
Research Article Antiosteoporotic Effects of Huangqi...
11
Research Article Antiosteoporotic Effects of Huangqi Sanxian Decoction in Cultured Rat Osteoblasts by Proteomic Characterization of the Target and Mechanism Chong-Chong Guo, 1 Li-Hua Zheng, 1 Jian-Ying Fu, 1 Jian-Hong Zhu, 2 Yan-Xing Zhou, 1 Tao Zeng, 3 and Zhi-Kun Zhou 1 1 Department of Pharmacy, Guangdong Medical College, No. 1, Xincheng Dadao, Songshan Lake Science and Technology Industry Park, Dongguan 523808, China 2 Department of Pharmacy, Sun Yat-Sen Memorial Hospital, Guangzhou 510120, China 3 Laboratory Medicine Center, Nanfang Hospital, Southern Medical University, Guangzhou, Guangdong 510515, China Correspondence should be addressed to Zhi-Kun Zhou; [email protected] Received 21 March 2015; Revised 5 June 2015; Accepted 24 June 2015 Academic Editor: Klaus Heese Copyright © 2015 Chong-Chong Guo et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Huangqi Sanxian decoction (HQSXD) is routinely used for the treatment of osteoporosis in the Chinese traditional healthcare system. However, the targets and mechanism underlying the effect of HQSXD on osteoporosis have not been documented. In the present study, seropharmacology and proteomic approaches (two-dimensional gel electrophoresis combined with mass spectrometry) were used to investigate the effects and possible target proteins of HQSXD on osteoblast. We found that HQSXD-treated rat serum significantly enhanced osteoblast proliferation, differentiation, and mineralization. In HQSXD-S- treated osteoblasts, there were increases in the expression of N-formyl peptide receptor 2 and heparan sulfate (glucosamine) 3- O-sulfotransferase 3A1 and reduction in the expression of alpha-spectrin, prohibitin, and transcription elongation factor B (SIII), polypeptide 1. e identified proteins are associated with cell proliferation, differentiation, signal transcription, and cell growth. ese findings might provide valuable insights into the mechanism of antiosteoporotic effect affected by HQSXD treatment in osteoblasts. 1. Introduction Osteoporosis is the most frequent bone remodeling dis- ease and its incidence increases with advancing age. is disease is characterized by a reduction in bone mass and microarchitectural deterioration of bone tissue, resulting in high risk of fractures [1]. Globally, osteoporosis and associated bone fractures have become a major health hazard afflicting millions of people [2]. Current treatment options for osteoporosis include bisphosphonates, estrogens, selective estrogen receptor modulators, calcitonin, denosumab, and teriparatide. However, many of these drugs generate side effects [3, 4] and their costs are too high to benefit a large population in the developing and developed countries, which may limit their applications. Traditional Chinese medicine has been a part of health- care in China for thousands of years and has recently been reevaluated for clinical approach [5]. e traditional Chinese medicine has fewer adverse reactions and is more suitable for long-term administration than synthetic drugs and antibiotics. Popular commercially available prescriptions include Jinkui Shenqi Wan (JKSQW), which exerts a thera- peutic effect on the kidney-Yang deficiency and osteoporosis indicated in Chinese Pharmacopoeia (2010), and Huangqi Sanxian decoction (HQSXD), a traditional Chinese formula, which is composed of Radix Astragali, Epimedii Folium, Cis- tanche Herba, Radix notoginseng, Radix Salviae Miltiorrhizae, Corydalis Rhizoma, Radix Angelicae Sinensis, and Radix Clematidis. Our previous study revealed that Huangqi Sanx- ian decoction treatment significantly increased sex estrogen Hindawi Publishing Corporation Evidence-Based Complementary and Alternative Medicine Volume 2015, Article ID 514063, 10 pages http://dx.doi.org/10.1155/2015/514063
Transcript of Research Article Antiosteoporotic Effects of Huangqi...

Research ArticleAntiosteoporotic Effects of Huangqi SanxianDecoction in Cultured Rat Osteoblasts by ProteomicCharacterization of the Target and Mechanism
Chong-Chong Guo1 Li-Hua Zheng1 Jian-Ying Fu1 Jian-Hong Zhu2
Yan-Xing Zhou1 Tao Zeng3 and Zhi-Kun Zhou1
1Department of Pharmacy Guangdong Medical College No 1 Xincheng Dadao Songshan Lake Science andTechnology Industry Park Dongguan 523808 China2Department of Pharmacy Sun Yat-Sen Memorial Hospital Guangzhou 510120 China3Laboratory Medicine Center Nanfang Hospital Southern Medical University Guangzhou Guangdong 510515 China
Correspondence should be addressed to Zhi-Kun Zhou zhikunzhou126com
Received 21 March 2015 Revised 5 June 2015 Accepted 24 June 2015
Academic Editor Klaus Heese
Copyright copy 2015 Chong-Chong Guo et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Huangqi Sanxian decoction (HQSXD) is routinely used for the treatment of osteoporosis in the Chinese traditional healthcaresystem However the targets and mechanism underlying the effect of HQSXD on osteoporosis have not been documentedIn the present study seropharmacology and proteomic approaches (two-dimensional gel electrophoresis combined with massspectrometry) were used to investigate the effects and possible target proteins of HQSXD on osteoblast We found thatHQSXD-treated rat serum significantly enhanced osteoblast proliferation differentiation and mineralization In HQSXD-S-treated osteoblasts there were increases in the expression of N-formyl peptide receptor 2 and heparan sulfate (glucosamine) 3-O-sulfotransferase 3A1 and reduction in the expression of alpha-spectrin prohibitin and transcription elongation factor B (SIII)polypeptide 1 The identified proteins are associated with cell proliferation differentiation signal transcription and cell growthThese findings might provide valuable insights into the mechanism of antiosteoporotic effect affected by HQSXD treatment inosteoblasts
1 Introduction
Osteoporosis is the most frequent bone remodeling dis-ease and its incidence increases with advancing age Thisdisease is characterized by a reduction in bone mass andmicroarchitectural deterioration of bone tissue resultingin high risk of fractures [1] Globally osteoporosis andassociated bone fractures have become a major health hazardafflicting millions of people [2] Current treatment optionsfor osteoporosis include bisphosphonates estrogens selectiveestrogen receptor modulators calcitonin denosumab andteriparatide However many of these drugs generate sideeffects [3 4] and their costs are too high to benefit a largepopulation in the developing and developed countries whichmay limit their applications
Traditional Chinese medicine has been a part of health-care in China for thousands of years and has recentlybeen reevaluated for clinical approach [5] The traditionalChinese medicine has fewer adverse reactions and is moresuitable for long-term administration than synthetic drugsand antibiotics Popular commercially available prescriptionsinclude Jinkui Shenqi Wan (JKSQW) which exerts a thera-peutic effect on the kidney-Yang deficiency and osteoporosisindicated in Chinese Pharmacopoeia (2010) and HuangqiSanxian decoction (HQSXD) a traditional Chinese formulawhich is composed of Radix Astragali Epimedii Folium Cis-tanche Herba Radix notoginseng Radix SalviaeMiltiorrhizaeCorydalis Rhizoma Radix Angelicae Sinensis and RadixClematidis Our previous study revealed that Huangqi Sanx-ian decoction treatment significantly increased sex estrogen
Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2015 Article ID 514063 10 pageshttpdxdoiorg1011552015514063
2 Evidence-Based Complementary and Alternative Medicine
level and bone mineral density (BMD) and repressed boneabsorption function in postmenopausal women [6]This sug-gested that HQSXD has beneficial effects in the treatment ofosteoporosis However little is known about the mechanismsand targets underlying the effects ofHQSXDon osteoporosis
The Chinese traditional medicine theory believes thatbone activities are controlled by the kidney Strong ldquokidneyrdquocan nourish bones but the weak ldquokidneyrdquo might hasten bonedeterioration [7] Kidney deficiency and blood stasis are themain pathological basis of osteoporosis Huangqi Sanxiandecoction is composed of eight Chinese medicinal herbsof these Epimedii Folium and Cistanche Herba strengthenkidneys while Radix Salviae Miltiorrhizae and Radix noto-ginseng invigorate the circulation of blood These herbs arean excellent combination for highlighting their superiority inthe treatment of osteoporosis [8]
Jinkui Shenqi Wan an ancient Chinese herbal formulais indicated in the Chinese Pharmacopoeia (2010) for thetreatment of Yang insufficiency of kidney weakness andsoreness of the loins and the knees cold feeling in the limbsand frequent urination Human clinical studies have certifiedthat Jinkui Shenqi Wan exerts a therapeutic effect on thekidney-Yang deficiency [9] Hence we used Jinkui ShenqiWan as a positive control in this study
In the present study seropharmacology and functionalproteomics technology were used to explore the multipleproteins associated with the antiosteoporotic effect Theresults suggest a basis for the clinical use of HQSXD in thetreatment of patients with osteoporosis
2 Materials and Methods
21 Animals Thirty Sprague-Dawley rats (fifteen male andfifteen female) weighing about 250ndash300 g were purchasedfrom the Animal Experimental Center of Guangdong Med-ical College (Dongguan China) The animals were housedindividually in a regulated environment (24 plusmn 05∘C) witha 12-hour lightdark cycle (under light 0800ndash2000 h) Foodand water were given ad libitum throughout the experimentAfter three days of acclimation male and female SD rats wererandomly divided into three groups blank-control groupexperimental group treated withHuangqi Sanxian decoctionand positive control group treated with Jinkui Shenqi WanThe Committee of Experimental Animal Administration ofthe University approved the study and the procedures ofthe experiment were in accordance with generally acceptedinternational rules and regulations
22 Preparation of HQSXD and JKSQW HQSXD includedeight plant extracts including Radix Astragali (root Chineseherbal name Huang-Qi) Epimedii Folium (leaf Chineseherbal name Yin-Yang-Huo) Cistanche Herba (succulentstem Chinese herbal name Rou-Cong-Rong) Radix noto-ginseng (root and rhizome Chinese herbal name San-Qi) Radix Salviae Miltiorrhizae (root and rhizome Chineseherbal name Dan-Shen) Corydalis Rhizoma (rhizome Chi-nese herbal name Yan-Hu-Suo) Radix Angelicae Sinensis
(root Chinese herbal name Dang-Gui) and Radix Clema-tidis (root and rhizome Chinese herbal name Wei-Ling-Xian) in a ratio of 15 10 10 5 1010 8 10 [6] The aboveeight medicinal extracts were obtained from DongguanSinopharm (Dongguan China) and identified by ProfessorZhou (Department of Pharmacy Guangdong Medical Col-lege Dongguan China) The eight medicinal plant materialsin the mixture (270 g) were powdered and coboiled with1000mLwater for 2 hoursThe extraction was repeated twiceThe filtrates were concentrated to 200mL under reducedpressure and kept at 4∘C
JKSQW consisted of Radix Rehmanniae Preparata (rootChinese herbal name Di-Huang) Dioscoreae Rhizome (rootChinese herbal name Shan-Yao) Fructus Corni Officinalis(fructus Chinese herbal name Shan-Zhu-Yu) SclerotiumPoriae Cocos (dried sclerotia Chinese herbal name Fu-Ling)CortexMoutan Radicis (root bark Chinese herbal nameMu-Dan-Pi) Rhizoma Alismatis Orientalis (stem Chinese herbalname Ze-Xie) Ramulus Cinnamomi Cassiae (twig Chineseherbal name Gui-Zhi) Radix Aconiti Lateralis Praeparata(root Chinese herbal name Fu-Zi)Radix Achyranthis Biden-tatae (root Chinese herbal name Niu-Xi) and PlantaginisSemen (seed Chinese herbal name Che-Qian-Zi) [9] Asper the instructions 5mg Jinkui Shenqi Wan (Beijing TongRen Tang Pharmaceutical Technology Development IncDongguan China) was dissolved in 100mL distilled waterbefore administration The experimental dose for HQSXDand JKSQW in the present study was equivalent to thecorresponding clinical prescription dose for a human subjectweighing 60 kg
23 Drug Administration and Sample Collections Rats wererandomly divided into three groups of 10 animals each(each group included five males and five females) Chinesemedicine HQSXD solution (14 gkg) and medicine JKSQWsolution (0520 gkg) were administered orally every day forthree days twice a day The blank-control group was orallyadministered distilled water and they were monitored con-currently with the HQSXD-experimental groups At the endof the experiment sixty minutes after the last treatment theanimalswere exposed to ether anesthesia blood sampleswerecollected by heart puncture under aseptic conditions andthen centrifuged for 15min to obtain serum samples SerumofHQSXD-treated rats and control serumwere inactivated at56∘C in awater bath for 30min and filtered through a 022120583mfilter membrane termed HQSXD-S JKSQW-control-S andBLANK-control-S respectively and then stored at minus80∘C
24 Primary Osteoblasts Culture and Assay for OsteoblastProliferation Primary rat osteoblast cells were obtained from1-day-old neonatal Sprague-Dawley rats as described previ-ously [10] Primary osteoblasts were cultured by seeding 96-well plates with a density of 1 times 104 per well and incubatedfor 24 h After adhesion of cells Dulbeccorsquos Modified EagleMedium (DMEM) was added and incubated for another 24hours Next after discarding the medium test (HQSXD-S)and control (JKSQW-control-S BLANK-control-S) samples
Evidence-Based Complementary and Alternative Medicine 3
were added at a concentration of 10 (vv) and the cellswere incubated at 37∘C in a humid atmosphere containing 5CO2for 72 h Thereafter 5mgmL of 3-(45-dimethylthiazol-
2-yl)-25-diphenyltetrazolium bromide (MTT) was addedand incubated for 4 hours after which the medium wasdiscarded and dimethyl sulfoxide (DMSO) (150120583L) wasadded Absorbance was measured at 490 nm using a Syn-ergy 2 multifunctional microplate reader (Bio-Tek) to assessosteoblast proliferation
25 ALP Activity and Staining Assay Osteoblasts were sus-pended in DMEM to obtain a cell density of 5 times 105mL2mL aliquots of the cell suspension were added to 6-wellplates After 24 h of incubation the medium was changedand cells were incubated with test (HQSXD-S) and control(JKSQW-control-S BLANK-control-S) at a concentration of10 (vv) for 3 days Alkaline phosphatase (ALP) activitywas measured using an ALP assay kit (Sigma) as describedpreviously [11] For ALP staining after incubation withHQSXD-S JKSQW-control-S and BLANK-control-S (con-centration of 10 (vv)) for three days the cells were fixedin 70 ethanol for 15min washed and then incubatedwith ALP staining buffer nitro blue tetrazolium5-bromo-4-chloro-3-indolyl phosphate (NBTBCIP) (Beyotime Instituteof Biotechnology China) at 37∘C for 30min air dried andphotographed
26 Mineralization Assay After 21 days of differentiationthe mineralization of osteoblasts was analyzed as describedpreviously [12] Briefly the cells were washed with phosphatebuffered saline (PBS) and fixed with 70 ethanol for onehourThe cellswere then rinsed in distilledwater stainedwith05 Alizarin Red S (ARS) at pH 42 with rotation for 30minat 37∘C and subsequently washed with distilled water anddried in air Stained cultures were photographed To analyzeARS activity the ARS in stained cells was destained with10 cetylpyridinium chloride (CPC) monohydrate solution(Sigma) for 30min with shaking The absorbance was mea-sured at 562 nmusing a Synergy 2multifunctionalmicroplatereader
27 Protein Extraction Osteoblasts cells were incubated in50mL culture flasks and grown to subconfluence (approx-imately 60ndash70) and then treated with test (HQSXD-S) and control (JKSQW-control-S BLANK-control-S) at aconcentration of 10 (vv) for 72 h respectively At the end ofthe incubation period osteoblasts were collected by centrifu-gation at 1000 rpm (4∘C) The osteoblasts were washed twicewith cold PBS after discarding the medium Total proteinswere extracted in a chilled lysis buffer containing 7M urea2M thiourea 4 (wv)CHAPS 40mMdithiothreitol (DTT)2 (vv) IPG buffer pH 3ndash10 4 120583gmL protease inhibitormixture and 4 120583gmL phosphatase inhibitors After additionof the chilled lysis buffer the cell solution was kept oscillatingfor 1 h at 4∘C to solubilize the proteins The homogenatewas subsequently centrifuged for 15min at 14000 rpm at
08
06
04
02
00
OD
(490
nm)
lowastlowast lowastlowast
JKSQW-treatedgroup
Blank-control HQSXD-treatedgroupgroup
Figure 1 Effects of drugs on proliferation of osteoblasts Althoughthere was some improvement in the proliferation of osteoblastsin the HQSXD-treated group it was not statistically significantwhen compared to the values observed in the group given JKSQW-control-S Data are expressed as the mean plusmn SD
4∘C Total protein concentration was quantified using theBradford assay (Amresco)
28 Two-Dimensional Electrophoresis (2DE) and Image Anal-ysis Electrophoresis was carried out at least thrice forreproducibility Two-dimensional electrophoresis (2DE) wascarried out as described previously [13] with some mod-ifications For the first-dimension isoelectric focusing gel250 120583g of protein was used for isoelectric focusing (IEF)using 17 cm nonlinear (NL) IPG strips (pH 3ndash10) whichwere immersed in rehydration buffer (8M urea 2 CHAPS25mM DTT 02 (wv) Bio-Lyte 01 bromophenol blueand 1 IPG buffer) The strips were covered with mineraloil (GE Healthcare) to prevent samples from evaporationand rehydrated for 10ndash20 hours Isoelectric focusing gelelectrophoresis was performed by using an electrophoresisapparatus (GE Healthcare) The running conditions of theIEF process were as follows 100V 1 h 200V 1 h 300V 1 h500V 1 h 1000V 1 h 2000V 1 h and 8000V up 60000VAfter the IEF the IPG strips were equilibrated with equilibra-tion buffer-1 (6M urea 2 SDS 20 glycerol 0375M Tris-HCl [pH 88] bromophenol blue dye and 1 [wv] DTT) for15min and then were repeated for an additional 15 minutes in5mL of equilibration buffer-2 except that DTT was replacedby 25 [wv] iodoacetamide The second dimension wasdone with a 125 acrylamide gel at 10mAgel for onehour and then at 38mAgel until the bromophenol blue dyereached the bottom of the gel After electrophoresis the gelswere visualized by silver nitrate staining Gels were scannedusing the ImageScanner (GE Healthcare) The images wereanalyzed with ImageMaster 2D Platinum v70 software (GEHealthcare San Francisco CA) including spot detectionbackground subtraction gel matching and normalizationSpots were detected and matched automatically to a mastergel and then edited manually Matched spots from triplicategel sets that showed overlap ratio with an absolute valuege2 were recognized as differentially expressed Differentially
4 Evidence-Based Complementary and Alternative Medicine
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
ALP staining
(a)
025
020
015
010
005
000
ALP
(Ug
prot
)
lowastlowast
lowastlowast
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
(b)
Figure 2 HQSXD-S enhances primary osteoblast differentiation (a) Primary osteoblasts were treated with various drug-treated rat serumsfor 3 days ALP-positive cells were stained with ALP solution (b) Effect of HQSXD-S on alkaline phosphatase activity in primary cultureosteoblasts Data are means plusmn SD of six replicates lowastlowast119901 lt 001 versus control
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
ARS staining
(a)
025
020
015
010
005
000
ARS
activ
ity (562
nm)
lowastlowastlowastlowastlowast
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
(b)
Figure 3 HQSXD-S promotes bone mineralization during osteoblastogenesis (a) Calcium deposits stained with Alizarin Red solution seenin primary osteoblasts treated with various drug-treated rat serums for 21 days (b) Stained calcium deposits were destained with 10 CPCbuffer to measure the level of staining Data are means plusmn SD of six replicates lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001 versus control
abundant spots were selected for mass spectrometry (MS)analysis
29 Mass Spectrometry and Database Search Protein spotswere excised from the 2DE gels using a pipette tip Gel pieceswere destained in a solution of 15mMpotassium ferricyanideand 50mM sodium thiosulfate (1 1) washed with deionizedwater and dehydrated in 100 acetonitrile (ACN) Sampleswere rehydrated for digestion with trypsin (125mgmL) at4∘C for 30min Excess trypsin solution was replaced with25mM ammonium bicarbonateThe samples were incubatedovernight at 37∘C Peptides were then extracted twice with50 ACN5 TFA followed by 100 ACN for 15min eachAfter drying the peptide extracts were desalted with ZipTipPipette Tips (Millipore) Mass spectrometry was done usingan ultraflex III MALDI-TOFTOF-MS (Bruker) with a highvoltage of 20 kV and spectra were externally calibrated
using the peptide standard Maker Protein identification wasdetermined by matching the peptide mass fingerprinting(PMF) and MALDI-TOFTOF-MS results via MASCOT(version 22 Matrix Science) against NCBInr database withBioTools software Database searches were performed usingthe following parameters taxonomy rice enzyme trypsinand one missed cleavage allowed Carbamidomethylationwas selected as a fixed modification and the oxidation wasallowed as a variable PMF tolerance set to 100 ppm andMSMS tolerance set to 07Da A protein was regarded asidentified if the MASCOT protein score was above the 5significance threshold for the database (score gt64)
210 Western Blotting Analysis To verify the results of 2DEof the identified proteins we randomly chose four proteinsfor Western blot FPR2 TCEB1 PHB and alpha-spectrinCytosolic extractswere prepared fromcells and the protein in
Evidence-Based Complementary and Alternative Medicine 5
50
Mr (
kDa)
37
25
20
14
pH 3 10
Blank-control group
(a)
pH 3 10
HQSXD-treated group
50
Mr (
kDa)
37
25
20
14
(b)
pH 3 10
JKSQW-treated group
50
Mr (
kDa)
37
25
20
14
(c)
Figure 4 Representative silver nitrate stained gels showing two-dimensional electrophoresis protein profiles of (a) blank-control group (b)HQSXD-treated group and (c) JKSQW-treated group Molecular weight (MW kDa) and isoelectric point (119875
119868
) are indicated along the 119910- and119909-axes respectively
the supernatant was quantified using the BCA protein assaykit (Beyotime Institute of Biotechnology China) A sample(50120583g) was electrophoresed on a 10 SDS-polyacrylamidegel and subsequently transferred onto a PVDF membrane(Millipore) After blocking with 5 nonfat dry milk themembranes were incubated with anti-FPR2 (M-73 sc-66901Santa Cruz Biotechnology) anti-alpha-spectrin (C-11 sc-46696 Santa Cruz Biotechnology) anti-TCEB1 (ProteinTechGroup Inc China) and anti-PHB (ProteinTech GroupInc China) The bound antibodies were detected using ahorseradish peroxidase- (HRP-) conjugated secondary anti-body and visualized by an enhanced chemiluminescencedetection system followed by quantification using the ImageJ2x
211 Statistical Analysis Data are presented as mean plusmn SD oftriplicate samples Comparisons were performed using one-way analysis of variance (ANOVA) followed byDunnettrsquos testand the difference was considered statistically significant if119901 lt 005
3 Results
31 Effects of HQSXD-Treated Rat Serum on Proliferation ofOsteoblasts The proliferation of primary osteoblasts showedan upward trend compared to that of blank control Howeverthere was no significant difference detected in the prolifer-ation between HQSXD-treated and JKSQW-treated groups
6 Evidence-Based Complementary and Alternative Medicine
Table 1 Effects of drugs on proliferation of osteoblasts
Groups 119873Oral administration
dose (gsdotkgminus1)Serum additivevolume () OD value
Blank-control 5 mdash 10 050 plusmn 003HQSXD-treated 5 14 10 0634 plusmn 0028lowastlowast
JKSQW-treated 5 0520 10 063 plusmn 004lowastlowast
Note lowastlowastp lt 001 compared with BLANK-control-S
Similar changes in proliferation between JKSQW-treatedgroup andHQSXD-treated group indicated that HQSXDhadan influence on osteoblasts (Table 1 Figure 1)
32 HQSXD-Treated Rat Serum Enhances Primary OsteoblastDifferentiation ALP is an important biochemical marker ofdifferentiated osteoblasts and the effects of drug on ALPactivities in osteoblasts were first determined Results ofALP staining showed that HQSXD-S and JKSQW-control-S stimulated osteoblast differentiation (Figure 2(a)) Thecells cultured with HQSXD-S showed a significantly higherALP activity than that cultured with BLANK-control-S(Figure 2(b)) ARS staining in osteoblasts was assessed after21 days of incubation to examine whether HQSXD enhancedbonemineralization during osteoblastogenesis In osteoblaststreated with BLANK-control-S the calcium deposition in themineralized matrix was minimal The proportional areas ofAlizarin Red-positive staining in the HQSXD-treated groupand JKSQW-treated group were higher than that in theBlank-control group (Figure 3(a)) As shown in Figure 3(b)the level and intensity of ARS staining indicated the extentof mineralization which increased upon treatment withHQSXD-S
33 Protein Expression Profile in HQSXD-Treated andHQSXD-Untreated Osteoblasts Two-dimensional electro-phoresis and gel sliver nitrate staining were conducted tofurther investigate the differential protein expression betweenHQSXD-S-treated and HQSXD-S-untreated osteoblastsAfter optimization of the 2DE gels with representative2DE gel images shown in Figure 4 approximately 938 plusmn 26875 plusmn 34 and 904 plusmn 22 protein spots were detected in blankprotein sample HQSXD protein sample and JKSQWproteinsample respectively During analysis with ImageMaster 2DPlatinum spots with an overlap ratio absolute value ge2were recognized as differentially expressed Thirty-eightprotein spots were found to be significantly regulated amongthree groups of which 15 spots were downregulated and23 spots upregulated Ten of these 38 spots exhibited amore than twofold increase or decrease in abundance asobserved in all replicate gels These 10 regulated proteinswere indicated by the circle in Figure 5 and the selectedregions that showed significant differences in proteinexpression profile of osteoblasts among three groups wereshown in Figure 6 All of them were excised from thegels for further identification by MALDI-TOFTOF-MSanalysis
50
Mr (
kDa)
37
25
20
14
pH 3 10
87
3
6
5
2 10
4
1
9
Figure 5 Representative 2DE gel image Ten spots that were statis-tically significant (119901 lt 005) are shown in the map and indicatedby numbers All were cut from the gels for further identification byMALDI-TOFTOF-MS analysis outlined in Table 2
34 Identification of the Differentially Expressed ProteinsProteins were identified by MALDI-TOFTOF-MS Ten pep-tide mass fingerprints (PMFs) and 50 peptide fragmentfingerprints (PFF) were successfully obtained A selectedPMF of protein spot 6 is displayed in Figure 7(a) and theTOFTOF analysis is shown in Figures 7(b)ndash7(f) All PMFswere evaluated with the Mascot software in NCBInr databaseto identify the protein spots The result had high confidenceif the protein was ranked as the best hit with a significantscore and high sequence coverage Finally we identified eightproteins in these spots Properties of the identification of eightselected protein spots are summarized in Table 2
35 Effect of HQSXD on the Expression of Proteins That Regu-late Antiosteoporotic Activity To further investigate the influ-ence of HQSXD on the expression of antiosteoporotic pro-teins we examined the expression of FPR2 alpha-spectrinPHB andTCEB1 byWestern blottingThe expression of FPR2was increased by treatment with HQSXD-S in comparisonto the blank-control group However HQSXD remarkablydecreased alpha-spectrin PHB and TCEB1 protein levelscompared with blank-control group (Figure 8) Results fromWestern blot manifested the same trend as from proteomicanalysis
Evidence-Based Complementary and Alternative Medicine 7
Spot 1 Spot 2 Spot 3 Spot 4 Spot 5
Spot 6 Spot 7 Spot 8 Spot 9 Spot 10
Blank-control group
HQSXD-treated group
JKSQW-treated group
Blank-control group
HQSXD-treated group
JKSQW-treated group
Figure 6 The ten protein spots of osteoblasts that were treated with and without HQSXD-S Selected regions showed significant differencesin the protein expression profile of osteoblasts among the three groups Upregulated spots are indicated by green circles and downregulatedones by red circles
Table 2 Summary of differentially expressed proteins in osteoblasts treated with HQSXD
Spotnumbera
Proteinscoreb
Matchingpeptides(number)
Theoretical119875119868
cTheoreticalMr (Da)c Target protein Species
1 70 6 927 39299 N-Formyl peptide receptor 2 Rattus norvegicus3 67 7 565 54851 Alpha-spectrin Rattus norvegicus
5 76 6 1008 43712 Heparan sulfate (glucosamine)3-O-sulfotransferase 3A1 Rattus norvegicus
6 173 12 557 29859 Prohibitin Rattus norvegicus
7 67 4 459 12752 Transcription elongationfactor B (SIII) polypeptide 1 Rattus norvegicus
8 70 11 498 127444 Chromosome segregationprotein Rattus norvegicus
9 101 4 691 17386 Nucleoside diphosphatekinase Rattus norvegicus
10 84 12 886 48199 Mast cell carboxypeptidase A Rattus norvegicusaProtein spot number according to Figure 3bProtein scores were based on combined mass and massmass spectra from MALDI-TOFTOF identification MScTheoretical molecular mass (Mr) and isoelectric point (119875
119868) from the NCBInr database
8 Evidence-Based Complementary and Alternative Medicine
724502
995615
861066
1023645
13968251149581
1198660
17947921606823 19980611460637929573 16698152284146
2225096
23712291876991 2119118
00
05
10
15
20
Inte
nsity
(au
)
1000 1500 2000 2500 3000 3500mz
times104
(a)
1023635
687769
832885
643812271987
453127352094621199 772789 979506
174985 322097 876761111971 665734
709626552159385049
86016 295124139982
0
1000
2000
3000
Inte
nsity
(au
)
200 400 600 800 1000 1200mz
(b)
Inte
nsity
(au
)
1037662
254948
467075
989061366041 635224
174962111956 703694338068 886923
80082486005 567816
1012722
200937
287991
0
500
1000
1500
2000
200 400 600 800 1000 1200mz
(c)
Inte
nsity
(au
)
1149574
887334
174919
953820
519998 1105500
633033111931386933
937990
842341486004 595064 785790302933254963
69965
142902
0
1000
2000
3000
200 400 600 800 1000 1200mz
(d)
Inte
nsity
(au
)
1198658
174936
2619201154530
111976440964358918 932243609086
84213585996 325947 741110572984515028287928
400987
228936
200907
0
500
1000
1500
2000
2500
3000
200 400 600 800 1000 1200 1400mz
(e)
Inte
nsity
(au
)
1396813
12406611354732
174968
1070439867233227008111958 286957 1202691
513068417073354995
0
500
1000
1500
2000
2500
200 400 600 800 1000 1200 1400 1600mz
(f)
Figure 7The results of the MALDI-TOFTPF-MS analysis of protein spot 6 (a) Peptide mass fingerprinting and ((b)ndash(f)) peptide fragmentfingerprinting of spot 6
4 Discussion
In this study we found that the proliferative activities ofosteoblasts between HQSXD-S-treated group and JKSQW-control-S-treated group were similar This investigationdemonstrates that HQSXD can significantly facilitate boneformation through increasing the number of osteoblastswhich is beneficial to the treatment of osteoporosis By mea-suring ALP activity HQSXD was first screened for its abilityto induce osteogenesis HQSXD is capable of significantly
promoting osteoblast differentiation as well as increasingosteoblast mineralization
Results of the identification of the selected protein spotsare summarized in Table 2 The molecular weight (Mr) andisoelectric point (119875
119868) of each protein spot shown in Table 2
are theoretical values The eight protein spots were identifiedas (1) N-formyl peptide receptor 2 (FPR2) (2) alpha-spectrin(3) heparan sulfate (glucosamine) 3-O-sulfotransferase 3A1(HS3ST3A1) (4) prohibitin (PHB) (5) transcription elonga-tion factor B (SIII) polypeptide 1 (TCEB1) (6) chromosome
Evidence-Based Complementary and Alternative Medicine 9
TCEB1
PHB
FPR2
Blank HQSXD JKSQW
120573-actin
Alpha-spectrin
Figure 8 HQSXD-S treatment decreased alpha-spectrin PHBand TCEB1 expression and increased FPR2 expression Cell lysates(50 120583g)were processed forWestern blot analysis Normalization per-formed to120573-actinThe bands shown here were from a representativeexperiment repeated three times
segregation protein (7) nucleoside diphosphate kinase and(8) mast cell carboxypeptidase AWe detected some proteinsrelated to transcription cell proliferation differentiationand apoptosis such as FPR2 prohibitin alpha-spectrinheparan sulfate (glucosamine) 3-O-sulfotransferase 3A1 andtranscription elongation factor B (SIII) polypeptide 1 whichvaried greatly after HQSXD treatment
Our results suggest that HQSXD can upregulate theexpression of FPR2 N-Formyl peptide receptor (FPR) is aG protein-coupled receptor which modulates stromal celldifferentiation [14] and binds to N-formyl peptides suchas N-formyl-methionyl-leucyl-phenylalanine (fMLP) Ourstudy has shown that fMLP enhances the differentiationof MSCs into osteoblasts via an FPR-mediated signalingpathway and results in bone formation [15] The FPR2receptor belongs to the formyl peptide receptor family thatis involved in signaling stem cell adhesion migration andhoming for injured and inflamed tissues awaiting repair thiscould potentially be exploited to direct the stem cells to targetspecific tissue site [16] Therefore the current results implythat treatment with HQSXDmight promote the formation ofosteoblasts by upregulating the expression level of FPR2
We found that HQSXD could inhibit the expression ofalpha-spectrin Alpha-spectrin includes two genes encodingfor alpha-I subunit and alpha-II subunit each of thempresenting its specific cellular expression pattern Alpha-II spectrin deficiency is associated with cell proliferationdefects due to cell cycle arrest in theG1 phase (first gap phase)[17] Spectrin and protein kinase C theta were observed inaggregates during the early stage of apoptosis [18] Hencewe presume that alpha-spectrin is involved in the antiosteo-porotic effect of HQSXD
In this study PHB was downregulated in HQSXD-treated osteoblasts Prohibitin a highly conservative proteinregulates the cohesion of sister chromatids cellular signalingmitochondrial biogenesis [19] cell proliferation differentia-tion apoptosis and gene transcription [20 21] PHB blocksthe transition of cells from G1 phase to S phase of the cellcycle thereby arresting cell proliferation [22 23] PHB hasbeen reported to affect the apoptotic pathways by repressingthe transcriptional activity of E2F1 [24 25] As mentionedabove it is possible that HQSXD induces proliferationdifferentiation and apoptosis partly through downregulatingexpression of PHB in osteoblasts It is likely that PHB mightbe the novel candidate in the new antiosteoporotic drugscreening
The expression of TCEB1 was downregulated in HQSXD-treatment osteoblasts TCEB1 a 13 kDa protein also namedelongin C was originally identified as a member of themammalian transcription factor SIII that increases the rateof transcription by suppressing RNA polymerase II pausing[26] As part of a family of separate complexes containingelongin B and various substrate specificity factors it acts asan E3 ubiquitin ligase [27] Elongin B (ELB 1) and elonginC (ELC1) form a stable complex and that depletion of eithergene product by RNA-mediated interference (RNAi) causespronounced defects in the second meiotic division and arrestof germ cell proliferation in G1 [28] Therefore it is possiblethat that TCEB1 is involved in the antiosteoporotic effect ofHQSXD
Although this study has thrown some light on themechanism of HQSXD action we failed to characterize thewell-identified protein (eg HS3ST3A1) closely involved inosteoporosis HS3ST3A1 a member of the heparan sulfatebiosynthetic enzyme family possesses heparan sulfate glu-cosaminyl 3-O-sulfotransferase activity Depletion of Hs3st-A in enterocytes results in increased intestinal stem cellproliferation and tissue homeostasis loss [29]
5 Conclusions
The results confirm that HQSXD has a beneficial effect onosteoblasts and alters the expression level of some proteinsin osteoblasts The protein expressed by osteoblasts treatedwith HQSXD may be involved in cell proliferation anddifferentiation and other physiological processes and in theregulation of cell activation Further study is needed toinvestigate the effects of major active constituents in HQSXDon protein expression on osteoblast so as to demonstrate theinteraction and synergistic mechanism
Conflict of Interests
All the authors state that they have no conflict of interests todeclare
Authorsrsquo Contribution
Chong-Chong Guo and Li-Hua Zheng contributed equally tothis work
10 Evidence-Based Complementary and Alternative Medicine
Acknowledgments
The authors thank Cheng-Ming Liu for her excellent tech-nical assistance This work was supported by the grantfrom the National Natural Science foundation of China (no81273779)
References
[1] S Khosla and B L Riggs ldquoPathophysiology of age-related boneloss and osteoporosisrdquo Endocrinology andMetabolism Clinics ofNorth America vol 34 no 4 pp 1015ndash1030 2005
[2] P Sambrook andCCooper ldquoOsteoporosisrdquoTheLancet vol 367no 9527 pp 2010ndash2018 2006
[3] M R Allen and D B Burr ldquoThree years of alendronate treat-ment results in similar levels of vertebral microdamage as afterone year of treatmentrdquo Journal of Bone and Mineral Researchvol 22 no 11 pp 1759ndash1765 2007
[4] S Davison and S R Davis ldquoHormone replacement therapycurrent controversiesrdquo Clinical Endocrinology vol 58 no 3 pp249ndash261 2003
[5] W-Y Jiang ldquoTherapeutic wisdom in traditional Chinesemedicine a perspective from modern sciencerdquo Trends in Phar-macological Sciences vol 26 no 11 pp 558ndash563 2005
[6] Z Xu and Z K Zhou ldquoClinical study on the treatment of 36cases of postmenopausal osteoporosis with HuangQiSanXianTangrdquo Guiding Journal of Traditional Chinese Medicine andPharmacy vol 15 no 1 pp 9ndash11 2009
[7] H Zhang W-W Xing Y-S Li et al ldquoEffects of a traditionalChinese herbal preparation on osteoblasts and osteoclastsrdquoMaturitas vol 61 no 4 pp 334ndash339 2008
[8] L Qin T Han Q Zhang et al ldquoAntiosteoporotic chemical con-stituents from Er-Xian Decoction a traditional Chinese herbalformulardquo Journal of Ethnopharmacology vol 118 no 2 pp 271ndash279 2008
[9] M Millikan A Kolasani and H Xu ldquoDetermination and com-parison of mineral elements in traditional Chinese herbalformulae at different decoction times used to improve kidneyfunctionmdashchemometric approachrdquo African Journal of Tradi-tional Complementary and Alternative Medicines vol 8 no 5pp 191ndash197 2011
[10] I R Orriss S E B Taylor and T R Arnett ldquoRat osteoblastculturesrdquoMethods inMolecular Biology vol 816 pp 31ndash41 2012
[11] JM Baek J-Y Kim Y-H Cheon et al ldquoDual effect ofChrysan-themum indicum extract to stimulate osteoblast differentia-tion and inhibit osteoclast formation and resorption in vitrordquoEvidence-Based Complementary and Alternative Medicine vol2014 Article ID 176049 13 pages 2014
[12] A I Idris I R Greig E Bassonga-Landao S H Ralston andR J Vanrsquot Hof ldquoIdentification of novel biphenyl carboxylic acidderivatives as novel antiresorptive agents that do not impairparathyroid hormone-induced bone formationrdquo Endocrinologyvol 150 no 1 pp 5ndash13 2009
[13] M Otani J Tabata T Ueki K Sano and S Inouye ldquoHeat-shock-induced proteins from Myxococcus xanthusrdquo Journal ofBacteriology vol 183 no 21 pp 6282ndash6287 2001
[14] M E Nuttall and J M Gimble ldquoIs there a therapeutic opportu-nity to either prevent or treat osteopenic disorders by inhibitingmarrow adipogenesisrdquo Bone vol 27 no 2 pp 177ndash184 2000
[15] M K Shin Y H Jang H J Yoo et al ldquoN-formyl-methionyl-leucyl-phenylalanine (fMLP) promotes osteoblast differentia-tion via the N-formyl peptide receptor 1-mediated signaling
pathway in humanmesenchymal stem cells from bonemarrowrdquoThe Journal of Biological Chemistry vol 286 no 19 pp 17133ndash17143 2011
[16] A Viswanathan R G Painter N A Lanson Jr and G WangldquoFunctional expression ofN-formyl peptide receptors in humanbone marrow-derived mesenchymal stem cellsrdquo Stem Cells vol25 no 5 pp 1263ndash1269 2007
[17] S Metral B Machnicka S Bigot Y Colin D Dhermy and M-C Lecomte ldquo120572II-spectrin is critical for cell adhesion and cellcyclerdquo Journal of Biological Chemistry vol 284 no 4 pp 2409ndash2418 2009
[18] P M Dubielecka M Grzybek A Kolondra et alldquoAggregation of spectrin and PKCtheta is an early hallmarkof fludarabinemitoxantronedexamethasone-induced apopto-sis in Jurkat T and HL60 cellsrdquo Molecular and CellularBiochemistry vol 339 no 1-2 pp 63ndash77 2010
[19] C Merkwirth and T Langer ldquoProhibitin function withinmitochondria essential roles for cell proliferation and cristaemorphogenesisrdquo Biochimica et Biophysica Acta vol 1793 no 1pp 27ndash32 2009
[20] A L Theiss and S V Sitaraman ldquoThe role and therapeuticpotential of prohibitin in diseaserdquoBiochimica et BiophysicaActavol 1813 no 6 pp 1137ndash1143 2011
[21] V Sanchez-Quiles E Santamarıa V Segura L Sesma J Prietoand F J Corrales ldquoProhibitin deficiency blocks proliferationand induces apoptosis in human hepatoma cells molecularmechanisms and functional implicationsrdquo Proteomics vol 10no 8 pp 1609ndash1620 2010
[22] S Manjeshwar D E Branam M R Lerner D J Brackettand E R Jupe ldquoTumor suppression by the prohibitin gene3rsquountranslated region RNA in human breast cancerrdquo CancerResearch vol 63 no 17 pp 5251ndash5256 2003
[23] S Manjeshwar M R Lerner X-P Zang et al ldquoExpressionof prohibitin 31015840 untranslated region suppressor RNA altersmorphology and inhibits motility of breast cancer cellsrdquo Journalof Molecular Histology vol 35 no 6 pp 639ndash646 2004
[24] M Lacroix R-A Toillon and G Leclercq ldquop53 and breastcancer an updaterdquo Endocrine-Related Cancer vol 13 no 2 pp293ndash325 2006
[25] G Fusaro P Dasgupta S Rastogi B Joshi and S ChellappanldquoProhibitin induces the transcriptional activity of p53 andis exported from the nucleus upon apoptotic signalingrdquo TheJournal of Biological Chemistry vol 278 no 48 pp 47853ndash47861 2003
[26] T Aso W S Lane J W Conaway and R C ConawayldquoElongin (SIII) a multisubunit regulator of elongation by RNApolymerase IIrdquo Science vol 269 no 5229 pp 1439ndash1443 1995
[27] T D CumminsM DMendenhall M N Lowry et al ldquoElonginC is a mediator of Notch4 activity in human renal tubule cellsrdquoBiochimica et Biophysica Acta vol 1814 no 12 pp 1748ndash17572011
[28] Y Sasagawa K Kikuchi K Dazai and A HigashitanildquoCaenorhabditis elegans ElonginBC complex is essential for cellproliferation and chromosome condensation and segregationduring mitosis and meiotic division IIrdquo Chromosome Researchvol 13 no 4 pp 357ndash375 2005
[29] Y Guo Z Li and X Lin ldquoHs3st-A and Hs3st-B regulateintestinal homeostasis in Drosophila adult midgutrdquo CellularSignalling vol 26 no 11 pp 2317ndash2325 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Evidence-Based Complementary and Alternative Medicine
level and bone mineral density (BMD) and repressed boneabsorption function in postmenopausal women [6]This sug-gested that HQSXD has beneficial effects in the treatment ofosteoporosis However little is known about the mechanismsand targets underlying the effects ofHQSXDon osteoporosis
The Chinese traditional medicine theory believes thatbone activities are controlled by the kidney Strong ldquokidneyrdquocan nourish bones but the weak ldquokidneyrdquo might hasten bonedeterioration [7] Kidney deficiency and blood stasis are themain pathological basis of osteoporosis Huangqi Sanxiandecoction is composed of eight Chinese medicinal herbsof these Epimedii Folium and Cistanche Herba strengthenkidneys while Radix Salviae Miltiorrhizae and Radix noto-ginseng invigorate the circulation of blood These herbs arean excellent combination for highlighting their superiority inthe treatment of osteoporosis [8]
Jinkui Shenqi Wan an ancient Chinese herbal formulais indicated in the Chinese Pharmacopoeia (2010) for thetreatment of Yang insufficiency of kidney weakness andsoreness of the loins and the knees cold feeling in the limbsand frequent urination Human clinical studies have certifiedthat Jinkui Shenqi Wan exerts a therapeutic effect on thekidney-Yang deficiency [9] Hence we used Jinkui ShenqiWan as a positive control in this study
In the present study seropharmacology and functionalproteomics technology were used to explore the multipleproteins associated with the antiosteoporotic effect Theresults suggest a basis for the clinical use of HQSXD in thetreatment of patients with osteoporosis
2 Materials and Methods
21 Animals Thirty Sprague-Dawley rats (fifteen male andfifteen female) weighing about 250ndash300 g were purchasedfrom the Animal Experimental Center of Guangdong Med-ical College (Dongguan China) The animals were housedindividually in a regulated environment (24 plusmn 05∘C) witha 12-hour lightdark cycle (under light 0800ndash2000 h) Foodand water were given ad libitum throughout the experimentAfter three days of acclimation male and female SD rats wererandomly divided into three groups blank-control groupexperimental group treated withHuangqi Sanxian decoctionand positive control group treated with Jinkui Shenqi WanThe Committee of Experimental Animal Administration ofthe University approved the study and the procedures ofthe experiment were in accordance with generally acceptedinternational rules and regulations
22 Preparation of HQSXD and JKSQW HQSXD includedeight plant extracts including Radix Astragali (root Chineseherbal name Huang-Qi) Epimedii Folium (leaf Chineseherbal name Yin-Yang-Huo) Cistanche Herba (succulentstem Chinese herbal name Rou-Cong-Rong) Radix noto-ginseng (root and rhizome Chinese herbal name San-Qi) Radix Salviae Miltiorrhizae (root and rhizome Chineseherbal name Dan-Shen) Corydalis Rhizoma (rhizome Chi-nese herbal name Yan-Hu-Suo) Radix Angelicae Sinensis
(root Chinese herbal name Dang-Gui) and Radix Clema-tidis (root and rhizome Chinese herbal name Wei-Ling-Xian) in a ratio of 15 10 10 5 1010 8 10 [6] The aboveeight medicinal extracts were obtained from DongguanSinopharm (Dongguan China) and identified by ProfessorZhou (Department of Pharmacy Guangdong Medical Col-lege Dongguan China) The eight medicinal plant materialsin the mixture (270 g) were powdered and coboiled with1000mLwater for 2 hoursThe extraction was repeated twiceThe filtrates were concentrated to 200mL under reducedpressure and kept at 4∘C
JKSQW consisted of Radix Rehmanniae Preparata (rootChinese herbal name Di-Huang) Dioscoreae Rhizome (rootChinese herbal name Shan-Yao) Fructus Corni Officinalis(fructus Chinese herbal name Shan-Zhu-Yu) SclerotiumPoriae Cocos (dried sclerotia Chinese herbal name Fu-Ling)CortexMoutan Radicis (root bark Chinese herbal nameMu-Dan-Pi) Rhizoma Alismatis Orientalis (stem Chinese herbalname Ze-Xie) Ramulus Cinnamomi Cassiae (twig Chineseherbal name Gui-Zhi) Radix Aconiti Lateralis Praeparata(root Chinese herbal name Fu-Zi)Radix Achyranthis Biden-tatae (root Chinese herbal name Niu-Xi) and PlantaginisSemen (seed Chinese herbal name Che-Qian-Zi) [9] Asper the instructions 5mg Jinkui Shenqi Wan (Beijing TongRen Tang Pharmaceutical Technology Development IncDongguan China) was dissolved in 100mL distilled waterbefore administration The experimental dose for HQSXDand JKSQW in the present study was equivalent to thecorresponding clinical prescription dose for a human subjectweighing 60 kg
23 Drug Administration and Sample Collections Rats wererandomly divided into three groups of 10 animals each(each group included five males and five females) Chinesemedicine HQSXD solution (14 gkg) and medicine JKSQWsolution (0520 gkg) were administered orally every day forthree days twice a day The blank-control group was orallyadministered distilled water and they were monitored con-currently with the HQSXD-experimental groups At the endof the experiment sixty minutes after the last treatment theanimalswere exposed to ether anesthesia blood sampleswerecollected by heart puncture under aseptic conditions andthen centrifuged for 15min to obtain serum samples SerumofHQSXD-treated rats and control serumwere inactivated at56∘C in awater bath for 30min and filtered through a 022120583mfilter membrane termed HQSXD-S JKSQW-control-S andBLANK-control-S respectively and then stored at minus80∘C
24 Primary Osteoblasts Culture and Assay for OsteoblastProliferation Primary rat osteoblast cells were obtained from1-day-old neonatal Sprague-Dawley rats as described previ-ously [10] Primary osteoblasts were cultured by seeding 96-well plates with a density of 1 times 104 per well and incubatedfor 24 h After adhesion of cells Dulbeccorsquos Modified EagleMedium (DMEM) was added and incubated for another 24hours Next after discarding the medium test (HQSXD-S)and control (JKSQW-control-S BLANK-control-S) samples
Evidence-Based Complementary and Alternative Medicine 3
were added at a concentration of 10 (vv) and the cellswere incubated at 37∘C in a humid atmosphere containing 5CO2for 72 h Thereafter 5mgmL of 3-(45-dimethylthiazol-
2-yl)-25-diphenyltetrazolium bromide (MTT) was addedand incubated for 4 hours after which the medium wasdiscarded and dimethyl sulfoxide (DMSO) (150120583L) wasadded Absorbance was measured at 490 nm using a Syn-ergy 2 multifunctional microplate reader (Bio-Tek) to assessosteoblast proliferation
25 ALP Activity and Staining Assay Osteoblasts were sus-pended in DMEM to obtain a cell density of 5 times 105mL2mL aliquots of the cell suspension were added to 6-wellplates After 24 h of incubation the medium was changedand cells were incubated with test (HQSXD-S) and control(JKSQW-control-S BLANK-control-S) at a concentration of10 (vv) for 3 days Alkaline phosphatase (ALP) activitywas measured using an ALP assay kit (Sigma) as describedpreviously [11] For ALP staining after incubation withHQSXD-S JKSQW-control-S and BLANK-control-S (con-centration of 10 (vv)) for three days the cells were fixedin 70 ethanol for 15min washed and then incubatedwith ALP staining buffer nitro blue tetrazolium5-bromo-4-chloro-3-indolyl phosphate (NBTBCIP) (Beyotime Instituteof Biotechnology China) at 37∘C for 30min air dried andphotographed
26 Mineralization Assay After 21 days of differentiationthe mineralization of osteoblasts was analyzed as describedpreviously [12] Briefly the cells were washed with phosphatebuffered saline (PBS) and fixed with 70 ethanol for onehourThe cellswere then rinsed in distilledwater stainedwith05 Alizarin Red S (ARS) at pH 42 with rotation for 30minat 37∘C and subsequently washed with distilled water anddried in air Stained cultures were photographed To analyzeARS activity the ARS in stained cells was destained with10 cetylpyridinium chloride (CPC) monohydrate solution(Sigma) for 30min with shaking The absorbance was mea-sured at 562 nmusing a Synergy 2multifunctionalmicroplatereader
27 Protein Extraction Osteoblasts cells were incubated in50mL culture flasks and grown to subconfluence (approx-imately 60ndash70) and then treated with test (HQSXD-S) and control (JKSQW-control-S BLANK-control-S) at aconcentration of 10 (vv) for 72 h respectively At the end ofthe incubation period osteoblasts were collected by centrifu-gation at 1000 rpm (4∘C) The osteoblasts were washed twicewith cold PBS after discarding the medium Total proteinswere extracted in a chilled lysis buffer containing 7M urea2M thiourea 4 (wv)CHAPS 40mMdithiothreitol (DTT)2 (vv) IPG buffer pH 3ndash10 4 120583gmL protease inhibitormixture and 4 120583gmL phosphatase inhibitors After additionof the chilled lysis buffer the cell solution was kept oscillatingfor 1 h at 4∘C to solubilize the proteins The homogenatewas subsequently centrifuged for 15min at 14000 rpm at
08
06
04
02
00
OD
(490
nm)
lowastlowast lowastlowast
JKSQW-treatedgroup
Blank-control HQSXD-treatedgroupgroup
Figure 1 Effects of drugs on proliferation of osteoblasts Althoughthere was some improvement in the proliferation of osteoblastsin the HQSXD-treated group it was not statistically significantwhen compared to the values observed in the group given JKSQW-control-S Data are expressed as the mean plusmn SD
4∘C Total protein concentration was quantified using theBradford assay (Amresco)
28 Two-Dimensional Electrophoresis (2DE) and Image Anal-ysis Electrophoresis was carried out at least thrice forreproducibility Two-dimensional electrophoresis (2DE) wascarried out as described previously [13] with some mod-ifications For the first-dimension isoelectric focusing gel250 120583g of protein was used for isoelectric focusing (IEF)using 17 cm nonlinear (NL) IPG strips (pH 3ndash10) whichwere immersed in rehydration buffer (8M urea 2 CHAPS25mM DTT 02 (wv) Bio-Lyte 01 bromophenol blueand 1 IPG buffer) The strips were covered with mineraloil (GE Healthcare) to prevent samples from evaporationand rehydrated for 10ndash20 hours Isoelectric focusing gelelectrophoresis was performed by using an electrophoresisapparatus (GE Healthcare) The running conditions of theIEF process were as follows 100V 1 h 200V 1 h 300V 1 h500V 1 h 1000V 1 h 2000V 1 h and 8000V up 60000VAfter the IEF the IPG strips were equilibrated with equilibra-tion buffer-1 (6M urea 2 SDS 20 glycerol 0375M Tris-HCl [pH 88] bromophenol blue dye and 1 [wv] DTT) for15min and then were repeated for an additional 15 minutes in5mL of equilibration buffer-2 except that DTT was replacedby 25 [wv] iodoacetamide The second dimension wasdone with a 125 acrylamide gel at 10mAgel for onehour and then at 38mAgel until the bromophenol blue dyereached the bottom of the gel After electrophoresis the gelswere visualized by silver nitrate staining Gels were scannedusing the ImageScanner (GE Healthcare) The images wereanalyzed with ImageMaster 2D Platinum v70 software (GEHealthcare San Francisco CA) including spot detectionbackground subtraction gel matching and normalizationSpots were detected and matched automatically to a mastergel and then edited manually Matched spots from triplicategel sets that showed overlap ratio with an absolute valuege2 were recognized as differentially expressed Differentially
4 Evidence-Based Complementary and Alternative Medicine
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
ALP staining
(a)
025
020
015
010
005
000
ALP
(Ug
prot
)
lowastlowast
lowastlowast
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
(b)
Figure 2 HQSXD-S enhances primary osteoblast differentiation (a) Primary osteoblasts were treated with various drug-treated rat serumsfor 3 days ALP-positive cells were stained with ALP solution (b) Effect of HQSXD-S on alkaline phosphatase activity in primary cultureosteoblasts Data are means plusmn SD of six replicates lowastlowast119901 lt 001 versus control
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
ARS staining
(a)
025
020
015
010
005
000
ARS
activ
ity (562
nm)
lowastlowastlowastlowastlowast
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
(b)
Figure 3 HQSXD-S promotes bone mineralization during osteoblastogenesis (a) Calcium deposits stained with Alizarin Red solution seenin primary osteoblasts treated with various drug-treated rat serums for 21 days (b) Stained calcium deposits were destained with 10 CPCbuffer to measure the level of staining Data are means plusmn SD of six replicates lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001 versus control
abundant spots were selected for mass spectrometry (MS)analysis
29 Mass Spectrometry and Database Search Protein spotswere excised from the 2DE gels using a pipette tip Gel pieceswere destained in a solution of 15mMpotassium ferricyanideand 50mM sodium thiosulfate (1 1) washed with deionizedwater and dehydrated in 100 acetonitrile (ACN) Sampleswere rehydrated for digestion with trypsin (125mgmL) at4∘C for 30min Excess trypsin solution was replaced with25mM ammonium bicarbonateThe samples were incubatedovernight at 37∘C Peptides were then extracted twice with50 ACN5 TFA followed by 100 ACN for 15min eachAfter drying the peptide extracts were desalted with ZipTipPipette Tips (Millipore) Mass spectrometry was done usingan ultraflex III MALDI-TOFTOF-MS (Bruker) with a highvoltage of 20 kV and spectra were externally calibrated
using the peptide standard Maker Protein identification wasdetermined by matching the peptide mass fingerprinting(PMF) and MALDI-TOFTOF-MS results via MASCOT(version 22 Matrix Science) against NCBInr database withBioTools software Database searches were performed usingthe following parameters taxonomy rice enzyme trypsinand one missed cleavage allowed Carbamidomethylationwas selected as a fixed modification and the oxidation wasallowed as a variable PMF tolerance set to 100 ppm andMSMS tolerance set to 07Da A protein was regarded asidentified if the MASCOT protein score was above the 5significance threshold for the database (score gt64)
210 Western Blotting Analysis To verify the results of 2DEof the identified proteins we randomly chose four proteinsfor Western blot FPR2 TCEB1 PHB and alpha-spectrinCytosolic extractswere prepared fromcells and the protein in
Evidence-Based Complementary and Alternative Medicine 5
50
Mr (
kDa)
37
25
20
14
pH 3 10
Blank-control group
(a)
pH 3 10
HQSXD-treated group
50
Mr (
kDa)
37
25
20
14
(b)
pH 3 10
JKSQW-treated group
50
Mr (
kDa)
37
25
20
14
(c)
Figure 4 Representative silver nitrate stained gels showing two-dimensional electrophoresis protein profiles of (a) blank-control group (b)HQSXD-treated group and (c) JKSQW-treated group Molecular weight (MW kDa) and isoelectric point (119875
119868
) are indicated along the 119910- and119909-axes respectively
the supernatant was quantified using the BCA protein assaykit (Beyotime Institute of Biotechnology China) A sample(50120583g) was electrophoresed on a 10 SDS-polyacrylamidegel and subsequently transferred onto a PVDF membrane(Millipore) After blocking with 5 nonfat dry milk themembranes were incubated with anti-FPR2 (M-73 sc-66901Santa Cruz Biotechnology) anti-alpha-spectrin (C-11 sc-46696 Santa Cruz Biotechnology) anti-TCEB1 (ProteinTechGroup Inc China) and anti-PHB (ProteinTech GroupInc China) The bound antibodies were detected using ahorseradish peroxidase- (HRP-) conjugated secondary anti-body and visualized by an enhanced chemiluminescencedetection system followed by quantification using the ImageJ2x
211 Statistical Analysis Data are presented as mean plusmn SD oftriplicate samples Comparisons were performed using one-way analysis of variance (ANOVA) followed byDunnettrsquos testand the difference was considered statistically significant if119901 lt 005
3 Results
31 Effects of HQSXD-Treated Rat Serum on Proliferation ofOsteoblasts The proliferation of primary osteoblasts showedan upward trend compared to that of blank control Howeverthere was no significant difference detected in the prolifer-ation between HQSXD-treated and JKSQW-treated groups
6 Evidence-Based Complementary and Alternative Medicine
Table 1 Effects of drugs on proliferation of osteoblasts
Groups 119873Oral administration
dose (gsdotkgminus1)Serum additivevolume () OD value
Blank-control 5 mdash 10 050 plusmn 003HQSXD-treated 5 14 10 0634 plusmn 0028lowastlowast
JKSQW-treated 5 0520 10 063 plusmn 004lowastlowast
Note lowastlowastp lt 001 compared with BLANK-control-S
Similar changes in proliferation between JKSQW-treatedgroup andHQSXD-treated group indicated that HQSXDhadan influence on osteoblasts (Table 1 Figure 1)
32 HQSXD-Treated Rat Serum Enhances Primary OsteoblastDifferentiation ALP is an important biochemical marker ofdifferentiated osteoblasts and the effects of drug on ALPactivities in osteoblasts were first determined Results ofALP staining showed that HQSXD-S and JKSQW-control-S stimulated osteoblast differentiation (Figure 2(a)) Thecells cultured with HQSXD-S showed a significantly higherALP activity than that cultured with BLANK-control-S(Figure 2(b)) ARS staining in osteoblasts was assessed after21 days of incubation to examine whether HQSXD enhancedbonemineralization during osteoblastogenesis In osteoblaststreated with BLANK-control-S the calcium deposition in themineralized matrix was minimal The proportional areas ofAlizarin Red-positive staining in the HQSXD-treated groupand JKSQW-treated group were higher than that in theBlank-control group (Figure 3(a)) As shown in Figure 3(b)the level and intensity of ARS staining indicated the extentof mineralization which increased upon treatment withHQSXD-S
33 Protein Expression Profile in HQSXD-Treated andHQSXD-Untreated Osteoblasts Two-dimensional electro-phoresis and gel sliver nitrate staining were conducted tofurther investigate the differential protein expression betweenHQSXD-S-treated and HQSXD-S-untreated osteoblastsAfter optimization of the 2DE gels with representative2DE gel images shown in Figure 4 approximately 938 plusmn 26875 plusmn 34 and 904 plusmn 22 protein spots were detected in blankprotein sample HQSXD protein sample and JKSQWproteinsample respectively During analysis with ImageMaster 2DPlatinum spots with an overlap ratio absolute value ge2were recognized as differentially expressed Thirty-eightprotein spots were found to be significantly regulated amongthree groups of which 15 spots were downregulated and23 spots upregulated Ten of these 38 spots exhibited amore than twofold increase or decrease in abundance asobserved in all replicate gels These 10 regulated proteinswere indicated by the circle in Figure 5 and the selectedregions that showed significant differences in proteinexpression profile of osteoblasts among three groups wereshown in Figure 6 All of them were excised from thegels for further identification by MALDI-TOFTOF-MSanalysis
50
Mr (
kDa)
37
25
20
14
pH 3 10
87
3
6
5
2 10
4
1
9
Figure 5 Representative 2DE gel image Ten spots that were statis-tically significant (119901 lt 005) are shown in the map and indicatedby numbers All were cut from the gels for further identification byMALDI-TOFTOF-MS analysis outlined in Table 2
34 Identification of the Differentially Expressed ProteinsProteins were identified by MALDI-TOFTOF-MS Ten pep-tide mass fingerprints (PMFs) and 50 peptide fragmentfingerprints (PFF) were successfully obtained A selectedPMF of protein spot 6 is displayed in Figure 7(a) and theTOFTOF analysis is shown in Figures 7(b)ndash7(f) All PMFswere evaluated with the Mascot software in NCBInr databaseto identify the protein spots The result had high confidenceif the protein was ranked as the best hit with a significantscore and high sequence coverage Finally we identified eightproteins in these spots Properties of the identification of eightselected protein spots are summarized in Table 2
35 Effect of HQSXD on the Expression of Proteins That Regu-late Antiosteoporotic Activity To further investigate the influ-ence of HQSXD on the expression of antiosteoporotic pro-teins we examined the expression of FPR2 alpha-spectrinPHB andTCEB1 byWestern blottingThe expression of FPR2was increased by treatment with HQSXD-S in comparisonto the blank-control group However HQSXD remarkablydecreased alpha-spectrin PHB and TCEB1 protein levelscompared with blank-control group (Figure 8) Results fromWestern blot manifested the same trend as from proteomicanalysis
Evidence-Based Complementary and Alternative Medicine 7
Spot 1 Spot 2 Spot 3 Spot 4 Spot 5
Spot 6 Spot 7 Spot 8 Spot 9 Spot 10
Blank-control group
HQSXD-treated group
JKSQW-treated group
Blank-control group
HQSXD-treated group
JKSQW-treated group
Figure 6 The ten protein spots of osteoblasts that were treated with and without HQSXD-S Selected regions showed significant differencesin the protein expression profile of osteoblasts among the three groups Upregulated spots are indicated by green circles and downregulatedones by red circles
Table 2 Summary of differentially expressed proteins in osteoblasts treated with HQSXD
Spotnumbera
Proteinscoreb
Matchingpeptides(number)
Theoretical119875119868
cTheoreticalMr (Da)c Target protein Species
1 70 6 927 39299 N-Formyl peptide receptor 2 Rattus norvegicus3 67 7 565 54851 Alpha-spectrin Rattus norvegicus
5 76 6 1008 43712 Heparan sulfate (glucosamine)3-O-sulfotransferase 3A1 Rattus norvegicus
6 173 12 557 29859 Prohibitin Rattus norvegicus
7 67 4 459 12752 Transcription elongationfactor B (SIII) polypeptide 1 Rattus norvegicus
8 70 11 498 127444 Chromosome segregationprotein Rattus norvegicus
9 101 4 691 17386 Nucleoside diphosphatekinase Rattus norvegicus
10 84 12 886 48199 Mast cell carboxypeptidase A Rattus norvegicusaProtein spot number according to Figure 3bProtein scores were based on combined mass and massmass spectra from MALDI-TOFTOF identification MScTheoretical molecular mass (Mr) and isoelectric point (119875
119868) from the NCBInr database
8 Evidence-Based Complementary and Alternative Medicine
724502
995615
861066
1023645
13968251149581
1198660
17947921606823 19980611460637929573 16698152284146
2225096
23712291876991 2119118
00
05
10
15
20
Inte
nsity
(au
)
1000 1500 2000 2500 3000 3500mz
times104
(a)
1023635
687769
832885
643812271987
453127352094621199 772789 979506
174985 322097 876761111971 665734
709626552159385049
86016 295124139982
0
1000
2000
3000
Inte
nsity
(au
)
200 400 600 800 1000 1200mz
(b)
Inte
nsity
(au
)
1037662
254948
467075
989061366041 635224
174962111956 703694338068 886923
80082486005 567816
1012722
200937
287991
0
500
1000
1500
2000
200 400 600 800 1000 1200mz
(c)
Inte
nsity
(au
)
1149574
887334
174919
953820
519998 1105500
633033111931386933
937990
842341486004 595064 785790302933254963
69965
142902
0
1000
2000
3000
200 400 600 800 1000 1200mz
(d)
Inte
nsity
(au
)
1198658
174936
2619201154530
111976440964358918 932243609086
84213585996 325947 741110572984515028287928
400987
228936
200907
0
500
1000
1500
2000
2500
3000
200 400 600 800 1000 1200 1400mz
(e)
Inte
nsity
(au
)
1396813
12406611354732
174968
1070439867233227008111958 286957 1202691
513068417073354995
0
500
1000
1500
2000
2500
200 400 600 800 1000 1200 1400 1600mz
(f)
Figure 7The results of the MALDI-TOFTPF-MS analysis of protein spot 6 (a) Peptide mass fingerprinting and ((b)ndash(f)) peptide fragmentfingerprinting of spot 6
4 Discussion
In this study we found that the proliferative activities ofosteoblasts between HQSXD-S-treated group and JKSQW-control-S-treated group were similar This investigationdemonstrates that HQSXD can significantly facilitate boneformation through increasing the number of osteoblastswhich is beneficial to the treatment of osteoporosis By mea-suring ALP activity HQSXD was first screened for its abilityto induce osteogenesis HQSXD is capable of significantly
promoting osteoblast differentiation as well as increasingosteoblast mineralization
Results of the identification of the selected protein spotsare summarized in Table 2 The molecular weight (Mr) andisoelectric point (119875
119868) of each protein spot shown in Table 2
are theoretical values The eight protein spots were identifiedas (1) N-formyl peptide receptor 2 (FPR2) (2) alpha-spectrin(3) heparan sulfate (glucosamine) 3-O-sulfotransferase 3A1(HS3ST3A1) (4) prohibitin (PHB) (5) transcription elonga-tion factor B (SIII) polypeptide 1 (TCEB1) (6) chromosome
Evidence-Based Complementary and Alternative Medicine 9
TCEB1
PHB
FPR2
Blank HQSXD JKSQW
120573-actin
Alpha-spectrin
Figure 8 HQSXD-S treatment decreased alpha-spectrin PHBand TCEB1 expression and increased FPR2 expression Cell lysates(50 120583g)were processed forWestern blot analysis Normalization per-formed to120573-actinThe bands shown here were from a representativeexperiment repeated three times
segregation protein (7) nucleoside diphosphate kinase and(8) mast cell carboxypeptidase AWe detected some proteinsrelated to transcription cell proliferation differentiationand apoptosis such as FPR2 prohibitin alpha-spectrinheparan sulfate (glucosamine) 3-O-sulfotransferase 3A1 andtranscription elongation factor B (SIII) polypeptide 1 whichvaried greatly after HQSXD treatment
Our results suggest that HQSXD can upregulate theexpression of FPR2 N-Formyl peptide receptor (FPR) is aG protein-coupled receptor which modulates stromal celldifferentiation [14] and binds to N-formyl peptides suchas N-formyl-methionyl-leucyl-phenylalanine (fMLP) Ourstudy has shown that fMLP enhances the differentiationof MSCs into osteoblasts via an FPR-mediated signalingpathway and results in bone formation [15] The FPR2receptor belongs to the formyl peptide receptor family thatis involved in signaling stem cell adhesion migration andhoming for injured and inflamed tissues awaiting repair thiscould potentially be exploited to direct the stem cells to targetspecific tissue site [16] Therefore the current results implythat treatment with HQSXDmight promote the formation ofosteoblasts by upregulating the expression level of FPR2
We found that HQSXD could inhibit the expression ofalpha-spectrin Alpha-spectrin includes two genes encodingfor alpha-I subunit and alpha-II subunit each of thempresenting its specific cellular expression pattern Alpha-II spectrin deficiency is associated with cell proliferationdefects due to cell cycle arrest in theG1 phase (first gap phase)[17] Spectrin and protein kinase C theta were observed inaggregates during the early stage of apoptosis [18] Hencewe presume that alpha-spectrin is involved in the antiosteo-porotic effect of HQSXD
In this study PHB was downregulated in HQSXD-treated osteoblasts Prohibitin a highly conservative proteinregulates the cohesion of sister chromatids cellular signalingmitochondrial biogenesis [19] cell proliferation differentia-tion apoptosis and gene transcription [20 21] PHB blocksthe transition of cells from G1 phase to S phase of the cellcycle thereby arresting cell proliferation [22 23] PHB hasbeen reported to affect the apoptotic pathways by repressingthe transcriptional activity of E2F1 [24 25] As mentionedabove it is possible that HQSXD induces proliferationdifferentiation and apoptosis partly through downregulatingexpression of PHB in osteoblasts It is likely that PHB mightbe the novel candidate in the new antiosteoporotic drugscreening
The expression of TCEB1 was downregulated in HQSXD-treatment osteoblasts TCEB1 a 13 kDa protein also namedelongin C was originally identified as a member of themammalian transcription factor SIII that increases the rateof transcription by suppressing RNA polymerase II pausing[26] As part of a family of separate complexes containingelongin B and various substrate specificity factors it acts asan E3 ubiquitin ligase [27] Elongin B (ELB 1) and elonginC (ELC1) form a stable complex and that depletion of eithergene product by RNA-mediated interference (RNAi) causespronounced defects in the second meiotic division and arrestof germ cell proliferation in G1 [28] Therefore it is possiblethat that TCEB1 is involved in the antiosteoporotic effect ofHQSXD
Although this study has thrown some light on themechanism of HQSXD action we failed to characterize thewell-identified protein (eg HS3ST3A1) closely involved inosteoporosis HS3ST3A1 a member of the heparan sulfatebiosynthetic enzyme family possesses heparan sulfate glu-cosaminyl 3-O-sulfotransferase activity Depletion of Hs3st-A in enterocytes results in increased intestinal stem cellproliferation and tissue homeostasis loss [29]
5 Conclusions
The results confirm that HQSXD has a beneficial effect onosteoblasts and alters the expression level of some proteinsin osteoblasts The protein expressed by osteoblasts treatedwith HQSXD may be involved in cell proliferation anddifferentiation and other physiological processes and in theregulation of cell activation Further study is needed toinvestigate the effects of major active constituents in HQSXDon protein expression on osteoblast so as to demonstrate theinteraction and synergistic mechanism
Conflict of Interests
All the authors state that they have no conflict of interests todeclare
Authorsrsquo Contribution
Chong-Chong Guo and Li-Hua Zheng contributed equally tothis work
10 Evidence-Based Complementary and Alternative Medicine
Acknowledgments
The authors thank Cheng-Ming Liu for her excellent tech-nical assistance This work was supported by the grantfrom the National Natural Science foundation of China (no81273779)
References
[1] S Khosla and B L Riggs ldquoPathophysiology of age-related boneloss and osteoporosisrdquo Endocrinology andMetabolism Clinics ofNorth America vol 34 no 4 pp 1015ndash1030 2005
[2] P Sambrook andCCooper ldquoOsteoporosisrdquoTheLancet vol 367no 9527 pp 2010ndash2018 2006
[3] M R Allen and D B Burr ldquoThree years of alendronate treat-ment results in similar levels of vertebral microdamage as afterone year of treatmentrdquo Journal of Bone and Mineral Researchvol 22 no 11 pp 1759ndash1765 2007
[4] S Davison and S R Davis ldquoHormone replacement therapycurrent controversiesrdquo Clinical Endocrinology vol 58 no 3 pp249ndash261 2003
[5] W-Y Jiang ldquoTherapeutic wisdom in traditional Chinesemedicine a perspective from modern sciencerdquo Trends in Phar-macological Sciences vol 26 no 11 pp 558ndash563 2005
[6] Z Xu and Z K Zhou ldquoClinical study on the treatment of 36cases of postmenopausal osteoporosis with HuangQiSanXianTangrdquo Guiding Journal of Traditional Chinese Medicine andPharmacy vol 15 no 1 pp 9ndash11 2009
[7] H Zhang W-W Xing Y-S Li et al ldquoEffects of a traditionalChinese herbal preparation on osteoblasts and osteoclastsrdquoMaturitas vol 61 no 4 pp 334ndash339 2008
[8] L Qin T Han Q Zhang et al ldquoAntiosteoporotic chemical con-stituents from Er-Xian Decoction a traditional Chinese herbalformulardquo Journal of Ethnopharmacology vol 118 no 2 pp 271ndash279 2008
[9] M Millikan A Kolasani and H Xu ldquoDetermination and com-parison of mineral elements in traditional Chinese herbalformulae at different decoction times used to improve kidneyfunctionmdashchemometric approachrdquo African Journal of Tradi-tional Complementary and Alternative Medicines vol 8 no 5pp 191ndash197 2011
[10] I R Orriss S E B Taylor and T R Arnett ldquoRat osteoblastculturesrdquoMethods inMolecular Biology vol 816 pp 31ndash41 2012
[11] JM Baek J-Y Kim Y-H Cheon et al ldquoDual effect ofChrysan-themum indicum extract to stimulate osteoblast differentia-tion and inhibit osteoclast formation and resorption in vitrordquoEvidence-Based Complementary and Alternative Medicine vol2014 Article ID 176049 13 pages 2014
[12] A I Idris I R Greig E Bassonga-Landao S H Ralston andR J Vanrsquot Hof ldquoIdentification of novel biphenyl carboxylic acidderivatives as novel antiresorptive agents that do not impairparathyroid hormone-induced bone formationrdquo Endocrinologyvol 150 no 1 pp 5ndash13 2009
[13] M Otani J Tabata T Ueki K Sano and S Inouye ldquoHeat-shock-induced proteins from Myxococcus xanthusrdquo Journal ofBacteriology vol 183 no 21 pp 6282ndash6287 2001
[14] M E Nuttall and J M Gimble ldquoIs there a therapeutic opportu-nity to either prevent or treat osteopenic disorders by inhibitingmarrow adipogenesisrdquo Bone vol 27 no 2 pp 177ndash184 2000
[15] M K Shin Y H Jang H J Yoo et al ldquoN-formyl-methionyl-leucyl-phenylalanine (fMLP) promotes osteoblast differentia-tion via the N-formyl peptide receptor 1-mediated signaling
pathway in humanmesenchymal stem cells from bonemarrowrdquoThe Journal of Biological Chemistry vol 286 no 19 pp 17133ndash17143 2011
[16] A Viswanathan R G Painter N A Lanson Jr and G WangldquoFunctional expression ofN-formyl peptide receptors in humanbone marrow-derived mesenchymal stem cellsrdquo Stem Cells vol25 no 5 pp 1263ndash1269 2007
[17] S Metral B Machnicka S Bigot Y Colin D Dhermy and M-C Lecomte ldquo120572II-spectrin is critical for cell adhesion and cellcyclerdquo Journal of Biological Chemistry vol 284 no 4 pp 2409ndash2418 2009
[18] P M Dubielecka M Grzybek A Kolondra et alldquoAggregation of spectrin and PKCtheta is an early hallmarkof fludarabinemitoxantronedexamethasone-induced apopto-sis in Jurkat T and HL60 cellsrdquo Molecular and CellularBiochemistry vol 339 no 1-2 pp 63ndash77 2010
[19] C Merkwirth and T Langer ldquoProhibitin function withinmitochondria essential roles for cell proliferation and cristaemorphogenesisrdquo Biochimica et Biophysica Acta vol 1793 no 1pp 27ndash32 2009
[20] A L Theiss and S V Sitaraman ldquoThe role and therapeuticpotential of prohibitin in diseaserdquoBiochimica et BiophysicaActavol 1813 no 6 pp 1137ndash1143 2011
[21] V Sanchez-Quiles E Santamarıa V Segura L Sesma J Prietoand F J Corrales ldquoProhibitin deficiency blocks proliferationand induces apoptosis in human hepatoma cells molecularmechanisms and functional implicationsrdquo Proteomics vol 10no 8 pp 1609ndash1620 2010
[22] S Manjeshwar D E Branam M R Lerner D J Brackettand E R Jupe ldquoTumor suppression by the prohibitin gene3rsquountranslated region RNA in human breast cancerrdquo CancerResearch vol 63 no 17 pp 5251ndash5256 2003
[23] S Manjeshwar M R Lerner X-P Zang et al ldquoExpressionof prohibitin 31015840 untranslated region suppressor RNA altersmorphology and inhibits motility of breast cancer cellsrdquo Journalof Molecular Histology vol 35 no 6 pp 639ndash646 2004
[24] M Lacroix R-A Toillon and G Leclercq ldquop53 and breastcancer an updaterdquo Endocrine-Related Cancer vol 13 no 2 pp293ndash325 2006
[25] G Fusaro P Dasgupta S Rastogi B Joshi and S ChellappanldquoProhibitin induces the transcriptional activity of p53 andis exported from the nucleus upon apoptotic signalingrdquo TheJournal of Biological Chemistry vol 278 no 48 pp 47853ndash47861 2003
[26] T Aso W S Lane J W Conaway and R C ConawayldquoElongin (SIII) a multisubunit regulator of elongation by RNApolymerase IIrdquo Science vol 269 no 5229 pp 1439ndash1443 1995
[27] T D CumminsM DMendenhall M N Lowry et al ldquoElonginC is a mediator of Notch4 activity in human renal tubule cellsrdquoBiochimica et Biophysica Acta vol 1814 no 12 pp 1748ndash17572011
[28] Y Sasagawa K Kikuchi K Dazai and A HigashitanildquoCaenorhabditis elegans ElonginBC complex is essential for cellproliferation and chromosome condensation and segregationduring mitosis and meiotic division IIrdquo Chromosome Researchvol 13 no 4 pp 357ndash375 2005
[29] Y Guo Z Li and X Lin ldquoHs3st-A and Hs3st-B regulateintestinal homeostasis in Drosophila adult midgutrdquo CellularSignalling vol 26 no 11 pp 2317ndash2325 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
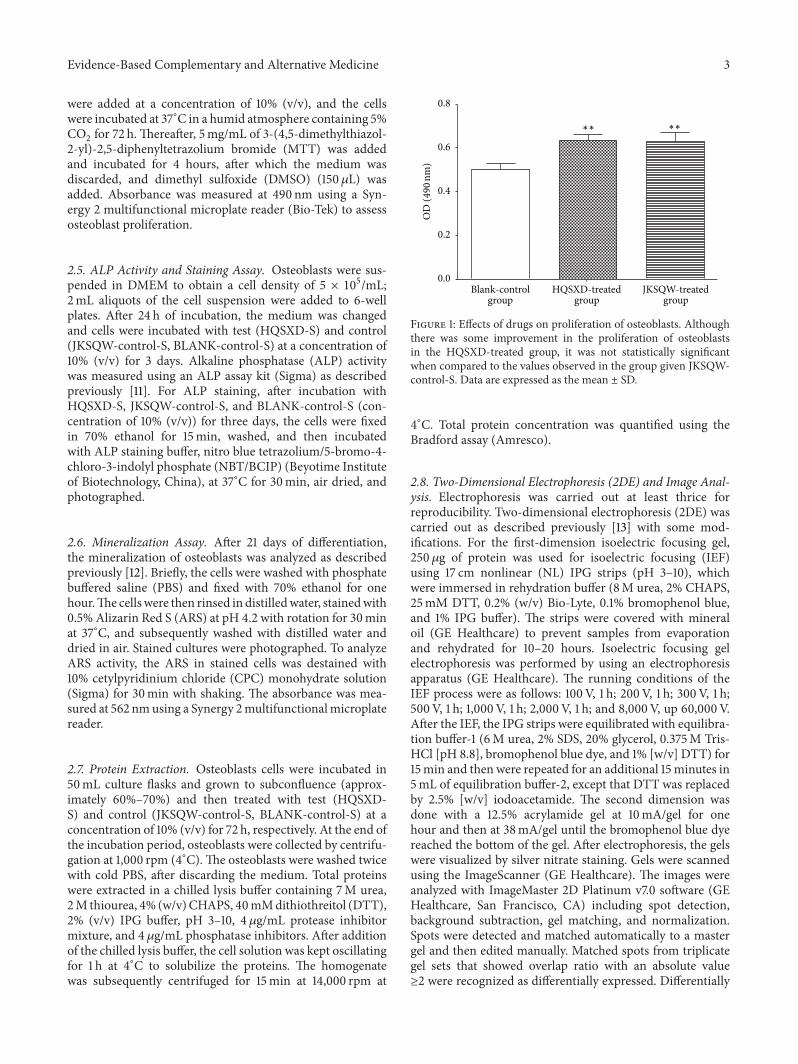
Evidence-Based Complementary and Alternative Medicine 3
were added at a concentration of 10 (vv) and the cellswere incubated at 37∘C in a humid atmosphere containing 5CO2for 72 h Thereafter 5mgmL of 3-(45-dimethylthiazol-
2-yl)-25-diphenyltetrazolium bromide (MTT) was addedand incubated for 4 hours after which the medium wasdiscarded and dimethyl sulfoxide (DMSO) (150120583L) wasadded Absorbance was measured at 490 nm using a Syn-ergy 2 multifunctional microplate reader (Bio-Tek) to assessosteoblast proliferation
25 ALP Activity and Staining Assay Osteoblasts were sus-pended in DMEM to obtain a cell density of 5 times 105mL2mL aliquots of the cell suspension were added to 6-wellplates After 24 h of incubation the medium was changedand cells were incubated with test (HQSXD-S) and control(JKSQW-control-S BLANK-control-S) at a concentration of10 (vv) for 3 days Alkaline phosphatase (ALP) activitywas measured using an ALP assay kit (Sigma) as describedpreviously [11] For ALP staining after incubation withHQSXD-S JKSQW-control-S and BLANK-control-S (con-centration of 10 (vv)) for three days the cells were fixedin 70 ethanol for 15min washed and then incubatedwith ALP staining buffer nitro blue tetrazolium5-bromo-4-chloro-3-indolyl phosphate (NBTBCIP) (Beyotime Instituteof Biotechnology China) at 37∘C for 30min air dried andphotographed
26 Mineralization Assay After 21 days of differentiationthe mineralization of osteoblasts was analyzed as describedpreviously [12] Briefly the cells were washed with phosphatebuffered saline (PBS) and fixed with 70 ethanol for onehourThe cellswere then rinsed in distilledwater stainedwith05 Alizarin Red S (ARS) at pH 42 with rotation for 30minat 37∘C and subsequently washed with distilled water anddried in air Stained cultures were photographed To analyzeARS activity the ARS in stained cells was destained with10 cetylpyridinium chloride (CPC) monohydrate solution(Sigma) for 30min with shaking The absorbance was mea-sured at 562 nmusing a Synergy 2multifunctionalmicroplatereader
27 Protein Extraction Osteoblasts cells were incubated in50mL culture flasks and grown to subconfluence (approx-imately 60ndash70) and then treated with test (HQSXD-S) and control (JKSQW-control-S BLANK-control-S) at aconcentration of 10 (vv) for 72 h respectively At the end ofthe incubation period osteoblasts were collected by centrifu-gation at 1000 rpm (4∘C) The osteoblasts were washed twicewith cold PBS after discarding the medium Total proteinswere extracted in a chilled lysis buffer containing 7M urea2M thiourea 4 (wv)CHAPS 40mMdithiothreitol (DTT)2 (vv) IPG buffer pH 3ndash10 4 120583gmL protease inhibitormixture and 4 120583gmL phosphatase inhibitors After additionof the chilled lysis buffer the cell solution was kept oscillatingfor 1 h at 4∘C to solubilize the proteins The homogenatewas subsequently centrifuged for 15min at 14000 rpm at
08
06
04
02
00
OD
(490
nm)
lowastlowast lowastlowast
JKSQW-treatedgroup
Blank-control HQSXD-treatedgroupgroup
Figure 1 Effects of drugs on proliferation of osteoblasts Althoughthere was some improvement in the proliferation of osteoblastsin the HQSXD-treated group it was not statistically significantwhen compared to the values observed in the group given JKSQW-control-S Data are expressed as the mean plusmn SD
4∘C Total protein concentration was quantified using theBradford assay (Amresco)
28 Two-Dimensional Electrophoresis (2DE) and Image Anal-ysis Electrophoresis was carried out at least thrice forreproducibility Two-dimensional electrophoresis (2DE) wascarried out as described previously [13] with some mod-ifications For the first-dimension isoelectric focusing gel250 120583g of protein was used for isoelectric focusing (IEF)using 17 cm nonlinear (NL) IPG strips (pH 3ndash10) whichwere immersed in rehydration buffer (8M urea 2 CHAPS25mM DTT 02 (wv) Bio-Lyte 01 bromophenol blueand 1 IPG buffer) The strips were covered with mineraloil (GE Healthcare) to prevent samples from evaporationand rehydrated for 10ndash20 hours Isoelectric focusing gelelectrophoresis was performed by using an electrophoresisapparatus (GE Healthcare) The running conditions of theIEF process were as follows 100V 1 h 200V 1 h 300V 1 h500V 1 h 1000V 1 h 2000V 1 h and 8000V up 60000VAfter the IEF the IPG strips were equilibrated with equilibra-tion buffer-1 (6M urea 2 SDS 20 glycerol 0375M Tris-HCl [pH 88] bromophenol blue dye and 1 [wv] DTT) for15min and then were repeated for an additional 15 minutes in5mL of equilibration buffer-2 except that DTT was replacedby 25 [wv] iodoacetamide The second dimension wasdone with a 125 acrylamide gel at 10mAgel for onehour and then at 38mAgel until the bromophenol blue dyereached the bottom of the gel After electrophoresis the gelswere visualized by silver nitrate staining Gels were scannedusing the ImageScanner (GE Healthcare) The images wereanalyzed with ImageMaster 2D Platinum v70 software (GEHealthcare San Francisco CA) including spot detectionbackground subtraction gel matching and normalizationSpots were detected and matched automatically to a mastergel and then edited manually Matched spots from triplicategel sets that showed overlap ratio with an absolute valuege2 were recognized as differentially expressed Differentially
4 Evidence-Based Complementary and Alternative Medicine
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
ALP staining
(a)
025
020
015
010
005
000
ALP
(Ug
prot
)
lowastlowast
lowastlowast
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
(b)
Figure 2 HQSXD-S enhances primary osteoblast differentiation (a) Primary osteoblasts were treated with various drug-treated rat serumsfor 3 days ALP-positive cells were stained with ALP solution (b) Effect of HQSXD-S on alkaline phosphatase activity in primary cultureosteoblasts Data are means plusmn SD of six replicates lowastlowast119901 lt 001 versus control
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
ARS staining
(a)
025
020
015
010
005
000
ARS
activ
ity (562
nm)
lowastlowastlowastlowastlowast
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
(b)
Figure 3 HQSXD-S promotes bone mineralization during osteoblastogenesis (a) Calcium deposits stained with Alizarin Red solution seenin primary osteoblasts treated with various drug-treated rat serums for 21 days (b) Stained calcium deposits were destained with 10 CPCbuffer to measure the level of staining Data are means plusmn SD of six replicates lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001 versus control
abundant spots were selected for mass spectrometry (MS)analysis
29 Mass Spectrometry and Database Search Protein spotswere excised from the 2DE gels using a pipette tip Gel pieceswere destained in a solution of 15mMpotassium ferricyanideand 50mM sodium thiosulfate (1 1) washed with deionizedwater and dehydrated in 100 acetonitrile (ACN) Sampleswere rehydrated for digestion with trypsin (125mgmL) at4∘C for 30min Excess trypsin solution was replaced with25mM ammonium bicarbonateThe samples were incubatedovernight at 37∘C Peptides were then extracted twice with50 ACN5 TFA followed by 100 ACN for 15min eachAfter drying the peptide extracts were desalted with ZipTipPipette Tips (Millipore) Mass spectrometry was done usingan ultraflex III MALDI-TOFTOF-MS (Bruker) with a highvoltage of 20 kV and spectra were externally calibrated
using the peptide standard Maker Protein identification wasdetermined by matching the peptide mass fingerprinting(PMF) and MALDI-TOFTOF-MS results via MASCOT(version 22 Matrix Science) against NCBInr database withBioTools software Database searches were performed usingthe following parameters taxonomy rice enzyme trypsinand one missed cleavage allowed Carbamidomethylationwas selected as a fixed modification and the oxidation wasallowed as a variable PMF tolerance set to 100 ppm andMSMS tolerance set to 07Da A protein was regarded asidentified if the MASCOT protein score was above the 5significance threshold for the database (score gt64)
210 Western Blotting Analysis To verify the results of 2DEof the identified proteins we randomly chose four proteinsfor Western blot FPR2 TCEB1 PHB and alpha-spectrinCytosolic extractswere prepared fromcells and the protein in
Evidence-Based Complementary and Alternative Medicine 5
50
Mr (
kDa)
37
25
20
14
pH 3 10
Blank-control group
(a)
pH 3 10
HQSXD-treated group
50
Mr (
kDa)
37
25
20
14
(b)
pH 3 10
JKSQW-treated group
50
Mr (
kDa)
37
25
20
14
(c)
Figure 4 Representative silver nitrate stained gels showing two-dimensional electrophoresis protein profiles of (a) blank-control group (b)HQSXD-treated group and (c) JKSQW-treated group Molecular weight (MW kDa) and isoelectric point (119875
119868
) are indicated along the 119910- and119909-axes respectively
the supernatant was quantified using the BCA protein assaykit (Beyotime Institute of Biotechnology China) A sample(50120583g) was electrophoresed on a 10 SDS-polyacrylamidegel and subsequently transferred onto a PVDF membrane(Millipore) After blocking with 5 nonfat dry milk themembranes were incubated with anti-FPR2 (M-73 sc-66901Santa Cruz Biotechnology) anti-alpha-spectrin (C-11 sc-46696 Santa Cruz Biotechnology) anti-TCEB1 (ProteinTechGroup Inc China) and anti-PHB (ProteinTech GroupInc China) The bound antibodies were detected using ahorseradish peroxidase- (HRP-) conjugated secondary anti-body and visualized by an enhanced chemiluminescencedetection system followed by quantification using the ImageJ2x
211 Statistical Analysis Data are presented as mean plusmn SD oftriplicate samples Comparisons were performed using one-way analysis of variance (ANOVA) followed byDunnettrsquos testand the difference was considered statistically significant if119901 lt 005
3 Results
31 Effects of HQSXD-Treated Rat Serum on Proliferation ofOsteoblasts The proliferation of primary osteoblasts showedan upward trend compared to that of blank control Howeverthere was no significant difference detected in the prolifer-ation between HQSXD-treated and JKSQW-treated groups
6 Evidence-Based Complementary and Alternative Medicine
Table 1 Effects of drugs on proliferation of osteoblasts
Groups 119873Oral administration
dose (gsdotkgminus1)Serum additivevolume () OD value
Blank-control 5 mdash 10 050 plusmn 003HQSXD-treated 5 14 10 0634 plusmn 0028lowastlowast
JKSQW-treated 5 0520 10 063 plusmn 004lowastlowast
Note lowastlowastp lt 001 compared with BLANK-control-S
Similar changes in proliferation between JKSQW-treatedgroup andHQSXD-treated group indicated that HQSXDhadan influence on osteoblasts (Table 1 Figure 1)
32 HQSXD-Treated Rat Serum Enhances Primary OsteoblastDifferentiation ALP is an important biochemical marker ofdifferentiated osteoblasts and the effects of drug on ALPactivities in osteoblasts were first determined Results ofALP staining showed that HQSXD-S and JKSQW-control-S stimulated osteoblast differentiation (Figure 2(a)) Thecells cultured with HQSXD-S showed a significantly higherALP activity than that cultured with BLANK-control-S(Figure 2(b)) ARS staining in osteoblasts was assessed after21 days of incubation to examine whether HQSXD enhancedbonemineralization during osteoblastogenesis In osteoblaststreated with BLANK-control-S the calcium deposition in themineralized matrix was minimal The proportional areas ofAlizarin Red-positive staining in the HQSXD-treated groupand JKSQW-treated group were higher than that in theBlank-control group (Figure 3(a)) As shown in Figure 3(b)the level and intensity of ARS staining indicated the extentof mineralization which increased upon treatment withHQSXD-S
33 Protein Expression Profile in HQSXD-Treated andHQSXD-Untreated Osteoblasts Two-dimensional electro-phoresis and gel sliver nitrate staining were conducted tofurther investigate the differential protein expression betweenHQSXD-S-treated and HQSXD-S-untreated osteoblastsAfter optimization of the 2DE gels with representative2DE gel images shown in Figure 4 approximately 938 plusmn 26875 plusmn 34 and 904 plusmn 22 protein spots were detected in blankprotein sample HQSXD protein sample and JKSQWproteinsample respectively During analysis with ImageMaster 2DPlatinum spots with an overlap ratio absolute value ge2were recognized as differentially expressed Thirty-eightprotein spots were found to be significantly regulated amongthree groups of which 15 spots were downregulated and23 spots upregulated Ten of these 38 spots exhibited amore than twofold increase or decrease in abundance asobserved in all replicate gels These 10 regulated proteinswere indicated by the circle in Figure 5 and the selectedregions that showed significant differences in proteinexpression profile of osteoblasts among three groups wereshown in Figure 6 All of them were excised from thegels for further identification by MALDI-TOFTOF-MSanalysis
50
Mr (
kDa)
37
25
20
14
pH 3 10
87
3
6
5
2 10
4
1
9
Figure 5 Representative 2DE gel image Ten spots that were statis-tically significant (119901 lt 005) are shown in the map and indicatedby numbers All were cut from the gels for further identification byMALDI-TOFTOF-MS analysis outlined in Table 2
34 Identification of the Differentially Expressed ProteinsProteins were identified by MALDI-TOFTOF-MS Ten pep-tide mass fingerprints (PMFs) and 50 peptide fragmentfingerprints (PFF) were successfully obtained A selectedPMF of protein spot 6 is displayed in Figure 7(a) and theTOFTOF analysis is shown in Figures 7(b)ndash7(f) All PMFswere evaluated with the Mascot software in NCBInr databaseto identify the protein spots The result had high confidenceif the protein was ranked as the best hit with a significantscore and high sequence coverage Finally we identified eightproteins in these spots Properties of the identification of eightselected protein spots are summarized in Table 2
35 Effect of HQSXD on the Expression of Proteins That Regu-late Antiosteoporotic Activity To further investigate the influ-ence of HQSXD on the expression of antiosteoporotic pro-teins we examined the expression of FPR2 alpha-spectrinPHB andTCEB1 byWestern blottingThe expression of FPR2was increased by treatment with HQSXD-S in comparisonto the blank-control group However HQSXD remarkablydecreased alpha-spectrin PHB and TCEB1 protein levelscompared with blank-control group (Figure 8) Results fromWestern blot manifested the same trend as from proteomicanalysis
Evidence-Based Complementary and Alternative Medicine 7
Spot 1 Spot 2 Spot 3 Spot 4 Spot 5
Spot 6 Spot 7 Spot 8 Spot 9 Spot 10
Blank-control group
HQSXD-treated group
JKSQW-treated group
Blank-control group
HQSXD-treated group
JKSQW-treated group
Figure 6 The ten protein spots of osteoblasts that were treated with and without HQSXD-S Selected regions showed significant differencesin the protein expression profile of osteoblasts among the three groups Upregulated spots are indicated by green circles and downregulatedones by red circles
Table 2 Summary of differentially expressed proteins in osteoblasts treated with HQSXD
Spotnumbera
Proteinscoreb
Matchingpeptides(number)
Theoretical119875119868
cTheoreticalMr (Da)c Target protein Species
1 70 6 927 39299 N-Formyl peptide receptor 2 Rattus norvegicus3 67 7 565 54851 Alpha-spectrin Rattus norvegicus
5 76 6 1008 43712 Heparan sulfate (glucosamine)3-O-sulfotransferase 3A1 Rattus norvegicus
6 173 12 557 29859 Prohibitin Rattus norvegicus
7 67 4 459 12752 Transcription elongationfactor B (SIII) polypeptide 1 Rattus norvegicus
8 70 11 498 127444 Chromosome segregationprotein Rattus norvegicus
9 101 4 691 17386 Nucleoside diphosphatekinase Rattus norvegicus
10 84 12 886 48199 Mast cell carboxypeptidase A Rattus norvegicusaProtein spot number according to Figure 3bProtein scores were based on combined mass and massmass spectra from MALDI-TOFTOF identification MScTheoretical molecular mass (Mr) and isoelectric point (119875
119868) from the NCBInr database
8 Evidence-Based Complementary and Alternative Medicine
724502
995615
861066
1023645
13968251149581
1198660
17947921606823 19980611460637929573 16698152284146
2225096
23712291876991 2119118
00
05
10
15
20
Inte
nsity
(au
)
1000 1500 2000 2500 3000 3500mz
times104
(a)
1023635
687769
832885
643812271987
453127352094621199 772789 979506
174985 322097 876761111971 665734
709626552159385049
86016 295124139982
0
1000
2000
3000
Inte
nsity
(au
)
200 400 600 800 1000 1200mz
(b)
Inte
nsity
(au
)
1037662
254948
467075
989061366041 635224
174962111956 703694338068 886923
80082486005 567816
1012722
200937
287991
0
500
1000
1500
2000
200 400 600 800 1000 1200mz
(c)
Inte
nsity
(au
)
1149574
887334
174919
953820
519998 1105500
633033111931386933
937990
842341486004 595064 785790302933254963
69965
142902
0
1000
2000
3000
200 400 600 800 1000 1200mz
(d)
Inte
nsity
(au
)
1198658
174936
2619201154530
111976440964358918 932243609086
84213585996 325947 741110572984515028287928
400987
228936
200907
0
500
1000
1500
2000
2500
3000
200 400 600 800 1000 1200 1400mz
(e)
Inte
nsity
(au
)
1396813
12406611354732
174968
1070439867233227008111958 286957 1202691
513068417073354995
0
500
1000
1500
2000
2500
200 400 600 800 1000 1200 1400 1600mz
(f)
Figure 7The results of the MALDI-TOFTPF-MS analysis of protein spot 6 (a) Peptide mass fingerprinting and ((b)ndash(f)) peptide fragmentfingerprinting of spot 6
4 Discussion
In this study we found that the proliferative activities ofosteoblasts between HQSXD-S-treated group and JKSQW-control-S-treated group were similar This investigationdemonstrates that HQSXD can significantly facilitate boneformation through increasing the number of osteoblastswhich is beneficial to the treatment of osteoporosis By mea-suring ALP activity HQSXD was first screened for its abilityto induce osteogenesis HQSXD is capable of significantly
promoting osteoblast differentiation as well as increasingosteoblast mineralization
Results of the identification of the selected protein spotsare summarized in Table 2 The molecular weight (Mr) andisoelectric point (119875
119868) of each protein spot shown in Table 2
are theoretical values The eight protein spots were identifiedas (1) N-formyl peptide receptor 2 (FPR2) (2) alpha-spectrin(3) heparan sulfate (glucosamine) 3-O-sulfotransferase 3A1(HS3ST3A1) (4) prohibitin (PHB) (5) transcription elonga-tion factor B (SIII) polypeptide 1 (TCEB1) (6) chromosome
Evidence-Based Complementary and Alternative Medicine 9
TCEB1
PHB
FPR2
Blank HQSXD JKSQW
120573-actin
Alpha-spectrin
Figure 8 HQSXD-S treatment decreased alpha-spectrin PHBand TCEB1 expression and increased FPR2 expression Cell lysates(50 120583g)were processed forWestern blot analysis Normalization per-formed to120573-actinThe bands shown here were from a representativeexperiment repeated three times
segregation protein (7) nucleoside diphosphate kinase and(8) mast cell carboxypeptidase AWe detected some proteinsrelated to transcription cell proliferation differentiationand apoptosis such as FPR2 prohibitin alpha-spectrinheparan sulfate (glucosamine) 3-O-sulfotransferase 3A1 andtranscription elongation factor B (SIII) polypeptide 1 whichvaried greatly after HQSXD treatment
Our results suggest that HQSXD can upregulate theexpression of FPR2 N-Formyl peptide receptor (FPR) is aG protein-coupled receptor which modulates stromal celldifferentiation [14] and binds to N-formyl peptides suchas N-formyl-methionyl-leucyl-phenylalanine (fMLP) Ourstudy has shown that fMLP enhances the differentiationof MSCs into osteoblasts via an FPR-mediated signalingpathway and results in bone formation [15] The FPR2receptor belongs to the formyl peptide receptor family thatis involved in signaling stem cell adhesion migration andhoming for injured and inflamed tissues awaiting repair thiscould potentially be exploited to direct the stem cells to targetspecific tissue site [16] Therefore the current results implythat treatment with HQSXDmight promote the formation ofosteoblasts by upregulating the expression level of FPR2
We found that HQSXD could inhibit the expression ofalpha-spectrin Alpha-spectrin includes two genes encodingfor alpha-I subunit and alpha-II subunit each of thempresenting its specific cellular expression pattern Alpha-II spectrin deficiency is associated with cell proliferationdefects due to cell cycle arrest in theG1 phase (first gap phase)[17] Spectrin and protein kinase C theta were observed inaggregates during the early stage of apoptosis [18] Hencewe presume that alpha-spectrin is involved in the antiosteo-porotic effect of HQSXD
In this study PHB was downregulated in HQSXD-treated osteoblasts Prohibitin a highly conservative proteinregulates the cohesion of sister chromatids cellular signalingmitochondrial biogenesis [19] cell proliferation differentia-tion apoptosis and gene transcription [20 21] PHB blocksthe transition of cells from G1 phase to S phase of the cellcycle thereby arresting cell proliferation [22 23] PHB hasbeen reported to affect the apoptotic pathways by repressingthe transcriptional activity of E2F1 [24 25] As mentionedabove it is possible that HQSXD induces proliferationdifferentiation and apoptosis partly through downregulatingexpression of PHB in osteoblasts It is likely that PHB mightbe the novel candidate in the new antiosteoporotic drugscreening
The expression of TCEB1 was downregulated in HQSXD-treatment osteoblasts TCEB1 a 13 kDa protein also namedelongin C was originally identified as a member of themammalian transcription factor SIII that increases the rateof transcription by suppressing RNA polymerase II pausing[26] As part of a family of separate complexes containingelongin B and various substrate specificity factors it acts asan E3 ubiquitin ligase [27] Elongin B (ELB 1) and elonginC (ELC1) form a stable complex and that depletion of eithergene product by RNA-mediated interference (RNAi) causespronounced defects in the second meiotic division and arrestof germ cell proliferation in G1 [28] Therefore it is possiblethat that TCEB1 is involved in the antiosteoporotic effect ofHQSXD
Although this study has thrown some light on themechanism of HQSXD action we failed to characterize thewell-identified protein (eg HS3ST3A1) closely involved inosteoporosis HS3ST3A1 a member of the heparan sulfatebiosynthetic enzyme family possesses heparan sulfate glu-cosaminyl 3-O-sulfotransferase activity Depletion of Hs3st-A in enterocytes results in increased intestinal stem cellproliferation and tissue homeostasis loss [29]
5 Conclusions
The results confirm that HQSXD has a beneficial effect onosteoblasts and alters the expression level of some proteinsin osteoblasts The protein expressed by osteoblasts treatedwith HQSXD may be involved in cell proliferation anddifferentiation and other physiological processes and in theregulation of cell activation Further study is needed toinvestigate the effects of major active constituents in HQSXDon protein expression on osteoblast so as to demonstrate theinteraction and synergistic mechanism
Conflict of Interests
All the authors state that they have no conflict of interests todeclare
Authorsrsquo Contribution
Chong-Chong Guo and Li-Hua Zheng contributed equally tothis work
10 Evidence-Based Complementary and Alternative Medicine
Acknowledgments
The authors thank Cheng-Ming Liu for her excellent tech-nical assistance This work was supported by the grantfrom the National Natural Science foundation of China (no81273779)
References
[1] S Khosla and B L Riggs ldquoPathophysiology of age-related boneloss and osteoporosisrdquo Endocrinology andMetabolism Clinics ofNorth America vol 34 no 4 pp 1015ndash1030 2005
[2] P Sambrook andCCooper ldquoOsteoporosisrdquoTheLancet vol 367no 9527 pp 2010ndash2018 2006
[3] M R Allen and D B Burr ldquoThree years of alendronate treat-ment results in similar levels of vertebral microdamage as afterone year of treatmentrdquo Journal of Bone and Mineral Researchvol 22 no 11 pp 1759ndash1765 2007
[4] S Davison and S R Davis ldquoHormone replacement therapycurrent controversiesrdquo Clinical Endocrinology vol 58 no 3 pp249ndash261 2003
[5] W-Y Jiang ldquoTherapeutic wisdom in traditional Chinesemedicine a perspective from modern sciencerdquo Trends in Phar-macological Sciences vol 26 no 11 pp 558ndash563 2005
[6] Z Xu and Z K Zhou ldquoClinical study on the treatment of 36cases of postmenopausal osteoporosis with HuangQiSanXianTangrdquo Guiding Journal of Traditional Chinese Medicine andPharmacy vol 15 no 1 pp 9ndash11 2009
[7] H Zhang W-W Xing Y-S Li et al ldquoEffects of a traditionalChinese herbal preparation on osteoblasts and osteoclastsrdquoMaturitas vol 61 no 4 pp 334ndash339 2008
[8] L Qin T Han Q Zhang et al ldquoAntiosteoporotic chemical con-stituents from Er-Xian Decoction a traditional Chinese herbalformulardquo Journal of Ethnopharmacology vol 118 no 2 pp 271ndash279 2008
[9] M Millikan A Kolasani and H Xu ldquoDetermination and com-parison of mineral elements in traditional Chinese herbalformulae at different decoction times used to improve kidneyfunctionmdashchemometric approachrdquo African Journal of Tradi-tional Complementary and Alternative Medicines vol 8 no 5pp 191ndash197 2011
[10] I R Orriss S E B Taylor and T R Arnett ldquoRat osteoblastculturesrdquoMethods inMolecular Biology vol 816 pp 31ndash41 2012
[11] JM Baek J-Y Kim Y-H Cheon et al ldquoDual effect ofChrysan-themum indicum extract to stimulate osteoblast differentia-tion and inhibit osteoclast formation and resorption in vitrordquoEvidence-Based Complementary and Alternative Medicine vol2014 Article ID 176049 13 pages 2014
[12] A I Idris I R Greig E Bassonga-Landao S H Ralston andR J Vanrsquot Hof ldquoIdentification of novel biphenyl carboxylic acidderivatives as novel antiresorptive agents that do not impairparathyroid hormone-induced bone formationrdquo Endocrinologyvol 150 no 1 pp 5ndash13 2009
[13] M Otani J Tabata T Ueki K Sano and S Inouye ldquoHeat-shock-induced proteins from Myxococcus xanthusrdquo Journal ofBacteriology vol 183 no 21 pp 6282ndash6287 2001
[14] M E Nuttall and J M Gimble ldquoIs there a therapeutic opportu-nity to either prevent or treat osteopenic disorders by inhibitingmarrow adipogenesisrdquo Bone vol 27 no 2 pp 177ndash184 2000
[15] M K Shin Y H Jang H J Yoo et al ldquoN-formyl-methionyl-leucyl-phenylalanine (fMLP) promotes osteoblast differentia-tion via the N-formyl peptide receptor 1-mediated signaling
pathway in humanmesenchymal stem cells from bonemarrowrdquoThe Journal of Biological Chemistry vol 286 no 19 pp 17133ndash17143 2011
[16] A Viswanathan R G Painter N A Lanson Jr and G WangldquoFunctional expression ofN-formyl peptide receptors in humanbone marrow-derived mesenchymal stem cellsrdquo Stem Cells vol25 no 5 pp 1263ndash1269 2007
[17] S Metral B Machnicka S Bigot Y Colin D Dhermy and M-C Lecomte ldquo120572II-spectrin is critical for cell adhesion and cellcyclerdquo Journal of Biological Chemistry vol 284 no 4 pp 2409ndash2418 2009
[18] P M Dubielecka M Grzybek A Kolondra et alldquoAggregation of spectrin and PKCtheta is an early hallmarkof fludarabinemitoxantronedexamethasone-induced apopto-sis in Jurkat T and HL60 cellsrdquo Molecular and CellularBiochemistry vol 339 no 1-2 pp 63ndash77 2010
[19] C Merkwirth and T Langer ldquoProhibitin function withinmitochondria essential roles for cell proliferation and cristaemorphogenesisrdquo Biochimica et Biophysica Acta vol 1793 no 1pp 27ndash32 2009
[20] A L Theiss and S V Sitaraman ldquoThe role and therapeuticpotential of prohibitin in diseaserdquoBiochimica et BiophysicaActavol 1813 no 6 pp 1137ndash1143 2011
[21] V Sanchez-Quiles E Santamarıa V Segura L Sesma J Prietoand F J Corrales ldquoProhibitin deficiency blocks proliferationand induces apoptosis in human hepatoma cells molecularmechanisms and functional implicationsrdquo Proteomics vol 10no 8 pp 1609ndash1620 2010
[22] S Manjeshwar D E Branam M R Lerner D J Brackettand E R Jupe ldquoTumor suppression by the prohibitin gene3rsquountranslated region RNA in human breast cancerrdquo CancerResearch vol 63 no 17 pp 5251ndash5256 2003
[23] S Manjeshwar M R Lerner X-P Zang et al ldquoExpressionof prohibitin 31015840 untranslated region suppressor RNA altersmorphology and inhibits motility of breast cancer cellsrdquo Journalof Molecular Histology vol 35 no 6 pp 639ndash646 2004
[24] M Lacroix R-A Toillon and G Leclercq ldquop53 and breastcancer an updaterdquo Endocrine-Related Cancer vol 13 no 2 pp293ndash325 2006
[25] G Fusaro P Dasgupta S Rastogi B Joshi and S ChellappanldquoProhibitin induces the transcriptional activity of p53 andis exported from the nucleus upon apoptotic signalingrdquo TheJournal of Biological Chemistry vol 278 no 48 pp 47853ndash47861 2003
[26] T Aso W S Lane J W Conaway and R C ConawayldquoElongin (SIII) a multisubunit regulator of elongation by RNApolymerase IIrdquo Science vol 269 no 5229 pp 1439ndash1443 1995
[27] T D CumminsM DMendenhall M N Lowry et al ldquoElonginC is a mediator of Notch4 activity in human renal tubule cellsrdquoBiochimica et Biophysica Acta vol 1814 no 12 pp 1748ndash17572011
[28] Y Sasagawa K Kikuchi K Dazai and A HigashitanildquoCaenorhabditis elegans ElonginBC complex is essential for cellproliferation and chromosome condensation and segregationduring mitosis and meiotic division IIrdquo Chromosome Researchvol 13 no 4 pp 357ndash375 2005
[29] Y Guo Z Li and X Lin ldquoHs3st-A and Hs3st-B regulateintestinal homeostasis in Drosophila adult midgutrdquo CellularSignalling vol 26 no 11 pp 2317ndash2325 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 Evidence-Based Complementary and Alternative Medicine
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
ALP staining
(a)
025
020
015
010
005
000
ALP
(Ug
prot
)
lowastlowast
lowastlowast
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
(b)
Figure 2 HQSXD-S enhances primary osteoblast differentiation (a) Primary osteoblasts were treated with various drug-treated rat serumsfor 3 days ALP-positive cells were stained with ALP solution (b) Effect of HQSXD-S on alkaline phosphatase activity in primary cultureosteoblasts Data are means plusmn SD of six replicates lowastlowast119901 lt 001 versus control
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
ARS staining
(a)
025
020
015
010
005
000
ARS
activ
ity (562
nm)
lowastlowastlowastlowastlowast
JKSQW-treatedgroup
HQSXD-treatedgroup
Blank-controlgroup
(b)
Figure 3 HQSXD-S promotes bone mineralization during osteoblastogenesis (a) Calcium deposits stained with Alizarin Red solution seenin primary osteoblasts treated with various drug-treated rat serums for 21 days (b) Stained calcium deposits were destained with 10 CPCbuffer to measure the level of staining Data are means plusmn SD of six replicates lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001 versus control
abundant spots were selected for mass spectrometry (MS)analysis
29 Mass Spectrometry and Database Search Protein spotswere excised from the 2DE gels using a pipette tip Gel pieceswere destained in a solution of 15mMpotassium ferricyanideand 50mM sodium thiosulfate (1 1) washed with deionizedwater and dehydrated in 100 acetonitrile (ACN) Sampleswere rehydrated for digestion with trypsin (125mgmL) at4∘C for 30min Excess trypsin solution was replaced with25mM ammonium bicarbonateThe samples were incubatedovernight at 37∘C Peptides were then extracted twice with50 ACN5 TFA followed by 100 ACN for 15min eachAfter drying the peptide extracts were desalted with ZipTipPipette Tips (Millipore) Mass spectrometry was done usingan ultraflex III MALDI-TOFTOF-MS (Bruker) with a highvoltage of 20 kV and spectra were externally calibrated
using the peptide standard Maker Protein identification wasdetermined by matching the peptide mass fingerprinting(PMF) and MALDI-TOFTOF-MS results via MASCOT(version 22 Matrix Science) against NCBInr database withBioTools software Database searches were performed usingthe following parameters taxonomy rice enzyme trypsinand one missed cleavage allowed Carbamidomethylationwas selected as a fixed modification and the oxidation wasallowed as a variable PMF tolerance set to 100 ppm andMSMS tolerance set to 07Da A protein was regarded asidentified if the MASCOT protein score was above the 5significance threshold for the database (score gt64)
210 Western Blotting Analysis To verify the results of 2DEof the identified proteins we randomly chose four proteinsfor Western blot FPR2 TCEB1 PHB and alpha-spectrinCytosolic extractswere prepared fromcells and the protein in
Evidence-Based Complementary and Alternative Medicine 5
50
Mr (
kDa)
37
25
20
14
pH 3 10
Blank-control group
(a)
pH 3 10
HQSXD-treated group
50
Mr (
kDa)
37
25
20
14
(b)
pH 3 10
JKSQW-treated group
50
Mr (
kDa)
37
25
20
14
(c)
Figure 4 Representative silver nitrate stained gels showing two-dimensional electrophoresis protein profiles of (a) blank-control group (b)HQSXD-treated group and (c) JKSQW-treated group Molecular weight (MW kDa) and isoelectric point (119875
119868
) are indicated along the 119910- and119909-axes respectively
the supernatant was quantified using the BCA protein assaykit (Beyotime Institute of Biotechnology China) A sample(50120583g) was electrophoresed on a 10 SDS-polyacrylamidegel and subsequently transferred onto a PVDF membrane(Millipore) After blocking with 5 nonfat dry milk themembranes were incubated with anti-FPR2 (M-73 sc-66901Santa Cruz Biotechnology) anti-alpha-spectrin (C-11 sc-46696 Santa Cruz Biotechnology) anti-TCEB1 (ProteinTechGroup Inc China) and anti-PHB (ProteinTech GroupInc China) The bound antibodies were detected using ahorseradish peroxidase- (HRP-) conjugated secondary anti-body and visualized by an enhanced chemiluminescencedetection system followed by quantification using the ImageJ2x
211 Statistical Analysis Data are presented as mean plusmn SD oftriplicate samples Comparisons were performed using one-way analysis of variance (ANOVA) followed byDunnettrsquos testand the difference was considered statistically significant if119901 lt 005
3 Results
31 Effects of HQSXD-Treated Rat Serum on Proliferation ofOsteoblasts The proliferation of primary osteoblasts showedan upward trend compared to that of blank control Howeverthere was no significant difference detected in the prolifer-ation between HQSXD-treated and JKSQW-treated groups
6 Evidence-Based Complementary and Alternative Medicine
Table 1 Effects of drugs on proliferation of osteoblasts
Groups 119873Oral administration
dose (gsdotkgminus1)Serum additivevolume () OD value
Blank-control 5 mdash 10 050 plusmn 003HQSXD-treated 5 14 10 0634 plusmn 0028lowastlowast
JKSQW-treated 5 0520 10 063 plusmn 004lowastlowast
Note lowastlowastp lt 001 compared with BLANK-control-S
Similar changes in proliferation between JKSQW-treatedgroup andHQSXD-treated group indicated that HQSXDhadan influence on osteoblasts (Table 1 Figure 1)
32 HQSXD-Treated Rat Serum Enhances Primary OsteoblastDifferentiation ALP is an important biochemical marker ofdifferentiated osteoblasts and the effects of drug on ALPactivities in osteoblasts were first determined Results ofALP staining showed that HQSXD-S and JKSQW-control-S stimulated osteoblast differentiation (Figure 2(a)) Thecells cultured with HQSXD-S showed a significantly higherALP activity than that cultured with BLANK-control-S(Figure 2(b)) ARS staining in osteoblasts was assessed after21 days of incubation to examine whether HQSXD enhancedbonemineralization during osteoblastogenesis In osteoblaststreated with BLANK-control-S the calcium deposition in themineralized matrix was minimal The proportional areas ofAlizarin Red-positive staining in the HQSXD-treated groupand JKSQW-treated group were higher than that in theBlank-control group (Figure 3(a)) As shown in Figure 3(b)the level and intensity of ARS staining indicated the extentof mineralization which increased upon treatment withHQSXD-S
33 Protein Expression Profile in HQSXD-Treated andHQSXD-Untreated Osteoblasts Two-dimensional electro-phoresis and gel sliver nitrate staining were conducted tofurther investigate the differential protein expression betweenHQSXD-S-treated and HQSXD-S-untreated osteoblastsAfter optimization of the 2DE gels with representative2DE gel images shown in Figure 4 approximately 938 plusmn 26875 plusmn 34 and 904 plusmn 22 protein spots were detected in blankprotein sample HQSXD protein sample and JKSQWproteinsample respectively During analysis with ImageMaster 2DPlatinum spots with an overlap ratio absolute value ge2were recognized as differentially expressed Thirty-eightprotein spots were found to be significantly regulated amongthree groups of which 15 spots were downregulated and23 spots upregulated Ten of these 38 spots exhibited amore than twofold increase or decrease in abundance asobserved in all replicate gels These 10 regulated proteinswere indicated by the circle in Figure 5 and the selectedregions that showed significant differences in proteinexpression profile of osteoblasts among three groups wereshown in Figure 6 All of them were excised from thegels for further identification by MALDI-TOFTOF-MSanalysis
50
Mr (
kDa)
37
25
20
14
pH 3 10
87
3
6
5
2 10
4
1
9
Figure 5 Representative 2DE gel image Ten spots that were statis-tically significant (119901 lt 005) are shown in the map and indicatedby numbers All were cut from the gels for further identification byMALDI-TOFTOF-MS analysis outlined in Table 2
34 Identification of the Differentially Expressed ProteinsProteins were identified by MALDI-TOFTOF-MS Ten pep-tide mass fingerprints (PMFs) and 50 peptide fragmentfingerprints (PFF) were successfully obtained A selectedPMF of protein spot 6 is displayed in Figure 7(a) and theTOFTOF analysis is shown in Figures 7(b)ndash7(f) All PMFswere evaluated with the Mascot software in NCBInr databaseto identify the protein spots The result had high confidenceif the protein was ranked as the best hit with a significantscore and high sequence coverage Finally we identified eightproteins in these spots Properties of the identification of eightselected protein spots are summarized in Table 2
35 Effect of HQSXD on the Expression of Proteins That Regu-late Antiosteoporotic Activity To further investigate the influ-ence of HQSXD on the expression of antiosteoporotic pro-teins we examined the expression of FPR2 alpha-spectrinPHB andTCEB1 byWestern blottingThe expression of FPR2was increased by treatment with HQSXD-S in comparisonto the blank-control group However HQSXD remarkablydecreased alpha-spectrin PHB and TCEB1 protein levelscompared with blank-control group (Figure 8) Results fromWestern blot manifested the same trend as from proteomicanalysis
Evidence-Based Complementary and Alternative Medicine 7
Spot 1 Spot 2 Spot 3 Spot 4 Spot 5
Spot 6 Spot 7 Spot 8 Spot 9 Spot 10
Blank-control group
HQSXD-treated group
JKSQW-treated group
Blank-control group
HQSXD-treated group
JKSQW-treated group
Figure 6 The ten protein spots of osteoblasts that were treated with and without HQSXD-S Selected regions showed significant differencesin the protein expression profile of osteoblasts among the three groups Upregulated spots are indicated by green circles and downregulatedones by red circles
Table 2 Summary of differentially expressed proteins in osteoblasts treated with HQSXD
Spotnumbera
Proteinscoreb
Matchingpeptides(number)
Theoretical119875119868
cTheoreticalMr (Da)c Target protein Species
1 70 6 927 39299 N-Formyl peptide receptor 2 Rattus norvegicus3 67 7 565 54851 Alpha-spectrin Rattus norvegicus
5 76 6 1008 43712 Heparan sulfate (glucosamine)3-O-sulfotransferase 3A1 Rattus norvegicus
6 173 12 557 29859 Prohibitin Rattus norvegicus
7 67 4 459 12752 Transcription elongationfactor B (SIII) polypeptide 1 Rattus norvegicus
8 70 11 498 127444 Chromosome segregationprotein Rattus norvegicus
9 101 4 691 17386 Nucleoside diphosphatekinase Rattus norvegicus
10 84 12 886 48199 Mast cell carboxypeptidase A Rattus norvegicusaProtein spot number according to Figure 3bProtein scores were based on combined mass and massmass spectra from MALDI-TOFTOF identification MScTheoretical molecular mass (Mr) and isoelectric point (119875
119868) from the NCBInr database
8 Evidence-Based Complementary and Alternative Medicine
724502
995615
861066
1023645
13968251149581
1198660
17947921606823 19980611460637929573 16698152284146
2225096
23712291876991 2119118
00
05
10
15
20
Inte
nsity
(au
)
1000 1500 2000 2500 3000 3500mz
times104
(a)
1023635
687769
832885
643812271987
453127352094621199 772789 979506
174985 322097 876761111971 665734
709626552159385049
86016 295124139982
0
1000
2000
3000
Inte
nsity
(au
)
200 400 600 800 1000 1200mz
(b)
Inte
nsity
(au
)
1037662
254948
467075
989061366041 635224
174962111956 703694338068 886923
80082486005 567816
1012722
200937
287991
0
500
1000
1500
2000
200 400 600 800 1000 1200mz
(c)
Inte
nsity
(au
)
1149574
887334
174919
953820
519998 1105500
633033111931386933
937990
842341486004 595064 785790302933254963
69965
142902
0
1000
2000
3000
200 400 600 800 1000 1200mz
(d)
Inte
nsity
(au
)
1198658
174936
2619201154530
111976440964358918 932243609086
84213585996 325947 741110572984515028287928
400987
228936
200907
0
500
1000
1500
2000
2500
3000
200 400 600 800 1000 1200 1400mz
(e)
Inte
nsity
(au
)
1396813
12406611354732
174968
1070439867233227008111958 286957 1202691
513068417073354995
0
500
1000
1500
2000
2500
200 400 600 800 1000 1200 1400 1600mz
(f)
Figure 7The results of the MALDI-TOFTPF-MS analysis of protein spot 6 (a) Peptide mass fingerprinting and ((b)ndash(f)) peptide fragmentfingerprinting of spot 6
4 Discussion
In this study we found that the proliferative activities ofosteoblasts between HQSXD-S-treated group and JKSQW-control-S-treated group were similar This investigationdemonstrates that HQSXD can significantly facilitate boneformation through increasing the number of osteoblastswhich is beneficial to the treatment of osteoporosis By mea-suring ALP activity HQSXD was first screened for its abilityto induce osteogenesis HQSXD is capable of significantly
promoting osteoblast differentiation as well as increasingosteoblast mineralization
Results of the identification of the selected protein spotsare summarized in Table 2 The molecular weight (Mr) andisoelectric point (119875
119868) of each protein spot shown in Table 2
are theoretical values The eight protein spots were identifiedas (1) N-formyl peptide receptor 2 (FPR2) (2) alpha-spectrin(3) heparan sulfate (glucosamine) 3-O-sulfotransferase 3A1(HS3ST3A1) (4) prohibitin (PHB) (5) transcription elonga-tion factor B (SIII) polypeptide 1 (TCEB1) (6) chromosome
Evidence-Based Complementary and Alternative Medicine 9
TCEB1
PHB
FPR2
Blank HQSXD JKSQW
120573-actin
Alpha-spectrin
Figure 8 HQSXD-S treatment decreased alpha-spectrin PHBand TCEB1 expression and increased FPR2 expression Cell lysates(50 120583g)were processed forWestern blot analysis Normalization per-formed to120573-actinThe bands shown here were from a representativeexperiment repeated three times
segregation protein (7) nucleoside diphosphate kinase and(8) mast cell carboxypeptidase AWe detected some proteinsrelated to transcription cell proliferation differentiationand apoptosis such as FPR2 prohibitin alpha-spectrinheparan sulfate (glucosamine) 3-O-sulfotransferase 3A1 andtranscription elongation factor B (SIII) polypeptide 1 whichvaried greatly after HQSXD treatment
Our results suggest that HQSXD can upregulate theexpression of FPR2 N-Formyl peptide receptor (FPR) is aG protein-coupled receptor which modulates stromal celldifferentiation [14] and binds to N-formyl peptides suchas N-formyl-methionyl-leucyl-phenylalanine (fMLP) Ourstudy has shown that fMLP enhances the differentiationof MSCs into osteoblasts via an FPR-mediated signalingpathway and results in bone formation [15] The FPR2receptor belongs to the formyl peptide receptor family thatis involved in signaling stem cell adhesion migration andhoming for injured and inflamed tissues awaiting repair thiscould potentially be exploited to direct the stem cells to targetspecific tissue site [16] Therefore the current results implythat treatment with HQSXDmight promote the formation ofosteoblasts by upregulating the expression level of FPR2
We found that HQSXD could inhibit the expression ofalpha-spectrin Alpha-spectrin includes two genes encodingfor alpha-I subunit and alpha-II subunit each of thempresenting its specific cellular expression pattern Alpha-II spectrin deficiency is associated with cell proliferationdefects due to cell cycle arrest in theG1 phase (first gap phase)[17] Spectrin and protein kinase C theta were observed inaggregates during the early stage of apoptosis [18] Hencewe presume that alpha-spectrin is involved in the antiosteo-porotic effect of HQSXD
In this study PHB was downregulated in HQSXD-treated osteoblasts Prohibitin a highly conservative proteinregulates the cohesion of sister chromatids cellular signalingmitochondrial biogenesis [19] cell proliferation differentia-tion apoptosis and gene transcription [20 21] PHB blocksthe transition of cells from G1 phase to S phase of the cellcycle thereby arresting cell proliferation [22 23] PHB hasbeen reported to affect the apoptotic pathways by repressingthe transcriptional activity of E2F1 [24 25] As mentionedabove it is possible that HQSXD induces proliferationdifferentiation and apoptosis partly through downregulatingexpression of PHB in osteoblasts It is likely that PHB mightbe the novel candidate in the new antiosteoporotic drugscreening
The expression of TCEB1 was downregulated in HQSXD-treatment osteoblasts TCEB1 a 13 kDa protein also namedelongin C was originally identified as a member of themammalian transcription factor SIII that increases the rateof transcription by suppressing RNA polymerase II pausing[26] As part of a family of separate complexes containingelongin B and various substrate specificity factors it acts asan E3 ubiquitin ligase [27] Elongin B (ELB 1) and elonginC (ELC1) form a stable complex and that depletion of eithergene product by RNA-mediated interference (RNAi) causespronounced defects in the second meiotic division and arrestof germ cell proliferation in G1 [28] Therefore it is possiblethat that TCEB1 is involved in the antiosteoporotic effect ofHQSXD
Although this study has thrown some light on themechanism of HQSXD action we failed to characterize thewell-identified protein (eg HS3ST3A1) closely involved inosteoporosis HS3ST3A1 a member of the heparan sulfatebiosynthetic enzyme family possesses heparan sulfate glu-cosaminyl 3-O-sulfotransferase activity Depletion of Hs3st-A in enterocytes results in increased intestinal stem cellproliferation and tissue homeostasis loss [29]
5 Conclusions
The results confirm that HQSXD has a beneficial effect onosteoblasts and alters the expression level of some proteinsin osteoblasts The protein expressed by osteoblasts treatedwith HQSXD may be involved in cell proliferation anddifferentiation and other physiological processes and in theregulation of cell activation Further study is needed toinvestigate the effects of major active constituents in HQSXDon protein expression on osteoblast so as to demonstrate theinteraction and synergistic mechanism
Conflict of Interests
All the authors state that they have no conflict of interests todeclare
Authorsrsquo Contribution
Chong-Chong Guo and Li-Hua Zheng contributed equally tothis work
10 Evidence-Based Complementary and Alternative Medicine
Acknowledgments
The authors thank Cheng-Ming Liu for her excellent tech-nical assistance This work was supported by the grantfrom the National Natural Science foundation of China (no81273779)
References
[1] S Khosla and B L Riggs ldquoPathophysiology of age-related boneloss and osteoporosisrdquo Endocrinology andMetabolism Clinics ofNorth America vol 34 no 4 pp 1015ndash1030 2005
[2] P Sambrook andCCooper ldquoOsteoporosisrdquoTheLancet vol 367no 9527 pp 2010ndash2018 2006
[3] M R Allen and D B Burr ldquoThree years of alendronate treat-ment results in similar levels of vertebral microdamage as afterone year of treatmentrdquo Journal of Bone and Mineral Researchvol 22 no 11 pp 1759ndash1765 2007
[4] S Davison and S R Davis ldquoHormone replacement therapycurrent controversiesrdquo Clinical Endocrinology vol 58 no 3 pp249ndash261 2003
[5] W-Y Jiang ldquoTherapeutic wisdom in traditional Chinesemedicine a perspective from modern sciencerdquo Trends in Phar-macological Sciences vol 26 no 11 pp 558ndash563 2005
[6] Z Xu and Z K Zhou ldquoClinical study on the treatment of 36cases of postmenopausal osteoporosis with HuangQiSanXianTangrdquo Guiding Journal of Traditional Chinese Medicine andPharmacy vol 15 no 1 pp 9ndash11 2009
[7] H Zhang W-W Xing Y-S Li et al ldquoEffects of a traditionalChinese herbal preparation on osteoblasts and osteoclastsrdquoMaturitas vol 61 no 4 pp 334ndash339 2008
[8] L Qin T Han Q Zhang et al ldquoAntiosteoporotic chemical con-stituents from Er-Xian Decoction a traditional Chinese herbalformulardquo Journal of Ethnopharmacology vol 118 no 2 pp 271ndash279 2008
[9] M Millikan A Kolasani and H Xu ldquoDetermination and com-parison of mineral elements in traditional Chinese herbalformulae at different decoction times used to improve kidneyfunctionmdashchemometric approachrdquo African Journal of Tradi-tional Complementary and Alternative Medicines vol 8 no 5pp 191ndash197 2011
[10] I R Orriss S E B Taylor and T R Arnett ldquoRat osteoblastculturesrdquoMethods inMolecular Biology vol 816 pp 31ndash41 2012
[11] JM Baek J-Y Kim Y-H Cheon et al ldquoDual effect ofChrysan-themum indicum extract to stimulate osteoblast differentia-tion and inhibit osteoclast formation and resorption in vitrordquoEvidence-Based Complementary and Alternative Medicine vol2014 Article ID 176049 13 pages 2014
[12] A I Idris I R Greig E Bassonga-Landao S H Ralston andR J Vanrsquot Hof ldquoIdentification of novel biphenyl carboxylic acidderivatives as novel antiresorptive agents that do not impairparathyroid hormone-induced bone formationrdquo Endocrinologyvol 150 no 1 pp 5ndash13 2009
[13] M Otani J Tabata T Ueki K Sano and S Inouye ldquoHeat-shock-induced proteins from Myxococcus xanthusrdquo Journal ofBacteriology vol 183 no 21 pp 6282ndash6287 2001
[14] M E Nuttall and J M Gimble ldquoIs there a therapeutic opportu-nity to either prevent or treat osteopenic disorders by inhibitingmarrow adipogenesisrdquo Bone vol 27 no 2 pp 177ndash184 2000
[15] M K Shin Y H Jang H J Yoo et al ldquoN-formyl-methionyl-leucyl-phenylalanine (fMLP) promotes osteoblast differentia-tion via the N-formyl peptide receptor 1-mediated signaling
pathway in humanmesenchymal stem cells from bonemarrowrdquoThe Journal of Biological Chemistry vol 286 no 19 pp 17133ndash17143 2011
[16] A Viswanathan R G Painter N A Lanson Jr and G WangldquoFunctional expression ofN-formyl peptide receptors in humanbone marrow-derived mesenchymal stem cellsrdquo Stem Cells vol25 no 5 pp 1263ndash1269 2007
[17] S Metral B Machnicka S Bigot Y Colin D Dhermy and M-C Lecomte ldquo120572II-spectrin is critical for cell adhesion and cellcyclerdquo Journal of Biological Chemistry vol 284 no 4 pp 2409ndash2418 2009
[18] P M Dubielecka M Grzybek A Kolondra et alldquoAggregation of spectrin and PKCtheta is an early hallmarkof fludarabinemitoxantronedexamethasone-induced apopto-sis in Jurkat T and HL60 cellsrdquo Molecular and CellularBiochemistry vol 339 no 1-2 pp 63ndash77 2010
[19] C Merkwirth and T Langer ldquoProhibitin function withinmitochondria essential roles for cell proliferation and cristaemorphogenesisrdquo Biochimica et Biophysica Acta vol 1793 no 1pp 27ndash32 2009
[20] A L Theiss and S V Sitaraman ldquoThe role and therapeuticpotential of prohibitin in diseaserdquoBiochimica et BiophysicaActavol 1813 no 6 pp 1137ndash1143 2011
[21] V Sanchez-Quiles E Santamarıa V Segura L Sesma J Prietoand F J Corrales ldquoProhibitin deficiency blocks proliferationand induces apoptosis in human hepatoma cells molecularmechanisms and functional implicationsrdquo Proteomics vol 10no 8 pp 1609ndash1620 2010
[22] S Manjeshwar D E Branam M R Lerner D J Brackettand E R Jupe ldquoTumor suppression by the prohibitin gene3rsquountranslated region RNA in human breast cancerrdquo CancerResearch vol 63 no 17 pp 5251ndash5256 2003
[23] S Manjeshwar M R Lerner X-P Zang et al ldquoExpressionof prohibitin 31015840 untranslated region suppressor RNA altersmorphology and inhibits motility of breast cancer cellsrdquo Journalof Molecular Histology vol 35 no 6 pp 639ndash646 2004
[24] M Lacroix R-A Toillon and G Leclercq ldquop53 and breastcancer an updaterdquo Endocrine-Related Cancer vol 13 no 2 pp293ndash325 2006
[25] G Fusaro P Dasgupta S Rastogi B Joshi and S ChellappanldquoProhibitin induces the transcriptional activity of p53 andis exported from the nucleus upon apoptotic signalingrdquo TheJournal of Biological Chemistry vol 278 no 48 pp 47853ndash47861 2003
[26] T Aso W S Lane J W Conaway and R C ConawayldquoElongin (SIII) a multisubunit regulator of elongation by RNApolymerase IIrdquo Science vol 269 no 5229 pp 1439ndash1443 1995
[27] T D CumminsM DMendenhall M N Lowry et al ldquoElonginC is a mediator of Notch4 activity in human renal tubule cellsrdquoBiochimica et Biophysica Acta vol 1814 no 12 pp 1748ndash17572011
[28] Y Sasagawa K Kikuchi K Dazai and A HigashitanildquoCaenorhabditis elegans ElonginBC complex is essential for cellproliferation and chromosome condensation and segregationduring mitosis and meiotic division IIrdquo Chromosome Researchvol 13 no 4 pp 357ndash375 2005
[29] Y Guo Z Li and X Lin ldquoHs3st-A and Hs3st-B regulateintestinal homeostasis in Drosophila adult midgutrdquo CellularSignalling vol 26 no 11 pp 2317ndash2325 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 5
50
Mr (
kDa)
37
25
20
14
pH 3 10
Blank-control group
(a)
pH 3 10
HQSXD-treated group
50
Mr (
kDa)
37
25
20
14
(b)
pH 3 10
JKSQW-treated group
50
Mr (
kDa)
37
25
20
14
(c)
Figure 4 Representative silver nitrate stained gels showing two-dimensional electrophoresis protein profiles of (a) blank-control group (b)HQSXD-treated group and (c) JKSQW-treated group Molecular weight (MW kDa) and isoelectric point (119875
119868
) are indicated along the 119910- and119909-axes respectively
the supernatant was quantified using the BCA protein assaykit (Beyotime Institute of Biotechnology China) A sample(50120583g) was electrophoresed on a 10 SDS-polyacrylamidegel and subsequently transferred onto a PVDF membrane(Millipore) After blocking with 5 nonfat dry milk themembranes were incubated with anti-FPR2 (M-73 sc-66901Santa Cruz Biotechnology) anti-alpha-spectrin (C-11 sc-46696 Santa Cruz Biotechnology) anti-TCEB1 (ProteinTechGroup Inc China) and anti-PHB (ProteinTech GroupInc China) The bound antibodies were detected using ahorseradish peroxidase- (HRP-) conjugated secondary anti-body and visualized by an enhanced chemiluminescencedetection system followed by quantification using the ImageJ2x
211 Statistical Analysis Data are presented as mean plusmn SD oftriplicate samples Comparisons were performed using one-way analysis of variance (ANOVA) followed byDunnettrsquos testand the difference was considered statistically significant if119901 lt 005
3 Results
31 Effects of HQSXD-Treated Rat Serum on Proliferation ofOsteoblasts The proliferation of primary osteoblasts showedan upward trend compared to that of blank control Howeverthere was no significant difference detected in the prolifer-ation between HQSXD-treated and JKSQW-treated groups
6 Evidence-Based Complementary and Alternative Medicine
Table 1 Effects of drugs on proliferation of osteoblasts
Groups 119873Oral administration
dose (gsdotkgminus1)Serum additivevolume () OD value
Blank-control 5 mdash 10 050 plusmn 003HQSXD-treated 5 14 10 0634 plusmn 0028lowastlowast
JKSQW-treated 5 0520 10 063 plusmn 004lowastlowast
Note lowastlowastp lt 001 compared with BLANK-control-S
Similar changes in proliferation between JKSQW-treatedgroup andHQSXD-treated group indicated that HQSXDhadan influence on osteoblasts (Table 1 Figure 1)
32 HQSXD-Treated Rat Serum Enhances Primary OsteoblastDifferentiation ALP is an important biochemical marker ofdifferentiated osteoblasts and the effects of drug on ALPactivities in osteoblasts were first determined Results ofALP staining showed that HQSXD-S and JKSQW-control-S stimulated osteoblast differentiation (Figure 2(a)) Thecells cultured with HQSXD-S showed a significantly higherALP activity than that cultured with BLANK-control-S(Figure 2(b)) ARS staining in osteoblasts was assessed after21 days of incubation to examine whether HQSXD enhancedbonemineralization during osteoblastogenesis In osteoblaststreated with BLANK-control-S the calcium deposition in themineralized matrix was minimal The proportional areas ofAlizarin Red-positive staining in the HQSXD-treated groupand JKSQW-treated group were higher than that in theBlank-control group (Figure 3(a)) As shown in Figure 3(b)the level and intensity of ARS staining indicated the extentof mineralization which increased upon treatment withHQSXD-S
33 Protein Expression Profile in HQSXD-Treated andHQSXD-Untreated Osteoblasts Two-dimensional electro-phoresis and gel sliver nitrate staining were conducted tofurther investigate the differential protein expression betweenHQSXD-S-treated and HQSXD-S-untreated osteoblastsAfter optimization of the 2DE gels with representative2DE gel images shown in Figure 4 approximately 938 plusmn 26875 plusmn 34 and 904 plusmn 22 protein spots were detected in blankprotein sample HQSXD protein sample and JKSQWproteinsample respectively During analysis with ImageMaster 2DPlatinum spots with an overlap ratio absolute value ge2were recognized as differentially expressed Thirty-eightprotein spots were found to be significantly regulated amongthree groups of which 15 spots were downregulated and23 spots upregulated Ten of these 38 spots exhibited amore than twofold increase or decrease in abundance asobserved in all replicate gels These 10 regulated proteinswere indicated by the circle in Figure 5 and the selectedregions that showed significant differences in proteinexpression profile of osteoblasts among three groups wereshown in Figure 6 All of them were excised from thegels for further identification by MALDI-TOFTOF-MSanalysis
50
Mr (
kDa)
37
25
20
14
pH 3 10
87
3
6
5
2 10
4
1
9
Figure 5 Representative 2DE gel image Ten spots that were statis-tically significant (119901 lt 005) are shown in the map and indicatedby numbers All were cut from the gels for further identification byMALDI-TOFTOF-MS analysis outlined in Table 2
34 Identification of the Differentially Expressed ProteinsProteins were identified by MALDI-TOFTOF-MS Ten pep-tide mass fingerprints (PMFs) and 50 peptide fragmentfingerprints (PFF) were successfully obtained A selectedPMF of protein spot 6 is displayed in Figure 7(a) and theTOFTOF analysis is shown in Figures 7(b)ndash7(f) All PMFswere evaluated with the Mascot software in NCBInr databaseto identify the protein spots The result had high confidenceif the protein was ranked as the best hit with a significantscore and high sequence coverage Finally we identified eightproteins in these spots Properties of the identification of eightselected protein spots are summarized in Table 2
35 Effect of HQSXD on the Expression of Proteins That Regu-late Antiosteoporotic Activity To further investigate the influ-ence of HQSXD on the expression of antiosteoporotic pro-teins we examined the expression of FPR2 alpha-spectrinPHB andTCEB1 byWestern blottingThe expression of FPR2was increased by treatment with HQSXD-S in comparisonto the blank-control group However HQSXD remarkablydecreased alpha-spectrin PHB and TCEB1 protein levelscompared with blank-control group (Figure 8) Results fromWestern blot manifested the same trend as from proteomicanalysis
Evidence-Based Complementary and Alternative Medicine 7
Spot 1 Spot 2 Spot 3 Spot 4 Spot 5
Spot 6 Spot 7 Spot 8 Spot 9 Spot 10
Blank-control group
HQSXD-treated group
JKSQW-treated group
Blank-control group
HQSXD-treated group
JKSQW-treated group
Figure 6 The ten protein spots of osteoblasts that were treated with and without HQSXD-S Selected regions showed significant differencesin the protein expression profile of osteoblasts among the three groups Upregulated spots are indicated by green circles and downregulatedones by red circles
Table 2 Summary of differentially expressed proteins in osteoblasts treated with HQSXD
Spotnumbera
Proteinscoreb
Matchingpeptides(number)
Theoretical119875119868
cTheoreticalMr (Da)c Target protein Species
1 70 6 927 39299 N-Formyl peptide receptor 2 Rattus norvegicus3 67 7 565 54851 Alpha-spectrin Rattus norvegicus
5 76 6 1008 43712 Heparan sulfate (glucosamine)3-O-sulfotransferase 3A1 Rattus norvegicus
6 173 12 557 29859 Prohibitin Rattus norvegicus
7 67 4 459 12752 Transcription elongationfactor B (SIII) polypeptide 1 Rattus norvegicus
8 70 11 498 127444 Chromosome segregationprotein Rattus norvegicus
9 101 4 691 17386 Nucleoside diphosphatekinase Rattus norvegicus
10 84 12 886 48199 Mast cell carboxypeptidase A Rattus norvegicusaProtein spot number according to Figure 3bProtein scores were based on combined mass and massmass spectra from MALDI-TOFTOF identification MScTheoretical molecular mass (Mr) and isoelectric point (119875
119868) from the NCBInr database
8 Evidence-Based Complementary and Alternative Medicine
724502
995615
861066
1023645
13968251149581
1198660
17947921606823 19980611460637929573 16698152284146
2225096
23712291876991 2119118
00
05
10
15
20
Inte
nsity
(au
)
1000 1500 2000 2500 3000 3500mz
times104
(a)
1023635
687769
832885
643812271987
453127352094621199 772789 979506
174985 322097 876761111971 665734
709626552159385049
86016 295124139982
0
1000
2000
3000
Inte
nsity
(au
)
200 400 600 800 1000 1200mz
(b)
Inte
nsity
(au
)
1037662
254948
467075
989061366041 635224
174962111956 703694338068 886923
80082486005 567816
1012722
200937
287991
0
500
1000
1500
2000
200 400 600 800 1000 1200mz
(c)
Inte
nsity
(au
)
1149574
887334
174919
953820
519998 1105500
633033111931386933
937990
842341486004 595064 785790302933254963
69965
142902
0
1000
2000
3000
200 400 600 800 1000 1200mz
(d)
Inte
nsity
(au
)
1198658
174936
2619201154530
111976440964358918 932243609086
84213585996 325947 741110572984515028287928
400987
228936
200907
0
500
1000
1500
2000
2500
3000
200 400 600 800 1000 1200 1400mz
(e)
Inte
nsity
(au
)
1396813
12406611354732
174968
1070439867233227008111958 286957 1202691
513068417073354995
0
500
1000
1500
2000
2500
200 400 600 800 1000 1200 1400 1600mz
(f)
Figure 7The results of the MALDI-TOFTPF-MS analysis of protein spot 6 (a) Peptide mass fingerprinting and ((b)ndash(f)) peptide fragmentfingerprinting of spot 6
4 Discussion
In this study we found that the proliferative activities ofosteoblasts between HQSXD-S-treated group and JKSQW-control-S-treated group were similar This investigationdemonstrates that HQSXD can significantly facilitate boneformation through increasing the number of osteoblastswhich is beneficial to the treatment of osteoporosis By mea-suring ALP activity HQSXD was first screened for its abilityto induce osteogenesis HQSXD is capable of significantly
promoting osteoblast differentiation as well as increasingosteoblast mineralization
Results of the identification of the selected protein spotsare summarized in Table 2 The molecular weight (Mr) andisoelectric point (119875
119868) of each protein spot shown in Table 2
are theoretical values The eight protein spots were identifiedas (1) N-formyl peptide receptor 2 (FPR2) (2) alpha-spectrin(3) heparan sulfate (glucosamine) 3-O-sulfotransferase 3A1(HS3ST3A1) (4) prohibitin (PHB) (5) transcription elonga-tion factor B (SIII) polypeptide 1 (TCEB1) (6) chromosome
Evidence-Based Complementary and Alternative Medicine 9
TCEB1
PHB
FPR2
Blank HQSXD JKSQW
120573-actin
Alpha-spectrin
Figure 8 HQSXD-S treatment decreased alpha-spectrin PHBand TCEB1 expression and increased FPR2 expression Cell lysates(50 120583g)were processed forWestern blot analysis Normalization per-formed to120573-actinThe bands shown here were from a representativeexperiment repeated three times
segregation protein (7) nucleoside diphosphate kinase and(8) mast cell carboxypeptidase AWe detected some proteinsrelated to transcription cell proliferation differentiationand apoptosis such as FPR2 prohibitin alpha-spectrinheparan sulfate (glucosamine) 3-O-sulfotransferase 3A1 andtranscription elongation factor B (SIII) polypeptide 1 whichvaried greatly after HQSXD treatment
Our results suggest that HQSXD can upregulate theexpression of FPR2 N-Formyl peptide receptor (FPR) is aG protein-coupled receptor which modulates stromal celldifferentiation [14] and binds to N-formyl peptides suchas N-formyl-methionyl-leucyl-phenylalanine (fMLP) Ourstudy has shown that fMLP enhances the differentiationof MSCs into osteoblasts via an FPR-mediated signalingpathway and results in bone formation [15] The FPR2receptor belongs to the formyl peptide receptor family thatis involved in signaling stem cell adhesion migration andhoming for injured and inflamed tissues awaiting repair thiscould potentially be exploited to direct the stem cells to targetspecific tissue site [16] Therefore the current results implythat treatment with HQSXDmight promote the formation ofosteoblasts by upregulating the expression level of FPR2
We found that HQSXD could inhibit the expression ofalpha-spectrin Alpha-spectrin includes two genes encodingfor alpha-I subunit and alpha-II subunit each of thempresenting its specific cellular expression pattern Alpha-II spectrin deficiency is associated with cell proliferationdefects due to cell cycle arrest in theG1 phase (first gap phase)[17] Spectrin and protein kinase C theta were observed inaggregates during the early stage of apoptosis [18] Hencewe presume that alpha-spectrin is involved in the antiosteo-porotic effect of HQSXD
In this study PHB was downregulated in HQSXD-treated osteoblasts Prohibitin a highly conservative proteinregulates the cohesion of sister chromatids cellular signalingmitochondrial biogenesis [19] cell proliferation differentia-tion apoptosis and gene transcription [20 21] PHB blocksthe transition of cells from G1 phase to S phase of the cellcycle thereby arresting cell proliferation [22 23] PHB hasbeen reported to affect the apoptotic pathways by repressingthe transcriptional activity of E2F1 [24 25] As mentionedabove it is possible that HQSXD induces proliferationdifferentiation and apoptosis partly through downregulatingexpression of PHB in osteoblasts It is likely that PHB mightbe the novel candidate in the new antiosteoporotic drugscreening
The expression of TCEB1 was downregulated in HQSXD-treatment osteoblasts TCEB1 a 13 kDa protein also namedelongin C was originally identified as a member of themammalian transcription factor SIII that increases the rateof transcription by suppressing RNA polymerase II pausing[26] As part of a family of separate complexes containingelongin B and various substrate specificity factors it acts asan E3 ubiquitin ligase [27] Elongin B (ELB 1) and elonginC (ELC1) form a stable complex and that depletion of eithergene product by RNA-mediated interference (RNAi) causespronounced defects in the second meiotic division and arrestof germ cell proliferation in G1 [28] Therefore it is possiblethat that TCEB1 is involved in the antiosteoporotic effect ofHQSXD
Although this study has thrown some light on themechanism of HQSXD action we failed to characterize thewell-identified protein (eg HS3ST3A1) closely involved inosteoporosis HS3ST3A1 a member of the heparan sulfatebiosynthetic enzyme family possesses heparan sulfate glu-cosaminyl 3-O-sulfotransferase activity Depletion of Hs3st-A in enterocytes results in increased intestinal stem cellproliferation and tissue homeostasis loss [29]
5 Conclusions
The results confirm that HQSXD has a beneficial effect onosteoblasts and alters the expression level of some proteinsin osteoblasts The protein expressed by osteoblasts treatedwith HQSXD may be involved in cell proliferation anddifferentiation and other physiological processes and in theregulation of cell activation Further study is needed toinvestigate the effects of major active constituents in HQSXDon protein expression on osteoblast so as to demonstrate theinteraction and synergistic mechanism
Conflict of Interests
All the authors state that they have no conflict of interests todeclare
Authorsrsquo Contribution
Chong-Chong Guo and Li-Hua Zheng contributed equally tothis work
10 Evidence-Based Complementary and Alternative Medicine
Acknowledgments
The authors thank Cheng-Ming Liu for her excellent tech-nical assistance This work was supported by the grantfrom the National Natural Science foundation of China (no81273779)
References
[1] S Khosla and B L Riggs ldquoPathophysiology of age-related boneloss and osteoporosisrdquo Endocrinology andMetabolism Clinics ofNorth America vol 34 no 4 pp 1015ndash1030 2005
[2] P Sambrook andCCooper ldquoOsteoporosisrdquoTheLancet vol 367no 9527 pp 2010ndash2018 2006
[3] M R Allen and D B Burr ldquoThree years of alendronate treat-ment results in similar levels of vertebral microdamage as afterone year of treatmentrdquo Journal of Bone and Mineral Researchvol 22 no 11 pp 1759ndash1765 2007
[4] S Davison and S R Davis ldquoHormone replacement therapycurrent controversiesrdquo Clinical Endocrinology vol 58 no 3 pp249ndash261 2003
[5] W-Y Jiang ldquoTherapeutic wisdom in traditional Chinesemedicine a perspective from modern sciencerdquo Trends in Phar-macological Sciences vol 26 no 11 pp 558ndash563 2005
[6] Z Xu and Z K Zhou ldquoClinical study on the treatment of 36cases of postmenopausal osteoporosis with HuangQiSanXianTangrdquo Guiding Journal of Traditional Chinese Medicine andPharmacy vol 15 no 1 pp 9ndash11 2009
[7] H Zhang W-W Xing Y-S Li et al ldquoEffects of a traditionalChinese herbal preparation on osteoblasts and osteoclastsrdquoMaturitas vol 61 no 4 pp 334ndash339 2008
[8] L Qin T Han Q Zhang et al ldquoAntiosteoporotic chemical con-stituents from Er-Xian Decoction a traditional Chinese herbalformulardquo Journal of Ethnopharmacology vol 118 no 2 pp 271ndash279 2008
[9] M Millikan A Kolasani and H Xu ldquoDetermination and com-parison of mineral elements in traditional Chinese herbalformulae at different decoction times used to improve kidneyfunctionmdashchemometric approachrdquo African Journal of Tradi-tional Complementary and Alternative Medicines vol 8 no 5pp 191ndash197 2011
[10] I R Orriss S E B Taylor and T R Arnett ldquoRat osteoblastculturesrdquoMethods inMolecular Biology vol 816 pp 31ndash41 2012
[11] JM Baek J-Y Kim Y-H Cheon et al ldquoDual effect ofChrysan-themum indicum extract to stimulate osteoblast differentia-tion and inhibit osteoclast formation and resorption in vitrordquoEvidence-Based Complementary and Alternative Medicine vol2014 Article ID 176049 13 pages 2014
[12] A I Idris I R Greig E Bassonga-Landao S H Ralston andR J Vanrsquot Hof ldquoIdentification of novel biphenyl carboxylic acidderivatives as novel antiresorptive agents that do not impairparathyroid hormone-induced bone formationrdquo Endocrinologyvol 150 no 1 pp 5ndash13 2009
[13] M Otani J Tabata T Ueki K Sano and S Inouye ldquoHeat-shock-induced proteins from Myxococcus xanthusrdquo Journal ofBacteriology vol 183 no 21 pp 6282ndash6287 2001
[14] M E Nuttall and J M Gimble ldquoIs there a therapeutic opportu-nity to either prevent or treat osteopenic disorders by inhibitingmarrow adipogenesisrdquo Bone vol 27 no 2 pp 177ndash184 2000
[15] M K Shin Y H Jang H J Yoo et al ldquoN-formyl-methionyl-leucyl-phenylalanine (fMLP) promotes osteoblast differentia-tion via the N-formyl peptide receptor 1-mediated signaling
pathway in humanmesenchymal stem cells from bonemarrowrdquoThe Journal of Biological Chemistry vol 286 no 19 pp 17133ndash17143 2011
[16] A Viswanathan R G Painter N A Lanson Jr and G WangldquoFunctional expression ofN-formyl peptide receptors in humanbone marrow-derived mesenchymal stem cellsrdquo Stem Cells vol25 no 5 pp 1263ndash1269 2007
[17] S Metral B Machnicka S Bigot Y Colin D Dhermy and M-C Lecomte ldquo120572II-spectrin is critical for cell adhesion and cellcyclerdquo Journal of Biological Chemistry vol 284 no 4 pp 2409ndash2418 2009
[18] P M Dubielecka M Grzybek A Kolondra et alldquoAggregation of spectrin and PKCtheta is an early hallmarkof fludarabinemitoxantronedexamethasone-induced apopto-sis in Jurkat T and HL60 cellsrdquo Molecular and CellularBiochemistry vol 339 no 1-2 pp 63ndash77 2010
[19] C Merkwirth and T Langer ldquoProhibitin function withinmitochondria essential roles for cell proliferation and cristaemorphogenesisrdquo Biochimica et Biophysica Acta vol 1793 no 1pp 27ndash32 2009
[20] A L Theiss and S V Sitaraman ldquoThe role and therapeuticpotential of prohibitin in diseaserdquoBiochimica et BiophysicaActavol 1813 no 6 pp 1137ndash1143 2011
[21] V Sanchez-Quiles E Santamarıa V Segura L Sesma J Prietoand F J Corrales ldquoProhibitin deficiency blocks proliferationand induces apoptosis in human hepatoma cells molecularmechanisms and functional implicationsrdquo Proteomics vol 10no 8 pp 1609ndash1620 2010
[22] S Manjeshwar D E Branam M R Lerner D J Brackettand E R Jupe ldquoTumor suppression by the prohibitin gene3rsquountranslated region RNA in human breast cancerrdquo CancerResearch vol 63 no 17 pp 5251ndash5256 2003
[23] S Manjeshwar M R Lerner X-P Zang et al ldquoExpressionof prohibitin 31015840 untranslated region suppressor RNA altersmorphology and inhibits motility of breast cancer cellsrdquo Journalof Molecular Histology vol 35 no 6 pp 639ndash646 2004
[24] M Lacroix R-A Toillon and G Leclercq ldquop53 and breastcancer an updaterdquo Endocrine-Related Cancer vol 13 no 2 pp293ndash325 2006
[25] G Fusaro P Dasgupta S Rastogi B Joshi and S ChellappanldquoProhibitin induces the transcriptional activity of p53 andis exported from the nucleus upon apoptotic signalingrdquo TheJournal of Biological Chemistry vol 278 no 48 pp 47853ndash47861 2003
[26] T Aso W S Lane J W Conaway and R C ConawayldquoElongin (SIII) a multisubunit regulator of elongation by RNApolymerase IIrdquo Science vol 269 no 5229 pp 1439ndash1443 1995
[27] T D CumminsM DMendenhall M N Lowry et al ldquoElonginC is a mediator of Notch4 activity in human renal tubule cellsrdquoBiochimica et Biophysica Acta vol 1814 no 12 pp 1748ndash17572011
[28] Y Sasagawa K Kikuchi K Dazai and A HigashitanildquoCaenorhabditis elegans ElonginBC complex is essential for cellproliferation and chromosome condensation and segregationduring mitosis and meiotic division IIrdquo Chromosome Researchvol 13 no 4 pp 357ndash375 2005
[29] Y Guo Z Li and X Lin ldquoHs3st-A and Hs3st-B regulateintestinal homeostasis in Drosophila adult midgutrdquo CellularSignalling vol 26 no 11 pp 2317ndash2325 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 Evidence-Based Complementary and Alternative Medicine
Table 1 Effects of drugs on proliferation of osteoblasts
Groups 119873Oral administration
dose (gsdotkgminus1)Serum additivevolume () OD value
Blank-control 5 mdash 10 050 plusmn 003HQSXD-treated 5 14 10 0634 plusmn 0028lowastlowast
JKSQW-treated 5 0520 10 063 plusmn 004lowastlowast
Note lowastlowastp lt 001 compared with BLANK-control-S
Similar changes in proliferation between JKSQW-treatedgroup andHQSXD-treated group indicated that HQSXDhadan influence on osteoblasts (Table 1 Figure 1)
32 HQSXD-Treated Rat Serum Enhances Primary OsteoblastDifferentiation ALP is an important biochemical marker ofdifferentiated osteoblasts and the effects of drug on ALPactivities in osteoblasts were first determined Results ofALP staining showed that HQSXD-S and JKSQW-control-S stimulated osteoblast differentiation (Figure 2(a)) Thecells cultured with HQSXD-S showed a significantly higherALP activity than that cultured with BLANK-control-S(Figure 2(b)) ARS staining in osteoblasts was assessed after21 days of incubation to examine whether HQSXD enhancedbonemineralization during osteoblastogenesis In osteoblaststreated with BLANK-control-S the calcium deposition in themineralized matrix was minimal The proportional areas ofAlizarin Red-positive staining in the HQSXD-treated groupand JKSQW-treated group were higher than that in theBlank-control group (Figure 3(a)) As shown in Figure 3(b)the level and intensity of ARS staining indicated the extentof mineralization which increased upon treatment withHQSXD-S
33 Protein Expression Profile in HQSXD-Treated andHQSXD-Untreated Osteoblasts Two-dimensional electro-phoresis and gel sliver nitrate staining were conducted tofurther investigate the differential protein expression betweenHQSXD-S-treated and HQSXD-S-untreated osteoblastsAfter optimization of the 2DE gels with representative2DE gel images shown in Figure 4 approximately 938 plusmn 26875 plusmn 34 and 904 plusmn 22 protein spots were detected in blankprotein sample HQSXD protein sample and JKSQWproteinsample respectively During analysis with ImageMaster 2DPlatinum spots with an overlap ratio absolute value ge2were recognized as differentially expressed Thirty-eightprotein spots were found to be significantly regulated amongthree groups of which 15 spots were downregulated and23 spots upregulated Ten of these 38 spots exhibited amore than twofold increase or decrease in abundance asobserved in all replicate gels These 10 regulated proteinswere indicated by the circle in Figure 5 and the selectedregions that showed significant differences in proteinexpression profile of osteoblasts among three groups wereshown in Figure 6 All of them were excised from thegels for further identification by MALDI-TOFTOF-MSanalysis
50
Mr (
kDa)
37
25
20
14
pH 3 10
87
3
6
5
2 10
4
1
9
Figure 5 Representative 2DE gel image Ten spots that were statis-tically significant (119901 lt 005) are shown in the map and indicatedby numbers All were cut from the gels for further identification byMALDI-TOFTOF-MS analysis outlined in Table 2
34 Identification of the Differentially Expressed ProteinsProteins were identified by MALDI-TOFTOF-MS Ten pep-tide mass fingerprints (PMFs) and 50 peptide fragmentfingerprints (PFF) were successfully obtained A selectedPMF of protein spot 6 is displayed in Figure 7(a) and theTOFTOF analysis is shown in Figures 7(b)ndash7(f) All PMFswere evaluated with the Mascot software in NCBInr databaseto identify the protein spots The result had high confidenceif the protein was ranked as the best hit with a significantscore and high sequence coverage Finally we identified eightproteins in these spots Properties of the identification of eightselected protein spots are summarized in Table 2
35 Effect of HQSXD on the Expression of Proteins That Regu-late Antiosteoporotic Activity To further investigate the influ-ence of HQSXD on the expression of antiosteoporotic pro-teins we examined the expression of FPR2 alpha-spectrinPHB andTCEB1 byWestern blottingThe expression of FPR2was increased by treatment with HQSXD-S in comparisonto the blank-control group However HQSXD remarkablydecreased alpha-spectrin PHB and TCEB1 protein levelscompared with blank-control group (Figure 8) Results fromWestern blot manifested the same trend as from proteomicanalysis
Evidence-Based Complementary and Alternative Medicine 7
Spot 1 Spot 2 Spot 3 Spot 4 Spot 5
Spot 6 Spot 7 Spot 8 Spot 9 Spot 10
Blank-control group
HQSXD-treated group
JKSQW-treated group
Blank-control group
HQSXD-treated group
JKSQW-treated group
Figure 6 The ten protein spots of osteoblasts that were treated with and without HQSXD-S Selected regions showed significant differencesin the protein expression profile of osteoblasts among the three groups Upregulated spots are indicated by green circles and downregulatedones by red circles
Table 2 Summary of differentially expressed proteins in osteoblasts treated with HQSXD
Spotnumbera
Proteinscoreb
Matchingpeptides(number)
Theoretical119875119868
cTheoreticalMr (Da)c Target protein Species
1 70 6 927 39299 N-Formyl peptide receptor 2 Rattus norvegicus3 67 7 565 54851 Alpha-spectrin Rattus norvegicus
5 76 6 1008 43712 Heparan sulfate (glucosamine)3-O-sulfotransferase 3A1 Rattus norvegicus
6 173 12 557 29859 Prohibitin Rattus norvegicus
7 67 4 459 12752 Transcription elongationfactor B (SIII) polypeptide 1 Rattus norvegicus
8 70 11 498 127444 Chromosome segregationprotein Rattus norvegicus
9 101 4 691 17386 Nucleoside diphosphatekinase Rattus norvegicus
10 84 12 886 48199 Mast cell carboxypeptidase A Rattus norvegicusaProtein spot number according to Figure 3bProtein scores were based on combined mass and massmass spectra from MALDI-TOFTOF identification MScTheoretical molecular mass (Mr) and isoelectric point (119875
119868) from the NCBInr database
8 Evidence-Based Complementary and Alternative Medicine
724502
995615
861066
1023645
13968251149581
1198660
17947921606823 19980611460637929573 16698152284146
2225096
23712291876991 2119118
00
05
10
15
20
Inte
nsity
(au
)
1000 1500 2000 2500 3000 3500mz
times104
(a)
1023635
687769
832885
643812271987
453127352094621199 772789 979506
174985 322097 876761111971 665734
709626552159385049
86016 295124139982
0
1000
2000
3000
Inte
nsity
(au
)
200 400 600 800 1000 1200mz
(b)
Inte
nsity
(au
)
1037662
254948
467075
989061366041 635224
174962111956 703694338068 886923
80082486005 567816
1012722
200937
287991
0
500
1000
1500
2000
200 400 600 800 1000 1200mz
(c)
Inte
nsity
(au
)
1149574
887334
174919
953820
519998 1105500
633033111931386933
937990
842341486004 595064 785790302933254963
69965
142902
0
1000
2000
3000
200 400 600 800 1000 1200mz
(d)
Inte
nsity
(au
)
1198658
174936
2619201154530
111976440964358918 932243609086
84213585996 325947 741110572984515028287928
400987
228936
200907
0
500
1000
1500
2000
2500
3000
200 400 600 800 1000 1200 1400mz
(e)
Inte
nsity
(au
)
1396813
12406611354732
174968
1070439867233227008111958 286957 1202691
513068417073354995
0
500
1000
1500
2000
2500
200 400 600 800 1000 1200 1400 1600mz
(f)
Figure 7The results of the MALDI-TOFTPF-MS analysis of protein spot 6 (a) Peptide mass fingerprinting and ((b)ndash(f)) peptide fragmentfingerprinting of spot 6
4 Discussion
In this study we found that the proliferative activities ofosteoblasts between HQSXD-S-treated group and JKSQW-control-S-treated group were similar This investigationdemonstrates that HQSXD can significantly facilitate boneformation through increasing the number of osteoblastswhich is beneficial to the treatment of osteoporosis By mea-suring ALP activity HQSXD was first screened for its abilityto induce osteogenesis HQSXD is capable of significantly
promoting osteoblast differentiation as well as increasingosteoblast mineralization
Results of the identification of the selected protein spotsare summarized in Table 2 The molecular weight (Mr) andisoelectric point (119875
119868) of each protein spot shown in Table 2
are theoretical values The eight protein spots were identifiedas (1) N-formyl peptide receptor 2 (FPR2) (2) alpha-spectrin(3) heparan sulfate (glucosamine) 3-O-sulfotransferase 3A1(HS3ST3A1) (4) prohibitin (PHB) (5) transcription elonga-tion factor B (SIII) polypeptide 1 (TCEB1) (6) chromosome
Evidence-Based Complementary and Alternative Medicine 9
TCEB1
PHB
FPR2
Blank HQSXD JKSQW
120573-actin
Alpha-spectrin
Figure 8 HQSXD-S treatment decreased alpha-spectrin PHBand TCEB1 expression and increased FPR2 expression Cell lysates(50 120583g)were processed forWestern blot analysis Normalization per-formed to120573-actinThe bands shown here were from a representativeexperiment repeated three times
segregation protein (7) nucleoside diphosphate kinase and(8) mast cell carboxypeptidase AWe detected some proteinsrelated to transcription cell proliferation differentiationand apoptosis such as FPR2 prohibitin alpha-spectrinheparan sulfate (glucosamine) 3-O-sulfotransferase 3A1 andtranscription elongation factor B (SIII) polypeptide 1 whichvaried greatly after HQSXD treatment
Our results suggest that HQSXD can upregulate theexpression of FPR2 N-Formyl peptide receptor (FPR) is aG protein-coupled receptor which modulates stromal celldifferentiation [14] and binds to N-formyl peptides suchas N-formyl-methionyl-leucyl-phenylalanine (fMLP) Ourstudy has shown that fMLP enhances the differentiationof MSCs into osteoblasts via an FPR-mediated signalingpathway and results in bone formation [15] The FPR2receptor belongs to the formyl peptide receptor family thatis involved in signaling stem cell adhesion migration andhoming for injured and inflamed tissues awaiting repair thiscould potentially be exploited to direct the stem cells to targetspecific tissue site [16] Therefore the current results implythat treatment with HQSXDmight promote the formation ofosteoblasts by upregulating the expression level of FPR2
We found that HQSXD could inhibit the expression ofalpha-spectrin Alpha-spectrin includes two genes encodingfor alpha-I subunit and alpha-II subunit each of thempresenting its specific cellular expression pattern Alpha-II spectrin deficiency is associated with cell proliferationdefects due to cell cycle arrest in theG1 phase (first gap phase)[17] Spectrin and protein kinase C theta were observed inaggregates during the early stage of apoptosis [18] Hencewe presume that alpha-spectrin is involved in the antiosteo-porotic effect of HQSXD
In this study PHB was downregulated in HQSXD-treated osteoblasts Prohibitin a highly conservative proteinregulates the cohesion of sister chromatids cellular signalingmitochondrial biogenesis [19] cell proliferation differentia-tion apoptosis and gene transcription [20 21] PHB blocksthe transition of cells from G1 phase to S phase of the cellcycle thereby arresting cell proliferation [22 23] PHB hasbeen reported to affect the apoptotic pathways by repressingthe transcriptional activity of E2F1 [24 25] As mentionedabove it is possible that HQSXD induces proliferationdifferentiation and apoptosis partly through downregulatingexpression of PHB in osteoblasts It is likely that PHB mightbe the novel candidate in the new antiosteoporotic drugscreening
The expression of TCEB1 was downregulated in HQSXD-treatment osteoblasts TCEB1 a 13 kDa protein also namedelongin C was originally identified as a member of themammalian transcription factor SIII that increases the rateof transcription by suppressing RNA polymerase II pausing[26] As part of a family of separate complexes containingelongin B and various substrate specificity factors it acts asan E3 ubiquitin ligase [27] Elongin B (ELB 1) and elonginC (ELC1) form a stable complex and that depletion of eithergene product by RNA-mediated interference (RNAi) causespronounced defects in the second meiotic division and arrestof germ cell proliferation in G1 [28] Therefore it is possiblethat that TCEB1 is involved in the antiosteoporotic effect ofHQSXD
Although this study has thrown some light on themechanism of HQSXD action we failed to characterize thewell-identified protein (eg HS3ST3A1) closely involved inosteoporosis HS3ST3A1 a member of the heparan sulfatebiosynthetic enzyme family possesses heparan sulfate glu-cosaminyl 3-O-sulfotransferase activity Depletion of Hs3st-A in enterocytes results in increased intestinal stem cellproliferation and tissue homeostasis loss [29]
5 Conclusions
The results confirm that HQSXD has a beneficial effect onosteoblasts and alters the expression level of some proteinsin osteoblasts The protein expressed by osteoblasts treatedwith HQSXD may be involved in cell proliferation anddifferentiation and other physiological processes and in theregulation of cell activation Further study is needed toinvestigate the effects of major active constituents in HQSXDon protein expression on osteoblast so as to demonstrate theinteraction and synergistic mechanism
Conflict of Interests
All the authors state that they have no conflict of interests todeclare
Authorsrsquo Contribution
Chong-Chong Guo and Li-Hua Zheng contributed equally tothis work
10 Evidence-Based Complementary and Alternative Medicine
Acknowledgments
The authors thank Cheng-Ming Liu for her excellent tech-nical assistance This work was supported by the grantfrom the National Natural Science foundation of China (no81273779)
References
[1] S Khosla and B L Riggs ldquoPathophysiology of age-related boneloss and osteoporosisrdquo Endocrinology andMetabolism Clinics ofNorth America vol 34 no 4 pp 1015ndash1030 2005
[2] P Sambrook andCCooper ldquoOsteoporosisrdquoTheLancet vol 367no 9527 pp 2010ndash2018 2006
[3] M R Allen and D B Burr ldquoThree years of alendronate treat-ment results in similar levels of vertebral microdamage as afterone year of treatmentrdquo Journal of Bone and Mineral Researchvol 22 no 11 pp 1759ndash1765 2007
[4] S Davison and S R Davis ldquoHormone replacement therapycurrent controversiesrdquo Clinical Endocrinology vol 58 no 3 pp249ndash261 2003
[5] W-Y Jiang ldquoTherapeutic wisdom in traditional Chinesemedicine a perspective from modern sciencerdquo Trends in Phar-macological Sciences vol 26 no 11 pp 558ndash563 2005
[6] Z Xu and Z K Zhou ldquoClinical study on the treatment of 36cases of postmenopausal osteoporosis with HuangQiSanXianTangrdquo Guiding Journal of Traditional Chinese Medicine andPharmacy vol 15 no 1 pp 9ndash11 2009
[7] H Zhang W-W Xing Y-S Li et al ldquoEffects of a traditionalChinese herbal preparation on osteoblasts and osteoclastsrdquoMaturitas vol 61 no 4 pp 334ndash339 2008
[8] L Qin T Han Q Zhang et al ldquoAntiosteoporotic chemical con-stituents from Er-Xian Decoction a traditional Chinese herbalformulardquo Journal of Ethnopharmacology vol 118 no 2 pp 271ndash279 2008
[9] M Millikan A Kolasani and H Xu ldquoDetermination and com-parison of mineral elements in traditional Chinese herbalformulae at different decoction times used to improve kidneyfunctionmdashchemometric approachrdquo African Journal of Tradi-tional Complementary and Alternative Medicines vol 8 no 5pp 191ndash197 2011
[10] I R Orriss S E B Taylor and T R Arnett ldquoRat osteoblastculturesrdquoMethods inMolecular Biology vol 816 pp 31ndash41 2012
[11] JM Baek J-Y Kim Y-H Cheon et al ldquoDual effect ofChrysan-themum indicum extract to stimulate osteoblast differentia-tion and inhibit osteoclast formation and resorption in vitrordquoEvidence-Based Complementary and Alternative Medicine vol2014 Article ID 176049 13 pages 2014
[12] A I Idris I R Greig E Bassonga-Landao S H Ralston andR J Vanrsquot Hof ldquoIdentification of novel biphenyl carboxylic acidderivatives as novel antiresorptive agents that do not impairparathyroid hormone-induced bone formationrdquo Endocrinologyvol 150 no 1 pp 5ndash13 2009
[13] M Otani J Tabata T Ueki K Sano and S Inouye ldquoHeat-shock-induced proteins from Myxococcus xanthusrdquo Journal ofBacteriology vol 183 no 21 pp 6282ndash6287 2001
[14] M E Nuttall and J M Gimble ldquoIs there a therapeutic opportu-nity to either prevent or treat osteopenic disorders by inhibitingmarrow adipogenesisrdquo Bone vol 27 no 2 pp 177ndash184 2000
[15] M K Shin Y H Jang H J Yoo et al ldquoN-formyl-methionyl-leucyl-phenylalanine (fMLP) promotes osteoblast differentia-tion via the N-formyl peptide receptor 1-mediated signaling
pathway in humanmesenchymal stem cells from bonemarrowrdquoThe Journal of Biological Chemistry vol 286 no 19 pp 17133ndash17143 2011
[16] A Viswanathan R G Painter N A Lanson Jr and G WangldquoFunctional expression ofN-formyl peptide receptors in humanbone marrow-derived mesenchymal stem cellsrdquo Stem Cells vol25 no 5 pp 1263ndash1269 2007
[17] S Metral B Machnicka S Bigot Y Colin D Dhermy and M-C Lecomte ldquo120572II-spectrin is critical for cell adhesion and cellcyclerdquo Journal of Biological Chemistry vol 284 no 4 pp 2409ndash2418 2009
[18] P M Dubielecka M Grzybek A Kolondra et alldquoAggregation of spectrin and PKCtheta is an early hallmarkof fludarabinemitoxantronedexamethasone-induced apopto-sis in Jurkat T and HL60 cellsrdquo Molecular and CellularBiochemistry vol 339 no 1-2 pp 63ndash77 2010
[19] C Merkwirth and T Langer ldquoProhibitin function withinmitochondria essential roles for cell proliferation and cristaemorphogenesisrdquo Biochimica et Biophysica Acta vol 1793 no 1pp 27ndash32 2009
[20] A L Theiss and S V Sitaraman ldquoThe role and therapeuticpotential of prohibitin in diseaserdquoBiochimica et BiophysicaActavol 1813 no 6 pp 1137ndash1143 2011
[21] V Sanchez-Quiles E Santamarıa V Segura L Sesma J Prietoand F J Corrales ldquoProhibitin deficiency blocks proliferationand induces apoptosis in human hepatoma cells molecularmechanisms and functional implicationsrdquo Proteomics vol 10no 8 pp 1609ndash1620 2010
[22] S Manjeshwar D E Branam M R Lerner D J Brackettand E R Jupe ldquoTumor suppression by the prohibitin gene3rsquountranslated region RNA in human breast cancerrdquo CancerResearch vol 63 no 17 pp 5251ndash5256 2003
[23] S Manjeshwar M R Lerner X-P Zang et al ldquoExpressionof prohibitin 31015840 untranslated region suppressor RNA altersmorphology and inhibits motility of breast cancer cellsrdquo Journalof Molecular Histology vol 35 no 6 pp 639ndash646 2004
[24] M Lacroix R-A Toillon and G Leclercq ldquop53 and breastcancer an updaterdquo Endocrine-Related Cancer vol 13 no 2 pp293ndash325 2006
[25] G Fusaro P Dasgupta S Rastogi B Joshi and S ChellappanldquoProhibitin induces the transcriptional activity of p53 andis exported from the nucleus upon apoptotic signalingrdquo TheJournal of Biological Chemistry vol 278 no 48 pp 47853ndash47861 2003
[26] T Aso W S Lane J W Conaway and R C ConawayldquoElongin (SIII) a multisubunit regulator of elongation by RNApolymerase IIrdquo Science vol 269 no 5229 pp 1439ndash1443 1995
[27] T D CumminsM DMendenhall M N Lowry et al ldquoElonginC is a mediator of Notch4 activity in human renal tubule cellsrdquoBiochimica et Biophysica Acta vol 1814 no 12 pp 1748ndash17572011
[28] Y Sasagawa K Kikuchi K Dazai and A HigashitanildquoCaenorhabditis elegans ElonginBC complex is essential for cellproliferation and chromosome condensation and segregationduring mitosis and meiotic division IIrdquo Chromosome Researchvol 13 no 4 pp 357ndash375 2005
[29] Y Guo Z Li and X Lin ldquoHs3st-A and Hs3st-B regulateintestinal homeostasis in Drosophila adult midgutrdquo CellularSignalling vol 26 no 11 pp 2317ndash2325 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 7
Spot 1 Spot 2 Spot 3 Spot 4 Spot 5
Spot 6 Spot 7 Spot 8 Spot 9 Spot 10
Blank-control group
HQSXD-treated group
JKSQW-treated group
Blank-control group
HQSXD-treated group
JKSQW-treated group
Figure 6 The ten protein spots of osteoblasts that were treated with and without HQSXD-S Selected regions showed significant differencesin the protein expression profile of osteoblasts among the three groups Upregulated spots are indicated by green circles and downregulatedones by red circles
Table 2 Summary of differentially expressed proteins in osteoblasts treated with HQSXD
Spotnumbera
Proteinscoreb
Matchingpeptides(number)
Theoretical119875119868
cTheoreticalMr (Da)c Target protein Species
1 70 6 927 39299 N-Formyl peptide receptor 2 Rattus norvegicus3 67 7 565 54851 Alpha-spectrin Rattus norvegicus
5 76 6 1008 43712 Heparan sulfate (glucosamine)3-O-sulfotransferase 3A1 Rattus norvegicus
6 173 12 557 29859 Prohibitin Rattus norvegicus
7 67 4 459 12752 Transcription elongationfactor B (SIII) polypeptide 1 Rattus norvegicus
8 70 11 498 127444 Chromosome segregationprotein Rattus norvegicus
9 101 4 691 17386 Nucleoside diphosphatekinase Rattus norvegicus
10 84 12 886 48199 Mast cell carboxypeptidase A Rattus norvegicusaProtein spot number according to Figure 3bProtein scores were based on combined mass and massmass spectra from MALDI-TOFTOF identification MScTheoretical molecular mass (Mr) and isoelectric point (119875
119868) from the NCBInr database
8 Evidence-Based Complementary and Alternative Medicine
724502
995615
861066
1023645
13968251149581
1198660
17947921606823 19980611460637929573 16698152284146
2225096
23712291876991 2119118
00
05
10
15
20
Inte
nsity
(au
)
1000 1500 2000 2500 3000 3500mz
times104
(a)
1023635
687769
832885
643812271987
453127352094621199 772789 979506
174985 322097 876761111971 665734
709626552159385049
86016 295124139982
0
1000
2000
3000
Inte
nsity
(au
)
200 400 600 800 1000 1200mz
(b)
Inte
nsity
(au
)
1037662
254948
467075
989061366041 635224
174962111956 703694338068 886923
80082486005 567816
1012722
200937
287991
0
500
1000
1500
2000
200 400 600 800 1000 1200mz
(c)
Inte
nsity
(au
)
1149574
887334
174919
953820
519998 1105500
633033111931386933
937990
842341486004 595064 785790302933254963
69965
142902
0
1000
2000
3000
200 400 600 800 1000 1200mz
(d)
Inte
nsity
(au
)
1198658
174936
2619201154530
111976440964358918 932243609086
84213585996 325947 741110572984515028287928
400987
228936
200907
0
500
1000
1500
2000
2500
3000
200 400 600 800 1000 1200 1400mz
(e)
Inte
nsity
(au
)
1396813
12406611354732
174968
1070439867233227008111958 286957 1202691
513068417073354995
0
500
1000
1500
2000
2500
200 400 600 800 1000 1200 1400 1600mz
(f)
Figure 7The results of the MALDI-TOFTPF-MS analysis of protein spot 6 (a) Peptide mass fingerprinting and ((b)ndash(f)) peptide fragmentfingerprinting of spot 6
4 Discussion
In this study we found that the proliferative activities ofosteoblasts between HQSXD-S-treated group and JKSQW-control-S-treated group were similar This investigationdemonstrates that HQSXD can significantly facilitate boneformation through increasing the number of osteoblastswhich is beneficial to the treatment of osteoporosis By mea-suring ALP activity HQSXD was first screened for its abilityto induce osteogenesis HQSXD is capable of significantly
promoting osteoblast differentiation as well as increasingosteoblast mineralization
Results of the identification of the selected protein spotsare summarized in Table 2 The molecular weight (Mr) andisoelectric point (119875
119868) of each protein spot shown in Table 2
are theoretical values The eight protein spots were identifiedas (1) N-formyl peptide receptor 2 (FPR2) (2) alpha-spectrin(3) heparan sulfate (glucosamine) 3-O-sulfotransferase 3A1(HS3ST3A1) (4) prohibitin (PHB) (5) transcription elonga-tion factor B (SIII) polypeptide 1 (TCEB1) (6) chromosome
Evidence-Based Complementary and Alternative Medicine 9
TCEB1
PHB
FPR2
Blank HQSXD JKSQW
120573-actin
Alpha-spectrin
Figure 8 HQSXD-S treatment decreased alpha-spectrin PHBand TCEB1 expression and increased FPR2 expression Cell lysates(50 120583g)were processed forWestern blot analysis Normalization per-formed to120573-actinThe bands shown here were from a representativeexperiment repeated three times
segregation protein (7) nucleoside diphosphate kinase and(8) mast cell carboxypeptidase AWe detected some proteinsrelated to transcription cell proliferation differentiationand apoptosis such as FPR2 prohibitin alpha-spectrinheparan sulfate (glucosamine) 3-O-sulfotransferase 3A1 andtranscription elongation factor B (SIII) polypeptide 1 whichvaried greatly after HQSXD treatment
Our results suggest that HQSXD can upregulate theexpression of FPR2 N-Formyl peptide receptor (FPR) is aG protein-coupled receptor which modulates stromal celldifferentiation [14] and binds to N-formyl peptides suchas N-formyl-methionyl-leucyl-phenylalanine (fMLP) Ourstudy has shown that fMLP enhances the differentiationof MSCs into osteoblasts via an FPR-mediated signalingpathway and results in bone formation [15] The FPR2receptor belongs to the formyl peptide receptor family thatis involved in signaling stem cell adhesion migration andhoming for injured and inflamed tissues awaiting repair thiscould potentially be exploited to direct the stem cells to targetspecific tissue site [16] Therefore the current results implythat treatment with HQSXDmight promote the formation ofosteoblasts by upregulating the expression level of FPR2
We found that HQSXD could inhibit the expression ofalpha-spectrin Alpha-spectrin includes two genes encodingfor alpha-I subunit and alpha-II subunit each of thempresenting its specific cellular expression pattern Alpha-II spectrin deficiency is associated with cell proliferationdefects due to cell cycle arrest in theG1 phase (first gap phase)[17] Spectrin and protein kinase C theta were observed inaggregates during the early stage of apoptosis [18] Hencewe presume that alpha-spectrin is involved in the antiosteo-porotic effect of HQSXD
In this study PHB was downregulated in HQSXD-treated osteoblasts Prohibitin a highly conservative proteinregulates the cohesion of sister chromatids cellular signalingmitochondrial biogenesis [19] cell proliferation differentia-tion apoptosis and gene transcription [20 21] PHB blocksthe transition of cells from G1 phase to S phase of the cellcycle thereby arresting cell proliferation [22 23] PHB hasbeen reported to affect the apoptotic pathways by repressingthe transcriptional activity of E2F1 [24 25] As mentionedabove it is possible that HQSXD induces proliferationdifferentiation and apoptosis partly through downregulatingexpression of PHB in osteoblasts It is likely that PHB mightbe the novel candidate in the new antiosteoporotic drugscreening
The expression of TCEB1 was downregulated in HQSXD-treatment osteoblasts TCEB1 a 13 kDa protein also namedelongin C was originally identified as a member of themammalian transcription factor SIII that increases the rateof transcription by suppressing RNA polymerase II pausing[26] As part of a family of separate complexes containingelongin B and various substrate specificity factors it acts asan E3 ubiquitin ligase [27] Elongin B (ELB 1) and elonginC (ELC1) form a stable complex and that depletion of eithergene product by RNA-mediated interference (RNAi) causespronounced defects in the second meiotic division and arrestof germ cell proliferation in G1 [28] Therefore it is possiblethat that TCEB1 is involved in the antiosteoporotic effect ofHQSXD
Although this study has thrown some light on themechanism of HQSXD action we failed to characterize thewell-identified protein (eg HS3ST3A1) closely involved inosteoporosis HS3ST3A1 a member of the heparan sulfatebiosynthetic enzyme family possesses heparan sulfate glu-cosaminyl 3-O-sulfotransferase activity Depletion of Hs3st-A in enterocytes results in increased intestinal stem cellproliferation and tissue homeostasis loss [29]
5 Conclusions
The results confirm that HQSXD has a beneficial effect onosteoblasts and alters the expression level of some proteinsin osteoblasts The protein expressed by osteoblasts treatedwith HQSXD may be involved in cell proliferation anddifferentiation and other physiological processes and in theregulation of cell activation Further study is needed toinvestigate the effects of major active constituents in HQSXDon protein expression on osteoblast so as to demonstrate theinteraction and synergistic mechanism
Conflict of Interests
All the authors state that they have no conflict of interests todeclare
Authorsrsquo Contribution
Chong-Chong Guo and Li-Hua Zheng contributed equally tothis work
10 Evidence-Based Complementary and Alternative Medicine
Acknowledgments
The authors thank Cheng-Ming Liu for her excellent tech-nical assistance This work was supported by the grantfrom the National Natural Science foundation of China (no81273779)
References
[1] S Khosla and B L Riggs ldquoPathophysiology of age-related boneloss and osteoporosisrdquo Endocrinology andMetabolism Clinics ofNorth America vol 34 no 4 pp 1015ndash1030 2005
[2] P Sambrook andCCooper ldquoOsteoporosisrdquoTheLancet vol 367no 9527 pp 2010ndash2018 2006
[3] M R Allen and D B Burr ldquoThree years of alendronate treat-ment results in similar levels of vertebral microdamage as afterone year of treatmentrdquo Journal of Bone and Mineral Researchvol 22 no 11 pp 1759ndash1765 2007
[4] S Davison and S R Davis ldquoHormone replacement therapycurrent controversiesrdquo Clinical Endocrinology vol 58 no 3 pp249ndash261 2003
[5] W-Y Jiang ldquoTherapeutic wisdom in traditional Chinesemedicine a perspective from modern sciencerdquo Trends in Phar-macological Sciences vol 26 no 11 pp 558ndash563 2005
[6] Z Xu and Z K Zhou ldquoClinical study on the treatment of 36cases of postmenopausal osteoporosis with HuangQiSanXianTangrdquo Guiding Journal of Traditional Chinese Medicine andPharmacy vol 15 no 1 pp 9ndash11 2009
[7] H Zhang W-W Xing Y-S Li et al ldquoEffects of a traditionalChinese herbal preparation on osteoblasts and osteoclastsrdquoMaturitas vol 61 no 4 pp 334ndash339 2008
[8] L Qin T Han Q Zhang et al ldquoAntiosteoporotic chemical con-stituents from Er-Xian Decoction a traditional Chinese herbalformulardquo Journal of Ethnopharmacology vol 118 no 2 pp 271ndash279 2008
[9] M Millikan A Kolasani and H Xu ldquoDetermination and com-parison of mineral elements in traditional Chinese herbalformulae at different decoction times used to improve kidneyfunctionmdashchemometric approachrdquo African Journal of Tradi-tional Complementary and Alternative Medicines vol 8 no 5pp 191ndash197 2011
[10] I R Orriss S E B Taylor and T R Arnett ldquoRat osteoblastculturesrdquoMethods inMolecular Biology vol 816 pp 31ndash41 2012
[11] JM Baek J-Y Kim Y-H Cheon et al ldquoDual effect ofChrysan-themum indicum extract to stimulate osteoblast differentia-tion and inhibit osteoclast formation and resorption in vitrordquoEvidence-Based Complementary and Alternative Medicine vol2014 Article ID 176049 13 pages 2014
[12] A I Idris I R Greig E Bassonga-Landao S H Ralston andR J Vanrsquot Hof ldquoIdentification of novel biphenyl carboxylic acidderivatives as novel antiresorptive agents that do not impairparathyroid hormone-induced bone formationrdquo Endocrinologyvol 150 no 1 pp 5ndash13 2009
[13] M Otani J Tabata T Ueki K Sano and S Inouye ldquoHeat-shock-induced proteins from Myxococcus xanthusrdquo Journal ofBacteriology vol 183 no 21 pp 6282ndash6287 2001
[14] M E Nuttall and J M Gimble ldquoIs there a therapeutic opportu-nity to either prevent or treat osteopenic disorders by inhibitingmarrow adipogenesisrdquo Bone vol 27 no 2 pp 177ndash184 2000
[15] M K Shin Y H Jang H J Yoo et al ldquoN-formyl-methionyl-leucyl-phenylalanine (fMLP) promotes osteoblast differentia-tion via the N-formyl peptide receptor 1-mediated signaling
pathway in humanmesenchymal stem cells from bonemarrowrdquoThe Journal of Biological Chemistry vol 286 no 19 pp 17133ndash17143 2011
[16] A Viswanathan R G Painter N A Lanson Jr and G WangldquoFunctional expression ofN-formyl peptide receptors in humanbone marrow-derived mesenchymal stem cellsrdquo Stem Cells vol25 no 5 pp 1263ndash1269 2007
[17] S Metral B Machnicka S Bigot Y Colin D Dhermy and M-C Lecomte ldquo120572II-spectrin is critical for cell adhesion and cellcyclerdquo Journal of Biological Chemistry vol 284 no 4 pp 2409ndash2418 2009
[18] P M Dubielecka M Grzybek A Kolondra et alldquoAggregation of spectrin and PKCtheta is an early hallmarkof fludarabinemitoxantronedexamethasone-induced apopto-sis in Jurkat T and HL60 cellsrdquo Molecular and CellularBiochemistry vol 339 no 1-2 pp 63ndash77 2010
[19] C Merkwirth and T Langer ldquoProhibitin function withinmitochondria essential roles for cell proliferation and cristaemorphogenesisrdquo Biochimica et Biophysica Acta vol 1793 no 1pp 27ndash32 2009
[20] A L Theiss and S V Sitaraman ldquoThe role and therapeuticpotential of prohibitin in diseaserdquoBiochimica et BiophysicaActavol 1813 no 6 pp 1137ndash1143 2011
[21] V Sanchez-Quiles E Santamarıa V Segura L Sesma J Prietoand F J Corrales ldquoProhibitin deficiency blocks proliferationand induces apoptosis in human hepatoma cells molecularmechanisms and functional implicationsrdquo Proteomics vol 10no 8 pp 1609ndash1620 2010
[22] S Manjeshwar D E Branam M R Lerner D J Brackettand E R Jupe ldquoTumor suppression by the prohibitin gene3rsquountranslated region RNA in human breast cancerrdquo CancerResearch vol 63 no 17 pp 5251ndash5256 2003
[23] S Manjeshwar M R Lerner X-P Zang et al ldquoExpressionof prohibitin 31015840 untranslated region suppressor RNA altersmorphology and inhibits motility of breast cancer cellsrdquo Journalof Molecular Histology vol 35 no 6 pp 639ndash646 2004
[24] M Lacroix R-A Toillon and G Leclercq ldquop53 and breastcancer an updaterdquo Endocrine-Related Cancer vol 13 no 2 pp293ndash325 2006
[25] G Fusaro P Dasgupta S Rastogi B Joshi and S ChellappanldquoProhibitin induces the transcriptional activity of p53 andis exported from the nucleus upon apoptotic signalingrdquo TheJournal of Biological Chemistry vol 278 no 48 pp 47853ndash47861 2003
[26] T Aso W S Lane J W Conaway and R C ConawayldquoElongin (SIII) a multisubunit regulator of elongation by RNApolymerase IIrdquo Science vol 269 no 5229 pp 1439ndash1443 1995
[27] T D CumminsM DMendenhall M N Lowry et al ldquoElonginC is a mediator of Notch4 activity in human renal tubule cellsrdquoBiochimica et Biophysica Acta vol 1814 no 12 pp 1748ndash17572011
[28] Y Sasagawa K Kikuchi K Dazai and A HigashitanildquoCaenorhabditis elegans ElonginBC complex is essential for cellproliferation and chromosome condensation and segregationduring mitosis and meiotic division IIrdquo Chromosome Researchvol 13 no 4 pp 357ndash375 2005
[29] Y Guo Z Li and X Lin ldquoHs3st-A and Hs3st-B regulateintestinal homeostasis in Drosophila adult midgutrdquo CellularSignalling vol 26 no 11 pp 2317ndash2325 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
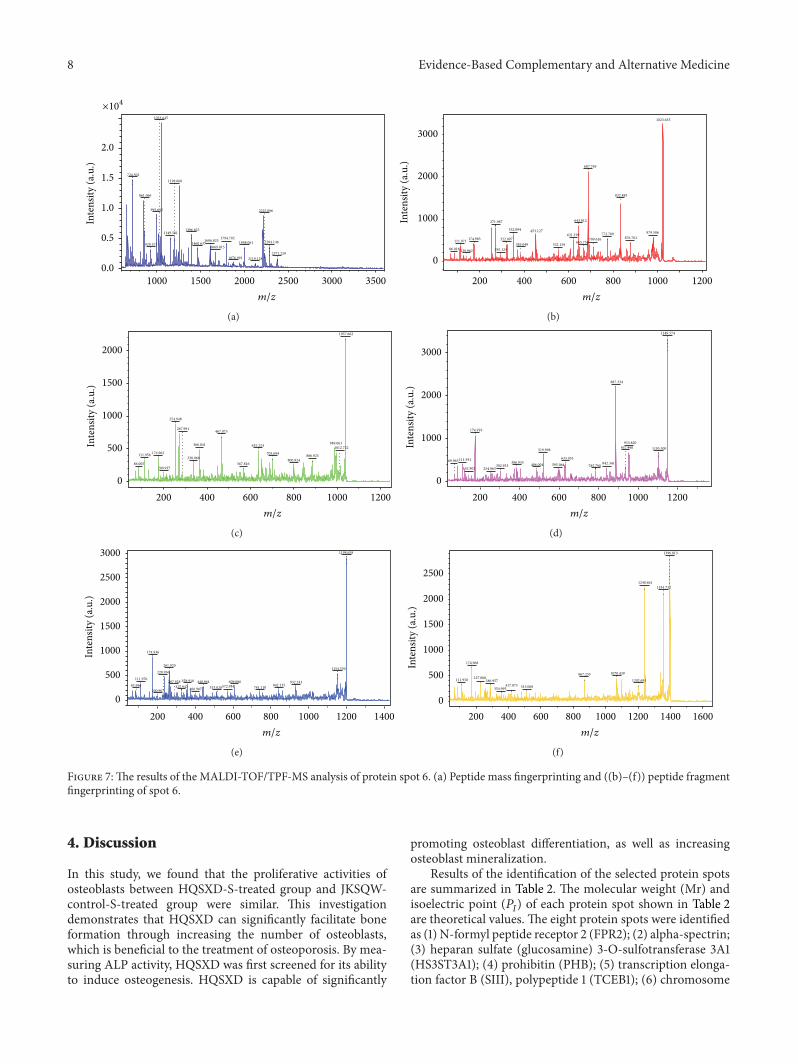
8 Evidence-Based Complementary and Alternative Medicine
724502
995615
861066
1023645
13968251149581
1198660
17947921606823 19980611460637929573 16698152284146
2225096
23712291876991 2119118
00
05
10
15
20
Inte
nsity
(au
)
1000 1500 2000 2500 3000 3500mz
times104
(a)
1023635
687769
832885
643812271987
453127352094621199 772789 979506
174985 322097 876761111971 665734
709626552159385049
86016 295124139982
0
1000
2000
3000
Inte
nsity
(au
)
200 400 600 800 1000 1200mz
(b)
Inte
nsity
(au
)
1037662
254948
467075
989061366041 635224
174962111956 703694338068 886923
80082486005 567816
1012722
200937
287991
0
500
1000
1500
2000
200 400 600 800 1000 1200mz
(c)
Inte
nsity
(au
)
1149574
887334
174919
953820
519998 1105500
633033111931386933
937990
842341486004 595064 785790302933254963
69965
142902
0
1000
2000
3000
200 400 600 800 1000 1200mz
(d)
Inte
nsity
(au
)
1198658
174936
2619201154530
111976440964358918 932243609086
84213585996 325947 741110572984515028287928
400987
228936
200907
0
500
1000
1500
2000
2500
3000
200 400 600 800 1000 1200 1400mz
(e)
Inte
nsity
(au
)
1396813
12406611354732
174968
1070439867233227008111958 286957 1202691
513068417073354995
0
500
1000
1500
2000
2500
200 400 600 800 1000 1200 1400 1600mz
(f)
Figure 7The results of the MALDI-TOFTPF-MS analysis of protein spot 6 (a) Peptide mass fingerprinting and ((b)ndash(f)) peptide fragmentfingerprinting of spot 6
4 Discussion
In this study we found that the proliferative activities ofosteoblasts between HQSXD-S-treated group and JKSQW-control-S-treated group were similar This investigationdemonstrates that HQSXD can significantly facilitate boneformation through increasing the number of osteoblastswhich is beneficial to the treatment of osteoporosis By mea-suring ALP activity HQSXD was first screened for its abilityto induce osteogenesis HQSXD is capable of significantly
promoting osteoblast differentiation as well as increasingosteoblast mineralization
Results of the identification of the selected protein spotsare summarized in Table 2 The molecular weight (Mr) andisoelectric point (119875
119868) of each protein spot shown in Table 2
are theoretical values The eight protein spots were identifiedas (1) N-formyl peptide receptor 2 (FPR2) (2) alpha-spectrin(3) heparan sulfate (glucosamine) 3-O-sulfotransferase 3A1(HS3ST3A1) (4) prohibitin (PHB) (5) transcription elonga-tion factor B (SIII) polypeptide 1 (TCEB1) (6) chromosome
Evidence-Based Complementary and Alternative Medicine 9
TCEB1
PHB
FPR2
Blank HQSXD JKSQW
120573-actin
Alpha-spectrin
Figure 8 HQSXD-S treatment decreased alpha-spectrin PHBand TCEB1 expression and increased FPR2 expression Cell lysates(50 120583g)were processed forWestern blot analysis Normalization per-formed to120573-actinThe bands shown here were from a representativeexperiment repeated three times
segregation protein (7) nucleoside diphosphate kinase and(8) mast cell carboxypeptidase AWe detected some proteinsrelated to transcription cell proliferation differentiationand apoptosis such as FPR2 prohibitin alpha-spectrinheparan sulfate (glucosamine) 3-O-sulfotransferase 3A1 andtranscription elongation factor B (SIII) polypeptide 1 whichvaried greatly after HQSXD treatment
Our results suggest that HQSXD can upregulate theexpression of FPR2 N-Formyl peptide receptor (FPR) is aG protein-coupled receptor which modulates stromal celldifferentiation [14] and binds to N-formyl peptides suchas N-formyl-methionyl-leucyl-phenylalanine (fMLP) Ourstudy has shown that fMLP enhances the differentiationof MSCs into osteoblasts via an FPR-mediated signalingpathway and results in bone formation [15] The FPR2receptor belongs to the formyl peptide receptor family thatis involved in signaling stem cell adhesion migration andhoming for injured and inflamed tissues awaiting repair thiscould potentially be exploited to direct the stem cells to targetspecific tissue site [16] Therefore the current results implythat treatment with HQSXDmight promote the formation ofosteoblasts by upregulating the expression level of FPR2
We found that HQSXD could inhibit the expression ofalpha-spectrin Alpha-spectrin includes two genes encodingfor alpha-I subunit and alpha-II subunit each of thempresenting its specific cellular expression pattern Alpha-II spectrin deficiency is associated with cell proliferationdefects due to cell cycle arrest in theG1 phase (first gap phase)[17] Spectrin and protein kinase C theta were observed inaggregates during the early stage of apoptosis [18] Hencewe presume that alpha-spectrin is involved in the antiosteo-porotic effect of HQSXD
In this study PHB was downregulated in HQSXD-treated osteoblasts Prohibitin a highly conservative proteinregulates the cohesion of sister chromatids cellular signalingmitochondrial biogenesis [19] cell proliferation differentia-tion apoptosis and gene transcription [20 21] PHB blocksthe transition of cells from G1 phase to S phase of the cellcycle thereby arresting cell proliferation [22 23] PHB hasbeen reported to affect the apoptotic pathways by repressingthe transcriptional activity of E2F1 [24 25] As mentionedabove it is possible that HQSXD induces proliferationdifferentiation and apoptosis partly through downregulatingexpression of PHB in osteoblasts It is likely that PHB mightbe the novel candidate in the new antiosteoporotic drugscreening
The expression of TCEB1 was downregulated in HQSXD-treatment osteoblasts TCEB1 a 13 kDa protein also namedelongin C was originally identified as a member of themammalian transcription factor SIII that increases the rateof transcription by suppressing RNA polymerase II pausing[26] As part of a family of separate complexes containingelongin B and various substrate specificity factors it acts asan E3 ubiquitin ligase [27] Elongin B (ELB 1) and elonginC (ELC1) form a stable complex and that depletion of eithergene product by RNA-mediated interference (RNAi) causespronounced defects in the second meiotic division and arrestof germ cell proliferation in G1 [28] Therefore it is possiblethat that TCEB1 is involved in the antiosteoporotic effect ofHQSXD
Although this study has thrown some light on themechanism of HQSXD action we failed to characterize thewell-identified protein (eg HS3ST3A1) closely involved inosteoporosis HS3ST3A1 a member of the heparan sulfatebiosynthetic enzyme family possesses heparan sulfate glu-cosaminyl 3-O-sulfotransferase activity Depletion of Hs3st-A in enterocytes results in increased intestinal stem cellproliferation and tissue homeostasis loss [29]
5 Conclusions
The results confirm that HQSXD has a beneficial effect onosteoblasts and alters the expression level of some proteinsin osteoblasts The protein expressed by osteoblasts treatedwith HQSXD may be involved in cell proliferation anddifferentiation and other physiological processes and in theregulation of cell activation Further study is needed toinvestigate the effects of major active constituents in HQSXDon protein expression on osteoblast so as to demonstrate theinteraction and synergistic mechanism
Conflict of Interests
All the authors state that they have no conflict of interests todeclare
Authorsrsquo Contribution
Chong-Chong Guo and Li-Hua Zheng contributed equally tothis work
10 Evidence-Based Complementary and Alternative Medicine
Acknowledgments
The authors thank Cheng-Ming Liu for her excellent tech-nical assistance This work was supported by the grantfrom the National Natural Science foundation of China (no81273779)
References
[1] S Khosla and B L Riggs ldquoPathophysiology of age-related boneloss and osteoporosisrdquo Endocrinology andMetabolism Clinics ofNorth America vol 34 no 4 pp 1015ndash1030 2005
[2] P Sambrook andCCooper ldquoOsteoporosisrdquoTheLancet vol 367no 9527 pp 2010ndash2018 2006
[3] M R Allen and D B Burr ldquoThree years of alendronate treat-ment results in similar levels of vertebral microdamage as afterone year of treatmentrdquo Journal of Bone and Mineral Researchvol 22 no 11 pp 1759ndash1765 2007
[4] S Davison and S R Davis ldquoHormone replacement therapycurrent controversiesrdquo Clinical Endocrinology vol 58 no 3 pp249ndash261 2003
[5] W-Y Jiang ldquoTherapeutic wisdom in traditional Chinesemedicine a perspective from modern sciencerdquo Trends in Phar-macological Sciences vol 26 no 11 pp 558ndash563 2005
[6] Z Xu and Z K Zhou ldquoClinical study on the treatment of 36cases of postmenopausal osteoporosis with HuangQiSanXianTangrdquo Guiding Journal of Traditional Chinese Medicine andPharmacy vol 15 no 1 pp 9ndash11 2009
[7] H Zhang W-W Xing Y-S Li et al ldquoEffects of a traditionalChinese herbal preparation on osteoblasts and osteoclastsrdquoMaturitas vol 61 no 4 pp 334ndash339 2008
[8] L Qin T Han Q Zhang et al ldquoAntiosteoporotic chemical con-stituents from Er-Xian Decoction a traditional Chinese herbalformulardquo Journal of Ethnopharmacology vol 118 no 2 pp 271ndash279 2008
[9] M Millikan A Kolasani and H Xu ldquoDetermination and com-parison of mineral elements in traditional Chinese herbalformulae at different decoction times used to improve kidneyfunctionmdashchemometric approachrdquo African Journal of Tradi-tional Complementary and Alternative Medicines vol 8 no 5pp 191ndash197 2011
[10] I R Orriss S E B Taylor and T R Arnett ldquoRat osteoblastculturesrdquoMethods inMolecular Biology vol 816 pp 31ndash41 2012
[11] JM Baek J-Y Kim Y-H Cheon et al ldquoDual effect ofChrysan-themum indicum extract to stimulate osteoblast differentia-tion and inhibit osteoclast formation and resorption in vitrordquoEvidence-Based Complementary and Alternative Medicine vol2014 Article ID 176049 13 pages 2014
[12] A I Idris I R Greig E Bassonga-Landao S H Ralston andR J Vanrsquot Hof ldquoIdentification of novel biphenyl carboxylic acidderivatives as novel antiresorptive agents that do not impairparathyroid hormone-induced bone formationrdquo Endocrinologyvol 150 no 1 pp 5ndash13 2009
[13] M Otani J Tabata T Ueki K Sano and S Inouye ldquoHeat-shock-induced proteins from Myxococcus xanthusrdquo Journal ofBacteriology vol 183 no 21 pp 6282ndash6287 2001
[14] M E Nuttall and J M Gimble ldquoIs there a therapeutic opportu-nity to either prevent or treat osteopenic disorders by inhibitingmarrow adipogenesisrdquo Bone vol 27 no 2 pp 177ndash184 2000
[15] M K Shin Y H Jang H J Yoo et al ldquoN-formyl-methionyl-leucyl-phenylalanine (fMLP) promotes osteoblast differentia-tion via the N-formyl peptide receptor 1-mediated signaling
pathway in humanmesenchymal stem cells from bonemarrowrdquoThe Journal of Biological Chemistry vol 286 no 19 pp 17133ndash17143 2011
[16] A Viswanathan R G Painter N A Lanson Jr and G WangldquoFunctional expression ofN-formyl peptide receptors in humanbone marrow-derived mesenchymal stem cellsrdquo Stem Cells vol25 no 5 pp 1263ndash1269 2007
[17] S Metral B Machnicka S Bigot Y Colin D Dhermy and M-C Lecomte ldquo120572II-spectrin is critical for cell adhesion and cellcyclerdquo Journal of Biological Chemistry vol 284 no 4 pp 2409ndash2418 2009
[18] P M Dubielecka M Grzybek A Kolondra et alldquoAggregation of spectrin and PKCtheta is an early hallmarkof fludarabinemitoxantronedexamethasone-induced apopto-sis in Jurkat T and HL60 cellsrdquo Molecular and CellularBiochemistry vol 339 no 1-2 pp 63ndash77 2010
[19] C Merkwirth and T Langer ldquoProhibitin function withinmitochondria essential roles for cell proliferation and cristaemorphogenesisrdquo Biochimica et Biophysica Acta vol 1793 no 1pp 27ndash32 2009
[20] A L Theiss and S V Sitaraman ldquoThe role and therapeuticpotential of prohibitin in diseaserdquoBiochimica et BiophysicaActavol 1813 no 6 pp 1137ndash1143 2011
[21] V Sanchez-Quiles E Santamarıa V Segura L Sesma J Prietoand F J Corrales ldquoProhibitin deficiency blocks proliferationand induces apoptosis in human hepatoma cells molecularmechanisms and functional implicationsrdquo Proteomics vol 10no 8 pp 1609ndash1620 2010
[22] S Manjeshwar D E Branam M R Lerner D J Brackettand E R Jupe ldquoTumor suppression by the prohibitin gene3rsquountranslated region RNA in human breast cancerrdquo CancerResearch vol 63 no 17 pp 5251ndash5256 2003
[23] S Manjeshwar M R Lerner X-P Zang et al ldquoExpressionof prohibitin 31015840 untranslated region suppressor RNA altersmorphology and inhibits motility of breast cancer cellsrdquo Journalof Molecular Histology vol 35 no 6 pp 639ndash646 2004
[24] M Lacroix R-A Toillon and G Leclercq ldquop53 and breastcancer an updaterdquo Endocrine-Related Cancer vol 13 no 2 pp293ndash325 2006
[25] G Fusaro P Dasgupta S Rastogi B Joshi and S ChellappanldquoProhibitin induces the transcriptional activity of p53 andis exported from the nucleus upon apoptotic signalingrdquo TheJournal of Biological Chemistry vol 278 no 48 pp 47853ndash47861 2003
[26] T Aso W S Lane J W Conaway and R C ConawayldquoElongin (SIII) a multisubunit regulator of elongation by RNApolymerase IIrdquo Science vol 269 no 5229 pp 1439ndash1443 1995
[27] T D CumminsM DMendenhall M N Lowry et al ldquoElonginC is a mediator of Notch4 activity in human renal tubule cellsrdquoBiochimica et Biophysica Acta vol 1814 no 12 pp 1748ndash17572011
[28] Y Sasagawa K Kikuchi K Dazai and A HigashitanildquoCaenorhabditis elegans ElonginBC complex is essential for cellproliferation and chromosome condensation and segregationduring mitosis and meiotic division IIrdquo Chromosome Researchvol 13 no 4 pp 357ndash375 2005
[29] Y Guo Z Li and X Lin ldquoHs3st-A and Hs3st-B regulateintestinal homeostasis in Drosophila adult midgutrdquo CellularSignalling vol 26 no 11 pp 2317ndash2325 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 9
TCEB1
PHB
FPR2
Blank HQSXD JKSQW
120573-actin
Alpha-spectrin
Figure 8 HQSXD-S treatment decreased alpha-spectrin PHBand TCEB1 expression and increased FPR2 expression Cell lysates(50 120583g)were processed forWestern blot analysis Normalization per-formed to120573-actinThe bands shown here were from a representativeexperiment repeated three times
segregation protein (7) nucleoside diphosphate kinase and(8) mast cell carboxypeptidase AWe detected some proteinsrelated to transcription cell proliferation differentiationand apoptosis such as FPR2 prohibitin alpha-spectrinheparan sulfate (glucosamine) 3-O-sulfotransferase 3A1 andtranscription elongation factor B (SIII) polypeptide 1 whichvaried greatly after HQSXD treatment
Our results suggest that HQSXD can upregulate theexpression of FPR2 N-Formyl peptide receptor (FPR) is aG protein-coupled receptor which modulates stromal celldifferentiation [14] and binds to N-formyl peptides suchas N-formyl-methionyl-leucyl-phenylalanine (fMLP) Ourstudy has shown that fMLP enhances the differentiationof MSCs into osteoblasts via an FPR-mediated signalingpathway and results in bone formation [15] The FPR2receptor belongs to the formyl peptide receptor family thatis involved in signaling stem cell adhesion migration andhoming for injured and inflamed tissues awaiting repair thiscould potentially be exploited to direct the stem cells to targetspecific tissue site [16] Therefore the current results implythat treatment with HQSXDmight promote the formation ofosteoblasts by upregulating the expression level of FPR2
We found that HQSXD could inhibit the expression ofalpha-spectrin Alpha-spectrin includes two genes encodingfor alpha-I subunit and alpha-II subunit each of thempresenting its specific cellular expression pattern Alpha-II spectrin deficiency is associated with cell proliferationdefects due to cell cycle arrest in theG1 phase (first gap phase)[17] Spectrin and protein kinase C theta were observed inaggregates during the early stage of apoptosis [18] Hencewe presume that alpha-spectrin is involved in the antiosteo-porotic effect of HQSXD
In this study PHB was downregulated in HQSXD-treated osteoblasts Prohibitin a highly conservative proteinregulates the cohesion of sister chromatids cellular signalingmitochondrial biogenesis [19] cell proliferation differentia-tion apoptosis and gene transcription [20 21] PHB blocksthe transition of cells from G1 phase to S phase of the cellcycle thereby arresting cell proliferation [22 23] PHB hasbeen reported to affect the apoptotic pathways by repressingthe transcriptional activity of E2F1 [24 25] As mentionedabove it is possible that HQSXD induces proliferationdifferentiation and apoptosis partly through downregulatingexpression of PHB in osteoblasts It is likely that PHB mightbe the novel candidate in the new antiosteoporotic drugscreening
The expression of TCEB1 was downregulated in HQSXD-treatment osteoblasts TCEB1 a 13 kDa protein also namedelongin C was originally identified as a member of themammalian transcription factor SIII that increases the rateof transcription by suppressing RNA polymerase II pausing[26] As part of a family of separate complexes containingelongin B and various substrate specificity factors it acts asan E3 ubiquitin ligase [27] Elongin B (ELB 1) and elonginC (ELC1) form a stable complex and that depletion of eithergene product by RNA-mediated interference (RNAi) causespronounced defects in the second meiotic division and arrestof germ cell proliferation in G1 [28] Therefore it is possiblethat that TCEB1 is involved in the antiosteoporotic effect ofHQSXD
Although this study has thrown some light on themechanism of HQSXD action we failed to characterize thewell-identified protein (eg HS3ST3A1) closely involved inosteoporosis HS3ST3A1 a member of the heparan sulfatebiosynthetic enzyme family possesses heparan sulfate glu-cosaminyl 3-O-sulfotransferase activity Depletion of Hs3st-A in enterocytes results in increased intestinal stem cellproliferation and tissue homeostasis loss [29]
5 Conclusions
The results confirm that HQSXD has a beneficial effect onosteoblasts and alters the expression level of some proteinsin osteoblasts The protein expressed by osteoblasts treatedwith HQSXD may be involved in cell proliferation anddifferentiation and other physiological processes and in theregulation of cell activation Further study is needed toinvestigate the effects of major active constituents in HQSXDon protein expression on osteoblast so as to demonstrate theinteraction and synergistic mechanism
Conflict of Interests
All the authors state that they have no conflict of interests todeclare
Authorsrsquo Contribution
Chong-Chong Guo and Li-Hua Zheng contributed equally tothis work
10 Evidence-Based Complementary and Alternative Medicine
Acknowledgments
The authors thank Cheng-Ming Liu for her excellent tech-nical assistance This work was supported by the grantfrom the National Natural Science foundation of China (no81273779)
References
[1] S Khosla and B L Riggs ldquoPathophysiology of age-related boneloss and osteoporosisrdquo Endocrinology andMetabolism Clinics ofNorth America vol 34 no 4 pp 1015ndash1030 2005
[2] P Sambrook andCCooper ldquoOsteoporosisrdquoTheLancet vol 367no 9527 pp 2010ndash2018 2006
[3] M R Allen and D B Burr ldquoThree years of alendronate treat-ment results in similar levels of vertebral microdamage as afterone year of treatmentrdquo Journal of Bone and Mineral Researchvol 22 no 11 pp 1759ndash1765 2007
[4] S Davison and S R Davis ldquoHormone replacement therapycurrent controversiesrdquo Clinical Endocrinology vol 58 no 3 pp249ndash261 2003
[5] W-Y Jiang ldquoTherapeutic wisdom in traditional Chinesemedicine a perspective from modern sciencerdquo Trends in Phar-macological Sciences vol 26 no 11 pp 558ndash563 2005
[6] Z Xu and Z K Zhou ldquoClinical study on the treatment of 36cases of postmenopausal osteoporosis with HuangQiSanXianTangrdquo Guiding Journal of Traditional Chinese Medicine andPharmacy vol 15 no 1 pp 9ndash11 2009
[7] H Zhang W-W Xing Y-S Li et al ldquoEffects of a traditionalChinese herbal preparation on osteoblasts and osteoclastsrdquoMaturitas vol 61 no 4 pp 334ndash339 2008
[8] L Qin T Han Q Zhang et al ldquoAntiosteoporotic chemical con-stituents from Er-Xian Decoction a traditional Chinese herbalformulardquo Journal of Ethnopharmacology vol 118 no 2 pp 271ndash279 2008
[9] M Millikan A Kolasani and H Xu ldquoDetermination and com-parison of mineral elements in traditional Chinese herbalformulae at different decoction times used to improve kidneyfunctionmdashchemometric approachrdquo African Journal of Tradi-tional Complementary and Alternative Medicines vol 8 no 5pp 191ndash197 2011
[10] I R Orriss S E B Taylor and T R Arnett ldquoRat osteoblastculturesrdquoMethods inMolecular Biology vol 816 pp 31ndash41 2012
[11] JM Baek J-Y Kim Y-H Cheon et al ldquoDual effect ofChrysan-themum indicum extract to stimulate osteoblast differentia-tion and inhibit osteoclast formation and resorption in vitrordquoEvidence-Based Complementary and Alternative Medicine vol2014 Article ID 176049 13 pages 2014
[12] A I Idris I R Greig E Bassonga-Landao S H Ralston andR J Vanrsquot Hof ldquoIdentification of novel biphenyl carboxylic acidderivatives as novel antiresorptive agents that do not impairparathyroid hormone-induced bone formationrdquo Endocrinologyvol 150 no 1 pp 5ndash13 2009
[13] M Otani J Tabata T Ueki K Sano and S Inouye ldquoHeat-shock-induced proteins from Myxococcus xanthusrdquo Journal ofBacteriology vol 183 no 21 pp 6282ndash6287 2001
[14] M E Nuttall and J M Gimble ldquoIs there a therapeutic opportu-nity to either prevent or treat osteopenic disorders by inhibitingmarrow adipogenesisrdquo Bone vol 27 no 2 pp 177ndash184 2000
[15] M K Shin Y H Jang H J Yoo et al ldquoN-formyl-methionyl-leucyl-phenylalanine (fMLP) promotes osteoblast differentia-tion via the N-formyl peptide receptor 1-mediated signaling
pathway in humanmesenchymal stem cells from bonemarrowrdquoThe Journal of Biological Chemistry vol 286 no 19 pp 17133ndash17143 2011
[16] A Viswanathan R G Painter N A Lanson Jr and G WangldquoFunctional expression ofN-formyl peptide receptors in humanbone marrow-derived mesenchymal stem cellsrdquo Stem Cells vol25 no 5 pp 1263ndash1269 2007
[17] S Metral B Machnicka S Bigot Y Colin D Dhermy and M-C Lecomte ldquo120572II-spectrin is critical for cell adhesion and cellcyclerdquo Journal of Biological Chemistry vol 284 no 4 pp 2409ndash2418 2009
[18] P M Dubielecka M Grzybek A Kolondra et alldquoAggregation of spectrin and PKCtheta is an early hallmarkof fludarabinemitoxantronedexamethasone-induced apopto-sis in Jurkat T and HL60 cellsrdquo Molecular and CellularBiochemistry vol 339 no 1-2 pp 63ndash77 2010
[19] C Merkwirth and T Langer ldquoProhibitin function withinmitochondria essential roles for cell proliferation and cristaemorphogenesisrdquo Biochimica et Biophysica Acta vol 1793 no 1pp 27ndash32 2009
[20] A L Theiss and S V Sitaraman ldquoThe role and therapeuticpotential of prohibitin in diseaserdquoBiochimica et BiophysicaActavol 1813 no 6 pp 1137ndash1143 2011
[21] V Sanchez-Quiles E Santamarıa V Segura L Sesma J Prietoand F J Corrales ldquoProhibitin deficiency blocks proliferationand induces apoptosis in human hepatoma cells molecularmechanisms and functional implicationsrdquo Proteomics vol 10no 8 pp 1609ndash1620 2010
[22] S Manjeshwar D E Branam M R Lerner D J Brackettand E R Jupe ldquoTumor suppression by the prohibitin gene3rsquountranslated region RNA in human breast cancerrdquo CancerResearch vol 63 no 17 pp 5251ndash5256 2003
[23] S Manjeshwar M R Lerner X-P Zang et al ldquoExpressionof prohibitin 31015840 untranslated region suppressor RNA altersmorphology and inhibits motility of breast cancer cellsrdquo Journalof Molecular Histology vol 35 no 6 pp 639ndash646 2004
[24] M Lacroix R-A Toillon and G Leclercq ldquop53 and breastcancer an updaterdquo Endocrine-Related Cancer vol 13 no 2 pp293ndash325 2006
[25] G Fusaro P Dasgupta S Rastogi B Joshi and S ChellappanldquoProhibitin induces the transcriptional activity of p53 andis exported from the nucleus upon apoptotic signalingrdquo TheJournal of Biological Chemistry vol 278 no 48 pp 47853ndash47861 2003
[26] T Aso W S Lane J W Conaway and R C ConawayldquoElongin (SIII) a multisubunit regulator of elongation by RNApolymerase IIrdquo Science vol 269 no 5229 pp 1439ndash1443 1995
[27] T D CumminsM DMendenhall M N Lowry et al ldquoElonginC is a mediator of Notch4 activity in human renal tubule cellsrdquoBiochimica et Biophysica Acta vol 1814 no 12 pp 1748ndash17572011
[28] Y Sasagawa K Kikuchi K Dazai and A HigashitanildquoCaenorhabditis elegans ElonginBC complex is essential for cellproliferation and chromosome condensation and segregationduring mitosis and meiotic division IIrdquo Chromosome Researchvol 13 no 4 pp 357ndash375 2005
[29] Y Guo Z Li and X Lin ldquoHs3st-A and Hs3st-B regulateintestinal homeostasis in Drosophila adult midgutrdquo CellularSignalling vol 26 no 11 pp 2317ndash2325 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Evidence-Based Complementary and Alternative Medicine
Acknowledgments
The authors thank Cheng-Ming Liu for her excellent tech-nical assistance This work was supported by the grantfrom the National Natural Science foundation of China (no81273779)
References
[1] S Khosla and B L Riggs ldquoPathophysiology of age-related boneloss and osteoporosisrdquo Endocrinology andMetabolism Clinics ofNorth America vol 34 no 4 pp 1015ndash1030 2005
[2] P Sambrook andCCooper ldquoOsteoporosisrdquoTheLancet vol 367no 9527 pp 2010ndash2018 2006
[3] M R Allen and D B Burr ldquoThree years of alendronate treat-ment results in similar levels of vertebral microdamage as afterone year of treatmentrdquo Journal of Bone and Mineral Researchvol 22 no 11 pp 1759ndash1765 2007
[4] S Davison and S R Davis ldquoHormone replacement therapycurrent controversiesrdquo Clinical Endocrinology vol 58 no 3 pp249ndash261 2003
[5] W-Y Jiang ldquoTherapeutic wisdom in traditional Chinesemedicine a perspective from modern sciencerdquo Trends in Phar-macological Sciences vol 26 no 11 pp 558ndash563 2005
[6] Z Xu and Z K Zhou ldquoClinical study on the treatment of 36cases of postmenopausal osteoporosis with HuangQiSanXianTangrdquo Guiding Journal of Traditional Chinese Medicine andPharmacy vol 15 no 1 pp 9ndash11 2009
[7] H Zhang W-W Xing Y-S Li et al ldquoEffects of a traditionalChinese herbal preparation on osteoblasts and osteoclastsrdquoMaturitas vol 61 no 4 pp 334ndash339 2008
[8] L Qin T Han Q Zhang et al ldquoAntiosteoporotic chemical con-stituents from Er-Xian Decoction a traditional Chinese herbalformulardquo Journal of Ethnopharmacology vol 118 no 2 pp 271ndash279 2008
[9] M Millikan A Kolasani and H Xu ldquoDetermination and com-parison of mineral elements in traditional Chinese herbalformulae at different decoction times used to improve kidneyfunctionmdashchemometric approachrdquo African Journal of Tradi-tional Complementary and Alternative Medicines vol 8 no 5pp 191ndash197 2011
[10] I R Orriss S E B Taylor and T R Arnett ldquoRat osteoblastculturesrdquoMethods inMolecular Biology vol 816 pp 31ndash41 2012
[11] JM Baek J-Y Kim Y-H Cheon et al ldquoDual effect ofChrysan-themum indicum extract to stimulate osteoblast differentia-tion and inhibit osteoclast formation and resorption in vitrordquoEvidence-Based Complementary and Alternative Medicine vol2014 Article ID 176049 13 pages 2014
[12] A I Idris I R Greig E Bassonga-Landao S H Ralston andR J Vanrsquot Hof ldquoIdentification of novel biphenyl carboxylic acidderivatives as novel antiresorptive agents that do not impairparathyroid hormone-induced bone formationrdquo Endocrinologyvol 150 no 1 pp 5ndash13 2009
[13] M Otani J Tabata T Ueki K Sano and S Inouye ldquoHeat-shock-induced proteins from Myxococcus xanthusrdquo Journal ofBacteriology vol 183 no 21 pp 6282ndash6287 2001
[14] M E Nuttall and J M Gimble ldquoIs there a therapeutic opportu-nity to either prevent or treat osteopenic disorders by inhibitingmarrow adipogenesisrdquo Bone vol 27 no 2 pp 177ndash184 2000
[15] M K Shin Y H Jang H J Yoo et al ldquoN-formyl-methionyl-leucyl-phenylalanine (fMLP) promotes osteoblast differentia-tion via the N-formyl peptide receptor 1-mediated signaling
pathway in humanmesenchymal stem cells from bonemarrowrdquoThe Journal of Biological Chemistry vol 286 no 19 pp 17133ndash17143 2011
[16] A Viswanathan R G Painter N A Lanson Jr and G WangldquoFunctional expression ofN-formyl peptide receptors in humanbone marrow-derived mesenchymal stem cellsrdquo Stem Cells vol25 no 5 pp 1263ndash1269 2007
[17] S Metral B Machnicka S Bigot Y Colin D Dhermy and M-C Lecomte ldquo120572II-spectrin is critical for cell adhesion and cellcyclerdquo Journal of Biological Chemistry vol 284 no 4 pp 2409ndash2418 2009
[18] P M Dubielecka M Grzybek A Kolondra et alldquoAggregation of spectrin and PKCtheta is an early hallmarkof fludarabinemitoxantronedexamethasone-induced apopto-sis in Jurkat T and HL60 cellsrdquo Molecular and CellularBiochemistry vol 339 no 1-2 pp 63ndash77 2010
[19] C Merkwirth and T Langer ldquoProhibitin function withinmitochondria essential roles for cell proliferation and cristaemorphogenesisrdquo Biochimica et Biophysica Acta vol 1793 no 1pp 27ndash32 2009
[20] A L Theiss and S V Sitaraman ldquoThe role and therapeuticpotential of prohibitin in diseaserdquoBiochimica et BiophysicaActavol 1813 no 6 pp 1137ndash1143 2011
[21] V Sanchez-Quiles E Santamarıa V Segura L Sesma J Prietoand F J Corrales ldquoProhibitin deficiency blocks proliferationand induces apoptosis in human hepatoma cells molecularmechanisms and functional implicationsrdquo Proteomics vol 10no 8 pp 1609ndash1620 2010
[22] S Manjeshwar D E Branam M R Lerner D J Brackettand E R Jupe ldquoTumor suppression by the prohibitin gene3rsquountranslated region RNA in human breast cancerrdquo CancerResearch vol 63 no 17 pp 5251ndash5256 2003
[23] S Manjeshwar M R Lerner X-P Zang et al ldquoExpressionof prohibitin 31015840 untranslated region suppressor RNA altersmorphology and inhibits motility of breast cancer cellsrdquo Journalof Molecular Histology vol 35 no 6 pp 639ndash646 2004
[24] M Lacroix R-A Toillon and G Leclercq ldquop53 and breastcancer an updaterdquo Endocrine-Related Cancer vol 13 no 2 pp293ndash325 2006
[25] G Fusaro P Dasgupta S Rastogi B Joshi and S ChellappanldquoProhibitin induces the transcriptional activity of p53 andis exported from the nucleus upon apoptotic signalingrdquo TheJournal of Biological Chemistry vol 278 no 48 pp 47853ndash47861 2003
[26] T Aso W S Lane J W Conaway and R C ConawayldquoElongin (SIII) a multisubunit regulator of elongation by RNApolymerase IIrdquo Science vol 269 no 5229 pp 1439ndash1443 1995
[27] T D CumminsM DMendenhall M N Lowry et al ldquoElonginC is a mediator of Notch4 activity in human renal tubule cellsrdquoBiochimica et Biophysica Acta vol 1814 no 12 pp 1748ndash17572011
[28] Y Sasagawa K Kikuchi K Dazai and A HigashitanildquoCaenorhabditis elegans ElonginBC complex is essential for cellproliferation and chromosome condensation and segregationduring mitosis and meiotic division IIrdquo Chromosome Researchvol 13 no 4 pp 357ndash375 2005
[29] Y Guo Z Li and X Lin ldquoHs3st-A and Hs3st-B regulateintestinal homeostasis in Drosophila adult midgutrdquo CellularSignalling vol 26 no 11 pp 2317ndash2325 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom


















