
Reduced Expression of Ribosomal Proteins Relieves MicroRNA ...
17
Reduced Expression of Ribosomal Proteins Relieves MicroRNA-Mediated Repression The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation Janas, Maja M., Eric Wang, Tara Love, Abigail S. Harris, Kristen Stevenson, Karlheinz Semmelmann, Jonathan M. Shaffer, et al. “Reduced Expression of Ribosomal Proteins Relieves MicroRNA- Mediated Repression.” Molecular Cell 46, no. 2 (April 2012): 171-186. © 2012 Elsevier. As Published http://dx.doi.org/10.1016/j.molcel.2012.04.008 Publisher Elsevier B.V. Version Final published version Citable link http://hdl.handle.net/1721.1/83590 Terms of Use Article is made available in accordance with the publisher's policy and may be subject to US copyright law. Please refer to the publisher's site for terms of use.
Transcript of Reduced Expression of Ribosomal Proteins Relieves MicroRNA ...

Reduced Expression of Ribosomal ProteinsRelieves MicroRNA-Mediated Repression
The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters.
Citation Janas, Maja M., Eric Wang, Tara Love, Abigail S. Harris, KristenStevenson, Karlheinz Semmelmann, Jonathan M. Shaffer, et al.“Reduced Expression of Ribosomal Proteins Relieves MicroRNA-Mediated Repression.” Molecular Cell 46, no. 2 (April 2012):171-186. © 2012 Elsevier.
As Published http://dx.doi.org/10.1016/j.molcel.2012.04.008
Publisher Elsevier B.V.
Version Final published version
Citable link http://hdl.handle.net/1721.1/83590
Terms of Use Article is made available in accordance with the publisher'spolicy and may be subject to US copyright law. Please refer to thepublisher's site for terms of use.

Molecular Cell
Article
Reduced Expression of Ribosomal ProteinsRelieves MicroRNA-Mediated RepressionMaja M. Janas,1,3,4 Eric Wang,5,6 Tara Love,1,3,4,8 Abigail S. Harris,7 Kristen Stevenson,2 Karlheinz Semmelmann,7
Jonathan M. Shaffer,7 Po-Hao Chen,1,3,4 John G. Doench,4 Subrahmanyam V.B.K. Yerramilli,7 Donna S. Neuberg,2
Dimitrios Iliopoulos,1,3 David E. Housman,5,6 Christopher B. Burge,5 and Carl D. Novina1,3,4,*1Department of Cancer Immunology and AIDS, Dana-Farber Cancer Institute2Department of Biostatistics and Computational Biology, Dana-Farber Cancer Institute3Department of Microbiology and Immunobiology
Harvard Medical School, Boston, MA 02115, USA4Broad Institute of Harvard and MIT, Cambridge, MA 02141, USA5Department of Biology6The Koch Institute
Massachusetts Institute of Technology, Cambridge MA 02139, USA7Qiagen, Frederick, MD 21703, USA8Present address: Correlagen Diagnostics, 307 Waverly Oaks Road, Suite 101, Waltham, MA 02452, USA
*Correspondence: [email protected]
DOI 10.1016/j.molcel.2012.04.008
SUMMARY
MicroRNAs (miRNAs) regulate physiological andpathological processes by inducing posttranscrip-tional repression of target messenger RNAs (mRNAs)via incompletely understood mechanisms. To dis-cover factors required for human miRNA activity, weperformed an RNAi screen using a reporter cell lineof miRNA-mediated repression of translation initia-tion. We report that reduced expression of ribosomalprotein genes (RPGs) dissociated miRNA complexesfrom target mRNAs, leading to increased polysomeassociation, translation, and stability of miRNA-tar-geted mRNAs relative to untargeted mRNAs. RNAsequencing of polysomes indicated substantial over-lap in sets of genes exhibiting increasedor decreasedpolysomal association after Argonaute or RPGknockdowns, suggesting similarity in affected path-ways.miRNAprofiling ofmonosomesandpolysomesdemonstrated that miRNAs cosediment with ribo-somes. RPG knockdowns decreased miRNAs inmonosomes and increased their target mRNAs inpolysomes.Our data show thatmostmiRNAs represstranslation and that the levels of RPGs modulatemiRNA-mediated repression of translation initiation.
INTRODUCTION
MicroRNA (miRNA)-mediated translational repression of
messenger RNA (mRNA) targets was first described in
C. elegans, where lin-4 and let-7 were reported to reduce pro-
tein levels without affecting mRNA stability (Olsen and Ambros,
1999; Wightman et al., 1993). Untargeted and miRNA-targeted
mRNAs demonstrated similar polysomal association as as-
sessed by density gradient centrifugation, suggesting that
miRNAs repressed translation after initiation (Olsen and Am-
bros, 1999; Seggerson et al., 2002). In mammals, polysome
profiling studies led to contradictory models of miRNA-
mediated translational repression. Although miRNA-targeted
mRNAs had reduced polysomal association compared to
unrepressed mRNAs in some studies, suggesting repression
of initiation (Bhattacharyya et al., 2006a; Pillai et al., 2005),
miRNAs and their mRNA targets had robust polysomal associ-
ation in other studies, suggesting repression after initiation
(Maroney et al., 2006; Nelson et al., 2004; Nottrott et al.,
2006; Petersen et al., 2006). Additionally, treatment with trans-
lation elongation inhibitors shifted miRNAs and miRNA-targeted
mRNAs from polysomes toward monosomes, indicating that
miRNAs target actively translating mRNAs (Maroney et al.,
2006; Nottrott et al., 2006).
Subsequent studies demonstrated that miRNAs could also
reduce mRNA stability. In C. elegans, lin-4 and let-7 decreased
target mRNA levels (Bagga et al., 2005). In both C. elegans and
mammals, miRNAs were shown to promote deadenylation,
leading to reduced target mRNA stability (Behm-Ansmant
et al., 2006; Wu and Belasco, 2005; Wu et al., 2006). In
mammals, transfected miRNAs globally reduced levels of target
mRNAs (Lim et al., 2005), and ribosome footprinting determined
that miRNA targeting resulted in a decrease in ribosome density
that was only modestly (�15%) greater than the decrease in
mRNA level, suggesting that miRNAs act primarily to destabilize
mRNAs (Guo et al., 2010). More recent ribosome footprinting
analysis at earlier time points demonstrated that repression of
translation initiation precedes deadenylation for the majority of
miR-430-targeted mRNAs in zebrafish and that mRNA decay is
likely a consequence of these earlier activities (Bazzini et al.,
2012).
Here we report that �70% of all miRNA species expressed
in HeLa cells are detected in monosomal and polysomal frac-
tions, implying that the majority of miRNA species can repress
translation. We also demonstrate that ribosomal protein genes
Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc. 171

(RPGs) preferentially regulate miRNA-mediated repression of
translation initiation. Specifically, RPG depletion led to
decreased monosomal association of a subset of miRNAs and
increased polysomal association of mRNA targets of those
miRNAs, indicating relief of translation initiation block. This
study establishes a framework for understanding ribosomes as
global regulators of miRNA-mediated repression of translation
initiation.
RESULTS
A Reporter of miRNA-Mediated Repression ofTranslation InitiationWe engineered a clonal HeLa cell line we called D8 that stably
expresses two reporter genes: Firefly Luciferase (FL) with six
binding sites for endogenous miR-21 in its 30 untranslated region
(30 UTR), and Renilla Luciferase (RL) lacking miRNA binding sites
(Figure 1A). Inhibition of miR-21 with a miR-21-specific antago-
mir resulted in up to 7-fold increase in FL relative to RL as
assessed by the dual luciferase assay (Figure 1B). The miR-21-
specific antagomir increased FL protein levels more than FL
mRNA levels, even after 144 hr of antagomir treatment (Fig-
ure S1A available online). These results show that in D8, FL
was strongly repressed by miR-21 and suggest that the mecha-
nism involved was repression of translation.
Additionally, knockdown of miRNA effector genes (Argo-
nautes [Agos] or eIF6) yielded a 2- to 10-fold increase in FL
protein levels accompanied by a proportional yet smaller
increase in FL mRNA levels (Figure 1C). Knockdown of miRNA
biogenesis (Drosha and DGCR8) or mRNA turnover (DCP2 and
PARN) genes also increased FL levels in a small interfering
RNA (siRNA) dose-dependent manner (Figure S1B). These
data show that D8 has a large dynamic range and high sensitivity
to reduced levels of miRNA pathway proteins and thus is
an appropriate tool to discover new factors and regulators of
miRNA-mediated repression by gain-of-signal RNA interference
(RNAi) screening.
To determine which step of FL translation is inhibited by
miR-21 in D8, we performed polysome profiling of control,
Ago1- or eIF6-depleted cells (Figures 1D and 1E). In control
cells, the majority of miR-21-targeted FL mRNA was in mono-
somes, and the majority of untargeted RL mRNA was in poly-
somes. Knockdown of Ago1 or eIF6 led to a striking shift of
FL mRNA from monosomes to polysomes, while RL mRNA
exhibited only a modest shift toward heavier polysome frac-
tions but remained polysomal in all conditions. Consistent
with RT-qPCR data in Figure 1C and with previously reported
miRNA effects on mRNA stability (Bagga et al., 2005; Guo
et al., 2010; Behm-Ansmant et al., 2006; Wu and Belasco,
2005; Wu et al., 2006), knockdown of Ago1 or eIF6 stabilized
miR-21-targeted FL mRNA, leading to a higher combined signal
for FL but not RL mRNA across gradients relative to the Scr
control. Consistent with its role in 60S ribosomal subunit
biogenesis (Basu et al., 2001), eIF6 knockdown strongly
reduced the levels of 60S ribosomal subunits without affecting
the levels of 40S subunits as assessed by A254 absorbance
traces (Figure 1D). These data demonstrate that in D8, miR-
21 repressed initiation of FL translation.
An RNAi Screen Identifies RPGs as Regulatorsof the miRNA PathwayRNAi screens for RNAi pathway genes have been used effec-
tively in simpler eukaryotes (Dorner et al., 2006; Eulalio et al.,
2007; Kim et al., 2005; Parry et al., 2007; Saleh et al., 2006; Ulvila
et al., 2006), leading to identification of many effectors of small
RNA function, such as RNA binding proteins, cytoskeletal
proteins, and one RPG in C. elegans (Parry et al., 2007), as
well as several RPGs and proteasomal components in
D. melanogaster (Zhou et al., 2008). However, similar screens
have not yet been reported in mammals. Therefore, we used
D8 to perform an RNAi screen for effectors and regulators of
the human miRNA pathway (Figure 2A). In all, �7,000 genes
were targeted with four individual siRNAs per gene from the
Druggable Genome Library of siRNAs (QIAGEN), and effects of
knockdowns were assessed with the dual luciferase assay.
Using stringent criteria (at least two siRNAs per gene causing
derepression of FL R eIF6 knockdown), 314 genes (4.3%)
scored as hits (Table S1, part A). RPGs were enriched more
than 13-fold in this screen: seven out of 12 RPGs (58%) scored
as hits. Closer inspection of the screening data showed that all
12 RPG knockdowns derepressed FL to some extent. Gene
ontology analysis revealed that the ribosomal pathway was
one of the most significantly enriched pathways in the screen
(Figure 2B and Table S1, part B). Thus, we hypothesized that
as a class RPGs might regulate miRNA repression.
To determine whether depletion of every RPG inhibits
miRNA-mediated repression, we targeted all 79 RPGs with
four individual siRNAs per gene in D8. Knockdown of virtually
all 40S (Figure 2C) and 60S (Figure 2D) RPGs with at least
one siRNA led to derepression of FL that was greater than
derepression after eIF6 knockdown. After three days (the point
of strongest FL derepression), most RPG knockdowns reduced
cell numbers by 20%–50%. The Y chromosome-expressed
S4Y1 and S4Y2 provide natural negative controls in female
cells and did not score in our HeLa cell validation screen (Fig-
ure 2C). The consistent FL derepression after depletion of
almost every 40S and 60S RPG suggests that altered ribo-
somal integrity or altered ribosomal subunit stoichiometry
and not extraribosomal functions of RPGs led to decreased
miRNA activity. Supporting this conclusion, knockdown of
factors involved in either 40S (Bms1 and Tsr1) or 60S (Bop1
and Nip7) subunit biogenesis also led to derepression of
miRNA-targeted mRNAs (Figure S2A), and double knockdown
of 40S and 60S RPGs led to greater derepression of miRNA-
targeted mRNAs compared to single RPG knockdowns
(Figure S2B).
To validate the screening results, we chose five representative
40S (S5, S11, S12, S15, and S18) and 60S (L5, L11, L12, L35A,
and LP2) RPGs for detailed biochemical analysis. Knockdown
of each RPG in D8 (Figure S2C) derepressed FL mostly at the
protein levels, although proportional increases in FL mRNA
levels were also observed (Figure S2D), consistent with knock-
downs of known miRNA pathway genes (Figure 1C). In the lucif-
erase screens, which were not normalized to total protein, deple-
tion of RPGs increased FL levels and decreased RL levels
(Figure S2E). These trends were affected by reduced cell
numbers with, on average, half the number of cells after 72 hr
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
172 Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc.

Firefly
Renilla
miR-21A
B
miR-21
tRNA
0 5 20 100
0
2
4
6
8
10
12
Fol
d D
erep
ress
ion
ProteinmRNA
C
miR-21 antagomir [nM]
Fol
d D
erep
ress
ion
Ago1Actin
Ago2Actin
eIF6Actin
Scr
Ago
1
Scr
Ago
2
Scr
eIF
6
ProteinmRNA
D
0 5 20 100
Scr Ago1 Ago2 eIF6
Scr
Ago1
eIF6
FL
RL
28S
18S
A254
FL
RL
28S
18S
A254
FL
RL
28S
18S
A254
0
1
2
3
4
5
6
7
8
CMV
CMV
0102030405060708090
1 2 3 4 5 6 7 8 9 10 11 12Fraction number
% F
L m
RN
A
ScrAgo1eIF6
E
0
10
20
30
40
50
60
70
1 2 3 4 5 6 7 8 9 10 11 12Fraction number
% R
L m
RN
A
ScrAgo1eIF6
40S 60S 80S Polysomes
40S 60S 80S Polysomes
40S 60S 80S Polysomes
Figure 1. D8 Is a Highly Sensitive Reporter of miR-21-Mediated Repression of Translation Initiation
(A) Schematic representation of CMV-driven luciferase reporters stably expressed in D8. Firefly Luciferase (FL) possesses six imperfect miR-21 binding sites in its
30 UTR, while Renilla Luciferase (RL) lacks miRNA binding sites.
(B) D8 reports on miR-21-mediated repression. A miR-21-specific antagomir was transfected into D8 at indicated concentrations, and FL and RL protein and
mRNA levels were assessed after 24 hr by the dual luciferase assay and RT-qPCR, respectively. Fold derepression was calculated as (FLtest/RLtest)/(FLcontrol/
RLcontrol). The antagomir reduced miR-21 levels in a dose-dependent fashion as assessed by northern blotting.
(C) D8 reports on miRNA effector function. D8 was transfected with Scrambled (Scr), Ago1-, Ago2-, or eIF6-specific siRNAs, and FL and RL protein and mRNA
levels were assessed after 72 hr. Knockdowns of Ago1, Ago2, and eIF6 were confirmed by western blotting.
(D) Polysomeprofiling demonstrates that knockdown ofAgo1 or eIF6 shifts FLmRNA frommonosomes to polysomes. Lysates fromD8 transfected either with Scr
or with Ago1- or eIF6-specific siRNAs were fractionated while absorbance was monitored at 254 nm (A254). FL and RL mRNAs were assessed by northern
blotting, and 28S and 18S rRNAs were assessed by ethidium bromide staining.
(E) Quantification of northern blots with ImageQuant. FL and RLmRNA detected in each fraction is represented as the percent of the total mRNA detected in all
fractions across the gradient.
Bar graphs show the mean ± SD from three independent experiments. See also Figure S1.
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc. 173
A
B
C
Scr
eIF
6L
3L
4L
5L
6L
7L
7A
L8
L9
L1
0L
11
L1
2L
13
L1
3A
L1
4L
15
L1
7L
18
L1
8A
L1
9L
21
L2
2L
23
AL
24
L2
6L
27
L2
7A
L2
8L
29
L3
1L
32
L3
4L
35
L3
5A
L3
6L
36
AL
37
L3
7A
L3
8L
39
L4
0L
41
LP
0L
P1
LP
2
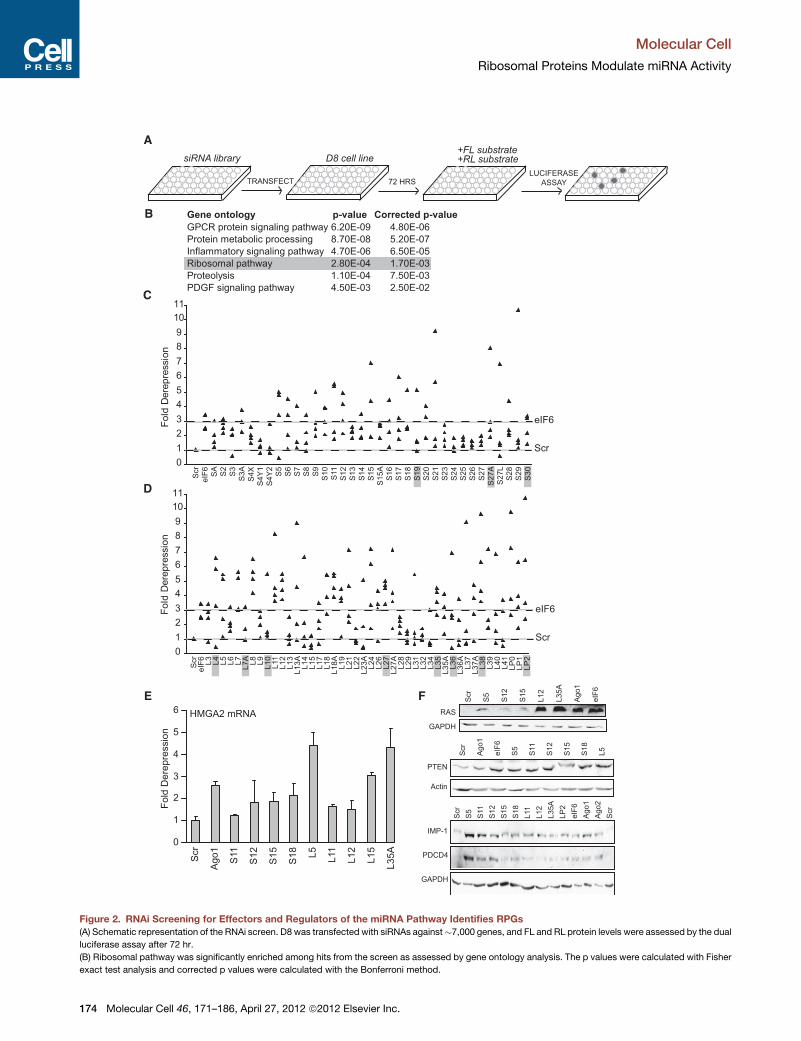
Gene ontology p-value Corrected p-value
GPCR protein signaling pathway 6.20E-09 4.80E-06
Protein metabolic processing 8.70E-08 5.20E-07
Inflammatory signaling pathway 4.70E-06 6.50E-05
Ribosomal pathway 2.80E-04 1.70E-03
Proteolysis 1.10E-04 7.50E-03
PDGF signaling pathway 4.50E-03 2.50E-02
TRANSFECT
Fo
ldD
ere
pre
ssio
n
Scr
eIF
6
SA
S2
S3
S3
A
S4
X
S4
Y1
S4
Y2
S5
S6
S7
S8
S9
S1
0
S11
S1
2
S1
3
S1
4
S1
5
S1
5A
S1
6
S1
7
S1
8
S1
9
S2
0
S2
1
S2
3
S2
4
S2
5
S2
6
S2
7
S2
7A
S2
7L
S2
8
S2
9
S3
0
D
Fo
ldD
ere
pre
ssio
nsiRNA library D8 cell line
72 HRS
+FL substrate
+RL substrate
LUCIFERASE
ASSAY
eIF6
Scr
0
1
2
3
4
5
6
7
8
9
10
11
0
1
2
3
4
5
6
7
8
9
10
11
eIF6
Scr
E F
0
1
2
3
4
5
6
Fo
ldD
ere
pre
ssio
n
HMGA2 mRNA
Scr
Ag
o1
S11
S1
2
S1
5
S1
8
L5
L11
L1
2
L1
5
L3
5A
PTEN
Actin
Scr
Ag
o1
eIF
6
S5
S11
S1
2
S1
5
S1
8
L5
Scr
S5
S11
S1
2
S1
5
S1
8
L11
L1
2
L3
5A
LP
2
eIF
6
Ag
o1
Ag
o2
Scr
Scr
S5
S1
2
S1
5
L1
2
L3
5A
Ag
o1
eIF
6
RAS
GAPDH
IMP-1
PDCD4
GAPDH
Figure 2. RNAi Screening for Effectors and Regulators of the miRNA Pathway Identifies RPGs
(A) Schematic representation of the RNAi screen. D8 was transfected with siRNAs against�7,000 genes, and FL and RL protein levels were assessed by the dual
luciferase assay after 72 hr.
(B) Ribosomal pathway was significantly enriched among hits from the screen as assessed by gene ontology analysis. The p values were calculated with Fisher
exact test analysis and corrected p values were calculated with the Bonferroni method.
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
174 Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc.

of RPG knockdown, presumably because RPG depletion
reduced cellular proliferation rates as previously reported (Fly-
gare et al., 2005; Kirn-Safran et al., 2007;Miller et al., 2003; Oliver
et al., 2004; Oristian et al., 2009; Pani�c et al., 2007; Volarevic
et al., 2000). When normalized to total protein by western blot-
ting, we observed increased FL and unchanged RL levels (Fig-
ure S2F). To further validate these findings and to control for
possible siRNA off-target effects, we expressed five siRNA-
resistant isoforms of RPGs (S5, S12, S15, L12, and LP2) in D8
cells knocking down these RPGs. In all cases, transfection of
constructs expressing RPGs with silent mutations in regions
complementary to siRNAs partially rescued the FL derepression
(Figure S2G), demonstrating that RPG depletion directly led to
miRNA-targeted mRNA derepression.
We next assessed the effect of RPG depletion on various
reporter and endogenous miRNA-targeted mRNAs. FL and RL
reporters containing six imperfectly complementary binding
sites for the artificial CXCR4 miRNA were derepressed in RPG
knockdowns only in the presence of the CXCR4 miRNA (Figures
S2H and S2I). However, an FL reporter containing one perfectly
complementary site for the CXCR4 miRNA was only modestly
affected by RPG knockdowns (Figure S2J), indicating that
RPGs preferentially regulate miRNA-mediated translational
repression and not siRNA-mediated mRNA cleavage. The RL
reporter containing HMGA2 30 UTR with seven seed matches
to the let-7 miRNA (Mayr and Bartel, 2009) was derepressed
upon RPG knockdowns (Figure S2K). Consistent with previous
studies showing that repression of HMGA2 correlates with its
mRNA level (Lee and Dutta, 2007), endogenous HMGA2
mRNA levels increased afterRPG knockdowns (Figure 2E). Simi-
larly, an RL reporter containing the KRAS 30 UTRwith seven let-7
sites (Johnson et al., 2005) was derepressed in HeLa cells upon
RPG knockdowns (Figure S2L), as was endogenous RAS protein
(Figure 2F). Other validated endogenous miRNA-targeted
mRNAs, such as miR-21-targeted PTEN (Lewis et al., 2003;
Talotta et al., 2009), let-7-targeted IMP-1 (Boyerinas et al.,
2008), and miR-21-targeted PDCD4 (Asangani et al., 2008)
were all derepressed upon RPG knockdowns (Figure 2F).
Although the degree and pattern of derepression varied between
different miRNA-targeted mRNAs, as a class RPGs regulated
miRNA repression.
RPG Knockdowns Relieve Repression of TranslationInitiationTo explore the mechanism of derepression of miRNA-targeted
mRNAs in RPG knockdown cells, we first examined miRNA
biogenesis. We did not detect any defects in the biogenesis of
any miRNAs tested (Figures S3A and S3B). Additionally, RPG
knockdowns did not decrease the levels of Ago1 or Ago2
proteins (Figure S3C). Polysome profiling of D8 cells knocking
down S15 (Figures 3A and 3C) or L12 (Figures 3B and 3C)
demonstrated a strong shift of FL mRNA but not RL mRNA
from monosomes to polysomes, indicating that RPG knock-
downs relieve miRNA-mediated repression of translation initia-
tion. Consistent with knockdowns of miRNA pathway genes
(Figure 1D), RPG knockdowns increased the combined signal
for FL but not RL mRNA across gradients relative to the Scr
control, indicating preferential stabilization of FL mRNA. Heavy
complexes formed on miRNA-targeted mRNAs after RPG
depletion were actively translating polysomes because FL
mRNA shifted to lighter fractions after puromycin treatment
which dissociates only ribosomes actively translocating on
mRNAs (Figures S3D and S3E). Thus, reduced levels of RPGs
increased loading of active ribosomes onto miRNA-targeted FL
mRNA but not untargeted RL mRNA.
Importantly, the effects of RPG knockdowns on ribosomal
subunit levels were similar among all tested 40S and 60S
RPGs. Knockdown of 40S RPGs consistently decreased the
levels of free 40S subunits and 18S ribosomal RNA (rRNA) and
increased the levels of free 60S subunits (Figures 3A, S4A, and
S4B). On the other hand, knockdown of 60S RPGs or eIF6
consistently decreased the levels of free 60S subunits and 28S
rRNA (Figures 3B, S4A, and S4B). These data are consistent
with established contributions of 40S and 60S RPGs to biogen-
esis and/or stability of ribosomal subunits (Lempiainen and
Shore, 2009) and further indicate that defects in ribosome
biogenesis rather than extraribosomal functions of RPGs led to
reduced miRNA-mediated repression.
To resolve the 40S, 60S, and 80S peaks, polysome profiling
was performed with lower-density gradients, higher-speed
spins, and longer times, which resulted in reduced resolution
of the highest-density complexes (Figures 3A and 3B). These
conditions enabled detection of reduced 40S and 60S peaks
and thus verified efficient and functional RPG knockdowns. To
resolve the highest density polysomes and prevent pelletting
of the heaviest complexes, we performed polysome profiling
using higher-density gradients, lower-speed spins, and shorter
times. Under these conditions, RPG knockdowns led to a con-
sistent shift of FL but not RLmRNAs to polysomes (Figure S4C).
Together, these data show that RPG knockdowns inhibited
miRNA-mediated repression of translation initiation and that
miRNA-targeted mRNA derepression could be a result of
perturbed stoichiometry between 40S and 60S ribosomal
subunits, either directly through physical interactions with
miRNP complexes or indirectly through signaling pathways
that sense ribosome biogenesis.
(C and D) Subgenomic RNAi screen in D8 identifies almost all RPGs as hits in themiRNApathway. Small and large ribosomal subunitRPGswere knocked down in
D8 and the dual luciferase assay was performed after 72 hr. Each point represents an individual siRNA. RPGs included in the original screen are highlighted.
Dotted horizontal gray lines represent derepression of FL in the negative control (Scr) and the positive control (eIF6) transfections.
(E) Knockdown of RPGs derepresses endogenous HMGA2 at the mRNA level. HeLa was transfected with siRNAs against indicated genes, and after 72 hr,
endogenousHMGA2mRNA levels were assessed by RT-qPCR and normalized toGAPDHmRNA levels. Bar graphs show themean ± SD from three independent
experiments.
(F) Knockdown of RPGs derepresses endogenous miRNA targets at the protein level. HeLa was transfected with siRNAs against the indicated genes, and RAS,
PTEN, IMP-1, PDCD4, and GAPDH protein levels were assessed by western blotting after 72 hr.
See also Figure S2.
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc. 175
A
BScr L12
Scr S15
FL
RL
A254
40S 60S 80S Polysomes
FL
RL
28S
18S
A254
C
0
10
20
30
40
50
60
70
80
1 2 3 4 5 6 7 8 9 10Fraction number
% F
L m
RN
A
S15ScrL12
Scr
0
10
20
30
40
50
60
70
% R
L m
RN
A
Scr
S1
5
L1
2
Scr
S1
5
L1
2
SCR CXCR4
Input
35S-FLAG IP
E
S
Scr
Scr+
CH
X
S5
S11
S1
2
S1
5
S1
8
L5
L11
L1
2
L3
5A
LP
2
eIF
6
Ag
o1
Co
om
assie
35
FLAG-FL6X
35S-FLAG IP/Input 1.0 1.3 1.5 0.3 0.8 0.9
D
28S
18S
40S 60S 80S Polysomes
40S 60S 80S Polysomes 40S 60S 80S Polysomes
35S/Coomassie 1.0 0.2 1.1 1.1 1.2 1.2 1.0 0.9 0.7 0.9 0.9 0.9 1.1 1.1
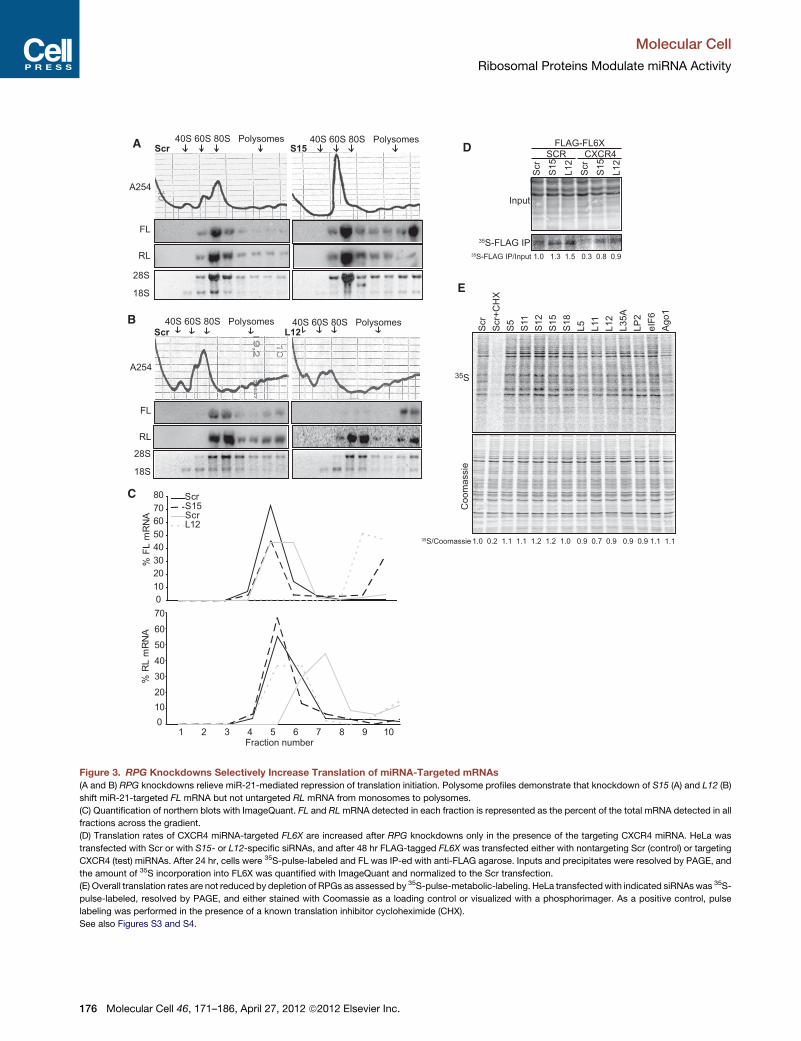
Figure 3. RPG Knockdowns Selectively Increase Translation of miRNA-Targeted mRNAs
(A and B) RPG knockdowns relieve miR-21-mediated repression of translation initiation. Polysome profiles demonstrate that knockdown of S15 (A) and L12 (B)
shift miR-21-targeted FL mRNA but not untargeted RL mRNA from monosomes to polysomes.
(C) Quantification of northern blots with ImageQuant. FL and RLmRNA detected in each fraction is represented as the percent of the total mRNA detected in all
fractions across the gradient.
(D) Translation rates of CXCR4 miRNA-targeted FL6X are increased after RPG knockdowns only in the presence of the targeting CXCR4 miRNA. HeLa was
transfected with Scr or with S15- or L12-specific siRNAs, and after 48 hr FLAG-tagged FL6X was transfected either with nontargeting Scr (control) or targeting
CXCR4 (test) miRNAs. After 24 hr, cells were 35S-pulse-labeled and FL was IP-ed with anti-FLAG agarose. Inputs and precipitates were resolved by PAGE, and
the amount of 35S incorporation into FL6X was quantified with ImageQuant and normalized to the Scr transfection.
(E) Overall translation rates are not reduced by depletion of RPGs as assessed by 35S-pulse-metabolic-labeling. HeLa transfectedwith indicated siRNAswas 35S-
pulse-labeled, resolved by PAGE, and either stained with Coomassie as a loading control or visualized with a phosphorimager. As a positive control, pulse
labeling was performed in the presence of a known translation inhibitor cycloheximide (CHX).
See also Figures S3 and S4.
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
176 Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc.
To further validate that RPG knockdowns increased transla-
tion rates of miRNA-targeted but not untargeted mRNAs, we
performed 35S pulse labeling followed by immunoprecipitation
(IP) of FLAG-tagged FL protein targeted by the CXCR4 miRNA.
AS
cr
S1
5
Scr
S1
5
Scr
S1
5
Scr
S1
5
FLAG-Ago1 FLAG-Ago2
Input IP
RN
A
Pro
tein
FLAG
Actin
FL
miR-21
let-7a
tRNA
B
Ago1
Ago2
Scr
S1
5
L1
2
eIF
6
Input
Scr
S1
5
L1
2
eIF
6
Ribosomal Pellet
S6
L7a
Actin
FLAG-Ago1 FLAG-Ago2
0
0.5
1
1.5
2
2.5
3
3.5
Fo
ldC
han
ge
FLmiR-21let-7a
0
0.2
0.4
0.6
0.8
1
1.2
1.4
0
1
2
3
Fo
ldC
han
ge
Ago1Ago2
0
1
2
3
4
*
*
**
*
** ** ** **
**
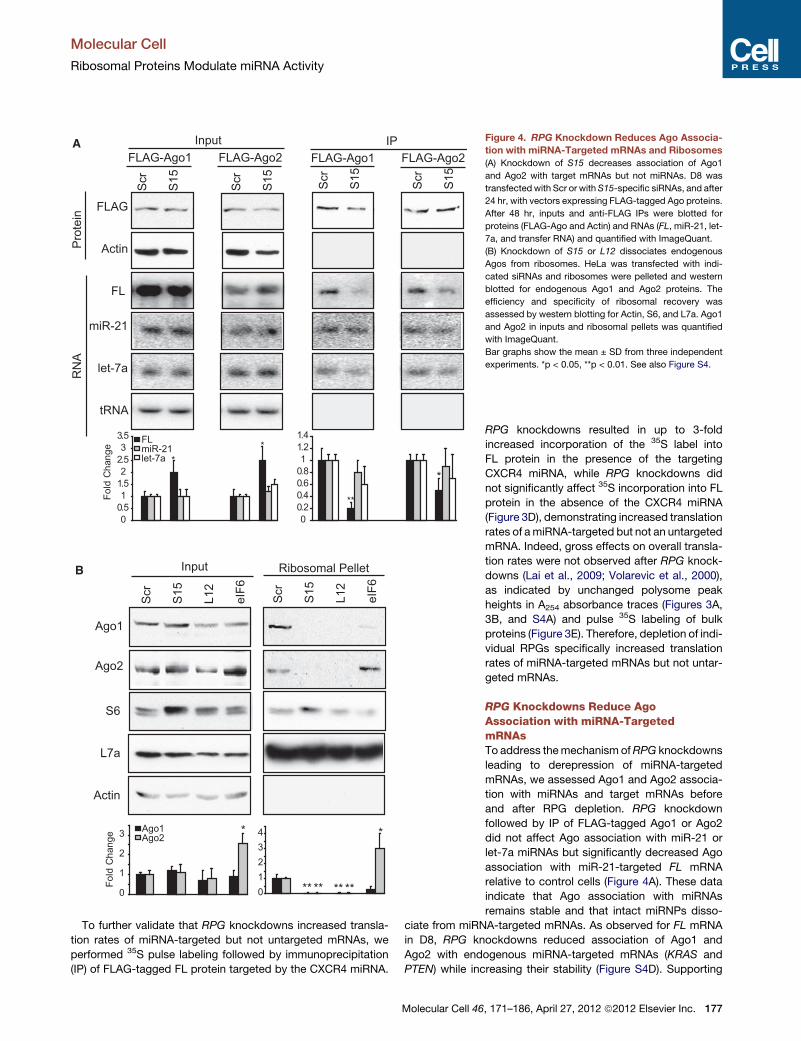
Figure 4. RPG Knockdown Reduces Ago Associa-
tion with miRNA-Targeted mRNAs and Ribosomes
(A) Knockdown of S15 decreases association of Ago1
and Ago2 with target mRNAs but not miRNAs. D8 was
transfectedwith Scr or withS15-specific siRNAs, and after
24 hr, with vectors expressing FLAG-tagged Ago proteins.
After 48 hr, inputs and anti-FLAG IPs were blotted for
proteins (FLAG-Ago and Actin) and RNAs (FL, miR-21, let-
7a, and transfer RNA) and quantified with ImageQuant.
(B) Knockdown of S15 or L12 dissociates endogenous
Agos from ribosomes. HeLa was transfected with indi-
cated siRNAs and ribosomes were pelleted and western
blotted for endogenous Ago1 and Ago2 proteins. The
efficiency and specificity of ribosomal recovery was
assessed by western blotting for Actin, S6, and L7a. Ago1
and Ago2 in inputs and ribosomal pellets was quantified
with ImageQuant.
Bar graphs show the mean ± SD from three independent
experiments. *p < 0.05, **p < 0.01. See also Figure S4.
RPG knockdowns resulted in up to 3-fold
increased incorporation of the 35S label into
FL protein in the presence of the targeting
CXCR4 miRNA, while RPG knockdowns did
not significantly affect 35S incorporation into FL
protein in the absence of the CXCR4 miRNA
(Figure 3D), demonstrating increased translation
rates of amiRNA-targeted but not an untargeted
mRNA. Indeed, gross effects on overall transla-
tion rates were not observed after RPG knock-
downs (Lai et al., 2009; Volarevic et al., 2000),
as indicated by unchanged polysome peak
heights in A254 absorbance traces (Figures 3A,
3B, and S4A) and pulse 35S labeling of bulk
proteins (Figure 3E). Therefore, depletion of indi-
vidual RPGs specifically increased translation
rates of miRNA-targeted mRNAs but not untar-
geted mRNAs.
RPG Knockdowns Reduce AgoAssociation with miRNA-TargetedmRNAsTo address themechanism ofRPG knockdowns
leading to derepression of miRNA-targeted
mRNAs, we assessed Ago1 and Ago2 associa-
tion with miRNAs and target mRNAs before
and after RPG depletion. RPG knockdown
followed by IP of FLAG-tagged Ago1 or Ago2
did not affect Ago association with miR-21 or
let-7a miRNAs but significantly decreased Ago
association with miR-21-targeted FL mRNA
relative to control cells (Figure 4A). These data
indicate that Ago association with miRNAs
remains stable and that intact miRNPs disso-
ciate from miRNA-targeted mRNAs. As observed for FL mRNA
in D8, RPG knockdowns reduced association of Ago1 and
Ago2 with endogenous miRNA-targeted mRNAs (KRAS and
PTEN) while increasing their stability (Figure S4D). Supporting
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc. 177
Polysome / Input RatioNo. of genes up-regulated > 2-fold
Polysome / Input RatioNo. of genes down-regulated > 2-fold
1.51.20.90.60.30.0-0.3-0.6-0.9-1.2-1.5
A
B
C
D
No. of poly
som
e-
up
-re
gu
late
d g
en
es
eIF6
L11
L12
S15
Ago1
Ago2
Ago3
Ago4
Ag
o4
Ag
o3
Ag
o2
Ag
o1
S1
5
L1
2
L11
eIF
6
Ag
o4
Ag
o3
Ag
o2
Ag
o1
S1
5
L1
2
L11
eIF
6
vs. S
cr1
vs. Scr2
log
2 (
Ob
se
rve
d/E
xp
ecte
d)
10
8
6
4
2
0
log
2 (
Ob
se
rve
d/E
xp
ecte
d)
2 3 4 5 6 7 8No. of knockdowns
1800
1600
1400
1200
1000
800
600
400
200
0
Observed
Expected
Observed/Expected
10
8
6
4
2
0
log
2 (
Ob
se
rve
d/E
xp
ecte
d)
No. of knockdowns
1800
1600
1400
1200
1000
800
600
400
200
0
Observed
Expected
Observed/Expected
Fraction of polysome up-regulated genes with
HeLa miRNA sites (conserved 8mers)
Observed
Expected
No
. o
f sh
uffle
d t
ria
ls 60
50
40
30
20
10
0
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35 0.40
Fraction of polysome down-regulated genes with
HeLa miRNA sites (conserved 8mers)
Observed
Expected
No
. o
f sh
uffle
d t
ria
ls
90
80
70
60
50
40
30
20
10
0
Observed
Expected
No. of knockdowns
0.35
0.30
0.25
0.20
0.15
0.10
No. of knockdowns
0.35
0.30
0.25
0.20
0.15
0.10
Observed
Expected
Fra
ction o
f poly
som
e
do
wn
-re
gu
late
d g
en
es w
ith
He
La
miR
NA
site
s
2 3 4 5 6 7 8
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35 0.40
2 3 4 5 6 7 8 2 3 4 5 6 7 8
No. of poly
som
e-
do
wn
-re
gu
late
d g
en
es
Fra
ction o
f poly
som
e
up
-re
gu
late
d g
en
es w
ith
He
La
miR
NA
site
s
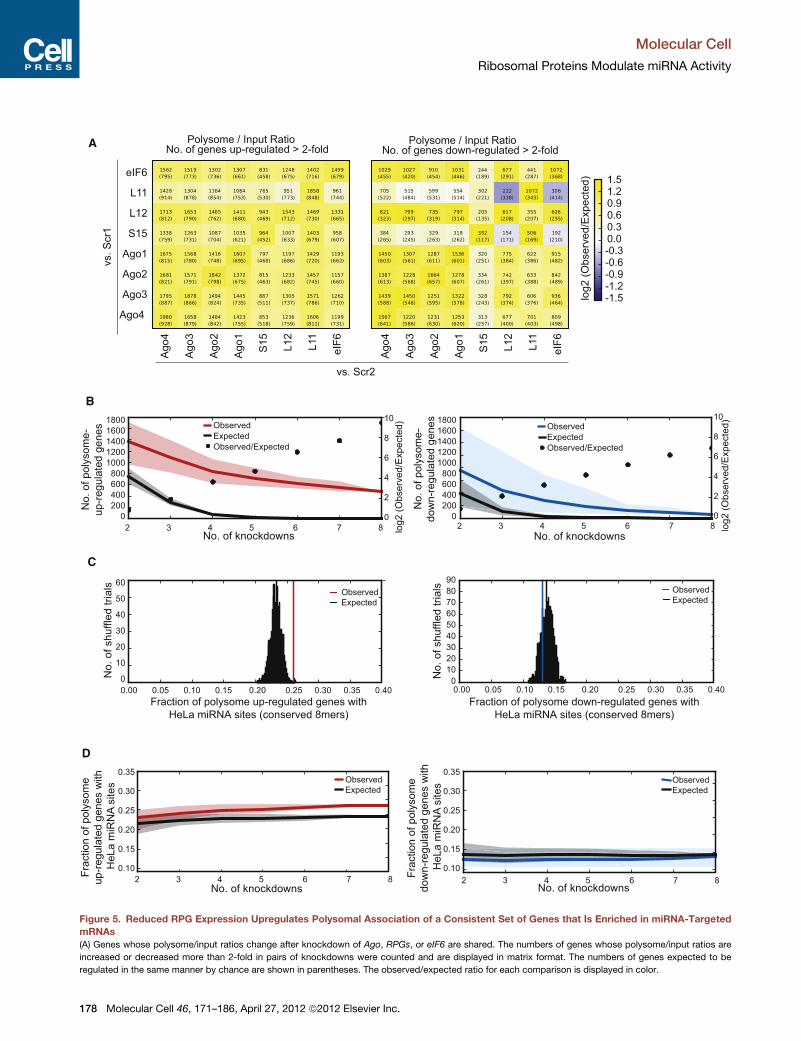
Figure 5. Reduced RPG Expression Upregulates Polysomal Association of a Consistent Set of Genes that Is Enriched in miRNA-Targeted
mRNAs
(A) Genes whose polysome/input ratios change after knockdown of Ago, RPGs, or eIF6 are shared. The numbers of genes whose polysome/input ratios are
increased or decreased more than 2-fold in pairs of knockdowns were counted and are displayed in matrix format. The numbers of genes expected to be
regulated in the same manner by chance are shown in parentheses. The observed/expected ratio for each comparison is displayed in color.
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
178 Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc.

the model of miRNP dissociation from miRNA-targeted mRNAs,
RPG depletions reduced Ago1 and Ago2 association with ribo-
somes (Figure 4B). Together, these data demonstrate that RPG
knockdown dissociates miRNPs from miRNA-targeted mRNAs
and does not affect miRNP assembly or stability.
RPG, eIF6, and Ago Knockdowns Alter PolysomeAssociation of Common mRNAsTo globally assess effects on translation of endogenous mRNAs,
we conducted RNA sequencing (RNA-Seq) analysis of polyso-
mal fractions and total RNA (‘‘input’’ to the gradient) before
and after knockdowns of RPGs, Agos, or eIF6. A remarkably
large number of common genes demonstrated a 2-fold or
greater increase in polysome association (polysome/input ratio)
after knockdown of RPGs, Agos, or eIF6 (Figure 5A). Many
common genes also demonstrated a 2-fold or greater decrease
in polysome association for many pairs of knockdowns, particu-
larly between knockdowns of differentAgos. These observations
suggested the surprising conclusion that large and small subunit
RPGs, Agos, and eIF6 have related effects on translational
regulation globally, perhaps through common regulatory path-
ways. This idea was further supported by analysis of mRNAs
impacted in three or more of these knockdowns. Instead of
decreasing rapidly to zero, as expected if the knockdowns
affected independent sets of genes, the numbers of mRNAs
with at least 2-fold increased polysome association remained
in the several hundred range, even as the number of compared
knockdowns increased from three to eight, with 479 mRNAs
exhibiting an increase in all eight knockdowns (Figure 5B and
Table S2, part A). A smaller but still highly significant set of 57
mRNAs with consistently decreased polysome association in
all knockdowns was also observed (Figure 5B and Table S2,
part B). Thus RPGs, Agos, and eIF6 inhibited polysome associ-
ation of a large common cohort of mRNAs, and promoted poly-
some association of a smaller common cohort of mRNAs.
miRNA Target Sites Are Enriched in Polysome-ShiftedmRNAsTo explore potential connections to the miRNA pathway, we
analyzed the fraction of mRNAs containing conserved 8-mer
seed matches to HeLa-expressed miRNAs, a relatively stringent
set of putative targets that likely excludesmany authentic targets
with weaker or less stringently conserved seed matches (Fried-
man et al., 2009). Significant enrichment of conserved target
sites relative to controls was observed in the set of mRNAs
with consistently increased polysomal association in all knock-
downs (Figure 5C). The extent of target site enrichment
increased as the number of intersected knockdowns increased
(Figure 5D). The observed enrichment for miRNA target sites in
this common set of polysome-increased mRNAs suggested
that miRNAs are involved in repression of this gene set via
a mechanism involving translation, and that the perturbations
introduced by RPG, Ago, and eIF6 knockdowns alter ribosome
loading onto a cohort of mRNAs that is strongly enriched
for miRNA-targeted mRNAs. Mechanisms involving miRNA-
directed changes in stability of polysome-associated versus
non-polysome-associatedmRNAs are also possible. In contrast,
the set of genes with consistently decreased polysomal associ-
ation in the knockdowns was not enriched for conserved miRNA
target sites (Figures 5C and 5D), suggesting that this set of
mRNAs is less often directly impacted by miRNAs. Although
RPG, Ago, and eIF6 knockdowns also altered mRNA levels of
a common cohort of genes, these sets of genes were not en-
riched for conserved 8-mer seed matches to HeLa-expressed
miRNAs (Figure S5). Together, these data show that RPGs,
Agos, and eIF6 contributed to reducing polysome association
of a common subset of mRNAs enriched for miRNA-targeted
mRNAs. Because miRNA-targeted mRNAs were not enriched
in a common subset of mRNAs exhibiting increased mRNA
levels after knockdowns, we observed translational derepres-
sion and not stabilization of miRNA-targeted mRNAs.
miRNAs Predominantly Cosediment with PolysomesThe preceding studies demonstrated that miRNA-targeted
mRNAs associate with monosomes and that RPG knockdowns
increase association of miRNA-targetedmRNAswith polysomes
without changing miRNA levels or miRNA association with Ago
proteins. To gain additional insights into RPG regulation of
miRNA-mediated repression, we performed global miRNA
profiling of total, monosomal, and polysomal RNA after sucrose
gradient fractionation of HeLa cells before and after RPG
knockdown. Out of 669 unique miRNA species expressed in
HeLa cells, 462 miRNAs (70%) were reliably detected in
monosomal and polysomal fractions at steady state (Table S3,
part A). The majority of miRNAs were equally abundant in
monosomal and polysomal fractions. Specifically, out of 462
miRNAs detected in monosomes and polysomes, 265 miRNAs
(57%) had no statistically significant enrichment in either frac-
tion. Thus, the majority of miRNAs are equally distributed across
monosomal and polysomal fractions and may contribute to
(B) A common set of mRNAs shifts to and away from polysomes in Ago, RPG, and eIF6 knockdowns. The numbers of genes whose polysome/input ratios are
increased or decreased more than 2-fold in two-way, three-way, . eight-way comparisons of knockdowns are displayed in red or blue, respectively. The
numbers of genes expected to be regulated in the same manner by chance are shown in black. Ninety percent confidence intervals are shown in pale red, blue,
and black.
(C) The upregulated genes have a significantly increased proportion of HeLa-expressed miRNA binding sites (p < 0.003). The fraction of genes with conserved 8-
mermiRNA sites was obtained for genes whose polysome expression is upregulated (red) or downregulated (blue) in all eight knockdowns (Ago1–Ago4, L11, L12,
S15, and eIF6). For assessment of significance, the same fraction was determined for 1,000 sets of control genes that matched the original up- or downregulated
sets in 7-mer conservation and UTR length (black histogram).
(D) The proportion of upregulated genes with HeLa-expressed miRNA-targeted mRNAs increases as the number of knockdowns used to define the set of
coregulated genes increases. The fraction of genes with HeLa-expressed conserved 8-mer miRNA sites was obtained for those genes whose polysome
expression is upregulated (red) or downregulated (blue) in two, three, . eight knockdowns. Control genes (black) were selected to match the up- or down-
regulated genes.
See also Figure S5.
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc. 179
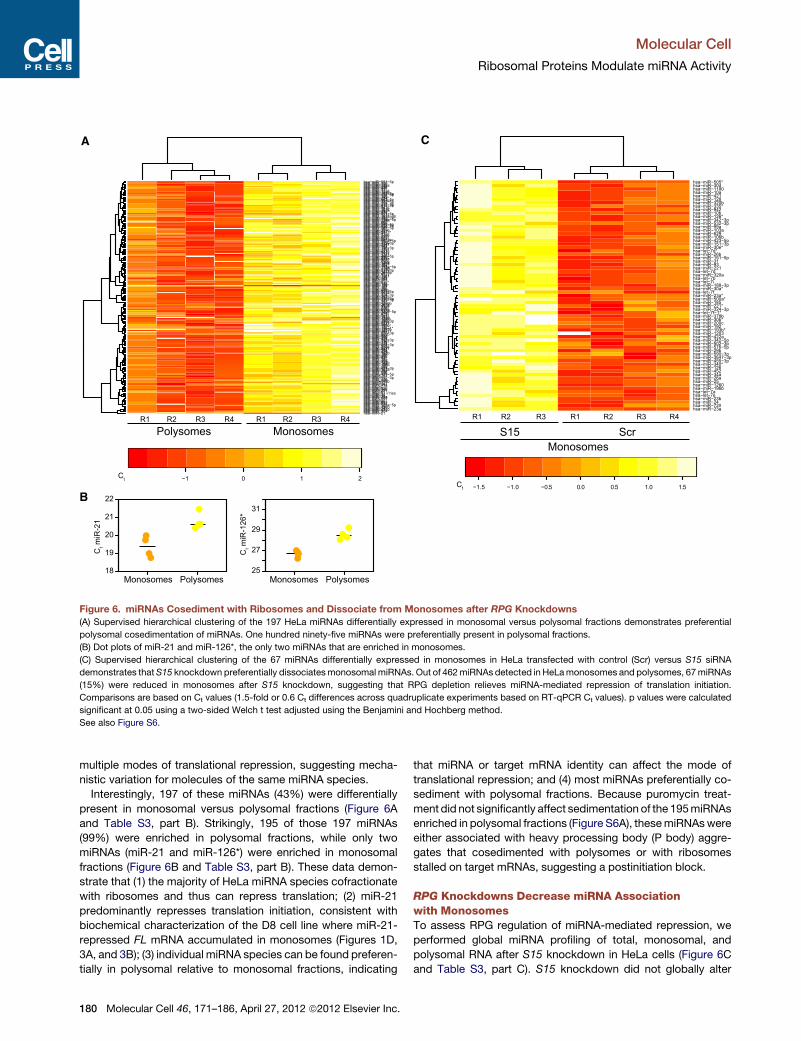
multiple modes of translational repression, suggesting mecha-
nistic variation for molecules of the same miRNA species.
Interestingly, 197 of these miRNAs (43%) were differentially
present in monosomal versus polysomal fractions (Figure 6A
and Table S3, part B). Strikingly, 195 of those 197 miRNAs
(99%) were enriched in polysomal fractions, while only two
miRNAs (miR-21 and miR-126*) were enriched in monosomal
fractions (Figure 6B and Table S3, part B). These data demon-
strate that (1) the majority of HeLa miRNA species cofractionate
with ribosomes and thus can repress translation; (2) miR-21
predominantly represses translation initiation, consistent with
biochemical characterization of the D8 cell line where miR-21-
repressed FL mRNA accumulated in monosomes (Figures 1D,
3A, and 3B); (3) individual miRNA species can be found preferen-
tially in polysomal relative to monosomal fractions, indicating
that miRNA or target mRNA identity can affect the mode of
translational repression; and (4) most miRNAs preferentially co-
sediment with polysomal fractions. Because puromycin treat-
ment did not significantly affect sedimentation of the 195miRNAs
enriched in polysomal fractions (Figure S6A), thesemiRNAswere
either associated with heavy processing body (P body) aggre-
gates that cosedimented with polysomes or with ribosomes
stalled on target mRNAs, suggesting a postinitiation block.
RPG Knockdowns Decrease miRNA Associationwith MonosomesTo assess RPG regulation of miRNA-mediated repression, we
performed global miRNA profiling of total, monosomal, and
polysomal RNA after S15 knockdown in HeLa cells (Figure 6C
and Table S3, part C). S15 knockdown did not globally alter
A
hsa−miR−21hsa−miR−7hsa−miR−92ahsa−miR−1280hsa−miR−24hsa−miR−1260hsa−miR−30chsa−miR−125a−5phsa−miR−224hsa−miR−25hsa−miR−26ahsa−let−7ghsa−miR−191hsa−miR−222hsa−miR−20ahsa−miR−16hsa−miR−31hsa−miR−17 // hsahsa−let−7dhsa−miR−221hsa−miR−365hsa−miR−92bhsa−let−7ihsa−miR−27ahsa−miR−454hsa−miR−197hsa−let−7bhsa−miR−148bhsa−miR−26bhsa−miR−27bhsa−miR−151−3phsa−let−7chsa−miR−182hsa−miR−423−5phsa−miR−19bhsa−miR−29chsa−miR−29ahsa−miR−103ahsa−miR−423−3phsa−miR−18ahsa−miR−20bhsa−miR−106bhsa−miR−19ahsa−miR−125bhsa−miR−30ahsa−miR−30ehsa−miR−195hsa−miR−93hsa−miR−99bhsa−miR−100hsa−miR−186hsa−miR−193bhsa−miR−425hsa−miR−34ahsa−miR−484hsa−miR−4291hsa−miR−4286hsa−miR−505hsa−miR−574−3phsa−miR−128hsa−miR−378hsa−miR−1307hsa−miR−342−3phsa−miR−424hsa−miR−30bhsa−miR−30dhsa−miR−99ahsa−miR−200bhsa−miR−876−3phsa−miR−16−1*hsa−miR−335hsa−miR−1255bhsa−miR−19b−1*hsa−miR−132*hsa−miR−1909*hsa−miR−589hsa−miR−1227hsa−miR−3916hsa−miR−296−3phsa−miR−196b*hsa−miR−1249hsa−miR−3909hsa−miR−183*hsa−miR−7−1*hsa−miR−425*hsa−miR−29a*hsa−miR−3200−5phsa−miR−4323hsa−miR−550ahsa−miR−27b*hsa−miR−33a*hsa−miR−450ahsa−miR−3618hsa−miR−1260bhsa−miR−22hsa−miR−18a*hsa−miR−339−5phsa−miR−330−3phsa−miR−4289hsa−miR−107hsa−miR−324−3phsa−miR−378*hsa−miR−301ahsa−miR−324−5phsa−miR−500ahsa−miR−29bhsa−miR−185hsa−miR−101hsa−miR−126*hsa−miR−10bhsa−miR−132hsa−miR−335*hsa−let−7a*hsa−miR−183hsa−miR−942hsa−miR−126hsa−miR−1281hsa−miR−1271hsa−miR−452hsa−miR−28−5phsa−miR−1233hsa−miR−532−3phsa−miR−1180hsa−miR−374ahsa−miR−193a−5phsa−miR−10ahsa−miR−3651hsa−miR−148ahsa−miR−130ahsa−miR−200chsa−miR−877*hsa−miR−422ahsa−miR−92b*hsa−miR−486−5phsa−miR−1287hsa−miR−3131hsa−miR−941hsa−miR−1226hsa−miR−221*hsa−miR−1301hsa−miR−140−3phsa−miR−194hsa−miR−629*hsa−miR−1247hsa−miR−455−3phsa−miR−130bhsa−miR−3607−5phsa−miR−28−3phsa−miR−20a*hsa−miR−675*hsa−miR−195*hsa−miR−378bhsa−miR−31*hsa−miR−24−2*hsa−miR−497hsa−miR−1296hsa−miR−96hsa−miR−188−5phsa−miR−920hsa−miR−502−5phsa−miR−140−5phsa−miR−660hsa−miR−662hsa−miR−1285hsa−miR−146b−5phsa−miR−340hsa−miR−361−3phsa−miR−193a−3phsa−miR−1231hsa−miR−491−5phsa−miR−874hsa−miR−99b*hsa−miR−181ahsa−miR−181chsa−miR−3676hsa−miR−22*hsa−miR−615−3phsa−miR−331−3phsa−miR−15ahsa−miR−330−5phsa−miR−301bhsa−miR−582−5phsa−miR−224*hsa−miR−149hsa−miR−105hsa−miR−532−5phsa−miR−362−5phsa−miR−135bhsa−miR−1238hsa−miR−17*hsa−miR−93*hsa−miR−326hsa−miR−596hsa−miR−135ahsa−miR−190hsa−miR−370hsa−miR−501−5p
−1 0 1 2
Polysomes Monosomes
R1 R2 R3 R4 R1 R2 R3 R4
Ct
B
18
19
20
21
22
Ct m
iR-2
1
25
27
29
31
Ct m
iR-1
26
*
C
hsa−miR−23ahsa−miR−720hsa−miR−24hsa−miR−23bhsa−let−7ehsa−let−7ahsa−miR−196bhsa−miR−1260hsa−miR−25hsa−miR−26ahsa−miR−34ahsa−miR−452hsa−miR−126hsa−miR−340*hsa−miR−455−3phsa−miR−3647−3phsa−miR−590−3phsa−miR−29bhsa−miR−576−5phsa−miR−502−3phsa−miR−342−5phsa−miR−422ahsa−miR−1203hsa−miR−193b*hsa−miR−192hsa−miR−200chsa−miR−30b*hsa−miR−378bhsa−let−7f−1*hsa−miR−324−3phsa−miR−221*hsa−miR−185hsa−miR−500a*hsa−miR−23a*hsa−let−7fhsa−miR−30a*hsa−miR−188−3phsa−let−7ihsa−let−7bhsa−miR−320ahsa−let−7dhsa−miR−221hsa−miR−93hsa−miR−31hsa−miR−151−5phsa−miR−26bhsa−let−7d*hsa−miR−30e*hsa−miR−151−3phsa−miR−423−5phsa−miR−106bhsa−miR−629hsa−miR−103ahsa−miR−99ahsa−miR−532−3phsa−miR−342−3phsa−miR−126*hsa−miR−10bhsa−miR−942hsa−miR−335*hsa−miR−320bhsa−miR−128hsa−miR−421hsa−miR−10ahsa−miR−1180hsa−miR−30dhsa−miR−505*
−1.5 −1.0 −0.5 0.0 0.5 1.0 1.5Ct
S15 Scr
R1 R2 R3 R1 R2 R3 R4
Monosomes
Monosomes Polysomes Monosomes Polysomes
Figure 6. miRNAs Cosediment with Ribosomes and Dissociate from Monosomes after RPG Knockdowns
(A) Supervised hierarchical clustering of the 197 HeLa miRNAs differentially expressed in monosomal versus polysomal fractions demonstrates preferential
polysomal cosedimentation of miRNAs. One hundred ninety-five miRNAs were preferentially present in polysomal fractions.
(B) Dot plots of miR-21 and miR-126*, the only two miRNAs that are enriched in monosomes.
(C) Supervised hierarchical clustering of the 67 miRNAs differentially expressed in monosomes in HeLa transfected with control (Scr) versus S15 siRNA
demonstrates thatS15 knockdown preferentially dissociatesmonosomal miRNAs. Out of 462miRNAs detected in HeLamonosomes and polysomes, 67miRNAs
(15%) were reduced in monosomes after S15 knockdown, suggesting that RPG depletion relieves miRNA-mediated repression of translation initiation.
Comparisons are based on Ct values (1.5-fold or 0.6 Ct differences across quadruplicate experiments based on RT-qPCR Ct values). p values were calculated
significant at 0.05 using a two-sided Welch t test adjusted using the Benjamini and Hochberg method.
See also Figure S6.
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
180 Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc.
miRNA levels (Figure S6B), consistent with results from northern
blotting experiments (Figure S3B). S15 knockdown also did not
affect miRNA abundance in polysomes, where the average
mean fold change (0.99 with a range 0.95–1.06) was not statisti-
cally significant according to the p values adjusted for multiple
comparisons. These data indicate that RPG depletion does not
affect miRNA-mediated repression of translation after initiation
or the abundance of miRNAs in P bodies.
In contrast, S15 knockdown reduced monosome association
of 67 out of 462 (15%) miRNAs detected in monosomes
and polysomes, demonstrating that RPGs regulate miRNA-
mediated repression of translation initiation (Figure 6C). Impor-
tantly, all 67 of these miRNAs were reduced in monosomes after
S15 knockdown and none of these miRNAs were increasingly
associated with polysomes. These data suggest that miRNPs
containing these miRNAs dissociated from target mRNAs,
consistent with reduced association of miRNA-targeted
mRNAs with Agos after RPG knockdowns (Figure 4A, 4B,
and S4D). Notably, multiple let-7 family members were among
the 67 miRNAs with reduced expression in monosomes
after S15 knockdown, consistent with derepression of validated
let-7 targets (Figures 2E and 2F). miR-21 was consistently
reduced in monosomes after S15 knockdown (average 3.5-
fold decrease) and was not affected in polysomes (average
1.1-fold increase). However, we note that the decreased
association of miR-21 with monosomes was not considered
AAAAAAA
60S
40S
miRNP
AUG
miRNP
AAAAAAA
60S
40S
60S60S
40S 40S
p53/63/73 p53/63/73
Normal Nucleolar Stress
pre-40S
pre-60S
A
0
0.5
1
1.5
2
2.5
Scr S5 S15 L11 L12
Fo
ld D
ere
pre
ssio
n
Scrp53
p53
GAPDH
Scr p53
Scr
S5
S1
5
L11
L1
2
Scr
S5
S1
5
L11
L1
2
0
0.5
1
1.5
2
2.5
3
3.5
Fold
Change
FLRL
B
p53
GAPDH
DM
SO
0.0
01
0.0
02
5.0
10
ActD DRB
DMSO 0.0005 0.001 0.002 2.5 5.0 10
ActD DRB
C
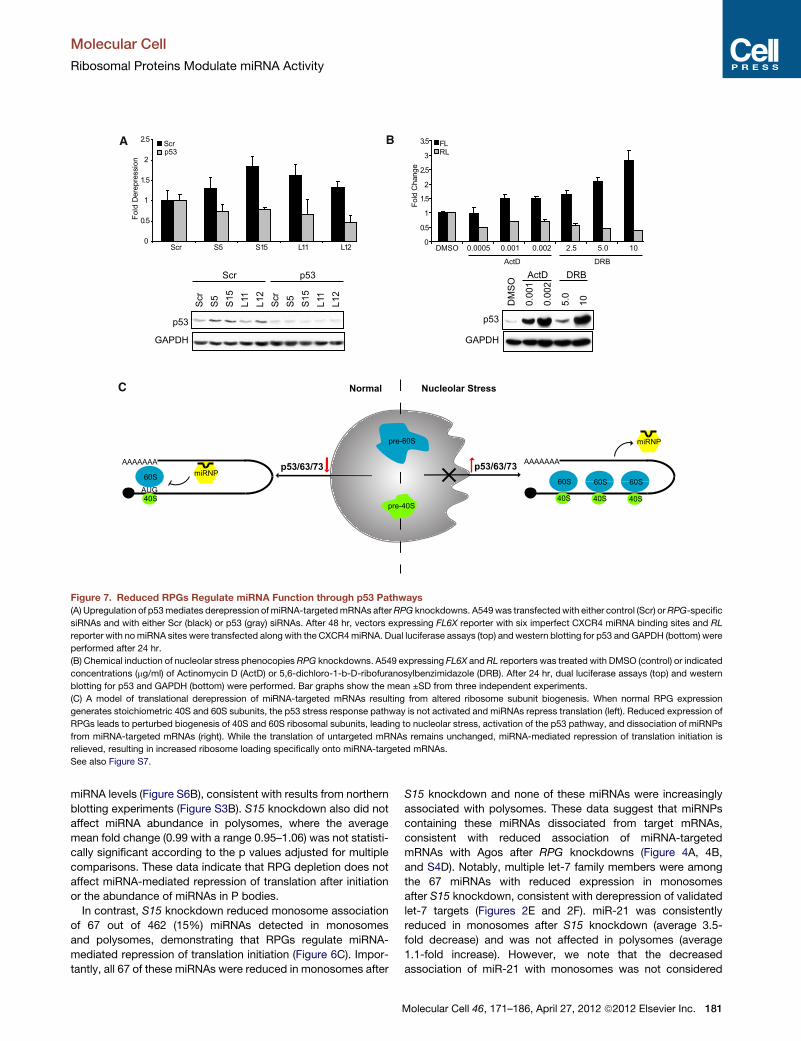
Figure 7. Reduced RPGs Regulate miRNA Function through p53 Pathways
(A) Upregulation of p53mediates derepression ofmiRNA-targetedmRNAs afterRPG knockdowns. A549was transfected with either control (Scr) orRPG-specific
siRNAs and with either Scr (black) or p53 (gray) siRNAs. After 48 hr, vectors expressing FL6X reporter with six imperfect CXCR4 miRNA binding sites and RL
reporter with nomiRNA sites were transfected along with the CXCR4miRNA. Dual luciferase assays (top) and western blotting for p53 and GAPDH (bottom) were
performed after 24 hr.
(B) Chemical induction of nucleolar stress phenocopies RPG knockdowns. A549 expressing FL6X and RL reporters was treated with DMSO (control) or indicated
concentrations (mg/ml) of Actinomycin D (ActD) or 5,6-dichloro-1-b-D-ribofuranosylbenzimidazole (DRB). After 24 hr, dual luciferase assays (top) and western
blotting for p53 and GAPDH (bottom) were performed. Bar graphs show the mean ±SD from three independent experiments.
(C) A model of translational derepression of miRNA-targeted mRNAs resulting from altered ribosome subunit biogenesis. When normal RPG expression
generates stoichiometric 40S and 60S subunits, the p53 stress response pathway is not activated and miRNAs repress translation (left). Reduced expression of
RPGs leads to perturbed biogenesis of 40S and 60S ribosomal subunits, leading to nucleolar stress, activation of the p53 pathway, and dissociation of miRNPs
from miRNA-targeted mRNAs (right). While the translation of untargeted mRNAs remains unchanged, miRNA-mediated repression of translation initiation is
relieved, resulting in increased ribosome loading specifically onto miRNA-targeted mRNAs.
See also Figure S7.
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc. 181

statistically significant using strict limits (the raw p value for Scr
versus S15 siRNA was 0.03, but the FDR p value [q value] was
0.07).
To determine whether dissociation of the 67 miRNAs from
monosomes after S15 knockdown affected repression, we per-
formed miRNA target prediction analysis using TargetScan on
the set of 479 mRNAs that commonly shift toward polysomes
after RPG, Ago, or eIF6 knockdowns as assessed by RNA-Seq
(Figure 5B and Table S2, part A). Only favorable targets, defined
asmRNAswith TargetScan context score in the top two quartiles
for each miRNA, were considered. We observed a highly sig-
nificant enrichment of mRNA targets of those 67 miRNAs (Chi-
square value of 103.107; p value of 1.973 10�24) amongmRNAs
that shifted to polysomes following RPG knockdowns (Table S3,
part D). These data indicate that RPG knockdowns relieve
repression of translation initiation by decreasing miRNP associ-
ation and increasing ribosome association of these miRNA-
targeted mRNAs.
p53 Pathway Activation Relieves miRNA-MediatedRepressionGlobal analysis of transcriptome changes (Figure S5) identified
robust effects on p53 pathway genes after RPG (S15, L11, and
L12) but not Ago knockdowns (Figures S7A and S7B and Table
S4). Specifically, gene network analysis of mRNAs commonly
dysregulated in RPG knockdowns revealed enrichment for
a DNA repair network (p value of 6.2 3 10�41; Figure S7B) and
a cell-cycle network (p value of 8.1 10�28; Figure S7B). Because
perturbation of ribosome subunit biogenesis induces nucleolar
stress and leads to p53 pathway activation (Deisenroth and
Zhang, 2010; Holzel et al., 2010; Pani�c et al., 2007; Rudra and
Warner, 2004; Volarevic et al., 2000), we hypothesized that
reduced RPG expression led to p53 pathway activation, which,
in turn, reduced repression of translation initiation of miRNA-
targeted mRNAs. Notably, the human papillomavirus 16/18 E6
protein degrades endogenous p53 in HeLa cells (Scheffner
et al., 1990; Werness et al., 1990). Thus, RPG depletion activates
p53 pathway genes even in the absence of the p53 protein,
possibly by activating other p53 family members (such as p63
and p73).
To test the hypothesis that reduced RPG expression induces
p53 pathways that inhibit miRNA-mediated repression, we
used the A549 human lung cancer cell line which expresses
wild-type p53. RPG knockdowns upregulated p53 at the protein
level (Figures 7A and S7C), suggesting induction of the nucleolar
stress response. Importantly, simultaneous knockdown of RPG
and p53 reversed derepression of miRNA-targeted mRNAs
(Figure 7A), directly implicating the p53 pathway in RPG regula-
tion of miRNA activity.
To test the effects of nucleolar stress on miRNA activity
independently of RPG knockdowns, we chemically induced
nucleolar stress with low concentrations of Actinomycin
D (ActD) or 5,6-dichloro-1-b-D-ribofuranosylbenzimidazole
(DRB) (David-Pfeuty et al., 2001; Holzel et al., 2010). Similar to
RPG knockdowns, ActD andDRB treatments led to dose-depen-
dent p53 activation, reduced cell numbers, and specifically
increased the expression of miRNA-targeted mRNAs without
affecting miRNA levels (Figure 7B and S7D). Derepression of
CXCR4 miRNA-targeted FL mRNA was observed at 8 hr after
treatment (Figure S7E), suggesting that decreased miRNA
activity is a direct result of nucleolar stress induction and not an
indirect result of altered cell metabolism. Taken together, our
data demonstrate that induction of nucleolar stress (triggered
by RPG knockdowns or by small molecules) activates the p53
pathway, which leads to decreased miRNA activity (Figure 7C).
DISCUSSION
RPGs Regulate miRNA-Mediated Repressionof Translation InitiationAn RNAi screen for effectors and regulators of miRNA function
identified an unexpected role for RPGs in regulating miRNA-
mediated repression of translation initiation. Previous reports
have implicated RPGs in small RNA pathways in other organ-
isms, suggesting RPG regulation of small RNA pathways may
be conserved across phyla. In Drosophila, 11 RPGs scored in
a screen of small RNA pathways, constituting 6% of all 177
hits (Zhou et al., 2008). However, RPG knockdown did not affect
miRNA-mediated repression but instead increased endo-siRNA-
mediated repression. Depending upon the RPG tested, siRNA-
mediated repression increased in some cases but decreased
in others. In C. elegans, a genome-wide RNAi screen for miRNA
pathway genes identified one RPG, though this hit was not
confirmed (Parry et al., 2007). In contrast to these model organ-
isms, our data functionally implicates RPGs (as a class) as regu-
lators of miRNA-mediated repression of translation initiation in
human cells.
We showed that RPG depletion reduced the association of 67
miRNAs with monosomes without increasingmiRNA association
with polysomes or reducing total miRNA levels. Importantly,
RPG depletion specifically increased polysomal association
of mRNAs targeted by these miRNAs. Together, these high-
throughput data independently confirm the biochemical data
and support a model in which RPG depletion inhibits repression
of translation initiation mediated by miRNAs. Moreover, we
propose that parallel miRNA and mRNA expression profiling
from monosomes and polysomes could be an accurate method
of target mRNA identification.
miRNAs have been shown to activate translation under certain
stress conditions. Steitz and colleagues reported that cells
forced into quiescence can switch miRNA activity from transla-
tional repression to translational activation (Vasudevan et al.,
2007). It is important to note that RPG knockdowns did not acti-
vate translation but rather derepressed miRNA-targeted
mRNAs. In D8, it was impossible to distinguish derepression
from activation because FL was constitutively repressed by
endogenous miR-21, and thus the expression level of unre-
pressed FL was unknown. Therefore, we knocked down RPGs
in HeLa cells transiently transfected with luciferase reporters
with imperfect binding sites to the artificial CXCR4 miRNA
(Figures S2H and S2I). The expression of unrepressed luciferase
(in the absence of CXCR4 miRNA) was consistently higher than
the expression of derepressed luciferase (in the presence of
CXCR4 miRNA) after RPG knockdowns. These data demon-
strate that RPG depletion derepressed but did not activate trans-
lation of miRNA-targeted mRNAs.
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
182 Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc.

p53 Rathways Regulate miRNA FunctionRPG and eIF6 knockdowns led to substantial changes in abun-
dance of ribosomal subunits. The p53 pathway and nucleolar
stress have been implicated in sensing perturbed ribosomal
subunit stoichiometry, leading to cell-cycle arrest (Bachand
et al., 2006; Deisenroth and Zhang, 2010; Holzel et al., 2010;
Pani�c et al., 2007; Rudra and Warner, 2004; Volarevic et al.,
2000). Interestingly, gene ontology analysis of transcriptional
changes revealed that cell cycle, DNA replication, and p53
signaling were affected in all RPG knockdowns but not Ago
knockdowns (Figures S7A and S7B and Table S4). Although
the connection between ribosomal subunit imbalance, nucleolar
stress, and the p53 pathway activation was already known, the
connection between ribosomal subunit imbalance and global
miRNA activity has not been established prior to this study.
We propose that global reduction in miRNA-mediated repres-
sion may be an adaptive response allowing cells to increase
translation of subsets of mRNAs in response to nucleolar stress.
miRNA-mediated repression has been linked to oxidative-,
endoplasmic reticulum-, and nutrient deprivation-stress re-
sponses (Bhattacharyya et al., 2006a, 2006b). In contrast to
nucleolar stress, these stress responses do not induce the
p53 pathway and are distinguished by their effects on general
translation. For instance, amino acid starvation increases eIF2a
phosphorylation and leads to global translation inhibition (Bhat-
tacharyya et al., 2006a, 2006b), which we did not observe in
RPG knockdowns (Figure S6C). Amino acid starvation also
does not affect miRNA activity unless an AU-rich element is
present. In contrast, RPG knockdowns derepress both reporter
and endogenous miRNA-targeted mRNAs even in the absence
of AU-rich elements.
TheMajority of miRNA Species Can Repress TranslationPrevious studies have identified miRNA-targeted mRNAs cose-
dimenting with polysomes suggesting repression of translation
postinitiation (Kim et al., 2004; Maroney et al., 2006; Nelson
et al., 2004; Nottrott et al., 2006; Olsen and Ambros, 1999;
Petersen et al., 2006; Seggerson et al., 2002). In agreement
with more recent studies (Bhattacharyya et al., 2006a; Pillai
et al., 2005), we detected miR-21-targeted FL mRNAs predom-
inantly in monosomes, suggesting repression of translation
initiation. There has been much speculation about the causes
and implications of these different results (reviewed in Filipowicz
et al., 2008). One possible explanation may be the efficiency of
repression. Less efficient repression of translation initiation
may result in a larger population of miRNA-targeted mRNAs in
polysomes, whereas more efficient repression of translation
initiation might result in a larger population of miRNA-targeted
mRNAs in monosomes. In D8, where FL is repressed by miR-
21 via six target sites, strong repression and monosome
association was observed.
We report global miRNA expression profiles from HeLa
monosomal and polysomal fractions. Detection of all miRNAs
in both monosomal and polysomal fractions suggests that the
mechanisms of miRNA-mediated translational repression are
not uniform. Differential sedimentation of particular miRNAs
implies that miRNA and/or mRNA identity may affect the mech-
anism of translational repression. miRNAs cosedimenting with
polysomes did not appear to associate with actively translating
ribosomes. Puromycin treatment reduced polysomes (Fig-
ure S3D) but did not affect sedimentation of these miRNAs (Fig-
ure S6A), suggesting that miRNAs may associate with stalled
ribosomes or non-translating multimegadalton complexes
(e.g., P bodies).
eIF6 Indirectly Affects the Human miRNA PathwayOur screen identified eIF6, a eukaryotic translation initiation
factor that has been implicated in growth and transformation
(Gandin et al., 2008). eIF6 binds to 60S subunits and functions
as an antiassociation factor by preventing 60S subunits from
joining with 40S subunits (Ceci et al., 2003). eIF6 has also been
implicated in miRNA-mediated translational repression in worms
and human cells (Chendrimada et al., 2007). Human eIF6 has
been shown to associate with Agos, miRNAs, and 60S subunits
(Chendrimada et al., 2007). In contrast to these observations, we
did not detect any interaction between eIF6 and Agos, miRNAs,
or miRNA-targeted mRNAs (data not shown). Indeed, we did not
detect any association of eIF6 with Agos even by the highly
sensitive Multidimensional Protein Identification Technology
analysis (data not shown). Our data suggest that eIF6 may affect
miRNA-targeted mRNA repression indirectly by altering ribo-
some subunit stoichiometry. eIF6 is required for 18S and 5.8S
rRNA maturation which, in turn, is required for generating the
60S ribosomal subunit (Basu et al., 2001). Indeed, depletion of
eIF6 reduced 60S ribosomal subunits (Figure 1D), the same
phenotype as in 60S RPG knockdowns.
Reduced RPGs in Ribosomopathies and CancersCancer pathway genes were significantly enriched in polysomes
after RPG, Ago, and eIF6 knockdowns (Table S4). Downregula-
tion ofRPGs has been identified in precancerous states, cancers
in situ, and metastatic cancers (van Riggelen et al., 2010). Addi-
tionally, several genetic diseases that predispose patients to
cancers (Avondo et al., 2009; Campagnoli et al., 2008; Gazda
et al., 2008) are characterized by mutations in or reduced
expression of RPGs, including dyskeratosis congenita (DKC1),
cartilage-hair hypoplasia (RMRP), Shwachman-Diamond
syndrome (SBDS), Turner syndrome (S4X), Noonan syndrome
(L6), Camurati-Englemann disease (S18), and, most notably,
Diamond-Blackfan anemia (S7, S15, S17, S19, S24, S27A, L5,
L11, L35A, and L36). Interestingly, decreased expression of
individual RPGs in zebrafish (Amsterdam et al., 2004; Lai et al.,
2009; MacInnes et al., 2008) and flies (Stewart and Denell,
1993; Watson et al., 1992) promotes tumorigenesis, suggesting
that RPGs may act as haploinsufficient tumor suppressors.
This relationship is counterintuitive because rapid growth and
proliferation of tumors must require robust translational activity.
Our data offer a possible resolution in that reduced levels of
RPGs may preferentially derepress protein production from a
cohort of miRNA-targeted messages, many of which contribute
to cellular proliferation and oncogenesis.
EXPERIMENTAL PROCEDURES
Detailed experimental procedures can be found in the Supplemental Experi-
mental Procedures.
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc. 183

Polysome Profiling
HeLa cells were incubated with 100 mg/ml cycloheximide (CHX) for 5 min at
37�C and washed on ice twice with 5 ml cold PBS containing 100 ug/ml
CHX. Cells were scraped in 500 ml lysis buffer (15 mM Tris [pH 7.4], 15 mM
MgCl2, 150 mM NaCl, 1% Triton X-100, 100 mg/ml CHX, and 1 mg/ml heparin)
and centrifuged at 12,000 g for 5 min at 4�C. Supernatant was loaded onto
12 ml 4.5%–45% sucrose gradients in 15 mM Tris (pH 7.4), 15 mM MgCl2,
150mMNaCl, and 100 mg/ml CHX. Gradients were centrifuged in SW41Ti rotor
at 39,000 rpm for 2.5 hr at 4�C and 1 ml fractions were collected. RNA was
extracted with Trizol LS (Invitrogen) or by addition of 750 ml guanidinium
hydrochloride and 800 ml isopropanol to 500 ml of a fraction and incubation
at �20�C overnight. Samples were centrifuged at 10,000 rpm for 25 min at
4�C, pellets washed with 70% ethanol, and resuspended in 180 ml TE buffer.
After addition of 20 ml 3 M sodium acetate (pH 5.2) and 600 ml 100% ethanol,
RNA was precipitated at �80�C overnight. Samples were centrifuged at
10,000 rpm for 25 min at 4�C, pellets washed with 70% ethanol, and RNA
was resuspended in water.
RNA-Seq Read Mapping and RPKM Calculation
Short reads were mapped to the human genome and a precomputed set of
splice junctions using Bowtie (Langmead et al., 2009). Reads per kilobase of
exon model per million mapped reads (RPKM) for each Entrez gene was
computed by using all reads mapping to constitutive Refseq exons.
Global miRNA Expression Profiling
RNA from unfractionated HeLa cells and from fractionated monosomes and
polysomes was extracted with Trizol LS (Invitrogen). Complementary DNA
(cDNA) synthesis, cDNA preamplification, and real-time PCR were performed
with the miScript PCR System (QIAGEN). miRNA Ct measures were compared
between monosomes and polysomes over all samples and separately for Scr
and S15 knockdown groups using a two-sided Welch t test. p values were
adjusted with the method of Benjamini and Hochberg and considered sig-
nificant at the 0.05 level. Supervised hierarchical clustering was performed
based on the Euclidean distance function with the complete linkage agglom-
eration method for the miRNAs found to be differentially expressed between
groups. Only comparisons based on Ct were used, rather than DCt, because
expression of the 5S control varies between monosomes and polysomes,
and is altered in S15 knockdown.
SUPPLEMENTAL INFORMATION
Supplemental Information includes four tables, Supplemental Experimental
Procedures, and seven figures and can be found with this article online at
doi:10.1016/j.molcel.2012.04.008.
ACKNOWLEDGMENTS
Wewould like to thank Dr. Steffen Schubert and The Institute of Chemistry and
Cell Biology (ICCB) Screening Facility at Harvard Medical School for helping
with designing and performing the RNAi screen. We also thank Dr. Stephen
Buratowski, Dr. Danesh Moazed, Dr. Kai Wucherpfennig, and Dustin Grie-
semer for insightful discussions. This work was supported by a Distinguished
Young Scholars Award from the W.M. Keck Foundation to C.D.N.
Received: March 17, 2011
Revised: February 16, 2012
Accepted: April 5, 2012
Published online: April 26, 2012
REFERENCES
Amsterdam, A., Sadler, K.C., Lai, K., Farrington, S., Bronson, R.T., Lees, J.A.,
and Hopkins, N. (2004). Many ribosomal protein genes are cancer genes in
zebrafish. PLoS Biol. 2, E139.
Asangani, I.A., Rasheed, S.A., Nikolova, D.A., Leupold, J.H., Colburn, N.H.,
Post, S., and Allgayer, H. (2008). MicroRNA-21 (miR-21) post-transcriptionally
downregulates tumor suppressor Pdcd4 and stimulates invasion, intravasa-
tion and metastasis in colorectal cancer. Oncogene 27, 2128–2136.
Avondo, F., Roncaglia, P., Crescenzio, N., Krmac, H., Garelli, E., Armiraglio,
M., Castagnoli, C., Campagnoli, M.F., Ramenghi, U., Gustincich, S., et al.
(2009). Fibroblasts from patients with Diamond-Blackfan anaemia show
abnormal expression of genes involved in protein synthesis, amino acid
metabolism and cancer. BMC Genomics 10, 442.
Bachand, F., Lackner, D.H., Bahler, J., and Silver, P.A. (2006). Autoregulation
of ribosome biosynthesis by a translational response in fission yeast. Mol. Cell.
Biol. 26, 1731–1742.
Bagga, S., Bracht, J., Hunter, S., Massirer, K., Holtz, J., Eachus, R., and
Pasquinelli, A.E. (2005). Regulation by let-7 and lin-4 miRNAs results in target
mRNA degradation. Cell 122, 553–563.
Basu, U., Si, K., Warner, J.R., and Maitra, U. (2001). The Saccharomyces
cerevisiae TIF6 gene encoding translation initiation factor 6 is required for
60S ribosomal subunit biogenesis. Mol. Cell. Biol. 21, 1453–1462.
Bazzini, A.A., Lee, M.T., and Giraldez, A.J. (2012). Ribosome Profiling Shows
That miR-430 Reduces Translation Before Causing mRNA Decay in
Zebrafish. Science, in press. Published online March 15, 2012.
Behm-Ansmant, I., Rehwinkel, J., Doerks, T., Stark, A., Bork, P., and
Izaurralde, E. (2006). mRNA degradation by miRNAs and GW182 requires
both CCR4:NOT deadenylase and DCP1:DCP2 decapping complexes.
Genes Dev. 20, 1885–1898.
Bhattacharyya, S.N., Habermacher, R., Martine, U., Closs, E.I., and Filipowicz,
W. (2006a). Relief of microRNA-mediated translational repression in human
cells subjected to stress. Cell 125, 1111–1124.
Bhattacharyya, S.N., Habermacher, R., Martine, U., Closs, E.I., and Filipowicz,
W. (2006b). Stress-induced reversal of microRNA repression and mRNA
P-body localization in human cells. Cold Spring Harb. Symp. Quant. Biol. 71,
513–521.
Boyerinas, B., Park, S.M., Shomron, N., Hedegaard, M.M., Vinther, J.,
Andersen, J.S., Feig, C., Xu, J., Burge, C.B., and Peter, M.E. (2008).
Identification of let-7-regulated oncofetal genes. Cancer Res. 68, 2587–
2591.
Campagnoli, M.F., Ramenghi, U., Armiraglio, M., Quarello, P., Garelli, E.,
Carando, A., Avondo, F., Pavesi, E., Fribourg, S., Gleizes, P.E., et al. (2008).
RPS19 mutations in patients with Diamond-Blackfan anemia. Hum. Mutat.
29, 911–920.
Ceci, M., Gaviraghi, C., Gorrini, C., Sala, L.A., Offenhauser, N., Marchisio, P.C.,
and Biffo, S. (2003). Release of eIF6 (p27BBP) from the 60S subunit allows 80S
ribosome assembly. Nature 426, 579–584.
Chendrimada, T.P., Finn, K.J., Ji, X., Baillat, D., Gregory, R.I., Liebhaber, S.A.,
Pasquinelli, A.E., and Shiekhattar, R. (2007). MicroRNA silencing through
RISC recruitment of eIF6. Nature 447, 823–828.
David-Pfeuty, T., Nouvian-Dooghe, Y., Sirri, V., Roussel, P., and Hernandez-
Verdun, D. (2001). Common and reversible regulation of wild-type p53 function
and of ribosomal biogenesis by protein kinases in human cells. Oncogene 20,
5951–5963.
Deisenroth, C., and Zhang, Y. (2010). Ribosome biogenesis surveillance:
probing the ribosomal protein-Mdm2-p53 pathway. Oncogene 29, 4253–
4260.
Dorner, S., Lum, L., Kim, M., Paro, R., Beachy, P.A., and Green, R. (2006). A
genomewide screen for components of the RNAi pathway in Drosophila
cultured cells. Proc. Natl. Acad. Sci. USA 103, 11880–11885.
Eulalio, A., Rehwinkel, J., Stricker, M., Huntzinger, E., Yang, S.F., Doerks, T.,
Dorner, S., Bork, P., Boutros, M., and Izaurralde, E. (2007). Target-specific
requirements for enhancers of decapping in miRNA-mediated gene silencing.
Genes Dev. 21, 2558–2570.
Filipowicz, W., Bhattacharyya, S.N., and Sonenberg, N. (2008). Mechanisms of
post-transcriptional regulation by microRNAs: are the answers in sight? Nat.
Rev. Genet. 9, 102–114.
Flygare, J., Kiefer, T., Miyake, K., Utsugisawa, T., Hamaguchi, I., Da Costa, L.,
Richter, J., Davey, E.J., Matsson, H., Dahl, N., et al. (2005). Deficiency of
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
184 Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc.

ribosomal protein S19 in CD34+ cells generated by siRNA blocks erythroid
development and mimics defects seen in Diamond-Blackfan anemia. Blood
105, 4627–4634.
Friedman, R.C., Farh, K.K., Burge, C.B., and Bartel, D.P. (2009). Most
mammalian mRNAs are conserved targets of microRNAs. Genome Res. 19,
92–105.
Gandin, V., Miluzio, A., Barbieri, A.M., Beugnet, A., Kiyokawa, H., Marchisio,
P.C., and Biffo, S. (2008). Eukaryotic initiation factor 6 is rate-limiting in
translation, growth and transformation. Nature 455, 684–688.
Gazda, H.T., Sheen, M.R., Vlachos, A., Choesmel, V., O’Donohue, M.F.,
Schneider, H., Darras, N., Hasman, C., Sieff, C.A., Newburger, P.E., et al.
(2008). Ribosomal protein L5 and L11 mutations are associated with cleft
palate and abnormal thumbs in Diamond-Blackfan anemia patients. Am. J.
Hum. Genet. 83, 769–780.
Guo, H., Ingolia, N.T., Weissman, J.S., and Bartel, D.P. (2010). Mammalian
microRNAs predominantly act to decrease target mRNA levels. Nature 466,
835–840.
Holzel, M., Orban, M., Hochstatter, J., Rohrmoser, M., Harasim, T.,
Malamoussi, A., Kremmer, E., Langst, G., and Eick, D. (2010). Defects in
18 S or 28 S rRNA processing activate the p53 pathway. J. Biol. Chem. 285,
6364–6370.
Johnson, S.M., Grosshans, H., Shingara, J., Byrom, M., Jarvis, R., Cheng, A.,
Labourier, E., Reinert, K.L., Brown, D., and Slack, F.J. (2005). RAS is regulated
by the let-7 microRNA family. Cell 120, 635–647.
Kim, J., Krichevsky, A., Grad, Y., Hayes, G.D., Kosik, K.S., Church, G.M., and
Ruvkun, G. (2004). Identification of many microRNAs that copurify with
polyribosomes in mammalian neurons. Proc. Natl. Acad. Sci. USA 101,
360–365.
Kim, J.K., Gabel, H.W., Kamath, R.S., Tewari, M., Pasquinelli, A., Rual, J.F.,
Kennedy, S., Dybbs, M., Bertin, N., Kaplan, J.M., et al. (2005). Functional
genomic analysis of RNA interference in C. elegans. Science 308, 1164–
1167.
Kirn-Safran, C.B., Oristian, D.S., Focht, R.J., Parker, S.G., Vivian, J.L., and
Carson, D.D. (2007). Global growth deficiencies in mice lacking the ribosomal
protein HIP/RPL29. Dev. Dyn. 236, 447–460.
Lai, K., Amsterdam, A., Farrington, S., Bronson, R.T., Hopkins, N., and Lees,
J.A. (2009). Many ribosomal protein mutations are associated with growth
impairment and tumor predisposition in zebrafish. Dev. Dyn. 238, 76–85.
Langmead, B., Trapnell, C., Pop, M., and Salzberg, S.L. (2009). Ultrafast and
memory-efficient alignment of short DNA sequences to the human genome.
Genome Biol. 10, R25.
Lee, Y.S., and Dutta, A. (2007). The tumor suppressor microRNA let-7
represses the HMGA2 oncogene. Genes Dev. 21, 1025–1030.
Lempiainen, H., and Shore, D. (2009). Growth control and ribosome biogen-
esis. Curr. Opin. Cell Biol. 21, 855–863.
Lewis, B.P., Shih, I.H., Jones-Rhoades, M.W., Bartel, D.P., and Burge, C.B.
(2003). Prediction of mammalian microRNA targets. Cell 115, 787–798.
Lim, L.P., Lau, N.C., Garrett-Engele, P., Grimson, A., Schelter, J.M., Castle, J.,
Bartel, D.P., Linsley, P.S., and Johnson, J.M. (2005). Microarray analysis
shows that some microRNAs downregulate large numbers of target mRNAs.
Nature 433, 769–773.
MacInnes, A.W., Amsterdam, A., Whittaker, C.A., Hopkins, N., and Lees, J.A.
(2008). Loss of p53 synthesis in zebrafish tumors with ribosomal protein gene
mutations. Proc. Natl. Acad. Sci. USA 105, 10408–10413.
Maroney, P.A., Yu, Y., Fisher, J., and Nilsen, T.W. (2006). Evidence that
microRNAs are associated with translating messenger RNAs in human cells.
Nat. Struct. Mol. Biol. 13, 1102–1107.
Mayr, C., and Bartel, D.P. (2009). Widespread shortening of 30UTRs by
alternative cleavage and polyadenylation activates oncogenes in cancer cells.
Cell 138, 673–684.
Miller, S.A., Brown, A.J., Farach-Carson, M.C., and Kirn-Safran, C.B. (2003).
HIP/RPL29 down-regulation accompanies terminal chondrocyte differentia-
tion. Differentiation 71, 322–336.
Nelson, P.T., Hatzigeorgiou, A.G., and Mourelatos, Z. (2004). miRNP:mRNA
association in polyribosomes in a human neuronal cell line. RNA 10,
387–394.
Nottrott, S., Simard, M.J., and Richter, J.D. (2006). Human let-7a miRNA
blocks protein production on actively translating polyribosomes. Nat. Struct.
Mol. Biol. 13, 1108–1114.
Oliver, E.R., Saunders, T.L., Tarle, S.A., and Glaser, T. (2004). Ribosomal
protein L24 defect in belly spot and tail (Bst), a mouse Minute. Development
131, 3907–3920.
Olsen, P.H., and Ambros, V. (1999). The lin-4 regulatory RNA controls develop-
mental timing in Caenorhabditis elegans by blocking LIN-14 protein synthesis
after the initiation of translation. Dev. Biol. 216, 671–680.
Oristian, D.S., Sloofman, L.G., Zhou, X., Wang, L., Farach-Carson, M.C., and
Kirn-Safran, C.B. (2009). Ribosomal protein L29/HIP deficiency delays
osteogenesis and increases fragility of adult bone in mice. J. Orthop. Res.
27, 28–35.
Pani�c, L., Montagne, J., Cokari�c, M., and Volarevi�c, S. (2007). S6-haploinsuf-
ficiency activates the p53 tumor suppressor. Cell Cycle 6, 20–24.
Parry, D.H., Xu, J., and Ruvkun, G. (2007). A whole-genome RNAi Screen for
C. elegans miRNA pathway genes. Curr. Biol. 17, 2013–2022.
Petersen, C.P., Bordeleau, M.E., Pelletier, J., and Sharp, P.A. (2006). Short
RNAs repress translation after initiation in mammalian cells. Mol. Cell 21,
533–542.
Pillai, R.S., Bhattacharyya, S.N., Artus, C.G., Zoller, T., Cougot, N., Basyuk, E.,
Bertrand, E., and Filipowicz, W. (2005). Inhibition of translational initiation by
Let-7 MicroRNA in human cells. Science 309, 1573–1576.
Rudra, D., and Warner, J.R. (2004). What better measure than ribosome
synthesis? Genes Dev. 18, 2431–2436.
Saleh, M.C., van Rij, R.P., Hekele, A., Gillis, A., Foley, E., O’Farrell, P.H., and
Andino, R. (2006). The endocytic pathway mediates cell entry of dsRNA to
induce RNAi silencing. Nat. Cell Biol. 8, 793–802.
Scheffner, M.,Werness, B.A., Huibregtse, J.M., Levine, A.J., and Howley, P.M.
(1990). The E6 oncoprotein encoded by human papillomavirus types 16 and
18 promotes the degradation of p53. Cell 63, 1129–1136.
Seggerson, K., Tang, L., and Moss, E.G. (2002). Two genetic circuits repress
the Caenorhabditis elegans heterochronic gene lin-28 after translation initia-
tion. Dev. Biol. 243, 215–225.
Stewart, M.J., and Denell, R. (1993). Mutations in the Drosophila gene encod-
ing ribosomal protein S6 cause tissue overgrowth. Mol. Cell Biol. 13, 2524–
2535.
Talotta, F., Cimmino, A., Matarazzo, M.R., Casalino, L., De Vita, G.,
D’Esposito, M., Di Lauro, R., and Verde, P. (2009). An autoregulatory loop
mediated by miR-21 and PDCD4 controls the AP-1 activity in RAS transforma-
tion. Oncogene 28, 73–84.
Ulvila, J., Parikka, M., Kleino, A., Sormunen, R., Ezekowitz, R.A., Kocks, C.,
and Ramet, M. (2006). Double-stranded RNA is internalized by scavenger
receptor-mediated endocytosis in Drosophila S2 cells. J. Biol. Chem. 281,
14370–14375.
van Riggelen, J., Yetil, A., and Felsher, D.W. (2010). MYC as a regulator of
ribosome biogenesis and protein synthesis. Nat. Rev. Cancer 10, 301–309.
Vasudevan, S., Tong, Y., and Steitz, J.A. (2007). Switching from repression
to activation: microRNAs can up-regulate translation. Science 318, 1931–
1934.
Volarevic, S., Stewart, M.J., Ledermann, B., Zilberman, F., Terracciano, L.,
Montini, E., Grompe, M., Kozma, S.C., and Thomas, G. (2000). Proliferation,
but not growth, blocked by conditional deletion of 40S ribosomal protein S6.
Science 288, 2045–2047.
Watson, K.L., Konrad, K.D., Woods, D.F., and Bryant, P.J. (1992). Drosophila
homolog of the human S6 ribosomal protein is required for tumor suppression
in the hematopoietic system. Proc. Natl. Acad. Sci. USA 89, 11302–11306.
Werness, B.A., Levine, A.J., and Howley, P.M. (1990). Association of human
papillomavirus types 16 and 18 E6 proteins with p53. Science 248, 76–79.
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc. 185

Wightman, B., Ha, I., and Ruvkun, G. (1993). Posttranscriptional regulation of
the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in
C. elegans. Cell 75, 855–862.
Wu, L., and Belasco, J.G. (2005). Micro-RNA regulation of the mammalian lin-
28 gene during neuronal differentiation of embryonal carcinoma cells. Mol. Cell
Biol. 25, 9198–9208.
Wu, L., Fan, J., and Belasco, J.G. (2006). MicroRNAs direct rapid deadenyla-
tion of mRNA. Proc. Natl. Acad. Sci. USA 103, 4034–4039.
Zhou, R., Hotta, I., Denli, A.M., Hong, P., Perrimon, N., and Hannon, G.J.
(2008). Comparative analysis of argonaute-dependent small RNA pathways
in Drosophila. Mol. Cell 32, 592–599.
Molecular Cell
Ribosomal Proteins Modulate miRNA Activity
186 Molecular Cell 46, 171–186, April 27, 2012 ª2012 Elsevier Inc.


















