
PTHrP regulates growth plate chondrocyte differentiation and proliferation in a Gli3 dependent...
12
PTHrP regulates growth plate chondrocyte differentiation and proliferation in a Gli3 dependent manner utilizing hedgehog ligand dependent and independent mechanisms Elaine Mau a,b,1 , Heather Whetstone a,1 , Chunying Yu a , Sevan Hopyan a,b,c,e , Jay S. Wunder b,c,d , Benjamin A. Alman a,b,c,e, ⁎ a The Program in Developmental Biology of the Hospital for Sick Children, Toronto, Ontario, Canada b The Institute of Medical Sciences, University of Toronto, Canada c Department of Surgery, University of Toronto, Canada d The Orthopaedic Oncology Unit of the Mount Sinai Hospital, Toronto, Ontario, Canada e The Division of Orthopaedic Surgery of the Hospital for Sick Children, Toronto, Ontario, Canada Received for publication 30 March 2006; revised 11 January 2007; accepted 23 January 2007 Available online 27 January 2007 Abstract Growth plate chondrocytes undergo a tightly regulated process of differentiation, allowing for the longitudinal growth of bones. Although it is known that parathyroid hormone related protein (PTHrP) and Indian hedgehog regulate the differentiation of growth plate chondrocytes, how these pathways interact to regulate chondrocyte development is not fully elucidated. We examined how the interaction between PTHrP and the hedgehog activated transcription factors, Gli2 and Gli3, regulates growth plate chondrocyte differentiation and proliferation. Analysis of fetal limbs showed that Gli2 is a negative regulator and Gli3 a positive regulator of type X collagen expression. Limb explant cultures showed that PTHrP treatment inhibited type X collagen expression and increased chondrocyte proliferation. This effect was substantially enhanced in Gli2-/- limbs, was blocked in Gli3-/- limbs, and was only partially inhibited by hedgehog ligand blockade. PTHrP negatively regulated Gli mediated transcription in cell cultures, and regulated the level of the repressor form of Gli3 in a PKA dependent manner. These results show that PTHrP regulates growth plate chondrocyte proliferation and differentiation in part through the activity of Gli3, suggesting a crucial role for Gli3 in growth plate chondrocyte development. © 2007 Elsevier Inc. All rights reserved. Keywords: Hedgehog; Growth plate; PTHrP; Gli; Organ culture; Proliferation; Differentiation Introduction The long bones of the body form by enchondral ossification, a process in which a cartilage is replaced by osseous tissue. During embryonic development, mesenchymal progenitor cells condense to form a cartilaginous template of the long bones. Blood vessels and osteoblasts appear in the middle of the cartilage to form a primary center of ossification. The chondrocytes at the end of the bone differentiate to form articular chondrocytes that ultimately form the joint lining surface, and growth plate chondrocytes that are located towards the center of the bone. A secondary ossification center forms between these two populations of chondrocytes. The growth plate is responsible for the longitudinal growth of the bone, and growth plate chondrocytes undergo an orderly process of differentiation, in which resting cells proliferate, hypertrophy, and undergo terminal differentiation, during which they lay the scaffolding for new bone formation (Linsenmayer et al., 1988; Olsen et al., 2000). A variety of signaling pathways are crucial for normal growth plate chondrocyte differentiation, including parathyroid hormone related protein (PTHrP) and Indian hedgehog (Ihh). These two signaling pathways act in a feedback loop regulating Developmental Biology 305 (2007) 28 – 39 www.elsevier.com/locate/ydbio ⁎ Corresponding author. 555 University Avenue, Toronto, Ontario, Canada M5G1X8. Fax: +1 416 813 6414. E-mail address: [email protected] (B.A. Alman). 1 Contributed equally to this work. 0012-1606/$ - see front matter © 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.ydbio.2007.01.031
-
Upload
elaine-mau -
Category
Documents
-
view
213 -
download
0
Transcript of PTHrP regulates growth plate chondrocyte differentiation and proliferation in a Gli3 dependent...

305 (2007) 28–39www.elsevier.com/locate/ydbio
Developmental Biology
PTHrP regulates growth plate chondrocyte differentiation and proliferation ina Gli3 dependent manner utilizing hedgehog ligand dependent and
independent mechanisms
Elaine Mau a,b,1, Heather Whetstone a,1, Chunying Yu a, Sevan Hopyan a,b,c,e,Jay S. Wunder b,c,d, Benjamin A. Alman a,b,c,e,⁎
a The Program in Developmental Biology of the Hospital for Sick Children, Toronto, Ontario, Canadab The Institute of Medical Sciences, University of Toronto, Canada
c Department of Surgery, University of Toronto, Canadad The Orthopaedic Oncology Unit of the Mount Sinai Hospital, Toronto, Ontario, Canada
e The Division of Orthopaedic Surgery of the Hospital for Sick Children, Toronto, Ontario, Canada
Received for publication 30 March 2006; revised 11 January 2007; accepted 23 January 2007Available online 27 January 2007
Abstract
Growth plate chondrocytes undergo a tightly regulated process of differentiation, allowing for the longitudinal growth of bones. Although it isknown that parathyroid hormone related protein (PTHrP) and Indian hedgehog regulate the differentiation of growth plate chondrocytes, howthese pathways interact to regulate chondrocyte development is not fully elucidated. We examined how the interaction between PTHrP and thehedgehog activated transcription factors, Gli2 and Gli3, regulates growth plate chondrocyte differentiation and proliferation. Analysis of fetallimbs showed that Gli2 is a negative regulator and Gli3 a positive regulator of type X collagen expression. Limb explant cultures showed thatPTHrP treatment inhibited type X collagen expression and increased chondrocyte proliferation. This effect was substantially enhanced in Gli2−/−limbs, was blocked in Gli3−/− limbs, and was only partially inhibited by hedgehog ligand blockade. PTHrP negatively regulated Gli mediatedtranscription in cell cultures, and regulated the level of the repressor form of Gli3 in a PKA dependent manner. These results show that PTHrPregulates growth plate chondrocyte proliferation and differentiation in part through the activity of Gli3, suggesting a crucial role for Gli3 in growthplate chondrocyte development.© 2007 Elsevier Inc. All rights reserved.
Keywords: Hedgehog; Growth plate; PTHrP; Gli; Organ culture; Proliferation; Differentiation
Introduction
The long bones of the body form by enchondral ossification,a process in which a cartilage is replaced by osseous tissue.During embryonic development, mesenchymal progenitor cellscondense to form a cartilaginous template of the long bones.Blood vessels and osteoblasts appear in the middle of thecartilage to form a primary center of ossification. Thechondrocytes at the end of the bone differentiate to form
⁎ Corresponding author. 555 University Avenue, Toronto, Ontario, CanadaM5G1X8. Fax: +1 416 813 6414.
E-mail address: [email protected] (B.A. Alman).1 Contributed equally to this work.
0012-1606/$ - see front matter © 2007 Elsevier Inc. All rights reserved.doi:10.1016/j.ydbio.2007.01.031
articular chondrocytes that ultimately form the joint liningsurface, and growth plate chondrocytes that are located towardsthe center of the bone. A secondary ossification center formsbetween these two populations of chondrocytes. The growthplate is responsible for the longitudinal growth of the bone, andgrowth plate chondrocytes undergo an orderly process ofdifferentiation, in which resting cells proliferate, hypertrophy,and undergo terminal differentiation, during which they lay thescaffolding for new bone formation (Linsenmayer et al., 1988;Olsen et al., 2000).
A variety of signaling pathways are crucial for normalgrowth plate chondrocyte differentiation, including parathyroidhormone related protein (PTHrP) and Indian hedgehog (Ihh).These two signaling pathways act in a feedback loop regulating

29E. Mau et al. / Developmental Biology 305 (2007) 28–39
growth plate chondrocyte differentiation, with PTHrP stimula-tion acting to delay terminal differentiation (Karp et al., 2000;Kobayashi et al., 2002; Lanske et al., 1996; Minina et al., 2001).PTHrP signals though PTHR1, which is a 7 transmembranetype II G-protein coupled receptor. Ligand binding activatesseveral secondary messenger pathway including PKA, IP3, andPKC (Gensure et al., 2005). Studies in-vitro suggest that PKAmediates much of the effect of PTHrP on chondrocytematuration (Zhang et al., 2003). The importance of PTHR1 ingrowth plate function is highlighted by the demonstration thatmutations in this receptor cause metaphyseal chondrodyspla-sias, conditions that are characterized by abnormal growth platechondrocyte maturation (Schipani et al., 1996).
Hedgehog (Hh) ligands bind to the Patched (Ptch1) receptor.This binding inactivates smoothened inhibition, which results inhedgehog mediated transcriptional factor(s) becoming active inthe nucleus. In Drosophila, hedgehog signaling activates thetranscription factor cubitus interruptus (Ci). In the absence ofhedgehog binding, Ci is tethered to microtubules where it iscleaved, resulting in a portion that acts as a transcriptionalrepressor. With hedgehog receptor activation, unprocessed Cifunctions as a transcriptional activator. In mammals, there arethree transcription factors: Gli1, Gli2 and Gli3. The regulationof the Gli transcription factors by hedgehog is more complex.Gli1 has a transcriptional activating domain, while Gli2 andGli3 contain both transcriptional activating and repressingdomains. Although this raises the possibility that Gli1 is aconstitutively active, mice deficient in Gli1 show no readilyapparent phenotype, suggesting that this factor is not requiredfor normal physiologic function. Gli2 and Gli3 transcriptionalactivation are regulated by hedgehog signaling, however, thecontrol of transcriptional activation does not appear to be asimple as for Ci. Hedgehog signaling inhibits the formation ofthe Gli3 repressor form but apparently not that of Gli2. This,and findings from mice deficient in Gli2 or Gli3, led to thenotion that Gli2 has acts primarily to activate hedgehogmediated transcription, while Gli3 can act as either a repressoror activator depending on the cell specific context (Cohen,2003; Dominguez et al., 1996; Matise and Joyner, 1999; Mulloret al., 2002; Nieuwenhuis and Hui, 2005). Interestingly, Glimediated transcription results in elevation of Gli1 and Ptch1transcripts, potentially acting as a transcriptional feedback loopin hedgehog signaling activation (Johnson et al., 1995; Marigoand Tabin, 1996; Schipani et al., 1997). Besides their regulationby hedgehog ligand signaling, the Gli proteins may also beregulated by other signaling pathways. For example, Gli2 andGli3, are implicated in fibroblast growth factor signaling inmesodermal development in the frog (Brewster et al., 2000).This raises the possibility that Gli transcription factors act asintegrators of multiple signaling pathways.
Given the important role of hedgehog signaling in bonedevelopment, it is not surprising that mice deficient in Gli2develop substantially abnormal bones. Gli2−/− mice are notviable, but have been investigated during embryonic develop-ment. They show an increase in the length of cartilaginousgrowth plates, increased numbers of hypertrophic growth platechondrocyte cells, and reduced bone formation. Theses changes
were accompanied by a decreased expression of Ptch1 andPTHrP, consistent with decreased hedgehog transcriptionalactivation (Miao et al., 2004). Genetic studies in mice show thatGli3 acts as an important transcription factor regulating theeffect of Ihh on growth plate function. Deficiency of Gli3 willrescue the phenotype imparted by Ihh deficiency, giving supportto the notion that Gli3 acts as a negative regulator of Hhmediated signaling in growth plate chondrocytes (Koziel et al.,2005).
Phosphorylation is required in the processing and transcrip-tional activation of Ci, and PKA is one of a variety of factorswhich plays a role in this phosphorylation. PKA activation willalso inhibit Gli mediated transcription (Epstein et al., 1996;Riobo et al., 2006). As such, PKA regulation by PTHrP mightdirectly influence Gli mediated transcription, even in theabsence of hedgehog ligand. In this study, we examine howPTHrP regulates the Gli transcriptional activation and the effecton terminal differentiation in growth plate chondrocytes.
Methods
Mice
The extra toes mutant mouse, which harbors heterozygous inactivatingmutation in Gli3 (Hui and Joyner, 1993), was used to generate fetal Gli3−/−mice. Homozygous mice are not viable, but will develop past embryonic day18.5. To investigate the role of Gli2 deficiency in growth plate function, a mouseharboring a targeted deletion in Gli2 was utilized (Mo et al., 1997). Like theextra toes mutant mouse, homozygous Gli2 deficient mice are not viable, butwill develop past embryonic day 18.5. Embryonic mice were obtained fromtimed pregnancies and the genotype of the various mice determined aspreviously reported (Hui and Joyner, 1993; Mo et al., 1997). In all cases,littermate wild-type mice were used as controls.
Cell lines and Gli transcriptional activation analysis
Mesenchymal progenitor cell lines, C2C12 and C3H10T1/2, were used toanalyze Gli transcriptional activation, as these cells exhibit some properties ofgrowth plate chondrocytes (Denker et al., 1995; Gu et al., 2004). These cellslines expressed PTHR1 at only very low levels, and for this reason, we generatedcell lines stably transfected with a Flag-tagged PTHR1. Epitope mapping studieshave previously shown minimal alterations in PTHR1 signaling due to theaddition of a Flag tag (Xie and Abou-Samra, 1998). We used PCR to tag aPTHR1-pcDNA1/AMP construct (obtained from Dr. H. Juppner, MassachusettsGeneral Hospital, Boston) with a Flag epitope. In-vitro transcription translationwas used to verify that this construct would produce a protein of predicted size.Fugene-6 (Roche) was used for transfections, and antibiotic suppression (G418,Multicell), was used to select cells that had been successfully transected. Toverify successful transfection of PTHR1, immunofluorescent staining to the Flagepitope was utilized. Cells were grown on coverslips, rinse twice with sterilePBS, fixed in 4% paraformaldehyde, followed methanol. The sections werestained using an anti-Flag mouse monoclonal M2 from Sigma followed bysecondary fluorescent dye-conjugated antibody (DAKO). To determine ifPTHrP could regulate Gli mediated signaling with constitutive activation ofhedgehog signaling, we utilized mouse embryonic fibroblatsts derived fromPtch1−/− mice (Goodrich et al., 1997; Bailey et al., 2002).
The cell cultures were examined for Gli transcriptional activity by transienttransfection with a Gli-reporter construct, in which the Gli promoter consensussequence 5′-GACCACCCA-3′ regulates expression of luciferase protein(Sasaki et al., 1997), and using real time PCR for the hedgehog target gene,Ptch1, and Gapdh as a control. The Gli reporter construct was transfectedalong with a β-galactosidase expression construct as a control for transfectionefficiency. In order to determine the effect of hedgehog and PTHrP signaling onGli activation, the cell cultures were examined under a variety of treatment

30 E. Mau et al. / Developmental Biology 305 (2007) 28–39
conditions, including the use of the PKA-specific inhibitor, H-89 dihydrochlor-ide (Alexis Biochemicals), at 20 μM; the hedgehog ligand, Shh-N at 10 μg/ml(Ontogeny Inc., Cambridge, Massachusetts, USA); 10−7 M PTHrP (Bachem,King of Prussia, Pennsylvania, USA); the hedgehog inhibitor cyclopamine at10−4 M (Biomol Research Laboratories, Inc., Plymouth Meeting PA); thecyclopamine analogue lacking hedgehog blocking activity, tomatidine, as anadditional control at 10−4 M (Sigma, St. Louis, Missouri, USA); or 10 μg/mlneutralizing 5E1 anti-hedgehog monoclonal antibody (Hybridoma Bank,University of Iowa).
β-Galactosidase activity and luciferase activity were measured as previouslyreported (Meng et al., 2001), and Gli transcriptional activity was represented byluciferase activity normalized for β-galactosidase. To assay for PKA activity anddetermine effective inhibition at the H-89 dosages used in our studies, 5 μl ofcell culture lysate was assayed using a commercially available PKA activityassay kit (Promega Biosciences, San Luis Obispo, CA). Each experiment wasperformed at least in triplicate. The results obtained using cyclopamine wereverified using the neutralizing antibody.
Explant cultures
Embryonic hindlimb organ cultures from 16.5 dpc limbs were established aspreviously described (Lanske et al., 1996; Minina et al., 2001). Limbs wereplaced on a Nucleopore filter inside a well of a 24-well tissue culture plate, withthe limb at the liquid/air interface. The limb explants were then cultured for4 days at 37 °C in a 5% CO2 incubator, initially in media containing 5% fetal calfserum. After 1 day in culture, the media was changed to 0.1% FBS, with variousagents added to the media. 10−7 M PTHrP (Bachem, King of Prussia,Pennsylvania, USA) was added to activate PTHrP signaling. To block hedgehogligand mediated singling, cyclopamine was utilized at 10−4 M concentration. Asa control for cyclopamine, an analogue which lacks hedgehog signalingproperties, tomatidine (Sigma, St. Louis, Missouri, USA) was utilized (Tiet etal., 2006). Each explant experiment was undertaken six times using limbs fromsix different fetal mice and littermate controls. As an additional control forblocking hedgehog ligand, 10 μg/ml of neutralizing 5E1 anti-SHH monoclonalantibody (Hybridoma Bank, University of Iowa) was utilized in a single explantculture to verify the results obtained using cyclopamine.
Proliferation was measured using Bromdeoxyuridine (BrdU) incorporation.Explant specimens were incubated with BrdU for the final 12 h of culture. Ananti-BrdU antibody was used as previously reported to detect cells which hadtaken up BrdU during this 12 h period (Alman et al., 1996). The proportion ofBrdU positive cells was calculated in the proliferative zone of the growth plateby counting the number of positive and negative cells over 10 high poweredfields. Apoptosis was measured using a TUNEL assay as previously reported(Tiet et al., 2006). Positively stained cells were analyzed in a similar manner asfor BrdU incorporation.
Immunohistochemistry
Fetal limbs at embryonic day 16.5 or 18.5, or explant cultures wereexamined for expression of Gli2, Gli3, and type X collagen by immunohis-tochemistry using previously reported techniques and antibodies (Hu et al.,2006; Linsenmayer et al., 1988; Saika et al., 2004; Tiet et al., 2006; Wang et al.,2000). Type X collagen is an exclusive marker of growth plate chondrocyteterminal differentiation, and was used as a marker for hypertrophic growth platechondrocytes. Since growth plate morphology varies with the anatomic location,we used the proximal tibial growth plate for all analyses.
Western analysis for Gli3 processing
Nuclear extracts were prepared from cell cultures using centrifugation aspreviously reported (Meng et al., 2001). A Gli3 antibody (H-280 antibody fromSanta Cruz) that recognizes both the full length and truncated form of the proteinwas used in Western analysis to determine expression level of the Gli3 repressor(Hu et al., 2006; Saika et al., 2004; Wang et al., 2000). The membranes for theWestern were stripped and probed using an antibody to actin as a loadingcontrol. Densitometry was used to compare protein levels between the fulllength activator and truncated repressor forms of Gli3.
Statistical analysis
In the explant studies, at least six limbs from different mice of a givengenotype were examined, along with six limbs from wild-type littermates ascontrols. For cell culture experiments, each was performed at least intriplicate. Sections for immunohistochemistry, TUNEL staining, and BrdUimportation studies were observed under light microscopy. The percent BrdUincorporation, percent TUNEL stained cells, or the height of the zone of thegrowth plate staining for type X collagen was measured by a blinded observer.For all of the data, the mean, 95% confidence intervals, and standarddeviations were calculated for each condition. These were compared using theStudent t-test.
Results
Expression of Gli proteins in the growth plate
Fetal limbs from wild-type, Gli2−/−, and Gli3−/− mice wereanalyzed for expression of the Gli2 and Gli3 transcriptionfactors using immunohistochemistry. The proximal tibialgrowth plates from E16.5 limbs were examined. In wild-typemice, there was diffuse expression of Gli2 throughout thegrowth plate, although there was slightly less expression interminally differentiating, hypertrophic, growth plate chondro-cytes (Fig. 1A). In contrast, Gli3 was expressed at substantiallylower levels in the hypertrophic zone (Fig. 1D). Gli2 expressionwas lacking in limbs from Gli2 null mice, and expression ofGli3 was lacking in mice null for Gli3 (Figs. 1C and F).Although there was not an obvious change in the distribution ofGli2 in Gli3−/− mice, in Gli2−/− mice, there was an extendedrange of expression of Gli3, extending into the hypertrophiczone (Figs. 1B and E). This expression pattern of Gli3 is similarto that reported in earlier stage embryonic limbs using in-situhybridization (Koziel et al., 2005), with the transcription factorprimarily expressed by cells which have not yet entered thehypertrophic zone.
Type X collagen expression in growth plate chondrocytes isregulated by the Gli genes
To determine what role Gli genes might play in theregulation of growth plate chondrocyte differentiation, theregion of expression of type X collagen, a marker ofhypertrophic growth plate chondrocytes, was comparedbetween mutant and wild type fetal mice at 16.5 and18.5 dpc. In 16.5 dpc mice there was a decreased height tothe region of type X collagen expression in the Gli3−/− micecompared to wild-type littermates, and an increased height tothe region of type X collagen expression in Gli2−/− micecompared to wild-type littermates (Fig. 2). These differenceswere, however, much more dramatic in the 18.5 dpc limbscompared to 16.5 dpc limbs. This pattern suggests a positiverole for Gli3 and a negative role for Gli2 in the regulation of theheight size of the zone of type X collagen expression in thegrowth plate. The more dramatic difference in the slightly olderfetal limbs illustrates that the regulation of hypertrophicchondrocytes by Gli genes in the growth plate depends on thestage of development.

Fig. 1. Expression of Gli proteins in the growth plate. Immunohistochemistry for Gli2 (panels A, B and C), and Gli3 (panels D, E, and F). The proximal tibial growthplate from E16.5 wild-type, Gli3−/−, and Gli2−/− limbs were examined. In wild-type mice (panel A), there was diffuse expression of Gli2 throughout the growthplate, although slightly less expression in terminally differentiating, hypertrophic growth plate chondrocytes. In Gli3−/−mice, there was a slightly elevated expressionlevel in chondrocytes in the zone of terminal differentiation (panel B). There was no expression in growth plates from Gli2−/−mice (panel C). In wild-type mice, Gli3was expressed at lower levels in the zone of terminal differentiation (panel D). Gli2−/−mice showed an extended range of expression of Gli3, extending into the zoneof terminal differentiation (panel E). Gli3−/− mice lacked expression of Gli3 in their growth plates (panel F).
31E. Mau et al. / Developmental Biology 305 (2007) 28–39
PTHrP treatment inhibits hedgehog mediated transcription incell lines
Cell cultures derived from mesenchymal progenitors wereused to investigate the regulation of Gli mediated transcrip-tion by PTHrP. Because these cell lines did not expressPTHR1, we generated cells transfected with a constructdriving expression of the receptor. The ability of the constructto produce PTHR1 protein was confirmed using an in-vitrotranscription translation assay (Fig. 3A), and the expression incells was determined using immunohistochemistry to the Flagtag (Fig. 3B). The regulation of PTHR1 mediated signalingby PTHrP was assayed by measuring PKA activity. Theability to regulate this activity was abolished with the additionof PKA-specific inhibitor, H-89 (Figs. 3C and D). In both celllines, there was little regulation of PKA activity in theabsence of transfection with PTHR1, and treatment with H-89brought PKA levels to baseline. PKA activity was alsoincreased in the Ptch1−/− mouse embryonic fibroblasts withPTHrP treatment, showing that these cells are responsive toPTHrP (Fig. 3E).
To determine the regulation of PTHrP on Gli mediatedtranscription, the cells were assayed for activity of a Gli reporterconstruct and expression level of the hedgehog target gene,
Ptch1. Activity was compared between stimulation withhedgehog ligand, with the hedgehog signaling inhibitorcyclopamine, with PTHrP, and the PKA inhibitor H-89. Doseresponse studies were undertaken to ensure that these agentswere able to completely block the effect of hedgehog ligandstimulation. As expected, hedgehog ligand stimulationincreased Gli transcriptional activation. PTHrP, on the otherhand, decreased Gli mediated transcription to roughly 50% ofcontrol levels. Intriguingly, PTHrP was able to cause thisdecrease even in the presence of hedgehog ligand activation orhedgehog signaling blockade by cyclopamine (Figs. 4A and B).The data from the use of the neutralizing antibody was identicalto that with use of cyclopamine (data not shown). Thesedifferences were significant with a p<0.05. Real time RT-PCRdata confirmed that the level of expression of Ptch1 correlatedwith Gli transcriptional activation level (data not shown). Theeffect of PTHrP stimulation on Gli transcriptional activationwas inhibited by treatment with the PKA inhibitor. This effectwas noted not only in the C2C12 and C3H10T1/2 cell lines, butalso in the Ptch1−/− mouse embryonic fibroblasts (Fig. 4C).Since the Ptch1−/− cells exhibit constitutive hedgehog signal-ing activation, there was a high baseline to the level of Glitranscriptional activation, but PTHrP treatment was still able todecrease the level of Gli transcriptional activation. These

Fig. 2. Expression of type X collagen in the growth plate of Gli mutant mice. Immunohistochemistry using an antibody to type X collagen was utilized to compare theheight of the zone of hypertrophic growth plate chondrocytes between Gli2−/−, Gli3−/−, wild type littermate fetal mice at 16.5 and 18.5 dpc. There was a decreasedheight to the region of type X collagen expression in the Gli3−/− mice compared to wild-type littermates, and an increased height to the region of type X collagenexpression in Gli2−/− mice compared to wild-type littermates. The bar in the figures is 1 mm long. The means and 95 confidence intervals for the height of the zonestaining for type X collagen (cm) are shown in graphic form. A line with an asterisk over it indicates a statistically significantly difference (p<0.05) in the height of thetype X collagen zone compared to the wild-type limbs.
32 E. Mau et al. / Developmental Biology 305 (2007) 28–39
findings demonstrate an inhibitory role for PTHrP in theregulation of Gli mediated transcription that is at least partiallyindependent of the regulation of hedgehog ligand, anddependent on PKA.
PTHrP stimulation regulates type X collagen expression ingrowth plate chondrocytes in a Gli dependent manner
To determine how PTHrP regulates growth plate chondro-cyte differentiation in the various Gli deficient limbs, weestablished fetal limb organ cultures, and compared the effectof PTHrP and hedgehog blockade to treatment with carrier orcontrols (tomatidine, a cyclopamine analogue that lacks theability to block hedgehog ligand mediated signaling). Asexpected, in wild type mice PTHrP treatment resulted in adecline in the size of the area of type X collagen staining (Figs.5A through C). Surprisingly, in Gli2−/− mice there was asubstantially exaggerated effect of PTHrP on type X collagen
staining, while in Gli3−/− mice the effect was substantiallyblunted (Fig. 5I though L and Q through T). These resultssuggest that Gli3 and Gli2 act opposite each other in theregulation of growth plate chondrocyte differentiation and thelack of a change in the size of the zone of terminaldifferentiation in Gli3−/− limbs implicates Gli3 as thepredominant transcription factor regulating the effect ofPTHrP on type X collagen expression.
In the presence of hedgehog blockade with cyclopamine,PTHrP was still able to exert an effect on the size of the zoneof cells expressing type X collagen. However, the effect ofPTHrP was blunted compared to tomatidine treated controls(Fig. 5E through H and M through P). In Gli3−/− mice, therewas little effect of PTHrP stimulation on type X collagenstaining in the presence of hedgehog blockade (Fig. 5Uthrough X). Taken together, these results suggest that Gli3plays an important role in how PTHrP regulates the zone oftype X collagen expression, and that PTHrP can regulate

Fig. 3. PTHrP regulation of Gli mediated transcription. An in-vitro transcription translation assay (panel A) showed that the Flag tagged construct produced PTHR1protein of predicted size. Immunofluorescence using an anti-flag antibody showed that the cells were efficiently transfected with the construct, a representativephotomicrograph from C3H10T1/2 cells is shown (panel B). PTHrP regulated PKA activity, which was blocked by the PKA-specific inhibitor, H-89. Means and 95%confidence intervals for PKA activity under different conditions are given in panel C, D, and E for the C2C12, C3H10T1/2 cell lines, and Ptch1−/− cells respectively.PTHrP regulates PKA activity in C2C12 and C3H10T1/2 cell lines expressing PTHR1 (pcDNA is cells transfected with an empty vector as a control), as well as in thecells from Ptch1−/− mice.
33E. Mau et al. / Developmental Biology 305 (2007) 28–39
growth plate chondrocyte differentiation in a hedgehog ligandindependent manner.
PTHrP regulates growth plate chondrocyte proliferation in aGli3 dependent manner
Since PTHrP and hedgehog signaling also regulate growthplate chondrocyte proliferation, we examined limb explantcultures to determine how PTHrP and the Gli genes interact inthe regulation of growth plate chondrocyte proliferation. Weinvestigated the bromodeoxyuridine (BrdU) incorporation ratein limb explants treated with PTHrP or carrier. There was ahigher rate of proliferation in Gli3−/− mice compared toGli2−/− mice, suggesting that the level of hedgehog transcrip-tional activation regulates growth plate chondrocyte prolifera-tion (Figs. 6A, C, and E). We found an increased BrdUincorporation rate with PTHrP treatment in wild-type limbs. Incontrast, the increase in proliferation rate was substantiallygreater in Gli2−/− mice, while we found little change inGli3−/−mice (Figs. 6B, D, and F). Taken together with our dataon the interaction of PTHrP and Gli in regulating the size of thezone of type X collagen expression in the growth plate, thissuggests a tight coordination between growth plate chondrocyteproliferation and differentiation, regulated at least in part by Glitranscriptional activation.
In contrast to our data on proliferation and type X collagenstaining, we saw only small numbers of TUNEL positive cellsin the explants and there was no change in the proportion ofpositively stained cells under any of the treatment conditions.
PTHrP stimulation regulates Gli3 processing
To determine how PTHrP regulates Gli3 activity in theabsence of hedgehog blockade, we examined our cell culturesfor the level of expression of Gli3 in its full length activator, andtruncated repressor form. Since our data suggests that PTHrPregulates Gli transcriptional activity and growth plate chon-drocytes terminal differentiation even in the presence ofhedgehog receptor inhibition, we examined the ability ofPTHrP to regulate Gli3 processing independent of hedgehogligand. Using Western analysis, we determined the relativeamount of Gli3 that is present in the repressor form in nuclearfractions compared to the full length activator form. We foundthat PTHrP treatment increased the amount of Gli3 repressorform present in the C3HT101/2, Ptch1−/−, and C2C12 celllines (Fig. 7). In the presence of hedgehog inhibition, PTHrPtreatment was still able to regulate the amount of GLI3 presentas the inhibitory form. Hedgehog treatment substantiallyreduced the amount of Gli3 repressor present, and PKAblockade, using H-89, abolished the ability of PTHrP to

Fig. 4. PTHrP regulation of Gli transcriptional activity. Data from the C2C12 cell are shown in panel A, and from the C3H10T1/2 cell line in panel B. Glitranscriptional activity in the presence of cyclopamine (panels B and C). Means for relative Gli activity and 95% confidence intervals for different cell cultureconditions are shown. PTHrP has an effect of Gli activity even in the presence of hedgehog signaling blockade or hedgehog ligand stimulation. Data in panel C fromthe C2C12 cell line, the C3H10T1/2 cell line, and from Ptch1−/− cells shows that the downregulation of Gli transcriptional activation, which is present in all three celllines, is dependent on PKA.
34 E. Mau et al. / Developmental Biology 305 (2007) 28–39
regulate the Gli3 repressor. This demonstrates a hedgehogligand independent process by which PTHrP can process Gli3that is dependent on PKA regulation.
Discussion
PTHrP and Hedgehog signaling play crucial roles regulatinggrowth plate chondrocyte differentiation. Genetic and biochem-ical studies show that these pathways regulate each other, butthe mechanism by which this regulation occurs has not beencompletely elucidated. In our study, we examined interactionsbetween PTHrP and the hedgehog mediated transcriptionfactors Gli2 and Gli3. PTHrP is known to inhibit growth platechondrocyte terminal differentiation and regulate chondrocyteproliferation (Harrington et al., 2004; Kobayashi et al., 2002).We found that the ability of PTHrP to regulate growth platechondrocyte proliferation and differentiation depends on the Gligenes present. Intriguingly, this regulation was only partiallydependent on hedgehog ligand activity, identifying a hedgehogligand independent mechanism by which PTHrP directlyregulates Gli activity.
Our data supports a crucial role for the Gli transcriptionfactors in the regulation of chondrocyte differentiation andproliferation. This is not unexpected, given the role of Ihh in theregulation of growth plate chondrocytes, and previous data onthe Gli genes in skeletal development (Miao et al., 2004; Hilton
et al., 2005; Koziel et al., 2005; Mo et al., 1997). Gli2 and Gli3often act as positive and negative regulators of hedgehogmediated signaling respectively (Mo et al., 1997; Motoyama etal., 2003). Since mice deficient in Gli2 have a larger zone ofexpression of type X collagen, and mice deficient in Gli3 have asmaller zone of expression of type X collagen, these transcrip-tion factors act in opposing ways to regulate growth platechondrocyte differentiation. In addition, organ explant culturesfrom Gli2−/− mice have a lower proliferation rate than organcultures from wild type or Gli3−/− mice. Thus Gli2 acts toinhibit differentiation and promote proliferation, while Gli3 actsin the opposite manner.
The expression pattern of the Gli2 and Gli3 suggest that theyplay a role in the chondrocytes prior to entering the hypertrophiczone. This finding, and the negative relationship between theproliferation rate and the size of the zone of terminaldifferentiation, provides further evidence that proliferation anddifferentiation are tightly coupled in the growth plate.Furthermore, the effect of Gli2 and Gli3 deficiency on terminaldifferentiation is dependent on the age of development, with amore predominant effect in 18.5 dpc limbs than in 16.5 dpclimbs. This finding is consistent with the notion that growthplate chondrocytes are regulated by the Gli genes in both atemporal and special manner.
The exaggerated effect of PTHrP treatment on terminaldifferentiation and proliferation in Gli2−/− mice, in concert

Fig. 5. PTHrP regulation of type X collagen in growth plate chondrocytes. Fetal limb organ cultures from Gi2−/−, Gli3−/−, or wild type littermates were treated withcyclopamine, tomatidine (cyclopamine analogue that lacks the ability to block hedgehog ligand mediated signaling), PTHrP, or carrier. The brown color is stained fortype X collagen. The whole tibia is shown in panels A, C, E, G, I, K, M, O, Q, S, U, and W, while higher magnification views of the proximal tibial growth plate isshown in panels B, D, F, H, J, L, N, P, R, T, W, and X. The bar in these figures is 0.01 mm long. PTHrP treatment resulted in a decline in the size of the zone of type Xcollagen expression in wild-type mice (panels A through D). Gli2−/− showed a substantially exaggerated effect of PTHrP on type X collagen expression, while inGli3−/−mice the effect was substantially blunted (panels I though L and Q though T). In the presence of hedgehog blockade with cyclopamine, PTHrP was still able toexert an effect on the size of the zone of cells expressing type X collagen. However, the effect of PTHrP was blunted compared to tomatidine treated controls (panels Ethough H and M though P). In Gli3−/− mice, there was no effect of PTHrP stimulation on terminal differentiation in the presence of hedgehog blockade (panels Uthough X). The mean and 95% confidence intervals for the height of the zone of type X collagen staining from the proximal tibial growth plate is shown in the graph inpanel Y (length in mm). A line with an asterisk above it indicates a statistically significant difference (p<0/05). The full tibia and proximal tibial growth platephotomicrographs are taken from different explant cultures.
35E. Mau et al. / Developmental Biology 305 (2007) 28–39

Fig. 6. PTHrP regulates growth plate chondrocyte proliferation in a Gli3dependent manner. Bromodeoxyuridine (BrdU) incorporation in Gli3−/−,Gli2−/−, and wild-type littermate explant limb cultures. There were a higherproportion of cells incorporating BrdU in Gli3−/− mice compared to Gli2−/−mice (panels A, C, and E). The ability of PTHrP to increase the BrdUincorporation rate was enhanced inGli2−/− limbs (panels B and D), while it wassubstantially blunted in Gli3−/− limbs (panel F). Graphs show the mean and95% confidence intervals for the percent of cells in the proliferative zone of thegrowth plate that stained for BrdU. A line with an asterisk above it indicates astatistically significant difference (p<0/05).
36 E. Mau et al. / Developmental Biology 305 (2007) 28–39
with the blunted effect in Gli3−/− mice suggests that thePTHrP mediated effect on differentiation is regulated by Gli-3. Gli3 can be act as either a repressor or activator oftranscription, while Gli2 seems to function primarily as atranscriptional activator. PTHrP treatment alters the amount
of Gli3 repressor in the nucleus through ligand dependentand independent processes. By decreasing the expression ofhedgehog ligands, the cells decrease their Gli mediatedtranscriptional activation through regulation of hedgehog'smembrane receptor. However, we also found a ligandindependent regulation that is mediated by PKA. Theincrease in PKA level associated with PTHrP signalingalso plays a role directly regulating the processing of Gli3.As such, the PTHrP regulation of differentiation through Glimediated transcription likely functions through regulation ofGli3, with its cleavage changing it from a transcriptionalactivator to a repressor.
The interpretation of the results of any study examininggenetically modified mice can be confounded by the geneticchange altering the population of cells present in the tissuesexamined. While the possibility that the changes we observedin our explant experiments is due to a shift in cell populationscan not be completely ruled out, the complimentary findingsfrom the various mutant mice makes this possibility lesslikely.
Besides functioning in growth plate development, PTHrPplays an important role in other physiologic and pathologicprocesses. It is possible that it could affect such processes bymodulating hedgehog mediated transcription, in either thepresence or absence of hedgehog ligand. For instance, thereis improved survival in patients with breast cancersexpressing PTHrP (Henderson et al., 2001, 2006). Further-more, breast cancers exhibit evidence of activated hedgehogsignaling, and hedgehog signaling blockade inhibits growthin breast cancer cell lines (Kubo et al., 2004). It is possiblethat PTHrP imparts its effects on breast cancer by regulatingHedgehog signaling in a similar manner as it does in growthplate chondrocytes.
Several studies found that PTHrP and Ihh have differingeffects on different chondrocyte populations (Deckelbaum etal., 2002; Iwasaki et al., 1997; Kindblom et al., 2002;Kobayashi et al., 2002, 2005; Lanske et al., 1996; Yoshida etal., 2001). Our data showing that PTHrP exerts its regulation ofdifferentiation through Gli3, and that Gli3 expression is limitedto only select growth plate chondrocytes, explains the restrictionof the effect of PTHrP to only select populations of growth platechondrocytes. In addition, the mild compensatory increase inthe region of growth plate chondrocytes expressing Gli3 in micedeficient in Gli2, may partially explain the exaggeratedphenotypic effect observed from PTHrP treatment in the Gli2mutant mice.
We previously found that transgenic mice over-expressingGli2 in the growth plate develop rests of chondrocytes in themetaphysic that fail to undergo terminal differentiation (Tiet etal., 2006). These metaphyseal cartilage rests are similar inappearance to a condition in humans in which benign cartilagetumours are found in the metaphysis, called enchondroma-tosis. The tumours that occur in this condition are associatedwith hedgehog signaling activation (Tiet et al., 2006). Ourcurrent findings are consistent with our supposition that Gli2activation in the transgenic mice inhibits terminal andpromotes chondrocyte proliferation. Taken together with our
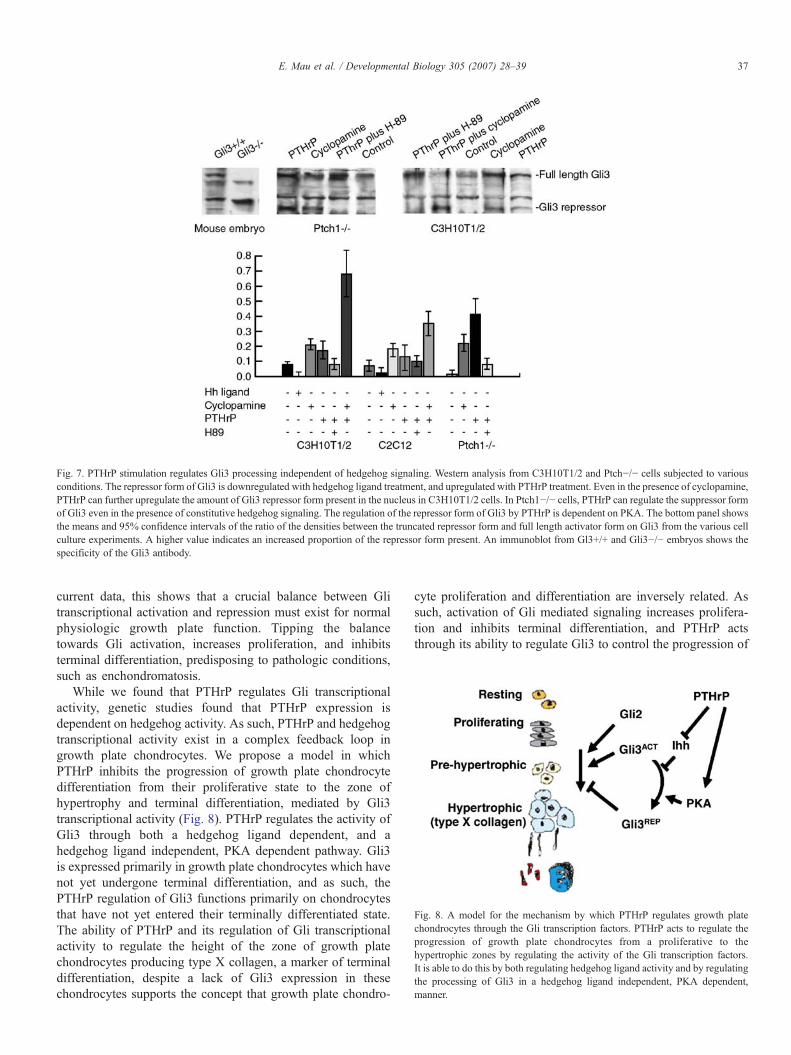
Fig. 7. PTHrP stimulation regulates Gli3 processing independent of hedgehog signaling. Western analysis from C3H10T1/2 and Ptch−/− cells subjected to variousconditions. The repressor form of Gli3 is downregulated with hedgehog ligand treatment, and upregulated with PTHrP treatment. Even in the presence of cyclopamine,PTHrP can further upregulate the amount of Gli3 repressor form present in the nucleus in C3H10T1/2 cells. In Ptch1−/− cells, PTHrP can regulate the suppressor formof Gli3 even in the presence of constitutive hedgehog signaling. The regulation of the repressor form of Gli3 by PTHrP is dependent on PKA. The bottom panel showsthe means and 95% confidence intervals of the ratio of the densities between the truncated repressor form and full length activator form on Gli3 from the various cellculture experiments. A higher value indicates an increased proportion of the repressor form present. An immunoblot from Gl3+/+ and Gli3−/− embryos shows thespecificity of the Gli3 antibody.
Fig. 8. A model for the mechanism by which PTHrP regulates growth platechondrocytes through the Gli transcription factors. PTHrP acts to regulate theprogression of growth plate chondrocytes from a proliferative to thehypertrophic zones by regulating the activity of the Gli transcription factors.It is able to do this by both regulating hedgehog ligand activity and by regulatingthe processing of Gli3 in a hedgehog ligand independent, PKA dependent,manner.
37E. Mau et al. / Developmental Biology 305 (2007) 28–39
current data, this shows that a crucial balance between Glitranscriptional activation and repression must exist for normalphysiologic growth plate function. Tipping the balancetowards Gli activation, increases proliferation, and inhibitsterminal differentiation, predisposing to pathologic conditions,such as enchondromatosis.
While we found that PTHrP regulates Gli transcriptionalactivity, genetic studies found that PTHrP expression isdependent on hedgehog activity. As such, PTHrP and hedgehogtranscriptional activity exist in a complex feedback loop ingrowth plate chondrocytes. We propose a model in whichPTHrP inhibits the progression of growth plate chondrocytedifferentiation from their proliferative state to the zone ofhypertrophy and terminal differentiation, mediated by Gli3transcriptional activity (Fig. 8). PTHrP regulates the activity ofGli3 through both a hedgehog ligand dependent, and ahedgehog ligand independent, PKA dependent pathway. Gli3is expressed primarily in growth plate chondrocytes which havenot yet undergone terminal differentiation, and as such, thePTHrP regulation of Gli3 functions primarily on chondrocytesthat have not yet entered their terminally differentiated state.The ability of PTHrP and its regulation of Gli transcriptionalactivity to regulate the height of the zone of growth platechondrocytes producing type X collagen, a marker of terminaldifferentiation, despite a lack of Gli3 expression in thesechondrocytes supports the concept that growth plate chondro-
cyte proliferation and differentiation are inversely related. Assuch, activation of Gli mediated signaling increases prolifera-tion and inhibits terminal differentiation, and PTHrP actsthrough its ability to regulate Gli3 to control the progression of

38 E. Mau et al. / Developmental Biology 305 (2007) 28–39
growth plate chondrocytes from their proliferative state intoterminal differentiation.
Acknowledgments
This work was funded by the Canadian Institutes of HealthResearch. BAAwas supported by the Canadian Research ChairsProgram. EM was supported by the Research Training Centre ofthe Hospital for Sick Children. The Gli3+/− and Gli2+/− micewere generously contributed by C.C. Hui (Hospital for SickChildren, Toronto), and the PTHrP construct was generouslycontributed byHarald Juppner (Massachusetts General Hospital,Boston).
References
Alman, B.A., Greel, D.A., Ruby, L.K., Goldberg, M.J., Wolfe, H.J., 1996.Regulation of proliferation and platelet-derived growth factor expressionin palmar fibromatosis (Dupuytren contracture) by mechanical strain.J. Orthoptera Res. 14, 722–728.
Bailey, E.C., Milenkovic, L., Scott, M.P., Collawn, J.F., Johnson, R.L., 2002.Several PATCHED1 missense mutations display activity in patched1-deficient fibroblasts. J. Biol. Chem. 277, 33632–33640.
Brewster, R., Mullor, J.L., Ruiz i Altaba, A., 2000. Gli2 functions inFGF signaling during antero-posterior patterning. Development 127,4395–4405.
Cohen Jr., M.M., 2003. The hedgehog signaling network. Am. J. Med. Genet., A123, 5–28.
Deckelbaum, R.A., Chan, G., Miao, D., Goltzman, D., Karaplis, A.C., 2002. Ihhenhances differentiation of CFK-2 chondrocytic cells and antagonizesPTHrP-mediated activation of PKA. J. Cell Sci. 115, 3015–3025.
Denker, A.E., Nicoll, S.B., Tuan, R.S., 1995. Formation of cartilage-likespheroids by micromass cultures of murine C3H10T1/2 cells upon treatmentwith transforming growth factor-beta 1. Differentiation 59, 25–34.
Dominguez, M., Brunner, M., Hafen, E., Basler, K., 1996. Sending andreceiving the hedgehog signal: control by the Drosophila Gli protein Cubitusinterruptus. Science 272, 1621–1625.
Epstein, D.J., Marti, E., Scott, M.P., McMahon, A.P., 1996. AntagonizingcAMP-dependent protein kinase A in the dorsal CNS activates a conservedSonic hedgehog signaling pathway. Development 122, 2885–2894.
Gensure, R.C., Gardella, T.J., Juppner, H., 2005. Parathyroid hormone andparathyroid hormone-related peptide, and their receptors. Biochem.Biophys. Res. Commun. 328, 666–678.
Goodrich, L.V., Milenkovic, L., Higgins, K.M., Scott, M.P., 1997. Alteredneural cell fates and medulloblastoma in mouse patched mutants. Science277, 1109–1113.
Gu, K., Zhang, L., Jin, T., Rutherford, R.B., 2004. Identification of potentialmodifiers of Runx2/Cbfa1 activity in C2C12 cells in response to bonemorphogenetic protein-7. Cells Tissues Organs 176, 28–40.
Harrington, E.K., Lunsford, L.E., Svoboda, K.K., 2004. Chondrocyte terminaldifferentiation, apoptosis, and type X collagen expression are downregulatedby parathyroid hormone. Anat. Rec. A. Discov. Mol. Cell Evol. Biol. 281,1286–1295.
Henderson, M., Danks, J., Moseley, J., Slavin, J., Harris, T., McKinlay, M.,Hopper, J., Martin, T., 2001. Parathyroid hormone-related protein produc-tion by breast cancers, improved survival, and reduced bone metastases.J. Natl. Cancer Inst. 93, 234–237.
Henderson, M.A., Danks, J.A., Slavin, J.L., Byrnes, G.B., Choong, P.F.,Spillane, J.B., Hopper, J.L., Martin, T.J., 2006. Parathyroid hormone-relatedprotein localization in breast cancers predict improved prognosis. CancerRes. 66, 2250–2256.
Hilton, M.J., Tu, X., Cook, J., Hu, H., Long, F., 2005. Ihh controls cartilagedevelopment by antagonizing Gli3, but requires additional effectors toregulate osteoblast and vascular development. Development 132,4339–4351.
Hu, M.C., Mo, R., Bhella, S., Wilson, C.W., Chuang, P.T., Hui, C.C.,Rosenblum, N.D., 2006. GLI3-dependent transcriptional repression of Gli1,Gli2 and kidney patterning genes disrupts renal morphogenesis. Develop-ment 133, 569–578.
Hui, C.C., Joyner, A.L., 1993. A mouse model of greig cephalopolysyndactylysyndrome: the extra-toes J mutation contains an intragenic deletion of theGli3 gene. Nat. Genet. 3, 241–246.
Iwasaki, M., Le, A.X., Helms, J.A., 1997. Expression of Indian hedgehog, bonemorphogenetic protein 6 and gli during skeletal morphogenesis. Mech. Dev.69, 197–202.
Johnson, R.L., Grenier, J.K., Scott, M.P., 1995. Patched overexpression alterswing disc size and pattern: transcriptional and post-transcriptional effects onhedgehog targets. Development 121, 4161–4170.
Karp, S.J., Schipani, E., St-Jacques, B., Hunzelman, J., Kronenberg, H.,McMahon, A.P., 2000. Indian hedgehog coordinates endochondral bonegrowth and morphogenesis via parathyroid hormone related-protein-dependent and -independent pathways. Development 127, 543–548.
Kindblom, J.M., Nilsson, O., Hurme, T., Ohlsson, C., Savendahl, L., 2002.Expression and localization of Indian hedgehog (Ihh) and parathyroidhormone related protein (PTHrP) in the human growth plate during pubertaldevelopment. J. Endocrinol. 174, R1–R6.
Kobayashi, T., Chung, U.I., Schipani, E., Starbuck, M., Karsenty, G., Katagiri,T., Goad, D.L., Lanske, B., Kronenberg, H.M., 2002. PTHrP and Indianhedgehog control differentiation of growth plate chondrocytes at multiplesteps. Development 129, 2977–2986.
Kobayashi, T., Soegiarto, D.W., Yang, Y., Lanske, B., Schipani, E., McMahon,A.P., Kronenberg, H.M., 2005. Indian hedgehog stimulates periarticularchondrocyte differentiation to regulate growth plate length independently ofPTHrP. J. Clin. Invest. 115, 1734–1742.
Koziel, L., Wuelling, M., Schneider, S., Vortkamp, A., 2005. Gli3 acts as arepressor downstream of Ihh in regulating two distinct steps of chondrocytedifferentiation. Development 132, 5249–5260.
Kubo, M., Nakamura, M., Tasaki, A., Yamanaka, N., Nakashima, H., Nomura,M., Kuroki, S., Katano, M., 2004. Hedgehog signaling pathway is a newtherapeutic target for patients with breast cancer. Cancer Res. 64,6071–6074.
Lanske, B., Karaplis, A.C., Lee, K., Luz, A., Vortkamp, A., Pirro, A., Karperien,M., Defize, L.H., Ho, C., Mulligan, R.C., Abou-Samra, A.B., Juppner, H.,Segre, G.V., Kronenberg, H.M., 1996. PTH/PTHrP receptor in earlydevelopment and Indian hedgehog-regulated bone growth. Science 273,663–666.
Linsenmayer, T.F., Eavey, R.D., Schmid, T.M., 1988. Type X collagen: ahypertrophic cartilage-specific molecule. Pathol. Immunopathol. Res. 7,14–19.
Marigo, V., Tabin, C.J., 1996. Regulation of patched by sonic hedgehog in thedeveloping neural tube. Proc. Natl. Acad. Sci. U. S. A. 93, 9346–9351.
Miao, D., Liu, H., Plut, P., Niu, M., Huo, R., Goltzman, D., Henderson, J.E.,2004. Impaired endochondral bone development and osteopenia in Gli2-deficient mice. Exp. Cell Res. 294, 210–222.
Matise, M.P., Joyner, A.L., 1999. Gli genes in development and cancer.Oncogene 18, 7852–7859.
Meng, X., Poon, R., Zhang, X., Cheah, A., Ding, Q., Hui, C.C., Alman, B.,2001. Suppressor of fused negatively regulates beta-catenin signaling.J. Biol. Chem. 276, 40113–40119.
Minina, E., Wenzel, H.M., Kreschel, C., Karp, S., Gaffield, W., McMahon,A.P., Vortkamp, A., 2001. BMP and Ihh/PTHrP signaling interact tocoordinate chondrocyte proliferation and differentiation. Development 128,4523–4534.
Mo, R., Freer, A.M., Zinyk, D.L., Crackower, M.A., Michaud, J., Heng,H.H., Chik, K.W., Shi, X.M., Tsui, L.C., Cheng, S.H., Joyner, A.L.,Hui, C., 1997. Specific and redundant functions of Gli2 and Gli3 zincfinger genes in skeletal patterning and development. Development 124,113–123.
Motoyama, J., Milenkovic, L., Iwama, M., Shikata, Y., Scott, M.P., Hui, C.C.,2003. Differential requirement for Gli2 and Gli3 in ventral neural cell fatespecification. Dev. Biol. 259, 150–161.
Mullor, J.L., Sanchez, P., Altaba, A.R., 2002. Pathways and consequences:Hedgehog signaling in human disease. Trends Cell Biol. 12, 562–569.

39E. Mau et al. / Developmental Biology 305 (2007) 28–39
Nieuwenhuis, E., Hui, C.C., 2005. Hedgehog signaling and congenitalmalformations. Clin. Genet. 67, 193–208.
Olsen, B.R., Reginato, A.M., Wang, W., 2000. Bone development. Annu. Rev.Cell Dev. Biol. 16, 191–220.
Riobo, N.A., Lu, K., Ai, X., Haines, G.M., Emerson Jr., C.P., 2006.Phosphoinositide 3-kinase and Akt are essential for sonic hedgehogsignaling. Proc. Natl. Acad. Sci. U. S. A.
Saika, S., Muragaki, Y., Okada, Y., Miyamoto, T., Ohnishi, Y., Ooshima, A.,Kao, W.W., 2004. Sonic hedgehog expression and role in healing cornealepithelium. Invest. Ophthalmol. Visual Sci. 45, 2577–2585.
Sasaki, H., Hui, C., Nakafuku, M., Kondoh, H., 1997. A binding site for Gliproteins is essential for HNF-3beta floor plate enhancer activity intransgenics and can respond to Shh in vitro. Development 124, 1313–1322.
Schipani, E., Langman, C.B., Parfitt, A.M., Jensen, G.S., Kikuchi, S., Kooh,S.W., Cole, W.G., Juppner, H., 1996. Constitutively activated receptors forparathyroid hormone and parathyroid hormone-related peptide in Jansen'smetaphyseal chondrodysplasia. N. Engl. J. Med. 335, 708–714.
Schipani, E., Lanske, B., Hunzelman, J., Luz, A., Kovacs, C.S., Lee, K., Pirro,A., Kronenberg, H.M., Juppner, H., 1997. Targeted expression ofconstitutively active receptors for parathyroid hormone and parathyroidhormone-related peptide delays endochondral bone formation and rescues
mice that lack parathyroid hormone-related peptide. Proc. Natl. Acad. Sci.U. S. A. 94, 13689–13694.
Tiet, T.D., Hopyan, S., Nadesan, P., Gokgoz, N., Poon, R., Lin, A.C., Yan, T.,Andrulis, I.L., Alman, B.A., Wunder, J.S., 2006. Constitutive hedgehogsignaling in chondrosarcoma up-regulates tumor cell proliferation. Am. J.Pathol. 168, 321–330.
Wang, B., Fallon, J.F., Beachy, P.A., 2000. Hedgehog-regulated processing ofGli3 produces an anterior/posterior repressor gradient in the developingvertebrate limb. Cell 100, 423–434.
Xie, L.Y., Abou-Samra, A.B., 1998. Epitope tag mapping of the extracellularand cytoplasmic domains of the rat parathyroid hormone (PTH)/PTH-relatedpeptide receptor. Endocrinology 139, 4563–4567.
Yoshida, E., Noshiro, M., Kawamoto, T., Tsutsumi, S., Kuruta, Y., Kato,Y., 2001. Direct inhibition of Indian hedgehog expression byparathyroid hormone (PTH)/PTH-related peptide and up-regulation byretinoic acid in growth plate chondrocyte cultures. Exp. Cell Res. 265,64–72.
Zhang, D., Schwarz, E.M., Rosier, R.N., Zuscik, M.J., Puzas, J.E., O'Keefe,R.J., 2003. ALK2 functions as a BMP type I receptor and induces Indianhedgehog in chondrocytes during skeletal development. J. Bone Miner.Res. 18, 1593–1604.


















