
Regulation ofAlkane Oxidation in Pseudomonas putida · whole-cell oxidation measure the total...
11
JOURNAL OF BACTERIOLOGY, Aug. 1975, p. 546-556 Copyright © 1975 American Society for Microbiology Vol. 123, No. 2 Printed in U.S.A. Regulation of Alkane Oxidation in Pseudomonas putida ALAN GRUND, JAMES SHAPIRO,* MICHAEL FENNEWALD, PATRICIA BACHA, JOHN LEAHY, KEITH MARKBREITER, MATTHEW NIEDER, AND MELANY TOEPFER Department of Microbiology* and Collegiate Division of Biological Sciences, University of Chicago, Chicago, Illinois 60637 Received for publication 18 April 1975 We have studied the appearance of whole-cell oxidizing activity for n-alkanes and their oxidation products in strains of Pseudomonas putida carrying the OCT plasmid. Our results indicate that the OCT plasmid codes for inducible alkane-hydroxylating and primary alcohol-dehydrogenating activities and that the chromosome codes for constitutive oxidizing activities for primary alcohols, aliphatic aldehydes, and fatty acids. Mutant isolation confirms the presence of an alcohol dehydrogenase locus on the OCT plasmid and indicates the presence of multiple alcohol and aldehyde dehydrogenase loci on the P. putida chromo- some. Induction tests with various compounds indicate that inducer recognition has specificity for chain length and can be affected by the degree of' oxidation of' the carbon chain. Some inducers are neither growth nor respiration substrates. Growth tests with and without a gratuitous inducer indicate that undecane is not a growth substrate because it does not induce alkane hydroxylase activity. Using a growth test for determining induction of the plasmid alcohol dehydrogenase it is possible to show that heptane induces this activity in hydroxylase-negative mutants. This suggests that unoxidized alkane molecules are the physiological inducers of both plasmid activities. Several strains of' Pseudomonas are capable of growth on n-alkanes and oxidize these com- pounds by the following pathway: R-CH,3 - R-CH2OH - R-CHO - R-COOH - beta oxida- tion (1, 7, 10, 16-18). Pseudomonas putida strains grow on n-alkanes of 6 to 10 carbon atoms by virtue of the transmissible OCT plasmid (2, 5, 10). This substrate range is strain specific and is due to the capacity of OCT+ P. putida strains to hydroxylate these alkanes to the corresponding primary alcohols (10). Whole-cell alkane-oxidizing activity is induc- ible in P. aeruginosa strains (1, 16) as well as in other alkane-utilizing bacterial species (11). In P. aeruginosa 473 the effective inducers include alkane growth substrates, aliphatic diols, straight-chain diethers, and dicyclopropyl com- pounds (20, 22). There has, however, been no detailed analysis of' the induction of alkane-oxi- dizing activity in P. putida strains. This paper reports the results of a series of physiological and genetic experiments with al- kane-utilizing P. putida strains designed to elucidate the nature of OCT plasmid-coded oxidizing activities, identify the inducers of these activities, and define the role that induc- tion plays in determining growth on alkanes and their oxidation products. MATERIALS AND METHODS Bacterial strains. The Pseudomonas strains used are listed in Table 1. Microbiological methods. Media, culture condi- tions, growth tests, and mutagenesis procedures are as described by Nieder and Shapiro (10). Chemicals. These are generally as described by Nieder and Shapiro (10). n-Heptane from Fisher Chemical Co. ("Spectranalyzed") was used as a carbon source in some experiments, but induction experiments were performed with n-heptane (99.9%) from Chemical Samples Co. Dicyclopropyl ketone (DCPK) and dicyclopropryl methanol (DCPM) were purchased from K and K Laboratories, Inc. Rifampin and chloramphenicol were obtained from Sigma Chemical Co. [1-14C]Nonane (3.67 mCi/mmol) was purchased from ICN Pharmaceuticals, Inc. and di- luted in cold nonane (99.9%) from Chemical Samples Co. Silicic acid (silicAR cc-7) was purchased from Mallinckrodt. Silicic acid chromatography performed af'ter the experiments reported here revealed that approxi- mately 3% of the radioactivity in our labeled nonane is retained on the column after elution with petroleum ether. These counts could be eluted with methanol and presumably represent contamination by oxidized substances. It is possible that some of the background level of nonane-oxidizing activity detected in unin- duced and OCT cultures is due to the oxidation of' this contaminating material. 546 on February 27, 2020 by guest http://jb.asm.org/ Downloaded from
Transcript of Regulation ofAlkane Oxidation in Pseudomonas putida · whole-cell oxidation measure the total...

JOURNAL OF BACTERIOLOGY, Aug. 1975, p. 546-556Copyright © 1975 American Society for Microbiology
Vol. 123, No. 2Printed in U.S.A.
Regulation of Alkane Oxidation in Pseudomonas putidaALAN GRUND, JAMES SHAPIRO,* MICHAEL FENNEWALD, PATRICIA BACHA, JOHN LEAHY,
KEITH MARKBREITER, MATTHEW NIEDER, AND MELANY TOEPFERDepartment of Microbiology* and Collegiate Division of Biological Sciences, University of Chicago,
Chicago, Illinois 60637
Received for publication 18 April 1975
We have studied the appearance of whole-cell oxidizing activity for n-alkanesand their oxidation products in strains of Pseudomonas putida carrying the OCTplasmid. Our results indicate that the OCT plasmid codes for induciblealkane-hydroxylating and primary alcohol-dehydrogenating activities and thatthe chromosome codes for constitutive oxidizing activities for primary alcohols,aliphatic aldehydes, and fatty acids. Mutant isolation confirms the presence ofan alcohol dehydrogenase locus on the OCT plasmid and indicates the presenceof multiple alcohol and aldehyde dehydrogenase loci on the P. putida chromo-some. Induction tests with various compounds indicate that inducer recognitionhas specificity for chain length and can be affected by the degree of' oxidation of'the carbon chain. Some inducers are neither growth nor respiration substrates.Growth tests with and without a gratuitous inducer indicate that undecane is nota growth substrate because it does not induce alkane hydroxylase activity. Usinga growth test for determining induction of the plasmid alcohol dehydrogenase itis possible to show that heptane induces this activity in hydroxylase-negativemutants. This suggests that unoxidized alkane molecules are the physiologicalinducers of both plasmid activities.
Several strains of' Pseudomonas are capableof growth on n-alkanes and oxidize these com-pounds by the following pathway: R-CH,3 -R-CH2OH - R-CHO - R-COOH - beta oxida-tion (1, 7, 10, 16-18). Pseudomonas putidastrains grow on n-alkanes of 6 to 10 carbonatoms by virtue of the transmissible OCTplasmid (2, 5, 10). This substrate range is strainspecific and is due to the capacity of OCT+ P.putida strains to hydroxylate these alkanes tothe corresponding primary alcohols (10).
Whole-cell alkane-oxidizing activity is induc-ible in P. aeruginosa strains (1, 16) as well as inother alkane-utilizing bacterial species (11). InP. aeruginosa 473 the effective inducers includealkane growth substrates, aliphatic diols,straight-chain diethers, and dicyclopropyl com-pounds (20, 22). There has, however, been nodetailed analysis of' the induction of alkane-oxi-dizing activity in P. putida strains.
This paper reports the results of a series ofphysiological and genetic experiments with al-kane-utilizing P. putida strains designed toelucidate the nature of OCT plasmid-codedoxidizing activities, identify the inducers ofthese activities, and define the role that induc-tion plays in determining growth on alkanes andtheir oxidation products.
MATERIALS AND METHODS
Bacterial strains. The Pseudomonas strains usedare listed in Table 1.
Microbiological methods. Media, culture condi-tions, growth tests, and mutagenesis procedures are asdescribed by Nieder and Shapiro (10).
Chemicals. These are generally as described byNieder and Shapiro (10). n-Heptane from FisherChemical Co. ("Spectranalyzed") was used as acarbon source in some experiments, but inductionexperiments were performed with n-heptane (99.9%)from Chemical Samples Co. Dicyclopropyl ketone(DCPK) and dicyclopropryl methanol (DCPM) werepurchased from K and K Laboratories, Inc. Rifampinand chloramphenicol were obtained from SigmaChemical Co. [1-14C]Nonane (3.67 mCi/mmol) waspurchased from ICN Pharmaceuticals, Inc. and di-luted in cold nonane (99.9%) from Chemical SamplesCo. Silicic acid (silicAR cc-7) was purchased fromMallinckrodt.
Silicic acid chromatography performed af'ter theexperiments reported here revealed that approxi-mately 3% of the radioactivity in our labeled nonane isretained on the column after elution with petroleumether. These counts could be eluted with methanoland presumably represent contamination by oxidizedsubstances. It is possible that some of the backgroundlevel of nonane-oxidizing activity detected in unin-duced and OCT cultures is due to the oxidation of'this contaminating material.
546
on February 27, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

PSEUDOMONAS ALKANE REGULATION 547
TABLE 1. Bacterial strainsa
Strain Genotype Source
P. aeruginosaPAS48 PAO leu- (CAM-OCT) A. Chakrabarty
P. putidaPpG1 Wild type (no plasmid) A. ChakrabartyPpG6 Wild type (OCT+) I. C. GunsalusPpG1737 alcA - str I. C. GunsalusPpS4 OCT- (PpG6 cured by mitomycin) I. C. GunsalusPpS5 met- (OCT+) A. ChakrabartyPpS69 alcA -ald -str NTG mutant of PpG1737PpS70 alcA - str (OCT+) PpS5 x PpG1737PpS76 alcA - NTG mutant of PpG1PpS81 alcA - NTG mutant of PpG1PpS84 alcA str (OCT alk- alcO+) NTG mutant of PpS70PpS88 alcA - str (OCT alk- alcO+) NTG mutant of PpS70PpS124 alcA+ str+ (CAM-OCT+) PAS48 x PpG1PpS125 alcA str+ (CAM-OCT+) PAS48 x PpS76PpS126 alcA- str+ (CAM-OCT+) PAS48 x PpS81PpS181 alcA str+ (CAM-OCT alk alcO+) NTG mutant of PpS125PpS191 alcA str+ (CAM-OCT alk alcO+) j NTG mutant of PpS126
a OCT and CAM-OCT are, respectively, the wild-type octane plasmid (5) and the fused plasmid (4) carryingthe replication, transfer, and camphor utilization loci of the P. putida CAM plasmid (13) and the alkaneutilization loci of the OCT plasmid (5). The genotypic symbols are explained in the legend to Table 9. We havedesignated the chromosomal locus for octanol utilization alcA; this has been previously referred to as ocl or ool(5).
Assay of whole-cell oxidizing activity. Warburgmeasurements were performed as described by Um-breit et al. (19). The flasks were incubated at 30.5 Cand contained: 1 ml of bacteria suspended in PA saltsplus 200 Asg of chloramphenicol per ml to preventenzyme synthesis during the measurements, 1 ml ofsubstrate, and 0.2 ml of 20% KOH in the center well.Substrates were either dissolved in PA salts at theconcentrations indicated or suspended in PA salts bysonication according to Robinson (14). High-vacuumsilicone grease (Dow) was used to lubricate all jointsbecause lanolin absorbs alkane vapors (14). For mea-surements with volatile substrates, we found it neces-sary to use a barometer containing 1 ml of substrateand 1 ml of PA salts in place of the bacterialsuspension. All QO2 values given are corrected forendogenous respiration.A more convenient and sensitive method for de-
terming whole-cell alkane oxidation is based on theevolution of 14CO2 from labeled nonane. One milliliterof cell suspension was incubated for 60 min at 32 Cwith gentle agitation in a 50-ml Erlenmeyer flaskcontaining 1 ml of a [1-14C Inonane suspension in PAsalts prepared by sonication. The flask was sealedwith a serum vial stopper, and a plastic cup wassuspended from the stopper. The cup contained apleated piece of filter paper saturated with 20% KOH.After the incubation, the assay was terminated byopening the flask, removing the filter paper, andmeasuring the "4CO2 evolved by counting the filterpaper in a Packard Tri-Carb scintillation counter.Residual KOH solution in the cups was soaked upwith a second piece of filter paper and counted
together with the first. The quenching for all sampleswas virtually the same. The alkalinity of the samplecauses considerable chemiluminescence of the scintil-lation fluid, and the vials must be kept dark andallowed to equilibrate for at least 1 h before counting.Activity is expressed as counts/min of 14CO, detectedper milligram (dry weight) of cells. The backgroundradioactivity due to nonane vapors adsorbing to thefilter paper was measured by incubating a flaskwithout bacteria; this was generally less than 100counts/min and has been subtracted from all thevalues given below.
Preliminary experiments indicated that the radi-oassay is linear with respect to bacterial concentra-tion, time, and substrate concentration in the range of0.005 to 0.05%. The experiments presented below wereall performed with a final concentration of 0.05%[1-"'C]nonane (5.63 umol/flask) at a specific activityof 100 MCi of nonane per ml.
In all experiments dry weight of the cells wascalculated from optical density measurements in aKlett colorimeter which had previously been cali-brated.
Induction procedure. In most of the experimentsreported here, cells were exposed to inducer duringexponential growth in PA salts plus carbon source at32 C on a rotary shaker with vigorous aeration.Inducing compounds of low solubility were dispensedin a small volume of PA salts by sonication beforebeing added to the culture. At the appropriate time,cell samples were withdrawn, chloramphenicol (200jtg/ml) was added to prevent further protein synthe-sis, the samples were centrifuged at low speed, and
VOL. 123, 1975
on February 27, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
548 GRUND ET AL.
the pellet was suspended in PA salts plus chloram-phenicol (200 qg/ml). In several of the experiments,the cells were suspended in PA salts lacking NH4Cl.
For the experiments reported in Table 2, cells weregrown overnight at 32 C as confluent lawns on PAagar with 0.4% glucose incorporated in the agar oralkanes were added in the vapor phase.
RESULTSInducibility of whole-cell alkane-oxidizing
activity. Growth of P. putida PpG6 on purifiedn-alkanes results in the appearance of alkane-oxidizing activity which is not found in glucose-grown cells (Table 2). Growth on a single alkanesubstrate (octane) induces oxidizing activity forall five alkanes which support the growth of thisstrain (experiment 13). This indicates a singleinducible enzyme or group of enzymes for theoxidation of all alkane growth substrates. Thisconclusion agrees with the genetic data ofNieder and Shapiro (10), which show that strainPpG6 has a single alkane hydroxylase activity.The fact that growth on or exposure to heptane,octane, or decane induces this activity (Table 2)further indicates that there is a single inductionmechanism which responds to any one of thealkane growth substrates. Thus, it appears thatany one of these alkanes is equivalent to anyother both as inducer and as oxidation sub-strate.
Induction of alkane-oxidizing activity ingrowing cultures. Van Eyk and Bartels (22)had demonstrated that cultures of Pseudomonasaeruginosa 473 growing in enriched minimal
TABLE 2. Inducibility of alkane-oxidizing activity inP. putida PpG6a
Growth QO2Expt substrate Hex- Hep- Oc- No- De-
ane tane tane nane cane
4 Glucose 5Octane 172
6 Glucose 0Heptane 97
13 Octane 131 104 146 138 92
15" None 5Decane 156 132 48
a Cultures of strain PpG6 were grown on -solidmedium and assayed as described in Materials andMethods. Alkane substrates in the Warburg assaywere present at a final concentration of 0.5%.
"In experiment 15, glucose-grown cultures werestarved with aeration in PA salts for 4 h at 32 C withor without 0.4% decane before washing and assay.
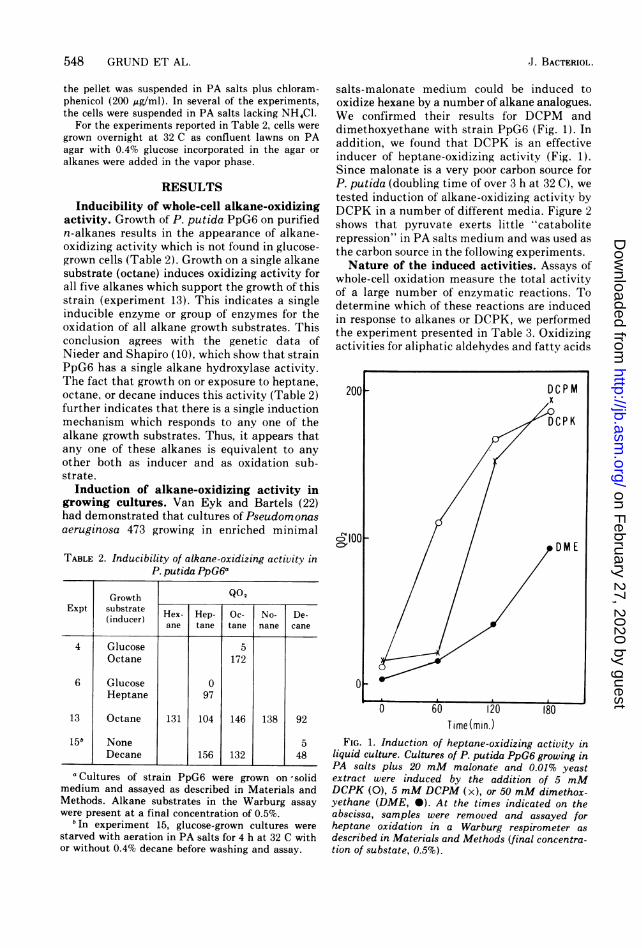
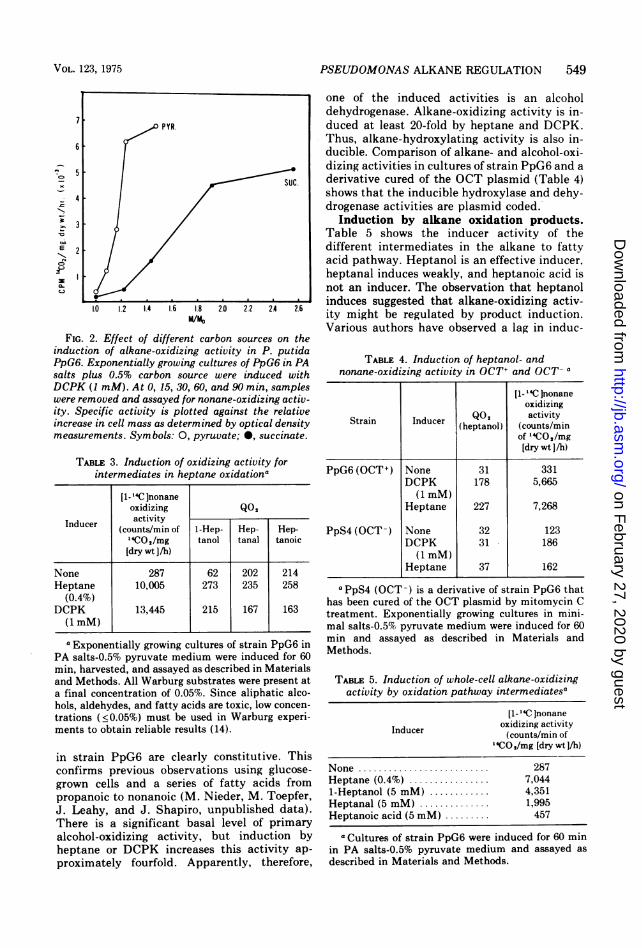
salts-malonate medium could be induced tooxidize hexane by a number of alkane analogues.We confirmed their results for DCPM anddimethoxyethane with strain PpG6 (Fig. 1). Inaddition, we found that DCPK is an effectiveinducer of heptane-oxidizing activity (Fig. 1).Since malonate is a very poor carbon source forP. putida (doubling time of over 3 h at 32 C), wetested induction of alkane-oxidizing activity byDCPK in a number of different media. Figure 2shows that pyruvate exerts little "cataboliterepression" in PA salts medium and was used asthe carbon source in the following experiments.Nature of the induced activities. Assays of
whole-cell oxidation measure the total activityof a large number of enzymatic reactions. Todetermine which of these reactions are inducedin response to alkanes or DCPK, we performedthe experiment presented in Table 3. Oxidizingactivities for aliphatic aldehydes and fatty acids
200
100o[
0
0 60 120Time (min.)
180
FIG. 1. Induction of heptane-oxidizing activity inliquid culture. Cultures of P. putida PpG6 growing inPA salts plus 20 mM malonate and 0.01% yeastextract were induced by the addition of 5 mMDCPK (0), 5 mM DCPM (x), or 50 mM dimethox-yethane (DME, 0). At the times indicated on theabscissa, samples were removed and assayed forheptane oxidation in a Warburg respirometer as
described in Materials and Methods (final concentra-tion of substate, 0.5%).
D M E
J. BACTERIOL.
on February 27, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
PSEUDOMONAS ALKANE REGULATION 549
5
4-
1.0 1.2 1.4 1.6 1.8 2.0 2.2 24 2.6M/A0
FIG. 2. Effect of different carbon sources on theinduction of alkane-oxidizing activity in P. putidaPpG6. Exponentially growing cultures of PpG6 in PAsalts plus 0.5% carbon source were induced withDCPK (1 mM). At 0, 15, 30, 60, and 90 min, sampleswere removed and assayed for nonane-oxidizing activ-ity. Specific activity is plotted against the relativeincrease in cell mass as determined by optical densitymeasurements. Symbols: 0, pyruvate; 0, succinate.
TABLE 3. Induction of oxidizing activity forintermediates in heptane oxidationa
[1-14C Jnonaneoxidizing Q02
Inducer activity(counts/min of 1-Hep- Hep- Hep-4CO,/mg tanol tanal tanoic
[dry wt l/h)
None 287 62 202 214Heptane 10,005 273 235 258
(0.4%)DCPK 13,445 215 167 163
(1 mM)a Exponentially growing cultures of strain PpG6 in
PA salts-0.5% pyruvate medium were induced for 60min, harvested, and assayed as described in Materialsand Methods. All Warburg substrates were present ata final concentration of 0.05%. Since aliphatic alco-hols, aldehydes, and fatty acids are toxic, low concen-trations (<0.05%) must be used in Warburg experi-ments to obtain reliable results (14).
in strain PpG6 are clearly constitutive. Thisconfirms previous observations using glucose-grown cells and a series of fatty acids frompropanoic to nonanoic (M. Nieder, M. Toepfer,J. Leahy, and J. Shapiro, unpublished data).There is a significant basal level of primaryalcohol-oxidizing activity, but induction byheptane or DCPK increases this activity ap-proximately fourfold. Apparently, therefore,
one of the induced activities is an alcoholdehydrogenase. Alkane-oxidizing activity is in-duced at least 20-fold by heptane and DCPK.Thus, alkane-hydroxylating activity is also in-ducible. Comparison of alkane- and alcohol-oxi-dizing activities in cultures of strain PpG6 and aderivative cured of the OCT plasmid (Table 4)shows that the inducible hydroxylase and dehy-drogenase activities are plasmid coded.
Induction by alkane oxidation products.Table 5 shows the inducer activity of thedifferent intermediates in the alkane to fattyacid pathway. Heptanol is an effective inducer,heptanal induces weakly, and heptanoic acid isnot an inducer. The observation that heptanolinduces suggested that alkane-oxidizing activ-ity might be regulated by product induction.Various authors have observed a lag in induc-
TABLE 4. Induction of heptanol- andnonane-oxidizing activity in OCT+ and OCT- a
[1 14C Inonaneoxidizing
Strain Inducer QO. activity(heptanol) (counts/min
of "'CO/mg[dry wt ]/h)
PpG6 (OCT+) None 31 331DCPK 178 5,665
(1 mM)Heptane 227 7,268
PpS4 (OCT-) None 32 123DCPK 31 186
(1 mM)Heptane 37 162
a PpS4 (OCT-) is a derivative of strain PpG6 thathas been cured of the OCT plasmid by mitomycin Ctreatment. Exponentially growing cultures in mini-mal salts-0.5% pyruvate medium were induced for 60min and assayed as described in Materials andMethods.
TABLE 5. Induction of whole-cell alkane-oxidizingactivity by oxidation pathway intermediatesa
[1- '4C ]nonaneoxidizing activity(counts/mmn of
TCO,/mg [dry wtJ/h)
None ........................ 287Heptane (0.4%) ................ 7,0441-Heptanol (5 mM) ............ 4,351Heptanal (5 mM) .............. 1,995Heptanoic acid (5 mM) ......... 457
a Cultures of strain PpG6 were induced for 60 minin PA salts-0.5% pyruvate medium and assayed asdescribed in Materials and Methods.
VOL. 123, 1975
on February 27, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

550 GRUND ET AL.
tion by alkane substrates (1, 22), and this lagcould be explained as the time needed for basalhydroxylase activity to form sufficient primaryalcohol for induction to occur. Accordingly, wedetermined the kinetics of induction by heptaneand 1-heptanol (Fig. 3). Both compounds dis-play the same 10-min lag. Thus, the observedlag cannot be due to the formation of an alcoholinducer. Further evidence that recognition ofthe primary alcohol inducer does not involve thehydroxylated end of the molecule comes fromthe experiment presented in Table 6,A. Thepresence of terminal or subterminal hydroxylgroups at both ends of a carbon chain effectivelyprevents induction. As the control experimentwith DCPK plus hexanediol demonstrates(Table 6,B), the absence of activity after expo-sure to the diol cannot be due to irreversibleinhibition of hydroxylase activity or cell toxic-ity. Similar results are obtained with hex-anediol concentrations in the range of 1 to 20mM. Thus, it appears that primary alcohols are
effective inducers, because one end of the car-
bon chain mimics the unoxidized alkane mole-
;o
C.')
x
C-
C-.'
Time(min)
FIG. 3. Kinetics of induction of alkane-oxidizingactivity in P. putida PpG6. An exponentially growingculture of strain PpG6 in PA salts-0.5% pyruvatemedium was divided and induced with 0.4% heptane(-) or 5 mM 1-heptanol (0). At the times indicatedon the abscissa, samples were removed and assayedfor nonane-oxidizing activity as described in Mate-rials and Methods.
cule. The induction specificity of P. putidaPpG6 is different from that of P. aeruginosa 473because the latter is inducible by 1,6-hexanediol(20). Table 7 shows that there is a chain-length
TABLE 6. Induction of alkane-oxidizing activity bysix-carbon alcohols and diolsa
[1- 14C jnonaneoxidizing activity(counts/m In of
'4C02/mg [dry wt]/h)
ANone ...................... 2351-Hexanol (1 mM) ........... 4,9331,6-Hexanediol (5 mM) ....... 5412-Hexanol (3 mM) ........... 4,8922,5-Hexanediol (5 mM) ....... 387
B1,6-Hexanediol (5 mM) ....... 437DCPK (1 mM) ............... 5,8551,6-Hexanediol (5 mM)+ DCPK (1 mM) .......... 6,093
aCultures of strain PpG6 were induced for 60 minin PA salts-0.5% pyruvate medium and assayed asdescribed in Materials and Methods.
TABLE 7. Induction of alkane-oxidizing activity byvarious compoundsa
[1- 14C jnonaneoxidizing
Compound Inducer activity(counts/minof 14CO,mg[dry wt h)b
Primary None 388alcohols 1-Propanol (3 mM) 434 (2)
1-Butanol (3 mM) 285 (2)1-Pentanol (3 mM) 4,737 (2)1-Dodecanol (1 mM) 159 (2)
Miscellaneous Diethoxyethane 4,335compounds (10 mM)
DCPK (1 mM) 9,022DCPM (5 mM) 7,0671,7-Octadiene (0.4%) 7,0302-Methylhexane 2,706
(0.4%)3-Hexanol (3 mM) 8,1622-Heptanone (0.4%) 6,2552-Octanone (0.4%) 7,128
Alkanes that n-Undecane (0.4%) 243 (2)are not n-Dodecane (0.4%) 328 (2)growthsubstrates
a Cultures of strain PpG6 were induced for 60 minin PA salts-0.5% pyruvate medium and assayed asdescribed in Materials and Methods.
b The numbers in parentheses indicate the numberof independent determinations averaged to give thevalues presented here.
J. BACTERIOL.
on February 27, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

VPSEUDOMONAS ALKANE REGULATION 551
specificity to induction by primary alcohols;propanol, butanol, and 1-dodecanol do not actas inducers.
Induction by other compounds. In additionto DCPK, DCPM, dimethoxyethane, and pri-mary alcohols, several compounds are effectiveinducers of alkane-oxidizing activity (Table 7,miscellaneous compounds). Many of these in-ducers do not serve as substrates either forgrowth or respiration (Table 8). (The low QO2values observed for DCPK and 2-hexanol areprobably not significant.) Many of these induc-ers, moreover, are toxic to P. putida cells abovea certain concentration, and it is sometimesnecessary to determine the optimum inducingconcentration. Use of too much inducer (e.g., 10mM 1-hexanol, 50 mM DCPM, or 50 mMdiethoxyethane) as well as too little (e.g., 1 mMDCPM or 5 mM diethoxyethane) can lead tofalse-negative results (A. Grund, unpublisheddata). The data in Table 7 show that twon-alkanes which do not serve as growth sub-strates, undecane and dodecane (10), are alsonot inducers of alkane-oxidizing activity.
Induction requires transcription. Van Eykand Bartels (22) have shown that exposure tohexane induces oxidizing activity detectable incrude-cell extracts of P. aeruginosa 473, and we
TABLE 8. Alkane analogues as substrates for growthand respirationa
Ability Ability to serve asto substrate for respiration
Compound support by P. putida PpG6 culturesgrowth of iueP. putidaPpG6 Concn QO, Inducer
DCPK 1 mM 0 None5 mM 0 None1 mM 4.7 DCPK ( mM)5 mM 0 DCPK ( mM)10mM 1.6 DCPK ( mM)
DCPM - 5 mM 0 DCPM (5 mM)Diethoxyethane - 10 mM 33 Diethoxyethane
(10 mM)2-Methylhexane - 0.4% 0 None
0.4% 0 DCPK ( mM)2-Hexanol - 3 mM 1 None
3 mM 4 DCPK ( mM)3-Hexanol - 3 mM 11 None
3 mM 0 DCPK (1 mM)1,7-Octadiene + 0.4% 4 None
0.4% 36 DCPK (1 mM)2-Heptanone _ NT2-Octanone NT
a Growth tests and Warburg assays were performed as
described in Materials and Methods. Induced cultures were
induced for 60 min in PA salts-0.5% pyruvate medium; theinducer used is indicated in the last column. Substrates forthe Warburg assays were added at the concentration indi-cated in the third column. NT, Not tested.
have found that either heptane or DCPK in-duces alkane hydroxylase activity detectable incell-free extracts of strain PpG6 (3). Thus,induction results in enzyme synthesis and is notdue to a permeability change. Figure 4 showsthat rifampin blocks the synthesis of alkane-oxidizing activity. Hence, induction requiresthe synthesis of ribonucleic acid.
Isolation of alkane oxidation pathwaymutants. Our physiological data demonstratedthe existence of inducible plasmid-coded oxidiz-ing activities for alkanes and primary alcoholsand constitutive chromosomally coded oxidiz-ing activities for primary alcohols, aldehydes,and fatty acids (Tables 3 and 4). To confirmthese conclusions, we isolated mutants unableto grow on alkanes or their oxidation productsand then determined the step at which theywere blocked by growth tests. The results for anumber of different strains are summarized in
7
6
C.,
I
2x
C-
C-
b-
EC4
5
4
3
2
0
0 6030Time(min.)
FIG. 4. Effect of rifampin on the induction ofalkane-oxidizing activity. An exponentially growingculture of strain PpG6 in PA salts-0.5% pyruvatemedium was induced with 1 mMDCPK (0). At 0 min(0) 200 ,g of rifampin per ml was added to a portion ofthe culture and incubation was continued. At the
times indicated on the abscissa, samples were re-
moved and assayed for nonane-oxidizing activity.
-Rif
+Rif
VOL. 123, 1975
on February 27, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
552 GRUND ET AL.
Table 9. These data lead to the following con-clusions. (i) The chromosome codes for at leasttwo alcohol dehydrogenase activities; mutantsof strain PpG1 with no plasmid selected for in-ability to grow on octanol or nonanol can stillgrow on shorter-chain alcohols. (ii) The chromo-some codes for two aldehyde dehydrogenaseactivities; mutants of strain PpG1 or PpS70which cannot grow on nonanal are negative onlyfor primary alcohols of seven or more carbonatoms. (iii) The chromosome codes for at leastone activity common to the oxidation of allstraight-chain fatty acids because oic- mutantsare unable to grow on all the alcohols tested. (iv)The plasmid codes for a redundant alcoholdehydrogenase activity because no alcohol-nega-tive mutants were found among mutants se-lected from strain PpG6 for inability to grow onoctane or nonane, but such mutants are readilyisolated from strain PpS70, which carries achromosomal mutation blocking growth on
octanol and nonanol. We have confirmed that atleast one of these mutants is hydroxylasepositive and dehydrogenase negative by crossingthe mutant plasmid to strain PpG1 (alcA+)and obtaining alkane-positive exconjugants,whereas a control cross to strains PpS81 (alcA -)yielded no such exconjugants. (v) The plasmiddoes not code for an aldehyde dehydrogenaseactivity because aldehyde-negative mutants arefound in strain PpS70, which has both the OCTplasmid and the chromosomal loci for aldehydeutilization. This conclusion was confirmed bycrossing the OCT+ met- strain PpS5 with analdehyde-negative OCT- mutant and selectingnonanal-positive prototrophic recombinants;less than 2%c of the recombinants had inheritedthe OCT plasmid, indicating that inheritance ofthe plasmid cannot correct the chromosomalblock in aldehyde utilization. (In contrast, thesame cross with an alcA - recipient strain yieldsmore than 50% OCT+ recombinants when they
TABLE 9. Isolation of P. putida mutants affected in the metabolism of alkanes and their oxidation productsa
Parent PpG1 (WT, OCT-) PpG6 (WT, OCT+) PpS70 (alcA - str, OCT t)
Selected phenotype Octanol-negative Octane- or Heptane-negative____________ ~~~nonane-negative Hpaengtv
Mutant types Parent alcA aldA oic ace- parent alk ace parent alk a1c(l aldA oic
No. of mutantsisolated ineach class 5 2 1 1 6 1 6 7 1 1
Growth on alkanesHexane | + -
Heptane +- - + + -
Octane + - +Nonane +Decane + -
Growth on primaryalcohols
1-Dodecanol + _ +1-Nonanol + + + + + + t1-Octanol + _ _ _ + + + +1-Heptanol + ? _ + ± ±1-Hexanol + + + - _ + ± +1-Pentanol + + + + +± + +1-Butanol + + + _ + t + +1-Propanol + + + + + + + +
Nonanal I- + - -v ± -+ -
Nonanoic acid + + + + + ± + t +Octanoic acid + + + + + -t + +
Glucose + + + + + + L ± + I _ + + +
aMutants were isolated and tested as described previously (10). The phenotypic symbols employed are: alc, growth onprimary alcohols; ald, growth on aliphatic aldehydes; oic, growth on straight-chain fatty acids; ace, growth on acetate; and alk,growth on alkanes. The alcA locus is the chromosomal locus which codes for growth on octanol, nonanol, and dodecanol; this haspreviously been called ocl or ool (5). The alcO locus is the plasmid locus which permits growth on octanol and nonanol in thepresence of an alcA- mutation. Mutants negative on nonanal and primary alcohols of seven or more carbon atoms aredesignated aldA- because we presume that there is at least one other ald locus which codes for growth on shorter aldehydes. Thedata for mutants from strain PpG6 is taken from Nieder and Shapiro (10). WT, Wild type.
J. BACTERIOL.
on February 27, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

PSEUDOMONAS ALKANE REGULATION 553
are selected for growth on nonanol.) (vi) Thespecificity of the plasmid-coded alcohol de-hydrogenase appears to differ from that of thecorresponding chromosomal enzyme. StrainsPpGl and PpG6 with the chromosomal enzyme
are able to grow on 1-dodecanol, whereasstrain PpS70, which has only the plasmidenzyme, is negative on 1-dodecanol. Strainscarrying the fused CAM-OCT plasmid yield thesame mutant types after nitrosoguanidinemutagenesis.Role of induction in determining growth
specificity. P. putida strains carrying the OCTor fused CAM-OCT plasmid cannot grow on
undecane or dodecane but can grow on dodeca-nol if they carry the alcA + allele on the chrom-some (10; Table 9). Thus, the growth-negativephenotype on these alkanes is due to lack ofhydroxylating activity (10). This can be ex-
plained either by the substrate specificity of thehydroxylase enzyme or by lack of induction.Two lines of evidence suggested that lack ofinduction is the correct explanation. First,McKenna and Coon (9) have shown that theOCT-coded alkane hydroxylase has 47% of max-imal activity in vitro with dodecane as sub-strate. Although in vitro and in vivo substratespecificities may differ, this level of activityshould support growth if present in the cells.Second, undecane and dodecane do not act as
inducers of whole-cell alkane-oxidizing activity(Table 7). To show that lack of induction isindeed the reason that OCT+ and CAM-OCT+strains cannot grow on undecane, we performedthe experiment presented in Table 10. Additionof DCPK to the medium permits slow growth on
undecane by OCT+ and CAM-OCT+ strains.(We had previously shown that strains carryingthe fused plasmid are heptane and DCPKinducible for alkane-oxidizing activity.) Thesame experiment performed with dodecane sug-
gests that OCT+ and CAM-OCT+ strains can
grow very slowly on this alkane in the presenceof DCPK; however, the growth is very faint after96 h of incubation, and the plates are difficult toscore reliably.A similar explanation could account for the
inability of alcA- (OCT+) and alcA- (CAM-OCT+) strains to grow on 1-dodecanol (Table9), since this compound is not an inducer ofwhole-cell alkane-oxidizing activity (Table 7).The results in Table 10 show that induction ofthe plasmid alcohol dehydrogenase activity byDCPK does permit growth on dodecanol.
Induction of plasmid alcohol-oxidizing ac-
tivity in hydroxylase-negative mutants. Theresults in Fig. 3 and Table 6 do not provide
TABLE 10. Effect of DCPK on the substrate range ofstrains carrying the CAM-OCTplasmida
Growth on PA agar plus
Strain 1-Dodecanol n-UndecaneDCPKb
None' DCPK None DCPK
PpG6 a1cA+ - + + NT - +(OCT)
PpS124 alcA' - + + + + - +(CAM-OCT)
PpS125 alcA - + + - +(CAM-OCT)
PpS126 alcA _ + + - +(CAM-OCT) __
a The strains were streaked on PA agar and PA agarcontaining 5 x 10-i M DCPK and exposed to sub-strate vapors for 48 h (dodecanol) or 96 h (undecane)at 32 C in sealed tins. Growth was scored by visualinspection: + +, normal dense bacterial growth yield-ing isolated colonies 1 to 2 mm in diameter; +, clearlyvisible growth with microcolonies. Genotypic symbolsare explained in the footnote to Table 9. NT, Nottested.
b Substrate.c Inducer.
evidence for the product induction model andindicate that inducer recognition is not specificfor the hydroxyl group of aliphatic alcohols. Ourresults remain consistent with any of threemodels for the induction of the two plasmid-coded activities: coordinate induction by theinitial substrate (alkane), coordinate inductionby the product (primary alcohol), or sequentialinduction. One prediction unique to the firstmodel is that an alkane inducer should induceplasmid alcohol-oxidizing activity in the ab-sence of hydroxylation to the primary alcohol.The results in Table 10 suggested a method oftesting this prediction. This is to determinewhether alkane induction would permit growthon 1-dodecanol by alk- mutants isolated fromalcA- strains lacking the chromosomal alcoholdehydrogenase activity. Accordingly, we per-formed the growth tests summarized in Table11. The heptane used in this experiment waspurified by passage through a silicic acid col-umn to eliminate any possible contaminationby alkane oxidation products. The results indi-cate that purified heptane can induce the plas-mid alcohol-oxidizing activity in the absence ofhydroxylation and therefore support the modelof coordinate induction by unoxidized alkanesubstrate. A similar result for induction ofalkane hydroxylase proteins is reported in theaccompanying paper (3).
VOL. 123, 1975
on February 27, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
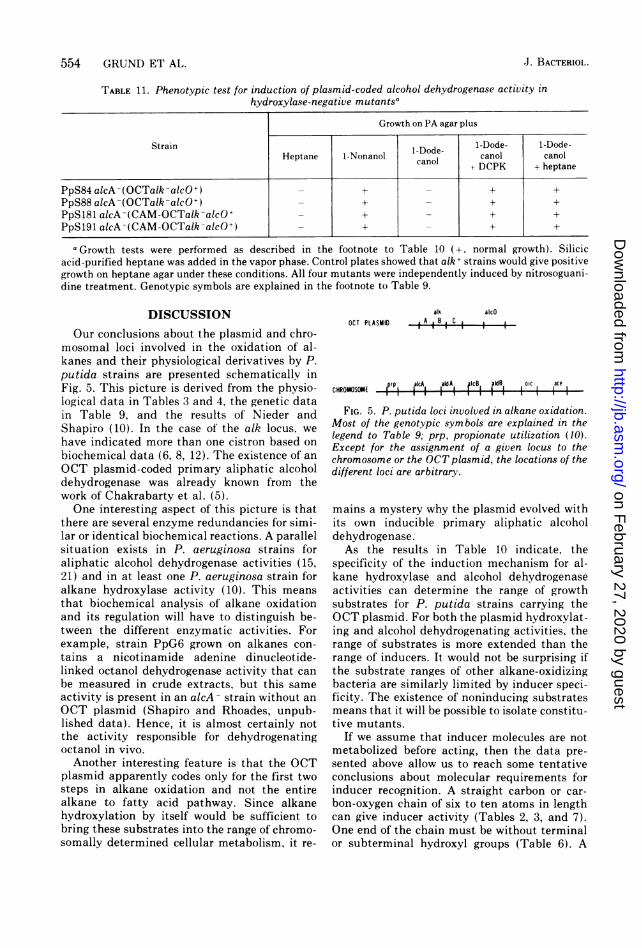
TABLE 11. Phenotypic test for induction of plasmid-coded alcohol dehydrogenase activity inhydroxylase-negative mutantsa
Growth on PA agar plus
Strain 1-Dode- 1-Dode-Heptane 1-Nonanol nDode canol canol
+ DCPK + heptane
PpS84 alcA-(OCTalk-alcO+) - + + +PpS88 alcA-(OCTalk-alc0+) - + _ + +PpS181 alcA-(CAM-OCTalk-alcO+ - + _ + +PpS191 alcA-(CAM-OCTalk-alcO+) - + + +
a Growth tests were performed as described in the footnote to Table 10 (+, normal growth). Silicicacid-purified heptane was added in the vapor phase. Control plates showed that alk + strains would give positivegrowth on heptane agar under these conditions. All four mutants were independently induced by nitrosoguani-dine treatment. Genotypic symbols are explained in the footnote to Table 9.
DISCUSSIONOur conclusions about the plasmid and chro-
mosomal loci involved in the oxidation of al-kanes and their physiological derivatives by P.putida strains are presented schematically inFig. 5. This picture is derived from the physio-logical data in Tables 3 and 4, the genetic datain Table 9, and the results of Nieder andShapiro (10). In the case of the alk locus, wehave indicated more than one cistron based onbiochemical data (6, 8, 12). The existence of anOCT plasmid-coded primary aliphatic alcoholdehydrogenase was already known from thework of Chakrabarty et al. (5).One interesting aspect of this picture is that
there are several enzyme redundancies for simi-lar or identical biochemical reactions. A parallelsituation exists in P. aeruginosa strains foraliphatic alcohol dehydrogenase activities (15,21) and in at least one P. aeruginosa strain foralkane hydroxylase activity (10). This meansthat biochemical analysis of alkane oxidationand its regulation will have to distinguish be-tween the different enzymatic activities. Forexample, strain PpG6 grown on alkanes con-tains a nicotinamide adenine dinucleotide-linked octanol dehydrogenase activity that canbe measured in crude extracts, but this sameactivity is present in an alcA - strain without anOCT plasmid (Shapiro and Rhoades, unpub-lished data). Hence, it is almost certainly notthe activity responsible for dehydrogenatingoctanol in vivo.Another interesting feature is that the OCT
plasmid apparently codes only for the first twosteps in alkane oxidation and not the entirealkane to fatty acid pathway. Since alkanehydroxylation by itself would be sufficient tobring these substrates into the range of chromo-somally determined cellular metabolism, it re-
alkOCT PLASMID I AA B I C
CHROMOSOME
alcO
?pp I lIcA, ?aIdAa fIcB L IdBL oic ace
FIG. 5. P. putida loci involved in alkane oxidation.Most of the genotypic symbols are explained in thelegend to Table 9; prp, propionate utilization (10).Except for the assignment of a given locus to thechromosome or the OCT plasmid, the locations of thedifferent loci are arbitrary.
mains a mystery why the plasmid evolved withits own inducible primary aliphatic alcoholdehydrogenase.As the results in Table 10 indicate, the
specificity of the induction mechanism for al-kane hydroxylase and alcohol dehydrogenaseactivities can determine the range of growthsubstrates for P. putida strains carrying theOCT plasmid. For both the plasmid hydroxylat-ing and alcohol dehydrogenating activities, therange of substrates is more extended than therange of inducers. It would not be surprising ifthe substrate ranges of other alkane-oxidizingbacteria are similarly limited by inducer speci-ficity. The existence of noninducing substratesmeans that it will be possible to isolate constitu-tive mutants.
If we assume that inducer molecules are notmetabolized before acting, then the data pre-sented above allow us to reach some tentativeconclusions about molecular requirements forinducer recognition. A straight carbon or car-bon-oxygen chain of six to ten atoms in lengthcan give inducer activity (Tables 2, 3, and 7).One end of the chain must be without terminalor subterminal hydroxyl groups (Table 6). A
J. BACTERIOL.554 GRUND ET AL.
on February 27, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

PSEUDOMONAS ALKANE REGULATION 555
ketone, hydroxyl, or methyl group on the sub-terminal carbon does not prevent induction,although the latter reduces effectiveness (Ta-bles 6 and 7). An aldehyde group also reducesinducer activity, and a carboxyl group elimi-nates it (Table 5). This may be related to thehigh consitutitive level of aldehyde and fattyacid oxidation (Table 3). From Table 7 it isclear that the length of straight-chain moleculescan determine whether or not they act asinducers. In other words, the inducer recogni-tion site can somehow measure the length of thecarbon chain. This suggests that the recognitionmolecule contains a "pocket" into which bothends of the inducer molecule fit. DCPK andDCPM may act as inducers because they resem-ble alkane chains folded inside this pocket.Our assumption that inducer molecules are
not metabolized before acting remains to berigorously demonstrated. It is based on resultswhich show that some inducers are not growthsubstrates and which indicate that some arealso not respiration substrates (Table 8). TheWarburg experiments are rather imprecise, buteven if our interpretation is correct the resultsdo not preclude metabolic transformationswhich stop before the inducer molecule canenter carbon- or energy-yielding pathways. Theresults in Table 11 indicate that alkanes caninduce the plasmid alcohol-oxidizing activity inthe absence of hydroxylation. Trivial explana-tions for these results (such as a residual level ofhydroxylation insufficent for growth) have notbeen excluded, but the results nonetheless sug-gest that both plasmid activities are undercoordinate control and that the alkane is thephysiological inducer. This is consistent withthe results in Fig. 3 and Table 6, which showthat induction is not alcohol specific, and withthe fact that a molecule like DCPK inducesboth plasmid activities. Induction requirestranscription of ribonucleic acid (Fig. 4) andpresumably involves one or more regulatoryproteins. Conclusive evidence for coordinatecontrol and the elucidation of whether thepostulated regulatory proteins act in a negativeor positive fashion will require the isolation ofconstitutive and noninducible mutants. We arecurrently searching for these mutants.
ACKNOWLEDGMENTSWe wish to thank Wolfgang Epstein, David Rhoades, and
Spencer Benson for helpful discussions and Lucia Rothman-Denes for her many valuable suggestions and for reading themanuscript.
Alan Grund was the recipient of a summer researchstipend from the Collegiate Division of Biological Sciences ofthe University of Chicago. Michael Fennewald is the recipientof a Public Health Service predoctoral traineeship. This work
was begun during an undergraduate course supported by theCollegiate Division of Biological Sciences of the University ofChicago and has been supported in part by grants from theLouis Block Fund of the University of Chicago and theNational Science Foundation.
LITERATURE CITED
1. Azoulay, E., and J. C. Senez. 1960. Degradation bac-terienne des hydrocarbures paraffiniques. II. Determi-nation des produits intermediaires par la methode desadaptations simultanees. Ann. Inst. Pasteur (Paris)98:868-879.
2. Baptist, J. N., R. K. Gholson, and M. J. Coon. 1963.Hydrocarbon oxidation by a bacterial enzyme system.I. Products of octane oxidation. Biochim. Biophys.Acta 69:40-47.
3. Benson, S., and J. Shapiro. 1975. Induction of alkanehydroxylase proteins by unoxidized alkane in Pseudo-monas putida. J. Bacteriol. 123:759-760.
4. Chakrabarty, A. M. 1973. Genetic fusion of incompatibleplasmids in Pseudomonas. Proc. Natl. Acad. Sci.U.S.A. 70:1641-1644.
5. Chakrabarty, A. M., G. Chou, and I. C. Gunsalus. 1973.Genetic regulation of octane dissimilation plasmid inPseudomonas. Proc. Natl. Acad. Sci. U.S.A.70:1137-1140.
6. Gholson, R. K., J. N. Baptist, and M. J. Coon. 1963.Hydrocarbon oxidation by a bacterial enzyme system.II. Cofactor requirements for octanol formation fromoctane. Biochemistry 2:1155-1159.
7. Heringa, J. W., R. Huybregste, and A. C. van der Linden.1961. n-Alkane oxidation by a Pseudomonas. Forma-tion and ,B-oxidation of intermediate fatty acids. Anto-nie van Leeuwenhoek J. Microbiol. Serol. 27:51-58.
8. Kusunose, M., E. Kusunose, and M. J. Coon. 1964.Enzymatic w-oxidation of fatty acids. II. Substratespecificity and other properties of the enzyme system.J. Biol. Chem. 239:2135-2139.
9. McKenna, E. J., and M. J. Coon. 1970. Enzymaticw-oxidation. IV. Purification and properties of thew-hydroxylase of Pseudomonas oleovorans. J. Biol.Chem. 245:3882-3889.
10. Nieder, M., and J. Shapiro. 1975. Physiological functionof the Pseudomonas putida PpG6 (Pseudomonasoleovorans) alkane hydroxylase: monoterminal oxida-tion of alkanes and fatty acids. J. Bacteriol. 122:93-98.
11. Perry, J. J., and H. W. Scheld. 1968. Oxidation ofhydrocarbons by microorganisms isolated from soil.Can. J. Microbiol. 14:403-407.
12. Peterson, J. A., M. Kusunose, E. Kusunose, and M. J.Coon. 1967. Enzymatic w-hydroxylation. J. Biol.Chem. 242:4334-4340.
13. Rheinwald, J. G., A. M. Chakrabarty, and I. C. Gunsalus.1973. A transmissible plasmid controlling camphoroxidation in Pseudomonas putida. Proc. Natl. Acad.Sci. U.S.A. 70:885-889.
14. Robinson, D. S. 1964. Oxidation of selected alkanes andrelated compounds by a Pseudomonas strain. Antonievan Leeuwenhoek J. Microbiol. Serol. 30:303-316.
15. Tassin, J. P., C. Celier, and J. P. Vandecasteele. 1973.Purification and properties of a membrane-bound alco-hol dehydrogenase involved in oxidation of long-chainhydrocarbons by Pseudomonas aeruginosa. Biochim.Biophys. Acta 315:220.
16. Thijsse, G. J. E., and A. C. van der Linden. 1958. n-Alkaneoxidation by a Pseudomonas. Studies on the intermedi-ate metabolism. Antonie van Leeuwenhoek J. Micro-biol. Serol. 24:298-308.
17. Thijsse, G. J. E., and A. C. van der Linden. 1961.Iso-alkane oxidation by a Pseudomonas. I. Metabolismof 2-methylhexane. Antonie van Leeuwenhoek J. Mi-
VOL. 123, 1975
on February 27, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

556 GRUND ET AL.
crobiol. Serol. 27:171-179.18. Thijsse, G. J. E., and A. C. van der Linden. 1963.
Pathways of hydrocarbon dissimilation by a
Pseudomonas as revealed by chloramphenicol. Antonievan Leeuwenhoek J. Microbiol. Serol. 29:89-100.
19. Umbreit, W. W., R. H. Burris, and J. F. Stauffer. 1964.Manometric techniques. Burgess Publishing Co., Min-neapolis.
20. van der Linden, A. C., and R. Huybregste. 1967. Induc-tion of alkane-oxidizing and a-olefin-epoxidizing en-
zymes by a non-hydrocarbon in Pseudomonas. Antonievan Leeuwenhoek J. Microbiol. Serol. 33:381-385.
21. van der Linden, A. C., and R. Huybregste. 1969. Occur-rence of inducible and NAD(P)-independent primaryalcohol dehydrogenases in an alkane-oxidizing Pseudo-monas. Antonie van Leeuwenhoek J. Microbiol. Serol.35:344-360.
22. van Eyk, J., and T. J. Bartels. 1968. Paraffin oxidation inPseudomonas aeruginosa. I. Induction of paraffin oxi-dation. J. Bacteriol. 96:706-712.
J. BACTERIOL.
on February 27, 2020 by guest
http://jb.asm.org/
Dow
nloaded from


















