
Regulation of Actin Dynamics in Animal Cells: the...
59
1 Regulation of Actin Dynamics in Animal Cells: the Role of ADF/cofilin and Twinfilin Maria Vartiainen Institute of Biotechnology Department of Biosciences, Division of Biochemistry, Helsinki Graduate School in Biotechnology And Molecular Biology, Faculty of Science University of Helsinki Finland Academic dissertation To be presented, with the permission of the Faculty of Science of the University of Helsinki, for public criticism in the auditorium 1041 at Viikki Biocenter (Viikinkaari 5, Helsinki) on October 4 th , 2002, at 12 o’clock.
Transcript of Regulation of Actin Dynamics in Animal Cells: the...

1
Regulation of Actin Dynamics in Animal Cells:
the Role of ADF/cofilin and Twinfilin
Maria Vartiainen
Institute of Biotechnology Department of Biosciences, Division of Biochemistry,
Helsinki Graduate School in Biotechnology And Molecular Biology,
Faculty of Science University of Helsinki
Finland
Academic dissertation
To be presented, with the permission of the Faculty of Science of the University of Helsinki, for public criticism in the auditorium 1041 at Viikki
Biocenter (Viikinkaari 5, Helsinki) on October 4th, 2002, at 12 o’clock.

2
Supervised by Docent Pekka Lappalainen Institute of Biotechnology University of Helsinki Reviewed by Docent Olli Carpen Department of Pathology University of Helsinki Professor John Eriksson Department of Biology University of Turku Opponent Professor Shoichiro Ono Department of Pathology Emory University, Atlanta Cover figure: NIH 3T3 cell stained with rhodamine-phalloidin (red) to visualize actin filaments and with FITC-DNAseI (green) to visualize DNA and actin monomers. ISBN 952-10-0317-0 ISBN 952-10-0318-9 (ethesis) ISSN 1239-9469 Gummerus Kirjapaino OY Saarijärvi, 2002

3
To my Mother and Father

4
Contents
CONTENTS .....................................................................................................2
ABBREVIATIONS ...........................................................................................6
ORIGINAL PUBLICATIONS ...........................................................................7
ABSTRACT .....................................................................................................8
INTRODUCTION .............................................................................................9 1. THE CYTOSKELETON ..................................................................................9 2. ACTIN........................................................................................................9 3. THE ROLE OF ACTIN IN VARIOUS CELL BIOLOGICAL PROCESSES....................11
3.1 Actin filament structures in cells.........................................................11 3.2 Cell movements rely on the actin cytoskeleton .................................12
4. ACTIN-BINDING PROTEINS .........................................................................13 4.1 The role of actin-binding proteins in actin filament dynamics.............15 4.2 Actin monomer-binding proteins .......................................................16
4.2.1 Profilin ..........................................................................................16 4.2.2 Thymosin β4 ................................................................................17 4.2.3 ADF-H domain proteins................................................................18 4.2.4 ADF/Cofilin...................................................................................19
4.2.4.1 Interactions with actin.............................................................19 4.2.4.2 Cell biological properties ........................................................20 4.2.4.3 Regulation by pH, phosphoinositides and phosphorylation...21 4.2.4.4 ADF/cofilin isoforms ..............................................................21
4.2.5 Twinfilin .......................................................................................22 4.2.5.1 Twinfilin is composed of two ADF-H domains ........................22 4.2.5.2 Biochemical and cell biological activities of yeast twinfilin......23
5. SIGNALING TO THE ACTIN CYTOSKELETON ..................................................23 5.1. Rho family GTPases ........................................................................24
5.1.1. Rho regulates actomyosin-based structures ...............................24 5.1.2. Rac and Cdc42 regulate the formation of lamellipodia and filopodia ................................................................................................25
5.2. PI(4,5)P2 ...........................................................................................26 AIMS OF THE STUDY...................................................................................28
MATERIALS AND METHODS ......................................................................29
RESULTS ......................................................................................................30 6. THE THREE MOUSE ADF/COFILINS HAVE DISTINCT EXPRESSION PATTERNS AS WELL AS DIFFERENT BIOCHEMICAL PROPERTIES (ΙΙΙΙ) ..........................................30 7. THE ROLE OF TWINFILIN IN DROSOPHILA DEVELOPMENT (ΙΙΙΙΙΙΙΙ).........................31 8. THE ROLE OF TWINFILIN IN MAMMALS (ΙΙΙ(ΙΙΙ(ΙΙΙ(ΙΙΙ ) .................................................31 9. THE TWO MAMMALIAN TWINFILIN ISOFORMS HAVE SIMILAR BIOCHEMICAL PROPERTIES BUT DISTINCT EXPRESSION PATTERNS (UNPUBLISHED) ..................32

5
DISCUSSION.................................................................................................35 10. THE BIOCHEMICAL PROPERTIES OF MOUSE ADF/COFILINS ARE ADAPTED TO THEIR EXPRESSION PATTERNS ........................................................................35 11. TWINFILIN IS ESSENTIAL FOR THE DEVELOPMENT OF DROSOPHILA..............38 12. MOUSE TWINFILIN ISOFORMS HAVE SIMILAR BIOCHEMICAL PROPERTIES BUT DISTINCT EXPRESSION PATTERNS....................................................................39 13. THE CELL BIOLOGICAL ROLE OF TWINFILIN.................................................41
CONCLUSIONS AND FUTURE PERSPECTIVES........................................43
ACKNOWLEDGEMENTS .............................................................................46
REFERENCES ..............................................................................................46

6
Abbreviations Abp1 actin binding protein 1 ADP adenosine diphosphate ADF actin depolymerizing factor ADF-H actin depolymerizing factor-homology Aip1 actin interacting protein 1 Arp actin related protein ATP adenosine triphosphate cAMP cyclic adenosine monophosphate CAP adenylyl cyclase associated protein Cc critical concentration E embryonic day ERM ezrin/radixin/moesin F-actin filamentous actin FH formin homology FtsZ filamentous temperature sensitive protein Z G-actin globular actin, actin monomer GAP GTPase activating protein GDI guanosine nucleotide dissociation inhibitor GDP guanosine diphosphate GEF guanosine nucleotide exchange factor GST glutathione S-transferase GTP guanosine triphosphate ICAM intercellular adhesion molecule KD dissociation constant MLC myosin light chain MRCK myotonic dystrophy kinase-related Cdc42-binding kinase MreB murein cluster e B mRNA messenger RNA NHE1 Na+/H+ exchange protein 1 NLS nuclear localization signal PAK p21-activated kinase Pi pyrophosphate PI phosphoinositide RNA ribonucleic acid ROK Rho associated kinase Scar suppressor of cAMP receptor mutation (WASp homolog) SH Src homology TESK testicular kinase WASp Wiscott-Aldrich syndrome protein WH WASP homology WIP WASP-interacting protein Note in nomenclature:
gene mRNA protein organism

7
Original publications This thesis is based on the following articles, which are referred to in the text by their Roman numerals. Unpublished material is also presented. Ι Vartiainen, M.K., Mustonen, T., Mattila, P.K., Ojala, P.J., Thesleff, I.,
Partanen, J., and Lappalainen, P. (2002). The three mouse Actin-depolymerizing factor/cofilins evolved to fulfill cell-type-specific requirements for actin dynamics. Mol. Biol. Cell 13:183-194.
ΙΙ Wahlström, G., Vartiainen, M., Yamamoto, L., Mattila, P.K.,
Lappalainen, P., and Heino, T.I. (2001). Twinfilin is required for actin-dependent developmental processes in Drosophila. J. Cell Biol. 155:787-796.
ΙΙΙ Vartiainen, M., Ojala, P.J., Auvinen, P., Peränen, J., and Lappalainen,
P. (2000). Mouse A6/twinfilin is an actin monomer-binding protein that localizes to the regions of rapid actin dynamics. Mol. Cell. Biol. 20:1772-1783.
Note in references:
Unpublished data presented in the results are referred to by “(unpublished)”, whereas other unpublished data is referred to by “(author et al., unpublished)”.

8
Abstract Actin is an essential, conserved protein that is necessary for a large number of cellular functions including cell division, movement, secretion and endocytosis. In cells, actin filaments are dynamic structures that assemble and disassemble rapidly in a co-ordinated fashion. This process of actin filament dynamics is tightly regulated both spatially and temporally, and therefore cells contain a large array of actin-binding proteins. One important aspect of actin filament turnover is the regulation of the size and dynamics of the actin monomer pool. This study has focused at analysis of two actin monomer-binding proteins ADF/cofilin and twinfilin, both of which share a common building block, the ADF-homology domain. ADF/cofilins are essential proteins whose function is to enhance actin filament turnover by depolymerizing actin filaments from the pointed ends. Unicellular organisms contain only one ADF/cofilin protein, whereas in multicellular organisms several isoforms exist. We compared the expression patterns and biochemical properties of the three mouse ADF/cofilin isoforms, which have 70-80% sequence identity. We found that these proteins have distinct expression patterns: cofilin-2 is a muscle and ADF an epithelial specific isoform, whereas cofilin-1 has a rather ubiquitous expression pattern. These mouse ADF/cofilin isoforms also exhibit differences in their biochemical properties. ADF is very efficient in enhancing actin filament disassembly and turnover, whereas cofilin-2 rather promotes the assembly of filaments. Cofilin-1 is an intermediate between cofilin-2 and ADF. Therefore, the biochemical properties of each isoform are well adapted to promote actin filament dynamics in the specific tissues where these proteins are expressed. Twinfilin is a recently identified actin monomer-binding protein that has been shown to regulate actin filament dynamics in yeast. We identified a Drosophila homologue of twinfilin and demonstrated that it is also an actin monomer-sequestering protein. Moreover, our studies showed that twinfilin is essential for actin-dependent developmental processes in Drosophila, providing the first in vivo evidence for twinfilin’s importance in multicellular organisms. We also identified and characterized two murine twinfilin isoforms and showed that these proteins have distinct expression patterns, but very similar biochemical properties. Furthermore, twinfilin’s localization as well as its overexpression phenotype in murine cells establishes twinfilin’s role as a universal regulator of actin filament turnover. Combined with previous results, this study allows us to present a model of how various ADF/cofilin and twinfilin isoforms contribute to the regulation of the actin monomer pool in animal cells.

9
Introduction
1. The Cytoskeleton The cytoskeleton, together with internal membranes and the nucleus, are considered to be hallmarks of eukaryotic cells. The cytoskeleton is an extensive system of protein filaments that enables eukaryotic cells to organize their interiors and to perform various directed movements. The most abundant components of the cytoskeleton are microtubules, actin filaments and intermediate filaments. Microtubules are long, stiff polymers that extend throughout the cytoplasm, determining the overall shape of the cell as well as governing the location of membrane bound organelles and directing intracellular traffic. Intermediate filaments are tough and durable protein fibres that provide mechanical stability to animal cells. Actin filaments are thin and flexible, and their mission is to shape the plasma membrane and drive its movements (reviewed in Bray et al., 2001). The development of the cytoskeleton during evolution gave cells many advantages, which eventually led to the appearance of multicellular organisms. The cytoskeleton enabled cells to change their shape, and the surface plasticity might have made possible the engulfment of endosymbiotic organelles, which led to the evolution of mitochondria and chloroplasts. Appearance of the mitotic spindle based on microtubules could have allowed the genome to split into multiple chromosomes, allowing the expansion of genetic information. The cytoskeleton also brought along the capacity to perform internal movements, which made it possible to expand cell size and complexity (reviewed in Bray et al., 2001). Although prokaryotic organisms do not have a true cytoskeleton like eukaryotes, proteins distantly related to actin and tubulin have been found in bacteria. FtsZ (filamentous temperature-sensitive protein Z) and MreB (murein cluster e B) have only low sequence homology to tubulin and actin, respectively, but their three-dimensional structures resemble their eukaryotic counterparts. These proteins are also able to form filaments at least in vitro, and they function in such processes as cell division. Therefore, it is possible that actin and tubulin have a prokaryotic origin and that also prokaryotes organize their interior through mechanisms that are similar to, but less sophisticated than those of eukaryotes (reviewed in van den Ent et al., 2001).
2. Actin Actin is an essential, globular protein with a molecular weight of approximately 43 kDa that is highly abundant in all eukaryotic cells. It is extremely conserved, and even actins from different species can complement each other’s functions. Lower eukaryotes, such as yeast, have only one actin gene encoding a single protein, but higher eukaryotes, like mammals, have multiple isoforms of actin that are selectively expressed in different tissues. In addition,

10
a single cell can contain several isoforms of actin that segregate to different structures within cells. Although actin isoforms display some differences in their thermodynamic stability, polymerization kinetics and affinity for actin- binding proteins, the importance of the different isoforms in vivo is not clear (reviewed in Sheterline et al., 1998). The actin molecule is a relatively flat, two-lobed molecule with a central cleft between the two lobes (Figure 1A). Each lobe-shaped domain is further divided into two subdomains. The central cleft contains binding sites for a nucleotide (ADP or ATP) and a divalent cation, which are essential cofactors of actin (Kabsch & Holmes, 1995). Actin monomers (G-actin) are able to spontaneously form filaments (F-actin) under the ionic conditions believed to exist in cells (Figure 1B). Several models for the F-actin structure have been proposed based on the structure of the actin monomer and other ultrastructural data. The actin filament can therefore be described either as a single-start left-handed helix or a two-start right-handed double helix. The typical time course of actin polymerization in vitro first shows a short delay, called a lag phase, due to the instability of actin dimers and trimers, which are necessary intermediates in actin polymerization. The lag phase is followed by a rapid polymerization period until the concentration of actin monomers reaches the critical concentration (Cc). At Cc, actin monomers exist in equilibrium with actin filaments, and actin polymerization balances depolymerization (reviewed in Steinmetz et al., 1997).
Figure 1. Overview of actin. (A) Actin monomer with a bound nucleotide (PDB ID: 1ATM). (B) Model of actin filament (PDB ID:1ALM). Panels A and B were created using programs Molscript (Kraulis, 1991) and Raster 3D (Merritt et al., 1994) (C) Actin filaments in NRK (normal rat kidney) cells visualized by rhodamine-phalloidin.

11
The turnover of actin filaments in cells is rapid, and individual actin filaments assemble and disassemble over a period of minutes. The chemical switch between polymerization and depolymerization is the hydrolysis of ATP, which brings energy to the system. Each actin monomer catalyzes the hydrolysis of an ATP molecule during a single cycle of assembly and disassembly. In rapidly assembling filaments, hydrolysis lags behind the assembly. Thus the hydrolysis of ATP is uncoupled from assembly site, and the hydrolysis occurs in the filament yielding first ADP.Pi-F-actin and after the release of the pyrophosphate, ADP-F-actin. A conformational switch accompanies this process. Because ADP-actin does not pack well into filaments, actin filaments are disassembled (reviewed in Welch et al., 1997). Due to the polar nature of each actin monomer, actin filaments also have a unique polarity. This polarity can be detected by decorating actin filaments with fragments of myosin. Projections of myosin are tilted in one direction and appear as arrowheads. Thus the two ends of actin filaments are called the barbed end (or the plus end) and the pointed end (minus end). These two ends grow at different rates, due to different association and dissociation rates and the difference in CC between the two ends. Under physiological conditions the CC for the barbed end is 0.1 µM and for the pointed end 0.7 µM. For these reasons, barbed ends tend to grow, while pointed ends shrink, and individual actin monomers can move steadily from the plus end to the minus end in a process called treadmilling (reviewed in Sheterline et al., 1998).
3. The role of actin in various cell biological processes 3.1 Actin filament structures in cells Actin filaments are the “functional” forms of actin within cells (Figure 1C). However, the filaments seldom function as isolated units, but are instead organized into higher-order assemblies. These can be divided into three categories (Figure 2). The first class includes antiparallel or bipolar actin bundles (Figure 2A), found in their most organized form in striated muscle. In muscle fibers, actin filaments are kept precisely interdigitated with myosin filaments, which then walk along actin filaments, thus producing the contractile force. Similar, but less obviously organized structures can be found in smooth muscle cells and non-muscle cells. For example stress fibers, adhesion belts and contractile rings of dividing animal cells are antiparallel actin bundles with contractile properties (reviewed in Sheterline et al., 1998). The second category is parallel actin bundles (Figure 2B), which often function to support or stabilize cellular protrusions, such as the neurosensory bristles of Drosophila, brush border microvilli and hair cell stereocilia (reviewed in Bartles et al., 2000). Also protrusions involved in cell motility, like filopodia, microspikes and lamellipodia, display unipolar actin filaments, having the fast-growing barbed ends oriented outwards (reviewed in Small et al., 2002). In lamellipodia, actin forms branched structures rather than

12
Figure 2. Schematic representation of actin filament structures in cells. Actin filaments are in red. (A) Antiparallel actin bundles of the contractile ring (B) Parallel actin filaments of filopodia and lamellipodia in a migrating cell (C) Isotropic array of actin filaments in the cell cortex.
bundles. These branches always grow at 70o angles from the mother filament, and the branching frequency is highest near the leading edge (Svitkina et al., 1997). The third category of actin filament structures comprises the isotropic array under the plasma membrane, also known as the cell cortex (Figure 2C). Within the cortex, actin filaments are linked together in a three-dimensional network with small pores. This layer gives the outer surface of the cell mechanical strength and enables it to move (reviewed in Bray et al., 2001).
3.2 Cell movements rely on the actin cytoskeleton Actin has been implicated in numerous cell biological processes. The actin cytoskeleton ramifies throughout the whole cytoplasm and provides a large surface area on which proteins can anchor themselves. For example, proteins of the glycolytic pathway are often bound to actin filaments. In addition, signaling complexes can use actin filaments to position themselves in the cytoplasm. Several ion transporters associate with actin, and manipulation of the actin cytoskeleton can modulate channel function. Actin is also an important localizator of mRNA molecules, and, for example, actin mRNA itself is associated with actin filaments, and the protein synthesis of actin is intimately linked to the actin cytoskeleton (reviewed in Sheterline et al., 1998). Most of the well-defined functions of actin have been localized to the cytoplasm, but actin can also be detected in the nucleus. The physiological

13
role of nuclear actin is still unclear, but there is some evidence that it is a component of the “nuclear matrix”, which provides the organizational scaffold upon which nuclear events can take place. Actin has also been implicated in the regulation of transcription, RNA splicing and chromatin remodelling (reviewed in Rando et al., 2000). The best-characterized functions of actin are, however, related to cell movement and can be divided into two categories. First, actin filaments serve as tracks along which motor proteins called myosins can walk in a directed fashion. Myosins constitute a large superfamily of proteins that share a common domain that is able to interact with actin, to hydrolyze ATP and to produce movement. Myosins are typically constructed of three functional subdomains: the motor domain, which interacts with actin and ATP, the neck domain, which is involved in the regulation of myosin activity, and the tail domain, which anchors the myosin to its cargo. This cargo can be, for example another actin filament, plasma membrane or a vesicle (reviewed in Sellers et al., 2000). Secondly, actin filaments are also able to generate movement in the absence of myosins by inducing changes in the structure of cross-linked actin gels or by polymerizing against a surface. Selective breakage or formation of crosslinks can drive the remodelling of the cortical actin cytoskeleton, which may underlie such processes as phagocytosis, endocytosis and secretion (reviewed in Bray et al., 2001). The small intracellular pathogen Listeria monocytogenes is an excellent model for polymerization-driven movement. This bacterium induces the formation of an actin comet tail on its surface, which then propels the bacterium forward in the cytoplasm and assists its spreading to adjacent cells (Tilney and Portnoy, 1989). In addition, some vesicles are reported to use a similar comet tail mechanism to move within the cell (Rozelle et al., 2000; Taunton et al., 2000). The protrusion of the lamellipodium during cell locomotion is thought to be accomplished by a mechanism similar to that of polymerizing actin pushing the plasma membrane. The same key players are present both in the comet tail of Listeria and in the lamellipodium of a migrating cell (reviewed in Cameron et al., 2000).
4. Actin-binding proteins The diversity of actin filament structures within cells is the result of the association of actin with actin-binding proteins (Table 1). Some of these proteins facilitate the organization of actin into higher-order structures, while others regulate the dynamics of filament turnover and allow remodelling of the actin cytoskeleton in response to appropriate signals, ensuring that proper structures are formed in the right place and at the right time. Over 100 ligands for actin are currently known (Sheterline et al., 1998). Nature usually employs a highly economical strategy in which a relatively small number of fundamental protein modules perform specific catalytic and

14
Table 1. Actin-binding proteins. Classification and cellular functions of those actin-binding proteins that are mentioned in the text.
Class Proteins Function
Motor proteins myosins
Move along actin filaments carrying a cargo
Cross-linking proteins α-actinin filamin vinculin etc. ERM proteins
Cross-link actin into higher- order assemblies or to other structures e.g. plasma membrane
Regulators of actin polymerization
- actin monomer binding
- nucleating
- capping
- actin filament binding
ADF/cofilin twinfilin profilin thymosin β4 actobindin-family Arp2/3 capping protein gelsolin Abp1/drebrin Aip1 tropomyosin
Depolymerizes actin filaments Sequesters ADP-actin monomer Controls barbed end polymerization Sequesters ATP-actin monomers Initiates formation of new filaments as branched networks Cap filament ends and prevent polymerization Activates Arp2/3 Enhances ADF/cofilin’s activity Stabilizes filaments
Signaling WASPs
Link Rho GTPase signaling to Arp2/3 activation
recognition functions. These modules are then modified, shuffled and combined to form a spectrum of proteins able to carry out the broadest possible array of biological functions. Therefore, evolution has also produced only a relatively small number of actin-binding units. The four most common actin-binding modules are the calponin homology domain, the WH2 domain, the gelsolin homology domain and the actin-depolymerizing factor homology (ADF-H) domain (Lappalainen et al., 1998; Puius et al., 1998; Van Troys et al., 1999; Paunola et al., 2002).

15
4.1 The role of actin-binding proteins in actin filament dynamics The actin polymerization and depolymerization cycle can be dissected into smaller tasks, which must all be coordinated in order to create actin-based motility, for example at the leading edge of the cell. First of all, polymerization of actin requires a free filament end, preferably a barbed end, onto which actin monomers can be added. Free barbed ends can be derived from capped filaments. Uncapping of filaments can occur, for example, in response to phosphatidyl inositol (4,5)bisphosphate [PI(4.5)P2], which can remove the two most abundant barbed end cappers, capping protein and gelsolin, from the filament ends (Jamney, 1994; Shafer et al., 1996). It is also possible to create new filament ends by severing/fragmenting actin filaments. Gelsolin can sever actin filaments in response to an increase in the cytoplasmic Ca2+ level, which occurs, for example, during platelet activation (Barkalow et al., 1996). ADF/cofilins also exhibit a weak severing activity (see paragraph 4.2.4.1). However, in many cell types, actin polymerization appears to be initiated de novo. Arp2/3 complex is the first cellular factor to be identified possessing barbed-end nucleating activity. Arp2/3 is a stable complex of seven subunits, two of which are actin-related proteins that are thought to act as a template for actin polymerization (reviewed in Cooper et al., 2001). Arp2/3 can nucleate the formation of actin filaments from the side of another filament, producing a 70o branch that can be detected in the lamellipodia of moving cells (Svitkina and Borisy, 1999). As the Arp2/3 complex is an inefficient nucleator on its own, it requires activators. WASp/Scar family proteins act as primary activators, and actin filaments are powerful secondary activators of the Arp2/3 complex. WASp/Scar proteins also link the Arp2/3 complex to several signaling pathways (Machesky and Insall, 1999; Takenawa and Miki, 2001). Other proteins capable of activating the Arp2/3 complex include the Listeria protein ActA (Welch et al., 1998), Abp1 (Goode et al., 2001), cortactin (Uruno et al., 2001), Pan1p (Duncan et al., 2001) and myosin (Lee et al., 2000). Actin filament polymerization also requires a supply of actin monomers, the “building blocks”. The pool of unpolymerized actin in non-muscle cells is approximately 50% (~ 100 µM) of the total cytoplasmic actin. This is well above the Cc for actin polymerization (~ 0.2 µM), and cells have evolved two basic mechanisms to maintain such a large actin monomer pool. Capping proteins can block the addition of actin monomers onto filaments. Barbed ends are capped by the capping protein Cap1/2 and pointed ends by the Arp2/3 complex. Both exist in cells at micromolar concentrations and have nanomolar affinity for filament ends, thus efficiently capping most filament ends (reviewed in Pollard et al. 2000). Cells also contain proteins that bind directly to actin monomers and affect their polymerization properties. These proteins include profilin, thymosin β4, ADF/cofilin and twinfilin, which are discussed more thoroughly in the next chapter. Once free barbed ends are created and cells contain an adequate supply of actin monomers, actin filaments can elongate. For the barbed ends this is a diffusion-limited process that is influenced by high concentrations of

16
macromolecules in the cell and the excluded water molecules (Drenckhahn and Pollard, 1986; Luby-Phelps, 1994). Capping the barbed end terminates filament growth. Capping protein prevents both association and dissociation of monomers at the barbed end. It has been estimated that under cellular concentrations of capping protein, a free barbed end will have a half life of about 0.25 seconds during which it can elongate by >200 monomers. Unless actively uncapped or severed, a capped filament will probably never elongate again (reviewed in Pollard et al., 2000). Finally, old actin filaments must be depolymerized. Hydrolysis of ATP after the addition of actin subunits to the filament and the dissociation of the pyrophosphate are postulated to mark filaments for depolymerization by ADF/cofilins, which are discussed more thoroughly in section 4.2.4. Some filaments may be protected from depolymerization in order to create the stable actin assemblies found in cells. For example, tropomyosin can prevent ADF/cofilin-induced actin depolymerization (Ono and Ono, 2002).
4.2 Actin monomer-binding proteins The importance of the maintenance of a large cellular actin monomer pool was already explained in section 4.1. Besides preventing the association of actin monomers to the ends of filaments by capping them, cells contain actin monomer-binding proteins that directly affect the polymerization capacity of the monomers. Three classes of actin monomer-binding proteins are conserved throughout evolution, existing in organisms as diverse as yeast and mammals. These are ADF/cofilins, profilins, and twinfilins. In addition to these ubiquitous molecules, a fourth class of small actin monomer-binding proteins, β-thymosins, is present in vertebrates. Some additional proteins also contain actin monomer-binding motifs, among other domains (for example WASPs and Ena/VASP proteins), but these proteins are not as abundant as the above-mentioned small actin-binding proteins, and their main function is the signaling to the actin cytoskeleton. These large multifunctional proteins are not further discussed here. 4.2.1 Profilin Profilin is a small (12-16 kDa) actin-binding protein that has been identified in all eukaryotic cells investigated so far. Several organisms, including maize, cow, mouse and man, also express more than one isoform of profilin. The importance of profilins for cell proliferation has been demonstrated in genetic studies: deletion of the profilin gene is either lethal or the organisms are impaired in growth and motility (reviewed in Schluter et al., 1997). Profilin interacts directly with actin, acidic phospholipids and several proteins containing polyproline stretches. It binds actin monomers in a 1:1 stoichiometric complex with a micromolar affinity, preferring ATP-actin to ADP-actin. Profilin can affect actin dynamics in several ways. In the absence

17
of a free filament barbed end, profilin functions as an actin monomer-sequestering protein, because profilin-G-actin complexes do not participate in polymerization at the pointed end. Profilin also inhibits nucleation of actin filaments. On the contrary, in the presence of free barbed ends, profilin promotes barbed end assembly (Pantaloni & Carlier, 1993). Most profilins also enhance the nucleotide exchange on actin, and, at least in yeasts S. cerevisiae and S. pombe, this activity is essential in vivo (Wolven et al., 2000; Lu and Pollard, 2001). Profilin optimizes the directionality of treadmilling, because profilin-G-actin complexes associate only with the barbed ends, whereas free actin monomers can associate with both ends of actin filaments (Didry et al., 1998). The binding sites of actin and phospholipids overlap on profilin, and the binding of these ligands is mutually exclusive. Binding to PI(4,5)P2 can localize profilin to the plasma membrane. This interaction also links profilin to the phosphatidylinositol cycle, because profilin can protect various phosphatidylinositols from hydrolysis (Schluter et al., 1997). Binding to poly-L-proline is essential for the viability of at least fission yeast (Lu and Pollard, 2001). Potential protein ligands with proline rich sequences include the Ena/VASP family, N-WASP, Scar/WAVE, verprolin/WIP, p140mDia, cappuccino, Bni1p and Bnr1p, cdc12, drebrin, gephyrin, SMN and aczonin (Schuter et al., 1997). Therefore, profilin can link several signaling pathways to actin polymerization. In mammals, there are at least three genes encoding for profilin. The profilin Ι gene encodes the major isoform, having a rather ubiquitous expression pattern (Witke et al., 1998). The profilin ΙΙ gene is transcribed as two splice variants, profilin ΙΙa and profilin ΙΙb. Profilin ΙΙa is the major profilin isoform in brain tissue, while profilin ΙΙb is expressed in only a few tissues (Di Nardo et al., 2000; Lambrechts et al., 2000). These profilin isoforms appear to have similar affinities for actin, but different affinities for phosphoinositides and proline-rich sequences (Lambrechts et al., 1997). Recently, a third profilin gene, profilin ΙΙΙ, was identified. This isoform seems to be selectively expressed in the testis (Braun et al., 2002). 4.2.2 Thymosin β4 The β-thymosins comprise a family of small peptides originally thought to be thymic hormones. Currently, 14 β-thymosins have been described from various vertebrate species, and they form a family of highly conserved polar 5 kDa peptides consisting of 40-44 amino acid residues. In most mammalian tissues investigated, two β-thymosins are expressed. Thymosin β4 is usually the main peptide, and its cellular concentration can be up to 0.4 mM (reviewed in Huff et al., 2001). Thymosin β4 and the other β-thymosins examined so far, form a 1:1 complex with G-actin and inhibit actin polymerization at approximately equimolar concentrations (Hannappel and Wartenberger, 1993; Yu et al., 1993; Heinz et al., 1993). Thymosin β4 binds about 50-fold more strongly to ATP-actin than

18
to ADP-actin, and the dissociation constants for the complex are approximately 0.5-2.5 µM. (Pantaloni and Carlier, 1993). It has been suggested that thymosin β4 competes with profilin for the overlapping binding site on actin, because profilin can shuttle actin away from thymosin β4 onto the barbed end of the actin filament. The exchange is rapid, owing to the high dissociation rate constants. Therefore, ATP-actin monomers would partition between profilin and thymosin β4 based on the affinities and concentrations of these proteins (reviewed in Pollard et al., 2000). Recent results indicate, however, that profilin, thymosin β4 and actin can form a ternary complex. This would result in a very large amplification mechanism by which profilin and thymosin β4 can sequester much more actin than would be possible for either protein acting alone (Yarmola et al., 2001). There is also some evidence that thymosin β4 is not just a simple G-actin sequestering protein. At least in vitro, the ability of thymosin β4 to depolymerize actin filaments decreases with increasing concentrations of thymosin β4. Furthermore, at concentrations above 100 µM, thymosin β4 can be crosslinked to actin filaments, and at low molar ratios it can be incorporated into filaments (Carlier et al., 1996). Interestingly, thymosin β4 has also been detected outside the cells in blood plasma and in wound fluid. Although the role of the extracellular thymosin β4 is not known, several functions have been attributed to this fraction. Among these are induction of metallo-proteinases, chemotaxis, angiogenesis and inhibition of inflammation and bone marrow stem cell proliferation (Huff et al., 2001). A β-thymosin-like motif is called the WH2 domain (WASP-homology domain 2). This 35 residue motif has been found in many proteins regulating the actin cytoskeleton, such as WASP, WIP (WASP-interacting protein), Srv2/CAP (adenylyl cyclase associated protein), actobindin and in several currently uncharacterized proteins (Paunola et al., 2001). The actobindin family is interesting because it consists of proteins that are entirely composed of two to four copies of WH2 domains. Interestingly, these proteins bind actin in a 1:1 complex similarly to β-thymosins, but they are able to participate in barbed end growth similarly to profilins. Furthermore, proteins of the actobindin family interact with the pointed end of the actin filament and thus prevent pointed end growth (Boquet et al., 2000; Hertzog et al., 2002). 4.2.3 ADF-H domain proteins The ADF-H domain is a widely occurring actin-binding module present in three phylogenetically distinct protein classes: the ADF/cofilins, twinfilins and Abp1/drebrins. Each class has existed in eukaryotic organisms for more than a billion years since the divergence of fungi and animals. The ADF-H domain can be considered as a building block that is arranged differently in the three protein classes (Figure 3). ADF/cofilins are the prototypes, consisting solely of a single ADF-H domain. Twinfilins, on the other hand, are composed of two ADF-H domains, while Abp1/drebrins have a single ADF-H domain in their N-terminus, followed by a non-conserved central region and a C-terminal Src homology 3 (SH3) domain. Intriguingly, each class seems to possess distinct biochemical functions. While ADF/cofilins are able to interact with both actin

19
monomers and filaments, twinfilins bind only monomers and Abp1/drebrins only filaments. Therefore, ancient duplication of the ADF-H domain and combination with other protein modules have allowed the development of functional diversity (Lappalainen et al., 1998). The properties of ADF/cofilins and twinfilins are discussed below, since they participate in the regulation of the actin monomer pool, whereas Abp1/drebrins have been implicated as links between the actin cytoskeleton and endocytosis (Kessels et al., 2001).
Figure 3. Domain structures of the ADF-H domain containing proteins. Adapted from Lappalainen et al., (1998).
4.2.4 ADF/Cofilin ADF/cofilins are small (15-19 kDa), globular proteins composed of a single ADF-H domain (Figure 3). They are ubiquitous among eukaryotes, but unicellular organisms, such as yeast, have only one ADF/cofilin protein, whereas multicellular organisms can have several ADF/cofilin isoforms (Lappalainen et al., 1998). Three dimensional structures of human destrin (Hatanaka et al., 1996), Saccharomyces cerevisiae cofilin (Fedorov et al., 1997), Acanthamoeba actophorin (Leonard et al., 1997) and Arabidopsis thaliana ADF1 (Bowman et al., 2000), have been determined either by x-ray crystallography or by NMR. The ADF/cofilin fold consists of five central β-sheets flanked by three to four α-helices. A comparative analysis of the different ADF/cofilin structures shows that the actin-binding surface has remained more conserved than other regions of the surface (Bowman et al., 2000). 4.2.4.1 Interactions with actin ADF/cofilins interact with both actin monomers and filaments. Under physiological conditions, ADF/cofilins bind ADP-actin monomers and filaments with higher affinity than ATP-actin (Maciver and Weeds, 1994; Carlier et al., 1997; Blanchoin and Pollard, 1998, 1999). ADF/cofilins also inhibit nucleotide exchange on actin. The binding of ADF/cofilins to actin filaments is cooperative (Hawkins et al., 1993; Hayden et al., 1993), and the binding induces a twist in the filament, changing its thermodynamic stability (McGough et al., 1997; McGough and Chiu, 1999). ADF/cofilins are also able to sever actin filaments. The extent of severing depends on the concentration of ADF/cofilin and on time, but the number of breaks is much lower than the

20
number of ADF/cofilins bound (Blanchoin and Pollard, 1999), making ADF/cofilin a weak severer, as compared to, for example, gelsolin. The most important physiological function of ADF/cofilins is to depolymerize actin filaments from their pointed ends, thereby increasing actin filament turnover. ADF/cofilins can increase the depolymerization rate from the pointed ends by about 30-folds. (Carlier et al., 1997). The actin filament disassembly activity is enhanced by actin-interacting protein 1 (Aip1), which interacts with both actin filaments and ADF/cofilin (Rodal et al., 1999). This protein also regulates the localization of ADF/cofilin in cells (Ono, 2001). Profilin competes with ADF/cofilins for binding to actin monomers, and physiological concentrations of profilin can overcome the inhibition of nucleotide exchange by ADF/cofilins (Blanchoin and Pollard, 1998). Profilin rapidly recycles ADP-actin monomers to the profilin-ATP-actin monomer pool, which is readily polymerized, and thus profilin synergizes with ADF/cofilins to enhance the turnover of actin filaments to a value 125-fold higher than in pure F-actin solutions (Didry et al., 1998). Tropomyosin inhibits binding of ADF/cofilins to actin filaments due to a difference in filament structure when tropomyosin or ADF/cofilin is bound (McGough, 1998). Tropomyosin also acts as a physiological inhibitor of ADF/cofilin-induced actin dynamics (Ono and Ono, 2002). Therefore, cells seem to contain two F-actin pools: stable actin filaments associated with tropomyosin and dynamic filaments accessible to ADF/cofilins. 4.2.4.2 Cell biological properties The activity of ADF/cofilins is fundamental to cells, because ADF/cofilin-inactivating mutations are lethal in every organisms tested so far (Moon et al., 1993; McKim et al., 1994; Gunsalus et al., 1995). ADF/cofilins are essential for those cellular processes that require a dynamic actin cytoskeleton, such as cell movement (Chen et al., 2001), cytokinesis (Moon et al., 1993; Gunsalus et al., 1995), phagocytosis (Nagaishi et al., 1999) and fluid phase endocytosis (Lappalainen and Drubin, 1997). Furthermore, the importance of ADF/cofilins as key regulators of actin dynamics is demonstrated by the fact that they are indispensable for the motility of the small intracellular pathogen Listeria monocytogenes (Loisel et al., 1999). In cells, ADF/cofilins are predominantly cytoplasmic, but upon activation they concentrate at the leading edge and ruffling membranes of motile cells. ADF/cofilins also localize to other structures displaying high actin filament turnover rates, such as the contractile ring and neuronal growth cones (reviewed in Bamburg, 1999). ADF/cofilins contain a putative nuclear localization signal (NLS), and they can be transported to the nucleus in response to various stress factors (Abe et al., 1993). In the nucleus ADF/cofilins form rods with actin. As actin itself does not contain a NLS, ADF/cofilins could be the transporters that mediate the localization of actin to the nucleus. It has been postulated that rod formation is a mechanism to conserve ATP in stress situations by decreasing actin filament dynamics, which is a major consumer of ATP (reviewed in Bamburg, 1999).

21
4.2.4.3 Regulation by pH, phosphoinositides and phosphorylation Some, but not all ADF/cofilin isoforms are regulated by pH. The pH dependency is the result of an increase in critical concentration for the assembly of the ADF/cofilin-actin complex to actin filaments at elevated pH (Ressad et al., 1998). The actin-binding and depolymerizing activities of ADF/cofilins can also be regulated by PI(4)P and PI(4,5)P2, which inhibit ADF/cofilin binding to actin in vitro (Yonezava et al., 1990), because the binding site for the phospholipid and actin overlap on the surface of ADF/cofilins (Ojala, et al., 2001). Most ADF/cofilins are specifically phosphorylated at serine-3 (Agnew et al., 1995). This phosphorylation inhibits the activity of ADF/cofilin by reducing its affinity for actin by 20-fold (Ressad et al., 1998). Several kinases responsible for this inactivation have been characterized. LIM kinases are composed of N-terminal LIM domains and a C-terminal kinase domain. LIM kinase 1 is expressed mainly in the central nervous system, but LIM kinase 2 has a more ubiquitous expression pattern. LIM kinases connect the Rho family of small GTPases, which are discussed in section 5.1, to the regulation of actin filament depolymerization by ADF/cofilins (Arber et al., 1998; Yang et al., 1998). Testicular protein kinases (TESK) 1 and 2 also phosphorylate ADF/cofilin on the same serine residue (Toshima et al., 2001a;b), but these kinases seem to be controlled by integrin mediated signaling pathways through 14-3-3 proteins. These proteins bind both TESK and integrins, and they can regulate the kinase activity, as well as subcellular localization of TESK (Toshima et al., 2001c). Other kinases phosphorylating ADF/cofilins include a calmodulin-like domain protein kinase (Allwood et al., 2001) and a neutrophil-specific kinase (Lian et al., 2000). The activation of ADF/cofilins is induced by dephosphorylation, which occurs rapidly in response to various stimuli, such as growth factors, chemotactic peptides, and agents increasing the intracellular levels of Ca2+ and cAMP. The dephosphorylation could result from down-regulation of a kinase or up-regulation of a phosphatase, or both (reviewed in Moon and Drubin, 1995). Also an ADF/cofilin-specific phosphatase, called Slingshot, has been recently idenfied in Drosophila and humans (Niwa et al., 2002). However, it is still not known how the activity of this phosphatase is regulated. 4.2.4.4 ADF/cofilin isoforms Multicellular organisms can have several isoforms of ADF/cofilins, either as a result of alternative splicing of a single gene or due to several genes encoding ADF/cofilins. For example, Xenopus laevis has two allelic variants of ADF/cofilins that differ only by 12 amino acids (Abe et al., 1996). Also Caenorhabditis elegans produces two ADF/cofilin transcripts by alternative splicing, resulting in proteins Unc60A and Unc60B, which are 36% identical. Unc60A is essential for the viability of C.elegans, and Unc-60B is required for the development of muscle. These isoforms also have distinct biochemical properties. Unc60A tends to depolymerize actin filaments and inhibits polymerization, whereas Unc60B binds strongly to actin filaments and can

22
change the rate of actin polymerization (Ono & Benian, 1998). Plants are peculiar organisms, because they can contain over ten ADF/cofilin proteins (Maciver and Hussey, 2002), but lack altogether the other two ADF-H domain- containing proteins twinfilins and Abp1/drebrins. For example in maize, there are three ADF/cofilin genes; two are expressed solely in pollen, and the third is expressed in vegetative tissues (Lopez et al., 1996). In previous studies, two ADF/cofilin isoforms have been reported to exist in mice, pigs and chicken (Lappalainen et al., 1998), whereas three ADF/cofilins are found in humans (Ogawa et al., 1990; Hawkins et al., 1993; Thirion et al., 2001). The two porcine isoforms display some biochemical differences in actin co-sedimentation assays, and they are both widely distributed among tissues (Moriyama et al., 1990). During muscle development in mouse, there is a switch from the rather ubiquitously expressed cofilin-1 to cofilin-2, which is a mainly muscle- and testes-specific isoform (Ono et al., 1994). The exact biochemical differences between the mammalian isoforms are not known. 4.2.5 Twinfilin Twinfilin was originally identified in yeast Saccharomyces cerevisiae through its sequence homology to ADF/cofilin proteins (Lappalainen et al., 1998, Goode et al., 1998). The human homologue of twinfilin, on the other hand, was characterized as a novel tyrosine kinase A6 (Beeler et al., 1994). Homologues of twinfilin have also been found in S. pombe, Drosophila melanogaster, Caenorhabditis elegans, and mice, suggesting that twinfilins are present across the entire spectrum of eukaryotic organisms (Lappalainen et al., 1998). Curiously, a homologue of twinfilin has not been detected in plants to date. 4.2.5.1 Twinfilin is composed of two ADF-H domains Twinfilin is a 37-40 kDa protein composed of two ADF-H domains that are connected by a short linker region and followed by a C-terminal tail (Figure 3). The two ADF-H domains are ~ 20% homologous to ADF/cofilin and to each other (Lappalainen et al., 1998). The individual twinfilin ADF-H domains are more similar across species than the N- and C-terminal ADF-H domains are within species, indicating that the ADF-H domain was duplicated once before the divergence of fungal and animal lineages (Palmgren et al., 2002). Both ADF-H domains of twinfilin probably have tertiary structures similar to those of ADF/cofilins because the insertions in the twinfilin ADF-H domains are located in predicted loop regions. Also, the residues important for actin monomer-binding are well conserved in both twinfilin ADF-H domains (Lappalainen et al., 1998). Mutagenesis studies on yeast twinfilin have demonstrated that these corresponding residues are important for actin monomer binding (Palmgren et al., 2001), suggesting that twinfilin and ADF/cofilin interact with actin monomers with at least partly overlapping interfaces. In contrast, the residues that are important for actin filament binding in ADF/cofilins, are not conserved in twinfilins (Lappalainen et al.,1998).

23
4.2.5.2 Biochemical and cell biological activities of yeast twinfilin Yeast twinfilin is an actin monomer-binding protein that, unlike ADF/cofilin, does not bind to actin filaments. Instead it forms a 1:1 complex with actin monomers and prevents actin filament assembly by sequestering the monomers. Yeast twinfilin also inhibits spontaneous nucleotide exchange on actin monomers (Goode et al., 1998). Like ADF/cofilins, twinfilin binds ADP-actin monomers with higher affinity than ATP-actin monomers (Palmgren et al., 2001). Deletion of the twinfilin gene in yeast does not produce a clear phenotype, but results in abnormal cortical actin patches and defects in bipolar budding pattern (Goode et al.,1998). This deletion also shows synthetic lethality with certain cofilin and profilin mutants that exhibit diminished actin filament depolymerization and defects in actin nucleotide exchange activity, respectively (Lappalainen & Drubin, 1997; Goode et al., 1998; Wolven et al., 2000). Moreover, overexpression of twinfilin in yeast results in depolarization of the actin cytoskeleton and accumulation of actin bars in the cytoplasm (Goode et al., 1998). In yeast, twinfilin localizes to the cytoplasm but is also enriched in the actin patches, which are sites of active filament dynamics. Localization to the actin patches depends on an intact actin cytoskeleton. A mutant twinfilin that is no longer able to bind to actin monomers does not localize to actin patches, suggesting that the ability to interact with actin monomers is essential for the correct localization of yeast twinfilin. The localization of twinfilin to cortical actin patches is also disrupted in yeast strains where genes encoding for the two subunits of capping protein, Cap1/2, are deleted. Based on native gel electrophoresis and co-immunopresipitation assays, twinfilin and capping protein also interact directly with each other. These results suggest that the role of twinfilin may be to localize actin monomers, in their inactive ADP-form, to the sites of actin assembly in cells (Palmgren et al., 2001).
5. Signaling to the actin cytoskeleton An animal cell contains an elaborate system of proteins that enables the cell to respond to signals from other cells. This allows each cell to determine its position and specialized role in the body of a multicellular organism. This system includes cell surface and intracellular receptor proteins, protein kinases and phosphatases, GTP-binding proteins and secondary messengers as well as a vast number of intracellular proteins with which these components interact (reviewed in Barritt, 1996). Many signaling pathways converge on the actin cytoskeleton. The targets of these signaling pathways are often actin-binding proteins, but also actin itself can be, for example phosphorylated or ADP-ribosylated, although the in vivo role of these post-translational modifications is not known (reviewed in Sheterline et al., 1998). The two main signaling systems that regulate the actin cytoskeleton are the

24
Rho family of small GTPases and the phosphoinositide lipid PI(4,5)P2 (Figure 4).
5.1. Rho family GTPases Rho GTPases are members of the Ras superfamily of monomeric GTP-binding proteins. Ten different mammalian Rho GTPases have been identified to date, and the most studied members are Rho, Rac and Cdc42. These proteins cycle between an inactive GDP-bound, and active GTP-bound state. In the GTP-bound form, Rho GTPases can interact with effector molecules to initiate a downstream response. The intrinsic GTPase activity then returns the proteins to the GDP-bound status. Numerous cellular proteins regulate cycling between the GTP- and GDP-bound states. Guanosine nucleotide exchange factors (GEFs) facilitate the exchange of GDP for GTP, and thus act as activators. GTPase-activating proteins (GAPs) increase the intrinsic rate of GTP hydrolysis of Rho GTPases facilitating the inactivation process. Rho GTPases are able to interact with membranes via post-translational lipid modifications, but they can also be sequestered to the cytoplasm in their inactivated state by a protein called RhoGDI, which also inhibits the exchange of GDP to GTP (reviewed in Hall, 2000). 5.1.1. Rho regulates actomyosin-based structures The major function of Rho GTPases is to link membrane receptors to the assembly and organization of the actin cytoskeleton. Each Rho GTPase has unique effects on actin, due to different effector proteins, of which over 30 have been identified. Rho affects the actin cytoskeleton by stimulating the formation of actomyosin-based structures and by regulating their contractility. Activation of Rho causes the accumulation of stress fibers in cells, and Rho is in fact needed for the formation of stress fibres induced by both soluble factors and integrin engagement. This is also accompanied by the formation of focal adhesions (reviewed in Bishop and Hall, 2000). ROK (Rho-associated kinase) is a Rho effector thought to be a key player in actomyosin assembly. ROK can induce the phosphorylation of myosin light chains (MLC), both by inhibiting the MLC phosphatase and by phosphorylating the MLC itself. Phosphorylation of the MLC stimulates the actin-activated ATPase activity of myosin II and promotes the assembly of actomyosin filaments (reviewed in Bresnick, 1999). Another target of ROK is LIM kinase, which is able to phosphorylate and inactivate ADF/cofilins, leading to stabilization of actin filament structures (Maekawa et al., 1999). ROK also activates Na+/H+ exchange protein (NHE1) and this contributes, via an unknown mechanism, to stress fiber and focal adhesion formation (Vexler et al., 1996). Other ROK substrates contributing to actin assembly are the adducin and ERM (ezrin/radixin/moesin) family of proteins (reviewed in Bishop and Hall, 2000).

25
Figure 3. Signaling to the actin cytoskeleton. The two main signaling pathways regulating the structure and dynamics of the actin cytoskeleton are the Rho-GTPases and PI(4,5)P2. Proteins that are able to integrate signals from both pathways are boxed. Actin-binding proteins that are targets of these signaling pathways are in bold. Modified from Bishop and Hall, (2000), Sechi and Wehland, (2000) and Ridley et al., (2001). ROK alone is not able to stimulate the formation of stress fibers, since it requires the assistance of another Rho effector, Dia. Dia is a member of the formin-homology (FH) family of proteins, and it is able to bind to the actin monomer-binding protein profilin. This allows Dia to promote actin polymerization in cells (Watanabe et al., 1999). 5.1.2. Rac and Cdc42 regulate the formation of lamellipodia and filopodia Rac regulates the formation of lamellipodia and membrane ruffles in a variety of cell types, while Cdc42 controls the formation of filopodia. Rac and Cdc42 have several common effectors, but some are unique to either GTPase (reviewed in Bishop and Hall, 2000). Cdc42 is able to bind to WASP and N-WASP, which are activators of the Arp2/3 complex. The activated Arp2/3 can then nucleate polymerization of actin filaments (Rohatgi et al., 1999). Cdc42 can also induce phosphorylation of MLC, via myotonic dystrophy kinase-related Cdc42-binding kinase (MRCK) (Leung et al., 1998). Furthermore, MRCKα is able to activate LIM kinases 1 and 2 and thus induce inactivation of ADF/cofilins (Sumi et al., 2001). Also Cdc42 effector PAK4, a novel PAK family member, can activate LIM kinase 1 (Dan et al., 2001). Only a few Rac-

26
specific effectors are known. Por-1 has been implicated in Rac-induced lamellipodia formation, but the mechanism is unknown. Another Rac-specific effector is p140Sra-1 which is able to co-sediment with actin filaments (Bishop and Hall, 2000). Also Rac can activate Arp2/3 through proteins of the Scar/WAVE family, but it is not known if the interaction is direct (Miki et al., 1998). Common targets for Rac and Cdc42 include PAK kinases 1, 2 and 3. As PAK4, also these PAKs can activate LIM kinase (Arber et al., 1998, Edwards et al., 1999) and they decrease the MLC phosphorylation by inactivating the MLC kinase (Sanders et al., 1999). This leads to reduced assembly of actomyosin structures and is antagonistic to the effects of Rho. IQGAPs are able to bind both calmodulin and actin filaments, and it has been suggested that binding to Cdc42 or Rac could facilitate the crosslinking activity of these proteins (Brill et al., 1996; Hart et al., 1996)
5.2. PI(4,5)P2 Initially, PI(4,5)P2 was regarded merely as a precursor of the second messengers inositol 1,4,5-trisphosphate and diacylglycerol, which are formed in response to agonist-mediated hydrolysis of PI(4,5)P2. Recent studies have, however, established PI(4,5)P2 itself as a key factor in transmitting signals to the actin cytoskeleton as well as in vesicle trafficking. PI(4,5)P2 can accumulate at membrane rafts and promote local recruitment and activation of signaling components at the cell membrane, which may promote for example actin filament accumulation and dynamics (reviewed in Sechi and Wehland, 2000; Caroni, 2001). In general, PI(4,5)P2 promotes actin polymerization and enhances crosslinking of actin filaments as well as their linkage to the plasma membrane. PI(4,5)P2 has been reported to affect the function of over 15 actin-binding proteins. In the case of actin monomer-binding proteins ADF/cofilin (Yonezava et al., 1990), profilin (Sohn et al., 1995) and twinfilin (Palmgren et al., 2001), PI(4,5)P2 causes dissociation of the actin monomer, thereby preventing the sequestering activities of these proteins near the plasma membrane. PI(4,5)P2 can also promote actin polymerization by uncapping the filaments. It is able to dissociate both capping protein and gelsolin from the barbed end of the filament (Heiss & Cooper, 1991; Jamney & Stossel, 1987), creating readily polymerizable filament ends. The actin crosslinking proteins, α-actinin and filamin, are differentially regulated by PI(4,5)P2. The crosslinking activity of α-actinin is enhanced and that of filamin reduced in response to PI(4,5)P2. PI(4,5)P2 also regulates the linkage of actin filaments to the plasma membrane via talin and induces a conformational change in vinculin, regulating its binding to other cytoskeletal proteins (Sechi and Wehland, 2000). Cells contain proteins that can integrate both PI(4,5)P2 and Rho GTPase-mediated signals and transmit them to the actin cytoskeleton. WASP proteins were already mentioned as effectors of Cdc42 and as activators of the actin nucleation machinery, the Arp2/3 complex. In their N-terminal region these proteins contain a putative PH-domain, which binds PI(4,5)P2 (Rohatgi et al.,

27
1999). Both this domain as well as the GTPase-binding domain appear to be critical for regulation of the activity of WASP-family members. Therefore a model has been proposed in which WASPs are activated by a GTPase and the activation is enhanced by binding PI(4,5)P2, which could also target WASPs to the plasma membrane (Higgs and Pollard, 2000). ERM proteins have been considered to be good candidates for linking the actin cytoskeleton to the plasma membrane because of their capability to interact both with actin filaments and membrane components such as CD44, CD43 as well as intercellular adhesion molecules (ICAMs). ERM proteins can exist either in an active state or in a dormant state, where the important binding sites are masked due to interactions between the N- and C-terminus of the protein. Rho is able to influence the phosphorylation state, activation and localization of ERM proteins. ERM proteins can also bind to PI(4,5)P2 and this seems to enhance the ability of these proteins to interact with plasma membrane proteins (reviewed in Tsukita and Yonemura, 1999). Besides interacting with the same effector proteins, PI(4,5)P2 and Rho GTPases can directly affect each other’s functions. Rac (and possibly also Rho) is able to act upstream of PI(4)P 5-kinase to induce PI(4,5)P2 synthesis. Rac is also able to interact with diacylglycerol kinase, which generates phosphatidic acid, a strong activator of PI(4)P 5-kinase, further increasing the production of PI(4,5)P2. On the other hand, PI(4,5)P2 is able to disrupt the interactions between Rac and RhoGDI, thereby allowing Rac to be activated and associated with membranes. PI(4,5)P2 has also been shown to stimulate the release of GDP from Cdc42 in vitro. Therefore, an initial increase in PI(4,5)P2 levels might activate the small GTPases, which in turn could stimulate the production of more PI(4,5)P2. The sustained production of PI(4,5)P2 may then cause further activation of the Rho GTPases (reviewed in Sechi and Wehland, 2000).

28
Aims of the study The actin cytoskeleton plays pivotal roles in various essential cell biological processes. The regulation of the structure and dynamics of the actin cytoskeleton consists of interdependent processes, all of which must take place in a temporally and spatially coordinated manner. A large array of actin- binding proteins govern this complex network in cells. The contribution of the individual components of the actin regulating machinery is still not completely understood, and even less is known of the complex interplay between these proteins. This study aims to elucidate one aspect of the actin filament dynamics, namely the regulation of size and dynamics of the actin monomer pool. The focus is on two proteins, ADF/cofilin and twinfilin, which share a common ADF-H domain. Specific aims were:
1. To understand why several ADF/cofilin isoforms exist in mammals
- Unicellular organisms have only one ADF/cofilin protein, whereas several isoforms exist in multicellular organisms. The biochemical differences of the mammalian ADF/cofilin isoforms are not known, and detailed expression analysis is also lacking. Moreover, different research groups have used distinct isoforms in their studies, and this has made it difficult to compare individual results and to elucidate the true biological role of this essential protein family.
2. To elucidate the role of twinfilin in the regulation of actin dynamics in multicellular organisms
- Twinfilin is a small, ubiquitous actin monomer-binding protein
that is conserved throughout evolution. This protein has previously been studied only in the unicellular yeast S. cerevisiae, but the lack of phenotype in the twinfilin deletion strain has hampered our understanding of the cell biological role of twinfilin. Database searches have identified homologues of yeast twinfilin in mammals and in Drosophila, but the biochemical and cell biological activities of these proteins have not been characterized.

29
Materials and methods The experimental methods used in this PhD-thesis project are listed in Table 2. The detailed description of each method is found in the original publication. Table 2. Experimental methods used in this study. Method Publication Plasmid construction Ι, ΙΙ, ΙΙΙ SDS-PAGE Ι, ΙΙ, ΙΙΙ Western blotting ΙΙ, ΙΙΙ Northern blotting Ι, ΙΙΙ Whole Mount In Situ Hybridization Ι Radioactive In Situ Hybridization Ι Cell culture Ι, ΙΙΙ Transfection of cultured animal cells Ι, ΙΙΙ Affinity purification of antibodies ΙΙΙ Immunofluorescence Ι, ΙΙΙ Protein expression and purification Ι, ΙΙ, ΙΙΙ Actin co-sedimentation assay Ι, ΙΙ, ΙΙΙ Depolymerization of Alexa-actin Ι Actin treadmilling assay Ι NBD-actin assay Ι Actin filament assembly assays using pyrene actin ΙΙ

30
Results
6. The three mouse ADF/cofilins have distinct expression patterns as well as different biochemical properties (ΙΙΙΙ) In our database searches we identified three mouse ADF/cofilin isoforms, of which two were previously characterized muscle and non-muscle cofilins. The third one had the highest sequence homology to chicken and porcine ADF and to human destrin. Furthermore, phylogenetic analysis of all known mammalian and avian ADF/cofilins grouped these proteins into three distinct classes, which we named cofilin-1, cofilin-2 and ADF. We used in situ hybridization and Northern blotting to analyze the expression patterns of the three mouse ADF/cofilin isoforms during development and in adult mice. Whole mount in situ hybridization of embryonic day (E) 9.5 mice revealed that the only isoform expressed at this stage of development is cofilin-1. In E14 embryos cofilin-2 and ADF expression were also detected in the developing muscle and epithelial tissues, respectively, while cofilin-1 was still expressed throughout the embryo. Based on Northern blot analysis of adult mouse tissues, all three ADF/cofilins are expressed at variable amounts in many organs, but the only isoform expressed in mature skeletal muscle is cofilin-2. Closer examination of the whisker pad, skin and testes by in situ hybridization revealed that the isoforms are selectively expressed in different tissues within these organs. Cofilin-2 appears to be expressed mainly in muscle cells, although it is also found in seminiferous tubules of the testis. The expression of ADF is mainly restricted to epithelial tissues, while cofilin-1 can be detected in many tissues and most of their cell types. To study the biochemical properties of the three mouse ADF/cofilin isoforms, we expressed them as GST-fusion proteins in E.coli. Both muscle and platelet actins were used in these experiments. The actin cosedimentation assay showed that all isoforms bind equally well to actin filaments and do not show any actin isoform specificity. The same assay was applied to study the abilities of the ADF/cofilins to disassemble actin filaments. In this assay, both cofilin-1 and ADF were able to increase significantly the amount of monomeric actin, while in contrast cofilin-2 increased the amount of actin filaments. Although the ADF/cofilins did not display any actin isoform specificity in this assay, they all shifted platelet actin more easily to the monomeric fraction than muscle actin. We also tested the effect of pH in a similar assay, and found that the only isoform regulated by pH is ADF. Next we examined why cofilin-2 was not able to disassemble actin filaments in the cosedimentation assay. First we used a visual assay to examine whether this was due to a defect in actin depolymerization or due to fragmenting activities of the protein. This assay demonstrated that all isoforms were able to shorten fluorescent Alexa 488-actin filaments to a similar extent. In a more quantitative actin turnover assay, cofilin-2 was the least efficient isoform in increasing actin filament turnover, while ADF was the most efficient. We also measured the affinities of these mouse ADF/cofilins for actin monomers using

31
NBD-actin. In agreement with previous results, we found that all mouse ADF/cofilins have higher affinity for ADP-actin monomers than for ATP-actin monomers. The affinities for ADP-actin monomers were similar for each isoform, but cofilin-2 displayed 5-10 times higher affinity for ATP-actin monomers than the other two isoforms.
7. The role of twinfilin in Drosophila development (ΙΙΙΙΙΙΙΙ ) Twinfilin was originally characterized in yeast, where it has since been extensively studied (Goode et al., 1998; Palmgren et al., 2001). The biological role of twinfilin has, however, remained unclear, since the deletion of the twinfilin gene in yeast has only a mild phenotype. We identified a gene encoding a putative protein of 343 amino acids from the complete Drosophila genome, and showed that it is 26% identical to yeast twinfilin. We cloned the full-length open reading frame of Drosophila twinfilin from an embryonic cDNA library and expressed it as a GST-fusion protein in E.coli. After the cleavage of GST and purification with gel filtration column, the protein was monomeric and fully soluble. To examine whether Drosophila twinfilin has biochemical properties that are similar to those of yeast twinfilin we used an actin cosedimentation assay and assembly/disassembly assays with pyrene actin. Drosophila twinfilin did not copellet with actin filaments, but it was able to increase the amount actin monomers in the actin cosedimentation assay. Furthermore, Drosophila twinfilin prevented the assembly and promoted the disassembly of pyrene actin filaments, indicating that Drosophila twinfilin is an actin monomer-sequestering protein and a functional homolog of yeast twinfilin. The recombinant Drosophila twinfilin was also used to raise an antiserum in a rabbit. The antiserum proved to be specific recognizing only a single ~ 40 kDa band from the extracts of Drosophila larvae. This antiserum was then used to study the expression of twinfilin during fly development. A fly strain was characterized in which there is a P-element insertion in the first exon of the twinfilin gene. This strain is a strong and specific hypomorphic twinfilin mutant strain, and the expression of both twinfilin mRNA and protein are significantly reduced. Twinfilin mutant flies have several developmental defects, including rough-eye phenotype and aberrant bristle morphology. In the absence of twinfilin, the bristle actin bundles are misoriented and ectopic actin polymerization at the bristle surface between the main bundles takes place. Twinfilin also interacts genetically with Drosophila ADF/cofilin in bristle morphogenesis.
8. The role of twinfilin in mammals ((((ΙΙΙΙΙΙΙΙΙΙΙΙ ) Database searches revealed also mammalian homologues of twinfilin that had a 25% sequence identity to yeast twinfilin at the amino acid level (Lappalainen et al., 1998). Previous studies with these mouse and human proteins had not revealed any actin-binding activities in these proteins. Instead the studies

32
suggested that twinfilins constitute a novel class of protein tyrosine kinases and they were therefore named A6 protein kinases (Beeler et al., 1994, 1997). We expressed mouse twinfilin-1 as a GST-fusion protein in E. coli. Since yeast twinfilin does not possess any tyrosine kinase activity (Goode et al., 1998), we carried out an in vitro kinase assay under similar conditions as described by Beeler et al. (1994). No tyrosine kinase activity was detected with our recombinant twinfilin-1. Furthermore, we found that in an actin cosedimentation assay, twinfilin did not pellet with actin filaments, but it was able to increase the amount of actin monomers. This indicates that mouse twinfilin-1 is also an actin monomer-sequestering protein like yeast and Drosophila twinfilin. We also confirmed with rate zonal centrifugation that mouse twinfilin-1 forms a 1:1 complex with actin monomers. To examine the expression pattern of mouse twinfilin-1, a multiple tissue Northern blot analysis was carried out using twinfilin-1 cDNA as a probe. Twinfilin-1 was found to be expressed at variable amount in many tissues, but no expression of twinfilin was detected in the spleen or in skeletal muscle. To generate antibodies against mouse twinfilin-1, recombinant protein was used in immunization of rabbits. The rabbit antiserum was then affinity purified and its specificity assessed with Western blotting. The antibody recognized the recombinant mouse twinfilin and only a single band of 40 kDa in NIH 3T3 and N18 cell extracts, indicating that the antibody was specific for twinfilin. In NIH 3T3 cells, twinfilin-1 localized both to the cytoplasm and to the cell processes. These cell processes were also rich in both actin filaments and monomers, as revealed by phalloidin and DNAseΙ stainings, respectively. In addition, in N18 neuroblastoma cells, twinfilin-1 showed strong perinuclear staining as well as localization to actin-rich filopodia. Overexpression of twinfilin-1 in NIH 3T3 cells resulted in loss of stress fibers and appearance of abnormal actin filament structures. In addition, we found that the small GTPase Rac1, which regulates the formation of membrane ruffles and lamellipodia in cells (Ridley et al., 1992), regulates the localization of twinfilin-1 in NIH 3T3 cells.
9. The two mammalian twinfilin isoforms have similar biochemical properties but distinct expression patterns (unpublished) Our database searches revealed two twinfilin isoforms in mammals. Mouse twinfilin-1 and twinfilin-2 have 64% sequence identity. The human homolog of twinfilin-2 protein called A6-related protein had previously been characterized as a binding partner of the catalytic domain of the atypical protein kinase C ξ (Rohwer et al., 1999). We compared the expression patterns of the two mouse twinfilin isoforms. Based on Northern blot analysis (Figure 5A) and whole mount in situ hybridization of 9.5-day-old mouse embryos (Figure 5B), both twinfilin isoforms are expressed at constant levels during mouse development and
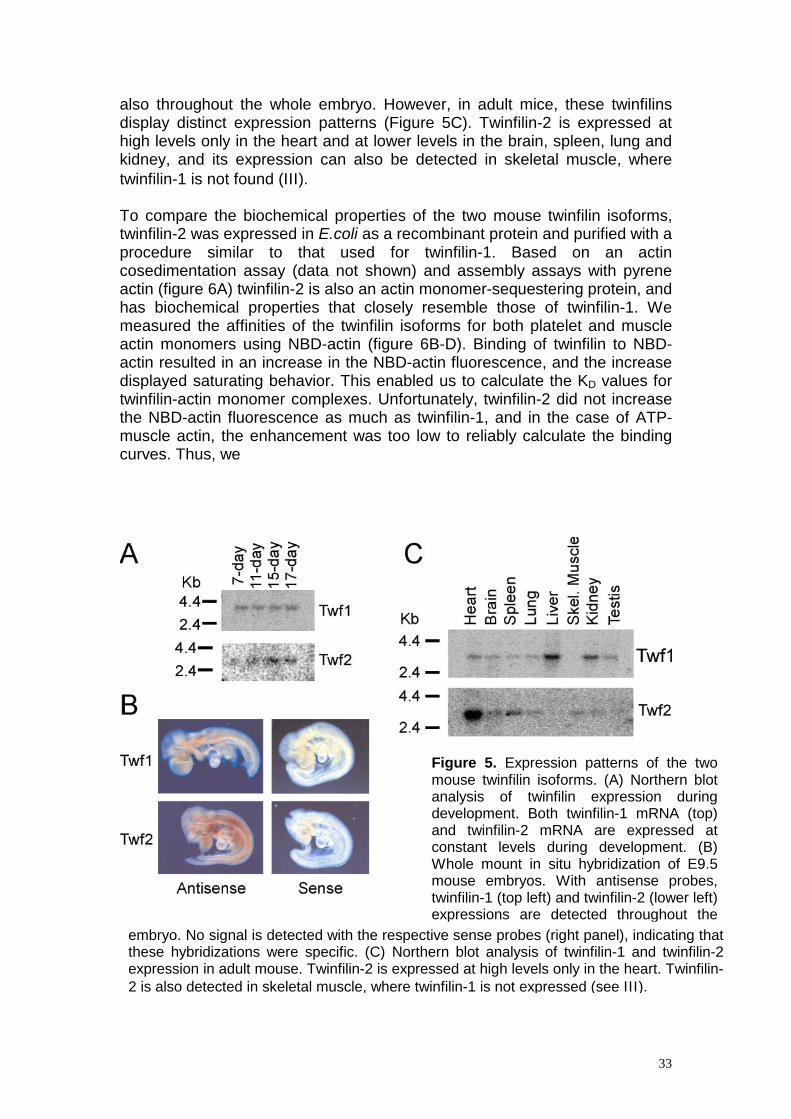
also throughout the whole embryo. However, in adult mice, these twinfilins display distinct expression patterns (Figure 5C). Twinfilin-2 is expressed at high levels only in the heart and at lower levels in the brain, spleen, lung and kidney, and its expression can also be detected in skeletal muscle, where twinfilin-1 is not found (ΙΙΙ ). To compare the biochemical properties of the two mouse twinfilin isoforms, twinfilin-2 was expressed in E.coli as a recombinant protein and purified with a procedure similar to that used for twinfilin-1. Based on an actin cosedimentation assay (data not shown) and assembly assays with pyrene actin (figure 6A) twinfilin-2 is also an actin monomer-sequestering protein, and has biochemical properties that closely resemble those of twinfilin-1. We measured the affinities of the twinfilin isoforms for both platelet and muscle actin monomers using NBD-actin (figure 6B-D). Binding of twinfilin to NBD-actin resulted in an increase in the NBD-actin fluorescence, and the increase displayed saturating behavior. This enabled us to calculate the KD values for twinfilin-actin monomer complexes. Unfortunately, twinfilin-2 did not increase the NBD-actin fluorescence as much as twinfilin-1, and in the case of ATP-muscle actin, the enhancement was too low to reliably calculate the binding curves. Thus, we
Figure 5. Expression patterns of the twomouse twinfilin isoforms. (A) Northern blotanalysis of twinfilin expression duringdevelopment. Both twinfilin-1 mRNA (top)and twinfilin-2 mRNA are expressed atconstant levels during development. (B)Whole mount in situ hybridization of E9.5mouse embryos. With antisense probes,twinfilin-1 (top left) and twinfilin-2 (lower left)expressions are detected throughout the
tive sense probes (right panel), indicating tha
embryo. No signal is detected with the respec tthese hybridizations were specific. (C) Northern blot analysis of twinfilin-1 and twinfilin-2expression in adult mouse. Twinfilin-2 is expressed at high levels only in the heart. Twinfilin-2 is also detected in skeletal muscle, where twinfilin-1 is not expressed (see ΙΙΙ ).33

34
Figure 6. Biochemical comparison of the two mouse twinfilin isoforms. (A) Both twinfilins prevent the assembly of actin filaments. Polymerization of 3 µM actin (1:6 pyrene actin:human platelet actin) in the presence and absence of 0, 1.5 µM or 3 µM twinfilin was initiated by the addition of 0.1 M KCl, 2 mM MgCl2 and 0.5 mM ATP. Polymerization of filaments was followed by an increase in pyrene fluorescence. (B-D) Interaction of twinfilins with actin monomers. The increase in fluorescence of 0.2 µM NBD-labeled Mg-ADP/ATP platelet/muscle actin was measured at different concentrations of twinfilin under physiological ionic conditions at pH 8.0. Symbols are data points, which are mean values of three independent experiments. Lines are calculated binding curves. were unable to obtain the KD value for twinfilin-2-muscle ATP-actin complex. In agreement with previous results (Palmgren et al., 2001) we found that both twinfilins bind 8-10 times more strongly to ADP-actin monomers (KD=0.04-0.12 µM, Figure 5B and D) than to ATP-actin monomers (KD=1.76-1.92 µM, Figure 5C). While the affinities for platelet ADP-actin monomers are very similar for both isoforms (twinfilin-1 KD=0.09 µM and twinfilin-2 KD=0.12 µM), twinfilin-2 (KD=0.04 µM) shows two times higher affinity for muscle ADP-actin monomers than twinfilin-1 (KD=0.08 µM).

35
Discussion The focus of this study was to elucidate the role of two actin monomer-binding proteins, ADF/cofilin and twinfilin, in regulating actin dynamics in multicellular organisms. We show that mice have three isoforms of ADF/cofilin, which display distinct expression patterns as well as different biochemical properties (Table 3). This has allowed the individual ADF/cofilins to adapt to regulate actin dynamics in different cell types. We also show that twinfilin, a novel actin monomer-binding protein, is an important regulator of actin dynamics both in mammalian cells (Table 3) and in Drosophila.
10. The biochemical properties of mouse ADF/cofilins are adapted to their expression patterns Unicellular organisms contain only one ADF/cofilin protein, whereas several ADF/cofilin isoforms exist in multicellular organisms. Our phylogenetic analysis of all known mammalian and avian ADF/cofilins grouped these proteins into three subgroups, which we named cofilin-1, cofilin-2 and ADF (Ι). These three classes probably exist in all mammals and birds, although only cofilin-2 and ADF are currently found in chickens (Abe et al., 1990, Adams et al., 1990). The isoforms probably arose through two duplications of the ancestral ADF/cofilin gene. The first duplication yielded ADF and “cofilin”, after which the latter further duplicated into non-muscle (cofilin-1) and muscle (cofilin-2) cofilins (Ι; Lappalainen et al., 1998). Our expression analysis indicates that the three mouse ADF/cofilins have distinct expression patterns. In agreement with previous studies, we found that although cofilin-1 is present in embryonic skeletal muscle, it is absent from mature skeletal muscle (Ι). Previous studies have shown that in terminally differentiated myogenic mouse cells, cofilin-1 is replaced by cofilin-2 (Ono et al., 1994; Obinata et al., 1997; Mohri et al., 2000). Northern blot analysis suggested that all three mouse ADF/cofilins would be expressed in many organs, but in situ hybridizations showed that these isoforms are confined to distinct tissues within these organs. We found ADF to be specific for epithelial and nervous tissues, whereas cofilin-2 was mainly found in muscle. Cofilin-1 was expressed rather ubiquitously (Ι). Therefore, Northern blot analysis gave only partial information about the expression patterns of the ADF/cofilin isoforms, and in situ hybridizations were necessary to determine the actual tissue distribution of these proteins. Our expression analysis also shows that more than one ADF/cofilin isoform can be expressed in the same cell. This raises questions about the differences in localization, regulation and biochemical properties between the isoforms present within one cell. Our studies indicate that mouse ADF/cofilins do not differ substantially in their basic cell biology. We found that the localizations of myc-tagged ADF/cofilins in cultured cells were similar to each other (Ι) and to the localizations of endogenous ADF and cofilin-1 (Vartiainen et al., unpublished). These localizations agree with those of previous studies

36
(Bamburg and Bray, 1987; Yonezawa et al., 1987). However, it is possible that the ADF/cofilin isoforms may behave differently in specific situations. For example during cytokinesis, microinjection of cofilin, but not of ADF, causes disassembly of actin filaments and their accumulation in the cleavage furrow in dividing myoblasts (Obinata et al., 1997). The Rho family of small GTPases are able to inactivate ADF/cofilins by inducing their phosphorylation through LIM and TES kinases (Arber et al., 1998; Yang et al., 1998; Toshima et al., 2001a-c). Our data indicate that the Rho GTPases RhoA, Cdc42 and Rac1 have similar effects on all mouse ADF/cofilin isoforms (Vartiainen et al., unpublished). This is supported by the fact that no reports have been published suggesting that the LIM- or TES-kinase isoforms prefer a certain ADF/cofilin isoform. Beside phosphorylation, the activity of most of the ADF/cofilin family members is regulated by pH (Yonezawa et al., 1985; Hawkins et al., 1993; Hayden et al., 1993; Maciver et al., 1998). We found that ADF was the only mouse isoform whose actin disassembling activity was affected by an increase in pH (Ι). This is also supported by the notion that only ADF’s distribution in Swiss 3T3 cells is sensitive to changes in the pH (Bernstein et al., 2000). The distinct tissue distribution of the mouse ADF/cofilin isoforms could also reflect specificity towards a certain actin isoform. In our actin filament binding and disassembly assays we did not detect any preference towards platelet or muscle actin with our mouse ADF/cofilin isoforms. However, we found that all ADF/cofilin isoforms (Ι) and also both twinfilins (unpublished) were able to disassemble platelet actin more efficiently than muscle actin. This indicates that platelet and muscle actins have quantitative differences in their dynamics, and agrees with the finding that muscle actin filaments are less dynamic than actin in other cells (Sanger et al., 1984). In agreement with recent studies on human ADF and cofilin-1 (Yeoh et al., 2002) we found that all mouse ADF/cofilins bind equally well to actin filaments. However, in other respects the mouse ADF/cofilin isoforms are biochemically different from each other. ADF and cofilin-1 disassemble actin filaments very efficiently, whereas cofilin-2 promotes filament formation (Ι). Similarly, the muscle-specific ADF/cofilin from C. elegans, Unc-60B, is less efficient in disassembling actin filaments than the ubiquitously expressed Unc60A (Ono and Benian, 1998). At higher concentrations, ADF was more efficient in increasing the amount of actin monomers than cofilin-1 (Ι). This could be due to the fact the ADP-actin monomer nucleates two times more readily when complexed to cofilin-1 than to ADF, which results in higher steady state depolymerization activity of ADF (Yeoh et al., 2002). We measured actin filament disassembling activity by a cosedimentation assay, which is a steady-state assay. This implies that the results obtained from these assays are combinations of many different parameters, such as actin filament binding, depolymerization, and severing, actin monomer sequestering, as well as the ability to promote filament assembly. Therefore, due to its inability to increase the amount of actin monomers in these assays, cofilin-2 may have differences in any of these aspects. Our visual actin depolymerization/severing assay showed that all mouse ADF/cofilins are able to depolymerize actin. This assay also indicated that all mouse isoforms have

37
some actin filament-severing activity, since they shortened actin filaments more than what could be regarded as mere actin depolymerization (Ι). In agreement with these results, recent studies showed that human ADF and cofilin-1 are also able to sever actin filaments to a similar extent (Yeoh et al., 2002). The three mouse ADF/cofilin isoforms, however, increased actin filament treadmilling to different extents. Cofilin-2 was the least efficient isoform (Ι), which can at least partly explain why cofilin-2 could not shift actin to the monomeric form in the actin filament sedimentation assay. Our results agree with findings that the ADF/cofilins have higher affinity for ADP-actin monomers than for ATP-actin monomers (Ι, Carlier et al., 1997; Blanchoin and Pollard, 1998, Ressad et al., 1998). However, the affinities we obtained are ~ 5-10 times greater than previously reported values. This discrepancy may be due to the protein constructs used in different studies. The thrombin cleavage of our recombinant GST-ADF/cofilin added an N-terminal Ser-Gly to the native ADF/cofilins. In other studies, ADF/cofilins were expressed from vectors in which translation starts from the methionine of ADF/cofilins. In E.coli, this N-terminal methionine is usually removed (Hawkins et al., 1993), which may affect the actin-binding properties of these recombinant proteins, since the first four amino acids have been shown to be important in interactions between actin and ADF/cofilins (Lappalainen et al., 1997; Wriggers et al., 1998). A significant difference that we found between the isoforms was that cofilin-2 showed smaller difference between its affinities for ADP- and ATP-actin monomers, due to its 5-10 times greater affinity for ATP-actin monomers (Ι). Computer simulations and experimental data suggest that the association rate constant of ADF/cofilin-ATP-actin is higher than that of ATP-actin monomer alone (Carlier et al., 1997; Sept et al., 1999). This suggests, that because it has a higher affinity for ATP-actin monomers, cofilin-2 might be able promote filament assembly more efficiently than ADF and cofilin-1. Specialized cells are a hallmark of multicellular organisms. These cells express distinct genes, have unique morphologies and different requirements for their actin dynamics. Our results indicate that the three mouse ADF/cofilins have adapted to promote actin filament turnover in these specific cell types. ADF is the most “active” ADF/cofilin. It is expressed in epithelial and nervous tissues (Ι), where cells often are highly polarized. Polarization is strongly dependent on the dynamic nature of the actin cytoskeleton, and would therefore benefit from an efficient promoter of actin turnover. Furthermore, epithelial cells often contain specialized cell surface modifications and cell junctions, both of which are intimately linked to the actin cytoskeleton. Maintenance and modification of these structures therefore rely on the dynamic nature of the actin cytoskeleton. Epithelial and endothelial cells are often found in those areas of the body that are subjected to physical damage and wounding. Cells need to be able to migrate in order to repair such sites, and cell movement requires rapid changes in the actin cytoskeleton. Therefore, the “active” ADF is ideally located in those cell types that undergo rapid actin dynamics. Cofilin-2 is the only ADF/cofilin isoform expressed in mature skeletal muscle (Ι, Obinata et al., 1997). Recent results indicate that actin dynamics at thin filament ends in cardiac myocytes is comparable to

38
actin dynamics in relatively stable actin structures in other cells (Littlefield et al., 2001). The turnover of the entire actin filament in muscle fibers is, however, likely to be very slow, and treadmilling probably does not take place (Littlefield et al., 2001). Therefore, muscle cells do not require an ADF/cofilin that would promote actin dynamics very efficiently. The biochemical properties of cofilin-2 are perfectly tuned for muscle cells, because, unlike cofilin-1 and ADF, cofilin-2 promotes filament assembly rather than disassembly. This suggests that in contractile cells, the specific isoforms of different proteins are adapted to keep the sarcomeric structure intact. Moreover, also the muscle actin itself is more resistant to depolymerization (Ι). The ubiquitous cofilin-1 is the “normal” ADF/cofilin, which is a biochemical intermediate between ADF and cofilin-2. Cofilin-1 may promote actin filament dynamics in less specialized cell types, and it might also assist ADF in more dynamic cell types. In the future, knock-out mice and RNAi knock-down cell lines of the ADF/cofilin isoforms will be very valuable in efforts to evaluate the importance of the distinct ADF/cofilin isoforms for different cell types.
11. Twinfilin is essential for the development of Drosophila The lack of a clear phenotype in the twinfilin deletion yeast strains (Goode et al., 1998) has hampered the understanding of the biological role of this ubiquitous actin monomer-binding protein. We identified the Drosophila homologue of twinfilin and showed that a mutation in the Drosophila twinfilin gene leads to several morphological defects. This provides the first genetic evidence for a biological role of twinfilin in the development of multicellular organisms (ΙΙ). Drosophila bristles are sensory organs formed as a cytoplasmic extension of a single cell supported by actin filament bundles. Each bundle is composed of successive repeated units attached end to end. As the bristle elongates, actin filaments are formed at the tip of the bristle, and then progressively packed to form new units that have maximally crosslinked filaments (Tilney et al., 2000a). For this reason, Drosophila bristles have been extensively used as a model to study actin filament bundling. In twinfilin mutant flies, the adult macrochaeta possess a similar phenotype (ΙΙ) as that seen in singed and forked mutants, which arise from mutations in actin filament bundling proteins (Cant et al., 1994; Petersen et al., 1994; Tilney et al., 1995; 1998). The fact that twinfilin is an actin monomer-binding protein and does not bind to actin filaments suggests that the accurate regulation of the size and dynamics of the actin monomer pool is essential for actin assembly in the developing bristles. This is supported by observations that mutations in Drosophila cofilin (Chen et al., 2001), profilin (Verheyen and Cooley, 1994) and the β-subunit of capping protein (Hopmann et al., 1996) genes also lead to defects in bristle morphology. Furthermore, loss of function of slingshot, a phosphatase that is able to activate ADF/cofilin, results in aberrant bristle morphology (Niwa et al., 2002). A unique feature of twinfilin mutant bristles is the presence of ectopic actin filament-containing spots and perpendicular tiny bundles in fully elongated

39
mutant bristles (ΙΙ). This could result from uncontrolled polymerization in the absence of twinfilin from pre-existing bundles, which would then split lengthwise and fragment further. A more plausible explanation is, however, that loss of twinfilin’s function would result in spontaneous nucleation of actin filaments, which would not be incorporated into the main bundles. This is supported by observations that the ectopic spots resemble the actin filament- containing spots in young bristles, but are clearly different from the fragmented actin bundles observed after cytochalasin D treatment (Tilney et al., 2000b). Our immunostainings show that Drosophila twinfilin localizes both to the cytoplasm and to the actin filament bundles in the bristles (ΙΙ). Yeast twinfilin (Palmgren et al., 2001) and murine twinfilin (ΙΙΙ ) also show similar localizations. In yeast, capping protein is needed for the correct localization of twinfilin (Palmgren et al., 2001). In fully elongated bristles, twinfilin localizes along actin filaments in spots that may be barbed ends of actin filaments. Therefore, it is possible that also in Drosophila, the localization of twinfilin to actin bundles is mediated through capping protein. Intriguingly, the bristle phenotypes of twinfilin (ΙΙ) and capping protein mutants (Hopmann et al., 1996) are almost identical. We have also shown that Drosophila twinfilin and the ADF/cofilin-encoding gene twinstar interact genetically in bristle morphogenesis (ΙΙ). A similar genetic interaction between twinfilin and ADF/cofilin was previously shown in yeast, where a mutation in the cofilin gene that affects its actin depolymerization activity rate shows synthetic lethality with a twinfilin deletion (Lappalainen and Drubin, 1997; Goode et al., 1998). This suggests that during actin bundle formation, the decreased actin filament depolymerization due to the ADF/cofilin mutation, together with uncontrolled actin assembly resulting from the twinfilin mutation, lead to defects in bristle morphogenesis.
12. Mouse twinfilin isoforms have similar biochemical properties but distinct expression patterns Mammalian twinfilin-1 and -2 were isolated with an anti-phosphotyrosine antibody (Beeler et al., 1994) and as a binding partner for protein kinase Cζ (Rohwer et al., 1999), respectively. No actin-related activities were reported for these proteins. We were, however, able to show that both mouse twinfilins are actin monomer-binding proteins with biochemical properties that are similar to each other (Table 3) and to yeast and Drosophila twinfilins. Cell biological characterization revealed that twinfilin is an important regulator of actin dynamics in murine cells as well. Previous biochemical study of the human homologue of twinfilin-1, A6, suggested that this protein is a protein kinase (Beeler et al.,1994). We re-examined the putative tyrosine kinase activity of mouse twinfilin-1, which is 96% identical to the human protein, because twinfilin does not have detectable sequence homology with known protein kinases and it does not
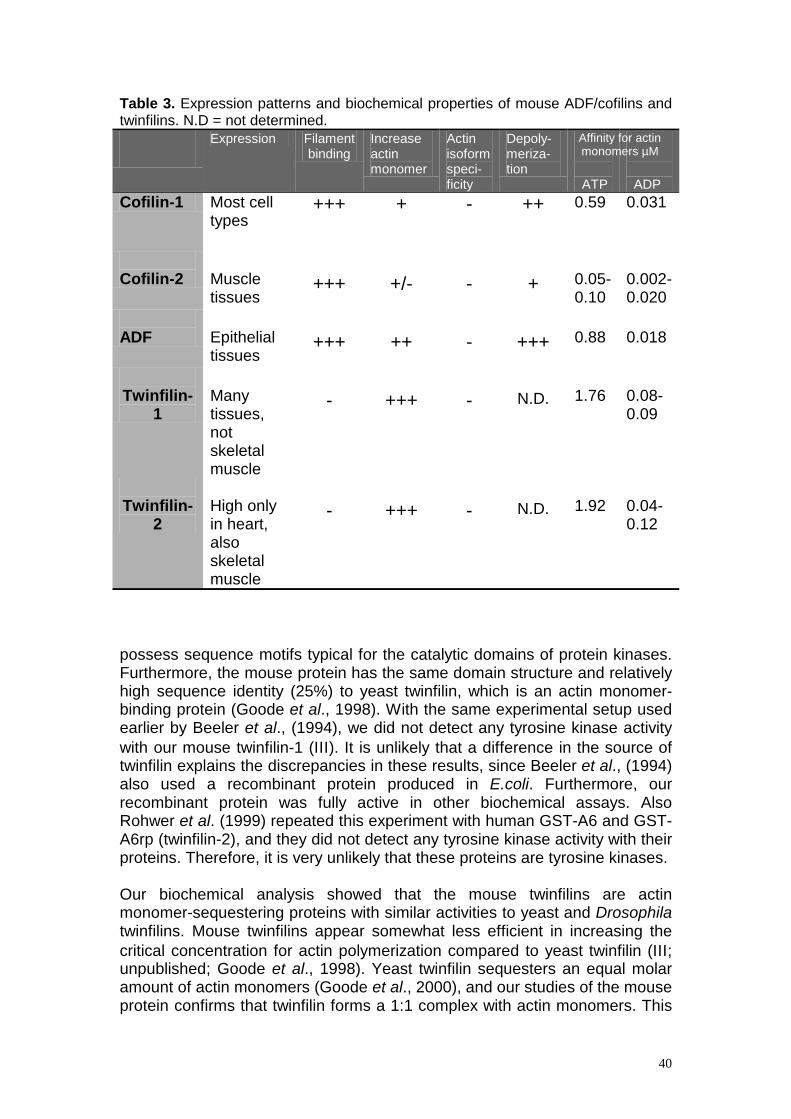
40
Table 3. Expression patterns and biochemical properties of mouse ADF/cofilins and twinfilins. N.D = not determined.
Expression Filament binding
Increase actin monomer
Actin isoform speci- ficity
Depoly-meriza- tion
ATP
ADP
Cofilin-1 Most cell types
+++ + - ++ 0.59 0.031
Cofilin-2
Muscle tissues
+++
+/-
-
+
0.05-0.10
0.002-0.020
ADF
Epithelial tissues
+++
++
-
+++
0.88
0.018
Twinfilin-
1
Many tissues, not skeletal muscle
-
+++
-
N.D.
1.76
0.08-0.09
Twinfilin-
2
High only in heart, also skeletal muscle
-
+++
-
N.D.
1.92
0.04-0.12
possess sequence motifs typical for the catalytic domains of protein kinases. Furthermore, the mouse protein has the same domain structure and relatively high sequence identity (25%) to yeast twinfilin, which is an actin monomer-binding protein (Goode et al., 1998). With the same experimental setup used earlier by Beeler et al., (1994), we did not detect any tyrosine kinase activity with our mouse twinfilin-1 (ΙΙΙ ). It is unlikely that a difference in the source of twinfilin explains the discrepancies in these results, since Beeler et al., (1994) also used a recombinant protein produced in E.coli. Furthermore, our recombinant protein was fully active in other biochemical assays. Also Rohwer et al. (1999) repeated this experiment with human GST-A6 and GST-A6rp (twinfilin-2), and they did not detect any tyrosine kinase activity with their proteins. Therefore, it is very unlikely that these proteins are tyrosine kinases. Our biochemical analysis showed that the mouse twinfilins are actin monomer-sequestering proteins with similar activities to yeast and Drosophila twinfilins. Mouse twinfilins appear somewhat less efficient in increasing the critical concentration for actin polymerization compared to yeast twinfilin (ΙΙΙ ; unpublished; Goode et al., 1998). Yeast twinfilin sequesters an equal molar amount of actin monomers (Goode et al., 2000), and our studies of the mouse protein confirms that twinfilin forms a 1:1 complex with actin monomers. This
Affinity for actin monomers µM

41
is intriguing, because twinfilin contains two ADF-H domains, which are both theoretically capable of interacting with one actin monomer. In addition, the residues important for actin monomer binding in ADF/cofilins are conserved in both domains of twinfilin, indicating that each of them is able to bind actin (Lappalainen et al., 1998). In agreement with previous data on yeast twinfilin (Palmgren et al., 2001), we found that both mouse twinfilins have a higher affinity for ADP- than for ATP-actin monomers. We did not find large differences in the biochemical properties of the mouse twinfilin isoforms (unpublished). Also different mammalian profilin isoforms appear to have similar actin-related activities, but they show differences in binding to other ligands, e.g. to polyphosphoinositides and proline-rich motifs (Lambrechts et al., 1997). It is therefore possible that the twinfilin isoforms may have different affinities for their other ligands. It remains to be shown for example, whether the twinfilin isoforms prefer a certain capping protein isoform. Beeler et al., (1994) found that mouse twinfilin-1 is expressed at high levels in the liver, kidney and lung and at moderate levels in the heart, brain, skeletal muscle and testis. They did not detect twinfilin-1 in the spleen, which agrees with our results. Our results also agree with the high expression levels of twinfilin-1 in the liver and kidney, but we found only a small amount of twinfilin-1 in the lung and we did not detect any twinfilin-1 in skeletal muscle (ΙΙΙ ). These small inconsistencies could be due to different probes and to the quality of the blots used in these studies. Our preliminary in situ hybridizations on adult mouse confirm that twinfilin-1 is expressed at high levels in the liver and is absent from skeletal muscle. Using the same method we found high twinfilin-2 expression in the heart (Vartiainen et al., unpublished). We are currently generating knock-out mice of both twinfilin isoforms. The knock-out construct will drive the expression of LacZ in the place of twinfilin, which allows us obtain detailed expression patterns of these proteins.
13. The cell biological role of twinfilin In mammalian cells twinfilin localizes both to the cytoplasm and to the cortical actin assemblies (ΙΙΙ ). Both yeast (Palmgren et al., 2001) and Drosophila twinfilins (ΙΙ) show similar localizations. It is notable that twinfilin does not colocalize with all actin filament structures: twinfilin is not found for example in stress fibers, but the staining overlaps significantly with the actin monomer staining. In cells, these high actin monomer concentrations are found in areas where actin filament turnover is rapid (Carlier and Pantaloni, 1997). Therefore, twinfilin’s intracellular localization supports its role as a regulator of actin dynamics. In yeast, interactions with actin monomer and capping protein are essential for the correct localization of twinfilin (Palmgren et al., 2001). Also mouse twinfilin-1 interacts directly with capping protein (Palmgren et al., 2001), and this interaction might drive the localization of mouse twinfilin to the cell cortex. Further evidence of twinfilin’s involvement in the regulation of actin dynamics comes from the overexpression data, which show that a high concentration of twinfilin causes a decrease in the amount of stress fibers and appearance of abnormal actin filament structures (III). Also in yeast,

42
overexpression of twinfilin results in the accumulation of abnormal actin structures (Goode et al., 1998). We also found that twinfilin-1 colocalizes with the activated forms of Rac1 and Cdc42 at the membrane ruffles and cell-cell contacts, respectively (ΙΙΙ ). These GTPases are known to be regulators of the actin cytoskeleton and could thus regulate the localization of twinfilin-1 to certain cellular compartments. This view is also supported by the observation that activated Rac1 causes accumulation of twinfilin to the membrane ruffles, whereas expression of the dominant negative form of Rac1 localizes twinfilin-1 to the cytoplasm (ΙΙΙ ). Interestingly, the small GTPases, including Rac1 and Cdc42, are able to induce phosphorylation and consequent inactivation of ADF/cofilins (Arber et al., 1998; Yang et al., 1998). We have recently learned that twinfilin-1 is phosphorylated on both serine and threonine residues (Vartiainen et al., unpublished), and it remains to be elucidated whether the small GTPases control twinfilin’s activity and localization through phosphorylation.

43
Conclusions and future perspectives The results of this work demonstrate that Drosophila and mouse twinfilins are biochemical homologs of the previously characterized yeast twinfilin (ΙΙ, ΙΙΙ , Goode et al., 1998). Furthermore, the overexpression and mutant phenotypes as well as the localization of twinfilin in Drosophila (ΙΙ) and mouse cells (ΙΙΙ ), suggest that animal twinfilins play cell biological roles that are similar to those of the yeast twinfilin. In yeast, a “mailman” model suggests that twinfilin’s function is to localize the inactive ADP-actin monomers to the sites of rapid actin assembly through interactions with the capping protein. Therefore twinfilin would act as a link between the ADF/cofilin-induced actin filament depolymerization and profilin-mediated actin assembly (Palmgren et al., 2001; 2002). The uncontrolled actin polymerization in the twinfilin mutant Drosophila bristles supports this model (ΙΙ). Moreover, capping protein interacts directly with mouse twinfilin-1 (Palmgren et al., 2001) and possibly mediates the localization of Drosophila twinfilin as well (ΙΙ). However, in higher eukaryotes, twinfilin’s biological function is likely to be more complex than in yeast. The phenotype of twinfilin mutants in Drosophila indicates that twinfilin is required especially in processes where actin polymerization occurs in a localized fashion, such as during the development of Drosophila bristles. This supports twinfilin’s role as a localizer of actin monomers, but could also pinpoint twinfilin’s role in preventing actin filament polymerization at unwanted sites. These two processes are very interdependent. In mammalian cells twinfilin could therefore also be important for the development and maintenance of cell surface protrusions, such as brush border microvilla, hair cell strereocilia and even cell motility organs like filopodia. Indeed, in N18 cells twinfilin localizes to the filopodia (ΙΙΙ ). Despite the fact that twinfilin is clearly involved in regulation of the actin filament turnover in mammalian cells (ΙΙΙ ), its exact in vivo function is not yet known. Knock-out mice of both twinfilin isoforms are now being developed, and they will give valuable information on the biological function of these proteins as well as on the relative importance of the two isoforms. The phenotypes of these knock-out mice are impossible to predict, but the phenotypes of Drosophila twinfilin mutants (ΙΙ) indicate that at least the double knock-out should have severe developmental defects. The “mailman” model predicts that twinfilin and ADF/cofilin function sequentially to regulate the actin monomer pool (Figure 7). Indeed, these proteins interact genetically during bristle development in Drosophila (ΙΙ). In the future, it will be important to elucidate how the activities of these two proteins are co-ordinated in cells. An excellent candidate to regulate this process is the small GTPase Rac1, which can inactivate the ADF/cofilins (Yang et al., 1998) and affect the localization of twinfilin in mammalian cells (ΙΙΙ ).
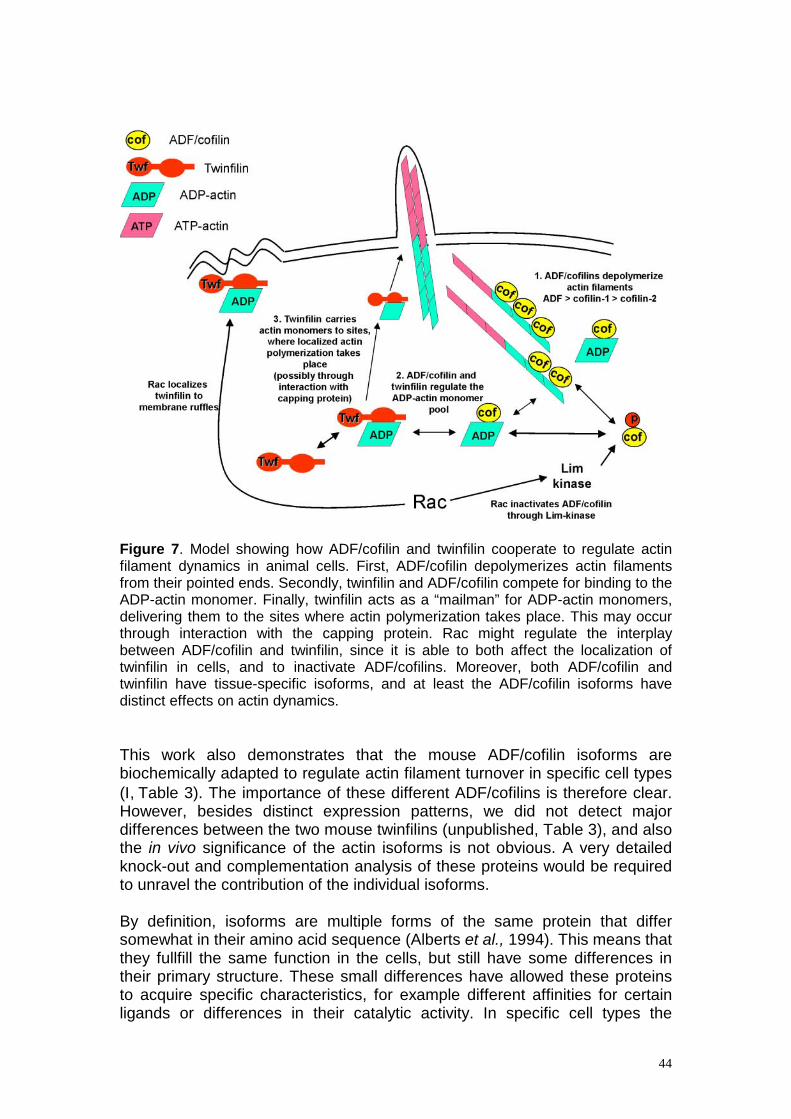
44
Figure 7. Model showing how ADF/cofilin and twinfilin cooperate to regulate actin filament dynamics in animal cells. First, ADF/cofilin depolymerizes actin filaments from their pointed ends. Secondly, twinfilin and ADF/cofilin compete for binding to the ADP-actin monomer. Finally, twinfilin acts as a “mailman” for ADP-actin monomers, delivering them to the sites where actin polymerization takes place. This may occur through interaction with the capping protein. Rac might regulate the interplay between ADF/cofilin and twinfilin, since it is able to both affect the localization of twinfilin in cells, and to inactivate ADF/cofilins. Moreover, both ADF/cofilin and twinfilin have tissue-specific isoforms, and at least the ADF/cofilin isoforms have distinct effects on actin dynamics. This work also demonstrates that the mouse ADF/cofilin isoforms are biochemically adapted to regulate actin filament turnover in specific cell types (Ι, Table 3). The importance of these different ADF/cofilins is therefore clear. However, besides distinct expression patterns, we did not detect major differences between the two mouse twinfilins (unpublished, Table 3), and also the in vivo significance of the actin isoforms is not obvious. A very detailed knock-out and complementation analysis of these proteins would be required to unravel the contribution of the individual isoforms. By definition, isoforms are multiple forms of the same protein that differ somewhat in their amino acid sequence (Alberts et al., 1994). This means that they fullfill the same function in the cells, but still have some differences in their primary structure. These small differences have allowed these proteins to acquire specific characteristics, for example different affinities for certain ligands or differences in their catalytic activity. In specific cell types the

45
appearance of the actin cytoskeleton is different due to unique compositions of actin filament assemblies. For example, most of the actin in muscle cells is organized to sarcomers, whereas motile cells have a very prominent cell cortex with highly dynamic actin filaments in their lamellipodia. Nevertheless, some cells have enormous cell surface structures supported by actin bundles. These specialized assemblies set specific requirements for the dynamics of the actin cytoskeleton, but at the same time, the cells must be also able to perform “normal” actin-dependent cellular functions, such as secretion and endocytosis. The regulation of the actin cytoskeleton is therefore an amenable field for isoforms, because although cytoskeletal regulation basically occurs in a similar manner in every cell, specific requirements are nevertheless posed by different cell types. It can be concluded that the isoforms of actin and actin-binding proteins have evolved to fine-tune the regulation of actin dynamics to best suite each individual cell type.

46
Acknowledgements This study was carried out at the Institute of Biotechnology with the financial support of Academy of Finland and Sigrid Juselius Foundation. I wish to express my greatest gratitude to my supervisor, docent Pekka Lappalainen, for the opportunity to work in his group and for introducing me to the fascinating world of the actin cytoskeleton. His enthusiasm and positive attitude have made it a great pleasure to work in the actin group. I wish to thank Professor Mart Saarma, the head of the Institute of Biotechnology, for providing excellent research and core facilities. Professor Heikki Rauvala, the head of the Helsinki Graduate School in Biotechnology and Molecular Biology, as well as Dr. Erkki Raulo and Anita Tienhaara are thanked for organizing excellent courses. I wish to thank Professor Carl G. Gahmberg, the head of the Division of Biochemistry, for the positive attitude when helping me with my studies. I am extremely grateful to Docent Olli Carpen and Professor John Eriksson for the critical reading of this manuscript. I wish to thank my collaborators, Professor Irma Thesleff and Dr. Juha Partanen for sharing their expertize in mouse development. Tuija Mustonen is thanked for teaching me how to “build” a mouse. Dr. Tapio Heino and Gudrun Wahlström are thanked for excellent work regarding the Drosophila twinfilin. I am also grateful to Dr. Petri Auvinen and Dr. Johan Peränen for their numerous practical tips and reagents. All former and present members of the dynamic actin group, Eija, Enni, Elisa, Hanna, Pauli, Pieta, Pirta, Sandra and Ville, are deeply thanked for creating such a fun and warm working atmosphere. It has been a pleasure to spend time with you both at work and off work. I wish to thank Pauli for teaching me biochemistry and providing me with his exceptional ideas. Ville is acknowledged for his assistance with computers. I am especially thankful to Sandra for suggestions regarding this manuscript and for many fruitful discussions during these years. This applies to Johanna as well. I wish to thank all my friends for the relaxing moments and wild parties that we have had together. I am especially grateful to Jenni and Hanna for their long-lasting friendships. I owe special thanks to my parents, Mona and Ahti, who have patiently loved and supported me in every (im)possible situation. Also my dog, Kossu, is acknowledged for taking me out every day. Finally, I want to thank Petri from the bottom of my heart for always being there for me. Helsinki, September 2002

47
References Abe, H., Endo, T., Yamamoto, K., and Obinata, T. (1990). Sequence of cDNAs encoding actin depolymerizing factor and cofilin of embryonic chicken skeletal muscle: two functionally distinct actin-regulatory proteins exhibit high structural homology. Biochemistry 29:7420-7425. Abe, H., Nagaoka, R., and Obinata, T. (1993). Cytoplasmic localization and nuclear transport of cofilin in cultured myotubes. Exp. Cell Res. 206:1-10. Abe, H., Obinata, T., Minamide, L.S., and Bamburg, J.R. (1996). Xenopus laevis actin-depolymerizing factor/cofilin: a phosphorylation-regulated protein essential for development. J. Cell Biol. 132:871-885. Adams, M.E., Minamide, L.S., Duester, G., and Bamburg, J.R. (1990). Nucleotide sequence and expression of a cDNA encoding chick brain actin depolymerizing factor. Biochemistry 29:7414-7420. Agnew, B.J., Minamide, L.S., and Bamburg, J.R. (1995). Reactivation of phosphorylated actin depolymerization factor and identification of the regulatory site. J. Biol. Chem. 270:17582-17587. Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K., and Watson, J.D. (1994). Molecular biology of the cell. Third edition. Garland Publishing Inc., New York. Allwood, E.G., Smertenko, A.P., and Hussey, P.J. (2001). Phosphorylation of plant actin-depolymerizing factor by calmodulin-like domain protein kinase. FEBS Lett. 24929:1-4. Arber, S., Barbayannis, F.A., Hanser, H., Schneider, C., Stanyon, C.A., Bernard, O., and Caroni, P. (1998). Regulation of actin dynamics through phosphorylation of cofilin by LIM-kinase. Nature 393:805-809. Bamburg, J.R., and Bray, D. (1987). Distribution and cellular localizations of actin depolymerizing factor. J. Cell Biol. 105:2817-2825. Bamburg, J.R. (1999). Proteins of the ADF/cofilin family: essential regulators of actin dynamics. Annu. Rev. Cell Dev. Biol. 15:185-230. Barkalow, K., Witke, W., Kwiatkowski, D.J., and Hartwig, J.H. (1996). Coordinated regulation of platelet actin filament barbed ends by gelsolin and capping protein. J. Cell Biol. 134:389-399. Bartles, J.R. (2000). Parallel actin bundles and their multiple actin bundling proteins. Curr. Opin. Cell Biol. 12:72-78. Barritt, G.J. (1996). Communication within animal cells. Oxford University Press, Oxford.

48
Beeler, J.F., LaRochelle, W.J., Chedid, M., Tronick, S.R., and Aaronson, S.A. (1994). Prokaryotic expression cloning of a novel human tyrosine kinase. Mol. Cell. Biol. 14:982-988. Beeler, J.F., Patel, B.K.R., Chedid, M., and LaRochelle, W.J. (1997). Cloning and characterization of the mouse homolog of the human A6 gene. Gene 193:31-37. Bernstein, B.W., Painter, W.B., Chen, H., Minamide, L.S., Abe, H., and Bamburg, J.R. (2000). Intracellular pH modulation of ADF/cofilin proteins. Cell Motil. 47:319-336. Bishop, A.L., and Hall, A. (2000). Rho GTPases and their effector proteins. Biochem. J. 348:241-255. Blanchoin, L., and Pollard, T.D. (1998). Interaction of actin monomers with Acanthamoeba actophorin (ADF/cofilin) and profilin. J. Biol. Chem. 273:25106-25111. Blanchoin, L., and Pollard, T.D. (1999). Mechanism of interaction of Acanthamoeba actophorin (ADF/cofilin) with actin filaments. J. Biol. Chem. 274:15538-15546. Boquet, I., Boujemaa, R., Carlier, M.-F., and Preat, T. (2000). Ciboulot regulates actin assembly during drosophila brain metamorphosis. Cell 102:797-808. Bowman, G.D., Nodelman, I.M., Hong, Y., Chua, N.-H., Lindberg, U., and Schutt, C.E. (2000). A comparative structural analysis of the ADF/cofilin family. Proteins 41:374-384. Braun, A., Aszodi, A., Hellebrand, H., Berna, A., Fässler, R., and Brandau, O. (2002). Genomic organization of profilin-III and evidence for a transcript expressed exclusively in testis. Gene 283:219-225. Bray, D. (2001). Cell movements: from molecules to motility. Second edition. Garland Publishing, New York. Bresnick, A.R. (1999). Molecular mechanisms of non-muscle myosin II regulation. Curr. Opin Cell Biol. 11:26-33. Brill, S., Li, S., Lyman, C.W., Church, D.M., Wasmuth, J.J., Weissbach, L., Bernards, A., and Snijders, A.J. (1996). The ras GTPase-activating-protein-related human protein IQGAP2 harbors potential actin binding domain and interacts with calmodulin and rho family GTPases. Mol. Cell. Biol. 16: 4869-1878. Cameron, L.A., Giardini, P.A., Soo, F.S., and Theriot, J. (2000). Secrets of actin-based motility revealed by a bacterial pathogen. Nat. Rev. Mol. Cell Biol. 1:110-119.

49
Cant, K., Knowles, B.A., Mooseker, M.S., and Cooley, L. (1994). Drosophila singed, a fascin homologue, is required for actin bundle formation during oogenesis and bristle extension. J. Cell Biol. 125:369-380. Carlier, M.-F., Didry, D., Erk, I., Lepault, J., Van Troys, M.L., Vandekerckhove, J., Perelroizen, I., Yin, H., Doi, Y., and Pantaloni, D. (1996). Tβ4 is not a simple G-actin sequestering protein and interacts with F-actin at high concentrations. J. Biol. Chem. 271:9231-9239. Carlier, M.F., Laurent, V., Santolini, J., Melki, R., Didry, D., Xia, G.X., Hong, Y., Chua, N.H., and Pantaloni, D. (1997). Actin depolymerizing factor (ADF/cofilin) enhances the rate of filament turnover: implication in actin-based motility. J. Cell Biol. 136:1307-1322. Carlier, M.F., and Pantaloni, D. (1997). Control of actin dynamics in cell motility. J. Mol. Biol. 269:459-467. Caroni, P. (2001). Actin cytoskeleton regulation through modulation of PI(4,5)P2 rafts. EMBO J. 20:4332-4336. Chen, J., Godt, D., Gunsalus, K., Kiss, I., Goldberg, M., and Laski, F.A. (2001). Cofilin/ADF is required for cell motility during Drosophila ovary development and oogenesis. Nat. Cell Biol. 3:204-209. Cooper, J.A., Wear, M.A., and Weaver, A.M. (2001). Arp2/3 complex: advances on the inner workings of a molecular machine. Cell 107:703-705. Dan, C., Kelly, A., Bernard, O., and Minden, A. (2001). Cytoskeletal changes regulated by the PAK4 serine/threonine kinase are mediated by LIM kinase 1 and cofilin. J. Biol. Chem. 276:32115-32121. Didry, D., Carlier, M.F., and Pantaloni, D. (1998). Synergy between actin depolymerizing factor/cofilin and profilin in increasing actin filament turnover. J. Biol. Chem. 273:25602-25611. Di Nardo, A., Gareus, R., Kwiatkowski, D., and Witke, W. (2000). Alternative splicing of the mouse profilin II gene generates functionally different profilin isoforms. J. Cell Sci. 113: 3795-3803. Drenckhahn, D., and Pollard, T.D. (1986). Elongation of actin filaments is a diffusion-limited reaction at the barbed end and is accelerated by inert macromolecules. J. Biol. Chem. 261:12754-12758. Duncan, M.C., Cope, M.J., Goode, B.L., and Wehland, B., and Drubin, D.G. (2001). Yeast Eps15-like endocytic protein, Pan1p, activates the Arp2/3 complex. Nat. Cell Biol. 3:687-690. Edwards, D.C., Sanders, L.C., Bokoch, G.M., and Gill, G.N. (1999). Activation of LIM-kinase by Pak1 couples Rac/Cdc42 GTPase signaling to actin dynamics. Nat. Cell Biol. 1:253-259.

50
Fedorov, A.A., Lappalainen, P., Fedorov, E.V., Drubin, D.G., and Almo, S.C. (1997). Structure determination of yeast cofilin. Nat. Struct. Biol. 4:366-369. Goode, B.L., Drubin, D.G., and Lappalainen, P. (1998). Regulation of the cortical actin cytoskeleton in budding yeast by twinfilin, a ubiquitous actin monomer-sequestering protein. J. Cell. Biol. 142:723-733. Goode, B.L., Rodal, A.A., Barnes, G., and Drubin, D.G. (2001). Activation of the Arp2/3 complex by the actin filament binding protein Abp1p. J. Cell Biol. 153:627-634. Gunsalus, K.C., Bonaccorsi, S., Williams, E., Verni, F., Gatti, M., and Goldberg, M.L. (1995). Mutations in twinstar, a Drosophila gene encoding a cofilin/ADF homologue, results in defects in centrosome migration and cytokinesis. J. Cell Biol. 131:1243-1259. Hannappel, E., and Wartenberg, F. (1993). Actin-sequestering ability of thymosin β4, thymosin β4 fragments, and thymosin β4-like peptides as assessed by the Dnase I inhibition assay. Biol. Chem. Hoppe-Seyler 374:117-122. Hall, A. (2000). Frontiers in molecular biology: GTPases. Oxford University Press Inc., New York. Hart, M.J., Callow, M.G., Souza, B., and Polakis, P. (1996). IQGAP1, a calmodulin-binding protein with a rasGAP-related domain, is potential effector of cdc42Hs. EMBO J. 15:2997-3005. Hatanaka, H., Ogura, K., Moriyama, K., Ichikawa, S., Yahara, I., and Inagi, F. (1996). Tertiary structure of destrin and structural similarity between two actin-regulating protein families. Cell 85:1047-1055. Hawkins, M., Pope, B., Maciver, S.K., and Weeds, A.G. (1993). Human actin depolymerizing factor mediates a pH sensitive destruction of actin filaments. Biochemistry 32:9985-9993. Hayden, S.M., Miller, P.S., Brauweiler, A., and Bamburg, J.R. (1993). Analysis of the interactions of actin depolymerization factor with G- and F-actin. Biochemistry 32:9994-10004. Heintz, D., Reichert, A., Mihelic, M., Voelter, W., and Faulstich, H. (1993). Use of bimanyl actin derivative (TMB-actin) for studying complexation of β-thymosins. Inhibition of actin polymerization of thymosin β9. FEBS Lett. 329:9-12. Heiss, S.G., and Cooper, J.A. (1991). Regulation of CapZ, an actin capping protein of chicken muscle, by anionic phospholipids. Biochemistry 30:8753-8758.

51
Hertzog, M., Yarmola, E.G., Didry, D., Bubb, M.R., and Carlier, M.-F. (2002). Control of actin dynamics by proteins made of β-thymosin repeats: the actobindin family. J. Biol. Chem. 277:14786-14792. Higgs, H.N., and Pollard, T.D. (2000). Activation by Cdc42 and PIP2 of Wiskott-Aldrich syndrome protein (WASp) stimulates actin nucleation by Arp2/3. J. Cell Biol. 150:1311-1320. Hoppman, R., Cooper, J.A., and Miller, K.G. (1996). Actin organization, bristle morphology, and viability are affected by actin capping protein mutations in Drosophila. J. Cell Biol. 133:1293-1305. Huff, T., Muller, C.S.G., Otto, A.M., Netzker, R., and Hannappel, E. (2001). Β-thymosins, small acidic peptides with multiple functions. Int. J. Biochem. Cell Biol. 33:205-220. Jamney, P.A., and Stossel, T.P. (1987). Modulation of gelsolin function by phosphatidylinositol 4,5-bisphosphate. Nature 325:362-364. Jamney, P.A. (1994). Phosphoinositides and calcium as regulators of cellular actin assembly and disassembly. Annu. Rev. Physiol. 56:169-191. Kabsch, W., and Holmes, K.C. (1995). The actin fold. FASEB. J. 9:167-174. Kessels, M.M., Engqvist-Goldstein, Å.E.Y., Drubin, D.G., and Qualmann, B. (2001). Mammalian Abp1, a signal-responsive F-actin-binding protein, links the actin cytoskeleton to endocytosis via the GTPase dynamin. J. Cell Biol. 153:351-366. Kraulis, P.J. (1991). Molscript: a program to produce both detailed and schematic plots of protein stuctures. J. appl. Cryst. 24:946-950. Lambrechts, A., Verschelde, J.-L., Jonckheere, V., Goethals, M., Vandekerckhove, J., and Ampe, C. (1997). The mammalian profilin isoforms display complementary affinities for PIP2 and proline-rich sequences. EMBO J. 16:484-494. Lambrechts, A., Braun, A., Jonckheere, V., Aszodi, A., Lanier, L.M., Robbens, J., Van Colen, I., Vandekerckhove, J., Fässler, R., and Ampe, C. (2000). Profilin II is alternatively spliced, resulting in profilin isoforms that are differentially expressed and have distinct biochemical properties. Mol. Cell. Biol. 20:8209-8219. Lappalainen, P., and Drubin, D.G. (1997). Cofilin promotes rapid actin filament turnover in vivo. Nature 388:78-82. Lappalainen, P., Fedorov, E.V., Fedorov, A.A., Almo, S.C., and Drubin, D.G. (1997). Essential functions and actin-binding surfaces of yeast cofilin revealed by systematic mutagenesis. EMBO J. 16:5520-5530.

52
Lappalainen, P., Kessels, M.M., Cope, M.J.T.V., and Drubin, D.G. (1998). The ADF homology (ADF-H) domain: a highly exploited actin-binding module. Mol. Biol. Cell 9:1951-1959. Lee, W.-L., Bezanilla, M., and Pollard, T.D. (2000). Fission yeast myosin-I, Myo1p, stimulates actin assembly by Arp2/3 complex and shares functions with WASp. J. Cell Biol. 151:789-799. Leonard, S.A., Gittis, A.G., Petrella, E.C., Pollard, T.D., and Lattman, E.E. (1997). Crystal structure of the actin-binding protein actophorin from Acanthamoeba. Nat. Struct. Biol. 4:369-373. Leung, T., Chen, X.Q., Tan, I., Manser, E., and Lim, L. (1998). Myotonic dystrophy kinase-related Cdc42-binding kinase acts as a Cdc42 effector in promoting cytoskeletal reorganization. Mol. Cell. Biol. 18:130-140. Lian, J.P., Marks, P.G., Wang, J.Y., Falls, D.L., and Badway, J.A. (2000). A protein kinase from neutrophils that specifically recognizes Ser-3 in cofilin. J. Biol. Chem. 275:2869-2876. Littlefield, R., Almenar-Queralt, A., and Fowler, V.M. (2001). Actin dynamics at pointed ends regulates thin filament length in striated muscle. Nat. Cell Biol. 3:544-551. Loisel, T.P., Boujemaa, R., Pantaloni, D., and Carlier, M.F. (1999). Reconstitution of actin-based motility of Listeria and Shigella using pure proteins. Nature 401:613-616. Lopez, I., Anthony, R.G., Maciver, S.K., Jiang, C.-J., Khan, S., Weeds, A.G., and Hussey, P.J. (1996). Pollen specific expression of maize genes encoding actin depolymerizing factor-like proteins. Proc. Natl. Acad. Sci. 93:7415-7420. Lu, J., and Pollard, T.D. (2001). Profilin binding to poly-L-proline and actin monomers along with ability to catalyze actin nucleotide exchange is required for viability of fission yeast. Mol. Biol. Cell 12:1161-1175. Luby-Phelps, K. (1994). Physical properties of cytoplasm. Curr. Opin. Cell Biol. 6:3-9. Machesky, L.M., and Insall, R. (1999). Signaling to actin dynamics. J. Cell Biol. 146:267-272. Maciver, S.K., and Weeds, A.G. (1994). Actophorin preferentially binds monomeric ADP-actin over ATP-bound actin: consequences for cell locomotion. FEBS Lett. 347:251-256. Maciver, S.K., Pope, B.J., Whytock, S., and Weeds, A.G. (1998). The effects of two actin depolymerizing factors (ADF/cofilins) on actin filament turnover: pH sensitivity of F-actin binding by human, but not Acanthamoeba actophorin. Eur. J. Biochem. 256:388-397.

53
Maciver, S.K., and Hussey, P.J. (2002). The ADF/cofilin family: actin remodeling proteins. Genome Biol. 3:3007.1-3007.12. Maekawa, M., Ishizaki, T., Boku, S., Watanabe, N., Fujita, A., Iwamatsu, A., Obinata, T., Ohashi, K., Mizuno, K., and Narumiya, S. (1999). Signaling from Rho to the actin cytoskeleton through protein kinases ROCK and LIM-kinase. Science 285:895-898. McGough, A., Pope, B., Chiu, W., and Weeds, A. (1997). Cofilin changes the twist of F-actin: implications for actin filament dynamics and cellular function. J. Cell Biol. 138:771-781. McGough, A. (1998). F-actin-binding proteins. Curr. Opin. Struct. Biol. 8:166-176. McGough, A., and Chiu, W. (1999). ADF/cofilin weakens lateral contacts in the actin filament. J. Mol. Biol. 291:513-519. McKim, K.S., Matheson, C., Marra, M.A., Wakarchuk, M.F., and Baillie, D.L. (1994). The Caenorhabditis elegans unc-60 gene encodes proteins homologous to a family of actin binding proteins. Mol. Gen. Genet. 242:346-357. Merritt, E.A., Murphy, and Michael, E.P. (1994). Raster 3D version 2.0: a program for photorealistic molecular graphics. Acta Cryst. D50:869-873. Miki, H., Suetsugu, S., and Takenawa, T. (1998). WAVE, a novel WASP-family protein involved in actin reorganization induced by Rac. EMBO J. 17:6932-6941. Mohri, K., Takano-Ohmuro, H., Nakashima, H., Hayakawa, K., Endo, T., Hanaoka, K., and Obinata, T. (2000). Expression of cofilin isoforms during development of mouse striated muscles. J. Musc. Res. Cell Motil. 21:49-57. Moon, A.L., Jamney, P.A., Louie, K.A., and Drubin, D.G. (1993). Cofilin is essential component of the yeast cortical cytoskeleton. J. Cell Biol. 120:421-435. Moon, A., and Drubin, D.G. (1995). The ADF/cofilin proteins: stimulus-responsive modulators of actin dynamics. Mol. Biol. Cell 6:1423-1431. Moriyama, K., Nishida, E., Yonezawa, N., Sakai, H., Matsumoto, S., Iida, K., and Yahara, I. (1990). Destrin, a mammalian actin-depolymerizing protein is closely related to cofilin. Cloning and expression of porcine brain destrin cDNA. J. Biol. Chem. 265:5768-5773. Nagaishi, K., Adachi, R., Matsui, S., Yamaguchi, T., Kasahar, T., and Suzuki, K. (1999). Herbimycin A inhibits both dephosphorylation and translocation of cofilin induced by opsonized zymosan in macrophagelike U937 cell. J. Cell Physiol. 180:345-354.

54
Niwa, R., Nagata-Ohashi, K., Takeichi, M., Mizuno, K., and Uemura, T. (2002). Control of actin reorganization by Slingshot, a family of phosphatases that dephosphorylate ADF/cofilin. Cell 108:233-246. Obinata, T., Nagaoka-Yasuda, R., Ono, S., Kusano, K., Mohri, K., Ohtaka, Y., Yamashiro, S., Okada, K., and Abe, H. (1997). Low molecular-weight G-actin binding proteins involved in the regulation of actin assembly during myogenesis. Cell Struct. Funct. 22:181-189. Ogawa, K., Tashima, M., Yumoto, Y., Okuda, T., Sawada, H., Okuma, M., and Maruyama, Y. (1990). Coding sequence of human placenta cofilin cDNA. Nucleic Acids Res. 18:7169. Ojala, P.J., Paavilainen, V., and Lappalainen, P. (2001). Identification of yeast cofilin residues specific for actin monomer and PIP2 binding. Biochemistry 40:15562-15569. Ono, S., Minami, N., Abe, H., and Obinata, T. (1994). Characterization of a novel cofilin isoform that is predominantly expressed in mammalian skeletal muscle. J. Biol. Chem. 269:15280-15286. Ono, S., and Benian, G.M. (1998). Two Caenorhabditis elegans actin depolymerizing factor/cofilin proteins, encoded by the unc-60 gene, differentially regulate actin dynamics. J. Biol. Chem. 273:3778-3783. Ono, S. (2001). The Caenorhabditis elegans unc-78 gene encodes a homologue of actin-interacting protein 1 required for organized assenbly of muscle actin filaments. J. Cell Biol. 152:1313-1319. Ono, S., and Ono, K. (2002). Tropomyosin inhibits ADF/cofilin-dependent actin filament dynamics. J. Cell Biol. 156:1065-1076. Palmgren, S., Ojala, P.J., Wear, M.A., Cooper, J.A., and Lappalainen, P. (2001). Interactions with PIP2, ADP-actin monomers, and capping protein regulate the activity and localization of yeast twinfilin. J. Cell Biol. 155:251-260. Palmgren, S., Vartiainen, M., and Lappalainen, P. (2002). Twinfilin, a molecular mailman for actin monomers. J. Cell Sci. 115:881-886. Pantaloni, D., and Carlier, M.F. (1993). How profilin promotes actin filament assembly in the presence of thymosin β4. Cell 75:1007-1014. Paunola, E., Mattila, P.K., and Lappalainen, P. (2001). WH2 domain: a small versatile adapter for actin monomers. FEBS Lett. 25626:1-6.

55
Petersen, N.S., Lankenau, D.-H., Mitchell, H.K., Young, P., and Corces, V.G. (1994). Forked proteins are components of fiber bundles present in developing bristles of Drosophila melanogaster. Genetics 136:173-182. Pollard, T.D., Blanchoin, L., and Mullins, R.D. (2000). Molecular mechanisms controlling actin filament dynamics in nonmuscle cells. Ann. Rev. Biophys. Biomol. Struct. 29:545-576. Puius, Y.A., Mahoney, N.M., and Almo, S.C. (1998). The modular structure of actin-regulatory proteins. Curr. Opin. Cell Biol. 10:23-34. Rando, O.J., Zhao, K., and Crabtree, G.R. (2000). Searching for a function for nuclear actin. Trends Cell Biol. 10:92-97. Ressad, F., Didry, D., Xia, G.-X., Hong, Y., Chua, N.-H., Pantaloni, D., and Carlier, M.-F. (1998). Kinetic analysis of the interaction of actin-depolymerizing factor (ADF)/cofilin with G- and F-actins. J. Biol. Chem. 273:20894-20902. Ridley, A.J., Paterson, H.F., Johnson, C.L., Diekman, D., and Hall, A. (1992). The small GTP-binding protein Rac regulates growth factor induced membrane ruffling. Cell 70:401-410. Ridley, A.J. (2001). Rho GTPases and cell migration. J. Cell Sci. 114:2713-2722. Rodal, A., Tetreault, J.W., Lappalainen, P., Drubin, D.G., and Amberg, D.C. (1999). Aip1p interacts with cofilin to disassemble actin filaments. J. Cell Biol. 145:1251-1264. Rohatgi, R., Ma, L., Miki, H., Lopez, M., Kirchhausen, T., Takenawa, T., and Kirschner, M.W. (1999). The interaction between N-WASP and the Arp2/3 complex links Cdc42-dependent signals to actin assembly. Cell 97:221-231. Rohwer, A., Kittstein, W., Marks, F., and Gschwendt, M. (1999). Cloning expression and characterization of an A6-related protein. Eur. J. Biochem. 263:518-525. Rozelle, A.L., Machesky, L.M., Yamamoto, M., Driessens, M.H.E., Insall, R.H., Roth, M.G., Luby-Phelps, K., Marriott, G., Hall, A., and Yin, H.L. (2000). Phosphatidyl 4,5-bisphosphate induces actin based movement of raft-enriched vesicles through WASP-Arp2/3. Curr. Biol. 10:311-320. Sanders, L.C., Matsumura, F., Bokoch, G.M., and de Lanerolle, P. (1999). Inhibition of myosin light chain kinase by p21-activated kinase. Science 283:2083-2085. Sanger, J.W., Mittal, B., and Sanger, J.M. (1984). Analysis of myofibrillar structure and assembly using fluorescently labeled contractile proteins. J. Cell Biol. 98:825-833.

56
Schafer, D.A., Jennings, P.B., and Cooper, J.A. (1996). Dynamics of capping protein and actin assembly in vitro: uncapping barbed ends by polyphosphoinositides. J. Cell Biol. 135:169-179. Schluter, K., Jockusch, B.M., and Rothkegel, M. (1997). Profilins as regulators of actin dynamics. Biochim. Biophys. Acta 1359:97-109. Sechi, A.S. and Wehland, J. (2000). The actin cytoskeleton and plasma membrane connection: PtdIns(4,5)P2 influences cytoskeletal protein activity at the plasma membrane. J. Cell Sci. 113:3685-3695. Sellers, J.R. (2000). Myosins: a diverse superfamily. Biochim. Biophys. Acta 1496:3-22. Sept, D., Elcock, A.H., and McCammon, J.A. (1999). Computer simulations of actin polymerization can explain the barbed-pointed end asymmetry. J. Mol. Biol. 294:1181-1189. Sheterline, P., Clayton, J., and Sparrow, J.C. (1994). Protein profile: actin. Fourth edition, Oxford University Press, Oxford. Small, J.V., Stradal, T., Vignal, E., and Rottner, K. (2002). The lamellipodium: where motility begins. Trends Cell Biol. 12:112-120. Sohn, R.H., Chen, J., Koblan, K.S., Bray, P.F., and Goldschmidt-Clermont, P.J. (1995). Localization of a binding site for phosphatidylinositol 4,5-bisphosphate on human profilin. J. Biol. Chem. 270:21114-21120. Steinmetz, M.O., Stoffler, D., Hoenger, A., Bremer, A., and Aebi, U. (1997). Actin: from cell biology to atomic detail. J. Struct. Biol. 119:295-320. Sumi, T., Matsumoto, K., Shibuya, A., and Nakamura, T. (2001). Activation of LIM-kinases by myotonic dystrophy kinase-related Cdc42-binding kinase α. J. Biol. Chem. 276:23092-23096. Svitkina, T.M., Verkhovsky, A.B., McQuade, K.M., and Borisy, G.G. (1997). Analysis of the actin-myosin II system in fish epidermal keratocytes: mechanism of cell body translocation. J. Cell Biol. 139:397-415. Svitkina, T.M., and Borisy, G.G. (1999). Arp2/3 complex and actin depolymerizing factor/cofilin in dendritic organization and treadmilling of actin filament array in lamellipodia. J. Cell Biol. 145:1009-1026. Takenawa, T., and Miki, H. (2001). WASP and WAVE family proteins: key molecules for rapid rearrangement of cortical actin filaments and cell movement. J. Cell Sci. 114:1801-1809.

57
Taunton, J., Rowning, B.A., Coughlin, M.L., Wu, M., Moon, R.T., Mitchison, T.J., and Larabell, C.A. (2000). Actin-dependent propulsion of endosomes and lysosomes by recruitment of N-WASP. J. Cell Biol. 148:519-530. Thirion, C., Stucka, R., Mendel, B., Gruhler, A., Jaksch, M., Nowak, K.J., Binz, N., Laing, N.G., and Lochmuller, H. (2001). Characterization of human muscle type cofilin (CFL2) in normal and regenerating muscle. Eur. J. Biochem. 268:3473-3482. Tilney, L. G., and Portnoy, D.A. (1989). Actin filaments and the growth, movement, and spread of the intracellular bacterial parasite, Listeria monocytogenes. J. Cell Biol. 109:1597-1608. Tilney, L.G., Tilney, M.S., and Guild, G.M. (1995). F-actin bundles in Drosophila bristles. I. Two filament cross-links are involved in bundling. J. Cell Biol. 130:629-638. Tilney, L.G., Connelly, P.S., Vranich, K.A., Shaw, M.K., and Guild, G.M. (1998). Why are two different cross-linkers necessary for actin bundle formation in vivo and what does each cross-link contribute? J. Cell Biol. 143:121-133. Tilney, L.G., Connelly, P.S., Vranich, K.A., Shaw, M.K., and Guild, G.M. (2000a). Regulation of actin filament cross-linking and bundle shape in Drosophila bristles. J. Cell Biol. 148:87-99. Tilney, L.G., Connelly, P.S., Vranich, K.A., Shaw, M.K., and Guild, G.M. (2000b). Actin filaments and microtubules play different roles during bristle elongation in Drosophila. J. Cell Sci. 113:1255-1265. Toshima, J., Toshima, J.Y., Amano, T., Yang, N., Narumiya, S., and Mizuno, K. (2001a). Cofilin phosphorylation by protein kinase testicular protein kinase 1 and its role in integrin mediated actin reorganization and focal adhesion formation. Mol. Biol. Cell 12:1131-1145. Toshima, J., Toshima, J.Y., Takeuchi, K., Mori, R., and Mizuno, K. (2001b). Cofilin phosphorylation and actin reorganization activities of testicular protein kinase 2 and its predominant expression in testicular sertoli cells. J. Biol. Chem. 276:31449-31458. Toshima, J.Y., Toshima, J., Watanabe, T., and Mizuno, K. (2001c). Binding of 14-3-3b regulates the kinase activity and subcellular localization of testicular protein kinase 1. J. Biol. Chem. 276:43471-43481. Tsukita, S., and Yonemura, S. (1999). Cortical actin organization: lessons from ERM (ezrin/radixin/moesin) proteins. J. Biol. Chem. 274:34507-34510. Uruno, T., Liu, J., Zhang, P., Fan, Y.-x., Egile, C., Li, R., Mueller, S.C., and Zhan, X. (2001). Activation of Arp2/3 complex-mediated actin polymerization by cortactin. Nat. Cell Biol. 3:259-266.

58
Van den Ent, F., Amos, L., and Löwe, J. (2001). Bacterial ancestry of actin and tubulin. Curr. Opin. Microbiol. 4: 634-638. Van Troys, M., Vandekerckhove, J., and Ampe, C. (1999). Structural modules in actin-binding proteins: towards a new classification. Biochim. Biophys. Acta 1448:323-348. Verheyen, E.M., and Cooley, L. (1994). Profilin mutations disrupt multiple actin dependent processes during Drosophila development. Development 120:717-728. Vexler, Z.S., Symons, M., and Barber, D.L. (1996). Activation of Na+/H+ exchange is necessary for RhoA-induced stress fiber formation. J. Biol. Chem. 271: 22281-22284. Watanabe, N., Kato, T., Fujita, A., Ishizaki, T., and Narumiya, S. (1999). Cooperation between mDia and ROCK in Rho-induced actin reorganization. Nat. Cell Biol. 1:136-143. Welch, M., Mallavarapu, A., Rosenblatt, J., and Mitchison, T. (1997). Actin dynamics in vivo. Curr. Opin. Cell Biol. 9:54-61. Welch, M.D., Rosenblatt, J., Skoble, J., Portnoy, D.A., and Mitchison, T.J. (1998). Interaction of human Arp2/3 complex and the Listeria monocytogenes ActA protein in actin filament nucleation. Science 281:105-108. Witke, W., Podtelejnikov, A.V., Di Nardo, A., Sutherland, J.D., Gurniak, C.G., Dotti, C., and Mann, M. (1998). In mouse brain profilin I and profilin II associate with regulators of the endocytic pathway and actin assembly. EMBO J. 17:967-976. Wolven, A.K., Belmont, L.D., Mahoney, N.M., Almo, S.C., and Drubin, D.G. (2000). In vivo importance of actin nucleotide exchange catalyzed by profilin. J. Cell Biol. 150:895-903. Wriggers, W., Tang, J.X., Azuma, T., Marks, P.W., and Jamney, P. (1998). Cofilin and gelsolin segment-1: molecular dynamics simulation and biochemical analysis predict a similar actin binding mode. J. Mol. Biol. 282:921-932. Yang, N., Higuchi, O., Ohashi, K., Nagata, K., Wada, A., Kangawa, K., Nishida, E., and Mizuno, K. (1998). Cofilin phosphorylation by LIM-kinase 1 and its role in Rac-mediated actin reorganization. Nature 393:809-812. Yarmola, E.G., Parikh, S., and Bubb, M.R. (2001). Formation of a ternary complex of profilin, thymosin β4, and actin. J. Biol. Chem. 276:45555-45563.

59
Yeoh, S., Pope, B., Mannherz, H.G., and Weeds, A. (2002). Determining the differences in actin binding by human ADF and cofilin. J. Mol. Biol. 315:911-925. Yonezawa, N., Nishida, E., Koyasu, S., Maekawa, S., Ohta, Y., Yahara, I., and Sakai, H. (1987). Distribution among tissues and intracellular localization of cofilin, a 21 kDa actin-binding protein. Cell Struct. Funct. 12:443-452. Yonezawa, N., Nishida, E., Iida, K., Yahara, I., and Sakai, H. (1990). Inhibition of interactions of cofilin, destrin and deoxyribonuclease I with actin by phosphoinositides. J. Biol. Chem. 265:8382-8386. Yu, F.-X., Lin, S.-C., Morrison-Bogorad, M., Atkinson, M.A.L., and Yin, H.L. (1993). Thymosin β10 and thymosin β4 are both actin monomer sequestering proteins. J. Biol. Chem 268:502-509.


















