
Purification and antimicrobial activity studies of the N-terminal fragment of ubiquitin from human...
6
Purification and antimicrobial activity studies of the N-terminal fragment of ubiquitin from human amniotic fluid Jin-Young Kim a,1 , Sun Young Lee a,1 , Seong-Cheol Park a,e , Song Yub Shin a,b , Sang Joon Choi d , Yoonkyung Park a,c, ⁎ , Kyung-Soo Hahm a,b, ⁎ a Research Center for Proteineous Materials (RCPM), Chosun University, Kwangju 501-759, Korea b Department of Cellular·Molecular Medicine School of Medicine, Chosun University, Kwangju 501-759, Korea c Department of Biotechnology and BK21 Research Team for Protein Activity Control, Chosun University, Kwangju 501-759, Korea d Department of Obstetrics and Gynecology, School of Medicine, Chosun University, Kwangju 501-759, Korea e Division of Applied Life Sciences (BK21 program), Gyeongsang National University, Jinju, 660-701, Korea Received 12 May 2007; received in revised form 28 June 2007; accepted 28 June 2007 Available online 10 July 2007 Abstract A 4.3-kDa antimicrobial peptide was isolated from human amniotic fluid by dialysis, ultrafiltration, and C18 reversed-phase high performance liquid chromatography. This peptide, which we named Amniotic Fluid Peptide-1 (AFP-1), possessed antimicrobial activity but lacked hemolytic activity. In addition, AFP-1 potently inhibited the growth of a variety of bacteria (Escherichia coli, Salmonella typhimurium, Listeria monocytogenes and Staphylococcus aureus), filamentous fungi (Botrytis cinerea, Aspergillus fumigatus, Neurospora crassa and Fusarium oxysporum) and yeast cells (Candida albicans and Cryptococcus neoformans). Automated Edman degradation showed that the N-terminal sequence of AFP-1 was NH 2 -Met-Gln-Ile-Phe-Val-Lys-Thr-Leu-Thr-Gly-Lys-Thr-Ile-Thr-Leu-Glu-Val-Glu-. The partial sequence had 100% homology to the N-terminal sequence of ubiquitin. Matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry revealed that the molecular mass of AFP-1 was 4280.2 Da. Our data show an antimicrobial activity of ubiquitin N-terminal derived peptide that makes it suitable for use as an antimicrobial agent. © 2007 Elsevier B.V. All rights reserved. Keywords: Amniotic Fluid Peptide-1 (AFP-1); Antimicrobial activity; Ubiquitin; Antimicrobial agent 1. Introduction The successful development of a healthy fetus is reliant upon normal amniotic fluid (AF). The AF forms a sac around the fetus to protect it from both mechanical and thermal shock. It has some antibacterial activity and may contain some nutritional factors [1]. The AF volume increases from 50 ml at 12 weeks to 1 l at 38 weeks [2]. Among other functions, the strategic location of vernix between the fetal skin surface and the amniotic fluid suggests a potential role in host defense. This is supported by recent reports of antimicrobial proteins present in the vernix caseosa and amniotic fluid [3–5] as well as in pulmonary surfactant [6–8]. However, the integral composition and exact role of AF remain unknown. Vernix reflects intimate maternal and fetal interactions, and during the third trimester, vernix on the skin surface detaches into the amniotic fluid under the putative influence of pulmonary surfactant [9]. This suggests that AF may contain antimicrobial factors [10–15]. Neonatal bacterial infec- tions are a serious cause of morbidity and mortality among newborns. Because the newborn adaptive immunity is immature, antimicrobial peptides as effectors of innate immunity may play a critical role in newborn defense reactions. Ubiquitin (Ub) is a highly conserved small regulatory protein that is ubiquitous in eukaryotes. Ubiquitin was first identified in 1975 as an 8.5-kDa protein of unknown function and was found to be universally expressed in living cells [16–18]. It is recently reported that ubiquitin protein plays an antimicrobial role [19– Biochimica et Biophysica Acta 1774 (2007) 1221 – 1226 www.elsevier.com/locate/bbapap Abbreviations: SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; RP-HPLC, reverse phase-high performance liquid chromato- graphy; BCA, bicinchoninic acid; CFU, colony forming unit ⁎ Corresponding authors. Tel.: +82 62 230 6854; fax: +82 62 227 8345. E-mail addresses: [email protected] (Y. Park), [email protected] (K.-S. Hahm). 1 These authors contributed equally to this work. 1570-9639/$ - see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.bbapap.2007.06.013
-
Upload
jin-young-kim -
Category
Documents
-
view
214 -
download
1
Transcript of Purification and antimicrobial activity studies of the N-terminal fragment of ubiquitin from human...

1774 (2007) 1221–1226www.elsevier.com/locate/bbapap
Biochimica et Biophysica Acta
Purification and antimicrobial activity studies of the N-terminal fragmentof ubiquitin from human amniotic fluid
Jin-Young Kim a,1, Sun Young Lee a,1, Seong-Cheol Park a,e, Song Yub Shin a,b, Sang Joon Choi d,Yoonkyung Park a,c,⁎, Kyung-Soo Hahm a,b,⁎
a Research Center for Proteineous Materials (RCPM), Chosun University, Kwangju 501-759, Koreab Department of Cellular·Molecular Medicine School of Medicine, Chosun University, Kwangju 501-759, Korea
c Department of Biotechnology and BK21 Research Team for Protein Activity Control, Chosun University, Kwangju 501-759, Koread Department of Obstetrics and Gynecology, School of Medicine, Chosun University, Kwangju 501-759, Koreae Division of Applied Life Sciences (BK21 program), Gyeongsang National University, Jinju, 660-701, Korea
Received 12 May 2007; received in revised form 28 June 2007; accepted 28 June 2007Available online 10 July 2007
Abstract
A 4.3-kDa antimicrobial peptide was isolated from human amniotic fluid by dialysis, ultrafiltration, and C18 reversed-phase high performanceliquid chromatography. This peptide, which we named Amniotic Fluid Peptide-1 (AFP-1), possessed antimicrobial activity but lacked hemolyticactivity. In addition, AFP-1 potently inhibited the growth of a variety of bacteria (Escherichia coli, Salmonella typhimurium, Listeriamonocytogenes and Staphylococcus aureus), filamentous fungi (Botrytis cinerea, Aspergillus fumigatus, Neurospora crassa and Fusariumoxysporum) and yeast cells (Candida albicans and Cryptococcus neoformans). Automated Edman degradation showed that the N-terminalsequence of AFP-1 was NH2-Met-Gln-Ile-Phe-Val-Lys-Thr-Leu-Thr-Gly-Lys-Thr-Ile-Thr-Leu-Glu-Val-Glu-. The partial sequence had 100%homology to the N-terminal sequence of ubiquitin. Matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometryrevealed that the molecular mass of AFP-1 was 4280.2 Da. Our data show an antimicrobial activity of ubiquitin N-terminal derived peptide thatmakes it suitable for use as an antimicrobial agent.© 2007 Elsevier B.V. All rights reserved.
Keywords: Amniotic Fluid Peptide-1 (AFP-1); Antimicrobial activity; Ubiquitin; Antimicrobial agent
1. Introduction
The successful development of a healthy fetus is reliant uponnormal amniotic fluid (AF). The AF forms a sac around the fetusto protect it from both mechanical and thermal shock. It has someantibacterial activity and may contain some nutritional factors [1].The AF volume increases from 50 ml at 12 weeks to 1 l at38 weeks [2]. Among other functions, the strategic location ofvernix between the fetal skin surface and the amniotic fluidsuggests a potential role in host defense. This is supported by
Abbreviations: SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gelelectrophoresis; RP-HPLC, reverse phase-high performance liquid chromato-graphy; BCA, bicinchoninic acid; CFU, colony forming unit⁎ Corresponding authors. Tel.: +82 62 230 6854; fax: +82 62 227 8345.E-mail addresses: [email protected] (Y. Park),
[email protected] (K.-S. Hahm).1 These authors contributed equally to this work.
1570-9639/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.bbapap.2007.06.013
recent reports of antimicrobial proteins present in the vernixcaseosa and amniotic fluid [3–5] as well as in pulmonarysurfactant [6–8]. However, the integral composition and exactrole of AF remain unknown. Vernix reflects intimatematernal andfetal interactions, and during the third trimester, vernix on the skinsurface detaches into the amniotic fluid under the putativeinfluence of pulmonary surfactant [9]. This suggests that AF maycontain antimicrobial factors [10–15]. Neonatal bacterial infec-tions are a serious cause of morbidity and mortality amongnewborns. Because the newborn adaptive immunity is immature,antimicrobial peptides as effectors of innate immunity may play acritical role in newborn defense reactions.
Ubiquitin (Ub) is a highly conserved small regulatory proteinthat is ubiquitous in eukaryotes. Ubiquitin was first identified in1975 as an 8.5-kDa protein of unknown function and was foundto be universally expressed in living cells [16–18]. It is recentlyreported that ubiquitin protein plays an antimicrobial role [19–

Fig. 1. Elution profile of AFP-1. (A) The fraction containing the major retainedpeak was separated by reversed-phase HPLC using a C18 column. (B) Thepurified antimicrobial peptide was then subjected again to C18 reversed-phaseHPLC. In (A and B), the peptides were eluted using a gradient of 5–65%acetonitrile for 60 min at a flow rate of 1 ml/min. To obtain highly purifiedhomogenous proteins, peak fractions with antibacterial activity were re-subjected to an RP-C18 column (5 μm, 300 Å, 2.1×150 mm, Vydac, Hesperia,CA) with a slower flow rate of 0.2 ml/min. The effluent from the column wasmonitored by measuring the absorbance at 214 nm.
Fig. 2. Tricine gel electrophoresis of purified AFP-1. Lane 1: molecular sizemarker (26.6, Triosephosphate isomerase; 17, Myoglobin; 14.2, α-Lactalbumin;6.5, Aprotinin; 3.5, Insulin chain B; 1, Bradykinin). Lane 2: AFP-1.
1222 J.-Y. Kim et al. / Biochimica et Biophysica Acta 1774 (2007) 1221–1226
23] in the living organisms. Ubiquitin conjugates to intracellulartarget proteins, in a process that appears to be the initiation stepfor selective protein degradation by the 26S proteasome andserves as a signal for the endocytosis of plasma membraneproteins [24]. It is now clear that ubiquitin can act as a regulatedsorting signal at different steps in the endosomal and bio-synthetic pathways [25].
Here, in our studies using screening methods to assessantimicrobial activity, we report the N-terminal fragment ofubiquitin in human amniotic fluid. We describe the purificationand characterization of Amniotic Fluid Peptide (AFP-1), anantimicrobial peptide corresponding to the N-terminalsequence of ubiquitin, from human amniotic fluid. AFP-1inhibits human pathogenic fungal and bacterial strains, and itshows sequence homology with the N-terminal sequence ofubiquitin. Finally, we determined the molecular mass of AFP-1by mass spectrometry.
2. Materials and methods
2.1. Materials
2.1.1. Amniotic fluidsAmniotic fluids were obtained from ten pregnant women in their third
trimester who underwent trans-abdominal amniocentesis in order to achieve
fetal lung maturity. All amniotic fluid samples selected for purification andanalysis were free of contamination from blood components and amnioticcells. The amniotic fluid samples were immediately centrifuged and thesupernatants were then filtered with a disposable membrane filter (poresize=0.45 μm, Millifore) for sterilization, aliquoted and stored at −80 °C untilneeded.
2.1.2. Microbial strainsBotrytis cinerea (KACC 40573) and Neurospora crassa (KACC 40996)
were obtained from the Korean Agricultural Culture Collection, and Aspergillusfumigates (ATCC 9197), Candida albicans (ATCC 18804), Escherichia coli(ATCC 25922), Listeria monocytogenes (ATCC 19115) and Staphylococcusaureus (ATCC 25923) were obtained from American Type Culture Collection.Fusarium oxysporum (KCTC 16909), Cryptococcus neoformans (KCTC7003) and Salmonella typhimurium (KCTC 1926) were obtained from KoreanCollection for Type Cultures.
2.2. Purification and characterization of antimicrobial proteins
The amniotic fluids were dialyzed against sodium acetate buffer (10 mM, pH6.0) to remove urine and salts. The dialyzed samples were then passed throughan ultrafiltration membrane with molecular-mass cutoffs of 10 kDa. Aliquots ofthe ultrafiltrates were subjected to an RP-C18 column (5 μm, 300 Å,4.6×250 mm, Vydac, Hesperia, CA) and separated with a gradient of 5–65%acetonitrile for 60 min at a flow rate of 1 ml/min. Each fraction was dried undervacuum, resuspended in potassium phosphate buffer (10 mM, pH 7.4) andassayed for antibacterial activity. To obtain highly purified homogenousproteins, the peak fractions with antibacterial activity were re-subjected to anRP-C18 column (5 μm, 300 Å, 2.1×150 mm, Vydac, Hesperia, CA) with aslower flow rate of 0.2 ml/min. The purity of the purified peptide was analyzed

Table 1Antimicrobial activities of purified protein against microbial strains
Pathogens Protein (MIC) a (μM)
Filamentous fungiBotrytis cinerea 16Aspergillus fumigatus 64Neurospora crassa 32Fusarium oxysporum 16Yeast cellsCandida albicans 8Cryptococcus neoformans 8BacteriaEscherichia coli 32Salmonella typhimurium 32Listeria monocytogenes 8Staphylococcus aureus 8
The bacteria was suspended (final concentration=2×105 colony formationunits/ml) in 100 μl of growth medium and mixed with 100 μl of the test peptidesolution in a microtiter plate well. Each condition was tested in triplicate.Microbial growth was assessed by measuring the increase in optical densityat 620 nm after 10-h incubation at 37 °C. The fungal cells (1×104 CFU/ml)were seeded in 100 μl of potato dextrose broth per well in 96-well micro-titer plates, mixed with 10 μl of the serially diluted peptide solution, andincubated for 24 h at 28 °C. The turbidity of each well was measured at570 nm using an Emax microtiter plate reader. All assays were performed intriplicate.a MIC is the minimal concentration inhibiting bacterial and fungal growth.
1223J.-Y. Kim et al. / Biochimica et Biophysica Acta 1774 (2007) 1221–1226
by 16.5% Tricine SDS-PAGE and analytical reversed-phase HPLC. Proteinconcentration was determined using a BCA protein assay (Pierce, Rockford)with BSA as a standard.
2.3. Antibacterial activity
The bacteria were grown to mid-logarithmic phase in a medium containing10 g/l bactotryptone, 5 g/l yeast extract, and 10 g/l NaCl at pH 7.0. Thepeptide was serially diluted in 1% bactopeptone. The test organism wassuspended (final concentration=2×105 colony formation units/ml) in 100 μlof growth medium and mixed with 100 μl of the test peptide solution in amicrotiter plate well. Each condition was tested in triplicate. Microbial growthwas assessed by measuring the increase in optical density at 620 nm after 10-hincubation at 37 °C.
Fig. 3. Effect of the peptide on the viability of the C. albicans (A), S. aureus (B) andcounts after 24 h incubation.
2.4. Antifungal activity
The fungal strains were grown at 28 °C in Potato Dextrose Broth (PDB)medium. The fungal cells (1×104 CFU/ml) were seeded in 100 μl of potatodextrose broth per well in 96-well microtiter plates, mixed with 10 μl of theserially diluted peptide solution, and incubated for 24 h at 28 °C. Ten μl of a 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) solu-tion [5 mg/ml MTT in phosphate-buffered saline (PBS), pH 7.4] was added toeach well, and the plates were incubated further at 37 °C for 4 h. Thirtymicroliters of 20% (w/v) sodium dodecyl sulfate (SDS) containing 0.02 M HClwas then added, and the plates were incubated at 37 °C for 16 h in order todissolve the formazan crystals that had formed [26]. The turbidity of each wellwas measured at 570 nm using an Emax microtiter plate reader (MolecularDevices Emax, California, USA). All assays were performed in triplicate.
2.5. Antimicrobial activity assay by CFU
Two-fold serial dilutions of each peptide, ranging from 1 to 64 μM, werearranged in sterile 96-well plates, and then an aliquot of cell suspension wasadded to each well. The cell counts were 1×104 CFU (colony forming units)/mland 2×105 CFU/ml in bacteria and fungi, respectively. The plates wereincubated for 2 h at 28 °C for fungal cell and 37 °C for bacterial cells. At the endof the incubation, 50 μl of 20-fold diluted samples were plated on appropriateagar plates, depending on the microorganism tested. Plates were incubated for24 h, and then colonies were counted [27].
2.6. Preparation of human red blood cells and hemolytic activity assay
Human red blood cells were centrifuged and washed three times withphosphate-buffered saline (PBS; 35 mM phosphate buffer with 0.15 M NaCl,pH 7.0). The hemolytic activities of the AFP-1 and melittin (positive control)were evaluated by measuring the amount of hemoglobin released from freshhuman erythrocytes. Aliquots (100 μl) of an 8% suspension of red blood cellswere transferred to 96-well plates, and hemolysis was determined by measuringthe absorbance at 414 nm with an Emax plate reader. No hemolysis (0%) wasobserved in the presence of PBS, while full hemolysis (100%) was observed inthe presence of 0.1% Triton X-100. The percent hemolysis was calculated usingthe following equation: % hemolysis= [(A414 nm with peptide solution−A414 nm
in PBS) / (A414 nm with 0.1% Triton-X 100−A414 nm in PBS)]×100 [28].
2.7. Amino acid sequencing of the isolated peptide
The N-terminal amino acid sequence of the purified protein was analyzed bymeans of automated Edman degradation on a pulse liquid automatic sequencer
L. monocytogenes (C). The peptide concentrations are plotted against the viable
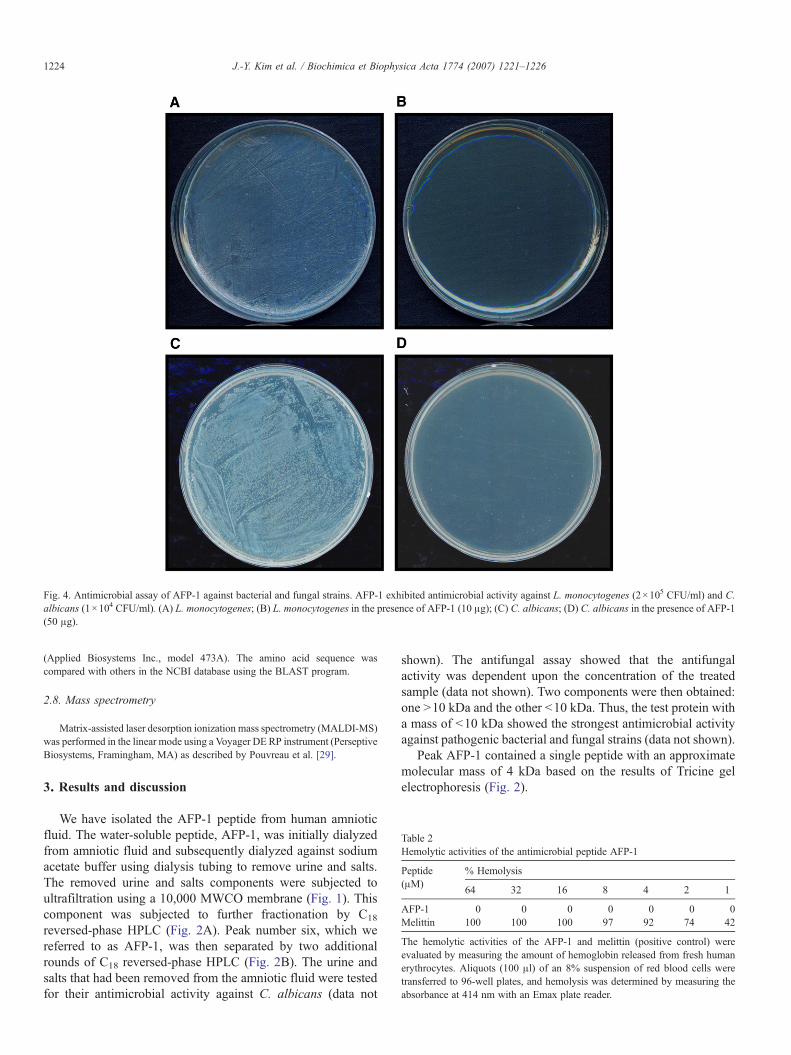
Fig. 4. Antimicrobial assay of AFP-1 against bacterial and fungal strains. AFP-1 exhibited antimicrobial activity against L. monocytogenes (2×105 CFU/ml) and C.albicans (1×104 CFU/ml). (A) L. monocytogenes; (B) L. monocytogenes in the presence of AFP-1 (10 μg); (C) C. albicans; (D) C. albicans in the presence of AFP-1(50 μg).
Table 2Hemolytic activities of the antimicrobial peptide AFP-1
Peptide(μM)
% Hemolysis
64 32 16 8 4 2 1
AFP-1 0 0 0 0 0 0 0Melittin 100 100 100 97 92 74 42
The hemolytic activities of the AFP-1 and melittin (positive control) wereevaluated by measuring the amount of hemoglobin released from fresh humanerythrocytes. Aliquots (100 μl) of an 8% suspension of red blood cells weretransferred to 96-well plates, and hemolysis was determined by measuring theabsorbance at 414 nm with an Emax plate reader.
1224 J.-Y. Kim et al. / Biochimica et Biophysica Acta 1774 (2007) 1221–1226
(Applied Biosystems Inc., model 473A). The amino acid sequence wascompared with others in the NCBI database using the BLAST program.
2.8. Mass spectrometry
Matrix-assisted laser desorption ionization mass spectrometry (MALDI-MS)was performed in the linear mode using a Voyager DE RP instrument (PerseptiveBiosystems, Framingham, MA) as described by Pouvreau et al. [29].
3. Results and discussion
We have isolated the AFP-1 peptide from human amnioticfluid. The water-soluble peptide, AFP-1, was initially dialyzedfrom amniotic fluid and subsequently dialyzed against sodiumacetate buffer using dialysis tubing to remove urine and salts.The removed urine and salts components were subjected toultrafiltration using a 10,000 MWCO membrane (Fig. 1). Thiscomponent was subjected to further fractionation by C18
reversed-phase HPLC (Fig. 2A). Peak number six, which wereferred to as AFP-1, was then separated by two additionalrounds of C18 reversed-phase HPLC (Fig. 2B). The urine andsalts that had been removed from the amniotic fluid were testedfor their antimicrobial activity against C. albicans (data not
shown). The antifungal assay showed that the antifungalactivity was dependent upon the concentration of the treatedsample (data not shown). Two components were then obtained:one N10 kDa and the other b10 kDa. Thus, the test protein witha mass of b10 kDa showed the strongest antimicrobial activityagainst pathogenic bacterial and fungal strains (data not shown).
Peak AFP-1 contained a single peptide with an approximatemolecular mass of 4 kDa based on the results of Tricine gelelectrophoresis (Fig. 2).

Fig. 6. MALDI-MS determination of AFP-1. Matrix-assisted laser desorptionionization mass spectrometry (MALDI-MS) was performed in the linear modeusing a Voyager DE RP.
1225J.-Y. Kim et al. / Biochimica et Biophysica Acta 1774 (2007) 1221–1226
We next examined the antimicrobial activity of the purifiedAFP-1 peptide against various human and plant pathogenicmicrobial cells by assessing the minimum inhibition concentra-tion of AFP-1 (Table 1). As shown in Table 1, AFP-1 exhibitedantimicrobial activity against the human pathogenic filamen-tous fungi B. cinerea, Aspergillus fumigatus, N. crassa and F.oxysporum; the human pathogenic yeast cells C. albicans andC. neoformans; and the human pathogenic bacterial strains E.coli, S. typhimurium, L. monocytogenes and S. aureus. Also,AFP-1 induced a decrease in viable counts of all the strainstested to the limit of detection when incubated for 24 h (Fig. 3).The microbicidal effect of AFP-1 was further visualized bytreating L. monocytogenes and C. albicans with AFP-1 andthen spreading the cells on agar plates. As shown in Fig. 4,AFP-1 inhibited the growth of L. monocytogens (Fig. 4A and B)and C. albicans (Fig. 4C and D).
We next examined the cytotoxicity of AFP-1 against mam-malian cells by measuring the lysis of human erythrocytes.AFP-1 had no hemolytic activity, while the melittin, used as apositive control for lysis, was strongly hemolytic (Table 2).These results demonstrated that AFP-1 possesses remarkableantimicrobial activity against various microbial cells but has nohemolytic activity.
Amino acid sequencing revealed that AFP-1 has an N-terminal amino acid sequence of NH2-Met-Gln-Ile-Phe-Val-Lys-Thr-Leu-Thr-Gly-Lys-Thr-Ile-Thr-Leu-Glu-Val-Glu-. Wefound that this sequence was 100% identical to that of ubiquitin(Fig. 5), which was purified and sequenced by Valueva et al.[30].
MALDI-MS (Fig. 6) showed that AFP-1 has a molecularmass of 4280.2 Da. This is in close agreement with the relativemolecular weight calculated from the protein sequence ofAFP-1 (4280.2).
Ubiquitin consists of 76 amino acids and has a molecularmass of about 8500 Da. It is highly conserved among eukaryoticspecies: human and yeast ubiquitin share 96% sequence identity[17,18]. The primary function of ubiquitin is to mark otherproteins for destruction via a process known as proteolysis.Several ubiquitin molecules attach to the condemned protein(polyubiquitination), and the polyubiquitinated protein is thensubjected to the proteasome, a barrel-shaped structure whereproteolysis occurs. Ubiquitin can also target transmembraneproteins (for example, receptors) for removal from membranes,and it plays multiple roles in cellular signaling. The key featuresof the ubiquitin are its C-terminal tail and lysine residues [31,32].
Ubiquitin and lipopolysaccharide were found to synergisti-cally increase the production of tumor necrosis factor α inmurine macrophage cell line RAW 264, suggesting thatubiquitin plays a role in the modulatory mechanisms of theimmune response [33].
Fig. 5. N-terminal amino acid sequence of AFP-1. Amino acids identical to the N-acid sequence of the purified protein was analyzed by means of automated Edmanprogram.
Ubiquitin was also shown to possess antimicrobial propertiesin a recent report. An antibacterial ubiquitin-derived peptidewas isolated from the bladder of a rat in a previous report [34].Several ubiquitin-derived peptides have been tested in order tocharacterize the structural features implicated in the antimicro-bial properties of ubiquitin. The C-terminal peptide Ub65-76displays antifungal activities at ìM range [19]. This cationicpeptide (charge +3) includes the tail of the last four residues ofubiquitin, Leu73, Arg74, Gly75, and Gly76, and is in the formof a β-strand [35]. It interacts with the cell wall, crosses thefungal membrane, and accumulates in microorganisms.
The sequence of AFP-1 was determined to be the N-terminalsequence of ubiquitin and AFP-1 exhibits antimicrobial activityagainst various pathogens. In this study, we examined theantifungal activity of AFP-1 against the filamentous plantpathogens B. cinerea and F. oxysporum, the filamentous humanpathogen A. fumigates and the yeast C. albicans, which is themost common cause of oral, esophageal, vaginal, and urinarycandidiasis [36]. We also showed that AFP-1 inhibitedfoodborne filamentous fungus N. crassa and the foodbornebacterial pathogen L. monocytogenes, which is a Shiga toxin-producing bacterium [37].
Therefore, AFP-1, the N-terminal partial sequence ofubiquitin, is a potent antimicrobial peptide with activity againsta broad-spectrum of pathogens, and it may have potential as alead compound for the development of therapeutics oragrochemicals.
Acknowledgements
This work was supported by grants from the Ministry ofScience and Technology, Korea and the Korea Science andEngineering Foundation through the Research Center for
terminal sequence of ubiquitin are indicated by boxes. The N-terminal aminodegradation and compared with others in the NCBI database using the BLAST

1226 J.-Y. Kim et al. / Biochimica et Biophysica Acta 1774 (2007) 1221–1226
Proteineous Materials and was supported (in part) by researchfunds from Chosun University, 2007.
References
[1] P. Axemo, E. Rwamushaija, M. Pettersson, L. Eriksson, S. Bergstrom,Amniotic fluid antibacterial activity and nutritional parameters in termMozambican and Swedish pregnant women, Gynecol. Obstet. Invest. 42(1996) 24–27.
[2] D.R. Ostergard, The physiology and clinical importance of amniotic fluid,a review, Obs. Gyn. Surv. 25 (1970) 297–319.
[3] J. Espinoza, T. chaiworapongsa, R. Romero, S. Edwin, C. Rathnasabapathy,R. Gomex, E. Bujold, N. Camacho, Y.M. Kim, S. Hassan, S. Blackwell,J. Whitty, S. Berman, M. Redman, B.H. Yoon, Y. sorokin, Antimicrobialpeptides in amniotic fluid: defensins calprotectin and bacterial/perme-ability-increasing protein in patients with microbial invasion of theamniotic cavity, intra-amniotic inflammation, preterm labor and pre-mature rupture of membranes, J. Matern.-Fetal Neonatal Med. 13 (2003)2–21.
[4] V. Narendran, W. Hull, H. Akinbi, J. Whitsett, W. Pickens, D. Lambers,et al., Vernix caseosa contains surfactant proteins: potential role in innateimmune function in the fetus, Pediatr. Res. 47 (2000) 420.
[5] H. Yoshio, M. Tollin, G.H. Gudmundsson, H. Lagercrantz, H. Jonrvall,G. Marchini, B. Agerberth, Antimicrobial polypeptides of born innatedefense, Pediatr. Res. 53 (2003) 211–216.
[6] A.M. LeVine, J.A. Whitsett, Pulmonary collectins and innate host defenseof the lung, Microbes. Infect. 3 (2001) 161–166.
[7] M.I.B. Bautista, R.R. Wickett, M.O. Visscher, W.L. Pickens, S.B. Hoath,Characterization of vernix caseosa as a natural biofilm: comparison tostandard oil-based ointments, Pediatr. Dermatol. 17 (2000) 253–260.
[8] F.A. Okah, R.R. Wickett, K. Pompa, S.B. Hoath, Human newborn skin: theeffect of isopropanol on skin surface hydrophobicity, Pediatr. Res. 35(1994) 443–446.
[9] V. Narendran, R.R. Wickett, W.L. Pickens, S.B. Hoath, Interactionbetween pulmonary surfactant and vernix: a potential mechanism forinduction of amniotic fluid turbidity, Pediatr. Res. 48 (2000) 120–124.
[10] T.M. Scane, D.F. Hawkins, Antibacterial activity in human amniotic fluid:relationship to zinc and phosphate, Br. J. Obstet. Gynaecol. 91 (1984)342–348.
[11] M.A. Ismail, G.I. Salti, A.H. Moawad, The effect of filtration of fluid onthe growth of Chlamydia trachomatis and Escherichia coli, Am. J.Perinatol. 8 (1991) 50–52.
[12] P. Axemo, E. Rwamushaija, M. Pettersson, L. Eriksson, S. Bergstrom,Amniotic fluid antibacterial activity and nutritional parameters in termMozambican and Swedish pregnant women, Gynecol. Obstet. Invest. 42(1996) 24–27.
[13] M.A. Nazir, G.A. Pankuch, J.J. Botti, P.C. Appelbaum, Antibacterialactivity of amniotic fluid in the early third trimester. Its association withpreterm labor and delivery, Am. J. Perinatol. 4 (1987) 59–62.
[14] K. Otsuki, A. Yoda, H. Saito, Y. Mitsuhashi, Y. Toma, Y. Shimizu,T. Yanaihara, Amniotic fluid lactoferrin in intrauterine infection, Placenta20 (1999) 175–179.
[15] M. Koyama, S. Ito, A. Nakajima, K. Shimoya, C. Azuma, N. Suehara,Y. Murata, H. Tojo, Elevations of group II phospholipase A2 concentra-tions in serum and amniotic fluid in association with preterm labor, Am. J.Obstet. Gynecol. 183 (2000) 1537–1543.
[16] D.H. Schlesinger, G. Goldstein, H.D. Niall, The complete aminoacid sequence of ubiquitin, an adenylate cyclase stimulating poly-peptide probably universal in living cells, Biochemistry 14 (1975)2214–2218.
[17] D.H. Schlesinger, G. Goldstein, Molecular conservation of 74 amino acid
sequence of ubiquitin between cattle and man, Nature 255 (1975)423–424.
[18] M.J. Cox, R. Shapira, K.D. Wilkinson, Tryptic peptide mapping ofubiquitin and derivatives using reverse-phase high performance liquidchromatography, Anal. Biochem. 154 (1986) 345–352.
[19] A.E. Kieffer, Y. Goumon, O. Ruh, S. Chasserot-Golaz, G. Nullans,C. Gasnier, D. Aunis, M.H. Metz-Boutigue, The N- and C-terminalfragments of ubiquitin are important for the antimicrobial activities,FASEB J. 17 (2003) 776–778.
[20] S.J. Howell, D. Wilk, S.P. Yadav, C.L. Bevins, Antimicrobial polypeptidesof the human colonic epithelium, Peptides 24 (2003) 1763–1770.
[21] I. Svensson, K. Calles, E. Lindskog, H. Henriksson, U. Eriksson,L. Häggström, Antimicrobial activity of conditioned medium fractionsfrom Spodoptera frugiperda Sf9 and Trichoplusia in Hi5 insect cells,Appl. Microbiol. Biotechnol. 69 (2005) 92–98.
[22] Y. Wang, W.J. Griffiths, H. Jörnvall, B. Agerberth, J. Johansson,Antibacterial peptides in stimulated human granulocytes: characterizationof ubiquitinated histone H1A, Eur. J. Biochem. 269 (2002) 512–518.
[23] S. Alonso, K. Pethe, D.G. Russell, G.E. Purdy, Lysosomal killing ofMycobacterium mediated by ubiquitin-derived peptides is enhanced byautophagy, Proc. Natl. Acad. Sci. U. S. A. 104 (2007) 6031–6036.
[24] L. Hicke, Protein regulation by monoubiquitin, Nat. Rev., Mol. Cell Biol. 2(2001) 195–201.
[25] L. Hicke, A new ticket for entry into budding vesicles-ubiquitin, Cell 106(2001) 527–530.
[26] Y. Park, D.G. Lee, P.I. Kim, E.-R. Woo, G.-W. Cheong, C.-H. Choi, K.-S.Hahm, A Leu-Lys-rich antimicrobial peptide: activity and mechanism,Biochim. Biophys. Acta 1645 (2003) 172–182.
[27] O. Marchetti, P. Moreillon, M.P. Glauser, J. Bille, D. Sanglard, Potentsynergism of the combination of fluconazole and cyclosporine in Candidaalbicans, Antimicrob. Agents Chemother. 44 (2000) 2373–2381.
[28] Y. Park, S.N. Park, S.-C. Park, J.Y. Park, Y.H. Park, J.S. Hahm, K.-S.Hahm, Antibiotic activity and synergistic effect of antimicrobial peptideagainst pathogens from a patient with gallstones, Biochem. Biophys. Res.Commun. 321 (2004) 631–637.
[29] L. Pouvreau, H. Gruppen, S.R. Piersma, L.A.M. van den Broek, G.A. vanKoningsveld, A.G.J. Voragen, Relative abundance and inhibitorydistribution of protease inhibitors in potato juice from cv. Elkana,J. Agric. Food Chem. 49 (2001) 2864–2874.
[30] T.A. Valueva, T.A. Revina, V.V. Mosolov, R. Mentele, Primary structure ofpotato Kunitz-type serine proteinase inhibitor, Biol. Chem. Hoppe-Seyler381 (2000) 1215–1221.
[31] M. Hochstrasser, Ubiquitin-dependent protein degradation, Annu. Rev.Genet. 30 (1996) 405–439.
[32] A.M. Weissman, Themes and variations on ubiquitylation, Nat. Rev., Mol.Cell Biol. 2 (2001) 169–178.
[33] T. Nabika, M. Terashima, I. Momose, Y. Hosokawa, N. Nagasue,Y. Tanigawa, Synergistic effect of ubiquitin on lipopolysaccharide-induced TNF-alpha production in murine macrophage cell line RAW 264.7cells, Biochim. Biophys. Acta 1450 (1999) 25–34.
[34] B. Wang, S.S. Harwig, R.I. Lehrer, Rat bladder ubiquitin-like molecule:isolation, purification and N-terminal sequencing, Hua. Xi. Yi. Ke. Da.Xue. Xue. Bao. 24 (1993) 127–130.
[35] S. Vijay-Kumar, C.E. Bugg, K.D. Wilkinson, W.J. Cook, Three-dimensional structure of ubiquitin at 2.8 A resolution, Proc. Natl. Acad.Sci. U. S. A. 82 (1985) 3582–3585.
[36] A. Benchekroun, M. Alami, M. Ghadouan, A. Lachkar, H. Kasmaoui,M. Marzouk, M. Faik, Urinary candidiasis revealed by ureteralobstruction: report of 2 cases, Ann. Urol. 34 (2000) 171–174.
[37] P.Gerner-Smidt,K.Hise, J.Kincaid, S.Hunter, S.Rolando,E.Hyytia-Trees,E.M. Ribot, B. Swaminathan, Pulsenet taskforce, pulsenet USA: a five-year update, Foodborne, Pathog. Dis. 3 (2006) 9–19.







![Ubiquitin and Ubiquitin-like Modifications in Viral ...1].pdf · Ubiquitin and Ubiquitin-like Modifications in Viral Infection and Immunity Abstracts of papers presented at the AUGUST](https://static.fdocuments.in/doc/165x107/5e2d68ba2a69b505b71e58fa/ubiquitin-and-ubiquitin-like-modifications-in-viral-1pdf-ubiquitin-and-ubiquitin-like.jpg)










