
Regulation of VIP production and secretion by murine lymphocytes
13
Ž . Journal of Neuroimmunology 93 1999 126–138 Regulation of VIP production and secretion by murine lymphocytes Carmen Martinez a,1 , Mario Delgado a,1 , Catalina Abad a , Rosa P. Gomariz a , Doina Ganea b , Javier Leceta a, ) a Departamento de Biologıa Celular, Facultad de Biologıa, UniÕersidad Complutense, 28040 Madrid, Spain ´ ´ b Department of Biological Sciences, Rutgers UniÕersity, Newark, NJ 07102, USA Received 10 June 1998; revised 17 September 1998; accepted 17 September 1998 Abstract Ž . Vasoactive intestinal peptide VIP is a neuropeptide present in the lymphoid microenvironment with a multiplicity of actions. Two sources for VIP have been described in the immune system, the terminals present in central and peripheral lymphoid organs and the immune cells. Although VIP is synthesized by lymphocytes, there is no evidence demonstrating that VIP is released, and which stimuli are able to induce VIP production and secretion. In this study, we demonstrated for the first time, that agents that mediate important Ž . Ž immune functions, such as proliferation and antigenic stimulation Con A, LPS, and anti-TCR antibody , inflammation LPS, TNFa , IL-6 . Ž . and IL-1b or apoptosis dexamethasone induce the production and release of VIP to the lymphoid microenvironment. We conclude that VIP is produced and secreted by lymphocytes and propose that during an immune response, the timely release of VIP within the lymphoid organs and peritoneum should influence the differentiation andror downregulation of the ongoing response. q 1999 Elsevier Science B.V. All rights reserved. Keywords: Vasoactive intestinal peptide; Lymphocyte; Lymphoid organs; ELISA; Neuroimmunomodulation 1. Introduction The immune and neuroendocrine systems represent an integrated network sharing the same molecular signals and Ž their receptors. In 1980 Blalock and Smith Blalock and . Smith, 1980 demonstrated that, in this network, neurons and endocrine cells were not the exclusive sources of hormones and regulatory peptides, describing for the first time that macrophages and lymphocytes were also sources for ACTH and endorphins. Actually, there are at least 20 different neuroendocrine peptides that can be synthesized Ž by cells of the immune system Blalock, 1992; Dardenne . and Savino, 1994 . Thus, the presence of hormones and Ž . neuropeptides including prolactin O’Neal et al., 1992 Ž .Ž . growth hormone GH Weigent et al., 1988 , GHRH Ž . Ž Weigent and Blalock, 1990 , LHRH Emanuele et al., . Ž . 1990 arginine–vasopressin Geenen et al., 1987 , sub- Ž . Ž stance P Weinstock et al., 1989 , somatostatin Fuller and ) Corresponding author. Tel.: q34-1-3944971; Fax: q34-1-3944981; E-mail: [email protected] 1 The first two authors have contributed in equally form in the present work. . Ž . Verity, 1989 , proenkephalin A Rosen et al., 1989 , nerve Ž . growth factor Santambrogio et al., 1994 and atrial natri- Ž . uretic peptide Vollmar et al., 1990 has been described in different cells of the immune system. Ž . Vasoactive intestinal peptide VIP is a 28 aminoacid Ž peptide isolated in 1970 from pig intestinal extracts Said . and Mutt, 1970 . It is known today that VIP is an ubiqui- tous peptide, with a multiplicity of actions and highly Ž . conserved sequence in vertebrates Dimaline, 1989 . VIP is a signal molecule of the neuroendocrine-immune net- work produced by neurons in different areas of the central and peripheral nervous system and by endocrine cells as the pituitary lactotropes and cells of the endocrine pan- Ž . creas Davidson et al., 1996 . In the immune system, two sources for VIP have been described, the nerve terminals Ž present in central and peripheral lymphoid organs Be- . llinger et al., 1997 , and the inflammatory and immune Ž cells Cutz et al., 1978; Goetzl et al., 1988; Gomariz et al., . 1990; Weinstock and Blum, 1990 . The first identification of VIP immunoreactivity in cells other than neurons was in Ž . peritoneal rat mast cells Cutz et al., 1978 , followed by Ž . eosinophils Weinstock and Blum, 1990 . The first evi- dence that lymphocytes produced VIP was the presence of 0165-5728r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved. Ž . PII: S0165-5728 98 00216-1
-
Upload
carmen-martinez -
Category
Documents
-
view
213 -
download
0
Transcript of Regulation of VIP production and secretion by murine lymphocytes

Ž .Journal of Neuroimmunology 93 1999 126–138
Regulation of VIP production and secretion by murine lymphocytes
Carmen Martinez a,1, Mario Delgado a,1, Catalina Abad a, Rosa P. Gomariz a, Doina Ganea b,Javier Leceta a,)
a Departamento de Biologıa Celular, Facultad de Biologıa, UniÕersidad Complutense, 28040 Madrid, Spain´ ´b Department of Biological Sciences, Rutgers UniÕersity, Newark, NJ 07102, USA
Received 10 June 1998; revised 17 September 1998; accepted 17 September 1998
Abstract
Ž .Vasoactive intestinal peptide VIP is a neuropeptide present in the lymphoid microenvironment with a multiplicity of actions. Twosources for VIP have been described in the immune system, the terminals present in central and peripheral lymphoid organs and theimmune cells. Although VIP is synthesized by lymphocytes, there is no evidence demonstrating that VIP is released, and which stimuliare able to induce VIP production and secretion. In this study, we demonstrated for the first time, that agents that mediate important
Ž . Žimmune functions, such as proliferation and antigenic stimulation Con A, LPS, and anti-TCR antibody , inflammation LPS, TNFa , IL-6. Ž .and IL-1b or apoptosis dexamethasone induce the production and release of VIP to the lymphoid microenvironment. We conclude that
VIP is produced and secreted by lymphocytes and propose that during an immune response, the timely release of VIP within the lymphoidorgans and peritoneum should influence the differentiation andror downregulation of the ongoing response. q 1999 Elsevier ScienceB.V. All rights reserved.
Keywords: Vasoactive intestinal peptide; Lymphocyte; Lymphoid organs; ELISA; Neuroimmunomodulation
1. Introduction
The immune and neuroendocrine systems represent anintegrated network sharing the same molecular signals and
Žtheir receptors. In 1980 Blalock and Smith Blalock and.Smith, 1980 demonstrated that, in this network, neurons
and endocrine cells were not the exclusive sources ofhormones and regulatory peptides, describing for the firsttime that macrophages and lymphocytes were also sourcesfor ACTH and endorphins. Actually, there are at least 20different neuroendocrine peptides that can be synthesized
Žby cells of the immune system Blalock, 1992; Dardenne.and Savino, 1994 . Thus, the presence of hormones and
Ž .neuropeptides including prolactin O’Neal et al., 1992Ž . Ž .growth hormone GH Weigent et al., 1988 , GHRH
Ž . ŽWeigent and Blalock, 1990 , LHRH Emanuele et al.,. Ž .1990 arginine–vasopressin Geenen et al., 1987 , sub-
Ž . Žstance P Weinstock et al., 1989 , somatostatin Fuller and
) Corresponding author. Tel.: q34-1-3944971; Fax: q34-1-3944981;E-mail: [email protected]
1 The first two authors have contributed in equally form in the presentwork.
. Ž .Verity, 1989 , proenkephalin A Rosen et al., 1989 , nerveŽ .growth factor Santambrogio et al., 1994 and atrial natri-Ž .uretic peptide Vollmar et al., 1990 has been described in
different cells of the immune system.Ž .Vasoactive intestinal peptide VIP is a 28 aminoacid
Žpeptide isolated in 1970 from pig intestinal extracts Said.and Mutt, 1970 . It is known today that VIP is an ubiqui-
tous peptide, with a multiplicity of actions and highlyŽ .conserved sequence in vertebrates Dimaline, 1989 . VIP
is a signal molecule of the neuroendocrine-immune net-work produced by neurons in different areas of the centraland peripheral nervous system and by endocrine cells asthe pituitary lactotropes and cells of the endocrine pan-
Ž .creas Davidson et al., 1996 . In the immune system, twosources for VIP have been described, the nerve terminals
Žpresent in central and peripheral lymphoid organs Be-.llinger et al., 1997 , and the inflammatory and immune
Žcells Cutz et al., 1978; Goetzl et al., 1988; Gomariz et al.,.1990; Weinstock and Blum, 1990 . The first identification
of VIP immunoreactivity in cells other than neurons was inŽ .peritoneal rat mast cells Cutz et al., 1978 , followed by
Ž .eosinophils Weinstock and Blum, 1990 . The first evi-dence that lymphocytes produced VIP was the presence of
0165-5728r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved.Ž .PII: S0165-5728 98 00216-1
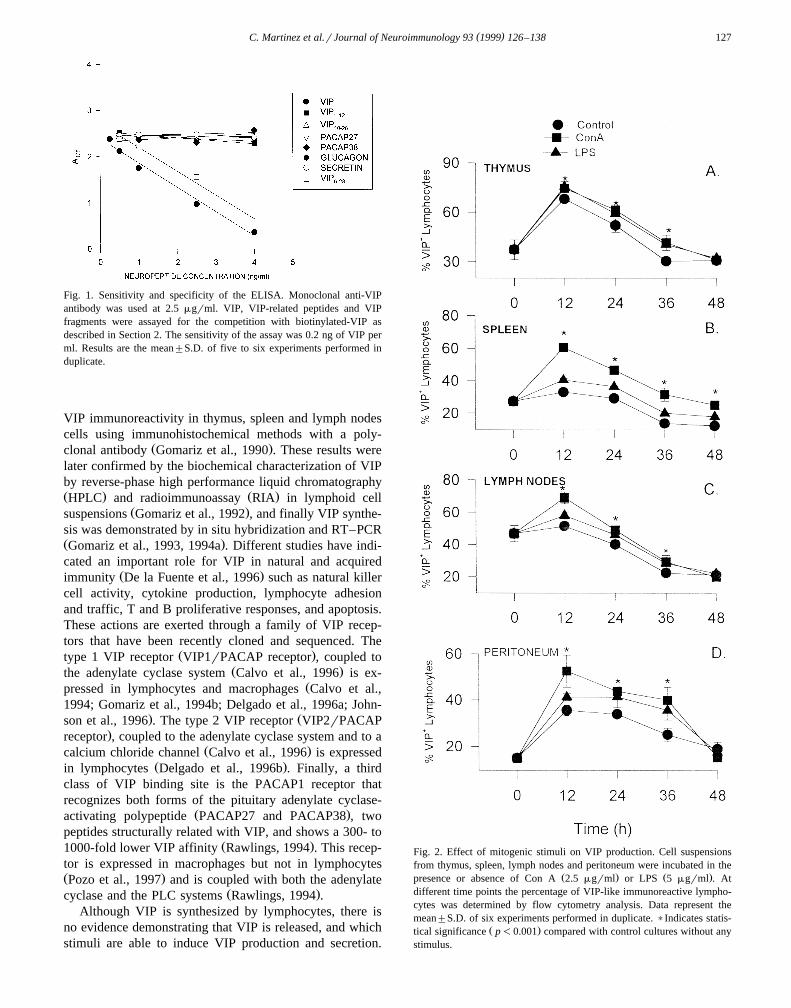
( )C. Martinez et al.rJournal of Neuroimmunology 93 1999 126–138 127
Fig. 1. Sensitivity and specificity of the ELISA. Monoclonal anti-VIPantibody was used at 2.5 mgrml. VIP, VIP-related peptides and VIPfragments were assayed for the competition with biotinylated-VIP asdescribed in Section 2. The sensitivity of the assay was 0.2 ng of VIP perml. Results are the mean"S.D. of five to six experiments performed induplicate.
VIP immunoreactivity in thymus, spleen and lymph nodescells using immunohistochemical methods with a poly-
Ž .clonal antibody Gomariz et al., 1990 . These results werelater confirmed by the biochemical characterization of VIPby reverse-phase high performance liquid chromatographyŽ . Ž .HPLC and radioimmunoassay RIA in lymphoid cell
Ž .suspensions Gomariz et al., 1992 , and finally VIP synthe-sis was demonstrated by in situ hybridization and RT–PCRŽ .Gomariz et al., 1993, 1994a . Different studies have indi-cated an important role for VIP in natural and acquired
Ž .immunity De la Fuente et al., 1996 such as natural killercell activity, cytokine production, lymphocyte adhesionand traffic, T and B proliferative responses, and apoptosis.These actions are exerted through a family of VIP recep-tors that have been recently cloned and sequenced. The
Ž .type 1 VIP receptor VIP1rPACAP receptor , coupled toŽ .the adenylate cyclase system Calvo et al., 1996 is ex-
Žpressed in lymphocytes and macrophages Calvo et al.,1994; Gomariz et al., 1994b; Delgado et al., 1996a; John-
. Žson et al., 1996 . The type 2 VIP receptor VIP2rPACAP.receptor , coupled to the adenylate cyclase system and to a
Ž .calcium chloride channel Calvo et al., 1996 is expressedŽ .in lymphocytes Delgado et al., 1996b . Finally, a third
class of VIP binding site is the PACAP1 receptor thatrecognizes both forms of the pituitary adenylate cyclase-
Ž .activating polypeptide PACAP27 and PACAP38 , twopeptides structurally related with VIP, and shows a 300- to
Ž .1000-fold lower VIP affinity Rawlings, 1994 . This recep-tor is expressed in macrophages but not in lymphocytesŽ .Pozo et al., 1997 and is coupled with both the adenylate
Ž .cyclase and the PLC systems Rawlings, 1994 .Although VIP is synthesized by lymphocytes, there is
no evidence demonstrating that VIP is released, and whichstimuli are able to induce VIP production and secretion.
Fig. 2. Effect of mitogenic stimuli on VIP production. Cell suspensionsfrom thymus, spleen, lymph nodes and peritoneum were incubated in the
Ž . Ž .presence or absence of Con A 2.5 mgrml or LPS 5 mgrml . Atdifferent time points the percentage of VIP-like immunoreactive lympho-cytes was determined by flow cytometry analysis. Data represent themean"S.D. of six experiments performed in duplicate. )Indicates statis-
Ž .tical significance p-0.001 compared with control cultures without anystimulus.

( )C. Martinez et al.rJournal of Neuroimmunology 93 1999 126–138128
Therefore, the present study was designed to determinewhether agents that mediate important immune functions,such as proliferation, inflammation or apoptosis, also in-duce the production and release of VIP to the lymphoidmicroenvironment. Our results demonstrate, for the firsttime, that VIP secretion from lymphocytes of lymphoidorgans and peritoneum is promoted by proliferative stim-uli, proinflammatory cytokines and glucocorticoids.
2. Materials and methods
2.1. Animals
Ž .Adult male Wistar rats Iffa Credo , weighing 200–250Ž .g, were maintained at constant temperature 23"28C on
a 12 h lightrdark cycle and food and water ad libitum.
2.2. Antibodies and reagents
Ž .Monoclonal anti-VIP antibody clone CURE.V55 wasŽa generous gift from Dr. Wong UCLA School of Medicine,
. Ž X.Los Angeles, CA . FITC-labeled F ab fragments of2Ž .sheep anti-mouse IgG was from Sigma St. Louis, MO .
Synthetic VIP, PACAP27, PACAP38, VIP and1 – 12ŽVIP were purchased from Novabiochem Laufelfin-¨10 – 28
.gen, Switzerland , and VIP from Sigma. Secretin,6 – 28
glucagon, and biotinylated-VIP were obtained from Penin-Ž .sula Laboratories Belmont, CA . Murine recombinants
mrIL-1b, mrTNFa , mrIL-6, mrIL-2, and monoclonalanti-murine TNFa and IL-6 antibodies were purchased
Ž . Žfrom PharMingen San Diego, CA . LPS from E. Coli. X Ž0111:B4 , retinal, 2,2 -Azino-bis 3-ethylbenzthiazoline-6-
. Ž . Žsulfonic acid ABTS substrate , Concanavaline A Con.A , forskolin, PMA and avidin-peroxidase were purchased
w Ž . xfrom Sigma and N- 2- p-bromocinnamyl-amino ethyl -5-Ž .iso-quinolinesulfonamide H-89 from ICN Pharmaceuti-
Ž .cals Costa Mesa, CA .
2.3. Preparation of cell suspensions
Each animal was killed by cervical dislocation, theabdomen cleaned with 70% ethanol, the abdominal skincarefully dissected without opening the peritoneum and 10
Žml of RPMI 1640 medium Gibco, BRL Laboratories,.Grand Island, NY injected intraperitoneally. Then the
abdomen was massaged and about 90% of the RPMI
Ž .medium was extracted containing lymphocytes 44% ,Ž . Ž .macrophages 52% , and granulocytes and mast cells 4% .
ŽNext, the thymus, spleen and lymph nodes containing 90,70, 85% lymphocytes and 10, 30, 15% nonlymphoid cells,
.respectively were removed and single-cell suspensionsŽ .were prepared by tapping organs on a wire mesh Sigma .
Contaminating red blood cells from spleen cell suspen-sions were removed by ammonium chloride lysis bufferŽ .NH Cl 0.17 M . Cells were washed twice and resus-4
wpended at the appropriate dilution in R5 medium RPMIŽ1640 medium containing 5% fetal calf serum FCS; Gibco,
.BRL , 100 Urml penicillin, 100 mgrml streptomycinŽ .xGibco, BRL . Cell viability was determined by the trypanblue exclusion test and was always higher than 95% at thetime of culture and 92–94% at the end of the cultureperiod.
2.4. Cell culture and stimulation
ŽCells were cultured in 24-well culture plates Corning. 6Glass, Corning, NY at a concentration of 5=10 cellsrml
Žin R5 medium,. Different stimuli Con A, 2.5 mgrml;LPS, 5 mgrml; anti-TCR, 1 mgrml; mrIL-1b, mrTNFa ,mrIL-2, 10 ngrml; mrIL-6, 20 ngrml; anti-IL-6, anti-TNFa , 50 mgrml; VIP 10y8 ; dexamethasone, forskolin,
.H-89, retinal, at various concentrations were added first,followed by the cell suspension. Controls with mediumalone were included. The culture plates were maintained at378C in a humidified incubator with 5% CO . At the2
indicated time points, cells incubated with the designatedstimuli were harvested and processed for immunofluo-rescence staining. Cell-free supernatants were harvested
Ž .when indicated and kept frozen y208C until assayed forVIP production by ELISA.
2.5. Cell staining procedure
Single staining with monoclonal antibody raised againstŽ 6VIP was performed as follows. Briefly, cells 1=10 per
.sample were fixed for 1 h at 48C with 1% paraformal-Ž . Ž .dehyde in phosphate buffer solution PBS pH 7.4 . After
washing, cells were treated with normal sheep serum for30 min to reduce nonspecific staining and incubated 1 h at
Ž48C with the monoclonal anti-VIP antibody clone. Ž . Ž X .CURE.V55 1:1000 followed by FITC-labeled F ab 2
Žfragments of sheep anti-mouse IgG 1:100 dilution in PBS.containing 1% normal rat serum for 45 min at 48C.
Ž .Fig. 3. page 129 Stimulation of VIP expression in lymphocytes. Cell suspensions from thymus, spleen, lymph nodes and peritoneum were cultured in theŽ . Ž .presence or absence of the mitogens Con A 2.5 mgrml or LPS 5 mgrml . Cells were collected at different time points and processed for
Ž .immunofluorescence staining by using an anti-VIP monoclonal antibody. A Histograms show the mean of fluorescence intensity for unstimulated andmitogen stimulated cultures, using as negative control the anti-VIP monoclonal antibody absorbed with 10 mM of VIP, from one representative experiment
Ž . Ž .out of six. B Differences in fluorescence units FU between control and experimental cultures after 12 h of incubation. Data represent the mean"S.D.of six experiments performed in duplicate.

( )C. Martinez et al.rJournal of Neuroimmunology 93 1999 126–138 129

( )C. Martinez et al.rJournal of Neuroimmunology 93 1999 126–138130
Negative controls were performed to verify the specificityof the staining by preabsorbing the VIP monoclonal anti-body overnight at 48C with 10 mM of VIP, as well asusing PBS instead of the primary antiserum.
2.6. Flow cytometry
Flow cytometry was performed after single immuno-fluorescence staining with a FACScan flow cytometerŽ .Becton-Dickinson, San Jose, CA . Lymphocytes wereelectronically gated and scored by their forward scatterŽ . Ž .FS vs. side scatter SS profiles. Samples in which theprimary antibody was omitted were used as negative con-trols to determine the proper region or window setting.
2.7. Assay for VIP production
The amount of VIP present in the supernatants wasdetermined by using a competitive ELISA. Spleen, thy-
Ž 6.mus, lymph node and peritoneal cell suspensions 5=10were incubated with different stimuli as indicated above.
Ž .At different times, the supernatants 1 ml were collectedand lyophilized. A capture monoclonal anti-VIP antibodyŽ .clone CURE.V55 was used to coat microtitter platesŽ .Corning ELISA plates at 2.5 mgrml at 48C for 16 h.After washing and blocking with PBS containing 3%bovine serum albumin, the lyophilized culture supernatantsresuspended in PBS containing 10% FCS and 0.5 ngrml
Ž .biotinylated-VIP volume of 100 ml were added to eachwell, and incubated for 12 h at 48C. The bound biotinyl-
Ž .ated-VIP was detected with avidin-peroxidase 2.5 mgrmlfor 30 min followed by addition of the ABTS substrate
Ž .solution 0.3 ngrml . Absorbances at 405 nm were mea-sured 30 min after the addition of the substrate. A standardcurve was constructed using various dilutions of VIP inPBS containing 10% FCS. The specificity of the antibodywas assessed by the binding of biotinilated-VIP in thepresence of increasing concentrations of secretin, glucagon,PACAP27, PACAP38, VIP , VIP and VIP .1 – 12 6 – 28 10 – 28
Fig. 1 shows no significant cross-reactivity with any testedneuropeptides but VIP . The amount of VIP in the6 – 28
culture supernatants was determined by extrapolation ofabsorbances to the standard curve. The sensitivity of theassay was 0.2 ng of VIP per ml. The concentration of VIPpresent in R5 medium was -0.2 ngrml.
2.8. Statistical analysis
All values are expressed as the mean"S.D. of thenumber of experiments performed in duplicate. The datawas evaluated statistically by Student’s test followed byScheffe’s F-test with P-0.05 as the minimum significantlevel.
3. Results
3.1. VIP expression in lymphocytes of central and periph-eral lymphoid organs and peritoneum after mitogenic stim-ulation
In the present study we have used a recently synthe-Ž .sized monoclonal anti-VIP antibody Wong et al., 1996 to
improve previous results obtained in our laboratory with apolyclonal antibody, as well as to develop a competitiveELISA in order to measure the VIP released to the lym-phoid microenvironment. Flow cytometry analysis infreshly isolated cells with the monoclonal anti-VIP anti-body demonstrates that 37% of thymocytes, 27% ofsplenocytes, 47% of lymph node lymphocytes, and 15% ofperitoneal lymphocytes show intracytoplasmic VIP-like
Ž .immunoreactivity Fig. 2 . Several different groups havereported an increase in neuropeptide production by im-
Žmune cells after mitogenic stimulation in culture Zuraw-.ski et al., 1986; Clarke et al., 1993 . To determine whether
the VIP expression could be modulated by cell activation,cell suspensions from thymus, spleen, lymph nodes andperitoneum were cultured in the presence or absence of the
Žmitogens ConA or LPS for different time periods 12, 24,.36 and 48 h and processed for immunofluorescence stain-
ing using a monoclonal anti-VIP antibody. We evaluatedthe proportion of lymphocytes bearing VIP as well as theamount of VIP expression by flow cytometry analysis. Thetime course for VIP expression is shown in Fig. 2. Inunstimulated cultures, we found a significant increase in
Ž .VIP positive cells especially at 12 h for thymocyte 68"2Ž .and peritoneal cell 36"2 cultures. Treatment with Con
A or LPS resulted in significant stimulatory effects fromŽ12 to 36 h, with maximal percentages at 12 h 75"1%,
50"7%, 69"4%, and 53"7% after ConA-stimulation,and 76"3%, 41"1%, 55"2%, and 41"1% afterLPS-stimulation, in thymus, spleen, lymph node and peri-
. Ž .toneal cell cultures, respectively Fig. 2 . The proportionof labeled lymphocytes at this time point was increased,depending of the tissue studied, from 1.2 to 1.7 fold and1.2 to 1.4 fold for ConA and LPS activation, respectively,by comparison to the unstimulated lymphocytes. The stim-ulatory effect of both stimuli was higher in spleen and
Ž .peritoneum Fig. 2B and D . In thymocytes both stimuliq Ž .induced similar percentages of VIP cells Fig. 2A ; how-
ever, Con A induced higher numbers of labeled lympho-cytes in spleen, lymph nodes and peritoneal cells as com-
Ž .pared to LPS Fig. 2B–D .In addition, to estimate the amount of VIP expression in
lymphocytes, we evaluated the difference in mean fluores-Ž .cence intensity FU, fluorescence units between the con-
trol and experimental histograms. We found that lympho-cytes stimulated with Con A or LPS showed a higherfluorescence intensity than the untreated lymphocytes forall the lymphoid organs studied, including peritoneumŽ .Fig. 3 . The major differences were observed at 12–24 h
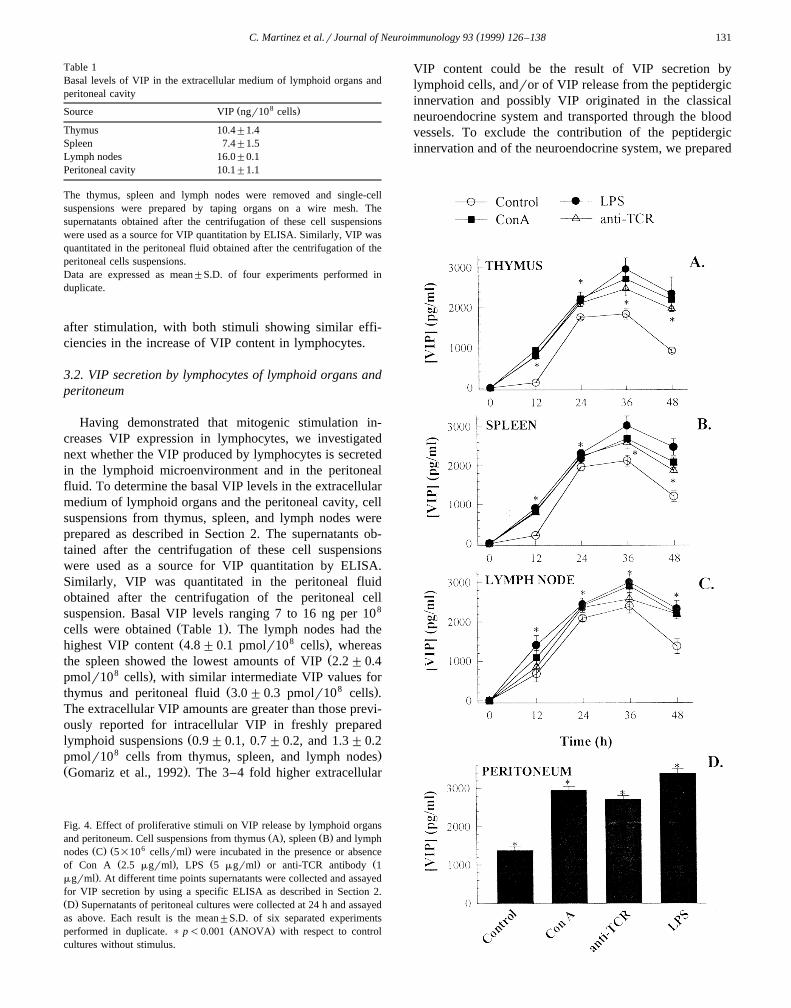
( )C. Martinez et al.rJournal of Neuroimmunology 93 1999 126–138 131
Table 1Basal levels of VIP in the extracellular medium of lymphoid organs andperitoneal cavity
8Ž .Source VIP ngr10 cells
Thymus 10.4"1.4Spleen 7.4"1.5Lymph nodes 16.0"0.1Peritoneal cavity 10.1"1.1
The thymus, spleen and lymph nodes were removed and single-cellsuspensions were prepared by taping organs on a wire mesh. Thesupernatants obtained after the centrifugation of these cell suspensionswere used as a source for VIP quantitation by ELISA. Similarly, VIP wasquantitated in the peritoneal fluid obtained after the centrifugation of theperitoneal cells suspensions.Data are expressed as mean"S.D. of four experiments performed induplicate.
after stimulation, with both stimuli showing similar effi-ciencies in the increase of VIP content in lymphocytes.
3.2. VIP secretion by lymphocytes of lymphoid organs andperitoneum
Having demonstrated that mitogenic stimulation in-creases VIP expression in lymphocytes, we investigatednext whether the VIP produced by lymphocytes is secretedin the lymphoid microenvironment and in the peritonealfluid. To determine the basal VIP levels in the extracellularmedium of lymphoid organs and the peritoneal cavity, cellsuspensions from thymus, spleen, and lymph nodes wereprepared as described in Section 2. The supernatants ob-tained after the centrifugation of these cell suspensionswere used as a source for VIP quantitation by ELISA.Similarly, VIP was quantitated in the peritoneal fluidobtained after the centrifugation of the peritoneal cellsuspension. Basal VIP levels ranging 7 to 16 ng per 108
Ž .cells were obtained Table 1 . The lymph nodes had theŽ 8 .highest VIP content 4.8"0.1 pmolr10 cells , whereas
Žthe spleen showed the lowest amounts of VIP 2.2"0.48 .pmolr10 cells , with similar intermediate VIP values for
Ž 8 .thymus and peritoneal fluid 3.0"0.3 pmolr10 cells .The extracellular VIP amounts are greater than those previ-ously reported for intracellular VIP in freshly prepared
Žlymphoid suspensions 0.9"0.1, 0.7"0.2, and 1.3"0.28 .pmolr10 cells from thymus, spleen, and lymph nodes
Ž .Gomariz et al., 1992 . The 3–4 fold higher extracellular
Fig. 4. Effect of proliferative stimuli on VIP release by lymphoid organsŽ . Ž .and peritoneum. Cell suspensions from thymus A , spleen B and lymph
Ž . Ž 6 .nodes C 5=10 cellsrml were incubated in the presence or absenceŽ . Ž . Žof Con A 2.5 mgrml , LPS 5 mgrml or anti-TCR antibody 1
.mgrml . At different time points supernatants were collected and assayedfor VIP secretion by using a specific ELISA as described in Section 2.Ž .D Supernatants of peritoneal cultures were collected at 24 h and assayedas above. Each result is the mean"S.D. of six separated experiments
Ž .performed in duplicate. ) p-0.001 ANOVA with respect to controlcultures without stimulus.
VIP content could be the result of VIP secretion bylymphoid cells, andror of VIP release from the peptidergicinnervation and possibly VIP originated in the classicalneuroendocrine system and transported through the bloodvessels. To exclude the contribution of the peptidergicinnervation and of the neuroendocrine system, we prepared

( )C. Martinez et al.rJournal of Neuroimmunology 93 1999 126–138132
thymocyte, splenocyte, lymph node and peritoneal cellsuspensions and cultured them in the presence or absenceof various stimuli. Culture supernatants were collected atdifferent time points and assayed for VIP content byELISA.
3.2.1. Effect of proliferatiÕe stimuli on VIP releaseUnstimulated lymphocytes showed a time-dependent
increase in the secretion of VIP, with maximum VIPrelease at 24–36 h in thymus, spleen, and lymph node
Ž .cultures Fig. 4A–C . We observed increases in VIP secre-tion when the lymphocytes were treated with proliferative
Žstimuli, such as ConA, LPS, and anti-TCR antibody Fig..4A–C . The time curves indicate that the VIP release was
significantly increased by all stimuli at 12 h. Progressivelyincreased VIP contents were present in the supernatants ofboth stimulated and unstimulated cultures, with maximal
Žlevels at 36 h for thymus, spleen and lymph node Fig..4A–C . Since preliminary experiments indicated that the
maximum VIP production in peritoneal cells occurs at 24 hŽ .data not shown , and due to limitations in cell numbers,we investigated the effect of the different stimuli only at24 h, showing that all stimuli increased VIP production in
Ž .peritoneal cells Fig. 4D .The different treatments resulted in a significant stimu-
latory effect at all time points and in all cultures. Theability to increase the VIP release was slightly differentdepending on the stimulus, with LPS and Con A being the
Ž .most potent Fig. 4 .
3.2.2. Effect of cytokines on VIP productionTo gain insight into the factors involved in VIP produc-
tion by lymphoid cells, we investigated whether cytokinesalso play a role in the secretion of VIP. Treatment ofthymus, spleen, lymph node and peritoneal cell cultureswith IL-1b, IL-6 and TNFa elicited a time-dependentincrease in the content of VIP for all the organs studiedŽ .Fig. 5A–C . In thymus, spleen, and lymph node themaximal effect was reached at 36 h and in peritoneal cells
Ž .at 24 h Fig. 5D . IL-1b, IL-6 and TNFa showed similarpotencies in inducing VIP production. In all the organsassayed, exposure to IL-2 had no significant effect on VIPproduction compared to controls, indicating that IL-2 can-
Ž .not induce VIP production Fig. 5 . These findings indicatethat several inflammatory mediators are able to regulatethe production of VIP in the immune system.
To elucidate whether the stimulatory effect of LPS, aknown B-cell mitogen as well as a potent activator of themacrophage response, on VIP production is mediatedthrough the elevation of proinflammatory cytokines se-creted by macrophages, we stimulated thymus, spleen,lymph node and peritoneal cells with LPS plus anti-TNFa
antibody, LPS plus anti-IL-6 antibody or LPS plus anti-TNFa and anti-IL-6 antibodies. When endogenous IL-6and TNFa are blocked by their specific antibodies, thestimulatory effect of LPS on VIP release is partially
Fig. 5. Effects of cytokines on VIP production. Cell suspensions fromŽ . Ž . Ž . Ž 6 .thymus A , spleen B and lymph nodes C 5=10 cellsrml were
Ž .incubated in the presence or absence of mrTNFa 10 ngrml , mrIL-1 ߎ . Ž . Ž .10 ngrml , mrIL-2 10 ngrml or mrIL-6 20 ngrml . At different timepoints supernatants were collected and assayed for VIP secretion by using
Ž .a specific ELISA as described in Section 2. D Supernatants of peri-toneal cultures were collected at 24 h and assayed as above. Each result isthe mean"S.D. of six separated experiments performed in duplicate.) p-0.001 with respect to control cultures without stimulus.

( )C. Martinez et al.rJournal of Neuroimmunology 93 1999 126–138 133
Ž .reversed in all the organs assayed Fig. 6 . In thymus,spleen and lymph nodes simultaneous treatment with LPSand both anti-IL-6 and anti-TNFa antibodies reduced VIPproduction almost to control levels suggesting that LPSinduces VIP secretion mainly through IL-6 and TNFa
Ž .production Fig. 6A–C . In contrast, anti-TNFa and anti-IL-6 had less effect in peritoneum, and the simultaneoustreatment with the two antibodies reduced VIP levels but
Ž .without bringing them to control levels Fig. 6D . Thissuggests that the LPS-induced VIP release from peritonealcells is only partially mediated through TNFa and IL-6.
3.2.3. Effect of glucocorticoids on VIP release by thymo-cytes
In a previous report from our laboratory we demon-strated that VIP protects thymocytes from dexamethasone-
Ž .induced apoptosis Delgado et al., 1996c . To investigatewhether glucocorticoids influence VIP production in thy-mocytes, we treated thymocytes with 5=10y7 M dexa-methasone for 12, 24, 36 and 48 h. Results indicated thatthe VIP production was elevated, showing the highest
Ž .increase after 24 h 1.6 fold and decreasing thereafterŽ .Fig. 7A . The effect of dexamethasone was dose-depen-
Ž .dent Fig. 7B . Significant stimulation was observed fordexamethasone in the concentration range 10y7 to 10y6 MŽ .Fig. 7B .
3.2.4. PositiÕe feedback of VIPIn order to find out if VIP is involved in the regulation
of its own secretion by cells of the immune system,thymus, spleen, lymph node and peritoneal cells were
Ž . y8cultured in the absence control or presence of 10 MVIP. Exogenous VIP stimulates indeed the VIP produc-tion, with the highest stimulatory effect at 12 h for all theorgans studied. At this time point, the levels of VIP were
Ž .increased at least fivefold compared to controls Fig. 8 .
3.2.5. Intracellular signal pathways inÕolÕed in the stimu-lation of VIP production
Finally, in order to elucidate the molecular mechanismunderlying the regulation of VIP release in the immune
Žsystem, we measured the effect of PMA an activator of. Žprotein kinase C , of retinal an inhibitor of protein kinase
. Ž . ŽC Taffet et al., 1983 , of forskolin an enhancer of cAMP. Ž . Žintracellular levels Boige et al., 1984 and of H-89 a
. Žpotent and selective protein kinase A inhibitor Chijiwa et.al., 1990 on the VIP production by splenocytes. Fig. 9A
Ž . Ž y6shows the effect of PMA 10 ngrml and forskolin 10.M on the VIP production by splenocytes. When spleno-
cytes were incubated with PMA or forskolin, an augmenta-tion in the VIP release occurred compared to untreated
Ž .samples controls , in a time-dependent manner. Both acti-
Ž 6 .Fig. 6. The effect of LPS on VIP production could be mediated through of proinflammatory cytokines. Cell suspensions 5=10 cellsrml from thymus,Ž . Ž . Ž .spleen, lymph nodes and peritoneum were incubated with LPS 5 mgrml , LPS plus anti-TNF antibody 1:250 , LPS plus anti-IL-6 antibody 1:250 , LPS
Ž .plus anti-IL-6 and anti-TNF antibodies, or medium alone controls . Supernatants were collected at 36 h or 24 h for lymphoid organs and peritoneumcultures, respectively, and assayed for VIP production by ELISA as described in Section 2. Results are the mean"S.D. of four to six experimentsperformed in duplicate.

( )C. Martinez et al.rJournal of Neuroimmunology 93 1999 126–138134
Ž .Fig. 7. Effect of glucocorticoids on VIP release by thymocytes. AŽ 6 .Thymocytes 5=10 cellsrml were incubated in the presence or ab-
sence of 5=10y7 M of dexamethasone. At different time points super-Ž .natants were collected and assayed for VIP production by ELISA. B
Dose-response curve for the stimulatory effect of dexamethasone on VIPŽ 6 .production. Thymocytes 5=10 cellsrml were incubated with different
Ž y8 y6 .concentrations of dexamethasone from 10 to 10 M . Supernatantswere collected at 24 h and assayed for VIP production by ELISA. Eachresult is the mean"S.D. of four to six separated experiments performedin duplicate. ) p-0.001 with respect to control cultures.
vators increased significantly the VIP levels after 12 h ofincubation, with maximal effects at 24–36 h. Stimulationof splenocytes for 36 h with forskolin or PMA resulted ina dose-dependent VIP release showing the strongest stimu-lation in cultures treated with 10y6 M forskolin or 10
Ž .ngrml PMA Fig. 9B and C . The stimulatory effect washigher for forskolin than for PMA at all time points and
Ž .concentrations assayed Fig. 9A–C . On the other hand,incubation with increasing concentrations of H89 or retinalproduced a progressively inhibitory effect on the VIP
Ž .production by splenocytes Fig. 9D and E . Both inhibitorsreduced in a dose-dependent manner VIP production.However, H-89 was more efficient than retinal, concentra-
Ž .Fig. 8. Positive feedback of VIP. Cell suspensions from thymus A ,Ž . Ž . Ž 6 .spleen B and lymph nodes C 5=10 cellsrml were incubated in the
presence or absence of VIP 10y8 M. The neuropeptide was removedŽ .after 2 h of incubation by washing three times with serum-free medium ,
and the cells were then resuspended in complete medium. At differenttime points supernatants were collected and assayed for VIP secretion by
Ž .using a specific ELISA as described in Section 2. D Supernatants ofperitoneal cultures were collected at 24 h and assayed as above. Eachresult is the mean"S.D. of six separated experiments performed induplicate. ) p-0.001 with respect to control cultures without stimulus.

( )C. Martinez et al.rJournal of Neuroimmunology 93 1999 126–138 135
Ž . Ž 6 .Fig. 9. Comparative effects of PMA, forskolin, retinal, and H-89 on VIP production by splenocytes. A Splenocytes 5=10 cellsrml were incubated inŽ . Ž y6 .presence or absence of PMA 10 ngrml or forskolin 10 M . Supernatants were collected at different time points and assayed for VIP production by
Ž . Ž 6 .ELISA. B–E Dose-response curves for the effect of forskolin, PMA, H-89 and retinal on VIP release. Splenocytes 5=10 cellsrml were cultured withŽ .different concentrations of forskolin, PMA, H-89 and retinal for 36 h. Supernatants were collected and assayed for VIP production by ELISA. F
Ž 6 . y7 y6Splenocytes 5=10 cellsrml were incubated in the presence or absence of 10 M forskolin or 10 M retinal in combination with differentconcentrations of PMA or retinal, respectively. Supernatants were collected 36 h later and assayed for VIP production by ELISA. Each result is themean"S.D. of six separated experiments performed in duplicate. ) p-0.05, )) p-0.01 and ))) p-0.001 with respect to control cultures withoutstimulus. ap-0.001 compared with samples treated with 10y7 M FK. §p-0.001 with respect to 10y6 M H-89.
Ž .tion wise Fig. 9D and E . Simultaneous incubations withforskolin 10y7 M and increasing PMA concentrationsshowed a progressive increase in VIP production com-pared with samples treated separately with the two activa-
Ž . y6tors Fig. 9F . In addition, combinations of 10 M H89and increasing retinal concentrations resulted in an addi-tive inhibitory effect on VIP release showing statisticallysignificant differences at 10y7 –10y6 M retinal–H89 com-
Ž .binations Fig. 9F .
4. Discussion
Several different groups have reported an increase inneuropeptide production by immune cells after mitogenic
Žstimulation in culture Zurawski et al., 1986; Clarke et al.,.1993 , and we provided evidence that lymphocytes can
Ž .produce VIP Gomariz et al., 1994a . In the present reportwe also demonstrate their ability to synthesize VIP in
response to different stimuli and to secrete this neuropep-tide to the culture medium ‘in vitro’. We have developedan ELISA that was specific and highly sensitive to detectVIP in cell cultures from lymphoid organs. The antibody,however, cross-reacts with VIP fragments that result fromproteolitic cleavage and that could be achieved by en-
Ždopeptidase activity present in lymphoid cells Goetzl et.al., 1989 . The identification and molecular characteriza-
tion of the VIP produced by cells of the lymphoid organsneeds to be elucidated by other techniques.
Flow cytometry analysis of the number of VIP positivelymphocytes in rat lymphoid organs reported here by usinga monoclonal antibody correlates with that reported previ-
Ž .ously with polyclonal antisera Leceta et al., 1994 . Thehigher proportion of positive thymocytes in the presentreport could be attributed to the higher titer and affinity ofthe monoclonal antibody used. A limited capacity of spleenand lymph node lymphocytes to produce VIP unless theyare stimulated can be inferred from data presented here,

( )C. Martinez et al.rJournal of Neuroimmunology 93 1999 126–138136
since the proportion of positive cells was not altered incontrol cultures. However, thymocytes seem to be capableof synthesizing VIP without specific stimulation. Evidencefor the production of VIP by thymocytes has been demon-strated ‘in vivo’ by the presence of VIP specific mRNAŽ .Gomariz et al., 1993 . In situ hybridization localized theVIP mRNA in cells with a recognizable lymphoid mor-phology in the corticomedullary and medullary regions ofthe rat thymus. The precise distribution of thymocytescontaining VIP mRNA suggests that thymocytes expressVIP at specific differentiation stages. VIP synthesis by
Žthymocytes was also confirmed by RT–PCR Gomariz et.al., 1994a; Delgado et al., 1996d; Leceta et al., 1996 . We
have reported that the VIP specific PCR product appears inŽ q q.the double-positive CD4 CD8 and single-positive
Ž q q.CD4 and CD8 subsets. Northern blot analysis ofmRNA from rat thymocytes showed two transcripts forVIP of approximately 1.7 and 1.0 kb, with a predominant
Ž .1.0 kb transcript Leceta et al., 1996 .The capacity of peripheral lymphoid cells to produce
VIP has been pointed out by the presence of mRNA byŽPCR in different subpopulations Gomariz et al., 1994a;
.Delgado et al., 1996d . Although immunoreactive-VIP haveŽbeen detected in mast cells and also in neutrophils Cutz et
.al., 1978; O’Dorisio et al., 1980 , lymphocytes representthe predominant cell type in which the peptide is detectedŽ .Leceta et al., 1994 . In this report we have shown thatlymphocytes from peripheral lymphoid organs produceVIP after stimulation with both T and B cell mitogens. Thehigher number of positive lymphocytes seen after Con Astimulation could reflect the proportion of T and B cellsubsets, or the presence in the medium of factors thatincrease the production of VIP by lymphocytes. The peakresponse in the cellular synthesis of VIP in lymphocytesseems to occur at 12–24 h after stimulation, according tothe number of positive cells and fluorescence intensitydetermined by flow cytometry. In this respect, VIP synthe-sis after stimulation seems to be delayed compared withthe synthesis of other neuropeptides and peptide hormonesby lymphocytes. For example, the production of GH whichplays an autocrinerparacrine role in lymphocyte replica-tion and cell cycle progression shows a very intense
Žresponse as early as 2–4 h after stimulation Weigent et.al., 1988 .
An important finding from our results is that cells fromthe lymphoid organs are able to secrete VIP, which couldsubsequently interact with the VIP receptors. VIP secretionin lymphoid organs ‘in vivo’ has been detected in thepresent study, since up to 3r4 of the total VIP content inthe different lymphoid organs was detected extracellularly.‘In vitro’, nanomolar concentrations determined by a spe-cific ELISA have been measured in the culture medium atthe peak response, a concentration at which maximumeffects are seen on different functions of the immune cells.While the spontaneous secretion of other neuropeptidesand hormones is induced very early, VIP secretion in the
absence of a specific stimulus seems to be delayed in boththymus and spleen cells in culture. The presence of VIP inthe culture medium can only be detected after 12 h. On theother hand, in lymph node and peritoneal cell cultures,extracellular VIP appears earlier indicating that the sponta-neous release of VIP is dependent on the cell type consid-ered. In any case the amounts of VIP secreted by lymphoidcells in culture, as detected in the present study are equiva-
Žlent to those reported for chromaffin cells ‘in vitro’ Eskay.and Eiden, 1992 .
All mitogenic stimuli tested induced a significant in-crease in the secretion of VIP compared with unstimulatedpopulations; an approximate 2-fold increase in the amountof secreted VIP was measured at the peak response instimulated thymocytes, with less amounts in the otherorgans. This increase is similar in range to that reported forother peptide hormones and neuropeptides after stimula-
Ž .tion Gutierrez et al., 1995 . However, while Con A in-duced the higher proportion of VIP positive lymphocytes,the maximum secretion was achieved with LPS stimula-tion. These data correlate with the ‘in vivo’ observationsof an increase in the circulating levels of VIP during the
Ž .endotoxic shock syndrome Brandtzaeg et al., 1989 . Also,an augmentation in VIP has also been observed in theextracellular medium during local immune responsesŽ .Kaltreider et al., 1997 .
Another important aspect to be pointed out is thatcytokines produced by activated macrophages, such asIL-1, IL-6 and TNF, stimulate VIP secretion, while IL-2has no effect. These factors may represent the majorstimuli for VIP secretion in thymus, spleen and lymphnodes, since anti-IL-6 and anti-TNF antibodies counteractthe stimulatory effect of LPS on VIP secretion. Speciallyin the thymus, the activation of the small population of
Ž .macrophages 10% could explain the stimulation in VIPproduction by LPS, a B-cell mitogen and macrophageactivator. However, blocking of IL-6 and TNF inhibitsonly partially the LPS-induced VIP secretion in peri-toneum, probably reflecting that different cell types orfactors are involved in the induction of VIP secretion byLPS.
The physiological significance of the VIP secretion bylymphoid cells remains elusive. An insight in this aspectcould be provided by the cellular target for this neuropep-tide in the immune system. Research from our lab indi-cated the constitutive expression of VIP-R1 in differentmurine lymphocyte subpopulations, and the induction of
ŽVIP-R2 48 h after stimulation in T cells Delgado et al.,.1996b . A recruitment of VIP receptor-bearing leukocytes
have also been described 2 days after antigen challengeŽduring local immune responses in the lung Kaltreider et
.al., 1997 . This is the time of the peak response reportedfor VIP secretion found in the present study. Anotherpossible target during local immune responses could be themacrophages, since this cells have VIP receptors. Expo-sure to VIP have been shown to influence macrophage

( )C. Martinez et al.rJournal of Neuroimmunology 93 1999 126–138 137
functions, cell mobility, cytokine production and adher-Ž .ence De la Fuente et al., 1996; Ganea, 1996 . In light of
the above observations VIP secreted during lymphocyteactivation could act as an autocrinerparacrine factor influ-encing the differentiation or downregulation of the im-mune response.
It has been shown that the expression of the VIP gene isregulated by various agents that cause activation of signaltransduction pathways mediated by cAMP, Ca2q and pro-
Žtein kinase C Eiden and Hotchkiss, 1983; Tsukada et al.,.1987; Waschek et al., 1988; Fink et al., 1991 . Results
presented in this report indicate that VIP secretion inlymphoid cells is under cAMP and PKC regulation as inendocrine and neuroblastoma cells. An almost 2-fold in-crease in secretion was seen after FK and PMA treatment.Although a larger increase has been reported in other cellsystems by the activation of PKA and PKC, the role ofthese two transduction pathways in lymphoid cells is indi-cated by the inhibitory effect of H89 and retinal. Regula-tion of the VIP expression is mediated by a CRE and a
Ž .cytokine response element CyRE found in the VIP geneŽ .promoter Waschek et al., 1988; Symes et al., 1994 . The
CyRE could mediate the regulation of VIP gene expressionby IL-1, TNF and IL-6 reported in this study. Thesecytokines also induce a response of the same magnitude in
Žthe secretion of VIP in endocrine cells Eskay and Eiden,.1992 .
It has been reported that VIP stimulates its own secre-Žtion in the central nervous system Gozes and Brenneman,
.1989 . Data presented here indicate that exogenous VIPcontrols its own release from the lymphoid cells shortlyafter exposure to this neuropeptide. The rapid VIP increasein the medium could be partially attributed to the secreta-gogen activity of VIP on lymphoid cells.
It has been demonstrated that corticosteroids regulatethe production of VIP in the nervous and endocrine sys-
Ž .tems Chew et al., 1997 . We have also demonstrated thatVIP protects thymocytes from dexamethasone induced
Ž .apoptosis Delgado et al., 1996c . Although dexametha-sone induces thymocyte apoptosis we have reported herean increase in VIP secretion, which suggests that glucocor-ticoids stimulate the production of VIP in the survivingmore mature population. This could indicate the impor-tance of combined effects of hormones and neuropeptidesfrom both the thymic microenvironment and the lymphoidcells in thymocyte differentiation.
We conclude from the results of this study that VIP isproduced and secreted by lymphocytes and could play animportant role in the development of the immune response.We propose that during an immune response, the timelyrelease of VIP within the lymphoid organs should influ-ence the differentiation andror downregulation of the on-going response. Additional information about the mecha-nisms of these regulatory processes should lead to noveltherapeutic strategies for the treatment of inflammatorydiseases and certain immune disorders.
Acknowledgements
This work was supported by grant PB94-0310 from theSpanish Department of Education and Science. We aregrateful to Dr. H.C. Wong for the generous gift of the
Žanti-VIP monoclonal antibody UCLA School of Medicine,.Los Angeles, CA .
References
Bellinger, D.L., Lorton, D., Horn, L., Brouxhon, S., Felten, S.Y., Felten,Ž .D.L., 1997. Vasoactive intestinal polypeptide VIP innervation of rat
spleen thymus and lymph nodes. Peptides 8, 1139–1149.Blalock, J.E., Smith, E.M., 1980. Human leukocyte interferon: structural
and biological relatedness to adrenocorticotropic hormone and endor-phins. Proc. Natl. Acad. Sci. USA 77, 5972–5974.
Blalock, J.E., 1992. Production of peptide hormones and neurotrans-mitters by the immune system. Chem. Immunol. 52, 1–24.
Boige, N., Aminaroff, B., Munk, A., Laburthe, M., 1984. Forskolinstimulates adenylate cyclase in human colonic crypts: Interaction withVIP. Eur. J. Pharmacol. 101, 111–117.
Brandtzaeg, P., Oktedalen, O., Kierulf, P., Opstad, P.K., 1989. ElevatedVIP and endotoxin plasma levels in human gram-negative septicshock. Regul. Pept. 24, 37–44.
Calvo, J.R., Guerrero, J.M., Lopez-Gonzalez, A., Osuna, C., Segura, J.J.,´ ´1994. Characteristics of receptors for VIP in rat peritonealmacrophages. Peptides 15, 309–315.
Calvo, J.R., Pozo, D., Guerrero, J.M., 1996. Functional and molecularcharacterization of VIP receptor and signal transduction in humanrodent immune systems. Adv. Neuroimmunol. 6, 39–47.
Chew, L.J., Burke, Z.D., Morgan, H., Gozes, I., Murphy, D., Carter,D.A., 1997. Transcription of the vasoactive intestinal peptide gene inresponse to glucocorticoids: differential regulation of alternative tran-scripts is modulated by a labile protein in rat anterior pituitary. Mol.Cell Endocrinol. 130, 83–91.
Chijiwa, T., Mishima, A., Hagiwara, M., Sano, M., Hayashi, K., Inoue,T., Naito, K., Toshioka, T., Hidaka, H., 1990. Inhibition of forskolin-induced neurite outgrowth and protein phosphorylation by a newly
wsynthesized inhibitor of cyclic AMP-dependent protein kinase, N- 2-Ž . x Ž .p-bromocinnamylamino ethyl -5-isoquinoleinsulfonamide H89 , ofPC12D pheochro-mocytoma cells. J. Biol. Chem. 265, 5267–5272.
Clarke, B.L., Gebhardt, B.M., Blalock, J.E., 1993. Mitogen-stimulatedlymphocytes release biologically active corticotrophin. Endocrinology132, 983–988.
Cutz, E., Chan, W., Track, N.S., Gooth, A., Said, S.I., 1978. Release ofvasoactive intestinal polypeptide in mast cells by histamine liberators.Nature 275, 661–662.
Dardenne, M., Savino, W., 1994. Control of thymus physiology bypeptidic hormones and neuropeptides. Immunol. Today 15, 518–523.
Davidson, A., Moody, T.W., Gozes, I., 1996. Regulation of VIP geneexpression in general. J. Mol. Neurosci. 7, 99–110.
De la Fuente, M., Delgado, M., Gomariz, R.P., 1996. VIP modulation ofimmune cell functions. Adv. Neuroimmunol. 6, 75–91.
Delgado, M., Pozo, D., Martınez, C., Garrido, E., Leceta, J., Calvo, J.R.,´Gomariz, R.P., 1996a. Characterization of gene expression of VIP andVIP1-receptor in rat peritoneal lymphocytes and macrophages. Regul.Pept. 62, 161–166.
Delgado, M., Martınez, C., Johson, M.C., Gomariz, R.P., Ganea, D.,´1996b. Differential expression of vasoactive intestinal peptide recep-
Ž .tor 1 and 2 VIP-R1 and VIP-R2 mRNA in murine lymphocytes. J.Neuroimmunol. 68, 27–38.
Delgado, M., Garrido, E., Martınez, C., Leceta, J., Gomariz, R.P., 1996c.´Vasoactive intestinal peptide and pituitary adenylate cyclase-activat-
Ž .ing polypeptides PACAP27 and PACAP38 protect CD4qCD8q

( )C. Martinez et al.rJournal of Neuroimmunology 93 1999 126–138138
thymocytes from glucocorticoid-induced apoptosis. Blood 12, 5152–5161.
Delgado, M., Martınez, C., Leceta, J., Garrido, E., Gomariz, R.P., 1996d.´Differential VIP and VIP receptor gene expression in rat thymocyte1
subsets. Peptides 17, 803–807.Dimaline, R., 1989. Vasoactive intestinal polypeptide. In: Holmgren, S.
Ž .Ed. , Comparative Physiology of Regulatory Peptides. Chapman andHall, Beckknham, England, pp. 150–173.
Eiden, L.E., Hotchkiss, A.J., 1983. Cyclic adenosine monophosphateregulates vasoactive intestinal polypeptide and enkephalin biosynthe-sis in cultured chromaffin cells. Neuropeptides 4, 1–9.
Emanuele, N.V., Emanuele, M.A., Tentler, J., Kirsteins, L., Azad, N.,Lawrence, A.M., 1990. Rat spleen lymphocytes contain an immunore-active and bioactive luteinizing hormone-releasing hormone. En-docrinology 126, 2482–2486.
Eskay, R.L., Eiden, L.E., 1992. Interleukin-1a and tumor necrosis factor-a differentially regulate enkephalin, Vasoactive Intestinal Polypep-tide, Neurotensin, and Substance P biosynthesis in chromaffin cells.Endocrinology 130, 2252–2258.
Fink, J.S., Verhave, M., Walton, K., Mandel, G., Goodman, R.H., 1991.Cyclic AMP- and phorbol ester-induced transcriptional activation aremediated by the same enhancer element in the human vasoactiveintestinal peptide gene. J. Biol. Chem. 266, 3881–3887.
Fuller, P.J., Verity, K., 1989. Somatostatin gene expression in the thymusgland. J. Immunol. 143, 1015–1017.
Ganea, D., 1996. Regulatory effects of vasoactive intestinal peptide oncytokine production in central and peripheral lymphoid organs. Adv.Neuroimmunol. 6, 61–74.
Geenen, V., Legros, J.J., Franchimont, P., Defresne, M.P., Boniver, J.,Ivell, R., Richter, D., 1987. The thymus as a neuroendocrine organ:Synthesis of vasopressin and oxytocin in human thymic epithelium.Ann. Acad. NY Acad. Sci. 496, 56–66.
Goetzl, E.J., Sreedharan, S.P., Turck, D.W., 1988. Structurally distinctivevasoactive intestinal peptides from rat basophilic leukemia cell. J.Biol. Chem. 263, 9083–9086.
Goetzl, E.J., Kodama, K.T., Turck, C.W., Schiogolev, A.S., Sreedharan,S.P., 1989. Unique pattern of cleavage of vasoactive intestinal peptideby human lymphocytes. Immunology 66, 554–558.
Gomariz, R.P., Lorenzo, M.J., Cacicedo, L., Vicente, A., Zapata, A.,1990. Demonstration of immunoreactive vasoactive intestinal peptideŽ . Ž .IR-VIP and somatostatin IR-SOM in rat thymus. Brain Behav.Immun. 4, 151–161.
Gomariz, R.P., De la Fuente, M., Hernanz, A., Leceta, J., 1992. Occur-Ž .rence of vasoactive intestinal peptide VIP in lymphoid organs from
rat and mouse. Ann. NY Acad. Sci. 650, 13–18.Gomariz, R.P., Delgado, M., Naranjo, J.R., Mellstrom, B., Tormo, A.,
Mata, F., Leceta, J., 1993. VIP gene expression in rat thymus andspleen. Brain Behav. Immun. 7, 271–278.
Gomariz, R.P., Leceta, J., Garrido, E., Garrido, T., Delgado, M., 1994a.Ž .Vasoactive intestinal peptide VIP mRNA expression in rat T and B
lymphocytes. Regul. Pept. 50, 177–184.Gomariz, R.P., Garrido, E., Leceta, J., Martınez, C., Abalo, R., Delgado,´
M., 1994b. Gene expression of VIP receptor in rat lymphocytes.Biochem. Biophys. Res. Commun. 203, 1599–1604.
Gozes, I., Brenneman, D.E., 1989. VIP: Molecular biology and neurobio-logical function. Mol. Neurobiol. 3, 201–236.
Gutierrez, M.A., Molina, J.F., Cuellar, M.L., Garcıa, C., Gutierrez-Urena,´S., Charavi, A., Espinoza, L.R., 1995. Prolactin and systemic lupuserythematosus: prolactin secretion by SLE lymphocytes and prolifera-
Ž .tive autocrine activity. Lupus 4, 348–352.Johnson, M.C., McCormack, R.J., Delgado, M., Martınez, C., Ganea, D.,´
1996. Murine T lymphocytes express constitutive intestinal peptideŽ .receptor 1 VIP-R1 mRNA. J. Neuroimmunol. 68, 109–119.
Kaltreider, H.B., Ichikawa, S., Byrd, P.K., Ingram, D.A., Kishiyama,J.L., Sreedharan, S.P., Warnock, M.L., Beck, J.M., Goetzl, E.J., 1997.
Upregulation of neuropeptides and neuropeptide receptors in a murinemodel of immune inflammation in lung parenchyma. Am. J. Respir.Cell Mol. Biol. 16, 133–144.
Leceta, J., Martınez, M.C., Delgado, M., Garrido, E., Gomariz, R.P.,´1994. Lymphoid cell subpopulations containing vasoactive intestinalpeptide in rat. Peptides 15, 791–797.
Leceta, J., Martınez, C., Delgado, M., Garrido, E., Gomariz, R.P., 1996.´Expression of vasoactive intestinal peptide in lymphocytes: A possi-ble endogenous role in regulation of the immune system. Adv.Neuroimmunol. 6, 29–36.
O’Dorisio, M.S., O’Dorisio, T.M., Cataland, S., Balcerzak, S.P., 1980.Vasoactive intestinal polypeptide as a biochemical marker for poly-morphonuclear leukocytes. J. Lab. Clin. Med. 96, 666–672.
O’Neal, K.D., Montgomery, D.W., Truong, T.M., Yu-Lee, L.Y., 1992.Prolactin gene expression in human thymocytes. Mol. Cell. En-docrinol. 87, R19–R23.
Pozo, D., Delgado, M., Martınez, C., Gomariz, R.P., Guerrero, J.M.,´Calvo, J.R., 1997. Functional characterization and mRNA expression
Ž .of pituitary adenylate cyclase activating polypeptide PACAP type Ireceptors in rat peritoneal macrophages. Biochim. Biophys. Acta1359, 250–262.
Rawlings, S.R., 1994. PACAP, PACAP receptors, and intracellular sig-nalling. Mol. Cell. Endocrinol. 101, C5–C9.
Rosen, H., Behar, O., Abramsky, O., Ovadia, H., 1989. Regulatedexpression of proenkephalin A in normal lymphocytes. J. Immunol.143, 3703–3707.
Said, S.I., Mutt, V., 1970. Potent peripheral splanchnic vasodilator pep-tide from normal gut. Nature 225, 863–864.
Santambrogio, L., Benedetti, M., Chao, M.V., Muzaffar, R., Kulig, K.,Gabellini, N., Hochwald, G., 1994. Nerve growth factor productionby lymphocytes. J. Immunol. 153, 4488–4495.
Symes, A.J., Lewis, S.E., Corpus, L., Rajan, P., Hyman, S.E., Fink, J.S.,1994. STAT proteins participate in the regulation of the vasoactiveintestinal peptide gene by ciliary neurotrophic factor family of cy-tokines. Mol. Endocrinol. 8, 1750–1763.
Taffet, S.M., Greenfield, A.R.L., Haddox, M.K., 1983. Retinal inhibitsTPA activated calcium-dependent, phospholipid-dependent protein ki-nase C. Biochem. Biophys. Res. Commun. 114, 1194–1199.
Tsukada, T., Fink, J.S., Mandel, G., Goodman, R.H., 1987. Identificationof a region in the human vasoactive intestinal peptide gene responsi-ble for regulation by cyclic AMP. J. Biol. Chem. 262, 8743–8747.
Vollmar, A.K., Lang, R.E., Hanze, A., Schulz, R., 1990. The rat thymus:a site of atrial natriuretic peptide synthesis. Peptides 11, 33–37.
Waschek, J.A., Hsu, C.M., Eiden, L.E., 1988. Lineage-specific regulationof the vasoactive intestinal peptide gene in neuroblastoma cells isconferred by 5.2 kilobases of 5X-flanking sequence. Proc. Natl. Acad.Sci. USA 85, 9547–9551.
Weigent, D.A., Baxter, J.B., Wear, W.E., Smith, L.R., Bost, K.L.,Blalock, E., 1988. Production of immunoreactive growth hormone bymononuclear leukocytes. FASEB J. 2, 2812–2818.
Weigent, D.A., Blalock, E., 1990. Immunoreactive growth hormone-re-leasing factor in rat leukocytes. J. Neuroimmunol. 29, 1–13.
Weinstock, J.V., Blum, A., Walder, R., 1989. Eosinophils from granulo-mas in murine schistosomiasis mansoni produce substance P. J.Immunol. 141, 961–966.
Weinstock, J.V., Blum, A.M., 1990. Detection of vasoactive intestinalpeptide and localization of its mRNA within granulomas of murineschistosomiasis. Cell Immunol. 125, 291–300.
Wong, H.C., Sternini, C., Lloyd, K., De Giorgio, R., Walsh, J.H., 1996.Monoclonal antibody to VIP: Production, characterization, im-munoneutralizing activity, and usefulness in cytochemical staining.Hybridoma 15, 133–139.
Zurawski, G., Benedik, M., Kamb, B.J., Abrams, J.S., Zurawski, S.M.,Lee, F.D., 1986. Activation of mouse T-helper cells induces abundantpreproenkephalin mRNA synthesis. Science 232, 772–775.


















