserved that cisplatin significantly induced the apoptosis ofHela cells in a concentration-dependent manner Cisplatintreatment was accompanied by increased Fas-1 and Baxexpression and the increased enzyme activity of caspase-3 Meanwhile cisplatin inhibited Bcl-2 expression Theseobservations suggested that cisplatin-induced apoptosis ofHela cell may be mediated by both intrinsic and extrinsicpathways Indeed extensive cross talks between the two path-ways have been identified [24 25] Simultaneous activationof the two pathways has been observed in many types of cellsfollowing various treatments
Our previous study found that both ovarian cancer cellsand peritoneal mesothelial cells contribute to the elevatedLPA levels in ovarian cancer ascites Under normal cultureconditions ovarian cancer cells and prostate cancer cellsbut not breast cancer cell lines constitutively synthesize andrelease significant amounts of LPA to the culture medium Amajor finding of this study is that LPA was able to rescue theHela cells apoptosis induced by cisplatin The protective roleof LPA against apoptosis was observed in ovarian cancer cells[26]We observed that LPA treatment blocked the alterationsof Bcl-2 Fas-1 and Bax expression that were induced bycisplatin LPA also significantly inhibited the upregulationof the caspase-3 enzyme activity induced by cisplatin Ourobservations of LPA effect on cervical cancer were consistentwith other reports on LPA effects in different kinds of cellsMeng et al found that LPA may protect epithelial ovariancancer from immune cell attack and cytoskeleton disruptingreagents induced apoptosis through multiple pathways andLPA inhibited anti-Fas-induced apoptosis enhanced by actindepolymerization [26 27] Actin depolymerization acceler-ated caspase-8 activation while LPA inhibited the associationand activation of caspase-8 at the DISC in epithelial ovariancancer OVCAR3 cells Thus our observations have raisedan interesting question on the potential role of LPA forthe development of chemoresistance by cervical cancer cellsIn addition could LPA be an effective biomarker for theprediction of chemoresistance in cervical cancer Furtherstudies are required to answer these questions
In this study three specific kinase inhibitors were appliedto determine the signaling pathways involved in the pro-tection effect of LPA on apoptosis It was found that PI3Kinhibitor (LY290042) almost completely blocked the LPAprotection against apoptosis induced by cisplatin Subsequentexperiments confirmed that LPA treatment of Hela cells ledto increased phosphorylation of AKT in a dose- and time-dependent manner This indicated that LPA protected Helacells from apoptosis induced by cisplatin This effect of LPAmay be through the activation of PI3KAKT pathway Thisfinding appears to be consistent with published data showingthat PI3KAKT pathway plays a key role in the regulationof cell apoptosis [28 29] Wan et al observed that PI3Kinhibition augmented staurosporine-induced apoptosis in
the endometrial carcinoma cell line [28] In that experimentstaurosporine reduced the levels of phosphorylation in AKTand Bad In a study by Kim et al 24-bis(p-hydroxyphenyl)-2-butenal (HPB242) induced apoptosis via the inhibition ofPI3KAKT pathway in human cervical cancer cells HPB242treatment decreased phosphatidyl inositol 3-kinase and p-AKT expression levels demonstrating that this survivalpathway may also be inhibited by HPB242 [30] In anotherstudy Cui et al found that MiR-125b inhibited tumor growthand promoted apoptosis of cervical cancer cells by targetingphosphoinositide 3-kinase catalytic subunit delta Overex-pression of MiR-125b in Hela cervical cancer cells inducedapoptosis and downregulated the expression of PIK3CDand Phospho-AKT [31] Kang et al found that thioridazineinduces apoptosis by targeting the PI3KAKTmTOR path-way in cervical cancer cells [32] Thanaketpaisarn et alfound that Artesunate enhanced TRAIL-induced apoptosisin human cervical carcinoma cells through inhibition of thePI3KAKT and NF-120581B signaling pathways [33] Wang et alfound that Stathmin is involved in arsenic trioxide-inducedapoptosis in human cervical cancer cell lines via PI3K linkedsignal pathway As2O3-induced stathmin downregulation ismediated through the phosphatidylinositol-3-kinase (PI3K)signaling pathway PI3K inhibitor effectively attenuated thestathmin downregulation and cell apoptosis upon As2O3-treatment [34] Recurrent and metastatic cervical cancersoften acquire chemoresistance to cisplatin Our findingpointed to a possible sensitization scheme through combinedadministration of PI3K inhibitor and cisplatin
It should be pointed out that while our current studyfocused on the apoptotic pathways andmechanisms the LPAeffects on cell cycle control and cell proliferation should notbe excluded In fact the PI3KAKT pathway is known to playan active role in the control of cell cycle [35] We found thatMEK1 inhibitor (PD98059) significantly inhibited the effectof LPA on the proliferation of Hela cells RasRaf1MEKERKsignal transduction pathway is one of the most importantmembers of the mitogen-activated protein kinase pathwaysregulating cell proliferation [36] MEK1 a MAPK representsa key molecule of the RasRaf1MEKERK signal pathway[37] Several lines of evidence suggest that in cervical cancerthe RasRaf1MEKERK pathway but not the JNK pathwayor the p38 MAPK pathway is the major regulator of cellproliferation [38ndash40]
In summary we demonstrated that cisplatin inducedapoptosis of Hela cell via the downregulation of Bcl-2 andupregulation of Bax Fas-L and increased enzyme activityof caspase-3 LPA was able to block the alterations in theapoptotic factors caused by cisplatin and ultimately reversedthe apoptosis induced by cisplatin Cisplatin-induced cellapoptosis appeared to be partially dependent on PI3KAKTpathway and AKT inhibitor was able to reverse the pro-tective effects of LPA against cisplatin-induced apoptosisLPA stimulated the proliferation of Hela cells throughthe RasRaf1MEKERK pathway These data enriched ourknowledge regarding the LPA activity in cervical cancer cellsIn addition the finding on the positive impact of PI3KAKTinhibitor on the apoptotic effects of cisplatin has provideduseful information for the design of sensitization strategy in
BioMed Research International 11
cervical cancer patients especially those with high levels ofLPA
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Yanxia Sui and Ya Yang contributed equally to this work
Acknowledgments
This study is supported by the following funding sourcesThe National Natural Science Foundations of China (JuanRen 31201060C0709 Juan Ren 30973175H1621 and JuanRen 81172490H1621) Program for New Century ExcellentTalents in University (Juan Ren NCET-12-0440) Scientificand Technological Research Foundation of Shaanxi Province(Juan Ren 2012K13-01-06 Juan Ren 2007K09-09) Scien-tific Research Foundation for the Returned overseas Chi-nese Scholars of State Education Ministry (Juan Ren 0601-18920006) Research Foundation of Health Department ofShaanrsquoxi Province (Juan Ren 2010D41) Qing Nian Jiao ShiGenZongJiHua ofXirsquoan JiaotongUniversity (JuanRen 2012)Program for Changjiang Scholars and Innovative ResearchTeam inUniversity (Zongfang Li PCSIRT1171) andResearchFoundation of Xirsquoan Jiao Tong University of China (JuanRen)
References
[1] J R Erickson Y Hasegawa X Fang et al ldquoLysophosphatidicacid and ovarian cancer a paradigm for tumorogenesis andpatient managementrdquo Prostaglandins and Other Lipid Media-tors vol 64 no 1ndash4 pp 63ndash81 2001
[2] Y Xu Z Z Shen DWWiper et al ldquoLysophosphatidic acid as apotential biomarker for ovarian and other gynecologic cancersrdquoThe Journal of the American Medical Association vol 280 no 8pp 719ndash723 1998
[3] J S Karliner ldquoLysophospholipids and the cardiovascular sys-temrdquoBiochimica et Biophysica ActamdashMolecular and Cell Biologyof Lipids vol 1582 no 1ndash3 pp 216ndash221 2002
[4] C Schulze C Smales L L Rubin and J M StaddonldquoLysophosphatidic acid increases tight junction permeability incultured brain endothelial cellsrdquo Journal of Neurochemistry vol68 no 3 pp 991ndash1000 1997
[5] I Ishii N Fukushima X Ye and J Chun ldquoLysophospholipidreceptors signaling and biologyrdquo Annual Review of Biochem-istry vol 73 pp 321ndash354 2004
[6] T S Panetti ldquoDifferential effects of sphingosine 1-phosphateand lysophosphatidic acid on endothelial cellsrdquo Biochimica etBiophysica ActamdashMolecular and Cell Biology of Lipids vol 1582no 1ndash3 pp 190ndash196 2002
[7] Z Z Shen M Z Wu P Elson et al ldquoFatty acid composition oflysophosphatidic acid and lysophosphatidylinositol in plasmafrom patients with ovarian cancer and other gynecologicaldiseasesrdquo Gynecologic Oncology vol 83 no 1 pp 25ndash30 2001
[8] M M Murph ldquoAgonist-induced endocytosis of lysophospha-tidic acid-coupled LPA1EDG-2 receptors via a dynamin2- andRab5-dependent pathwayrdquo Journal of Cell Science vol 116 no10 pp 1969ndash1980 2003
[9] L M Baudhuin K L Cristina J Lu and Y Xu ldquoAkt activationinduced by lysophosphatidic acid and sphingosine-1-phosphaterequires both mitogen-activated protein kinase kinase andp38 mitogen-activated protein kinase and is cell-line specificrdquoMolecular Pharmacology vol 62 no 3 pp 660ndash671 2002
[10] R-J Chen S-U Chen C-H Chou andM-C Lin ldquoLysophos-phatidic acid receptor 23-mediated IL-8-dependent angiogen-esis in cervical cancer cellsrdquo International Journal of Cancer vol131 no 4 pp 789ndash802 2012
[11] J W Choi S Lim Y-S Oh et al ldquoSubtype-specific role ofphospholipase C-120573 in bradykinin and LPA signaling throughdifferential binding of different PDZ scaffold proteinsrdquo CellularSignalling vol 22 no 7 pp 1153ndash1161 2010
[12] Y Kishi S Okudaira M Tanaka et al ldquoAutotaxin is over-expressed in glioblastoma multiforme and contributes to cellmotility of glioblastoma by converting lysophosphatidylcholineTO lysophosphatidic acidrdquoThe Journal of Biological Chemistryvol 281 no 25 pp 17492ndash17500 2006
[13] A Eastman ldquoThe formation isolation and characterization ofDNA adducts produced by anticancer platinum complexesrdquoPharmacology andTherapeutics vol 34 no 2 pp 155ndash166 1987
[14] K Wozniak and J Błasiak ldquoRecognition and repair of DNA-cisplatin adductsrdquo Acta Biochimica Polonica vol 49 no 3 pp583ndash596 2002
[15] A Basu and S Krishnamurthy ldquoCellular responses to cisplatin-induced DNA damagerdquo Journal of Nucleic Acids vol 2010Article ID 201367 16 pages 2010
[16] A Efeyan and M Serrano ldquop53 guardian of the genome andpoliceman of the oncogenesrdquo Cell Cycle vol 6 no 9 pp 1006ndash1010 2007
[17] O Kutuk E D Arisan T Tezil M C Shoshan and H BasagaldquoCisplatin overcomesBcl-2-mediated resistance to apoptosis viapreferential engagement of Bak critical role of Noxa-mediatedlipid peroxidationrdquo Carcinogenesis vol 30 no 9 pp 1517ndash15272009
[18] C M Yan E Q Chai H Y Cai G Miao and W MaldquoOleuropein induces apoptosis via activation of caspases andsuppression of phosphatidylinositol 3-kinaseprotein kinaseB pathway in HepG2 human hepatoma cell linerdquo MolecularMedicine Reports vol 11 pp 4617ndash4624 2015
[19] X Y Wang X S Fan L Cai S Liu X F Cong and X ChenldquoLysophosphatidic acid rescues bone mesenchymal stem cellsfrom hydrogen peroxide-induced apoptosisrdquo Apoptosis vol 20no 3 pp 273ndash284 2015
[20] A Brozovic G Fritz M Christmann et al ldquoLong-term activa-tion of SAPKJNK p38 kinase and fas-L expression by cisplatinis attenuated in human carcinoma cells that acquired drugresistancerdquo International Journal of Cancer vol 112 no 6 pp974ndash985 2004
[21] Y Q Liu H Xing X B Han et al ldquoApoptosis of HeLa cellsinduced by cisplatin and its mechanismrdquo Journal of HuazhongUniversity of Science and TechnologymdashMedical Science vol 28no 2 pp 197ndash199 2008
[22] K L Jin J-Y Park E J Noh et al ldquoThe effect of combinedtreatment with cisplatin and histone deacetylase inhibitors onHeLa cellsrdquo Journal of Gynecologic Oncology vol 21 no 4 pp262ndash268 2010
12 BioMed Research International
[23] T Sakaeda T Nakamura M Hirai et al ldquoMDR1 up-regulatedby apoptotic stimuli suppresses apoptotic signalingrdquo Pharma-ceutical Research vol 19 no 9 pp 1323ndash1329 2002
[24] K Kandasamy S M Srinivasula E S Alnemri et al ldquoInvolve-ment of proapoptotic molecules Bax and Bak in tumor necro-sis factor-related apoptosis-inducing ligand (TRAIL)-inducedmitochondrial disruption and apoptosis differential regulationof cytochrome c and SmacDIABLO releaserdquo Cancer Researchvol 63 no 7 pp 1712ndash1721 2003
[25] H Toyota N Yanase T Yoshimoto M Moriyama T Sudoand J Mizuguchi ldquoCalpain-induced Bax-cleavage product is amore potent inducer of apoptotic cell death thanwild-type BaxrdquoCancer Letters vol 189 no 2 pp 221ndash230 2003
[26] Y R Meng S J Kang and D A Fishman ldquoLysophosphatidicacid inhibits anti-Fas-mediated apoptosis enhanced by actindepolymerization in epithelial ovarian cancerrdquo FEBS Lettersvol 579 no 5 pp 1311ndash1319 2005
[27] Y Meng S Kang J So S Reierstad and D A FishmanldquoTranslocation of Fas by LPA prevents ovarian cancer cells fromanti-Fas-induced apoptosisrdquo Gynecologic Oncology vol 96 no2 pp 462ndash469 2005
[28] X Wan Y Yokoyama A Shinohara Y Takahashi and TTamaya ldquoPTEN augments staurosporine-induced apoptosis inPTEN-null Ishikawa cells by downregulating PI3KAkt signal-ing pathwayrdquo Cell Death and Differentiation vol 9 no 4 pp414ndash420 2002
[29] J Yu Y Zhang J Mcilroy T Rordorf-Nikolic G A Orr and JM Backer ldquoRegulation of the p85p110 phosphatidylinositol 31015840-kinase stabilization and inhibition of the p110120572 catalytic subunitby the p85 regulatory subunitrdquoMolecular and Cellular Biologyvol 18 no 3 pp 1379ndash1387 1998
[30] M S Kim J H Kim Y Bak et al ldquo24-bis (p-hydroxyphenyl)-2-butenal (HPB242) induces apoptosis via modulating E7expression and inhibition of PI3KAkt pathway in SiHa humancervical cancer cellsrdquo Nutrition and Cancer vol 64 no 8 pp1236ndash1244 2012
[31] F Cui X L Li X Y Zhu et al ldquoMiR-125b inhibits tumorgrowth and promotes apoptosis of cervical cancer cells bytargeting phosphoinositide 3-kinase catalytic subunit deltardquoCellular Physiology and Biochemistry vol 30 no 5 pp 1310ndash1318 2012
[32] S Kang S M Dong B-R Kim et al ldquoThioridazine inducesapoptosis by targeting the PI3KAktmTOR pathway in cervicaland endometrial cancer cellsrdquo Apoptosis vol 17 no 9 pp 989ndash997 2012
[33] O Thanaketpaisarn P Waiwut H Sakurai and I Saiki ldquoArte-sunate enhances TRAIL-induced apoptosis in human cervicalcarcinoma cells through inhibition of the NF-120581B and PI3KAktsignaling pathwaysrdquo International Journal of Oncology vol 39no 1 pp 279ndash285 2011
[34] XWang J-H Ren F Lin et al ldquoStathmin is involved in arsenictrioxide-induced apoptosis in human cervical cancer cell linesvia PI3K linked signal pathwayrdquo Cancer Biology and Therapyvol 10 no 6 pp 632ndash643 2010
[35] C W Lee N H Kim H K Choi et al ldquoLysophosphatidicacid-induced c-fos up-regulation involves cyclic AMP responseelement-binding protein activated by mitogen- and stress-activated protein kinase-1rdquo Journal of Cellular Biochemistry vol104 no 3 pp 785ndash794 2008
[36] DW Hommes M P Peppelenbosch and S J H van DeventerldquoMitogen activated protein (MAP) kinase signal transduction
pathways and novel anti-inflammatory targetsrdquoGut vol 52 no1 pp 144ndash151 2003
[37] M Mukai F Imamura M Ayaki et al ldquoInhibition of tumorinvasion andmetastasis by a novel lysophosphatidic acid (cyclicLPA)rdquo International Journal of Cancer vol 81 no 6 pp 918ndash9221999
[38] E-Y Huang Y-F Chen Y-M Chen et al ldquoA novel radiore-sistant mechanism of galectin-1 mediated by H-Ras-dependentpathways in cervical cancer cellsrdquo Cell Death and Disease vol3 no 1 article e251 2012
[39] W-H Su P-C Chuang E-Y Huang and K D YangldquoRadiation-induced increase in cell migration and metastaticpotential of cervical cancer cells operates via theK-ras pathwayrdquoThe American Journal of Pathology vol 180 no 2 pp 862ndash8712012
[40] H Rincon-Arano R Rosales N Mora A Rodriguez-Castaneda and C Rosales ldquoR-Ras promotes tumor growthof cervical epithelial cellsrdquo Cancer vol 97 no 3 pp 575ndash5852003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 9
Fold
chan
ge o
f ban
d in
tens
ity
5
4
3
2
1
00 01 1 5 20 50
LPA (120583M)
0 01 1 5 20 50 LPA (120583M)
P-AKT
AKT
(a)
Fold
chan
ge o
f ban
d in
tens
ity
P-AKT
Total-AKT
4
35
3
25
2
15
1
05
00 5min 10min 30min 1h 2h 3h 4h
Time
0 5min 10min 30min 1h 2h 3h 4h Time
AKT activation
(b)
Figure 6 Western blotting analysis of phosphorylated AKT in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01120583M 1 120583M 5 120583M 20 120583M and 50 120583M for 1 hour Western blotting was conducted to detect the expressionof phosphorylated AKT Total AKT was detected and the results were used as protein input control Densitometry analysis was carried outfor quantification purpose LPA treatment led to increased phosphorylation of AKT in a dose-dependent manner (b) Time-response effectof LPA on the phosphorylation of AKT Hela cells were treated with LPA at 20 120583M for different times including 5min 10min 30min 1 hour2 hours 3 hours and 4 hours Western blotting was used to detect the expression of phosphorylated AKT Compared to the total AKT LPAtreatment activated the phosphorylation of AKT in a time-dependent manner
25
20
15
10
05
00
Fold
chan
ge
0 01 10 50 20 50
LPA (120583M)
0 01 10 05 20 50 LPA (120583M)
P-ERK
ERK
(a)
25
20
15
10
05
00
Fold
chan
ge
0 min5 min10 min30 1h 2h 3h 4hTime
0 min5 min10 min30 1h 2h 3h 4h Time
P-ERK
ERK
(b)
Figure 7 Western blotting analysis of phosphorylated ERK in cells treated with LPA (a) Hela cells were treated with LPA at differentconcentrations including 0 01 120583M 1 120583M 5120583M 20 120583M and 50 120583M for 1 hour Western blotting was performed to detect the levels ofphosphorylated ERK Total ERK was used as protein input control The density of bands was quantified and compared LPA activated thephosphorylation of ERK in a dose-dependent manner (b) Hela cells were treated with LPA at 20120583M for different times including 5min10min 30min 1 hour 2 hours 3 hours and 4 hours A time-dependent increase in the ERK phosphorylation was detected
10 BioMed Research International
Some studies found that cell apoptosis induced by cis-platinwasmediated by death receptors signaling pathway [2021] while others claimed that it was mediated by mitochon-drial pathway [22 23] The discrepancy in previous studiesmay be caused by the divergent cell lines methodologiesand conditions used by different groups In this study weobserved that cisplatin significantly induced the apoptosis ofHela cells in a concentration-dependent manner Cisplatintreatment was accompanied by increased Fas-1 and Baxexpression and the increased enzyme activity of caspase-3 Meanwhile cisplatin inhibited Bcl-2 expression Theseobservations suggested that cisplatin-induced apoptosis ofHela cell may be mediated by both intrinsic and extrinsicpathways Indeed extensive cross talks between the two path-ways have been identified [24 25] Simultaneous activationof the two pathways has been observed in many types of cellsfollowing various treatments
Our previous study found that both ovarian cancer cellsand peritoneal mesothelial cells contribute to the elevatedLPA levels in ovarian cancer ascites Under normal cultureconditions ovarian cancer cells and prostate cancer cellsbut not breast cancer cell lines constitutively synthesize andrelease significant amounts of LPA to the culture medium Amajor finding of this study is that LPA was able to rescue theHela cells apoptosis induced by cisplatin The protective roleof LPA against apoptosis was observed in ovarian cancer cells[26]We observed that LPA treatment blocked the alterationsof Bcl-2 Fas-1 and Bax expression that were induced bycisplatin LPA also significantly inhibited the upregulationof the caspase-3 enzyme activity induced by cisplatin Ourobservations of LPA effect on cervical cancer were consistentwith other reports on LPA effects in different kinds of cellsMeng et al found that LPA may protect epithelial ovariancancer from immune cell attack and cytoskeleton disruptingreagents induced apoptosis through multiple pathways andLPA inhibited anti-Fas-induced apoptosis enhanced by actindepolymerization [26 27] Actin depolymerization acceler-ated caspase-8 activation while LPA inhibited the associationand activation of caspase-8 at the DISC in epithelial ovariancancer OVCAR3 cells Thus our observations have raisedan interesting question on the potential role of LPA forthe development of chemoresistance by cervical cancer cellsIn addition could LPA be an effective biomarker for theprediction of chemoresistance in cervical cancer Furtherstudies are required to answer these questions
In this study three specific kinase inhibitors were appliedto determine the signaling pathways involved in the pro-tection effect of LPA on apoptosis It was found that PI3Kinhibitor (LY290042) almost completely blocked the LPAprotection against apoptosis induced by cisplatin Subsequentexperiments confirmed that LPA treatment of Hela cells ledto increased phosphorylation of AKT in a dose- and time-dependent manner This indicated that LPA protected Helacells from apoptosis induced by cisplatin This effect of LPAmay be through the activation of PI3KAKT pathway Thisfinding appears to be consistent with published data showingthat PI3KAKT pathway plays a key role in the regulationof cell apoptosis [28 29] Wan et al observed that PI3Kinhibition augmented staurosporine-induced apoptosis in
the endometrial carcinoma cell line [28] In that experimentstaurosporine reduced the levels of phosphorylation in AKTand Bad In a study by Kim et al 24-bis(p-hydroxyphenyl)-2-butenal (HPB242) induced apoptosis via the inhibition ofPI3KAKT pathway in human cervical cancer cells HPB242treatment decreased phosphatidyl inositol 3-kinase and p-AKT expression levels demonstrating that this survivalpathway may also be inhibited by HPB242 [30] In anotherstudy Cui et al found that MiR-125b inhibited tumor growthand promoted apoptosis of cervical cancer cells by targetingphosphoinositide 3-kinase catalytic subunit delta Overex-pression of MiR-125b in Hela cervical cancer cells inducedapoptosis and downregulated the expression of PIK3CDand Phospho-AKT [31] Kang et al found that thioridazineinduces apoptosis by targeting the PI3KAKTmTOR path-way in cervical cancer cells [32] Thanaketpaisarn et alfound that Artesunate enhanced TRAIL-induced apoptosisin human cervical carcinoma cells through inhibition of thePI3KAKT and NF-120581B signaling pathways [33] Wang et alfound that Stathmin is involved in arsenic trioxide-inducedapoptosis in human cervical cancer cell lines via PI3K linkedsignal pathway As2O3-induced stathmin downregulation ismediated through the phosphatidylinositol-3-kinase (PI3K)signaling pathway PI3K inhibitor effectively attenuated thestathmin downregulation and cell apoptosis upon As2O3-treatment [34] Recurrent and metastatic cervical cancersoften acquire chemoresistance to cisplatin Our findingpointed to a possible sensitization scheme through combinedadministration of PI3K inhibitor and cisplatin
It should be pointed out that while our current studyfocused on the apoptotic pathways andmechanisms the LPAeffects on cell cycle control and cell proliferation should notbe excluded In fact the PI3KAKT pathway is known to playan active role in the control of cell cycle [35] We found thatMEK1 inhibitor (PD98059) significantly inhibited the effectof LPA on the proliferation of Hela cells RasRaf1MEKERKsignal transduction pathway is one of the most importantmembers of the mitogen-activated protein kinase pathwaysregulating cell proliferation [36] MEK1 a MAPK representsa key molecule of the RasRaf1MEKERK signal pathway[37] Several lines of evidence suggest that in cervical cancerthe RasRaf1MEKERK pathway but not the JNK pathwayor the p38 MAPK pathway is the major regulator of cellproliferation [38ndash40]
In summary we demonstrated that cisplatin inducedapoptosis of Hela cell via the downregulation of Bcl-2 andupregulation of Bax Fas-L and increased enzyme activityof caspase-3 LPA was able to block the alterations in theapoptotic factors caused by cisplatin and ultimately reversedthe apoptosis induced by cisplatin Cisplatin-induced cellapoptosis appeared to be partially dependent on PI3KAKTpathway and AKT inhibitor was able to reverse the pro-tective effects of LPA against cisplatin-induced apoptosisLPA stimulated the proliferation of Hela cells throughthe RasRaf1MEKERK pathway These data enriched ourknowledge regarding the LPA activity in cervical cancer cellsIn addition the finding on the positive impact of PI3KAKTinhibitor on the apoptotic effects of cisplatin has provideduseful information for the design of sensitization strategy in
BioMed Research International 11
cervical cancer patients especially those with high levels ofLPA
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Yanxia Sui and Ya Yang contributed equally to this work
Acknowledgments
This study is supported by the following funding sourcesThe National Natural Science Foundations of China (JuanRen 31201060C0709 Juan Ren 30973175H1621 and JuanRen 81172490H1621) Program for New Century ExcellentTalents in University (Juan Ren NCET-12-0440) Scientificand Technological Research Foundation of Shaanxi Province(Juan Ren 2012K13-01-06 Juan Ren 2007K09-09) Scien-tific Research Foundation for the Returned overseas Chi-nese Scholars of State Education Ministry (Juan Ren 0601-18920006) Research Foundation of Health Department ofShaanrsquoxi Province (Juan Ren 2010D41) Qing Nian Jiao ShiGenZongJiHua ofXirsquoan JiaotongUniversity (JuanRen 2012)Program for Changjiang Scholars and Innovative ResearchTeam inUniversity (Zongfang Li PCSIRT1171) andResearchFoundation of Xirsquoan Jiao Tong University of China (JuanRen)
References
[1] J R Erickson Y Hasegawa X Fang et al ldquoLysophosphatidicacid and ovarian cancer a paradigm for tumorogenesis andpatient managementrdquo Prostaglandins and Other Lipid Media-tors vol 64 no 1ndash4 pp 63ndash81 2001
[2] Y Xu Z Z Shen DWWiper et al ldquoLysophosphatidic acid as apotential biomarker for ovarian and other gynecologic cancersrdquoThe Journal of the American Medical Association vol 280 no 8pp 719ndash723 1998
[3] J S Karliner ldquoLysophospholipids and the cardiovascular sys-temrdquoBiochimica et Biophysica ActamdashMolecular and Cell Biologyof Lipids vol 1582 no 1ndash3 pp 216ndash221 2002
[4] C Schulze C Smales L L Rubin and J M StaddonldquoLysophosphatidic acid increases tight junction permeability incultured brain endothelial cellsrdquo Journal of Neurochemistry vol68 no 3 pp 991ndash1000 1997
[5] I Ishii N Fukushima X Ye and J Chun ldquoLysophospholipidreceptors signaling and biologyrdquo Annual Review of Biochem-istry vol 73 pp 321ndash354 2004
[6] T S Panetti ldquoDifferential effects of sphingosine 1-phosphateand lysophosphatidic acid on endothelial cellsrdquo Biochimica etBiophysica ActamdashMolecular and Cell Biology of Lipids vol 1582no 1ndash3 pp 190ndash196 2002
[7] Z Z Shen M Z Wu P Elson et al ldquoFatty acid composition oflysophosphatidic acid and lysophosphatidylinositol in plasmafrom patients with ovarian cancer and other gynecologicaldiseasesrdquo Gynecologic Oncology vol 83 no 1 pp 25ndash30 2001
[8] M M Murph ldquoAgonist-induced endocytosis of lysophospha-tidic acid-coupled LPA1EDG-2 receptors via a dynamin2- andRab5-dependent pathwayrdquo Journal of Cell Science vol 116 no10 pp 1969ndash1980 2003
[9] L M Baudhuin K L Cristina J Lu and Y Xu ldquoAkt activationinduced by lysophosphatidic acid and sphingosine-1-phosphaterequires both mitogen-activated protein kinase kinase andp38 mitogen-activated protein kinase and is cell-line specificrdquoMolecular Pharmacology vol 62 no 3 pp 660ndash671 2002
[10] R-J Chen S-U Chen C-H Chou andM-C Lin ldquoLysophos-phatidic acid receptor 23-mediated IL-8-dependent angiogen-esis in cervical cancer cellsrdquo International Journal of Cancer vol131 no 4 pp 789ndash802 2012
[11] J W Choi S Lim Y-S Oh et al ldquoSubtype-specific role ofphospholipase C-120573 in bradykinin and LPA signaling throughdifferential binding of different PDZ scaffold proteinsrdquo CellularSignalling vol 22 no 7 pp 1153ndash1161 2010
[12] Y Kishi S Okudaira M Tanaka et al ldquoAutotaxin is over-expressed in glioblastoma multiforme and contributes to cellmotility of glioblastoma by converting lysophosphatidylcholineTO lysophosphatidic acidrdquoThe Journal of Biological Chemistryvol 281 no 25 pp 17492ndash17500 2006
[13] A Eastman ldquoThe formation isolation and characterization ofDNA adducts produced by anticancer platinum complexesrdquoPharmacology andTherapeutics vol 34 no 2 pp 155ndash166 1987
[14] K Wozniak and J Błasiak ldquoRecognition and repair of DNA-cisplatin adductsrdquo Acta Biochimica Polonica vol 49 no 3 pp583ndash596 2002
[15] A Basu and S Krishnamurthy ldquoCellular responses to cisplatin-induced DNA damagerdquo Journal of Nucleic Acids vol 2010Article ID 201367 16 pages 2010
[16] A Efeyan and M Serrano ldquop53 guardian of the genome andpoliceman of the oncogenesrdquo Cell Cycle vol 6 no 9 pp 1006ndash1010 2007
[17] O Kutuk E D Arisan T Tezil M C Shoshan and H BasagaldquoCisplatin overcomesBcl-2-mediated resistance to apoptosis viapreferential engagement of Bak critical role of Noxa-mediatedlipid peroxidationrdquo Carcinogenesis vol 30 no 9 pp 1517ndash15272009
[18] C M Yan E Q Chai H Y Cai G Miao and W MaldquoOleuropein induces apoptosis via activation of caspases andsuppression of phosphatidylinositol 3-kinaseprotein kinaseB pathway in HepG2 human hepatoma cell linerdquo MolecularMedicine Reports vol 11 pp 4617ndash4624 2015
[19] X Y Wang X S Fan L Cai S Liu X F Cong and X ChenldquoLysophosphatidic acid rescues bone mesenchymal stem cellsfrom hydrogen peroxide-induced apoptosisrdquo Apoptosis vol 20no 3 pp 273ndash284 2015
[20] A Brozovic G Fritz M Christmann et al ldquoLong-term activa-tion of SAPKJNK p38 kinase and fas-L expression by cisplatinis attenuated in human carcinoma cells that acquired drugresistancerdquo International Journal of Cancer vol 112 no 6 pp974ndash985 2004
[21] Y Q Liu H Xing X B Han et al ldquoApoptosis of HeLa cellsinduced by cisplatin and its mechanismrdquo Journal of HuazhongUniversity of Science and TechnologymdashMedical Science vol 28no 2 pp 197ndash199 2008
[22] K L Jin J-Y Park E J Noh et al ldquoThe effect of combinedtreatment with cisplatin and histone deacetylase inhibitors onHeLa cellsrdquo Journal of Gynecologic Oncology vol 21 no 4 pp262ndash268 2010
12 BioMed Research International
[23] T Sakaeda T Nakamura M Hirai et al ldquoMDR1 up-regulatedby apoptotic stimuli suppresses apoptotic signalingrdquo Pharma-ceutical Research vol 19 no 9 pp 1323ndash1329 2002
[24] K Kandasamy S M Srinivasula E S Alnemri et al ldquoInvolve-ment of proapoptotic molecules Bax and Bak in tumor necro-sis factor-related apoptosis-inducing ligand (TRAIL)-inducedmitochondrial disruption and apoptosis differential regulationof cytochrome c and SmacDIABLO releaserdquo Cancer Researchvol 63 no 7 pp 1712ndash1721 2003
[25] H Toyota N Yanase T Yoshimoto M Moriyama T Sudoand J Mizuguchi ldquoCalpain-induced Bax-cleavage product is amore potent inducer of apoptotic cell death thanwild-type BaxrdquoCancer Letters vol 189 no 2 pp 221ndash230 2003
[26] Y R Meng S J Kang and D A Fishman ldquoLysophosphatidicacid inhibits anti-Fas-mediated apoptosis enhanced by actindepolymerization in epithelial ovarian cancerrdquo FEBS Lettersvol 579 no 5 pp 1311ndash1319 2005
[27] Y Meng S Kang J So S Reierstad and D A FishmanldquoTranslocation of Fas by LPA prevents ovarian cancer cells fromanti-Fas-induced apoptosisrdquo Gynecologic Oncology vol 96 no2 pp 462ndash469 2005
[28] X Wan Y Yokoyama A Shinohara Y Takahashi and TTamaya ldquoPTEN augments staurosporine-induced apoptosis inPTEN-null Ishikawa cells by downregulating PI3KAkt signal-ing pathwayrdquo Cell Death and Differentiation vol 9 no 4 pp414ndash420 2002
[29] J Yu Y Zhang J Mcilroy T Rordorf-Nikolic G A Orr and JM Backer ldquoRegulation of the p85p110 phosphatidylinositol 31015840-kinase stabilization and inhibition of the p110120572 catalytic subunitby the p85 regulatory subunitrdquoMolecular and Cellular Biologyvol 18 no 3 pp 1379ndash1387 1998
[30] M S Kim J H Kim Y Bak et al ldquo24-bis (p-hydroxyphenyl)-2-butenal (HPB242) induces apoptosis via modulating E7expression and inhibition of PI3KAkt pathway in SiHa humancervical cancer cellsrdquo Nutrition and Cancer vol 64 no 8 pp1236ndash1244 2012
[31] F Cui X L Li X Y Zhu et al ldquoMiR-125b inhibits tumorgrowth and promotes apoptosis of cervical cancer cells bytargeting phosphoinositide 3-kinase catalytic subunit deltardquoCellular Physiology and Biochemistry vol 30 no 5 pp 1310ndash1318 2012
[32] S Kang S M Dong B-R Kim et al ldquoThioridazine inducesapoptosis by targeting the PI3KAktmTOR pathway in cervicaland endometrial cancer cellsrdquo Apoptosis vol 17 no 9 pp 989ndash997 2012
[33] O Thanaketpaisarn P Waiwut H Sakurai and I Saiki ldquoArte-sunate enhances TRAIL-induced apoptosis in human cervicalcarcinoma cells through inhibition of the NF-120581B and PI3KAktsignaling pathwaysrdquo International Journal of Oncology vol 39no 1 pp 279ndash285 2011
[34] XWang J-H Ren F Lin et al ldquoStathmin is involved in arsenictrioxide-induced apoptosis in human cervical cancer cell linesvia PI3K linked signal pathwayrdquo Cancer Biology and Therapyvol 10 no 6 pp 632ndash643 2010
[35] C W Lee N H Kim H K Choi et al ldquoLysophosphatidicacid-induced c-fos up-regulation involves cyclic AMP responseelement-binding protein activated by mitogen- and stress-activated protein kinase-1rdquo Journal of Cellular Biochemistry vol104 no 3 pp 785ndash794 2008
[36] DW Hommes M P Peppelenbosch and S J H van DeventerldquoMitogen activated protein (MAP) kinase signal transduction
pathways and novel anti-inflammatory targetsrdquoGut vol 52 no1 pp 144ndash151 2003
[37] M Mukai F Imamura M Ayaki et al ldquoInhibition of tumorinvasion andmetastasis by a novel lysophosphatidic acid (cyclicLPA)rdquo International Journal of Cancer vol 81 no 6 pp 918ndash9221999
[38] E-Y Huang Y-F Chen Y-M Chen et al ldquoA novel radiore-sistant mechanism of galectin-1 mediated by H-Ras-dependentpathways in cervical cancer cellsrdquo Cell Death and Disease vol3 no 1 article e251 2012
[39] W-H Su P-C Chuang E-Y Huang and K D YangldquoRadiation-induced increase in cell migration and metastaticpotential of cervical cancer cells operates via theK-ras pathwayrdquoThe American Journal of Pathology vol 180 no 2 pp 862ndash8712012
[40] H Rincon-Arano R Rosales N Mora A Rodriguez-Castaneda and C Rosales ldquoR-Ras promotes tumor growthof cervical epithelial cellsrdquo Cancer vol 97 no 3 pp 575ndash5852003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 BioMed Research International
Some studies found that cell apoptosis induced by cis-platinwasmediated by death receptors signaling pathway [2021] while others claimed that it was mediated by mitochon-drial pathway [22 23] The discrepancy in previous studiesmay be caused by the divergent cell lines methodologiesand conditions used by different groups In this study weobserved that cisplatin significantly induced the apoptosis ofHela cells in a concentration-dependent manner Cisplatintreatment was accompanied by increased Fas-1 and Baxexpression and the increased enzyme activity of caspase-3 Meanwhile cisplatin inhibited Bcl-2 expression Theseobservations suggested that cisplatin-induced apoptosis ofHela cell may be mediated by both intrinsic and extrinsicpathways Indeed extensive cross talks between the two path-ways have been identified [24 25] Simultaneous activationof the two pathways has been observed in many types of cellsfollowing various treatments
Our previous study found that both ovarian cancer cellsand peritoneal mesothelial cells contribute to the elevatedLPA levels in ovarian cancer ascites Under normal cultureconditions ovarian cancer cells and prostate cancer cellsbut not breast cancer cell lines constitutively synthesize andrelease significant amounts of LPA to the culture medium Amajor finding of this study is that LPA was able to rescue theHela cells apoptosis induced by cisplatin The protective roleof LPA against apoptosis was observed in ovarian cancer cells[26]We observed that LPA treatment blocked the alterationsof Bcl-2 Fas-1 and Bax expression that were induced bycisplatin LPA also significantly inhibited the upregulationof the caspase-3 enzyme activity induced by cisplatin Ourobservations of LPA effect on cervical cancer were consistentwith other reports on LPA effects in different kinds of cellsMeng et al found that LPA may protect epithelial ovariancancer from immune cell attack and cytoskeleton disruptingreagents induced apoptosis through multiple pathways andLPA inhibited anti-Fas-induced apoptosis enhanced by actindepolymerization [26 27] Actin depolymerization acceler-ated caspase-8 activation while LPA inhibited the associationand activation of caspase-8 at the DISC in epithelial ovariancancer OVCAR3 cells Thus our observations have raisedan interesting question on the potential role of LPA forthe development of chemoresistance by cervical cancer cellsIn addition could LPA be an effective biomarker for theprediction of chemoresistance in cervical cancer Furtherstudies are required to answer these questions
In this study three specific kinase inhibitors were appliedto determine the signaling pathways involved in the pro-tection effect of LPA on apoptosis It was found that PI3Kinhibitor (LY290042) almost completely blocked the LPAprotection against apoptosis induced by cisplatin Subsequentexperiments confirmed that LPA treatment of Hela cells ledto increased phosphorylation of AKT in a dose- and time-dependent manner This indicated that LPA protected Helacells from apoptosis induced by cisplatin This effect of LPAmay be through the activation of PI3KAKT pathway Thisfinding appears to be consistent with published data showingthat PI3KAKT pathway plays a key role in the regulationof cell apoptosis [28 29] Wan et al observed that PI3Kinhibition augmented staurosporine-induced apoptosis in
the endometrial carcinoma cell line [28] In that experimentstaurosporine reduced the levels of phosphorylation in AKTand Bad In a study by Kim et al 24-bis(p-hydroxyphenyl)-2-butenal (HPB242) induced apoptosis via the inhibition ofPI3KAKT pathway in human cervical cancer cells HPB242treatment decreased phosphatidyl inositol 3-kinase and p-AKT expression levels demonstrating that this survivalpathway may also be inhibited by HPB242 [30] In anotherstudy Cui et al found that MiR-125b inhibited tumor growthand promoted apoptosis of cervical cancer cells by targetingphosphoinositide 3-kinase catalytic subunit delta Overex-pression of MiR-125b in Hela cervical cancer cells inducedapoptosis and downregulated the expression of PIK3CDand Phospho-AKT [31] Kang et al found that thioridazineinduces apoptosis by targeting the PI3KAKTmTOR path-way in cervical cancer cells [32] Thanaketpaisarn et alfound that Artesunate enhanced TRAIL-induced apoptosisin human cervical carcinoma cells through inhibition of thePI3KAKT and NF-120581B signaling pathways [33] Wang et alfound that Stathmin is involved in arsenic trioxide-inducedapoptosis in human cervical cancer cell lines via PI3K linkedsignal pathway As2O3-induced stathmin downregulation ismediated through the phosphatidylinositol-3-kinase (PI3K)signaling pathway PI3K inhibitor effectively attenuated thestathmin downregulation and cell apoptosis upon As2O3-treatment [34] Recurrent and metastatic cervical cancersoften acquire chemoresistance to cisplatin Our findingpointed to a possible sensitization scheme through combinedadministration of PI3K inhibitor and cisplatin
It should be pointed out that while our current studyfocused on the apoptotic pathways andmechanisms the LPAeffects on cell cycle control and cell proliferation should notbe excluded In fact the PI3KAKT pathway is known to playan active role in the control of cell cycle [35] We found thatMEK1 inhibitor (PD98059) significantly inhibited the effectof LPA on the proliferation of Hela cells RasRaf1MEKERKsignal transduction pathway is one of the most importantmembers of the mitogen-activated protein kinase pathwaysregulating cell proliferation [36] MEK1 a MAPK representsa key molecule of the RasRaf1MEKERK signal pathway[37] Several lines of evidence suggest that in cervical cancerthe RasRaf1MEKERK pathway but not the JNK pathwayor the p38 MAPK pathway is the major regulator of cellproliferation [38ndash40]
In summary we demonstrated that cisplatin inducedapoptosis of Hela cell via the downregulation of Bcl-2 andupregulation of Bax Fas-L and increased enzyme activityof caspase-3 LPA was able to block the alterations in theapoptotic factors caused by cisplatin and ultimately reversedthe apoptosis induced by cisplatin Cisplatin-induced cellapoptosis appeared to be partially dependent on PI3KAKTpathway and AKT inhibitor was able to reverse the pro-tective effects of LPA against cisplatin-induced apoptosisLPA stimulated the proliferation of Hela cells throughthe RasRaf1MEKERK pathway These data enriched ourknowledge regarding the LPA activity in cervical cancer cellsIn addition the finding on the positive impact of PI3KAKTinhibitor on the apoptotic effects of cisplatin has provideduseful information for the design of sensitization strategy in
BioMed Research International 11
cervical cancer patients especially those with high levels ofLPA
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Yanxia Sui and Ya Yang contributed equally to this work
Acknowledgments
This study is supported by the following funding sourcesThe National Natural Science Foundations of China (JuanRen 31201060C0709 Juan Ren 30973175H1621 and JuanRen 81172490H1621) Program for New Century ExcellentTalents in University (Juan Ren NCET-12-0440) Scientificand Technological Research Foundation of Shaanxi Province(Juan Ren 2012K13-01-06 Juan Ren 2007K09-09) Scien-tific Research Foundation for the Returned overseas Chi-nese Scholars of State Education Ministry (Juan Ren 0601-18920006) Research Foundation of Health Department ofShaanrsquoxi Province (Juan Ren 2010D41) Qing Nian Jiao ShiGenZongJiHua ofXirsquoan JiaotongUniversity (JuanRen 2012)Program for Changjiang Scholars and Innovative ResearchTeam inUniversity (Zongfang Li PCSIRT1171) andResearchFoundation of Xirsquoan Jiao Tong University of China (JuanRen)
References
[1] J R Erickson Y Hasegawa X Fang et al ldquoLysophosphatidicacid and ovarian cancer a paradigm for tumorogenesis andpatient managementrdquo Prostaglandins and Other Lipid Media-tors vol 64 no 1ndash4 pp 63ndash81 2001
[2] Y Xu Z Z Shen DWWiper et al ldquoLysophosphatidic acid as apotential biomarker for ovarian and other gynecologic cancersrdquoThe Journal of the American Medical Association vol 280 no 8pp 719ndash723 1998
[3] J S Karliner ldquoLysophospholipids and the cardiovascular sys-temrdquoBiochimica et Biophysica ActamdashMolecular and Cell Biologyof Lipids vol 1582 no 1ndash3 pp 216ndash221 2002
[4] C Schulze C Smales L L Rubin and J M StaddonldquoLysophosphatidic acid increases tight junction permeability incultured brain endothelial cellsrdquo Journal of Neurochemistry vol68 no 3 pp 991ndash1000 1997
[5] I Ishii N Fukushima X Ye and J Chun ldquoLysophospholipidreceptors signaling and biologyrdquo Annual Review of Biochem-istry vol 73 pp 321ndash354 2004
[6] T S Panetti ldquoDifferential effects of sphingosine 1-phosphateand lysophosphatidic acid on endothelial cellsrdquo Biochimica etBiophysica ActamdashMolecular and Cell Biology of Lipids vol 1582no 1ndash3 pp 190ndash196 2002
[7] Z Z Shen M Z Wu P Elson et al ldquoFatty acid composition oflysophosphatidic acid and lysophosphatidylinositol in plasmafrom patients with ovarian cancer and other gynecologicaldiseasesrdquo Gynecologic Oncology vol 83 no 1 pp 25ndash30 2001
[8] M M Murph ldquoAgonist-induced endocytosis of lysophospha-tidic acid-coupled LPA1EDG-2 receptors via a dynamin2- andRab5-dependent pathwayrdquo Journal of Cell Science vol 116 no10 pp 1969ndash1980 2003
[9] L M Baudhuin K L Cristina J Lu and Y Xu ldquoAkt activationinduced by lysophosphatidic acid and sphingosine-1-phosphaterequires both mitogen-activated protein kinase kinase andp38 mitogen-activated protein kinase and is cell-line specificrdquoMolecular Pharmacology vol 62 no 3 pp 660ndash671 2002
[10] R-J Chen S-U Chen C-H Chou andM-C Lin ldquoLysophos-phatidic acid receptor 23-mediated IL-8-dependent angiogen-esis in cervical cancer cellsrdquo International Journal of Cancer vol131 no 4 pp 789ndash802 2012
[11] J W Choi S Lim Y-S Oh et al ldquoSubtype-specific role ofphospholipase C-120573 in bradykinin and LPA signaling throughdifferential binding of different PDZ scaffold proteinsrdquo CellularSignalling vol 22 no 7 pp 1153ndash1161 2010
[12] Y Kishi S Okudaira M Tanaka et al ldquoAutotaxin is over-expressed in glioblastoma multiforme and contributes to cellmotility of glioblastoma by converting lysophosphatidylcholineTO lysophosphatidic acidrdquoThe Journal of Biological Chemistryvol 281 no 25 pp 17492ndash17500 2006
[13] A Eastman ldquoThe formation isolation and characterization ofDNA adducts produced by anticancer platinum complexesrdquoPharmacology andTherapeutics vol 34 no 2 pp 155ndash166 1987
[14] K Wozniak and J Błasiak ldquoRecognition and repair of DNA-cisplatin adductsrdquo Acta Biochimica Polonica vol 49 no 3 pp583ndash596 2002
[15] A Basu and S Krishnamurthy ldquoCellular responses to cisplatin-induced DNA damagerdquo Journal of Nucleic Acids vol 2010Article ID 201367 16 pages 2010
[16] A Efeyan and M Serrano ldquop53 guardian of the genome andpoliceman of the oncogenesrdquo Cell Cycle vol 6 no 9 pp 1006ndash1010 2007
[17] O Kutuk E D Arisan T Tezil M C Shoshan and H BasagaldquoCisplatin overcomesBcl-2-mediated resistance to apoptosis viapreferential engagement of Bak critical role of Noxa-mediatedlipid peroxidationrdquo Carcinogenesis vol 30 no 9 pp 1517ndash15272009
[18] C M Yan E Q Chai H Y Cai G Miao and W MaldquoOleuropein induces apoptosis via activation of caspases andsuppression of phosphatidylinositol 3-kinaseprotein kinaseB pathway in HepG2 human hepatoma cell linerdquo MolecularMedicine Reports vol 11 pp 4617ndash4624 2015
[19] X Y Wang X S Fan L Cai S Liu X F Cong and X ChenldquoLysophosphatidic acid rescues bone mesenchymal stem cellsfrom hydrogen peroxide-induced apoptosisrdquo Apoptosis vol 20no 3 pp 273ndash284 2015
[20] A Brozovic G Fritz M Christmann et al ldquoLong-term activa-tion of SAPKJNK p38 kinase and fas-L expression by cisplatinis attenuated in human carcinoma cells that acquired drugresistancerdquo International Journal of Cancer vol 112 no 6 pp974ndash985 2004
[21] Y Q Liu H Xing X B Han et al ldquoApoptosis of HeLa cellsinduced by cisplatin and its mechanismrdquo Journal of HuazhongUniversity of Science and TechnologymdashMedical Science vol 28no 2 pp 197ndash199 2008
[22] K L Jin J-Y Park E J Noh et al ldquoThe effect of combinedtreatment with cisplatin and histone deacetylase inhibitors onHeLa cellsrdquo Journal of Gynecologic Oncology vol 21 no 4 pp262ndash268 2010
12 BioMed Research International
[23] T Sakaeda T Nakamura M Hirai et al ldquoMDR1 up-regulatedby apoptotic stimuli suppresses apoptotic signalingrdquo Pharma-ceutical Research vol 19 no 9 pp 1323ndash1329 2002
[24] K Kandasamy S M Srinivasula E S Alnemri et al ldquoInvolve-ment of proapoptotic molecules Bax and Bak in tumor necro-sis factor-related apoptosis-inducing ligand (TRAIL)-inducedmitochondrial disruption and apoptosis differential regulationof cytochrome c and SmacDIABLO releaserdquo Cancer Researchvol 63 no 7 pp 1712ndash1721 2003
[25] H Toyota N Yanase T Yoshimoto M Moriyama T Sudoand J Mizuguchi ldquoCalpain-induced Bax-cleavage product is amore potent inducer of apoptotic cell death thanwild-type BaxrdquoCancer Letters vol 189 no 2 pp 221ndash230 2003
[26] Y R Meng S J Kang and D A Fishman ldquoLysophosphatidicacid inhibits anti-Fas-mediated apoptosis enhanced by actindepolymerization in epithelial ovarian cancerrdquo FEBS Lettersvol 579 no 5 pp 1311ndash1319 2005
[27] Y Meng S Kang J So S Reierstad and D A FishmanldquoTranslocation of Fas by LPA prevents ovarian cancer cells fromanti-Fas-induced apoptosisrdquo Gynecologic Oncology vol 96 no2 pp 462ndash469 2005
[28] X Wan Y Yokoyama A Shinohara Y Takahashi and TTamaya ldquoPTEN augments staurosporine-induced apoptosis inPTEN-null Ishikawa cells by downregulating PI3KAkt signal-ing pathwayrdquo Cell Death and Differentiation vol 9 no 4 pp414ndash420 2002
[29] J Yu Y Zhang J Mcilroy T Rordorf-Nikolic G A Orr and JM Backer ldquoRegulation of the p85p110 phosphatidylinositol 31015840-kinase stabilization and inhibition of the p110120572 catalytic subunitby the p85 regulatory subunitrdquoMolecular and Cellular Biologyvol 18 no 3 pp 1379ndash1387 1998
[30] M S Kim J H Kim Y Bak et al ldquo24-bis (p-hydroxyphenyl)-2-butenal (HPB242) induces apoptosis via modulating E7expression and inhibition of PI3KAkt pathway in SiHa humancervical cancer cellsrdquo Nutrition and Cancer vol 64 no 8 pp1236ndash1244 2012
[31] F Cui X L Li X Y Zhu et al ldquoMiR-125b inhibits tumorgrowth and promotes apoptosis of cervical cancer cells bytargeting phosphoinositide 3-kinase catalytic subunit deltardquoCellular Physiology and Biochemistry vol 30 no 5 pp 1310ndash1318 2012
[32] S Kang S M Dong B-R Kim et al ldquoThioridazine inducesapoptosis by targeting the PI3KAktmTOR pathway in cervicaland endometrial cancer cellsrdquo Apoptosis vol 17 no 9 pp 989ndash997 2012
[33] O Thanaketpaisarn P Waiwut H Sakurai and I Saiki ldquoArte-sunate enhances TRAIL-induced apoptosis in human cervicalcarcinoma cells through inhibition of the NF-120581B and PI3KAktsignaling pathwaysrdquo International Journal of Oncology vol 39no 1 pp 279ndash285 2011
[34] XWang J-H Ren F Lin et al ldquoStathmin is involved in arsenictrioxide-induced apoptosis in human cervical cancer cell linesvia PI3K linked signal pathwayrdquo Cancer Biology and Therapyvol 10 no 6 pp 632ndash643 2010
[35] C W Lee N H Kim H K Choi et al ldquoLysophosphatidicacid-induced c-fos up-regulation involves cyclic AMP responseelement-binding protein activated by mitogen- and stress-activated protein kinase-1rdquo Journal of Cellular Biochemistry vol104 no 3 pp 785ndash794 2008
[36] DW Hommes M P Peppelenbosch and S J H van DeventerldquoMitogen activated protein (MAP) kinase signal transduction
pathways and novel anti-inflammatory targetsrdquoGut vol 52 no1 pp 144ndash151 2003
[37] M Mukai F Imamura M Ayaki et al ldquoInhibition of tumorinvasion andmetastasis by a novel lysophosphatidic acid (cyclicLPA)rdquo International Journal of Cancer vol 81 no 6 pp 918ndash9221999
[38] E-Y Huang Y-F Chen Y-M Chen et al ldquoA novel radiore-sistant mechanism of galectin-1 mediated by H-Ras-dependentpathways in cervical cancer cellsrdquo Cell Death and Disease vol3 no 1 article e251 2012
[39] W-H Su P-C Chuang E-Y Huang and K D YangldquoRadiation-induced increase in cell migration and metastaticpotential of cervical cancer cells operates via theK-ras pathwayrdquoThe American Journal of Pathology vol 180 no 2 pp 862ndash8712012
[40] H Rincon-Arano R Rosales N Mora A Rodriguez-Castaneda and C Rosales ldquoR-Ras promotes tumor growthof cervical epithelial cellsrdquo Cancer vol 97 no 3 pp 575ndash5852003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 11
cervical cancer patients especially those with high levels ofLPA
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Yanxia Sui and Ya Yang contributed equally to this work
Acknowledgments
This study is supported by the following funding sourcesThe National Natural Science Foundations of China (JuanRen 31201060C0709 Juan Ren 30973175H1621 and JuanRen 81172490H1621) Program for New Century ExcellentTalents in University (Juan Ren NCET-12-0440) Scientificand Technological Research Foundation of Shaanxi Province(Juan Ren 2012K13-01-06 Juan Ren 2007K09-09) Scien-tific Research Foundation for the Returned overseas Chi-nese Scholars of State Education Ministry (Juan Ren 0601-18920006) Research Foundation of Health Department ofShaanrsquoxi Province (Juan Ren 2010D41) Qing Nian Jiao ShiGenZongJiHua ofXirsquoan JiaotongUniversity (JuanRen 2012)Program for Changjiang Scholars and Innovative ResearchTeam inUniversity (Zongfang Li PCSIRT1171) andResearchFoundation of Xirsquoan Jiao Tong University of China (JuanRen)
References
[1] J R Erickson Y Hasegawa X Fang et al ldquoLysophosphatidicacid and ovarian cancer a paradigm for tumorogenesis andpatient managementrdquo Prostaglandins and Other Lipid Media-tors vol 64 no 1ndash4 pp 63ndash81 2001
[2] Y Xu Z Z Shen DWWiper et al ldquoLysophosphatidic acid as apotential biomarker for ovarian and other gynecologic cancersrdquoThe Journal of the American Medical Association vol 280 no 8pp 719ndash723 1998
[3] J S Karliner ldquoLysophospholipids and the cardiovascular sys-temrdquoBiochimica et Biophysica ActamdashMolecular and Cell Biologyof Lipids vol 1582 no 1ndash3 pp 216ndash221 2002
[4] C Schulze C Smales L L Rubin and J M StaddonldquoLysophosphatidic acid increases tight junction permeability incultured brain endothelial cellsrdquo Journal of Neurochemistry vol68 no 3 pp 991ndash1000 1997
[5] I Ishii N Fukushima X Ye and J Chun ldquoLysophospholipidreceptors signaling and biologyrdquo Annual Review of Biochem-istry vol 73 pp 321ndash354 2004
[6] T S Panetti ldquoDifferential effects of sphingosine 1-phosphateand lysophosphatidic acid on endothelial cellsrdquo Biochimica etBiophysica ActamdashMolecular and Cell Biology of Lipids vol 1582no 1ndash3 pp 190ndash196 2002
[7] Z Z Shen M Z Wu P Elson et al ldquoFatty acid composition oflysophosphatidic acid and lysophosphatidylinositol in plasmafrom patients with ovarian cancer and other gynecologicaldiseasesrdquo Gynecologic Oncology vol 83 no 1 pp 25ndash30 2001
[8] M M Murph ldquoAgonist-induced endocytosis of lysophospha-tidic acid-coupled LPA1EDG-2 receptors via a dynamin2- andRab5-dependent pathwayrdquo Journal of Cell Science vol 116 no10 pp 1969ndash1980 2003
[9] L M Baudhuin K L Cristina J Lu and Y Xu ldquoAkt activationinduced by lysophosphatidic acid and sphingosine-1-phosphaterequires both mitogen-activated protein kinase kinase andp38 mitogen-activated protein kinase and is cell-line specificrdquoMolecular Pharmacology vol 62 no 3 pp 660ndash671 2002
[10] R-J Chen S-U Chen C-H Chou andM-C Lin ldquoLysophos-phatidic acid receptor 23-mediated IL-8-dependent angiogen-esis in cervical cancer cellsrdquo International Journal of Cancer vol131 no 4 pp 789ndash802 2012
[11] J W Choi S Lim Y-S Oh et al ldquoSubtype-specific role ofphospholipase C-120573 in bradykinin and LPA signaling throughdifferential binding of different PDZ scaffold proteinsrdquo CellularSignalling vol 22 no 7 pp 1153ndash1161 2010
[12] Y Kishi S Okudaira M Tanaka et al ldquoAutotaxin is over-expressed in glioblastoma multiforme and contributes to cellmotility of glioblastoma by converting lysophosphatidylcholineTO lysophosphatidic acidrdquoThe Journal of Biological Chemistryvol 281 no 25 pp 17492ndash17500 2006
[13] A Eastman ldquoThe formation isolation and characterization ofDNA adducts produced by anticancer platinum complexesrdquoPharmacology andTherapeutics vol 34 no 2 pp 155ndash166 1987
[14] K Wozniak and J Błasiak ldquoRecognition and repair of DNA-cisplatin adductsrdquo Acta Biochimica Polonica vol 49 no 3 pp583ndash596 2002
[15] A Basu and S Krishnamurthy ldquoCellular responses to cisplatin-induced DNA damagerdquo Journal of Nucleic Acids vol 2010Article ID 201367 16 pages 2010
[16] A Efeyan and M Serrano ldquop53 guardian of the genome andpoliceman of the oncogenesrdquo Cell Cycle vol 6 no 9 pp 1006ndash1010 2007
[17] O Kutuk E D Arisan T Tezil M C Shoshan and H BasagaldquoCisplatin overcomesBcl-2-mediated resistance to apoptosis viapreferential engagement of Bak critical role of Noxa-mediatedlipid peroxidationrdquo Carcinogenesis vol 30 no 9 pp 1517ndash15272009
[18] C M Yan E Q Chai H Y Cai G Miao and W MaldquoOleuropein induces apoptosis via activation of caspases andsuppression of phosphatidylinositol 3-kinaseprotein kinaseB pathway in HepG2 human hepatoma cell linerdquo MolecularMedicine Reports vol 11 pp 4617ndash4624 2015
[19] X Y Wang X S Fan L Cai S Liu X F Cong and X ChenldquoLysophosphatidic acid rescues bone mesenchymal stem cellsfrom hydrogen peroxide-induced apoptosisrdquo Apoptosis vol 20no 3 pp 273ndash284 2015
[20] A Brozovic G Fritz M Christmann et al ldquoLong-term activa-tion of SAPKJNK p38 kinase and fas-L expression by cisplatinis attenuated in human carcinoma cells that acquired drugresistancerdquo International Journal of Cancer vol 112 no 6 pp974ndash985 2004
[21] Y Q Liu H Xing X B Han et al ldquoApoptosis of HeLa cellsinduced by cisplatin and its mechanismrdquo Journal of HuazhongUniversity of Science and TechnologymdashMedical Science vol 28no 2 pp 197ndash199 2008
[22] K L Jin J-Y Park E J Noh et al ldquoThe effect of combinedtreatment with cisplatin and histone deacetylase inhibitors onHeLa cellsrdquo Journal of Gynecologic Oncology vol 21 no 4 pp262ndash268 2010
12 BioMed Research International
[23] T Sakaeda T Nakamura M Hirai et al ldquoMDR1 up-regulatedby apoptotic stimuli suppresses apoptotic signalingrdquo Pharma-ceutical Research vol 19 no 9 pp 1323ndash1329 2002
[24] K Kandasamy S M Srinivasula E S Alnemri et al ldquoInvolve-ment of proapoptotic molecules Bax and Bak in tumor necro-sis factor-related apoptosis-inducing ligand (TRAIL)-inducedmitochondrial disruption and apoptosis differential regulationof cytochrome c and SmacDIABLO releaserdquo Cancer Researchvol 63 no 7 pp 1712ndash1721 2003
[25] H Toyota N Yanase T Yoshimoto M Moriyama T Sudoand J Mizuguchi ldquoCalpain-induced Bax-cleavage product is amore potent inducer of apoptotic cell death thanwild-type BaxrdquoCancer Letters vol 189 no 2 pp 221ndash230 2003
[26] Y R Meng S J Kang and D A Fishman ldquoLysophosphatidicacid inhibits anti-Fas-mediated apoptosis enhanced by actindepolymerization in epithelial ovarian cancerrdquo FEBS Lettersvol 579 no 5 pp 1311ndash1319 2005
[27] Y Meng S Kang J So S Reierstad and D A FishmanldquoTranslocation of Fas by LPA prevents ovarian cancer cells fromanti-Fas-induced apoptosisrdquo Gynecologic Oncology vol 96 no2 pp 462ndash469 2005
[28] X Wan Y Yokoyama A Shinohara Y Takahashi and TTamaya ldquoPTEN augments staurosporine-induced apoptosis inPTEN-null Ishikawa cells by downregulating PI3KAkt signal-ing pathwayrdquo Cell Death and Differentiation vol 9 no 4 pp414ndash420 2002
[29] J Yu Y Zhang J Mcilroy T Rordorf-Nikolic G A Orr and JM Backer ldquoRegulation of the p85p110 phosphatidylinositol 31015840-kinase stabilization and inhibition of the p110120572 catalytic subunitby the p85 regulatory subunitrdquoMolecular and Cellular Biologyvol 18 no 3 pp 1379ndash1387 1998
[30] M S Kim J H Kim Y Bak et al ldquo24-bis (p-hydroxyphenyl)-2-butenal (HPB242) induces apoptosis via modulating E7expression and inhibition of PI3KAkt pathway in SiHa humancervical cancer cellsrdquo Nutrition and Cancer vol 64 no 8 pp1236ndash1244 2012
[31] F Cui X L Li X Y Zhu et al ldquoMiR-125b inhibits tumorgrowth and promotes apoptosis of cervical cancer cells bytargeting phosphoinositide 3-kinase catalytic subunit deltardquoCellular Physiology and Biochemistry vol 30 no 5 pp 1310ndash1318 2012
[32] S Kang S M Dong B-R Kim et al ldquoThioridazine inducesapoptosis by targeting the PI3KAktmTOR pathway in cervicaland endometrial cancer cellsrdquo Apoptosis vol 17 no 9 pp 989ndash997 2012
[33] O Thanaketpaisarn P Waiwut H Sakurai and I Saiki ldquoArte-sunate enhances TRAIL-induced apoptosis in human cervicalcarcinoma cells through inhibition of the NF-120581B and PI3KAktsignaling pathwaysrdquo International Journal of Oncology vol 39no 1 pp 279ndash285 2011
[34] XWang J-H Ren F Lin et al ldquoStathmin is involved in arsenictrioxide-induced apoptosis in human cervical cancer cell linesvia PI3K linked signal pathwayrdquo Cancer Biology and Therapyvol 10 no 6 pp 632ndash643 2010
[35] C W Lee N H Kim H K Choi et al ldquoLysophosphatidicacid-induced c-fos up-regulation involves cyclic AMP responseelement-binding protein activated by mitogen- and stress-activated protein kinase-1rdquo Journal of Cellular Biochemistry vol104 no 3 pp 785ndash794 2008
[36] DW Hommes M P Peppelenbosch and S J H van DeventerldquoMitogen activated protein (MAP) kinase signal transduction
pathways and novel anti-inflammatory targetsrdquoGut vol 52 no1 pp 144ndash151 2003
[37] M Mukai F Imamura M Ayaki et al ldquoInhibition of tumorinvasion andmetastasis by a novel lysophosphatidic acid (cyclicLPA)rdquo International Journal of Cancer vol 81 no 6 pp 918ndash9221999
[38] E-Y Huang Y-F Chen Y-M Chen et al ldquoA novel radiore-sistant mechanism of galectin-1 mediated by H-Ras-dependentpathways in cervical cancer cellsrdquo Cell Death and Disease vol3 no 1 article e251 2012
[39] W-H Su P-C Chuang E-Y Huang and K D YangldquoRadiation-induced increase in cell migration and metastaticpotential of cervical cancer cells operates via theK-ras pathwayrdquoThe American Journal of Pathology vol 180 no 2 pp 862ndash8712012
[40] H Rincon-Arano R Rosales N Mora A Rodriguez-Castaneda and C Rosales ldquoR-Ras promotes tumor growthof cervical epithelial cellsrdquo Cancer vol 97 no 3 pp 575ndash5852003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

12 BioMed Research International
[23] T Sakaeda T Nakamura M Hirai et al ldquoMDR1 up-regulatedby apoptotic stimuli suppresses apoptotic signalingrdquo Pharma-ceutical Research vol 19 no 9 pp 1323ndash1329 2002
[24] K Kandasamy S M Srinivasula E S Alnemri et al ldquoInvolve-ment of proapoptotic molecules Bax and Bak in tumor necro-sis factor-related apoptosis-inducing ligand (TRAIL)-inducedmitochondrial disruption and apoptosis differential regulationof cytochrome c and SmacDIABLO releaserdquo Cancer Researchvol 63 no 7 pp 1712ndash1721 2003
[25] H Toyota N Yanase T Yoshimoto M Moriyama T Sudoand J Mizuguchi ldquoCalpain-induced Bax-cleavage product is amore potent inducer of apoptotic cell death thanwild-type BaxrdquoCancer Letters vol 189 no 2 pp 221ndash230 2003
[26] Y R Meng S J Kang and D A Fishman ldquoLysophosphatidicacid inhibits anti-Fas-mediated apoptosis enhanced by actindepolymerization in epithelial ovarian cancerrdquo FEBS Lettersvol 579 no 5 pp 1311ndash1319 2005
[27] Y Meng S Kang J So S Reierstad and D A FishmanldquoTranslocation of Fas by LPA prevents ovarian cancer cells fromanti-Fas-induced apoptosisrdquo Gynecologic Oncology vol 96 no2 pp 462ndash469 2005
[28] X Wan Y Yokoyama A Shinohara Y Takahashi and TTamaya ldquoPTEN augments staurosporine-induced apoptosis inPTEN-null Ishikawa cells by downregulating PI3KAkt signal-ing pathwayrdquo Cell Death and Differentiation vol 9 no 4 pp414ndash420 2002
[29] J Yu Y Zhang J Mcilroy T Rordorf-Nikolic G A Orr and JM Backer ldquoRegulation of the p85p110 phosphatidylinositol 31015840-kinase stabilization and inhibition of the p110120572 catalytic subunitby the p85 regulatory subunitrdquoMolecular and Cellular Biologyvol 18 no 3 pp 1379ndash1387 1998
[30] M S Kim J H Kim Y Bak et al ldquo24-bis (p-hydroxyphenyl)-2-butenal (HPB242) induces apoptosis via modulating E7expression and inhibition of PI3KAkt pathway in SiHa humancervical cancer cellsrdquo Nutrition and Cancer vol 64 no 8 pp1236ndash1244 2012
[31] F Cui X L Li X Y Zhu et al ldquoMiR-125b inhibits tumorgrowth and promotes apoptosis of cervical cancer cells bytargeting phosphoinositide 3-kinase catalytic subunit deltardquoCellular Physiology and Biochemistry vol 30 no 5 pp 1310ndash1318 2012
[32] S Kang S M Dong B-R Kim et al ldquoThioridazine inducesapoptosis by targeting the PI3KAktmTOR pathway in cervicaland endometrial cancer cellsrdquo Apoptosis vol 17 no 9 pp 989ndash997 2012
[33] O Thanaketpaisarn P Waiwut H Sakurai and I Saiki ldquoArte-sunate enhances TRAIL-induced apoptosis in human cervicalcarcinoma cells through inhibition of the NF-120581B and PI3KAktsignaling pathwaysrdquo International Journal of Oncology vol 39no 1 pp 279ndash285 2011
[34] XWang J-H Ren F Lin et al ldquoStathmin is involved in arsenictrioxide-induced apoptosis in human cervical cancer cell linesvia PI3K linked signal pathwayrdquo Cancer Biology and Therapyvol 10 no 6 pp 632ndash643 2010
[35] C W Lee N H Kim H K Choi et al ldquoLysophosphatidicacid-induced c-fos up-regulation involves cyclic AMP responseelement-binding protein activated by mitogen- and stress-activated protein kinase-1rdquo Journal of Cellular Biochemistry vol104 no 3 pp 785ndash794 2008
[36] DW Hommes M P Peppelenbosch and S J H van DeventerldquoMitogen activated protein (MAP) kinase signal transduction
pathways and novel anti-inflammatory targetsrdquoGut vol 52 no1 pp 144ndash151 2003
[37] M Mukai F Imamura M Ayaki et al ldquoInhibition of tumorinvasion andmetastasis by a novel lysophosphatidic acid (cyclicLPA)rdquo International Journal of Cancer vol 81 no 6 pp 918ndash9221999
[38] E-Y Huang Y-F Chen Y-M Chen et al ldquoA novel radiore-sistant mechanism of galectin-1 mediated by H-Ras-dependentpathways in cervical cancer cellsrdquo Cell Death and Disease vol3 no 1 article e251 2012
[39] W-H Su P-C Chuang E-Y Huang and K D YangldquoRadiation-induced increase in cell migration and metastaticpotential of cervical cancer cells operates via theK-ras pathwayrdquoThe American Journal of Pathology vol 180 no 2 pp 862ndash8712012
[40] H Rincon-Arano R Rosales N Mora A Rodriguez-Castaneda and C Rosales ldquoR-Ras promotes tumor growthof cervical epithelial cellsrdquo Cancer vol 97 no 3 pp 575ndash5852003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom






![Prohibitin (PHB) inhibits apoptosis in rat granulosa cells ...apoptosis of granulosa cells (GCs) during follicular growth and development [1, 2]. Ovarian GCs play an important physiological](https://static.fdocuments.in/doc/165x107/5f84f4a5739a256f3f64c746/prohibitin-phb-inhibits-apoptosis-in-rat-granulosa-cells-apoptosis-of-granulosa.jpg)











