
Pseudomonas aeruginosa Increases Formation of Multidrug … · made for phzA2-phzG2, which was then...
10
JOURNAL OF BACTERIOLOGY, Apr. 2010, p. 1946–1955 Vol. 192, No. 7 0021-9193/10/$12.00 doi:10.1128/JB.01231-09 Copyright © 2010, American Society for Microbiology. All Rights Reserved. Pseudomonas aeruginosa Increases Formation of Multidrug-Tolerant Persister Cells in Response to Quorum-Sensing Signaling Molecules † Nina Mo ¨ker, Charles R. Dean, and Jianshi Tao* Department of Infectious Diseases, Novartis Institutes for BioMedical Research, Inc., 500 Technology Square, Cambridge, Massachusetts 02139 Received 11 September 2009/Accepted 18 December 2009 Bacterial persister cells constitute a small portion of a culture which is tolerant to killing by lethal doses of bactericidal antibiotics. These phenotypic variants are formed in numerous bacterial species, including those with clinical relevance like the opportunistic pathogen Pseudomonas aeruginosa. Although persisters are believed to contribute to difficulties in the treatment of many infectious diseases, the underlying mechanisms affecting persister formation are not well understood. Here we show that even though P. aeruginosa cultures have a significantly smaller fraction of multidrug-tolerant persister cells than cultures of Escherichia coli or Staphylococcus aureus, they can increase persister numbers in response to quorum-sensing-related signaling molecules. The phenazine pyocyanin (and the closely related molecule paraquat) and the acyl-homoserine lactone 3-OC12-HSL significantly increased the persister numbers in logarithmic P. aeruginosa PAO1 or PA14 cultures but not in E. coli or S. aureus cultures. Over the last 50 years, Pseudomonas aeruginosa has emerged as a major cause of nosocomial infections in immunocompro- mised patients, including those relying on mechanical ventila- tion and suffering from neutropenia or severe burns, and is perhaps most well known as the agent primarily responsible for the decline in lung function leading to death in cystic fibrosis (CF) patients (8, 14, 16, 23, 38). P. aeruginosa possesses a multiplicity of virulence factors that are elicited upon access to susceptible individuals, including various toxins, secretion sys- tems, siderophores, surface appendages, endotoxin (lipopoly- saccharide [LPS]), alginate, and phenazines. These can gener- ate acute toxicity/injury, leading P. aeruginosa to be labeled as the “hyena” of the bacterial world (15). Treatment of infec- tions by P. aeruginosa is hindered by its high level of intrinsic resistance to antibiotics due primarily to a combination of the impermeable outer membrane and a number of broad-spec- trum efflux pumps (50, 51). P. aeruginosa is also thought to enter into a biofilm mode of growth in CF lung infections (36, 48, 63), contributing both to pathogenicity/colonization and resistance to therapeutic intervention. In P. aeruginosa, global regulation, mediated by at least 3 quorum-sensing (QS) systems, controls population behaviors and synthesis of the majority of these pathogenicity factors (59, 66). This bacterium possesses two N-acyl-homoserine lactone (HSL)- mediated quorum-sensing systems, las and rhl (26, 43, 46, 47), and a Pseudomonas quinolone signal (PQS) system mediated by 2-heptyl-3-hydroxy-4-quinolone (49). In the HSL-mediated sys- tems, the HSL synthases LasI and RhlI are responsible for the synthesis of the autoinducers N-(3-oxododecanoyl)-L-HSL (3- OC12-HSL) and N-butyryl-L-HSL (C4-HSL), respectively. Ex- pressions of the lasI and rhl genes are regulated by the tran- scriptional activators LasR and RhlR in response to their cognate HSL signal molecules. Among the numerous cellu- lar and secreted virulence factors in P. aeruginosa whose biosynthesis is regulated by quorum sensing is the phenazine pyocyanin (PYO). PYO is one of the predominant phena- zines secreted by P. aeruginosa in stationary phase (11) and has been shown to act as both an important virulence factor (27, 28) and a signaling molecule (11). Phenazines are re- dox-active secondary metabolites whose antibiotic effects on competing organisms have been of research interest for more than 50 years (28). More recently, the effect of PYO on P. aeruginosa itself has been explored (11, 52), showing that PYO acts as the final signal in the quorum-sensing system cascade and regulates at least 22 genes in P. aerugi- nosa (11). Phenazine synthesis is mediated by the gene prod- ucts of two homologous operons, phzA1B1C1D1E1F1G1 (phz1) and phzA2B2C2D2E2F2G2 (phz2) (37); however, the expression of phz1 accounts for the majority of phenazine production (7, 64). Expression of genes for phenazine bio- synthesis is regulated by several mechanisms in P. aerugi- nosa, including quorum-sensing systems. Whereas both the PQS and rhl systems positively regulate phz1 expression (12, 26), the orphan LuxR-type quorum-sensing regulator QscR negatively regulates the expression of phz1 and phz2 (29). In recent years, the resistance/tolerance of biofilms to anti- biotic treatment has been associated with the existence of a subpopulation of bacteria called persister cells (60, 61). Per- sisters were first described in 1944 by Joseph Bigger, who noted that penicillin could not completely sterilize a culture of Staph- ylococcus (3). This observation was generally ignored until Harris Moyed again examined persisters in the 1980s (40, 41, 55). Persisters constitute a small subpopulation of apparently nongrowing multidrug-tolerant cells present in all bacterial populations studied to date (31). In contrast to resistance caused by genetic mutations, tolerance to antibiotics by persis- ters occurs as phenotypic variance of the wild type (WT): * Corresponding author. Mailing address: Department of Infectious Diseases, Novartis Institutes for BioMedical Research, 500 Technol- ogy Square, Cambridge, MA 02139. Phone: (617) 871-7431. Fax: (617) 871-5791. E-mail: [email protected]. † Supplemental material for this article may be found at http://jb .asm.org/. Published ahead of print on 22 January 2010. 1946 on March 19, 2020 by guest http://jb.asm.org/ Downloaded from
Transcript of Pseudomonas aeruginosa Increases Formation of Multidrug … · made for phzA2-phzG2, which was then...

JOURNAL OF BACTERIOLOGY, Apr. 2010, p. 1946–1955 Vol. 192, No. 70021-9193/10/$12.00 doi:10.1128/JB.01231-09Copyright © 2010, American Society for Microbiology. All Rights Reserved.
Pseudomonas aeruginosa Increases Formation of Multidrug-TolerantPersister Cells in Response to Quorum-Sensing Signaling Molecules�†
Nina Moker, Charles R. Dean, and Jianshi Tao*Department of Infectious Diseases, Novartis Institutes for BioMedical Research, Inc.,
500 Technology Square, Cambridge, Massachusetts 02139
Received 11 September 2009/Accepted 18 December 2009
Bacterial persister cells constitute a small portion of a culture which is tolerant to killing by lethal doses ofbactericidal antibiotics. These phenotypic variants are formed in numerous bacterial species, including thosewith clinical relevance like the opportunistic pathogen Pseudomonas aeruginosa. Although persisters arebelieved to contribute to difficulties in the treatment of many infectious diseases, the underlying mechanismsaffecting persister formation are not well understood. Here we show that even though P. aeruginosa cultureshave a significantly smaller fraction of multidrug-tolerant persister cells than cultures of Escherichia coli orStaphylococcus aureus, they can increase persister numbers in response to quorum-sensing-related signalingmolecules. The phenazine pyocyanin (and the closely related molecule paraquat) and the acyl-homoserinelactone 3-OC12-HSL significantly increased the persister numbers in logarithmic P. aeruginosa PAO1 or PA14cultures but not in E. coli or S. aureus cultures.
Over the last 50 years, Pseudomonas aeruginosa has emergedas a major cause of nosocomial infections in immunocompro-mised patients, including those relying on mechanical ventila-tion and suffering from neutropenia or severe burns, and isperhaps most well known as the agent primarily responsible forthe decline in lung function leading to death in cystic fibrosis(CF) patients (8, 14, 16, 23, 38). P. aeruginosa possesses amultiplicity of virulence factors that are elicited upon access tosusceptible individuals, including various toxins, secretion sys-tems, siderophores, surface appendages, endotoxin (lipopoly-saccharide [LPS]), alginate, and phenazines. These can gener-ate acute toxicity/injury, leading P. aeruginosa to be labeled asthe “hyena” of the bacterial world (15). Treatment of infec-tions by P. aeruginosa is hindered by its high level of intrinsicresistance to antibiotics due primarily to a combination of theimpermeable outer membrane and a number of broad-spec-trum efflux pumps (50, 51). P. aeruginosa is also thought toenter into a biofilm mode of growth in CF lung infections (36,48, 63), contributing both to pathogenicity/colonization andresistance to therapeutic intervention.
In P. aeruginosa, global regulation, mediated by at least 3quorum-sensing (QS) systems, controls population behaviors andsynthesis of the majority of these pathogenicity factors (59, 66).This bacterium possesses two N-acyl-homoserine lactone (HSL)-mediated quorum-sensing systems, las and rhl (26, 43, 46, 47), anda Pseudomonas quinolone signal (PQS) system mediated by2-heptyl-3-hydroxy-4-quinolone (49). In the HSL-mediated sys-tems, the HSL synthases LasI and RhlI are responsible for thesynthesis of the autoinducers N-(3-oxododecanoyl)-L-HSL (3-OC12-HSL) and N-butyryl-L-HSL (C4-HSL), respectively. Ex-
pressions of the lasI and rhl genes are regulated by the tran-scriptional activators LasR and RhlR in response to theircognate HSL signal molecules. Among the numerous cellu-lar and secreted virulence factors in P. aeruginosa whosebiosynthesis is regulated by quorum sensing is the phenazinepyocyanin (PYO). PYO is one of the predominant phena-zines secreted by P. aeruginosa in stationary phase (11) andhas been shown to act as both an important virulence factor(27, 28) and a signaling molecule (11). Phenazines are re-dox-active secondary metabolites whose antibiotic effects oncompeting organisms have been of research interest formore than 50 years (28). More recently, the effect of PYOon P. aeruginosa itself has been explored (11, 52), showingthat PYO acts as the final signal in the quorum-sensingsystem cascade and regulates at least 22 genes in P. aerugi-nosa (11). Phenazine synthesis is mediated by the gene prod-ucts of two homologous operons, phzA1B1C1D1E1F1G1(phz1) and phzA2B2C2D2E2F2G2 (phz2) (37); however, theexpression of phz1 accounts for the majority of phenazineproduction (7, 64). Expression of genes for phenazine bio-synthesis is regulated by several mechanisms in P. aerugi-nosa, including quorum-sensing systems. Whereas both thePQS and rhl systems positively regulate phz1 expression (12,26), the orphan LuxR-type quorum-sensing regulator QscRnegatively regulates the expression of phz1 and phz2 (29).
In recent years, the resistance/tolerance of biofilms to anti-biotic treatment has been associated with the existence of asubpopulation of bacteria called persister cells (60, 61). Per-sisters were first described in 1944 by Joseph Bigger, who notedthat penicillin could not completely sterilize a culture of Staph-ylococcus (3). This observation was generally ignored untilHarris Moyed again examined persisters in the 1980s (40, 41,55). Persisters constitute a small subpopulation of apparentlynongrowing multidrug-tolerant cells present in all bacterialpopulations studied to date (31). In contrast to resistancecaused by genetic mutations, tolerance to antibiotics by persis-ters occurs as phenotypic variance of the wild type (WT):
* Corresponding author. Mailing address: Department of InfectiousDiseases, Novartis Institutes for BioMedical Research, 500 Technol-ogy Square, Cambridge, MA 02139. Phone: (617) 871-7431. Fax: (617)871-5791. E-mail: [email protected].
† Supplemental material for this article may be found at http://jb.asm.org/.
� Published ahead of print on 22 January 2010.
1946
on March 19, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

reinoculation (growth) of isolated persister cells regeneratesthe original sensitive population (3, 21, 67), indicating thatpersistence is an epigenetic phenomenon. Given the potentialclinical importance of multidrug-tolerant persister cells, it issurprising how little is known about them, in the cases of someof the more intractable pathogens confronting us today, andwhether one or more regulatory circuits mediate switching from anormal growth mode to a persistent cell. The phenomenon ofpersister formation so far has been characterized mainly with theorganism Escherichia coli (reviewed in reference 32), whereas lessis understood regarding persister cells of P. aeruginosa. One hy-pothesis about the underlying mechanism for persister formationis that persister cells are phenotypic variants which are not regu-lated per se but result from stochastic fluctuations of toxic proteins(32). However, several genes, including genes encoding the globalstationary phase-related regulators SpoT, RelA, DskA, and RpoS(42, 62), that appear to be involved in persistence in P. aeruginosahave been discovered (9, 42, 62). We undertook to investigate theformation of persister cells in P. aeruginosa cultures in a compre-hensive analysis under various conditions related to cidal andstatic antibiotic challenge, quorum-sensing-mediated signaling,and environmental stress conditions. Here, we demonstrate thateven though P. aeruginosa produces less persisters than does E.coli or S. aureus, it can increase the persister cell fraction up to90-fold in response to 3-OC12-HSL or PYO but not to a numberof tested antibiotics or general stress conditions like hyperosmoticor heat stress.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions. Bacterial strains and plas-mids that were used or constructed in this study are summarized in Table 1.
Bacteria were cultivated aerobically (220 rpm) at 37°C, unless otherwise indi-cated. LB broth (Difco) or LB agar (Difco, Remel) was used for routine cultur-ing. Cells were typically grown by diluting overnight cultures 1:1,000 in 15 ml LBusing 125-ml Erlenmeyer flasks with vented caps (Corning Inc.). Overnightcultures were prepared by inoculating 5 to 7 single colonies from the correspond-ing LB agar plates into 15 ml LB broth and incubating for 16 to 18 h at 37°C withshaking at 220 rpm.
DNA methods. Restriction digestion, ligation, transformation, and agarose gelelectrophoresis were done according to standard protocols (54). Plasmids were
prepared from E. coli using a QIAprep Spin miniprep kit from Qiagen according tothe manufacturer’s protocol. Electroporation of plasmids into P. aeruginosa cells wasperformed as previously described (6). Genomic DNA of P. aeruginosa was extractedusing the Puregene DNA purification kit (Gentra Systems) according to the man-ufacturer’s instructions. DNA fragments used for cloning were extracted from aga-rose gels using a QIAquick gel extraction kit from Qiagen. PCR products werepurified using a QIAquick PCR purification kit (Qiagen) and, when cloned, se-quenced to confirm the correct sequences (Agencourt Bioscience Corp., Beverly,MA). Oligonucleotide synthesis was carried out by Agencourt.
Construction of mutants. Unmarked deletions of the two redundant operonsfor phenazine biosynthesis, phz1 and phz2, were constructed in P. aeruginosaPA14 with a procedure based on that recently described by Dietrich et al.(11), by using the pEX18Tc suicide vector (19) for the sac-based selectionmethod described by H. Schweizer (56). Briefly, a single deletion was firstmade for phzA2-phzG2, which was then used for generating the �phz1 �phz2double knockout (the PA14 �phz mutant). For the deletion of phzA2-phzG2, theflanking regions of approximately 1 kb were amplified from the chromosome ofP. aeruginosa PA14 using the primer pairs phzA2-1000-s/as (upstream of phzA2;Table 2) and phzG2�1000-s/as (downstream of phzG2; Table 2). The resultingDNA fragments contained 21-bp overlaps and were joined by overlap extensionPCR. The PCR product contained the phzA2-phzG2 deletion and was first clonedinto pCR2.1TOPO (Invitrogen) and subsequently subcloned into pEX18Tc usingthe HindIII and BamHI restriction sites. The resulting plasmid, pEX18Tc-�phz2,was mobilized into P. aeruginosa from E. coli S17-1 (4). Briefly, P. aeruginosaPA14 was inoculated in 10 ml LB and incubated overnight at 42°C withoutshaking, while the donor E. coli/pEX18Tc-�phz2 was cultivated overnight at 37°Cwith shaking at 220 rpm. For matings, 300 �l of each culture were mixed, the cellscollected, and the pellets spread onto the center of an LB agar plate. After 18 hof growth at 37°C, the cells were plated on Pseudomonas isolation agar (PIA;Difco) containing 100 �g/ml tetracycline to select for merodiploids. After re-growth on PIA/tetracycline, single colonies were plated on LB agar containing5% (wt/vol) sucrose for selecting cointegrants that are sucrose resistant due tothe loss of the sacB gene. These were then screened for the appropriate deletionusing colony PCR. For the double deletion, the procedure was repeated startingfrom the amplification with the primer pairs phzA1-1000-s/as (upstream ofphzA1; Table 2) and phzG1�1000-s/as (downstream of phzG1; Table 2) andusing the single-deletion PA14 �phz2 mutant strain.
MICs. For MIC determination for different antibiotics, two independent P.aeruginosa cultures were grown to early logarithmic phase and treated with orwithout 2 mM PYO prior to MIC testing according to our persister assayconditions (see below). For the non-PYO samples, the same volume of dimethylsulfoxide (DMSO) was added. After cultivation in the presence of PYO/DMSOfor 1 h 15 min (37°C, 220 rpm), the cultures were diluted 1:1 in LB prior toaddition of 1.5 �l bacterial suspensions to 100 �l LB containing various concen-trations of carbenicillin, piperacillin, imipenem, meropenem, ciprofloxacin,ofloxacin, tobramycin, or kanamycin. The LB used for both dilution steps con-
TABLE 1. Bacterial strains and plasmids used in this study
Strain or plasmid Relevant characteristics Source orreference
E. coli strainsTOP10 F� mcrA �(mrr-hsdRMS-mcrBC) �80lacZ�M15 �lac�74 recA1 araD139
�(ara-leu)7697 galU galK rpsL (Strr) endA1 nupGInvitrogen
S17-1 thi pro hsdR recA Tra� 57K-12 AG100 argE3 thi-1 rpsL xyl mtl �(gal-uvrB) supE44 44
P. aeruginosa strainsPAO1 Wild type 35PAO1 �soxR mutant PAO1 with an in-frame deletion in gene PA2273 K. PoolePA14 Clinical isolate UCBPP-PA14 53PA14 �phz2 mutant �phzA2-phzG2 This studyPA14 �phz mutant �phzA2-phzG2, �phzA1-phzG1 This study
S. aureus RN4220 �rsbU �agr 25
PlasmidspEX18Tc Broad-host-range gene replacement vector; sacB, Tcr 19pEX18Tc-�phz2 pEX18Tc carrying an in-frame deletion in �phzA2-phzG2 This studypEX18Tc-�phz1 pEX18Tc carrying an in-frame deletion in �phzA1-phzG1 This study
VOL. 192, 2010 QS-MEDIATED INCREASE OF P. AERUGINOSA PERSISTER CELLS 1947
on March 19, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
tained an amount of PYO/DMSO identical to that in the cultures. The MICs ofPYO, PCA, 3-OC12-HSL, or paraquat were determined accordingly, without theadditional PYO/DMSO treatment. The MICs were determined after �24 h ofincubation at 37°C.
Persistence assay. Persisters were determined, following a modified procedurepreviously described (32), by exposure of logarithmic or stationary cultures toantibiotics using concentrations exceeding five times the MIC for each antibiotic.Persister numbers were determined by plating the antibiotic-treated cultures onLB agar plates and subsequent counting of CFU representing the cell numberswhich survived antibiotic exposure. Logarithmic cultures of P. aeruginosa werechallenged with different antibiotics after 3 to 3.5 h of cultivation, whereaslogarithmic cultures of E. coli or S. aureus were treated after 3 h or 2.5 h ofincubation at 37°C with shaking at 220 rpm, respectively. The difference in thetimes of incubation allows these bacterial species to be compared at similargrowth stages. Stationary cultures were generally challenged after 18 h of culti-vation. To kill all nonpersister cells, treatment with antibiotics was performed for
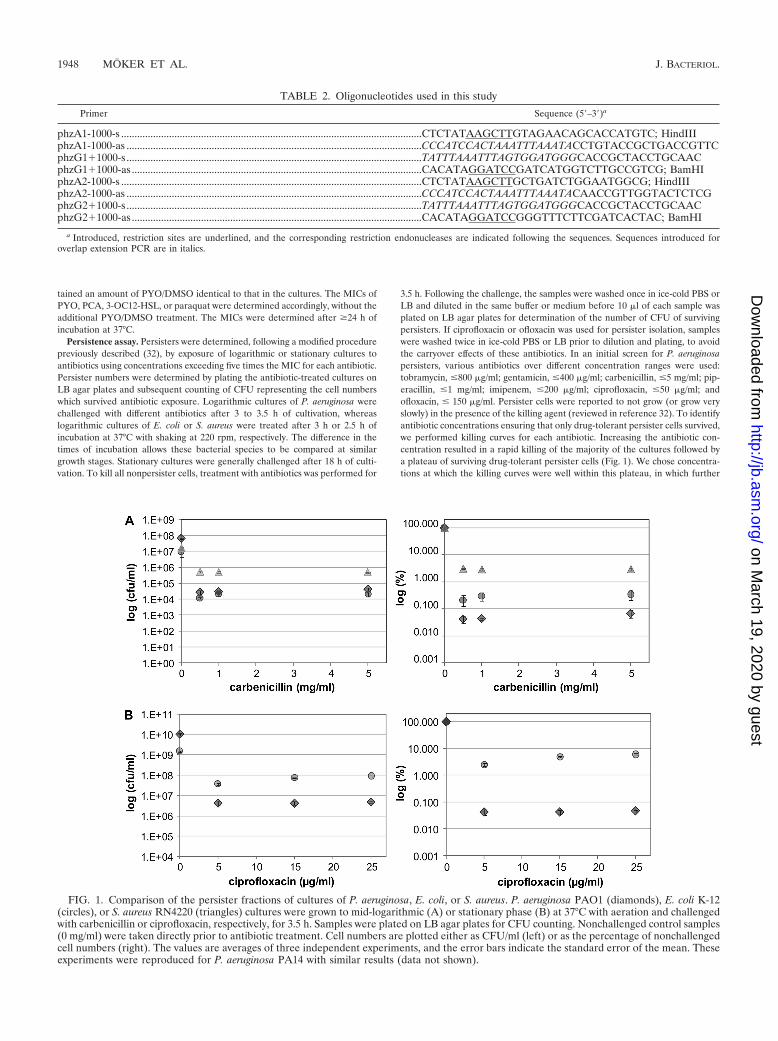
3.5 h. Following the challenge, the samples were washed once in ice-cold PBS orLB and diluted in the same buffer or medium before 10 �l of each sample wasplated on LB agar plates for determination of the number of CFU of survivingpersisters. If ciprofloxacin or ofloxacin was used for persister isolation, sampleswere washed twice in ice-cold PBS or LB prior to dilution and plating, to avoidthe carryover effects of these antibiotics. In an initial screen for P. aeruginosapersisters, various antibiotics over different concentration ranges were used:tobramycin, �800 �g/ml; gentamicin, �400 �g/ml; carbenicillin, �5 mg/ml; pip-eracillin, �1 mg/ml; imipenem, �200 �g/ml; ciprofloxacin, �50 �g/ml; andofloxacin, � 150 �g/ml. Persister cells were reported to not grow (or grow veryslowly) in the presence of the killing agent (reviewed in reference 32). To identifyantibiotic concentrations ensuring that only drug-tolerant persister cells survived,we performed killing curves for each antibiotic. Increasing the antibiotic con-centration resulted in a rapid killing of the majority of the cultures followed bya plateau of surviving drug-tolerant persister cells (Fig. 1). We chose concentra-tions at which the killing curves were well within this plateau, in which further
FIG. 1. Comparison of the persister fractions of cultures of P. aeruginosa, E. coli, or S. aureus. P. aeruginosa PAO1 (diamonds), E. coli K-12(circles), or S. aureus RN4220 (triangles) cultures were grown to mid-logarithmic (A) or stationary phase (B) at 37°C with aeration and challengedwith carbenicillin or ciprofloxacin, respectively, for 3.5 h. Samples were plated on LB agar plates for CFU counting. Nonchallenged control samples(0 mg/ml) were taken directly prior to antibiotic treatment. Cell numbers are plotted either as CFU/ml (left) or as the percentage of nonchallengedcell numbers (right). The values are averages of three independent experiments, and the error bars indicate the standard error of the mean. Theseexperiments were reproduced for P. aeruginosa PA14 with similar results (data not shown).
TABLE 2. Oligonucleotides used in this study
Primer Sequence (5�–3�)a
phzA1-1000-s .................................................................................................................CTCTATAAGCTTGTAGAACAGCACCATGTC; HindIIIphzA1-1000-as ...............................................................................................................CCCATCCACTAAATTTAAATACCTGTACCGCTGACCGTTCphzG1�1000-s ...............................................................................................................TATTTAAATTTAGTGGATGGGCACCGCTACCTGCAACphzG1�1000-as .............................................................................................................CACATAGGATCCGATCATGGTCTTGCCGTCG; BamHIphzA2-1000-s .................................................................................................................CTCTATAAGCTTGCTGATCTGGAATGGCG; HindIIIphzA2-1000-as ...............................................................................................................CCCATCCACTAAATTTAAATACAACCGTTGGTACTCTCGphzG2�1000-s ...............................................................................................................TATTTAAATTTAGTGGATGGGCACCGCTACCTGCAACphzG2�1000-as .............................................................................................................CACATAGGATCCGGGTTTCTTCGATCACTAC; BamHI
a Introduced, restriction sites are underlined, and the corresponding restriction endonucleases are indicated following the sequences. Sequences introduced foroverlap extension PCR are in italics.
1948 MOKER ET AL. J. BACTERIOL.
on March 19, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

increase of the concentration does not result in further killing. The compoundswere then used as listed: 0.5 to 5 mg/ml carbenicillin, 25 to 75 �g/ml imipenem,and 10 to 25 �g/ml ofloxacin or ciprofloxacin. Antibiotics were purchased fromSigma-Aldrich, except ciprofloxacin and imipenem, which were purchased fromUnited States Pharmacopeia (USP).
Determination of persister fractions under different conditions. To test theeffect of different agents on P. aeruginosa persister formation, we cultivated P.aeruginosa to early logarithmic phase (2 h 20 min to 2 h 25 min) and subsequentlyadded �2 mM PYO (Cayman Chemical) or phenazine-1-carboxylic acid (PCA;isolated by the Novartis Natural Products Unit from a Streptomyces strain), �1mM paraquat (Sigma-Aldrich), 0.5 mM 3-OC12-HSL (MP Biomedicals), �2.5%H2O2, �0.5 �g/ml ofloxacin, �0.12 �g/ml ciprofloxacin, or �32 �g/ml chloram-phenicol to the samples. Besides pyocyanin, which was diluted 1:40 into thecultures, all agents were prepared as 100� stock solutions. For the mock-treatedcontrol samples, the same amount of the solvent was added (DMSO for PYO,PCA, or 3-OC12-HSL controls, 0.7% ethanol for the chloramphenicol controls,and H2O for all other compounds). After further cultivation to mid-logarithmicgrowth phase (an additional 1 h 10 min to 1 h 15 min), the persister fraction ofthe cultures was determined as described above. To ensure that the differentbacterial strains were treated at similar growth stages, cultures of E. coli or S.aureus were cultivated for 1 h 20 min and subsequently treated for 1 h 10 min to1 h 15 min.
To investigate the effect of culture supernatants of different strains on persisterformation, the P. aeruginosa PAO1 and PA14 strains, the PA14 �phz mutant, E.coli K-12, and S. aureus RN4220 were grown for 16 to 18 h at 37°C, 220 rpm,before the cells were removed by centrifugation at 37°C. The resulting superna-tants were supplemented with 25 g/liter powdered LB (Difco) before sterilefiltration (0.2-�m-pore diameter; Millipore) to restore depleted nutrients. As thecontrol, the same concentration of LB was dissolved in H2O and sterile filtered.These supernatants, tested to ensure that no contaminating live cells werepresent and prewarmed at 37°C, were used to reculture bacterial cells collectedat early logarithmic phase for 1.5 h before they were challenged with 1 mg/mlcarbenicillin or piperacillin for 3.5 h.
The influence of hyperosmotic or heat stress conditions on P. aeruginosapersister formation was tested as follows. To test for hyperosmotic stress, the P.aeruginosa cells were collected at 37°C and resuspended in the same culturevolume of prewarmed (37°C) LB broth or LB broth supplemented with 0.2 M or0.5 M NaCl. Heat stress conditions were applied by splitting the cultures forcultivation at 37 and 45°C. After 1 h 20 min of stress exposure, the persisterfraction of the cultures was determined as described above.
RESULTS
P. aeruginosa cultures have a significantly smaller persisterfraction than that of E. coli or S. aureus. In order to charac-terize persister cells of P. aeruginosa, it was first determined iftreatment with antibiotics of various classes left a fraction ofsurviving cells and how large this proportion of persisters incultures of P. aeruginosa was. Logarithmic or stationary P.aeruginosa PAO1 or PA14 cultures were exposed to severalantibiotic classes over different concentration ranges includingaminoglycosides (tobramycin and gentamicin), penicillins (car-benicillin and piperacillin), carbapenems (imipenem), or quin-olones (ciprofloxacin and ofloxacin) for 3.5 h before determin-ing the fractions of surviving persister cells by plating for CFUcounting. Treatment with increasing concentrations of the dif-ferent antibiotics resulted in the expected biphasic killingcurves, showing a rapid killing of the majority of cells using upto the MIC of each antibiotic, followed by a plateau of persistercells surviving antibiotic concentrations of several times theMICs (data not shown). Persisters were previously reported toconstitute a remarkably high fraction of stationary (1%) aswell as logarithmic (0.1%) E. coli or S. aureus (�1%) cul-tures (reviewed in reference 32). In comparison, the fraction ofpersisters that we initially observed with P. aeruginosa cultureswas significantly lower. We therefore performed a direct com-parison of the persister fractions in mid-logarithmic and sta-
tionary cultures of P. aeruginosa PAO1 and PA14, E. coli K-12,and S. aureus RN4220 by treatment with up to 5 mg/ml car-benicillin or up to 25 �g/ml ciprofloxacin for 3.5 h. In all fourinstances, rapid killing of the majority of cells in logarithmiccultures occurred at carbenicillin concentrations of �0.5 mg/ml, followed by plateaus of cells surviving up to 5 mg/ml of thiscell wall inhibitor (Fig. 1). The concentration range we chosefor carbenicillin was therefore suitable to compare the frac-tions of persisters in these different bacterial species. Surpris-ingly, even though the P. aeruginosa cultures had higher cellnumbers in the untreated control samples, they had signifi-cantly lower persister fractions than did E. coli or S. aureus(Fig. 1). In logarithmic-phase cultures, the portion of P. aerugi-nosa survivor cells (0.04%) was almost 10-fold lower than thatof E. coli (0.3%) and at least 60-fold lower than that of S. aureus(2.5%). Comparison of persisters in logarithmic P. aeruginosaand E. coli cultures isolated by ciprofloxacin resulted in evenlower levels in P. aeruginosa cultures of 0.0005% compared tothe 0.15% found in identically treated E. coli cultures (data notshown). One possible explanation for the higher persister num-bers in the carbenicillin-treated Pseudomonas cultures is that weobserved the cultures still undergoing growth during carbenicillintreatment prior to lysis (data not shown), whereas ciprofloxacinappeared to more rapidly halt the growth of Pseudomonas cul-tures.
We found similar species-specific differences in persisternumbers in stationary cultures. In stationary E. coli cultures, upto 7% of the cells were persisters tolerant to ciprofloxacintreatment. In comparison, only 0.02 to 0.1% of drug-tolerantpersister cells survived ciprofloxacin treatment in stationary P.aeruginosa cultures (Fig. 1). To ensure that the higher persisternumbers in E. coli K-12 and S. aureus RN4220 cultures werenot a phenomenon specific to these laboratory-adapted strains,we confirmed these results using clinical isolates (data notshown). Taken together, these findings somewhat unexpect-edly show that P. aeruginosa forms persister fractions in loga-rithmic and stationary cultures which are significantly lowerthan those of the other tested bacteria.
P. aeruginosa increases the fraction of persisters in responseto exogenous pyocyanin. As P. aeruginosa has a comparativelysmall fraction of persisters, we speculated that certain environ-mental conditions or chemical treatments might increase thepersister level. P. aeruginosa PAO1 and PA14 persister forma-tion was investigated under hyperosmotic (�0.5 M NaCl), heat(45°C), and oxidative stress (�2.5% H2O2) conditions. We alsotested exposure to bacteriostatic or cidal antibiotics, using con-centration ranges from well below to up to the MICs (�0.5�g/ml ofloxacin, �0.12 �g/ml ciprofloxacin, and �32 �g/mlchloramphenicol), as well as �2 mM of the phenazine PYO(MIC of 2 mM). While general stress or the listed antibioticsshowed no significant impact on persister formation (data notshown), addition of PYO, a predominant phenazine secretedby P. aeruginosa at stationary phase, to early logarithmic cul-tures of P. aeruginosa PAO1 or PA14 substantially increasedthe proportion of cells surviving carbenicillin treatment ap-plied in mid-log phase (Fig. 2). This effect was dose dependent,reaching an almost 90-fold increase over untreated cells when2 mM PYO was used (Fig. 2A). To confirm that these survivorcells indeed represented persisters, we measured the effect of2 mM PYO over different carbenicillin concentrations. The
VOL. 192, 2010 QS-MEDIATED INCREASE OF P. AERUGINOSA PERSISTER CELLS 1949
on March 19, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
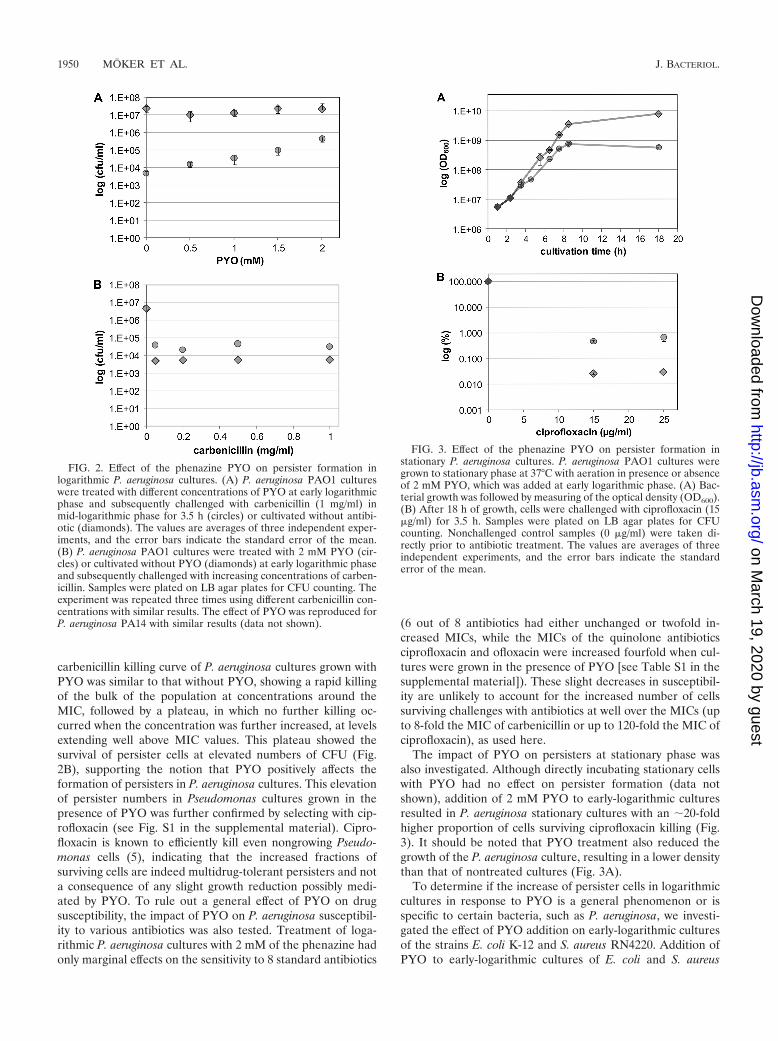
carbenicillin killing curve of P. aeruginosa cultures grown withPYO was similar to that without PYO, showing a rapid killingof the bulk of the population at concentrations around theMIC, followed by a plateau, in which no further killing oc-curred when the concentration was further increased, at levelsextending well above MIC values. This plateau showed thesurvival of persister cells at elevated numbers of CFU (Fig.2B), supporting the notion that PYO positively affects theformation of persisters in P. aeruginosa cultures. This elevationof persister numbers in Pseudomonas cultures grown in thepresence of PYO was further confirmed by selecting with cip-rofloxacin (see Fig. S1 in the supplemental material). Cipro-floxacin is known to efficiently kill even nongrowing Pseudo-monas cells (5), indicating that the increased fractions ofsurviving cells are indeed multidrug-tolerant persisters and nota consequence of any slight growth reduction possibly medi-ated by PYO. To rule out a general effect of PYO on drugsusceptibility, the impact of PYO on P. aeruginosa susceptibil-ity to various antibiotics was also tested. Treatment of loga-rithmic P. aeruginosa cultures with 2 mM of the phenazine hadonly marginal effects on the sensitivity to 8 standard antibiotics
(6 out of 8 antibiotics had either unchanged or twofold in-creased MICs, while the MICs of the quinolone antibioticsciprofloxacin and ofloxacin were increased fourfold when cul-tures were grown in the presence of PYO [see Table S1 in thesupplemental material]). These slight decreases in susceptibil-ity are unlikely to account for the increased number of cellssurviving challenges with antibiotics at well over the MICs (upto 8-fold the MIC of carbenicillin or up to 120-fold the MIC ofciprofloxacin), as used here.
The impact of PYO on persisters at stationary phase wasalso investigated. Although directly incubating stationary cellswith PYO had no effect on persister formation (data notshown), addition of 2 mM PYO to early-logarithmic culturesresulted in P. aeruginosa stationary cultures with an 20-foldhigher proportion of cells surviving ciprofloxacin killing (Fig.3). It should be noted that PYO treatment also reduced thegrowth of the P. aeruginosa culture, resulting in a lower densitythan that of nontreated cultures (Fig. 3A).
To determine if the increase of persister cells in logarithmiccultures in response to PYO is a general phenomenon or isspecific to certain bacteria, such as P. aeruginosa, we investi-gated the effect of PYO addition on early-logarithmic culturesof the strains E. coli K-12 and S. aureus RN4220. Addition ofPYO to early-logarithmic cultures of E. coli and S. aureus
FIG. 2. Effect of the phenazine PYO on persister formation inlogarithmic P. aeruginosa cultures. (A) P. aeruginosa PAO1 cultureswere treated with different concentrations of PYO at early logarithmicphase and subsequently challenged with carbenicillin (1 mg/ml) inmid-logarithmic phase for 3.5 h (circles) or cultivated without antibi-otic (diamonds). The values are averages of three independent exper-iments, and the error bars indicate the standard error of the mean.(B) P. aeruginosa PAO1 cultures were treated with 2 mM PYO (cir-cles) or cultivated without PYO (diamonds) at early logarithmic phaseand subsequently challenged with increasing concentrations of carben-icillin. Samples were plated on LB agar plates for CFU counting. Theexperiment was repeated three times using different carbenicillin con-centrations with similar results. The effect of PYO was reproduced forP. aeruginosa PA14 with similar results (data not shown).
FIG. 3. Effect of the phenazine PYO on persister formation instationary P. aeruginosa cultures. P. aeruginosa PAO1 cultures weregrown to stationary phase at 37°C with aeration in presence or absenceof 2 mM PYO, which was added at early logarithmic phase. (A) Bac-terial growth was followed by measuring of the optical density (OD600).(B) After 18 h of growth, cells were challenged with ciprofloxacin (15�g/ml) for 3.5 h. Samples were plated on LB agar plates for CFUcounting. Nonchallenged control samples (0 �g/ml) were taken di-rectly prior to antibiotic treatment. The values are averages of threeindependent experiments, and the error bars indicate the standarderror of the mean.
1950 MOKER ET AL. J. BACTERIOL.
on March 19, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

resulted in efficient killing of these bacterial species (Fig. 4),consistent with the fact that phenazines are a class of antibi-otics (1, 2, 18, 22). Nonetheless, the total numbers of persistersthat tolerated ampicillin killing in the PYO-treated samplesdid not increase. The killing of E. coli or S. aureus by PYOreached the plateau at 0.5 mM and higher, at which the smallfractions of cells that survived the PYO treatment were alsotolerant to ampicillin. This suggests that an increase in per-sister numbers in response to PYO is not a general phenom-enon and also that other bacterial persisters are in fact tolerantto killing by PYO.
To examine whether the positive effect on persister forma-tion in P. aeruginosa cultures is a common feature of phena-zines and related molecules or more specific to PYO, we testedthe effect of two structurally related compounds, paraquat andPCA, on logarithmic P. aeruginosa cultures.
It was recently demonstrated that paraquat and PYO inducepartially overlapping transcriptional changes in P. aeruginosa,including the induction of genes encoding the RND effluxpump MexGHI-OpmD, as well as PA2274, a putative flavin-dependent monooxygenase, and that this induction is mediatedby the transcriptional regulator SoxR (gene product ofPA2273) (11, 24, 45). The addition of �0.2 mM paraquat(MIC of 1 mM) increased persisters in P. aeruginosa PAO1 andPA14 cultures significantly compared to untreated cultures
(Fig. 5A). As in the case of PYO, cultivation of logarithmiccultures of E. coli K-12 in the presence of paraquat did notaffect the persister fraction (data not shown), showing that theparaquat-mediated increase may be specific for P. aeruginosa.A P. aeruginosa deletion mutant lacking the soxR gene (thePAO1 �soxR mutant) produced numbers of persisters in allgrowth stages similar to those of the parent strain and alsoexhibited the paraquat-mediated increase in logarithmic cul-ture persisters, suggesting that SoxR does not play a role in theformation of persisters in PAO1.
PCA was shown to be a second predominant phenazineexcreted at similar levels by stationary PA14 cultures (30 �Mwhen grown in minimal medium) (12). Interestingly, in con-trast to PYO and paraquat, addition of up to 2 mM of thephenazine molecule PCA (MIC of 2 mM) did not have aneffect on persister fractions in PAO1 or PA14 cultures (datanot shown). Hence, elevated persister numbers are not medi-ated by all phenazines but seem to be more specific to PYO (orparaquat). PCA, like paraquat, is structurally related to PYOand is also thought to have oxidative activity (52). The findingthat PYO, but not PCA, leads to increased persister numbersin P. aeruginosa cultures indicates that the underlying mecha-nism is more specific than exposure to oxidative stress, whichis in accordance with our observation that up to 2.5% H2O2
FIG. 4. Effect of the phenazine PYO on persister formation inlogarithmic cultures of E. coli and S. aureus. S. aureus RN4220 (A) orE. coli K-12 (B) cultures were treated with different concentrations ofPYO at early logarithmic phase and subsequently challenged withampicillin (1 mg/ml) in mid-logarithmic phase for 3.5 h (circles) orcultivated without antibiotic (diamonds). Samples were plated on LBagar plates for CFU counting. The values are averages of three inde-pendent experiments, and the error bars indicate the standard error ofthe mean.
FIG. 5. Effect of paraquat or 3-OC12-HSL on P. aeruginosa per-sister formation. P. aeruginosa PAO1 cultures were treated with dif-ferent concentrations of paraquat (A) or 3-OC12-HSL (B) at early logphase and subsequently challenged with carbenicillin (1 mg/ml) inmid-log phase (circles) for 3.5 h or cultivated without antibiotic (dia-monds). Samples were plated on LB agar plates for CFU counting.The effect of paraquat was reproduced for P. aeruginosa PA14 withsimilar results (data not shown). The values are averages of threeindependent experiments, and the error bars indicate the standarderror of the mean.
VOL. 192, 2010 QS-MEDIATED INCREASE OF P. AERUGINOSA PERSISTER CELLS 1951
on March 19, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

did not enhance persister formation (data not shown). Fur-ther, despite the close structural similarity, there seems tobe a differential recognition of PYO versus PCA.
The observation that the phenazine PYO can increase thefraction of persisters in logarithmic cultures suggested a directrole for phenazines in regulating persister numbers. To testwhether the loss of PYO production would alter the persistercell fraction, a �phz1 �phz2 double-deletion mutant, which waspreviously reported to exhibit a complete lack of phenazineproduction (11), was constructed in PA14 (the PA14 �phzmutant). The persister levels in logarithmic, early-stationary(18 h), and late-stationary (24 to 72 h) cultures of the PA14strain and PA14 �phz mutant were then examined. Interest-ingly, the deletion strain lacking phenazine production did notdiffer in its fraction of persister cells in all the cultures exam-ined (data not shown). These findings suggest that even thoughthe phenazine PYO is able to modulate persister formation inP. aeruginosa, an additional and possibly compensatory signal-ing system(s) capable of controlling the persister phenotype islikely also present in this organism.
P. aeruginosa persister formation is positively influenced bythe quorum-sensing-related signaling molecule acyl-homo-serine lactone 3-OC12-HSL. Since PYO acts as quorum-sens-ing signaling molecule in P. aeruginosa, we decided to furtherexamine the effect of another QS signal, the acyl-homoserinelactone 3-OC12-HSL, on persister formation. This signal mol-ecule is produced by LasI, which sits atop the currently pro-posed hierarchy of the QS circuit (10). The effect of increasingconcentrations of 3-OC12-HSL (MIC of 1 mM) on logarith-mic cultures of PAO1 and PA14 was determined. Similar to theeffect seen with PYO, 3-OC12-HSL significantly increased per-sister numbers in strain PAO1 (Fig. 5B). However, and incontrast to PYO, 3-OC12-HSL did not affect persister numbersin PA14 (data not shown). Hence, the signaling effects of3-OC12-HSL on early-logarithmic cultures seem to differ be-tween the two P. aeruginosa strains.
Since the phenazine PYO and 3-OC12-HSL, each a QSsignal, increased persister numbers in logarithmic cultures in
PAO1 when added exogenously, we then asked if these mole-cules potentiate each other to promote the persister formation.This was tested by coincubating the PAO1 cultures with 0.2mM PYO, which is closer to the reported physiological level(11) and alone does not affect persister numbers (Fig. 2A), andvarious amounts of 3-OC12-HSL (Fig. 6). It is clear that in thepresence of 0.2 mM PYO, even low concentrations of 3-OC12-HSL (�40 �M) resulted in increases of persister cells in thetreated PAO1 cultures (Fig. 6).
Effect of cell-free spent growth media on PA14 persisterformation. The PYO- and 3-OC12-HSL concentrations used inthe above-described experiments to trigger an increase in per-sisters are higher than physiological concentrations typicallyaccumulated in stationary P. aeruginosa cultures; in the super-natant of PA14, a strain that secrets 10-fold more PYO thanPAO1, about 60 �M PYO was detected after growth in LB(11), whereas 3-OC12-HSL was shown to be excreted at 5�M by PAO1 (47). However, it is clear that some signalingmolecules are able to potentiate each other to enhance thepersister phenotype (Fig. 6). To address if other unidentifiedsignaling molecule(s) in P. aeruginosa cultures would allowPYO to exert its persister-promoting activity at the physiolog-ical concentrations, spent media prepared from both wild-typePA14 and PA14 �phz mutant stationary cultures, which weresupplemented with fresh LB, were used to cultivate early-logarithmic P. aeruginosa cultures prior to antibiotic challenge.As shown in Fig. 7, the spent media from the stationary cul-tures of PA14 WT induced a significant increase of the per-sister fraction in the PA14 cultures. In contrast, the spentgrowth medium derived from the phenazine-deficient mutantdid not increase the persister fraction (Fig. 7). This suggeststhat physiological levels of the phenazine PYO excreted bystationary PA14 cultures may be sufficient to induce elevatedpersister formation in P. aeruginosa in the context of the entiresecretome. Persisters in E. coli or S. aureus logarithmic cultureswere not affected by the above-described spent medium pre-pared from PA14 (or by the spent medium prepared from
FIG. 6. Synergistic positive effects of PYO and 3-OC12-HSL on P. aeruginosa PAO1 persister formation. P. aeruginosa PAO1 cultures weretreated with different concentrations of 3-OC12-HSL and 0.2 mM PYO (circles) or without PYO (diamonds) at early log phase and subsequentlychallenged with carbenicillin (5 mg/ml, filled symbols) in mid-log phase or cultivated without antibiotic (open symbols). Samples were plated onLB agar plates for CFU counting. Cell numbers are either plotted as CFU/ml (left) or as the percentage of nonchallenged cell numbers (right).The values are averages of three independent experiments, and the error bars indicate the standard error of the mean.
1952 MOKER ET AL. J. BACTERIOL.
on March 19, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

stationary cultures of their own species; data not shown), con-sistent with the results obtained with the synthetic PYO.
DISCUSSION
Bacterial persisters comprise a small subpopulation of aculture that is tolerant to killing with lethal doses of bacteri-cidal antibiotics (3) and are believed to contribute to the dif-ficulties in effectively treating many infectious diseases (30, 32,33). Although the persister theory was proposed in the 1940s,there is still an incomplete understanding of the underlyingmechanisms that affect the persistence of bacterial cells toantibiotic killing. A current theory proposed for persister for-mation in E. coli postulates that stochastic fluctuation in theproduction of toxic proteins known to block cell metabolismmediates entrance of cells into a dormant state enabling thepersister cell to escape corruption of cellular functions by an-tibiotics (32). However, in both E. coli and P. aeruginosa anumber of the genes that were identified to affect the persisterphenotype encode global regulators, such as DnaK, DnaJ,DksA, histone-like protein, integration host factor (17), andPhoU (34) in E. coli or SpoT, RelA, DksA (62), and RpoS (42)in P. aeruginosa, suggesting that environmental signaling playsan important role in modulating persistence.
Here we demonstrate that even though cultures of the ubiq-uitous environmental organism and therapeutically challengingopportunistic pathogen P. aeruginosa have significantly lesspersisters than cultures of E. coli or S. aureus under standardlaboratory conditions, this phenotype can be affected by QS-linked molecules. The phenazine pyocyanin and the acyl-ho-moserine lactone 3-OC12-HSL, which are normally secretedby this bacterium at later stages of growth, as well as paraquat,significantly increased the persister numbers in logarithmic P.aeruginosa cultures. During the time we were preparing themanuscript, Kayama et al. (20) reported that overexpression of
LasR in P. aeruginosa, which should lead to elevated produc-tion of 3-OC12-HSL, resulted in increased tolerance to ofloxa-cin. This observation is consistent with our results obtainedfrom 3-OC12-HSL treatment. In contrast, overexpression ofRhlR in the same study did not show any effect on ofloxacintolerance.
Interestingly, testing the PA14 �phz mutant showed that acomplete lack of phenazine biosynthesis did not result in al-tered persister levels in logarithmic or stationary cultures.There is, however, a clear and specific response of log-phasecells to these molecules, which in various environmental orclinical niches would likely be present via production by neigh-boring stationary-phase cells. Our data showing that spent P.aeruginosa culture medium from wild-type in which QS mole-cules as a whole are present in physiological levels, but notphenazine-defective cultures, increased persister numbers sup-port this notion and also suggest that additional molecules maybe secreted that could contribute alone or together with PYOto the regulation of persister numbers in log-phase cultures.Consistent with this, PYO and 3-OC12-HSL, when addedalone, required high concentrations to exert persister-promot-ing activities but worked synergistically at lower concentrationsto increase persister fractions in log-phase cultures of strainPAO1.
Also, many phenomena involved in persister formation havebeen reported to show high redundancy, with prominent ex-amples given by the toxins RelE and MazF in E. coli (32);whereas plasmid-mediated overexpression of RelE or MazF inE. coli was shown to strongly increase tolerance to antibiotics,deletion of the relE or mazF gene was reported to have noeffection persister production phenotype.
It should be noted that under the conditions tested 3-OC12-HSL affected persister levels in PAO1 but not in PA14. Thismay be related to the fact that PAO1 produces much lowerlevels of PYO than PA14 (11). So in this case, it can bespeculated that additional acyl-HSL would have a more pro-found synergistic effect with a low concentration of PYOand/or other factors affecting the persister phenotype in PAO1.Indeed, the results obtained with the spent media also suggestthat additional signaling molecule(s) are likely involved inmodulating persister formation in P. aeruginosa, as the rela-tively lower concentration of PYO that accumulates in PA14stationary culture medium is sufficient to enhance P. aerugi-nosa persisters.
The observed persistence enhancement is a specific responseto PYO and 3-OC12-HSL treatments and is not associatedwith general environmental stress, growth arresting or cidalityexerted by the static or cidal antibiotics tested here, or oxida-tive stress. In addition, the ability of these signaling moleculesto increase persistence to antibiotic killing appears to be bac-teria specific. Clearly, when PYO alone, PYO in PA14 spentmedium, or the spent medium of stationary E. coli or S. aureuscultures were used to treat logarithmic E. coli or S. aureus cells,no significant change to the persistence of these bacteria couldbe observed. It is intriguing to speculate that phenazines (aloneor in synergy with other QS molecules) may not only inhibitother bacteria but also enhance persister formation to poten-tially protect the phenazine-producing P. aeruginosa popula-tion from complete killing by antibiotics produced by compet-ing organisms.
FIG. 7. Effect of cell-free supernatants of stationary cultures of thePA14 WT and the PA14 �phz mutant on persister formation. P. aerugi-nosa PA14 cultures were grown to early log phase, collected, andresuspended in LB (�) or cell-free supernatants of stationary PA14WT or PA14 �phz mutant cultures (supplemented with LB powder).After another 1 h 15 min of cultivation, the cultures were challengedwith carbenicillin or piperacillin (1 mg/ml) for 3.5 h (solid bars). Sam-ples were plated on LB agar plates for CFU counting. Nonchallengedcontrol samples (shaded bars) were taken directly prior to antibiotictreatment. The values are averages of three independent experiments,and the error bars indicate the standard error of the mean.
VOL. 192, 2010 QS-MEDIATED INCREASE OF P. AERUGINOSA PERSISTER CELLS 1953
on March 19, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

Interestingly, although we did not observe a difference in thelevels of persister production in cultures of the PA14 WT andthe phz mutant, the spent media of the two strains differed intheir effects. Since in our studies PYO increased persisternumbers when added to logarithmic but not to stationary cul-tures, it can be speculated that once they reach later stages ofgrowth, the cultures are unaffected. The effects of QS mole-cules on persister formation would then become importantwhen growing cells are exposed to exogenous QS molecules,for example, in a setting like the CF lung, where it was recentlyshown that the majority of the P. aeruginosa cells analyzedwere actively growing (68) and could therefore benefit fromenhancing effects on persister formation by PYO or HSL mol-ecules in their environment.
In this context it is intriguing to note that acyl-HSLs can bereadily detected in the sputa of CF patients (39, 58) and thatthe activities of the las and rhl systems were shown to beenhanced in the presence of sputum extracts from CF patients(13). Furthermore, PYO in the sputa of Pseudomonas-infectedCF patients has been detected in concentrations up to 100 �M(65). Future experiments will be required to delineate thesignaling pathway that leads to the increased persister forma-tion in response to PYO or acyl-HSL treatments and to betterunderstand the clinical relevance of this phenomenon.
ACKNOWLEDGMENTS
We thank Keith Poole for providing the PAO1 �soxR strain. Wethank E. Peter Greenberg for very helpful discussions and methodicalinput. We thank Steven J. Projan and Neil S. Ryder for careful readingof the manuscript and their suggestions for improvement. We thankEsther Schmitt for helpful advice on naturally produced phenazineanalogues.
REFERENCES
1. Baron, S. S., and J. J. Rowe. 1981. Antibiotic action of pyocyanin. Antimi-crob. Agents Chemother. 20:814–820.
2. Baron, S. S., G. Terranova, and J. J. Rowe. 1989. Molecular mechanism ofthe antimicrobial action of pyocyanin. Curr. Microbiol. 18:223–230.
3. Bigger, J. W. 1944. Treatment of staphylococcal infections with penicillin.Lancet ii:497–500.
4. Cao, L., R. Srikumar, and K. Poole. 2004. MexAB-OprM hyperexpression inNalC-type multidrug-resistant Pseudomonas aeruginosa: identification andcharacterization of the nalC gene encoding a repressor of PA3720-PA3719.Mol. Microbiol. 53:1423–1436.
5. Chalkley, L. J., and H. J. Koornhof. 1985. Antimicrobial activity of cipro-floxacin against Pseudomonas aeruginosa, Escherichia coli, and Staphylococ-cus aureus determined by the killing curve method: antibiotic comparisonsand synergistic interactions. Antimicrob. Agents Chemother. 28:331–342.
6. Choi, K. H., A. Kumar, and H. P. Schweizer. 2006. A 10-min method forpreparation of highly electrocompetent Pseudomonas aeruginosa cells: ap-plication for DNA fragment transfer between chromosomes and plasmidtransformation. J. Microbiol. Methods 64:391–397.
7. Chugani, S. A., M. Whiteley, K. M. Lee, D. D’Argenio, C. Manoil, and E. P.Greenberg. 2001. QscR, a modulator of quorum-sensing signal synthesis andvirulence in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. U. S. A. 98:2752–2757.
8. Crouch Brewer, S., R. G. Wunderink, C. B. Jones, and K. V. Leeper, Jr. 1996.Ventilator-associated pneumonia due to Pseudomonas aeruginosa. Chest109:1019–1029.
9. De Groote, V. N., N. Verstraeten, M. Fauvart, C. I. Kint, A. M. Verbeeck, S.Beullens, P. Cornelis, and J. Michiels. 2009. Novel persistence genes inPseudomonas aeruginosa identified by high-throughput screening. FEMS Mi-crobiol. Lett. 297:73–79.
10. de Kievit, T. R., and B. H. Iglewski. 2000. Bacterial quorum sensing inpathogenic relationships. Infect. Immun. 68:4839–4849.
11. Dietrich, L. E., A. Price-Whelan, A. Petersen, M. Whiteley, and D. K. New-man. 2006. The phenazine pyocyanin is a terminal signalling factor in thequorum sensing network of Pseudomonas aeruginosa. Mol. Microbiol. 61:1308–1321.
12. Diggle, S. P., K. Winzer, S. R. Chhabra, K. E. Worrall, M. Camara, and P.Williams. 2003. The Pseudomonas aeruginosa quinolone signal molecule
overcomes the cell density-dependency of the quorum sensing hierarchy,regulates rhl-dependent genes at the onset of stationary phase and can beproduced in the absence of LasR. Mol. Microbiol. 50:29–43.
13. Duan, K., and M. G. Surette. 2007. Environmental regulation of Pseudomo-nas aeruginosa PAO1 Las and Rhl quorum-sensing systems. J. Bacteriol.189:4827–4836.
14. Dunn, M., and R. G. Wunderink. 1995. Ventilator-associated pneumoniacaused by Pseudomonas infection. Clin. Chest Med. 16:95–109.
15. Fick, R. B. 1993. Pseudomonas aeruginosa: the microbial hyena and itsrole in disease: an introduction., p. 1–5. In R. B. Fick (ed.), Pseudomonasaeruginosa, the opportunist: pathogenesis and disease. CRC Press, BocaRaton, FL.
16. Govan, J. R., and V. Deretic. 1996. Microbial pathogenesis in cystic fibrosis:mucoid Pseudomonas aeruginosa and Burkholderia cepacia. Microbiol. Rev.60:539–574.
17. Hansen, S., K. Lewis, and M. Vulic. 2008. Role of global regulators andnucleotide metabolism in antibiotic tolerance in Escherichia coli. Antimi-crob. Agents Chemother. 52:2718–2726.
18. Hassan, H. M., and I. Fridovich. 1980. Mechanism of the antibiotic actionpyocyanine. J. Bacteriol. 141:156–163.
19. Hoang, T. T., R. R. Karkhoff-Schweizer, A. J. Kutchma, and H. P. Schweizer.1998. A broad-host-range Flp-FRT recombination system for site-specificexcision of chromosomally-located DNA sequences: application for isolationof unmarked Pseudomonas aeruginosa mutants. Gene 212:77–86.
20. Kayama, S., K. Murakami, T. Ono, M. Ushimaru, A. Yamamoto, K. Hirota,and Y. Miyake. 2009. The role of rpoS gene and quorum-sensing system inofloxacin tolerance in Pseudomonas aeruginosa. FEMS Microbiol. Lett.
21. Keren, I., N. Kaldalu, A. Spoering, Y. Wang, and K. Lewis. 2004. Persistercells and tolerance to antimicrobials. FEMS Microbiol. Lett. 230:13–18.
22. Kerr, J. R., G. W. Taylor, A. Rutman, N. Hoiby, P. J. Cole, and R. Wilson.1999. Pseudomonas aeruginosa pyocyanin and 1-hydroxyphenazine inhibitfungal growth. J. Clin. Pathol. 52:385–387.
23. Kluytmans, J. 1997. Surgical infections including burns, p. 841–865. In R. P.Wenzel (ed.), Prevention and control of nosocomial infections, 3rd ed. Wil-liams and Wilkins, Baltimore, MD.
24. Kobayashi, K., and S. Tagawa. 2004. Activation of SoxR-dependent tran-scription in Pseudomonas aeruginosa. J. Biochem. 136:607–615.
25. Kreiswirth, B. N., S. Lofdahl, M. J. Betley, M. O’Reilly, P. M. Schlievert,M. S. Bergdoll, and R. P. Novick. 1983. The toxic shock syndrome exotoxinstructural gene is not detectably transmitted by a prophage. Nature 305:709–712.
26. Latifi, A., M. K. Winson, M. Foglino, B. W. Bycroft, G. S. Stewart, A.Lazdunski, and P. Williams. 1995. Multiple homologues of LuxR and LuxIcontrol expression of virulence determinants and secondary metabolitesthrough quorum sensing in Pseudomonas aeruginosa PAO1. Mol. Microbiol.17:333–343.
27. Lau, G. W., D. J. Hassett, H. Ran, and F. Kong. 2004. The role of pyocyaninin Pseudomonas aeruginosa infection. Trends Mol. Med. 10:599–606.
28. Laursen, J. B., and J. Nielsen. 2004. Phenazine natural products: biosynthe-sis, synthetic analogues, and biological activity. Chem. Rev. 104:1663–1686.
29. Lequette, Y., J. H. Lee, F. Ledgham, A. Lazdunski, and E. P. Greenberg.2006. A distinct QscR regulon in the Pseudomonas aeruginosa quorum-sensing circuit. J. Bacteriol. 188:3365–3370.
30. Levin, B. R., and D. E. Rozen. 2006. Non-inherited antibiotic resistance. Nat.Rev. Microbiol. 4:556–562.
31. Lewis, K. 2001. Riddle of biofilm resistance. Antimicrob. Agents Chemother.45:999–1007.
32. Lewis, K. 2007. Persister cells, dormancy and infectious disease. Nat. Rev.Microbiol. 5:48–56.
33. Lewis, K. 2008. Multidrug tolerance of biofilms and persister cells. Curr.Top. Microbiol. Immunol. 322:107–131.
34. Li, Y., and Y. Zhang. 2007. PhoU is a persistence switch involved in persisterformation and tolerance to multiple antibiotics and stresses in Escherichiacoli. Antimicrob. Agents Chemother. 51:2092–2099.
35. Masuda, N., and S. Ohya. 1992. Cross-resistance to meropenem, cephems,and quinolones in Pseudomonas aeruginosa. Antimicrob. Agents Chemother.36:1847–1851.
36. Matsui, H., B. R. Grubb, R. Tarran, S. H. Randell, J. T. Gatzy, C. W. Davis,and R. C. Boucher. 1998. Evidence for periciliary liquid layer depletion, notabnormal ion composition, in the pathogenesis of cystic fibrosis airwaysdisease. Cell 95:1005–1015.
37. Mavrodi, D. V., R. F. Bonsall, S. M. Delaney, M. J. Soule, G. Phillips, andL. S. Thomashow. 2001. Functional analysis of genes for biosynthesis ofpyocyanin and phenazine-1-carboxamide from Pseudomonas aeruginosaPAO1. J. Bacteriol. 183:6454–6465.
38. Mendelson, M. H., A. Gurtman, S. Szabo, E. Neibart, B. R. Meyers, M.Policar, T. W. Cheung, D. Lillienfeld, G. Hammer, S. Reddy, et al. 1994.Pseudomonas aeruginosa bacteremia in patients with AIDS. Clin. Infect. Dis.18:886–895.
39. Middleton, B., H. C. Rodgers, M. Camara, A. J. Knox, P. Williams, and A.Hardman. 2002. Direct detection of N-acylhomoserine lactones in cysticfibrosis sputum. FEMS Microbiol. Lett. 207:1–7.
1954 MOKER ET AL. J. BACTERIOL.
on March 19, 2020 by guest
http://jb.asm.org/
Dow
nloaded from

40. Moyed, H. S., and K. P. Bertrand. 1983. hipA, a newly recognized gene ofEscherichia coli K-12 that affects frequency of persistence after inhibition ofmurein synthesis. J. Bacteriol. 155:768–775.
41. Moyed, H. S., and S. H. Broderick. 1986. Molecular cloning and expressionof hipA, a gene of Escherichia coli K-12 that affects frequency of persistenceafter inhibition of murein synthesis. J. Bacteriol. 166:399–403.
42. Murakami, K., T. Ono, D. Viducic, S. Kayama, M. Mori, K. Hirota, K.Nemoto, and Y. Miyake. 2005. Role for rpoS gene of Pseudomonas aerugi-nosa in antibiotic tolerance. FEMS Microbiol. Lett. 242:161–167.
43. Ochsner, U. A., A. Fiechter, and J. Reiser. 1994. Isolation, characterization,and expression in Escherichia coli of the Pseudomonas aeruginosa rhlABgenes encoding a rhamnosyltransferase involved in rhamnolipid biosurfac-tant synthesis. J. Biol. Chem. 269:19787–19795.
44. Okusu, H., D. Ma, and H. Nikaido. 1996. AcrAB efflux pump plays a majorrole in the antibiotic resistance phenotype of Escherichia coli multiple-anti-biotic-resistance (Mar) mutants. J. Bacteriol. 178:306–308.
45. Palma, M., J. Zurita, J. A. Ferreras, S. Worgall, D. H. Larone, L. Shi, F.Campagne, and L. E. Quadri. 2005. Pseudomonas aeruginosa SoxR does notconform to the archetypal paradigm for SoxR-dependent regulation of thebacterial oxidative stress adaptive response. Infect. Immun. 73:2958–2966.
46. Passador, L., J. M. Cook, M. J. Gambello, L. Rust, and B. H. Iglewski. 1993.Expression of Pseudomonas aeruginosa virulence genes requires cell-to-cellcommunication. Science 260:1127–1130.
47. Pearson, J. P., L. Passador, B. H. Iglewski, and E. P. Greenberg. 1995. Asecond N-acylhomoserine lactone signal produced by Pseudomonas aerugi-nosa. Proc. Natl. Acad. Sci. U. S. A. 92:1490–1494.
48. Pedersen, S. S. 1992. Lung infection with alginate-producing, mucoidPseudomonas aeruginosa in cystic fibrosis. APMIS Suppl. 28:1–79.
49. Pesci, E. C., J. B. Milbank, J. P. Pearson, S. McKnight, A. S. Kende, E. P.Greenberg, and B. H. Iglewski. 1999. Quinolone signaling in the cell-to-cellcommunication system of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci.U. S. A. 96:11229–11234.
50. Poole, K. 2004. Efflux-mediated multiresistance in Gram-negative bacteria.Clin. Microbiol. Infect. 10:12–26.
51. Poole, K. 2005. Efflux-mediated antimicrobial resistance. J. Antimicrob. Che-mother. 56:20–51.
52. Price-Whelan, A., L. E. Dietrich, and D. K. Newman. 2006. Rethinking‘secondary’ metabolism: physiological roles for phenazine antibiotics. Nat.Chem. Biol. 2:71–78.
53. Rahme, L. G., E. J. Stevens, S. F. Wolfort, J. Shao, R. G. Tompkins, andF. M. Ausubel. 1995. Common virulence factors for bacterial pathogenicity inplants and animals. Science 268:1899–1902.
54. Sambrook, J., and D. W. Russell. 2001. Molecular cloning: a laboratorymanual, 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Har-bor, NY.
55. Scherrer, R., and H. S. Moyed. 1988. Conditional impairment of cell divisionand altered lethality in hipA mutants of Escherichia coli K-12. J. Bacteriol.170:3321–3326.
56. Schweizer, H. P. 1992. Allelic exchange in Pseudomonas aeruginosa usingnovel ColE1-type vectors and a family of cassettes containing a portable oriTand the counter-selectable Bacillus subtilis sacB marker. Mol. Microbiol.6:1195–1204.
57. Simon, R., U. Priefer, and A. Puhler. 1983. A broad host range mobilizationsystem for in vivo genetic-engineering - transposon mutagenesis in gram-negative bacteria. Biotechnology (NY) 1:784–791.
58. Singh, P. K., A. L. Schaefer, M. R. Parsek, T. O. Moninger, M. J. Welsh, andE. P. Greenberg. 2000. Quorum-sensing signals indicate that cystic fibrosislungs are infected with bacterial biofilms. Nature 407:762–764.
59. Smith, R. S., and B. H. Iglewski. 2003. P. aeruginosa quorum-sensing systemsand virulence. Curr. Opin. Microbiol. 6:56–60.
60. Spoering, A. L., and K. Lewis. 2001. Biofilms and planktonic cells of Pseudo-monas aeruginosa have similar resistance to killing by antimicrobials. J.Bacteriol. 183:6746–6751.
61. Stewart, P. S., and J. W. Costerton. 2001. Antibiotic resistance of bacteria inbiofilms. Lancet 358:135–138.
62. Viducic, D., T. Ono, K. Murakami, H. Susilowati, S. Kayama, K. Hirota, andY. Miyake. 2006. Functional analysis of spoT, relA and dksA genes on quin-olone tolerance in Pseudomonas aeruginosa under nongrowing condition.Microbiol. Immunol. 50:349–357.
63. Wagner, V. E., and B. H. Iglewski. 2008. P. aeruginosa biofilms in CF infec-tion. Clin. Rev. Allergy Immunol. 35:124–134.
64. Whiteley, M., K. M. Lee, and E. P. Greenberg. 1999. Identification of genescontrolled by quorum sensing in Pseudomonas aeruginosa. Proc. Natl. Acad.Sci. U. S. A. 96:13904–13909.
65. Wilson, R., D. A. Sykes, D. Watson, A. Rutman, G. W. Taylor, and P. J. Cole.1988. Measurement of Pseudomonas aeruginosa phenazine pigments in spu-tum and assessment of their contribution to sputum Sol toxicity for respira-tory epithelium. Infect. Immun. 56:2515–2517.
66. Winzer, K., and P. Williams. 2001. Quorum sensing and the regulation ofvirulence gene expression in pathogenic bacteria. Int. J. Med. Microbiol.291:131–143.
67. Wiuff, C., R. M. Zappala, R. R. Regoes, K. N. Garner, F. Baquero, and B. R.Levin. 2005. Phenotypic tolerance: antibiotic enrichment of noninheritedresistance in bacterial populations. Antimicrob. Agents Chemother.49:1483–1494.
68. Yang, L., J. A. Haagensen, L. Jelsbak, H. K. Johansen, C. Sternberg, N.Hoiby, and S. Molin. 2008. In situ growth rates and biofilm development ofPseudomonas aeruginosa populations in chronic lung infections. J. Bacteriol.190:2767–2776.
VOL. 192, 2010 QS-MEDIATED INCREASE OF P. AERUGINOSA PERSISTER CELLS 1955
on March 19, 2020 by guest
http://jb.asm.org/
Dow
nloaded from
![Phase-matched scalable THz generation in two-color ... THz 10 THz 100 THz 1 PHz 10 PHz 300 m 30 m ... Kim presentation at Argonne 2012_no backup.ppt [Compatibility Mode] Author:](https://static.fdocuments.in/doc/165x107/5ac2b9eb7f8b9aca388e95a7/phase-matched-scalable-thz-generation-in-two-color-thz-10-thz-100-thz-1-phz.jpg)

















