
renewal of glycerol in the visual cells and pigment epithelium of the ...
12
RENEWALOFGLYCEROLINTHEVISUALCELLS ANDPIGMENTEPITHELIUMOFTHEFROGRETINA CAROLBIBBandRICHARDW .YOUNG FromtheDepartmentofAnatomyandthe Jules SteinEyeInstitute,UniversityofCaliforniaatLos AngelesSchoolofMedicine,LosAngeles,California 90024 ABSTRACT Therenewalofglycerolinthevisualcellsandpigmentepitheliumofthefrogretina wasstudiedbyautoradiographicanalysisofanimalsinjectedwith[2- 1 1]glycerol . Assayofchloroform :methanolextractsshowedthatthelabeledprecursorwasused mainlyinlipidsynthesis,althoughtherewasalsosomeutilizationintheformation ofprotein .Radioactiveglycerolwasinitiallyconcentratedinthemyoidportionof rodsandcones,indicatingthatthisisthesiteofphospholipidsynthesisinvisual cells .Theglycogenbodies(paraboloids)ofaccessoryconeswerealsoheavily labeled,suggestingthediversionofsomeglycerolintoglycogenicpathways .Inthe pigmentepithelium,onlytheoildropletsbecamesignificantlyradioactive .The outerplexiformlayer(whichcontainsthevisualcellsynapticbodies)andthecone oildropletsgraduallyaccumulatedconsiderableamountsoflabeledmaterial . Within1-4h,labeledmoleculesbegantoappearinthevisualcelloutersegments, evidentlyhavingbeentransportedtherefromthemyoidportionoftheinner segment .Mostofthesewerephospholipidmoleculeswhichbecamedistributed throughouttheoutersegments,presumablyreplacingcomparableconstituentsin existingmembranes .Inrodsonly,therewasalsoanaggregationoflabeled materialatthebaseoftheoutersegmentduetomembranebiogenesis .These highlyradioactivemembranes,containinglabeledmoleculesoflipidandprotein, weresubsequentlydisplacedalongtherodoutersegmentsduetorepeated membraneassemblyatthebase .Thedistributionofradioactivitysupportedthe conclusionthatmembranerenewalbymolecularreplacementismorerapidfor lipidthanitisforprotein . INTRODUCTION Renewalofthestackofdisk-shaped,light-sensitive membraneswhichcomprisestheoutersegmentof rodandconevisualcellshasbeenstudiedexten- sivelybyautoradiographyusingradioactiveamino acidsasproteinprecursors(5,9,13,29,30,32,33, 37,39) .Withinminutesafterinjectionintoverte- brateanimals,labeledaminoacidsbecomecon- centratedinthemyoidportionoftheinnerseg- 37 8 mentofthecell(Fig .1) .Thiscellcompartment . whichcontainsfreeandmembTane-bound ribo- somes, theGolgicomplex,andessentiallyallofthe cell'scytoplasmicRNA(3,31)isthemajorsiteof proteinsynthesisinbothrodsandcones .Protein destinedfortherenewaloftheoutersegment membranesistransportedpastthemitochondria massedintheellipsoidregion,andthenthrough THEJOURNALOFCELLBIOLOGY . VOLUME62,1974 • pages 378-389 on February 11, 2018 jcb.rupress.org Downloaded from
Transcript of renewal of glycerol in the visual cells and pigment epithelium of the ...

RENEWAL OF GLYCEROL IN THE VISUAL CELLSAND PIGMENT EPITHELIUM OF THE FROG RETINA
CAROL BIBB and RICHARD W . YOUNG
From the Department of Anatomy and the Jules Stein Eye Institute, University of California at LosAngeles School of Medicine, Los Angeles, California 90024
ABSTRACT
The renewal of glycerol in the visual cells and pigment epithelium of the frog retinawas studied by autoradiographic analysis of animals injected with [2- 11]glycerol .Assay of chloroform : methanol extracts showed that the labeled precursor was used
mainly in lipid synthesis, although there was also some utilization in the formationof protein. Radioactive glycerol was initially concentrated in the myoid portion ofrods and cones, indicating that this is the site of phospholipid synthesis in visualcells. The glycogen bodies (paraboloids) of accessory cones were also heavilylabeled, suggesting the diversion of some glycerol into glycogenic pathways . In thepigment epithelium, only the oil droplets became significantly radioactive . Theouter plexiform layer (which contains the visual cell synaptic bodies) and the cone
oil droplets gradually accumulated considerable amounts of labeled material .Within 1-4 h, labeled molecules began to appear in the visual cell outer segments,evidently having been transported there from the myoid portion of the innersegment. Most of these were phospholipid molecules which became distributedthroughout the outer segments, presumably replacing comparable constituents inexisting membranes . In rods only, there was also an aggregation of labeledmaterial at the base of the outer segment due to membrane biogenesis . Thesehighly radioactive membranes, containing labeled molecules of lipid and protein,were subsequently displaced along the rod outer segments due to repeatedmembrane assembly at the base . The distribution of radioactivity supported theconclusion that membrane renewal by molecular replacement is more rapid forlipid than it is for protein .
INTRODUCTION
Renewal of the stack of disk-shaped, light-sensitivemembranes which comprises the outer segment ofrod and cone visual cells has been studied exten-sively by autoradiography using radioactive aminoacids as protein precursors (5, 9, 13, 29, 30, 32, 33,37, 39) . Within minutes after injection into verte-brate animals, labeled amino acids become con-centrated in the myoid portion of the inner seg-
3 78
ment of the cell (Fig . 1) . This cell compartment .which contains free and membTane-bound ribo-somes, the Golgi complex, and essentially all of thecell's cytoplasmic RNA (3, 31) is the major site ofprotein synthesis in both rods and cones . Proteindestined for the renewal of the outer segmentmembranes is transported past the mitochondriamassed in the ellipsoid region, and then through
THE JOURNAL OF CELL BIOLOGY . VOLUME 62, 1974 • pages 378-389
on February 11, 2018
jcb.rupress.orgD
ownloaded from

FIGURE I Diagram of rod and cone visual cells andtheir relationship to the pigment epithelium in the frogretina . In the pigment epithelium, the oil droplets arestorage sites for vitamin A . Phagosomes contain groupsof phagocytized membranes shed from the tip of the rodouter segments . In the visual cells, the outer segmentconsists of a photosensitive stack of disk-shaped doublemembranes, derived from the outer cell membrane byinfolding . The ellipsoid is filled with mitochondria,whereas the myoid contains a granular endoplasmicreticulum, free ribosomes, and the Golgi complex . To-gether, the ellipsoid and myoid make up the innersegment of the cell . A double cone is shown . Theprincipal member contains an oil droplet (of unknownfunction) ; the accessory member contains a glycogenmass (paraboloid) . A few rods with short outer segmentsand some single cones bearing oil droplets are alsopresent in small numbers in the retina .
the hollow connecting cilium to the outer segment .In rods, much of this protein is used in thecontinual assembly of new lipoprotein membranesat the base of the outer segment . Membraneformation occurs by a process in which the outercell membrane folds inward, creating a circular,double-membrane disk, which subsequently losesits peripheral attachment, and is separated from
adjacent disks by a thin film of cytoplasm . Whenthe proteins in new disks are labeled as a result ofthe injection of radioactive amino acids, the fate ofthese labeled disks may be followed by autoradiog-raphy. Repeated formation of membranous disksat the base of the rod outer segment displacespreviously formed disks away from the base and
towards the pigment epithelium . To maintain thelength of the stack of outer segment membranes,continual membrane biogenesis at the base isbalanced by the intermittent shedding of groups ofold membranes from the apex . These discarded
membranes are phagocytized and destroyed by thepigment epithelium (34, 37) . This mechanism of
renewal by membrane replacement apparentlyoccurs only in rods (32, 35) .
In rod outer segments, protein which is not usedin the assembly of new membranes becomesdiffusely distributed in or among the previouslyformed disks . In cones, all of the new proteindelivered to the outer segment becomes diffuselydistributed in this manner (5, 32). This represents a
different type of renewal process-renewal bymolecular replacement-in which new moleculesreplace old ones in or among the membranes of theexisting membrane system (36) .
When labeled fatty acids are used as precursorsof the lipid components of the membranes in frogs(2), a different autoradiographic pattern is ob-
served . Instead of initially being concentrated inthe myoid part of the cell, in the manner of aminoacids, the fatty acids immediately appear through-out the cell, including the outer segments . Outersegment membranes contain abundant fatty acidsas components of phospholipid molecules, but lackthe enzymes to produce phospholipids (27) . Wetherefore concluded that the direct incorporationof fatty acids in the outer segments is due to fattyacid exchange in phospholipid molecules already
situated in the membranes (2) . Gradually, theconcentration of radioactive material in the visualcell outer segment membranes increases, appar-ently as a result of the addition of new phospho-lipid molecules containing labeled fatty acids,perhaps supplemented by the transfer to the outersegments from the pigment epithelium of vitaminA esterified with fatty acid . Within a few days,labeled fatty acid beomes intensely localized in thenew membranes which have formed at the base ofthe rod outer segments since the injection . But thisdiscrete concentration of radioactivity in newmembrane is short-lived, apparently due to therapid rate of fatty acid renewal . In contrast, when
C . BIBB AND R. W. YOUNG Glycerol Renewal in Outer Segment Membranes 379
on February 11, 2018
jcb.rupress.orgD
ownloaded from

proteins of the disk membranes are labeled, thereis no significant decline in their content of radioac-tivity as they are displaced along the outer segment(13, 29) .
These studies demonstrated that the lipids of thevisual cell outer segment membranes are continu-ally renewed, provided further evidence of bothmembrane and molecular replacement, and re-vealed some differences in the mechanisms bywhich the lipid and protein constituents of themembranes undergo turnover . However, the com-plexity of fatty acid metabolism in these cells,involving processes of exchange, vitamin A es-terification, and phospholipid synthesis-all ofwhich are superimposed in the autoradiographicimage-did not enable us to answer the question ofwhere phospholipid synthesis takes place .
Glycerol should serve as a useful lipid precursorin the study of phospholipid synthesis, as well asmembrane biogenesis and renewal . About 40% ofthe dry weight of the outer segment membranes islipid, most of which is phospholipid (1, 6, 11, 20) .Glycerol is the structural backbone of the phos-pholipid molecule, does not undergo exchangeonce it has been incorporated into phospholipid,and is not involved in vitamin A esterification .Accordingly, we have used tritiated glycerol in theautoradiographic analysis of lipid renewal in thevisual cells and pigment epithelium of the frog,with particular emphasis on the membranes of therod and cone outer segments .
MATERIALS AND METHODS
A utoradiograph yNine frogs (Rana pipiens), weighing between 2 .4 and
15 .0 g (average, 8 .0 g), were injected with [2- 3 H]glycerolin aqueous solution at a concentration of 100 mCi/ml .The labeled compound was obtained from New EnglandNuclear Corp., Boston, Mass ., at a specific activity of7 .85 to 8 .81 Ci/mmol . The animals were killed at 15 min,I h (2 frogs), 4 h, 15 h, I day, I wk, 5 wk, and 8 wk afterinjection . The dosage was I mCi/g body weight, exceptthat the frog killed at 15 min received 10 mCi/g . Allinjections were made intravenously, under anesthesia,except for the animals killed at I wk or later, which wereinjected in the dorsal lymph sac without anesthesia . Atsacrifice, the eyes were fixed in phosphate-buffered 4%formaldehyde, pH 7 .2, and left in that solution over-night . The posterior half of the eye was then cut intosmaller pieces, which were rinsed in buffer . Half of thesewere then fixed in 1% osmium tetroxide and embedded inAraldite using an abbreviated dehydration proceduredesigned to retain lipids (2) . The remaining specimenswere subjected to prolonged extraction with chloroform :
3 8 0
THE JOURNAL OF CELL BIOLOGY . VOLUME 62, 1974
methanol (2 : 1, vol/vol) to remove lipids . They were thenfixed in osmium tetroxide and embedded in Araldite (2) .
The autoradiograms were developed in Kodak Dektol(Eastman Kodak Co ., Rochester, N . Y .) for 2 min at17°C after an average exposure period (at 4°C underlow humidity) of I-2 mo .
Developed silver grains overlying components of thevisual cells and adjoining pigment epithelium werecounted at several intervals in order to obtain a quantita-tive estimate of the distribution of radioactivity . Autora-diograms of frogs killed at I h, 15 h, I wk, and 5 wk wereanalyzed, using preparations in which lipids had beenpreserved . The lipid-extracted specimens from the 5-wkanimal were similarly assessed . Counts were made usinga grid mounted in the ocular lens of the microscope, andwere expressed as grains/grid square . Each square hadan area of 25 um' at the magnification used . Thecytoplasm, oil droplets, nuclei, and phagosomes of thepigment epithelium were analyzed, as were the outersegments (rods only), ellipsoids, myoids, nuclei, andsynaptic bodies of the visual cells . Grains over the smallcone outer segments were not counted . However, thelabeling of the oil droplets and paraboloids (Fig . 1) incone visual cells was analyzed .
Radiobiochemistry45 frogs, with an average weight of 34.7 g, were
injected intravenously with 1 .7 µCi/g of [2- 3 H]glycerol(7 .85 Ci/mmol) and were then raised under normallaboratory lighting conditions . The frogs, divided intothree groups of 15, and dark-adapted 2 h before sacri-fice, were killed at I day, 7 days, and 14 days after injec-tion . Rod outer segments were prepared by sucrose floa-tation and repeated pH 7 .1 phosphate buffer washes(15) . Two-thirds of the outer segment preparation wasextracted with chloroform : methanol (2 : 1, vol/vol),washed with 0 .2 vol of 0 .1 M NaCI solution, and the up-per, water-methanol layer was discarded. The lowerlayer was taken to dryness, redissolved in chloroform-methanol, and the content of radioactivity determined byliquid scintillation spectrometry . The remaining materialwas solubilized in 10% hexadecyltrimethylammoniumbromide (CTAB) in 0 .067 M phosphate buffer, pH 7 .1,then fractionated on a column of agarose (l3) which sep-arates visual pigment from phospholipid . The visual pig-ment fraction was identified by measurement of absorp-tion at 280 nm and 500 nm, and the content of radioactiv-ity was determined by liquid scintillation spectrometry .These analyses were carried out in the laboratory of Dr .M . O . Hall, Jules Stein Eye Institute .
RESULTS
A utoradiograph yDespite the very high levels of radioactive
glycerol administered to the animal killed 15 minafter injection, very little radioactivity had accu-
on February 11, 2018
jcb.rupress.orgD
ownloaded from

mulated in the retina during that interval . Withprolonged auto radiographic exposure it could bedetermined that the most heavily labeled retinalstructures were the paraboloids of the accessorycones (Fig . 2) . Otherwise, the most intense concen-trations of radioactivity occurred in the myoidportion of the rod and cone inner segments . Aweak, scattered labeling was discernible in theouter nuclear layer, containing the rod and conenuclei, and in the outer plexiform layer, in whichare situated the synaptic endings of the visual cellsand adjoining neurons . Labeling of the ellipsoidswas negligible, and the outer segments were notreactive . A few grains were detected over thecytoplasm of the pigment epithelium, but thenuclei and phagosomes were not labeled . Occa-sionally an oil droplet was overlaid by 1 to 3 silvergrains .
By 1 h (Figs . 3-5, Table I) the intensity of theautoradiographic reaction had increased . In thepigment epithelium, radioactive material now hadaccumulated in significant amounts in the oildroplets, which were labeled to a considerablyhigher degree than the surrounding, weakly reac-tive cytoplasm . Even fewer grains were associatedwith the nuclei, and the phagosomes remainedunlabeled. In the visual cells, the accessory coneparaboloids were highly radioactive . The myoid
portion of the inner segment contained the nexthighest concentrations of radioactive material,although the synaptic bodies were now nearly asheavily labeled, due to the marked increase inlabeling which had taken place in the outer plex-iform layer . A few grains lay over the ellipsoids,often near the periphery . At the base of a few rodouter segments the suggestion of beginning accu-mulation of radioactive material was visible. Oth-erwise, the rod and cone outer segments weredevoid of radioactivity .
4 h after injection (Figs . 6-8), labeling formerlyconfined to the visual cell myoids appeared to bespreading through the ellipsoids towards the outersegments. Although the concentration of labelingin the ellipsoid rose only slightly, the preferentialaggregation of silver grains at the base of each rodouter segment was now apparent (Fig . 8) . Other-wise, there was only a weakly scattered labeling ofthe outer segments in both rods and cones . Incones with oil droplets, a few grains were associ-ated with the perimeter of these structures, al-though the lipid within the droplets showed noevidence of radioactivity . The paraboloids ofaccessory cones continued to contain the greatestconcentration of radioactivity in the visual cells,but the accumulation of labeled material in theouter plexiform layer had continued, so that the
TABLE IConcentration of Radioactivity* in Components of the Pigment Epithelium and Visual
Cells at Different Intervals after Injection of f'H]Glvcerol
* Silver grains per unit area in autoradiograms ; I mCi/g dose ; 2-mo exposure .$ Region scleral to the band of intensely labeled disks .§ Outer nuclear layer.11 Outer plexiform layer .
C. BIBB AND R. W. YOUNG Glycerol Renewal in Outer Segment Membranes 381
Time after injection
Ih 15 h I wk 5 wk
Pigment epitheliumCytoplasm 0 .5 2 .8 3 .1 1 .3Oil droplets 2 .7 10 .0 8 .9 2 .2Nucleus 0 .4 1 .8 1 .3 0 .4Phagosomes 0 .1 0 .5 1 .9 1 .2
Visual cellsOuter segment (rods)t 0 .2 0 .5 1 .5 1 .3
Band 22 .3 19 .2Ellipsoid 1 .4 1 .3 1 .4 0 .9Myoid 5 .5 6 .5 6 .1 1 .2Nucleus§ 1 .0 1 .9 2 .0 0 .8Synaptic bodyf f 4 .6 24 .7 11 .6 5 .3Paraboloid (cones) 28 .6 30 .1 25 .0 4 .6Oil droplet (cones) 0 .4 1 .6 7 .5 2 .0
on February 11, 2018
jcb.rupress.orgD
ownloaded from

synaptic bodies of both rods and cones were nowvery heavily reactive . In contrast, the nuclei werepractically devoid of labeling . What radioactivematerial was present in the outer nuclear layerappeared to reside predominantly between thenuclei, rather than in them (Fig . 8) . In the pigmentepithelium, there had been no increase of labelingin the cytoplasm or nuclei, but the amount ofreactive material in the oil droplets continued torise (Fig . 6). Phagosomes remained unreactive .
At 15-24 h after injection (Figs . 9, 10, Table 1) itwas apparent that the gradual rise in labeling hadcontinued. The greatest relative increase had oc-curred in the synaptic bodies, which were now as
heavily labeled as the paraboloids of accessorycones. The aggregation of radioactive material atthe base of the rod outer segments was more
382
FIGURES 2-8 Pigment epithelium and visual cells from frogs injected intravenously with [2-'H]glyceroland killed between 15 min and 4 h later . Autoradiograms ; 1/2 µm Araldite sections ; lipid-preservedspecimens; toluidine blue staining .
FIGURE 2 15 min after injection . Despite a tenfold increase in dosage, and prolonged (2-mo) autoradio-graphic exposure, there was only a weak reaction in autoradiograms prepared at this interval . The greatestaccumulation of radioactivity had taken place in the accessory cone paraboloids (arrow) . Next most heavilylabeled were the myoids of the rods and cones (m) . x 800 .
FIGURE 3 1 h after injection . The intense concentration of labeled material in the myoid portion of the rodand cone inner segments is readily apparent. The darkly stained, round structures (arrow) are cone oildroplets . Note the relative absence of significant labeling in the cytoplasm (c) of the pigment epithelium . x800 .
FIGURE 4 1 h after injection . The paraboloids of accessory cones (arrows) are highly radioactive,presumably due to the incorporation of the labeled precursor into glycogen . Heavy labeling of the myoid (m)is believed to be a result of the utilization of ['H]glycerol in the synthesis of protein and phospholipid . e,ellipsoid . Radioactive material has begun to aggregate in the outer plexiform layer (opl), which contains thesynaptic bodies of the visual cells . x 1,250 .
FIGURE 5 1 h after injection . Although there is relatively little incorporation of ['H]glycerol into thecytoplasm (c) and nuclei (n) of the pigment epithelium, there is appreciable deposition of the precursor in theoil droplets (arrows) . x 1,250 .
FIGURE 6 4 h after injection . In the pigment epithelium the oil droplets (arrow) continue to containradioactive material, but the phagosomes (p) are not labeled . x 1,250 .
FIGURE 7 4 h after injection . There has been a relative and absolute increase in labeling of the outerplexiform layer (opl) . The myoids (m) of the visual cells remain heavily reactive . Between these two layers,the nuclei of the rods and cones are practically devoid of radioactivity, although some exposed silver grainsare associated with the nuclear peripheries or internuclear material . Labeled molecules have begun toaccumulate at the base of the rod outer segments (arrow) . (See Fig . 8) . Otherwise, there is only a weakscattering of silver grains over the outer segments . In the pigment epithelium, only the oil droplets (od) arewell labeled . x 800.
FIGURE 8 4 h after injection . Note the heavy labeling of the visual cell myoids (m) and the outer plexiformlayer (opl) . The intervening nuclei have not used much of the injected ['H]glycerol . Labeled material,evidently including both protein and phospholipid, is aggregating at the base of each rod outer segment(arrow), presumably having been transported from the myoid region . This aggregation is due to theassembly of new membranes . x 1,250.
pronounced, and a weak, diffuse reaction was nowmore clearly visible over the rod and cone outersegments . Most of the cone oil droplets showedweak labeling. The grains formerly situated attheir perimeter were no longer present . The myoidsin both rods and cones were still prominently
labeled, but the nuclear reaction was very weak,and largely restricted to the sparse, perinuclearcytoplasm . Labeling of the pigment epithelium hadalso increased slightly . Highest concentrationsoccurred in the oil droplets . A few grains were nowobserved over some of the phagosomes .
In the frog killed I wk after injection of['H]glycerol (Fig . 11, Table I), the synaptic bodies
of the visual cells and the paraboloids of accessorycones continued to be the most heavily labeled ret-
inal structures, although both had declined in
THE JOURNAL OF CELL BIOLOGY . VOLUME 62, 1974
on February 11, 2018
jcb.rupress.orgD
ownloaded from

C. BIBB AND R . W. YOUNG Glycerol Renewal in Outer Segment Membranes
383
on February 11, 2018
jcb.rupress.orgD
ownloaded from

labeling intensity . The decrease in radioactivitywas most pronounced in the region of the synapticbodies, which now contained less than half of thelabeled material which had been present at 15-24h . In contrast, labeling of the cone oil droplets hadnoticeably increased . The diffuse labeling in rodand cone outer segments and in phagosomes withinthe pigment epithelium also had increased . In rods,the discrete, intense band of radioactive material,which had been located near the base of each outersegment at I day, was now slightly displaced fromthe base .
5 wk after injection, this band of labeled mate-rial was located near the apical ends of the rodouter segments (Fig . 12). There was a weak,diffuse labeling on the outer (scleral) side of theband and a slightly heavier labeling on the inner(vitreal) side, within the outer segment . Radioac-tive material had been lost from all other constitu-ents of the visual cells and pigment epithelium to
384
such a considerable extent that this group ofpreferentially radioactive disk membranes wasnow by far the most intensely reactive componentin the retina . Labeling of the synaptic bodies,myoids, and cone paraboloids, as well as that ofthe oil droplets in the cones and the pigmentepithelium, had declined to about 20% of its peakintensity .
By 8 wk, the intense reaction band, formerlypresent in the rod outer segments, had disap-peared. Only a weak, random labeling could bediscerned in association with the visual cells andpigment epithelium, except that a few phagosomesshowed evidence of significant residual content ofradioactivity .
Extraction of the tissue specimens with chloro-form-methanol solution before fixation with os-mium tetroxide removed practically all of theradioactivity from the oil droplets in cones and inthe pigment epithelium . Grain counts in the case of
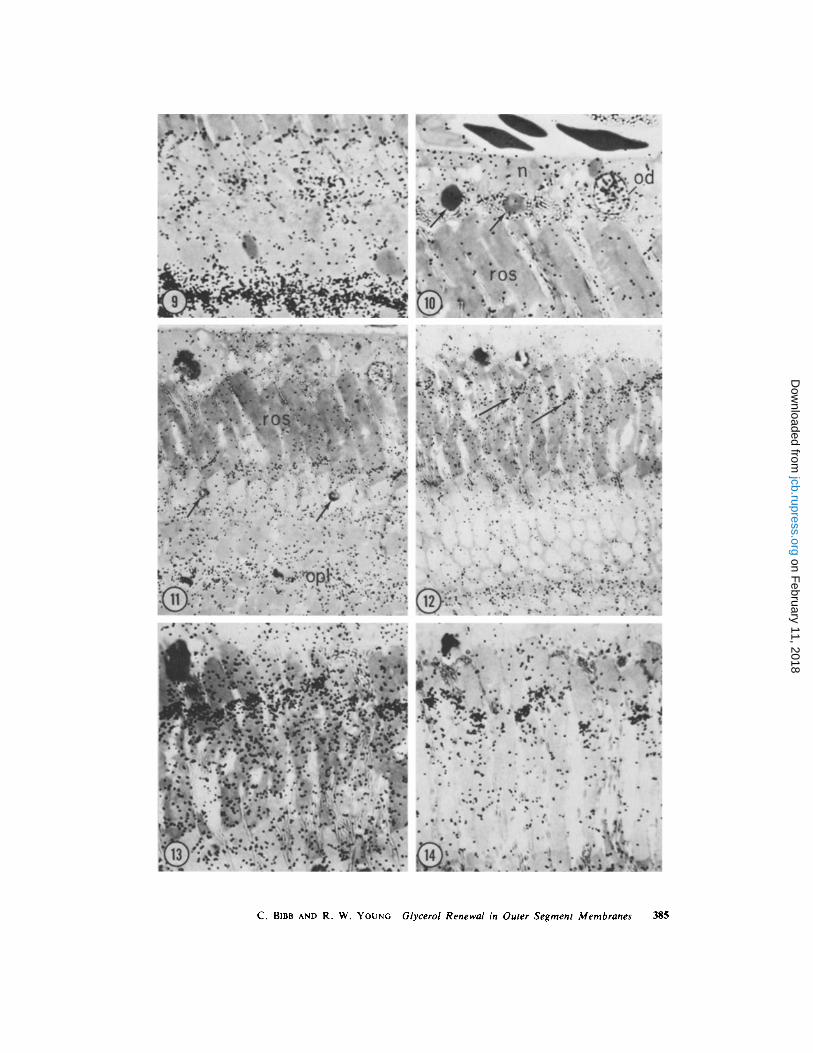
FIGURES 9-14 Pigment epithelium and visual cells from frogs injected with [2- 3 H]glycerol and killedbetween 15 h and 5 wk later. Lipid-preserved specimens, except for Fig . 14, which is lipid-extracted .Autoradiograms ; 1/z µm Araldite sections ; toluidine blue staining .
FIGURE 9 15 h after injection . The accumulation of radioactivity at the base of each rod outer segment andin the outer plexiform layer has increased . x 1,250.
FIGURE 10 15 h after injection . In the pigment epithelium, the nuclei (n) contain very little radioactivity,the cytoplasm is diffusely and rather lightly labeled, but the oil droplets (od) are intensely reactive . A weaklabeling of the rod outer segments (ros) and phagosomes (arrows) is now apparent . x 1,250 .
FIGURE I 1 I wk after injection . Labeling in the outer plexiform layer (opl) has declined . In contrast, thediffuse reaction over the rod outer segments (ros) has perceptibly increased . The highly radioactive diskmembranes, assembled at the base of the rod outer segments during the first hours after the injection, havenow been displaced from the base due to the repeated assembly of newer membranes . Note the grainsoverlying the cone oil droplets (arrows), which attain their peak labeling at about this time . x 800 .
FIGURE 12 5 wk after injection . Labeling in all structures has declined, except for the intensely radioactivemembranes in the rod outer segments (arrows), which are now the most heavily labeled components in theretina . Observe that they have been displaced more than half-way along the outer segments . x 800 .
FIGURE 13 5 wk after injection . This figure, when compared with Fig . 14, demonstrates the loss ofradioactive material which resulted from extraction of tissue specimens with chloroform-methanol solution .With maximal retention of lipid shown here, the outer segments are filled with radioactive material, which isparticularly concentrated in a few disk membranes which are situated in the outer half of the outer segment .The radioactive molecules which are located in the outermost portion of the outer segments (above the mostheavily labeled disks) have infiltrated membranes which had been assembled before the injection of[ 3 H]glycerol in the process of renewal by molecular replacement . 3-mo exposure, x 1,250 .
FIGURE 14 5 wk after injection . This autoradiogram was prepared from the same retina which yielded thespecimen shown in Fig. 13, except that the retinal fragment was extracted with chloroform : methanol (2 :1,vol/vol) before fixation in osmium tetroxide . Most of the radioactivity situated in the outermost portion ofthe rod outer segments has been extracted, demonstrating that it was in lipid . There has also been loss ofmaterial from the rest of the outer segment, with the least decrease occurring in the intensely labeledmembranes. 3-mo exposure, x 1,250 .
THE JOURNAL OF CELL BIOLOGY • VOLUME 62, 1974
on February 11, 2018
jcb.rupress.orgD
ownloaded from
C. BIBB AND R. W. YOUNG Glycerol Renewal in Outer Segment Membranes
385
on February 11, 2018
jcb.rupress.orgD
ownloaded from

the animal killed 5 wk after injection (Figs . 13, 14)showed a decrease of 93-95% in oil droplet label-ing . No other structure showed a greater loss . Theleast amount of radioactivity was removed fromthe discrete band of labeled material which movedalong the outer segments of rods between 4 h and 5wk after injection . At 5 wk, quantitative analysisshowed a loss of only 25% of the labeling from thisband as a result of extraction with chloroform-methanol solution . This extraction procedure re-moved about half of the radioactivity from othercompartments of the visual cells and pigmentepithelium (range, 36-67%). Labeling on the outer(scleral) side of the rod outer segment reactionband was reduced more than half (about 60%) . Onthe inner (vitreal) side, it was reduced by about54% .
Radiobiochemistry
Part of the purified rod outer segment prepara-tion was extracted with chloroform-methanol solu-tion to determine the amount of radioactivity inlipid . The remainder was solubilized in CTABsolution, then fractionated to separate the visualpigment (which consists of the protein, opsin,combined with vitamin A aldehyde) . The totaldpm/eye in the chloroform-methanol extract ex-pressed as a ratio of the total dpm/eye in purifiedvisual pigment at 1, 7, and 14 days after injectionwas 3.6, 3 .9, and 4 .8, respectively . This indicatesthat approximately four times as much glycerolwas incorporated into lipid as into visual pigmentprotein in the rod outer segment membranes .
DISCUSSION
Glycerol, a small, water-soluble molecule, wasadministered to frogs in radioactive form byintravenous or intralymphatic injection . The visualcells and pigment epithelium incorporated someof the tritiated glycerol which had passed from theblood stream into the retina and used it in thesynthesis of products which were thereby renderedradioactive. Prolonged initial fixation in aqueousformaldehyde solution and subsequent bufferrinses assured the extraction of unbound precursormolecules, so that residual tissue radioactivitycould be ascribed to the larger product moleculesretained by the aldehyde-osmium fixation proce-dure and the abbreviated dehydration-embeddingsequence which we used to assure maximal reten-tion of lipids .
One of our primary goals was to determine
386 THE JOURNAL OF CELL BIOLOGY • VOLUME 62, 1974
where phospholipids are produced in visual cells .Glycerol constitutes the structural backbone ofphospholipids (excluding sphingomyelin), whichare the major lipid constituents of the visual cellouter segment membranes . That glycerol wasactually used by the cells in producing outersegment phospholipids was shown by purifying rodouter segments from [ 3 H]glycerol-injected frogs,extracting the lipids from them, and then assayingthe content of radioactivity in the extract . The vastmajority of the radioactivity in these membraneswas indeed in the lipid fraction . We furtherdemonstrated the utilization of the injected glyc-erol in lipid synthesis by extracting samples oftissue with chloroform : methanol (2 : 1) solution .This solvent, which extracts lipids, removed asignificant part of the radioactivity from theretina . Having demonstrated that the glycerol wasused in the synthesis of lipids (probably almostentirely phospholipids), it remained to determinein what part of the cell the labeled precursormolecules were initially concentrated, because thesite at which labeled product molecules are firstdetected may be presumed to be the site at whichthey are synthesized . 15 min after injection, onlyone region of all the rod and cone visual cellsshowed an accumulation of radioactive ma-terial-the myoid portion of the inner segment .We may therefore tentatively conclude that it is inthe myoid region that phospholipids are produced .
Further support for this conclusion is providedby biochemical studies on the retina (27) and onother cell systems, particularly rat liver, whichhave revealed that the enzymes involved in phos-pholipid synthesis are localized in the "mi-crosome" fraction of cells (7, 10, 19, 28) . Thisfraction contains the endoplasmic reticulum, ribo-somes, and Golgi complex-all of which aresituated in the myoid region of visual cells . Auto-radiographic studies are also consistent with thisconclusion. These have indicated that fatty acidsare esterified with glycerol in the endoplasmicreticulum of mouse mammary gland (24), rat heartmuscle (25), and rat liver cells (22, 23), and that thephospholipids, phosphatidyl choline and phospha-tidyl ethanolamine, are synthesized mainly in theendoplasmic reticulum of rat liver cells (26) .
In summary, available evidence indicates thatthe incorporation of [ 3H]glycerol into the myoidportion of visual cells reflects the synthesis ofphospholipids. However, some of the labeled pre-cursor was used in protein synthesis, which also islocalized in the myoid portion of the cell (29, 39) .
on February 11, 2018
jcb.rupress.orgD
ownloaded from

Most of the glycerol which came to reside in therod outer segment membranes was recovered inlipids (which are predominantly phospholip-ids : 11), but a significant labeling of visual pigmentprotein was also recorded . Furthermore, on theaverage only half of the radioactivity was removedfrom most components of the visual cells andpigment epithelium by extraction with a lipidsolvent before autoradiography, indicating a sig-nificant utilization of the [ 3 H]glycerol in nonlipidsynthetic pathways. Labeling of cone paraboloids,which are glycogen deposits, further implies thatsome of the glycerol was used in glucose forma-tion . Because there is some glycogen in the myoidof frog visual cells (31), and because glucosamineis incorporated into the myoid region (4), a smallportion of the myoid labeling may also be due tocarbohydrate metabolism . It is evident that themyoid region is the visual cell's major syntheticcenter and appears capable of producing all of themajor constituents of the outer segment mem-branes .
Excess carbohydrate is stored in the retina asglycogen, particularly in avascular retinas such asthat of the frog (12, 17, 31) . In frog visual cells, themost prominent storage site is the paraboloid ofaccessory cones. However, glycogen is also abun-dant in the outer plexiform layer, where much of itis apparently located in the rod and cone synapticbodies (although some may be in Müller fibers) .After injection of [ 3 H]glycerol, the paraboloidswere already heavily labeled within 15 min. Evi-dently the synthesis of glycogen occurs in situ; thatis, in the region where it is stored . Much of theintense buildup of labeled material in the outerplexiform layer was probably due also to glycogenproduction .
The paraboloids had practically reached peaklabeling within I h, but the greatest content ofradioactivity in the outer plexiform layer was notattained until 15-24 h after injection . Further-more, labeling in this region dropped off rapidly .By I wk the concentration of radioactivity in theouter plexiform layer had declined to less than halfof that in the paraboloids, whereas at 15 h theywere at a similar level . This suggests a differentmetabolic function for the labeled material in thesetwo zones. This is also revealed by their differentresponses to temperature . A drop in temperatureincreases glycogen in the plexiform layers, butdiminishes it in the cone paraboloids ; with highertemperatures, the situation is reversed (18, 31) .
The rise in labeling of cone oil droplets was even
more gradual, first becoming significant at 15 h,and reaching a peak (among the intervals sampled)1 wk after injection . These colorless lipid droplets,of unknown function, apparently do not containprotein (39), but gradually become heavily la-beled after administration of fatty acids . Peaklabeling with fatty acids, as in the case of glycerol,is reached about a week after injection (2).
Neither the nuclei nor the ellipsoids of the visualcells were prominently labeled at any intervalexamined. A possible role for the nuclear mem-brane in phospholipid synthesis (8) cannot beexcluded, because the scattering of grains in theouter nuclear layer was heaviest between theclosely packed nuclei-but whether this was due tolabeling of the nuclear membrane, the thin rim ofperinuclear cytoplasm, or the slender interveningextensions of the Müller cells, we could not deter-mine . Nuclei in the pigment epithelium also failedto make significant use of the injected glycerol .Lack of glycerol uptake in nuclei has also beenobserved in liver cells (23) . Similarly, nuclei fail toincorporate appreciable quantities of fatty acids (2,23), indicating that these organelles do not partici-pate directly in a major way in lipid metabolism .
The outer segments of rods and cones failed totake up [ 3 H]glycerol from the tissue fluid, as didthe detached rod membranes present in the cyto-plasm of the pigment epithelium (phagosomes) .This is direct evidence that these membranes areincapable of synthesizing their own lipid constitu-ents, a conclusion supported by in vitro radiobio-chemical studies (27) . Nevertheless, after a delayof 1-4 h, radioactive material began to appear inthe outer segments of both classes of visual cells .As noted earlier, the source of these molecules isevidently the myoid region of the cell inner seg-ment. In rods, much of the labeled materialaggregated at the base of the outer segments, asign that new lipid (and protein) was being used inthe assembly of outer segment membranes . Thedisplacement of these new, heavily labeled mem-branes along the rod outer segments, due torepeated formation of newer disks (29), was ob-served at I and 5 wk after injection . By 8 wk thesehighly radioactive membranes were no longerpresent in the outer segments . They had been shedfrom the end of the cell and phagocytized by thepigment epithelium (37), within which a few well-labeled phagosomes remained as evidence of thisscavenging process .
Our failure to detect a small group of preferen-tially labeled membranes in cone outer segments at
C . BIBB AND R . W . YOUNG Glycerol Renewal in Outer Segment Membranes
387
on February 11, 2018
jcb.rupress.orgD
ownloaded from

any interval after injection is consistent with allprevious auto radiographic studies of this class ofvisual cells . Cones do not appear to form newmembranes and shed old ones in the same manneras rods (32, 35) .
Radiobiochemical assay indicated that approxi-mately four times as much glycerol was incorpo-rated into rod outer segment lipid as into visualpigment protein (which constitutes at least 80% oftotal protein in rod outer segments : 13, 14, 21) .
Autoradiograms revealed that approximately60% of the diffuse outer segment labeling wasextractable with a lipid solvent . The 60% loss isbased upon comparison with the "lipid-preserved"tissues, in which there is actually some loss oflipids-probably on the order of 10-15% (23) .These measurements indicate that most of thediffusely distributed rod outer segment labeling isdue to lipid .
In contrast, only about 25% of the radioactivitywas extracted (in autoradiograms) from the "reac-tion band," representing the heavily labeled newmembranes which were assembled in the first fewhours after the injection of radioactive glycerol . Inother words, more than half of the radioactivitydiffusely distributed among membranes which hadbeen assembled before the injection was in lipid,but only one-fourth of the label in disks assembledimmediately after the injection was in lipid. Thismeans that-compared to protein-a greater pro-portion of new lipid is used for molecular replace-ment, and a lesser proportion is used for mem-brane assembly .' However, we presume that a fullcomplement of lipid, as well as protein, must besupplied for the assembly of the new membranes .Therefore, renewal by molecular replacement isapparently more rapid for lipid than it is forprotein, a conclusion we also derived from ourstudies of fatty acid renewal (2) . In fact, there isevidence that visual pigment protein is not re-placed at all, once it has been inserted into themembrane (5, 13) . The rate of molecular replace-ment is even more rapid for fatty acids than for theintact phospholipid molecules, because they arereplaced by fatty acid exchange as well as bymolecular replacement of phospholipids (2) .
In the pigment epithelium, there was initiallyvery little incorporation of [ 3 H]glycerol, although
' Because some of the glycerol was apparently convertedto glucose, it is possible that the minor carbohydrateconstituent of visual pigment (16) may also have con-tained a small proportion of the nonlipid radioactivity .
388 THE JOURNAL OF CELL BIOLOGY - VOLUME 62, 1974
by I h it was apparent that radioactive materialwas gradually accumulating in the oil droplets .This was the only organelle in the pigment epithe-lium which attained significant labeling . The oildroplets also contain radioactive material afterinjection into frogs of tritiated galactose, vitaminA (38), and fatty acids (2) . The presence of aglycerol derivative and fatty acids in the oildroplets (which are completely extracted withchloroform-methanol) suggests that they may con-tain triglycerides and phospholipids as well asvitamin A esterified with fatty acid .
The technical assistance of Ms . Mirdza Lasmanis andMs . Zoja Trirogoff is gratefully acknowledged . Drs .Dean Bok and Michael Hall offered helpful suggestionsconcerning the manuscript .
This research was supported by United States PublicHealth Service Grants EY 00095 and EY 00444 .Received for publication 28 December 1973 .
REFERENCES
1 . ANDERSON, R . E ., and M . B. MAUDE . 1970 . Phos-pholipids of bovine rod outer segments . Biochemis-try 9:3624 .
2 . BIBB, C ., and R . W. YOUNG. 1974 . Renewal of fattyacids in the membranes of visual cell outer segments .J. Cell Biol . 61 :327 .
3 . Box, D . 1970 . The distribution and renewal of RNAin retinal rods . Invest . Ophthalmol. 9 :516 .
4 . BOK, D., S . F . BASINGER, and M. O. HALL . 1974 .Autoradiographic and radiobiochemical studies onthe incorporation of [6-'H]glucosamine into frogrhodopsin . Exp. Eye Res . 18.
5. BOK, D ., and R . W . YOUNG. 1972 . The renewal ofdiffusely distributed protein in the outer segments ofrods and cones . Vision Res . 12 :161 .
6 . BORGGREVEN, J . M . P . M ., F . J . M. DAEMEN, and S .L. BONTING . 1970 . Biochemical aspects of the visualprocess . V1. The lipid composition of native andhexane-extracted cattle rod outer segments . Bio-chim . Biophys . Acta . 202:374 .
7 . CARTER, J . R ., and E . P . KENNEDY . 1966 . Enzymaticsynthesis of cytidine diphosphate diglyceride . J .Lipid Res . 7 :678 .
8 . CHLAPOWSKI, F . J ., and R . N . BAND . 1971 . Assem-bly of lipids into membranes in AcanthamoebaPalestinensis . 11 . The origin and fate of glycerol-'H-labeled phospholipids of cellular membranes . J . CellBiol . 50:634 .
9 . DROz, B . 1963 . Dynamic conditions of proteins inthe visual cells of rats and mice as shown by radio-autography with labeled amino acids . Anal. Rec .145:157 .
on February 11, 2018
jcb.rupress.orgD
ownloaded from

11 . EICHBERG, J ., and H . H . HEss . 1967 . The lipidcomposition of frog retinal rod outer segments .Experientia (Basel) . 23 :993 .
12 . EICHNER, D., and H . THEMANN . 1962. Zur Frage desNetzhautglykogen beim Meerschweinchen . Z . Zell-forsch . Mikrosk . Anat . 56 :231 .
13 . HALL, M . O ., D. BOK, and A. D . E. BACHARACH .1969. Biosynthesis and assembly of the rod outersegment membrane system . Formation and fate ofvisual pigment in the frog retina . J . Mol. Biol.45 :397 .
14 . HEITZMANN, H . 1972 . Rhodopsin is the predominantprotein of rod outer segment membranes . Nat. NewBiol. 235 :114 .
15 . HELLER, J . 1968 . Structure of visual pigments . 1 .Purification, molecular weight, and composition ofbovine visual pigment 500 . Biochemistry 7:2906 .
16 . HELLER, J ., and M . A. LAWRENCE . 1970 . Structureof the glycopeptide from bovine visual pigment 500 .Biochemistry 9:864 .
17. KUWABARA, T ., and D. G. COGAN . 1961 . Retinalglycogen. Arch. Ophthalmol . 66:680.
18 . MAJIMA, K . 1924 . Studien über die Struktur derSehzellen and der Pigmentepithelzellen der Frosch-netzhaut . Albrecht von Graefes Arch. Klin . Exp .Ophthalmol. 115 :286 .
19. MCMURRAY, W . C ., and R . M. C. DAwsoN . 1969 .Phospholipid exchange reactions within the liver cell .Biochem . J . 112 :91 .
20. NIELSEN, N . C ., S . FLEISHER, and D . G .MCCONNELL . 1970 . Lipid composition of bovineretinal outer segment fragments . Biochim . Biophys .Aeta . 211 :10 .
21. ROBINSON, W . E., A. GORDON-WALKER, and D .BOWNDS. 1972 . Molecular weight of frog rhodopsin .Nat. New Biol. 235:112 .
22 . STEIN, O ., and Y . STEIN . 1966 . Electron microscopicautoradiography of [ 3 H]glycerol labeled lipid inethanol induced fatty liver . Exp. Cell Res . 42 :198 .
23 . STEIN, O., and Y. STEIN . 1967 . Lipid synthesis,intracellular transport, storage, and secretion . 1 .Electron microscopic radioautographic study ofliver after injection of tritiated palmitate or glycerolin fasted and ethanol-treated rats . J . Cell Biol .33:319.
24 . STEIN, O., and Y. STEIN . 1967 . Lipid synthesis,intracellular transport, storage, and secretion . 11 .Electron microscopic radioautographic study of the
microscopic radioautographic study of the rat heartperfused with tritiated oleic acid . J. Cell Biol . 36 :63 .
26. STEIN, O., and Y . STEIN . 1969 . Lecithin synthesis,intracellular transport, and secretion in rat liver . IV .A radioautographic and biochemical study of cho-line-deficient rats injected with choline- 3H . J . CellBiol . 40 :461 .
27 . SWARTZ, J . G ., and J . E . MITCHELL . 1970 . Biosyn-thesis of retinal phospholipids : incorporation ofradioactivity from labeled phosphorylcholine andcytidine diphosphate choline . J. Lipid Res . 11 :544.
28. WILGRAM, G . F ., and E . P. KENNEDY . 1963 . Intra-cellular distribution of some enzymes catalyzingreactions in the biosynthesis of complex lipids . J .Biol . Chem . 238:2615 .
29. YOUNG, R . W. 1967 . The renewal of photoreceptorcell outer segments . J . Cell Biol . 33 :61 .
30. YOUNG, R. W. 1968 . Passage of newly formedprotein through the connecting cilium of retinal rodsin the frog . J. Ultrastruct . Res . 23 :462 .
31. YOUNG, R . W. 1969 . The organization of vertebratephotoreceptor cells. In The Retina . B . R . Straatsma,R . A . Allen, F . Crescitelli, and M . O . Hall, editors .University of California Press, Berkeley, Calif. 177 .
32 . YOUNG, R. W . 1969 . A difference between rods andcones in the renewal of outer segment protein .Invest . Ophthalmol . 8 :222 .
33 . YOUNG, R . W . 1971 . The renewal of rod and coneouter segments in the rhesus monkey . J . Cell Biol.49 :303 .
34 . YOUNG, R. W . 1971 . Shedding of discs from rodouter segments in the rhesus monkey . J . Ultrastruct .Res . 34 :190 .
35 . YOUNG, R . W . 1971 . An hypothesis to account for abasic distinction between rods and cones . VisionRes . 11 :1 .
36 . YOUNG, R. W . 1974 . Biogenesis and renewal ofvisual cell outer segment membranes . Exp. Eye Res .18 .
37. YOUNG, R . W ., and D. BoK . 1969 . Participation ofthe retinal pigment epithelium in the rod outersegment renewal process . J. Cell Biol . 42 :392 .
38. YOUNG, R. W., and D. BOK. 1970 . Autoradio-graphic studies on the metabolism of the retinalpigment epithelium . Invest . Ophthalmol . 9 :524 .
39. YOUNG, R . W., and B . DROZ . 1968 . The renewal ofprotein in retinal rods and cones . J . Cell Biol .39 :169 .
C. BIBB AND R. W. YOUNG Glycerol Renewal in Outer Segment Membranes 389
10 . EIBL, H ., E . E. HILL, and W . E. M. LANDS . 1969 . mouse lactating mammary gland . J . Cell Biol.The subcellular distribution of acyltransferases 34 :251 .which catalyze the synthesis of phosphoglycerides .Eur. J. Biochem . 9 :250 .
25. STEIN, O., and Y. STEIN . 1968 . Lipid synthesis,intracellular transport, and storage . Ill . Electron
on February 11, 2018
jcb.rupress.orgD
ownloaded from


















