
Protein metabolism, protein and amino acid requirement and ... · Food intake is sensitive to...
41
Protein metabolism, protein and amino acid requirement and body functions Daniel TOME AGROPARISTECH – Departement of Life Sciences and Health INRA, UMR Nutrition physiology and ingestive behavior Paris, France 1 Food Valley Summit Sports & Nutrition, Ede, The Netherlands, 11 October 2018
Transcript of Protein metabolism, protein and amino acid requirement and ... · Food intake is sensitive to...

Protein metabolism, protein and amino
acid requirement and body functions
Daniel TOME
AGROPARISTECH – Departement of Life Sciences and Health INRA, UMR Nutrition physiology and ingestive behavior
Paris, France
1Food Valley Summit Sports & Nutrition, Ede, The
Netherlands, 11 October 2018

Introduction• Dietary proteins provide nitrogen and amino acid (including 9 indispensable
amino acid) to support protein synthesis and related physiological functions
• An adequate protein status is required to balance nitrogen losses and for the maintenance of physiological functions and survival
• Amino acid sufficiency is tightly controlled in the body and influences metabolism and food intake to maintain or restore an adequate protein status
2
62 % water17 % protein (50% Muscle)14 % fat 6 % minerals1 % carbohydrates
Body composition of adult human
Deamination
BodyProtein
Free amino acid pool
Nitrogenlosses
Carbon skeleton metabolism
Oxidation
Protein intake
Synthesis Breakdown

Food intake is sensitive to protein content in the diet
Generous protein intake : Subjects tend to reduce energy intake in relation with an increased fullness and satiety
Adequate protein intake : derived from the quantity of protein intake that balance protein (nitrogen) losses and meets amino acid needs to support body protein synthesis.
Moderate protein deficiency : Subjects try to increase their protein intake to compensate the deficiency, by choosing protein rich food or by increasing total food intake
Severe protein deficiency : induces an aversive response, decreases food intake and body weight (lean and adipose tissue)
% Energy as protein in the diet
25-50%
10-24%
1-2 %
5-9%
3
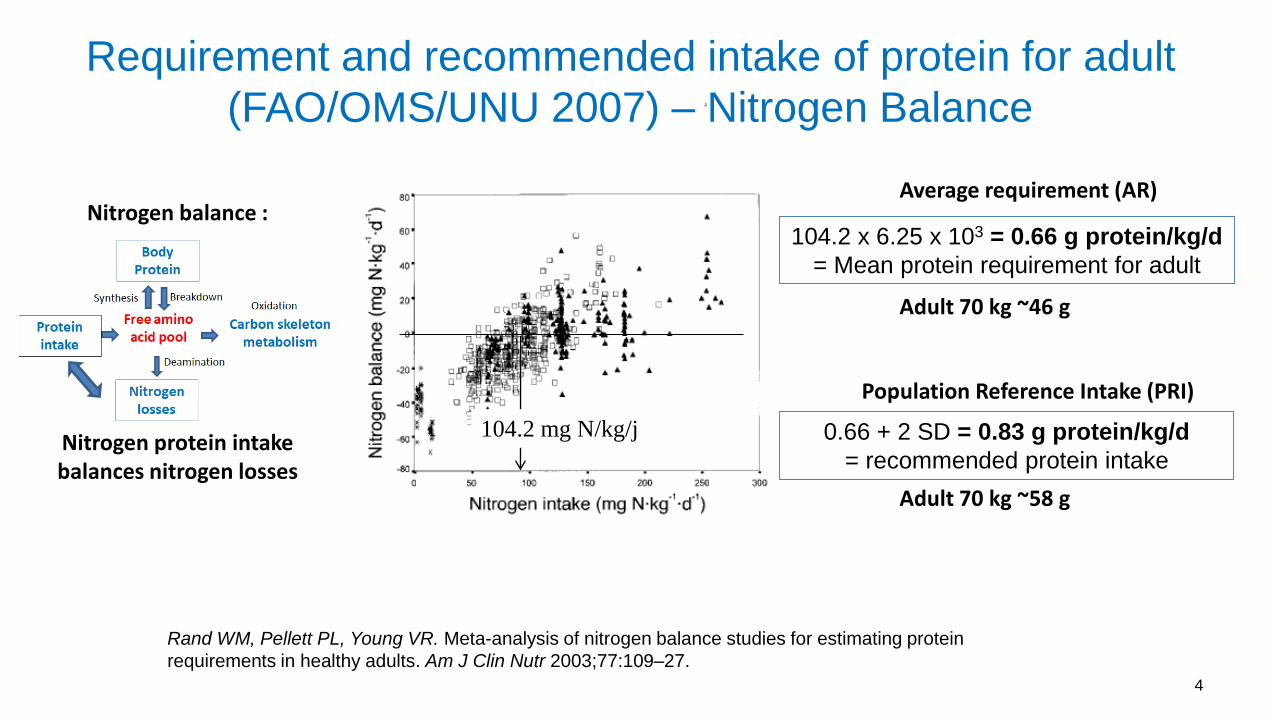
Rand WM, Pellett PL, Young VR. Meta-analysis of nitrogen balance studies for estimating protein
requirements in healthy adults. Am J Clin Nutr 2003;77:109–27.
104.2 mg N/kg/j
104.2 x 6.25 x 103 = 0.66 g protein/kg/d
= Mean protein requirement for adult
0.66 + 2 SD = 0.83 g protein/kg/d
= recommended protein intake
Requirement and recommended intake of protein for adult
(FAO/OMS/UNU 2007) – Nitrogen Balance
4
Adult 70 kg ~46 g
Adult 70 kg ~58 g
Nitrogen balance :
Nitrogen protein intake balances nitrogen losses
Average requirement (AR)
Population Reference Intake (PRI)
5
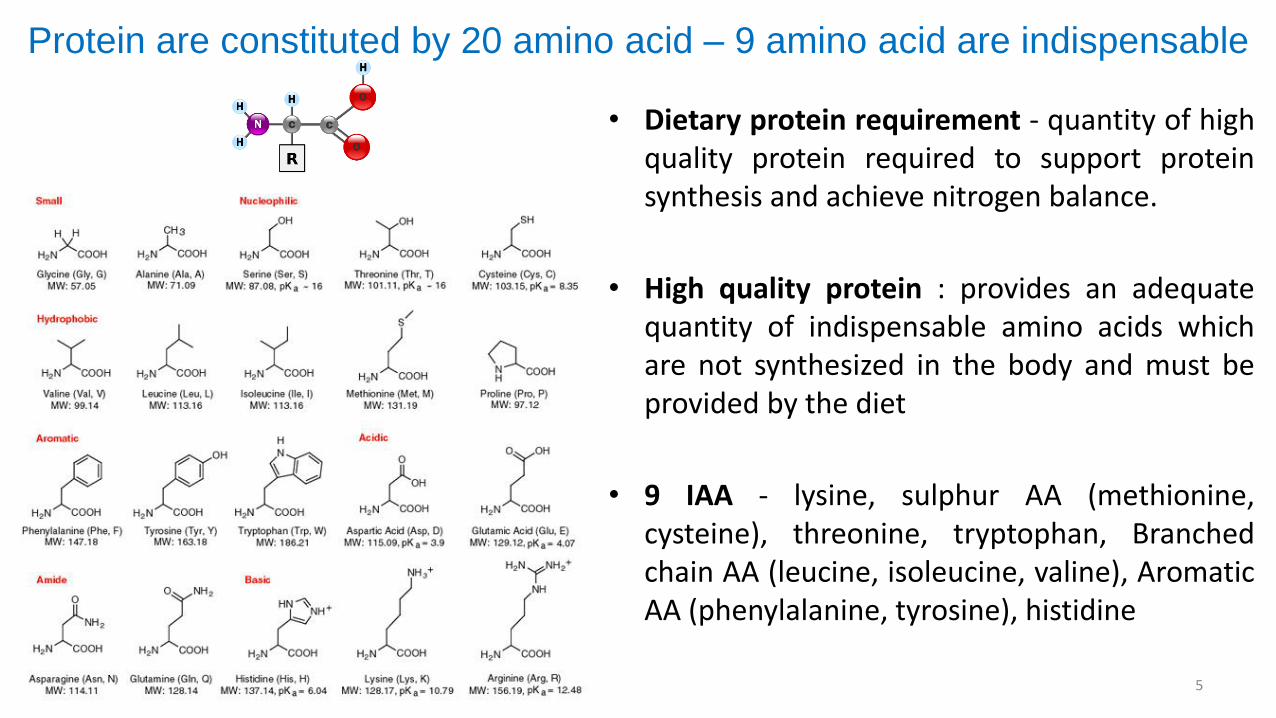
Protein are constituted by 20 amino acid – 9 amino acid are indispensable
• Dietary protein requirement - quantity of highquality protein required to support proteinsynthesis and achieve nitrogen balance.
• High quality protein : provides an adequatequantity of indispensable amino acids whichare not synthesized in the body and must beprovided by the diet
• 9 IAA - lysine, sulphur AA (methionine,cysteine), threonine, tryptophan, Branchedchain AA (leucine, isoleucine, valine), AromaticAA (phenylalanine, tyrosine), histidine
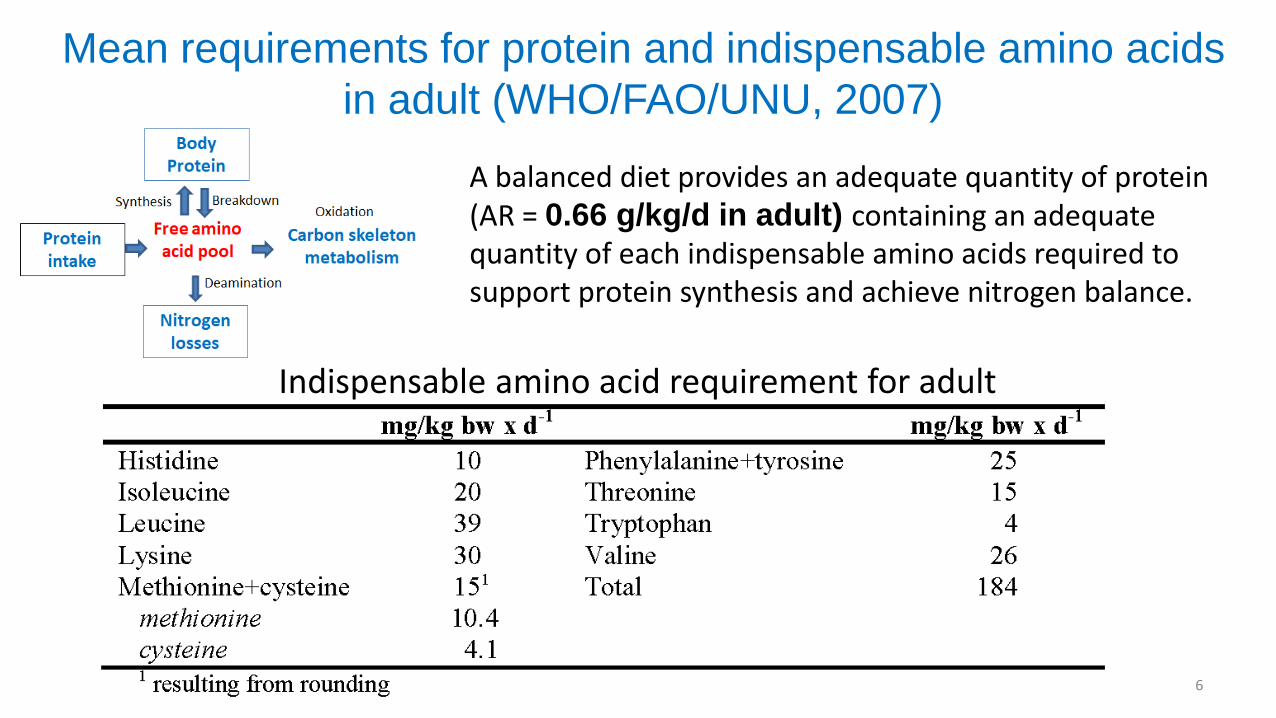
Mean requirements for protein and indispensable amino acids
in adult (WHO/FAO/UNU, 2007)
6
A balanced diet provides an adequate quantity of protein (AR = 0.66 g/kg/d in adult) containing an adequate quantity of each indispensable amino acids required to support protein synthesis and achieve nitrogen balance.
Indispensable amino acid requirement for adult

Recommendation for protein intake
• Protein requirement are derived from nitrogen balance but several
health outcomes associated with protein intake could also be
considered as criteria for setting DRVs for protein.
• Health outcomes associated with protein intake : muscle mass, body
weight, insulin sensitivity, bone health, kidney function, urea cycle,
protein tolerance.
• It is conceivable that in the event of sufficient evidence for a positive
effect on health, a requirement for protein above the value derived
from nitrogen balance would result.
• Potentially adverse effects should be taken into account when setting
DRVs for protein
7(EFSA 2012)

Protein status : protein synthesis and nitrogen losses in rats fed 2% to 20% (w/w) protein diet
Du et al, J. Nutr. 130: 514–521, 2000
Nitrogenlosses
Protein synthesis
An adequate protein intake provides amino acid, particularly indispensable amino acids, to support protein synthesis
Protein deficiency
Generous protein intake
adequate Protein intake
8
… And to balance nitrogen losses
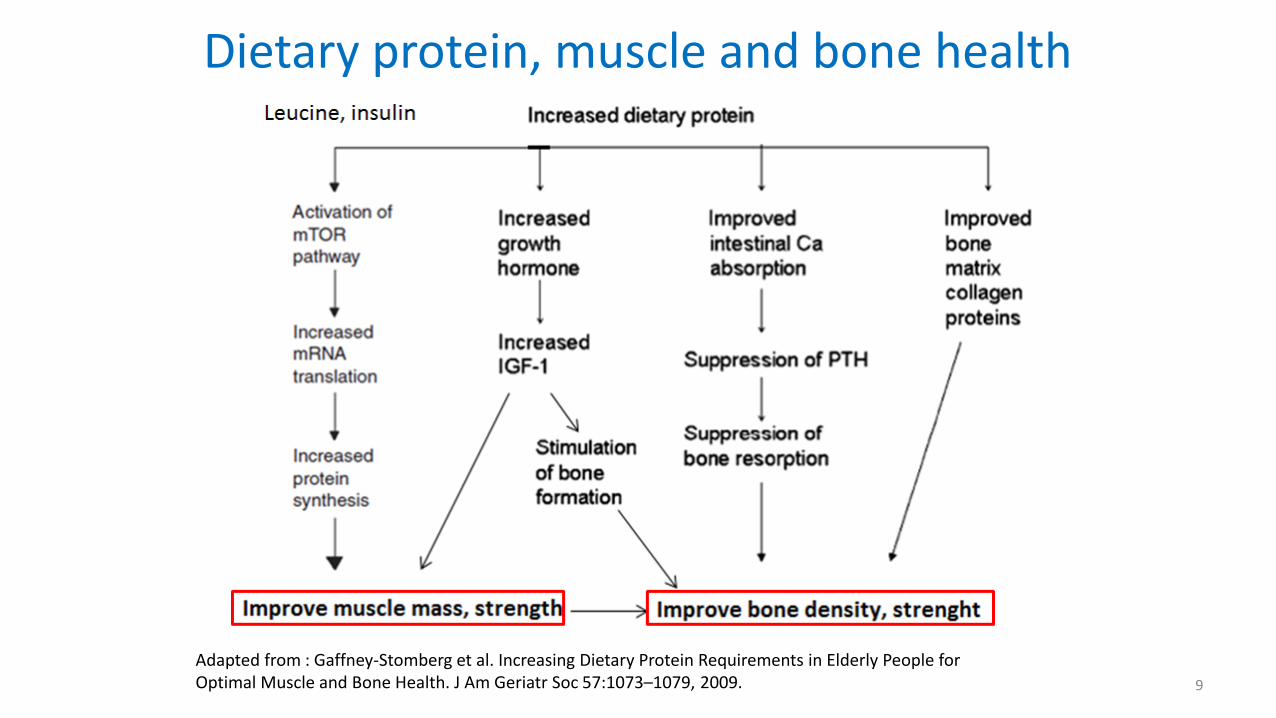
Dietary protein, muscle and bone health
Adapted from : Gaffney-Stomberg et al. Increasing Dietary Protein Requirements in Elderly People for Optimal Muscle and Bone Health. J Am Geriatr Soc 57:1073–1079, 2009. 9

Simplified overview of whole body oral protein utilization at rest
10
Of the protein ingested-derived amino acids :
- Approximately 50% is extracted by splanchnic tissues before entering peripheral circulation.
- About 40% is deaminated and catabolized, producing urea, CO2 and H2O.
- About 10% is utilized for skeletal muscle protein synthesis
Adapted from Stokes et al. Recent Perspectives Regarding the Role of Dietary Protein for the Promotion of
Muscle Hypertrophy with Resistance Exercise Training. Nutrients 2018, 10, 180
~10 g
~8 g
10% (~2 g)
~20 g

Muscle protein synthesis and breakdown
• The major anabolic influences on muscle are contractile activity and feeding.
• Ingestion of sufficient dietary energy and protein is a prerequisite for muscle protein synthesis and maintenance of muscle mass and function.
• As a result of feeding, anabolism occurs by an increase in protein synthesis. Insulin has a permissive role and amino acids are crucial for increasing synthesis and net anabolism.
• In vivo, amino acids display an anabolic effect and were shown to stimulate muscle protein synthesis.
11
(EFSA 2012)
Effect of balanced amino acidinfusion (164 mg/h/kg) on MuscleProtein Synthesis and Breakdown
SynthesisBreakdown
(Giordano et al., 1996; Volpi et al., 1996; Biolo et al, 1997; Bohe et al., 2003; Liu et al., 2002; Nygren and Nair, 2003; Nair and Short, 2005)

Postprandial plasma level of total AAs (A) and branched-chain AAs (B) measured by NMR spectroscopy after intake of intact casein (IC), hydrolyzed casein (HC), and intact whey (IW).
12
Subjects received approximately 30 g protein per protein shake for a diet corresponding to an energy expenditure of 10 MJ/day.
Each protein shake contained 26 energy percent (E%) from protein, of which HC, IC, and IW contributed to 22.3 E% in the respective shakes.
Schmedes et al. The effect of casein, hydrolyzed casein, and whey proteins on urinary and postprandial plasma metabolites in overweight and moderately obese human subjects. J Sci Food Agric (2018)
CaseinHydrolyzed Casein
Intact whey protein
Sum of isoleucine, valine, leucine, alanine, tyrosine, glutamine, histidine, lysine, and glycine
Sum of isoleucine, leucine, and valine

Protein stimulate insulin response
Santosa et al, Diabetologia, 2000
Correlation between daily protein intakeand acute insulin response to glucose as an estimate of insulin secretion capacity

Amino acid sensing recruits mTORC1 and initiates protein translation
14
4E-BP1/2, 4E binding protein 1/2; eiF4A/4G/4E, eukaryotic initiation factor 4A, 4G, or 4E; GATOR1/2, GTPase activating protein (GAP) activity toward Rags 1/2; GDP, guanosine 5’-diphosphate; GTP, guanosine 5’-triphosphate; KICSTOR, Kaptin (KPTN), Integrin alpha phenylalanyl-glycyl-glycyl-alanyl-prolyl (FG-GAP) repeat containing 2 (ITFG2), chromosome 12 open reading frame 66 (C12orf66) and seizure threshold 2 homolog (SZT2)-containing regulator of mTORC1;mTORC1, mechanistic target of Rapamycin complex 1; p70S6K1, 70-kDa ribosomal protein S6 kinase β-1; Rag, Ras-related GTP binding; Rheb, Ras homolog enriched in brain; SLC38A9, solute carrier family 38 member 9; v-ATPase; vacuolar ATPase.
1. Cytosolic amino acids (leucine, arginine) activate: - GATOR2 by CASTOR1 and Sestrin1/2, respectively, leading to the
inhibition of GATOR1, the GAP for the RagA/B small GTPases; - v-ATPase through SLC38A9, leading to the activation of Ragulator,
the guanine exchange factor (GEF) for RagA/B.
2. Upon activation of the Rag heterodimer, mTORC1 is recruited andtranslocated from the cytosol to the lysosomal membrane and isthen activated by the small Rheb GTPase
3. To initiate translation, the capped (m7G) mRNA forms a complex with eIF4A/G/E (termed eIF4F), which is facilitated by phosphorylation of eIF4E-BP 1 through the action of mTORC1.
(Tomé, 2018)

Proposed amino acid sensing mechanisms in skeletal muscle fibers
15
mTORC1 kinase activity is stimulated when it is translocated from the cytosol to the lysosomal membrane to interact with Rheb- Rag GTPases (RagA or RagB bound to RagC or RagD) respond to AAs within
the cell by altering their nucleotide state and their interaction with Ragulator(a lysosomal scaffold).
- GATOR1 is a complex of three proteins and acts as a GAP (GTPase activating protein) for RagA/B, which inhibits mTORC1 activation.
- Upstream of GATOR1 is a five-protein complex known as GATOR2 that is considered a positive regulator of mTORC1 due to its ability to inhibit GATOR1.
Leucine and arginine are stimulator of muscle protein synthesis and mTORC1 signaling in human skeletal muscle. - a leucine sensor (Sestrin2) has been identified. Leucine blocks the
interaction of Sestrin2 with the GATOR2 complex, which ultimately allows mTORC1 to be activated
- The lysosomal AA transporter SLC38A9 has been linked to sensing arginine sufficiency within the lysosome, and interacts with Ragulator to activate mTORC1
- A newly identified arginine sensor (CASTOR1). In this case, arginine binding to CASTOR1 disrupts the GATOR2-CASTOR1 complex thereby allowing GATOR2 to inhibit GATOR1 and eventually leading to activation of mTORC1.
Moro et al. Amino Acid Sensing in Skeletal Muscle Trends Endocrinol Metab. 2016 November ; 27(11): 796–806.
A hypothetical muscle fiber is shown that highlighting the recently identified AA sensing apparatus for leucine and arginine signaling to mTORC1

Muscle protein synthesis (MPS, blue curve) and muscle protein breakdown (MPB, red curve) rates in response to graded intake of protein.
16
With increasing protein ingestion, MPS increases fast but reaches a plateau (at approximately 20 g high quality protein)
Dideriksen et al, Influence of Amino Acids, Dietary Protein, and Physical Activity on Muscle Mass Development in Humans. Nutrients 2013, 5, 852-876
MPB is decreased slightly but continuously with increasing protein intakes.
The net muscle protein balance can possibly benefit from a greater protein intake than known to stimulate MPS maximally (illustrated by the extra net balance area marked with green).
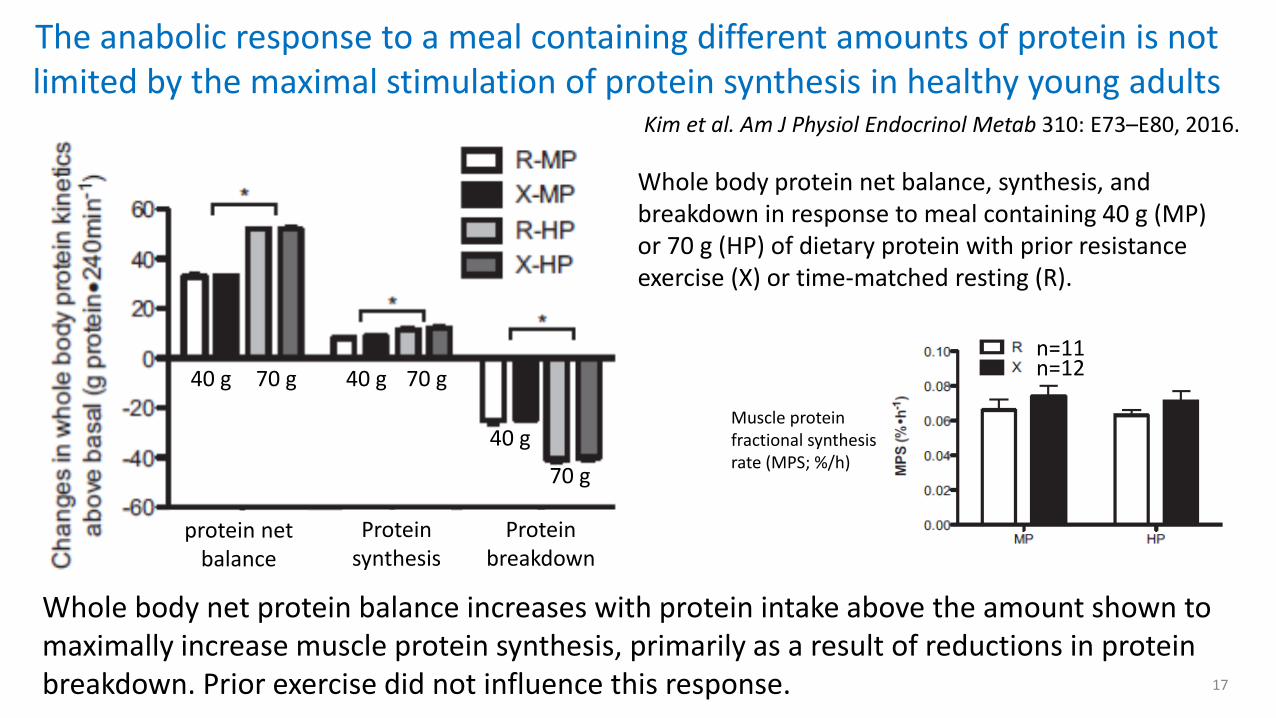
The anabolic response to a meal containing different amounts of protein is not limited by the maximal stimulation of protein synthesis in healthy young adults
17
Muscle protein fractional synthesis rate (MPS; %/h)
Kim et al. Am J Physiol Endocrinol Metab 310: E73–E80, 2016.
n=11n=12
Whole body net protein balance increases with protein intake above the amount shown to maximally increase muscle protein synthesis, primarily as a result of reductions in protein breakdown. Prior exercise did not influence this response.
Whole body protein net balance, synthesis, and breakdown in response to meal containing 40 g (MP) or 70 g (HP) of dietary protein with prior resistance exercise (X) or time-matched resting (R).
40 g 40 g
40 g
70 g
70 g70 g
protein net balance
Proteinsynthesis
Protein breakdown

Branched chain amino acid (BCAA) and Leucine promotesmuscle protein synthesis in adult
Correlation between muscle protein synthesis and arterialflux of leucine (higher values are obtained with aminoacid iv infusion) (Wolfe, 2000).
18
Holeček. Branched-chain amino acids in health and disease. Nutrition & Metabolism(2018) 15:33
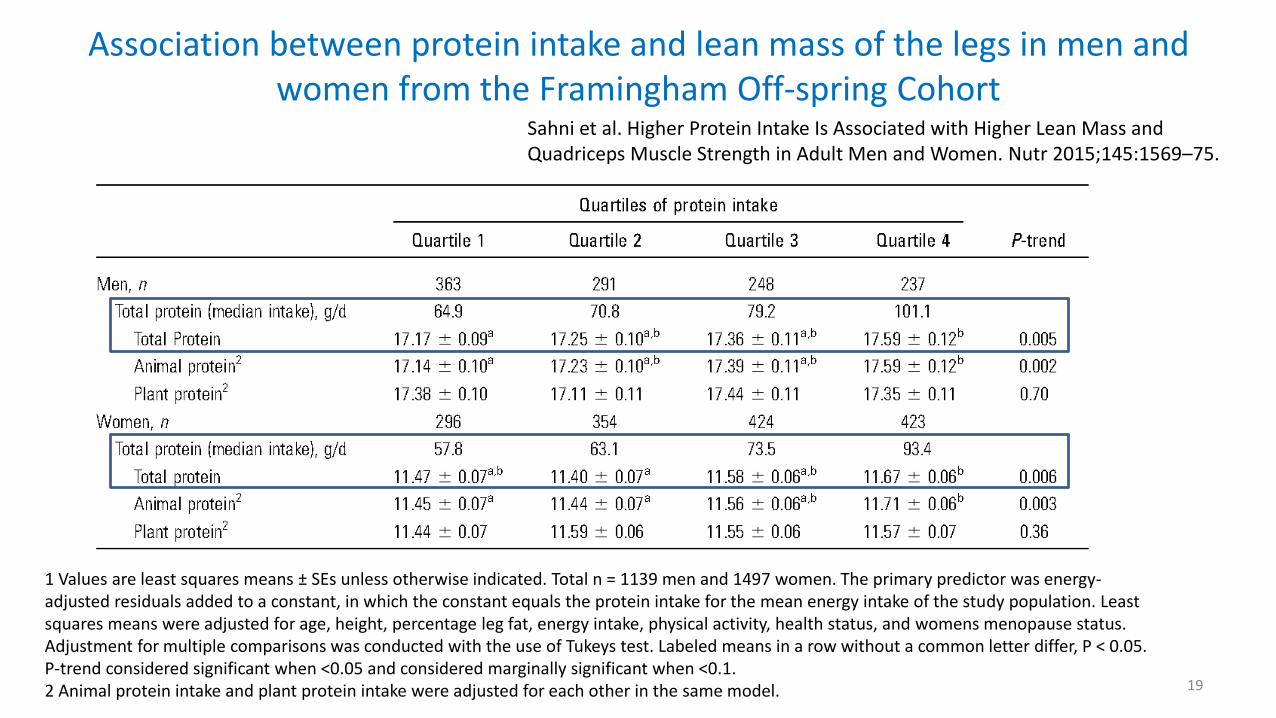
Association between protein intake and lean mass of the legs in men and women from the Framingham Off-spring Cohort
19
Sahni et al. Higher Protein Intake Is Associated with Higher Lean Mass and Quadriceps Muscle Strength in Adult Men and Women. Nutr 2015;145:1569–75.
1 Values are least squares means ± SEs unless otherwise indicated. Total n = 1139 men and 1497 women. The primary predictor was energy-adjusted residuals added to a constant, in which the constant equals the protein intake for the mean energy intake of the study population. Least squares means were adjusted for age, height, percentage leg fat, energy intake, physical activity, health status, and womens menopause status. Adjustment for multiple comparisons was conducted with the use of Tukeys test. Labeled means in a row without a common letter differ, P < 0.05. P-trend considered significant when <0.05 and considered marginally significant when <0.1. 2 Animal protein intake and plant protein intake were adjusted for each other in the same model.
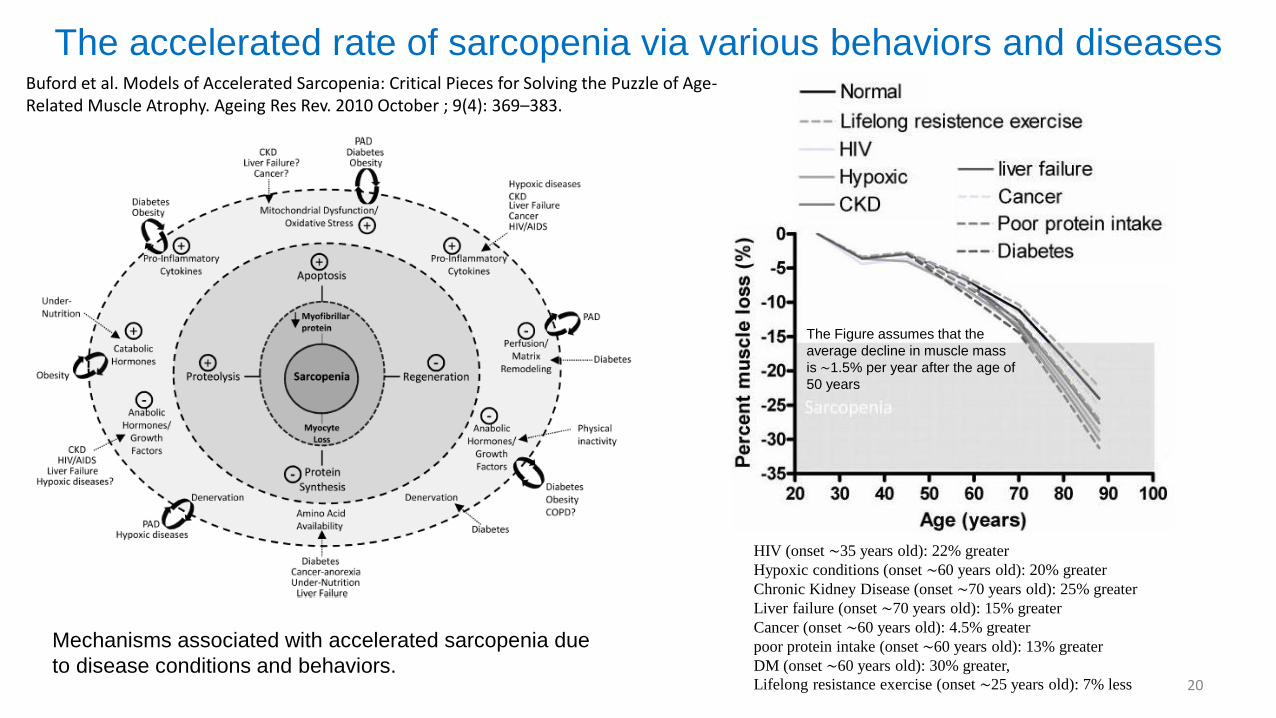
The accelerated rate of sarcopenia via various behaviors and diseases
20
HIV (onset ∼35 years old): 22% greater
Hypoxic conditions (onset ∼60 years old): 20% greater
Chronic Kidney Disease (onset ∼70 years old): 25% greater
Liver failure (onset ∼70 years old): 15% greater
Cancer (onset ∼60 years old): 4.5% greater
poor protein intake (onset ∼60 years old): 13% greater
DM (onset ∼60 years old): 30% greater,
Lifelong resistance exercise (onset ∼25 years old): 7% less
Buford et al. Models of Accelerated Sarcopenia: Critical Pieces for Solving the Puzzle of Age-Related Muscle Atrophy. Ageing Res Rev. 2010 October ; 9(4): 369–383.
The Figure assumes that the
average decline in muscle mass
is ∼1.5% per year after the age of
50 years
Mechanisms associated with accelerated sarcopenia due
to disease conditions and behaviors.

Does the muscle protein synthetic response to exercise and amino acid-based nutrition diminish with advancing age? A systematic review
21
Magnitude of the Muscle Protein Synthetic (MPS) response to provision of a source of amino acids (AA)/protein in young vs. older adults (expressed as %change from basal postabsorptive values).
Shad et al, Am J Physiol Endocrinol Metab 311: E803–E817, 2016.
(Obtained from 21 studies. )(NB: dose, protein source, leucine content, fractional synthesis rate (FSR) incorporation period, and route of administration differ between but not within studies.)
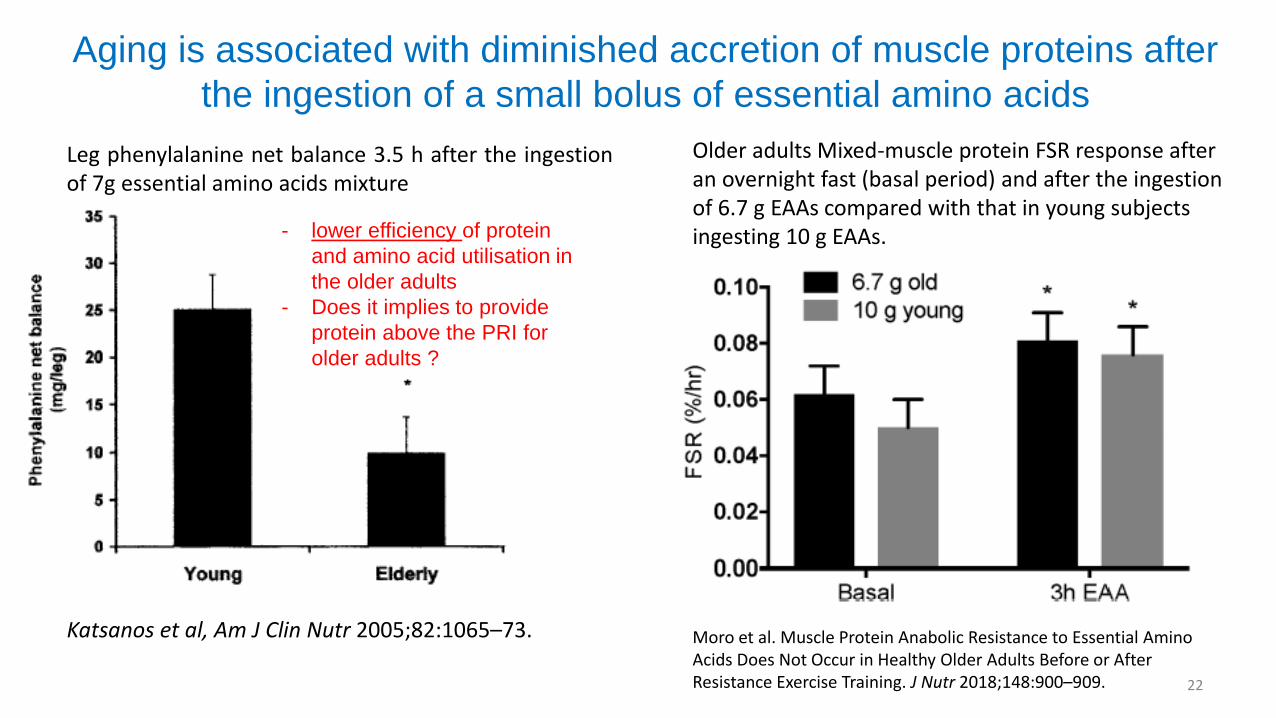
Aging is associated with diminished accretion of muscle proteins after
the ingestion of a small bolus of essential amino acids
Leg phenylalanine net balance 3.5 h after the ingestionof 7g essential amino acids mixture
Katsanos et al, Am J Clin Nutr 2005;82:1065–73.
- lower efficiency of protein
and amino acid utilisation in
the older adults
- Does it implies to provide
protein above the PRI for
older adults ?
22
Older adults Mixed-muscle protein FSR response after an overnight fast (basal period) and after the ingestion of 6.7 g EAAs compared with that in young subjects ingesting 10 g EAAs.
Moro et al. Muscle Protein Anabolic Resistance to Essential Amino Acids Does Not Occur in Healthy Older Adults Before or After Resistance Exercise Training. J Nutr 2018;148:900–909.

A Moderate Serving of High-Quality Protein Maximally Stimulates
Skeletal Muscle Protein Synthesis in Young and Elderly SubjectsSymons et al. J Am Diet Assoc. 2009;109:1582-1586
Mean mixed muscle fractional synthesis rate
before and after ingestions of 113 g (A), or
340 g (B) of 90% lean ground beef by young
and elderly subjects:
- Same increase from fasting (young and
elderly) after 113 g and 340 g lean beef
(P<0.008)
- 113g is sufficient to maximally stimulate
skeletal muscle protein synthesis
- No additional effect on protein synthesis
with 340g lean beef
23

Distribution of protein intake between meals
Protein intake between meal in aduls (≥19 ans) (US)
Paddon-Jones, Rasmussen. Curr Opin Clin Nutr MetabCare. 2009; 12(1): 86–90; Douglas Paddon-Jones et al. Am J Clin Nutr 2015;101:1339S-1345S
Distribution of protein intake in men and women from the Nutrition as a Determinant of Successful Aging study. Data are from Farsijani et al. (2015).

Optimal protein synthesis and daily protein distribution
25
In older adults, optimal protein synthesis in muscle is achieved at
a target intake of 30g or more of high-quality protein. The preponderance of data indicate that while consumption of higher protein doses (> 20 g) results in greater AA oxidation, this is not the fate for all the additional ingested AAs as some are utilized for tissue-building purposes.
Based on the current evidence, we conclude that to maximize anabolism one should consume protein at a target intake of 0.4 g/kg/meal across a minimum of four meals in order to reach a minimum of 1.6 g/kg/day.
Using the upper daily intake of 2.2 g/kg/day reported in the literature spread out over the same four meals would necessitate a maximum of 0.55 g/kg/meal.
Schoenfeld, Aragon. How much protein can the body use in a single meal for muscle-building? Implications for daily protein distribution. Journal of the International Society of Sports Nutrition (2018) 15:10
McDonald et al. MEAL-BASED ENHANCEMENT OF PROTEIN QUALITY AND QUANTITY DURING
WEIGHT LOSS IN OBESE OLDER ADULTS WITH MOBILITY LIMITATIONS: RATIONALE AND
DESIGN FOR THE MEASUR-UP TRIAL. Contemp Clin Trials. 2015 January ; 40: 112–123.

Protein intake distribution pattern does not affect anabolic response, lean body mass, muscle strength or function over 8 weeks in older adults: A randomized-controlled trial
26
Kim et al. Clinical Nutrition 37 (2018) 488e493
Interventional diet during the entire study periodB, breakfast; L, lunch; D, Dinner
EVEN group (n=7) - consumed dietary protein evenly throughout the day (33/33/33%).UNEVEN group (n=7) - consumed the majority of dietary protein with dinner (15/20/65%; breakfast, lunch, dinner),
In older adults consuming an average amount of protein, the distribution pattern of protein intake does not affect changes in lean body mass, muscle strength or muscle function, nor result in changes in whole body net protein balance or MPS after 8 weeks of dietary intervention.

Per meal dose and frequency of protein consumption is associated with lean mass and muscle performance
27
Loenneke t al. Clinical Nutrition 35 (2016) 1506e1511
Strength and leg lean mass results are reported for 1 (vs. 0) and 2 (vs. 0) meals above various protein thresholds (per meal).
With regard to 1 (vs. 0) meal, strength (grey line) and lean mass (grey dash) appear to level off at approximately 30 g/meal.
With regard to 2 (vs. 0) meals, strength (black line) and lean mass (black dash) appear to level off at approximately 45 g/meal.
1-2 daily meals with protein content from 30 to 45 g may be a strategy for increasing and/or maintaining lean body mass with aging.
Dose-response association between protein frequency (1 or 2 meals/day above various thresholds [e.g., 15 g/meal, 20 g/meal, 25 g/meal, etc.) and knee extensor strength and leg lean mass.

Representative schematic for the effect of post-exercise PRO/AA supplementation on the overall mTORC1 signaling and MPS response in human skeletal muscle.
28
Reidy, Rasmussen. Role of Ingested Amino Acids and Protein in the Promotion of Resistance Exercise–Induced Muscle Protein Anabolism. J Nutr 2016;146:155–83.
AA, amino acid; Akt, protein kinase B; AMPK, AMP-activated protein kinase; eEF2, eukaryotic elongation factor 2; eEF2K, eukaryotic elongation factor 2 kinase; eIF2B, eukaryotic initiation factor 2B; ERK1/2, extracellular-related kinase 1/2; Gator, GTPase-activating protein activity towardRags; MPS, muscle protein synthesis; mTOR, mammaliantarget of rapamycin; mTORC1, mammalian target of rapamycin complex 1; PA, phosphatidic acid; Phosph, phosphorylation; PRAS40, proline-rich Akt substrate 40; PRO, protein; Rags, recombination activating genes; Raptor, regulatory-associated protein of mTOR; RE, resistance exercise; Rheb, Ras homolog enriched in brain; rpS6, ribosomal protein S6; S6K1/p70S6K1, p70 ribosomal protein S6 kinase 1; TSC1, tuberous sclerosis complex 1; TSC2, tuberous sclerosis complex 2; 4E-BP1, eukaryotic initiation factor 4E binding protein 1.
Resistance Exercise promotes the anabolic effect of amino acid and insulin on muscle protein synthesis.

Muscle protein synthetic (MPS) and protein intake at rest (solid curves) and with prior resistance exercise (dashed curves) in young (green curves) and elderly (blue curves).
29
In the young, MPS is stimulated already at small doses (~5 g) of protein intake and the response reaches a plateau at approximately 20 g of protein intake. The MPS response to protein intake can be enhanced by prior completion of resistance exercise
In the elderly, a higher amount of protein intake is necessary to simulate MPS and to obtain the maximal MPS response. The effect of resistance exercise is somewhat reduced in elderly compared to young individuals.
The red curve illustrates that the MPS response to protein intake is reduced in immobilized muscle.
Dideriksen et al, Influence of Amino Acids, Dietary Protein, and Physical Activity on Muscle Mass Development in Humans. Nutrients 2013, 5, 852-876
Young
Older
Immobilized muscle
Exercise
Exercise
Rest
Rest

Gains in muscle mass and function due to exercise require regular training over an extended period of time.
Galvan et al. Protecting Skeletal Muscle with Protein and Amino Acid during Periods of DisuseNutrients. 2016 Jul; 8(7): 404. Twelve weeks of resistance exercise training result
in a modest gain (~1.5 kg) in musclemass in older adults
Loss of muscle health due to disuse occurs over a very short period of time; seven days of bed rest is sufficient to induce ~1 kg loss leg lean mass alone.
Leucine supplemented meals may partially and temporarily protect skeletal muscle during disuse by preserving anabolism and mitigating reductions in mass, function and metabolic homeostasis ?

Whey protein ingestion-induced increase in MPS in young men, percent change from 0 g
31Stokes et al. Recent Perspectives Regarding the Role of Dietary Protein for the Promotion of Muscle
Hypertrophy with Resistance Exercise Training. Nutrients 2018, 10, 180
At rest, consumption of 10 g or 20 g of protein results in a
rise of 19% and 52% respectively from 0g. Consumption of
40 g of whey protein does not result in superior stimulation
of MPS beyond consumption of 20 g
Following resistance exercise, consumption of 20 g of protein
increases MPS almost twice as much as consumption of 10 g,
while consumption of 40 g of protein results in a small stimulation
of MPS over and above that seen at 20 g

A systematic review, meta-analysis and meta-regression of the effect of protein supplementation on resistance training-induced gains in muscle mass and strength in healthy adults
Morton RW, et al. Br J Sports Med 2018;52:376–384
Random-effects univariate meta-regression between ageand the mean difference in fat-free mass (FFM) between groups. The regression prediction is represented by the solid line (−0.01 kg (−0.02,–0.00), p=0.02).
32
Dietary protein supplementation augments changes in muscle mass and strength during prolonged RET. Protein supplementation is more effective at improving FFM in young or resistance-trained individuals than in older or untrained individuals. Protein supplementation is sufficient at ~1.6 g/kg/day in healthy adults during RET.
Segmental linear regression between relative total protein intake (g/kg body mass/day) and the change in fat-free mass (ΔFFM) measured by dual energy X-ray absorptiometry. Dashed arrow indicates the break point=1.62 g protein/kg/day, p=0.079. Solid arrow indicates 95% CI, (1.03 to 2.20).
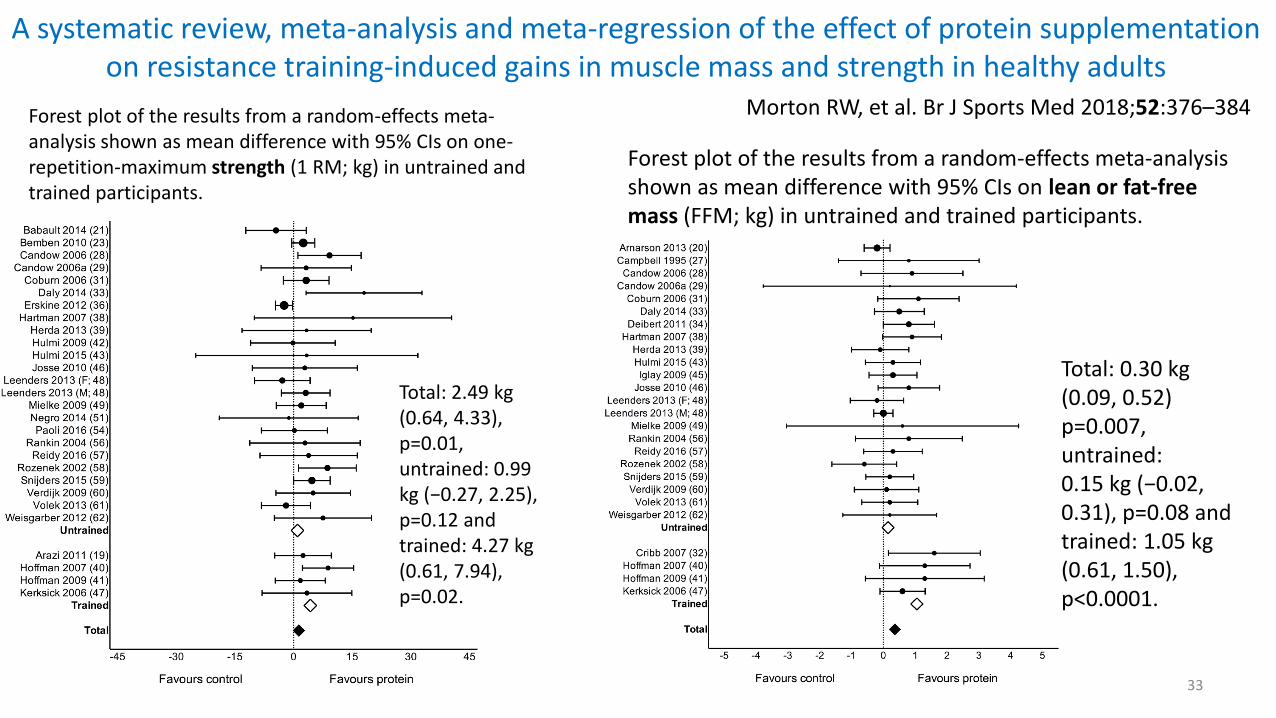
A systematic review, meta-analysis and meta-regression of the effect of protein supplementation on resistance training-induced gains in muscle mass and strength in healthy adults
33
Morton RW, et al. Br J Sports Med 2018;52:376–384Forest plot of the results from a random-effects meta-analysis shown as mean difference with 95% CIs on one-repetition-maximum strength (1 RM; kg) in untrained and trained participants.
Total: 2.49 kg (0.64, 4.33), p=0.01, untrained: 0.99 kg (−0.27, 2.25), p=0.12 and trained: 4.27 kg (0.61, 7.94), p=0.02.
Forest plot of the results from a random-effects meta-analysis shown as mean difference with 95% CIs on lean or fat-free mass (FFM; kg) in untrained and trained participants.
Total: 0.30 kg (0.09, 0.52) p=0.007, untrained:0.15 kg (−0.02, 0.31), p=0.08 and trained: 1.05 kg (0.61, 1.50), p<0.0001.

The Effect of Whey Protein Supplementation on the Temporal Recovery of Muscle Function Following Resistance Training: A Systematic Review and Meta-Analysis
34
Davies et al. Nutrients 2018, 10, 221
Forest plot of the temporal effect sizes (ESs) of the whey protein supplement for the recovery of muscle function following resistance training, compared to a control treatment. Data are mean 95% CI. ESs are ordered by time.
Overall small to medium ergogenic effects were observed for WP supplementation restoring muscle contractile function following RT from < 24 to 96 h (ES range = 0.4–0.7).
Half of the included studies reported a beneficial overall effect for WP, whilst the other half concluded no effect occurred compared to the control.
No study reported any overall negative effect for WP and only one negative effect (favouring the control) was reported out of the 44 temporal comparisons.
Although the RCTs were of reasonably high quality, significant heterogeneity was observed between studies.

Exercising before protein intake allows for greater use of dietary protein–derived amino acids for de novo muscle protein synthesis in both young and elderly men
35
Pennings et al. Am J Clin Nutr 2011;93:322–31.
Exercise effectP < 0.001
Exercise effectP < 0.05
Mixed-muscle protein fractional synthetic rates (FSRs) after protein ingestion in young (n = 24) and elderly (n = 24) men at rest (REST) and after exercise (EXC)
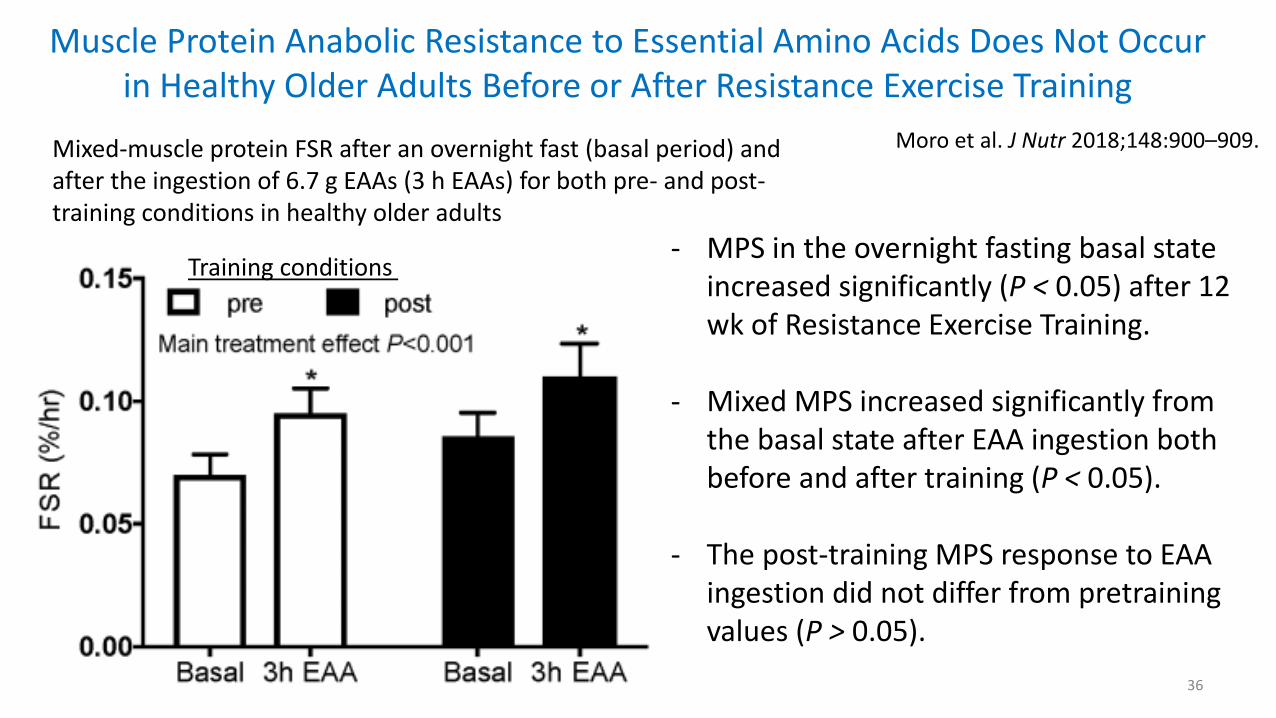
Muscle Protein Anabolic Resistance to Essential Amino Acids Does Not Occur in Healthy Older Adults Before or After Resistance Exercise Training
36
Moro et al. J Nutr 2018;148:900–909.
- MPS in the overnight fasting basal state increased significantly (P < 0.05) after 12 wk of Resistance Exercise Training.
- Mixed MPS increased significantly from the basal state after EAA ingestion both before and after training (P < 0.05).
- The post-training MPS response to EAA ingestion did not differ from pretraining values (P > 0.05).
Mixed-muscle protein FSR after an overnight fast (basal period) and after the ingestion of 6.7 g EAAs (3 h EAAs) for both pre- and post-training conditions in healthy older adults
Training conditions

Protein supplementation augments the adaptive response of skeletal muscle to resistance-type exercise training: a meta-analysis
37
Forest plot of the results of a random-effects meta-analysis shown as pooled mean differences with 95% CIs on fat-free mass in younger and older subjects (weighted mean difference: 0.69 kg; 95% CI: 0.47, 0.91 kg; P , 0.00001).
Forest plot of the results of a random-effects meta-analysis shown as pooled mean differences with 95% CIs on fat-free mass in younger untrained and younger trained subjects (weighted mean difference: 0.81 kg; 95% CI: 0.53, 1.1 kg; P , 0.00001).
The diamonds represent the subgroup mean difference ()) and pooled mean difference.
Protein supplementation increases muscle mass and strength gains during prolonged resistance-type exercise training in both younger and older subjects.
Cermak et al; Am J Clin Nutr 2012;96:1454–64.

Leucine, Not Total Protein, Content of a Supplement Is the Primary Determinant of Muscle Protein Anabolic Responses in Healthy Older Women
38
Devries et al. Nutr 2018;148:1088–1095
Acute response. = 11/group. *Different from fasting, P < 0.001; #different from rest, P < 0.001; †different from WPI, P = 0.04. FSR, fractional synthetic rate; LEU, leucine; myoPS, myofibrillar protein synthesis; WPI, whey protein isolate.
Integrated response. *Different from basal, P = 0.02; #different from rest, P < 0.001. FSR, fractional synthetic rate; LEU, leucine; myoPS, myofibrillar protein synthesis; WPI, whey protein isolate.
A lower-protein (10 g) beverage containing added leucine (3 g) elicited comparable muscle protein synthetic responses to a beverage containing 25 g protein and matched for leucine content in older women, both acutely and when incorporated into the habitual diet in the rested and exercised states.
Myofibrillar FSR in the fasting state and after consumption of LEU or WPI in the rested (Rest) and exercised (Exercise) leg in healthy, older women
6 d

The impact of protein quality on the promotion of resistance exercise-induced changes in muscle mass
39
Schematic showing the impact of proteins with differing leucine contents on skeletal muscle fibre intracellularconcentration of leucine and subsequent stimulation of muscle protein synthesis (MPS).
The concept is that certain proteins with high leucine content and digestibility would raise intracellular Leu concentration and bind with Sestrin2 (see text for details) to trigger activation of MPS of varying degrees (a).
The resultant MPS response (shown as a percent stimulation above resting) is shown in (b)
Phillips Nutrition & Metabolism (2016) 13:64

Conclusion
40
• The major anabolic influences on muscle are contractile activity and feeding.
• Ingestion of 15-30 g protein by meal promotes muscle protein synthesis and maintenance of muscle mass and function.
• Exercise potentiates the anabolic effect of feeding on muscle protein synthesis and maintenance of muscle mass and function
• the consumption of 1-2 daily meals with protein content above 20 g may be a strategy for increasing and/or maintaining lean body mass and muscle strength with aging
• Leucine remains an interesting strategy to promote muscle protein synthesis and maintenance of muscle mass with aging

Thank you
41


















