
Remanentes de bosque de roble en la vereda Capilla … de Pregrado.pdfinsectos asociados a hongos...
87
Transcript of Remanentes de bosque de roble en la vereda Capilla … de Pregrado.pdfinsectos asociados a hongos...


Remanentes de bosque de roble en la vereda Capilla (Villa de Leyva - Boyacá).

INSECTOS ASOCIADOS A HONGOS MACROMICETOS (BASIDIOMYCOTA) EN
ROBLEDALES DE LA REGION DE IGUAQUE (VILLA DE LEYVA-BOYACA-COLOMBIA)
EDUARDO CARLO AMAT GARCIA
GERMAN D AMAT G PROFESOR ASOCIADO,
INSTITUTO DE CIENCIAS NATURALES UNIVERSIDAD NACIONAL DE COLOMBIA
DIRECTOR
MARIBEL PINZON BIOLOGA
PROFESORA CATEDRA PONTIFICIA UNIVERSIDAD JAVERIANA
CO-DIRECTORA
TRABAJO DE GRADO Presentado como requisito parcial
para optar al título de
BIÓLOGO
FACULTAD DE CIENCIAS CARRERA DE BIOLOGIA
PONTIFICIA UNIVERSIDAD JAVERIANA Bogotá, D.C. Junio de 2002

NOTA DE ADVERTENCIA Artículo 23 de la Resolución No13 de julio de 1946: "La Universidad no se hace responsable por los conceptos emitidos por sus alumnos en sus trabajos de de grado".

INSECTOS ASOCIADOS A HONGOS MACROMICETOS (BASIDIOMYCOTA)
EN ROBLEDALES DE LA REGION DE IGUAQUE
(VILLA DE LEYVA-BOYACA-COLOMBIA)
EDUARDO CARLO AMAT GARCIA
APROBADO:
________________________________ _________________________________ GERMAN AMAT MARIBEL PINZON Biólogo Msc. Bióloga PROFESOR ASOCIADO PROFESORA CATEDRA UNIVERSIDAD JAVERIANA INSTITUTO DE CIENCIAS NATURALES PONTIFICIA UNIVERSIDAD JAVERIANA UNIERSIDAD NACIONAL DE COLOMBIA CO-DIRECTORA DIRECTOR
________________________________ _________________________________ CARLOS RIVERA R RODULFO OSPINA
DEPARTAMENTO DE BIOLOGÍA DEPARTAMENTO DE BIOLOGÍA FACULTAD DE CIENCIAS FACULTAD DE CIENCIAS
PONTIFICIA UNIVERSIDAD JAVERIANA UNIVERSIDAD NACIONAL DE COLOMBIA JURADO JURADO

INSECTOS ASOCIADOS A HONGOS MACROMICETOS (BASIDIOMYCOTA)
EN ROBLEDALES DE LA REGION DE IGUAQUE
(VILLA DE LEYVA-BOYACA-COLOMBIA)
EDUARDO CARLO AMAT GARCIA
APROBADO:
_________________________________ __________________________________
ANGELA UMAÑA LUZ MERCEDES SANTAMARIA DECANO ACADEMICO DIRECTOR DE CARRERA

Este trabajo esta especialmente dedicado a Cecilia, Domingo, German y Zoila.
a ellos gracias por brindarme lo mejor de sus corazones.

AGRADECIMIENTOS
Deseo agradecer a todas las personas e instituciones que hicieron posible la realización de este trabajo
- Dr. German Amat y a Luis Guillermo Henao por la valiosa dirección y asesoria de este trabajo. Además por haberme dado la oportunidad de explorar sobre la ecología de estos dos grandes taxones.
- A Maribel Pinzon por su colaboración y la co-dirección de este estudio. - Instituto Alexander Von Humboldt (en especial Fernando Gast y Yaneth Muñoz) por
haberme permitido desarrollar la parte de laboratorio en sus instalaciones en Villa de Leyva, y la utilización de sus equipos.
- A todas las personas de Entomología del Instituto Humboldt encabezadas por el
Profesor Fernando Fernández, Diego Campos, y asistentes de laboratorio por haberme permitido aprender acerca del oficio entomológico, además de su ayuda.
- Al Programa Nacional de Transferencia de Tecnología Agropecuaria (Pronatta) del
Ministerio de Agricultura, por la financiación del proyecto en el cual esta enmarcado este trabajo de grado.
- A Santiago Grillo por su valiosa colaboración en el análisis estadístico, a Angélica
Ruiz por la ayuda en la determinación de muestras fúngicas y a Andrea Olaya por su ayuda durante todas las fases del estudio.
- A doña Vitelvina Espitia, Jailer y sus hermanitas, a don Germán Borras por su especial
cortesía durante la fase de campo en la vereda Capilla. - A mi familia por su apoyo incondicional en todas las etapas de este trabajo
- A todos los amigos y conocidos que de una u otra forma estuvieron directamente
relacionados con este trabajo.

vi
TABLA DE CONTENIDOS Pág. 1. INTRODUCCION………………………………….……………………………………….. 10
2. MARCO TEORICO………………………..……………………………………………….. 12
2.1 Interacción hongo-insecto……...……………………………………………….............. 12
2.2 El hongo como microhábitat……………………………………………………………. 14
2.3 Estrategia de fungivoria de los insectos………………..………………………………. 15
2.4 Dimensión temporal de la interacción………………………………………………….. 16
2.5 Dimensión espacial de la interacción………..………………………………………….. 17
3. ANTECEDENTES………………………………………………………………………….. 17
4. JUSTIFICACION…………………………………………………………………………… 18
5. OBJETIVOS…………………………………………………………………………………. 20
5.1 Objetivo General……………………………………………………………………………. 20
5.2 Objetivos específicos............................................................................................................... 20
6. MATERIALES Y METODOS……………………………………………………………... 21
6.1 Área de estudio………………………………………………………………………….. 21
6.1.1 Localización………………………………………………………………………. 21
6.1.2 Clima y precipitación……………………………………………………………... 22
6.1.3 Vegetación……………………………………………………………………….. 23
6.2 Fase de Campo………………………………………………………………………….. 24
6.2.1 Colecta de hongos…………………………………………………….…………... 24
6.3 Fase de Laboratorio…………………………………………………….……………….. 24
6.3.1 Determinación taxonómica de insectos y hongos colectados…………………….. 26
6.3.2 Cría de insectos…………………………………………………………………… 27
6.4 Análisis de la información………………………………………………………………. 28
6.4.1 Relación especies-abundancia…………………………………………………..... 28
6.4.2 Riqueza esperada…………………………………………………........................ 29
6.4.3 Indices de riqueza, diversidad y dominancia……………………………………... 30
6.4.4 Relación precipitación-hongos-diversidad de insectos…………………………… 31
6.4.5 Selección de hospederos………………………………………………………….. 31
6.4.6 Amplitud de la dieta alimenticia …………………………………………………. 32
6.4.7 Categorías Tróficas……………………………………………………………….. 32
6.4.8 Relación disponibilidad de recurso-diversidad de insectos……………................ 33
6.4.9 Relación área-especies……………………………………………………………. 33

vii
6.4.10 Relación estados de desarrollo-diversidad de insectos…………….……………. 34
6.4.11 Patrones de sucesión de la entomofauna...……...……………………………... 34
7. RESULTADOS……………………………………………………………………………… 35
7.1 Caracterización de los insectos asociados………………………………………………. 35
7.1.1 Abundancia……………………………………………………………………….. 35
7.1.2 Relación especies-abundancia……………………………………………………. 35
7.1.3 Riqueza esperada………………………………………………............................. 39
7.1.4 Categorías tróficas………………………………………………………………... 42
7.2 Caracterización de los hongos hospedantes……...…………………………………….. 44
7.2.1 Abundancia de hongos macromicetos…….……………………………………… 44
7.2.2 Relación especies-abundancias……………………………………….................. 46
7.2.3 Riqueza esperada…………………………………………………….................... 47
7.2.4 Relación precipitación-hongos-insectos………………………………………….. 48
7.3 Interacción insecto-hongo………………………………………………………………. 49
7.3.1 selección de hospederos…………………………………………………………... 49
7.3.2 Amplitud de dieta………………………………………………………................ 51
7.3.3 Relación disponibilidad del recurso-diversidad de insectos……………………… 51
7.3.4 Relación área-especies……………………………………………………………. 52
7.3.5 Relación estados de desarrollo-diversidad de insectos…………………………… 54
7.3.6 patrones de sucesión de la entomofauna………………………………………… 55
8. DISCUSION…………………………………………………………………………………. 57
8.1 Ensamblaje de la comunidad de insectos asociados a hongos..………………………… 57
8.2 Ensamblaje de la comunidad de hongos hospedantes de insectos..…………………….. 58
8.3 Modelos espacio-temporales resultantes de la interacción insecto-hongo……………… 59
8.4 modelo trófico resultante de la interacción insecto-hongo……………………………… 60
9. CONCLUSIONES…………………………………………………………………………... 64
10. RECOMENDACIONES…………………………………………………………………… 65
11. REFERENCIAS……………………………………………………………………………. 66
12. ANEXOS………………………………………………………………………….............. 72

Insectos asociados a hongos macromicetos en robledales de Iguaque
10
1. INTRODUCCION
Hongos e insectos hacen parte del grupo de organismos megadiversos del planeta (Cowell
& Coddington 1994) y entre ellos existe una variada gama de relaciones, desde los hongos
entomopatogenos cuya dependencia es obligatoria por insectos como recurso alimentario,
hasta la estricta dependencia de los insectos por hongos con el mismo fin (Wilding 1989).
El consumo de alguna estructura fúngica como micelio, cuerpos fructíferos y esporas por
un insecto se define como micofágia (Lawrence 1989), La micofágia es la principal
relación que se establece entre insectos y carpóforos de macromicetos, también es la
encargada de moldear las características de la interacción entre estos dos grandes taxones,
en carpóforos especialmente del phyllum Basidiomycotina se ha caracterizado una fauna
particular de insectos micófagos principalmente representada por los ordenes Díptera y
Coleóptera.
Algunos autores consideran este patrón de alimentación como una situación análoga con
la herbivoría. Hanski (1989) plantea similitudes y diferencias, desde el punto de vista de
los insectos entre plantas y hongos, la inmovilidad, presencia de sustancias toxicas y falta
de defensas físicas son condiciones similares en estos dos tipos de sustratos. Sin embargo
la escasa duración temporal y los impredecibles patrones espaciales en los hongos hacen
que los insectos Micófagos se vean enfrentados a problemas muy diferentes, estas
condiciones afectan la ecología y evolución de los insectos asociados (Ashe 1987, Leschen
1994). Se destaca las claras diferencias entre la fauna de insectos Agaricales y
Aphylloporales hongos cuya persistencia temporal y consistencia física son muy
diferentes, como característica general acerca de la selección de sus hospederos cabe
mencionar que los insectos asociados a hongos exhiben un alto grado de generalismo
(Hanski 1989).
La mayoría de trabajos se han dirigido a estudiar la diversidad de especies integrantes de
esta relación, y en algunos casos la estructura poblacional y algunas relaciones ecológicas
como competencia, predación, parasitismo (Guevara & Dirzo 1999). Si tenemos en cuenta

Insectos asociados a hongos macromicetos en robledales de Iguaque
11
que en los trópicos las comunidades biológicas son altamente ricas en especies y que el
papel de los insectos es esencial en estudios sobre biología tropical (Janzen 1970). Además
de la falta de información de esta interacción en estas regiones; se esboza aquí una de las
primeras aproximaciones al conocimiento de las comunidades de insectos asociados a
hongos en nuestro país, se consideran algunos factores bióticos y abióticos que afectan la
dinámica de esta relación, teniendo como especial recurso la micota en bosques de roble,
bosque que son altamente ricos en macromicetos especialmente agaricales (Mueller &
Halling 1995).

Insectos asociados a hongos macromicetos en robledales de Iguaque
12
2. MARCO TEORICO
Cualquiera que sea la definición de biodiversidad, este concepto incluye todas o cada una de
las jerarquías de la organización biológica (genes, individuos, poblaciones, especies,
comunidades, ecosistemas), una de las tradicionales medidas de diversidad en un contexto
ecológico es la que engloba principalmente dos componentes: variedad y abundancias
relativas de las especies (Magurran 1988). Atendiendo a estas variables, el problema se enfoca
directamente en las propiedades de la comunidad en cuestión.
Estudios de la relación insecto-hongo han centrado especial interés en la identificación de la
diversidad de insectos que componen estas asociaciones, esto se debe a que son comunidades
claramente definidas (Klimaszewski y Peck 1987). De esta forma se delimita nuestra
comunidad; como el grupo de especies de insectos que mantienen algún tipo de relación o
asociación con los cuerpos fructíferos de hongos Macromicetos.
Una de las definiciones mas aceptadas de comunidad es la que incluye la presencia de
poblaciones de especies que se encuentran juntas en espacio y tiempo (Begon 1995). Los
investigadores interesados en la ecología de comunidades centran su atención en el modo de
distribución de las agrupaciones de especies y en la forma como están influenciadas por las
interacciones entre las mismas especies o los factores abióticos que las rodea. Así el estudio de
la ecología de comunidades ofrece instrumentos teóricos y experimentales convenientes para
aproximarse al conocimiento de esta interacción ecológica.
2.1 INTERACCION HONGO-INSECTO
En este trabajo el uso de la palabra hongo hace referencia exclusivamente al estado
reproductivo de macromicetos que llevan a cabo su reproducción por medio de carpóforos o
cuerpos fructíferos.

Insectos asociados a hongos macromicetos en robledales de Iguaque
13
En los macromicetos se ha caracterizado una fauna particular de insectos, existe registros de
algún tipo de relación para 13 ordenes, entre los principales Coleóptera con 47 familias,
Díptera con 44 familias e Hymenoptera con 9 (Hammond y Lawrence 1989).
Los hongos ofrecen una variada gama de recursos aprovechada por los insectos, gracias a esta
oferta se establecen diferentes tipos de relaciones. Los insectos pueden utilizar el hongo con
fines alimenticios, reproductivos, de refugio y ocasionalmente para buscar presas.
Varios autores han designado con el nombre de Micetócolos a los insectos que mantienen una
determinada relación con hongos, y dependiendo el grado de asociación se clasifican en tres
diferentes categorías: Micetobiontes, Micetófilos y Micetóxenos (Scheerpeltz y Hofler 1948,
Graves 1960, Klimaszewki y Peck 1979), otros autores los clasifican en cuatro: fungívoros
primarios, secundarios, detritívoros y predadores (Bruns 1984, Navarrete 1997, Koomonen
2001) estos últimos separan los detritívoros y depredadores en categorías diferentes, es común
encontrar en trabajos recientes una mezcla entre estas dos categorizaciones esto se debe
principalmente a la fauna local asociada.
Micetobiontes o fungívoros primarios: Insectos cuya asociación es obligatoria y dependen de
los cuerpos fructíferos para llevar su ciclo de vida, además de lugar de abrigo y oviposición;
por lo general se encuentran en todos los estados de maduración del carpóforo. Esta relación
obligatoria no implica que sea especialista, muchos micetobiontes están relacionados con un
amplio rango de especies fúngicas (Lawrence 1973).
Micetófilos o fungívoros secundarios: Insectos cuya dependencia por lo hongos no es
absoluta, también se pueden encontrar en otros microhábitats como excremento o materia
orgánica en descomposición. Este tipo de insectos usualmente se encuentran en estados
intermedios de desarrollo del carpóforo. Se reconocen dos tipos de Micetófilos con base en sus
hábitos alimentarios depredadores o saprófagos.
Depredadores: entomófagos y parasitóides de otros insectos micetócolos.

Insectos asociados a hongos macromicetos en robledales de Iguaque
14
Saprófagos o Detritívoros: insectos que además de hongos se alimentan de materia en
descomposición.
Micetóxenos: a esta categoría pertenecen los insectos que ocasionalmente se encuentran en los
carpóforos, no es clara la relación con estos o con otros insectos micetócolos por lo general
utilizan el hongo como refugio temporal.
2.2 EL HONGO COMO MICROHÁBITAT
Los Hongos ofrecen características particulares muy variables como hábitat para insectos,
disponibilidad temporal, espacial y diferentes estados de maduración cuentan como las
propiedades más importantes (Hanski 1989).
La consistencia y persistencia del recurso permiten agrupar los hongos en dos grupos bien
definidos, los hongos carnosos de un relativo corto tiempo de vida (Ej: Agaricales) y por otra
parte los leñosos que tienen un periodo de existencia mas largo (Ej: Aphylloporales) la tabla 1
resume las principales características de los hongos como microhábitat.
Tabla 1. Principales atributos de los hongos macromicetos como microhábitats Característica Descripción
Tamaño Generalmente pequeño (Volumenes de Cm3 ).
Distribución Aleatoria, en algunas ocasiones agregada
Densidad en un parche Baja ( bajo numero de individuos por unidad de área)
Duración Corta en hongos carnosos (en Agaricales desde dias hasta meses)
Larga en hongos leñosos (en Aphylloporales desde meses hasta
años)
Esta naturaleza afecta profundamente la ecología de la fauna asociada, además de las historias
evolutivas de algunos grupos. Algunos coleópteros micófagos que llevan su ciclo de vida en
carpóforos carnosos tienen un desarrollo larval mucho mas acelerado aproximadamente de 3 a

Insectos asociados a hongos macromicetos en robledales de Iguaque
15
11 días, mientras los que se encuentran asociados a hongos persistentes que pueden tardar de
16 a 72 días, muy posiblemente esto refleja una adaptación ante la naturaleza efímera del
hábitat además se enfrentan a diferentes presiones y sus respuestas exhiben patrones bien
definidos en el uso de este recurso (Ashe 1987).
Esta característica temporal y el lugar especifico en el hongo donde los insectos se alimentan
inciden en algunos aspectos etológicos; los comportamientos anti-depredaror muestran una
clara evidencia, especies de coleópteros que se alimentan en la superficie externa del hongo
parecen tener comportamientos más elaborados que aquellas que se alimentan en el interior.
La protección de los huevos esta correlacionada con la alimentación en hongos leñosos
persistentes, al igual que la existencia de algún cuidado parental. La pupación ocurre en el
hongo sólo para especies que viven en hongos duraderos mientras que la pupación de especies
que se relacionan con hongos efímeros se lleva en el suelo (Leschen 1994).
El desarrollo del hongo desde su aparición hasta su total descomposición presenta etapas
durante las cuales el tejido fúngico sufre cambios en sus características físicas y químicas,
como también cambia su fauna asociada (Klimaszewki y Peck 1986). algunas especies de
insectos muestran una preferencia por los diferentes estados, esta tendencia depende del tipo
de relación que el insecto busca del recurso (Hanski 1989) de esta manera puede haber un
recambio de especies a manera de sucesión ecológica.
2.3 ESTRATEGIAS DE FUNGIVORIA DE LOS INSECTOS
Este patrón alimenticio se puede considerar análogo a la interacción entre insectos fitófagos y
sus plantas hospederas, aunque se reconocen algunas diferencias (Hanski 1989). El daño
ocurrido por el insecto fitófago puede ser tal, que conduzca a la búsqueda de alguna defensa
por parte de la planta, lo que muy posiblemente y a lo largo de la historia desencadene en una
relación coevolutiva (Haukioja 1980). Por otra parte el daño causado por un insecto micófago
no afecta significativamente la producción y dispersión de esporas, objetivo principal de un
cuerpo fructífero (Hackman & Meinander 1979). Por el contrario algunos dípteros en
búsqueda de lugar para la oviposición y como consecuencia del contacto pueden llegar a

Insectos asociados a hongos macromicetos en robledales de Iguaque
16
favorecer la dispersión de esporas (Hackman & Meinander 1979). Otra diferencia con las
plantas vasculares es la variabilidad y poca predicción temporal y espacial de los hongos,
estos factores se discutirán mas adelante en como patrones espacio-temporales.
De acuerdo con el habito alimenticio los insectos Micófagos están mas estrechamente
relacionados con los saprófagos que con los herbívoros (Martin et al 1981), regularmente es
mas común encontrar insectos saprófagos o coprófagos alimentándose de estructuras fúngicas
que herbívoros. Hackman y Meinander (1979) contemplan una posible tendencia evolutiva de
algunos grupos de dípteros por estos hábitos alimenticios, posiblemente algunas especies de
saprófagos ocasionalmente se alimentaban de tejido fúngico en descomposición para luego
especializare en hábitos Micófagos y por tanto exhibir marcadas tendencias por la preferencia
hacia este sustrato.
2.4 DIMENSION TEMPORAL DE LA INTERACCION
Los patrones temporales de esta relación, incluyen diferentes escalas de tiempo, desde la
duración de un hongo, que fluctua de días a semanas hasta la estacionalidad de las especies
(fenología) y la dinámica de producción de carpóforos que comprenden largos periodos de
tiempo (Hanski 1989). Cada una de estas variaciones en la disponibilidad del hongo como
recurso condiciona de alguna manera el uso por parte del insecto, los patrones espaciales a
menor escala se pueden ver reflejados en los procesos de sucesión de especies. Los de mayor
escala abarcan las fluctuaciones poblacionales de los insectos que mantienen una relación más
estrecha (Wilding et al 1989). Es en este punto donde los factores ambientales juegan un papel
decisivo en la ecología de esta asociación si tenemos en cuenta que el desarrollo de los hongos
esta condicionado por características de humedad, temperatura, calidad del suelo y hojarasca,
entre otros.

Insectos asociados a hongos macromicetos en robledales de Iguaque
17
2.5 DIMENSION ESPACIAL DE LA INTERACCION
La distribución espacial de los hongos condiciona la aparición de insectos con hábitos
micófagos; aunque su aparición es azarosa, para este caso en el que la mayoría de hongos son
simbiotróficos con algunas especies de árboles, podría asegurarse la presencia tanto de hongos
como insectos asociados a lo largo de una amplia escala temporal en lugares con
características definidas.
Una escala espacial mas reducida de esta interacción es la que corresponde a la localización de
la oviposición y por consiguiente la distribución de las larvas en el carpóforo. Algunas
especies del género Pegomya (Diptera:Anthomidae) han especializado su localización en el
estípite mientras que un numero considerablemente mayor de dípteros utilizan varias partes
del píleo, o el interior de las lamelas (Hanski 1989). Es posible que la agregación espacial de
los hongos también condicione la presencia de insectos no se tiene información acerca de este
factor sobre los insectos asociados.
La relación del tamaño del hongo y la riqueza de especies de insectos asociados no ha sido un
tema muy común en estudios sobre esta interacción, sólo se conoce una relación positiva entre
estas variables gracias al trabajo con micro artrópodos asociados a hongos de O´connel &
Bolger (1997).
3. ANTECEDENTES
Este tipo de interacción ha sido principalmente estudiada en Europa, en menor proporción se
ha llevado a cabo estudios en Norteamérica (Bruns 1984) y México.
Los primeros estudios enfocaron principalmente sus objetivos en la descripción de la fauna de
insectos asociados a hongos (Pielou & Verna 1966, Graves & Graves 1966). En 1973
Lawrence después de hacer la revisión de la familia Ciidae (grupo de coleópteros encontrados

Insectos asociados a hongos macromicetos en robledales de Iguaque
18
comúnmente en hongos leñosos) propone las posibles causas de la selección de hospederos en
esta interacción.
Uno de los estudios clásicos en esta temática fue el realizado por Hackman y Meinander
(1979) en Finlandia, donde describen la importancia de los dípteros como plagas de hongos
comestibles además de proveer valiosa información ecológica de esta relación. Estudios como
el de Klimaszewki y Peck (1987) amplían el conocimiento de esta interacción y proponen un
patrón de sucesión de insectos en hongos polyporaceos. Hanski (1989) genera la mayor
información ecológica, compila varios trabajos y expone sus ideas sobre selección de
hospederos, ecología de poblaciones, caracterización y funcionalidad de gremios de insectos
fungívoros e importancia económica de estos. En 1994 Leschen establece correlaciones entre
comportamientos de coleópteros micófagos, encuentra que los comportamientos larvales más
diversos y conspicuos son los comportamientos anti-depredador; principalmente explicados
por la persistencia del hongo.
En los últimos años la relación insecto-hongo ha centrado principal interés por encontrar
respuestas sobre los factores que modelan las comunidades como son la repartición del
recurso, agregación espacial, competencia interespecífica y organización de sus niveles
tróficos (Yacovlev 1992, Komonen 2001).
Llama la atención la ausencia de información encontrada en la literatura para esta asociación
en el trópico (Hammond y Lawrence 1989). Para Colombia Gutierrez (1998) reporta 25
familias de insectos asociados a Agaricales en el departamento de Antioquia.
4. JUSTIFICACION
Trabajos dirigidos a investigar en dinámicas de poblaciones y comunidades, en historias de
vida de insectos, competencia interespecifica, selección de hospederos, diversidad genética
evolución de componentes químicos, entre otros, pueden encontrar en este tipo de interacción
una alternativa adecuada para el estudio de estos temas (Hanski 1989).

Insectos asociados a hongos macromicetos en robledales de Iguaque
19
Nuestra situación de país megadiverso hace importante estudiar la temática ecológica, es
prácticamente desconocida la diversidad de insectos asociados a cuerpos fructiferos de
macrohongos y aun menos conocidas las dinámicas de estas comunidades, el presente trabajo
representa una de las primeras aproximaciones al conocimiento de esta interacción desde el
punto de vista de la ecología de comunidades y uno de los primeros aportes para el estudio
funcional de está en el neotrópico.
Algunos grupos de insectos pueden representar una amenaza para los cultivos comerciales de
hongos, esperando la proyección de estos como alternativa alimentaría para el hombre, en
especial en nuestro país, se abre camino hacia el conocimiento de los mecanismos y dinámicas
que rigen esta asociación y así su posible manejo.

Insectos asociados a hongos macromicetos en robledales de Iguaque
20
4. OBJETIVOS
4.1 OBJETIVO GENERAL
Caracterizar y estudiar la estructura, composición y funcionamiento de la comunidad de
insectos asociados a macromicetos en bosques de roble (Quercus spp) de la región de
Iguaque, Boyacá.
4.2 OBJETIVOS ESPECIFICOS
• Estudiar la composición, riqueza y abundancia de las especies de insectos asociados
a hongos macromicetos en bosques de Roble (Quercus spp).
• Determinar el efecto de la disponibilidad del hongo como recurso alimentario y su
relación con el ensamblaje de la comunidad de insectos micófagos.
• Determinar el efecto del estado de desarrollo del hongo sobre la riqueza y
abundancia de las especies de insectos asociados.
• Determinar el efecto del tamaño del hongo sobre la riqueza y abundancia de las
especies de insectos asociados.
• Establecer un modelo de sucesión de las especies en la comunidad de insectos
asociados a hongos macromicetos, y caracterizar sus patrones micro temporales en
la interacción.

Insectos asociados a hongos macromicetos en robledales de Iguaque
21
6. MATERIALES Y METODOS
6.1 AREA DE ESTUDIO
6.1.1 Localización
El área de estudio se localiza en los 5° 39 y 5° 42 LN y 73°20 y 73°24 LO, a una altura de
2700 m. Esta situada en la vereda Capilla, municipio de Villa de Leiva, Boyacá, Colombia
(Figura 1), limita con el sector noroccidental del Santuario de Fauna y Flora de Iguaque;
esta región hace parte de la zona de amortiguación del parque.
Figura 1. Localización del área de estudio para el reconocimiento de insectos asociados a hongos macromicetos.

Insectos asociados a hongos macromicetos en robledales de Iguaque
22
6.1.2 Clima y Precipitación
La región de Villa de Leyva presenta tres diferentes tipos de unidades climáticas: zona
seca, zona subhúmeda y zona húmeda, (Schutze 1999). El área de estudio pertenece a la
zona subhúmeda, donde predomina un régimen bimodal con dos picos de precipitación y
una marcada temporada seca (Figura 2).
0
50
100
150
200
250
Ene Feb M ar Abr M ay Jun Jul Ago Sep O ct N ov D ic
Prec
ipita
ción
-mm
Figura 2. Precipitación promedio mensual en el área de estudio (estación Arcabuco, 1982-2000. Fuente: Ideam 2001.)
El promedio anual de precipitación es de 1760 mm registrado para 19 años, las épocas de
mayor precipitación son marzo-abril y octubre-noviembre siendo el mes mas lluvioso
Noviembre (220 mm). La temporada seca corresponde a los meses de julio y agosto siendo
agosto el mes mas seco (54 mm).
La precipitación en el área; esta condicionada a las masas de nubes provenientes de la
cuenca media y baja del río Suárez y del valle del Magdalena, las cuales ascienden hasta la
región y descargan torrenciales lluvias (Schutze 1999).

Insectos asociados a hongos macromicetos en robledales de Iguaque
23
Figura 3. Régimen de lluvias en el área de estudio (Estación Arcabuco, 1999-2001, Fuentes: Ideam 2001)
En la primera temporada lluviosa de 2001 (Figura 3), época que coincide con la del
muestreo, marzo y abril no alcanzan sus valores normales de lluvia y en general el año
presenta valores muy bajos de precipitación, exhibiendo el mes más seco de los últimos 9
años (Agosto con 9 mm).
6.1.3 Vegetación
Siguiendo la clasificación de zonas de vida propuesta por Holdridge, el área corresponde a
la zona de vida Bosque Húmedo Montano Bajo (BhMB), el paisaje se compone de
formaciones vegetales de pajonales y de matorrales cuya vegetación arbustiva esta
dominada por Maclania rupestres, Escallonia murtilloides (tibar) Senecios spp,
Gaiadendrum sp y Bucquetia glutinosa. La vegetación arbórea esta dominada Clusia spp y
Roble Quercus spp.
La presencia de formaciones herbáceas y arbustivas dominadas por una cobertura de
pastizales obedece en gran parte a la acción reiterada del fuego, tala y siembra de Pino y

Insectos asociados a hongos macromicetos en robledales de Iguaque
24
eucalipto, que a través de la historia fueron desplazando los bosques nativos de Roble.
(Schutze 1999).
El área de estudio comprende pequeños remanentes de estos bosques principalmente
asociaciones Roble-Aliso (Quercus spp- Alnus spp), Roble-Encenillo (Querqus spp-
Weinmania tomentosa) y Roble- Guaques (Quercus spp-Clusia sp) localmente conocidos
como Robledales; el paisaje se mezcla con áreas de reciente ocupación y transformación en
donde las principales actividades económicas son los cultivos de Papa y ganadería de leche.
Se presentan otras actividades agrícolas de subsistencia como el cultivo de maíz, cebolla,
arveja, y en menor grado uso de la tierra para pastoreo y cría de ovejas (IGAC 1989).
6.2 FASE DE CAMPO
6.2.1 Colecta de hongos
Durante el periodo comprendido entre noviembre de 2000 y julio del 2001 se realizaron 12
salidas de campo con muestreos en una área control de robledal de aproximadamente 10 ha.
Durante los recorridos de muestreo se colectaron los carpóforos o cuerpos fructíferos de
macromicetos encontrados, la duración por cada salida de campo varió dependiendo la
cantidad de carpóforos encontrados, (aproximadamente 1 hora por salida) Para cada
carpóforo se utilizó un envase plástico con el fin de no dejar escapar los insectos asociados.
El esfuerzo de muestreo se midió en minutos recorridos a lo largo del área control.
6.3 FASE DE LABORATORIO
El mismo día del muestreo y en el menor tiempo posible se procede a separar los hongos
en cuatro categorías dependiendo su estado de desarrollo (Tabla 2). Una vez separados los
hongos se completó la información con el examen de las muestras, características físicas,
color, altura, diámetro del píleo, presencia y número de insectos, el número de muestraza
fecha y algunas otras observaciones (anexo 1).

Insectos asociados a hongos macromicetos en robledales de Iguaque
25
En la preparación de las muestras de hongos se disectó una parte del carpóforo, esta se
introdujo en un sobre de papel y se secó en horno para muestras botánicas a una
temperatura aprox. de 25ºC, este manejo permite la preservación de la muestra para su
posterior determinación taxonómica.
Tabla 2. Descripción de cada estado de desarrollo en los carpóforos de hongos macromicetos (Modificado de Navarrete, 1977).
Estado de
desarrollo Descripción física
Estado I
Carpóforos inmaduros, con el
píleo poco extendido y el
himenio en ocasiones cubierto
por el velo. Fase previa a la
maduración.
Estado II
Carpóforos maduros. Píleo
extendido. Fase de liberación
de esporas por primera o única
vez.
Estado III
Carpóforos en descomposición
después de la liberación de
esporas. Húmedos o secos.

Insectos asociados a hongos macromicetos en robledales de Iguaque
26
Estado IV
Carpóforos en descomposición,
húmedos o secos formando
parte de los componentes del
suelo.
Los insectos adultos se preservaron en alcohol al 70%. Los carpóforos infestados de larvas
se llevaron a las cámaras de cría, para el monitoreo de su crecimiento y desarrollo (Figura
4).
Figura 4. Insectos asociados a macromicetos: selección de larvas a partir de carpóforos.
6.3.1 Determinación taxonómica de los insectos y hongos colectados.
Para la determinación taxonómica de los insectos se utilizaron las siguientes claves: Mc
alpine (1981) para Drosophilidae, Clhoropidae, Phoridae, Muscidae, Scatopsidae,
Tachinidae, Psychodidae, Ceratopogonidae; Lane (1948), Mc Alpine (1981) y Papavero
(1978) para Mycetophilidae; Mc Alpine (1981) y Amorin (1992) para Sciaridae; Ashe
(1998) para Staphylinidae y Borror (1981) y Lawrence (1999) para las demás familias de
Coleóptera.

Insectos asociados a hongos macromicetos en robledales de Iguaque
27
Los fragmentos de cuerpo fructífero fueron examinados en busca de estructuras
microscópicas que permitieran revelar su identidad. Observaciones de esporas de hongos
100 aumentos al microscopio, permitieron el reconocimiento de grandes grupos, familias y
géneros. Ocasionalmente se consideraron otras estructuras que facilitaron la determinación
a especie cuando el examen de las esporas no lo permitía.
La identificación micológica se basó en morfología de esporas (forma, tamaño,
ornamentación, grosor de la pared) de esporas, cistidios, elementos de la cutícula o hifas de
la trama de las lamelas, También se realizó un tratamiento con KOH al 2-5% y se
observaron las respuestas de reacción química del material fúngico frente al reactivo
Melzer con anotaciones del color final. El hidróxido de potasio permite reembeber las
estructuras, que han sido colapsadas en el proceso de secado; el reactivo de Melzer,
compuesto de yodo, reacciona de tres formas diferentes: si no existe reacción o sólo se dan
amarillos, se dice que la estructura es inamiloide (la mayoría); si la reacción es al rojo, es
dextrinoide (p. ej., las esporas de Rozites); y si es azul negruzco, amiloide (p. ej., las
esporas de los rusuláceos).
La identificación de agaricales se hizo a nivel de género siguiendo la diagnosis de Singer
(1986).
6.3.2 Cría de insectos
Las cámaras de cría se componen de recipientes de vidrio de boca ancha con una capa
aséptica de tierra negra (hasta formar una capa de aprox. de 2 cm.); esta con el fin de
albergar los insectos que después de alimentarse del tejido fúngico buscan suelo para seguir
su desarrollo al estadio de pupa. El montaje se cubre con una malla de nylon, o un tapón de
espuma (Figura 5).
Cada hongo se localizó en el interior sobre la capa de tierra, y se cerró herméticamente con
la malla o el tapón; el montaje se almacenó en un lugar oscuro y relativamente seco, debido
al comportamiento fotofóbico de las larvas y para evitar la contaminación por hongos.

Insectos asociados a hongos macromicetos en robledales de Iguaque
28
Periódicamente se revisaron las cámaras de cría para extraer los nuevos adultos que
emergían; estos se registraron, cuantificaron y posteriormente se preservaron en alcohol al
70%. Cada cámara se mantuvo por dos meses después de su montaje.
Figura 5. Insectos asociados a macromicetos; Fase de Laboratorio. Cámaras de cría para el monitoreo del desarrollo del los insectos.
Luego de determinados los insectos se creó una base de datos con información acerca de
fecha de colecta, estados de desarrollo del hongo, insectos asociados a cada muestra y
abundancias.
6.4 ANALISIS DE LA INFORMACION
6.4.1 Relación Especies – Abundancia.
Para determinar la estructura de la comunidad a partir de las distribuciones de abundancia
de las especies se organizó la información del número de individuos por especie en clases
de abundancia y se realizó un gráfico de rango abundancia en escala logarítmica. Los datos
resultantes para la comunidad fueron procesados para verificar su ajuste a los siguientes
modelos:

Insectos asociados a hongos macromicetos en robledales de Iguaque
29
Log Normal: S(R)= So exp(-a2R2), donde S(R) = número de especies en el Rth octavo a la
izquierda y derecha de la curva simetrica, So= número de especies en el octavo modal, y
a=(2α2)1/2
Series Logarítmicas: S/N = (1 – x)/x (-Ln (1 – x)); donde S = α[-Ln (1 – x)]N = número
total de individuos y α = N(1 – x)/x
Log Normal: S(R)= So exp(-a2R2), donde S(R) = número de especies en el Rth octavo a la
izquierda y derecha de la curva simétrica, So= número de especies en el octavo modal, y
a=(2σ2)1/2
Palo quebrado: S(n) = [S(S – 1)/N] (1 – n/N)S-2, donde S(n) es el numero de especies en
la clase de abundancia con n individuos.
Para establecer el modelo de abundancia al cual se ajustaban los datos, fueron realizadas
sus respectivas pruebas de bondad de ajuste Χ2, esta metodología básicamente prueba si las
frecuencias observadas están suficientemente próximas a las esperadas que podrían ocurrir
conforme a la Ho, en este caso Ho = los datos muestreados se ajustan al modelo de
distribución de abundancias.
6.4.2 Riqueza Esperada
Para calcular el numero de especies esperadas en la comunidad se utilizo estimadores no
parametricos Chao 1 y Chao, ACE estimador de riqueza de especies basado en la
abundancia e ICE estimador de riqueza de especies basado en la incidencia (Coddington
2000).
Se utilizo el programa estadístico EstimateS 6.0 b (Cowell 2000) que adopta un algoritmo
para estimar el promedio de la tasa a la cual las especies han sido colectadas. Finalmente se
construyó la curva de acumulación de especies, además de los modelos gráficos derivados
de Chao1, Chao2, ICE y ACE generados por el programa. Se interpreto el comportamiento
de la curvas de especies observadas, de únicos (singletons), de duplicados (dobletons) y el

Insectos asociados a hongos macromicetos en robledales de Iguaque
30
calculo de los estimativos no parametricos para deducir que tan representada esta la
entomofauna y la micota con el muestreo en este estudio.
6.4.3 Índices de Riqueza, Diversidad y Dominancia
Para medir la riqueza y la diversidad de especies tanto de insectos como de hongos se
empleó el programa estadístico “sp divers.bas” incluído en el programa Statistical Ecology
(Ludwig & Reynolds, 1989) que calcula los índices de riqueza de Margalef (Dmg) y
Menhinick (Dmn), el índice de abundancia de Shannon (H’) y el índice de dominancia de
Simpson.(D) Estos estimadores pueden dar una idea de la relación Riqueza, dominancia y
abundancia-especies en la comunidad.
Las fórmulas de los índices usados son:
Margalef: DMg = (S – 1) / Ln (N) (Ludwig & Reynolds, 1988, Magurran, 1989) donde N es
la abundancia y S es el numero de especies.
Menhinick: DMn = S / √N (Ludwig & Reynolds, 1988, Magurran, 1989)
donde S = Número de especies.
Shannon: H’ = - ∑pi Ln pi (Ludwig & Reynolds, 1988, Magurran, 1989) donde pi = ni /N;
ni = Número de individuos de la especie i, y N = Número de individuos totales.
Simpson: ∑=
=s
iipD
1
2(Ludwig & Reynolds, 1988, ) donde pi es una proporción de la
abundancia de la especie i.
Para comparar la riqueza de esta fauna con otros trabajos similares se utilizó la metodología
de rarefacción (Simberloff citado por Stiling 1996), se comparó la riqueza obtenida por
Gutiérrez (1998) en bosques de roble en Antioquia, Whertheim et al (2000) para bosques
deciduos y de confieras en Dwingelderveld-Holanda y Hackman y Meinander 1979 en
Finlandia, para un tamaño de muestra de 2000 individuos.
Rarefacción: E(s)=∑i=1s[1-((N-Ni/N)/(N/n))] (Stiling 1996)

Insectos asociados a hongos macromicetos en robledales de Iguaque
31
Donde E(s) es el numero esperado de especies en la muestra enrarecida, n es el tamaño de
muestra estándar (para este caso 2000 individuos) N es el numero total de individuos en la
muestra ha enrarecer, y Ni es el numero de individuos de la iesima especie. En la formula
las expresiones (N-Ni/N) y (N/n) corresponden a una combinación.
6.4.4 Relación precipitación-hongos-Diversidad de insectos
La presencia de cuerpos fructíferos esta relacionada principalmente con la humedad
disponible en el suelo, además de la calidad del mismo y cantidad de hojarasca. Para
evaluar el efecto de la precipitación en la abundancia de hongos se comparó el valor de
precipitación los siete días antes de la colecta y el número de carpóforos encontrados para
el día del muestreo, por medio del coeficiente r de correlación de Pearson se estableció la
relación entre el número de hongos presentes y los valores de precipitación.
Este mismo análisis se aplicó para evaluar el efecto del número de hongos sobre la
entomofauna así: comparación del número de hongos por día de colecta y la cantidad de
insectos asociados a estos. Igualmente se aplicaron índices de correlación para evaluar la
relación entre los valores de precipitación y la abundancia de insectos.
6.4.5. Selección de Hospederos
Con el propósito de establecer las relaciones entre insectos-hongos y la preferencia de
hospederos, se cuantificó las frecuencias de las familias de insectos encontrados en las
diferentes familias de hongos hospederos (anexo Nº6) y se realizó un análisis de
correspondencia simple. El análisis de correspondencia nos permite hacer una descripción
de los perfiles o distribuciones condicionales en la relación insectos-hongos representadas
en un plano factorial, este modelo grafico sugiere que la proximidad entre puntos; se puede
interpretar como dependencia estadística, esta metodología nos permite obtener
información acerca de la relación entre hongos hospederos e insectos asociados.

Insectos asociados a hongos macromicetos en robledales de Iguaque
32
A partir de las cuantificaciones obtenidas del análisis de correspondencia se aplicó un
análisis de clusters, el cual congrega a hongos e insectos en grupos, de tal forma que sea
homogénea la distribución dentro de los grupos y muy heterogénea entre grupos.
6.4.6. Amplitud de la dieta alimenticia
La amplitud de la dieta entre morfoespecies de insectos fue calculada como
piiB
2/1 ∑= donde pi es la fracción de individuos de la especie de insectos en el hongo
i (Southwood 1978) B se incrementa con la diversidad de hongos en la cual el insecto ha
sido encontrado, de manera se pretende evaluar la existencia de competencia interespecífica
por el hongo como recurso alimenticio. El sobrelapamiento de dietas entre pares de familias
de insectos fue calculado como )( , yixiixy ppMIN∑=α en donde Pxi es la proporción del
taxón x en el hongo i y MIN(pxi, pyi) es la mínima relación de pxi y pyi
6.4.7. Categorías Trófica
Según las preferencias por el hospedero, el grado de dependencia por el hongo y su
permanencia en este durante su desarrollo, la entomofauna se dividió en cuatro categorías
tróficas:
• Fungívoros primarios: insectos cuya asociación con el hongo es obligada, exhiben una
leve preferencia por algunos grupos de hongos, tienden a presentarse en los primeros
estadios del hongo aunque es común también encontrarlos en estados mas
desarrollados de maduración
• Fungívoros Secundarios: Insectos cuya asociación no es del todo dependiente; se
encuentran principalmente en los últimos estados de desarrollo del hongo especialmente
cuando este inicia su proceso de descomposición.

Insectos asociados a hongos macromicetos en robledales de Iguaque
33
• Detritívoros: se alimentan de una amplia variedad de material orgánico en
descomposición incluyendo hongos, al igual que los fungívoros secundarios, se
encuentran en los últimos estados avanzados de desarrollo del hongo.
• Predadores: este grupo se alimenta principalmente de los individuos inmaduros de los
grupos anteriormente descritos; en esta categoría se incluyen los insectos que
conforman el último escalón de la cadena trófica incluyendo insectos parasitóides.
6.4.8. Relación Disponibilidad del recurso - Diversidad de insectos
Para evaluar el efecto de la disponibilidad del hongo como recurso sobre la riqueza y
abundancia de insectos, se comparó el número de hongos encontrados para cada día de
colecta con la riqueza y abundancia a través de un análisis de varianza y el test de de
medias de Duncan. Se proponen tres categorías de disponibilidad de hongos por día de
colecta:
• Bajo nivel de disponibilidad: menos de 20 hongos el día de la colecta.
• Mediano nivel de disponibilidad: de 20 a 40 hongos el día de la colecta.
• Alto nivel de disponibilidad: mas de 40 hongos el día de la colecta.
6.4.9. Relación Área - Especies de Insectos
Teniendo en cuenta el modelo clásico área-especies (Mac Arthur & Wilson 1967) en el que
se relaciona el tamaño de una isla y su número de especies, se evaluó la relación entre el
tamaño del carpóforo y la riqueza de especies de la entomofauna presente
Se calculo el volumen aproximado del hongo teniendo en cuenta el diámetro del píleo y la
altura del estípite como si este fuera un cilindro, a este valor se le resto un cuarto del total
del volumen y se obtuvo un volumen final que representa una aproximación al volumen

Insectos asociados a hongos macromicetos en robledales de Iguaque
34
verdadero del hongo, este fue utilizado como un índice del tamaño del hongo y el cual
permitió comparar las variables tamaño, riqueza y abundancia de especies. Por medio del
coeficiente r de correlación de Pearson se compararon las variables tamaño-riqueza de
especies y Tamaño-abundancia. Los hongos se agruparon en tres categorías de tamaños;
con el fin de identificar la categoría de tamaño más importante con relación a la riqueza de
especies se utilizó el análisis de varianza y el test de comparación de medias de Kruskall-
Wallis
6.4.10. Relación Estados de desarrollo - Diversidad de insectos
En la evaluación del efecto del estado de desarrollo del hongo sobre la abundancia de la
entomofauna se comparó la abundancia relativa de insectos para cada uno de los cuatro
estados de desarrollo del hongo, se realizo un análisis de varianza y el test de Duncan para
establecer las diferencias dentro del mismo estado de desarrollo del hongo y entre los
cuatro estados de desarrollo.
De la misma manera se evaluó el efecto de cada estado de desarrollo con respecto a la
riqueza de especies por medio del análisis de varianza y el test de comparación de medias
de Kruskall-Wallis
6.4.11. Patrones de Sucesión de la entomofauna
Para identificar la existencia de algún patrón en el recambio o permanencia de grupos de
insectos que colonizan este tipo de sustrato, se registró la presencia de las familias de
insectos y las categorías ecológicas para cada estado del hongo en las familias
Cortinariacea y Russulaceae.

Insectos asociados a hongos macromicetos en robledales de Iguaque
35
7. RESULTADOS
7.1 CARACTERIZACION DE LOS INSECTOS ASOCIADOS
7.1.1 Abundancia
Se encontraron 1778 individuos en estado adulto y en laboratorio se criaron otros 3409 para un
total de 5187 insectos distribuidos en 48 morfoespecies (Anexo 2), 21 familias y 3 órdenes
(Tabla 3). Las familias más abundantes son Drosophilidae, Endomychidae Scatopsidae y
Derodontidae, las menos abundantes están representadas por una única especie con un número
bajo de individuos. La figura 6 describe la forma en que los individuos están distribuidos por
familias en orden de mayor a menor abundancia.
2558
1185
298248 229 227
13494 83 39 26 21 19 14 12
0
200
400
600
800
1000
1200
1400
1600
1800
2000
2200
2400
2600
Nº d
e in
divi
duos
Droso
philidae
Endomychidae
Scatopsid
ae
Derodontid
ae
Staphyli
nidae
Myceto
philidae
Sciarid
ae
Leiodidae
Phoridae
Psych
odidae
Otras f
amilia
s
Chloropidae
Ceratopogonidae
Procto
trupidae
Formici
dae
Figura 6. Distribución de abundancias por familia de insectos asociados a hongos macromicetos en robledales de Iguaque (Villa de Leyva-Boyacá).
7.1.2 Relación especies-abundancia
Según Magurran (1996) las distribuciones de abundancia de las especies emplean toda la
información reunida en una comunidad y dan la más completa descripción matemática de los

Insectos asociados a hongos macromicetos en robledales de Iguaque
36
Tabla 3. Composición de la entommofauna asociada a los hongos macromicetos en la region de Iguaque (Villa de Leyva,Boyacá). Se incluyen las abundancias absolutas y estilo porcentual de las especies presentes.
Orden Especie Nº de Individuos %
Sp.1 25 0,48
Sp.2 45 0,87
Sp.3 17 0,33
Sp.4 7 0,13
Sp.5 63 1,21
Sp.6 93 1,79
Sp.7 1886 36,36
Sp.8 351 6,77
Sp.9 46 0,89
Sp.10 25 0,48
Leia sp. 1 2 0,04
Mycetophila sp. 1 57 1,10
Mycetophila sp. 2 131 2,53
Mycetophila sp. 3 36 0,69
Zigomia sp. 1 1 0,02
Bradysia sp. 1 81 1,56
Scatopsciara sp. 1 49 0,94
Scatopsciara sp. 2 4 0,08
Sp.1 8 0,15
Sp.2 3 0,06
Sp.3 10 0,19
Sp.1 33 0,64
Sp.2 29 0,56
Sp.3 21 0,40
Sp.1 5 0,10
Silvicola sp. 1 2 0,04
Sp.1 3 0,06
Sp.1 298 5,75
Sp.1 6 0,12
Sp.1 39 0,75
Sp.1 19 0,37
Staphylinidae Sp.1 2 0,04
Sp.1 1 0,02
Sp.1 98 1,89
Sp.2 45 0,87
Sp.3 5 0,10
Sp.4 2 0,04
Sp.5 34 0,66
Sp.6 42 0,81
Sp.1 1167 22,50
Sp.2 18 0,35
Sp.1 248 4,78
Leptodirinae Sp.1 94 1,81
Aphodiinae Sp.1 2 0,04
Sp.1 2 0,04
Sp.1 6 0,12
Solenopsis sp. 1 12 0,23
Quintanus sp. 1 14 0,27
Familia-Subfamilia
Diptera Drosophilidae
Mycetophilidae
Sciaridae
Chloropidae
Phoridae
Muscidae
Anisopodidae
Tipulidae
Scatopsidae
Tachinidae
Psychodidae
Ceratopogonidae
Aleocharinae
Aleocharinae
Aleocharinae
Endomychidae
Dedorontidae
Leiodidae
Scarabidae
Pselaphidae
Elateridae
Hymenoptera Formicidae
Proctotrupidae
Coleoptera Osorinae
Oxitelinae
Aleocharinae
Aleocharinae
Aleocharinae

Insectos asociados a hongos macromicetos en robledales de Iguaque
37
datos, por lo que su estudio es una herramienta diagnóstico del ecosistema y el mejor
estimador de la diversidad.
Se observa que las especies en el total de la muestra se distribuyen normalmente, según
ordenes de abundancia; esto significa la existencia de pocas especies abundantes y un numero
alto de especies raras. La figura 7 relaciona las clases de abundancias con el número de
especies por clase.
0
2
4
6
8
10
12
14
0,0 1,0 1,3 1,4 1,5 1,6 1,6 1,7 1,8 1,9 2,0 2,1 2,5 3,1
Log Abundancia
Nº d
e sp
p
Figura 7. Distribución de abundancias en la comunidad de insectos asociados a hongos en robledales de Iguaque (Villa de Leyva-Boyacá).
La ordenación de las especies en una secuencia de mayor a menor rango también exhibe una
distribución normal de sus abundancias (Figura 8).

Insectos asociados a hongos macromicetos en robledales de Iguaque
38
1
10
100
1000
0 10 20 30 40 50 60
Secuencia de especies
Log
Abun
danc
ia
Figura 8. Distribución de abundancias por rangos de especies de insectos asociados a hongos en robledales de la region de Iguaque (Villa de Leyva-Boyacá).
Se realizaron varias pruebas para determinar el modelo de abundancia que mejor se ajustara
con los datos obtenidos. Según metodología planteada por Magurran (1988) se examinaron los
modelos log normal, logarítmica y palo quebrado utilizando como criterio de bondad de ajuste
el test de chi-cuadrado (Tabla 4).
Como se observa en la tabla 4, para la muestra de 5187 individuos el valor X2= 10.57 para
gl=10 obtenemos un P valor = 0.47. En vista que la probabilidad es mayor que el nivel de
significancia fijado (α=0.05). No podemos rechazar la Ho, por lo tanto con los datos obtenidos
en este estudio estadísticamente se puede concluir que los datos concuerdan con el modelo de
abundancias de especies log normal. Los datos de abundancias y sus transformaciones del
ajuste al modelo mencionado se consideran en el anexo 3.
Tabla 4. Valores de probabilidad y de prueba de bondad de ajuste a los modelos teóricos de distribución de abundancias (log normal, logaritmico y palo quebrado)
Modelo Gl X2 Valor de P
Log Normal 10 10.57 0.47
Serie logaritmica 10 3143.34 0
Palo quebrado 10 118.37 1,08332E-20

Insectos asociados a hongos macromicetos en robledales de Iguaque
39
Los valores calculados para cada índice de riqueza y diversidad utilizado (Shannon, Margalef,
Menhinick, Simpson y equitatividad) se expresan en la tabla 5.
Tabla 5. Valores calculados para los índices de diversidad y equitatividad en la comunidad de insectos asociados a hongos en robledales de Iguaque.
7.1.3 RIQUEZA ESPERADA
El número de especies encontradas depende principalmente del número de muestras que se
hayan tomado, así que las especies más comunes estarán representadas en las primeras
muestras y a medida que se incrementa el muestreo es posible que otras especies mas raras se
añadan a la cuenta, es por ello que la curva de acumulación de especies permite establecer que
tan completo fue el muestreo además de relacionar una medida de esfuerzo de muestreo con el
número acumulado de especies por cada incremento en el esfuerzo, es también útil para
comparar la riqueza esperada y la observada (Cowell y Coddington 1994 ).
La figura 9 representa la curva de acumulación de especies de insectos con respecto al
esfuerzo de muestreo (horas de colecta), las estimas para ICE, Chao1 y Jacknife 2 el
comportamiento de las curvas de singletons (únicos) y doubletons (duplicados).
INDICE VALOR
Shanon 2.28
Equitatividad 0.58
Margalef 5.61
Menhinick 0.66
Simpsom 0.19

Insectos asociados a hongos macromicetos en robledales de Iguaque
40
48
54,96
0
20
40
60
80
100
120
140
160
180
0 1 2 3 4 5 6 7 8 9 10 11 12Salidas de campo
Riq
ueza
de
espe
cies
Sobs Jack2 ICE Chao1 Singletons Doubletons
Figura 9. Curva de acumulación de especies de insectos asociados a hongos y estimas de riqueza para Chao1, ICE y Jacknife 2 con respecto a las salidas de campo en robledales de Iguaque.
De la misma manera se considera la curva de acumulación de especies y estimas de riqueza
para ACE, ICE, Chao1, Chao 2 con respecto al numero de hongos macromicetos hospederos
(figura 10).
La tabla 6 muestra los valores de riqueza esperada calculados para otros estimadores no
parametricos.

Insectos asociados a hongos macromicetos en robledales de Iguaque
41
0
10
20
30
40
50
60
0 10 20 30 40 50 60 70 80 90 100
110
120
130
140
150
160
170
180
190
200
210
220
230
240
250
260
270
280
290
300
Numero de Hongos
Riq
ueza
acu
mul
ada
de e
spec
ies
Sobs
Singletons
Doubletons
ACE
ICE
Chao1
Chao2
Figura 10. Curva de acumulación de especies de insectos y estimas de riqueza para Chao1, Chao 2, ICE, ACE con respecto al número de hongos muestreados.
Tabla 6. Estimas de riqueza para Chao1, Chao2, Jacknife de primer orden, Jacknife de segundo orden, Ace e ICE de la comunidad de insectos asociados a hongos en robledales de Iguaque
ESTIMADOR RIQUEZA
S obs 48
Chao 1 corregido 54.97
Chao 2 corregido 54.96
ICE 54.91
ACE 53.69
Jacknife 1 52.58
Jacknife 2 62.8
Las comparaciones bajo la técnica de rarefacción (Simberloff 1972 citado por Stiling 1996)
con otras entomofaunas de micetócolos indican que en este estudio se reportan valores de
diversidad mas altos(figura 11). Cabe mencionar que para el trabajo de Hackman y Meinander
1979 en Finlandia se encontró 112 especies pero se desconoce el numero total de individuos
en ese estudio.

Insectos asociados a hongos macromicetos en robledales de Iguaque
42
0
5
10
15
20
25
30
35
40
45
50
0 100 200 300 400 500 600 700 800 900 1000 1100 1200 1300 1400 1500 1600 1700 1800 1900 2000
Nº de individuos
Nº d
e es
peci
es
Boyacá Antioquia Dwingelderveld
Figura 11.Curvas de rarefacción y estimación de diversidad de insectos habitantes de macromicetos en bosques de roble de Colombia (Boyacá y Antioquia), comparada con la diversidad en un bosque de zona templada (Dwingelderveld-Holanda).
7.1.4 Categorías Tróficas
Sin duda los fungívoros primarios colonizan el sustrato en los primeros estados de desarrollo.
Sin embargo, se encontró un numero apreciable en estados avanzados de desarrollo, las
tendencias en la selección de hospederos por los fungívoros primarios en algunas ocasiones
contradicen su supuesta preferencia por algunos estados de desarrollo y grupos de hongos;
algunos fungívoros secundarios se encontraron en estados tempranos de desarrollo, esta forma
de clasificación puede representar un intento de aproximación si se tiene en cuenta los hábitos
alimenticios en los insectos (Bruns 1984).
Considerando que especies de una misma familia guardan similitud en sus patrones de
alimentación se encontró un total de 25 especies de Fungívoros primarios 13 especies de
fungívoros secundarios, 7 especies de Detritívoros y 3 especies de depredadores en este grupo
se incluye una especie de avispa parasitoide (tabla 7).

Insectos asociados a hongos macromicetos en robledales de Iguaque
43
Tabla 7. Caracterización trófica de las familias de insectos asociados a hongos macromicetos según sus hábitos alimenticios y grado de dependencia por el hongo.
Fungívoros
primarios
Fungívoros
secundarios
Detritívoros Depredadores
Drosophilidae
Mycetophilidae
Endomychidae
Staphylinidae
Leiodidae
Psychodidae
Derodontidae
Sciaridae
Chloropidae
Scatopsidae
Phoridae
Pselaphidae
Formicidae
Anisopodidae
Ceratopogonidae
Muscidae
Tipulidae
Scarabaeidae
Tachinidae
Elateridae
Proctotrupidae
Al cuantificar cada nivel trófico de acuerdo a la abundancia relativa, se obtiene que los
fungívoros primarios mantienen un predominio amplio, con respecto al resto de categorías.
(Figura 12).
18% 1%1%80%
Fungivoros primariosFungivoros secundariosDetritivorosDepredadores
Figura 12. Predominio ecológico de las categorías tróficas según la abundancia relativa de los insectos asociados a hongos macromicetos en robledales de Iguaque (Villa de Leyva-Colombia)

Insectos asociados a hongos macromicetos en robledales de Iguaque
44
7.2 CARACTERIZACION DE LOS HONGOS HOSPEDEROS
7.2.1 Abundancia de hongos macromicetos
Los resultados obtenidos para la micota presente consideran cada cuerpo fructífero como un
único individuo de esta forma se facilita la cuantificación, estudio y valoración de los datos
obtenidos. Se colectaron 309 hongos de los cuales 226 presentan identificación genérica o
específica, distribuidos en 12 familias y 48 morfoespecies (Anexo 4).
La mayor parte de hongos colectados son macromicetos simbiotróficos (anexo 5), toda la biota
de macromicetos está representada en el primer nivel, el mantillo; el horizonte de humus
contiene todos los grupos excepto Cortinarius. Shubin (2001) reporta en su trabajo a
Cortinarius como dominante en la biota de bosques vírgenes. La alta presencia de Cortinarius
en Colombia ya había sido registrada por Singer (1963) para bosques de roble en la cordillera
oriental. En la cordillera occidental también se da una gran cantidad de especies de
Cortinarius (Matallana, com. pers.).Este género es muy complejo y rico en especies, unas 400
según Hawksworth et al. (1995). Algunas especies de cortinarios poseen orelanina y
cortinarinas, responsables de fallas renales severas (Hawksworth et al., 1995). A diferencia de
otros macromicetos, ejemplares de este genero no son consumidos por los pobladores locales,
esto se asegura su presencia durante su fructificación (Henao com. pers).
En la caracterización de los hongos hospederos se aprecia una marcada presencia de especies
representadas por un solo individuo además se destaca la dominancia de las familias
Cortinariaceae y Russulaceae cuyas especies representan cerca del 50% del total de la micota
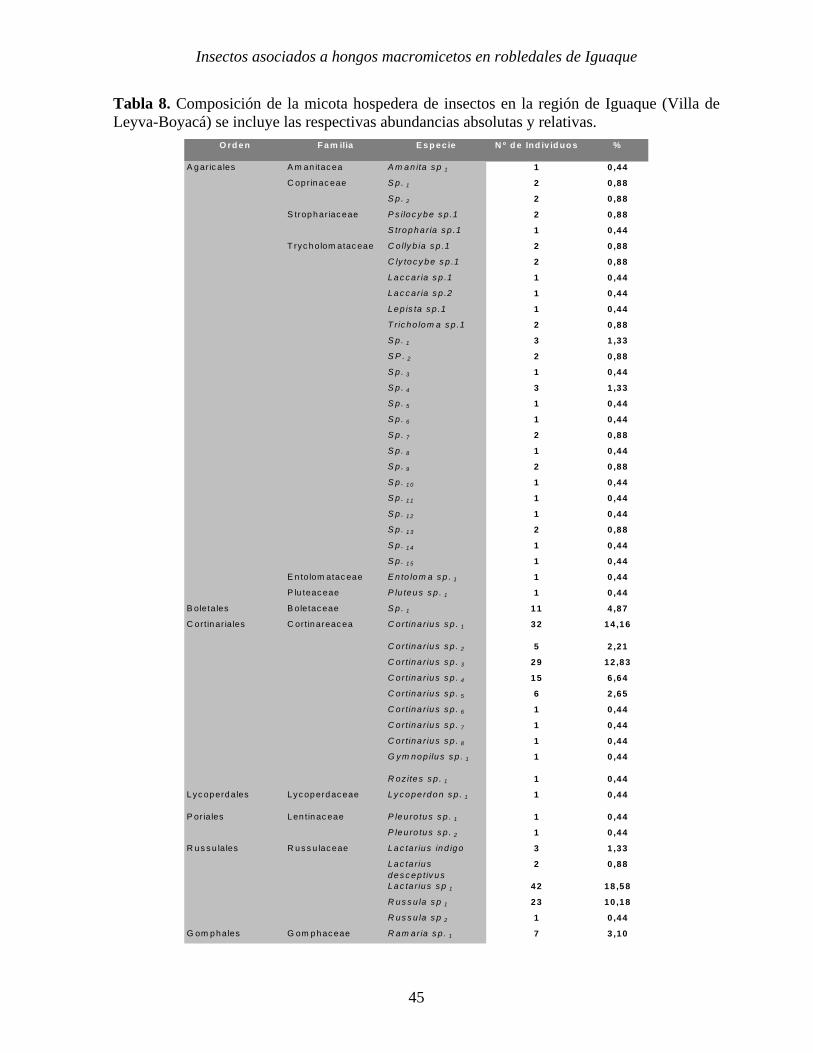
presente en el área muestreada (Tabla 8).
Insectos asociados a hongos macromicetos en robledales de Iguaque
45
Tabla 8. Composición de la micota hospedera de insectos en la región de Iguaque (Villa de Leyva-Boyacá) se incluye las respectivas abundancias absolutas y relativas.
O rd en F am ilia E sp ec ie N º d e Ind iv id uo s %
A m an itac ea A m an ita s p 1 1 0 ,44
S p . 1 2 0 ,88
S p . 2 2 0 ,88
P s iloc y be s p .1 2 0 ,88
S tropha r ia s p .1 1 0 ,44
C olly b ia s p .1 2 0 ,88
C ly toc y be s p .1 2 0 ,88
Lac c ar ia s p .1 1 0 ,44
Lac c ar ia s p .2 1 0 ,44
Lep is ta s p .1 1 0 ,44
T ric ho lom a s p .1 2 0 ,88
S p . 1 3 1 ,33
S P . 2 2 0 ,88
S p . 3 1 0 ,44
S p . 4 3 1 ,33
S p . 5 1 0 ,44
S p . 6 1 0 ,44
S p . 7 2 0 ,88
S p . 8 1 0 ,44
S p . 9 2 0 ,88
S p . 1 0 1 0 ,44
S p . 1 1 1 0 ,44
S p . 1 2 1 0 ,44
S p . 1 3 2 0 ,88
S p . 1 4 1 0 ,44
S p . 1 5 1 0 ,44
E n tolom atac eae E n to lom a s p . 1 1 0 ,44
P lu teac eae P lu teus s p . 1 1 0 ,44
B oletales B oletac eae S p . 1 11 4 ,87
C ortina r ius s p . 1 32 14 ,16
C ortina r ius s p . 2 5 2 ,21
C ortina r ius s p . 3 29 12 ,83
C ortina r ius s p . 4 15 6 ,64
C ortina r ius s p . 5 6 2 ,65
C ortina r ius s p . 6 1 0 ,44
C ortina r ius s p . 7 1 0 ,44
C ortina r ius s p . 8 1 0 ,44
G y m nop ilus s p . 1 1 0 ,44
R oz ites s p . 1 1 0 ,44
Lyc operdales Lyc operdac eae Ly c ope rdon s p . 1 1 0 ,44
P leu ro tus s p . 1 1 0 ,44
P leu ro tus s p . 2 1 0 ,44
Lac ta r ius ind igo 3 1 ,33
Lac ta r ius des c ep tiv us
2 0 ,88
Lac ta r ius s p 1 42 18 ,58
R us s u la s p 1 23 10 ,18
R us s u la s p 2 1 0 ,44
G om phales G om phac eae R am ar ia s p . 1 7 3 ,10
A garic ales
C opr inac eae
S trophar iac eae
T ryc holom atac eae
R us s u lales R us s u lac eae
C ortinar ia les C ortinareac ea
P oria les Len tinac eae

Insectos asociados a hongos macromicetos en robledales de Iguaque
46
La figura 13 describe la forma como se distribuyen los individuos por familias, en un orden de
mayor a menor abundancia.
92
71
32
11
74 3 2 1 1 1 1
0
10
20
30
40
50
60
70
80
90
100
Nº d
e in
divi
duos
Cortinari
acea
e
Russu
lacea
e
Trycho
lomatace
ae
Boletac
eae
Gomphacea
e
Coprinac
eae
Strophari
acea
e
Lentin
acea
e
Amanita
ceae
Entolomatace
ae
Plutea
ecea
e
Lycoperd
acea
e
Figura 13. Distribución de las abundancias por familia de hongo hospedante de insectos en robledales de Iguaque (Villa de Leyva-Boyacá).
7.2.2 Relación especie-abundancias
Al igual que para el análisis de la comunidad de insectos se calcularon los valores de los
índices de riqueza, diversidad y equitatividad (Tabla 9).
Tabla 9. Índices de riqueza, diversidad y equitatividad para la comunidad de hongos macromicetos hospedantes de insectos en robledales de Iguaque (Villa de Leyva-Boyacá)
Indice Valor
Margalef 8.6
Menhinick 3.19
Simpsom 8.818092-02
Shannon 2.93
Equitatividad 0.75

Insectos asociados a hongos macromicetos en robledales de Iguaque
47
También se graficó la distribución de las abundancias (figura 14) de las especies de hongos y
se observa que la mayoría de especies están representadas por un numero muy bajo de
individuos.
0
5
10
15
20
25
30
35
40
45
0,4 0,9 1,1 1,2 1,4 1,4 1,5 1,6 1,6
Log Abundancia
Nº d
e sp
p
Figura 14. Distribución de las abundancias para la comunidad de hongos macromicetos hospedantes de insectos en Robledales de Iguaque (Villa de Leyva-Boyacá).
7.2.3 Riqueza esperada
La tabla 10 presenta los valores de riqueza calculados para los estimadores Chao 1, Chao 2,
Jacknife de primer orden, Jacknife de segundo Orden, ACE e ICE.
Tabla 10. Estimas de riqueza esperada para Chao1, Chao2, Jacknife de primer orden, Jacknife de segundo orden, ACE e ICE de la comunidad de hongos macromicetos hospedantes de insectos en bosques de roble de Iguaque (Villa de Leyva-Boyacá).
Estimador Riqueza
S obs 32
Chao 1 78,28
Chao 2 60,76
ICE 66,71
ACE 69,33
Jacknife 1 49,27
Jacknife 2 61,14

Insectos asociados a hongos macromicetos en robledales de Iguaque
48
La figura 15 corresponde a la curva de acumulación de especies donde se compara los datos de
riqueza observada y esperada para los estimadores Chao 1, ICE y ACE.
0
10
20
30
40
50
60
70
80
90
0 1 2 3 4 5 6 7 8 9 10 11Horas de muestreo
Nº d
e es
peci
es
Sobs Singletons Doubletons Jack2 ICE Chao1
Figura 15. Curva de acumulación de especies y estimas de riqueza esperada para Chao1, ICE y Jacknife2 de la comunidad de hongos macromicetos hospedantes de insectos en robledales de Iguaque (Villa de leyva-Boyacá).
7.2.4 Relación Precipitación-hongos-diversidad de insectos
El coeficiente de correlación r de Pearson para las variables precipitación y hongos dió un
valor de 0.3256 y un valor de F de 0.3017 lo cual indica que no existe relación entre estas
variables, en contraste el valor de la correlación para hongos-insectos: 0.5807 y un valor P de
0.0242 lo que indica que hay evidencia estadística para afirmar la existencia de una relación
directa entre el numero de hongos y numero de insectos asociados. Las variables
precipitación-insectos cuyo coeficiente dio 0.5951 con un valor F de 0.0412 indica que existe
evidencia estadística para afirmar que existe alguna relación entre e lluvia y abundancia de
insectos (Tabla 11).

Insectos asociados a hongos macromicetos en robledales de Iguaque
49
Tabla 11. Coeficiente r de correlación de Pearson para las variables precipitación, Numero de hongos y Numero de Insectos
Precipitación Log Nº de hongos Log Nº de hongos Coeficiente r 0.3256
Valor de F 0.3017 Log Nº de insectos Coeficiente r 0.5951 Coeficiente r 0.5807
Valor de F 0.0412 Valor de F 0.0477
7.3 INTERACCION INSECTO-HONGO
7.3.1 Selección de Hospederos
De las 21 familias de insectos encontradas; 9 exhiben un alto número de individuos asociados
a las 5 principales familias de hongos hospederos, la tabla 12 presenta los valores de las
frecuencias de insectos encontradas en sus respectivos hongos.
Tabla 12. Numero de insectos por familia de hongo hospedero encontrados en robledales de Iguaque.
Cortinariaceae Russulaceae Trycholomataceae Boletaceae Gomphaceae
Drosophilidae 173 595 287 413 67
Endomychidae 0 413 0 0 15
Scatopsidae 39 50 78 11 0
Derodontidae 0 126 0 0 0
Staphylinidae 19 137 10 1 0
Mycetophilidae 52 80 23 0 3
Sciaridae 9 27 1 5 13
Leiodidae 7 49 1 0 0
Phoridae 50 1 21 0 0
Estos insectos presentan una alta preferencia por las familias Cortinariacea, Russulaceae y
Trycholomataceae, El mayor número de familias y de insectos se registro en la familia
Russulacea (tabla 13).

Insectos asociados a hongos macromicetos en robledales de Iguaque
50
Tabla 13. Principales familias de hongos hospederos por numero de familias e insectos asociados.
Familia de hongo Nº de insectos Nº de insectos
por hongo
Nº de familias de
insectos asociadas
Cortinariaceae 364 3.95 11
Russulaceae 1540 21.6 17
Trycholomataceae 424 13.6 8
Boletaceae 430 39 4
Gomphaceae 110 15.7 7
Durante las salidas de campo se registraron tendencias y fluctuaciones en las abundancias de
las familias de insectos y la presencia de carpóforos, los picos más altos se encuentran poco
después de la aparición de un considerable número de hongo (figura 16).
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0,45
0,5
1 2 3 4 5 6 7 8 9 10 11 12
Salida de Campo
Abu
ndan
cias
rela
tivas
de
inse
ctos
Drosophilidae Mycetophilidae Derodontidae Scatopsidae Abundancia Hongos
Figura 16. Abundancias relativas de las principales familias de insectos y total de hongos durante el periodo de muestreo.

Insectos asociados a hongos macromicetos en robledales de Iguaque
51
7.3.2 Amplitud de la dieta
Se calculo la amplitud y sobreposición de la dieta para cada especie y para las familias de
insectos sobre la utilización del hongo como recuso (anexo 7), el valor fluctúa entre 1 y 7, un
valor cercano a 1 se relaciona con especies que utilizan solo una pequeña proporción de
especies hongos disponibles y viceversa, el promedio de este valor para la comunidad es de
1.74 entre especies y 1.66 para familias. El calculo para sobreposición de nicho por parejas de
familias fluctúa entre 0 y 2, cuando dos grupos no tienen ninguna especie de hongo hospedero
en común su valor es 0, y 2 cuando comparten las mismas familias de hongos como recurso.
El promedio de sobreposición para la comunidad es de 0.96.
7.3.3 Relación disponibilidad del recurso-diversidad de insectos
El valor de P>chi- cuadrado para las fuentes de variabilidad (disponibilidad baja, media y alta)
con respecto a riqueza de especies fue mayor de 0.05, lo cual indica que no existen diferencias
significativas entre la categoría de disponibilidad del recurso y riqueza de especies de insectos
(anexo 7). En contraste el valor de P>chi-cuadrado con respecto a las abundancias por
categoría este valor es de 0.0903 lo cual indica que se encuentran diferencias en el numero de
insectos encontrados por categoría de disponibilidad de recurso (Tabla14).
El test de comparación de medias de Duncan agrupa las categorías disponibilidad baja y
media, se observa que estas no presentan diferencias significativas lo que indica un similar
comportamiento de las abundancias de insectos para estas dos categorías. El análisis indica
adicionalmente que la categoría disponibilidad alta del recurso es la más importante en cuanto
a la abundancia de insectos (Tabla 15).

Insectos asociados a hongos macromicetos en robledales de Iguaque
52
Tabla 14. Salida SAS análisis de varianza para abundancia de insectos con respecto a la categoría de disponibilidad del recurso.
Tabla 15.Test de comparación de medias de Duncan para abundancias de insectos encontradas por categoría de disponibilidad de hongos como recurso.
Duncan Grouping Disponibilidad Mean N
A Disponibilidad Alta 860.0 3
B Disponibilidad media 358.2 5
B Disponibilidad baja 167.3 4
7.3.4 Relación área-especies
El modelo clásico de área-especies (MacArthur y Wilson 1967) no se encuentra exitosamente
representado en la relación entre el tamaño del hongo y riqueza de especies de insectos;
adicionalmente los valores del coeficiente de correlación son muy bajos (anexo 9) esto indica
que no hay evidencia estadística para afirmar que haya una relación entre el tamaño del hongo
y la diversidad de insectos.
El análisis de varianza señala que existe evidencia para afirmar que hay diferencias entre la
riqueza de especies de insectos encontrada en cada una de las tres categorías de tamaño (Tabla
16), se evidencia también que principalmente en hongos de talla pequeña es donde se halla la
mayor riqueza de especies de insectos asociados.
Analisis de varianza para variable ABUNDANCIA DE INSECTOS Clasificado por DISPONIBILIDAD DISPONIB LIDAD N MEDIA Entre MS Dentro MS 427713.225 62493.2833 baja 4 167.25 mediana 5 358.20 F Value Prob > F alta 3 860.00 6.844 0.0156 Median Scores (Number of Points Above Median) for Variable INSECTOS Classified by Variable DISPONIB Sumaof Esperado Std Dev Mean DISPONIBILIDAD N Scores Under H0 Under H0 Score baja 4 0.0 2.00 0.85 0.00 mediana 5 3.0 2.50 0.89 0.60 alta 3 3.0 1.50 0.78 1.00 Median 1-Way Analysis (Chi-Square Approximation) CHISQ = 6.6000 DF = 2 Prob > CHISQ = 0.0369

Insectos asociados a hongos macromicetos en robledales de Iguaque
53
Tabla 16. Salida SAS para el análisis de la varianza de Krusskall Wallis para riqueza de especies con respecto al tamaño del hongo.
La figura 17 muestra la distribución de las abundancias y riquezas de insectos de acuerdo al
tamaño de los hongos
.
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
0,90
1,00
1,00 1,20 1,40 1,60 1,80 2,00 2,20 2,40 2,60 2,80
log Tamaño
Log(
rique
za)
0,00
0,50
1,00
1,50
2,00
2,50
3,00
Log(
Abun
danc
ia)
log(riqueza)log(abundundancia)
Figura 17. Correlación entre el tamaño del hongo con respecto a la abundancia y riqueza de especies de insectos asociados.
Analisis de varianza para la variable RIQUEZA DE INSECTOS Clasificado por la variable TAMAÑO AREA N MEDIA Entre MS Dentro MS 209.083333 37.2777778 1 4 20.25 2 4 15.25 F Value Prob > F 3 4 6.00 5.609 0.0262 Wilcoxon Scores (Rank Sums) for Variable RIQUEZA Classified by Variable AREA Sum of Expected Std Dev Mean AREA N Scores Under H0 Under H0 Score 1 4 36.00 26.0 5.87 9.00 2 4 31.50 26.0 5.87 7.87 3 4 10.50 26.0 5.87 2.62 Kruskal-Wallis Test (Chi-Square Approximation) CHISQ = 7.1500 DF = 2 Prob > CHISQ = 0.0280

Insectos asociados a hongos macromicetos en robledales de Iguaque
54
7.3.5 Relación Estado de desarrollo-diversidad de insectos
Existe evidencia estadística para afirmar diferencias entre las abundancias de insectos
econtrados para cada estado de desarrollo del hongo P=0.0001, α=0.001 (anexo 10).
La prueba con el test de comparación de medias de Duncan agrupa el estado 1 y 2, esto indica
que en estos estados de desarrollo no hay diferencia significativa en la abundancia de insectos
encontrados. También se deriva que el estado 3 de desarrollo del hongo se asocia con el mayor
número de insectos (Figura 18 y Anexo11).
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0 1 2 3 4 5Estados de desarrollo
Figura 18. Agrupación por el test de medias de Duncan para abundancias de insectos de acuerdo al estado de Desarrollo del hongo.
Debido a que se registró una baja cantidad de hongos en estados 1 y 2, en el estudio de
varianza y su agrupamiento se asumió como un solo estado de desarrollo; de esta forma el
valor de p>chi-cuadrado fue =0.0325, lo que indica una diferencia significativa entre el estado
del hongo y la riqueza de especies de insectos (Tabla 19). Es consistente en consecuencia
afirmar que el estado 3 se asocia con el mayor valor de riqueza de insectos asociados.

Insectos asociados a hongos macromicetos en robledales de Iguaque
55
Tabla 19. Salida SAS para análisis de la Varianza de Kruskall-Wallis para riqueza de especies con respecto a estados del hongo
7.3.6 Patrones de sucesión de la entomofauna
Para cada estado de desarrollo de las dos familias más abundantes en el estudio, se registró la
presencia de insectos teniendo en cuenta su categoría trófica, la figura 19 y 20 muestra las
ocupaciones de los grupos tróficos para cada estado de desarrollo de la familia Cortinariacea y
Russulacea respectivamente.
Analysis of Variance for Variable RIQUEZAClassified by Variable ESTADO DE DESARROLLO ESTADO N Mean Among MS Within MS 156.333333 40.6944444 1.2 4 7.2500000 3 4 19.7500000 F Value Prob > F 4 4 13.2500000 3.842 0.0622 Wilcoxon Scores (Rank Sums) for Variable RIQUEZA Classified by Variable ESTADO DE DESARROLLO Sum of Expected Std Dev Mean ESTADO N Scores Under H0 Under H0 Score 1.2 4 11.0 26.0 5.83614651 2.75000000 3 4 36.0 26.0 5.83614651 9.00000000 4 4 31.0 26.0 5.83614651 7.75000000 Kruskal-Wallis Test (Chi-Square Approximation) CHISQ = 6.8505 DF = 2 Prob > CHISQ = 0.0325

Insectos asociados a hongos macromicetos en robledales de Iguaque
56
Figura 19. Grupos de insectos de insectos asociados a cada estado de desarrollo para hongos de la familia Cortinariaceae encontrados en robledales de Iguaque (Villa de Leyva-Boyacá).
Figura 20. Grupos de insectos asociados en cada estado de desarrollo para hongos de la familia Russulaceae encontrados en robledales de Iguaque (Villa de Leyva-Boyacá).

Insectos asociados a hongos macromicetos en robledales de Iguaque
57
8. DISCUSION
8.1 Ensamblaje de la comunidad de insectos asociados a hongos
Una distribución con pocas especies dominantes y un gran número de especies raras es común
en comunidades de insectos asociados a Hongos (Pielou & Verna 1968) y en general en
muchas y comunidades (Krebs 1985) en especial en los trópicos, los índices aplicados
evidencian; la poca uniformidad en la distribución de las abundancias según Equitatividad.
Una baja diversidad o alta dominancia de algunas especies según Simpson, y una alta riqueza
y abundancia según Shannon y Margalef.
La comunidad de acuerdo a la distribución de las abundancias exhibe una distribución
especies-abundancia del tipo log normal, la mayoría de comunidades se ajustan a este modelo
(Krebs 1985), May en 1975 sugiere que este tipo de distribución se debe a un considerable
numero de especies en la comunidad y que sus abundancias dependen de múltiples factores
operando de manera independiente. Algunos de estos factores son discutidos mas adelante en
este estudio (disponibilidad y tamaño del recurso, competencia, selección de hospedero), sin
embargo quedaría por estudiarse una considerable cantidad de estos (por Ej. patrones
temporales, dinámicas poblacionales, predación, competencia intraespecifica, estrategias de
ciclos de vida, patrones químicos y físicos reguladores de la interacción entre otros) para
poder determinar su incidencia en la modelación de la composición y estructura de la
comunidad.
Teniendo en cuenta los estimadores de riqueza, se encuentra que las curvas en base a el
numero de salidas de campo y numero de hongos para las especies observadas en el modelo
llega a una asintota (Figuras 7 y 8) lo cual indica una baja probabilidad de encontrar mas
especies de insectos, (Cowell y Coddington 1994, Coddington 2000) la distancia entre los
estimadores y la curva de observados al final del muestreo es pequeña. El comportamiento de
los únicos describe un ascenso y luego tiende a 0 luego, la curva de duplicados se presenta un
poco retrasada en comparación con los únicos. Estos comportamientos de las curvas permiten
concluir que la comunidad se encuentra adecuadamente muestreada (Coddington 2000).

Insectos asociados a hongos macromicetos en robledales de Iguaque
58
Los datos enrarecidos permitieron comparar las faunas en trabajos similares, la fauna aquí
estudiada resulto ser la mas rica en especies resultado que era de esperarse por la situación
geográfica, clima, diversidad de hábitats, régimen de lluvias entre otros, pero hasta el
momento no estaba confirmado ya que la información de la riqueza de especies en estas
comunidades era del todo desconocidas en países tropicales, cabe mencionar los extensos
periodos de muestreo hechos en altas latitudes comparados con los relativamente cortos aquí
en el neotrópico, la relación de la riqueza de especies en estos lapsos es mayor en nuestros
bosques.
8.2 Ensamblaje de la comunidad de hongos hopedantes de insectos
Con respecto a los hongos hospederos se encontró una similar estructura en la composición de
la micota en comparación con la de insectos, los índices indican alta riqueza para Margalef,
una alta dominancia de algunas especies para Simpson y una alta uniformidad esto se debe
básicamente a que la mayoría de especies solo tienen uno o dos individuos que las representan.
Los estimadores arrojan un numero considerablemente mas alto con respecto a lo observado
en campo, al igual que el comportamiento de la curva en el modelo, esta sugiere que a un
mayor esfuerzo de muestreo en este caso expresado en horas es posible encontrar nuevas
especies de hongos antes no muestreadas, de esta manera la micota en esta área no esta
adecuadamente representada. Lo que para este estudio de Interacción no implica que sus
insectos asociados también estén subestimados ya que la curva de acumulación de especies de
insectos con respecto al número de hongos, nos indica que si se colecta un número mayor de
hongos es poco probable encontrar mas insectos asociados, la distancia final entre lo
observado y los esperado, el comportamiento de la curva de acumulación y la estabilización
de la asintota ratifica esta afirmación (Figura 8).

Insectos asociados a hongos macromicetos en robledales de Iguaque
59
8.3 Modelos espacio-temporales resultantes de la interacción insecto-hongo
A partir de la cuantificación del numero de insectos encontrados en cada familia de hongo
se reconocen principalmente 5 agrupaciones en la interaccion, la primera la componen los
insectos asociados a la familia Russulaceae, grupo que se caracteriza por la presencia de un
mayor numero de insectos a fines (17 familias) y la segunda los componen los asociados a la
familia Cortinariacea (11 familias), Trycholomatacea (8 familias) Gomphaceae (7 Familias) y
Boletacea (4 familias).
Merece la atención analizar que la presencia de un insecto en un determinado grupo no
necesariamente significa que se encontró exclusivamente en ese hongo. Drosophilidae por
ejemplo se localiza en el grupo afín con Trycholomataceae y Boletacea aunque es el insecto
mas abundante en Russulaceae, además de encontrarse presente en todos los grupos de hongos
(anexo 6).
La comunidad resulta ser dominada por familias de insectos generalistas (Drosophilidae,
Sciaridae, Scatopsidae y en menor grado Mycetophilidae, representan el 67% del total de
insectos asociados) cabe mencionar la presencia de varias familias poco frecuentes con algún
grado de selectividad.
Estos grupos conforman cinco asociaciones, de los que podemos sugerir la conformación de 2
importantes agrupaciones o subcomunidades, los insectos mas estrechamente asociados a
russulaceos y los relacionados con los cortinariáceos, Russulacea y Cortinariácea representan
familias de órdenes bien diferenciados, tanto en su composición química como sus
características ecológicas, la poca información química y ecológica no nos permite
profundizar sobre sus marcadas diferencias como para hacer afirmaciones mas concluyentes.
Las otras dos agrupaciones son incipientes por sus frecuencias tanto de hongos como de
insectos, además de estar representadas por insectos generalistas de la asociación. Este grado
de generalismo puede explicarse por una alta presión selectiva hacia este comportamiento y
por su bajo costo energético (Hanski 1989).

Insectos asociados a hongos macromicetos en robledales de Iguaque
60
Hanski 1989 plantea dos hipótesis para entender el marcado generalismo en estas
comunidades, la primera hace referencia a la estabilidad del recurso, cuando aumenta la
estabilidad temporal del hongo disminuye el grado de polifagia y viceversa. Y el segundo se
refiere a que las diferencias en la calidad del recurso son mínimas como para hacer uso
indistinto del tipo de recurso. La poca estabilidad temporal de los hongos encontrados en este
trabajo (Russsula sp de 4-a 7 días, Lactarius sp de 6 a 18 días y Ramaria sp de de 15 días a un
mes, (Ruiz com. pers 2002) y las preferencias de hospedero en los pocos grupos de insectos
selectivos como las diferencias químicas y biológicas entre los grupos de hongos, pueden dar
evidencia del cumplimiento de la primera hipótesis.
Con respecto a la amplitud en la dieta, este valor tan bajo se debe principalmente a que la
oferta de hongos en términos de especies es alta, hay un numero alto de especies que pueden
acoger a esta cantidad de especies de insectos, la relación es 48 morfoespecies de hongos y 48
morfoespecies de insectos, así que es poco probable que hayan valores altos. Para este caso la
explicación de una comunidad dominada por generalistas y un valor tan bajo de amplitud de
dieta se debe; principalmente a la fenología de las especies de hongos, temporalmente las
especies de hongos no están presentes de manera constante durante el muestreo (figura 14).
El valor de amplitud de dieta nos esta dando una idea sesgada acerca del uso de las especies de
hongos como alimento, este valor podría constituir una buena herramienta cuando las especies
estén compartiendo una misma dimensión temporal y espacial. Adicionalmente el valor de
sobrelapamiento sugiere una considerable sobreposición de nichos, en la grafica se evidencia
esta tendencia. Sin embargo, el comportamiento de los picos para la distribución de insectos
podría sugerir que están minimizando un efecto de competencia distribuyéndose de manera
diferente durante la dimensión temporal.
8.4 Modelo trófico de la interacción insecto-hongo
La gran mayoría de insectos que integra esta relación son fungívoros primarios, verdaderos
consumidores de hongos tanto en su estado larval como en estado adulto muy pocos insectos
con hábitos Detritívoros o descomponedores hacen parte de la comunidad, al igual que

Insectos asociados a hongos macromicetos en robledales de Iguaque
61
depredadores o parasitóides la figura 20 representa un modelo simplificado de la estructura
trófica de la comunidad.
Figura 20. Modelo simplificado de la estructura trófica encontrada en insectos asociados a macromicetos en robledales de Iguaque (Villa de Leyva-Boyaca).
No se establece algún tipo de estacionalidad en la interacción esto se debe a la irregularidad de
la precipitación en la época de muestreo y corto periodo destinado para este, en la figura 3
podemos observar que los datos de precipitación no superan los 150 mm promedio mensual,
promedio que para otros años son ampliamente superados en épocas lluviosas es decir no hubo
epoca lluviosa durante el muestreo. Esta razón también explica la poca relación entre
precipitación días antes del muestreo y el numero de hongos encontrados, es posible que la
presencia significativamente diferente de hongos se deba a patrones de precipitación para
periodos mas largos.
La relación positiva entre precipitación e insectos hace pensar en la existencia de algún factor
diferente a la cantidad de hongos que condiciona la mayor aparición de insectos en estas
temporadas lluviosas, la lluvia posiblemente puede activar la salida de adultos quiecentes en el
suelo, el apareamiento y oviposición. Es necesario muestreos más largos y periódicos para
establecer algún tipo de estacionalidad en esta relación.

Insectos asociados a hongos macromicetos en robledales de Iguaque
62
El modelo área- especies no se ajusta para esta comunidad lo que indica que los insectos hacen
uso indiferente de acuerdo al tamaño del hospedero, son claras las diferencias de la riqueza de
la entomofauna con respecto a cada categoría de tamaño. El mayor numero especies de
insectos contrario a lo esperado; se dio en el tamaño de hongo mas pequeño, esta tendencia se
podría explicar o refutar teniendo información acerca de los patrones de distribución espacial
de los hongos con relación a su tamaño y sus efectos sobre la atracción de insectos. Se sugiere
identificar un grupo de hongos específico para tratar de deducir un patrón mas claro. En este
estudio la utilización de un solo grupo específico de hongos no hubiese generado la cantidad
de datos suficiente para generar alguna conclusión.
El estado de desarrollo juega un papel importante en la relación, podemos afirmar la existencia
de tres diferentes estados de desarrollo; con respecto a la diversidad de insectos, el estado I y
II pueden considerarse el mismo, el estado de desarrollo preferido para los insectos es el
estado III, estado que posiblemente provee la mejor calidad del recurso el tejido fúngico esta
maduro y aun no se encuentra a merced de agentes descomponedores.
Es obvio pensar que mayor disponibilidad de hongos hay posibilidad de encontrar mas
insectos, de echo para este caso esta premisa se cumple, lo que no se cumple son los valores
de riqueza encontrada dependiendo la disponibilidad del hongo, lo que hace suponer que
siempre encontramos un mismo patrón en el numero de especies explotando los diferentes
grados de disponibilidad, el habito estrictamente micófago de la mayoría de los grupos de
insectos y la existencia de una estrategia de explotación generalista con el recurso puede
explicar la incidencia de estas especies en cada categoría de disponibilidad.
Si tenemos en cuenta que uno de los principales factores que afectan la riqueza de especies es
el estado de desarrollo, es de suponer ciertos cambios en la aparición de grupos de insectos
con relación al estado de desarrollo, el comportamiento temporal de estos grupos no sugiere
un recambio de grupos de insectos sencillamente algunos grupos llegan y se mantienen otras
aparecen y abandonan el hongo manteniendo un patrón según su nivel trófico. Los fungívoros

Insectos asociados a hongos macromicetos en robledales de Iguaque
63
primarios son los primeros en aparecer, a partir del estado III llegan los fungívoros
secundarios y los predadores no muestran ningún tipo de preferencia.
En este sentido la minimización de competencia y la abundancia del recurso permitiría los
solapamientos de grupos de insectos a través de los estados de desarrollo. Seria clave entender
estos comportamientos desde el punto de vista de las historias de vida de las especies.
Este estudio apenas constituye una primera idea general de la diversidad de insectos
Micófagos, ya que una acertada interpretación de las dinámicas de estas comunidades requiere
grupos de extensos datos que incluyan su naturaleza especial y temporal (Devries y Walla
2001).

Insectos asociados a hongos macromicetos en robledales de Iguaque
64
9. CONCLUSIONES
El presente estudio permite concluir los siguientes aspectos:
• La aproximación al conocimiento de las comunidades naturales especialmente
tropicales es una tarea exigente debido a que son entidades extremadamente complejas
donde se conjugan una infinita cantidad de factores que afectan su naturaleza.
• La fauna de insectos asociados a hongos en Iguaque esta conformada por un
considerable numero de especies, lo cual ratifica la presencia de una mayor
complejidad numérica y ecológica de las comunidades tropicales.
• La gran mayoría de insectos asociados a hongos en robledales de Iguaque exhiben
patrones de selección de hospedero generalistas, lo que ratifica la hipótesis de cantidad
expuesta para estas preferencias.
• La comunidad se encuentra subdividas en dos claros grupos o subcomunidades
diferenciadas por las características químicas y físicas del hospedero.
• Se encontró una clara estructura trófica, donde cabe resaltar la mayoría numérica de
Micófagos primarios y la escasa aparición de predadores.
• La abundancia de insectos esta siendo principalmente afectada por el tamaño, estado
de desarrollo y disponibilidad del hongo como recurso y en menor grado la
precipitación.
• La riqueza de especies esta siendo principalmente afectada por el estado de desarrollo
del hongo.
• No hay evidencia de un recambio de especies a manera de sucesión ecológica durante
los estados de desarrollo del hongo.

Insectos asociados a hongos macromicetos en robledales de Iguaque
65
10. RECOMENDACIONES
• La aparición de carpóforos es muy azarosa en términos espaciales, así que trabajar con
una metodología de muestreo como cuadrantes, parcelas o transectos es inadecuada
para estimar una riqueza por unidad de muestreo, se recomienda por su buen
desempeño en este trabajo el uso del esfuerzo de colecta por unidad de tiempo.
• Se recomienda dirigir este tipo de trabajo para comunidades de insectos en bosques
menos degradados, de esta manera se podría llegar a comparar el efecto de la
destrucción de hábitats con respecto a la diversidad de estos insectos en la interacción,
• Se recomienda el estudio de los ciclos de vida en estos insectos, de esta manera se
puede llegar a explorar el efecto de las características del hongo en la historia de vida
del insecto asociado
• Este tipo de relación esta mediada por algún mecanismo de atracción visual o químico,
se recomienda el estudio de estos mecanismos para evaluar el efecto de estos sobre la
interacción.
• El futuro uso de los hongos como recurso alimenticio en humanos y en especial su
producción con fines económicos señala la necesidad de estudiar los insectos que
potencialmente puedan llegarse a convertir en plaga, se recomienda estudiar la fauna
de insectos asociada a cultivos comerciales desconocido para nuestro país.

Insectos asociados a hongos macromicetos en robledales de Iguaque
66
11. REFERENCIAS
Amorin, D. 1992. A catalogue of the family Sciaridae (Diptera) of the Americas South of the
United States. Revista Brasilera de Entomología 36:55-77
Ashe. J.A. 1987. Egg Chamber production, egg protection and clutch size among fungivorous
beetles of the genus Eumicrota (Coleoptera:Staphylinidae) and their evolutionary implications.
Zoological Journal of the Linnean Society 90:255-273
Begon, M., J. Harper & C. Townsen. 1995. Ecología: individuos, poblaciones y comunidades.
Editorial Omega. Barcelona.
Bruns, T.D. 1984. Insect Mycophagy in the Boletales: Fungivore Diversity and the
Mushroom Habitat. Págs, 93 - 129 en: Wheeler, Q.D. & M. Blackwell (eds.) Fungus – Insect
relationships: Perspectives in Ecology and Evolution. Columbia University Press, New York.
Coddington, J. 2000. Criterios cuantitativos para el inventario adecuado utilizando
estimadores de riqueza. Memorias 1ºer Congreso Colombiano de Zoología, mayo 12 al 18.
I.C.N
Collwell, R. 2000 EstimateS, Estatistical estimation of species richness and shared species
from samples Version 6.01b. University of Connecticut. En linea < http:
www.viceroy.eeb.uconn.edu/estimates.
Colwell, R. & J. Coddington. 1994. Estimating terrestrial biodiversity through extrapolation.
Págs. 1-31. en: Hawksworth, D.L (ed.) The quantification and estimation of organismal
biodiversity. Phil. Trans. R. Soc. Lond. B. Londres.
Daniel, W. 1998. Bioestadística: Base para el análisis de las ciencias de la salud. Tercera
edición. Uteha Noriega Editores, México.

Insectos asociados a hongos macromicetos en robledales de Iguaque
67
Devries. J.P & Walla, T.R. 2001. Species diversity and community structure in neotropical
fruit-feeding butterflies. Biological journal of Linnean Society. 74: 1-15
Guevara. R, & Dirzo, R. 1999. Consumption of macro-fungi by invertebrates in Mexican
tropical cloud forest: do fruit body characteristics matter?. Journal of Tropical Ecology
15:603-617
Gutierrez, A.N. 1998. Entomofauna asociada a Agaricales de bosques de Querqus humboldtti
en algunos municipios del departamento de Antioquia. Trabajo de Grado (Biólogo) Facultad
de Ciencias Exactas y Naturales, carrera de biología. Medellín. 264 p.
Graves, R.C. 1960. Ecological observations on the insects and other inhabitants of woody
shelf fungi (Basidiomycetes: Polyporaceae) in the Chicago area. Annals of the Entomological
Society of America Vol. 53: 61-78
Graves R.C & Graves A.C. 1966 The insects and other inhabitants of shelf fungi in the souther
blue rigde region of western north Carolina. I. Introduction, description of the habitat, and host
fungi. Annals of the Entomological Society of America Vol.59: 381-390
Hackman, W. & M. Meinander. 1979. Diptera feeding as larvae macrofungi in Filand. Ann.
Zoolo. Fennici 16:50-83
Hammond, P.M. & J.F. Lawrance. 1989. Mycophagy in Insects: a Summary. Págs. 275-324
en: Wilding, N.M. Collins, P.M. Hammund, J.F. Webber (eds.) Insect – Fungus Interactions.
Academic Press. New York.
Hanski, I. 1989. Fungivory: Fungi, Insects and Ecology. Págs. 25-68 en: Wilding, N.M.
Collins, P.M. Hammund, J.F. Webber (eds.) Insect – Fungus Interactions. Academic Press.
New York.

Insectos asociados a hongos macromicetos en robledales de Iguaque
68
Hawksworth, D. L., P. M. Kirk, B. C. Sutton and D. N. Pegler 1995. Ainsworth and Bisby
Dictionary of the Fungi. 8th edition. IMI-CAB, Cambridge.
Haukioja, E. 1980.On the role of plant defences in the fluctuation of herbivore populations.
Oikos 35:202-289
IGAC, 1989. Levantamiento ecológico del santuario de fauna y flora de Iguaque. Villa de
Leyva-Departamento de Boyacá. Instituto Geográfico Agustín Codazzi. Subdirección de
Docencia e investigación. Bogotá DC.
Janzen, D.H. 1970. Herbivores and the number of tree species in tropical forests. American
Naturalist 104:501-528
Komonen, A. 2001. Structure of insect communities inhabiting old-growth forest specialist
bracket fungi. Ecological Entomology, Vol 26, 63-75.
Klimaszewski, J & S.B. Peck. 1987. Succession and phenology of beetle faunas (Coleoptera)
in the fungus Polyporellus squamosus (Huds.:Fr.) Karst. (Polyporaceae) in Silesia, Poland.
Can. J. Zoool. Vol. 65: 542-550
Krebs, J.C. 1985. Ecologia-Estudio de la distribución y la abundancia. Harla, S.A. Industria
Editorial Mexicana. Mexico D.F. Mexico. 753p.
Lawrence, J.F. 1973 . Host preference in Ciid Beetles (Coleoptera:Ciidae) Inhabiting the
fruiting Bodies of Basidiomycetes in North America. Bulletin of The Museum of Comparative
Zoology. Vol 145:163-211
Lawrence, J.F. 1989. Mycophagy in the Coleptera: Feeding Strategies and Morphological
Adaptations. Págs. 1-23 en: Wilding, N.M. Collins, P.M. Hammond, J.F. Webber (eds.)
Insect – Fungus Interactions. Academic Press. New York.

Insectos asociados a hongos macromicetos en robledales de Iguaque
69
Lawrence, J.F. A. M. Hastings, M. J. Dallwitz, T. A. Paine and E. J. Zurcher 1999. ‘Beetles of
the World: A Key and Information System for Families and Subfamilies.’ Version 1.0 for MS-
Windows (CSIRO Publishing: Melbourne).
Leschen, R.A. 1994. Ecological and Behavioral Correlates amog Mycophagous Coleoptera.
Folia. Entomol. Mex. 92: 9-19
Ludwing, J & J. Reynolds. 1988. Statistical Ecology: A primer on methods and computing.
Wiley – Interscience publications, New York.
Ludwing, J & J. Reynolds. 1989. Sp divers.bas. En: Statistical Ecology: A primer on
methods and computing. Wiley – Interscience publications, New York.
Mac Arthur, P.H & Wilson, E.O. 1967 the theory of island biogeography. Princeton
University Press.
Magurran, A. 1988. Ecological Diversity and Its Measurement. Princenton University Press.
Princenton, New Yersey.
Martin, M. M., Kukor, J.J., Martin, J.S., Matsumara, F., & Johnson, M.W. 1981. Digestive
enzymes of fungus-feeding beetles. Physiol. Zool. 54:137-145
May, R. M. 1975. Patterns of species abundances and diversity. En Ecology and Evolution of
Communities, Editado por M. L. Cody, and J. M. Diamond, 81-120. Harvard University Press.
Cambridge, Mass
Mueller, G. M. & Halling, R. E. 1995. Evidence for hig biodiversity of agaricales (Fungi) in
neotropical montane Querqus forest 302-313 en: Steven P. Curchill (ed) Biodiversiry and
Conservation of Neotropical Montane Forest, The New York Botanic Garden.

Insectos asociados a hongos macromicetos en robledales de Iguaque
70
Navarrete, J.L. & N.E. Galindo. 1997. Escarabajos asociados a Basidiomycetes en San Jose
de los Laureles, Morelos, México (Coleoptera: Scarabaeidae). Folia Entomol. Mex. 99:1-16.
O´connell, T & Bolger, T. 1997. Fungal fruiting bodies and the structure of fungus-micro-
arthropod assemblages. Proceedings of The Royal Irish Academy 97B: 249-262
Pielou, D.P & Verna, A.N. 1968 The arthropod fauna associated with the birch bracket
fungus, Polyporus betulinnus, in eastern Canada. Canadian Entomologist, 100. 1179-1199
SAS Institute. 1993. SAS System Analytical, Statistical. SAS/STAT Users guide. Version 6
Cary, NK.
Shubin, V. I. 2001. Features of organization of edible and medicinal symbiotrophic
macromycetes en forest ecosystems. International Journal of Medicinal Mushrooms 3:217
Shutze, K. 1999. La cobertura vegetal del municipio de Villa de Leyva. Primera edición.
Instituto de Investigaciones Biológicas, Alexander Von Humboldt. Villa de Leyva, Boyacá.
Colombia.
Singer, R. 1963. Oak mycorrhiza fungi in Colombia. Mycopathol. et Mycol. Appl. 20(3-4):239-252 Singer, R. 1986. The Agaricales in Modern Taxonomy. 4th ed. Koeltz Scientific Books, Koenigstein.
Southwood, T.R.E. 1978. Ecological Methods. Chapman & Hall. Segunda edición. Londres.
Stiling, P.D. 1996. Ecology: theories and applications. Segunda edición. Prentice Hall,
New Jersey. 537 p.
Wertheim, B., J. G. Sevenster., I.E. Eijs & J. Van Alphen. 2000. Species diversity in a
mycophagous insect community: the case of spatial aggregation vs. resource partitioning.
Journal of Animal Ecology 69: 335 – 351.

Insectos asociados a hongos macromicetos en robledales de Iguaque
71
Wilding, N. 1989. Introduction chapter. pág x en: Wilding, N.M. Collins, P.M. Hammond, J.F.
Webber (eds.) Insect – Fungus Interactions. Academic Press. New York
Wockeroth, J.R. 1981. Mycetophilidae. Págs: 223-246 en: Mc Alpine, J.F. Manual of the
Neartic Diptera. Vol 1. Biosystematic Research Institute. Otawa, Ontario.
Yakovlev, E.B. & V.I. Tobias. 1992. Braconidae (Hymenoptera) parasites of fungivorous
Diptera in Karelia. Entomologica Fennica Vol. 3: 139-148

Insectos asociados a hongos macromicetos en robledales de Iguaque
72
11. ANEXOS. Anexo 1.
Anexo 1. Formato para le recolección de información ecológica de los carpóforos.

Insectos asociados a hongos macromicetos en robledales de Iguaque
73
ANEXO 2
Principales insectos asociados a macromicetos en robledales de Iguaque: (1). Aleocharinae sp1 (2). Aleocharinae sp2 (3).Aleocharinae sp3 (4).Aleocharinae sp4 (5). Aleocharinae sp5 (6). Aleocharinae sp6

Insectos asociados a hongos macromicetos en robledales de Iguaque
74
Principales insectos asociados a macromicetos en robledales de Iguaque (Continuación): (7).Derodontidae sp1 (8).Elateridae sp1 (9).Endomychidae sp1 (10).Endomychidae sp2 (11). Leptodirinae sp1 (12).Osoriinae sp1

Insectos asociados a hongos macromicetos en robledales de Iguaque
75
¡Error! Principales insectos asociados a macromicetos en robledales de Iguaque. (Continuación): (13). Anisopodidae sp1 (14).Bradysia sp1 (15).Ceratopogonidae sp2 (16).Chloropidae sp1 (17).Chloropidae sp2 (18).Chloropidae sp3

Insectos asociados a hongos macromicetos en robledales de Iguaque
76
Principales insectos asociados a macromicetos en robledales de Iguaque. (Continuación): (19). Drosophilidae sp1 (20).Drosophilidae sp2 (21).Drosophilidae sp3 (22).Drosophilidae sp4 (23). Drosophilidae sp5 (24).Drosophilidae sp6

Insectos asociados a hongos macromicetos en robledales de Iguaque
77
Principales insectos asociados a macromicetos en robledales de Iguaque. (Continuación) (25). Drosophilidae sp7 (26).Drosophilidae sp9 (27).Leia sp1 (28).Mycetophilidae sp1 (29). Mycetophilidae sp2 (30). Mycetophilidae sp3

Insectos asociados a hongos macromicetos en robledales de Iguaque
78
Principales insectos asociados a macromicetos en robledales de Iguaque. (Continuación) (31).Phoridae sp1 (32).Phoridae sp2 (33).Psychodidae sp1 (34).Scatopsidae sp1 (35).Scatopsciara sp1 (36). Scatopsciara sp2

Insectos asociados a hongos macromicetos en robledales de Iguaque
79
Principales insectos asociados a macromicetos en robledales de Iguaque. (Continuación): (37).Tipulidae sp1 (38).Zigomia sp1

Insectos asociados a hongos macromicetos en robledales de Iguaque
80
Anexo 3.
Especie Abundancia Log10Dro.sp1 25 1,39794 0,00967 clase limite obs log10 estandar. esp esp esp acum. Ji Cuadrado
Dro.sp2 45 1,6532125 0,12504 - 0,5 - -0,3010 -1,346286 0,0901 4 4,744702 -Dro.sp3 17 1,2304489 0,00478 1 2,5 8 0,3979 -0,644576 0,2611 9 13,74963 0,1121476Dro.sp4 7 0,845098 0,20657 2 4,5 3 0,6532 -0,388303 0,3483 5 18,341617 0,55192279Dro.sp5 63 1,7993405 0,24974 3 8,5 6 0,9294 -0,111014 0,4562 6 24,023674 0,01779068Dro.sp6 93 1,9684829 0,4474 4 16,5 3 1,2175 0,1781796 0,5714 6 30,090153 1,55004068Dro.sp7 1886 3,2755417 3,90435 5 32,5 7 1,5119 0,4737329 0,6808 6 35,8512 0,26644505Dro.sp8 351 2,5453071 1,55179 6 64,5 11 1,8096 0,7725759 0,7794 5 41,043516 6,49598437Dro.sp9 46 1,6627578 0,13188 7 128,5 4 2,1089 1,0730926 0,8577 4 45,166825 0,00368762
Dro.sp10 25 1,39794 0,00967 8 256,5 2 2,4091 1,3744534 0,9147 3 48,168468 0,3342464Zig.sp1 1 0 1,68896 9 512,5 2 2,7097 1,6762381 0,9525 2 50,159031 4,4738E-05Myc.sp1 57 1,7558749 0,20819 10 1024,5 0 3,0105 1,9782352 0,9761 1 51,401816 1,24278544Myc.sp2 131 2,1172713 0,66859 11 2048,5 2 3,3114 2,2803387 0,9887 1 52,065337 2,6919659Myc.sp3 36 1,5563025 0,0659 10,5750954
Leia.sp1 2 0,30103 0,99714 P valor= 0,47Brad.sp1 81 1,908485 0,37074Scat.sp1 49 1,6901961 0,15257 AJUSTADOScat.sp2 4 0,60206 0,48656Cloro.sp1 8 0,90309 0,15722Clorosp2 3 0,4771213 0,67647Cloro.sp4 10 1 0,08976 σ2 γ θ μ VxSilv.sp1 2 0,30103 0,99714 0,58275 0,34507 0,1598 1,043902979 0,9922072Tip.sp1 3 0,4771213 0,67647 Zo Po S* l MEDIASca.sp1 298 2,4742163 1,37972 -1,3502 0,0885 52,66045 68,98344782 1,2996996Tac.sp1 6 0,7781513 0,27191 x 0,9985Psy.sp1 39 1,5910646 0,08495 α 7,792188Cera.sp1 19 1,2787536 0,00043 s/N 0,009254
Pho.sp1 33 1,5185139 0,04792Pho.sp2 29 1,462398 0,0265Pho.sp3 21 1,3222193 0,00051End.sp1 1167 3,0670709 3,12395End.sp2 18 1,2552725 0,00196Oso.sp1 2 0,30103 0,99714Oxt.sp1 1 0 1,68896Aleo.sp1 98 1,9912261 0,47835Aleo.sp2 45 1,6532125 0,12504Aleo.sp3 5 0,69897 0,36076Aleo.sp4 2 0,30103 0,99714Aleo.s5 34 1,5314789 0,05377
Aleo.sp6 42 1,6232493 0,10475Lpt.sp1 94 1,9731279 0,45364
Dero.sp1 248 2,3944517 1,1987Psela.sp1 2 0,30103 0,99714Musc.sp1 5 0,69897 0,36076Aph.sp1 2 0,30103 0,99714
Quint.sp1 14 1,146128 0,02355Ela.sp1 6 0,7781513 0,27191
Formi.sp1 12 1,0791812 0,048585187 62,385579 27,9718
Anexo3. Prueba de ajuste al modelo log-normal para la comunidad de insectos asociados a hongos en robledales de Iguaque.

Insectos asociados a hongos macromicetos en robledales de Iguaque
81
Anexo 4.
Taxon Fungico Nº de colecciones
Nº de insectos
asociados
Promedio de insectos por carpoforo
Cortinarius sp.1 32 180 5,625 Cortinarius sp.2 5 0 0 Cortinarius sp.3 29 78 2,6897 Cortinarius sp.4 15 96 6,4 Cortinarius sp.5 6 5 0,8333 Cortinarius sp.6 1 5 5 Cortinarius sp.7 1 0 0 Cortinarius sp.8 1 0 0 Gymnopilus sp.1 1 0 0
Cortinariaceae
Rozites sp.1 1 0 0 Russula sp.1 23 195 8,4783 Russula sp.2 1 38 38 Lactarius sp.1 42 1256 29,905 Lactarius indigo 3 51 17
Russulaceae
Lactarius desceptivus 2 0 0 Tricolomatáceo sp.1 3 29 9,6667 Tricolomatáceo sp.2 2 11 5,5 Tricolomatáceo sp.3 1 0 0 Tricolomatáceo sp.4 3 101 33,667 Tricolomatáceo sp.5 1 3 3 Tricolomatáceo sp.6 1 104 104 Tricolomatáceo sp.7 2 25 12,5 Tricolomatáceo sp.8 1 2 2 Tricolomatáceo sp.9 2 46 23 Tricolomatáceo sp.10 1 13 13 Tricolomatáceo sp.11 1 0 0 Tricolomatáceo sp.12 1 0 0 Tricolomatáceo sp.13 2 0 0 Tricolomatáceo sp.14 1 0 0 Tricolomatáceo sp.15 1 0 0 Tricholoma sp.1 2 82 41 Lepista sp.1 1 6 6 Collybia sp.1 2 0 0 Clitocybe sp.1 2 2 1 Laccaria sp.1 1 0 0
Trycholomataceae
Laccaria sp.2 1 0 0 Boletaceae Boletáceo sp.1 11 430 39,091 Gomphaceae Ramaria sp.1 7 110 15,714
Coprinaceae sp.1 2 0 0 Coprinaceae Coprinaceae sp.2 2 0 0 Psilocybe sp.1 2 0 0 Strophariaceae Stropharia sp.1 1 2 2 Pleurotus sp.1 1 0 0 Lentinaceae Pleurotus sp.2 1 6 6
Amanitaceae Amanita sp.1 1 0 0 Entolomataceae Entoloma sp.1 1 0 0 Pluteaceae Pluteus sp.1 1 0 0 Lycoperdaceae Lycoperdon sp.1 1 0 0
Anexo 4. Hongos Macromicetos hospederos de insectos en Robledales de Iguaque

Insectos asociados a hongos macromicetos en robledales de Iguaque
82
Anexo 5
Anexo 5. Cuerpos fructíferos de algunos hongos hospedantes de insectos en robledales de Iguaque (Villa de Leyva-Boyacá). (A) Cortinariaceae. (B) Russulaceae. (C) Gomphaceae. (D) Russulaceae Lactarius indigo

Insectos asociados a hongos macromicetos en robledales de Iguaque
83
Anexo 6.
Cortinariaceae Russulaceae Trycholomataceae Boletaceae Gomphaceae Lentinaceae Strophariaceae TotalDrosophilidae 173 595 287 413 67 0 0 1535Endomychidae 0 413 0 0 15 2 0 430 Scatopsidae 39 50 78 11 0 0 0 178 Derodontidae 0 126 0 0 0 0 0 126 Staphylinidae 19 137 10 1 0 0 0 167 Mycetophilidae 52 80 23 0 3 0 0 158 Sciaridae 9 27 1 5 13 4 0 59 Leiodidae 7 49 1 0 0 0 2 59 Phoridae 50 1 21 0 0 0 0 72 Psychodidae 7 8 0 0 0 0 0 15 Chloropidae 0 12 0 0 7 0 0 19 Ceratopogonidae 0 19 0 0 0 0 0 19 Proctotrupidae 3 7 0 0 0 0 0 10 Formicidae 0 0 0 0 0 0 0 0 Elateridae 1 1 1 0 0 0 0 3 Tachinidae 0 6 0 0 0 0 0 6 Muscidae 0 4 0 0 0 0 0 4 Tipulidae 0 0 0 0 3 0 0 3 Anisopodidae 0 0 0 0 2 0 0 2 Pselaphidae 0 2 0 0 0 0 0 2 Scarabidae 2 0 0 0 0 0 0 2 Total 362 1537 422 430 110 6 2
Anexo 6. Valores de las frecuencias de insectos por familia de hongo encontrados en robledales de Iguaque (Villa de Leyva-Boyacá)

Insectos asociados a hongos macromicetos en robledales de Iguaque
84
Anexo 7. Morfosp AmplitudDro.sp1 4,5Dro.sp2 2,19Dro.sp3 1,85Dro.sp4 1,8Dro.sp5 3,7Dro.sp6 1,17Dro.sp7 3,9Dro.sp8 2,89Dro.sp9 1Dro.sp10 2,21Zig.sp1 1Myc.sp1 3,08Myc.sp2 3,09Myc.sp3 4,17Leia.sp1 1Brad.sp1 3,09Scat.sp1 4,5Scat.sp2 1Cloro.sp1 2,33Clorosp2 1Cloro.sp4 1Silv.sp1 1Tip.sp1 1Sca.sp1 3,33Tac.sp1 1Psy.sp1 2,22Cera.sp1 1Pho.sp1 1,85Pho.sp2 1,56Pho.sp3 1End.sp1 2,06End.sp2 1,13Oso.sp1 1Oxt.sp1 1Aleo.sp1 2,23Aleo.sp2 1,59Aleo.sp3 2Aleo.sp4 1Aleo.s5 1Aleo.sp6 1Lpt.sp1 2,09Dero.sp1 1,01Psela.sp1 1Musc.sp1 4Aph.sp1 2Quint.sp1 1,72Ela.sp1 3
Familia Amplitud Min piAnisopodidae 1 1Ceratopogonidae 1 1Chloropidae 1,87 0,368Derodontidae 1 1 Amplitud de 1 a 7Drosophilidae 3,69 0,043 Sobrelapamiento de1 a 2Elateridae 3 0,333 Prom amplitud 1,747Endomychidae 1,08 0,004 Prom Sobrelap 0,964Formicidae 0 0Leiodidae 1,35 0,014Muscidae 1 1Mycetophilidae 2,53 0,02Phoridae 1,76 0,013Proctotrupidae 1,72 0,3Pselaphidae 1 1Psychodidae 1,99 0,466Scarabidae 1 1Scatopsidae 3,09 0,061Sciaridae 3,4 0,016Staphylinidae 1,46 0,006Tachinidae 1 1Tipulidae 1 1
Anisopodidae Ceratopogonidae Chloropidae Derodontidae Drosophilidae Elateridae Endomychidae Leiodidae Muscidae Mycetophilidae Phoridae Proctotrupidae Pselaphidae Psychodidae Scarabidae Scatopsidae Sciaridae Staphylinidae Tachinidae
Anisopodidae
Ceratopogonidae 2
Chloropidae 1,368 1,368
Derodontidae 2 2 1,368
Drosophilidae 1,043 1,043 0,411 1,043
Elateridae 1,333 1,333 0,701 1,333 0,376
Endomychidae 1,004 1,004 0,372 1,004 0,047 0,337
Leiodidae 1,014 1,014 0,382 1,014 0,057 0,347 0,018
Muscidae 2 2 1,368 2 1,043 1,333 1,004 1,014
Mycetophilidae 1,02 1,02 0,388 1,02 0,063 0,353 0,024 0,034 1,02
Phoridae 1,013 1,013 0,381 1,013 0,056 0,346 0,017 0,027 1,013 0,033
Proctotrupidae 1,3 1,3 0,668 1,3 0,343 0,633 0,304 0,314 1,3 0,32 0,313
Pselaphidae 2 2 1,368 2 1,043 1,333 1,004 1,014 2 1,02 1,013 1,3
Psychodidae 1,466 1,466 0,834 1,466 0,509 0,799 0,47 0,48 1,466 0,486 0,479 0,766 1,466
Scarabidae 2 2 1,368 2 1,043 1,333 1,004 1,014 2 1,02 1,013 1,3 2 1,466
Scatopsidae 1,061 1,061 0,429 1,061 0,104 0,394 0,065 0,075 1,061 0,081 0,074 0,361 1,061 0,527 1,061
Sciaridae 1,016 1,016 0,384 1,016 0,059 0,349 0,02 0,03 1,016 0,036 0,029 0,316 1,016 0,482 1,016 0,077
Staphylinidae 1,006 1,006 0,374 1,006 0,049 0,339 0,01 0,02 1,006 0,026 0,019 0,306 1,006 0,472 1,006 0,067 0,022
Tachinidae 2 2 1,368 2 1,043 1,333 1,004 1,014 2 1,02 1,013 1,3 2 1,466 2 1,061 1,016 1,006
Tipulidae 2 2 1,368 2 1,043 1,333 1,004 1,014 2 1,02 1,013 1,3 2 1,466 2 1,061 1,016 1,006 2
Anexo 7. Valores de amplitud y sobreposición de dieta para las especies y familias de insectos asociados a hongos en robledales de Iguaque (Villa de Leyva-Boyacá).

Insectos asociados a hongos macromicetos en robledales de Iguaque
85
Analisis de Varianza para la variable RIQUEZA Clasificado por DISPONIBILIDAD DE RECURSO DISPONIBILIDAD N MEDIA ENTRE MS DENTRO MS 58.1000000 46.2740741 baja 4 12.50 mediana 5 15.20 F Value Prob > F alta 3 20.66 1.256 0.3304 Wilcoxon Scores (Rank Sums) for Variable RIQUEZA Classified by Variable DISPONIB
Suma de Expected Std Dev Mean DISPONIBILIDAD N Scores Under H0 Under H0 Score baja 4 20.50 26.00 5.85 5.12 mediana 5 30.00 32.50 6.12 6.00 alta 3 27.50 19.50 5.37 9.16 Kruskal-Wallis Test (Chi-Square Approximation) CHISQ = 2.3435 DF = 2 Prob > CHISQ = 0.3098
Anexo 8. Salida SAS del análisis de la varianza de Krusskall-Wallis para riqueza de especies con respecto a la categoría e disponibilidad de recurso. Anexo 9. Coeficientes de correlación r de Pearson para abundancia y riqueza de especies con respecto al tamaño del hongo.
Abundancia Riqueza
Tamaño Coeficiente r 0.17175 Coeficiente r -0.02569
Valor de P 0.0127 Valor de P 0.7120
Anexo 10. Salida SAS, Análisis de varianza para abundancia de insectos con respecto al estado de desarrollo del hongo.
General Linear Models Procedure Dependent Variable: ABUNDANCIA Source DF Sum of Squares Mean Square F Value Pr > F Model 3 2.01286329 0.67095443 13.05 0.0001 Error 44 2.26236770 0.05141745 Corrected Total 47 4.27523099

Insectos asociados a hongos macromicetos en robledales de Iguaque
86
Anexo 11. Agrupación por el test de comparación Duncan para abundancias de insectos de acuerdo al estado de desarrollo del hongo.
Duncan Grouping Estado de desarrollo N Mean
A 3 12 0.54000
B 4 12 0.33380
C 2 12 0.12123
C 1 12 0.00498


















