![Acorus tatarinowii Schott extract reduces cerebral edema ......cerebral edema [11, 12]. Thus, the expression of glial fi-brillary acidic protein (GFAP), a marker of reactive astrogliosis,](https://static.fdocuments.in/doc/165x107/60f9fb03b1d27d0bb6581189/acorus-tatarinowii-schott-extract-reduces-cerebral-edema-cerebral-edema.jpg)
Research Article Anorexia Reduces GFAP+ Cell Density in...
12
Research Article Anorexia Reduces GFAP+ Cell Density in the Rat Hippocampus Daniel Reyes-Haro, Francisco Emmanuel Labrada-Moncada, Durairaj Ragu Varman, Janina Krüger, Teresa Morales, Ricardo Miledi, and Ataúlfo Martínez-Torres Departamento de Neurobiolog´ ıa Celular y Molecular, Instituto de Neurobiolog´ ıa, Universidad Nacional Aut´ onoma de M´ exico, Campus Juriquilla, Boulevard Universitario 3001, 76230 Juriquilla, QRO, Mexico Correspondence should be addressed to Daniel Reyes-Haro; [email protected] Received 15 February 2016; Revised 18 May 2016; Accepted 16 June 2016 Academic Editor: Christian Wozny Copyright © 2016 Daniel Reyes-Haro et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Anorexia nervosa is an eating disorder observed primarily in young women. e neurobiology of the disorder is unknown but recently magnetic resonance imaging showed a volume reduction of the hippocampus in anorexic patients. Dehydration-induced anorexia (DIA) is a murine model that mimics core features of this disorder, including severe weight loss due to voluntary reduction in food intake. e energy supply to the brain is mediated by astrocytes, but whether their density is compromised by anorexia is unknown. us, the aim of this study was to estimate GFAP+ cell density in the main regions of the hippocampus (CA1, CA2, CA3, and dentate gyrus) in the DIA model. Our results showed that GFAP+ cell density was significantly reduced (∼20%) in all regions of the hippocampus, except in CA1. Interestingly, DIA significantly reduced the GFAP+ cells/nuclei ratio in CA2 (−23%) and dentate gyrus (−48%). e reduction of GFAP+ cell density was in agreement with a lower expression of GFAP protein. Additionally, anorexia increased the expression of the intermediate filaments vimentin and nestin. Accordingly, anorexia increased the number of reactive astrocytes in CA2 and dentate gyrus more than twofold. We conclude that anorexia reduces the hippocampal GFAP+ cell density and increases vimentin and nestin expression. 1. Introduction Anorexia nervosa is an eating disorder characterized by excessively restricted caloric intake that induces profound weight loss, osteoporosis, and amenorrhea [1]. e onset of anorexia nervosa is commonly observed during puberty and adolescence, with 90–95% of the cases occurring among females [2]. Due to its complexity, the neurobiology of anorexia nervosa is unknown, but studies using magnetic resonance imaging showed hippocampal volume reduction [3–6]. e hippocampus is involved in spatial learning, cognition, and regulation of anxiety [7]. Alterations in these cognitive functions have been reported in patients and in experimen- tal models of anorexia. Murine models of anorexia such as dehydration-induced anorexia (DIA) or activity-based anorexia (ABA) mimic the characteristic weight loss and reduced food intake observed in anorexic patients [1, 2, 8– 13]. e smaller hippocampal volume in anorexic patients may reflect structural changes at the cellular level. In the ABA model, changes in the hippocampus included reduced cell proliferation in the dentate gyrus and decreased branching of dendrites in the stratum radiatum of CA1 [1, 11, 12]. Excessive physical activity alters food ingestion, which can result in nutrient and electrolyte imbalance. us, alterations in hippocampal function and structure could be the result of reduced caloric intake that compromises the supply of energy to the brain. Astrocytes, one of the major populations of cells in the central nervous system, play a crucial role in supplying energy to neurons; therefore, we tested if hippocampal astrocyte density and intermediate filament expression are affected by anorexia. 2. Materials and Methods 2.1. Animals and Housing. e Institutional Animal Care and Use of Laboratory Animals Committee of the UNAM Instituto de Neurobiolog´ ıa approved all the experimental protocols. Animals were handled in accordance with the National Institute of Health Guide for the Care and Use Hindawi Publishing Corporation Neural Plasticity Volume 2016, Article ID 2426413, 11 pages http://dx.doi.org/10.1155/2016/2426413
Transcript of Research Article Anorexia Reduces GFAP+ Cell Density in...

Research ArticleAnorexia Reduces GFAP+ Cell Density in the Rat Hippocampus
Daniel Reyes-Haro Francisco Emmanuel Labrada-Moncada Durairaj Ragu VarmanJanina Kruumlger Teresa Morales Ricardo Miledi and Atauacutelfo Martiacutenez-Torres
Departamento de Neurobiologıa Celular y Molecular Instituto de Neurobiologıa Universidad Nacional Autonoma de MexicoCampus Juriquilla Boulevard Universitario 3001 76230 Juriquilla QRO Mexico
Correspondence should be addressed to Daniel Reyes-Haro dharounammx
Received 15 February 2016 Revised 18 May 2016 Accepted 16 June 2016
Academic Editor Christian Wozny
Copyright copy 2016 Daniel Reyes-Haro et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Anorexia nervosa is an eating disorder observed primarily in young women The neurobiology of the disorder is unknown butrecently magnetic resonance imaging showed a volume reduction of the hippocampus in anorexic patients Dehydration-inducedanorexia (DIA) is amurinemodel thatmimics core features of this disorder including severe weight loss due to voluntary reductionin food intake The energy supply to the brain is mediated by astrocytes but whether their density is compromised by anorexia isunknownThus the aim of this study was to estimate GFAP+ cell density in the main regions of the hippocampus (CA1 CA2 CA3and dentate gyrus) in the DIAmodel Our results showed that GFAP+ cell density was significantly reduced (sim20) in all regions ofthe hippocampus except in CA1 Interestingly DIA significantly reduced the GFAP+ cellsnuclei ratio in CA2 (minus23) and dentategyrus (minus48) The reduction of GFAP+ cell density was in agreement with a lower expression of GFAP protein Additionallyanorexia increased the expression of the intermediate filaments vimentin and nestin Accordingly anorexia increased the numberof reactive astrocytes in CA2 and dentate gyrus more than twofold We conclude that anorexia reduces the hippocampal GFAP+cell density and increases vimentin and nestin expression
1 Introduction
Anorexia nervosa is an eating disorder characterized byexcessively restricted caloric intake that induces profoundweight loss osteoporosis and amenorrhea [1] The onsetof anorexia nervosa is commonly observed during pubertyand adolescence with 90ndash95 of the cases occurring amongfemales [2]
Due to its complexity the neurobiology of anorexianervosa is unknown but studies using magnetic resonanceimaging showed hippocampal volume reduction [3ndash6] Thehippocampus is involved in spatial learning cognition andregulation of anxiety [7] Alterations in these cognitivefunctions have been reported in patients and in experimen-tal models of anorexia Murine models of anorexia suchas dehydration-induced anorexia (DIA) or activity-basedanorexia (ABA) mimic the characteristic weight loss andreduced food intake observed in anorexic patients [1 2 8ndash13] The smaller hippocampal volume in anorexic patientsmay reflect structural changes at the cellular level In the ABA
model changes in the hippocampus included reduced cellproliferation in the dentate gyrus and decreased branchingof dendrites in the stratum radiatum of CA1 [1 11 12]Excessive physical activity alters food ingestion which canresult in nutrient and electrolyte imbalanceThus alterationsin hippocampal function and structure could be the result ofreduced caloric intake that compromises the supply of energyto the brain Astrocytes one of the major populations of cellsin the central nervous system play a crucial role in supplyingenergy to neurons therefore we tested if hippocampalastrocyte density and intermediate filament expression areaffected by anorexia
2 Materials and Methods
21 Animals and Housing The Institutional Animal Careand Use of Laboratory Animals Committee of the UNAMInstituto de Neurobiologıa approved all the experimentalprotocols Animals were handled in accordance with theNational Institute of Health Guide for the Care and Use
Hindawi Publishing CorporationNeural PlasticityVolume 2016 Article ID 2426413 11 pageshttpdxdoiorg10115520162426413
2 Neural Plasticity
of Laboratory Animals Wistar female rats (160ndash190 g) werehoused individually under 12-h12-h lightdark cycle andcontrolled temperature with food and water ad libitum
22 Dehydration-Induced Anorexia The protocol was per-formed as previously described [8 13 14] Briefly two inde-pendent experimental series of twelve animals were placedin individual cages for each series animals were randomlyselected to form three groups of fourThe first group receivedwater and food ad libitum (control) The DIA group receiveda 25 NaCl solution as their sole drinking liquid and hadunrestricted access to food The forced food-restricted (FFR)group a positive control to distinguish between starvationand dehydration effects received tap water ad libitum andthe same amount of food consumed by DIA animals Theexperimental protocol was conducted for five days and bodyweight and solid food intake were recorded daily at noonfor each experimental group The FFR group received thesame amount of food as that ingested by DIA animals (seeSupplemental Figure 1 in Supplementary Material availableonline at httpdxdoiorg10115520162426413)
23 Histology Rats were deeply anaesthetized with an over-dose of sodium pentobarbital (100mgKg) and transcardiallyperfused with 100mL of saline followed by 250mL of chilled4 paraformaldehyde in phosphate-buffered saline (PBS)(pH 74) Brains were removed postfixed overnight and thentransferred to a series of sucrose solutions (from 10 to 30)Coronal sections (30 120583m) including the dorsal hippocampuswere obtained on a freezing microtome collected and storedin cryoprotectant solution (30 ethylene glycol20 glycerolin PBS) at minus20∘C [13]
24 Immunohistofluorescence Glial fibrillary acidic protein(GFAP) immunoreactivity was performed on floating sec-tions [13] Briefly coronal sections were rinsed three timesin PBS buffer and treated with 3 hydrogen peroxide for10min followed by three rinses in PBS and incubation in 10sodium borohydride for 6ndash8min to reduce free aldehydesThe sections were incubated for 1 h in blocking solution (5horse serum albumin1 Triton X-100 in PBS) Sections wereincubated for 48 h with polyclonal rabbit anti-GFAP anti-body (dilution 1 1000 DakoCytomation Fort Collins COUSA 4∘C) After washing primary antibody was detectedwith Alexa 594 (1 500 Invitrogen) coupled goat anti-rabbitsecondary antibody The sections were counterstained with410158406-diamidino-2-phenylindole (DAPI) and mounted withVectashield H-1000 (Vector Laboratories Burlingame CAUSA)
Mounted coronal sections containing the hippocampuswere photographed with a digital camera (Photometrics CoolSnap FX USA) attached to a Nikon microscope (NikonEclipse E600 Tokyo Japan) and analyzed using IMAGEJ version 141 (NIH Bethesda MD USA) A Zeiss LSM780 Meta Confocal Microscope (Zeiss Gottingen Germany)was used for confocal images with Alexa 594 (excita-tionemission wavelength 590617 nm) and DAPI (excita-tionemission wavelength 350460 nm) (Figure 1)
CA1CA2
CA3
DG
spsr
so
SGZ
GCL
CA1CA2
CA3
SGZ
FFR
Control
CA1
CA2
CA3
SGZ
D
VL L
DIA
spsr
so
spsr
so
D
VL L
D
VL L
DG
DG
GCL
GCL
lowast
lowast
lowast
Figure 1 The rat hippocampus Coronal brain sections from youngfemale rats showing the hippocampus of control dehydration-induced anorexia (DIA) and forced food-restricted (FFR) ratsThe histological sections from the three experimental groups wereimmunostained for GFAP (green) and nuclei were labeled withDAPI (blue) The hippocampal regions for this study included thecornus ammonis 1ndash3 (CA1 CA2 and CA3) and the dentate gyrus(DG) Nuclear and astrocyte densities were estimated in the stratumoriens (so) and stratum radiatum (sr) of CA1 CA2 andCA3whilethe hilus region (lowast) was studied for the DG (squares) Other regionsare indicated for anatomical reference stratum pyramidale (sp)granular cell layer (GCL) and subgranular zone (SGZ) Arrowsshow anatomical orientation dorsal (D) ventral (V) and lateral (L)Scale bars correspond to 200 120583m
25 Cell Counting GFAP-immunolabeled cells were countedand contrasted with the number of DAPI-labeled nuclei foreach subfield of the hippocampus CA1 CA2 CA3 anddentate gyrus (DG) A total of 2-3 aleatory fields in each ofthe three tissue sections selected from each of 6ndash8 animalsper group were used for cell counting and ldquonrdquo refers to thetotal aleatory fields used from all the tissue sections from allthe animals of each experimental group A test square grid of100 times 100 120583m (001mm2) was used to estimate the number of
Neural Plasticity 3
Table 1 Regional changes in astrocyte density in DIA and FFR
Hippocampal region Control DIA FFR 119901
NucleiCA1 1099 plusmn 52 (119899 = 62) 924 plusmn 67 (119899 = 56) 932 plusmn 68 (119899 = 52) 0075CA2 1125 plusmn 101 (119899 = 40) 1007 plusmn 51 (119899 = 40) 1022 plusmn 56 (119899 = 36) 0467CA3 1288 plusmn 43 (119899 = 62) 1084 plusmn 41lowastlowastlowast (119899 = 56) 1117 plusmn 46lowastlowastlowast (119899 = 52) 0001DGSGZ 1433 plusmn 110 (119899 = 21) 1535 plusmn 142 (119899 = 20) 1268 plusmn 161 (119899 = 19) 0398
AstrocytesCA1 257 plusmn 14 (119899 = 82) 253 plusmn 14 (119899 = 72) 221 plusmn 15 (119899 = 66) 0172CA2 405 plusmn 19 (119899 = 40) 323 plusmn 18lowastlowastlowast (119899 = 40) 267 plusmn 19lowastlowastlowast (119899 = 36) lt0001CA3 332 plusmn 14 (119899 = 82) 269 plusmn 15lowastlowastlowast (119899 = 72) 258 plusmn 15lowastlowastlowast (119899 = 66) lt0001DGSGZ 476 plusmn 28 (119899 = 41) 294 plusmn 24lowastlowastlowast (119899 = 36) 321 plusmn 26lowastlowastlowast (119899 = 32) lt0001
Astrocytesnuclei ratioCA1 0277 plusmn 0025 (119899 = 62) 0324 plusmn 0025 (119899 = 56) 0286 plusmn 0029 (119899 = 52) 0417CA2 0447 plusmn 0053 (119899 = 40) 0345 plusmn 0021lowastlowastlowast (119899 = 40) 0282 plusmn 0024lowastlowastlowast (119899 = 36) 0008CA3 0292 plusmn 0016 (119899 = 62) 0262 plusmn 0016 (119899 = 56) 0264 plusmn 0016 (119899 = 52) 0347DGSGZ 0334 plusmn 0031 (119899 = 21) 0173 plusmn 0022lowastlowastlowast (119899 = 20) 0139 plusmn 0032lowastlowastlowast (119899 = 19) lt0001
DAPI-stained nuclei (equivalent to the total number of cells)and GFAP-labeled cells corresponding to astrocytes manu-ally and with Cell Profiler [13 15] Only process-bearing cellsshowing their soma in the plane of the analyzed area werecounted and their density was estimated (number per mm2)Six to eight randomly chosen fields located within each of theanalyzed hippocampal subfields were counted by progressivedisplacement of a test square gridThe astrocytesnuclei ratiowas calculated by dividing the number of GFAP+ cells bythe total number of nuclei labeled with DAPI GFAP+ cellswere identified as resting (with small somata bearing longthin and ramified processes) or reactive (with retraction ofprocesses to a length shorter than the diameter of the somata)[16] The number of reactive astrocytes was estimated for allthe experimental groups and normalized with the control
All the data corresponding to each treatment and regionwere pooled and are presented as the mean plusmn standard errorof the mean (SEM) Statistical analysis of data was performedusing a one-way ANOVA followed by a Bonferroni posttestwith Origin 70 software 119901 lt 005was considered statisticallysignificant
26Western Blot Thehippocampuswas dissected out as pre-viously described [17] The hippocampi from three differentrats for each experimental group (control DIA and FFR)were homogenized in ice cold lysis buffer (200mM glycine150mM NaCl 50mM EGTA 50mM EDTA and 300mMsucrose) with a protease inhibitor (Sigma-Aldrich USA)The homogenate was centrifuged at 10000timesg for 15 minutesat 4∘C and the supernatants were aliquoted and stored atminus80∘C Concentration of each protein sample was estimatedby Bradfordrsquos method [18] An equal concentration of protein(30 120583g) was resolved by 10 polyacrylamide gel electrophore-sis (PAGE)The proteins were transferred electrophoreticallyto PVDFmembrane then the membranes were blocked with5 nonfat dry milk in TBS-T for 3 h at room temperature
(RT) Membranes were incubated at 4∘C overnight withone of the following primary antibodies (a) polyclonalrabbit anti-GFAP antibody (dilution 1 2000 DakoCytoma-tion Fort Collins CO USA) (b) polyclonal rabbit anti-vimentin (dilution 1 2500 Cell Signaling Danvers MAUSA) (c) monoclonal mouse anti-nestin (dilution 1 5000BD Biosciences San Jose CA USA) and (d) polyclonal goatanti-actin (dilution 1 2500 Santacruz Dallas TX USA)The membranes were washed and bound antibodies weredetected by incubating for 3 h with either the goat anti-rabbitalkaline phosphatase conjugated antibody (dilution 1 2500Santacruz Dallas TX) goat anti-mouse alkaline phosphataseconjugated antibody (dilution 1 2500 Santacruz DallasTX) or rabbit anti-goat alkaline phosphatase conjugatedantibody (dilution 1 2500 Santacruz Dallas TX) Then themembrane was washed with TBS-T and the alkaline phos-phatase activity was detected with BCIPNBT AP-conjugatesubstrate reaction Kit (Bio-Rad USA) The intensity of eachband was measured using Image Lab 30 software
3 Results
The effects of DIA and FFR on the density of nuclei andastrocytes were estimated for the following hippocampalregions CA1 CA2 CA3 and dentate gyrus (DG) (Figure 1)All observations included the stratum radiatum and stratumoriens where astrocytes are preferentially located whilefor the DG the observations were performed in the hilus(Figure 1)
31 Anorexia Had No Effect on GFAP+ Cell Density inCA1 The hippocampal region CA1 showed no significantchange in nuclear density between the control (1099 plusmn 52nucleimm2 119899 = 62) DIA (924 plusmn 67 nucleimm2 119899 = 56)and FFR groups (932 plusmn 68 nucleimm2 119899 = 52) (119901 = 0075)(Figure 1 and Table 1) Additionally no significant changes
4 Neural Plasticity
500
1000
Nuc
leim
m 2
CTRL
DIA
FFR
DAPIso
sp
sp
sp
sr
so
sr
so
sr
Control DIA FFR(a)
GFAPso
sr
so
sr
so
sr
CTRL
DIA
FFR
100
200
300
Astr
ocyt
esm
m 2
Control DIA FFR
sp
sp
sp
(b)
Ratio
astro
cyte
snu
clei
02
04
Mergeso
sr
so
sr
so
sr
CTRL
DIA
FFR
Control DIA FFR
sp
sp
sp
(c)
Figure 2 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA1 Coronal sections of thehippocampus show nuclei labeled with DAPI (a) or immunostained with GFAP (b) The overlay of DAPI and GFAP is shown for controldehydration-induced anorexia (DIA) and forced food restricted (FFR) experimental groups (c)The densities of nuclei or GFAP+ cells as wellas the ratio of astrocytenuclei did not differ significantly among experimental groups (119901 = 0075 119901 = 0172 119901 = 0417 for the control DIAand FFR groups respectively see Table 1) Nuclear and astrocyte densities were estimated in the stratum oriens (so) and stratum radiatum(sr) stratum pyramidale (sp) is indicated as a reference Scale bar = 50 120583m Data are mean plusmn SEM
were observed for astrocyte density between the control(257 plusmn 14 nucleimm2 119899 = 82) DIA (253 plusmn 14 nucleimm2119899 = 72) and FFR groups (221 plusmn 15 nucleimm2 119899 = 66)(119901 = 0172) (Figure 2 and Table 1) We also determined theastrocytenuclei ratio by estimating the GFAP+ cells from thetotal nuclei The astrocytenuclei ratio for the control groupwas 0277 plusmn 0025 (119899 = 62) and it was not significantlydifferent for the DIA and FFR groups (0324 plusmn 0025 119899 = 56and 0286 plusmn 0029 119899 = 52 119901 = 0417) (Figure 2 and Table 1)
32 Anorexia Reduced GFAP+ Cell Density in CA2 Nucleardensity in CA2 was similar for the control (1125 plusmn 101nucleimm2 119899 = 40) DIA (1007 plusmn 51 nucleimm2 119899 = 40)
and FFR groups (1022 plusmn 56 nucleimm2 119899 = 36) (119901 = 0467)(Figure 3 Table 1) In contrast the astrocyte density estimatedfor the control group (405 plusmn 19 astrocytesmm2 119899 = 40) wassignificantly reduced for the DIA (323 plusmn 18 astrocytesmm2119899 = 40) and FFR groups (267 plusmn 19 astrocytemm2 119899 =36) (119901 lt 0001) (Figure 3 Table 1) Accordingly theastrocytenuclei ratio estimated for the control group (0447plusmn0053 119899 = 40) was also significantly reduced for the DIA(0345plusmn0021 119899 = 40) and FFR groups (0282plusmn0024 119899 = 36)(119901 = 0008) (Figure 3 Table 1)
33 Anorexia Reduced GFAP+ Cell Density in CA3 Thenuclear density of CA3 did not show significant changes
Neural Plasticity 5
DAPI
so
sp
sp
sp
sr
so
sr
so
sr
CTRL
DIA
FFR
500
1000
Nuc
leim
m 2
Control DIA FFR(a)
GFAP
so
sr
so
sr
so
sr
CTRL
DIA
FFR
200
400
Astr
ocyt
esm
m 2
lowastlowastlowast
lowastlowastlowast
Control DIA FFR
sp
sp
sp
(b)
Merge
so
sr
so
sr
so
sr
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
06
lowastlowastlowast
lowastlowastlowast
Control DIA FFR
sp
sp
sp
(c)
Figure 3 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA2 Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is shown for control dehydration-inducedanorexia (DIA) and forced-food restricted (FFR) experimental groups (c) The density of the nuclei was similar in all experimental groups(119901 = 0467) However a significant reduction of GFAP+ cells was observed for the DIA and FFR groups (119901 lt 0001) This reduction wasconsistently observed in the astrocytenuclei ratio (119901 = 0008) (see Table 1) Nuclear and astrocyte densities were estimated in the stratumoriens (so) and stratum radiatum (sr) stratum pyramidale (sp) is indicated as an anatomical reference Scale bar = 50120583m Data areexpressed as mean plusmn SEM
between the control (1288 plusmn 43 nucleimm2 119899 = 62) DIA(1084 plusmn 41 nucleimm2 119899 = 56) and FFR groups (1117 plusmn46 nucleimm2 119899 = 52) (119901 = 0001) (Figure 4 Table 1)However the astrocyte density of the control group (332plusmn 19astrocytesmm2 119899 = 82) was significantly reduced for theDIA (269plusmn15 astrocytesmm2 119899 = 72) and FFR groups (258plusmn15 astrocytemm2 119899 = 66) (119901 lt 0001) (Figure 4 Table 1)However the astrocytenuclei ratio estimated for the controlgroup (0292plusmn0016 119899 = 62) was not significantly reduced intheDIA (0262plusmn0016 119899 = 56) and FFR groups (0264plusmn0016119899 = 52) (119901 = 0347) (Figure 4 Table 1)
34 Anorexia Reduced GFAP+ Cell Density in DG Thenuclear density of the dentate gyrus did not differ signifi-cantly for the control (1433 plusmn 110 nucleimm2 119899 = 21)DIA (1535 plusmn 142 nucleimm2 119899 = 20) and FFR groups(1268 plusmn 161 nucleimm2 119899 = 19) (119901 = 0398) (Figure 5Table 1) The astrocyte density of the control group (476 plusmn 28astrocytesmm2 119899 = 41) was significantly reduced for theDIA (294 plusmn 124 astrocytesmm2 119899 = 36) and FFR groups(321 plusmn 26 astrocytemm2 119899 = 32) (119901 lt 0001) (Figure 5Table 1) Accordingly the astrocytenuclei ratio estimated forthe control group (0334 plusmn 0031 119899 = 21) was significantly
6 Neural Plasticity
DAPI
sosr
sosr
sosr
CTRL
DIA
FFR
500
1000
Nuc
leim
m 2
Control DIA FFR
sp
sp
sp
(a)
GFAP
sosr
sosr
sosr
CTRL
DIA
FFR
200
400
Astr
ocyt
esm
m 2
lowastlowastlowastlowastlowastlowast
FFRControl DIA
sp
sp
sp
(b)
Merge
sosr
sosr
sosr
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
FFRControl DIA
sp
sp
sp
(c)
Figure 4 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA3 Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is shown for control dehydration-inducedanorexia (DIA) and forced-food restricted (FFR) experimental groups (c) Neither the density of nuclei nor the astrocytenuclei ratio differedsignificantly among groups (119901 = 0931 and119901 = 0347 resp) However a significant reduction of GFAP+ cell density was observed (119901 lt 0001)(see Table 1) Nuclear and astrocyte densities were estimated in the stratum oriens (so) and stratum radiatum (sr) stratum pyramidale (sp)is indicated as an anatomical reference Scale bar = 50120583m Data are expressed as mean plusmn SEM
reduced for the DIA (0173 plusmn 0022 119899 = 20) and FFR groups(0139 plusmn 0032 119899 = 19) (119901 lt 0001) (Figure 5 Table 1)
341 Expression of Intermediate Filaments Is Affected byAnorexia The expression of intermediate filaments of astro-cytes was tested byWestern blot in the control DIA and FFRexperimental groups (Figure 6) The expression of GFAP wassignificantly reduced by DIA (048 plusmn 002 119899 = 3 119901 = 0003)and FFR (067plusmn004 119899 = 3119901 = 0006) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0668) (Figures 6(a) and6(b)) The opposite effect was observed for the expressionof vimentin and nestin The expression of vimentin was
significantly increased by DIA (151 plusmn 008 119899 = 3 119901 = 0009)and FFR (146plusmn011 119899 = 3119901 = 0008) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0896) (Figures 6(a)and 6(b)) Finally the expression of nestin was significantlyincreased by DIA (174 plusmn 01 119899 = 3 119901 = 0004) and FFR(161 plusmn 012 119899 = 3 119901 = 0001) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0468) (Figures 6(a) and6(b))
342 Anorexia Decreases GFAP Expression after the ThirdDay Our results showed a reduced GFAP+ cell density in
Neural Plasticity 7
DAPI
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
1000
1500
500
Nuc
leim
m 2
Control DIA FFR(a)
GFAP
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
200
400
600
Astr
ocyt
esm
m 2
lowastlowastlowastlowastlowastlowast
Control DIA FFR(b)
Merge
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
lowastlowastlowast
lowastlowastlowast
Control DIA FFR(c)
Figure 5 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal dentate gyrus (DG) Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is also shown for the control DIA and forcedfood-restricted (FFR) groups (c) The density of nuclei did not differ significantly among experimental groups (119901 = 0398) However asignificant reductionwas observed forGFAP+ cell density and the astrocytenuclei ratio (119901 lt 0001 and119901 lt 0001 resp) (see Table 1) Nuclearand astrocyte densities were estimated in the hilus subgranular zone (SGZ) and granular cell layer (GCL) are indicated as an anatomicalreference Scale bar = 50 120583m Data are expressed as mean plusmn SEM
the hippocampus that correlated with a lower expression ofthe protein Nevertheless GFAP expression may be affectedearlier than five daysThus the expression ofGFAPwas testedby Western blot after one and three days for control DIA orFFR experimental groups (Figures 7(a) and 7(b)) The resultsshowed that the expression of GFAP was unaffected after thefirst day for DIA (092 plusmn 004) and FFR (094 plusmn 003) whencompared to the normalized control (119901 gt 01) HoweverGFAP expression was significantly reduced after the third dayfor DIA (072 plusmn 001) and FFR (075 plusmn 004) when comparedto the normalized control (119901 lt 0001) (Figures 7(a) and7(b))
35 Anorexia Increases Reactive Astrocytes The reactiveastrocytes were estimated in CA2 andDG because astrocyte-nuclei ratio was significantly reduced in these regions Reac-tive astrocytes were defined as GFAP+ cells with processesshorter than the diameter of the somata [16] The numberof reactive astrocytes was estimated for all the experimentalgroups and normalized with their respective controls (seeSection 25) In CA2 reactive astrocytes increased 276 plusmn 07(119899 = 4 119901 = 0047) for DIA and 292 plusmn 075 (119899 = 4 119901 = 0032)for FFR while for dentate gyrus reactive astrocytes increased225plusmn015 (119899 = 4 119901 lt 00001) for DIA and 161plusmn029 (119899 = 4119901 = 0034) for FFR (Figure 8)
8 Neural Plasticity
GFAP
Vimentin
Actin
Nestin
RFFLRTC DIA
50kDa
57kDa
43kDa
200 kDa
(a)
CTRLDIA
FFR
GFAP Vimentin Nestin
10
20
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
lowastlowast
lowastlowast
(b)
Figure 6 Effect of anorexia on astrocyte intermediate filaments expression after the fifth day (a) Representative Western blot showing theexpression of GFAP vimentin and nestin for control DIA and FFR experimental groups actin was used as an internal control (b)The bandintensity of DIA and FFR groups was normalized to their respective controls Data were shown as mean plusmn SEM Significant differences wereconsidered as lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001
GFAP
Actin
After the first day
50kDa
43kDa
CTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
CTRL DIA FFR(a)
GFAP
Actin
50kDa
43kDa
After the third dayCTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
CTRL DIA FFR(b)
Figure 7 GFAP expression is reduced after the third day (a b) Representative Western blot showing the expression of GFAP after the first(a) and third day (b) of anorexia actin was used as an internal control The band intensity of DIA and FFR groups was normalized to theirrespective controls ((a b) under the Western blots) Data were shown as plusmn SEM Significant differences were considered as lowastlowastlowast119901 lt 0001
Neural Plasticity 9
RFFAIDlrtC
1
2
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in D
G
Ctrl DIA FFR
lowastlowastlowast
lowast
1
2
3
Ctrl DIA FFR
lowast
lowast
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in C
A2
Figure 8 Anorexia increases the density of reactive astrocytes Representative images of GFAP+ cells in the dentate gyrus (DG) for control(Ctrl) dehydration-induced anorexia (DIA) and forced food restriction (FFR) groupsThe density of reactive astrocytes (cells with processesshorter than the diameter of the somata) was estimated for all the experimental groups and normalized with their respective controls for DGand CA2 Data were shown as mean plusmn SEM Significant differences were considered as lowast119901 lt 005 lowastlowastlowast119901 lt 0001
4 Discussion
Anorexia nervosa is common among female adolescents andhas one of the highest mortality rates within psychiatric dis-orders (10ndash20) even surpassing depression [19ndash21] Duringpuberty mood swings and anxiety are common due to a sud-den rise of hormones which affects the hippocampus [11 1222] Moreover in the activity-based anorexia model (ABA)structural and functional alterations have been observedin the hippocampus accompanied by anxiety-like behavior[1 11 12 23] However the effect of anorexia on astrocytedensity has been barely explored Indeed previous studies inthe rat corpus callosum reported that cell proliferation wasreduced with the ABA model while astrocyte density wasregionally reduced in the DIA model [1 13] The ABA studyalso reported a reduced hippocampal cell proliferation in thedentate gyrus [1]
Results from the present study show that anorexiareduced astrocyte density in CA2 CA3 and DG whileno significant changes were observed in CA1 Accordinglyanorexia mutant mice (anxanx) showed lower expressionof c-Fos in the dentate gyrus and CA3 region of the hip-pocampus while no significant differences were found inCA1 [24] On the other hand the decrease in GFAP+ celldensity resulted in a reduced expression of GFAP proteinAstrocytes express three types of intermediate filament pro-teins GFAP vimentin and nestin [25] Nestin and vimentinare the main intermediate filaments in immature astrocyteswhereas maturing and adult astrocytes contain vimentin and
GFAP [25] Interestingly anorexia increased vimentin andnestin expression resembling reactive astrocytes observedafter injury [26 27] Accordingly GFAP+ cells deficit may bereplaced by vimentin+nestin+ cells in anorexia In supportof this hypothesis nestin reexpression has been observedfollowing experimental hippocampal lesions [25 28 29] andalso in human pathology such as multiple sclerosis [30]Moreover reactive astrocytes were increased by anorexia andforced food restriction indicating that GFAP+ cells react tosevere caloric restriction [31] Another factor to consider isthat female animals subjected to anorexia or chronic stressstop their ovarian cycles ending their exposure to hormonesthat may affect morphology and astrocyte density [32 33]Moreover hippocampal gliogenesis but not neurogenesisis reduced in the ABA model [1] Thus astrocytes possessdifferent levels of differentiation and maturation and theycan switch between these states even in the adult brain[26] Likewise astrocytes secrete gliotransmitters and growthfactors like BDNF required for synaptic plasticity synapto-genesis and neurogenesis [34 35] Thus a reduced densityof GFAP+ cells may affect hippocampal gliotransmission ofd-serine a gliotransmitter necessary for neuronal NMDA-mediated transmission and long term potentiation [34]Growth factors such as BDNF are also required for long termsynaptic plasticity of dendrites in the adult hippocampus [35]however decreased branching of dendrites was reported inan ABA model particularly in the stratum radiatum [11 12]Accordingly our results showed that GFAP+ cell density andGFAP expression are decreased by anorexia Similar results
10 Neural Plasticity
were reported in a rat model of depression where GFAPexpression was reduced [36] Thus we conclude that GFAP+cells and intermediate filament expression are affected byanorexia Our results add new information about the struc-tural changes observed in the hippocampus ofmurinemodelsof anorexia
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
The authors thank Dr P De Gortari (Insituto Nacionalde Psiquiatrıa Mexico) for introducing them to the DIAmodel The authors are grateful to M Macedo-MendozaM E Ramos-Aguilar E N Hernandez-Rıos A Castilla LCasanova A E Espino and M Garcıa-Servın for technicalsupport The authors thank Dr D Pless for reviewing thepaper The authors thank Dr G Ramırez-Rodrıguez (Insti-tuto Nacional de Psiquiatrıa Mexico) for donating the nestinand vimentin antibodies used in this study Janina Krugerwas an internship student supported by the DAAD-RISEprogram Durairaj Ragu Varman is a postdoctoral fellowsupported by DGAPA-UNAM This work was supported bygrants from Programa de Apoyo a Proyectos de Investigacione Innovacion Tecnologica de la UNAM (PAPIIT-UNAMIN200913 and IN201915) to Ataulfo Martınez-Torres andDaniel Reyes-Haro
References
[1] N C Barbarich-Marsteller C A Fornal L F Takase et alldquoActivity-based anorexia is associated with reduced hippocam-pal cell proliferation in adolescent female ratsrdquo BehaviouralBrain Research vol 236 no 1 pp 251ndash257 2013
[2] C Aoki N Sabaliauskas T Chowdhury et al ldquoAdolescentfemale rats exhibiting activity-based anorexia express elevatedlevels of GABAAreceptor 1205724 and 120575 subunits at the plasmamembrane of hippocampal CA1 spinesrdquo Synapse vol 66 no 5pp 391ndash407 2012
[3] G D Giordano P Renzetti R C Parodi et al ldquoVolume mea-surement with magnetic resonance imaging of hippocampus-amygdala formation in patients with anorexia nervosardquo Journalof Endocrinological Investigation vol 24 no 7 pp 510ndash514 2001
[4] F Connan F Murphy S E J Connor et al ldquoHippocampalvolume and cognitive function in anorexia nervosardquo PsychiatryResearchmdashNeuroimaging vol 146 no 2 pp 117ndash125 2006
[5] J N Beadle S Paradiso M Brumm M Voss K Halmiand L M McCormick ldquoLarger hippocampus size in womenwith anorexia nervosa who exercise excessively than healthywomenrdquo Psychiatry ResearchmdashNeuroimaging vol 232 no 2 pp193ndash199 2015
[6] N T Burkert KKoschutnig F Ebner andW Freidl ldquoStructuralhippocampal alterations perceived stress and coping deficien-cies in patients with anorexia nervosardquo International Journal ofEating Disorders vol 48 no 6 pp 670ndash676 2015
[7] S B McHugh M Fillenz J P Lowry J N P Rawlins and DM Bannerman ldquoBrain tissue oxygen amperometry in behavingrats demonstrates functional dissociation of dorsal and ventral
hippocampus during spatial processing and anxietyrdquo EuropeanJournal of Neuroscience vol 33 no 2 pp 322ndash337 2011
[8] A G Watts ldquoDehydration-associated anorexia developmentand rapid reversalrdquo Physiology and Behavior vol 65 no 4-5 pp871ndash878 1998
[9] L Jaimes-Hoy P Joseph-Bravo and P de Gortari ldquoDiffer-ential response of TRHergic neurons of the hypothalamicparaventricular nucleus (PVN) in female animals submittedto food-restriction or dehydration-induced anorexia and coldexposurerdquo Hormones and Behavior vol 53 no 2 pp 366ndash3772008
[10] A G Watts and C N Boyle ldquoThe functional architecture ofdehydration-anorexiardquo Physiology and Behavior vol 100 no 5pp 472ndash477 2010
[11] T G Chowdhury M B Rıos T E Chan D S Cassataro NC Barbarich-Marsteller and C Aoki ldquoActivity-based anorexiaduring adolescence disrupts normal development of the CA1pyramidal cells in the ventral hippocampus of female ratsrdquoHippocampus vol 24 no 12 pp 1421ndash1429 2014
[12] T G Chowdhury N C Barbarich-Marsteller T E Chan andCAoki ldquoActivity-based anorexia has differential effects on apicaldendritic branching in dorsal and ventral hippocampal CA1rdquoBrain Structure and Function vol 219 no 6 pp 1935ndash1945 2014
[13] D Reyes-Haro F E Labrada-Moncada R Miledi andA Martınez-Torres ldquoDehydration-induced anorexia reducesastrocyte density in the rat corpus callosumrdquo Neural Plasticityvol 2015 Article ID 474917 8 pages 2015
[14] P de Gortari K Mancera A Cote-Velez et al ldquoInvolvement ofCRH-R2 receptor in eating behavior and in the response of theHPT axis in rats subjected to dehydration-induced anorexiardquoPsychoneuroendocrinology vol 34 no 2 pp 259ndash272 2009
[15] M R Lamprecht DM Sabatini and A E Carpenter ldquoCellPro-filer free versatile software for automated biological imageanalysisrdquo BioTechniques vol 42 no 1 pp 71ndash75 2007
[16] B Rinaldi F Guida A Furiano et al ldquoEffect of prolongedmoderate exercise on the changes of nonneuronal cells in earlymyocardial infarctionrdquo Neural Plasticity vol 2015 Article ID265967 8 pages 2015
[17] K Chiu W M Lau H T Lau K-F So and R C-C ChangldquoMicro-dissection of rat brain for RNA or protein extractionfrom specific brain regionrdquo Journal of Visualized Experimentsno 7 article e269 2007
[18] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[19] P F Sullivan ldquoMortality in anorexia nervosardquo The AmericanJournal of Psychiatry vol 152 no 7 pp 1073ndash1074 1995
[20] C L Birmingham J Su J A Hlynsky E M Goldner and MGao ldquoThe mortality rate from anorexia nervosardquo InternationalJournal of Eating Disorders vol 38 no 2 pp 143ndash146 2005
[21] C M Bulik M C T Slof-Oprsquot Landt E F van Furth and P FSullivan ldquoThe genetics of anorexia nervosardquo Annual Review ofNutrition vol 27 pp 263ndash275 2007
[22] K J McLaughlin J O Wilson J Harman et al ldquoChronic 17120573-estradiol or cholesterol prevents stress-induced hippocampalCA3 dendritic retraction in ovariectomized female rats possi-ble correspondence between CA1 spine properties and spatialacquisitionrdquo Hippocampus vol 20 no 6 pp 768ndash786 2010
[23] K P Kinzig and S L Hargrave ldquoAdolescent activity-basedanorexia increases anxiety-like behavior in adulthoodrdquo Physi-ology and Behavior vol 101 no 2 pp 269ndash276 2010
Neural Plasticity 11
[24] S A Kim Y M Choi H J Park et al ldquoDecrease of c-Fosexpression in hippocampus of anorexia (anxanx)micerdquoKoreanJournal of Biological Sciences vol 5 no 2 pp 157ndash161 2001
[25] C Eliasson C Sahlgren C-H Berthold et al ldquoIntermediatefilament protein partnership in astrocytesrdquo The Journal ofBiological Chemistry vol 274 no 34 pp 23996ndash24006 1999
[26] R Schmidt-Kastner andCHumpel ldquoNestin expression persistsin astrocytes of organotypic slice cultures from rat cortexrdquoInternational Journal of Developmental Neuroscience vol 20 no1 pp 29ndash38 2002
[27] Y Takamiya S Kohsaka S Toya M Otani and Y TsukadaldquoImmunohistochemical studies on the proliferation of reactiveastrocytes and the expression of cytoskeletal proteins followingbrain injury in ratsrdquo Brain Research vol 466 no 2 pp 201ndash2101988
[28] S R Clarke A K Shetty J L Bradley and D A TurnerldquoReactive astrocytes express the embryonic intermediate neuro-filament nestinrdquoNeuroReport vol 5 no 15 pp 1885ndash1888 1994
[29] M C Geloso V Corvino V Cavallo et al ldquoExpression ofastrocytic nestin in the rat hippocampus during trimethyltin-induced neurodegenerationrdquo Neuroscience Letters vol 357 no2 pp 103ndash106 2004
[30] J E Holley D Gveric J Newcombe M L Cuzner and N JGutowski ldquoAstrocyte characterization in the multiple sclerosisglial scarrdquoNeuropathology andAppliedNeurobiology vol 29 no5 pp 434ndash444 2003
[31] C N Boyle S M Lorenzen D Compton and A G WattsldquoDehydration-anorexia derives from a reduction in meal sizebut not meal numberrdquo Physiology and Behavior vol 105 no 2pp 305ndash314 2012
[32] I AzcoitiaM Santos-GalindoMAArevalo and LMGarcıa-Segura ldquoRole of astroglia in the neuroplastic and neuroprotec-tive actions of estradiolrdquo European Journal of Neuroscience vol32 no 12 pp 1995ndash2002 2010
[33] L Eiland J Ramroop M N Hill J Manley and B S McEwenldquoChronic juvenile stress produces corticolimbic dendritic archi-tectural remodeling and modulates emotional behavior in maleand female ratsrdquo Psychoneuroendocrinology vol 37 no 1 pp39ndash47 2012
[34] C Henneberger T Papouin S H R Oliet and D A RusakovldquoLong-term potentiation depends on release of d-serine fromastrocytesrdquo Nature vol 463 no 7278 pp 232ndash236 2010
[35] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo The Journal ofNeuroscience vol 22 no 5 pp 1532ndash1540 2002
[36] Y Ye G Wang H Wang and X Wang ldquoBrain-derived neu-rotrophic factor (BDNF) infusion restored astrocytic plasticityin the hippocampus of a rat model of depressionrdquo NeuroscienceLetters vol 503 no 1 pp 15ndash19 2011
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

2 Neural Plasticity
of Laboratory Animals Wistar female rats (160ndash190 g) werehoused individually under 12-h12-h lightdark cycle andcontrolled temperature with food and water ad libitum
22 Dehydration-Induced Anorexia The protocol was per-formed as previously described [8 13 14] Briefly two inde-pendent experimental series of twelve animals were placedin individual cages for each series animals were randomlyselected to form three groups of fourThe first group receivedwater and food ad libitum (control) The DIA group receiveda 25 NaCl solution as their sole drinking liquid and hadunrestricted access to food The forced food-restricted (FFR)group a positive control to distinguish between starvationand dehydration effects received tap water ad libitum andthe same amount of food consumed by DIA animals Theexperimental protocol was conducted for five days and bodyweight and solid food intake were recorded daily at noonfor each experimental group The FFR group received thesame amount of food as that ingested by DIA animals (seeSupplemental Figure 1 in Supplementary Material availableonline at httpdxdoiorg10115520162426413)
23 Histology Rats were deeply anaesthetized with an over-dose of sodium pentobarbital (100mgKg) and transcardiallyperfused with 100mL of saline followed by 250mL of chilled4 paraformaldehyde in phosphate-buffered saline (PBS)(pH 74) Brains were removed postfixed overnight and thentransferred to a series of sucrose solutions (from 10 to 30)Coronal sections (30 120583m) including the dorsal hippocampuswere obtained on a freezing microtome collected and storedin cryoprotectant solution (30 ethylene glycol20 glycerolin PBS) at minus20∘C [13]
24 Immunohistofluorescence Glial fibrillary acidic protein(GFAP) immunoreactivity was performed on floating sec-tions [13] Briefly coronal sections were rinsed three timesin PBS buffer and treated with 3 hydrogen peroxide for10min followed by three rinses in PBS and incubation in 10sodium borohydride for 6ndash8min to reduce free aldehydesThe sections were incubated for 1 h in blocking solution (5horse serum albumin1 Triton X-100 in PBS) Sections wereincubated for 48 h with polyclonal rabbit anti-GFAP anti-body (dilution 1 1000 DakoCytomation Fort Collins COUSA 4∘C) After washing primary antibody was detectedwith Alexa 594 (1 500 Invitrogen) coupled goat anti-rabbitsecondary antibody The sections were counterstained with410158406-diamidino-2-phenylindole (DAPI) and mounted withVectashield H-1000 (Vector Laboratories Burlingame CAUSA)
Mounted coronal sections containing the hippocampuswere photographed with a digital camera (Photometrics CoolSnap FX USA) attached to a Nikon microscope (NikonEclipse E600 Tokyo Japan) and analyzed using IMAGEJ version 141 (NIH Bethesda MD USA) A Zeiss LSM780 Meta Confocal Microscope (Zeiss Gottingen Germany)was used for confocal images with Alexa 594 (excita-tionemission wavelength 590617 nm) and DAPI (excita-tionemission wavelength 350460 nm) (Figure 1)
CA1CA2
CA3
DG
spsr
so
SGZ
GCL
CA1CA2
CA3
SGZ
FFR
Control
CA1
CA2
CA3
SGZ
D
VL L
DIA
spsr
so
spsr
so
D
VL L
D
VL L
DG
DG
GCL
GCL
lowast
lowast
lowast
Figure 1 The rat hippocampus Coronal brain sections from youngfemale rats showing the hippocampus of control dehydration-induced anorexia (DIA) and forced food-restricted (FFR) ratsThe histological sections from the three experimental groups wereimmunostained for GFAP (green) and nuclei were labeled withDAPI (blue) The hippocampal regions for this study included thecornus ammonis 1ndash3 (CA1 CA2 and CA3) and the dentate gyrus(DG) Nuclear and astrocyte densities were estimated in the stratumoriens (so) and stratum radiatum (sr) of CA1 CA2 andCA3whilethe hilus region (lowast) was studied for the DG (squares) Other regionsare indicated for anatomical reference stratum pyramidale (sp)granular cell layer (GCL) and subgranular zone (SGZ) Arrowsshow anatomical orientation dorsal (D) ventral (V) and lateral (L)Scale bars correspond to 200 120583m
25 Cell Counting GFAP-immunolabeled cells were countedand contrasted with the number of DAPI-labeled nuclei foreach subfield of the hippocampus CA1 CA2 CA3 anddentate gyrus (DG) A total of 2-3 aleatory fields in each ofthe three tissue sections selected from each of 6ndash8 animalsper group were used for cell counting and ldquonrdquo refers to thetotal aleatory fields used from all the tissue sections from allthe animals of each experimental group A test square grid of100 times 100 120583m (001mm2) was used to estimate the number of
Neural Plasticity 3
Table 1 Regional changes in astrocyte density in DIA and FFR
Hippocampal region Control DIA FFR 119901
NucleiCA1 1099 plusmn 52 (119899 = 62) 924 plusmn 67 (119899 = 56) 932 plusmn 68 (119899 = 52) 0075CA2 1125 plusmn 101 (119899 = 40) 1007 plusmn 51 (119899 = 40) 1022 plusmn 56 (119899 = 36) 0467CA3 1288 plusmn 43 (119899 = 62) 1084 plusmn 41lowastlowastlowast (119899 = 56) 1117 plusmn 46lowastlowastlowast (119899 = 52) 0001DGSGZ 1433 plusmn 110 (119899 = 21) 1535 plusmn 142 (119899 = 20) 1268 plusmn 161 (119899 = 19) 0398
AstrocytesCA1 257 plusmn 14 (119899 = 82) 253 plusmn 14 (119899 = 72) 221 plusmn 15 (119899 = 66) 0172CA2 405 plusmn 19 (119899 = 40) 323 plusmn 18lowastlowastlowast (119899 = 40) 267 plusmn 19lowastlowastlowast (119899 = 36) lt0001CA3 332 plusmn 14 (119899 = 82) 269 plusmn 15lowastlowastlowast (119899 = 72) 258 plusmn 15lowastlowastlowast (119899 = 66) lt0001DGSGZ 476 plusmn 28 (119899 = 41) 294 plusmn 24lowastlowastlowast (119899 = 36) 321 plusmn 26lowastlowastlowast (119899 = 32) lt0001
Astrocytesnuclei ratioCA1 0277 plusmn 0025 (119899 = 62) 0324 plusmn 0025 (119899 = 56) 0286 plusmn 0029 (119899 = 52) 0417CA2 0447 plusmn 0053 (119899 = 40) 0345 plusmn 0021lowastlowastlowast (119899 = 40) 0282 plusmn 0024lowastlowastlowast (119899 = 36) 0008CA3 0292 plusmn 0016 (119899 = 62) 0262 plusmn 0016 (119899 = 56) 0264 plusmn 0016 (119899 = 52) 0347DGSGZ 0334 plusmn 0031 (119899 = 21) 0173 plusmn 0022lowastlowastlowast (119899 = 20) 0139 plusmn 0032lowastlowastlowast (119899 = 19) lt0001
DAPI-stained nuclei (equivalent to the total number of cells)and GFAP-labeled cells corresponding to astrocytes manu-ally and with Cell Profiler [13 15] Only process-bearing cellsshowing their soma in the plane of the analyzed area werecounted and their density was estimated (number per mm2)Six to eight randomly chosen fields located within each of theanalyzed hippocampal subfields were counted by progressivedisplacement of a test square gridThe astrocytesnuclei ratiowas calculated by dividing the number of GFAP+ cells bythe total number of nuclei labeled with DAPI GFAP+ cellswere identified as resting (with small somata bearing longthin and ramified processes) or reactive (with retraction ofprocesses to a length shorter than the diameter of the somata)[16] The number of reactive astrocytes was estimated for allthe experimental groups and normalized with the control
All the data corresponding to each treatment and regionwere pooled and are presented as the mean plusmn standard errorof the mean (SEM) Statistical analysis of data was performedusing a one-way ANOVA followed by a Bonferroni posttestwith Origin 70 software 119901 lt 005was considered statisticallysignificant
26Western Blot Thehippocampuswas dissected out as pre-viously described [17] The hippocampi from three differentrats for each experimental group (control DIA and FFR)were homogenized in ice cold lysis buffer (200mM glycine150mM NaCl 50mM EGTA 50mM EDTA and 300mMsucrose) with a protease inhibitor (Sigma-Aldrich USA)The homogenate was centrifuged at 10000timesg for 15 minutesat 4∘C and the supernatants were aliquoted and stored atminus80∘C Concentration of each protein sample was estimatedby Bradfordrsquos method [18] An equal concentration of protein(30 120583g) was resolved by 10 polyacrylamide gel electrophore-sis (PAGE)The proteins were transferred electrophoreticallyto PVDFmembrane then the membranes were blocked with5 nonfat dry milk in TBS-T for 3 h at room temperature
(RT) Membranes were incubated at 4∘C overnight withone of the following primary antibodies (a) polyclonalrabbit anti-GFAP antibody (dilution 1 2000 DakoCytoma-tion Fort Collins CO USA) (b) polyclonal rabbit anti-vimentin (dilution 1 2500 Cell Signaling Danvers MAUSA) (c) monoclonal mouse anti-nestin (dilution 1 5000BD Biosciences San Jose CA USA) and (d) polyclonal goatanti-actin (dilution 1 2500 Santacruz Dallas TX USA)The membranes were washed and bound antibodies weredetected by incubating for 3 h with either the goat anti-rabbitalkaline phosphatase conjugated antibody (dilution 1 2500Santacruz Dallas TX) goat anti-mouse alkaline phosphataseconjugated antibody (dilution 1 2500 Santacruz DallasTX) or rabbit anti-goat alkaline phosphatase conjugatedantibody (dilution 1 2500 Santacruz Dallas TX) Then themembrane was washed with TBS-T and the alkaline phos-phatase activity was detected with BCIPNBT AP-conjugatesubstrate reaction Kit (Bio-Rad USA) The intensity of eachband was measured using Image Lab 30 software
3 Results
The effects of DIA and FFR on the density of nuclei andastrocytes were estimated for the following hippocampalregions CA1 CA2 CA3 and dentate gyrus (DG) (Figure 1)All observations included the stratum radiatum and stratumoriens where astrocytes are preferentially located whilefor the DG the observations were performed in the hilus(Figure 1)
31 Anorexia Had No Effect on GFAP+ Cell Density inCA1 The hippocampal region CA1 showed no significantchange in nuclear density between the control (1099 plusmn 52nucleimm2 119899 = 62) DIA (924 plusmn 67 nucleimm2 119899 = 56)and FFR groups (932 plusmn 68 nucleimm2 119899 = 52) (119901 = 0075)(Figure 1 and Table 1) Additionally no significant changes
4 Neural Plasticity
500
1000
Nuc
leim
m 2
CTRL
DIA
FFR
DAPIso
sp
sp
sp
sr
so
sr
so
sr
Control DIA FFR(a)
GFAPso
sr
so
sr
so
sr
CTRL
DIA
FFR
100
200
300
Astr
ocyt
esm
m 2
Control DIA FFR
sp
sp
sp
(b)
Ratio
astro
cyte
snu
clei
02
04
Mergeso
sr
so
sr
so
sr
CTRL
DIA
FFR
Control DIA FFR
sp
sp
sp
(c)
Figure 2 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA1 Coronal sections of thehippocampus show nuclei labeled with DAPI (a) or immunostained with GFAP (b) The overlay of DAPI and GFAP is shown for controldehydration-induced anorexia (DIA) and forced food restricted (FFR) experimental groups (c)The densities of nuclei or GFAP+ cells as wellas the ratio of astrocytenuclei did not differ significantly among experimental groups (119901 = 0075 119901 = 0172 119901 = 0417 for the control DIAand FFR groups respectively see Table 1) Nuclear and astrocyte densities were estimated in the stratum oriens (so) and stratum radiatum(sr) stratum pyramidale (sp) is indicated as a reference Scale bar = 50 120583m Data are mean plusmn SEM
were observed for astrocyte density between the control(257 plusmn 14 nucleimm2 119899 = 82) DIA (253 plusmn 14 nucleimm2119899 = 72) and FFR groups (221 plusmn 15 nucleimm2 119899 = 66)(119901 = 0172) (Figure 2 and Table 1) We also determined theastrocytenuclei ratio by estimating the GFAP+ cells from thetotal nuclei The astrocytenuclei ratio for the control groupwas 0277 plusmn 0025 (119899 = 62) and it was not significantlydifferent for the DIA and FFR groups (0324 plusmn 0025 119899 = 56and 0286 plusmn 0029 119899 = 52 119901 = 0417) (Figure 2 and Table 1)
32 Anorexia Reduced GFAP+ Cell Density in CA2 Nucleardensity in CA2 was similar for the control (1125 plusmn 101nucleimm2 119899 = 40) DIA (1007 plusmn 51 nucleimm2 119899 = 40)
and FFR groups (1022 plusmn 56 nucleimm2 119899 = 36) (119901 = 0467)(Figure 3 Table 1) In contrast the astrocyte density estimatedfor the control group (405 plusmn 19 astrocytesmm2 119899 = 40) wassignificantly reduced for the DIA (323 plusmn 18 astrocytesmm2119899 = 40) and FFR groups (267 plusmn 19 astrocytemm2 119899 =36) (119901 lt 0001) (Figure 3 Table 1) Accordingly theastrocytenuclei ratio estimated for the control group (0447plusmn0053 119899 = 40) was also significantly reduced for the DIA(0345plusmn0021 119899 = 40) and FFR groups (0282plusmn0024 119899 = 36)(119901 = 0008) (Figure 3 Table 1)
33 Anorexia Reduced GFAP+ Cell Density in CA3 Thenuclear density of CA3 did not show significant changes
Neural Plasticity 5
DAPI
so
sp
sp
sp
sr
so
sr
so
sr
CTRL
DIA
FFR
500
1000
Nuc
leim
m 2
Control DIA FFR(a)
GFAP
so
sr
so
sr
so
sr
CTRL
DIA
FFR
200
400
Astr
ocyt
esm
m 2
lowastlowastlowast
lowastlowastlowast
Control DIA FFR
sp
sp
sp
(b)
Merge
so
sr
so
sr
so
sr
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
06
lowastlowastlowast
lowastlowastlowast
Control DIA FFR
sp
sp
sp
(c)
Figure 3 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA2 Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is shown for control dehydration-inducedanorexia (DIA) and forced-food restricted (FFR) experimental groups (c) The density of the nuclei was similar in all experimental groups(119901 = 0467) However a significant reduction of GFAP+ cells was observed for the DIA and FFR groups (119901 lt 0001) This reduction wasconsistently observed in the astrocytenuclei ratio (119901 = 0008) (see Table 1) Nuclear and astrocyte densities were estimated in the stratumoriens (so) and stratum radiatum (sr) stratum pyramidale (sp) is indicated as an anatomical reference Scale bar = 50120583m Data areexpressed as mean plusmn SEM
between the control (1288 plusmn 43 nucleimm2 119899 = 62) DIA(1084 plusmn 41 nucleimm2 119899 = 56) and FFR groups (1117 plusmn46 nucleimm2 119899 = 52) (119901 = 0001) (Figure 4 Table 1)However the astrocyte density of the control group (332plusmn 19astrocytesmm2 119899 = 82) was significantly reduced for theDIA (269plusmn15 astrocytesmm2 119899 = 72) and FFR groups (258plusmn15 astrocytemm2 119899 = 66) (119901 lt 0001) (Figure 4 Table 1)However the astrocytenuclei ratio estimated for the controlgroup (0292plusmn0016 119899 = 62) was not significantly reduced intheDIA (0262plusmn0016 119899 = 56) and FFR groups (0264plusmn0016119899 = 52) (119901 = 0347) (Figure 4 Table 1)
34 Anorexia Reduced GFAP+ Cell Density in DG Thenuclear density of the dentate gyrus did not differ signifi-cantly for the control (1433 plusmn 110 nucleimm2 119899 = 21)DIA (1535 plusmn 142 nucleimm2 119899 = 20) and FFR groups(1268 plusmn 161 nucleimm2 119899 = 19) (119901 = 0398) (Figure 5Table 1) The astrocyte density of the control group (476 plusmn 28astrocytesmm2 119899 = 41) was significantly reduced for theDIA (294 plusmn 124 astrocytesmm2 119899 = 36) and FFR groups(321 plusmn 26 astrocytemm2 119899 = 32) (119901 lt 0001) (Figure 5Table 1) Accordingly the astrocytenuclei ratio estimated forthe control group (0334 plusmn 0031 119899 = 21) was significantly
6 Neural Plasticity
DAPI
sosr
sosr
sosr
CTRL
DIA
FFR
500
1000
Nuc
leim
m 2
Control DIA FFR
sp
sp
sp
(a)
GFAP
sosr
sosr
sosr
CTRL
DIA
FFR
200
400
Astr
ocyt
esm
m 2
lowastlowastlowastlowastlowastlowast
FFRControl DIA
sp
sp
sp
(b)
Merge
sosr
sosr
sosr
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
FFRControl DIA
sp
sp
sp
(c)
Figure 4 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA3 Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is shown for control dehydration-inducedanorexia (DIA) and forced-food restricted (FFR) experimental groups (c) Neither the density of nuclei nor the astrocytenuclei ratio differedsignificantly among groups (119901 = 0931 and119901 = 0347 resp) However a significant reduction of GFAP+ cell density was observed (119901 lt 0001)(see Table 1) Nuclear and astrocyte densities were estimated in the stratum oriens (so) and stratum radiatum (sr) stratum pyramidale (sp)is indicated as an anatomical reference Scale bar = 50120583m Data are expressed as mean plusmn SEM
reduced for the DIA (0173 plusmn 0022 119899 = 20) and FFR groups(0139 plusmn 0032 119899 = 19) (119901 lt 0001) (Figure 5 Table 1)
341 Expression of Intermediate Filaments Is Affected byAnorexia The expression of intermediate filaments of astro-cytes was tested byWestern blot in the control DIA and FFRexperimental groups (Figure 6) The expression of GFAP wassignificantly reduced by DIA (048 plusmn 002 119899 = 3 119901 = 0003)and FFR (067plusmn004 119899 = 3119901 = 0006) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0668) (Figures 6(a) and6(b)) The opposite effect was observed for the expressionof vimentin and nestin The expression of vimentin was
significantly increased by DIA (151 plusmn 008 119899 = 3 119901 = 0009)and FFR (146plusmn011 119899 = 3119901 = 0008) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0896) (Figures 6(a)and 6(b)) Finally the expression of nestin was significantlyincreased by DIA (174 plusmn 01 119899 = 3 119901 = 0004) and FFR(161 plusmn 012 119899 = 3 119901 = 0001) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0468) (Figures 6(a) and6(b))
342 Anorexia Decreases GFAP Expression after the ThirdDay Our results showed a reduced GFAP+ cell density in
Neural Plasticity 7
DAPI
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
1000
1500
500
Nuc
leim
m 2
Control DIA FFR(a)
GFAP
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
200
400
600
Astr
ocyt
esm
m 2
lowastlowastlowastlowastlowastlowast
Control DIA FFR(b)
Merge
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
lowastlowastlowast
lowastlowastlowast
Control DIA FFR(c)
Figure 5 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal dentate gyrus (DG) Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is also shown for the control DIA and forcedfood-restricted (FFR) groups (c) The density of nuclei did not differ significantly among experimental groups (119901 = 0398) However asignificant reductionwas observed forGFAP+ cell density and the astrocytenuclei ratio (119901 lt 0001 and119901 lt 0001 resp) (see Table 1) Nuclearand astrocyte densities were estimated in the hilus subgranular zone (SGZ) and granular cell layer (GCL) are indicated as an anatomicalreference Scale bar = 50 120583m Data are expressed as mean plusmn SEM
the hippocampus that correlated with a lower expression ofthe protein Nevertheless GFAP expression may be affectedearlier than five daysThus the expression ofGFAPwas testedby Western blot after one and three days for control DIA orFFR experimental groups (Figures 7(a) and 7(b)) The resultsshowed that the expression of GFAP was unaffected after thefirst day for DIA (092 plusmn 004) and FFR (094 plusmn 003) whencompared to the normalized control (119901 gt 01) HoweverGFAP expression was significantly reduced after the third dayfor DIA (072 plusmn 001) and FFR (075 plusmn 004) when comparedto the normalized control (119901 lt 0001) (Figures 7(a) and7(b))
35 Anorexia Increases Reactive Astrocytes The reactiveastrocytes were estimated in CA2 andDG because astrocyte-nuclei ratio was significantly reduced in these regions Reac-tive astrocytes were defined as GFAP+ cells with processesshorter than the diameter of the somata [16] The numberof reactive astrocytes was estimated for all the experimentalgroups and normalized with their respective controls (seeSection 25) In CA2 reactive astrocytes increased 276 plusmn 07(119899 = 4 119901 = 0047) for DIA and 292 plusmn 075 (119899 = 4 119901 = 0032)for FFR while for dentate gyrus reactive astrocytes increased225plusmn015 (119899 = 4 119901 lt 00001) for DIA and 161plusmn029 (119899 = 4119901 = 0034) for FFR (Figure 8)
8 Neural Plasticity
GFAP
Vimentin
Actin
Nestin
RFFLRTC DIA
50kDa
57kDa
43kDa
200 kDa
(a)
CTRLDIA
FFR
GFAP Vimentin Nestin
10
20
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
lowastlowast
lowastlowast
(b)
Figure 6 Effect of anorexia on astrocyte intermediate filaments expression after the fifth day (a) Representative Western blot showing theexpression of GFAP vimentin and nestin for control DIA and FFR experimental groups actin was used as an internal control (b)The bandintensity of DIA and FFR groups was normalized to their respective controls Data were shown as mean plusmn SEM Significant differences wereconsidered as lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001
GFAP
Actin
After the first day
50kDa
43kDa
CTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
CTRL DIA FFR(a)
GFAP
Actin
50kDa
43kDa
After the third dayCTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
CTRL DIA FFR(b)
Figure 7 GFAP expression is reduced after the third day (a b) Representative Western blot showing the expression of GFAP after the first(a) and third day (b) of anorexia actin was used as an internal control The band intensity of DIA and FFR groups was normalized to theirrespective controls ((a b) under the Western blots) Data were shown as plusmn SEM Significant differences were considered as lowastlowastlowast119901 lt 0001
Neural Plasticity 9
RFFAIDlrtC
1
2
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in D
G
Ctrl DIA FFR
lowastlowastlowast
lowast
1
2
3
Ctrl DIA FFR
lowast
lowast
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in C
A2
Figure 8 Anorexia increases the density of reactive astrocytes Representative images of GFAP+ cells in the dentate gyrus (DG) for control(Ctrl) dehydration-induced anorexia (DIA) and forced food restriction (FFR) groupsThe density of reactive astrocytes (cells with processesshorter than the diameter of the somata) was estimated for all the experimental groups and normalized with their respective controls for DGand CA2 Data were shown as mean plusmn SEM Significant differences were considered as lowast119901 lt 005 lowastlowastlowast119901 lt 0001
4 Discussion
Anorexia nervosa is common among female adolescents andhas one of the highest mortality rates within psychiatric dis-orders (10ndash20) even surpassing depression [19ndash21] Duringpuberty mood swings and anxiety are common due to a sud-den rise of hormones which affects the hippocampus [11 1222] Moreover in the activity-based anorexia model (ABA)structural and functional alterations have been observedin the hippocampus accompanied by anxiety-like behavior[1 11 12 23] However the effect of anorexia on astrocytedensity has been barely explored Indeed previous studies inthe rat corpus callosum reported that cell proliferation wasreduced with the ABA model while astrocyte density wasregionally reduced in the DIA model [1 13] The ABA studyalso reported a reduced hippocampal cell proliferation in thedentate gyrus [1]
Results from the present study show that anorexiareduced astrocyte density in CA2 CA3 and DG whileno significant changes were observed in CA1 Accordinglyanorexia mutant mice (anxanx) showed lower expressionof c-Fos in the dentate gyrus and CA3 region of the hip-pocampus while no significant differences were found inCA1 [24] On the other hand the decrease in GFAP+ celldensity resulted in a reduced expression of GFAP proteinAstrocytes express three types of intermediate filament pro-teins GFAP vimentin and nestin [25] Nestin and vimentinare the main intermediate filaments in immature astrocyteswhereas maturing and adult astrocytes contain vimentin and
GFAP [25] Interestingly anorexia increased vimentin andnestin expression resembling reactive astrocytes observedafter injury [26 27] Accordingly GFAP+ cells deficit may bereplaced by vimentin+nestin+ cells in anorexia In supportof this hypothesis nestin reexpression has been observedfollowing experimental hippocampal lesions [25 28 29] andalso in human pathology such as multiple sclerosis [30]Moreover reactive astrocytes were increased by anorexia andforced food restriction indicating that GFAP+ cells react tosevere caloric restriction [31] Another factor to consider isthat female animals subjected to anorexia or chronic stressstop their ovarian cycles ending their exposure to hormonesthat may affect morphology and astrocyte density [32 33]Moreover hippocampal gliogenesis but not neurogenesisis reduced in the ABA model [1] Thus astrocytes possessdifferent levels of differentiation and maturation and theycan switch between these states even in the adult brain[26] Likewise astrocytes secrete gliotransmitters and growthfactors like BDNF required for synaptic plasticity synapto-genesis and neurogenesis [34 35] Thus a reduced densityof GFAP+ cells may affect hippocampal gliotransmission ofd-serine a gliotransmitter necessary for neuronal NMDA-mediated transmission and long term potentiation [34]Growth factors such as BDNF are also required for long termsynaptic plasticity of dendrites in the adult hippocampus [35]however decreased branching of dendrites was reported inan ABA model particularly in the stratum radiatum [11 12]Accordingly our results showed that GFAP+ cell density andGFAP expression are decreased by anorexia Similar results
10 Neural Plasticity
were reported in a rat model of depression where GFAPexpression was reduced [36] Thus we conclude that GFAP+cells and intermediate filament expression are affected byanorexia Our results add new information about the struc-tural changes observed in the hippocampus ofmurinemodelsof anorexia
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
The authors thank Dr P De Gortari (Insituto Nacionalde Psiquiatrıa Mexico) for introducing them to the DIAmodel The authors are grateful to M Macedo-MendozaM E Ramos-Aguilar E N Hernandez-Rıos A Castilla LCasanova A E Espino and M Garcıa-Servın for technicalsupport The authors thank Dr D Pless for reviewing thepaper The authors thank Dr G Ramırez-Rodrıguez (Insti-tuto Nacional de Psiquiatrıa Mexico) for donating the nestinand vimentin antibodies used in this study Janina Krugerwas an internship student supported by the DAAD-RISEprogram Durairaj Ragu Varman is a postdoctoral fellowsupported by DGAPA-UNAM This work was supported bygrants from Programa de Apoyo a Proyectos de Investigacione Innovacion Tecnologica de la UNAM (PAPIIT-UNAMIN200913 and IN201915) to Ataulfo Martınez-Torres andDaniel Reyes-Haro
References
[1] N C Barbarich-Marsteller C A Fornal L F Takase et alldquoActivity-based anorexia is associated with reduced hippocam-pal cell proliferation in adolescent female ratsrdquo BehaviouralBrain Research vol 236 no 1 pp 251ndash257 2013
[2] C Aoki N Sabaliauskas T Chowdhury et al ldquoAdolescentfemale rats exhibiting activity-based anorexia express elevatedlevels of GABAAreceptor 1205724 and 120575 subunits at the plasmamembrane of hippocampal CA1 spinesrdquo Synapse vol 66 no 5pp 391ndash407 2012
[3] G D Giordano P Renzetti R C Parodi et al ldquoVolume mea-surement with magnetic resonance imaging of hippocampus-amygdala formation in patients with anorexia nervosardquo Journalof Endocrinological Investigation vol 24 no 7 pp 510ndash514 2001
[4] F Connan F Murphy S E J Connor et al ldquoHippocampalvolume and cognitive function in anorexia nervosardquo PsychiatryResearchmdashNeuroimaging vol 146 no 2 pp 117ndash125 2006
[5] J N Beadle S Paradiso M Brumm M Voss K Halmiand L M McCormick ldquoLarger hippocampus size in womenwith anorexia nervosa who exercise excessively than healthywomenrdquo Psychiatry ResearchmdashNeuroimaging vol 232 no 2 pp193ndash199 2015
[6] N T Burkert KKoschutnig F Ebner andW Freidl ldquoStructuralhippocampal alterations perceived stress and coping deficien-cies in patients with anorexia nervosardquo International Journal ofEating Disorders vol 48 no 6 pp 670ndash676 2015
[7] S B McHugh M Fillenz J P Lowry J N P Rawlins and DM Bannerman ldquoBrain tissue oxygen amperometry in behavingrats demonstrates functional dissociation of dorsal and ventral
hippocampus during spatial processing and anxietyrdquo EuropeanJournal of Neuroscience vol 33 no 2 pp 322ndash337 2011
[8] A G Watts ldquoDehydration-associated anorexia developmentand rapid reversalrdquo Physiology and Behavior vol 65 no 4-5 pp871ndash878 1998
[9] L Jaimes-Hoy P Joseph-Bravo and P de Gortari ldquoDiffer-ential response of TRHergic neurons of the hypothalamicparaventricular nucleus (PVN) in female animals submittedto food-restriction or dehydration-induced anorexia and coldexposurerdquo Hormones and Behavior vol 53 no 2 pp 366ndash3772008
[10] A G Watts and C N Boyle ldquoThe functional architecture ofdehydration-anorexiardquo Physiology and Behavior vol 100 no 5pp 472ndash477 2010
[11] T G Chowdhury M B Rıos T E Chan D S Cassataro NC Barbarich-Marsteller and C Aoki ldquoActivity-based anorexiaduring adolescence disrupts normal development of the CA1pyramidal cells in the ventral hippocampus of female ratsrdquoHippocampus vol 24 no 12 pp 1421ndash1429 2014
[12] T G Chowdhury N C Barbarich-Marsteller T E Chan andCAoki ldquoActivity-based anorexia has differential effects on apicaldendritic branching in dorsal and ventral hippocampal CA1rdquoBrain Structure and Function vol 219 no 6 pp 1935ndash1945 2014
[13] D Reyes-Haro F E Labrada-Moncada R Miledi andA Martınez-Torres ldquoDehydration-induced anorexia reducesastrocyte density in the rat corpus callosumrdquo Neural Plasticityvol 2015 Article ID 474917 8 pages 2015
[14] P de Gortari K Mancera A Cote-Velez et al ldquoInvolvement ofCRH-R2 receptor in eating behavior and in the response of theHPT axis in rats subjected to dehydration-induced anorexiardquoPsychoneuroendocrinology vol 34 no 2 pp 259ndash272 2009
[15] M R Lamprecht DM Sabatini and A E Carpenter ldquoCellPro-filer free versatile software for automated biological imageanalysisrdquo BioTechniques vol 42 no 1 pp 71ndash75 2007
[16] B Rinaldi F Guida A Furiano et al ldquoEffect of prolongedmoderate exercise on the changes of nonneuronal cells in earlymyocardial infarctionrdquo Neural Plasticity vol 2015 Article ID265967 8 pages 2015
[17] K Chiu W M Lau H T Lau K-F So and R C-C ChangldquoMicro-dissection of rat brain for RNA or protein extractionfrom specific brain regionrdquo Journal of Visualized Experimentsno 7 article e269 2007
[18] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[19] P F Sullivan ldquoMortality in anorexia nervosardquo The AmericanJournal of Psychiatry vol 152 no 7 pp 1073ndash1074 1995
[20] C L Birmingham J Su J A Hlynsky E M Goldner and MGao ldquoThe mortality rate from anorexia nervosardquo InternationalJournal of Eating Disorders vol 38 no 2 pp 143ndash146 2005
[21] C M Bulik M C T Slof-Oprsquot Landt E F van Furth and P FSullivan ldquoThe genetics of anorexia nervosardquo Annual Review ofNutrition vol 27 pp 263ndash275 2007
[22] K J McLaughlin J O Wilson J Harman et al ldquoChronic 17120573-estradiol or cholesterol prevents stress-induced hippocampalCA3 dendritic retraction in ovariectomized female rats possi-ble correspondence between CA1 spine properties and spatialacquisitionrdquo Hippocampus vol 20 no 6 pp 768ndash786 2010
[23] K P Kinzig and S L Hargrave ldquoAdolescent activity-basedanorexia increases anxiety-like behavior in adulthoodrdquo Physi-ology and Behavior vol 101 no 2 pp 269ndash276 2010
Neural Plasticity 11
[24] S A Kim Y M Choi H J Park et al ldquoDecrease of c-Fosexpression in hippocampus of anorexia (anxanx)micerdquoKoreanJournal of Biological Sciences vol 5 no 2 pp 157ndash161 2001
[25] C Eliasson C Sahlgren C-H Berthold et al ldquoIntermediatefilament protein partnership in astrocytesrdquo The Journal ofBiological Chemistry vol 274 no 34 pp 23996ndash24006 1999
[26] R Schmidt-Kastner andCHumpel ldquoNestin expression persistsin astrocytes of organotypic slice cultures from rat cortexrdquoInternational Journal of Developmental Neuroscience vol 20 no1 pp 29ndash38 2002
[27] Y Takamiya S Kohsaka S Toya M Otani and Y TsukadaldquoImmunohistochemical studies on the proliferation of reactiveastrocytes and the expression of cytoskeletal proteins followingbrain injury in ratsrdquo Brain Research vol 466 no 2 pp 201ndash2101988
[28] S R Clarke A K Shetty J L Bradley and D A TurnerldquoReactive astrocytes express the embryonic intermediate neuro-filament nestinrdquoNeuroReport vol 5 no 15 pp 1885ndash1888 1994
[29] M C Geloso V Corvino V Cavallo et al ldquoExpression ofastrocytic nestin in the rat hippocampus during trimethyltin-induced neurodegenerationrdquo Neuroscience Letters vol 357 no2 pp 103ndash106 2004
[30] J E Holley D Gveric J Newcombe M L Cuzner and N JGutowski ldquoAstrocyte characterization in the multiple sclerosisglial scarrdquoNeuropathology andAppliedNeurobiology vol 29 no5 pp 434ndash444 2003
[31] C N Boyle S M Lorenzen D Compton and A G WattsldquoDehydration-anorexia derives from a reduction in meal sizebut not meal numberrdquo Physiology and Behavior vol 105 no 2pp 305ndash314 2012
[32] I AzcoitiaM Santos-GalindoMAArevalo and LMGarcıa-Segura ldquoRole of astroglia in the neuroplastic and neuroprotec-tive actions of estradiolrdquo European Journal of Neuroscience vol32 no 12 pp 1995ndash2002 2010
[33] L Eiland J Ramroop M N Hill J Manley and B S McEwenldquoChronic juvenile stress produces corticolimbic dendritic archi-tectural remodeling and modulates emotional behavior in maleand female ratsrdquo Psychoneuroendocrinology vol 37 no 1 pp39ndash47 2012
[34] C Henneberger T Papouin S H R Oliet and D A RusakovldquoLong-term potentiation depends on release of d-serine fromastrocytesrdquo Nature vol 463 no 7278 pp 232ndash236 2010
[35] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo The Journal ofNeuroscience vol 22 no 5 pp 1532ndash1540 2002
[36] Y Ye G Wang H Wang and X Wang ldquoBrain-derived neu-rotrophic factor (BDNF) infusion restored astrocytic plasticityin the hippocampus of a rat model of depressionrdquo NeuroscienceLetters vol 503 no 1 pp 15ndash19 2011
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
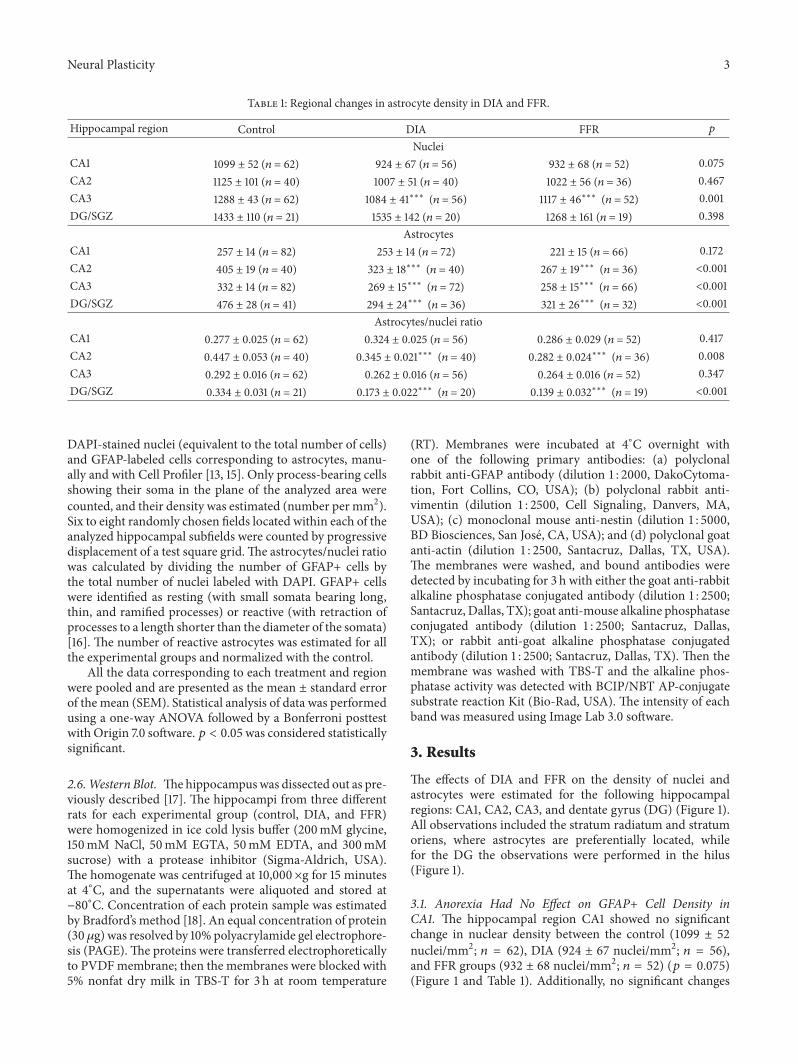
Neural Plasticity 3
Table 1 Regional changes in astrocyte density in DIA and FFR
Hippocampal region Control DIA FFR 119901
NucleiCA1 1099 plusmn 52 (119899 = 62) 924 plusmn 67 (119899 = 56) 932 plusmn 68 (119899 = 52) 0075CA2 1125 plusmn 101 (119899 = 40) 1007 plusmn 51 (119899 = 40) 1022 plusmn 56 (119899 = 36) 0467CA3 1288 plusmn 43 (119899 = 62) 1084 plusmn 41lowastlowastlowast (119899 = 56) 1117 plusmn 46lowastlowastlowast (119899 = 52) 0001DGSGZ 1433 plusmn 110 (119899 = 21) 1535 plusmn 142 (119899 = 20) 1268 plusmn 161 (119899 = 19) 0398
AstrocytesCA1 257 plusmn 14 (119899 = 82) 253 plusmn 14 (119899 = 72) 221 plusmn 15 (119899 = 66) 0172CA2 405 plusmn 19 (119899 = 40) 323 plusmn 18lowastlowastlowast (119899 = 40) 267 plusmn 19lowastlowastlowast (119899 = 36) lt0001CA3 332 plusmn 14 (119899 = 82) 269 plusmn 15lowastlowastlowast (119899 = 72) 258 plusmn 15lowastlowastlowast (119899 = 66) lt0001DGSGZ 476 plusmn 28 (119899 = 41) 294 plusmn 24lowastlowastlowast (119899 = 36) 321 plusmn 26lowastlowastlowast (119899 = 32) lt0001
Astrocytesnuclei ratioCA1 0277 plusmn 0025 (119899 = 62) 0324 plusmn 0025 (119899 = 56) 0286 plusmn 0029 (119899 = 52) 0417CA2 0447 plusmn 0053 (119899 = 40) 0345 plusmn 0021lowastlowastlowast (119899 = 40) 0282 plusmn 0024lowastlowastlowast (119899 = 36) 0008CA3 0292 plusmn 0016 (119899 = 62) 0262 plusmn 0016 (119899 = 56) 0264 plusmn 0016 (119899 = 52) 0347DGSGZ 0334 plusmn 0031 (119899 = 21) 0173 plusmn 0022lowastlowastlowast (119899 = 20) 0139 plusmn 0032lowastlowastlowast (119899 = 19) lt0001
DAPI-stained nuclei (equivalent to the total number of cells)and GFAP-labeled cells corresponding to astrocytes manu-ally and with Cell Profiler [13 15] Only process-bearing cellsshowing their soma in the plane of the analyzed area werecounted and their density was estimated (number per mm2)Six to eight randomly chosen fields located within each of theanalyzed hippocampal subfields were counted by progressivedisplacement of a test square gridThe astrocytesnuclei ratiowas calculated by dividing the number of GFAP+ cells bythe total number of nuclei labeled with DAPI GFAP+ cellswere identified as resting (with small somata bearing longthin and ramified processes) or reactive (with retraction ofprocesses to a length shorter than the diameter of the somata)[16] The number of reactive astrocytes was estimated for allthe experimental groups and normalized with the control
All the data corresponding to each treatment and regionwere pooled and are presented as the mean plusmn standard errorof the mean (SEM) Statistical analysis of data was performedusing a one-way ANOVA followed by a Bonferroni posttestwith Origin 70 software 119901 lt 005was considered statisticallysignificant
26Western Blot Thehippocampuswas dissected out as pre-viously described [17] The hippocampi from three differentrats for each experimental group (control DIA and FFR)were homogenized in ice cold lysis buffer (200mM glycine150mM NaCl 50mM EGTA 50mM EDTA and 300mMsucrose) with a protease inhibitor (Sigma-Aldrich USA)The homogenate was centrifuged at 10000timesg for 15 minutesat 4∘C and the supernatants were aliquoted and stored atminus80∘C Concentration of each protein sample was estimatedby Bradfordrsquos method [18] An equal concentration of protein(30 120583g) was resolved by 10 polyacrylamide gel electrophore-sis (PAGE)The proteins were transferred electrophoreticallyto PVDFmembrane then the membranes were blocked with5 nonfat dry milk in TBS-T for 3 h at room temperature
(RT) Membranes were incubated at 4∘C overnight withone of the following primary antibodies (a) polyclonalrabbit anti-GFAP antibody (dilution 1 2000 DakoCytoma-tion Fort Collins CO USA) (b) polyclonal rabbit anti-vimentin (dilution 1 2500 Cell Signaling Danvers MAUSA) (c) monoclonal mouse anti-nestin (dilution 1 5000BD Biosciences San Jose CA USA) and (d) polyclonal goatanti-actin (dilution 1 2500 Santacruz Dallas TX USA)The membranes were washed and bound antibodies weredetected by incubating for 3 h with either the goat anti-rabbitalkaline phosphatase conjugated antibody (dilution 1 2500Santacruz Dallas TX) goat anti-mouse alkaline phosphataseconjugated antibody (dilution 1 2500 Santacruz DallasTX) or rabbit anti-goat alkaline phosphatase conjugatedantibody (dilution 1 2500 Santacruz Dallas TX) Then themembrane was washed with TBS-T and the alkaline phos-phatase activity was detected with BCIPNBT AP-conjugatesubstrate reaction Kit (Bio-Rad USA) The intensity of eachband was measured using Image Lab 30 software
3 Results
The effects of DIA and FFR on the density of nuclei andastrocytes were estimated for the following hippocampalregions CA1 CA2 CA3 and dentate gyrus (DG) (Figure 1)All observations included the stratum radiatum and stratumoriens where astrocytes are preferentially located whilefor the DG the observations were performed in the hilus(Figure 1)
31 Anorexia Had No Effect on GFAP+ Cell Density inCA1 The hippocampal region CA1 showed no significantchange in nuclear density between the control (1099 plusmn 52nucleimm2 119899 = 62) DIA (924 plusmn 67 nucleimm2 119899 = 56)and FFR groups (932 plusmn 68 nucleimm2 119899 = 52) (119901 = 0075)(Figure 1 and Table 1) Additionally no significant changes
4 Neural Plasticity
500
1000
Nuc
leim
m 2
CTRL
DIA
FFR
DAPIso
sp
sp
sp
sr
so
sr
so
sr
Control DIA FFR(a)
GFAPso
sr
so
sr
so
sr
CTRL
DIA
FFR
100
200
300
Astr
ocyt
esm
m 2
Control DIA FFR
sp
sp
sp
(b)
Ratio
astro
cyte
snu
clei
02
04
Mergeso
sr
so
sr
so
sr
CTRL
DIA
FFR
Control DIA FFR
sp
sp
sp
(c)
Figure 2 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA1 Coronal sections of thehippocampus show nuclei labeled with DAPI (a) or immunostained with GFAP (b) The overlay of DAPI and GFAP is shown for controldehydration-induced anorexia (DIA) and forced food restricted (FFR) experimental groups (c)The densities of nuclei or GFAP+ cells as wellas the ratio of astrocytenuclei did not differ significantly among experimental groups (119901 = 0075 119901 = 0172 119901 = 0417 for the control DIAand FFR groups respectively see Table 1) Nuclear and astrocyte densities were estimated in the stratum oriens (so) and stratum radiatum(sr) stratum pyramidale (sp) is indicated as a reference Scale bar = 50 120583m Data are mean plusmn SEM
were observed for astrocyte density between the control(257 plusmn 14 nucleimm2 119899 = 82) DIA (253 plusmn 14 nucleimm2119899 = 72) and FFR groups (221 plusmn 15 nucleimm2 119899 = 66)(119901 = 0172) (Figure 2 and Table 1) We also determined theastrocytenuclei ratio by estimating the GFAP+ cells from thetotal nuclei The astrocytenuclei ratio for the control groupwas 0277 plusmn 0025 (119899 = 62) and it was not significantlydifferent for the DIA and FFR groups (0324 plusmn 0025 119899 = 56and 0286 plusmn 0029 119899 = 52 119901 = 0417) (Figure 2 and Table 1)
32 Anorexia Reduced GFAP+ Cell Density in CA2 Nucleardensity in CA2 was similar for the control (1125 plusmn 101nucleimm2 119899 = 40) DIA (1007 plusmn 51 nucleimm2 119899 = 40)
and FFR groups (1022 plusmn 56 nucleimm2 119899 = 36) (119901 = 0467)(Figure 3 Table 1) In contrast the astrocyte density estimatedfor the control group (405 plusmn 19 astrocytesmm2 119899 = 40) wassignificantly reduced for the DIA (323 plusmn 18 astrocytesmm2119899 = 40) and FFR groups (267 plusmn 19 astrocytemm2 119899 =36) (119901 lt 0001) (Figure 3 Table 1) Accordingly theastrocytenuclei ratio estimated for the control group (0447plusmn0053 119899 = 40) was also significantly reduced for the DIA(0345plusmn0021 119899 = 40) and FFR groups (0282plusmn0024 119899 = 36)(119901 = 0008) (Figure 3 Table 1)
33 Anorexia Reduced GFAP+ Cell Density in CA3 Thenuclear density of CA3 did not show significant changes
Neural Plasticity 5
DAPI
so
sp
sp
sp
sr
so
sr
so
sr
CTRL
DIA
FFR
500
1000
Nuc
leim
m 2
Control DIA FFR(a)
GFAP
so
sr
so
sr
so
sr
CTRL
DIA
FFR
200
400
Astr
ocyt
esm
m 2
lowastlowastlowast
lowastlowastlowast
Control DIA FFR
sp
sp
sp
(b)
Merge
so
sr
so
sr
so
sr
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
06
lowastlowastlowast
lowastlowastlowast
Control DIA FFR
sp
sp
sp
(c)
Figure 3 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA2 Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is shown for control dehydration-inducedanorexia (DIA) and forced-food restricted (FFR) experimental groups (c) The density of the nuclei was similar in all experimental groups(119901 = 0467) However a significant reduction of GFAP+ cells was observed for the DIA and FFR groups (119901 lt 0001) This reduction wasconsistently observed in the astrocytenuclei ratio (119901 = 0008) (see Table 1) Nuclear and astrocyte densities were estimated in the stratumoriens (so) and stratum radiatum (sr) stratum pyramidale (sp) is indicated as an anatomical reference Scale bar = 50120583m Data areexpressed as mean plusmn SEM
between the control (1288 plusmn 43 nucleimm2 119899 = 62) DIA(1084 plusmn 41 nucleimm2 119899 = 56) and FFR groups (1117 plusmn46 nucleimm2 119899 = 52) (119901 = 0001) (Figure 4 Table 1)However the astrocyte density of the control group (332plusmn 19astrocytesmm2 119899 = 82) was significantly reduced for theDIA (269plusmn15 astrocytesmm2 119899 = 72) and FFR groups (258plusmn15 astrocytemm2 119899 = 66) (119901 lt 0001) (Figure 4 Table 1)However the astrocytenuclei ratio estimated for the controlgroup (0292plusmn0016 119899 = 62) was not significantly reduced intheDIA (0262plusmn0016 119899 = 56) and FFR groups (0264plusmn0016119899 = 52) (119901 = 0347) (Figure 4 Table 1)
34 Anorexia Reduced GFAP+ Cell Density in DG Thenuclear density of the dentate gyrus did not differ signifi-cantly for the control (1433 plusmn 110 nucleimm2 119899 = 21)DIA (1535 plusmn 142 nucleimm2 119899 = 20) and FFR groups(1268 plusmn 161 nucleimm2 119899 = 19) (119901 = 0398) (Figure 5Table 1) The astrocyte density of the control group (476 plusmn 28astrocytesmm2 119899 = 41) was significantly reduced for theDIA (294 plusmn 124 astrocytesmm2 119899 = 36) and FFR groups(321 plusmn 26 astrocytemm2 119899 = 32) (119901 lt 0001) (Figure 5Table 1) Accordingly the astrocytenuclei ratio estimated forthe control group (0334 plusmn 0031 119899 = 21) was significantly
6 Neural Plasticity
DAPI
sosr
sosr
sosr
CTRL
DIA
FFR
500
1000
Nuc
leim
m 2
Control DIA FFR
sp
sp
sp
(a)
GFAP
sosr
sosr
sosr
CTRL
DIA
FFR
200
400
Astr
ocyt
esm
m 2
lowastlowastlowastlowastlowastlowast
FFRControl DIA
sp
sp
sp
(b)
Merge
sosr
sosr
sosr
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
FFRControl DIA
sp
sp
sp
(c)
Figure 4 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA3 Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is shown for control dehydration-inducedanorexia (DIA) and forced-food restricted (FFR) experimental groups (c) Neither the density of nuclei nor the astrocytenuclei ratio differedsignificantly among groups (119901 = 0931 and119901 = 0347 resp) However a significant reduction of GFAP+ cell density was observed (119901 lt 0001)(see Table 1) Nuclear and astrocyte densities were estimated in the stratum oriens (so) and stratum radiatum (sr) stratum pyramidale (sp)is indicated as an anatomical reference Scale bar = 50120583m Data are expressed as mean plusmn SEM
reduced for the DIA (0173 plusmn 0022 119899 = 20) and FFR groups(0139 plusmn 0032 119899 = 19) (119901 lt 0001) (Figure 5 Table 1)
341 Expression of Intermediate Filaments Is Affected byAnorexia The expression of intermediate filaments of astro-cytes was tested byWestern blot in the control DIA and FFRexperimental groups (Figure 6) The expression of GFAP wassignificantly reduced by DIA (048 plusmn 002 119899 = 3 119901 = 0003)and FFR (067plusmn004 119899 = 3119901 = 0006) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0668) (Figures 6(a) and6(b)) The opposite effect was observed for the expressionof vimentin and nestin The expression of vimentin was
significantly increased by DIA (151 plusmn 008 119899 = 3 119901 = 0009)and FFR (146plusmn011 119899 = 3119901 = 0008) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0896) (Figures 6(a)and 6(b)) Finally the expression of nestin was significantlyincreased by DIA (174 plusmn 01 119899 = 3 119901 = 0004) and FFR(161 plusmn 012 119899 = 3 119901 = 0001) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0468) (Figures 6(a) and6(b))
342 Anorexia Decreases GFAP Expression after the ThirdDay Our results showed a reduced GFAP+ cell density in
Neural Plasticity 7
DAPI
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
1000
1500
500
Nuc
leim
m 2
Control DIA FFR(a)
GFAP
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
200
400
600
Astr
ocyt
esm
m 2
lowastlowastlowastlowastlowastlowast
Control DIA FFR(b)
Merge
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
lowastlowastlowast
lowastlowastlowast
Control DIA FFR(c)
Figure 5 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal dentate gyrus (DG) Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is also shown for the control DIA and forcedfood-restricted (FFR) groups (c) The density of nuclei did not differ significantly among experimental groups (119901 = 0398) However asignificant reductionwas observed forGFAP+ cell density and the astrocytenuclei ratio (119901 lt 0001 and119901 lt 0001 resp) (see Table 1) Nuclearand astrocyte densities were estimated in the hilus subgranular zone (SGZ) and granular cell layer (GCL) are indicated as an anatomicalreference Scale bar = 50 120583m Data are expressed as mean plusmn SEM
the hippocampus that correlated with a lower expression ofthe protein Nevertheless GFAP expression may be affectedearlier than five daysThus the expression ofGFAPwas testedby Western blot after one and three days for control DIA orFFR experimental groups (Figures 7(a) and 7(b)) The resultsshowed that the expression of GFAP was unaffected after thefirst day for DIA (092 plusmn 004) and FFR (094 plusmn 003) whencompared to the normalized control (119901 gt 01) HoweverGFAP expression was significantly reduced after the third dayfor DIA (072 plusmn 001) and FFR (075 plusmn 004) when comparedto the normalized control (119901 lt 0001) (Figures 7(a) and7(b))
35 Anorexia Increases Reactive Astrocytes The reactiveastrocytes were estimated in CA2 andDG because astrocyte-nuclei ratio was significantly reduced in these regions Reac-tive astrocytes were defined as GFAP+ cells with processesshorter than the diameter of the somata [16] The numberof reactive astrocytes was estimated for all the experimentalgroups and normalized with their respective controls (seeSection 25) In CA2 reactive astrocytes increased 276 plusmn 07(119899 = 4 119901 = 0047) for DIA and 292 plusmn 075 (119899 = 4 119901 = 0032)for FFR while for dentate gyrus reactive astrocytes increased225plusmn015 (119899 = 4 119901 lt 00001) for DIA and 161plusmn029 (119899 = 4119901 = 0034) for FFR (Figure 8)
8 Neural Plasticity
GFAP
Vimentin
Actin
Nestin
RFFLRTC DIA
50kDa
57kDa
43kDa
200 kDa
(a)
CTRLDIA
FFR
GFAP Vimentin Nestin
10
20
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
lowastlowast
lowastlowast
(b)
Figure 6 Effect of anorexia on astrocyte intermediate filaments expression after the fifth day (a) Representative Western blot showing theexpression of GFAP vimentin and nestin for control DIA and FFR experimental groups actin was used as an internal control (b)The bandintensity of DIA and FFR groups was normalized to their respective controls Data were shown as mean plusmn SEM Significant differences wereconsidered as lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001
GFAP
Actin
After the first day
50kDa
43kDa
CTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
CTRL DIA FFR(a)
GFAP
Actin
50kDa
43kDa
After the third dayCTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
CTRL DIA FFR(b)
Figure 7 GFAP expression is reduced after the third day (a b) Representative Western blot showing the expression of GFAP after the first(a) and third day (b) of anorexia actin was used as an internal control The band intensity of DIA and FFR groups was normalized to theirrespective controls ((a b) under the Western blots) Data were shown as plusmn SEM Significant differences were considered as lowastlowastlowast119901 lt 0001
Neural Plasticity 9
RFFAIDlrtC
1
2
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in D
G
Ctrl DIA FFR
lowastlowastlowast
lowast
1
2
3
Ctrl DIA FFR
lowast
lowast
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in C
A2
Figure 8 Anorexia increases the density of reactive astrocytes Representative images of GFAP+ cells in the dentate gyrus (DG) for control(Ctrl) dehydration-induced anorexia (DIA) and forced food restriction (FFR) groupsThe density of reactive astrocytes (cells with processesshorter than the diameter of the somata) was estimated for all the experimental groups and normalized with their respective controls for DGand CA2 Data were shown as mean plusmn SEM Significant differences were considered as lowast119901 lt 005 lowastlowastlowast119901 lt 0001
4 Discussion
Anorexia nervosa is common among female adolescents andhas one of the highest mortality rates within psychiatric dis-orders (10ndash20) even surpassing depression [19ndash21] Duringpuberty mood swings and anxiety are common due to a sud-den rise of hormones which affects the hippocampus [11 1222] Moreover in the activity-based anorexia model (ABA)structural and functional alterations have been observedin the hippocampus accompanied by anxiety-like behavior[1 11 12 23] However the effect of anorexia on astrocytedensity has been barely explored Indeed previous studies inthe rat corpus callosum reported that cell proliferation wasreduced with the ABA model while astrocyte density wasregionally reduced in the DIA model [1 13] The ABA studyalso reported a reduced hippocampal cell proliferation in thedentate gyrus [1]
Results from the present study show that anorexiareduced astrocyte density in CA2 CA3 and DG whileno significant changes were observed in CA1 Accordinglyanorexia mutant mice (anxanx) showed lower expressionof c-Fos in the dentate gyrus and CA3 region of the hip-pocampus while no significant differences were found inCA1 [24] On the other hand the decrease in GFAP+ celldensity resulted in a reduced expression of GFAP proteinAstrocytes express three types of intermediate filament pro-teins GFAP vimentin and nestin [25] Nestin and vimentinare the main intermediate filaments in immature astrocyteswhereas maturing and adult astrocytes contain vimentin and
GFAP [25] Interestingly anorexia increased vimentin andnestin expression resembling reactive astrocytes observedafter injury [26 27] Accordingly GFAP+ cells deficit may bereplaced by vimentin+nestin+ cells in anorexia In supportof this hypothesis nestin reexpression has been observedfollowing experimental hippocampal lesions [25 28 29] andalso in human pathology such as multiple sclerosis [30]Moreover reactive astrocytes were increased by anorexia andforced food restriction indicating that GFAP+ cells react tosevere caloric restriction [31] Another factor to consider isthat female animals subjected to anorexia or chronic stressstop their ovarian cycles ending their exposure to hormonesthat may affect morphology and astrocyte density [32 33]Moreover hippocampal gliogenesis but not neurogenesisis reduced in the ABA model [1] Thus astrocytes possessdifferent levels of differentiation and maturation and theycan switch between these states even in the adult brain[26] Likewise astrocytes secrete gliotransmitters and growthfactors like BDNF required for synaptic plasticity synapto-genesis and neurogenesis [34 35] Thus a reduced densityof GFAP+ cells may affect hippocampal gliotransmission ofd-serine a gliotransmitter necessary for neuronal NMDA-mediated transmission and long term potentiation [34]Growth factors such as BDNF are also required for long termsynaptic plasticity of dendrites in the adult hippocampus [35]however decreased branching of dendrites was reported inan ABA model particularly in the stratum radiatum [11 12]Accordingly our results showed that GFAP+ cell density andGFAP expression are decreased by anorexia Similar results
10 Neural Plasticity
were reported in a rat model of depression where GFAPexpression was reduced [36] Thus we conclude that GFAP+cells and intermediate filament expression are affected byanorexia Our results add new information about the struc-tural changes observed in the hippocampus ofmurinemodelsof anorexia
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
The authors thank Dr P De Gortari (Insituto Nacionalde Psiquiatrıa Mexico) for introducing them to the DIAmodel The authors are grateful to M Macedo-MendozaM E Ramos-Aguilar E N Hernandez-Rıos A Castilla LCasanova A E Espino and M Garcıa-Servın for technicalsupport The authors thank Dr D Pless for reviewing thepaper The authors thank Dr G Ramırez-Rodrıguez (Insti-tuto Nacional de Psiquiatrıa Mexico) for donating the nestinand vimentin antibodies used in this study Janina Krugerwas an internship student supported by the DAAD-RISEprogram Durairaj Ragu Varman is a postdoctoral fellowsupported by DGAPA-UNAM This work was supported bygrants from Programa de Apoyo a Proyectos de Investigacione Innovacion Tecnologica de la UNAM (PAPIIT-UNAMIN200913 and IN201915) to Ataulfo Martınez-Torres andDaniel Reyes-Haro
References
[1] N C Barbarich-Marsteller C A Fornal L F Takase et alldquoActivity-based anorexia is associated with reduced hippocam-pal cell proliferation in adolescent female ratsrdquo BehaviouralBrain Research vol 236 no 1 pp 251ndash257 2013
[2] C Aoki N Sabaliauskas T Chowdhury et al ldquoAdolescentfemale rats exhibiting activity-based anorexia express elevatedlevels of GABAAreceptor 1205724 and 120575 subunits at the plasmamembrane of hippocampal CA1 spinesrdquo Synapse vol 66 no 5pp 391ndash407 2012
[3] G D Giordano P Renzetti R C Parodi et al ldquoVolume mea-surement with magnetic resonance imaging of hippocampus-amygdala formation in patients with anorexia nervosardquo Journalof Endocrinological Investigation vol 24 no 7 pp 510ndash514 2001
[4] F Connan F Murphy S E J Connor et al ldquoHippocampalvolume and cognitive function in anorexia nervosardquo PsychiatryResearchmdashNeuroimaging vol 146 no 2 pp 117ndash125 2006
[5] J N Beadle S Paradiso M Brumm M Voss K Halmiand L M McCormick ldquoLarger hippocampus size in womenwith anorexia nervosa who exercise excessively than healthywomenrdquo Psychiatry ResearchmdashNeuroimaging vol 232 no 2 pp193ndash199 2015
[6] N T Burkert KKoschutnig F Ebner andW Freidl ldquoStructuralhippocampal alterations perceived stress and coping deficien-cies in patients with anorexia nervosardquo International Journal ofEating Disorders vol 48 no 6 pp 670ndash676 2015
[7] S B McHugh M Fillenz J P Lowry J N P Rawlins and DM Bannerman ldquoBrain tissue oxygen amperometry in behavingrats demonstrates functional dissociation of dorsal and ventral
hippocampus during spatial processing and anxietyrdquo EuropeanJournal of Neuroscience vol 33 no 2 pp 322ndash337 2011
[8] A G Watts ldquoDehydration-associated anorexia developmentand rapid reversalrdquo Physiology and Behavior vol 65 no 4-5 pp871ndash878 1998
[9] L Jaimes-Hoy P Joseph-Bravo and P de Gortari ldquoDiffer-ential response of TRHergic neurons of the hypothalamicparaventricular nucleus (PVN) in female animals submittedto food-restriction or dehydration-induced anorexia and coldexposurerdquo Hormones and Behavior vol 53 no 2 pp 366ndash3772008
[10] A G Watts and C N Boyle ldquoThe functional architecture ofdehydration-anorexiardquo Physiology and Behavior vol 100 no 5pp 472ndash477 2010
[11] T G Chowdhury M B Rıos T E Chan D S Cassataro NC Barbarich-Marsteller and C Aoki ldquoActivity-based anorexiaduring adolescence disrupts normal development of the CA1pyramidal cells in the ventral hippocampus of female ratsrdquoHippocampus vol 24 no 12 pp 1421ndash1429 2014
[12] T G Chowdhury N C Barbarich-Marsteller T E Chan andCAoki ldquoActivity-based anorexia has differential effects on apicaldendritic branching in dorsal and ventral hippocampal CA1rdquoBrain Structure and Function vol 219 no 6 pp 1935ndash1945 2014
[13] D Reyes-Haro F E Labrada-Moncada R Miledi andA Martınez-Torres ldquoDehydration-induced anorexia reducesastrocyte density in the rat corpus callosumrdquo Neural Plasticityvol 2015 Article ID 474917 8 pages 2015
[14] P de Gortari K Mancera A Cote-Velez et al ldquoInvolvement ofCRH-R2 receptor in eating behavior and in the response of theHPT axis in rats subjected to dehydration-induced anorexiardquoPsychoneuroendocrinology vol 34 no 2 pp 259ndash272 2009
[15] M R Lamprecht DM Sabatini and A E Carpenter ldquoCellPro-filer free versatile software for automated biological imageanalysisrdquo BioTechniques vol 42 no 1 pp 71ndash75 2007
[16] B Rinaldi F Guida A Furiano et al ldquoEffect of prolongedmoderate exercise on the changes of nonneuronal cells in earlymyocardial infarctionrdquo Neural Plasticity vol 2015 Article ID265967 8 pages 2015
[17] K Chiu W M Lau H T Lau K-F So and R C-C ChangldquoMicro-dissection of rat brain for RNA or protein extractionfrom specific brain regionrdquo Journal of Visualized Experimentsno 7 article e269 2007
[18] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[19] P F Sullivan ldquoMortality in anorexia nervosardquo The AmericanJournal of Psychiatry vol 152 no 7 pp 1073ndash1074 1995
[20] C L Birmingham J Su J A Hlynsky E M Goldner and MGao ldquoThe mortality rate from anorexia nervosardquo InternationalJournal of Eating Disorders vol 38 no 2 pp 143ndash146 2005
[21] C M Bulik M C T Slof-Oprsquot Landt E F van Furth and P FSullivan ldquoThe genetics of anorexia nervosardquo Annual Review ofNutrition vol 27 pp 263ndash275 2007
[22] K J McLaughlin J O Wilson J Harman et al ldquoChronic 17120573-estradiol or cholesterol prevents stress-induced hippocampalCA3 dendritic retraction in ovariectomized female rats possi-ble correspondence between CA1 spine properties and spatialacquisitionrdquo Hippocampus vol 20 no 6 pp 768ndash786 2010
[23] K P Kinzig and S L Hargrave ldquoAdolescent activity-basedanorexia increases anxiety-like behavior in adulthoodrdquo Physi-ology and Behavior vol 101 no 2 pp 269ndash276 2010
Neural Plasticity 11
[24] S A Kim Y M Choi H J Park et al ldquoDecrease of c-Fosexpression in hippocampus of anorexia (anxanx)micerdquoKoreanJournal of Biological Sciences vol 5 no 2 pp 157ndash161 2001
[25] C Eliasson C Sahlgren C-H Berthold et al ldquoIntermediatefilament protein partnership in astrocytesrdquo The Journal ofBiological Chemistry vol 274 no 34 pp 23996ndash24006 1999
[26] R Schmidt-Kastner andCHumpel ldquoNestin expression persistsin astrocytes of organotypic slice cultures from rat cortexrdquoInternational Journal of Developmental Neuroscience vol 20 no1 pp 29ndash38 2002
[27] Y Takamiya S Kohsaka S Toya M Otani and Y TsukadaldquoImmunohistochemical studies on the proliferation of reactiveastrocytes and the expression of cytoskeletal proteins followingbrain injury in ratsrdquo Brain Research vol 466 no 2 pp 201ndash2101988
[28] S R Clarke A K Shetty J L Bradley and D A TurnerldquoReactive astrocytes express the embryonic intermediate neuro-filament nestinrdquoNeuroReport vol 5 no 15 pp 1885ndash1888 1994
[29] M C Geloso V Corvino V Cavallo et al ldquoExpression ofastrocytic nestin in the rat hippocampus during trimethyltin-induced neurodegenerationrdquo Neuroscience Letters vol 357 no2 pp 103ndash106 2004
[30] J E Holley D Gveric J Newcombe M L Cuzner and N JGutowski ldquoAstrocyte characterization in the multiple sclerosisglial scarrdquoNeuropathology andAppliedNeurobiology vol 29 no5 pp 434ndash444 2003
[31] C N Boyle S M Lorenzen D Compton and A G WattsldquoDehydration-anorexia derives from a reduction in meal sizebut not meal numberrdquo Physiology and Behavior vol 105 no 2pp 305ndash314 2012
[32] I AzcoitiaM Santos-GalindoMAArevalo and LMGarcıa-Segura ldquoRole of astroglia in the neuroplastic and neuroprotec-tive actions of estradiolrdquo European Journal of Neuroscience vol32 no 12 pp 1995ndash2002 2010
[33] L Eiland J Ramroop M N Hill J Manley and B S McEwenldquoChronic juvenile stress produces corticolimbic dendritic archi-tectural remodeling and modulates emotional behavior in maleand female ratsrdquo Psychoneuroendocrinology vol 37 no 1 pp39ndash47 2012
[34] C Henneberger T Papouin S H R Oliet and D A RusakovldquoLong-term potentiation depends on release of d-serine fromastrocytesrdquo Nature vol 463 no 7278 pp 232ndash236 2010
[35] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo The Journal ofNeuroscience vol 22 no 5 pp 1532ndash1540 2002
[36] Y Ye G Wang H Wang and X Wang ldquoBrain-derived neu-rotrophic factor (BDNF) infusion restored astrocytic plasticityin the hippocampus of a rat model of depressionrdquo NeuroscienceLetters vol 503 no 1 pp 15ndash19 2011
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

4 Neural Plasticity
500
1000
Nuc
leim
m 2
CTRL
DIA
FFR
DAPIso
sp
sp
sp
sr
so
sr
so
sr
Control DIA FFR(a)
GFAPso
sr
so
sr
so
sr
CTRL
DIA
FFR
100
200
300
Astr
ocyt
esm
m 2
Control DIA FFR
sp
sp
sp
(b)
Ratio
astro
cyte
snu
clei
02
04
Mergeso
sr
so
sr
so
sr
CTRL
DIA
FFR
Control DIA FFR
sp
sp
sp
(c)
Figure 2 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA1 Coronal sections of thehippocampus show nuclei labeled with DAPI (a) or immunostained with GFAP (b) The overlay of DAPI and GFAP is shown for controldehydration-induced anorexia (DIA) and forced food restricted (FFR) experimental groups (c)The densities of nuclei or GFAP+ cells as wellas the ratio of astrocytenuclei did not differ significantly among experimental groups (119901 = 0075 119901 = 0172 119901 = 0417 for the control DIAand FFR groups respectively see Table 1) Nuclear and astrocyte densities were estimated in the stratum oriens (so) and stratum radiatum(sr) stratum pyramidale (sp) is indicated as a reference Scale bar = 50 120583m Data are mean plusmn SEM
were observed for astrocyte density between the control(257 plusmn 14 nucleimm2 119899 = 82) DIA (253 plusmn 14 nucleimm2119899 = 72) and FFR groups (221 plusmn 15 nucleimm2 119899 = 66)(119901 = 0172) (Figure 2 and Table 1) We also determined theastrocytenuclei ratio by estimating the GFAP+ cells from thetotal nuclei The astrocytenuclei ratio for the control groupwas 0277 plusmn 0025 (119899 = 62) and it was not significantlydifferent for the DIA and FFR groups (0324 plusmn 0025 119899 = 56and 0286 plusmn 0029 119899 = 52 119901 = 0417) (Figure 2 and Table 1)
32 Anorexia Reduced GFAP+ Cell Density in CA2 Nucleardensity in CA2 was similar for the control (1125 plusmn 101nucleimm2 119899 = 40) DIA (1007 plusmn 51 nucleimm2 119899 = 40)
and FFR groups (1022 plusmn 56 nucleimm2 119899 = 36) (119901 = 0467)(Figure 3 Table 1) In contrast the astrocyte density estimatedfor the control group (405 plusmn 19 astrocytesmm2 119899 = 40) wassignificantly reduced for the DIA (323 plusmn 18 astrocytesmm2119899 = 40) and FFR groups (267 plusmn 19 astrocytemm2 119899 =36) (119901 lt 0001) (Figure 3 Table 1) Accordingly theastrocytenuclei ratio estimated for the control group (0447plusmn0053 119899 = 40) was also significantly reduced for the DIA(0345plusmn0021 119899 = 40) and FFR groups (0282plusmn0024 119899 = 36)(119901 = 0008) (Figure 3 Table 1)
33 Anorexia Reduced GFAP+ Cell Density in CA3 Thenuclear density of CA3 did not show significant changes
Neural Plasticity 5
DAPI
so
sp
sp
sp
sr
so
sr
so
sr
CTRL
DIA
FFR
500
1000
Nuc
leim
m 2
Control DIA FFR(a)
GFAP
so
sr
so
sr
so
sr
CTRL
DIA
FFR
200
400
Astr
ocyt
esm
m 2
lowastlowastlowast
lowastlowastlowast
Control DIA FFR
sp
sp
sp
(b)
Merge
so
sr
so
sr
so
sr
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
06
lowastlowastlowast
lowastlowastlowast
Control DIA FFR
sp
sp
sp
(c)
Figure 3 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA2 Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is shown for control dehydration-inducedanorexia (DIA) and forced-food restricted (FFR) experimental groups (c) The density of the nuclei was similar in all experimental groups(119901 = 0467) However a significant reduction of GFAP+ cells was observed for the DIA and FFR groups (119901 lt 0001) This reduction wasconsistently observed in the astrocytenuclei ratio (119901 = 0008) (see Table 1) Nuclear and astrocyte densities were estimated in the stratumoriens (so) and stratum radiatum (sr) stratum pyramidale (sp) is indicated as an anatomical reference Scale bar = 50120583m Data areexpressed as mean plusmn SEM
between the control (1288 plusmn 43 nucleimm2 119899 = 62) DIA(1084 plusmn 41 nucleimm2 119899 = 56) and FFR groups (1117 plusmn46 nucleimm2 119899 = 52) (119901 = 0001) (Figure 4 Table 1)However the astrocyte density of the control group (332plusmn 19astrocytesmm2 119899 = 82) was significantly reduced for theDIA (269plusmn15 astrocytesmm2 119899 = 72) and FFR groups (258plusmn15 astrocytemm2 119899 = 66) (119901 lt 0001) (Figure 4 Table 1)However the astrocytenuclei ratio estimated for the controlgroup (0292plusmn0016 119899 = 62) was not significantly reduced intheDIA (0262plusmn0016 119899 = 56) and FFR groups (0264plusmn0016119899 = 52) (119901 = 0347) (Figure 4 Table 1)
34 Anorexia Reduced GFAP+ Cell Density in DG Thenuclear density of the dentate gyrus did not differ signifi-cantly for the control (1433 plusmn 110 nucleimm2 119899 = 21)DIA (1535 plusmn 142 nucleimm2 119899 = 20) and FFR groups(1268 plusmn 161 nucleimm2 119899 = 19) (119901 = 0398) (Figure 5Table 1) The astrocyte density of the control group (476 plusmn 28astrocytesmm2 119899 = 41) was significantly reduced for theDIA (294 plusmn 124 astrocytesmm2 119899 = 36) and FFR groups(321 plusmn 26 astrocytemm2 119899 = 32) (119901 lt 0001) (Figure 5Table 1) Accordingly the astrocytenuclei ratio estimated forthe control group (0334 plusmn 0031 119899 = 21) was significantly
6 Neural Plasticity
DAPI
sosr
sosr
sosr
CTRL
DIA
FFR
500
1000
Nuc
leim
m 2
Control DIA FFR
sp
sp
sp
(a)
GFAP
sosr
sosr
sosr
CTRL
DIA
FFR
200
400
Astr
ocyt
esm
m 2
lowastlowastlowastlowastlowastlowast
FFRControl DIA
sp
sp
sp
(b)
Merge
sosr
sosr
sosr
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
FFRControl DIA
sp
sp
sp
(c)
Figure 4 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA3 Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is shown for control dehydration-inducedanorexia (DIA) and forced-food restricted (FFR) experimental groups (c) Neither the density of nuclei nor the astrocytenuclei ratio differedsignificantly among groups (119901 = 0931 and119901 = 0347 resp) However a significant reduction of GFAP+ cell density was observed (119901 lt 0001)(see Table 1) Nuclear and astrocyte densities were estimated in the stratum oriens (so) and stratum radiatum (sr) stratum pyramidale (sp)is indicated as an anatomical reference Scale bar = 50120583m Data are expressed as mean plusmn SEM
reduced for the DIA (0173 plusmn 0022 119899 = 20) and FFR groups(0139 plusmn 0032 119899 = 19) (119901 lt 0001) (Figure 5 Table 1)
341 Expression of Intermediate Filaments Is Affected byAnorexia The expression of intermediate filaments of astro-cytes was tested byWestern blot in the control DIA and FFRexperimental groups (Figure 6) The expression of GFAP wassignificantly reduced by DIA (048 plusmn 002 119899 = 3 119901 = 0003)and FFR (067plusmn004 119899 = 3119901 = 0006) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0668) (Figures 6(a) and6(b)) The opposite effect was observed for the expressionof vimentin and nestin The expression of vimentin was
significantly increased by DIA (151 plusmn 008 119899 = 3 119901 = 0009)and FFR (146plusmn011 119899 = 3119901 = 0008) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0896) (Figures 6(a)and 6(b)) Finally the expression of nestin was significantlyincreased by DIA (174 plusmn 01 119899 = 3 119901 = 0004) and FFR(161 plusmn 012 119899 = 3 119901 = 0001) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0468) (Figures 6(a) and6(b))
342 Anorexia Decreases GFAP Expression after the ThirdDay Our results showed a reduced GFAP+ cell density in
Neural Plasticity 7
DAPI
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
1000
1500
500
Nuc
leim
m 2
Control DIA FFR(a)
GFAP
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
200
400
600
Astr
ocyt
esm
m 2
lowastlowastlowastlowastlowastlowast
Control DIA FFR(b)
Merge
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
lowastlowastlowast
lowastlowastlowast
Control DIA FFR(c)
Figure 5 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal dentate gyrus (DG) Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is also shown for the control DIA and forcedfood-restricted (FFR) groups (c) The density of nuclei did not differ significantly among experimental groups (119901 = 0398) However asignificant reductionwas observed forGFAP+ cell density and the astrocytenuclei ratio (119901 lt 0001 and119901 lt 0001 resp) (see Table 1) Nuclearand astrocyte densities were estimated in the hilus subgranular zone (SGZ) and granular cell layer (GCL) are indicated as an anatomicalreference Scale bar = 50 120583m Data are expressed as mean plusmn SEM
the hippocampus that correlated with a lower expression ofthe protein Nevertheless GFAP expression may be affectedearlier than five daysThus the expression ofGFAPwas testedby Western blot after one and three days for control DIA orFFR experimental groups (Figures 7(a) and 7(b)) The resultsshowed that the expression of GFAP was unaffected after thefirst day for DIA (092 plusmn 004) and FFR (094 plusmn 003) whencompared to the normalized control (119901 gt 01) HoweverGFAP expression was significantly reduced after the third dayfor DIA (072 plusmn 001) and FFR (075 plusmn 004) when comparedto the normalized control (119901 lt 0001) (Figures 7(a) and7(b))
35 Anorexia Increases Reactive Astrocytes The reactiveastrocytes were estimated in CA2 andDG because astrocyte-nuclei ratio was significantly reduced in these regions Reac-tive astrocytes were defined as GFAP+ cells with processesshorter than the diameter of the somata [16] The numberof reactive astrocytes was estimated for all the experimentalgroups and normalized with their respective controls (seeSection 25) In CA2 reactive astrocytes increased 276 plusmn 07(119899 = 4 119901 = 0047) for DIA and 292 plusmn 075 (119899 = 4 119901 = 0032)for FFR while for dentate gyrus reactive astrocytes increased225plusmn015 (119899 = 4 119901 lt 00001) for DIA and 161plusmn029 (119899 = 4119901 = 0034) for FFR (Figure 8)
8 Neural Plasticity
GFAP
Vimentin
Actin
Nestin
RFFLRTC DIA
50kDa
57kDa
43kDa
200 kDa
(a)
CTRLDIA
FFR
GFAP Vimentin Nestin
10
20
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
lowastlowast
lowastlowast
(b)
Figure 6 Effect of anorexia on astrocyte intermediate filaments expression after the fifth day (a) Representative Western blot showing theexpression of GFAP vimentin and nestin for control DIA and FFR experimental groups actin was used as an internal control (b)The bandintensity of DIA and FFR groups was normalized to their respective controls Data were shown as mean plusmn SEM Significant differences wereconsidered as lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001
GFAP
Actin
After the first day
50kDa
43kDa
CTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
CTRL DIA FFR(a)
GFAP
Actin
50kDa
43kDa
After the third dayCTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
CTRL DIA FFR(b)
Figure 7 GFAP expression is reduced after the third day (a b) Representative Western blot showing the expression of GFAP after the first(a) and third day (b) of anorexia actin was used as an internal control The band intensity of DIA and FFR groups was normalized to theirrespective controls ((a b) under the Western blots) Data were shown as plusmn SEM Significant differences were considered as lowastlowastlowast119901 lt 0001
Neural Plasticity 9
RFFAIDlrtC
1
2
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in D
G
Ctrl DIA FFR
lowastlowastlowast
lowast
1
2
3
Ctrl DIA FFR
lowast
lowast
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in C
A2
Figure 8 Anorexia increases the density of reactive astrocytes Representative images of GFAP+ cells in the dentate gyrus (DG) for control(Ctrl) dehydration-induced anorexia (DIA) and forced food restriction (FFR) groupsThe density of reactive astrocytes (cells with processesshorter than the diameter of the somata) was estimated for all the experimental groups and normalized with their respective controls for DGand CA2 Data were shown as mean plusmn SEM Significant differences were considered as lowast119901 lt 005 lowastlowastlowast119901 lt 0001
4 Discussion
Anorexia nervosa is common among female adolescents andhas one of the highest mortality rates within psychiatric dis-orders (10ndash20) even surpassing depression [19ndash21] Duringpuberty mood swings and anxiety are common due to a sud-den rise of hormones which affects the hippocampus [11 1222] Moreover in the activity-based anorexia model (ABA)structural and functional alterations have been observedin the hippocampus accompanied by anxiety-like behavior[1 11 12 23] However the effect of anorexia on astrocytedensity has been barely explored Indeed previous studies inthe rat corpus callosum reported that cell proliferation wasreduced with the ABA model while astrocyte density wasregionally reduced in the DIA model [1 13] The ABA studyalso reported a reduced hippocampal cell proliferation in thedentate gyrus [1]
Results from the present study show that anorexiareduced astrocyte density in CA2 CA3 and DG whileno significant changes were observed in CA1 Accordinglyanorexia mutant mice (anxanx) showed lower expressionof c-Fos in the dentate gyrus and CA3 region of the hip-pocampus while no significant differences were found inCA1 [24] On the other hand the decrease in GFAP+ celldensity resulted in a reduced expression of GFAP proteinAstrocytes express three types of intermediate filament pro-teins GFAP vimentin and nestin [25] Nestin and vimentinare the main intermediate filaments in immature astrocyteswhereas maturing and adult astrocytes contain vimentin and
GFAP [25] Interestingly anorexia increased vimentin andnestin expression resembling reactive astrocytes observedafter injury [26 27] Accordingly GFAP+ cells deficit may bereplaced by vimentin+nestin+ cells in anorexia In supportof this hypothesis nestin reexpression has been observedfollowing experimental hippocampal lesions [25 28 29] andalso in human pathology such as multiple sclerosis [30]Moreover reactive astrocytes were increased by anorexia andforced food restriction indicating that GFAP+ cells react tosevere caloric restriction [31] Another factor to consider isthat female animals subjected to anorexia or chronic stressstop their ovarian cycles ending their exposure to hormonesthat may affect morphology and astrocyte density [32 33]Moreover hippocampal gliogenesis but not neurogenesisis reduced in the ABA model [1] Thus astrocytes possessdifferent levels of differentiation and maturation and theycan switch between these states even in the adult brain[26] Likewise astrocytes secrete gliotransmitters and growthfactors like BDNF required for synaptic plasticity synapto-genesis and neurogenesis [34 35] Thus a reduced densityof GFAP+ cells may affect hippocampal gliotransmission ofd-serine a gliotransmitter necessary for neuronal NMDA-mediated transmission and long term potentiation [34]Growth factors such as BDNF are also required for long termsynaptic plasticity of dendrites in the adult hippocampus [35]however decreased branching of dendrites was reported inan ABA model particularly in the stratum radiatum [11 12]Accordingly our results showed that GFAP+ cell density andGFAP expression are decreased by anorexia Similar results
10 Neural Plasticity
were reported in a rat model of depression where GFAPexpression was reduced [36] Thus we conclude that GFAP+cells and intermediate filament expression are affected byanorexia Our results add new information about the struc-tural changes observed in the hippocampus ofmurinemodelsof anorexia
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
The authors thank Dr P De Gortari (Insituto Nacionalde Psiquiatrıa Mexico) for introducing them to the DIAmodel The authors are grateful to M Macedo-MendozaM E Ramos-Aguilar E N Hernandez-Rıos A Castilla LCasanova A E Espino and M Garcıa-Servın for technicalsupport The authors thank Dr D Pless for reviewing thepaper The authors thank Dr G Ramırez-Rodrıguez (Insti-tuto Nacional de Psiquiatrıa Mexico) for donating the nestinand vimentin antibodies used in this study Janina Krugerwas an internship student supported by the DAAD-RISEprogram Durairaj Ragu Varman is a postdoctoral fellowsupported by DGAPA-UNAM This work was supported bygrants from Programa de Apoyo a Proyectos de Investigacione Innovacion Tecnologica de la UNAM (PAPIIT-UNAMIN200913 and IN201915) to Ataulfo Martınez-Torres andDaniel Reyes-Haro
References
[1] N C Barbarich-Marsteller C A Fornal L F Takase et alldquoActivity-based anorexia is associated with reduced hippocam-pal cell proliferation in adolescent female ratsrdquo BehaviouralBrain Research vol 236 no 1 pp 251ndash257 2013
[2] C Aoki N Sabaliauskas T Chowdhury et al ldquoAdolescentfemale rats exhibiting activity-based anorexia express elevatedlevels of GABAAreceptor 1205724 and 120575 subunits at the plasmamembrane of hippocampal CA1 spinesrdquo Synapse vol 66 no 5pp 391ndash407 2012
[3] G D Giordano P Renzetti R C Parodi et al ldquoVolume mea-surement with magnetic resonance imaging of hippocampus-amygdala formation in patients with anorexia nervosardquo Journalof Endocrinological Investigation vol 24 no 7 pp 510ndash514 2001
[4] F Connan F Murphy S E J Connor et al ldquoHippocampalvolume and cognitive function in anorexia nervosardquo PsychiatryResearchmdashNeuroimaging vol 146 no 2 pp 117ndash125 2006
[5] J N Beadle S Paradiso M Brumm M Voss K Halmiand L M McCormick ldquoLarger hippocampus size in womenwith anorexia nervosa who exercise excessively than healthywomenrdquo Psychiatry ResearchmdashNeuroimaging vol 232 no 2 pp193ndash199 2015
[6] N T Burkert KKoschutnig F Ebner andW Freidl ldquoStructuralhippocampal alterations perceived stress and coping deficien-cies in patients with anorexia nervosardquo International Journal ofEating Disorders vol 48 no 6 pp 670ndash676 2015
[7] S B McHugh M Fillenz J P Lowry J N P Rawlins and DM Bannerman ldquoBrain tissue oxygen amperometry in behavingrats demonstrates functional dissociation of dorsal and ventral
hippocampus during spatial processing and anxietyrdquo EuropeanJournal of Neuroscience vol 33 no 2 pp 322ndash337 2011
[8] A G Watts ldquoDehydration-associated anorexia developmentand rapid reversalrdquo Physiology and Behavior vol 65 no 4-5 pp871ndash878 1998
[9] L Jaimes-Hoy P Joseph-Bravo and P de Gortari ldquoDiffer-ential response of TRHergic neurons of the hypothalamicparaventricular nucleus (PVN) in female animals submittedto food-restriction or dehydration-induced anorexia and coldexposurerdquo Hormones and Behavior vol 53 no 2 pp 366ndash3772008
[10] A G Watts and C N Boyle ldquoThe functional architecture ofdehydration-anorexiardquo Physiology and Behavior vol 100 no 5pp 472ndash477 2010
[11] T G Chowdhury M B Rıos T E Chan D S Cassataro NC Barbarich-Marsteller and C Aoki ldquoActivity-based anorexiaduring adolescence disrupts normal development of the CA1pyramidal cells in the ventral hippocampus of female ratsrdquoHippocampus vol 24 no 12 pp 1421ndash1429 2014
[12] T G Chowdhury N C Barbarich-Marsteller T E Chan andCAoki ldquoActivity-based anorexia has differential effects on apicaldendritic branching in dorsal and ventral hippocampal CA1rdquoBrain Structure and Function vol 219 no 6 pp 1935ndash1945 2014
[13] D Reyes-Haro F E Labrada-Moncada R Miledi andA Martınez-Torres ldquoDehydration-induced anorexia reducesastrocyte density in the rat corpus callosumrdquo Neural Plasticityvol 2015 Article ID 474917 8 pages 2015
[14] P de Gortari K Mancera A Cote-Velez et al ldquoInvolvement ofCRH-R2 receptor in eating behavior and in the response of theHPT axis in rats subjected to dehydration-induced anorexiardquoPsychoneuroendocrinology vol 34 no 2 pp 259ndash272 2009
[15] M R Lamprecht DM Sabatini and A E Carpenter ldquoCellPro-filer free versatile software for automated biological imageanalysisrdquo BioTechniques vol 42 no 1 pp 71ndash75 2007
[16] B Rinaldi F Guida A Furiano et al ldquoEffect of prolongedmoderate exercise on the changes of nonneuronal cells in earlymyocardial infarctionrdquo Neural Plasticity vol 2015 Article ID265967 8 pages 2015
[17] K Chiu W M Lau H T Lau K-F So and R C-C ChangldquoMicro-dissection of rat brain for RNA or protein extractionfrom specific brain regionrdquo Journal of Visualized Experimentsno 7 article e269 2007
[18] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[19] P F Sullivan ldquoMortality in anorexia nervosardquo The AmericanJournal of Psychiatry vol 152 no 7 pp 1073ndash1074 1995
[20] C L Birmingham J Su J A Hlynsky E M Goldner and MGao ldquoThe mortality rate from anorexia nervosardquo InternationalJournal of Eating Disorders vol 38 no 2 pp 143ndash146 2005
[21] C M Bulik M C T Slof-Oprsquot Landt E F van Furth and P FSullivan ldquoThe genetics of anorexia nervosardquo Annual Review ofNutrition vol 27 pp 263ndash275 2007
[22] K J McLaughlin J O Wilson J Harman et al ldquoChronic 17120573-estradiol or cholesterol prevents stress-induced hippocampalCA3 dendritic retraction in ovariectomized female rats possi-ble correspondence between CA1 spine properties and spatialacquisitionrdquo Hippocampus vol 20 no 6 pp 768ndash786 2010
[23] K P Kinzig and S L Hargrave ldquoAdolescent activity-basedanorexia increases anxiety-like behavior in adulthoodrdquo Physi-ology and Behavior vol 101 no 2 pp 269ndash276 2010
Neural Plasticity 11
[24] S A Kim Y M Choi H J Park et al ldquoDecrease of c-Fosexpression in hippocampus of anorexia (anxanx)micerdquoKoreanJournal of Biological Sciences vol 5 no 2 pp 157ndash161 2001
[25] C Eliasson C Sahlgren C-H Berthold et al ldquoIntermediatefilament protein partnership in astrocytesrdquo The Journal ofBiological Chemistry vol 274 no 34 pp 23996ndash24006 1999
[26] R Schmidt-Kastner andCHumpel ldquoNestin expression persistsin astrocytes of organotypic slice cultures from rat cortexrdquoInternational Journal of Developmental Neuroscience vol 20 no1 pp 29ndash38 2002
[27] Y Takamiya S Kohsaka S Toya M Otani and Y TsukadaldquoImmunohistochemical studies on the proliferation of reactiveastrocytes and the expression of cytoskeletal proteins followingbrain injury in ratsrdquo Brain Research vol 466 no 2 pp 201ndash2101988
[28] S R Clarke A K Shetty J L Bradley and D A TurnerldquoReactive astrocytes express the embryonic intermediate neuro-filament nestinrdquoNeuroReport vol 5 no 15 pp 1885ndash1888 1994
[29] M C Geloso V Corvino V Cavallo et al ldquoExpression ofastrocytic nestin in the rat hippocampus during trimethyltin-induced neurodegenerationrdquo Neuroscience Letters vol 357 no2 pp 103ndash106 2004
[30] J E Holley D Gveric J Newcombe M L Cuzner and N JGutowski ldquoAstrocyte characterization in the multiple sclerosisglial scarrdquoNeuropathology andAppliedNeurobiology vol 29 no5 pp 434ndash444 2003
[31] C N Boyle S M Lorenzen D Compton and A G WattsldquoDehydration-anorexia derives from a reduction in meal sizebut not meal numberrdquo Physiology and Behavior vol 105 no 2pp 305ndash314 2012
[32] I AzcoitiaM Santos-GalindoMAArevalo and LMGarcıa-Segura ldquoRole of astroglia in the neuroplastic and neuroprotec-tive actions of estradiolrdquo European Journal of Neuroscience vol32 no 12 pp 1995ndash2002 2010
[33] L Eiland J Ramroop M N Hill J Manley and B S McEwenldquoChronic juvenile stress produces corticolimbic dendritic archi-tectural remodeling and modulates emotional behavior in maleand female ratsrdquo Psychoneuroendocrinology vol 37 no 1 pp39ndash47 2012
[34] C Henneberger T Papouin S H R Oliet and D A RusakovldquoLong-term potentiation depends on release of d-serine fromastrocytesrdquo Nature vol 463 no 7278 pp 232ndash236 2010
[35] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo The Journal ofNeuroscience vol 22 no 5 pp 1532ndash1540 2002
[36] Y Ye G Wang H Wang and X Wang ldquoBrain-derived neu-rotrophic factor (BDNF) infusion restored astrocytic plasticityin the hippocampus of a rat model of depressionrdquo NeuroscienceLetters vol 503 no 1 pp 15ndash19 2011
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
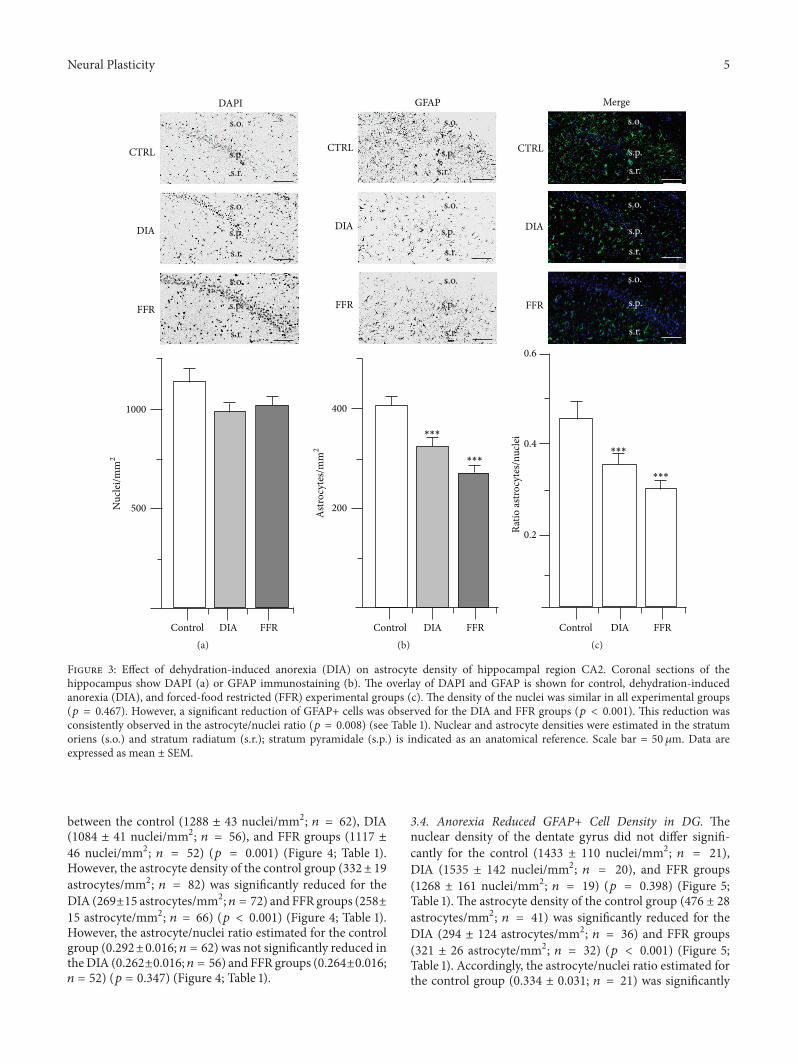
Neural Plasticity 5
DAPI
so
sp
sp
sp
sr
so
sr
so
sr
CTRL
DIA
FFR
500
1000
Nuc
leim
m 2
Control DIA FFR(a)
GFAP
so
sr
so
sr
so
sr
CTRL
DIA
FFR
200
400
Astr
ocyt
esm
m 2
lowastlowastlowast
lowastlowastlowast
Control DIA FFR
sp
sp
sp
(b)
Merge
so
sr
so
sr
so
sr
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
06
lowastlowastlowast
lowastlowastlowast
Control DIA FFR
sp
sp
sp
(c)
Figure 3 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA2 Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is shown for control dehydration-inducedanorexia (DIA) and forced-food restricted (FFR) experimental groups (c) The density of the nuclei was similar in all experimental groups(119901 = 0467) However a significant reduction of GFAP+ cells was observed for the DIA and FFR groups (119901 lt 0001) This reduction wasconsistently observed in the astrocytenuclei ratio (119901 = 0008) (see Table 1) Nuclear and astrocyte densities were estimated in the stratumoriens (so) and stratum radiatum (sr) stratum pyramidale (sp) is indicated as an anatomical reference Scale bar = 50120583m Data areexpressed as mean plusmn SEM
between the control (1288 plusmn 43 nucleimm2 119899 = 62) DIA(1084 plusmn 41 nucleimm2 119899 = 56) and FFR groups (1117 plusmn46 nucleimm2 119899 = 52) (119901 = 0001) (Figure 4 Table 1)However the astrocyte density of the control group (332plusmn 19astrocytesmm2 119899 = 82) was significantly reduced for theDIA (269plusmn15 astrocytesmm2 119899 = 72) and FFR groups (258plusmn15 astrocytemm2 119899 = 66) (119901 lt 0001) (Figure 4 Table 1)However the astrocytenuclei ratio estimated for the controlgroup (0292plusmn0016 119899 = 62) was not significantly reduced intheDIA (0262plusmn0016 119899 = 56) and FFR groups (0264plusmn0016119899 = 52) (119901 = 0347) (Figure 4 Table 1)
34 Anorexia Reduced GFAP+ Cell Density in DG Thenuclear density of the dentate gyrus did not differ signifi-cantly for the control (1433 plusmn 110 nucleimm2 119899 = 21)DIA (1535 plusmn 142 nucleimm2 119899 = 20) and FFR groups(1268 plusmn 161 nucleimm2 119899 = 19) (119901 = 0398) (Figure 5Table 1) The astrocyte density of the control group (476 plusmn 28astrocytesmm2 119899 = 41) was significantly reduced for theDIA (294 plusmn 124 astrocytesmm2 119899 = 36) and FFR groups(321 plusmn 26 astrocytemm2 119899 = 32) (119901 lt 0001) (Figure 5Table 1) Accordingly the astrocytenuclei ratio estimated forthe control group (0334 plusmn 0031 119899 = 21) was significantly
6 Neural Plasticity
DAPI
sosr
sosr
sosr
CTRL
DIA
FFR
500
1000
Nuc
leim
m 2
Control DIA FFR
sp
sp
sp
(a)
GFAP
sosr
sosr
sosr
CTRL
DIA
FFR
200
400
Astr
ocyt
esm
m 2
lowastlowastlowastlowastlowastlowast
FFRControl DIA
sp
sp
sp
(b)
Merge
sosr
sosr
sosr
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
FFRControl DIA
sp
sp
sp
(c)
Figure 4 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA3 Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is shown for control dehydration-inducedanorexia (DIA) and forced-food restricted (FFR) experimental groups (c) Neither the density of nuclei nor the astrocytenuclei ratio differedsignificantly among groups (119901 = 0931 and119901 = 0347 resp) However a significant reduction of GFAP+ cell density was observed (119901 lt 0001)(see Table 1) Nuclear and astrocyte densities were estimated in the stratum oriens (so) and stratum radiatum (sr) stratum pyramidale (sp)is indicated as an anatomical reference Scale bar = 50120583m Data are expressed as mean plusmn SEM
reduced for the DIA (0173 plusmn 0022 119899 = 20) and FFR groups(0139 plusmn 0032 119899 = 19) (119901 lt 0001) (Figure 5 Table 1)
341 Expression of Intermediate Filaments Is Affected byAnorexia The expression of intermediate filaments of astro-cytes was tested byWestern blot in the control DIA and FFRexperimental groups (Figure 6) The expression of GFAP wassignificantly reduced by DIA (048 plusmn 002 119899 = 3 119901 = 0003)and FFR (067plusmn004 119899 = 3119901 = 0006) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0668) (Figures 6(a) and6(b)) The opposite effect was observed for the expressionof vimentin and nestin The expression of vimentin was
significantly increased by DIA (151 plusmn 008 119899 = 3 119901 = 0009)and FFR (146plusmn011 119899 = 3119901 = 0008) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0896) (Figures 6(a)and 6(b)) Finally the expression of nestin was significantlyincreased by DIA (174 plusmn 01 119899 = 3 119901 = 0004) and FFR(161 plusmn 012 119899 = 3 119901 = 0001) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0468) (Figures 6(a) and6(b))
342 Anorexia Decreases GFAP Expression after the ThirdDay Our results showed a reduced GFAP+ cell density in
Neural Plasticity 7
DAPI
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
1000
1500
500
Nuc
leim
m 2
Control DIA FFR(a)
GFAP
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
200
400
600
Astr
ocyt
esm
m 2
lowastlowastlowastlowastlowastlowast
Control DIA FFR(b)
Merge
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
lowastlowastlowast
lowastlowastlowast
Control DIA FFR(c)
Figure 5 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal dentate gyrus (DG) Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is also shown for the control DIA and forcedfood-restricted (FFR) groups (c) The density of nuclei did not differ significantly among experimental groups (119901 = 0398) However asignificant reductionwas observed forGFAP+ cell density and the astrocytenuclei ratio (119901 lt 0001 and119901 lt 0001 resp) (see Table 1) Nuclearand astrocyte densities were estimated in the hilus subgranular zone (SGZ) and granular cell layer (GCL) are indicated as an anatomicalreference Scale bar = 50 120583m Data are expressed as mean plusmn SEM
the hippocampus that correlated with a lower expression ofthe protein Nevertheless GFAP expression may be affectedearlier than five daysThus the expression ofGFAPwas testedby Western blot after one and three days for control DIA orFFR experimental groups (Figures 7(a) and 7(b)) The resultsshowed that the expression of GFAP was unaffected after thefirst day for DIA (092 plusmn 004) and FFR (094 plusmn 003) whencompared to the normalized control (119901 gt 01) HoweverGFAP expression was significantly reduced after the third dayfor DIA (072 plusmn 001) and FFR (075 plusmn 004) when comparedto the normalized control (119901 lt 0001) (Figures 7(a) and7(b))
35 Anorexia Increases Reactive Astrocytes The reactiveastrocytes were estimated in CA2 andDG because astrocyte-nuclei ratio was significantly reduced in these regions Reac-tive astrocytes were defined as GFAP+ cells with processesshorter than the diameter of the somata [16] The numberof reactive astrocytes was estimated for all the experimentalgroups and normalized with their respective controls (seeSection 25) In CA2 reactive astrocytes increased 276 plusmn 07(119899 = 4 119901 = 0047) for DIA and 292 plusmn 075 (119899 = 4 119901 = 0032)for FFR while for dentate gyrus reactive astrocytes increased225plusmn015 (119899 = 4 119901 lt 00001) for DIA and 161plusmn029 (119899 = 4119901 = 0034) for FFR (Figure 8)
8 Neural Plasticity
GFAP
Vimentin
Actin
Nestin
RFFLRTC DIA
50kDa
57kDa
43kDa
200 kDa
(a)
CTRLDIA
FFR
GFAP Vimentin Nestin
10
20
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
lowastlowast
lowastlowast
(b)
Figure 6 Effect of anorexia on astrocyte intermediate filaments expression after the fifth day (a) Representative Western blot showing theexpression of GFAP vimentin and nestin for control DIA and FFR experimental groups actin was used as an internal control (b)The bandintensity of DIA and FFR groups was normalized to their respective controls Data were shown as mean plusmn SEM Significant differences wereconsidered as lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001
GFAP
Actin
After the first day
50kDa
43kDa
CTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
CTRL DIA FFR(a)
GFAP
Actin
50kDa
43kDa
After the third dayCTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
CTRL DIA FFR(b)
Figure 7 GFAP expression is reduced after the third day (a b) Representative Western blot showing the expression of GFAP after the first(a) and third day (b) of anorexia actin was used as an internal control The band intensity of DIA and FFR groups was normalized to theirrespective controls ((a b) under the Western blots) Data were shown as plusmn SEM Significant differences were considered as lowastlowastlowast119901 lt 0001
Neural Plasticity 9
RFFAIDlrtC
1
2
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in D
G
Ctrl DIA FFR
lowastlowastlowast
lowast
1
2
3
Ctrl DIA FFR
lowast
lowast
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in C
A2
Figure 8 Anorexia increases the density of reactive astrocytes Representative images of GFAP+ cells in the dentate gyrus (DG) for control(Ctrl) dehydration-induced anorexia (DIA) and forced food restriction (FFR) groupsThe density of reactive astrocytes (cells with processesshorter than the diameter of the somata) was estimated for all the experimental groups and normalized with their respective controls for DGand CA2 Data were shown as mean plusmn SEM Significant differences were considered as lowast119901 lt 005 lowastlowastlowast119901 lt 0001
4 Discussion
Anorexia nervosa is common among female adolescents andhas one of the highest mortality rates within psychiatric dis-orders (10ndash20) even surpassing depression [19ndash21] Duringpuberty mood swings and anxiety are common due to a sud-den rise of hormones which affects the hippocampus [11 1222] Moreover in the activity-based anorexia model (ABA)structural and functional alterations have been observedin the hippocampus accompanied by anxiety-like behavior[1 11 12 23] However the effect of anorexia on astrocytedensity has been barely explored Indeed previous studies inthe rat corpus callosum reported that cell proliferation wasreduced with the ABA model while astrocyte density wasregionally reduced in the DIA model [1 13] The ABA studyalso reported a reduced hippocampal cell proliferation in thedentate gyrus [1]
Results from the present study show that anorexiareduced astrocyte density in CA2 CA3 and DG whileno significant changes were observed in CA1 Accordinglyanorexia mutant mice (anxanx) showed lower expressionof c-Fos in the dentate gyrus and CA3 region of the hip-pocampus while no significant differences were found inCA1 [24] On the other hand the decrease in GFAP+ celldensity resulted in a reduced expression of GFAP proteinAstrocytes express three types of intermediate filament pro-teins GFAP vimentin and nestin [25] Nestin and vimentinare the main intermediate filaments in immature astrocyteswhereas maturing and adult astrocytes contain vimentin and
GFAP [25] Interestingly anorexia increased vimentin andnestin expression resembling reactive astrocytes observedafter injury [26 27] Accordingly GFAP+ cells deficit may bereplaced by vimentin+nestin+ cells in anorexia In supportof this hypothesis nestin reexpression has been observedfollowing experimental hippocampal lesions [25 28 29] andalso in human pathology such as multiple sclerosis [30]Moreover reactive astrocytes were increased by anorexia andforced food restriction indicating that GFAP+ cells react tosevere caloric restriction [31] Another factor to consider isthat female animals subjected to anorexia or chronic stressstop their ovarian cycles ending their exposure to hormonesthat may affect morphology and astrocyte density [32 33]Moreover hippocampal gliogenesis but not neurogenesisis reduced in the ABA model [1] Thus astrocytes possessdifferent levels of differentiation and maturation and theycan switch between these states even in the adult brain[26] Likewise astrocytes secrete gliotransmitters and growthfactors like BDNF required for synaptic plasticity synapto-genesis and neurogenesis [34 35] Thus a reduced densityof GFAP+ cells may affect hippocampal gliotransmission ofd-serine a gliotransmitter necessary for neuronal NMDA-mediated transmission and long term potentiation [34]Growth factors such as BDNF are also required for long termsynaptic plasticity of dendrites in the adult hippocampus [35]however decreased branching of dendrites was reported inan ABA model particularly in the stratum radiatum [11 12]Accordingly our results showed that GFAP+ cell density andGFAP expression are decreased by anorexia Similar results
10 Neural Plasticity
were reported in a rat model of depression where GFAPexpression was reduced [36] Thus we conclude that GFAP+cells and intermediate filament expression are affected byanorexia Our results add new information about the struc-tural changes observed in the hippocampus ofmurinemodelsof anorexia
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
The authors thank Dr P De Gortari (Insituto Nacionalde Psiquiatrıa Mexico) for introducing them to the DIAmodel The authors are grateful to M Macedo-MendozaM E Ramos-Aguilar E N Hernandez-Rıos A Castilla LCasanova A E Espino and M Garcıa-Servın for technicalsupport The authors thank Dr D Pless for reviewing thepaper The authors thank Dr G Ramırez-Rodrıguez (Insti-tuto Nacional de Psiquiatrıa Mexico) for donating the nestinand vimentin antibodies used in this study Janina Krugerwas an internship student supported by the DAAD-RISEprogram Durairaj Ragu Varman is a postdoctoral fellowsupported by DGAPA-UNAM This work was supported bygrants from Programa de Apoyo a Proyectos de Investigacione Innovacion Tecnologica de la UNAM (PAPIIT-UNAMIN200913 and IN201915) to Ataulfo Martınez-Torres andDaniel Reyes-Haro
References
[1] N C Barbarich-Marsteller C A Fornal L F Takase et alldquoActivity-based anorexia is associated with reduced hippocam-pal cell proliferation in adolescent female ratsrdquo BehaviouralBrain Research vol 236 no 1 pp 251ndash257 2013
[2] C Aoki N Sabaliauskas T Chowdhury et al ldquoAdolescentfemale rats exhibiting activity-based anorexia express elevatedlevels of GABAAreceptor 1205724 and 120575 subunits at the plasmamembrane of hippocampal CA1 spinesrdquo Synapse vol 66 no 5pp 391ndash407 2012
[3] G D Giordano P Renzetti R C Parodi et al ldquoVolume mea-surement with magnetic resonance imaging of hippocampus-amygdala formation in patients with anorexia nervosardquo Journalof Endocrinological Investigation vol 24 no 7 pp 510ndash514 2001
[4] F Connan F Murphy S E J Connor et al ldquoHippocampalvolume and cognitive function in anorexia nervosardquo PsychiatryResearchmdashNeuroimaging vol 146 no 2 pp 117ndash125 2006
[5] J N Beadle S Paradiso M Brumm M Voss K Halmiand L M McCormick ldquoLarger hippocampus size in womenwith anorexia nervosa who exercise excessively than healthywomenrdquo Psychiatry ResearchmdashNeuroimaging vol 232 no 2 pp193ndash199 2015
[6] N T Burkert KKoschutnig F Ebner andW Freidl ldquoStructuralhippocampal alterations perceived stress and coping deficien-cies in patients with anorexia nervosardquo International Journal ofEating Disorders vol 48 no 6 pp 670ndash676 2015
[7] S B McHugh M Fillenz J P Lowry J N P Rawlins and DM Bannerman ldquoBrain tissue oxygen amperometry in behavingrats demonstrates functional dissociation of dorsal and ventral
hippocampus during spatial processing and anxietyrdquo EuropeanJournal of Neuroscience vol 33 no 2 pp 322ndash337 2011
[8] A G Watts ldquoDehydration-associated anorexia developmentand rapid reversalrdquo Physiology and Behavior vol 65 no 4-5 pp871ndash878 1998
[9] L Jaimes-Hoy P Joseph-Bravo and P de Gortari ldquoDiffer-ential response of TRHergic neurons of the hypothalamicparaventricular nucleus (PVN) in female animals submittedto food-restriction or dehydration-induced anorexia and coldexposurerdquo Hormones and Behavior vol 53 no 2 pp 366ndash3772008
[10] A G Watts and C N Boyle ldquoThe functional architecture ofdehydration-anorexiardquo Physiology and Behavior vol 100 no 5pp 472ndash477 2010
[11] T G Chowdhury M B Rıos T E Chan D S Cassataro NC Barbarich-Marsteller and C Aoki ldquoActivity-based anorexiaduring adolescence disrupts normal development of the CA1pyramidal cells in the ventral hippocampus of female ratsrdquoHippocampus vol 24 no 12 pp 1421ndash1429 2014
[12] T G Chowdhury N C Barbarich-Marsteller T E Chan andCAoki ldquoActivity-based anorexia has differential effects on apicaldendritic branching in dorsal and ventral hippocampal CA1rdquoBrain Structure and Function vol 219 no 6 pp 1935ndash1945 2014
[13] D Reyes-Haro F E Labrada-Moncada R Miledi andA Martınez-Torres ldquoDehydration-induced anorexia reducesastrocyte density in the rat corpus callosumrdquo Neural Plasticityvol 2015 Article ID 474917 8 pages 2015
[14] P de Gortari K Mancera A Cote-Velez et al ldquoInvolvement ofCRH-R2 receptor in eating behavior and in the response of theHPT axis in rats subjected to dehydration-induced anorexiardquoPsychoneuroendocrinology vol 34 no 2 pp 259ndash272 2009
[15] M R Lamprecht DM Sabatini and A E Carpenter ldquoCellPro-filer free versatile software for automated biological imageanalysisrdquo BioTechniques vol 42 no 1 pp 71ndash75 2007
[16] B Rinaldi F Guida A Furiano et al ldquoEffect of prolongedmoderate exercise on the changes of nonneuronal cells in earlymyocardial infarctionrdquo Neural Plasticity vol 2015 Article ID265967 8 pages 2015
[17] K Chiu W M Lau H T Lau K-F So and R C-C ChangldquoMicro-dissection of rat brain for RNA or protein extractionfrom specific brain regionrdquo Journal of Visualized Experimentsno 7 article e269 2007
[18] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[19] P F Sullivan ldquoMortality in anorexia nervosardquo The AmericanJournal of Psychiatry vol 152 no 7 pp 1073ndash1074 1995
[20] C L Birmingham J Su J A Hlynsky E M Goldner and MGao ldquoThe mortality rate from anorexia nervosardquo InternationalJournal of Eating Disorders vol 38 no 2 pp 143ndash146 2005
[21] C M Bulik M C T Slof-Oprsquot Landt E F van Furth and P FSullivan ldquoThe genetics of anorexia nervosardquo Annual Review ofNutrition vol 27 pp 263ndash275 2007
[22] K J McLaughlin J O Wilson J Harman et al ldquoChronic 17120573-estradiol or cholesterol prevents stress-induced hippocampalCA3 dendritic retraction in ovariectomized female rats possi-ble correspondence between CA1 spine properties and spatialacquisitionrdquo Hippocampus vol 20 no 6 pp 768ndash786 2010
[23] K P Kinzig and S L Hargrave ldquoAdolescent activity-basedanorexia increases anxiety-like behavior in adulthoodrdquo Physi-ology and Behavior vol 101 no 2 pp 269ndash276 2010
Neural Plasticity 11
[24] S A Kim Y M Choi H J Park et al ldquoDecrease of c-Fosexpression in hippocampus of anorexia (anxanx)micerdquoKoreanJournal of Biological Sciences vol 5 no 2 pp 157ndash161 2001
[25] C Eliasson C Sahlgren C-H Berthold et al ldquoIntermediatefilament protein partnership in astrocytesrdquo The Journal ofBiological Chemistry vol 274 no 34 pp 23996ndash24006 1999
[26] R Schmidt-Kastner andCHumpel ldquoNestin expression persistsin astrocytes of organotypic slice cultures from rat cortexrdquoInternational Journal of Developmental Neuroscience vol 20 no1 pp 29ndash38 2002
[27] Y Takamiya S Kohsaka S Toya M Otani and Y TsukadaldquoImmunohistochemical studies on the proliferation of reactiveastrocytes and the expression of cytoskeletal proteins followingbrain injury in ratsrdquo Brain Research vol 466 no 2 pp 201ndash2101988
[28] S R Clarke A K Shetty J L Bradley and D A TurnerldquoReactive astrocytes express the embryonic intermediate neuro-filament nestinrdquoNeuroReport vol 5 no 15 pp 1885ndash1888 1994
[29] M C Geloso V Corvino V Cavallo et al ldquoExpression ofastrocytic nestin in the rat hippocampus during trimethyltin-induced neurodegenerationrdquo Neuroscience Letters vol 357 no2 pp 103ndash106 2004
[30] J E Holley D Gveric J Newcombe M L Cuzner and N JGutowski ldquoAstrocyte characterization in the multiple sclerosisglial scarrdquoNeuropathology andAppliedNeurobiology vol 29 no5 pp 434ndash444 2003
[31] C N Boyle S M Lorenzen D Compton and A G WattsldquoDehydration-anorexia derives from a reduction in meal sizebut not meal numberrdquo Physiology and Behavior vol 105 no 2pp 305ndash314 2012
[32] I AzcoitiaM Santos-GalindoMAArevalo and LMGarcıa-Segura ldquoRole of astroglia in the neuroplastic and neuroprotec-tive actions of estradiolrdquo European Journal of Neuroscience vol32 no 12 pp 1995ndash2002 2010
[33] L Eiland J Ramroop M N Hill J Manley and B S McEwenldquoChronic juvenile stress produces corticolimbic dendritic archi-tectural remodeling and modulates emotional behavior in maleand female ratsrdquo Psychoneuroendocrinology vol 37 no 1 pp39ndash47 2012
[34] C Henneberger T Papouin S H R Oliet and D A RusakovldquoLong-term potentiation depends on release of d-serine fromastrocytesrdquo Nature vol 463 no 7278 pp 232ndash236 2010
[35] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo The Journal ofNeuroscience vol 22 no 5 pp 1532ndash1540 2002
[36] Y Ye G Wang H Wang and X Wang ldquoBrain-derived neu-rotrophic factor (BDNF) infusion restored astrocytic plasticityin the hippocampus of a rat model of depressionrdquo NeuroscienceLetters vol 503 no 1 pp 15ndash19 2011
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

6 Neural Plasticity
DAPI
sosr
sosr
sosr
CTRL
DIA
FFR
500
1000
Nuc
leim
m 2
Control DIA FFR
sp
sp
sp
(a)
GFAP
sosr
sosr
sosr
CTRL
DIA
FFR
200
400
Astr
ocyt
esm
m 2
lowastlowastlowastlowastlowastlowast
FFRControl DIA
sp
sp
sp
(b)
Merge
sosr
sosr
sosr
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
FFRControl DIA
sp
sp
sp
(c)
Figure 4 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal region CA3 Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is shown for control dehydration-inducedanorexia (DIA) and forced-food restricted (FFR) experimental groups (c) Neither the density of nuclei nor the astrocytenuclei ratio differedsignificantly among groups (119901 = 0931 and119901 = 0347 resp) However a significant reduction of GFAP+ cell density was observed (119901 lt 0001)(see Table 1) Nuclear and astrocyte densities were estimated in the stratum oriens (so) and stratum radiatum (sr) stratum pyramidale (sp)is indicated as an anatomical reference Scale bar = 50120583m Data are expressed as mean plusmn SEM
reduced for the DIA (0173 plusmn 0022 119899 = 20) and FFR groups(0139 plusmn 0032 119899 = 19) (119901 lt 0001) (Figure 5 Table 1)
341 Expression of Intermediate Filaments Is Affected byAnorexia The expression of intermediate filaments of astro-cytes was tested byWestern blot in the control DIA and FFRexperimental groups (Figure 6) The expression of GFAP wassignificantly reduced by DIA (048 plusmn 002 119899 = 3 119901 = 0003)and FFR (067plusmn004 119899 = 3119901 = 0006) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0668) (Figures 6(a) and6(b)) The opposite effect was observed for the expressionof vimentin and nestin The expression of vimentin was
significantly increased by DIA (151 plusmn 008 119899 = 3 119901 = 0009)and FFR (146plusmn011 119899 = 3119901 = 0008) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0896) (Figures 6(a)and 6(b)) Finally the expression of nestin was significantlyincreased by DIA (174 plusmn 01 119899 = 3 119901 = 0004) and FFR(161 plusmn 012 119899 = 3 119901 = 0001) when compared to thenormalized control no significant differences were observedbetween DIA and FFR groups (119901 = 0468) (Figures 6(a) and6(b))
342 Anorexia Decreases GFAP Expression after the ThirdDay Our results showed a reduced GFAP+ cell density in
Neural Plasticity 7
DAPI
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
1000
1500
500
Nuc
leim
m 2
Control DIA FFR(a)
GFAP
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
200
400
600
Astr
ocyt
esm
m 2
lowastlowastlowastlowastlowastlowast
Control DIA FFR(b)
Merge
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
lowastlowastlowast
lowastlowastlowast
Control DIA FFR(c)
Figure 5 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal dentate gyrus (DG) Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is also shown for the control DIA and forcedfood-restricted (FFR) groups (c) The density of nuclei did not differ significantly among experimental groups (119901 = 0398) However asignificant reductionwas observed forGFAP+ cell density and the astrocytenuclei ratio (119901 lt 0001 and119901 lt 0001 resp) (see Table 1) Nuclearand astrocyte densities were estimated in the hilus subgranular zone (SGZ) and granular cell layer (GCL) are indicated as an anatomicalreference Scale bar = 50 120583m Data are expressed as mean plusmn SEM
the hippocampus that correlated with a lower expression ofthe protein Nevertheless GFAP expression may be affectedearlier than five daysThus the expression ofGFAPwas testedby Western blot after one and three days for control DIA orFFR experimental groups (Figures 7(a) and 7(b)) The resultsshowed that the expression of GFAP was unaffected after thefirst day for DIA (092 plusmn 004) and FFR (094 plusmn 003) whencompared to the normalized control (119901 gt 01) HoweverGFAP expression was significantly reduced after the third dayfor DIA (072 plusmn 001) and FFR (075 plusmn 004) when comparedto the normalized control (119901 lt 0001) (Figures 7(a) and7(b))
35 Anorexia Increases Reactive Astrocytes The reactiveastrocytes were estimated in CA2 andDG because astrocyte-nuclei ratio was significantly reduced in these regions Reac-tive astrocytes were defined as GFAP+ cells with processesshorter than the diameter of the somata [16] The numberof reactive astrocytes was estimated for all the experimentalgroups and normalized with their respective controls (seeSection 25) In CA2 reactive astrocytes increased 276 plusmn 07(119899 = 4 119901 = 0047) for DIA and 292 plusmn 075 (119899 = 4 119901 = 0032)for FFR while for dentate gyrus reactive astrocytes increased225plusmn015 (119899 = 4 119901 lt 00001) for DIA and 161plusmn029 (119899 = 4119901 = 0034) for FFR (Figure 8)
8 Neural Plasticity
GFAP
Vimentin
Actin
Nestin
RFFLRTC DIA
50kDa
57kDa
43kDa
200 kDa
(a)
CTRLDIA
FFR
GFAP Vimentin Nestin
10
20
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
lowastlowast
lowastlowast
(b)
Figure 6 Effect of anorexia on astrocyte intermediate filaments expression after the fifth day (a) Representative Western blot showing theexpression of GFAP vimentin and nestin for control DIA and FFR experimental groups actin was used as an internal control (b)The bandintensity of DIA and FFR groups was normalized to their respective controls Data were shown as mean plusmn SEM Significant differences wereconsidered as lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001
GFAP
Actin
After the first day
50kDa
43kDa
CTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
CTRL DIA FFR(a)
GFAP
Actin
50kDa
43kDa
After the third dayCTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
CTRL DIA FFR(b)
Figure 7 GFAP expression is reduced after the third day (a b) Representative Western blot showing the expression of GFAP after the first(a) and third day (b) of anorexia actin was used as an internal control The band intensity of DIA and FFR groups was normalized to theirrespective controls ((a b) under the Western blots) Data were shown as plusmn SEM Significant differences were considered as lowastlowastlowast119901 lt 0001
Neural Plasticity 9
RFFAIDlrtC
1
2
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in D
G
Ctrl DIA FFR
lowastlowastlowast
lowast
1
2
3
Ctrl DIA FFR
lowast
lowast
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in C
A2
Figure 8 Anorexia increases the density of reactive astrocytes Representative images of GFAP+ cells in the dentate gyrus (DG) for control(Ctrl) dehydration-induced anorexia (DIA) and forced food restriction (FFR) groupsThe density of reactive astrocytes (cells with processesshorter than the diameter of the somata) was estimated for all the experimental groups and normalized with their respective controls for DGand CA2 Data were shown as mean plusmn SEM Significant differences were considered as lowast119901 lt 005 lowastlowastlowast119901 lt 0001
4 Discussion
Anorexia nervosa is common among female adolescents andhas one of the highest mortality rates within psychiatric dis-orders (10ndash20) even surpassing depression [19ndash21] Duringpuberty mood swings and anxiety are common due to a sud-den rise of hormones which affects the hippocampus [11 1222] Moreover in the activity-based anorexia model (ABA)structural and functional alterations have been observedin the hippocampus accompanied by anxiety-like behavior[1 11 12 23] However the effect of anorexia on astrocytedensity has been barely explored Indeed previous studies inthe rat corpus callosum reported that cell proliferation wasreduced with the ABA model while astrocyte density wasregionally reduced in the DIA model [1 13] The ABA studyalso reported a reduced hippocampal cell proliferation in thedentate gyrus [1]
Results from the present study show that anorexiareduced astrocyte density in CA2 CA3 and DG whileno significant changes were observed in CA1 Accordinglyanorexia mutant mice (anxanx) showed lower expressionof c-Fos in the dentate gyrus and CA3 region of the hip-pocampus while no significant differences were found inCA1 [24] On the other hand the decrease in GFAP+ celldensity resulted in a reduced expression of GFAP proteinAstrocytes express three types of intermediate filament pro-teins GFAP vimentin and nestin [25] Nestin and vimentinare the main intermediate filaments in immature astrocyteswhereas maturing and adult astrocytes contain vimentin and
GFAP [25] Interestingly anorexia increased vimentin andnestin expression resembling reactive astrocytes observedafter injury [26 27] Accordingly GFAP+ cells deficit may bereplaced by vimentin+nestin+ cells in anorexia In supportof this hypothesis nestin reexpression has been observedfollowing experimental hippocampal lesions [25 28 29] andalso in human pathology such as multiple sclerosis [30]Moreover reactive astrocytes were increased by anorexia andforced food restriction indicating that GFAP+ cells react tosevere caloric restriction [31] Another factor to consider isthat female animals subjected to anorexia or chronic stressstop their ovarian cycles ending their exposure to hormonesthat may affect morphology and astrocyte density [32 33]Moreover hippocampal gliogenesis but not neurogenesisis reduced in the ABA model [1] Thus astrocytes possessdifferent levels of differentiation and maturation and theycan switch between these states even in the adult brain[26] Likewise astrocytes secrete gliotransmitters and growthfactors like BDNF required for synaptic plasticity synapto-genesis and neurogenesis [34 35] Thus a reduced densityof GFAP+ cells may affect hippocampal gliotransmission ofd-serine a gliotransmitter necessary for neuronal NMDA-mediated transmission and long term potentiation [34]Growth factors such as BDNF are also required for long termsynaptic plasticity of dendrites in the adult hippocampus [35]however decreased branching of dendrites was reported inan ABA model particularly in the stratum radiatum [11 12]Accordingly our results showed that GFAP+ cell density andGFAP expression are decreased by anorexia Similar results
10 Neural Plasticity
were reported in a rat model of depression where GFAPexpression was reduced [36] Thus we conclude that GFAP+cells and intermediate filament expression are affected byanorexia Our results add new information about the struc-tural changes observed in the hippocampus ofmurinemodelsof anorexia
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
The authors thank Dr P De Gortari (Insituto Nacionalde Psiquiatrıa Mexico) for introducing them to the DIAmodel The authors are grateful to M Macedo-MendozaM E Ramos-Aguilar E N Hernandez-Rıos A Castilla LCasanova A E Espino and M Garcıa-Servın for technicalsupport The authors thank Dr D Pless for reviewing thepaper The authors thank Dr G Ramırez-Rodrıguez (Insti-tuto Nacional de Psiquiatrıa Mexico) for donating the nestinand vimentin antibodies used in this study Janina Krugerwas an internship student supported by the DAAD-RISEprogram Durairaj Ragu Varman is a postdoctoral fellowsupported by DGAPA-UNAM This work was supported bygrants from Programa de Apoyo a Proyectos de Investigacione Innovacion Tecnologica de la UNAM (PAPIIT-UNAMIN200913 and IN201915) to Ataulfo Martınez-Torres andDaniel Reyes-Haro
References
[1] N C Barbarich-Marsteller C A Fornal L F Takase et alldquoActivity-based anorexia is associated with reduced hippocam-pal cell proliferation in adolescent female ratsrdquo BehaviouralBrain Research vol 236 no 1 pp 251ndash257 2013
[2] C Aoki N Sabaliauskas T Chowdhury et al ldquoAdolescentfemale rats exhibiting activity-based anorexia express elevatedlevels of GABAAreceptor 1205724 and 120575 subunits at the plasmamembrane of hippocampal CA1 spinesrdquo Synapse vol 66 no 5pp 391ndash407 2012
[3] G D Giordano P Renzetti R C Parodi et al ldquoVolume mea-surement with magnetic resonance imaging of hippocampus-amygdala formation in patients with anorexia nervosardquo Journalof Endocrinological Investigation vol 24 no 7 pp 510ndash514 2001
[4] F Connan F Murphy S E J Connor et al ldquoHippocampalvolume and cognitive function in anorexia nervosardquo PsychiatryResearchmdashNeuroimaging vol 146 no 2 pp 117ndash125 2006
[5] J N Beadle S Paradiso M Brumm M Voss K Halmiand L M McCormick ldquoLarger hippocampus size in womenwith anorexia nervosa who exercise excessively than healthywomenrdquo Psychiatry ResearchmdashNeuroimaging vol 232 no 2 pp193ndash199 2015
[6] N T Burkert KKoschutnig F Ebner andW Freidl ldquoStructuralhippocampal alterations perceived stress and coping deficien-cies in patients with anorexia nervosardquo International Journal ofEating Disorders vol 48 no 6 pp 670ndash676 2015
[7] S B McHugh M Fillenz J P Lowry J N P Rawlins and DM Bannerman ldquoBrain tissue oxygen amperometry in behavingrats demonstrates functional dissociation of dorsal and ventral
hippocampus during spatial processing and anxietyrdquo EuropeanJournal of Neuroscience vol 33 no 2 pp 322ndash337 2011
[8] A G Watts ldquoDehydration-associated anorexia developmentand rapid reversalrdquo Physiology and Behavior vol 65 no 4-5 pp871ndash878 1998
[9] L Jaimes-Hoy P Joseph-Bravo and P de Gortari ldquoDiffer-ential response of TRHergic neurons of the hypothalamicparaventricular nucleus (PVN) in female animals submittedto food-restriction or dehydration-induced anorexia and coldexposurerdquo Hormones and Behavior vol 53 no 2 pp 366ndash3772008
[10] A G Watts and C N Boyle ldquoThe functional architecture ofdehydration-anorexiardquo Physiology and Behavior vol 100 no 5pp 472ndash477 2010
[11] T G Chowdhury M B Rıos T E Chan D S Cassataro NC Barbarich-Marsteller and C Aoki ldquoActivity-based anorexiaduring adolescence disrupts normal development of the CA1pyramidal cells in the ventral hippocampus of female ratsrdquoHippocampus vol 24 no 12 pp 1421ndash1429 2014
[12] T G Chowdhury N C Barbarich-Marsteller T E Chan andCAoki ldquoActivity-based anorexia has differential effects on apicaldendritic branching in dorsal and ventral hippocampal CA1rdquoBrain Structure and Function vol 219 no 6 pp 1935ndash1945 2014
[13] D Reyes-Haro F E Labrada-Moncada R Miledi andA Martınez-Torres ldquoDehydration-induced anorexia reducesastrocyte density in the rat corpus callosumrdquo Neural Plasticityvol 2015 Article ID 474917 8 pages 2015
[14] P de Gortari K Mancera A Cote-Velez et al ldquoInvolvement ofCRH-R2 receptor in eating behavior and in the response of theHPT axis in rats subjected to dehydration-induced anorexiardquoPsychoneuroendocrinology vol 34 no 2 pp 259ndash272 2009
[15] M R Lamprecht DM Sabatini and A E Carpenter ldquoCellPro-filer free versatile software for automated biological imageanalysisrdquo BioTechniques vol 42 no 1 pp 71ndash75 2007
[16] B Rinaldi F Guida A Furiano et al ldquoEffect of prolongedmoderate exercise on the changes of nonneuronal cells in earlymyocardial infarctionrdquo Neural Plasticity vol 2015 Article ID265967 8 pages 2015
[17] K Chiu W M Lau H T Lau K-F So and R C-C ChangldquoMicro-dissection of rat brain for RNA or protein extractionfrom specific brain regionrdquo Journal of Visualized Experimentsno 7 article e269 2007
[18] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[19] P F Sullivan ldquoMortality in anorexia nervosardquo The AmericanJournal of Psychiatry vol 152 no 7 pp 1073ndash1074 1995
[20] C L Birmingham J Su J A Hlynsky E M Goldner and MGao ldquoThe mortality rate from anorexia nervosardquo InternationalJournal of Eating Disorders vol 38 no 2 pp 143ndash146 2005
[21] C M Bulik M C T Slof-Oprsquot Landt E F van Furth and P FSullivan ldquoThe genetics of anorexia nervosardquo Annual Review ofNutrition vol 27 pp 263ndash275 2007
[22] K J McLaughlin J O Wilson J Harman et al ldquoChronic 17120573-estradiol or cholesterol prevents stress-induced hippocampalCA3 dendritic retraction in ovariectomized female rats possi-ble correspondence between CA1 spine properties and spatialacquisitionrdquo Hippocampus vol 20 no 6 pp 768ndash786 2010
[23] K P Kinzig and S L Hargrave ldquoAdolescent activity-basedanorexia increases anxiety-like behavior in adulthoodrdquo Physi-ology and Behavior vol 101 no 2 pp 269ndash276 2010
Neural Plasticity 11
[24] S A Kim Y M Choi H J Park et al ldquoDecrease of c-Fosexpression in hippocampus of anorexia (anxanx)micerdquoKoreanJournal of Biological Sciences vol 5 no 2 pp 157ndash161 2001
[25] C Eliasson C Sahlgren C-H Berthold et al ldquoIntermediatefilament protein partnership in astrocytesrdquo The Journal ofBiological Chemistry vol 274 no 34 pp 23996ndash24006 1999
[26] R Schmidt-Kastner andCHumpel ldquoNestin expression persistsin astrocytes of organotypic slice cultures from rat cortexrdquoInternational Journal of Developmental Neuroscience vol 20 no1 pp 29ndash38 2002
[27] Y Takamiya S Kohsaka S Toya M Otani and Y TsukadaldquoImmunohistochemical studies on the proliferation of reactiveastrocytes and the expression of cytoskeletal proteins followingbrain injury in ratsrdquo Brain Research vol 466 no 2 pp 201ndash2101988
[28] S R Clarke A K Shetty J L Bradley and D A TurnerldquoReactive astrocytes express the embryonic intermediate neuro-filament nestinrdquoNeuroReport vol 5 no 15 pp 1885ndash1888 1994
[29] M C Geloso V Corvino V Cavallo et al ldquoExpression ofastrocytic nestin in the rat hippocampus during trimethyltin-induced neurodegenerationrdquo Neuroscience Letters vol 357 no2 pp 103ndash106 2004
[30] J E Holley D Gveric J Newcombe M L Cuzner and N JGutowski ldquoAstrocyte characterization in the multiple sclerosisglial scarrdquoNeuropathology andAppliedNeurobiology vol 29 no5 pp 434ndash444 2003
[31] C N Boyle S M Lorenzen D Compton and A G WattsldquoDehydration-anorexia derives from a reduction in meal sizebut not meal numberrdquo Physiology and Behavior vol 105 no 2pp 305ndash314 2012
[32] I AzcoitiaM Santos-GalindoMAArevalo and LMGarcıa-Segura ldquoRole of astroglia in the neuroplastic and neuroprotec-tive actions of estradiolrdquo European Journal of Neuroscience vol32 no 12 pp 1995ndash2002 2010
[33] L Eiland J Ramroop M N Hill J Manley and B S McEwenldquoChronic juvenile stress produces corticolimbic dendritic archi-tectural remodeling and modulates emotional behavior in maleand female ratsrdquo Psychoneuroendocrinology vol 37 no 1 pp39ndash47 2012
[34] C Henneberger T Papouin S H R Oliet and D A RusakovldquoLong-term potentiation depends on release of d-serine fromastrocytesrdquo Nature vol 463 no 7278 pp 232ndash236 2010
[35] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo The Journal ofNeuroscience vol 22 no 5 pp 1532ndash1540 2002
[36] Y Ye G Wang H Wang and X Wang ldquoBrain-derived neu-rotrophic factor (BDNF) infusion restored astrocytic plasticityin the hippocampus of a rat model of depressionrdquo NeuroscienceLetters vol 503 no 1 pp 15ndash19 2011
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
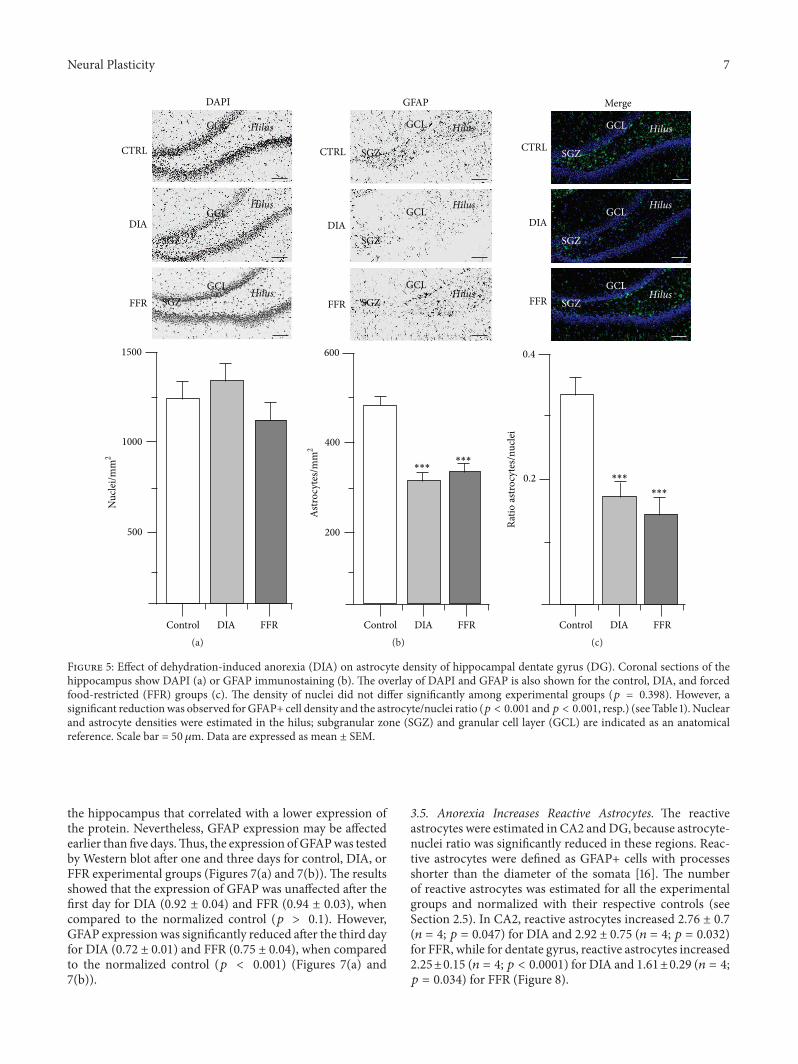
Neural Plasticity 7
DAPI
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
1000
1500
500
Nuc
leim
m 2
Control DIA FFR(a)
GFAP
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
200
400
600
Astr
ocyt
esm
m 2
lowastlowastlowastlowastlowastlowast
Control DIA FFR(b)
Merge
SGZ
SGZ
SGZ
GCL
GCL
GCL
Hilus
Hilus
Hilus
CTRL
DIA
FFR
Ratio
astro
cyte
snu
clei
02
04
lowastlowastlowast
lowastlowastlowast
Control DIA FFR(c)
Figure 5 Effect of dehydration-induced anorexia (DIA) on astrocyte density of hippocampal dentate gyrus (DG) Coronal sections of thehippocampus show DAPI (a) or GFAP immunostaining (b) The overlay of DAPI and GFAP is also shown for the control DIA and forcedfood-restricted (FFR) groups (c) The density of nuclei did not differ significantly among experimental groups (119901 = 0398) However asignificant reductionwas observed forGFAP+ cell density and the astrocytenuclei ratio (119901 lt 0001 and119901 lt 0001 resp) (see Table 1) Nuclearand astrocyte densities were estimated in the hilus subgranular zone (SGZ) and granular cell layer (GCL) are indicated as an anatomicalreference Scale bar = 50 120583m Data are expressed as mean plusmn SEM
the hippocampus that correlated with a lower expression ofthe protein Nevertheless GFAP expression may be affectedearlier than five daysThus the expression ofGFAPwas testedby Western blot after one and three days for control DIA orFFR experimental groups (Figures 7(a) and 7(b)) The resultsshowed that the expression of GFAP was unaffected after thefirst day for DIA (092 plusmn 004) and FFR (094 plusmn 003) whencompared to the normalized control (119901 gt 01) HoweverGFAP expression was significantly reduced after the third dayfor DIA (072 plusmn 001) and FFR (075 plusmn 004) when comparedto the normalized control (119901 lt 0001) (Figures 7(a) and7(b))
35 Anorexia Increases Reactive Astrocytes The reactiveastrocytes were estimated in CA2 andDG because astrocyte-nuclei ratio was significantly reduced in these regions Reac-tive astrocytes were defined as GFAP+ cells with processesshorter than the diameter of the somata [16] The numberof reactive astrocytes was estimated for all the experimentalgroups and normalized with their respective controls (seeSection 25) In CA2 reactive astrocytes increased 276 plusmn 07(119899 = 4 119901 = 0047) for DIA and 292 plusmn 075 (119899 = 4 119901 = 0032)for FFR while for dentate gyrus reactive astrocytes increased225plusmn015 (119899 = 4 119901 lt 00001) for DIA and 161plusmn029 (119899 = 4119901 = 0034) for FFR (Figure 8)
8 Neural Plasticity
GFAP
Vimentin
Actin
Nestin
RFFLRTC DIA
50kDa
57kDa
43kDa
200 kDa
(a)
CTRLDIA
FFR
GFAP Vimentin Nestin
10
20
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
lowastlowast
lowastlowast
(b)
Figure 6 Effect of anorexia on astrocyte intermediate filaments expression after the fifth day (a) Representative Western blot showing theexpression of GFAP vimentin and nestin for control DIA and FFR experimental groups actin was used as an internal control (b)The bandintensity of DIA and FFR groups was normalized to their respective controls Data were shown as mean plusmn SEM Significant differences wereconsidered as lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001
GFAP
Actin
After the first day
50kDa
43kDa
CTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
CTRL DIA FFR(a)
GFAP
Actin
50kDa
43kDa
After the third dayCTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
CTRL DIA FFR(b)
Figure 7 GFAP expression is reduced after the third day (a b) Representative Western blot showing the expression of GFAP after the first(a) and third day (b) of anorexia actin was used as an internal control The band intensity of DIA and FFR groups was normalized to theirrespective controls ((a b) under the Western blots) Data were shown as plusmn SEM Significant differences were considered as lowastlowastlowast119901 lt 0001
Neural Plasticity 9
RFFAIDlrtC
1
2
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in D
G
Ctrl DIA FFR
lowastlowastlowast
lowast
1
2
3
Ctrl DIA FFR
lowast
lowast
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in C
A2
Figure 8 Anorexia increases the density of reactive astrocytes Representative images of GFAP+ cells in the dentate gyrus (DG) for control(Ctrl) dehydration-induced anorexia (DIA) and forced food restriction (FFR) groupsThe density of reactive astrocytes (cells with processesshorter than the diameter of the somata) was estimated for all the experimental groups and normalized with their respective controls for DGand CA2 Data were shown as mean plusmn SEM Significant differences were considered as lowast119901 lt 005 lowastlowastlowast119901 lt 0001
4 Discussion
Anorexia nervosa is common among female adolescents andhas one of the highest mortality rates within psychiatric dis-orders (10ndash20) even surpassing depression [19ndash21] Duringpuberty mood swings and anxiety are common due to a sud-den rise of hormones which affects the hippocampus [11 1222] Moreover in the activity-based anorexia model (ABA)structural and functional alterations have been observedin the hippocampus accompanied by anxiety-like behavior[1 11 12 23] However the effect of anorexia on astrocytedensity has been barely explored Indeed previous studies inthe rat corpus callosum reported that cell proliferation wasreduced with the ABA model while astrocyte density wasregionally reduced in the DIA model [1 13] The ABA studyalso reported a reduced hippocampal cell proliferation in thedentate gyrus [1]
Results from the present study show that anorexiareduced astrocyte density in CA2 CA3 and DG whileno significant changes were observed in CA1 Accordinglyanorexia mutant mice (anxanx) showed lower expressionof c-Fos in the dentate gyrus and CA3 region of the hip-pocampus while no significant differences were found inCA1 [24] On the other hand the decrease in GFAP+ celldensity resulted in a reduced expression of GFAP proteinAstrocytes express three types of intermediate filament pro-teins GFAP vimentin and nestin [25] Nestin and vimentinare the main intermediate filaments in immature astrocyteswhereas maturing and adult astrocytes contain vimentin and
GFAP [25] Interestingly anorexia increased vimentin andnestin expression resembling reactive astrocytes observedafter injury [26 27] Accordingly GFAP+ cells deficit may bereplaced by vimentin+nestin+ cells in anorexia In supportof this hypothesis nestin reexpression has been observedfollowing experimental hippocampal lesions [25 28 29] andalso in human pathology such as multiple sclerosis [30]Moreover reactive astrocytes were increased by anorexia andforced food restriction indicating that GFAP+ cells react tosevere caloric restriction [31] Another factor to consider isthat female animals subjected to anorexia or chronic stressstop their ovarian cycles ending their exposure to hormonesthat may affect morphology and astrocyte density [32 33]Moreover hippocampal gliogenesis but not neurogenesisis reduced in the ABA model [1] Thus astrocytes possessdifferent levels of differentiation and maturation and theycan switch between these states even in the adult brain[26] Likewise astrocytes secrete gliotransmitters and growthfactors like BDNF required for synaptic plasticity synapto-genesis and neurogenesis [34 35] Thus a reduced densityof GFAP+ cells may affect hippocampal gliotransmission ofd-serine a gliotransmitter necessary for neuronal NMDA-mediated transmission and long term potentiation [34]Growth factors such as BDNF are also required for long termsynaptic plasticity of dendrites in the adult hippocampus [35]however decreased branching of dendrites was reported inan ABA model particularly in the stratum radiatum [11 12]Accordingly our results showed that GFAP+ cell density andGFAP expression are decreased by anorexia Similar results
10 Neural Plasticity
were reported in a rat model of depression where GFAPexpression was reduced [36] Thus we conclude that GFAP+cells and intermediate filament expression are affected byanorexia Our results add new information about the struc-tural changes observed in the hippocampus ofmurinemodelsof anorexia
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
The authors thank Dr P De Gortari (Insituto Nacionalde Psiquiatrıa Mexico) for introducing them to the DIAmodel The authors are grateful to M Macedo-MendozaM E Ramos-Aguilar E N Hernandez-Rıos A Castilla LCasanova A E Espino and M Garcıa-Servın for technicalsupport The authors thank Dr D Pless for reviewing thepaper The authors thank Dr G Ramırez-Rodrıguez (Insti-tuto Nacional de Psiquiatrıa Mexico) for donating the nestinand vimentin antibodies used in this study Janina Krugerwas an internship student supported by the DAAD-RISEprogram Durairaj Ragu Varman is a postdoctoral fellowsupported by DGAPA-UNAM This work was supported bygrants from Programa de Apoyo a Proyectos de Investigacione Innovacion Tecnologica de la UNAM (PAPIIT-UNAMIN200913 and IN201915) to Ataulfo Martınez-Torres andDaniel Reyes-Haro
References
[1] N C Barbarich-Marsteller C A Fornal L F Takase et alldquoActivity-based anorexia is associated with reduced hippocam-pal cell proliferation in adolescent female ratsrdquo BehaviouralBrain Research vol 236 no 1 pp 251ndash257 2013
[2] C Aoki N Sabaliauskas T Chowdhury et al ldquoAdolescentfemale rats exhibiting activity-based anorexia express elevatedlevels of GABAAreceptor 1205724 and 120575 subunits at the plasmamembrane of hippocampal CA1 spinesrdquo Synapse vol 66 no 5pp 391ndash407 2012
[3] G D Giordano P Renzetti R C Parodi et al ldquoVolume mea-surement with magnetic resonance imaging of hippocampus-amygdala formation in patients with anorexia nervosardquo Journalof Endocrinological Investigation vol 24 no 7 pp 510ndash514 2001
[4] F Connan F Murphy S E J Connor et al ldquoHippocampalvolume and cognitive function in anorexia nervosardquo PsychiatryResearchmdashNeuroimaging vol 146 no 2 pp 117ndash125 2006
[5] J N Beadle S Paradiso M Brumm M Voss K Halmiand L M McCormick ldquoLarger hippocampus size in womenwith anorexia nervosa who exercise excessively than healthywomenrdquo Psychiatry ResearchmdashNeuroimaging vol 232 no 2 pp193ndash199 2015
[6] N T Burkert KKoschutnig F Ebner andW Freidl ldquoStructuralhippocampal alterations perceived stress and coping deficien-cies in patients with anorexia nervosardquo International Journal ofEating Disorders vol 48 no 6 pp 670ndash676 2015
[7] S B McHugh M Fillenz J P Lowry J N P Rawlins and DM Bannerman ldquoBrain tissue oxygen amperometry in behavingrats demonstrates functional dissociation of dorsal and ventral
hippocampus during spatial processing and anxietyrdquo EuropeanJournal of Neuroscience vol 33 no 2 pp 322ndash337 2011
[8] A G Watts ldquoDehydration-associated anorexia developmentand rapid reversalrdquo Physiology and Behavior vol 65 no 4-5 pp871ndash878 1998
[9] L Jaimes-Hoy P Joseph-Bravo and P de Gortari ldquoDiffer-ential response of TRHergic neurons of the hypothalamicparaventricular nucleus (PVN) in female animals submittedto food-restriction or dehydration-induced anorexia and coldexposurerdquo Hormones and Behavior vol 53 no 2 pp 366ndash3772008
[10] A G Watts and C N Boyle ldquoThe functional architecture ofdehydration-anorexiardquo Physiology and Behavior vol 100 no 5pp 472ndash477 2010
[11] T G Chowdhury M B Rıos T E Chan D S Cassataro NC Barbarich-Marsteller and C Aoki ldquoActivity-based anorexiaduring adolescence disrupts normal development of the CA1pyramidal cells in the ventral hippocampus of female ratsrdquoHippocampus vol 24 no 12 pp 1421ndash1429 2014
[12] T G Chowdhury N C Barbarich-Marsteller T E Chan andCAoki ldquoActivity-based anorexia has differential effects on apicaldendritic branching in dorsal and ventral hippocampal CA1rdquoBrain Structure and Function vol 219 no 6 pp 1935ndash1945 2014
[13] D Reyes-Haro F E Labrada-Moncada R Miledi andA Martınez-Torres ldquoDehydration-induced anorexia reducesastrocyte density in the rat corpus callosumrdquo Neural Plasticityvol 2015 Article ID 474917 8 pages 2015
[14] P de Gortari K Mancera A Cote-Velez et al ldquoInvolvement ofCRH-R2 receptor in eating behavior and in the response of theHPT axis in rats subjected to dehydration-induced anorexiardquoPsychoneuroendocrinology vol 34 no 2 pp 259ndash272 2009
[15] M R Lamprecht DM Sabatini and A E Carpenter ldquoCellPro-filer free versatile software for automated biological imageanalysisrdquo BioTechniques vol 42 no 1 pp 71ndash75 2007
[16] B Rinaldi F Guida A Furiano et al ldquoEffect of prolongedmoderate exercise on the changes of nonneuronal cells in earlymyocardial infarctionrdquo Neural Plasticity vol 2015 Article ID265967 8 pages 2015
[17] K Chiu W M Lau H T Lau K-F So and R C-C ChangldquoMicro-dissection of rat brain for RNA or protein extractionfrom specific brain regionrdquo Journal of Visualized Experimentsno 7 article e269 2007
[18] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[19] P F Sullivan ldquoMortality in anorexia nervosardquo The AmericanJournal of Psychiatry vol 152 no 7 pp 1073ndash1074 1995
[20] C L Birmingham J Su J A Hlynsky E M Goldner and MGao ldquoThe mortality rate from anorexia nervosardquo InternationalJournal of Eating Disorders vol 38 no 2 pp 143ndash146 2005
[21] C M Bulik M C T Slof-Oprsquot Landt E F van Furth and P FSullivan ldquoThe genetics of anorexia nervosardquo Annual Review ofNutrition vol 27 pp 263ndash275 2007
[22] K J McLaughlin J O Wilson J Harman et al ldquoChronic 17120573-estradiol or cholesterol prevents stress-induced hippocampalCA3 dendritic retraction in ovariectomized female rats possi-ble correspondence between CA1 spine properties and spatialacquisitionrdquo Hippocampus vol 20 no 6 pp 768ndash786 2010
[23] K P Kinzig and S L Hargrave ldquoAdolescent activity-basedanorexia increases anxiety-like behavior in adulthoodrdquo Physi-ology and Behavior vol 101 no 2 pp 269ndash276 2010
Neural Plasticity 11
[24] S A Kim Y M Choi H J Park et al ldquoDecrease of c-Fosexpression in hippocampus of anorexia (anxanx)micerdquoKoreanJournal of Biological Sciences vol 5 no 2 pp 157ndash161 2001
[25] C Eliasson C Sahlgren C-H Berthold et al ldquoIntermediatefilament protein partnership in astrocytesrdquo The Journal ofBiological Chemistry vol 274 no 34 pp 23996ndash24006 1999
[26] R Schmidt-Kastner andCHumpel ldquoNestin expression persistsin astrocytes of organotypic slice cultures from rat cortexrdquoInternational Journal of Developmental Neuroscience vol 20 no1 pp 29ndash38 2002
[27] Y Takamiya S Kohsaka S Toya M Otani and Y TsukadaldquoImmunohistochemical studies on the proliferation of reactiveastrocytes and the expression of cytoskeletal proteins followingbrain injury in ratsrdquo Brain Research vol 466 no 2 pp 201ndash2101988
[28] S R Clarke A K Shetty J L Bradley and D A TurnerldquoReactive astrocytes express the embryonic intermediate neuro-filament nestinrdquoNeuroReport vol 5 no 15 pp 1885ndash1888 1994
[29] M C Geloso V Corvino V Cavallo et al ldquoExpression ofastrocytic nestin in the rat hippocampus during trimethyltin-induced neurodegenerationrdquo Neuroscience Letters vol 357 no2 pp 103ndash106 2004
[30] J E Holley D Gveric J Newcombe M L Cuzner and N JGutowski ldquoAstrocyte characterization in the multiple sclerosisglial scarrdquoNeuropathology andAppliedNeurobiology vol 29 no5 pp 434ndash444 2003
[31] C N Boyle S M Lorenzen D Compton and A G WattsldquoDehydration-anorexia derives from a reduction in meal sizebut not meal numberrdquo Physiology and Behavior vol 105 no 2pp 305ndash314 2012
[32] I AzcoitiaM Santos-GalindoMAArevalo and LMGarcıa-Segura ldquoRole of astroglia in the neuroplastic and neuroprotec-tive actions of estradiolrdquo European Journal of Neuroscience vol32 no 12 pp 1995ndash2002 2010
[33] L Eiland J Ramroop M N Hill J Manley and B S McEwenldquoChronic juvenile stress produces corticolimbic dendritic archi-tectural remodeling and modulates emotional behavior in maleand female ratsrdquo Psychoneuroendocrinology vol 37 no 1 pp39ndash47 2012
[34] C Henneberger T Papouin S H R Oliet and D A RusakovldquoLong-term potentiation depends on release of d-serine fromastrocytesrdquo Nature vol 463 no 7278 pp 232ndash236 2010
[35] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo The Journal ofNeuroscience vol 22 no 5 pp 1532ndash1540 2002
[36] Y Ye G Wang H Wang and X Wang ldquoBrain-derived neu-rotrophic factor (BDNF) infusion restored astrocytic plasticityin the hippocampus of a rat model of depressionrdquo NeuroscienceLetters vol 503 no 1 pp 15ndash19 2011
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
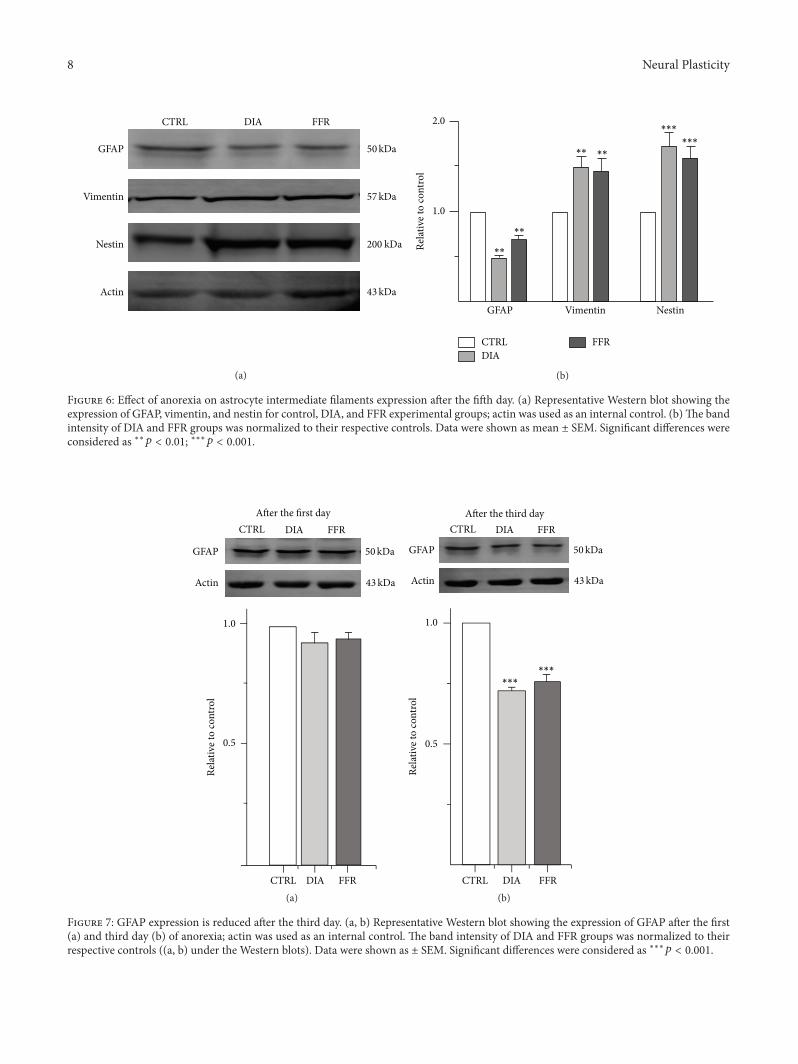
8 Neural Plasticity
GFAP
Vimentin
Actin
Nestin
RFFLRTC DIA
50kDa
57kDa
43kDa
200 kDa
(a)
CTRLDIA
FFR
GFAP Vimentin Nestin
10
20
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
lowastlowastlowastlowast
lowastlowast
lowastlowast
(b)
Figure 6 Effect of anorexia on astrocyte intermediate filaments expression after the fifth day (a) Representative Western blot showing theexpression of GFAP vimentin and nestin for control DIA and FFR experimental groups actin was used as an internal control (b)The bandintensity of DIA and FFR groups was normalized to their respective controls Data were shown as mean plusmn SEM Significant differences wereconsidered as lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001
GFAP
Actin
After the first day
50kDa
43kDa
CTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
CTRL DIA FFR(a)
GFAP
Actin
50kDa
43kDa
After the third dayCTRL DIA FFR
10
05
Relat
ive t
o co
ntro
l
lowastlowastlowast
lowastlowastlowast
CTRL DIA FFR(b)
Figure 7 GFAP expression is reduced after the third day (a b) Representative Western blot showing the expression of GFAP after the first(a) and third day (b) of anorexia actin was used as an internal control The band intensity of DIA and FFR groups was normalized to theirrespective controls ((a b) under the Western blots) Data were shown as plusmn SEM Significant differences were considered as lowastlowastlowast119901 lt 0001
Neural Plasticity 9
RFFAIDlrtC
1
2
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in D
G
Ctrl DIA FFR
lowastlowastlowast
lowast
1
2
3
Ctrl DIA FFR
lowast
lowast
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in C
A2
Figure 8 Anorexia increases the density of reactive astrocytes Representative images of GFAP+ cells in the dentate gyrus (DG) for control(Ctrl) dehydration-induced anorexia (DIA) and forced food restriction (FFR) groupsThe density of reactive astrocytes (cells with processesshorter than the diameter of the somata) was estimated for all the experimental groups and normalized with their respective controls for DGand CA2 Data were shown as mean plusmn SEM Significant differences were considered as lowast119901 lt 005 lowastlowastlowast119901 lt 0001
4 Discussion
Anorexia nervosa is common among female adolescents andhas one of the highest mortality rates within psychiatric dis-orders (10ndash20) even surpassing depression [19ndash21] Duringpuberty mood swings and anxiety are common due to a sud-den rise of hormones which affects the hippocampus [11 1222] Moreover in the activity-based anorexia model (ABA)structural and functional alterations have been observedin the hippocampus accompanied by anxiety-like behavior[1 11 12 23] However the effect of anorexia on astrocytedensity has been barely explored Indeed previous studies inthe rat corpus callosum reported that cell proliferation wasreduced with the ABA model while astrocyte density wasregionally reduced in the DIA model [1 13] The ABA studyalso reported a reduced hippocampal cell proliferation in thedentate gyrus [1]
Results from the present study show that anorexiareduced astrocyte density in CA2 CA3 and DG whileno significant changes were observed in CA1 Accordinglyanorexia mutant mice (anxanx) showed lower expressionof c-Fos in the dentate gyrus and CA3 region of the hip-pocampus while no significant differences were found inCA1 [24] On the other hand the decrease in GFAP+ celldensity resulted in a reduced expression of GFAP proteinAstrocytes express three types of intermediate filament pro-teins GFAP vimentin and nestin [25] Nestin and vimentinare the main intermediate filaments in immature astrocyteswhereas maturing and adult astrocytes contain vimentin and
GFAP [25] Interestingly anorexia increased vimentin andnestin expression resembling reactive astrocytes observedafter injury [26 27] Accordingly GFAP+ cells deficit may bereplaced by vimentin+nestin+ cells in anorexia In supportof this hypothesis nestin reexpression has been observedfollowing experimental hippocampal lesions [25 28 29] andalso in human pathology such as multiple sclerosis [30]Moreover reactive astrocytes were increased by anorexia andforced food restriction indicating that GFAP+ cells react tosevere caloric restriction [31] Another factor to consider isthat female animals subjected to anorexia or chronic stressstop their ovarian cycles ending their exposure to hormonesthat may affect morphology and astrocyte density [32 33]Moreover hippocampal gliogenesis but not neurogenesisis reduced in the ABA model [1] Thus astrocytes possessdifferent levels of differentiation and maturation and theycan switch between these states even in the adult brain[26] Likewise astrocytes secrete gliotransmitters and growthfactors like BDNF required for synaptic plasticity synapto-genesis and neurogenesis [34 35] Thus a reduced densityof GFAP+ cells may affect hippocampal gliotransmission ofd-serine a gliotransmitter necessary for neuronal NMDA-mediated transmission and long term potentiation [34]Growth factors such as BDNF are also required for long termsynaptic plasticity of dendrites in the adult hippocampus [35]however decreased branching of dendrites was reported inan ABA model particularly in the stratum radiatum [11 12]Accordingly our results showed that GFAP+ cell density andGFAP expression are decreased by anorexia Similar results
10 Neural Plasticity
were reported in a rat model of depression where GFAPexpression was reduced [36] Thus we conclude that GFAP+cells and intermediate filament expression are affected byanorexia Our results add new information about the struc-tural changes observed in the hippocampus ofmurinemodelsof anorexia
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
The authors thank Dr P De Gortari (Insituto Nacionalde Psiquiatrıa Mexico) for introducing them to the DIAmodel The authors are grateful to M Macedo-MendozaM E Ramos-Aguilar E N Hernandez-Rıos A Castilla LCasanova A E Espino and M Garcıa-Servın for technicalsupport The authors thank Dr D Pless for reviewing thepaper The authors thank Dr G Ramırez-Rodrıguez (Insti-tuto Nacional de Psiquiatrıa Mexico) for donating the nestinand vimentin antibodies used in this study Janina Krugerwas an internship student supported by the DAAD-RISEprogram Durairaj Ragu Varman is a postdoctoral fellowsupported by DGAPA-UNAM This work was supported bygrants from Programa de Apoyo a Proyectos de Investigacione Innovacion Tecnologica de la UNAM (PAPIIT-UNAMIN200913 and IN201915) to Ataulfo Martınez-Torres andDaniel Reyes-Haro
References
[1] N C Barbarich-Marsteller C A Fornal L F Takase et alldquoActivity-based anorexia is associated with reduced hippocam-pal cell proliferation in adolescent female ratsrdquo BehaviouralBrain Research vol 236 no 1 pp 251ndash257 2013
[2] C Aoki N Sabaliauskas T Chowdhury et al ldquoAdolescentfemale rats exhibiting activity-based anorexia express elevatedlevels of GABAAreceptor 1205724 and 120575 subunits at the plasmamembrane of hippocampal CA1 spinesrdquo Synapse vol 66 no 5pp 391ndash407 2012
[3] G D Giordano P Renzetti R C Parodi et al ldquoVolume mea-surement with magnetic resonance imaging of hippocampus-amygdala formation in patients with anorexia nervosardquo Journalof Endocrinological Investigation vol 24 no 7 pp 510ndash514 2001
[4] F Connan F Murphy S E J Connor et al ldquoHippocampalvolume and cognitive function in anorexia nervosardquo PsychiatryResearchmdashNeuroimaging vol 146 no 2 pp 117ndash125 2006
[5] J N Beadle S Paradiso M Brumm M Voss K Halmiand L M McCormick ldquoLarger hippocampus size in womenwith anorexia nervosa who exercise excessively than healthywomenrdquo Psychiatry ResearchmdashNeuroimaging vol 232 no 2 pp193ndash199 2015
[6] N T Burkert KKoschutnig F Ebner andW Freidl ldquoStructuralhippocampal alterations perceived stress and coping deficien-cies in patients with anorexia nervosardquo International Journal ofEating Disorders vol 48 no 6 pp 670ndash676 2015
[7] S B McHugh M Fillenz J P Lowry J N P Rawlins and DM Bannerman ldquoBrain tissue oxygen amperometry in behavingrats demonstrates functional dissociation of dorsal and ventral
hippocampus during spatial processing and anxietyrdquo EuropeanJournal of Neuroscience vol 33 no 2 pp 322ndash337 2011
[8] A G Watts ldquoDehydration-associated anorexia developmentand rapid reversalrdquo Physiology and Behavior vol 65 no 4-5 pp871ndash878 1998
[9] L Jaimes-Hoy P Joseph-Bravo and P de Gortari ldquoDiffer-ential response of TRHergic neurons of the hypothalamicparaventricular nucleus (PVN) in female animals submittedto food-restriction or dehydration-induced anorexia and coldexposurerdquo Hormones and Behavior vol 53 no 2 pp 366ndash3772008
[10] A G Watts and C N Boyle ldquoThe functional architecture ofdehydration-anorexiardquo Physiology and Behavior vol 100 no 5pp 472ndash477 2010
[11] T G Chowdhury M B Rıos T E Chan D S Cassataro NC Barbarich-Marsteller and C Aoki ldquoActivity-based anorexiaduring adolescence disrupts normal development of the CA1pyramidal cells in the ventral hippocampus of female ratsrdquoHippocampus vol 24 no 12 pp 1421ndash1429 2014
[12] T G Chowdhury N C Barbarich-Marsteller T E Chan andCAoki ldquoActivity-based anorexia has differential effects on apicaldendritic branching in dorsal and ventral hippocampal CA1rdquoBrain Structure and Function vol 219 no 6 pp 1935ndash1945 2014
[13] D Reyes-Haro F E Labrada-Moncada R Miledi andA Martınez-Torres ldquoDehydration-induced anorexia reducesastrocyte density in the rat corpus callosumrdquo Neural Plasticityvol 2015 Article ID 474917 8 pages 2015
[14] P de Gortari K Mancera A Cote-Velez et al ldquoInvolvement ofCRH-R2 receptor in eating behavior and in the response of theHPT axis in rats subjected to dehydration-induced anorexiardquoPsychoneuroendocrinology vol 34 no 2 pp 259ndash272 2009
[15] M R Lamprecht DM Sabatini and A E Carpenter ldquoCellPro-filer free versatile software for automated biological imageanalysisrdquo BioTechniques vol 42 no 1 pp 71ndash75 2007
[16] B Rinaldi F Guida A Furiano et al ldquoEffect of prolongedmoderate exercise on the changes of nonneuronal cells in earlymyocardial infarctionrdquo Neural Plasticity vol 2015 Article ID265967 8 pages 2015
[17] K Chiu W M Lau H T Lau K-F So and R C-C ChangldquoMicro-dissection of rat brain for RNA or protein extractionfrom specific brain regionrdquo Journal of Visualized Experimentsno 7 article e269 2007
[18] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[19] P F Sullivan ldquoMortality in anorexia nervosardquo The AmericanJournal of Psychiatry vol 152 no 7 pp 1073ndash1074 1995
[20] C L Birmingham J Su J A Hlynsky E M Goldner and MGao ldquoThe mortality rate from anorexia nervosardquo InternationalJournal of Eating Disorders vol 38 no 2 pp 143ndash146 2005
[21] C M Bulik M C T Slof-Oprsquot Landt E F van Furth and P FSullivan ldquoThe genetics of anorexia nervosardquo Annual Review ofNutrition vol 27 pp 263ndash275 2007
[22] K J McLaughlin J O Wilson J Harman et al ldquoChronic 17120573-estradiol or cholesterol prevents stress-induced hippocampalCA3 dendritic retraction in ovariectomized female rats possi-ble correspondence between CA1 spine properties and spatialacquisitionrdquo Hippocampus vol 20 no 6 pp 768ndash786 2010
[23] K P Kinzig and S L Hargrave ldquoAdolescent activity-basedanorexia increases anxiety-like behavior in adulthoodrdquo Physi-ology and Behavior vol 101 no 2 pp 269ndash276 2010
Neural Plasticity 11
[24] S A Kim Y M Choi H J Park et al ldquoDecrease of c-Fosexpression in hippocampus of anorexia (anxanx)micerdquoKoreanJournal of Biological Sciences vol 5 no 2 pp 157ndash161 2001
[25] C Eliasson C Sahlgren C-H Berthold et al ldquoIntermediatefilament protein partnership in astrocytesrdquo The Journal ofBiological Chemistry vol 274 no 34 pp 23996ndash24006 1999
[26] R Schmidt-Kastner andCHumpel ldquoNestin expression persistsin astrocytes of organotypic slice cultures from rat cortexrdquoInternational Journal of Developmental Neuroscience vol 20 no1 pp 29ndash38 2002
[27] Y Takamiya S Kohsaka S Toya M Otani and Y TsukadaldquoImmunohistochemical studies on the proliferation of reactiveastrocytes and the expression of cytoskeletal proteins followingbrain injury in ratsrdquo Brain Research vol 466 no 2 pp 201ndash2101988
[28] S R Clarke A K Shetty J L Bradley and D A TurnerldquoReactive astrocytes express the embryonic intermediate neuro-filament nestinrdquoNeuroReport vol 5 no 15 pp 1885ndash1888 1994
[29] M C Geloso V Corvino V Cavallo et al ldquoExpression ofastrocytic nestin in the rat hippocampus during trimethyltin-induced neurodegenerationrdquo Neuroscience Letters vol 357 no2 pp 103ndash106 2004
[30] J E Holley D Gveric J Newcombe M L Cuzner and N JGutowski ldquoAstrocyte characterization in the multiple sclerosisglial scarrdquoNeuropathology andAppliedNeurobiology vol 29 no5 pp 434ndash444 2003
[31] C N Boyle S M Lorenzen D Compton and A G WattsldquoDehydration-anorexia derives from a reduction in meal sizebut not meal numberrdquo Physiology and Behavior vol 105 no 2pp 305ndash314 2012
[32] I AzcoitiaM Santos-GalindoMAArevalo and LMGarcıa-Segura ldquoRole of astroglia in the neuroplastic and neuroprotec-tive actions of estradiolrdquo European Journal of Neuroscience vol32 no 12 pp 1995ndash2002 2010
[33] L Eiland J Ramroop M N Hill J Manley and B S McEwenldquoChronic juvenile stress produces corticolimbic dendritic archi-tectural remodeling and modulates emotional behavior in maleand female ratsrdquo Psychoneuroendocrinology vol 37 no 1 pp39ndash47 2012
[34] C Henneberger T Papouin S H R Oliet and D A RusakovldquoLong-term potentiation depends on release of d-serine fromastrocytesrdquo Nature vol 463 no 7278 pp 232ndash236 2010
[35] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo The Journal ofNeuroscience vol 22 no 5 pp 1532ndash1540 2002
[36] Y Ye G Wang H Wang and X Wang ldquoBrain-derived neu-rotrophic factor (BDNF) infusion restored astrocytic plasticityin the hippocampus of a rat model of depressionrdquo NeuroscienceLetters vol 503 no 1 pp 15ndash19 2011
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
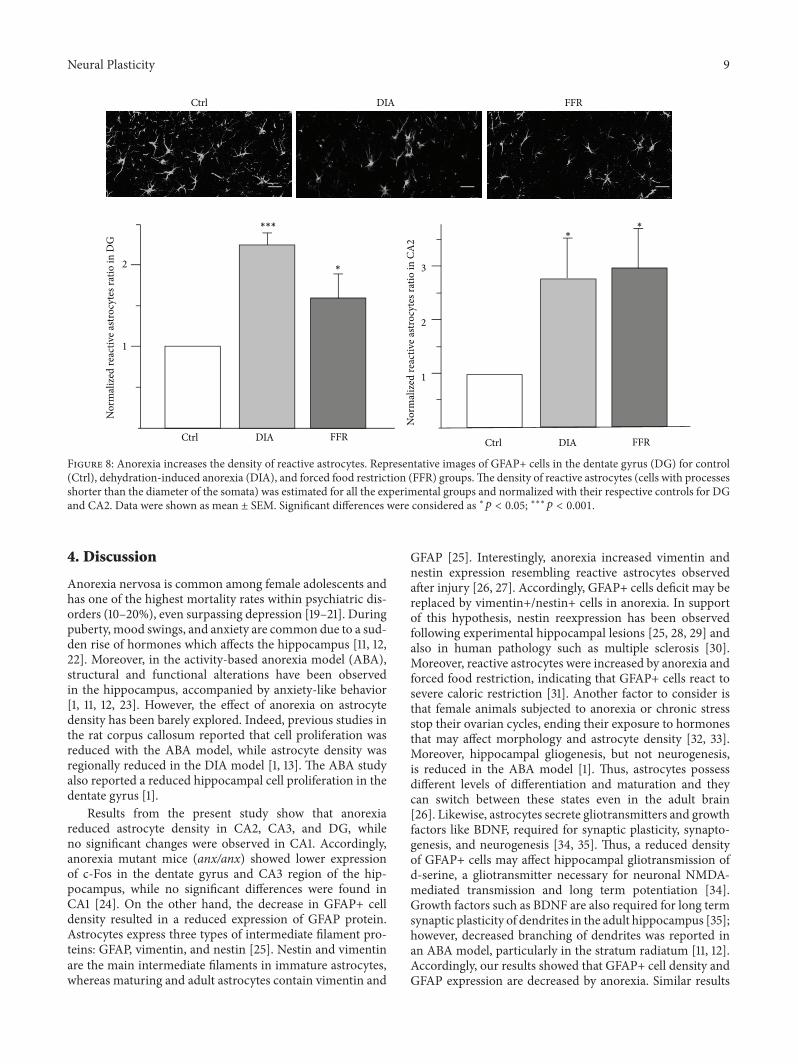
Neural Plasticity 9
RFFAIDlrtC
1
2
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in D
G
Ctrl DIA FFR
lowastlowastlowast
lowast
1
2
3
Ctrl DIA FFR
lowast
lowast
Nor
mal
ized
reac
tive a
stroc
ytes
ratio
in C
A2
Figure 8 Anorexia increases the density of reactive astrocytes Representative images of GFAP+ cells in the dentate gyrus (DG) for control(Ctrl) dehydration-induced anorexia (DIA) and forced food restriction (FFR) groupsThe density of reactive astrocytes (cells with processesshorter than the diameter of the somata) was estimated for all the experimental groups and normalized with their respective controls for DGand CA2 Data were shown as mean plusmn SEM Significant differences were considered as lowast119901 lt 005 lowastlowastlowast119901 lt 0001
4 Discussion
Anorexia nervosa is common among female adolescents andhas one of the highest mortality rates within psychiatric dis-orders (10ndash20) even surpassing depression [19ndash21] Duringpuberty mood swings and anxiety are common due to a sud-den rise of hormones which affects the hippocampus [11 1222] Moreover in the activity-based anorexia model (ABA)structural and functional alterations have been observedin the hippocampus accompanied by anxiety-like behavior[1 11 12 23] However the effect of anorexia on astrocytedensity has been barely explored Indeed previous studies inthe rat corpus callosum reported that cell proliferation wasreduced with the ABA model while astrocyte density wasregionally reduced in the DIA model [1 13] The ABA studyalso reported a reduced hippocampal cell proliferation in thedentate gyrus [1]
Results from the present study show that anorexiareduced astrocyte density in CA2 CA3 and DG whileno significant changes were observed in CA1 Accordinglyanorexia mutant mice (anxanx) showed lower expressionof c-Fos in the dentate gyrus and CA3 region of the hip-pocampus while no significant differences were found inCA1 [24] On the other hand the decrease in GFAP+ celldensity resulted in a reduced expression of GFAP proteinAstrocytes express three types of intermediate filament pro-teins GFAP vimentin and nestin [25] Nestin and vimentinare the main intermediate filaments in immature astrocyteswhereas maturing and adult astrocytes contain vimentin and
GFAP [25] Interestingly anorexia increased vimentin andnestin expression resembling reactive astrocytes observedafter injury [26 27] Accordingly GFAP+ cells deficit may bereplaced by vimentin+nestin+ cells in anorexia In supportof this hypothesis nestin reexpression has been observedfollowing experimental hippocampal lesions [25 28 29] andalso in human pathology such as multiple sclerosis [30]Moreover reactive astrocytes were increased by anorexia andforced food restriction indicating that GFAP+ cells react tosevere caloric restriction [31] Another factor to consider isthat female animals subjected to anorexia or chronic stressstop their ovarian cycles ending their exposure to hormonesthat may affect morphology and astrocyte density [32 33]Moreover hippocampal gliogenesis but not neurogenesisis reduced in the ABA model [1] Thus astrocytes possessdifferent levels of differentiation and maturation and theycan switch between these states even in the adult brain[26] Likewise astrocytes secrete gliotransmitters and growthfactors like BDNF required for synaptic plasticity synapto-genesis and neurogenesis [34 35] Thus a reduced densityof GFAP+ cells may affect hippocampal gliotransmission ofd-serine a gliotransmitter necessary for neuronal NMDA-mediated transmission and long term potentiation [34]Growth factors such as BDNF are also required for long termsynaptic plasticity of dendrites in the adult hippocampus [35]however decreased branching of dendrites was reported inan ABA model particularly in the stratum radiatum [11 12]Accordingly our results showed that GFAP+ cell density andGFAP expression are decreased by anorexia Similar results
10 Neural Plasticity
were reported in a rat model of depression where GFAPexpression was reduced [36] Thus we conclude that GFAP+cells and intermediate filament expression are affected byanorexia Our results add new information about the struc-tural changes observed in the hippocampus ofmurinemodelsof anorexia
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
The authors thank Dr P De Gortari (Insituto Nacionalde Psiquiatrıa Mexico) for introducing them to the DIAmodel The authors are grateful to M Macedo-MendozaM E Ramos-Aguilar E N Hernandez-Rıos A Castilla LCasanova A E Espino and M Garcıa-Servın for technicalsupport The authors thank Dr D Pless for reviewing thepaper The authors thank Dr G Ramırez-Rodrıguez (Insti-tuto Nacional de Psiquiatrıa Mexico) for donating the nestinand vimentin antibodies used in this study Janina Krugerwas an internship student supported by the DAAD-RISEprogram Durairaj Ragu Varman is a postdoctoral fellowsupported by DGAPA-UNAM This work was supported bygrants from Programa de Apoyo a Proyectos de Investigacione Innovacion Tecnologica de la UNAM (PAPIIT-UNAMIN200913 and IN201915) to Ataulfo Martınez-Torres andDaniel Reyes-Haro
References
[1] N C Barbarich-Marsteller C A Fornal L F Takase et alldquoActivity-based anorexia is associated with reduced hippocam-pal cell proliferation in adolescent female ratsrdquo BehaviouralBrain Research vol 236 no 1 pp 251ndash257 2013
[2] C Aoki N Sabaliauskas T Chowdhury et al ldquoAdolescentfemale rats exhibiting activity-based anorexia express elevatedlevels of GABAAreceptor 1205724 and 120575 subunits at the plasmamembrane of hippocampal CA1 spinesrdquo Synapse vol 66 no 5pp 391ndash407 2012
[3] G D Giordano P Renzetti R C Parodi et al ldquoVolume mea-surement with magnetic resonance imaging of hippocampus-amygdala formation in patients with anorexia nervosardquo Journalof Endocrinological Investigation vol 24 no 7 pp 510ndash514 2001
[4] F Connan F Murphy S E J Connor et al ldquoHippocampalvolume and cognitive function in anorexia nervosardquo PsychiatryResearchmdashNeuroimaging vol 146 no 2 pp 117ndash125 2006
[5] J N Beadle S Paradiso M Brumm M Voss K Halmiand L M McCormick ldquoLarger hippocampus size in womenwith anorexia nervosa who exercise excessively than healthywomenrdquo Psychiatry ResearchmdashNeuroimaging vol 232 no 2 pp193ndash199 2015
[6] N T Burkert KKoschutnig F Ebner andW Freidl ldquoStructuralhippocampal alterations perceived stress and coping deficien-cies in patients with anorexia nervosardquo International Journal ofEating Disorders vol 48 no 6 pp 670ndash676 2015
[7] S B McHugh M Fillenz J P Lowry J N P Rawlins and DM Bannerman ldquoBrain tissue oxygen amperometry in behavingrats demonstrates functional dissociation of dorsal and ventral
hippocampus during spatial processing and anxietyrdquo EuropeanJournal of Neuroscience vol 33 no 2 pp 322ndash337 2011
[8] A G Watts ldquoDehydration-associated anorexia developmentand rapid reversalrdquo Physiology and Behavior vol 65 no 4-5 pp871ndash878 1998
[9] L Jaimes-Hoy P Joseph-Bravo and P de Gortari ldquoDiffer-ential response of TRHergic neurons of the hypothalamicparaventricular nucleus (PVN) in female animals submittedto food-restriction or dehydration-induced anorexia and coldexposurerdquo Hormones and Behavior vol 53 no 2 pp 366ndash3772008
[10] A G Watts and C N Boyle ldquoThe functional architecture ofdehydration-anorexiardquo Physiology and Behavior vol 100 no 5pp 472ndash477 2010
[11] T G Chowdhury M B Rıos T E Chan D S Cassataro NC Barbarich-Marsteller and C Aoki ldquoActivity-based anorexiaduring adolescence disrupts normal development of the CA1pyramidal cells in the ventral hippocampus of female ratsrdquoHippocampus vol 24 no 12 pp 1421ndash1429 2014
[12] T G Chowdhury N C Barbarich-Marsteller T E Chan andCAoki ldquoActivity-based anorexia has differential effects on apicaldendritic branching in dorsal and ventral hippocampal CA1rdquoBrain Structure and Function vol 219 no 6 pp 1935ndash1945 2014
[13] D Reyes-Haro F E Labrada-Moncada R Miledi andA Martınez-Torres ldquoDehydration-induced anorexia reducesastrocyte density in the rat corpus callosumrdquo Neural Plasticityvol 2015 Article ID 474917 8 pages 2015
[14] P de Gortari K Mancera A Cote-Velez et al ldquoInvolvement ofCRH-R2 receptor in eating behavior and in the response of theHPT axis in rats subjected to dehydration-induced anorexiardquoPsychoneuroendocrinology vol 34 no 2 pp 259ndash272 2009
[15] M R Lamprecht DM Sabatini and A E Carpenter ldquoCellPro-filer free versatile software for automated biological imageanalysisrdquo BioTechniques vol 42 no 1 pp 71ndash75 2007
[16] B Rinaldi F Guida A Furiano et al ldquoEffect of prolongedmoderate exercise on the changes of nonneuronal cells in earlymyocardial infarctionrdquo Neural Plasticity vol 2015 Article ID265967 8 pages 2015
[17] K Chiu W M Lau H T Lau K-F So and R C-C ChangldquoMicro-dissection of rat brain for RNA or protein extractionfrom specific brain regionrdquo Journal of Visualized Experimentsno 7 article e269 2007
[18] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[19] P F Sullivan ldquoMortality in anorexia nervosardquo The AmericanJournal of Psychiatry vol 152 no 7 pp 1073ndash1074 1995
[20] C L Birmingham J Su J A Hlynsky E M Goldner and MGao ldquoThe mortality rate from anorexia nervosardquo InternationalJournal of Eating Disorders vol 38 no 2 pp 143ndash146 2005
[21] C M Bulik M C T Slof-Oprsquot Landt E F van Furth and P FSullivan ldquoThe genetics of anorexia nervosardquo Annual Review ofNutrition vol 27 pp 263ndash275 2007
[22] K J McLaughlin J O Wilson J Harman et al ldquoChronic 17120573-estradiol or cholesterol prevents stress-induced hippocampalCA3 dendritic retraction in ovariectomized female rats possi-ble correspondence between CA1 spine properties and spatialacquisitionrdquo Hippocampus vol 20 no 6 pp 768ndash786 2010
[23] K P Kinzig and S L Hargrave ldquoAdolescent activity-basedanorexia increases anxiety-like behavior in adulthoodrdquo Physi-ology and Behavior vol 101 no 2 pp 269ndash276 2010
Neural Plasticity 11
[24] S A Kim Y M Choi H J Park et al ldquoDecrease of c-Fosexpression in hippocampus of anorexia (anxanx)micerdquoKoreanJournal of Biological Sciences vol 5 no 2 pp 157ndash161 2001
[25] C Eliasson C Sahlgren C-H Berthold et al ldquoIntermediatefilament protein partnership in astrocytesrdquo The Journal ofBiological Chemistry vol 274 no 34 pp 23996ndash24006 1999
[26] R Schmidt-Kastner andCHumpel ldquoNestin expression persistsin astrocytes of organotypic slice cultures from rat cortexrdquoInternational Journal of Developmental Neuroscience vol 20 no1 pp 29ndash38 2002
[27] Y Takamiya S Kohsaka S Toya M Otani and Y TsukadaldquoImmunohistochemical studies on the proliferation of reactiveastrocytes and the expression of cytoskeletal proteins followingbrain injury in ratsrdquo Brain Research vol 466 no 2 pp 201ndash2101988
[28] S R Clarke A K Shetty J L Bradley and D A TurnerldquoReactive astrocytes express the embryonic intermediate neuro-filament nestinrdquoNeuroReport vol 5 no 15 pp 1885ndash1888 1994
[29] M C Geloso V Corvino V Cavallo et al ldquoExpression ofastrocytic nestin in the rat hippocampus during trimethyltin-induced neurodegenerationrdquo Neuroscience Letters vol 357 no2 pp 103ndash106 2004
[30] J E Holley D Gveric J Newcombe M L Cuzner and N JGutowski ldquoAstrocyte characterization in the multiple sclerosisglial scarrdquoNeuropathology andAppliedNeurobiology vol 29 no5 pp 434ndash444 2003
[31] C N Boyle S M Lorenzen D Compton and A G WattsldquoDehydration-anorexia derives from a reduction in meal sizebut not meal numberrdquo Physiology and Behavior vol 105 no 2pp 305ndash314 2012
[32] I AzcoitiaM Santos-GalindoMAArevalo and LMGarcıa-Segura ldquoRole of astroglia in the neuroplastic and neuroprotec-tive actions of estradiolrdquo European Journal of Neuroscience vol32 no 12 pp 1995ndash2002 2010
[33] L Eiland J Ramroop M N Hill J Manley and B S McEwenldquoChronic juvenile stress produces corticolimbic dendritic archi-tectural remodeling and modulates emotional behavior in maleand female ratsrdquo Psychoneuroendocrinology vol 37 no 1 pp39ndash47 2012
[34] C Henneberger T Papouin S H R Oliet and D A RusakovldquoLong-term potentiation depends on release of d-serine fromastrocytesrdquo Nature vol 463 no 7278 pp 232ndash236 2010
[35] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo The Journal ofNeuroscience vol 22 no 5 pp 1532ndash1540 2002
[36] Y Ye G Wang H Wang and X Wang ldquoBrain-derived neu-rotrophic factor (BDNF) infusion restored astrocytic plasticityin the hippocampus of a rat model of depressionrdquo NeuroscienceLetters vol 503 no 1 pp 15ndash19 2011
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

10 Neural Plasticity
were reported in a rat model of depression where GFAPexpression was reduced [36] Thus we conclude that GFAP+cells and intermediate filament expression are affected byanorexia Our results add new information about the struc-tural changes observed in the hippocampus ofmurinemodelsof anorexia
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
The authors thank Dr P De Gortari (Insituto Nacionalde Psiquiatrıa Mexico) for introducing them to the DIAmodel The authors are grateful to M Macedo-MendozaM E Ramos-Aguilar E N Hernandez-Rıos A Castilla LCasanova A E Espino and M Garcıa-Servın for technicalsupport The authors thank Dr D Pless for reviewing thepaper The authors thank Dr G Ramırez-Rodrıguez (Insti-tuto Nacional de Psiquiatrıa Mexico) for donating the nestinand vimentin antibodies used in this study Janina Krugerwas an internship student supported by the DAAD-RISEprogram Durairaj Ragu Varman is a postdoctoral fellowsupported by DGAPA-UNAM This work was supported bygrants from Programa de Apoyo a Proyectos de Investigacione Innovacion Tecnologica de la UNAM (PAPIIT-UNAMIN200913 and IN201915) to Ataulfo Martınez-Torres andDaniel Reyes-Haro
References
[1] N C Barbarich-Marsteller C A Fornal L F Takase et alldquoActivity-based anorexia is associated with reduced hippocam-pal cell proliferation in adolescent female ratsrdquo BehaviouralBrain Research vol 236 no 1 pp 251ndash257 2013
[2] C Aoki N Sabaliauskas T Chowdhury et al ldquoAdolescentfemale rats exhibiting activity-based anorexia express elevatedlevels of GABAAreceptor 1205724 and 120575 subunits at the plasmamembrane of hippocampal CA1 spinesrdquo Synapse vol 66 no 5pp 391ndash407 2012
[3] G D Giordano P Renzetti R C Parodi et al ldquoVolume mea-surement with magnetic resonance imaging of hippocampus-amygdala formation in patients with anorexia nervosardquo Journalof Endocrinological Investigation vol 24 no 7 pp 510ndash514 2001
[4] F Connan F Murphy S E J Connor et al ldquoHippocampalvolume and cognitive function in anorexia nervosardquo PsychiatryResearchmdashNeuroimaging vol 146 no 2 pp 117ndash125 2006
[5] J N Beadle S Paradiso M Brumm M Voss K Halmiand L M McCormick ldquoLarger hippocampus size in womenwith anorexia nervosa who exercise excessively than healthywomenrdquo Psychiatry ResearchmdashNeuroimaging vol 232 no 2 pp193ndash199 2015
[6] N T Burkert KKoschutnig F Ebner andW Freidl ldquoStructuralhippocampal alterations perceived stress and coping deficien-cies in patients with anorexia nervosardquo International Journal ofEating Disorders vol 48 no 6 pp 670ndash676 2015
[7] S B McHugh M Fillenz J P Lowry J N P Rawlins and DM Bannerman ldquoBrain tissue oxygen amperometry in behavingrats demonstrates functional dissociation of dorsal and ventral
hippocampus during spatial processing and anxietyrdquo EuropeanJournal of Neuroscience vol 33 no 2 pp 322ndash337 2011
[8] A G Watts ldquoDehydration-associated anorexia developmentand rapid reversalrdquo Physiology and Behavior vol 65 no 4-5 pp871ndash878 1998
[9] L Jaimes-Hoy P Joseph-Bravo and P de Gortari ldquoDiffer-ential response of TRHergic neurons of the hypothalamicparaventricular nucleus (PVN) in female animals submittedto food-restriction or dehydration-induced anorexia and coldexposurerdquo Hormones and Behavior vol 53 no 2 pp 366ndash3772008
[10] A G Watts and C N Boyle ldquoThe functional architecture ofdehydration-anorexiardquo Physiology and Behavior vol 100 no 5pp 472ndash477 2010
[11] T G Chowdhury M B Rıos T E Chan D S Cassataro NC Barbarich-Marsteller and C Aoki ldquoActivity-based anorexiaduring adolescence disrupts normal development of the CA1pyramidal cells in the ventral hippocampus of female ratsrdquoHippocampus vol 24 no 12 pp 1421ndash1429 2014
[12] T G Chowdhury N C Barbarich-Marsteller T E Chan andCAoki ldquoActivity-based anorexia has differential effects on apicaldendritic branching in dorsal and ventral hippocampal CA1rdquoBrain Structure and Function vol 219 no 6 pp 1935ndash1945 2014
[13] D Reyes-Haro F E Labrada-Moncada R Miledi andA Martınez-Torres ldquoDehydration-induced anorexia reducesastrocyte density in the rat corpus callosumrdquo Neural Plasticityvol 2015 Article ID 474917 8 pages 2015
[14] P de Gortari K Mancera A Cote-Velez et al ldquoInvolvement ofCRH-R2 receptor in eating behavior and in the response of theHPT axis in rats subjected to dehydration-induced anorexiardquoPsychoneuroendocrinology vol 34 no 2 pp 259ndash272 2009
[15] M R Lamprecht DM Sabatini and A E Carpenter ldquoCellPro-filer free versatile software for automated biological imageanalysisrdquo BioTechniques vol 42 no 1 pp 71ndash75 2007
[16] B Rinaldi F Guida A Furiano et al ldquoEffect of prolongedmoderate exercise on the changes of nonneuronal cells in earlymyocardial infarctionrdquo Neural Plasticity vol 2015 Article ID265967 8 pages 2015
[17] K Chiu W M Lau H T Lau K-F So and R C-C ChangldquoMicro-dissection of rat brain for RNA or protein extractionfrom specific brain regionrdquo Journal of Visualized Experimentsno 7 article e269 2007
[18] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein-dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[19] P F Sullivan ldquoMortality in anorexia nervosardquo The AmericanJournal of Psychiatry vol 152 no 7 pp 1073ndash1074 1995
[20] C L Birmingham J Su J A Hlynsky E M Goldner and MGao ldquoThe mortality rate from anorexia nervosardquo InternationalJournal of Eating Disorders vol 38 no 2 pp 143ndash146 2005
[21] C M Bulik M C T Slof-Oprsquot Landt E F van Furth and P FSullivan ldquoThe genetics of anorexia nervosardquo Annual Review ofNutrition vol 27 pp 263ndash275 2007
[22] K J McLaughlin J O Wilson J Harman et al ldquoChronic 17120573-estradiol or cholesterol prevents stress-induced hippocampalCA3 dendritic retraction in ovariectomized female rats possi-ble correspondence between CA1 spine properties and spatialacquisitionrdquo Hippocampus vol 20 no 6 pp 768ndash786 2010
[23] K P Kinzig and S L Hargrave ldquoAdolescent activity-basedanorexia increases anxiety-like behavior in adulthoodrdquo Physi-ology and Behavior vol 101 no 2 pp 269ndash276 2010
Neural Plasticity 11
[24] S A Kim Y M Choi H J Park et al ldquoDecrease of c-Fosexpression in hippocampus of anorexia (anxanx)micerdquoKoreanJournal of Biological Sciences vol 5 no 2 pp 157ndash161 2001
[25] C Eliasson C Sahlgren C-H Berthold et al ldquoIntermediatefilament protein partnership in astrocytesrdquo The Journal ofBiological Chemistry vol 274 no 34 pp 23996ndash24006 1999
[26] R Schmidt-Kastner andCHumpel ldquoNestin expression persistsin astrocytes of organotypic slice cultures from rat cortexrdquoInternational Journal of Developmental Neuroscience vol 20 no1 pp 29ndash38 2002
[27] Y Takamiya S Kohsaka S Toya M Otani and Y TsukadaldquoImmunohistochemical studies on the proliferation of reactiveastrocytes and the expression of cytoskeletal proteins followingbrain injury in ratsrdquo Brain Research vol 466 no 2 pp 201ndash2101988
[28] S R Clarke A K Shetty J L Bradley and D A TurnerldquoReactive astrocytes express the embryonic intermediate neuro-filament nestinrdquoNeuroReport vol 5 no 15 pp 1885ndash1888 1994
[29] M C Geloso V Corvino V Cavallo et al ldquoExpression ofastrocytic nestin in the rat hippocampus during trimethyltin-induced neurodegenerationrdquo Neuroscience Letters vol 357 no2 pp 103ndash106 2004
[30] J E Holley D Gveric J Newcombe M L Cuzner and N JGutowski ldquoAstrocyte characterization in the multiple sclerosisglial scarrdquoNeuropathology andAppliedNeurobiology vol 29 no5 pp 434ndash444 2003
[31] C N Boyle S M Lorenzen D Compton and A G WattsldquoDehydration-anorexia derives from a reduction in meal sizebut not meal numberrdquo Physiology and Behavior vol 105 no 2pp 305ndash314 2012
[32] I AzcoitiaM Santos-GalindoMAArevalo and LMGarcıa-Segura ldquoRole of astroglia in the neuroplastic and neuroprotec-tive actions of estradiolrdquo European Journal of Neuroscience vol32 no 12 pp 1995ndash2002 2010
[33] L Eiland J Ramroop M N Hill J Manley and B S McEwenldquoChronic juvenile stress produces corticolimbic dendritic archi-tectural remodeling and modulates emotional behavior in maleand female ratsrdquo Psychoneuroendocrinology vol 37 no 1 pp39ndash47 2012
[34] C Henneberger T Papouin S H R Oliet and D A RusakovldquoLong-term potentiation depends on release of d-serine fromastrocytesrdquo Nature vol 463 no 7278 pp 232ndash236 2010
[35] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo The Journal ofNeuroscience vol 22 no 5 pp 1532ndash1540 2002
[36] Y Ye G Wang H Wang and X Wang ldquoBrain-derived neu-rotrophic factor (BDNF) infusion restored astrocytic plasticityin the hippocampus of a rat model of depressionrdquo NeuroscienceLetters vol 503 no 1 pp 15ndash19 2011
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Neural Plasticity 11
[24] S A Kim Y M Choi H J Park et al ldquoDecrease of c-Fosexpression in hippocampus of anorexia (anxanx)micerdquoKoreanJournal of Biological Sciences vol 5 no 2 pp 157ndash161 2001
[25] C Eliasson C Sahlgren C-H Berthold et al ldquoIntermediatefilament protein partnership in astrocytesrdquo The Journal ofBiological Chemistry vol 274 no 34 pp 23996ndash24006 1999
[26] R Schmidt-Kastner andCHumpel ldquoNestin expression persistsin astrocytes of organotypic slice cultures from rat cortexrdquoInternational Journal of Developmental Neuroscience vol 20 no1 pp 29ndash38 2002
[27] Y Takamiya S Kohsaka S Toya M Otani and Y TsukadaldquoImmunohistochemical studies on the proliferation of reactiveastrocytes and the expression of cytoskeletal proteins followingbrain injury in ratsrdquo Brain Research vol 466 no 2 pp 201ndash2101988
[28] S R Clarke A K Shetty J L Bradley and D A TurnerldquoReactive astrocytes express the embryonic intermediate neuro-filament nestinrdquoNeuroReport vol 5 no 15 pp 1885ndash1888 1994
[29] M C Geloso V Corvino V Cavallo et al ldquoExpression ofastrocytic nestin in the rat hippocampus during trimethyltin-induced neurodegenerationrdquo Neuroscience Letters vol 357 no2 pp 103ndash106 2004
[30] J E Holley D Gveric J Newcombe M L Cuzner and N JGutowski ldquoAstrocyte characterization in the multiple sclerosisglial scarrdquoNeuropathology andAppliedNeurobiology vol 29 no5 pp 434ndash444 2003
[31] C N Boyle S M Lorenzen D Compton and A G WattsldquoDehydration-anorexia derives from a reduction in meal sizebut not meal numberrdquo Physiology and Behavior vol 105 no 2pp 305ndash314 2012
[32] I AzcoitiaM Santos-GalindoMAArevalo and LMGarcıa-Segura ldquoRole of astroglia in the neuroplastic and neuroprotec-tive actions of estradiolrdquo European Journal of Neuroscience vol32 no 12 pp 1995ndash2002 2010
[33] L Eiland J Ramroop M N Hill J Manley and B S McEwenldquoChronic juvenile stress produces corticolimbic dendritic archi-tectural remodeling and modulates emotional behavior in maleand female ratsrdquo Psychoneuroendocrinology vol 37 no 1 pp39ndash47 2012
[34] C Henneberger T Papouin S H R Oliet and D A RusakovldquoLong-term potentiation depends on release of d-serine fromastrocytesrdquo Nature vol 463 no 7278 pp 232ndash236 2010
[35] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo The Journal ofNeuroscience vol 22 no 5 pp 1532ndash1540 2002
[36] Y Ye G Wang H Wang and X Wang ldquoBrain-derived neu-rotrophic factor (BDNF) infusion restored astrocytic plasticityin the hippocampus of a rat model of depressionrdquo NeuroscienceLetters vol 503 no 1 pp 15ndash19 2011
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014


















