
Relationship between nutritional state and testes function, together with observations on patterns...
9
J. Zool., Lond. (1980) 192, 147-155 Relationship between nutritional state and testes function, together with observations on patterns of feeding, in the toad, Bufo bufo bufo KALLOL GUHA,* c. BARKER J0RGENSEN AND LIS OLESEN LARSEN Zoophysiological Laboratory A, August Krogh Institute, I3 Universitetsparken 13, DK-2100 Copenhugen, Denmark (Accepted 8 January 1980) (With 1 figure in the text) The effects of 10 weeks of starvation, restricted feeding, or od lib. feeding on nutritional state and testes functions were investigated in male toads that had artificially hibernated in a refrigerator before being placed at room temperature. Food intake increased for about four weeks in both the fed groups. After this initial period the toads on restricted food ate their ration for the remaining part of the experiment, and they developed fat bodies of sizes characteristic of male toads during late summer in nature. The toads fed ad lib. increased food intake to about twice that of those on restricted food. After two weeks of high feeding rate food intake rapidly declined and remained low to the end of the experi- ment, when the fat bodies were larger than normally seen in nature. At the end of the experiment spermatogenetic activity had declined to a low level in the starving toads, whereas it was high in the fed toads. Starvation for 10 weeks did not affect the late stages of spermatogenesis. Interstitial cells and thumbpads remained reduced in the starving toads, but were restored in the fed toads. The importance of the annual cycle in nutritional state for gonadal cycles in male and female toads is discussed. Contents Introduction ...... .. .. .. Material and methods .... .. .. .. Results .... .. .. .. .. .. Food intake . . .. .. .. .. .. Body weight and weight of fat bodies . . .. Testes weight .... .. .. .. .. Tubule diameter . . .. .. .. .. Spermatogenetic state . . .. .. .. Interstitium . . .. .. .. .... Thumbpads .. .. .. .... .. Discussion . . .. .. .. ...... Feeding pattern and fat bodies . . .. .. Spermatogenesis . . .. .. .. .. Interstitiurn and thumbpads .. .. .. Comparison with ovary .... .. .. Summary . . .... .... .. .. References . . .... ...... .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. Page 147 148 148 148 149 149 151 151 151 151 152 152 152 153 153 153 154 Introduction Temperature zone anurans exhibit annual gonadal cycles that are correlated with cycles in nutritional state. In female toads, Bufo bufb bufo, initiation of a new ovarian *Present address: Department of Anatomy, University of Kuopio, SF-70101, Kuopio 10, Finland. 147 0022-5160/80/100147 +09 $02.00.10 (Q 1980 The Zoological Society of London
-
Upload
kallol-guha -
Category
Documents
-
view
212 -
download
0
Transcript of Relationship between nutritional state and testes function, together with observations on patterns...

J. Zool., Lond. (1980) 192, 147-155
Relationship between nutritional state and testes function, together with observations on patterns of feeding, in the toad, Bufo bufo bufo
KALLOL G U H A , * c. B A R K E R J 0 R G E N S E N A N D LIS O L E S E N L A R S E N Zoophysiological Laboratory A, August Krogh Institute,
I3 Universitetsparken 13, DK-2100 Copenhugen, Denmark
(Accepted 8 January 1980)
(With 1 figure in the text)
The effects of 10 weeks of starvation, restricted feeding, or od lib. feeding on nutritional state and testes functions were investigated in male toads that had artificially hibernated in a refrigerator before being placed at room temperature. Food intake increased for about four weeks in both the fed groups. After this initial period the toads on restricted food ate their ration for the remaining part of the experiment, and they developed fat bodies of sizes characteristic of male toads during late summer in nature. The toads fed ad lib. increased food intake to about twice that of those on restricted food. After two weeks of high feeding rate food intake rapidly declined and remained low to the end of the experi- ment, when the fat bodies were larger than normally seen in nature. At the end of the experiment spermatogenetic activity had declined to a low level in the starving toads, whereas it was high in the fed toads. Starvation for 10 weeks did not affect the late stages of spermatogenesis. Interstitial cells and thumbpads remained reduced in the starving toads, but were restored in the fed toads. The importance of the annual cycle in nutritional state for gonadal cycles in male and female toads is discussed.
Contents
Introduction . . . . . . . . . . .. Material and methods . . . . . . . . . . Results . . . . . . . . . . . . ..
Food intake . . . . . . . . . . . . Body weight and weight of fat bodies . . . . Testes weight . . . . . . . . . . . . Tubule diameter . . . . . . . . . . Spermatogenetic state . . . . . . . . Interstitium . . . . . . . . . . . . Thumbpads . . . . . . . . . . . .
Discussion . . . . . . . . . . . . . . Feeding pattern and fat bodies . . . . . . Spermatogenesis . . . . . . . . . . Interstitiurn and thumbpads . . . . . . Comparison with ovary . . . . . . . .
Summary . . . . . . . . . . . . . . References . . . . . . . . . . . . . .
. .
..
..
. .
. .
..
. .
. .
. .
. .
..
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
..
. .
. .
. .
. .
. .
. .
. .
. .
..
..
. .
. .
. .
. .
. .
. .
Page 147 148 148 148 149 149 151 151 151 151 152 152 152 153 153 153 154
Introduction Temperature zone anurans exhibit annual gonadal cycles that are correlated with
cycles in nutritional state. In female toads, Bufo bufb bufo, initiation of a new ovarian *Present address: Department of Anatomy, University of Kuopio, SF-70101, Kuopio 10, Finland.
147 0022-5160/80/100147 +09 $02.00.10 (Q 1980 The Zoological Society of London

I48 K . G U H A , C. B A R K E R J 0 R G E N S E N A N D L. O L E S E N L A R S E N
cycle after breeding in spring presumably depends upon reestablishment of a satisfactory nutritional state after the period of starvation lasting throughout hibernation and breeding (Jrargensen et a/., 1979). In male toads initiation of a new spermatogenetic cycle seems to be less dependent upon the nutritional state, but little is known about the effects of starvation or feeding on functions of the testes in the toad, or other amphibians. The effects of starvation on testes in amphibians have previously been studied with regard to weight and gross histology of the testes, and colour of the thumbpads (urodeles: Adams & Rae, 1929; Champy, 1921 ; Ifft, 1942; anurans: Aron, 1926; Heusser, 1969; Kehl, 1944; Nussbaum, 1906a, b ; Ott, 1924; Penhos & Cardeza, 1950; Sluiter et al., 1950; Zander, 1951). In general, a marked weight loss of the testes was observed only after some months of starvation, and spermatogenesis might continue even after prolonged starvation, but detailed studies are lacking on spermatogenetic activity and secondary sex characters, the thumbpads, and the effects of various food levels d o not seem to have been studied. The present investigation was therefore performed to provide a quantitative estimation of changes in various parameters reflecting spermatogenetic activity and androgen produc- tion in toads that were starved or fed at two levels.
Material and methods Adult male toads, Bufo bufo bufo (L.), were collected on the island Falster, Denmark, i n
September. Prior to the experiment the toads were kept in a refrigerator at 5°C for 6 months, which corresponds to the duration of normal hibernation in Denmark. In March, four groups of five toads each were placed at 22-23°C in an artificially illuminated room (12L/12D). Each group of toads, which were marked individually by toe clipping, were kept in a 36 x 55 cm plastic box on filter paper and with access to water in a dish. One group of toads were killed 24 hours after being taken out of the refrigerator, and a second group was not provided with food through- out the experiment, which lasted 10 weeks. The third group was on restricted food regime, receiving 50 Tetiebrio molitor larvae (meal worms) per week for the first 3 weeks and 100 meal worms per week for the rest of the experiment. A fourth group was fed ad lib. being offered meal worms in excess of consumption for 24 hours every second day. Only on day 17 and 29 did they eat the whole ration. Each day the meal worms were inspected, and if dead meal worms were found, they were replaced by live ones. Numbers of meal worms eaten by the groups of toads were determined by counting those left.
At autopsy the fat bodies and testes were weighed. The testes and thumbpads were fixed in Bouin’s fluid, embedded in paraffin, cut at 7 pm and stained with Ehrlich’s hematoxylin-eosin or with Brookes’ hemalum-orange G-carmoisin-wool green. The functional state of the semin- iferous tubules and the development of the interstitial tissue and thumbpads were assessed as previously described (van Dongen et af., 1966). Thumbpad development reflects androgen secretion of the interstitial cells and can therefore be used to evaluate the functional state of the interstitium (see Rastogi, 1976).
Differences between mean values have been tested for statistical significance by means of Student’s t-test.
Results Food intake
In the ad fib. fed group transferred from the refrigerator to the laboratory, food intake increased for 25 days, followed by a sharp decline to one quarter of maximum food intake and then a gradual decrease to near zero. In the group fed limited amounts of meal worms
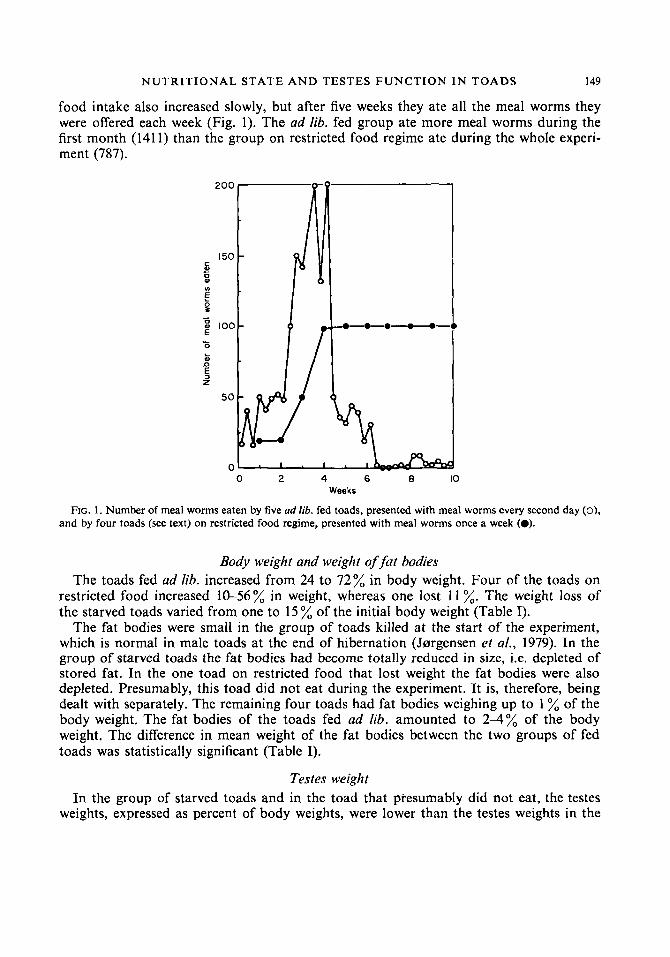
NUTRITIONAL STATE A N D TESTES F U N C T I O N I N TOADS 149
food intake also increased slowly, but after five weeks they ate all the meal worms they were offered each week (Fig. 1). The ad lib. fed group ate more meal worms during the first month (1411) than the group on restricted food regime ate during the whole experi- men t (787).
10 0 0 2 4 6 8
Weeks
FIG. 1 . Number of meal worms eaten by five ad lib. fed toads, presented with meal worms every second day (0), and by four toads (see text) on restricted food regime, presented with meal worms once a week (0).
Body weight and weight of fut bodies The toads fed ad lib. increased from 24 to 72% in body weight. Four of the toads on
restricted food increased 10-56% in weight, whereas one lost 11 %. The weight loss of the starved toads varied from one to 15 % of the initial body weight (Table I).
The fat bodies were small in the group of toads killed at the start of the experiment, which is normal in male toads at the end of hibernation (Jorgensen et al., 1979). In the group of starved toads the fat bodies had become totally reduced in size, i.e. depleted of stored fat. In the one toad on restricted food that lost weight the fat bodies were also depleted. Presumably, this toad did not eat during the experiment. It is, therefore, being dealt with separately. The remaining four toads had fat bodies weighing up to 1 % of the body weight, The fat bodies of the toads fed ad lib. amounted to 2 4 % of the body weight. The difference in mean weight of the fat bodies between the two groups of fed toads was statistically significant (Table I).
Testes weight In the group of starved toads and in the toad that piesumably did not eat, the testes
weights, expressed as percent of body weights, were lower than the testes weights in the

TA
BL
E I
Efle
cts o
f sta
rvat
ion
or fe
edin
g fo
r 10
wee
ks o
n bo
dy w
eigh
t, w
eigh
t of f
ar b
odie
s an
d te
stes
, trtb
rrle
diam
eter
, thr
onbp
nds,
nnd
inte
rstit
ial t
issi
re in
inal
e to
ads,
&fo
bufo bufo
(L.)
Num
ber
Gro
ups
of to
ads
Initi
al
body
wta
(8
)
Perc
ent
Fat b
ody
wt,
body
wtb
of
bod
y w
t"
chan
ge in
as
per
cent
Te
stes
wei
ght
as p
erce
nt
of b
ody
wt'l
Tubu
le
diam
eter
" (P
m)
Hei
ght
of
Con
ditio
n of
inte
rstit
ium
' th
umbp
ad
-
epith
eliu
m"
Inte
r-
Wel
l (w
) R
educ
ed
med
iate
de
velo
ped
Con
trols
at
sta
rt
5 St
arve
d 5
Food
ld
re
stric
ted
4 Fe
d ad
lib.
5
30k2
28
+ 1
23
30
22
29
k 1
0.09
f 0.
03
-7(-
1-15
) -0
-1
1 -0
31
(10-
56)
0.85
f 0.
17
50(2
4-72
) 2.
77 5
0.4
s
0.48
2 0.
07
0.40
f 0.
01
0.42
0.
5920
.10
0.63
10.
08 *
283
& 1
6 21
5&9
20 1
299
& 6
** *
324+
26*
*
112+
12
0 0
5 32
( -0-57jb
3 I
0
1845
13
1 1
2 2
32
fll*
0
0 5
3
-0
1
n m
"Mea
ns+s
.e.m
.; bM
eans
and
rang
e; 'N
umbe
r of
toa
ds; d
Not
eatin
g (s
ee te
xt).
Stat
istic
al s
igni
fican
ce o
f di
ffer
ence
s: *P<
0.05
; **P
<O
.OI ;
***P
<O
.OO
I. Si
gnifi
canc
es re
fer
to d
iffer
ence
s be
twee
n gr
oup
of s
tarv
ed t
oads
and
gr
oups
of f
ed to
ads,
exc
ept f
at b
ody
wei
ght a
nd h
eigh
t of
thum
bpad
s whi
ch re
fer t
o di
ffer
ence
s be
rwee
n to
ads f
ed u
dlib
. and
toa
ds o
n re
stric
ted
food
.

N U T R I T I O N A L STATE A N D TESTES F U N C T I O N IN T O A D S
fed toads, but only the difference between starved toads and toads fed ad lib. was statisti- cally significant.
151
Tubule diameter The tubule diameters followed closely the testes weights, being significantly larger in
the feeding toads than in the starving, with no clear difference between the toads at the two feeding levels (Table I).
Spermatogenetic state At the start of the experiment the seminiferous tubules contained only early spermato-
genetic stages, up to cysts of primary spermatocytes, and spermatozoan bundles. At autopsy the spermatogenetic wave had progressed, and all spermatogenetic stages were present, but the frequency of distribution of the stages varied between the starved and the fed toads. The spermatogenetic activity, expressed by the number of cysts of secondary spermatogonia, was significantly higher in the groups of feeding toads than in the starving toads. Also the numbers of spermatocytic cysts were significantly higher in the fed groups, whereas there was no clear difference in numbers of cysts of spermatid stages or of spermatozoan bundles between the groups. The spermatogenetic activity was the same in the toads at the two feeding levels (Table 11).
T A B L E I1 Effects of starvation or feeding for 10 weeks on functional state of seminiferous tubules in toad testes
Number of cysts per tubule cross section (Meansks.e.m.) Number Primary Secondary Spermatozoan
Groups of toads spermatogonia spermatogonia Spermatocytes Spermatides bundles
Controls at start 5 5.9 k 0.6 6.0k 1 . 1 I .9 f 0.4 0 7.8 f 1.3
Starved 5 3.9 2 0.4 0.6? 0.2 1.3 k 0.3 2.3 f 0.4 11.6f 1.0 Food 1 " 2.9 0.4 0.1 1.2 11.4
restricted 4 3.0 f 0.4 3.1 +0.5** 2.9 k 0.5 * 2.1 f0 .4 12.0f 1.7 Fed ad lib. 5 4.6k0.5 2.7 5 0.3 *** 3.9 f 0.8* 3.2 f 0.5 13.1 k1.3
"Not eating (see text). Statistical significance, see note below Table I.
In terstitium The interstitium was well developed in the controls at start. At the end of the experi-
ment the interstitium of the starved toads, including the one from the group on restricted food that did not eat, was in a reduced state, characterized by few cells with pycnotic nuclei. In the ud lib. fed toads the interstitium was well developed, with large cells con- taining large nuclei, whereas it was well developed in only two of the four toads feeding on the restricted food regime (Table I).
Thumbpads The controls at start had brown thumbpads with medium sized papillae. At the end
of the experiment the starved toads and the one that did not eat had reduced thumbpads,

152 K . G U H A , C . B A R K E R J 0 R G E N S E N A N D L . OLESEN LARSEN
whereas the feeding toads possessed well developed thumbpads. The thumbpad epithelium in the group of ad lib. fed toads was significantly higher than the thumbpads of toads on restricted food (Table I).
Discussion Feeding pattern and fat bodies
The food intake was low in the toads immediately after their transfer from the refriger- ator to room temperature. It increased slowly and did not reach maximum level until after three to four weeks. This gradual increase of food uptake in cold acclimated toads upon transfer to a high temperature agrees with studies on the feeding behaviour of toads taken from the refrigerator into the laboratory. I t takes such toads up to three weeks to reach maximum responsiveness in snapping at a meal worm or a dummy (Jytte Nielsen, pers. comm.).
Tt is noteworthy that toads fed ad lib. practically stopped feeding after about a month, whereas the toads on restricted food continued to eat their weekly ration throughout the experiment. The toads fed ad lib. had attained fat bodies amounting to about 3 % of the body weight at the end of the experiment despite their low feeding rate the month prior to autopsy. The mean weight of the fat bodies of the four toads that continued to eat their restricted ration was 0.9% of the body weight. In nature, the annual maximum size of the fat bodies in male toads is normally about 1 % of the body weight (Jarrgensen et a/., 1979). The toads on restricted food thus had fat bodies comparable to those attained by toads during natural feeding conditions in summer, whereas fat bodies of sizes observed in the toads fed ad lib. are seldom reached by adult male toads in nature in Denmark. Several other experiments made in this laboratory have confirmed that adult toads fed ad lib. tend to strongly reduce or discontinue eating after about a month. It is thus indi- cated that adult toads practically stop feeding when their energy depots have reached high levels.
Spernza t ogenesis At the start of the experiment the spermatogenetic condition in the seminiferous tubules
was normal for toads late in the hibernating period, prior to breeding in April, with an accumulation of early and final stages in spermatogenesis and lack of intermediate stages (Jrargensen et a/ . , 1979). Ten weeks later all spermatogenetic stages were present, but the frequency distribution of cysts of various stages differed between starved and fed toads. The late spermatogenetic stages were present in about equal numbers in all toads, whereas the number of cysts in early stages were lower in the starved toads. It has been found by means of injection of [3H]thymidine in male Bufo bufo kept at 22°C that two weeks after the pulse labelling of spermatogonial cysts the label had reached the meiotic stage of primary spermatocytes. After four weeks spermatid stages were labelled besides many cysts of spermatocytes. After six weeks only spermatid and spermatozoan stages were labelled, and after eight weeks only spermatozoan bundles (unpublished results). At high temperatures spermatid cysts thus result from spermatogenetic activity, i.e. production of secondary spermatogonia and their mitotic divisions, that took place one month or more previously. It is therefore indicated that spermatogenetic activity was about the same in starved and fed toads during the first half or more of the experiment. Moreover, even

N U T R I T I O N A L STATE A N D TESTES F U N C T I O N I N T O A D S 153
10 weeks of starvation had little effect on the late spermatogenetic stages, i.e. spermio- genesis. However, the prolonged starvation resulted in reduced spermatogenetic activity, as evidenced by the low numbers of secondary spermatogonial cysts in the starved toads at the end of the experiment. It thus seems that severe starvation is required to seriously reduce the spermatogenetic activity in toads. The secondary spermatogonial stage is the gonadotropin-dependent stage in spermatogenesis (Guha & Jerrgensen, 1978). Starvation may therefore depress spermatogenetic activity by reducing the rate of gonadotropin secretion from the pituitary.
Interstitiurn and thumbpads After the breeding period in spring the interstitium and thumbpads undergo atrophy
(Jerrgensen et al., 1979). The present experiment showed that in starved toads the inter- stitium and the thumbpads remained reduced, whereas they recovered in fed toads. The interstitium and thumbpads tended to become more developed in the ad lib. fed toads than in those on restricted food.
Presumably, thumbpad development depends upon gonadotropin-stimulated androgen secretion from the interstitial cells. It seems that the interstitial tissue is more affected by the reduced secretion of gonadotropin that results from starvation than is the spermato- genetic activity. This agrees with the findings that ectopic grafting of the pituitary is compatible with maintained spermatogenetic activity, whereas the interstitial tissue and thumbpads atrophy (van Dongen et al., 1966).
Comparison with ovary In the ovary of female toads starvation results in atresia of the vitellogenetic oocytes
(Jmgensen et al., 1979), in agreement with the high levels of gonadotropin required for oocyte growth and maintenance (van Dongen et al., 1966). Apparently, therefore, the two sexes differ in their requirements for gonadotropin to support normal gametogenesis. This difference is reflected in the finding that after breeding in spring the next gameto- genetic wave proceeds without delay in the male toads, but awaits improvement of the nutritional condition in the female, as does restoration of the secondary sex characters, the thumbpads, in the male (Jerrgensen et al., 1979).
Summary Temperate zone anurans exhibit annual gonadal cycles that are correlated with cycles
in nutritional state. The nutritional state again depends upon availability of food and pattern of feeding. In spent female toads, Bgfo bufo bufo (L.) , a new ovarian cycle is only initiated when a satisfactory nutritional state has become reestablished. In male toads initiation of a new spermatogenetic cycle seems less dependent upon the nutritional state, but little is known about the effects of starvation or feeding on function of the testes in the toad, or other amphibians. We therefore assessed the functional state of the testes in male toads emerging from simulated hibernation in a refrigerator, and kept at room temperature, starved or fed at two levels, representing a restricted food regime and ad lib. feeding. Moreover, the pattern of food intake was studied at the two levels of feeding.
When transferred to 22-23°C after a six months’ stay at 5°C in the refrigerator the toads increased food intake for about four weeks at both feeding levels, After this initial

154 K . G U H A , C. B A R K E R J 0 R G E N S E N A N D L. O L E S E N L A R S E N
period the toads on restricted food ate their ration for the remaining part of the experi- ment, and they developed fat bodies of sizes characteristic of male toads during late summer in nature. The toads fed ud lib. increased food intake to about twice that of toads on restricted food. After two weeks of high feeding rate food intake rapidly declined and remained low during the last month of the experiment. At autopsy the fat bodies were larger than normally seen in nature. It is indicated that adult toads regulate their energy balance by practically ceasing to eat when their depots have reached high levels.
At the end of the experiment spermatogenetic activity had declined to a low level in the starving toads, whereas it was high in the fed toads. Starvation for 10 weeks did not affect the later stages in spermatogenesis. Severe starvation was therefore required to seriously reduce spermatogenetic activity, as reflected in the low numbers of secondary spermatogonial cysts within the seminiferous tubules. The secondary spermatogonial stage is the gonadotropin-dependent stage in spermatogenesis in the toad. Lt is therefore suggested that starvation depress spermatogenetic activity by reducing the rate of gonado- tropin secretion from the pituitary gland.
Interstitial cells and thumbpads remained reduced in the starving toads, but were restored in the fed toads. The interstitium and thumbpads tended to become more developed in the od lib. fed toads than in those on restricted food. It is suggested that interstitial tissue is more affected by a reduced secretion of gonadotropin than is the s per ma t ogenet ic activity .
R E F E R E N C E S
Adanis, A. E. & Rae, E. (1929). An experimental study of the fat-bodies in Trirrrrrrs (DiernjY't>.lris) viridcscwrs.
Aron, M. (1926). Recherches morphologiques et exp6rimentales sur le determinisme des characttires sexuels secondaires miles chez les anoures. A r c h Biol. 36: 1-97.
Champy. C. (1921). Sur les correlations entre les characteres sexuels miles et les divers elements du testicule chez les Arnphibiens (Etude sur Triron ulpes/ri.s). C.r . hebd. SParrc. Acod. Sci., Puris 172: 482-484.
van Dongen, W. J.. Jorgensen, C. B., Larsen, L. O., Rosenkilde, P., Lofts, B. & van Oordt, P. G. W. J . (1966). Function and cytology of the normal and autotransplanted pars distalis of the hypophysis in the toad B i / b bu/h (L.). Gen. c o t p . Errdocr. 6: 491-518.
Ciuha, K. K. & Jorgensen. C. B. (1978). Effects of hypophysectoniy on structure and function of testis i n adult toads, Eu/b hrrfi, brtfo (L.). Gerr. conrp. E d o c r . 34: 201-210.
Heusser, H. (1969). Die Lebensweise der Erdkrote, Bujb bufo (L.): Nahrungsaufnahme und Pigmentierung der Daumencchwielen ini Jahreslauf. Biol. Zbl. 88: 457-467.
ItTt. J . D. (1942). The effect of environmental factors on the sperm cycle of Trirrirns virid~~sccws. Biol. Bull. mar'. hiol. Lobs Woods Hole 83: 1 I I - I 18.
Jsrgensen, C . B., Larsen, L. 0. & Lofts, B. (1979). Annual cycles of fat bodies and gonads in the toad Brifo hrc/i>
Kehl, R. (1944). Etudes de quelques problemes d'endocrinologie genitale chez u n batracien nord-africain (Disco-
Nussbaum, M. (19OGu). Ober den Einfluss der Jahreszeit, des Alters und der Ernahrung auf die Form der Hoden
Nussbaum. M. (1906h). Fortgesetzte Untersuchungen uber den Einfluss des Hungers auf die Entwicklung der
011, M. D. (1924). Changes in the weights of the various organs and parts of the leopard frog (Rurrc pipicm) at
Penhos, J . C. & Cardeza, A. F. (1950). lnfluencia del ayuno y alimentacion forzada en el sapo W?/o urc/r(rrr/iu.
AM^. R(.(.. 41 : 1 8 I -203.
brifi) (L. ) , compared with cycles in other temperate zone anurans. Biol. Skr. 22 (5 ) : 1-37,
glosse). Revue c m . Biol. 3: 29-95.
und Hodenzellen der Batrachier. Arch. mikr. Aimt. 68: 1-121.
miinnlichen Geschlechtsorgane der R u m fuscn. Anal. Arrz. 29: 3 15-316.
different stages of inanition. Am. J . A m / . 33: 17-56.
macho. Revto Sot. urgent. Biol. 26: 239-246.

N U T R I T I O N A L S T A T E A N D TESTES F U N C T I O N I N T O A D S 155
Rastogi, R. K. (1976). Seasonal cycle in anuran (Amphibia) testis: the endocrine and environmental controls.
Sluiter, J. W., van Oordt, G. J. &Grasveld, M. S. (1950). Spermatogenesis in normal and hypophysectomized frogs Boll. ZOO^. 43: 151-172.
( R u m temporaria), following gonadotrophin administration. I. Experiments with spring and summer frogs. Acta Endocr. 4: 1-15.
Zdnder, J. (1951). Uber die Grenzen des Schwangerschaftstestes nach Houssay auf Grund des periodischen Ablaufs der Spermiogenese bei der Erdkrote. Geburrsh. Fruuenheilk. 11: 61M22.


















