
Rapid identification of ocular dominance columns in...
14
Rapid identification of ocular dominance columns in macaques using cytochrome oxidase, Zif268, and dark-field microscopy JONATHAN C. HORTON, DAVINA R. HOCKING, and DANIEL L. ADAMS Beckman Vision Center, University of California—San Francisco, San Francisco (Received November 16, 1999; Accepted January 31, 2000) Abstract Strabismus induces an abnormal pattern of alternating light and dark columns of cytochrome oxidase (CO) activity in macaque striate cortex. This pattern may arise because visual perception is suppressed in one eye to avoid diplopia. To test whether CO activity is reduced in the ocular dominance columns of the suppressed eye, we performed monocular enucleation to co-label the ocular dominance columns with Zif268 immunohistochemistry in seven exotropic adult Macaca fascicularis. This approach was unsuccessful, for two reasons. First, Zif268 yielded inconsistent labelling, that was usually greater in the enucleated eye’s ocular dominance columns, but was sometimes greater in the intact eye’s columns. Therefore, Zif268 was not a reliable method for identifying the ocular dominance columns serving each eye. Second, in three control animals we found that a brief survival period following monocular enucleation (needed for Zif268 levels to change) was long enough to alter CO staining. For example, a survival time of only 3 h was sufficient to induce CO columns, indicating that the activity of this enzyme fluctuates more rapidly than realized previously. Independent of these findings, we have also discovered that acute monocular enucleation produces a vivid pattern of ocular dominance columns visible in unstained or CO-stained sections under dark-field illumination. The ocular dominance columns of the acutely enucleated eye appear dark. This was verified by labelling the ocular dominance columns with [ 3 H]proline. Dark-field illumination of the cortex after acute monocular enucleation offers a new, easy method for identifying the ocular dominance columns in macaques. Keywords: Intermediate early gene, EGR1, Krox24, Striate cortex, Strabismus, Border strip, Core zone, Stereopsis, Binocular, Diplopia, Dark field, Enucleation, Proline, Autoradiography, Suppression Introduction Experimental strabismus induces alternating dark and light col- umns of CO activity in macaque striate cortex (Tychsen & Burkhal- ter, 1997; Fenstemaker et al., 1997; Horton & Hocking, 1998a). This pattern may result from suppression of visual perception in one eye. To prove that metabolic activity is reduced in columns serving the suppressed eye, it is vital to employ a second label to visualize independently the ocular dominance columns in layer IVc. The traditional approach has been to rely on transneuronal autoradiography after [ 3 H]proline injection into one eye (Wiesel et al., 1974). This method, however, has disadvantages. The eye can be injured during injection, causing an artefactual change in cortical CO activity. Emulsion-coated slides must be exposed for months in the dark and disposal costs for radioactivity have be- come exorbitant. For these reasons, we have been eager to find an alternative way to compare CO strabismus patterns and ocular dominance columns in the same cortex. Immunostaining for the transcription factor Zif268 (Christy et al., 1988) has shown promise for this purpose. Levels of Zif268, also known as Krox24 (Lemaire et al., 1988) or EGR1 (Sukhatme et al., 1988), fluctuate with neuronal activity. These fluctuations occur rapidly, because Zif268 is the product of an immediate-early gene. Such genes regulate the expression of other genes, and hence are among the first to be transcribed differentially in response to sen- sory stimulation (Hunt et al., 1987; Sagar et al., 1988; Bullitt, 1990; Draisci & Iadarola 1989; Bravo, 1990). In rodents, injection of tetrodotoxin into one eye reduces Zif268 immunoreactivity in the contralateral visual cortex within 4 h (Worley et al., 1990). Just 1 h of visual experience in dark-reared kittens is enough to in- crease Zif268 mRNA levels in visual cortex (Rosen et al., 1992). In macaque striate cortex, dark and light columns of Zif268 im- munostaining appear only 2 h after monocular deprivation (Chau- duri & Cynader, 1993; Chaudhuri et al., 1995). In these monkey experiments, the dark columns were assigned to the ocular dom- inance columns of the open, stimulated eye. Address correspondence and reprint requests to: Jonathan C. Horton, Beckman Vision Center, 10 Kirkham Street, University of California—San Francisco, San Francisco, CA 94143-0730, USA. E-mail: horton@itsa. ucsf.edu Visual Neuroscience (2000), 17, 495–508. Printed in the USA. Copyright © 2000 Cambridge University Press 0952-5238000 $12.50 495
Transcript of Rapid identification of ocular dominance columns in...

Rapid identification of ocular dominance columnsin macaques using cytochrome oxidase, Zif268,and dark-field microscopy
JONATHAN C. HORTON, DAVINA R. HOCKING,and DANIEL L. ADAMSBeckman Vision Center, University of California—San Francisco, San Francisco
(Received November 16, 1999;Accepted January 31, 2000)
Abstract
Strabismus induces an abnormal pattern of alternating light and dark columns of cytochrome oxidase (CO) activityin macaque striate cortex. This pattern may arise because visual perception is suppressed in one eye to avoiddiplopia. To test whether CO activity is reduced in the ocular dominance columns of the suppressed eye, weperformed monocular enucleation to co-label the ocular dominance columns with Zif268 immunohistochemistry inseven exotropic adultMacaca fascicularis. This approach was unsuccessful, for two reasons. First, Zif268 yieldedinconsistent labelling, that was usually greater in the enucleated eye’s ocular dominance columns, but wassometimes greater in the intact eye’s columns. Therefore, Zif268 was not a reliable method for identifying theocular dominance columns serving each eye. Second, in three control animals we found that a brief survival periodfollowing monocular enucleation (needed for Zif268 levels to change) was long enough to alter CO staining. Forexample, a survival time of only 3 h was sufficient to induce CO columns, indicating that the activity of thisenzyme fluctuates more rapidly than realized previously. Independent of these findings, we have also discoveredthat acute monocular enucleation produces a vivid pattern of ocular dominance columns visible in unstained orCO-stained sections under dark-field illumination. The ocular dominance columns of the acutely enucleated eyeappear dark. This was verified by labelling the ocular dominance columns with [3H]proline. Dark-field illuminationof the cortex after acute monocular enucleation offers a new, easy method for identifying the ocular dominancecolumns in macaques.
Keywords: Intermediate early gene, EGR1, Krox24, Striate cortex, Strabismus, Border strip, Core zone, Stereopsis,Binocular, Diplopia, Dark field, Enucleation, Proline, Autoradiography, Suppression
Introduction
Experimental strabismus induces alternating dark and light col-umns of CO activity in macaque striate cortex (Tychsen & Burkhal-ter, 1997; Fenstemaker et al., 1997; Horton & Hocking, 1998a).This pattern may result from suppression of visual perception inone eye. To prove that metabolic activity is reduced in columnsserving the suppressed eye, it is vital to employ a second label tovisualize independently the ocular dominance columns in layerIVc. The traditional approach has been to rely on transneuronalautoradiography after [3H]proline injection into one eye (Wieselet al., 1974). This method, however, has disadvantages. The eyecan be injured during injection, causing an artefactual change incortical CO activity. Emulsion-coated slides must be exposed formonths in the dark and disposal costs for radioactivity have be-
come exorbitant. For these reasons, we have been eager to find analternative way to compare CO strabismus patterns and oculardominance columns in the same cortex.
Immunostaining for the transcription factor Zif268 (Christy et al.,1988) has shown promise for this purpose. Levels of Zif268, alsoknown as Krox24 (Lemaire et al., 1988) or EGR1 (Sukhatme et al.,1988), fluctuate with neuronal activity. These fluctuations occurrapidly, because Zif268 is the product of an immediate-early gene.Such genes regulate the expression of other genes, and hence areamong the first to be transcribed differentially in response to sen-sory stimulation (Hunt et al., 1987; Sagar et al., 1988; Bullitt,1990; Draisci & Iadarola 1989; Bravo, 1990). In rodents, injectionof tetrodotoxin into one eye reduces Zif268 immunoreactivity inthe contralateral visual cortex within 4 h (Worley et al., 1990). Just1 h of visual experience in dark-reared kittens is enough to in-crease Zif268 mRNA levels in visual cortex (Rosen et al., 1992).In macaque striate cortex, dark and light columns of Zif268 im-munostaining appear only 2 h after monocular deprivation (Chau-duri & Cynader, 1993; Chaudhuri et al., 1995). In these monkeyexperiments, the dark columns were assigned to the ocular dom-inance columns of the open, stimulated eye.
Address correspondence and reprint requests to: Jonathan C. Horton,Beckman Vision Center, 10 Kirkham Street, University of California—SanFrancisco, San Francisco, CA 94143-0730, USA. E-mail: [email protected]
Visual Neuroscience(2000),17, 495–508. Printed in the USA.Copyright © 2000 Cambridge University Press 0952-5238000 $12.50
495

We decided to use Zif268 histochemistry to label the oculardominance columns in macaques with experimental strabismus,using the following approach. The medial rectus muscle tendonswere cut to induce a divergent strabismus (exotropia). Next, wewaited at least 5 weeks to allow plenty of time for strabismus toproduce CO columns. Finally, one eye was enucleated 4–5 h be-fore perfusion to label the ocular dominance columns with Zif268.The timing of each step was designed to exploit the time windowbetween the rapid response of Zif268 and the slow response of COto experimental manipulation. Zif268 immunostaining changes inonly a few hours, but CO patterns have been thought to requiremany hours, if not days, to achieve a new level resolvable histo-chemically (Wong-Riley, 1989; Hevner & Wong-Riley, 1990). Thesurvival time after enucleation was intended to be long enough toproduce strong Zif268 labelling, but short enough to avoid cor-rupting CO patterns induced by strabismus.
This experimental strategy failed, for two reasons. First, in ourhands Zif268 could not be counted on to label the ocular domi-nance columns in a consistent fashion. In some animals, immuno-staining was darker in the ocular dominance columns of the intacteye, as expected, but in other animals it appeared darker in thecolumns of the enucleated eye. In a few instances, contradictorypatterns of staining were found in the same animal. Second, con-trol experiments unexpectedly showed that faint CO columns couldbe induced in normal macaques within 3–5.5 h of enucleation.Consequently, for practical purposes, the difference in the timingbetween Zif268 and CO responses proved illusory. CO patternscaused by strabismus could be confounded by the acute enucle-ation performed for Zif268 labelling.
After discovering these pitfalls, we abandoned Zif268 immu-nohistochemistry and returned to [3H]proline transneuronal auto-radiography. Using autoradiography, we have shown that the darkCO columns induced by strabismus are usually associated with thefixating eye. In addition, we have described a pattern of slenderpale bands, induced by loss of CO activity in the border strips thatrun along the boundaries of ocular dominance columns. This pat-tern prevails in cortex where perception alternates between theeyes, but binocularity is reduced by strabismus. A complete ac-count of these experiments has been published elsewhere (Hortonet al., 1999).
Our goal in reporting the present CO0Zif268 experiments,which ultimately fell short of our expectations, is to provide acautionary note for other investigators who may attempt the sameapproach. In addition, we describe a new method for visualizingocular dominance columns, discovered during these experiments.Ocular dominance columns can be seen clearly after acute enu-cleation by examining CO-stained or unstained sections underdark-field illumination. This technique provides an easy, rapidway to label ocular dominance columns, which may be usefulfor many applications.
Methods
Experimental animals
We used ten normal adult maleMacaca fascicularis. Table 1 sum-marizes the experimental procedures and findings for each animal.These results have not been reported previously, except in prelim-inary form (Horton & Hocking, 1998a; Hocking et al., 1999). Allexperiments were approved by the Committee on Animal Researchat the University of California, San Francisco. In seven monkeys,we induced strabismus by tenotomy of the medial rectus muscle in
both eyes. The procedure was performed under general anesthesiawith ketamine HCl (15 mg0kg, IM) and topical proparacaine HCl.The animals were checked daily after the operation to verify rapidhealing and to observe ocular alignment. We waited at least 5weeks before proceeding with the next step in each experiment.
The ocular dominance columns were labelled with [3H]prolinein one strabismic monkey and one control monkey (Table 1), usingmethods described previously (Horton & Hocking, 1998b). Im-mediately after tracer injection into the eye, the fundus was checkedwith an indirect ophthalmoscope for any sign of injury. Label wasallowed to transport for 7–9 days. Just prior to perfusion, the eyewas examined again with an indirect ophthalmoscope. No damagefrom the eye injection was detected in any monkey.
For Zif268 labelling, monkeys were dark-adapted overnight for12 h. The next morning we enucleated one eye in total darknesswith the aid of night vision goggles (Horton & Hocking, 1996a).The operation was performed under general anesthesia with keta-mine HCl (15 mg0kg, IM) and retrobulbar injection of 1% lidocaine0epinephrine (1:200,000). Ketamine is an NMDA receptor blocker,with the potential to reduce Zif268 levels. However, Chaudhuriet al. (1995) have obtained robust Zif268 labelling in animalsgiven ketamine, and, in any case, the drug should not affectrela-tive levels of Zif268 in the ocular dominance columns.
Immediately after enucleation, the room lights were turned onto stimulate the remaining eye. The monkey usually recoveredconsciousness in 30 min. Total survival time after enucleationvaried between 4.25–5.0 h. Thorough infiltration of the orbitaltissues with local anesthetic insured that the animal had no post-operative pain. Enucleation was performed, rather than eyelid su-ture, because it yields higher contrast Zif268 labelling (Chaudhuriet al., 1995). The animals were allowed to reawaken after monoc-ular enucleation, instead of being maintained under general anes-thesia, because visual stimulation of conscious animals yieldsstronger Zif268 labelling (Chaudhuri, personal communication,1996).
Histological procedures
After receiving a lethal injection of sodium pentobarbital into theperitoneal cavity, each monkey was perfused through the left ven-tricle with 1 liter of normal saline followed by 1 liter of 2%paraformaldehyde in 0.1 M phosphate buffer. Flat-mounts of stri-ate cortex were prepared from each hemisphere (Horton & Hock-ing, 1996b) and cut with a freezing microtome. Alternate sectionswere reacted on slides for CO (Wong-Riley, 1979) or free-floatingfor Zif268. In the two animals who received [3H]proline, everythird section was dipped in NTB-2 emulsion (Eastman Kodak,Rochester, NY) for autoradiography. The autoradiographs wereexposed for 10 weeks and developed with D19 developer.
Zif268 immunochemistry
Sections were incubated in the primary anti-Zif268 rabbit antibodydiluted at 1:10,000 in 3% normal goat serum, 0.02% sodium azide,and 0.1 M phosphate-buffered saline (PBS)(Herdegen et al., 1990;Chaudhuri et al., 1995). The anti-Zif268 antibody was a gift fromRodrigo Bravo. The sections were agitated at room temperature for48 h. After three washes in PBS containing 0.3% Triton X-100, thesections were processed using a rabbit Vectorstain Elite ABC kit(Vector Laboratories, Inc, Burlingame, CA). They were placedinto biotinylated goat anti-rabbit secondary antibody (1:1000 with3% normal goat serum in PBS1 0.3% Triton X-100) and agitated
496 J.C. Horton, D.R. Hocking, and Daniel L. Adams

Table 1. Summary of experimentsa
Results
Monkey No.Deviated eye0
Strabismus duration Techniques Procedures Cytochrome oxidase, dark field Darker Zif268 immunostaining
1 Alternating exotropia5 weeks
COZif268
Enucleated OS5-h survival
Left cortex: BS dark, ipsi columns darkRight cortex: BS light, ipsi columns light
Dark columns = enucleated eye
Left cortex: Enucleated eye’s columnsRight cortex: Not reacted
2 Left exotropia6 weeks
COZif268
Enucleated OD4.5-h survival
Left cortex: BS light, ipsi columns lightRight cortex: peripheral tissue damaged
Dark columns = enucleated eye
Left cortex: Enucleated eye’s columnsRight cortex: Not reacted
3 Left exotropia14 weeks
COZif268
Enucleated OD5-h survival
Left cortex: BS light, ipsi columns lightRight cortex: BS dark, ipsi columns dark
Dark columns = enucleated eye
Left cortex: Enucleated eye’s and intact eye’scolumns in different areasRight cortex: Enucleated eye’s and intact eye’scolumns in different areas
4 Right exotropia5 weeks
COZif268
Enucleated OS4.5-h survival
Left cortex: BS not visible, ipsi columns darkRight cortex: BS not visible, ipsi columns light
Dark columns = enucleated eye
Left cortex: Intact eye’s columnsRight cortex: Intact eye’s columns
5 Left exotropia6 weeks
COZif268
Enucleated OD5-hr survival
Left cortex: BS light, ipsi columns lightRight cortex: BS dark, ipsi columns dark
Dark columns = enucleated eye
Left cortex: Enucleated eye’s columnsRight cortex: Enucleated eye’s columns
6 Right exotropia7 weeks
CO,Zif268
Enucleated OD5-h survival
Left cortex: BS light, ipsi column lightRight cortex: BS dark, ipsi columns dark
Dark columns = enucleated eye
Left cortex: Enucleated eye’s columnsRight cortex: Enucleated eye’s columns
7 Left exotropia11 months
COZif268[3H]Proline
[3H]Proline injection ODEnucleated OS4.25-h survival
Left cortex: BS dark, ipsi columns darkRight cortex: BS light, ispi columns light
Dark columns = enucleated eye
Left cortex: Intact eye’s columnsRight cortex: Enucleated eye’s and intact eye’scolumns in different areas
8 Normal animal CO No dark adaptationEnucleated OD10-h survival
Left cortex: BS not visible, ipsi columns lightRight cortex: BS dark, ipsi columns dark
Dark columns = enucleated eye
Not reacted
9 Normal animal COZif268[3H]Proline
[3H]Proline injection ODEnucleated OS5.5-h survival
Left cortex: BS not visible, ipsi columns darkRight cortex: BS not visible, ipsi columns light
Dark columns = enucleated eye
Left cortex: Intact eye’s columnsRight cortex: Enucleated eye’s columns
10 Normal animal CO No dark adaptationEnucleated OD3-h survival under generalanesthesia
No CO dark-field columns Not reacted
aCO 5 cytochrome oxidase; OD5 right eye; OS5 left eye; BS5 blind spot.
Ra
pid
lab
ellin
go
fo
cula
rd
om
ina
nce
colu
mn
s497
at room temperature for 2 h. After washing in PBS1 0.3% TritonX-100, the sections were transferred to solution A (1:1000 in PBS10.3% Triton X-100 ) and solution B (1:1000 in PBS1 0.3% TritonX-100) mixed in equal volumes. Incubation in this solution lasted1 h at room temperature. The sections were then washed in PBSand reacted with a diaminobenzidine0nickel chromagen. The re-action was monitored frequently until it reached the desired inten-sity, usually within 20–30 min. The sections were washed in buffer,mounted onto gelatinized slides, and coverslipped. Control sec-tions from each hemisphere were processed in an identical manner,except for the omission of the anti-Zif268 primary antibody. Theyshowed no column patterns and only faint, nonspecific backgroundlabel.
Data analysis
Cortical CO flat-mounts were imaged at 600 dots per inch on anAgfa Arcus II flatbed scanner with a transparency adapter. Fordark-field illumination, CO flat-mounts and autoradiographs were
examined with an Olympus SZH10 microscope. To compare re-gions at high power, CO sections, Zif268 sections, and autoradio-graphs were photographed using Technical Pan film (EastmanKodak). Negatives were processed with D19 developer and scannedinto the computer using a Microtek Scanmaker 35t scanner (Re-dondo Beach, CA). Images were imported into Photoshop 5.0(Adobe Systems, San Jose, CA) to make adjustments in the con-trast and brightness of sections. In addition, we used the UnsharpMask filter to enhance the visibility of column patterns.
Results
Strabismic monkeys
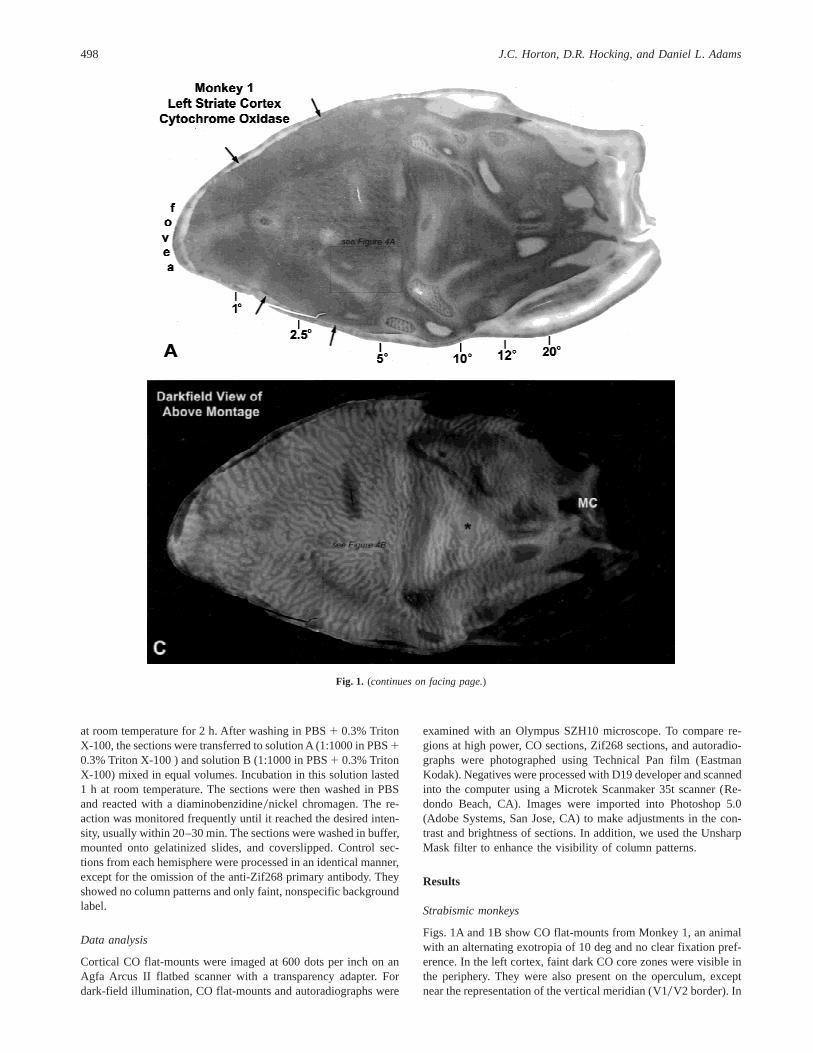
Figs. 1A and 1B show CO flat-mounts from Monkey 1, an animalwith an alternating exotropia of 10 deg and no clear fixation pref-erence. In the left cortex, faint dark CO core zones were visible inthe periphery. They were also present on the operculum, exceptnear the representation of the vertical meridian (V10V2 border). In
Fig. 1. (continues on facing page.)
498 J.C. Horton, D.R. Hocking, and Daniel L. Adams

this region, a pattern of pale border strips was seen (for definitionsof “core zones” and “border strips”, see Horton, 1984; Horton &Hocking, 1998b; Horton et al., 1999). In the right cortex, paleborder strips predominated throughout the operculum. In the pe-riphery, there were dark and light CO columns. The pattern of COstaining is summarized in Fig. 2. It was consistent with perceptionalternating between the eyes in the central visual field, and dom-inance of the ipsilateral eye in the periphery of each visual hemifield.
Several years ago we reported that ocular dominance columnscan be seen in wet, unstained sections viewed in dark-field illumi-nation (Horton & Hocking, 1997). These sections were from mon-keys that had survived many weeks after monocular enucleation. Thisfinding prompted us to examine the CO sections from Monkey 1under dark field. We were rewarded with a pattern of alternating darkand bright columns, resembling ocular dominance columns (Figs. 1C
and 1D). The dark columns coincided with the ocular dominancecolumns of the enucleated left eye. This was ascertained by exam-ining the blind spot “representation” in each cortex. It matched thecontrast of the dark columns on the left side and and the bright col-umns on the right side. The blind spot representation receives inputfrom only the ipsilateral eye, so one can infer that the dark oculardominance columns belonged to the left eye. This impression wasconfirmed by examining the ocular dominance columns in periph-eral cortex. In this region, LeVay and colleagues (1985) have shownthat the ocular dominance columns serving the ipsilateral eye be-come naturally shrunken and fragmented. In the left periphery(Fig. 1C), the shrunken columns were dark. Therefore, the dark col-umns corresponded to the ocular dominance columns of the ipsi-lateral (enucleated), left eye. In the right cortex, as expected, theshrunken peripheral columns were bright (Fig. 1D).
Fig. 1. Monkey 1 (10 deg exotropia, alternator). (A,B) Flat-mounts showing the pattern of CO staining in layer IVc. In the left cortex(montage of two sections), alternating pale and dark columns were seen everywhere, except near the V10V2 border (arrows). In thisregion, pale border strips were present, which are poorly visible at low power. In the right cortex (single section), there were pale borderstrips on the operculum and columns in the periphery. (C,D) Same tissue as above, viewed in dark field, showing hidden oculardominance columns induced after 5 h by enucleation of the left eye. In the left periphery, the blind spot representation (*) and theshrunken, fragmented columns were dark, indicating that the dark columns belonged to the enucleated eye. MC: monocular crescent.
Rapid labelling of ocular dominance columns 499
Seven strabismic monkeys underwent monocular enucleation,with survival times of 4.25–5.0 h. Dark-field illumination showedclear ocular dominance columns in layer IVc of every animal.Examination of the blind spots and shrunken peripheral columns ineach animal established, without exception, that the dark columnsbelonged to the enucleated eye (Table 1). This allowed us to usedark-field illumination of CO sections to identify reliably the oc-ular dominance columns serving the left eye and the right eye ineach animal.
To determine if the dark-field columns were due to strabismus,or to monocular enucleation, we used dark-field illumination toexamine CO sections from five strabismic monkeys used in aseparate study (Horton et al., 1999) who had not undergone mon-ocular enucleation. None showed dark-field columns. Therefore,enucleation was responsible for their emergence.
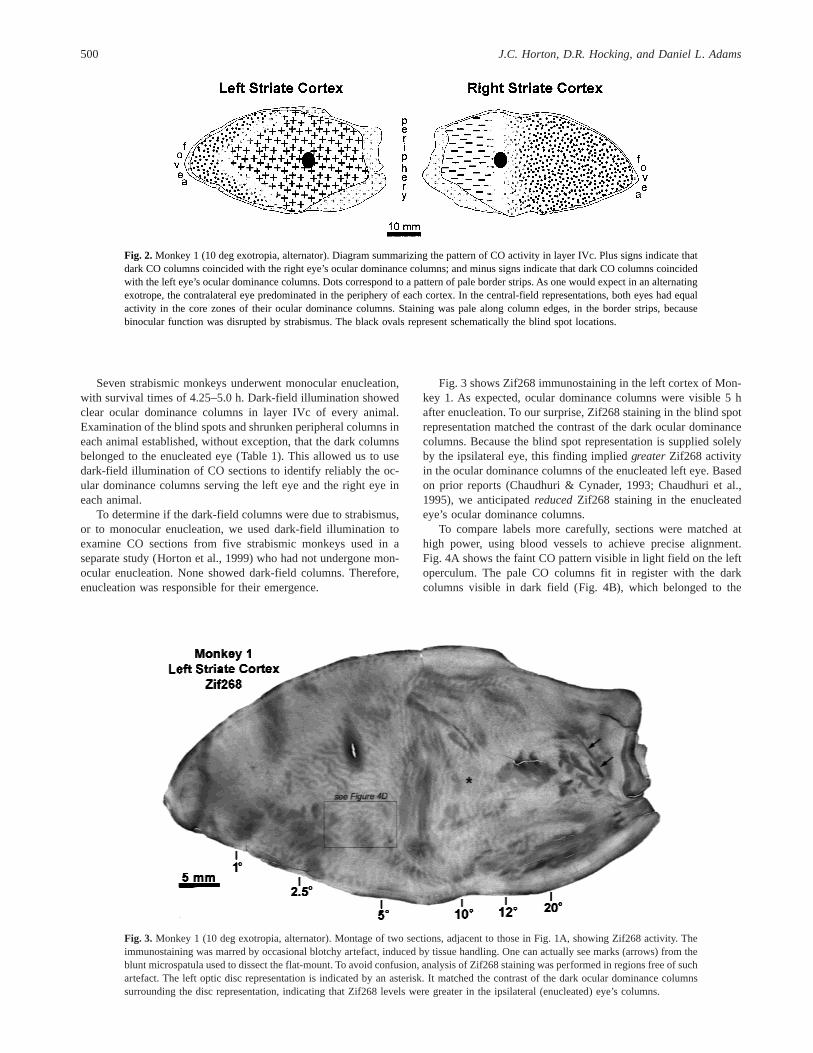
Fig. 3 shows Zif268 immunostaining in the left cortex of Mon-key 1. As expected, ocular dominance columns were visible 5 hafter enucleation. To our surprise, Zif268 staining in the blind spotrepresentation matched the contrast of the dark ocular dominancecolumns. Because the blind spot representation is supplied solelyby the ipsilateral eye, this finding impliedgreaterZif268 activityin the ocular dominance columns of the enucleated left eye. Basedon prior reports (Chaudhuri & Cynader, 1993; Chaudhuri et al.,1995), we anticipatedreducedZif268 staining in the enucleatedeye’s ocular dominance columns.
To compare labels more carefully, sections were matched athigh power, using blood vessels to achieve precise alignment.Fig. 4A shows the faint CO pattern visible in light field on the leftoperculum. The pale CO columns fit in register with the darkcolumns visible in dark field (Fig. 4B), which belonged to the
Fig. 2. Monkey 1 (10 deg exotropia, alternator). Diagram summarizing the pattern of CO activity in layer IVc. Plus signs indicate thatdark CO columns coincided with the right eye’s ocular dominance columns; and minus signs indicate that dark CO columns coincidedwith the left eye’s ocular dominance columns. Dots correspond to a pattern of pale border strips. As one would expect in an alternatingexotrope, the contralateral eye predominated in the periphery of each cortex. In the central-field representations, both eyes had equalactivity in the core zones of their ocular dominance columns. Staining was pale along column edges, in the border strips, becausebinocular function was disrupted by strabismus. The black ovals represent schematically the blind spot locations.
Fig. 3. Monkey 1 (10 deg exotropia, alternator). Montage of two sections, adjacent to those in Fig. 1A, showing Zif268 activity. Theimmunostaining was marred by occasional blotchy artefact, induced by tissue handling. One can actually see marks (arrows) from theblunt microspatula used to dissect the flat-mount. To avoid confusion, analysis of Zif268 staining was performed in regions free of suchartefact. The left optic disc representation is indicated by an asterisk. It matched the contrast of the dark ocular dominance columnssurrounding the disc representation, indicating that Zif268 levels were greater in the ipsilateral (enucleated) eye’s columns.
500 J.C. Horton, D.R. Hocking, and Daniel L. Adams
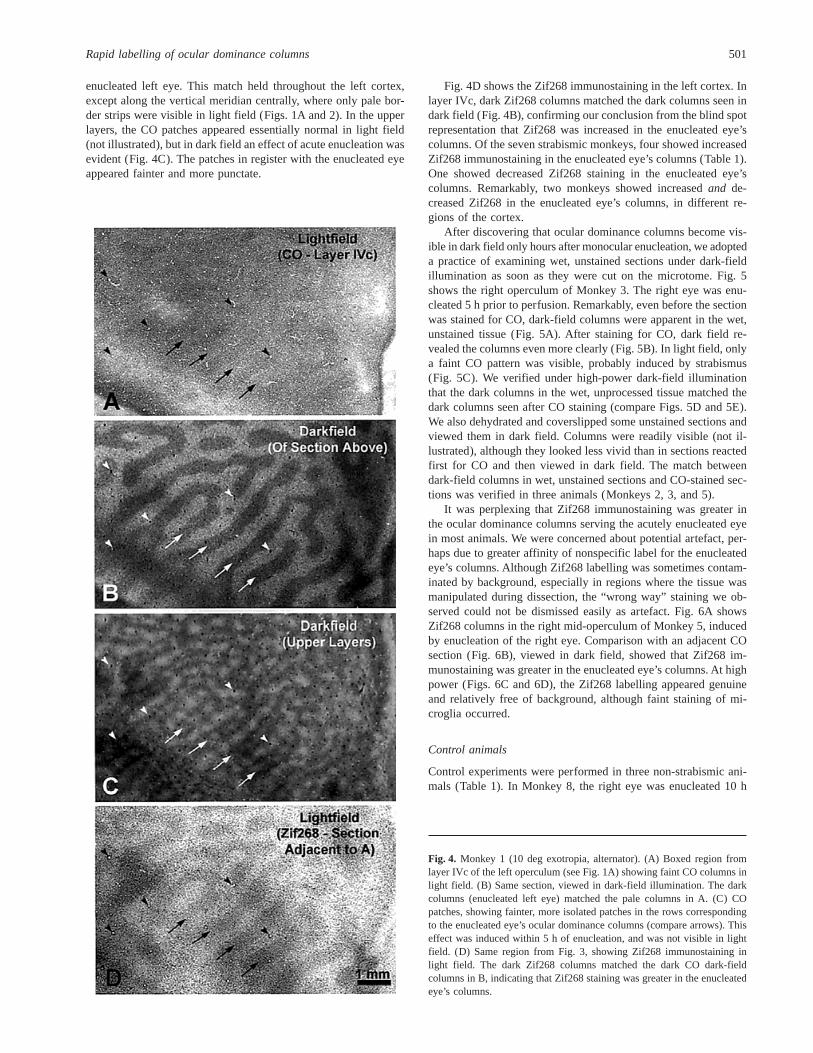
enucleated left eye. This match held throughout the left cortex,except along the vertical meridian centrally, where only pale bor-der strips were visible in light field (Figs. 1A and 2). In the upperlayers, the CO patches appeared essentially normal in light field(not illustrated), but in dark field an effect of acute enucleation wasevident (Fig. 4C). The patches in register with the enucleated eyeappeared fainter and more punctate.
Fig. 4D shows the Zif268 immunostaining in the left cortex. Inlayer IVc, dark Zif268 columns matched the dark columns seen indark field (Fig. 4B), confirming our conclusion from the blind spotrepresentation that Zif268 was increased in the enucleated eye’scolumns. Of the seven strabismic monkeys, four showed increasedZif268 immunostaining in the enucleated eye’s columns (Table 1).One showed decreased Zif268 staining in the enucleated eye’scolumns. Remarkably, two monkeys showed increasedand de-creased Zif268 in the enucleated eye’s columns, in different re-gions of the cortex.
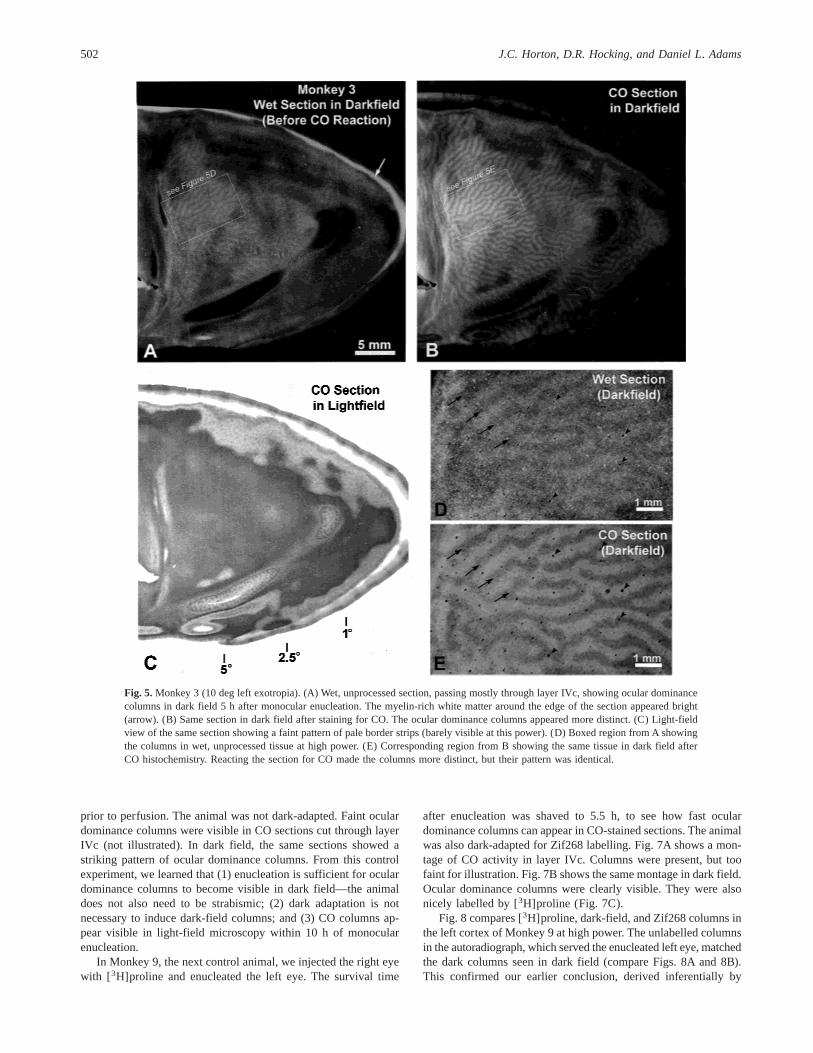
After discovering that ocular dominance columns become vis-ible in dark field only hours after monocular enucleation, we adopteda practice of examining wet, unstained sections under dark-fieldillumination as soon as they were cut on the microtome. Fig. 5shows the right operculum of Monkey 3. The right eye was enu-cleated 5 h prior to perfusion. Remarkably, even before the sectionwas stained for CO, dark-field columns were apparent in the wet,unstained tissue (Fig. 5A). After staining for CO, dark field re-vealed the columns even more clearly (Fig. 5B). In light field, onlya faint CO pattern was visible, probably induced by strabismus(Fig. 5C). We verified under high-power dark-field illuminationthat the dark columns in the wet, unprocessed tissue matched thedark columns seen after CO staining (compare Figs. 5D and 5E).We also dehydrated and coverslipped some unstained sections andviewed them in dark field. Columns were readily visible (not il-lustrated), although they looked less vivid than in sections reactedfirst for CO and then viewed in dark field. The match betweendark-field columns in wet, unstained sections and CO-stained sec-tions was verified in three animals (Monkeys 2, 3, and 5).
It was perplexing that Zif268 immunostaining was greater inthe ocular dominance columns serving the acutely enucleated eyein most animals. We were concerned about potential artefact, per-haps due to greater affinity of nonspecific label for the enucleatedeye’s columns. Although Zif268 labelling was sometimes contam-inated by background, especially in regions where the tissue wasmanipulated during dissection, the “wrong way” staining we ob-served could not be dismissed easily as artefact. Fig. 6A showsZif268 columns in the right mid-operculum of Monkey 5, inducedby enucleation of the right eye. Comparison with an adjacent COsection (Fig. 6B), viewed in dark field, showed that Zif268 im-munostaining was greater in the enucleated eye’s columns. At highpower (Figs. 6C and 6D), the Zif268 labelling appeared genuineand relatively free of background, although faint staining of mi-croglia occurred.
Control animals
Control experiments were performed in three non-strabismic ani-mals (Table 1). In Monkey 8, the right eye was enucleated 10 h
Fig. 4. Monkey 1 (10 deg exotropia, alternator). (A) Boxed region fromlayer IVc of the left operculum (see Fig. 1A) showing faint CO columns inlight field. (B) Same section, viewed in dark-field illumination. The darkcolumns (enucleated left eye) matched the pale columns in A. (C) COpatches, showing fainter, more isolated patches in the rows correspondingto the enucleated eye’s ocular dominance columns (compare arrows). Thiseffect was induced within 5 h of enucleation, and was not visible in lightfield. (D) Same region from Fig. 3, showing Zif268 immunostaining inlight field. The dark Zif268 columns matched the dark CO dark-fieldcolumns in B, indicating that Zif268 staining was greater in the enucleatedeye’s columns.
Rapid labelling of ocular dominance columns 501
prior to perfusion. The animal was not dark-adapted. Faint oculardominance columns were visible in CO sections cut through layerIVc (not illustrated). In dark field, the same sections showed astriking pattern of ocular dominance columns. From this controlexperiment, we learned that (1) enucleation is sufficient for oculardominance columns to become visible in dark field—the animaldoes not also need to be strabismic; (2) dark adaptation is notnecessary to induce dark-field columns; and (3) CO columns ap-pear visible in light-field microscopy within 10 h of monocularenucleation.
In Monkey 9, the next control animal, we injected the right eyewith [3H]proline and enucleated the left eye. The survival time
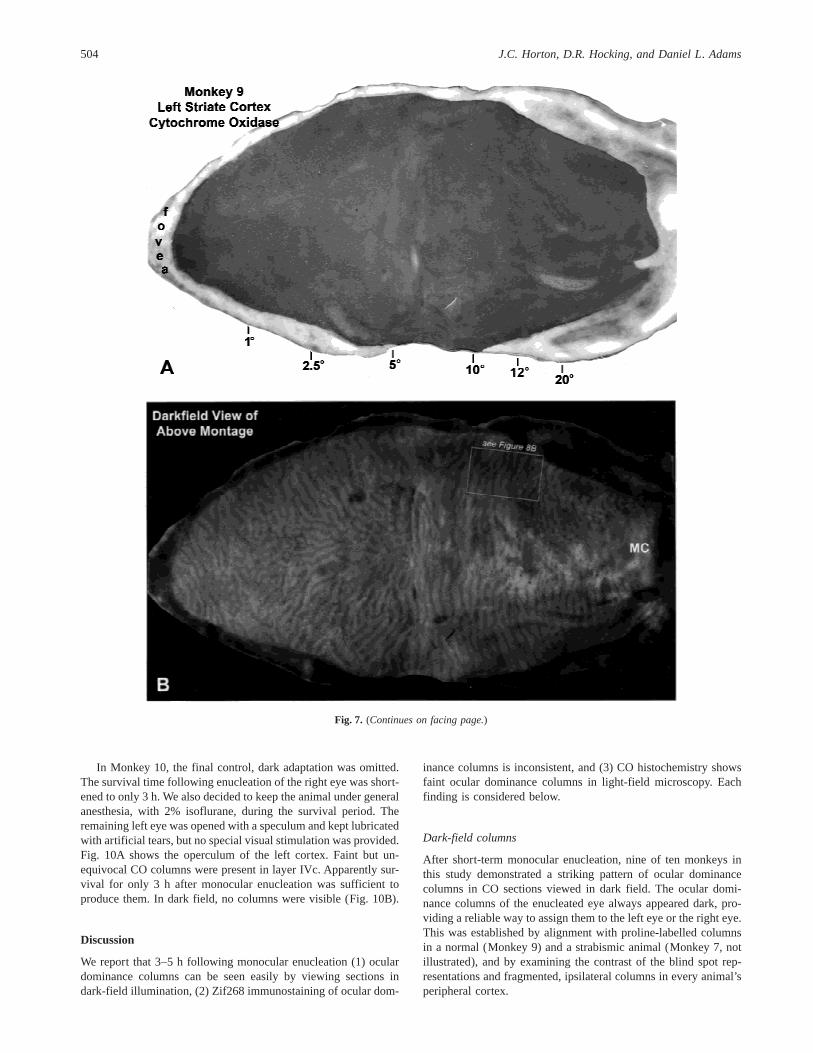
after enucleation was shaved to 5.5 h, to see how fast oculardominance columns can appear in CO-stained sections. The animalwas also dark-adapted for Zif268 labelling. Fig. 7A shows a mon-tage of CO activity in layer IVc. Columns were present, but toofaint for illustration. Fig. 7B shows the same montage in dark field.Ocular dominance columns were clearly visible. They were alsonicely labelled by [3H]proline (Fig. 7C).
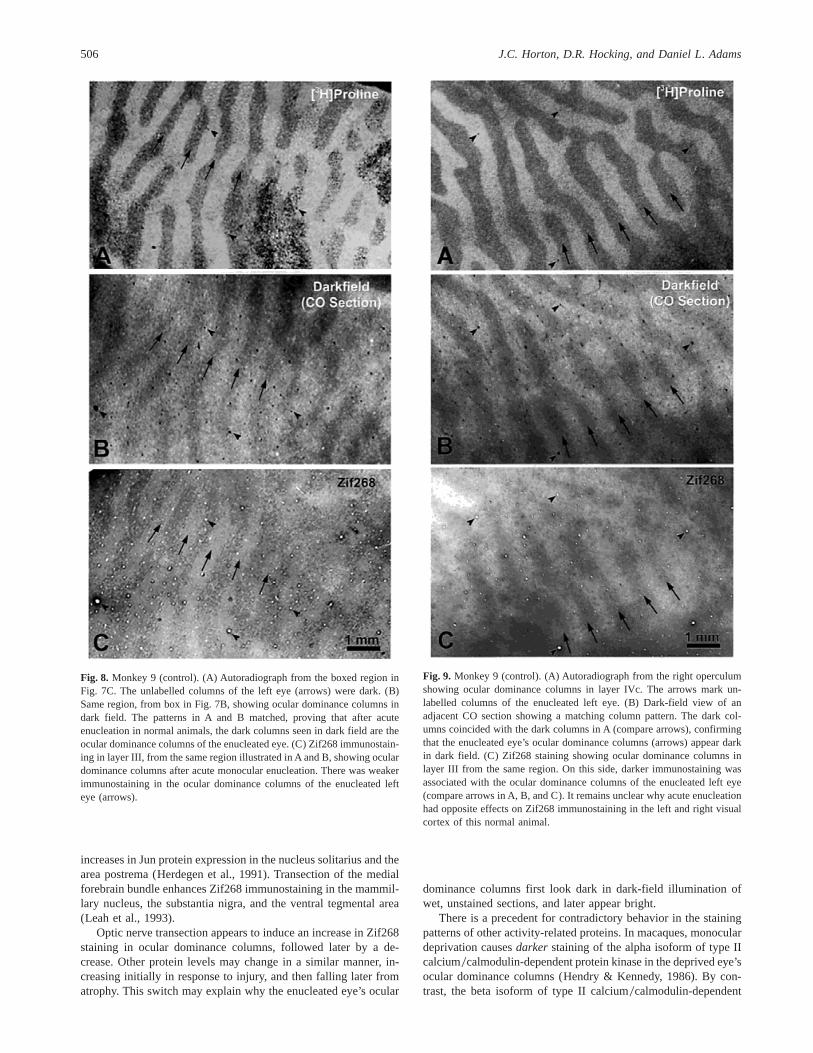
Fig. 8 compares [3H]proline, dark-field, and Zif268 columns inthe left cortex of Monkey 9 at high power. The unlabelled columnsin the autoradiograph, which served the enucleated left eye, matchedthe dark columns seen in dark field (compare Figs. 8A and 8B).This confirmed our earlier conclusion, derived inferentially by
Fig. 5. Monkey 3 (10 deg left exotropia). (A) Wet, unprocessed section, passing mostly through layer IVc, showing ocular dominancecolumns in dark field 5 h after monocular enucleation. The myelin-rich white matter around the edge of the section appeared bright(arrow). (B) Same section in dark field after staining for CO. The ocular dominance columns appeared more distinct. (C) Light-fieldview of the same section showing a faint pattern of pale border strips (barely visible at this power). (D) Boxed region from A showingthe columns in wet, unprocessed tissue at high power. (E) Corresponding region from B showing the same tissue in dark field afterCO histochemistry. Reacting the section for CO made the columns more distinct, but their pattern was identical.
502 J.C. Horton, D.R. Hocking, and Daniel L. Adams

examining the blind spot representation and peripheral columns,that the enucleated eye’s columns appear dark in dark field. Fig. 8Cshows the pattern of Zif268 labelling. The columns of the left eyewere pale, indicating that Zif268 activity was weaker in the enu-cleated eye’s columns.
Fig. 9 compares [3H]proline, dark-field, and Zif268 columnsin the right cortex of Monkey 9. As in the left cortex, theunlabelled columns in the autoradiograph matched the dark col-umns visible in dark field (compare Figs. 9A and 9B), confirm-ing that the dark columns belonged to the enucleated left eye.
Surprisingly, however, on this side Zif268 activity was greater inthe enucleated left eye’s columns (compare Figs. 9A–9C). Insummary, this animal showed that (1) faint ocular dominancecolumns emerge in CO light-field staining only 5.5 h after mon-ocular enucleation, and (2) inconsistent Zif268 staining can beseen even in normal monkeys. We also confirmed, using auto-radiography, the assignment of the dark CO dark-field columnsto the enucleated eye. This match was verified at high powerin numerous regions, in addition to those illustrated in Figs. 8and 9.
Fig. 6. Monkey 5 (5 deg left exotropia). (A) Zif268 staining in layer IVc of the right mid-operculum, showing ocular dominancecolumns in light field 5 h after monocular enucleation. (B) Adjacent CO dark-field section showing the same pattern of oculardominance columns. Comparison with A shows that Zif268 immunoreactivity was greater in the dark columns in B, which belongedto the enucleated right eye. (C) High-power view of an intact eye’s column containing weak Zif268 staining (upper box in A). (D) Forcomparison, dense nuclear Zif268 immunostaining within an enucleated eye’s ocular dominance column (lower box in A).
Rapid labelling of ocular dominance columns 503
In Monkey 10, the final control, dark adaptation was omitted.The survival time following enucleation of the right eye was short-ened to only 3 h. We also decided to keep the animal under generalanesthesia, with 2% isoflurane, during the survival period. Theremaining left eye was opened with a speculum and kept lubricatedwith artificial tears, but no special visual stimulation was provided.Fig. 10A shows the operculum of the left cortex. Faint but un-equivocal CO columns were present in layer IVc. Apparently sur-vival for only 3 h after monocular enucleation was sufficient toproduce them. In dark field, no columns were visible (Fig. 10B).
Discussion
We report that 3–5 h following monocular enucleation (1) oculardominance columns can be seen easily by viewing sections indark-field illumination, (2) Zif268 immunostaining of ocular dom-
inance columns is inconsistent, and (3) CO histochemistry showsfaint ocular dominance columns in light-field microscopy. Eachfinding is considered below.
Dark-field columns
After short-term monocular enucleation, nine of ten monkeys inthis study demonstrated a striking pattern of ocular dominancecolumns in CO sections viewed in dark field. The ocular domi-nance columns of the enucleated eye always appeared dark, pro-viding a reliable way to assign them to the left eye or the right eye.This was established by alignment with proline-labelled columnsin a normal (Monkey 9) and a strabismic animal (Monkey 7, notillustrated), and by examining the contrast of the blind spot rep-resentations and fragmented, ipsilateral columns in every animal’speripheral cortex.
Fig. 7. (Continues on facing page.)
504 J.C. Horton, D.R. Hocking, and Daniel L. Adams

It is unclear why the ocular dominance columns appeared vis-ible in dark field. The phenomenon probably does not reflect COactivity, because the dark-field and light-field patterns were oftendissociated. For example, in strabismic monkeys, the enucleatedeye’s ocular dominance columns always appeared dark in darkfield, but they could contain pale columns, dark columns, or paleborder strips when viewed in light field (Fig. 1). Furthermore, COstaining was not necessary to see the dark-field columns. Theywere present in wet, unstained sections and in dehydrated, un-stained sections coverslipped with Permount. However, CO stain-ing did improve the clarity of the dark-field columns.
In a previous report (Horton & Hocking, 1997), we describedocular dominance columns in wet, unprocessed sections viewed indark field. In those experiments, the minimum survival time aftermonocular enucleation was 3 weeks. The ocular dominance col-umns of the enucleated eye appeared bright in dark field. They alsostained more intensely for myelin basic protein, implying thatgreater myelin content made the columns visible in dark field. Inthe present experiments, the survival time following enucleationwas only a few hours. It seems unlikely that myelin content couldchange so rapidly.
Following acute enucleation, the dark-field columns of the enu-cleated eye appeared dark. As stated above, in a separate set ofexperiments,weeksafter monocular enucleation the dark-field col-umns of the enucleated eye appeared bright. These findings meanthat the contrast of the dark-field columns switches sometime be-tween hours and weeks following monocular enucleation.
Of the ten monkeys monocularly enucleated in this study, onlythe last control lacked dark-field columns. It was the only monkeymaintained under general anesthesia during the survival periodafter enucleation. This might explain the absence of columns. Thebrevity of the survival period (3 h) may also have been a factor.Further experiments would be required to settle this issue.
Dark-field illumination of sections after acute enucleation pro-vides a simple, rapid method for labelling the ocular dominance
columns. In some animals (but not all), columns were visible evenin wet, unstained tissue cut in tangential or cross section, whichmay be useful forin vitro applications.
Inconsistent Zif268 immunostaining
In previous studies, Zif268 columns were observed in macaquestriate cortex at 5 h, 1 day, 3 days, 5 days, and 3 months aftermonocular enucleation (Chaudhuri & Cynader, 1993; Chaudhuriet al., 1995). Comparison of Zif268 columns with ocular domi-nance columns (using CO as a second label) was performed onlyfor survival times of 5 days and 3 months. Comparison was alsomade at 24 h following intraocular tetrodotoxin injection. In everycase, a match was reported between the dark Zif268 columns andthe ocular dominance columns serving the intact eye. The corre-lations made by Chaudhuri and colleagues are difficult to evaluatebecause they usually examined cross sections of striate cortex.Comparisons between different labels are more reliable in the tan-gential plane, because blood vessels passing through the corticallayers are easy to identify, allowing precise registration of adjacentsections. This reservation aside, we have tested a single monkey6 days after monocular enucleation and concur that Zif268 immu-nostaining is greater in the ocular dominance columns of the intacteye (unpublished observation).
Chaudhuri and colleagues (1993, 1995) did not compare Zif268patterns and ocular dominance columns at 5 h after enucleation.They assumed that Zif268 immunostaining was reduced in theenucleated eye’s columns. However, after short-term enucleation(e.g. 5 h), most of the animals we tested showed greater Zif268immunostaining in the ocular dominance columns of the enucle-ated eye. This result was surprising, but not unprecedented. Infra-orbital nerve section increases c-Fos immunoreactivity in the spinaltrigeminal nucleus, beginning 4 h after injury (Sharp et al., 1989).Within 2 h, sciatic nerve crush increases Zif268 immunostaining indorsal horn neurons (Leah et al., 1991). Vagotomy induces rapid
Fig. 7. Monkey 9 (control). (A) Montage of layer IVc showing CO staining 5.5 h after enucleation of the left eye. Columns were toofaint to see in this illustration, but they could be detected in the actual sections. (B) Montage of the same sections showing columnsin dark field. The blind spot region was not seen clearly, but the thin, fragmented ipsilateral columns in the left periphery (near MC)were dark, consistent with their assignment to the enucleated eye. (C) Autoradiographic montage of adjacent sections prepared after[3H]proline injection into the right eye. In dark field, the radioactive label appeared bright.
Rapid labelling of ocular dominance columns 505
increases in Jun protein expression in the nucleus solitarius and thearea postrema (Herdegen et al., 1991). Transection of the medialforebrain bundle enhances Zif268 immunostaining in the mammil-lary nucleus, the substantia nigra, and the ventral tegmental area(Leah et al., 1993).
Optic nerve transection appears to induce an increase in Zif268staining in ocular dominance columns, followed later by a de-crease. Other protein levels may change in a similar manner, in-creasing initially in response to injury, and then falling later fromatrophy. This switch may explain why the enucleated eye’s ocular
dominance columns first look dark in dark-field illumination ofwet, unstained sections, and later appear bright.
There is a precedent for contradictory behavior in the stainingpatterns of other activity-related proteins. In macaques, monoculardeprivation causesdarker staining of the alpha isoform of type IIcalcium0calmodulin-dependent protein kinase in the deprived eye’socular dominance columns (Hendry & Kennedy, 1986). By con-trast, the beta isoform of type II calcium0calmodulin-dependent
Fig. 8. Monkey 9 (control). (A) Autoradiograph from the boxed region inFig. 7C. The unlabelled columns of the left eye (arrows) were dark. (B)Same region, from box in Fig. 7B, showing ocular dominance columns indark field. The patterns in A and B matched, proving that after acuteenucleation in normal animals, the dark columns seen in dark field are theocular dominance columns of the enucleated eye. (C) Zif268 immunostain-ing in layer III, from the same region illustrated in A and B, showing oculardominance columns after acute monocular enucleation. There was weakerimmunostaining in the ocular dominance columns of the enucleated lefteye (arrows).
Fig. 9. Monkey 9 (control). (A) Autoradiograph from the right operculumshowing ocular dominance columns in layer IVc. The arrows mark un-labelled columns of the enucleated left eye. (B) Dark-field view of anadjacent CO section showing a matching column pattern. The dark col-umns coincided with the dark columns in A (compare arrows), confirmingthat the enucleated eye’s ocular dominance columns (arrows) appear darkin dark field. (C) Zif268 staining showing ocular dominance columns inlayer III from the same region. On this side, darker immunostaining wasassociated with the ocular dominance columns of the enucleated left eye(compare arrows in A, B, and C). It remains unclear why acute enucleationhad opposite effects on Zif268 immunostaining in the left and right visualcortex of this normal animal.
506 J.C. Horton, D.R. Hocking, and Daniel L. Adams

protein kinase results inlighter staining of the deprived eye’socular dominance columns (Tighilet et al., 1998). Clearly, levels ofsome activity-related proteins decrease following monocular enu-cleation, whereas levels of other activity-related proteins mayincrease.
From a practical standpoint, Zif268 was unsatisfactory as alabel for ocular dominance columns in our strabismus experimentsbecause it was inconsistent. In two animals, we found opposite setsof ocular dominance columns preferentially labelled in differentregions of the same cortex (Table 1). We assume this must beartefactual, but we cannot find a technical error to explain it.Manipulation of tissue during dissection does cause nonspecificbackground staining, but it is hard to see how such artefact wouldlabel opposite sets of ocular dominance columns. One might blamestrabismus for creating local differences in Zif268 levels within thesame cortex, but our 12-h period of dark adaptation prior to mon-ocular enucleation was intended to erase any such gradients. More-over, in a control (Monkey 9) experiment, Zif268 preferentiallylabelled opposite sets of ocular dominance columns on each side.
Another curiosity was that Zif268 did not label preferentiallythe CO patches in striate cortex. After monocular enucleation, ityielded ocular dominance columns rather than light and dark rowsof Zif268-labelled patches in layers II and III (Figs. 8 and 9). Thecontrast and sharpness of the ocular dominance columns was sim-ilar in layer III and layer IVc.
Rapid changes in CO histochemistry after enucleation
Wong-Riley and Carroll (1984) have reported that 24 h followingtetrodotoxin injection into one eye, CO “changes in layer IV weresubtle, with minor fluctuations in staining within lamina IVC”.Later, Wong-Riley (1994) described CO columns only 14 h aftermonocular tetrodotoxin injection. To our knowledge, shorter sur-vival times have never been tested.
Given that the CO patterns induced by strabismus are subtle, itwas crucial for the design of our experiments to prove that short-
term monocular enucleation did not affect them. Unexpectedly, inthree normal monkeys, with survival times of 10, 5.5, and 3 h, wesaw CO columns in layer IVc. The columns were so faint that weoverlooked them initially, leading us to report erroneously in apreliminary abstract that “no CO pattern was seen in light field”following acute monocular enucleation in a normal monkey (Hock-ing et al., 1999). Although faint, the columns were undeniable(Fig. 10A). It appears that CO staining can change within hours ofmonocular enucleation.
This rapid change was surprising, given that the half-life of COis 7–10 days, at least in rat liver (Ip et al., 1974). It should be noted,however, that no one has ever proved that the deposition of di-aminobenzidine reaction product in CO histochemistry correlatesstrictly with CO content in mitochrondria. The CO stain is gener-ally used empirically, without a profound understanding of itsunderlying biochemistry. Wong-Riley and colleagues (1997) haveshown that monocular enucleation induces a 35% drop in COsubunit III mRNA signal in rat lateral geniculate body within 24 h,the shortest time they tested. This finding suggests that reductionof CO expression following monocular enucleation may be quiterapid. Cells may also possess other mechanisms for regulating COactivity, short of enzyme turnover, which may affect CO histo-chemistry on a timescale of hours.
The design of our experiments was predicated on the assump-tion that CO staining would not be affected within 5 h of acutemonocular enucleation. Our control experiments proved this as-sumption wrong. For this reason, we have not presented the dataregarding patterns of CO activity in our stabismic monkeys (exceptto provide Monkey 1 as an example), because the effects of stra-bismus may have been confounded by monocular enucleation.Instead, we have used [3H]proline as a second label (without mon-ocular enucleation) to test how strabismus affects CO staining inmacaque striate cortex. [3H]proline injection, if done carefully,does not alter CO staining in striate cortex. A definitive account ofthose experiments, describing the CO patterns produced by sup-pression scotomas, has appeared elsewhere (Horton et al., 1999).
Fig. 10. Monkey 10 (control). (A) Single section processed for CO histochemistry showing faint ocular dominance columns in layerIVcb (arrows) only 3 h after enucleation of the right eye. (B) Dark-field view of the same section showing absence of columns in layerIVc. Rows of CO patches are visible at the far left in layer V.
Rapid labelling of ocular dominance columns 507

Acknowledgments
This work was supported by the National Eye Institute (Grant RO1 EY10217and Core Crant EY02162), That Man May See, and Research to PreventBlindness. We thank Dr. Douglas R. Fredrick and Dr. Creig S. Hoyt fortheir help in evaluating the animals’ ocular fixation preferences, and RobinTroyer for her assistance with animal care and surgery. We also thank theCalifornia Regional Primate Research Center (especially Jenny Short andDavid Robb). The California Primate Center is supported by NationalInstitutes of Health Base Grant RR00169. Eric P. Hornstein, ChristopherTrepel, and Lynne Kiorpes provided valuable comments on the manuscript.
References
Bravo, R. (1990). Growth factor inducible genes in fibroblasts. InGrowthFactors, Differentiation Factors and Cytokines, ed. Habenicht A.,pp. 324–343. Berlin: Springer.
Bullitt, E. (1990). Expression of c-fos-like protein as a marker for neu-ronal activity following noxious stimulation in the rat.Journal of Com-parative Neurology296, 517–530.
Chaudhuri, A. & Cynader, M.S. (1993). Activity-dependent expressionof the transcription factor Zif268 reveals ocular dominance columns inmonkey visual cortex.Brain Research605, 349–353.
Chaudhuri, A., Matsubara, J.A. & Cynader, M.S. (1995). Neuronalactivity in primate visual cortex assessed by immunostaining for thetranscription factor Zif268.Visual Neuroscience12, 35–50.
Christy, B.A., Lau, L.F. & Nathans, D. (1988). A gene activated inmouse 3T3 cells by serum growth factors encodes a protein with “zincfinger” sequences.Proceedings of the National Academy of Sciences ofthe U.S.A.85, 7857–7861.
Draisci, G. & Iadarola, M.J. (1989). Temporal analysis of increases inc-fos, preprodynorphin and preproenkephalin mRNAs in rat spinal cord.Molecular Brain Research6, 31–37.
Fenstemaker, S.B., George, K.H., Kiorpes, L. & Movshon, J.A. (1997).Tangential organization of eye dominance in area V1 of macaques withstrabismic amblyopia.Society for Neuroscience Abstracts23, 920.16.
Hendry, S.H.C. & Kennedy, M.B. (1986). Immunoreactivity for acalmodulin-dependent protein kinase is selectively increased in ma-caque striate cortex after monocular deprivation.Proceedings of theNational Academy of Sciences of the U.S.A.83, 1536–1540.
Herdegen, T., Walker, T., Leah, J.D., Bravo, R. & Zimmermann, M.(1990). The KROX-24 protein, a new transcription regulating factor:Expression in the rat central nervous system following afferent soma-tosensory stimulation.Neuroscience Letters120, 21–24.
Herdegen, T., Kummer, W., Fiallos, C.E., Leah, J. & Bravo, R. (1991).Expression of c-JUN, JUN B and JUN D proteins in rat nervous systemfollowing transection of vagus nerve and cervical sympathetic trunk.Neuroscience45, 413–422.
Hevner, R.F. & Wong-Riley, M.T.T. (1990). Regulation of cytochromeoxidase protein levels by functional activity in the macaque monkeyvisual system.Journal of Neuroscience10, 1331–1340.
Hocking, D.R., Adams, D.L. & Horton, J.C. (1999). A new method forrapid visualization of ocular dominance columns in macaque striatecortex.Society for Neuroscience Abstracts25, 572.15.
Horton, J.C. (1984). Cytochrome oxidase patches: A new cytoarchitec-tonic feature of monkey visual cortex.Philosophical Transactions ofthe Royal Society B(London)304, 199–253.
Horton, J.C. & Hocking, D.R. (1996a). An adult-like pattern of oculardominance columns in striate cortex of newborn monkeys prior tovisual experience.Journal of Neuroscience16, 1791–1807.
Horton, J.C. & Hocking, D.R. (1996b). Intrinsic variability of oculardominance column periodicity in normal macaque monkeys.Journal ofNeuroscience16, 7228–7239.
Horton, J.C. & Hocking, D.R. (1997). Myelin patterns in V1 and V2 ofnormal and monocularly enucleated monkeys.Cerebral Cortex7,166–177.
Horton, J.C. & Hocking, D.R. (1998a). Experimental strabismus reducesmetabolic activity in ocular dominance columns serving the deviatedeye.Society for Neuroscience Abstracts24, 105.5.
Horton, J.C. & Hocking, D.R. (1998b). Monocular core zones and bin-ocular border strips in primate striate cortex revealed by the contrasting
effects of enucleation, eyelid suture, and retinal laser lesions on cyto-chrome oxidase activity.Journal of Neuroscience18, 5433–5455.
Horton J.C., Hocking, D.R. & Adams, D.L. (1999). Metabolic mappingof suppression scotomas in striate cortex of macaques with experimen-tal strabismus.Journal of Neuroscience19, 7111–7129.
Hunt, S.P., Pini, A. & Evan, G. (1987). Induction of c-fos-like protein inspinal cord neurons following sensory stimulation.Nature328, 632–634.
Ip, M.M., Chee, P.Y. & Swick, R.W. (1974). Turnover of hepatic mito-chondrial ornithine aminotransferase and cytochrome oxidase using[14C]carbonate as tracer.Biochimica et Biophysica Acta354, 29–38.
Leah, J.D., Herdegen, T. & Bravo, R. (1991). Selective expression ofJun proteins following axotomy and axonal transport block in periph-eral nerves in the rat: Evidence for a role in the regeneration process.Brain Research566, 198–207.
Leah, J.D., Herdegen, T., Murashov, A., Dragunow, M. & Bravo, R.(1993). Expression of immediate early gene proteins following axot-omy and inhibition of axonal transport in the rat central nervous sys-tem.Neuroscience57, 53–66.
Lemaire, P., Revelant, O., Bravo, R. & Charnay, P. (1988). Twomouse genes encoding potential transcription factors with identicalDNA-binding domains are activated by growth factors in cultured cells.Proceedings of the National Academy of Sciences of the U.S.A.85,4691–4695.
LeVay, S., Connolly, M., Houde, J. & Van Essen, D.C. (1985). Thecomplete pattern of ocular dominance stripes in the striate cortex andvisual field of the macaque monkey.Journal of Neuroscience5, 486–501.
Rosen, K.M., McCormack, M.A., Villa-Komaroff, L. & Mower, G.D.(1992). Brief visual experience induces immediate early gene expres-sion in the cat visual cortex.Proceedings of the National Academy ofSciences of the U.S.A.89, 5437–5441.
Sagar, S.M., Sharp, F.R. & Curran, T. (1988). Expression of c-fosprotein in brain: Metabolic mapping at the cellular level.Science240,1328–1331.
Sharp, F.R., Griffith, J., Gonzalez, M.F. & Sagar, S.M. (1989). Tri-geminal nerve section induces Fos-like immunoreactivity (FLI) in brain-stem and decreases FLI in sensory cortex.Molecular Brain Research6,217–220.
Sukhatme, V.P., Cao, X., Chang, L.C., Tsai-Morris, C.H., Stamenk-ovich, D., Ferreira, P.C.P., Cohen, D.R., Edwards, S.A., Shows,T.B. & Curran, T. (1988). A zinc finger-encoding gene coregulatedwith c-fos during growth and differentiation, and after cellular depo-larization.Cell 53, 37–43.
Tighilet, B., Hashikawa, T. & Jones, E.G. (1998). Cell- and lamina-specific expression and activity-dependent regulation of type II calcium0calmodulin-dependent protein kinase isoforms in monkey visual cortex.Journal of Neuroscience18, 2129–2146.
Tychsen, L. & Burkhalter, A. (1997). Nasotemporal asymmetries inV1: Ocular dominance columns of infant, adult, and strabismic ma-caque monkeys.Journal of Comparative Neurology388, 32–46.
Wiesel, T.N., Hubel, D.H. & Lam, D.M.K. (1974). Autoradiographicdemonstration of ocular-dominance columns in the monkey striate cor-tex by means of transneuronal transport.Brain Research79, 273–279.
Wong-Riley, M.T.T. (1979). Changes in the visual system of monocularlysutured or enucleated kittens demonstrable with cytochrome oxidasehistochemistry.Brain Research171, 11–28.
Wong-Riley, M.T.T. & Carroll, E.W. (1984). Effect of impulse block-age on cytochrome oxidase activity in monkey visual system.Nature307, 262–264.
Wong-Riley, M.T.T. (1994). Primate visual cortex: Dynamic metabolicorganization and plasticity revealed by cytochrome oxidase. InCere-bral Cortex, ed. Peters, A. & Rockland, K.S., pp. 141–200. NewYork: Plenum Press.
Wong-Riley, M.T.T. (1989). Cytochrome oxidase: An endogenous meta-bolic marker for neuronal activity.Trends in Neurosciences12, 94–101.
Wong-Riley, M.T.T., Mullen, M.A., Huang, Z. & Guyer, C. (1997).Brain cytochrome oxidase subunit complementary DNAs: Isolation,subcloning, sequencing, light and electron microscopicin situ hybrid-ization of transcripts, and regulation by neuronal activity.Neuroscience76, 1035–1055.
Worley, P.F., Cole, A.J., Murphy, T.H., Christy, B.A., Nakabeppu, Y.& Baraban, J.M. (1990). Synaptic regulation of immediate-early genesin brain. Cold Spring Harbor Symposia on Quantitative Biology55,213–223.
508 J.C. Horton, D.R. Hocking, and Daniel L. Adams


















