
Présentée par - univ-oran1.dz · recommended to store the samples in low-temperature s to protect...
130
Mémoire présenté en vue de l’obtention du diplôme de MAGISTER en Gestion des Ressources Aquatiques LABORATOIRE D’AQUACULTURE ET DE BIOREMEDIATION (AQUABIOR) Présentée par: M elle . BENCHEGRA Khadidja Devant la commission du jury : Président : BENSAHLA-TALET Ahmed Professeur à l’université d’Oran Examinateur : ALI MEHIDI Smail M.C.A. à l’université d’Oran Examinateur : AIT YAHIA Dalila Professeur à l’université d’Oran Promoteur : ABI AYAD S.-M. El-amine Professeur à l’université d’Oran Co-promoteur: DRICI Habiba M.C.A. à l’université d’Oran Soutenu le : Faculté des Sciences Département de Biotechnologie Dynamique dans la formation de l’amine biogène, histamine, des hydroperoxydes, des TBA-rs et le suivi de la qualité microbiologique chez la sardine (Sardina pilchardus) Année universitaire : 2011 - 2012
Transcript of Présentée par - univ-oran1.dz · recommended to store the samples in low-temperature s to protect...

Mémoire présenté en vue de l’obtention du diplôme de
MAGISTER
en Gestion des Ressources Aquatiques
LABORATOIRE D’AQUACULTURE ET DE BIOREMEDIATION (AQUABIOR)
Présentée par:
Melle. BENCHEGRA Khadidja
Devant la commission du jury :
Président : BENSAHLA-TALET Ahmed Professeur à l’université d’Oran
Examinateur : ALI MEHIDI Smail M.C.A. à l’université d’Oran
Examinateur : AIT YAHIA Dalila Professeur à l’université d’Oran
Promoteur : ABI AYAD S.-M. El-amine Professeur à l’université d’Oran
Co-promoteur: DRICI Habiba M.C.A. à l’université d’Oran
Soutenu le :
Faculté des Sciences
Département de Biotechnologie
Dynamique dans la formation de l’amine biogène, histamine,
des hydroperoxydes, des TBA-rs et le suivi de la qualité
microbiologique chez la sardine (Sardina pilchardus)
Année universitaire : 2011 - 2012

Remerciements

Remerciement
Ce travail est le fruit d’une aventure de 3 ans qui n’aurait pas pu voir le jour sans le soutien
de nombreuses personnes que je tiens à remercier.
Je remercie en premier lieu le Créateur des cieux et des terres, notre Grand Dieu Tout
Puissant qui par Sa Volonté m’a permis d’affronter les difficultés rencontrées et aboutir à la
réalisation de ce travail.
Je souhaite adresser mes remerciements les plus sincères à mon encadreur, Professeur
Abi-Ayad Sidi Mohammed El-amine. Sa gentillesse, sa patience et sa grande humanité ont
fait de ces années de travail à ses côtés un réel plaisir. Je n’oublierai jamais la qualité de son
encadrement. Merci de m’avoir donné l'opportunité de faire une maîtrise. Merci pour vos
bons conseils et votre grande disponibilité tout au long des expérimentations et de la
rédaction, et aussi pour l'inspiration, l'aide et le temps que vous avez bien voulu me
consacrer.
Un profond respect et un merci tout spécial au Docteur Drici Habiba mon co-encadreur
pour son soutien permanent, sa confiance, son esprit scientifique, sa patience et pour avoir su
répondre à mes innombrables questions existentielles. Je souhaite être à la hauteur de votre
attente et votre espérance.
Je remercie également Monsieur BENSAHLA TALET Ahmed, Professeur a la faculté
des sciences d'Oran pour sa gentillesse, pour l'encouragement qu’il m’a apporté tout au long
de ces trois années. J'ai apprécié son enthousiasme qui a constitué un précieux soutien et qui
m’a fait l’honneur d'avoir accepté de présider ce jury.
La présence dans mon jury de Professeur AIT YAHIA Dalila m’honore très
sincèrement. Elle a très aimablement accepté d’examiner ce travail. Qu’elle veuille trouver ici
ma profonde gratitude.
J’exprime mes sincères remerciements à Monsieur ALI MEHIDI Smail pour avoir bien
voulu examiner ce travail je le remercie pour l’honneur qu’il me fait en acceptant de le juger.
Je le remercie également pour les nombreuses questions que j’ai du lui poser depuis six ans
maintenant dès les années de l’ingéniorat et qu’il a pris le temps gentiment pour me répondre
et m’expliquer…….
Je tiens également à remercier chaleureusement les membres de l’équipe du laboratoire
d’AQUABIOR qui ont su allumer et nourrir la petite flamme que j’avais pour la recherche, je
ne citerai pas de noms par peur d’oublier quelques uns leur place est bien réservée dans mon
cœur. Mais je veux que vous sachiez que vous m’avez tous apporté quelque chose pour la

Remerciement
réalisation de ce travail. Mais c’est surtout sur le plan humain que je voudrais vous remercier.
Il est vrai que je n’oublierai pas les discussions scientifiques, et encore moins l’entraide, tous
les services, les petits mots de réconfort…et parfois les petites tensions qui font que les
semaines se suivent sans se ressembler…c’était excellent !!!…
Je tiens aussi à exprimer ma gratitude et ma reconnaissance aux chercheurs séniors du
laboratoire d’AQUABIOR, qui nous ont encadré pendant notre année théorique et nous ont tant
assisté pendant la phase expérimentale tout particulièrement : Professeur BENSAHLA,
Docteur ALI MEHIDI, Docteur LAMARA, Professeur BABA HAMED, Docteur
TALEB et je ne saurai dépasser ce stade sans remercier Professeur BOUKORTT.
Un grand merci à tous les membres d’administration et techniciens du département de
Biotechnologie, tout particulièrement Louisa, Atika et Kader pour les nombreuses heures
d’aide pendant la réalisation de ce travail.
Je ne pourrai jamais assez remercier ma famille, tout particulièrement mon G.P.S qui me
guide dans ma vie : ma maman et mes deux sœurs Wafaa et Rachida. Ainsi que ma grand-
mère, mes tantes : hbiba Ramzia, Houria, Malika et mes oncles : hbibi Hafid, hbibi Mustapha,
Hassen et leurs enfants tout particulièrement ma petite nièce Iness, qui m’ont tant soutenu
tout au long de mes années d’étude et qui ont cru en moi, chose qui n’a fait que me pousser
pour donner encore plus.
Un merci tout spécial à la famille Kabouche : Baraat, Fateh, Malak, Nidal et également
Soraya et Amineainsi que Fatiha et ses enfants : Bouchra, Meriem et Mouhamed pour les
nombreux moments de pure délire qui m’ont aidé à dépasser des moments difficiles : MERCI
BEAUCOUP.
Je remercie toutes les personnes qui ont contribué de loin ou de près à la réalisation du
présent mémoire.

Remerciement
A mes parents
A Rachida
A Wafaa
A tous ceux que j’aime

Résumé
L’objectif de cette étude est la mise en évidence et le suivi de l’évolution temporelle des
changements post-mortem que subit le muscle de poisson par une approche biochimique et
microbiologique et leur corrélation avec l’altération sensorielle. Ce travail expérimental porte
sur l’étude de l’effet du mode (température ambiante : 25° C et réfrigérée : 4° C) et de la
durée de conservation sur l’altération de la qualité chez la sardine (Sardina pilchardus).
La première partie de ce mémoire est consacrée aux paramètres biochimiques; la
dynamique de production des hydroperoxydes et des TBA-rs, l’apparition de l’azote basique
volatile total (ABVT) et la mesure de la quantité des protéines totales. Ces paramètres
montrent des différences significatives (p<0,05) en fonction du temps et du mode de
conservation.
Dans un deuxième temps, l’altération d’origine microbienne a été étudiée, ainsi la flore
mésophile aérobie totale, les coliformes totaux, les coliformes fécaux, les streptocoques
thermotolérants, les staphylocoques, les bactéries sulfito-réductrices, les pseudomonas, les
salmonelles et les shigelles ont été dénombrées ou isolées. Enfin, certaines activités
enzymatiques microbiennes d’altération comme la lipolyse et la gélatinolyse sont aussi
abordées. L’utilisation des milieux sélectifs ont permis de démontrer la présence de ces
activités qui apparaissent et disparaissent à des moments différents en fonction de la période
et du mode de conservation.
Le suivi de la cinétique de production de l’amine biogène, histamine, nous a permis de
voir le moment de son apparition et de sa disparition ainsi que sa quantification et d’isoler
également les bactéries responsables de sa production.
Nous observons l’existence d’une synergie entre l’évolution de l’altération biochimique
et la détérioration microbiologique. Ces dernières, sont en corrélation positive avec la perte de
la qualité sensorielle. Par ailleurs, il s’avère que la réfrigération retarde ces activités
d’altération en faveur de cette dernière. Il est, donc, conseillé de stocker les échantillons à
basse température afin de préserver leur qualité, et de les consommer, au plus tard, dans les
deux jours qui suivent leur capture.
Mots-clé : Hydroperoxydes, TBA-rs, ABVT, flore bactérienne, activité lipolytique,
activité gélatinolytique, histamine, Sardina pilchardus.

Abstract
The objective of this experimental work was to study the time evolution of the
post-mortem changes taking place in the fish muscle by a biochemical and microbiological
approach and their correlation with the sensory change. The work concerned the temperature
(25 ° C, 4 ° C) and the time effect on changes of sardina (Sardina pilchardus) quality.
The first part of this report is consecrated to the biochemical parameters; the dynamics
of production of hydroperoxydes and TBA-rs, appearance of the total volatile basic nitrogen
(TVBN) and the measure of the quantity of total proteins. These parameters show significant
differences (p < 0,05) according to the preservation.
Secondly, the microbial spoiling was studied, so the aerobic mesophilic total flora, the
total coliformes, the faecal coliformes, the thermotolerants streptococci, the staphylococci, the
sulfito-reducing bacteria, the pseudomonas, the salmonellas and shigelles was counted or
isolated. Finally, certain spoilage microbial enzymatic activities as the lipolysis and the
gelatinolysis are also approached. The use of the selective mediums allowed demonstrating
the presence of these activities which appear and disappear at the different moments
according to period and of the mode of preservation.
The follow-up of the kinetic production of the biogenic amine histamine, allowed us to
determine the moment of its appearance and its disappearance as well as its quantification and
to isolate also the bacteria responsible for its production.
We observe the existence of a synergy between the evolution of the biochemical change
and the microbiological spoilage. They were in positive correlation with the loss of the
sensory quality. Besides, it turns out that the refrigeration delays these activities. It is
recommended to store the samples in low-temperature s to protect their quality, and consume
them, at the latest, in two days follow their capture.
Keywords: Hydroperoxydes, TBA-rs, proteins, TVBN, bacterial flora, lipolytic
activity, gelatinolytic activity, histamine, Sardina pilchardus.

ملخص
بعذ يح رنك عضم انسك انذف ي زا انعم انكشف انخببعت انزييت نهخطساث انبجت في
.بقبسبت بي كييبئيت ييكشبينجيت عالقخب ببنخغيش انحسي
فيsardina pilchardus))طشيقت انحفبظ عهي عيت انسشدي حأثيشيقو زا انعم انخجشيبي عهي دساست
.)دسجت يئيت 4دسجت يئيت ،دسجت انخبشيذ 52:دسجت حشاسة انغشفت ) األخيشحغييش عيت زا
إخبجدساست انخغيشاث انبيكييبئيت ديبييكيت إنيي زا انخقشيش يخصص األلانجزء
حػش ز انخغيشاث اخخالف ,،كيت انبشحيبث انكهيتABVT إخبجكزانك TBA-rsانيذسبيشكسيذ
.( p< 0,05)كبيش يهحظ
انخغيشاث انذسست .نيكشبينجيت انجزء انثبي ي انخقشيش خصص نذساست انخغيشاث في انعيت ا
إجبنيت coliformesانخي ح في دسجت حشاسة يخسطت، اإلجبنيتحخص انكبئبث انجشيت انائيت
Staphylocoques ,Streptocoque thermotolérants , coliformes fécaux. بكخيشيب يخجت
Shigelles , Salmonelles, Pseudomonas.نهكبشيج
انسؤل ع ذو انذيبث اإلزيى: انسؤنت ع انخهف انيكشبينجي اإلزييتانغبئف أخيشا
غس اخخفبء ز انغبئف .الحع يى نهبكخيشيب انذسست .انسؤل ع ذو انجيالحي اإلزيى
انسؤنت ع سحج نب بعزل انبيكخيشيب سخذي إخبجيخببعت حشكيت .كزا يشاحم انخخزي يشحبط بكيفيت
.اإلخبجزا
الحع جد عالقت بي انخهف انبيكيبئي انيكشبينجي ،زي االخيشي بذسب يحثب ايجببيب
غس عاليبث انخهف نزا صح ببنخخزي في الحع كزانك ا انخبيشد يقص ايؤخش.انخهف انحسي
.دسجت حشاسة يخفضت
:بث انفخبحيت انكه
غيفت ازييت نذو انذ ،غيفت ايزييت نذو ،ABVT, ،بشحيبثTBA-rs،يذسبيشكسيذ
. Sardina picharrdus , سخذي, ،بيكخيشيب انجيالحي

SOMMAIRE
Page
1. INTRODUCTION………………………………………………………………....
2. ETUDE BIBLIOGRAPHIQUE……………………………………………………
2.1. Présentation de l’espèce cible : La sardine commune (Sardina pilchardus) …...
2.1.1. Position systématique…………………………………………………………..
2.1.2. Biologie de la sardine………………………………………………………….
2.2. Caractéristiques de la chair de poisson ………………………………………….
2.2.1. Généralités……………………………………………………………………..
2.2.1.a. Structure physique …………………………………………………………...
2.2.1.b. Composition chimique …………..…………………………………………..
Composés lipidiques………………………………………………………………..
Composés protéiques……………………………………………………………….
Vitamines et sels minéraux…………………………………………………............
2.2.2. Caractéristiques de la chair de sardine………………………………………...
Acides gras (n-3)…………………………………………………………………...
Protéines……………………………………………………………………............
2.3. Altération de la qualité sensorielle du poisson…………………………………...
2.4. Oxydation des lipides…………………………………………………………….
2.5. Altération microbienne de la chair de poisson………………………………......
2.6. Formation de l’amine biogène (Histamine)……………………………………...
2.6.1. Généralités sur les amines biogènes et l’histamine…………………………….
2.6.2. Les bactéries productrices d’histamine ………………………………………..
2.6.3. Intoxication histaminique………………………………………………………
a. Dose toxique……………………………………………………………………….
b. Aspect réglementaire………………………………………………………………
3. MATERIEL ET METHODES ……………………………………………………
3.1. Echantillonnage et conditionnement…………………………………………….
3.2. Appréciation sensorielle…………………………………………………………
3.3 .Analyses biochimiques…………………………………………………………..
3.3.1. Analyses physico-chimiques…………………………………………………...
3.3.1.a. pH ……………………………………………………………………………
3.3.1. b. Teneur en eau……………………………………………………………….
1
3
3
3
4
6
6
6
7
7
8
8
9
9
9
10
11
13
15
15
18
19
19
20
21
21
21
22
22
22
22

3.3.2. Etude de la peroxydation lipidique…………………………………………….
3.3.2. a. Teneur en hydroperoxydes ………………………………………………….
3.3.2.b. Teneur en TBA-rs …………………………………………………………...
3.3.3. Concentration en protéines totales……………………………………………..
3.3.4. Teneur en azote basique volatil total (ABVT)…………………………………
3.3.5. Teneur en histamine……………………………………………………………
3.4. Analyse microbiologique………………………………………………………..
3.4.1. Echantillonnage………………………………………………………………..
3.4.2. Traitement de l’échantillon : préparation de la solution mère…………………
3.4.3. Numération de la flore aérobie mésophile totale………………………………
3.4.4. Numération des coliformes totaux……………………………………………..
3.4.5. Numération des coliformes fécaux ou coliformes thermotolérants……………
3.4.6. Numération des Streptocoques fécaux ou thermotolérants…………………….
3.4.6.a. Etape 1 ou test présomptif……………………………………………………
3.4.6.a. Etape 2 ou test confirmatif…………………………………………………...
3.4.7. Numération des Clostridium sulfito-réducteurs……………………………….
3.4.8. Recherche de Staphylococcus aureus………………………………………….
3.4.9. Recherche de Salmonella et de Shigella ………………………………………
3.4.10. Recherche de Pseudomonas…………………………………………………..
3.5. Activité enzymatique d’altération d’origine bactérienne………………………..
3.5.1. Recherche de l’activité lipolytique…………………………………………….
3.5. 2. Recherche de l’activité gélatinolytique………………………………………..
3.6. Bactéries productrices d’histamine………………………………………………
3.6.1. Isolement des bactéries productrices d’histamine……………………………...
3.6.2. Purification et essai d’identification……………………………………………
3.7. Etude statistique………………………………………………………………….
4. RESULTATS………………………………………………………………………
4.1. Analyse sensorielle………………………………………………………………
4.2. Caractéristiques biochimiques………………………………………………….
4.2.1. Variation du pH ……………………………………………………………….
4.2.1.1. Effet de la durée de conservation….…………………………………………
4.1.1.2. Effet du mode de conservation………………………………………………
4.2.2. Détermination de la teneur en eau …………………………………………….
22
22
23
23
23
24
24
24
25
25
25
26
26
26
27
27
27
28
28
29
29
29
29
29
30
30
31
31
33
33
33
33
34

4.2.2.a. Effet de la durée de conservation…………………………………………….
4.2.2.b. Effet du mode de conservation……………………………………………….
4.2.3. Evolution du niveau d’oxydation des lipides…………………………………..
4.2.3.a. Teneur en hydroperoxydes…………………………………………………...
* Effet de la durée de conservation…………………………………………………..
* Effet du mode de conservation……………………………………………………..
4.2.3.b. Teneur en TBA-rs……………………………………………………………
* Effet de la durée de conservation…………………………………………………..
* Effet du mode de conservation……………………………………………………..
4.2.4. Estimation de la concentration des protéines totales ………………………….
4.2.4.a. Effet de la durée de conservation…………………………………………….
4.2.4.b. Effet du mode de conservation……………………………………………….
4.2.5. Concentration de l’azote basique volatil total (ABVT) ………………………
4.2.5.a. Effet de la durée de conservation…………………………………………….
4.2.5.b. Effet du mode de conservation………………………………………………
4.2.6. Teneur en histamine……………………………………………………………
4.2.6.a. Effet du temps de conservation………………………………………………
4.2.6.b. Effet du mode de conservation………………………………………………
4.3. Analyse microbiologique………………………………………………….....….
4.3.1. Numération de la flore aérobie mésophile totale………………………………
4.3.2. Numération des coliformes totaux …………………………………………….
4.3.3. Numération des coliformes fécaux ou coliformes thermotolérants……………
4.3.4. Numération des Streptocoques fécaux ou thermotolérants…………………….
4.3.5. Numération des Clostridium sulfito-réducteurs ……………………………….
4.3.6. Présence / numération des Staphylocoques ……………………………………
4.3.7. Présence / numération Salmonelles et Shigelles ………………………………
4.3.8. Présence / numération de Pseudomonas……………………………………….
4.3.9. Activités enzymatiques bactériennes d’altération ……………………………..
4.3.9.a. Activité lipolytique …………………………………………………………..
4.3.9.b. Activité gélatinolytique ……………………………………………………...
4.3.10. Bactéries productrices d’histamine ……………………………………..........
4.3.10.a. Cinétique d’apparition des bactéries histaminogènes………………………
5. DISCUSSION ……………………………………………………………………..
34
35
36
36
36
37
38
38
39
40
40
41
42
42
43
44
44
45
46
46
48
49
50
51
53
56
58
61
61
62
64
64
66

6. CONCLUSION ET PERSPECTIVES …………………………………………….
7. REFERENCES BIBLIOGRAPHIQUES ………………………………………….
8. ANNEXES ………………………………………………………………………...
74
76
94

LISTE DES FIGURES ET DES PLANCHES
Planche 1 : Aspect extérieur de la sardine pendant la conservation.
Figure 1 : La sardine commune : Sardina pilchardus (Walbaum, 1792).
Figure 2 : Estimation de la capture globale mondiale de Sardina pilchardus
(FAO, 2011).
Figure 3 : Auto-oxydation des lipides polyinsaturés (Al-Sayed, 2007).
Figure 5 : Dégradation enzymatique des acides aminés.
Figure 6: Structure de l’histamine.
Figure 7: Evolution temporelle du pH chez la sardine (S. pilchardus). Les
histogrammes portant des lettres et des chiffres différents sont significativement
différents (p < 0,05).
Figure 8 : Effet du mode de conservation sur le pH chez la sardine (S. pilchardus).
Les histogrammes portant des lettres et des chiffres différents sont significativement
différents (p < 0,05).
Figure 9 : Evolution temporelle de la teneur en eau chez la sardine (S. pilchardus).
Les histogrammes portant des lettres et des chiffres différents sont significativement
différents (p < 0,05).
Figure 10 : Effet du mode de conservation sur la teneur en eau chez la sardine (S.
pilchardus). Les histogrammes portant des lettres et différents sont
significativement différents (p < 0,05).
Figure 11 : Evolution temporelle de la teneur en hydroperoxydes (mmoles Eq
CuOOH/g M.H.) chez la sardine (S. pilchardus). Les histogrammes portant des
lettres et des chiffres différents sont significativement différents (p < 0,05).
Figure 12 : effet du mode conservation sur la teneur en hydroperoxydes (mmoles
Eq CuOOH/g M.H. chez la sardine (S. pilchardus). Les histogrammes portant des
lettres et des chiffres différents sont significativement différents (p < 0,05).
Figure 13 : Evolution temporelle de la teneur en TBA rs (mg Eq MDA/ g de M.F.)
chez la sardine (S. pilchardus). Les histogrammes portant des lettres et des chiffres
différents sont significativement différents (p < 0,05).
Figure 14 : effet du mode de conservation sur la teneur en TBA rs (mg Eq MDA/ g
de M.F.) chez la sardine (S. pilchardus). Les histogrammes portant des lettres et
chiffres différents sont significativement différents (p < 0,05).

Figure 15 : Evolution temporelle de la teneur en protéines (g de protéines/ 100 g de
chair) chez la sardine (S. pilchardus). Les histogrammes portant des lettres et des
chiffres différents sont significativement différents (p < 0,05).
Figure 16: Effet du mode de conservation sur la teneur en protéines (g de
protéines/ 100 g de chair) chez la sardine (S. pilchardus). Les histogrammes portant
des lettres et des chiffres différents sont significativement différents (p < 0,05).
Figure 17 : Evolution temporelle de la teneur en ABVT (mg d’ABVT / 100 g de
chair) chez la sardine (S. pilchardus). Les histogrammes portant des lettres et des
chiffres différents sont significativement différents (p < 0,05).
Figure 18 : Effet du mode de conservation sur la teneur en ABVT (mg d’ABVT /
100 g de chair) chez la sardine (S. pilchardus). Les histogrammes portant des lettres
et chiffres différents sont significativement différents (p < 0,05).
Figure 19 : Evolution temporelle de la teneur en histamine (mg/100gM.H.) chez la
sardine (S. pilchardus). Les histogrammes portant des lettres et des chiffres
différents sont significativement différents (p < 0,05).
Figure 20 : Effet du mode de conservation sur la teneur en histamine
(mg/100gM.H.) chez la sardine (S. pilchardus). Les histogrammes portant des
lettres et chiffres différents sont significativement différents (p < 0,05).
Figure 21 : Evolution temporelle de la flore aérobie mésophile totale chez la
sardine (S. pilchardus).
Figure 22 : Aspect des colonies de bactéries isolées sur milieu Plate Count Agar
(P.C.A) pour dénombrement.
Figure 23 : Evolution temporelle de la teneur des coliformes totaux chez la sardine
(S. pilchadus).
Figure 24 : Aspect du bouillon au pourpre de bromocrésol (BCPL) avant et après
incubation (virage du milieu au jaune et production de gaz récolté dans la cloche de
Durham).
Figure 25 : Evolution temporelle de la teneur des coliformes fécaux chez la sardine
(S. pilchardus).
Figure 26: Evolution temporelle de la teneur des streptocoques thermotolérants
chez la sardine (S. pilchardus).
Figure 27 : Aspect des colonies de bactéries isolées sur milieu Chapman.
Figure 28 : Aspect des colonies de bactéries isolées sur milieu King A.

Figure 29 : Aspect des colonies de bactéries isolées sur milieu King B.
Figure 30: Evolution temporelle de l’activité lipolytique bactérienne chez la
sardine (S. pilchardus).
Figure 31 : Aspect macroscopique de la manifestation de l’activité lipolytique.
Figure 32: Evolution temporelle de l’activité gélatinolytique bactérienne chez la
sardine (S. pilchardus).
Figure 33 : Aspect macroscopique de la manifestation de l’activité gélatinolytique.
Figure 34 : Aspect d’une colonie de bactéries aminogènes.

LISTE DES TABLEAUX
Tableau 1 : composition proximale d’une portion de sardine (Santé Canada, 2005)
Tableau 2 : Dénombrement bactérien sur la sardine fraiche (cfu/g) selon
Abbabouch, 1996.
Tableau 3: Quelques espèces de poissons pouvant causer une intoxication
histaminique (Boucher et Djabali, 1998)
Tableau 4 : Principales bactéries productrices d’amines biogènes (Boulay, 1991).
Tableau 5 : Evolution temporelle de la concentration cellulaire des clostridium
sulfito-réducteurs chez la sardine (S. pilchardus) conservée à température ambiante
/ réfrigérée et réfrigérée.
Tableau 6 : Evolution temporelle des Staphylocoques sur milieu Chapman chez la
sardine (S. pilchardus) conservée à température ambiante.
Tableau 7 : Evolution temporelle des Staphylocoques sur milieu Chapman chez la
sardine (S. pilchardus) à réfrigérée.
Tableau 8 : Evolution temporelle des isolats sur milieu S-S chez la sardine (S.
pilchadus) à température ambiante et réfrigérée.
Tableau 9 : Evolution temporelle des Pseudomonas chez la sardine (S. pilchardus)
à température ambiante.
Tableau 10 : Evolution temporelle des Pseudomonas chez la sardine (S. pilchardus)
conservée à température réfrigérée.
Tableau 11 : Tableau récapitulatif de l’évolution temporelle de l’apparition et de la
disparition des bactéries productrices d’amines biogènes chez la sardine (S.
pilchardus).

LISTE DES ABREVIATIONS
ABVT : Azote basique volatil total.
A.G.: Acide gras.
A.G.P.I.: Acide gras polyinsaturé.
A.D.P.: Adénosine diphosphate.
A.H.A: American Heart Association.
A.M.P. : Adénosine monophosphate.
AOCA : Association Officielle des Chimistes Analystes.
A.T.P. : Adénosine triphosphate.
B.H.T.: 3,5-ditertiobutyl-4-hydroxytoluène.
D.H.A : Docosahexaécoique.
E.P.A : Eicosapentaénoique.
F.A.O.: Food and Agriculture Organization.
FDA: Food and Drug Administration.
H2S: Sulfure d’hydrogène.
Hx : Hypoxantine.
I.M.P. : Inosine monophosphate.
M.D.A. : Malondialdéhyde.
M.H. : Matière humide.
M.S. : Matière sèche.
O.T.M.A. : Oxyde de triméthylamine.
P.C.: Phosphatidylcholine.
P.L.: Phospholipide.
T.G.: Triglyceride.

TBA-rs : Thiobarbituric acid reactive substances (substances réactives à l’acide
thiobarbiturique).
T.M.A. : Triméthylamine.
U.F.C : Unité formant colonie.
U.F.T : Unité formant trouble.

Introduction

Introduction
1
1. INTRODUCTION
Depuis toujours, dans de nombreuses régions du monde, les produits de la mer font
partie du régime alimentaire quand ils ne constituent pas, comme c’est le cas dans certains
pays, la principale source de protéines animales. De nos jours, de plus en plus nombreux sont
ceux qui voient dans le poisson un substitut à la viande rouge, jugée meilleure pour la santé
(Martin, 2001 ; AFSSA, 2003). La faible teneur en matières grasses de plusieurs espèces de
poissons et les effets sur les cardiopathies coronariennes des acides gras (n-3) polyinsaturés
(Dumay, 2006), constituent des caractéristiques extrêmement précieuses d’un point de vue
sanitaire, notamment dans les pays où la mortalité par les maladies cardiovasculaires est la
plus élevée. Les protéines très digestes et la grande variété de sels minéraux, d'oligo-éléments
et des vitamines sont des constantes caractéristiques de la chair de poisson qui en font un
produit unique dans le monde animal.
La qualité et la sécurité des produits de la pêche et de l’aquaculture sont des éléments
cruciaux tout au long de la chaîne, depuis la production primaire jusqu’à l’usager final. Les
produits de la mer englobent une grande diversité d’espèces qui va des poissons aux
mollusques et crustacés. Différents risques sont liés à chacune des espèces et peuvent en
influencer la qualité et la sécurité du consommateur. L'amélioration de la qualité des produits
de la pêche et de l’aquaculture est devenue une préoccupation majeure des pouvoirs publics et
de tous les acteurs opérant dans ce domaine. Le contrôle des taux de substances générées dans
les tissus des poissons, destinées à la consommation, telles que l'histamine, l'azote basique
volatil total « ABVT » et récemment les métaux lourds (Hg, Cd, ...), est devenu obligatoire.
La demande des consommateurs a conduit à la mise en place de nouveaux procédés,
comme le traitement minimal des produits et la production d’aliments prêts à l’emploi. Une
profonde connaissance de la composition du poisson et des modifications qui surviennent lors
de la manipulation, de la transformation et du stockage est un élément essentiel dans la prise
de décisions concernant les méthodologies à mettre en place pour l’évaluation de la qualité et
de la sécurité.
En effet, les produits de la pêche subissent une dégradation naturelle post mortem,
résultante de réactions endogènes et exogènes. Ces dernières sont de natures chimiques,
enzymatiques et microbiologiques (Koutsoumanis et al., 2002). Cette altération est
dépendante en grande partie des conditions de stockage qui favorisent davantage ces
réactions. L’absence d’infrastructures adéquates accentue la détérioration des produits et

Introduction
2
réduit de manière conséquente leur valeur. On assiste à des hydrolyses lipidiques,
protéiques,….etc, dont la résultante est une multitude de produits responsables de l’altération
de la qualité organoleptique, sanitaire, nutritionnelle du produits, dont certains constituent un
danger mortel telles que les intoxications histaminiques (Parente et al., 2001) .
Nous assistons partout dans le monde à un développement rapide des méthodes
microbiologiques, chimiques et autres, ce qui a permis d’améliorer notablement les
performances des analyses en matière de qualité des produits de la mer.
C’est dans ce contexte que s’insère notre travail expérimental qui vise à développer des
outils de diagnostic permettant d’évaluer les niveaux d’altération et de déterminer les points
critiques de la conservation des produits de la pêche et de l’aquaculture. Dans notre travail
nous nous sommes intéressés à:
L’évaluation dans le temps, de la dynamique de production des produits
d’oxydation lipidique primaire et secondaire à savoir ; les hydroperoxydes et les
TBA-rs.
La détermination de l’état hygiénique microbiologique de nos échantillons.
L’estimation des teneurs des protéines ainsi que de l’ABVT produit au cours du
processus de conservation.
L’isolement des bactéries productrices de l’histamine, leur purification et essai
de leur identification.
Le suivi dans le temps de la dynamique de production de l’histamine par HPLC.
La corrélation entre les paramètres organoleptiques, biochimiques et
microbiologiques dans l’appréciation de la qualité de l’espèce cible.

ETUDE
BIBLIOGRAPHIQUE

Etude bibliographique
3
2. ETUDE BIBLIOGRAPHIQUE
2.1. Présentation de l’espèce cible : La sardine commune (Sardina pilchardus) (figure 1)
2.1.1. Position systématique
Embranchement : Vertebrés
Sous embranchement : Gnathostomes
Super classe : Poissons
Classe : Ostéichtyens
Sous classe : Téléostéens
Super ordre : Clupéiformes
Ordre : Clupéoides
Famille : Clupéidés
Genre : Sardina
Espèce : Sardina pilchardus (Walbaum, 1792)
Figure 1 : La sardine commune : Sardina pilchardus (Walbaum, 1792)
1cm
AQUABIOR, 2011

Etude bibliographique
4
2.1.2. Biologie de la sardine
La sardine (S. pilchardus) est un poisson de la famille des Clupeidae, qui comprend
également le hareng et l'alose. Selon la région, elle prend les noms de célan, célerin, pilchard,
sarda, sardinyola. Son nom provient de la Sardaigne car les Grecs avaient remarqué qu'elle
abondait dans ces eaux côtières. Ce n'est pas une espèce considérée comme menacée, mais à
la suite d'une surpêche elle a fortement régressé dans une grande partie de son aire de
répartition où elle a été très abondante jusqu'au début du XXe siècle.
La sardine commune (S. pilchardus) est un poisson pélagique vivant dans les eaux côtières
et jusqu'à 120 m de profondeur. Elle vit en bancs parfois très compacts, près de la surface la
nuit et plus en profondeur le jour, entre 10 et 50 mètres. Elle mesure au plus une vingtaine de
centimètres de long, et possède un ventre argenté et un dos bleuté. Elle se caractérise par un
opercule strié, des tâches sombres sur le dos, une carène ventrale peu aigüe, des écailles
sessiles, les deux derniers rayons de l’arête anale plus longs.
Ce poisson vit en Méditerranée et dans presque tout l'Atlantique nord, de l'Irlande jusqu'aux
Açores, en zone tropicale (le Sénégal et la Mauritanie) aux côtes Atlantiques Marocaines et
européennes en zone pélagique côtière de 15 à 35 m de profondeur. En méditerranée, elle
constitue le second poisson le plus pêché (16 %) parmi les petits pélagiques (qui constituent
50 % de la pêche totale), loin derrière l'anchois (Engraulis encrasicolus) qui constitue 59 %
des captures de petits pélagiques (FAO, 2009). En Méditerranée, la situation dans son
ensemble est restée stable. Les principaux stocks de petits pélagiques (sardines et anchois)
sont pleinement exploités (figure 2) (FAO, 2011).

Etude bibliographique
5
Figure 2 : Estimation de la capture globale mondiale de Sardina pilchardus (FAO, 2011)
La reproduction a lieu en haute mer et peut advenir toute l'année, avec une période de ponte
variant en fonction de la répartition géographique (Dumay, 2006). Au large de l'Afrique, la
reproduction semble maximale de novembre à février (Amenzoui, 2005). La sardine se
reproduit dans une eau dont la température varie de 16°C à 19°C. La réussite de la
reproduction dépend, aussi, des remontées d'eau froide ou upwellings. Après une phase
planctonique, les alevins rejoignent les côtes au printemps, et y restent jusqu'au début de
l'hiver.
La pêche à la sardine remonte à l'Antiquité et est une activité fortement influencée par les
conditions hydrologiques. En effet, la température agit directement sur les migrations ainsi
que sur l’importance et la localisation des concentrations de sardines et, donc sur leur
accessibilité aux flottilles de pêche. La sardine est pêchée à la boliche et de plus en plus, au
chalut pélagique. Elle est consommée fraîche, salée, parfois fumée mais principalement en
conserve.
La sardine est un poisson de grande consommation. Elle est commercialisée dans sa majeure
partie en Algérie dans les centres urbains les plus importants (Alger, Annaba, Oran,
Constantine…). En effet, elle jouit d’excellents apports nutritionnels ainsi que d’un prix
1950 1960 1970 1980 1990 2000 2010
1600
1400
1200
1000
800
600
400
200
0
Tonnes (x1000)
Année

Etude bibliographique
6
raisonnable et convenable à toutes les bourses. Cependant, nous assistons récemment à une
diminution des débarquements accompagnée d’une flambée des prix atteignant jusqu’à
600 D.A. le kilogramme.
2.2. Caractéristiques de la chair de poisson
2.2.1. Généralités
La chair de poisson présente une structure similaire à celle des autres animaux. Toutefois,
elle est particulière de part la structure physique et de la composition chimique spécifique des
poissons.
2.2.1.a. Structure physique
Le muscle de poisson se compose essentiellement de deux types de muscles : le muscle blanc
et le muscle rouge.
La plus grande partie du tissu musculaire est blanche. Cependant, on rencontre chez de
nombreux poissons une certaine quantité de tissu brun ou rougeâtre. Le muscle rouge forme
une lame sous-cutanée et dans le cas de certaines espèces actives, une bande près de l’épine
dorsale. La proportion du muscle rouge (type oxydatif) par rapport au muscle blanc (type
glycolytique) varie selon l’activité du poisson. Ainsi, chez les poissons pélagiques tels que la
sardine, le muscle rouge peut représenter jusqu’à 48% du poids du poisson (Love, 1980).
Chez les poissons dérmersaux, qui se nourrissent au fond des mers et ne se déplacent que
périodiquement, ce taux est trop faible. La composition chimique des deux types de muscles
est également différente. Les muscles rouges contiennent des taux élevés de lipides,
d’hémoglobines, de glycogène et de la plupart des vitamines. La teneur élevée en lipides
présente un intérêt technologique particulier en raison des problèmes de rancidité.
Les muscles rouges et les muscles blancs possèdent des fonctions différentes. Il est estimé que
les premiers sont essentiellement des muscles de croisière, c'est-à-dire utilisés pour les
mouvements continus et lents alors que les seconds sont des muscles de propulsion servant
aux mouvements rapides et soudains.

Etude bibliographique
7
2.2.1.b. Composition chimique
La composition chimique du poisson varie considérablement d’une espèce et d’un individu à
l’autre selon l’âge, le sexe, l’environnement et la saison. Les variations de la composition
chimique du poisson sont étroitements liées à son alimentation, aux déplacements migratoires
et aux changements sexuels en rapport avec la ponte.
Composés lipidiques
Les lipides présents dans les espèces de poissons téléostéens peuvent être divisés en deux
groupes principaux: les phospholipides et les triglycérides. Les phospholipides constituent la
structure intégrale des membranes cellulaires et sont de ce fait appelés souvent lipides
structuraux. Les triglycérides sont des lipides utilisés pour entreposer l’énergie dans les dépôts
de graisse, habituellement à l’intérieur des cellules grasses spéciales entourées d’une
membrane de phospholipides et d’un réseau assez faible de collagène.
Les poissons peuvent être classés en espèces maigres ou grasses selon l’organe de stockage.
Les poissons maigres utilisent le foie comme réservoir d’énergie tandis que les poissons gras
répartissent leurs lipides dans les cellules grasses situées au niveau du tissu sous-cutané, les
muscles de la paroi abdominale et dans les muscles animant les nageoires et la queue. Chez
certaines espèces, qui stockent des quantités extrêmement importantes de lipides, la graisse
est localisée dans la cavité abdominale.
Le contenu élevé en acides gras (n-3) dans les poissons, leur confère des avantages
incontestables pour la santé. La littérature scientifique abonde à ce sujet, et l’impact de la
consommation de poissons sur la diminution du risque de maladies cardiovasculaires fait
maintenant l’unanimité auprès des chercheurs (Calder, 2004). Des études ont démontré que
les personnes consommant plus de poissons présentaient moins de cas de dépression (Ness,
2003) et moins de risques d’être atteints de la maladie d’Alzheimer (Morris, 2003). De plus,
d’autres études ont montré un lien entre la consommation de poissons gras et la diminution de
l’incidence de l’arthrite (Pedersen, 2005 et Pattison, 2004). L’American Heart Association
(AHA) recommande aux adultes de consommer au moins deux repas de poisson par semaine,
principalement les poissons gras, afin de profiter de leurs effets bénéfiques sur la santé
(Krauss, 1992).

Etude bibliographique
8
Composés protéiques
Les protéines des tissus musculaires du poisson peuvent être divisées en trois groupes:
Les protéines structurelles (actine, myosine, tropomyosine et actomyosine),
constituent de 70 à 80 % de la teneur totale en protéines (comparée à 40 % chez les
mammifères).
Les protéines sarcoplasmiques (myoalbumine, globuline et enzymes) représentent
25 à 30 % des protéines.
Les protéines du tissu conjonctif (collagène) constituent environ 3 % des protéines
chez les téléostéens et environ 10 % chez les élasmobranches (comparé à 17 % chez
les mammifères).
La composition en acides aminés du muscle des poissons est approximativement la même que
pour les protéines correspondantes dans le muscle des mammifères bien que les propriétés
physiques puissent être légèrement différentes. En effet, le point iso-électrique (pI) se situe
aux environs d’un pH de 4,5 à 5,5.
Par ailleurs, plusieurs études ont révélé que la consommation de protéine de poisson, en
l’occurrence la protéine de morue, améliorerait la sensibilité à l'insuline et augmenterait
l’absorption du glucose par l’organisme (Lavigne, 2001).
Chez certaines espèces, on note la présence de protéines basiques (protéines riches en acides
aminés basiques) qui jouent un rôle dans l’inhibition de la croissance bactérienne (Kamal-
Eldin, 2002), ainsi qu’à l’amélioration des propriétés fonctionnelles des autres protéines
alimentaires (Philips et al., 2002).
Vitamines et sels minéraux
La teneur en vitamines et sels minéraux est spécifique aux espèces et peut, varier selon la
saison. En général, la chair du poisson est une source en vitamines B et également, dans le cas
des espèces grasses, en vitamines A et D. En ce qui concerne les éléments minéraux, la chair
du poisson est considérée comme une source appréciable en calcium et en phosphore en
particulier mais également en fer, cuivre et sélénium. Les poissons d’eau de mer ont une forte
teneur en iode.

Etude bibliographique
9
2.2.2. Caractéristiques de la chair de sardine
Acides gras (n-3)
La sardine est une excellente source d’acide eicosapentaénoïque (EPA) et d’acide
docosahexaénoïque (DHA) (tableau1). Ces acides gras agissent comme précurseurs de
messagers chimiques favorisant le bon fonctionnement du système immunitaire, circulatoire
et hormonal. Plusieurs études épidémiologiques et cliniques ont démontré que la
consommation d’acides gras (n-3) exerçait des effets favorables sur la santé et réduisait la
mortalité par les maladies cardiovasculaires (Calder, 2004). Ces acides gras sont connus pour
agir de plusieurs façons sur l’organisme, notamment en réduisant la tension artérielle, les
triglycérides sanguins et la formation de caillots sanguins, diminuant ainsi les risques
d’athérosclérose. La consommation régulière d’acides gras (n-3) diminuerait l’arythmie
cardiaque et pourrait même inhiber la croissance des cellules cancéreuses (Horrocks, 1999 et
Larsson, 2004).
La sardine est l’un des six poissons les plus riches en acides gras EPA et DHA, avec la truite
(Oncorhynchus mykiss) , le maquereau (Scomber scombrus), le thon (Thunnus alalunga), le
hareng (Clupea harengus) et le saumon (Salumo salar). Il est bon de noter que le contenu en
lipides et en acides gras (n-3) de la sardine varie considérablement selon la saison. Elles sont
plus riches en lipides en été et moins en hiver (Macciola, 2004).
Protéines
La protéine de sardine a été isolée et distribuée à des animaux afin d’évaluer son impact sur le
sang (Murata, 2004). Les résultats ont montré que la fibrinolyse et le temps de coagulation du
sang augmentaient. Ces effets pourraient être bénéfiques pour les individus à risque de
thromboses.

Etude bibliographique
10
Tableau 1 : Composition proximale d’une portion de sardine (Santé Canada, 2005).
* EPA, DHA et acide alpha-linolénique(LNA)
2.3. Altération de la qualité sensorielle du poisson
L'évaluation sensorielle permet de déterminer l’état de fraîcheur des produits à consommer et
d’estimer le moment de rejet organoleptique.
L’évaluation sensorielle du poisson frais sur les marchés et aux débarcadères se fait en
vérifiant l’aspect, la texture et l’odeur. La plupart des systèmes d’évaluation se basent sur les
modifications qui se produisent pendant le stockage dans la glace fondante. Il faut rappeler
que les changements dépendent des méthodes de stockage. L’aspect du poisson stocké au
froid sans glace ne change pas autant que celui du poisson sous glace. De ce fait, une
connaissance des conditions temps/température de conservation s’avère nécessaire au
débarquement.
Le changement le plus important est l’établissement de la rigor-mortis. Immédiatement après
la mort, le muscle est totalement détendu et la texture élastique et souple dure habituellement
quelques heures, après quoi le muscle se contracte. Quand il durcit, le corps se raidit et le
poisson est alors en état de rigor-mortis. Cet état dure habituellement un jour ou plus et alors
la rigor disparaît, ce qui détend le muscle à nouveau et le rend souple mais il n’est plus aussi
Poids/volume Pour 100 g de sardine
Calories 208 k cal
Protéines 24,6 g
Glucides 0,0 g
Lipides 11,5 g
- saturés 1,5 g
- monoinsaturés 3,9 g
- polyinsaturés 5,2 g
- oméga-3* 1,5 g
Cholestérol 142 mg
Fibres alimentaires 0,0 g

Etude bibliographique
11
élastique qu’avant la rigor. Le rapport entre l’apparition et la disparition de la rigor varie
d’une espèce à l’autre et est affecté par la température, la manutention, la taille et la condition
physique du poisson. Les directives de la CEE, élaborées à cet effet donnent une description
générale pour l’examen de la qualité du poisson (voir annexe : p102).
2.4. Oxydation des lipides
La stabilité des lipides vis-à-vis de l’oxydation dépend de leur localisation dans les différents
tissus (Eymard, 2003). Lorsque le tissu est vivant, il existe des mécanismes naturels de
contrôle qui permettent de prévenir la destruction oxydative des lipides membranaires, des
protéines et des acides nucléiques. Le dérèglement intervient lors de la destruction du tissu
(Hultin, 1994), notamment lors de la dégradation des membranes (Hwang et al., 1993).
Concernant les lipides, les facteurs qui influencent l‘oxydation sont nombreux : composition
en acides gras, nombre et position des insaturations, présence de pro-oxydant (ions
métalliques, enzymes) ou d’antioxydants naturels (tocophérols, caroténoides….), la surface
exposée à l’oxygène, la température, la lumière, etc….
L’oxydation des lipides peut résulter de causes multiples. Ainsi l’auto-oxydation est catalysée
par la température, les ions métalliques, les radicaux libres. La photo-oxydation est initiée par
la lumière en présence de photo-sensibilisateurs. L’oxydation enzymatique est initiée par la
lipoxygénase.
Il s’agit dans tous les cas d’une réaction auto-catalytique, par enchainement de réactions
radicalaires se déroulant en trois étapes. Une première réaction produit un radical libre par
élimination d’un hydrogène de l’acide gras en présence de lumière ou d’oxygène (initiation).
Elle se poursuit par une étape de propagation (réaction en chaine) et s’arrête par une phase de
terminaison quand les substrats radicalaires sont épuisés (figure 3).

Etude bibliographique
12
H° : Hydrogène
L° : Radical libre de lipide
AH : Antioxydant
A° : Radical libre
Figure 3 : Auto-oxydation des lipides polyinsaturés (Al-Sayed, 2007)
Pendant les phases d’initiation et de propagation, des hydroperoxydes apparaissent. Ces
molécules sont des intermédiaires importants et sont systématiquement dosés lors des
analyses d’oxydation. Cependant, ce sont des composés instables qui se dissocient par
scission homolytique de la liaison O-O pour former un radical alcoyl et un radical hydroxyl
qui interviennent dans une série de réactions complexes aboutissant notamment à la formation
de composés secondaires, tels que des aldéhydes et des hydrocarbures. L’analyse des
peroxydes n’est donc pas un critère suffisant pour déterminer l’état oxydatif du milieu (Al-
Sayed, 2007).
LH (chaine acyle d’acide gras)
L° LOO° Propagation LOOH
LOOH
(hydroperoxydes)
LH
Terminaison
Produits secondaires
(Aldéhydes, cétones, alcools à chaine
courte, alcanes)
H° Initiation
(Antioxydant)
AH A°

Etude bibliographique
13
D’autres indicateurs doivent être utilisés. C’est le cas des diènes, de polyènes et d’autres
méthodes indirectes, comme la détermination du taux de caroténoïdes (astaxanthine, par
exemple) et de l’évolution de la couleur. Une technique prometteuse, quoique coûteuse, est
l’analyse par spectrométrie infra-rouge à transformée de Fourier. Une investigation fine de
certaines bandes met en évidence des caractéristiques extrêmement précises de l’état
d’oxydation des molécules.
Le phénomène d’oxydation des acides gras insaturés peut également être d’origine
enzymatique. La lipoxygénase et la cyclo-oxygénase sont les deux enzymes principales
impliquées (Hultin, 1994). Les substrats privilégiés de la lipoxygénase de poisson sont les
acides gras polyinsaturés.
L’oxydation se produit même aux températures les plus basses. En effet, à l’état congelé
cette activité est très faible. Une fois la décongélation amorcée et à des températures variant
entre 0°C et 4°C, cette activité reprend et s’accentue (Frankel, 1998). Les lipoxygénases du
poisson sont inactivées à des températures supérieures à 60°C, mais c’est, alors, l’oxydation
non enzymatique qui est favorisée.
2.5. Altération microbienne de la chair de poisson
On doit faire la distinction entre les termes flore d’altération et bactérie d’altération. La
première décrit simplement les bactéries présentes sur le poisson quand il s’altère tandis que
la seconde constitue le groupe spécifique qui produit les odeurs et les goûts désagréables
associés à la dégradation. Une grande partie des bactéries présentes sur le poisson altéré ne
joue aucun rôle dans son altération. Chaque poisson possède ses propres bactéries spécifiques
d’altération dont le nombre à l'opposé du nombre total de bactéries, sera en rapport avec la
durée de conservation.
La chair du poisson sain, vivant ou fraîchement pêché, est stérile car le système immunitaire
du poisson empêche les bactéries de se multiplier et proliférer dans la chair. A la mort du
poisson, le système immunitaire s’effondre et les bactéries peuvent proliférer librement. A la
surface de la peau, les bactéries colonisent largement les alvéoles des écailles. Pendant le
stockage, elles envahissent la chair en se déplaçant entre les fibres musculaires. Murray et
Shewan (2000) ont montré que seul un nombre très limité de bactéries envahissent la chair
pendant la conservation sous glace. Ruskol et Bendsen (1992) ont montré que les bactéries
peuvent être détectées au microscope dans la chair, stockée sous glace ou à température
ambiante, quand le nombre d’organismes à la surface de la peau dépasse 106 UFC/cm².

Etude bibliographique
14
Aucune différence du mode d'invasion n'a été détectée dans les bactéries spécifiques
d’altération (S. putrefaciens) et les autres bactéries.
Du fait que seul un nombre limité d’organismes envahit réellement la chair et que le
développement microbien (tableau 2) se situe essentiellement à la surface, l’altération s’avère
probablement, pour une grande part, être une conséquence de la diffusion des enzymes
bactériennes dans la chair et des nutriments résultants de leur activité à l’extérieur.
Tableau 2 : Dénombrement bactérien sur la sardine fraiche (cfu/g) selon Abbabouch, 1996.
Flore
Bactérienne
Sardines conservées dans la glace Sardines non conservées dans la glace
Muscle branchies Peau Viscères Muscle branchies Peau viscères
Mésophile 2.104 10
7 4.10
4 4.10
3 10
5 6.10
6 1,6.10
6 3.10
5
Psychrophile 2.104 9.10
6 3.10
5 4,3.10
4 1,6.10
5 5.10
6 2.10
6 4.10
5
Le poisson s’altère à des vitesses très variables. Certains auteurs expliquent ce fait par des
différences dans les propriétés de la surface du poisson. La peau des poissons est de texture
différente selon l’espèce. Le merlan (Merlangius merlangus) et la morue (Gadus
morhua) s’abiment très rapidement, en raison de leur tégument fragile, comparativement aux
poissons plats. En effet, le carrelet (Pleuronectes platessa) possède un derme et un épiderme
très robustes. Ce dernier a de plus une couche épaisse de mucus qui comprend plusieurs
substances antibactériennes tels que des anticorps et des enzymes bactériolytiques (Murray et
Fletcher, 1976).

Etude bibliographique
15
2.6. Formation de l’amine biogène (Histamine)
2.6.1. Généralités sur les amines biogènes et l’histamine
Les amines biogènes sont, des composés azotés basiques non volatils, formés essentiellement
par décarboxylation des acides aminés (figure 5), par amination et transamination des
aldéhydes et des cétones (Maijala et al., 1993) ou hydrolyse de composés azotés par des
enzymes endogènes ou exogènes. Ce sont des bases organiques avec un poids moléculaire
faible et sont des métabolites normaux de microorganismes, de plantes et d’animaux (Brink
et al., 1990)
Figure 5 : Dégradation enzymatique des acides aminés.
On peut considérer que ces molécules sont des traceurs de la dégradation des aliments,
particulièrement celle des protéines, et traduisent, ainsi, la fraîcheur des aliments. La structure
chimique des amines biogènes peut être: aliphatique (putrescine, cadaverine, spermine,
spermidine), aromatique (tyramine, phenylethylamine) ou hétérocyclique (histamine,
tryptamine). Les amines telles que la putrescine, la spermidine, la spermine et la cadavérine
sont des composés indispensables à la survie cellulaire et sont importants dans la régulation
du fonctionnement des acides nucléiques et la synthèse des protéines et probablement dans la
stabilisation membranaire (Bardóz et al., 1993; Maijala et al., 1993; Halász et al.,1994).
L'histamine a été découverte par Akerman en 1910 dans les produits résultant de la
putréfaction bactérienne. Les biochimistes ont cependant pu la mettre en évidence dans tous
les tissus animaux. Elle fait partie des amines biogènes, définies comme des molécules
biologiquement actives sur le système nerveux central et le système vasculaire, c’est une
molécule thermostable.
Mis à part celle qui existe naturellement en faible quantité dans les tissus, qui est préformée et
d'origine physiologique, et celle qui se forme pendant l'autolyse, l'histamine provient
essentiellement de la décarboxylation bactérienne de l'histidine. Ainsi Kimata et Kawai
(1951) ont montré que, dans les conditions les plus favorables, dans les poissons à chair
rouge, riches en histidine, la formation d'histamine par autolyse ne dépasse pas 15 mg/100 g
R-CH-COOH
NH2
Acide aminé
R-CH2-NH2
Amine biogène
Décarboxylation
(amino-acide-décarboxylase)

Etude bibliographique
16
et n'explique pas la disparition d'histidine observée. Parmi les nombreuses souches de
bactéries provoquant l'altération du poisson, Kimata et Kawai (1951) n'en ont trouvé qu'une,
Achromobactec histamineum, qui produise de l'histamine de façon notable à partir de
l'histidine.
Figure 6: Structure de l’histamine
Les éléments essentiels à la formation de l’histamine par les microorganismes sont :
- La disponibilité de l’histidine (Marklinder et Lönner, 1992).
- Présence de microorganismes décarboxylase-positive (Brink et al.,1990).
- Conditions favorables à la croissance bactérienne (notamment la température adéquate et le
pH).
Bien que de nombreuses amines biogènes aient été trouvées dans les poissons, seules
l'histamine, la cadavérine et la putrescine ont été principalement décrites comme ayant un
impact significatif sur la sécurité et de la qualité des produits. L'histamine présente dans le
poisson (tableau 3) est bien évidemment liée à la scombrotoxicité, mais elle ne saurait être le
seul agent responsable de cette intoxication alimentaire. La cadavérine et la putrescine
joueraient un rôle potentialisant son effet toxique. De manière générale on ne peut établir un
seuil de putréfaction à partir de la seule concentration d’une amine biogène. Selon Valle et al.
(2004) l’indice de putréfaction doit intégrer donc les amines volatiles (TMA, ammoniaque) et
les amines biogènes (histamine, tyramine, spermine, spermidine,….)

Etude bibliographique
17
Tableau 3: Quelques espèces de poissons pouvant causer une intoxication histaminique
(Boucher et Djabali, 1998)
Famille Espèces Noms communs en français
Ampidae Ampis trutta Loup de mer
Carangidae Seriola dumerili
Seriola lalandii
Seriole, limon
Coryphaenidae Coryphaena hippurus Coryphène, mahimahi
Clupeidae Sardinella sirm
Amblygaster sirm
Sardinops sp.
Sardina pilchardus
Clupea harengus
Anchois de norvège
Sprat
Sardinelle tachetée
Pilchard ou sardine
Hareng
Istiophoridae Makaira audax Makaire
Pomatomidae Pomatomus saltatris Poisson-serre
Scomberesocidae Cololabis saira Balaou japomais, scombérésoce,
samana
Scombridae Euthynus alleteratus
Katsuwonus pelamis
Sarda sarda
Scomber japonicus
Scomber scombrus
Scomberomorus cavalla
Scomber maculatus
Scomber regalis
Thunnus alalunga
Thunnus albacores
Thunnus obesus
Thunnus thynnus
Auxide
Bonitou
Thonine
Maquereau espagnol
Maquereau
Maquereau roi
Tassard tacheté
Tassard royal
Thon blanc
Albacore
Patudo
Thon rouge
Salmonidae Salmo salar
Oncorhynchus mykiss
Saumon
Truite arc-en-ciel
Siluridae Siturus glanis Poisson-chat, silure
Xiphiidae Xiphia gladius Espadon

Etude bibliographique
18
2.6.2. Les bactéries productrices d’histamine
L’histamine résulte essentiellement de l’activité des enzymes bactériennes. En effet,
Fernandez et Mackie (1979) montrent que l’ajout d’agents antimicrobiens (chloroforme à 3%
et toluène à 8%) sur des échantillons de maquereau empêche la formation d’histamine. La
flore aminogène (tableau 4) est très variée dans sa composition : des bacilles Gram-, des
bacilles Gram+ et des coques Gram+, mais ce sont les entérobactéries qui prédominent. Par
ailleurs, ces bactéries sont détectables quelle que soit la température de conservation du
poisson.
Tableau 4 : Principales bactéries productrices d’amines biogènes (Boulay, 1991).
Entérobactéries Autres bactéries
Bactéries fortes
productrices
Proteus morganii
Klebsielle pneumoniae
Klebsiella oxytoca
Enterobacter aerogenes
Clostridium perfringens
Staphylococcus sp.
Bactéries faibles
productrices
Hafnia alvei
Citrobacter freundii
Serratia spp.
Plesiomonas shigelloides
Escherichia coli
Vibrio spp.
Photobacterium phosphoreum
Pseudomonas spp.
Lactobacillus sp.
Bacillus sp.
Bactéries
potentiellement
productrices
Proteus mirabilis Streptococcus faecalis
Acinetobacter lwoffi
Aeromonas sp.

Etude bibliographique
19
2.6.3. Intoxication histaminique
L’intoxication histaminique, ou syndrome de pseudo-allergie alimentaire, provient de la
consommation d’aliments renfermant de fortes quantités d’histamine qui peut induire des
effets toxiques dans l’organisme.
a. Dose toxique
La dose seuil entraînant le débordement des systèmes de détoxication est très difficile à
déterminer. Elle dépend de multiples facteurs dont la variabilité individuelle. La
réglementation (le règlement CE n° 2073/2005 du 15 novembre 2005 concernant les critères
microbiologiques applicables aux denrées alimentaires qui définit les limites de
concentrations à ne pas dépasser pour l’histamine) fixe les règles sanitaires régissant la
production et la mise sur le marché des produits de la pêche comme suit :
Lors d’un plan de surveillance, neuf échantillons sont prélevés sur chaque lot :
- la teneur moyenne ne doit pas dépasser 100 mg.kg-1
- deux échantillons peuvent dépasser 100 mg.kg-1 sans atteindre 200 mg.kg-1
- aucun échantillon ne doit dépasser 200 mg.kg-1
.
Ces limites s’appliquent seulement aux poissons des familles suivantes :
Scombridés : maquereaux, auxides, thonites, thons, thazards, palomettes, bonites.
Clupeidés : sardines, harengs, menhadens, ethmaloses, harengules, chardins, sardinelles
sardinops, sprats, shadines.
Engraulidés : anchois.
Coryphaenidés : coryphènes.
Toutefois, les poissons de ces familles qui ont subi un traitement de maturation enzymatique
dans la saumure peuvent avoir des teneurs en histamine plus élevées mais ne dépassant pas le
double des valeurs indiquées ci-dessus.
Les difficultés pour fixer une valeur limite toxique pour l’histamine sont dues à l’action de
potentialisateurs, imparfaitement élucidée. Les seuils retenus ne prennent pas en compte le
rôle synergique exercé par d’autres composés que l’histamine (présence d’autres amines
biogènes dans la denrée). La présence d’inhibiteurs d’enzymes de détoxification permet à
l’histamine d’exercer ses effets toxiques. L’alcool et certains médicaments, notamment les

Etude bibliographique
20
antidépresseurs, auraient un effet inhibiteur sur ces enzymes (notamment les
aminesoxydases).
b. Aspect réglementaire
Au niveau national: En Algérie, le seuil recommandé est de 10mg/100g (Journal
Officiel de la République Algérienne, 2006).
Au niveau international : les normes du Codex alimentarus ont une approche
différente et fixent deux seuils :
-Le premier est un seuil de qualité, indicateur d’altération du produit = 100 mg / kg.
-Le second est un critère de santé publique qui ne doit pas être dépassé =200 mg / kg.

MATERIEL
ET
METHODES

Matériels & Méthodes
21
3. MATERIEL ET METHODES
3.1. Echantillonnage et conditionnement
L’étude expérimentale porte sur la sardine (Sardina pilchardus), dont le poids moyen est de
32,48 ± 5,89 g et la longueur totale moyenne est de 17,67±0,76 cm. L’échantillonnage est
réalisé lors de la criée matinale au niveau du port de pêche d’Oran. Les échantillons sont
ensuite acheminés sous glace au laboratoire d’Aquaculture et Bioremédiation (AQUABIOR)
au département de biotechnologie de l’Université d’Oran.
Pour les différentes expérimentations réalisées lors de la présente étude, un effectif de dix
individus est retenu par mode. Les poissons sont répartis en deux lots, selon le mode de
conservation comme suit:
Lot A : poissons entiers (non éviscérés), conservés à température ambiante (25±5°C) /
réfrigérée (+4°C) : (8 h / 16 h).
Lot B : poissons entiers (non éviscérés), conservés au réfrigérateur (+4°C) : (24 h / 24 h).
Le traitement des échantillons (analyses sensorielles, biochimiques et microbiologiques)
s’effectue le premier jour sur des spécimens fraichement capturés toutes les trois heures : H0,
H3, H6, H9. Ensuite aux jours : j2(H24), j3(H48), j4(H72), j5(H96) sur les lots A et B et aux
jours J6(H120) et J7 (H144) sur le lot B uniquement.
3.2. Appréciation sensorielle
Cette analyse a été réalisée sur les poissons entiers non filetés. La méthode utilisée est le
système de cotation spécifique selon les directives codex alimentarius pour l’évaluation
organoleptique des poissons, des mollusques et des crustacés CAC/GL 31-1999, Volume 9A
- 2001. Ce système distingue trois catégories de fraîcheur : E, A et B, correspondant aux
niveaux divers d’altération. La catégorie E (Extra) correspond au niveau de qualité le plus
élevé, suivi de la qualité A et B, tandis qu'au-dessous de B, on considère le poisson impropre
à la consommation.
Le test sensoriel a été effectué quotidiennement par les membres scientifiques du laboratoire.
Chaque membre a donné un score pour les paramètres utilisés dans ce système d'évaluation,
afin de déterminer le temps de rejet organoleptique du poisson. De plus, nous avons essayé
d’établir une corrélation avec l’analyse biochimique et le profil microbiologique.

Matériels & Méthodes
22
3.3 .Analyses biochimiques
3.3.1. Analyses physico-chimiques
3.3.1.a. pH
La mesure du pH a été réalisée selon la procédure de Wang (2002). Dix grammes
d'échantillon ont été homogénéisés avec 10 ml d'eau déionisée à pH ≈ 7 dans les proportions
1 / 1(w/v). Le pH a été mesuré à l’aide d’un pH mètre.
3.3.1.b. Teneur en eau
3 g d’échantillon de muscle sont pesés (balance portée : 200 g, précision 0,01g) et placé à
l’étuve toute une nuit à une température variant entre 105 et 110°C pour séchage. Après
refroidissement totale, le muscle séché est pesé à nouveau. Ce test est réalisé sur dix individus
en même temps et dans les mêmes conditions.
La teneur en matière sèche (g / 100g) est donnée par la formule:
Teneur en eau (g/100g) = (M1-M2) x100/M1.
Matière sèche : (MS) (g/100g) =M2x100/M1.
Où : M1 : Poids initial de l’échantillon
M2 : Poids de l’échantillon après séchage.
MS : Poids de la matière sèche.
3.3.2. Etude de la peroxydation lipidique
3.3.2.a. Teneur en hydroperoxydes
La technique adoptée dans ce travail est proposée par Hermes-Lima et al., (1995) et améliorée
par Eymard et Genot, (2003). Elle consiste en l’extraction des hydroperoxydes par le
méthanol. Ces derniers oxydent le Fe2+
en Fe3+
, qui forme un complexe coloré avec le xylénol
orange possèdant un maximum d’absorption à 560 nm.
Les hydroperoxydes présents dans l’extrait sont quantifiés par spectrophotométrie faisant
référence à l’équation (y = 0,036 x) de la droite de la courbe étalon d’hydroperoxydes de
cumène (CuOOH), qui est réalisée dans le méthanol pour des concentrations variant de
0 à 20 μM. Les résultats sont exprimés en mmoles Eq CuOOH/kg MH.

Matériels & Méthodes
23
3.3.2.b. Teneur en TBA-rs
La technique utilisée pour le dosage des TBA-rs a été mise au point par Genot (1996) qui est
une adaptation des méthodes de Salih et al., (1987) et Bostoglou et al., (1994).
L’acide thiobarbiturique (TBA) réagit avec le malonaldéhyde (MDA) pour former un
complexe de couleur rose et/ou jaune possédant un maximum d’absorption à une longueur
d’onde de 532 nm. La lecture de l’absorbance des TBA-rs extraits des échantillons par l’acide
thiobarbiturique (TCA) est effectuée au spectrophotomètre aux longueurs d’onde : 508, 532 et
600 nm. Dans le but de minimiser le bruit de fond observé sur les spectres d’absorbance et
d’améliorer la sensibilité de la méthode, la mesure d’absorbance à λ max=532 nm est
corrigée selon la formule (Genot, 1996):
A532corrigée=A532-[((A508-A600)x(600-532))/(600-508)]-A600
La conversion de l’absorbance, mesurée à λ max =532 nm, en équivalents de malonaldéhyde
(mg/kg d’échantillon) est obtenue en utilisant le coefficient d’extinction molaire du complexe
MDA-TBA : 1,56.105 M
-1.cm
-1 (Buedge et al., 1978). Le mode de calcul est le suivant :
mg équivalent MDA/Kg = (Acorrigée x V TCA x 2 x M.10-2
) / (1,56 xm)
Avec V TCA : volume du solvant d’extraction (16 mL)
m : masse de l’échantillon analysée (g)
M : masse moléculaire du malonaldéhyde = 72 g.mol-1
3.3.3. Concentration en protéines totales
La teneur en protéines totales est déterminée selon la méthode de Bradford (1976), faisant
référence à l’équation (y = 0,013 x) de la droite de la courbe étalon réalisée à partir de
l’albumine sérique bovine (BSA).
3.3.4. Teneur en azote basique volatil total (ABVT)
Les analyses d’ABVT ont été effectuées selon la technique basée sur la distillation proposée
par Woyewoda et al., (1986) et modifiée par Uriarte-Montoya et Villalba (2010). 5 g de
muscle ont été mélangés avec 300 ml d'eau et homogénéisés pendant 2 minutes à l’aide d’un
Ultra-Turrax. On rajoute 2 ml d'huile comestible et 2 g d'oxyde de magnésium. Le mélange
est mis dans un ballon, placé dans le dispositif de distillation et mis à chauffer jusqu’à
l’ébullition. Le distillat est récolté pendant 25 minutes dans un bécher contenant 25 ml d'acide
borique à 2 % (w/v) additionné de quelques gouttes de rouge de méthyle et est titré
ultérieurement avec l’acide sulfurique à 0,05 N. Un témoin a été préparé de la même manière
sans échantillon.

Matériels & Méthodes
24
Les données ont été exprimées en mg d’ABVT/ 100 g d'échantillon, utilisant l'équation
suivante :
mg ABVT=[(V4-V5) x N2 x 100 x 14]/W2
Où: V4 : Quantité (ml) de H2SO4 utilisés pour l’échantillon.
V5 : Quantité (ml) de H2SO4 utilisés pour le témoin.
N2 : Normalité de H2SO4.
W2 : Poids de l’échantillon.
3.3.5. Teneur en histamine
La détermination de la teneur en histamine est réalisée selon la méthode colorimétrique
proposée par Patange et al. (2005). Cette méthode est basée sur la formation d’un complexe
coloré entre le noyau imidazole de l’histamine et le p-phenyldiazonium sulfonate (réactif).
L’absorbance de la couleur produite par le complexe est mesurée après 5min à λ=496 nm.
Une gamme étalon est réalisée avec de l’histamine dihydrochloride dans de l’eau distillée
pour des concentrations allant de 0 à 50 μg/ml.
La concentration de l’histamine dans l’échantillon est obtenue en faisant référence à
l’équation de la droite (y = 0,03x + 0,018) de la courbe étalon et estimée par la formule
suivante :
Histamine (mg/100g) =
= A mg/100g
Où A est la valeur d’histamine obtenue en μg/ml de la courbe étalon.
Cette méthode possède un seuil de détection de 1mg/100g (1μg/ml). Toutes les concentrations
en dessous de cette limite sont considérées comme des traces d’histamine.
3.4. Analyse microbiologique
3.4.1. Echantillonnage
Le contrôle de la qualité et de la salubrité du poisson s’effectue par sondage. La méthode
d’échantillonnage utilisée est celle préconisée par La Commission Internationale des Normes
Microbiologiques Relatives aux Denrées Alimentaires ou ICMSF (International Commission
on Microbiological Specifications for Foods), qui a défini des méthodes d’échantillonnages
pour l’analyse systématique des produits alimentaires.
Un échantillon de dix poissons est utilisé. 25 g de chair de chaque poisson sont prélevés
aseptiquement.

Matériels & Méthodes
25
3.4.2. Traitement de l’échantillon : préparation de la solution mère.
L’analyse microbiologique des différents échantillons s’effectue à partir d’une solution mère
dont la composition est la suivante : 25 g de chair de poisson prélevée est homogénéisée
(30000 tour/min) dans 225 ml de solution de TSE ( Tryptone, sel et eau distillée).
3.4.3. Numération de la flore aérobie mésophile totale.
Le milieu préconisé pour ce test est le Plate Count Agar le (PCA), proposé en 1973 par
l’AFNOR (association française de normalisation).
Méthodologie
Deux dilutions décimales successives sont utilisées. 1 ml de chaque dilution est réparti en
goutte au fond de la boîte correspondante (pour chaque dilution, deux boites de Pétri sont
utilisées). Les gouttes sont ensuite recouvertes d'une couche de gélose PCA en surfusion (45-
47°C), et le tout est homogénéisé avec des mouvements circulaires. Une fois la gélose
refroidie, elle est recouverte d’une seconde couche de gélose PCA, ce qui a pour effet
d'immobiliser les bactéries, et de former des colonies bien définies. Les boites de Pétri ainsi
ensemencées sont incubées à 30°C pendant 72h.
Les colonies sont dénombrables si leur nombre est compris entre 30 et 300. Au-dessus de 300,
elles sont indénombrables, et en dessous de 30 on considère qu'elles sont trop rares pour être
dénombrées. La formule mathématique suivante est utilisée :
N=∑ colonies/ VmL x (n1 + 0,1 n2) x d1
Où : N : nombre d’UFC par gramme de produit initial ;
∑ Colonies : Somme des colonies de boites interprétables ;
VmL : Volume de la solution déposé (1mL) ;
n1 : Nombre de boites considérées à la première dilution retenue ;
n2 : Nombre de boites considérées à la seconde dilution retenue ;
d1 : Facteur de la première dilution retenue.
3.4.4. Numération des coliformes totaux
Les coliformes totaux peuvent être indirectement associés à une pollution d’origine fécale. Ce
sont des bactéries en forme de bâtonnet, aérobies ou anaérobies facultatives, possédant
l’enzyme ß-galactosidase. Celle-ci permet l’hydrolyse du lactose à 35 °C afin de produire
l’acide lactique et du gaz sur milieu approprié (Archibald, 2000; CEAEQ, 2000; Edberg et al.,
2000).

Matériels & Méthodes
26
La méthode utilisée est celle de 3-3-3. Cette méthode préconise l’utilisation de neuf tubes
remplis (9 ml) avec le milieu bouillon lactosé pourpre de bromocrésol (BCPL) à différentes
concentrations. Trois tubes sont à double concentration (D/C) et six tubes à simple
concentration (S/C).
10 ml, 1 ml et 0,1 ml de la solution mère sont rajoutés, respectivement, aux trois tubes double
concentration, à trois tubes simple concentration et aux trois tubes simple concentration
restant. Les tubes sont ensuite mélangés, le gaz est chassé des cloches et les tubes sont ensuite
incuber à 37°C pendant 24 – 48 heures.
Sont considérés comme positifs les tubes présentant un dégagement de gaz (recueilli
dans les cloches) et un virage de la couleur du milieu au jaune (témoin de la
fermentation du lactose présent dans le milieu). L’utilisation de la table de Mac Grady
est nécessaire pour le dénombrement des troubles formant colonies ou TFC.
3.4.5. Numération des coliformes fécaux ou coliformes thermotolérants
Les coliformes fécaux, ou coliformes thermotolérants, sont un sous-groupe des coliformes
totaux capables de fermenter le lactose à une température de 44,5 °C.
A partir des tubes positifs du test précédent, prélever 1ml et l’ensemencer dans 9 ml de milieu
Schubert stérile et l’incuber à 44°C pendant 24 heures. Sont considérés positifs les tubes
présentant un anneau rouge en surface du tube, témoin de la production d’indole dans le
milieu après ajout de 2 à 3 gouttes du réactif d’Erlich de Kovacs.
3.4.6. Numération des Streptocoques fécaux ou thermotolérants
3.4.6.a. Etape 1 ou test présomptif
Ce test est effectué par ensemencement d’un milieu de Rothe S/C (AFNOR, 1960). Ce milieu
contient de l’azide de sodium (NaN3) qui inhibe la plupart des microorganismes. Il est peu
favorable à la croissance des Streptocoques fécaux et la plupart des autres bactéries ne s’y
cultivent pas. Le milieu de Rothe est cependant moins sélectif que le milieu de Litzky, ce qui
nous mène d’abord à l’utiliser dans la première étape, et les germes adaptés à l’effet
inhibiteur de l’azide de sodium s’adaptent à la présence d’éthyl violet.
1 ml de la suspension mère et des dilutions décimales de cette dernière sont
ensemencé dans 9 ml de milieu. Après 24 heures à 48 heures d’incubation à 37°C,
sont considérés positifs les tubes présentant un trouble bactérien. Ces tubes sont
soumis au test confirmatif.

Matériels & Méthodes
27
3.4.6.a. Etape 2 ou test confirmatif
L’addition d’Ethyl violet au milieu de Rothe le rend sélectif et spécifique des seuls
streptocoques fécaux.
Ce milieu est ensemencé à partir des tubes positifs du milieu de Rothe. Après 24 h à 48 h
d’incubation à 37°C, sont considérés positifs les tubes présentant un trouble bactérien avec
parfois formation d’un culot violet.
Le nombre de Streptocoques fécaux est exprimé par le NPP selon la table de Mac
Grady.
3.4.7. Numération des Clostridium sulfito-réducteurs
Parmi les Clostridium sulfito-réducteurs, C. perfringens occupe une place très importante. En
effet, ce germe est très souvent à l’origine de toxi-infections d’origine alimentaire.
Porter aseptiquement 1ml de chaque dilution de la solution mère en double dans deux tubes.
Ajouter 15ml de gélose viande-foie sulfitée (sulfite de sodium) et 4 à 5 gouttes d’Alun de fer.
Couvrir les tubes avec une couche de paraffine afin d’empêcher l’introduction d’oxygène.
Laisser solidifier pendant 30 minutes.
La première lecture doit se faire impérativement à 16 heures, car les colonies de Clostridium
sulfito-reducteur sont envahissantes et on pourrait se trouver en face d’un tube noir dont
l’interprétation devient impossible. Il faut repérer toute colonie noire d’un diamètre supérieur
à 0,5mm ainsi que compter les tubes positifs et calculer le nombre le plus probable (NPP).
3.4.8. Recherche de Staphylococcus aureus
Tous les staphylocoques présents dans les aliments ne sont pas entérotoxinogènes. Ce n’est
que depuis peu que l’on admet le rôle de S. aureus comme germe indicateur de contamination
et il figure dans un certain nombre de critères microbiologiques.
Ce groupe bactérien est recherché sur milieu gélosé Chapman par ensemencement en surface
de 0,1 ml de la solution mère. Les boîtes sont incubées 30 heures à 37°C. Les colonies de
Staphylococcus aureus sont rondes, régulières, bombées, opaques et pigmentées en jaune-
doré. Elles sont entourées d’un halo jaune correspondant à une acidification à partir de la
dégradation du mannitol (mannitol +). La seule exception est faite pour S. epidermidis qui
donne des colonies blanches (mannitol -).

Matériels & Méthodes
28
3.4.9. Recherche de Salmonella et de Shigella
Il s’agit d’un problème très important en microbiologie alimentaire. La transmission
s’effectue par des voies multiples et parfois complexes. Le nombre de germes pouvant
provoquer une maladie infectieuse est parfois très faible (de l’ordre de 1 à 50 par 100 g
produit) et dépend de l’espèce et du consommateur. La recherche de ces germes s’effectue par
des tests de présence ou d’absence et la norme est de 0 germe par g (AFNOR V 08 013).
Enrichissement sélectif :
Le milieu d’enrichissement utilisé est le bouillon au sélénite et à la cystine (milieu le mieux
adapté au contrôle des aliments). 10 ml de la solution mère sont ajoutés à 100 ml de bouillon
au sélénite. Les tubes sont incubés à 37°C pendant 24 h.
Isolement :
A partir d’une öse ou d’une goutte du milieu d’enrichissement on réalise un isolement sur
milieu SS (Salmonelle-Shigelle). Après 48 h d’incubation à 37°C, les colonies se présentent
sous les aspects suivants:
colonies rouges lactose + (Enterobacter, Klebsiella)
colonies incolores lactose- (Salmonella, Shigella, Serratia, Proteus morganii, E.
hafniae)
colonies à centre orangé (Proteus rettgeri, Providencia)
colonies à centre noir (Salmonella, Citrobacter, Proteus vulgaris, Proteus mirabilis).
3.4.10. Recherche de Pseudomonas
Ce sont des bactéries très répandues dans la nature contaminant les produits alimentaires
qu’elles peuvent dégrader de façon importante. Elles peuvent être l’agent de toxi-infections
alimentaires.
Les milieux gélosés King A et King B sont ensemencés à partir de la solution mère. Après
incubation à 35°C pendant 24 à 48 heures, apparaissent sur le milieu des colonies plates ou
légèrement surélevées, opaques à bords irréguliers ou finement dentelées de couleur
blanchâtre à légèrement beige. Après 2 - 3 jours on assiste à un bleuissement ou verdissement
du milieu de culture dû aux pigments fabriqués et diffusés par la bactérie (pyocyanine et
pyoverdine par Pseudomonas aerogenosa et Pseudomonas fluoresens, respectivement).

Matériels & Méthodes
29
3.5. Activité enzymatique d’altération d’origine bactérienne
3.5.1. Recherche de l’activité lipolytique
Les enzymes lipolytiques d’origine bactérienne permettent de dégrader les composants
lipidiques, dans le but de démontrer une éventuelle corrélation entre l’activité bactérienne et
l’activité autolytique propre au muscle. Le test suivant est réalisé : le milieu de culture utilisé
est celui préconisé par Sierra (1957). Ce dernier, gélosé au tween 80 (oléate de sorbitol), est
ensemencé par touche, et incubé à 37°C pendant 24 à 72 heures. L’activité lipasique se
manifeste par l’apparition d’un halo opaque autour des colonies, halo dû à la précipitation
d’oléate de calcium.
3.5. 2. Recherche de l’activité gélatinolytique
Cette activité est recherchée par la technique de Frazier (1950) et modifiée par Feuer (1996).
Dans le même contexte que celui de l’activité lipolityque, ce test nous permet de déterminer
s’il existe une relation entre cette activité et la concentration brute en protéines ainsi que la
structure de la chair de poisson. Une gélose nutritive ordinaire est additionnée de 4% de
gélatine et répartie en boite de Pétri qui sont ensuite ensemencées en touche. Après incubation
à 37°C, l’hydrolyse de la gélatine est mise en évidence par vaporisation de quelques ml de
réactif de Frazier. Un précipité opaque se produit sauf aux endroits où la gélatine est
hydrolysée.
3.6. Bactéries productrices des amines biogènes
3.6.1. Isolement des bactéries productrices des amines biogènes
Le milieu de culture utilisé est celui préconisé par Joosten et Northold (1989) et modifié par
Bover-Cid en 1999. Ce milieu nous permet de mettre en évidence l’activité des amino-acides
décarboxylases d’origine bactérienne, responsables de la production des amines biogènes, en
fonction de l’acide aminé présent dans le milieu.
1 ml de la solution mère est ensemencé sur le milieu de culture gélosé. L’incubation à 35°C
dure pendant 4 jours minimum. Ce traitement est toujours réalisé en double et on prévoit,
aussi, un témoin exempt d’acide aminé.
Sont considérées positives les colonies dont la couleur vire au violet ainsi que la zone qui les
entoure. Ceci est synonyme de production d’amines biogènes (l’indicateur de pH coloré est
jaune à pH acide et violet à pH alcalin).

Matériels & Méthodes
30
3.6.2. Purification et essai d’identification
Les colonies positives feront l’objet d’une série de repiquage en vue de leur purification. Les
souches pures feront à leurs tours l’objet d’une série d’examens :
Macroscopique : L’aspect des colonies (couleur, contour, forme…).
Microscopique : La forme, le mode d’association et le Gram
Test de la catalase : Action directe de l’enzyme mise en évidence dans la masse
bactérienne. Une goutte d’eau oxygénée à 10 volumes est placée sur une lame et une
partie d’une colonie y est émulsionnée. L’activité positive de l’enzyme se traduit par
un dégagement gazeux résultant de la dégradation du peroxyde d’hydrogène.
3.7. Etude statistique
Les différents dosages sont effectués sur 10 spécimens (n=10). Dans tous les cas, les
statistiques descriptives (moyennes ± écarts types) sont utilisées pour décrire l’ensemble des
résultats.
Avant toute analyse statistique, l’homogénéité des variances a été vérifiée. Lorsque les
variances sont homogènes, l’analyse consiste en un test paramétrique d’Anova 1 (p< 0,05).
Lorsqu’elles ne sont pas homogènes, l’analyse consiste en un test non paramétrique de
Kruskal - Wallis.
Pour déterminer si les différences, entre les moyennes obtenues, sont significatives, nous
utilisons le test de Tukey (H.S.D) dans le cas de l’Anova et celui de Mann -Withney dans le
cas de Kruskal - Wallis.

RESULTATS

Résultats
31
4. RESULTATS
4.1. Analyse sensorielle
Arrivée au laboratoire, la sardine fraîche est caractérisée par un ventre argenté et un
dos bleuté, un opercule strié, des taches sombres sur le dos, une carène ventrale peu aigüe et
des écailles sessiles.
A J1 et durant les premières heures de conservation (H0, H3, H6, H9), la sardine est
caractérisée par une fine odeur d’algues iodées. Le corps présente un aspect luisant et des
écailles bien ancrées, avec présence d’un mucus transparent de consistance visqueuse. L’œil
est convexe et brillant avec une cornée translucide. Les branchies sont de couleur rouge vif
uniforme. L’abdomen du poisson reste intact. Après ouverture du poisson, nous remarquons
que les organes internes sont indemnes et restent bien différenciés. La chair est ferme et
élastique avec une nette différenciation entre le muscle rouge et le muscle blanc (Planche 1).
A température ambiante, Après 24 heures (J2) de conservation, nous observons les premiers
signes de la perte de qualité. En effet, la peau perd de sa brillance, de sa pigmentation et des
plaques de sang apparaissent rapidement sous l’opercule et l’œil. A J3, l’odeur du poisson
devient aigre et rance, l’œil devient opaque, laiteux et concave. Les viscères éclatent et leur
contenu liquide se dissémine dans la chair lui procurant un aspect mou et pâteux, accélèrant,
ainsi, son altération. Le poisson perd de sa fermeté et se détache difficilement de la colonne
vertébrale. A J5, la peau devient totalement terne, dépourvue d’écailles et totalement
recouverte d’un liquide visqueux jaunâtre et l’odeur nauséabonde devient très présente et
insupportable (planche1).
Au réfrigérateur, la qualité sensorielle de la sardine (S. pilchardus) présente un décalage
d’environ 48 heures par rapport à celui observé chez la sardine conservée à température
ambiante. À J2, la sardine préserve des qualités sensorielles acceptables et satisfaisantes.
L’odeur du poisson devient forte à J3, répulsive et repoussante à J5 et J7. La peau, très terne,
est pratiquement dépourvue d’écailles et est recouverte d’un mucus visqueux de couleur
foncée à J5. A J7 la décoloration est très avancée. La peau est molle et pâteuse. A
l’éviscération du poisson, les organes internes sont indifférenciés. Le poisson atteint son
maximum d’altération et est au stade de putréfaction (Planche 1).

Résultats
32
A= Température ambiante R= Température réfrigérée
Planche 1 : Aspect extérieur de la sardine pendant la conservation
J3A
J1
J2A J2R
J3R J3A
J7R J5A
AQUABIOR, 2011
AQUABIOR, 2011 AQUABIOR, 2011
AQUABIOR, 2011 AQUABIOR, 2011
AQUABIOR, 2011 AQUABIOR, 2011

Résultats
33
4.2. Caractéristiques biochimiques
4.2.1. Variation du pH
4.2.1.1. Effet de la durée de conservation
Quel que soit le mode de conservation, l’étude statistique montre une augmentation
significative (p<0,05) du pH de la chair de sardine pendant toute la durée de conservation. Le
pH passe de 5,91 ± 0,19 à7,11 ± 0,29 à J5 et 7,21±0,07 à J7, respectivement à température
ambiante et réfrigérée (figure 7).
Figure 7 : Evolution temporelle du pH chez la sardine (S. pilchardus). Les histogrammes
portant des lettres et des chiffres différents sont significativement différents (p < 0,05).
4.1.1.2. Effet du mode de conservation
De manière générale, le pH mesuré dans le muscle de sardine conservée à température
ambiante (Figure 8) est significativement (p<0,05) plus élevé que celui mesuré chez la sardine
conservée à température réfrigérée.
0
1
2
3
4
5
6
7
8
H0 H3 H6 H9 H24 H48 H72 H96 H120 H144
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
pH
pH
Durée (heures)
a 1 ab
1 ab 1.2 bc 2.3
cd 3.4 d 4.5 e 5 f 6
6 6

Résultats
34
Figure 8 : Effet du mode de conservation sur le pH chez la sardine (S. pilchardus). Les
histogrammes portant des lettres et des chiffres différents sont significativement différents (p
< 0,05).
4.2.2. Détermination de la teneur en eau
4.2.2.a. Effet de la durée de conservation
Quel que soit le mode de conservation, la teneur en eau dans le muscle de la sardine diminue
(<0,05) progressivement et continuellement, atteignant un minimum après 24 et 96 heure de
conservation, respectivement, à température ambiante et réfrigérée.
0
1
2
3
4
5
6
7
8
H24 H48 H96
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
Durée (heures)
pH
a b A B
1 2

Résultats
35
Figure 9 : Evolution temporelle de la teneur en eau chez la sardine (S. pilchardus). Les
histogrammes portant des lettres et des chiffres différents sont significativement différents
(p < 0,05).
4.2.2.b. Effet du mode de conservation
L’analyse statistique montre un effet significatif (p < 0,05) du mode de conservation sur la
teneur en eau chez la sardine en faveur de la réfrigération. En effet, nous remarquons que
cette teneur est plus élevée chez la sardine conservée au réfrigérateur que chez celle conservée
à température ambiante (figure 10).
0
10
20
30
40
50
60
70
80
90
100
H0 H3 H6 H9 H24 H48 H96 H144
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
Durée (heures)
Eau
( % M.F.)
a 1 a 1 1
2 2
2 3 3
d c
c b
a

Résultats
36
Figure 10 : Effet du mode de conservation sur la teneur en eau chez la sardine (S.
pilchardus). Les histogrammes portant des lettres et différents sont significativement
différents (p < 0,05).
4.2.3. Evolution du niveau d’oxydation des lipides
4.2.3.a. Teneur en hydroperoxydes
* Effet de la durée de conservation
Quel que soit le mode de conservation, l’analyse statistique Anova 1 montre un effet
significatif (p < 0,05) de la conservation sur la teneur en hydroperoxydes du muscle de la
sardine.
En effet, à température ambiante, la teneur en hydroperoxydes augmente progressivement
et significativement (p < 0,05) dès les premières heures de conservation et continue son
évolution, passant de 4,29 ± 0,52 mmoles/g M.H. à H0 à une valeur maximale de 26,56 ±
0,99 mmoles Eq CuOOH/g M.H. à H48 (figure 11). Entre H48 etH96 nous mesurons une
diminution significative (<0,05) de la teneur de ce produit.
En mode réfrigéré (figure 11), la teneur en hydroperoxydes augmente de manière
significative (p < 0,05) et atteint son maximum à J3 (32,18 ± 4,37 mmoles Eq CuOOH/g
M.H.). Ensuite, la teneur de ce produit diminue (<0,05) atteignant la valeur 21,58 ± 2,53
mmoles Eq CuOOH/g M.H. à J7.
0
10
20
30
40
50
60
70
80
90
H24 H48 H96
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
Durée (heures)
Eau
(% M.F.)
Eau
(% M.F.)
b B 2
a A
1
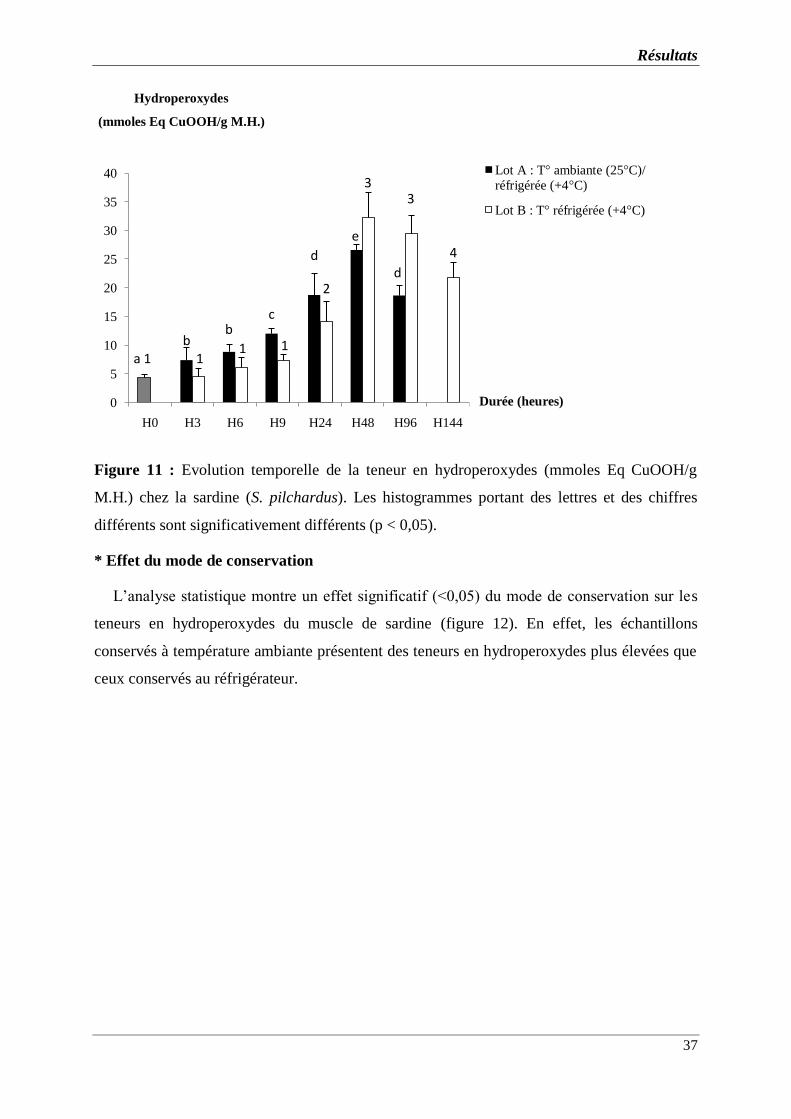
Résultats
37
Figure 11 : Evolution temporelle de la teneur en hydroperoxydes (mmoles Eq CuOOH/g
M.H.) chez la sardine (S. pilchardus). Les histogrammes portant des lettres et des chiffres
différents sont significativement différents (p < 0,05).
* Effet du mode de conservation
L’analyse statistique montre un effet significatif (<0,05) du mode de conservation sur les
teneurs en hydroperoxydes du muscle de sardine (figure 12). En effet, les échantillons
conservés à température ambiante présentent des teneurs en hydroperoxydes plus élevées que
ceux conservés au réfrigérateur.
0
5
10
15
20
25
30
35
40
H0 H3 H6 H9 H24 H48 H96 H144
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
Durée (heures)
Hydroperoxydes
(mmoles Eq CuOOH/g M.H.)
a 1
3
4
b b
c
d
3
d
e
2
1 1 1

Résultats
38
Figure 12 : effet du mode conservation sur la teneur en hydroperoxydes (mmoles Eq
CuOOH/g M.H. chez la sardine (S. pilchadus). Les histogrammes portant des lettres et des
chiffres différents sont significativement différents (p < 0,05).
4.2.3.b. Teneur en TBA-rs
* Effet de la durée de conservation
Chez la sardine fraîche, et jusqu’à 6 heures de conservation la teneur en TBA-rs est
comparable et extrêmement faible (≈0,3 mg Eq MDA/ g de M.F.).
A température ambiante (figure 13) et après 9 heures de conservation, nous mesurons une
augmentation significative (<0,05) et continue dans la teneur en TBA-rs, passant de 0,29 ±
0,08 mg Eq MDA/ g de M.F. à H0 à 1,8 ± 0,07 mg Eq MDA/ g de M.F.
De même chez la sardine réfrigérée (figure 13), les TBA-rs augmentent significativement
(p < 0,05) et progressivement à partir de H24 et atteignent le maximum après 7 jours de
conservation (2,91 ± 0,37 mg Eq MDA/ g de M.F.).
0
5
10
15
20
25
30
35
40
H24 H48 H96
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
Durée (heures)
Hydroperoxydes
(mmoles Eq CuOOH/g M.H.)
B
a
2
b
A
1

Résultats
39
Figure 13 : Evolution temporelle de la teneur en TBA rs (mg Eq MDA/ g de M.F.) chez la
sardine (S. pilchardus). Les histogrammes portant des lettres et des chiffres différents sont
significativement différents (p < 0,05).
* Effet du mode de conservation
L’analyse statistique montre une différence significative (p< 0,05) dans la concentration en
TBA-rs entre la sardine conservée à température ambiante et celle conservée à température
réfrigérée (figure 14).
0
0,5
1
1,5
2
2,5
3
3,5
H0 H3 H6 H9 H24 H48 H96 H144
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
Durée (heures)
TBA rs
(mg Eq MDA/ g de M.F.)
a 1 a 1 a 1
b 1
2 2 2
3
c
d e

Résultats
40
Figure 14 : effet du mode de conservation sur la teneur en TBA rs (mg Eq MDA/ g de M.F.)
chez la sardine (S. pilchadus). Les histogrammes portant des lettres et chiffres différents sont
significativement différents (p < 0,05).
4.2.4. Estimation de la concentration des protéines totales
4.2.4.a. Effet de la durée de conservation
La sardine fraiche présente une teneur initiale en protéines de 28,91 ± 0,9 g de protéines par
100 g de chair de poisson (figure 15).
Quel que soit le mode de conservation, l’analyse statistique anova 1 montre une diminution
significative (p <0,05) progressive et continue de la concentration protéique jusqu’à la fin de
l’expérimentation, atteignant les valeurs (24,12± 0,23 g / 100 g) et (23,71± 0,35 g / 100 g) à
J5 et J7, respectivement, à température ambiante et réfrigérée (figure 15).
0
0,5
1
1,5
2
2,5
3
H24 H48 H96
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
Durée (heures)
TBA rs
(mg Eq MDA/ g de M.F.)
b 2
a
1

Résultats
41
Figure 15 : Evolution temporelle de la teneur en protéines (g de protéines/ 100 g de chair)
chez la sardine (S. pilchardus). Les histogrammes portant des lettres et des chiffres différents
sont significativement différents (p < 0,05).
4.2.4.b. Effet du mode de conservation
L’analyse statistique montre un effet significatif (p < 0,05) du mode de conservation sur la
concentration protéique chez la sardine en faveur de la réfrigération (figure 16).
0
5
10
15
20
25
30
35
H0 H3 H6 H9 H24 H48 H72 H96 H120 H144
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
lot B (à + 4° C)
Durée (heures)
g protéines / 100 g de chair
a 1 b 1.2 c 2.3
cd 3.4 cd 4 de 4 f 4 g 5 6 6

Résultats
42
Figure 16: Effet du mode de conservation sur la teneur en protéines (g de protéines/ 100 g de
chair) chez la sardine (S. pilchardus). Les histogrammes portant des lettres et des chiffres
différents sont significativement différents (p < 0,05).
4.2.5. Concentration de l’azote basique volatil total (ABVT)
4.2.5.a. Effet de la durée de conservation
A l’état initial la sardine fraiche présente une valeur en azote basique volatil total de 3,11 ±
0,93 mg / 100 g de chair de poisson.
A température ambiante (figure 17), nous enregistrons une augmentation significative (p <
0,05) et continuelle de la teneur en azote basique volatil total jusqu’au 4ème
jour où elle atteint
sa valeur maximale (40,74 ± 2,50 mg / 100 g de chair de poisson), ensuite se stabilise.
Au réfrigérateur, l’évolution de la concentration en azote basique volatil total suit le même
schéma que pour la température ambiante où nous observons une augmentation continue
jusqu’à J6 (46,48 ± 3,94 mg / 100 g de chair de poisson).
22
23
24
25
26
27
28
H24 H48 H96
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
g protéines / 100 g de chair
Durée (heures)
2
B A
1

Résultats
43
Figure 17 : Evolution temporelle de la teneur en ABVT (mg d’ABVT / 100 g de chair) chez
la sardine (S. pilchardus). Les histogrammes portant des lettres et des chiffres différents sont
significativement différents (p < 0,05).
4.2.5.b. Effet du mode de conservation
Lors de l’analyse statistique Anova 1, nous notons une différence significative (p < 0,05)
nette entre les échantillons conservés à température ambiante et à température réfrigérée. En
effet, nous remarquons que cette teneur est plus élevée chez la sardine conservée à
température ambiante que chez celle conservée au réfrigérateur (figure 18).
0
10
20
30
40
50
60
H0 H3 H6 H9 H24 H48 H72 H96 H120 H144
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
Durée (heures)
Mg d’ABVT / 100
g de chair
a 1
b
c c
d
e
f
f 5
5
1 1
4
3
2 2 2

Résultats
44
Figure 18 : Effet du mode de conservation sur la teneur en ABVT (mg d’ABVT / 100 g de
chair) chez la sardine (S. pilchardus). Les histogrammes portant des lettres et chiffres
différents sont significativement différents (p < 0,05).
4.2.6. Teneur en histamine
4.2.6.a. Effet du temps de conservation
Chez la sardine fraiche, la teneur moyenne en histamine est de 3,44 ± 0,35 mg/100g reste
plus ou moins stable aux premières heures de conservation. Quel que soit le mode de
conservation, la teneur en histamine n’augmente significativement (p < 0,05) qu’à partir de
neufs heures de conservation, et persiste jusqu’en fin d’expérimentation. Le seuil
réglementaire de 10mg/100g de muscle est rapidement dépassé au bout de 24heures, à
température ambiante. Alors qu’au réfrigérateur, cette teneur limite n’est dépassée qu’après
72 heures de conservation (figure19).
0
5
10
15
20
25
30
35
40
45
50
H24 H48 H96
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
Durée (heures)
Mg d’ABVT / 100
g de chair
1
A
a
b
2
B

Résultats
45
Figure 19 : Evolution temporelle de la teneur en histamine (mg/100gM.H.) chez la sardine (S.
pilchardus). Les histogrammes portant des lettres et des chiffres différents sont
significativement différents (p < 0,05).
4.2.6.b. Effet du mode de conservation
En comparant les modes de conservation du lot A et B (figure 20), l’analyse statistique
montre un effet significatif du mode de conservation sur la teneur en histamine chez la sardine
(figure 20) en faveur des poissons conservés au réfrigérateur.
0
5
10
15
20
25
30
H0 H3 H6 H9 H24 H48 H72 H96 H120 H144
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
Durée (heures)
Histamine
(mg/100gM.H.)
a 1 a 1 a 1
b
2
c
3
d
4
e
5
f
6 7
8

Résultats
46
Figure 20 : Effet du mode de conservation sur la teneur en histamine (mg/100gM.H.) chez la
sardine (S. pilchardus). Les histogrammes portant des lettres et chiffres différents sont
significativement différents (p < 0,05).
4.3. Analyse microbiologique
4.3.1. Numération de la flore aérobie mésophile totale
La numération des germes totaux chez la sardine fraiche donne une valeur initiale d’unités
formant une colonie de 1,4.104 UFC/ml (figure 21).
A température ambiante, nous notons une croissance faible aux premières heures de
conservation et une croissance accélérée à partir de J 3, pour atteindre une valeur maximale en
fin d’expérimentation (3,1.1010
UFC / ml) à J 5.
Au réfrigérateur, le schéma évolutif de la croissance emprunte la même voie que celle à
température ambiante. Cependant, nous observons un décalage de 24 heures et l’accélération
de la croissance à J4. La valeur maximale est atteinte au bout du 7ème
jour de conservation
(3,8.1010
UFC / ml).
0
5
10
15
20
25
30
H24 H48 H96
Lot A : T° ambiante (25°C)/ réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
Histamine
(mg/100gM.H.)
Durée (heures)
a
b
1
2
A
B

Résultats
47
Figure 21 : Evolution temporelle de la flore aérobie mésophile totale chez la sardine
(S. pilchardus).
Figure 22 : Aspect des colonies de bactéries isolées sur milieu Plate Count Agar (P.C.A) pour
dénombrement.
0
2
4
6
8
10
12
H0 H3 H6 H9 H24 H48 H72 H96 H120 H144
Lot A : T° ambiante
(25°C)/ réfrigérée (+4°C)
Lot B : T° réfrigérée
(+4°C)
Durée (heures)
Log UFC/ g
AQUABIOR,2011

Résultats
48
4.3.2. Numération des coliformes totaux
La présence des coliformes totaux est observée dès les premières heures de conservation à H0.
On estime que ces germes sont présents avec une valeur inferieure à 300 UFT / ml. Les
échantillons conservés à température ambiante (figure 23) montrent une croissance importante
qui s’accélère dès les premières heures de conservation jusqu’à la fin de l’expérience pour
atteindre une valeur maximale de 1,1.107 UFT / ml.
En mode réfrigéré (figure 23), la croissance démarre lentement puis augmente
progressivement jusqu’à J3 où nous assistons à une accélération rapide de la croissance pour
atteindre une valeur maximale de 1,5.107
UFT / ml au bout du 5ème
jour. Au-delà de J5, la
croissance se stabilise jusqu’à la fin de l’expérimentation (J7) où elle atteint 5.107 UFT / ml.
Figure 23 : Evolution temporelle de la teneur des coliformes totaux chez la sardine (S.
pilchardus).
0
1
2
3
4
5
6
7
8
9
H0 H3 H6 H9 H24 H48 H72 H96 H120 H144
Lot A : T° ambiante
(25°C)/ réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
Durée (heures)
Log UFT/ g

Résultats
49
Figure 24 : Aspect du bouillon au pourpre de bromocrésol (BCPL) avant et après incubation
(virage du milieu au jaune et production de gaz récolté dans la cloche de Durham)
4.3.3. Numération des coliformes fécaux ou coliformes thermotolérants
Nous remarquons que les coliformes fécaux sont présents dès le début d’expérience (H0) avec
un taux faible inférieur à 300 UFT / ml. A température ambiante (figure 25), cette valeur reste
stable jusqu’à H3. Ensuite, nous assistons à une accélération continue et régulière jusqu’à J5
pour atteindre la valeur maximale de 1,1.107 UFT /ml.
Au réfrigérateur (figure 25), la valeur initiale reste stable jusqu’à H6 puis subit une forte
accélération pour se stabiliser à nouveau aprés 9 heures de conservation. La croissance
augmente à nouveau progressivement jusqu’à J3 puis se stabilise jusqu’à J5 où nous notons
une forte accélération qui persiste jusqu’à la fin de l’expérience pour atteindre la valeur
maximale de 2,5.107 UFT / ml (figure 25).
AQUABIOR,2011

Résultats
50
Figure 25 : Evolution temporelle de la teneur des coliformes fécaux chez la sardine (S.
pilchardus).
4.3.4. Numération des Streptocoques fécaux ou thermotolérants
Sur les échantillons de sardine fraiche, les tests révèlent la présence de streptocoques
thermotolérants mais à faibles concentrations ne dépassant pas 300 UFT/ml (figure 26).
En mode ambiant, la croissance est faible jusqu’à H9. Ensuite, elle subit une forte
accélération jusqu’ à la fin des expériences où nous enregistrons une valeur maximale de
l’ordre de 1,5.107 UFT/ml.
Conservés au réfrigérateur, les échantillons de poissons subissent plus ou moins le même
schéma évolutif de croissance qu’en mode ambiant avec un décalage de 24 heures. Le
dénombrement révèle une valeur maximale de 1,3.107
UFT/ml au bout du 7ème
jour de
conservation à 4°C.
0
1
2
3
4
5
6
7
8
H0 H3 H6 H9 H24 H48 H72 H96 H120 H144
Lot A : T° ambiante
(25°C)/ réfrigérée (+4°C)
Lot B : T° réfrigérée
(+4°C)
Durée (heures)
Log UFT/ g

Résultats
51
Figure 26: Evolution temporelle de la teneur des streptocoques thermotolérants chez la
sardine (S. pilchardus).
4.3.5. Numération des Clostridium sulfito-réducteurs
Sur les échantillons frais il y a absence des clostridium sulfito-réducteurs aux premières
heures jusqu’à H3. Concernant les échantillons réfrigérés la présence de Clostridium ne se
manifeste qu’après 6 heures de conservation.
Le tableau 5 récapitule les résultats obtenus sur la numération des bactéries Clostridium
sulfito-réductrices dans les échantillons de sardine conservés à température ambiante et
réfrigérée.
0
1
2
3
4
5
6
7
8
H0 H3 H6 H9 H24 H48 H72 H96 H120 H144
Lot A : T° ambiante
(25°C)/ réfrigérée (+4°C)
Lot B : T° réfrigérée
(+4°C)
Durée (heures)
Log UFT/ g

Résultats
52
Tableau 5 : Evolution temporelle de la concentration cellulaire des clostridium sulfito-
réducteurs chez la sardine (S. pilchardus) conservée à température ambiante / réfrigérée et
réfrigérée.
TEMPS
Température ambiante Température réfrigéré
Description
des colonies
Diamètre Nombre des
colonies
Description
des colonies
Diamètre Nombre des
colonies
H°
Absence
H3
Colonies
noires.
Colonies
blanches.
< 1mm
1-3mm
Présence. Absence
H6
Colonies
noires.
< 1mm
Peu
nombreuses.
Colonies
noires.
Colonies
blanchâtre à
grisâtre.
1mm
2-3mm
Nombreuses.
H9
Colonies
grises à
noires.
2mm
Peu
nombreuses.
Colonies
noires.
< 1mm
Nombreuses.
H24
Colonies
noires.
<1-3mm
Peu
nombreuses
*colonies
noires
*colonies
blanches
2 mm
< 1mm
Nombreuses.
H48
Colonies
noires.
<1mm
Nombreuses. Colonies
grisâtres.
1-2mm
Peu
nombreuses.
H72
Colonies
noires
associées en
paires.
<2-3mm
Nombreuses. Colonies
noires.
< 1mm Peu
Nombreuses.
H96
Colonies
noire
présentes au
fond du tube.
2-3mm
Nombreuses. Colonies
grisâtre à
noires.
< 1mm Peu
nombreuses.
H120
Colonies
noires.
2-3mm
Nombreuse.
H144
Colonies
noires. Envahissant
le tube.

Résultats
53
4.3.6. Présence / numération des Staphylocoques
Le résultat d’isolement de colonies sur milieu Chapman a montré la présence de divers types
de colonies correspondant à différents groupes bactériens et ceci depuis les premières heures
de conservation et ce quel que soit le mode de conservation.
Les tableaux 6 et 7 récapitulent les résultats obtenus sur la recherche des bactéries
Staphylocoques dans les échantillons de sardine conservés à température ambiante et
réfrigérée.

Résultats
54
Tableau 6 : Evolution temporelle des Staphylocoques sur milieu Chapman chez la sardine (S.
pilchardus) conservée à température ambiante.
TEMPS Température ambiante
Description des colonies Diamètre Nombre
H°
Colonies de couleur saumon: bombées à contours
réguliers, brillantes.
Colonies blanches légèrement surélevées et à contours
légèrement irréguliers.
Petite colonies de couleur jaune, à contours réguliers,
bombées et brillantes.
Colonies de couleur rosâtre, plates, brillantes et de
contour régulier.
1-8 mm
<1-3 mm
<1-1 mm
<1-2 mm
15
11
5
34
H3
Colonies de couleur jaune, de contours réguliers,
bombées et brillantes.
Colonies de couleur légèrement rosâtre centre surélevé,
brillantes, rondes.
Colonies de couleur orangée, brillantes, à contours
irréguliers, à centre foncé.
1 mm
<1-6 mm
<1-6 mm
11
61
3
H6
Colonies jaunes bambées, à contours irréguliers et
brillantes.
Colonies de couleur blanchâtre à contours réguliers,
plates, aspect crémeux.
1-3mm
1-4mm
9
26
H9
Colonies de couleur saumon: bombées à contours
réguliers, brillantes.
Colonies de couleur jaune, de contours réguliers,
bombées et brillantes.
Colonies de couleur blanchâtre à contours réguliers,
plates, brillantes.
3mm
<1-2 mm
1 mm
15
5
38
H24
Colonies blanchâtres, bombées, brillantes, à contours
réguliers.
Colonies jaunâtres, bombées, très brillantes à contours
irréguliers, légèrement transparentes.
Colonies orange bombées, à centre surélevé, brillantes.
1-5 mm
2-7 mm
1-2 mm
48
11
27
H48
Un tapis de colonies jaunes avec virages de la couleur
du milieu au jaune.
H72
Colonies jaunes, brillantes, rondes à contours réguliers,
à centre surélevé.
2-3mm
37
H96
Colonies beiges, rondes, brillantes, à aspect crémeux. <1-2 mm
16

Résultats
55
Tableau 7 : Evolution temporelle des Staphylocoques sur milieu Chapman chez la sardine
(S. pilchardus) à réfrigérée.
TEMPS Température réfrigérée
Description des colonies Diamètre Nombre
H3
Colonies de couleur blanchâtres à contours irréguliers
bombées, brillantes, au centre surélevé.
Colonies de couleur beige à contours irréguliers
bombées, brillantes, légèrement surélevées.
<1-7 mm
1-5 mm
45
19
H6
Colonies jaunes à contours légèrement irréguliers,
bombées, brillantes.
Colonies blanches à contours légèrement dentelés,
bombées, ternes.
2 mm
<1-5 mm
8
47
H9
Colonies de couleur rosâtre, plate, brillantes et de
contours réguliers.
Colonies blanches, bombées, contours réguliers, au
centre surélevé.
2-3 mm
<2 mm
24
27
H24
Colonies jaunes, brillantes, à contours réguliers, à centre
surélevé.
2 mm
59
H48
Colonies jaunes, brillantes, à contours réguliers, à centre
surélevé.
1 – 2 mm 28
H72
Colonies jaunes, brillantes, à contours réguliers, à centre
surélevé.
< 1 – 3
mm
45
H96
Colonies blanches, rondes, aspect crémeux, peu
brillantes.
2-3 mm 18
H120
Colonie orange, brillante, transparente.
Colonies jaunes, brillantes, à contours réguliers.
1mm
1-3mm
1
24
H144
Colonies beiges, rondes, aspect crémeux, peu brillantes.
Colonies jaunes, brillantes, à contours réguliers, à centre
surélevé.
1-2 mm
2 mm
41
17

Résultats
56
Figure 27 : Aspect des colonies de bactéries isolées sur milieu Chapman.
4.3.7. Présence / numération Salmonelles et Shigelles
Quel que soit le mode de conservation nous remarquons la présence de colonies de différents
aspects sur milieu S-S dès les premières heures d’échantillonnage,
Le tableau 8 récapitule les résultats obtenus sur la recherche des bactéries isolées sur milieu
S-S dans les échantillons de sardine conservés à température ambiante et réfrigérée.
AQUABIOR,2011 AQUABIOR,2011

Résultats
57
Tableau 8 : Evolution temporelle des isolats sur milieu S-S chez la sardine (S. pilchadus) à
température ambiante et réfrigérée.
TEMPS
Température ambiante
Température réfrigérée
Description des
colonies
Diamètre Nbre
Description des
colonies
Diamètre Nbre
H°
Colonies grises
1-2mm 24
H3
Colonies saumon à
centre noir
<1-2mm 8 Colonies rouge brique 1-3mm 16
H6
Colonies rosâtres à
centre noir
1-3mm 31 Colonies saumon à
centre noire
1-2mm 54
H9
Colonies saumon à
centre gris
<1-3mm 19 Colonies rosâtres à
saumon
2mm 30
H24
Colonies saumon à
rouge brique
1-2mm 45 Colonies rouge brique
à centre noir diffus
1-3mm 44
H48
Colonies roses
1-4mm 27 Colonies rouge brique
à centre noir fin
1-2mm 26
H72
Colonies saumon
Colonies grises à
bleuâtres
1-2mm
2-3mm
45
18
Colonies saumon à
centre noir
2-3mm 44
H96
Colonies saumon à
rosâtres
1-2mm 36 Colonies beiges à
rosâtres
<1-2mm 12
H120
Colonies saumon à
centre noir
2-3mm 62
H144
Colonies saumon à
rosâtre
1-2mm 48

Résultats
58
.3.8. Présence / numération de Pseudomonas
Quel que soit le mode de conservation nous remarquons les aspects suivants sur les milieux
King A et King B:
Tableau 9 : Evolution temporelle des Pseudomonas chez la sardine (S. pilchardus) à
température ambiante.
TEMPS Température ambiante
Description des colonies Diamètre Nombre
H0
King B:
Colonies beiges, plates rondes
King A:
Petites colonies jaunes, rondes, bombées et brillantes
Colonie orange, transparante, bombée
Colonies blanches, de contours légèrement
irréguliers, plates, non brillantes
Colonies beiges, de contours irréguliers, légèrement
surélevées brillantes
1mm
2mm
2mm
2-3mm
tapis
3
1
21
34
H3
King B:
Colonies beiges légèrement transparentes
Colonie orange, ronde, brillante, plate
King A:
Colonies jaunes de contours irréguliers plates, ternes
Colonies beiges, transparentes, brillantes
5mm
5mm
Tapis
Tapis
H6
King B:
Colonies blanches, de contours légèrement
irréguliers, bombées, crémeuses
King A:
Colonies blanchâtres à beiges plus ou moins
transparentes à contours légèrement irréguliers,
plates, peu brillantes
2-3mm
<1-3mm
nombreuses
nombreuses
H9
King B:
Colonies jaunes, contours régulier, bombées, non
brillantes, crémeuses
Colonies blanches, de contours irréguliers, brillantes
King A:
Colonies beiges, de contours régulier, légèrement
surélevées brillantes
2-3mm
<1-3mm
<1-4mm
24
nombreuses
56
H24
King B:
Colonies blanches, à contours régulier, plates, ternes
Colonies jaunes, à contours irréguliers, bombées, peu brillantes
King A:
Colonies beiges, transparentes, brillantes, plates à
centre surélevé, contours
<1-5mm
1-3mm
1-3mm
Nombreuses
17
67

Résultats
59
H48
King B et king A:
Un tapis bactérien de colonies beiges à aspect
crémeux.
H72
King B:
Colonies blanches, de contours légèrement dentelée,
bombées, crémeuses
King A:
Colonies blanches à beiges, contours réguliers,
brillantes
1-2mm
<2mm
Nombreuses
82
H96
King B:
Colonies blanches, à contours réguliers, plates,
brillantes
King A:
Colonies beiges, de contours irréguliers, au centre
surélevé brillantes
<1-3mm
2 mm
54
61
Tableau 10 : Evolution temporelle des Pseudomonas chez la sardine (S. pilchadus) conservée
à température réfrigérée.
TEMPS Température réfrigérée
Description des colonies Diamètre Nombre
H3
King B:
Colonies blanchâtres à contours réguliers, au centre
surélevé, légèrement brillantes
Colonies jaunes, contours non réguliers, plates, non
brillantes
Colonie orange brillante, transparente à contours
régulier
King A:
Colonies blanchâtres, de contours irréguliers, ternes
au centre surélevé
2mm
<2mm
1mm
<3mm
34
16
1
Nombreuses
H6
King B:
Colonies jaunes, contours réguliers, plates,
brillantes
Colonies beiges, de contours irréguliers, au centre
surélevé brillantes
King A:
Colonies blanches, de contours réguliers, plates,
crémeuses
2-3mm
<1-2mm
1-3mm
27
Nombreuses
Nombreuses
H9
King B:
Colonies blanches, de contours réguliers, bombées,
crémeuses
King A:
Colonies jaunes, contours réguliers, plates,
brillantes
Colonie orange brillante, transparente à contours
réguliers
<1-3mm
2mm
3mm
Nombreuses
14
5

Résultats
60
H24
King B:
Colonies beiges à contours légèrement irrégulier,
ternes, plates
Colonie jaunes, à contours réguliers, bombées,
brillantes
Colonie orange, plate, peu brillante, bombée
King A:
Colonies beiges, plates, peu brillantes à contours
régulier
Colonies jaunes, à contours irrégulier peu élevé,
brillantes
Colonie blanche peu brillante, bombée à contours
régulier
2mm
1-5mm
3mm
2mm
1-3mm
3mm
11
8
1
29
13
1
H48
King B et King A:
Colonies blanchâtres et beiges diffuses sur la boite
H72
King B et King A:
Colonies beiges diffuses jusqu'à formation d'un tapis
bactérien.
H96
King B:
Colonie blanche brillante, bombée à contours
réguliers
King A:
Colonies beiges à contours légèrement irréguliers,
ternes, plates
2mm
1-3mm
51
46
H120
King B:
Colonies jaunes, contours non régulier, plates, non
brillantes
Colonies blanches, à contours régulier, plates, ternes
King A:
Colonies beiges, plates, brillantes à contours régulier
2mm
1-2mm
<1-3mm
14
38
Nombreuses
H144 King A:
Colonies blanches, de contours régulier, bombées,
crémeuses
<1-2mm
Nombreuses

Résultats
61
4.3.9. Activités enzymatiques bactériennes d’altération
4.3.9.a. Activité lipolytique
La sardine fraiche (H0) ne présente aucune activité enzymatique bactérienne lipolytique. Cette
dernière ne se manifeste qu’après 6 heures et 24heures de conservation, respectivement, à
température ambiante et réfrigérée.
A température ambiante, l’activité lipolytique est maximale au début de son apparition à H6 et
H9 puis diminue à H24 et se stabilise à H48. A H72, cette activité augmente de nouveau et
atteint sa valeur maximale. A partir de H96, nous ne mesurons aucune activité lipolytique.
A température réfrigérée, l’activité lipolytique ne se manifeste qu’à H24, et reste stable
jusqu’à H72. A partir de H96, aucune activité lipolytique n’est détectée (figure 31).
Figure 28 : Aspect des colonies de
bactéries isolées sur milieu King A.
Figure 29 : Aspect des colonies de
bactéries isolées sur milieu King B.
AQUABIOR,2011 AQUABIOR,2011

Résultats
62
Figure 30: Evolution temporelle de l’activité lipolytique bactérienne chez la sardine
(S. pilchardus).
Figure 31 : Aspect macroscopique de la manifestation de l’activité lipolytique.
4.3.9.b. Activité gélatinolytique
L’activité gélatinolytique n’est pas détectée dans les échantillons frais. A température
ambiante, cette activité apparait après 6 heures de conservation, elle diminue jusqu’à H24 puis
augmente à nouveau jusqu’à la fin de l’expérimentation où elle disparait à H96.
0
1
2
3
4
5
6
7
H0 H3 H6 H9 H24 H48 H72 H96 H120 H144
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
Øhalo/Øcolonie
Durée (heures)
AQUABIOR, 2011
B.L

Résultats
63
Au réfrigérateur, l’activité gélatinolytique des échantillons se manifeste à la 9ème
heure de
conservation, cette activité baisse à H24 puis augmente progressivement et se stabilise à H72
jusqu’à la fin de l’expérience où elle disparait à H120.
Au cours de cette étude, nous notons un décalage de 3 heures pour l’apparition et de 24 heures
pour la disparition de l’activité gélatinolytique entre les échantillons conservés à température
ambiante et ceux au réfrigérateur (figure32).
Figure 32: Evolution temporelle de l’activité gélatinolytique bactérienne chez la sardine
(S. pilchardus).
Figure 33 : Aspect macroscopique de la manifestation de l’activité gélatinolytique.
0
1
2
3
4
5
6
H0 H3 H6 H9 H24 H48 H72 H96 H120 H144
Lot A : T° ambiante (25°C)/
réfrigérée (+4°C)
Lot B : T° réfrigérée (+4°C)
Øhalo/Øcolonie
Durée (heures)
AQUABIOR,2011
B.G
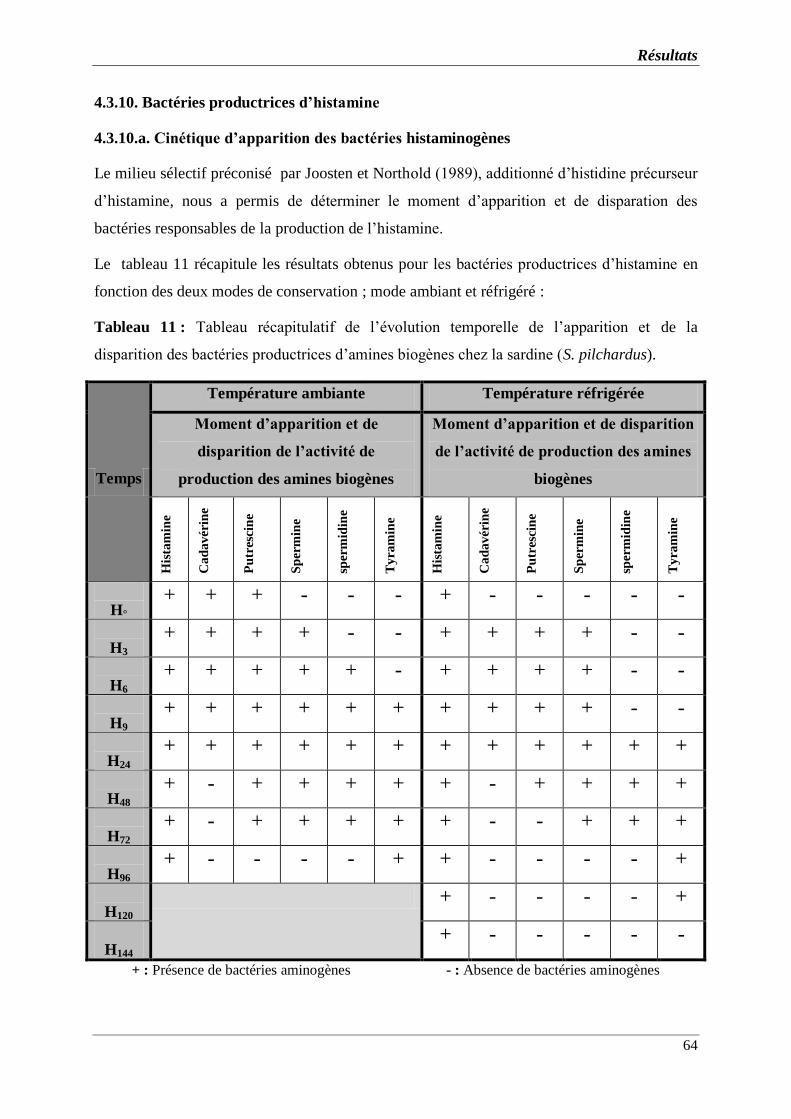
Résultats
64
4.3.10. Bactéries productrices d’histamine
4.3.10.a. Cinétique d’apparition des bactéries histaminogènes
Le milieu sélectif préconisé par Joosten et Northold (1989), additionné d’histidine précurseur
d’histamine, nous a permis de déterminer le moment d’apparition et de disparation des
bactéries responsables de la production de l’histamine.
Le tableau 11 récapitule les résultats obtenus pour les bactéries productrices d’histamine en
fonction des deux modes de conservation ; mode ambiant et réfrigéré :
Tableau 11 : Tableau récapitulatif de l’évolution temporelle de l’apparition et de la
disparition des bactéries productrices d’amines biogènes chez la sardine (S. pilchardus).
Temps
Température ambiante Température réfrigérée
Moment d’apparition et de
disparition de l’activité de
production des amines biogènes
Moment d’apparition et de disparition
de l’activité de production des amines
biogènes
His
tam
ine
Cad
avér
ine
Pu
tres
cin
e
Sp
erm
ine
sper
mid
ine
Tyra
min
e
His
tam
ine
Cad
avér
ine
Pu
tres
cin
e
Sp
erm
ine
sper
mid
ine
Tyra
min
e
H°
+ + + - - - + - - - - -
H3
+ + + + - - + + + + - -
H6
+ + + + + - + + + + - -
H9
+ + + + + + + + + + - -
H24
+ + + + + + + + + + + +
H48
+ - + + + + + - + + + +
H72
+ - + + + + + - - + + +
H96
+ - - - - + + - - - - +
H120
+ - - - - +
H144
+ - - - - -
+ : Présence de bactéries aminogènes - : Absence de bactéries aminogènes

Résultats
65
La figure suivante montre l’aspect macroscopique observé lors de la réalisation de ce test et le
virage du milieu sous ces colonies au violet qui est dû à la production d’amines biogènes.
Figure 34 : Aspect d’une colonie de bactéries aminogènes.
AQUABIOR,2011
B.A

41
DISCUSSION

Discussion
66
5. DISCUSSION
Le poisson subit immédiatement après sa mort un processus naturel de décomposition
qui est le résultat de la superposition de réactions chimiques, enzymatiques et bactériennes
(Aubourg, 2005). La conséquence en est une rapide altération des propriétés organoleptiques,
une réduction de la valeur nutritive et la formation de substances toxiques.
La qualité des produits de la pêche est définie par une série de caractéristiques
impliquant leur composition initiale, leur valeur nutritionnelle, les conditions de capture, de
stockage, de distribution et de commercialisation (Aguilar &Martinez, 2001). Les conditions
de manutention post-mortem influencent directement la fraicheur du produit (Aguilar,
Sanchez & Burgueno, 2000). Les modifications sensorielles affectent l’apparence, la texture,
l’odeur et la flaveur (Adolphe, 2006). Ces dernières s’installent très rapidement après la rigor-
mortis. Les basses températures exercent un effet protecteur sur la préservation de la qualité
organoleptique du poisson. En effet, nous observons un décalage dans le temps dans
l’apparition des signes manifestes de la détérioration de la qualité avec un effet de
préservation en faveur de la réfrigération.
Dans la présente étude, menée sur la sardine (S. pilchardus), le temps du rejet
organoleptique est à 24 H et 72 H, respectivement, à température ambiante et réfrigérée. Des
observations similaires ont été signalées chez la même espèce (Barhoumi, 1980; El Marrakchi
et al., 1990; Ababouch et al., 1996 ; Rouabhi, 2009). L’altération sensorielle s’accompagne
par l’installation progressive d’odeurs désagréables. En effet, la sardine est un poisson gras,
l’hydrolyse induite par les lipases et les phospholipases produit les acides gras qui sont très
insaturés qui feront l’objet d’un rancissement oxydatif responsable de la production de
composés de faibles poids moléculaire, cause de la flaveur désagréable de rancidité
(Burgueno, 2000). La dégradation de l’ATP permet également, la formation d’inosine
monophosphate (IMP), d’ammoniaque (NH3), de disulfure (H2S), d’acetalaldehyde et de
diacetyl responsables du développement d’odeurs désagréable.
L’installation de microorganismes accélère la dégradation du poisson (Gram, 1989 ;
Jorgensen et al., 2000). Cette dernière débute par la disparition de l’odeur de l’algue marine
laissant place à des odeurs aigres ou acides puis aminées ou soufrés enfin, ammoniacale ou
fécale à l’état putride. L’oxyde de triméthylamine (OTMA) inodore est réduit en TMA à
odeur d’ammoniac et en diméthyl amine (DMA) ou MMA et la production de NH3 croit
(Eymard, 2003).

Discussion
67
Durant la conservation à température ambiante et réfrigérée, les résultats montrent une
perte en eau probablement due au fait que les membranes cytoplasmiques du tissu des
poissons perdent leur rôle de barrière. De ce fait, l’accroissement de la concentration des
solutés dans l’espace extracellulaire provoque la sortie de l’eau (osmose) puis son évaporation
(Eymard, 2003). La capacité de rétention de l’eau est liée également à la capacité des
protéines myofibrillaires à conserver leur liaison avec l’H2O (Savage et al., 1990; Bremner,
2002). De plus les poissons gras présentent une forte proportion de muscle brun. Or ce
dernier peut rétrécir jusqu’à 52 % alors que le muscle blanc ne perd que 15 % de sa longuer
(Penfield & Campbell, 1990).
Chez la sardine vivante, le pH du muscle est à 7,2. Après les 2 et 8 premières heures de
la rigor-mortis, il atteint, respectivement, 6,8 et 5,8 (Watabe et al., 1991). La sardine contient
une forte proportion de muscle brun riche en glycogène (Shimizu et al., 1992). L’acidification
du muscle est due en grande partie à la glycogénolyse anaérobie et à la formation d’acide
lactique suite à l’épuisement des réserves d’oxygène des cellules musculaires (Huss, 1986).
Dans la présente étude, les valeurs basses du pH laisse suggérer que la sardine était déjà en
phase de post rigor-mortis (Aguilar et al., 2000). L’augmentation du pH mesurée chez la
sardine à J5 et J7 s’explique par la production de bases volatiles (NH3, TMA et DMA) et par
l’accumulation de peptides et d’amines dans le muscle (El Marrakchi et al., 1990).
Le suivi de l’oxydation des lipides est un outil indispensable dans l’évaluation de la
qualité du poisson (Melton, 1983). Cependant, les produits de l’oxydation des lipides
interagissent rapidement avec d’autres constituants qui ne sont pas sensibles au dosage
(Aubourg et al., 2007). La cinétique de formation des hydroperoxydes dépend du contenu
lipidique. Les espèces à taux lipidique élevé, présentent une augmentation de la concentration
en hydroperoxydes (Undeland et al., 1999). Les résultats enregistrés lors de notre étude
montrent la présence d’hydroperoxydes sur les échantillons frais. En effet, dès la mort de
l’animal, la baisse du pH due à la glycogénolyse représente des conditions favorables pour
initier la peroxydation lipidique. Quel que soit le mode de conservation, le même schéma
évolutif de production d’hydroperoxydes est observé avec un décalage dans le temps en
faveur de la réfrigération. En effet, le froid n’inhibe pas mais ralentit cette production
(Rodriguez, 2008). L’augmentation de la teneur en hydroperoxydes persiste et ne diminue
qu’en fin d’expérience. Il semble que cette période corresponde à la phase de propagation.
Chez la sardine, cette phase est longue en raison du pH post mortem qui est acide, favorisant

Discussion
68
la présence des formes actives de myoglobines (puissant catalyseur de lipoperoxydation pH
dépendante) (Baron & Anderson, 2002) et des ions métalliques.
Lors des premières heures de stockage, la teneur en TBA-rs reste plus ou moins stable
puis augmente jusqu’à la fin de l’expérience. La teneur et la rapidité de la formation des TBA-
rs sont dues au contenu en phospholipides du poisson. Ces substances sont le substrat de
formation des hydroperoxydes, eux-mêmes précurseurs de la formation des substances
réactives à l’acide thiobarbiturique (Ke & Nash, 1977). La perte des antioxydants naturels
durant la période de conservation pourrait contribuer à l’augmentation de l’oxydation
(Turksuban, 2003). Eymard (2003) et Aubourg (2001, 2004) ont montré que l’étude des
hydroperoxydes, des TBA-rs ainsi que l’analyse sensorielle constituent une combinaison
pertinente pour l’évaluation de la qualité du poisson et la formation d’odeur de rance liée
majoritairement aux TBA-rs.
La teneur en protéines totales décroit progressivement et continuellement quel que soit
le mode de conservation. L’effet des enzymes protéolytiques dans la chair de poisson
contribue à dégrader les tissus conduisant au ramollissement du muscle (Chéret, 2005).
L’effet de ces enzymes est contrôlé par des cofacteurs, le pH et la température environnante.
De plus, l’activité protéolytique varie énormément selon l’espèce, le sexe, la saison de pêche,
la période de frai, la maturité sexuelle et d’autres variables (Jiang, 2000). Les principales
protéases, les enzymes lysosomales, les calpaines et les cathépsines D (Hurley et al., 2000)
jouent un role important dans l’évolution de la qualité post-mortem du poisson.
Plusieurs études ont montré l’effet de la température sur le développement des bactéries
impliquées dans l’augmentation de la teneur en ABVT. Nishimoto et al. (1985) rapportent des
valeurs d’ABVT de 30 mg / 100 g de chair pour S. melanostica après 24 heures de
conservation à température ambiante. Erkan et Ozden (2008) trouvent des valeurs de 11,11
mg / 100g et 29,23 mg /100g dans le muscle de sardine (S. pilchardus) conservée,
respectivement, sous glace à J0 et J9. Dans la présente étude, les valeurs de l’ABVT passent
de 3,11 ± 0,93 à 40,52 ± 2,50 et à 46,48 ± 3,94 mg/ 100g, respectivement, à température
ambiante et réfrigérée. Au réfrigérateur, les teneurs en ABVT sont moins élevées que celles
observées à température ambiante. Ceci peut être expliqué par la différence qualitative et
quantitative de la flore microbienne d’altération observée. La limite maximale admissible en
ABVT est limitée à 35mg / 100g de chair de poisson. Au delà le poisson est considéré
impropre à la consommation (El Marrakchi, 1990). Nos échantillons sont donc jugés

Discussion
69
impropres à la consommation à H72 et H120 respectivement à température ambiante et
réfrigérée.
L’altération des poissons est due aux enzymes tissulaires ainsi qu’aux microorganismes.
Ces derniers jouent un rôle très important. De nombreux facteurs conditionnent les modalités
de l’altération microbienne :
Variété de poisson, pH de la chair, richesse en graisse.
Habitat du poisson, type et étendue de la contamination bactérienne.
Conditions de pêche et de stockage : conditionnement en milieu aérobie et
anaérobie pouvant entrainer un écrasement des tissus, présence ou non d’un
étêtage et éviscération, température de stockage,etc…….
L’implantation de germes sur le poisson est responsable du mauvais goût, des
mauvaises odeurs, du surissement, de la coloration / décoloration, de la dégradation et de la
putréfaction du produit. L’altération aboutit souvent à la formation d’amines (TMA,
ABVT…) et d’ammoniaque. Il y a également formation d’H2S, de méthyl mercaptan et
d’autres composés nauséabonds, ainsi que des amines toxiques comme l’histamine formées à
partir d’acide aminé, l’histidine.
Nous avons étudié en premier lieu la flore d’altération qui s’installe dans la chair de
poisson et son évolution dans le temps. L’estimation de la flore mésophile aérobie totale
(FMAT) montre une valeur initiale plus ou moins faible, dans la limite de l’acceptable. La
circulaire du 4 avril 1980 qui commente l’arrêté du 21 Décembre 1979 sur les denrées
d’origine animale éditée par CNERA, propose que la charge de la FMAT soit inférieure à 105
UFC par gramme de chair pour un niveau de qualité acceptable. Nous supposons donc que la
sardine fraiche faisant l’objet de notre analyse répond à ce critère et est donc salubre. Ce
niveau est maintenu jusqu’à H6 pour les deux modes de conservation. Au-delà nos
échantillons dépassent le taux réglementaire et représenteraient un éventuel danger
(Ababouch et al., 1996 et Kakli et al., 2006).
La recherche et la numération des coliformes totaux montrent leur présence dans la
chair de poissons dès le début. Ces bactéries regroupent de nombreuses espèces dont les
animaux constituent des hôtes normaux et représentent 10 % de la flore totale. Ce sont des
contaminants alimentaires très fréquents, se développent de manière très abondante et
dégradent le produit.

Discussion
70
Les Coliformes fécaux, quant à eux, ne peuvent dépasser 10 germes / g de chair selon
l’arrêté du 21 Décembre 1979. Nos échantillons dépassent largement cette limite, et sont donc
impropres à la consommation. Ceci ne correspond pas à ce qui a été observé par Ababouch
(1996) et El Marrakchi (1990). En effet, la croissance de ces bactéries sur nos échantillons est
plus rapide et plus importante. On suppose que la charge initiale du lot retenu pour
l’expérimentation était importante à laquelle s’ajoutent les conditions de stockage et de
manipulation du produit. En effet, l’utilisation de caisses en bois dans un état hygiénique
inadéquat augmente le risque de contamination. Ces caisses sont déjà chargées en germes qui
adhéreront à nos échantillons et fausseront donc l’estimation de la charge initiale. Selon la
réglementation en vigueur en Algérie (janvier, 2010), l’utilisation de caisse en plastique pour
la commercialisation des poissons est obligatoire et ce pour des raisons sanitaire, des
sanctions et des amandes sont prévues pour le non respect de cette loi.
Les Streptocoques sont des germes très répandus dans la nature en raison de leur grande
résistance aux conditions défavorables du milieu (température : 0-50°C, pH 5-9,6 la bile à
40 % et 6,5 % NaCl). Selon Lancefield (1958), ces germes sont moins associés aux
pathogènes que les coliformes fécaux. Nous pensons que les conditions de stockage que nous
avons adoptées au cours de notre expérimentation ne constituent pas des éléments pouvant
ralentir la croissance des streptocoques. Il faut penser à d’autres moyens de transformation et
de traitement des échantillons pour les éliminer. Leur présence dans l’aliment n’indiquera pas
une contamination fécale forcément.
Un certain nombre de germes pathogènes jugés dangereux pour la santé est recherché :
La présence de bactéries sulfito-rédutrices se manifeste aux premières heures de
conservation et restent jusqu’en fin d’expérience. Vu leur caractère de résistance grâce à leurs
spores, ces microorganismes persistent dans l’aliment et sont responsables de production
d’H2S provoquant ainsi une altération organoleptique conséquente. Dans nos échantillons, les
bactéries sulfito-réductrices présentent une faible proportion et augmentent faiblement comme
observé par Huss (1988) chez les poissons. Les groupes prédominants sont S.
putrefaciens et certaines Enterobacteriaceae comme montré par Liston (1992). Le lot est
rejeté si les germes dépassent 10 / g de chair selon le même arrêté du 21 Décembre 1979.
Tous les staphylocoques présents dans les aliments ne sont pas enterotoxinogènes. Ce
n’est que depuis peu que l’on admet le rôle de ces derniers comme germes indicateurs de
contamination humaine, animale ou originelle dans les aliments, crus en particulier. Cette
notion permet donc d’accepter des staphylocoques en petit nombre dans les aliments. Ces

Discussion
71
germes sont naturellement présents sur la peau et les muqueuses des humains et des animaux
avec des charges qui peuvent largement dépasser les 100000 / cm2. Dans les aliments, le
risque devient grand quand leur nombre dépasse cette valeur. D’après les résultats, nos
échantillons ne présentent pas un danger d’ordre sanitaire et ce quel que soit le mode de
conservation.
Les Pseudomonas sont fréquents dans les produits alimentaires et survivent même aux
faibles températures. Ils sont dotés d’activités métaboliques très développées, ont souvent un
pouvoir protéolytique parfois lipolytique et peuvent dégrader des substances très variées. Les
groupes les plus fréquents incriminés dans la détérioration des aliments sont : P. fluoresens, P.
putida, P. ludensis, etc. Ces microorganismes ne sont pas généralement pathogènes pour
l’homme, mais certains peuvent exceptionnellement être associés à des troubles digestifs tel
que P. aerogenosa suspecté dans des gastro-entérites et P. cocovenenans qui produit des
substances toxiques. Par ailleurs, Ababouch (1996) a montré qu’il existe une relation directe
dans la prolifération de ces germes dans la perte de la qualité chez la sardine atlantique (S.
pilchardus).
L’association française de normalisation (AFNOR, 1973) définit un échantillon sain par
absence de salmonelles dans 25 g d’aliment et selon l’arrêté du 21 Décembre 1979 qui impose
l’absence de tout germe pathogène et de ses toxines. Notre échantillon montre la présence de
divers germes, correspondant probablement à différents groupes : Klebsiella, Serratia,
Salmonella, Proteus, Shigella, etc. Pour leur identification, il faudrait passer par une série de
tests biochimiques, immunologiques et moléculaire. Par ailleurs, leur présence dès le début
laisse supposer une matière première déjà contaminée, ce qui présente un réel danger pour le
consommateur. Nos résultats concordent aves ceux trouvés par Gennari (1999) chez S.
pilchardus.
Nous avons isolé des germes dotés d’activité enzymatique d’altération sur les deux lots
d’échantillons conservés en modes ambiant et réfrigéré :
Nous avons noté la présence d’activité lipolytique avec un décalage horaire pour son
apparition, Il semblerait que la réfrigération préserve de la lypolyse. Toutefois, nous
enregistrons un temps de disparition similaire pour les deux modes, on peut éventuellement
émettre l’hypothèse suivante :
Les germes impliqués dans cette activité sont nombreux, nous rencontrons
principalement les Pseudomonas et les Psychrobacter se développant à basse température et

Discussion
72
dotées dans grand pouvoir lipolytique (Macrae, 1975 ; Liston, 1980). Nous supposons, donc,
que l’activité du premier lot est le résultat de la flore présente à l’état initiale tandis que pour
le second, la flore incriminée s’installe probablement au cours du stockage au froid
(contamination croisée), vu le caractère psychrotrophe des germes les plus lipolytiques, nous
supposons qu’ils épuisent leur substrat rapidement et que la disparition de l’activité coïncide
dans le temps pour les deux lots (Shewan, 1960).
Concernant l’activité gélatinolytique, nous notons un décalage horaire pour l’apparition
et la disparition de ce dernier, la réfrigération retarde cette dernière mais sans diminuer de son
pouvoir. La gélatinolyse et la collagénolyse se succèdent dans le temps (Haard, 1992). Ces
deux enzymes attaquent les structures permettant le maintien des liaisons préservant une
bonne texture (Chéret, 2005), cette activité dépend aussi de la matière première qui dépend
elle-même des facteurs biologiques de l’espèce : l’âge, le sexe et aussi de la zone et la saison
de pêche. Les microorganismes protéolytiques présents dans nos échantillons augmentent en
fonction du temps, l’arrêt de cette activité est justifié par l’épuisement de son substrat (Olson,
2003).
L’activité de l’histidine décarboxylase est présente dès le début et jusqu’en fin
d’expérience. En effet, la sardine appartient au groupe des espèces les plus fréquemment
impliquées dans la scombrotoxité due au fait que leur muscle contient une teneur élevée en
histidine libre (Kim et al., 2003). La présence de l’histamine dans le muscle de la sardine
fraiche est un indice de traitement inadéquat des échantillons (mauvaise gestion de la
température lors du stockage ainsi que les conditions hygiéniques inappropriées lors de la
pêche). Les résultats trouvés lors de la présente étude rejoignent ceux montrés par Ababouch
et al., (1996), Pacheco-Aguilar et al.(2000) et Chaouqy & El Marrakchi (2005).
Cette production excessive d’histamine d’origine microbienne est probablement due à la
présence d’un taux élevé d’histidine dans le muscle rouge des poissons, des conditions
optimales de température (26°C) et un pH approprié à la synthèse et l’activité de l’histidine
décarboxylase. Les résultats obtenus lors de l’estimation chimique des teneurs en histamine
viennent consolider ceux obtenus lors de l’analyse microbiologique. Nous avons pu isoler des
bactéries histaminogènes dès les premières heures de conservation et jusqu’en fin
d’expérimentation. Plusieurs études ont montré que la flore productrice d’histamine est
mésophile, et qu’à des températures comprises entre 0 et 20°C, cette production est massive
(Ababouch et al., 1996 ; Bover-Cid, 2005). Cependant, Photobacterium phosphorum et
photobacterium histaminum montrent une activité décarboxylase à +4°C et même à -20°C

Discussion
73
(Okuzumi et al., 1981 ; Fujii et al., 1994). Les bactéries psychrophiles histaminogènes que
l'on connaît conduisent à l'accumulation de grandes quantités d’histamine dans la chair de
poisson (Olofsson et al., 2007), il est préférable de consommer le poisson rapidement après sa
capture, même s'il est conservé sous glace.

CONCLUSION
ET
PERSPECTIVES

Conclusion & Perspectives
74
6. CONCLUSION ET PERSPECTIVES
La présente étude apporte des informations importantes sur les changements sensoriels,
biochimiques et microbiologiques intervenant lors de la conservation de la sardine (S.
pilchardus) en fonction de la durée et de la température.
En conclusion, Le poisson est un système dynamique dans lequel les changements
arrivent dans le pH, la composition nutritive et la microflore au fil du temps. La croissance et
le métabolisme de microorganismes sont les éléments impliqués dans la manifestation de
l’altération sensorielle du produit le rendant peu convenable pour la consommation humaine.
La réfrigération a montré un ralentissement de la dégradation autolytique, l'hydrolyse et
l’oxydation des lipides, la modification du profil protéique et l’altération microbiologique.
La mesure des produits primaires de la peroxydation lipidique notamment les hydroperoxydes
fournirait l’information la plus correcte sur l’intensité de la lipoperoxydation, (Laguerre et al.,
2007). Ceci doit être complété par la quantification des produits secondaires de l’oxydation
des lipides (TBA-rs), composés plus stables, nécessaire pour estimer de façon plus fiable le
niveau d’oxydation des lipides (Eymard, 2003). Néanmoins, il semble que les pics en TBA-rs
enregistrés correspondent à une diminution des hydroperoxydes et à des niveaux d’altérations
sensorielles très avancées chez la sardine.
Selon Sato et al. (1995); Mendes (1999); Özogul et al. (2004) ; Chaouqy & El Marrakchi
(2005), l’absence de corrélation entre accumulation d’histamine et altération organoleptique,
explique que le paramètre de l’histamine est beaucoup plus considéré par les hygiénistes
comme un indice sanitaire qu’un indice fiable d’altération du poisson. De plus, nous
constatons une similitude dans la dynamique de synthèse de l’histamine et des produits de la
peroxydation lipidique, particulièrement les TBA-rs. Selon Uchida et al.(1990), l’histamine
peut induire l’oxydation lipidique et par conséquent le rancissement pendant la transformation
et la conservation du poisson (Mendes, 2009). En effet, il affirme que l'histamine exerce un
effet stimulant sur la peroxydation de l'acide linoléique par le groupement amine et imidazole
de l'histamine.
L’estimation chimique de certains métabolites peut être utilisée comme des indices de
qualité, comparée à des méthodes microbiologiques, longues et coûteuses. Ces indices de
qualité incluent l’estimation de l'azote volatil total, TMA, la dégradation des protéines et
l’hypoxanthine. Les ratios entre des produits de dégradation de l’ATP (valeur K) et les amines
biogènes ont aussi été utilisés pour quelques temps comme des indices de qualité, ces derniers
sont représentatifs qu’en combinant plusieurs métabolites, des propriétés sensorielles et

Conclusion & Perspectives
75
l’analyse microbiologique. L’estimation de la FMAT et les coliformes totaux et fécaux nous
renseignent sur l’état hygiénique de l’échantillon. La recherche de germes pathogènes dans
nos échantillons nous a révélé leur présence due certainement à un défaut d’hygiène tout au
long de la filière de pêche.
Il serait intéressant de se baser sur des dosages de métabolites qui se manifestent sur
l’aspect organoleptique et seront plus facilement perçus par les consommateurs. Le but de ces
travaux vise de proposer des combinaisons faciles à réaliser, peu onéreuses et rapides pour
l’évaluation du niveau d’altération des produits de la pêche : en se basant sur les résultats
enregistrés lors de la présente étude nous pouvons proposer la combinaison incluant l’analyse
sensorielle, dosage des produits d’oxydations lipidique, notamment les produits secondaires,
l’ABVT et une estimation de la charge microbienne.
La recherche de germes pathogènes ne se fera que si l’aliment est incriminé dans une
intoxication alimentaire pour l’éliminer du marché.
Pour compléter cette étude, il serait intéressant de :
Quantifier le substrat des amino-acides adécarboxylases des bactéries aminogènes, et
l’établissement du profil en acides aminés libres présents dans le muscle et son suivi
dans le temps.
Suivre la cinétique de production des autres amines biogènes (cadaverine, putrescine,
spermine, spermidine, tyramine, tryptamine) et les microorganismes responsables de
leur production.
Approfondir cette étude par l’identification moléculaire des souches histaminogènes
isolées lors du présent travail et tester leur capacité à produire l’histamine sur milieu
liquide et doser cette dernière par techniques chromatographiques.
Compléter l’étude sur les activités gélatinase et collagénase par une étude chimique
des produits correspondant.
Etudier le rôle des microorganismes dans la dégradation des phospholipides.
Penser à la formation professionnelle des pêcheurs. En effet, il faudrait procéder à une
sensibilisation ciblée sur les dysfonctionnements et ce qu’ils peuvent engendrer sur le
plan sanitaire, multiplier les systèmes et les fréquences de contrôles, unifier
l'application des dispositions réglementaires et imposer la possession d'une traçabilité
de la production.

REFERENCES
BIBLIOGRAPHIQUES

Références bibliographiques
76
7. REFERENCES BIBLIOGRAPHIQUES
A
Ababouch L.H., Afilal M.E., Rhafiri S. & Busta F.F., 1991a. Identification of
histamineproducing bacteria isolated from sardines (Sardina pilchardus) stored in ice and at
ambient temperature (25°C). Food Microbiology, 8: 127-136.
Ababouch L.H., Afilal M.E., Benabdeljelil H. & Busta F.F., 1991b. Quantitative changes
in bacteria, amino acids and biogenic amines in sardine (Sardina pilchardus) stored at
ambient temperature (25–28°C) and in ice. International Journal of Food Science and
Technology, 26: 297-306.
Abi-ayad S. M., E.-A., 1998- Etude expérimentale de la biologie de la reproduction de la
perche fluviatile (Perca fluviatilis). Effet de la composition en acides gras de la série (n-3) de
l’alimentation des géniteurs sur la qualité des œufs et des larves. Thèse de doctorat, faculté
des sciences, Université de Liège, Belgique, 147p.
Adolphe Y., 2006. Etude du fonctionnement du système lactoperoxydasique et validation de
son effet inhibiteur vis-à-vis de flores du poisson. Thèse de Doctorat en procédés
biotechnologiques et alimentaires. Institut National Polytechnique de Lorraine, 132p.
Afilal M.A., Daoudi H., Jdaini S., Asehraou A. & Bouali A., 2006- Study of the histamine
production in the red flesh fish (Sardina pilchardus) and a with flesh fish (Dicentrarchus
ounctatus). Turkish journal of fisheries and aquatic sciences. 6 :43-48.
Afilal M.E. & Zlaiji E., 1997. Isolement de bactéries mésophiles productrices d’amines
biogènes dans Sardina pilchardus. Industries Alimentaires et Agricoles, 114(5): 274-277.
Agence canadienne d’inspection des aliments, 2005. Liste des noms communs acceptables
au Canada pour les espèces de poissons et de fruits de mer : Sardine.
Agence canadienne d’inspection des aliments, 2005. Poisson. Un des neuf allergènes
alimentaires les plus courants.
Aguilar R., Lugo-Sanchez M.E. & Robles-Burgueno M.R., 2000. Post-mortem
biochemical and functional characteristic of Monterey sardine muscle stored at 0 °C. Journal
of Food Science, 65(1): 40-47.
Al-Sayed Mahmoud Kassem, 2007. Extraction, fractionnement et caractérisation des lipides
polyinsaturés d'oeufs de la truite arc-en-ciel (Oncorhynchus mykiss), thèse de doctorat.
Université de Nantes. p143.

Références bibliographiques
77
Antonacopoulos N., Vyncke W., 1989. Determination of volatile basic nitrogen in fish: a
third collaborative study by West European Fish Technologists Association (WEFTA).
Zeitschrift fur Lebensmittel-Untersuchung und-Forschung A, 189, 309-316.
AOAC, 1995c. Histamine in seafood: fluorometric method. Sec. 35.1.32, Method 977.13. In:
Cunniff P.A. (Ed.), Official Methods of Analysis of AOAC International, 16th
Ed. AOAC
International, Gaithersburg, MD. pp. 16-17.
Archibald F., 2000. The presence of coliform bacteria in Canadian pulp and paper mill water
systems - a cause for concern. Water Qual Res J. Canada, 35:1-22.
Aubourg, S. P., 1993- Review: interaction of malondialdehyde with biological molecules-
new trends about reactivity and significance. International Journal of Food Science and
Technology. 28: 323–335.
Aubourg S., Medina I. & Gallardo J., 1998a. Quality assessment of blue whiting
(Micromesistius poutassou) during chilled storage by monitoring lipid damages. Journal of
the Agricultural and Food Chemistry. 46: 3662–3666.
Aubourg S., Sotelo C. & Perez-Martin R., 1998b. Assessment of quality changes in frozen
sardine (Sardina pilchardus) by fluorescence detection. Journal of the American Oil Chemists
Society. 75: 575–580.
Aubourg S., & Medina I., 1999a. Influence of storage time and temperature on lipid
deterioration during cod (Gadus morhua) and haddock (Melanogrammus aeglefinus) frozen
storage. Journal of the Science of Food and Agriculture.79: 1943–1948.
Aubourg S., 1999b. Review: recent advances in assessment of marine lipid oxidation by
using fluorescence. J. of the American Oil Chemists Society. 76: 409-419.
Aubourg S., 2001. Damage detection in horse mackerel (Trachurus trachurus) during chilled
storage. Journal of the American Oil Chemists Society. 78: 857–862.
Aubourg S, Carmen Pineiro , Jose M. Gallardo & Jorge Barros-Velazquez, 2005.
Biochemical changes and quality loss during chilled storage of farmed turbot (Psetta
maxima), Food Chemistry. 90: 445–452
Aubourg S., Quitral V., Larraın A., Rodrıguez A., Gomez J., Maier L., & Vinagre J.,
2007. Autolytic degradation and microbiological activity in farmed Coho salmon
(Oncorhynchus kisutch) during chilled storage, Food Chemistry. 104: 369-375.

Références bibliographiques
78
B
Bandarra, N.M., Batista, I., Nunes, M.L., Empis, J.M. and Christie, W., 1997. Seasonal
changes in lipid composition of sardine (Sardina pilchardus). Journal of Food Science. 62:
40-42.
Bandarra N.M., Batista I., Nunes M.L. & Empis J.M., 2001. Seasonal variation in the
chemical composition of horse-mackerel (Trachurus trachurus). European Food Research
and Technology, 212: 535-539.
Bardóz S., Grant G., Brown D.S., Ralph A. & Pusztai A., 1993. Polyamines in
foodimplications for growth and health. Journal of Nutritional Biochemistry, 4: 66-71.
Barhoumi M., 1980. Essai de conservation de la sardine par la glace et l’eau de mer refroidie.
Institut Scientifique des pêches Maritimes. Travaux et Documents N°26. Casablanca,
Morocco.
Barhoumi M ., Faye A., Teutscher, Lima Dos Santos, Vike E., 1982. Storage
characteristics of sardines (Sardina pilchardus) stored on ice. FAO Fish Rep N 268. pp 32-39.
Baron, C.P. & Andersen, H.J., 2002- Myoglobin-induced lipid oxidation. A review.
Journal of Agricultural and Food Chemistry. 50: 3887-3897.
Bennour M., El Marrakchi A., Bouchriti N., Hamama A. & El Ouadaa M., 1991.
Chemical and microbiogical assessments of mackerel (Scomber scombrus) stored in ice.
Journal. Food Product. 54: 789-792.
Berset C. & Cuvelier M. E., 1996. Methods of estimating the degree of lipid oxidation and
of measuring antioxidizing power. Sciences des Aliments. Vol. 16, pp 219-245.
Bostoglou N.A., Fletouris D.J., Papageorgiou G.E., Vassilopoulos V.N., Mantis A.J. &
Trakatellis A.G., 1994. Rapid sensitive and specific thiobarbituric acid method for
measuring lipid peroxidation in animal tissue, food and feedstuff samples. Journal of
Agricultural and Food Chemistry, 42: 1931-1937.
Bradford M. M., 1976. A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry.
72, 248-254.
Bremner H.A., 2002. Safety and quality issues in fish processing. Cambridge Woodhead
Publishing Limited, 507 p.
Brink B.T., Damink C., Joosten H., 1990. Occurrence and formation of biologically active
amines in foods International Journal of Food Microbiology, 11, pp. 73–84.

Références bibliographiques
79
Boulay C., 1999. Intoxications histaminiques : poissons en cause, conditions de survenue et
mécanismes pathogéniques actuellement connus. Thèse de doctorat. Ecole nationale de
vétérinaire de Nantes. 122p.
Bourgeois, C.M. & Leveau, J.Y., 1991. Techniques d’analyse et de contrôle dans les
industries agro-alimentaires. Le contrôle microbiologique, Lavoisier, Apria.
Bourgeois, C.M. Mescle, G.F. & Zucca, J., 1996. Microbiologie alimentaire. Aspect
microbiologique de la sécurité et de la qualité des aliments. Londres, Paris.
Bourre JM, 2005. Dietary omega-3 Fatty acids and psychiatry: mood, behaviour, stress,
depression, dementia and aging. J. Nutr Health Aging;9(1):31-8.
Bover-Cid S., Holzapfel W. H., 1999. Improved screening procedure for biogenic amine
production by lactic acid bacteria. International Journal of Food Microbiology. 53:33–41.
Burgueño T.,Pedro J-N. , 2000. 13C NMR substituent chemical shifts in hydroxy-p-
benzoquinones. Magnetic resonance in chemestry-Volume 38, issue 5, p390-393.
C
Calder PC, 2004. n-3 Fatty acids and cardiovascular disease: evidence explained and
mechanisms explored. Clin Sci (Lond) July;107(1):1-11.
Careche M. & Tejada M., 2002. The effect of neutral and oxidized lipids on functionality in
hake (Merlucciuc merluccius): A dimethylamine and formaldehyde forming species during
frozen storage. Food Chemistry. 36: 113-128.
Castell C.H., 1950. The influence of trimethylamine oxide on the bacterial reduction of redox
indicators. J. Fish. Res. Bd. Can., 7 : 567-575.
Castrillon, A., Alvarez-Pontes, E., Garcıa, M., & Navarro, P., 1996. Influence of frozen
storage and defrosting on the chemical and nutritional quality of sardine (Clupea pilchardus).
Journal of the Science of Food and Agriculture. 70 : 29-34.
CEAEQ, 2000. Recherche et dénombrement des coliformes totaux; méthode par filtration sur
membrane. Centre d’expertise en analyse environnementale, Gouvernement du Québec, p 25.
Chang, S. C., Kung, H. F., Chen, H. C., Lin, C. S., & Tsai, Y. H., 2008. Determination of
histamine and bacterial isolation in swordfish fillets (Xiphias gladius) implicated in a food
borne poisoning. Food Control, 19, 16–21.

Références bibliographiques
80
Chaouqy N.E. & El Marrakchi A., 2005. Aspects chimiques et bactériologiques de
l’anchois (Engraulis encrasicholus) entreposé sous glace et à moyenne température (20-
25°C). Revue de Médecine Vétérinaire, 156(6): 341-349
Chaveron H., 1999. Molécules toxiques. Molécules formées au cours des réactions
d’oxydation ed. Introduction à la toxicologie nutritionnelle. Londres : Tec Et Doc, p 175-83.
Cheftel J. & Cheftel H., 1976- Introduction to the biochemistry and technology of foods.
Editorial acribia, Zaragoza.
Chen, H. C., Kung, H. F., Chen, W. C., Lin, W. F., Hwang, D. F., Lee, Y. C., et al., 2008.
Determination of histamine and histamine-forming bacteria in tuna dumpling implicated in a
food-borne poisoning. Food Chemistry, 106, 612–618.
Chen, H. C., Huang, Y. R., Hsu, H. H., Lin, C. S., Chen, W. C., Lin, C. M., et al., 2010.
Determination of histamine and biogenic amines in fish cubes (Tetrapturus angustirostris)
implicated in a food-borne poisoning. Food Control, 21, 13–18.
Chéret R., 2005. Effet des hautes pressions sur les indicateurs de maturation de la viande et
d’altération du muscle de poisson. Thèse de doctorat. Université de Nantes. 176p.
Cheriot S., 2007. Role des produits de la réaction de Maillard dans l’inhibition de l’oxydation
enzymatique des phenols et des lipides. Thèse de doctorat. Université de Paris, Agro Paris
Tech. p239.
Cho S.-Y., Endo Y., Fujimoto K. & Kaneda T., 1989. Oxidative deterioration of lipids in
salted and dried sardine during storage at 5°C. Bulletin of the Japanese Society of Scientific
Fisheries, 55(3): 541-544.
Cobb, B. F., Aoaniz, I., & Thompson, C. A., 1973. Biochemical and microbial studies
on shrimp: Volatile nitrogen and amino nitrogen analysis. Journal of Food Science, 38, 431–
435.
Codex Alimentarius., 1993. Section 7.5, HACCP Guidelines, Vol. 1, sup.1 : 95-103.
D
Davies C., 2004. Chapter 8: lipolysis in lipid oxidation. In: Steele R. (Ed.), Understanding
and measuring the shelf-life of food. Woodhead Publishing Limited (England) and CRC Press
LLC (USA). p. 142-161.
Decker E.A. & Hultin H.O., 1990. Nonenzymic Catalysts of Lipid Oxidation in Mackerel
Ordinary Muscle. Journal of Food Science. 55 : 951-953.

Références bibliographiques
81
De las Rivas, B., González, R., Landete, J. M., & Muñoz, R., 2008. Characterization of
a second ornithine decarboxylase isolated from Morganella morganii. Journal of Food
Protection, 71, 657-661.
De las Rivas, B., Marcobal, A., Carrascosa, A. V., & Muñoz, R., 2006. PCR detection of
Food borne bacteria producing the biogenic amines histamine, tyramine, putrescine, and
cadaverine. Journal of Food Protection, 69, 2509e2514.
Doyle M.P., Beuchat L.R. & MontVille T.J., 1997. Food Microbiology: Fondamentals and
Frontiers. Washington DC: ASM Press. p 94.
Duflos G., Dervin C., Malle P. and Bouquelet S., 1999. Use of biogenic amines to evaluate
spoilage in plaice and whiting. Journal of AOAC International. 82 : 1357-1363.
Dumay J., 2006. Extraction des lipides en voie aqueuse par bioréacteur enzymatique combiné
à l’ultrafiltration: Application à la valorisation de co-produits de poisson (Sardina
pilchardus). Thèse de doctorat de l’université de Nantes. p318.
Dyer, W.J. & Fraser, D.I., 1959. Proteins in Fish Muscle. 13. Lipid Hydrolysis. Journal of
Fisheries Research Board of Canada. 16: 43-52.
E
Edberg, Rice, Karlin & Allen, 2000. Escherichia coli: the best biological drinking water
indicator for public health protection. Journal of Applied Microbiology, 88: 106-116.
El Marrakchi A., Bouchriti N., Bennour N., Hmama A. & Tagafit H., 1990. Sensory,
chemical and microbiological assessment of moroccan sardines (Sardina pilchardus) stored in
ice. J. Food Prot. 53: 600-605.
E.U., 1996. Règlement (CE) n° 2406/96 du conseil du 26 novembre 1996 fixant des normes
communes de commercialisation pour certains produits de la pêche. Official Journal of the
European Union L 334. pp. 1-15.
E.U., 2005. Règlement (CE) n°2073/2005 de la commission du 15 novembre 2005,
concernant les critères microbiologiques applicables aux denrées alimentaires. Official
Journal of the European Union L 338. pp. 22.
Eymard S., 2003. Mise en évidence et suivi de l'oxydation des lipides au cours de la
conservation et de la transformation du chinchard (Trachurus trachurus): choix des procédés.
Thèse de doctorat de l'Université de Nantes, p143.
Eymard S. & Genot C., 2003. A modified xylenol orange method to evaluate formation of
lipid hydroperoxides during storage and processing of small pelagic fish. European Journal of
Lipid and science technology.105: 497-501.

Références bibliographiques
82
F
F.A.O., 2009. The State of World Fisheries and Aquaculture-2008. In: FAO Fisheries
and Aquaculture Department. Food and Agriculture Organization of the United
Nations. Rome.
F.A.O., 2010a. Fish and fishery products: world apparent consumption statistics based on
food balance sheets. FAO Yearbook 2008. Food and Agriculture Organization of United
Nations, Rome, Italy. pp. 215-219.
F.A.O., 2010b. Capture production. FAO Yearbook 2008. Food and Agriculture Organization
of United Nations, Rome, Italy. pp. 28-30
F.A.O. 2011. The State of World Fisheries and Aquaculture. Table 1. Rome, FAO. 2011.
197p.
F.D.A (US Food and Drug Administration), 2001. Fish and Fishery Products Hazards and
Controls Guide (3rd Ed.). FDA, Center for Food Safety and Applied Nutrition, Office of
Seafood, Washington DC, USA.
Fernandez S.J. & Mackie I., 1987. Comparative rates of spoilage of fillets and whole fish
during storage of haddock (Melanogrammus aeglefinus) and herring (Clupea harengus) as
determined by the formation of non-volatile and volatile amines. Int. J. Food Sci. Technol.,
22: 385-390.
Folch J., Lees M. & Sloane Stanley G.H.S., 1957- A simple method for the isolation and
purification of total lipides from animal tissues. Journal of Biological Chemistry. 226 : 497-
509.
Frank H.A., Yoshinaga D.H., 1984. Histamine production in Tuna. Seafood toxins.ACS
symposium series N°.262. American Chem. SOC., Pp. 443-451. E.P. Ragelis edition,
Washington, D.C.
Frankel, E.N., 1998 - Lipid oxidation. The Oily Press.10. Dundee, Scotland. 10.
Frankel E.N., Neff W.E. & Weisleder D., 1990. Determination of methyl linoleate
hydroperoxides by 13C nuclear magnetic resonance spectroscopy. Methods in Enzymology
186 :380-387.
Fujii T., Kurikara K. & Okuzumi M., 1994. Viability and histidine decarboxylase activity
of halophilic histamine-forming bacteria during frozen storage. Journal of Food Protection,
57(7): 611-613.

Références bibliographiques
83
G
Genot C., 1996. Some factors influencing TBA test. Report of diet-ox project. p.92-1577.
Genot C., 2000. Congélation et qualité de la viande. Techniques et pratiques, INRA Ed.
Paris.
Genot C., Eymard S. & Viau M., 2004. Comment protéger les acides gras polyinsaturés à
longues chaînes oméga 3 (AGPI- LC ω3) vis-à-vis de l’oxydation. Oléagineux, Corps gras,
Lipides, 11(2): 133-141.
Genot C., Meynier A., Riaublanc A. & Chobert J.M., 2003. Protein Alterations Due to
Lipid Oxidation in Multiphase Systems. In: Kamal-Eldin A. (Ed.), Lipid oxidation pathways.
AOACS Press Champaign. p. 265-292.
German, J.B., 1992. Lipid oxidation in fish tissue. enzymatic initiation via lipoxygenase.
Journal of Agricultural and Food Chemistry. 33: 680-683.
Gharbaoui N., 2010. Mise en évidence de l’oxydation lipidique au cours de la conservation
des produits de la pêche, mémoire de Magister en gestion des ressources aquatiques. Faculté
des Sciences. Université d’Oran, 68 p.
Gram L., 1989. Identification, characterization and inhibition of bacteria isolated from
tropical fish. PhD thesis. Royal Veterinary and Agricultural University. Denmark
Gram L. & Dalgaard P., 2002. Fish spoilage bacteria B problems and solutions. Current
Opinion in Biotechnology, 13: 262-266.
Grau C., Sanchez D., Zerpa A., Vallenilla O. & Berti O., 2003. Study of microflora
associated with histamine formation in sardines (Sardinella aurita). Revista Cientifica-
Facultad de Ciencias Veterinarias, 13: 199-204.
Guillén-Sans R. & Guzmàn-Chozas G., 1998- The thiobarbituric acid (TBA) reaction in
foods: A review. Critical Reviews in Food science and Nutrition. 38: 315-330.
H
Haard N. F., 1992. Biochemical reactions in fish muscle during frozen storage. In: Bligh
E.G. (Ed.), Seafood Science and Technology. Fishing News Books, Oxford. p. 176-209.
Halász A., Barath A., Simon-Sakardi L. & Holzapfel W., 1994. Biogenic amines and their
production by microorganisms in food. Trends in Food Science & Technology, 5: 42-48.
Hermes-Lima M., Willmore W.G. &Storey K.B., 1995- Quantification of lipid
peroxidation in tissue extracts based on Fe(III) xylenol orange complex formation. Free
Radical Biology & Medicine. 19: 271-280.

Références bibliographiques
84
Hidalgo J., Zamora R. & Giron J., 1992. Grasas Acietes. 28: 43-97/100. In. Aubourg S.P.,
1998a.
Horrocks LA, Yeo YK, 1999. Health benefits of docosahexaenoic acid (ADH). Pharmacol
Res September;40(3):211-25.
Hughes, R.B. & Jones, N.R., 1966 - Measurement of hypoxantine concentration in canned
herring as an index of the freshness of the raw material, with a comment on the flavor
relations. J. Sea. Food Agree.17: 434-436.
Hurley M.J., Larsen L.B., Kelly A.L. & Mc Sweeney P.L.H., 2000. Cathepsin D activity in
Quarg. Int. Dairy J., 10 : 453-8.
Huss HH (1988). Fresh fish quality and quality changes. FAO Fisheries Series, No. 29, FAO,
Rome.
Hwang B.-S., Wang J.-T., Choong Y.-M., 2003. A rapid gas chromatographic method for
the determination of histamine in fish and fish products. Food Chemistry. 82, pp. 329–334.
Hwang, K. & Regeinstein, J., 1993. Characteristics of mackerel mince lipid hydrolysis.
Journal of Food Science. 58: 79-83.
Hwang, K., & Regenstein, J.,1995. Hydrolysis and oxidation of mackerel (Scomber
scombrus) mince lipids with NaCl treatments. Journal of Aquatic Food Production and
Technology. 4, 19-30.
I
Ingemansson T., Olsson N.U., Herslöf B.G., Ekstrand B., 1991- J. Sci. Food Agric. 57:
443-447.
J
Jiang S.T., 2000. Effect of proteinases on the meat texture and seafood quality. Food Science
and Agricultural Chemistry, 2 : 55-74.
Joosten, H.M.L.J., Northold, M.D., 1989. Detection, growth and amine-producing capacity
of lactobacilli in cheese. Appl. Environ. Microbiol. 55 (9), 2356–2359.
Jorgensen L., Huss H. & Dalgard P., 2000. The effect of biogenic amine production by
single bacteria cultures and metabolisis on cold-smoked salmon. Journal of Applied
Microbiology, 56: 920-934.
Journal Officiel de la République Algérienne, 2006. Arrêté du 12 Joumada Ethania 1427
correspondant au 8 juillet 2006 rendant obligatoire la méthode de détermination de la teneur
en histamine dans les produits de la pêche par chromatographie liquide haute performance.
J.O.R.A. N°58. p. 19-20.

Références bibliographiques
85
K
Kamal-Eldin A. & Yanishlieva N.V., 2002. N-3 fatty acids for human nutrition: stability
considerations. European Journal of Lipid Sciences and Technology. 104: 825-836.
Kawasaki T, Seki E, Osajima K et al., 2000. Antihypertensive effect of valyl-tyrosine, a
short chain peptide derived from sardine muscle hydrolyzate, on mild hypertensive subjects. J
Hum Hypertens August.14(8):519-23.
Ke P.J., Ackman R.G., Linke B.A. & Nash D.M., 1977. Differential lipid oxidation in
various parts of frozen mackerel. Journal of Food Technology. 12: 37-47.
Kim, C., Hearnsberger, J., & Eun, J., 1995. Gram-negative bacteria in refrigerated catfish
fillets treated with lactic culture and lactic acid. Journal of Food Protection.58, 639-643.
Kołakowski, E., & Bednarczyk, B., 2003. Changes in headed and gutted Baltic herring
immersed salting in brine with the addition of acetic acid Part 2. Intensity of proteolysis.
Electronic Journal of Polish Agricultural Universities.
Kołakowski E., 2005. Analysis of proteins, peptides, and amino acids in foods. In S. Otles_
(Ed.), Methods of analysis of food components and additives. p. 59–96.
Kondjoyan N., & Berdagué J. L., 1996. A compilation of relative retention indices for
analysis of aromatic compounds. In I:Laboratoire Flaveur (Ed.). Clermont-Ferrand.
Koutsoumanis K.P., Sofos J.N., 2004. Comparative acid stress response of Listeria
monocytogenes, Escherichia coli O157:H7 and Salmonella typhimurium after habituation at
different pH conditions. Letters in Applied Microbiology. 38, pp. 321–326.
Kraus, L. (1992). Refrigerated sea water treatment of herring and mackerel for human
consumption. In J. Burt, R. Hardy, & K. Whittle (Eds.), Pelagic fish. The resource and its
exploitation (pp. 73e81). Aberdeen, Scotland, UK: Fishing News Books.
Krauss RM, Eckel RH, Howard B et al., 2000. AHA Dietary Guidelines: revision 2000: A
statement for healthcare professionals from the Nutrition Committee of the American Heart
Association. Stroke November; 31(11):2751-66.
Kung, H. F., Tsai Y. H., & Wei C. I., 2007. Histamine and other biogenic amines and
histamine-forming bacteria in miso products. Food Chemistry. 101, 351–356.
Kung H. F., Wang T. Y., Huang Y. R., Lin C. S., Wu W. S., LinC. M. et al., 2009.
Isolation and identification of histamine-forming bacteria in tuna sandwiches. Food Control.
20, 1013–1017.

Références bibliographiques
86
J
Jorgensen L., Huss H. & Dalgard P., 2000. The effect of biogenic amine production by
single bacteria cultures and metabolisis on cold-smoked salmon. Journal of Applied
Microbiology. 56: 920-934.
L
Laguerre M., Lopez-Giraldo L.J., Lecomte J., Pina M. & Villeneuve P., 2007. Outils
d’évaluation in vitro de la capacité antioxydante. Oléagineux, Corps gras, Lipides, 14(5):
278-292.
Larsson SC, Kumlin M, Ingelman-Sundberg M, Wolk A, 2004. Dietary long-chain n-3
fatty acids for the prevention of cancer: a review of potential mechanisms. Am J Clin Nutr
June.79(6):935-45.
Lavigne C, Marette A, Jacques H, 2000. Cod and soy proteins compared with casein
improve glucose tolerance and insulin sensitivity in rats. Am J Physiol Endocrinol Metab
March.278(3):E491-E500.
Lavigne C, Tremblay F, Asselin G, Jacques H, Marette A, 2001. Prevention of skeletal
muscle insulin resistance by dietary cod protein in high fat-fed rats. Am J Physiol Endocrinol
Metab July.281(1):E62-E71.
Liston J., Samadpour, Ongerth J.E., Tran N., Nguyen D., Wittam T.S., 1994.Occurrence
of shiga-like toxinproducing Escherichia coli in retail fresh seafood, beef, lamb, pork and
poultry from grocery stores in Seattle, Washington. Appl Environ Microbiol. 60: 1038-1040.
Lopez-Sabater E.I., JJ. Rodriguez- Jerez, A.X. Roig-Sagues, M.A.T. Mora- Ventura,
1994. Bacteriological quality of tunafish (Thunnus) destinedfor canning : effect oftuna
handling on présence of histidine decarboxylase bacteria and histamine level. J. Food-Prot.
57 (4 ) 318-323.
Love R.M., 1980 -The Chemical Biology of Fishes. Vol.2. Academic Press London. 1968-
1977.
M
Macciola V, 2004. A study on the lipid fraction of Adriatic sardine filets (Sardina
pilchardus). Nahrung June.48(3):209-12.
Mahmoud, S & Benkakaa, K., 2007. Effet du mode de conservation sur l’oxydation des
lipides ; concentration en malodialdéhyde (TBARS) chez Sardina pilchardus (Walbaum,
1792) et Merluccius merluccius (Linnaeus, 1758). Mémoire d’ingéniorat. Univ. D’Oran, Es
Sénia. 29p.

Références bibliographiques
87
Maijala R.L., Eerola S.H., Aho M.A. & Hirn J.A., 1993. The effect of GDL-induced pH
decrease on the formation of biogenic amines in meat. Journal of Food Protection, 50: 125-
129.
Malle P., 2006. Histamine. Fiche de description de danger transmissible par les aliments.
Fiche AFSSA. pp. 1-5.
Malle P., Valle M. and Bouquelet S., 1996. Assay of biogenic amines involved in fish
decomposition. Journal of AOAC International. 79 : 43-49.
Marklinder I. & Lönner C., 1992. Fermentation properties of intestinal strains of
Lactobacillus, of a sour dough and of a yoghurt starter culture in an oat-based nutritive
solution. Food Microbiology, 9: 197-205.
Martin A., 2001. Apports nutritionnels conseillés pour la population française. Editions Tec
& Doc, Paris. 650 p.
Mazorra-Manzano M.A., PachecoAguilar R., Diaz-Rojas E.I., & Lugo-Sanchez H.E.,
2000. Post mortem changes in black skipjack muscle during storage in ice. J. Food Sci. 65:
774-779.
Melton S.L., 1983. Methodology for following lipid oxidation in muscle foods. Food
Technol., 37 : 105-111,116.
Mendes R., 1999. Changes in biogenic amines of major Portuguese bluefish species during
storage at different temperatures. Journal of Food Biochemistry. 23: 33-43.
Mendes R., 2009. Chapter 3: Biogenic amines. In: Rehbein H. & Oehlenschläger J. (Eds.),
Fishery products: quality, safety and authenticity. Wiley-Blackwell publication. p. 42-67.
Meyer V., 1965. Marinades. In G. Borgstrom (Ed.), Fish as Food (Vol. III, part 1) (p. 165–
193). New York: Academic Press.
Mori TA, Beilin LJ, 2004. Omega-3 fatty acids and inflammation. Curr Atheroscler Rep
November; 6(6):461-7.
Morris MC, Evans DA, Bienias JL et al., 2003. Consumption of fish and n-3 fatty acids and
risk of incident Alzheimer disease. Arch Neurol July;60(7):940-6.
Murata M, Sano Y, Bannai S, Ishihara K, Matsushima R, Uchida M, 2004. Fish protein
stimulated the fibrinolysis in rats. Ann Nutr Metab September;48(5):348-56.
Murray & Burt, 1969. The composition of fish. J. Fish Biol. 9: 329-334.
Murray , Burt & Shewan , 2000. The microbial spoilage of fish with special reference to
the role of psychrotrophs. In: Russell, A.D. and R. Fuller (eds.) Cold tolerant microbes in
spoilage and the environment, Academic Press, 117-136.

Références bibliographiques
88
N
Naganawa, H., & Ochi, K., 2002. Bacilysocin, a novel phospholipid antibiotic produced by
Bacillus subtilis 168. Antimicrob. Agents Chemother. 46: 315-20.
Ness AR, Gallacher JE, Bennett PD et al., 2003. Advice to eat fish and mood: a randomised
controlled trial in men with angina. Nutr Neurosci February;6(1):63-5.
Ngah E., Génot C., Meynier A., Viau M. & Gandemer G. 1993. Phospholipid oxidation in
Turkey precooked meat. 39th
Int Congress Meat Sci Technol, Proceeding on disks, 5, August
1-8, Calgary, Canada.
Niven, C. F., Jeffreg, M. B., & Corlett, D. A., 1981. Differential plating medium
forquantitative detection of histamine-producing bacteria. Applied and Environmental
Microbiology, 41, 321–322.
O
Official Methods of Analysis of AOAC international (AOAC 16th ed.), Method 977.13
(1995). Histamine in seafood, Fluorimetric method. Sec.35.1.32. 6-17. P.A. Cunniff Edition,
AOAC International Gaithersburg, MD.
Okuzumi M., Hiraishi A., Kobayashi T., & Fujii T., 1994. Photobacterium histaminum
sp. nov., a histamine-producing marine bacterium. International Journal of Systemic
Bacteriology. 44, 631–636.
Okuzumi M., Okuda S. & Awano M., 1981. Isolation of psychrophilic and halophilic
histamine-forming bacteria from Scomber japonicus. Bulletin of the Japanese Society of
Scientific Fisheries, 47(12): 1591-1598.
Olafsdottir G., & Fleurence J., 1997. Evaluation of fish freshness using volatile compounds-
classification of volatile compounds in fish. Methods to determine the freshness of fish in
research and industry In Proceedings of the final meeting of the concerted action Evaluation
of fish freshness AIR3CT94 2283, Nantes, France.
Olson J.C. and Mocquot G., 1980. Milkand milkproduct . In:International Commission on
Microbiological Specification for Foods (Ed.), Microbial Ecology of Foods: Food
Commodities,Vol. 2. Academic Press, New York, pp. 470–490.
Özogul F., Polat A. & Özogul Y., 2004. The effects of modified atmosphere packaging and
vacuum packaging on chemical, sensory and microbiological changes of sardines (Sardina
pilchardus). Food Chemistry, 85(1): 49-57.

Références bibliographiques
89
Özogul F. & Özogul Y., 2006. Biogenic amine content and biogenic amine quality indices of
sardines (Sardina pilchardus) stored in modified atmosphere packaging and vacuum
packaging. Food Chemistry, 99: 574-578.
Özyurt G., Kuley E., Ozkutuk S. & Özogul F., 2009. Sensory, microbiological and
chemical assessment of the freshness of red mullet (Mullus barbatus) and goldband goatfish
(Upeneus moluccensis) during storage in ice. Food Chemistry, 114: 505-510.
Ozyurt G., Ozogul Y., Ozogul F., Kuley E. & Polat A., 2005. Freshness assessment of
European eel (Anguilla Anguilla) by sensory, chemical and microbiological methods. Food
Chemistry. 92: 745-751.
Ozyurt, G.n Polat, A. & Tokur, B., 2007. Chemical and sensory changes in frozen ()18 _C)
wild sea bass (Dicentrarchus labrax) captured at different fishing seasons. International
Journal of Food Science and Technology. 42: 887–893.
P
Parente E., Matuscelli M., Gadrini F., Grieco S., Crudele M.A., Suzzi G., 2001. Evolution
of microbial populations and biogenic amines production in dry sausages produced in
southern Italy Journal of Applied Microbiology, 90 (2001), p. 882–891.
Patange S.B., Mukundan M.K. & Ashok Kumar K., 2005. A simple and rapid method for
colorimetric determination of histamine in fish flesh. Food Control, 16(5): 465-472.
Pattison DJ, Harrison RA, Symmons DP, 2004. The role of diet in susceptibility to
rheumatoid arthritis: a systematic review. J Rheumatol July;31(7):1310-9.
Pedersen M, Stripp C, Klarlund M, Olsen SF, Tjonneland AM, Frisch M, 2005. Diet and
risk of rheumatoid arthritis in a prospective cohort. J Rheumatol July;32(7):1249-52.
Penfield M.P. & Campbell A.M., 1990. Fats and their lipids constituents. Chemical
reactions of lipids eds. Experimental food science, 3e ed. San Diego: Academic Press, p 3407.
Phillips B., Wang B., Pace R.D., Dessai A.P., Bovell-Benjamin A. &, 2002. Modified
extraction method for determining 2-thiobarbituric acid values in meat with increased
specificity and simplicity. Journal of Food Science, 67: 2833-2836.
Pigott G. & Tucker B., 1987. Science opens new horizons for marine lipids in human
nutrition. Food Rev. Int. 3: 105-138.
Poulter, N.H. & Nicolaides L., 1985b. Studies of the iced storage characteristics and
composition of a variety of Bolivian freshwater fish. 2. Parana and Amazon Basins fish. J.
Food Technol. 20: 451-465.
Prescott, Harley & Klein (1995). Microbiologie.DeBoeck Université, 1014 p.

Références bibliographiques
90
R
Rachidi H., 1998. Mise au point d'un système de cotation pour l'appréciation sensorielle de la
fraîcheur des produits de la pêche. Mémoire de fin d'étude halieutique. Inst. Agro. Vet. Hassan
II. Rabat, Maroc.
Rodrıguez A., Carriles N., Cruz J.M. & Aubourg S.P., 2008. Changes in the flesh of
cooked farmed salmon (Oncorhynchus kisutch) with previous storage in slurry ice (_1.5 _C).
LWT - Food Science and Technology. 41: 1726-1732.
Rodrıguez, O., Barros-Velazquez, J., Ojea, A., Pineiro, C., Gallardo, J. M., & Aubourg,
S., 2003. Effect of chilled storage in flow ice on the microbial quality and shelf life of farmed
turbot (P. maxima), isolation and identification of major proteolytic bacteria. In: Proceedings
first joint trans-atlantic fisheries technology conference, TAFT 2003. 33rd WEFTA and 48th
AFTC meetings, 11–14 June 2003, Reykjavik, Iceland (p. 73–74).
Rossell, J. B., 1989. Measurement of rancidity. In J. C. Allen & R. J. Hamilton (Eds.),
Rancidity in foods (p. 23–52). New York: Elsevier.
Rouabhi I.F., 2009. Effet du mode de conservation sur la qualité sensorielle et biochimique
des poissons : la sardine commune (Sardina pilchardus), le rouget de roche (Mullus
surmuletus) et le merlan bleu (Micromesistius poutassou). Mémoire de magister. Gestion des
ressources aquatique et control qualité. Faculté des sciences. Université d’Oran Es-Sénia,
78p.
S
Saeed S. & Howell N. K., 2001. 12-Lipoxygenase activity in the muscle tissue of atlantic
mackerel (scomber scombrus) and its prevention by antioxidants. Journal of the science of
Food and Agriculture. 81 :745-750.
Saeed S. & Howell N., 2002. Effect of lipid oxidation and frozen storage on the muscle
proteins of Atlantic mackerel (Scomber scombrus). J. Scie. Food.Agric. 82: 579-586.
Salih A.M., Smith D.M., Price J.F. & Dawson L.E. , 1987. Modified extraction 2
thiobarbituric acid method for measuring lipid oxidation in poutry. Poultry science. 66 :1483-
1488.
Santos S. & Regenstein J., 1990. Effects of vacuum packaging, glazing and erythorbic acid
on the shelf-life of frozen white hake and mackerel. J. Food Sci. 55: 64-70.
Sarma J, Reddy G.V.S. & Srikar L.N., 2000. Effect of frozen storage on lipids and
functional properties of proteins Indian oil sardine (Sardinella longiceps). Food Research
International, 33: 815-820.

Références bibliographiques
91
Sato T., Fujii T., Masuda T. & Okuzumi M., 1994. Changes in numbers of
histaminemetabolic bacteria and histamine content during storage of common mackerel.
Fisheries Science, 60(3): 299-302.
Sato T., Okuzumi M. & Fujii T., 1995. Evaluation of polyamines of Common Mackerel
during storage as indicators of decomposition. Journal of the Food Hygienic Society of Japan,
36(6): 743-747.
Shewan J.M., 1977. The bacteriology of fresh and spoiling fish and the biochemical changes
induced by bacterial action. In: Sutcliffe P. & Disney J. (Eds.), Proceedings of the Conference
on Handling, Processing and Marketing of Tropical Fish. Tropical Products Institute, London.
pp. 51-66.
Shewfelt, R.L. 1981- Fish muscle lipolysis-A review. Journal of food Biochemistry. 5: 79-
100.
Shimizu Y., Toyohara H. & Lanier T.C., 1992. Surimi production from fatty and
darkfleshed fish species. In: Lanier T.C. & Lee C.M. (Eds), Surimi Technology. Marcel
Dekker Inc, New York. p. 181-207.
Siebert G., 1961. Enzymes of marine fish muscle and their role in fish spoilage. In : Fish in
nutrition, Fishing News Books Ltd, 80 - 82.
Sierra, G. 1957. Antonie van Leeuwenhoek 23, 15.
Sikorski Z., & Kolakowska A., 1994. Changes in protein in frozen stored fish. In Z. Sikorski,
B. Sun Pan, & F. Shahidi (Eds.), Seafood proteins (p. 99e112). New York, USA: Chapman
and Hall.
Silla M. H., 1996. Biogenic amines: their importance in foods. International Journal of Food
Microbiology, 29, 213-231.
T
Taylor S.L., 1991. Histamine food poisoning: toxicology and clinical aspects. Critical
Reviews in Toxicology. 17 : 91-128.
U
Uchida K., Haraguchi K., Mitsui M. & Kawakishi S., 1990. Stimulatory effect of
histamine on the peroxidation of linoleic acid. Journal of Agricultural and Food Chemistry,
38: 1491-1493.

Références bibliographiques
92
Undeland B., Ekstrand H. & Lingnert I., 1998. Lipid oxidation in minced herring (Clupea
harengus) during frozen storage. Effect of washing and precooking. J. Agric. Food Chem. 46:
2319-2338.
Undeland I., Kelleher S.D., Hultin H.O., McClement J., & Thongraung,
2003.Consistency and solubility changes in herring (Clupea harengus) light muscle
homogenates as a function of pH. Journal of Agricultural and Food Chemistry.51,
3992–3998.
Undeland I., Hultin H.O. & Richards M.P., 2002. An aqueous fraction of cod muscle
inhibits hemoglobin-mediated oxidation of cod muscle membrane lipids. 32nd Annual
WEFTA Meeting, May 13th-15th, Galway (Ireland) 90.
Uriarte-Montoya M.H., Villalba-Villalba A.G., Pacheco-Aguilar * R., Ramirez-Suarez J.
C., Lugo-Sánchez M. E., García-Sánchez G., Carvallo-Ruíz M. G., 2010. Changes in
quality parameters of Monterey sardine (Sardinops sagax caerulea) muscle during the
canning process. Food Chemistry, 2010, Vol. 122 (3), p. 482-487.
V
Valle M., Malle P. and Bouquelet S., 2004. Optimization of a Liquid Chromatographic
Method for Determination of Amines in Fish. Journal of AOAC International. 80 : 49-56.
Verma J., Srikar L, Sudhakara N. & Sarma J., 1995. Effects of frozen storage on lipid
freshness parameters and some functional properties of oil sardine (Sardina pilchardus)
mince. Food Research International. 28:87-90.
Vyncke W., 1970. Direct determination of the thiobarbituric acid value in thrichloroacetic
acid extracts of fish as a measure of oxidative rancidity. Fette Seifen Anstrichm. 72: 1084-
1096.
Vyncke W., 1996. Comparison of the official EC method for the determination of total
volatile bases in fish with routinemethods. Archiv for Lebensmittel hygiene, 47:110–112
W
Wang B., Pace R.D., Dessai A.P., Bovell-Benjamin A. & Phillips B., 2002. Modified
extraction method for determining 2-thiobarbituric acid values in meat with increased
specificity and simplicity. Journal of Food Science, 67: 2833-2836.
Wang J., Miyazawa T. & Fujimoto K., 1991. Inhibition of methyl linoleate peroxidation by
maize zein in powder model system at high water activity. Agricultural Biology and
Chemistry, 55: 1531-1536.

Références bibliographiques
93
Watanabe T., Aoki H., Shimoto K., Hadzuma M., Maita M., Yamagata Y., KironV.,
Satoh S., 1999. A trial to culture yellowtail with non-fishmeal diets. Fisheries Science, 64,
505-512.
Watanabe T., Verakunpiriya V., Watanabe K., Viswanath K., Satoh S., 1997. Feeding of
rainbow trout with non-fish meal diets. Fisheries Science, 63,258-266.
X
Xiong S., Xiong Y. L., Blanchard S. P., Wang B., & d Tidwell J. H., 2002. Evaluation
of tenderness in prawns (Machrobrachium rosenbergii) marinated in various salt and acid
solutions. International Journal of Food Science and Technology. 37, 291–296.


ANNEXES

Annexe
94
8. ANNEXES
Annexe I : Composition des milieux de culture en microbiologie
La composition donnée est pour un volume final d'un litre et l’autoclavage se fait à 121°C
pendant 20 minutes:
Milieu PCA
Peptone 5,0 g
Extrait de levure 2,5 g
Glucose 1,0 g
Agar 15,0 g
pH = 7
Milieu BCPL
Peptone 5,0 g
Extrait de viande de bœuf 3,0 g
Lactose 10,0 g
Pourpre de bromocrésol 25 mg
pH = 6,8
Milieu Schubert
Tryptone 12 g
Tryptophane 1 g
Mannitol 9 g
Bile de boeuf déshydratée 25 g
Chlorure de sodium 5 g
Phosphate monopotassique 1,5 g
Phosphate di-potassique 3,5 g
Lauryl-sulfate de sodium 0,2 g
pH = 7

Annexe
95
Milieu Rothe S/C
Peptone 20,0 g
Glucose 5,0 g
Azide 0,2 g
NaCl 5,0 g
Hydrogénophosphate de potassium 2,7 g
Dihydrogénophosphate de potassium 2,7 g
pH = 6,8
Milieu Eva- Litzky
Peptone 20,0 g
Glucose 5,0 g
Azide 0,2 g
Ethyl-violet 0,5 g
NaCl 5,0 g
Hydrogénophosphate de potassium 2,7 g
Dihydrogénophosphate de potassium 2,7 g
pH= 6,8
Milieu gélose viande-foie
base viande foie 30,0 g
glucose 2,0 g
agar 6,0 g
pH = 7,4
http://fr.wikipedia.org/w/index.php?title=Hydrog%C3%A9nophosphate_de_potassium&action=edit&redlink=1

Annexe
96
Milieu Chapman
Peptone 10,0 g
Extrait de viande de bœuf 1,0 g
Chlorure de sodium 75,0 g
Mannitol 10,0 g
Rouge de phénol 0,025 g
Agar-Agar 15,0 g
pH = 7,4
Milieu bouillon sélinite
Tryptone 5g
Lactose 4g
Sélénite 4g
Hydrogénosélénite de sodium 4,0g
pH=7
Milieu Salmonelle-Shigelle
Peptone 5,0 g
Extrait de viande 5,0 g
Lactose 10,0 g
Citrate de sodium 10,0 g
Citrate de fer III 1,0 g
Sels biliaires 8,5 g
Vert brillant 3,3 mg
Rouge neuter 25 mg
Thiosulfate de sodium 8,5 g
Agar 12,0 g
pH = 7,3 (il faut dissoudre ce milieu par ébullition et ne pas l’autoclaver)

Annexe
97
Milieu King A
Peptone dite 20,0 g
Glycérol 10,0 g
Sulfate de potassium 10,0 g
Chlorure de magnésium 1,4 g
Agar purifié 12,0 g
pH = 7,2
Milieu King B
Peptone 20,0 g
Glycérol 10,0 g
Hydrogénophosphate de potassium 1,5 g
Sulfate de magnésium heptahydraté 1,5 g
Agar 12,0 g
pH = 7,2
Gélose nutritive
Extrait de viande 1,0g
Extrait de levure 2.5g
Peptone 5,0g
Chlorure de sodium 5,0g
Agar 15,0g
pH = 7,0

Annexe
98
Milieu sélectif pour les bactéries aminogènes
Tryptone 5 g
Extrait de levure 5 g
Extrait de viande 5 g
Chlorure de sodium 2,5 g
Glucose 5 g
Tween 80 1g
MgSO4 0,2 g
MnSO4 0,05 g
FeSO4 0,04 g
Citrate d’ammonium 2 g
Thiamine 0,01g
K2PO4 2 g
CaCO3 0,1 g
Pyridoxal phosphate 0,05 g
Acide aminé 10 g
Pourpre de crésol 0,06 g
Agar 20 g
pH=5,3
Réactif de Frazier
Chlorure de sodium 67,0g
Qsp d’eau 100 ml

Annexe
99
Annexe II
Barème de cotation de la fraicheur selon directives du codex alimentarius pour l’évaluation
organoleptique au laboratoire du poisson, mollusques et crustacés (CAC/GL 31-1999)
E A B C
Peau
Pigmentation
vive et
chatoyante (sauf
pour les
sébastes) ou
opalescente, pas
de décoloration
Pigmentation
vive mais sans
lustre ; très
légère
décoloration
Pigmentation
terne en voie de
décoloration
Pigmentation
terne décolorée
grisâtre
Mucus cutané
Aqueux,
transparent Laiteux
Gris jaunâtre et
un peu
grumeleux
Brun jaunâtre
très grumeleux
et épais
Œil
Convexe
(bombé)
Pupille noire
Brillante,
transparente
Pupille
légèrement
opaque, cornée
opalescente
Légèrement
concave, pupille
grise cornée
opaque
Très concave
pupille grise
décoloré,
opaque
Branchies
Rouge foncé ou
rouge brillante
mucus
transparent
Rouge au rose :
mucus
légèrement
opaque
Brun/ gris se
décolorant :
mucus opaque et
épais
Brun ou
décoloré :
mucus jaunâtre
et grumeleux
Péritoines
Lissé, brillant et
difficile à
détacher de la
chair
Un peu terni,
difficile à
détacher de la
chair
Grumeleux se
détachant assez
facilement de la
chair
Grumeleux se
détachant très
facilement de la
chair
odeur
Fraiche, d’algue
marine de fruit
de mer
Absence
d’odeur : neutre
ou légèrement
douceâtre
Fermenté :
légèrement aigre
Aigre,
ammoniaque,
putride

Annexe
100
Annexe III
Exemple de formulaire d’évaluation :
Paramètre de qualité Caractéristiques Classement
Peau Aspect de surface
0
1
2
3
Brillant luisant
Cireux, perte d’éclat
Terne, décoloration
Terne, râpeux
Mucus
0
1
2
3
Transparent
Laiteux
Gris jaunâtre
Brun jaunâtre
Rigidité 0
1 Ferme
Mou
Chair Rigidité 0
1 Rigor
Post-rigor
Yeux Clarté
0
1
2
Cornée translucide
Cornée opalescente
Cornée opaque
Forme de la pupille
0
1
2
Convexe
Plane
Concave
Branchies Couleur
0
1
2
Rouge vif
Rose
Pâle / décoloré
Odeur
0
1
2
3
Frais / algues
Douteux
Pas frais
Gâté
Mucus
0
1
2
Absent
Modéré
Excessif
Couleur de la chair Surfaces découverte
0
1
2
Translucide
Gris
Brun-jaune
Sang Dans la coupe de la
gorge
0
1
2
Rouge
Rouge foncé
Brun
Total des notes

Annexe
101
Annexe IV
Méthodes de dosage des amines biogènes dans la chair de poisson
Méthode Principe de la
méthode
Seuil de
détection
Avantages Inconvénients
Méthodes
biologiques :
- Contraction de
l’iléon de
cobaye
- Injection au
cobaye
-Test Daphnies
Mise en évidence
d’effet physiologique
de l’histamine sur un
organe ou un
organisme vivant.
Simplicité et rapidité de
la réalisation
Faible coût
Résultats semi-quantitatif
Sensibilité variable
Chromatographi
e sur couche
mince
Extraction de l’amine
et migration sur phase
solide
Visualisation par
méthode
colorimétrique
2 à 5 mg
d’histamine
pour 100g
d’échantillon
selon les
méthodes
Simplicité et rapidité
couramment utilisée en
dosage de routine
Résultat semi-quantitatif
ou quantitatif si couplage à
une autre méthode
Spécificité passable
Chromatographi
e par colonne
échangeuse
d’ions
Extraction de l’amine
par échaude d’ions
Visualisation et dosage
après réaction
colorimétrique
De l’ordre
du mg pour
100g
Simplicité et rapidité
Spécificité meilleure que
lors de l’utilisation de la
CCM
Utilisable en dosage
d’expertise
Dosage d’une seule amine
seulement
Méthode
enzymatiqque
Oxydation de
l’histamine par DAO
Dosage colorimétrique
ou volumétrique
1 -2 mg
/100g
Simplicité, rapidité,
faible coût, bonne
sensibilité et spécificité
et utilisable pour des
contrôles de routine
Fluorimétrie Couplage de l’amine à
un réactif donnant un
composé fluorescent
associé à une méthode
de chromatographie
Dépend de la
méthode de
chromatogra
phie
employée
Simplicité, spécificité
variable (dépend du
réactif fluorogène utilisé)
Nécessité d’un
appareillage sophistiqué
Chromatographi
e en phase
liquide
Migration des amines
sur phase mobile avant
ou après la formation
de composés
fluorescents
<1mg/ 1 kg
soit 1 ppm
Simplicité, sensibilité,
spécificité, plusieurs
amines peuvent être
séparées et dosées en
même temps, utilisable
Nécessité un appareillage
sophistiqué

Annexe
102
lors d’expertise
Chromatographi
e en phase
gazeuse
Conversion de l’amine
en composés volatil
Méthode rarement
employée
Méthodes radio-
enzymatique
Détection d’un
composé radio-actif
après alkylation
enzymatique de
l’amine
De l’ordre
de 1ng/ml
Dosage avec une grande
précision
Emploi de radio-isotope
Méthode
immunologique
Fixation de l’amine à
un anticorps
monoclonal
Dosage immuno-
enzymatique ou radio-
immunologique
De 0.1 à
2ng/ml
d’échantillon
Simplicité et spécifité
Electrophorèse
sur capillaire
Migration de l’amine
entre deux électrodes et
détection dans le
spectre UV
1ppm Simplicité, spécificité et
sensibilité
Nécessite un appareillage
sophistiqué
spectrophotomét
rie
Mesure de la densité
optique après couplage
de l’amine au
diazonium
1mg/100g Spécificité variable et
risque de faux négatifs

Annexe
103
Annexe V
Mise en évidence des réactions substrat / réactif
A : Réaction des hydroperoxydes avec le réactif FOX 2.
B : Formation d’un complexe rose entre le M.D.A et le T.B.A-rs.
C : Réaction des protéines avec le réactif de Brad ford
D : Réaction de l’histamine avec le réactif p-phenyldiazonium sulfonate.
T : Témoin
AQUABIOR, 2011
T C
AQUABIOR, 2011 AQUABIOR, 2011
T T
B A
T C
AQUABIOR, 2011 AQUABIOR, 2011


















