
PROTEIN METABOLISM: NITROGEN CYCLE; DIGESTION OF...
53
PROTEIN PROTEIN METABOLISM: METABOLISM: NITROGEN CYCLE; NITROGEN CYCLE; DIGESTION OF DIGESTION OF PROTEINS PROTEINS Red meat is an important dietary source of protein nitrogen
Transcript of PROTEIN METABOLISM: NITROGEN CYCLE; DIGESTION OF...

PROTEIN PROTEIN METABOLISM: METABOLISM:
NITROGEN CYCLE; NITROGEN CYCLE; DIGESTION OF DIGESTION OF
PROTEINS PROTEINS
Red meat is an important dietary source of protein nitrogen

The Nitrogen Cycle and Nitrogen Fixation
• Nitrogen is needed for amino acids, nucleotides, etc
• Atmospheric N2 is the ultimate source of biological nitrogen
• Nitrogen fixation: biosynthetic process of the reduction of N2 to NH3 (ammonia)
• Higher organisms are unable to fix nitrogen.• Some bacteria and archaea can fix nitrogen.

Archaea
Symbiotic Rhizobium bacteria invade the roots of leguminous plants and form root nodules.
Rhizobium bacteria fix nitrogen supplying both the bacteria and the plants.
Nodules of Rhizobium bacteria

The amount of N2 fixed by nitrogen-fixing microorganisms is about 60% of Earth's newly fixed nitrogen.
25% is fixed by industrial processes (fertilizer factories)
Lightning and ultraviolet radiation fix 15%
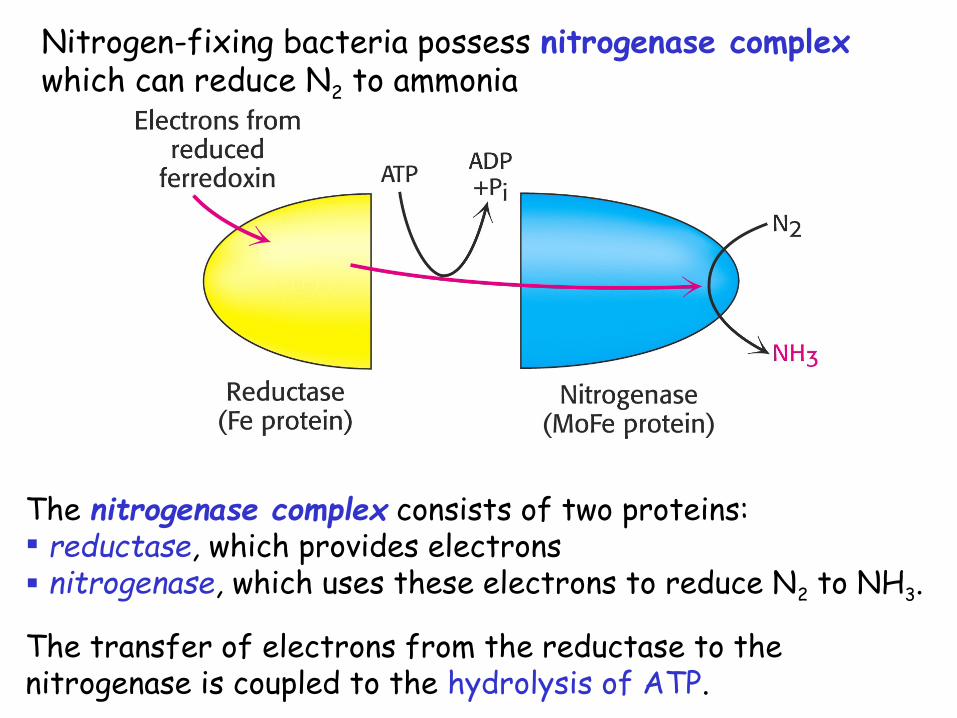
Nitrogen-fixing bacteria possess nitrogenase complex which can reduce N2 to ammonia
The nitrogenase complex consists of two proteins: reductase, which provides electrons nitrogenase, which uses these electrons to reduce N2 to NH3.
The transfer of electrons from the reductase to the nitrogenase is coupled to the hydrolysis of ATP.

• Nitrogenase reaction: N2 + 8 H+ + 8 e- + 16 ATP 2 NH3 + H2 + 16 ADP + 16 Pi
The high-potential electrons come from protein ferredoxin, generated by photosynthesis or oxidative processes.
16 molecules of ATP are hydrolyzed for each molecule of N2 reduced.
Reductase – dimer containing Fe-S clusters and ATP-binding site

The nitrogenase component is an 22 tetramer.
Contains P cluster (Fe-S) and FeMo cofactor.
FeMo cofactor is the site of nitrogen fixation.

Nitrosomonas
Ammonia in the presence of water becomes NH4
+ which can be used by plants
NH4+ can be rapidly oxidized
by soil bacteria Nitrosomonas and Nitrobacter to NO2
- and NO3
- (nitrification)
NO2- and NO3
- are used by higher plants
Another soil bacteria can reverse the nitrification process and convert NO2
- and NO3- back to nitrogen
Nitrogen from plants and animals is recycled to soil (excretion of nitrogen in the form of urea or uric acid; decay of plants and animals) - nitrogen cycle


Assimilation of Ammonia
• Ammonia generated from N2 is assimilated into amino acids such as glutamate or glutamine

A. Ammonia Is Incorporated into Glutamate
• Reductive amination of a-ketoglutarate by glutamate dehydrogenase occurs in plants, animals and microorganisms
This reaction establishes the stereochemistry of the -carbon atom in glutamate. Only the L isomer of glutamate is synthesized.
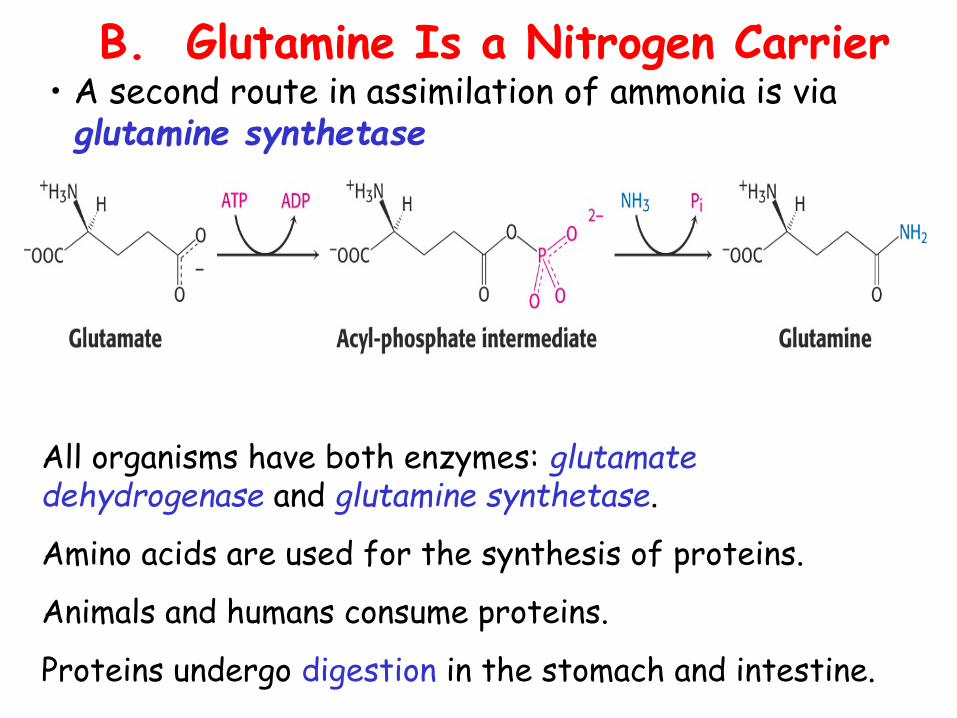
B. Glutamine Is a Nitrogen Carrier• A second route in assimilation of ammonia is via glutamine synthetase
All organisms have both enzymes: glutamate dehydrogenase and glutamine synthetase.
Amino acids are used for the synthesis of proteins.
Animals and humans consume proteins.
Proteins undergo digestion in the stomach and intestine.

Protein digestionDigestion in Stomach
Stimulated by food acetylcholine, histamine and gastrin are released onto the cells of the stomach The combination of acetylcholine, histamine and gastrin cause the release of the gastric juice.
Mucin - is always secreted in the stomach HCl - pH 0.8-2.5 (secreted by parietal cells) Pepsinogen (a zymogen, secreted by the chief cells)
Hydrochloric acid: Creates optimal pH for pepsin Denaturates proteins Kills most bacteria and other foreign cells

Pepsinogen (MW=40,000) is activated by the enzyme pepsin present already in the stomach and the stomach acid. Pepsinogen cleaved off to become the enzyme pepsin (MW=33,000) and a peptide fragment to be degraded. Pepsin partially digests proteins by cleaving the peptide bond formed by aromatic amino acids: Phe, Tyr, Trp

Digestion in the Duodenum Stimulated by food secretin and cholecystokinin regulate the secretion of bicarbonate and zymogens trypsinogen, chymotrypsinogen, proelastase and procarboxypeptidase by pancreas into the duodenum
Bicarbonate changes the pH to about 7
The intestinal cells secrete an enzyme called enteropeptidase that acts on trypsinogen cleaving it into trypsin

Trypsin converts chymotrypsinogen into chymotrypsin, procarboxypeptidase into carboxypeptidase and proelastase into elastase, and trypsinogen into more trypsin.
Trypsin which cleaves peptide bonds between basic amino acids Lys and Arg
Chymotrypsin cleaves the bonds between aromatic amino acids Phe, Tyr and Trp Carboxypeptidase which cleaves one amino acid at a time from the carboxyl sideAminopeptidase is secreted by the small intestine and cleaves off the N-terminal amino acids one at a time

Most proteins are completely digested to free amino acids
Amino acids and sometimes short oligopeptides are absorbed by the secondary active transport
Amino acids are transported via the blood to the cells of the body.

The ways of entry and using of amino acids in tissue
The sources of amino acids:
1) absorption in the intestine; 2) protein decomposition; 3) synthesis from the carbohydrates and lipids.
Using of amino acids: 1) for protein synthesis;2) for synthesis of other nitrogen containing
compounds (creatine, purines, choline, pyrimidine);3) as the source of energy; 4) for the gluconeogenesis.

PROTEIN PROTEIN METABOLISM: METABOLISM:
PROTEIN TURNOVER; PROTEIN TURNOVER; GENERAL WAYS OF GENERAL WAYS OF
AMINO ACIDS AMINO ACIDS METABOLISM METABOLISM

PROTEIN TURNOVER
How can a cell distinguish proteins that are meant for degradation?
Protein turnover — the degradation and resynthesis of proteinsHalf-lives of proteins – from several minutes to many years
Structural proteins – usually stable (lens protein crystallin lives during the whole life of the organism)Regulatory proteins - short lived (altering the amounts of these proteins can rapidly change the rate of metabolic processes)

Ubiquitin - is the tag that marks proteins for destruction ("black spot" - the signal for death)
Ubiquitin - a small (8.5-kd) protein present in all eukaryotic cells
Structure: extended carboxyl terminus (glycine) that is linked to other proteins; lysine residues for linking additional ubiquitin molecules

Ubiquitin covalently binds to -amino group of lysine residue on a protein destined to be degraded.
Isopeptide bond is formed.

E1 - ubiquitin-activating enzyme (attachment of ubiquitin to a sulfhydryl group of E1; ATP-driven reaction)
E2 - ubiquitin-conjugating enzyme (ubiquitin is shuttled to a sulfhydryl group of E2) E3 - ubiquitin-protein ligase (transfer of ubiquitin from E2 to -amino group on the target protein)
Mechanism of the binding of ubiquitin to target protein

Attachment of a single molecule of ubiquitin - weak signal for degradation.
Chains of ubiquitin are generated.
Linkage – between -amino group of lysine residue of one ubiquitin to the terminal carboxylate of another.
Chains of ubiquitin molecules are more effective in signaling degradation.

What determines ubiquitination of the protein?
1. The half-life of a protein is determined by its amino-terminal residue (N-terminal rule). E3 enzymes are the readers of N-terminal residues.
2. Cyclin destruction boxes - specific amino acid sequences (proline, glutamic acid, serine, and threonine –PEST)

What is the executioner of the protein death?
A large protease complex proteasome or the 26S proteasome digests the ubiquitinated proteins.
26S proteasome - ATP-driven multisubunit protease.
26S proteasome consists of two components: 20S - catalytic subunit 19S - regulatory subunit
Digestion of the Ubiquitin-Tagged Proteins

20S subunit resembles a barrel is constructed from 28 polipeptide chains which are arranged in four rings (two and two ) active sites are located in rings on the interior of the barrel degrades proteins to peptides (seven-nine residues)

made up of 20 polipeptide chains
controls the access to interior of 20S barrel
binds to both ends of the 20S proteasome core
binds to polyubiquitin chains and cleaves them off
possesses ATPase activity
unfold the substrate
induce conformational changes in the 20S proteasome (the substrate can be passed into the center of the complex)
19S subunit

GENERAL WAYS OF AMINO ACIDS METABOLISM
The fates of amino acids: 1) for protein synthesis;
2) for synthesis of other nitrogen containing compounds (creatine, purines, choline, pyrimidine);
3) as the source of energy;
4) for the gluconeogenesis.

The general ways of amino acids degradation: Deamination Transamination Decarboxilation
The major site of amino acid degradation - the liver.
Deamination of amino acidsDeamination - elimination of amino group from amino
acid with ammonia formation.
Four types of deamination: - oxidative (the most important for higher animals), - reduction, - hydrolytic, and - intramolecular

Reduction deamination:
R-CH(NH2)-COOH + 2H+ R-CH2-COOH + NH3
amino acid fatty acid
Hydrolytic deamination:
R-CH(NH2)-COOH + H2O R-CH(OH)-COOH + NH3
amino acid hydroxyacid
Intramolecular deamination:
R-CH(NH2)-COOH R-CH-CH-COOH + NH3
amino acid unsaturated fatty acid

Oxidative deaminationL-Glutamate dehydrogenase plays a central role in amino acid deaminationIn most organisms glutamate is the only amino acid that has active dehydrogenasePresent in both the cytosol and mitochondria of the liver

Transamination of amino acids
Transamination - transfer of an amino group from an -amino acid to an -keto acid (usually to -ketoglutarate)
Enzymes: aminotransferases (transaminases).
-amino acid-keto acid -keto acid-amino acid

There are different transaminases The most common: alanine aminotransferase alanine + -ketoglutarate pyruvate + glutamateaspartate aminotransferase aspartate + -ketoglutarate oxaloacetate + glutamate
Aminotransferases funnel -amino groups from a variety of amino acids to -ketoglutarate with glutamate formationGlutamate can be deaminated with NH4
+ release

Mechanism of transaminationAll aminotransferases require the prosthetic group pyridoxal phosphate (PLP), which is derived from pyridoxine (vitamin B6).
First step: the amino group of amino acid is transferred to pyridoxal phosphate, forming pyridoxamine phosphate and releasing ketoacid.
Second step: -ketoglutarate reacts with pyridoxamine phosphate forming glutamate
Ping-pong kinetic mechanism
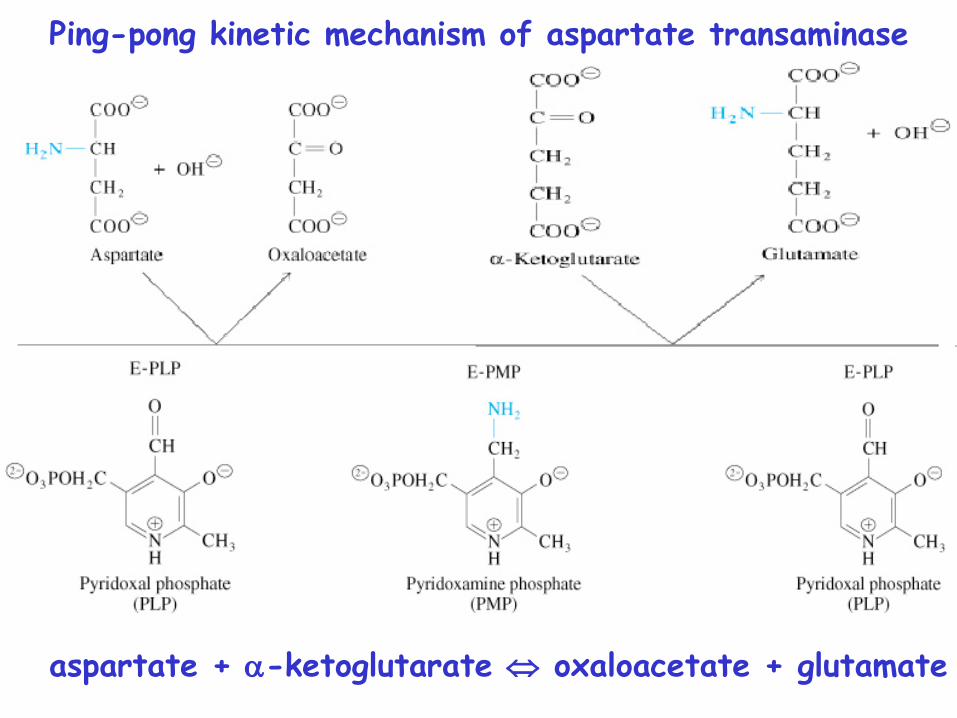
Ping-pong kinetic mechanism of aspartate transaminase
aspartate + -ketoglutarate oxaloacetate + glutamate

Decarboxylation – removal of carbon dioxide from amino acid with formation of amines.
Usually amines have high physiological activity (hormones, neurotransmitters etc).
amine
Enzyme: decarboxylases Coenzyme – pyrydoxalphosphate
Decarboxylation of amino acids

Significance of amino acid decarboxylation1. Formation of physiologically active compounds
glutamate gamma-aminobutyric acid (GABA)
GABA – mediator of nervous system
histaminehistidine
Histamine – mediator of inflammation, allergic reaction.

2. Catabolism of amino acids during the decay of proteins
ornithine putrescine
lysine cadaverine
Enzymes of microorganisms (in colon; dead organisms) decarboxylate amino acids with the formation of diamines.

PROTEIN METABOLISM: PROTEIN METABOLISM: UTILIZATION OF AMMONIA UTILIZATION OF AMMONIA
IONS; UREA CYCLEIONS; UREA CYCLE
The basic features of nitrogen metabolism were elucidated initially in pigeons

AMMONIA METABOLISMThe ways of ammonia formation
1. Oxidative deamination of amino acids
2. Deamination of physiologically active amines and nitrogenous bases.
3. Absorption of ammonia from intestine (degradation of proteins by intestinal microorganisms results in the ammonia formation).
4. Hydrolytic deamination of AMP in the brain (enzyme – adenosine deaminase)

Ammonia is a toxic substance to plants and animals (especially for brain)
Normal concentration: 25-40 mol/l (0.4-0.7 mg/l)
Ammonia must be removed from the organism
Terrestrial vertebrates synthesize urea (excreted by the kidneys) - ureotelic organisms
Birds, reptiles synthesize uric acid
Urea formation takes place in the liver

Peripheral Tissues Transport Nitrogen to the Liver
Two ways of nitrogen transport from peripheral tissues (muscle) to the liver:
1. Alanine cycle. Glutamate is formed by transamination reactions
Glutamate is not deaminated in
peripheral tissues

Nitrogen is then transferred to pyruvate to form alanine, which is released into the blood.
The liver takes up the alanine and converts it back into pyruvate by transamination. The glutamate formed in the liver is deaminated and ammonia is utilized in urea cycle.
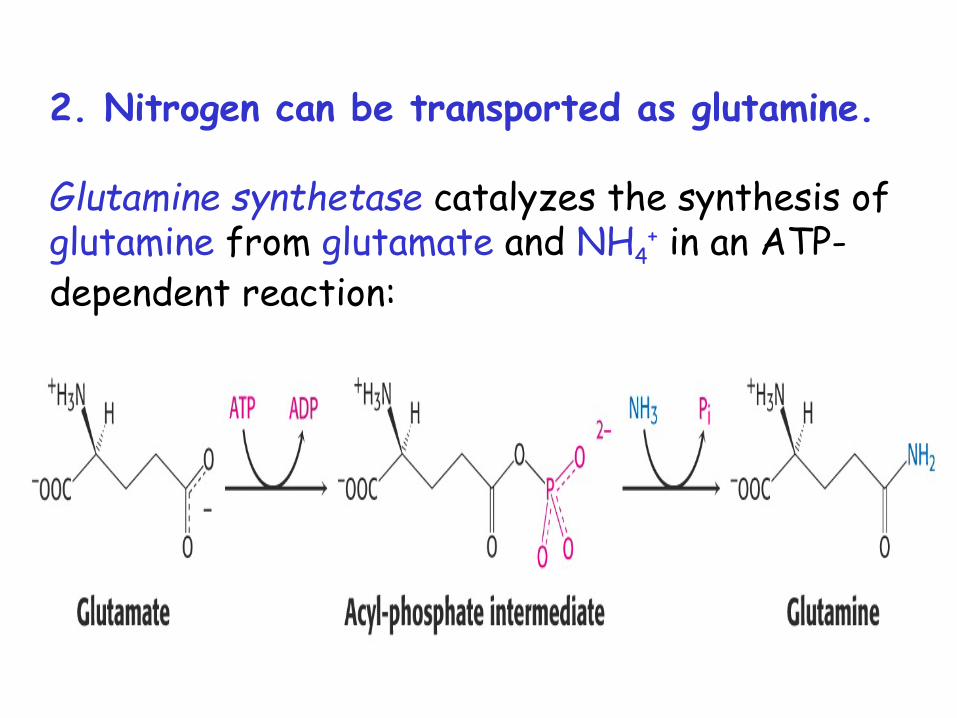
2. Nitrogen can be transported as glutamine.
Glutamine synthetase catalyzes the synthesis of glutamine from glutamate and NH4
+ in an ATP-dependent reaction:

Urea cycle - a cyclic pathway of urea synthesis first postulated by H.Krebs
THE UREA CYCLE
The sources of nitrogen atoms in urea molecule:- aspartate;- NH4
+.
Carbon atom comes from CO2.

The free ammonia is coupling with carbon dioxide to form carbamoyl phosphate
Two molecules of ATP are requiredReaction takes place in the matrix of liver
mitochondriaEnzyme: carbamoyl phosphate synthetase (20 % of
the protein of mitochondrial matrix)

Carbamoyl phosphate donates carbamoyl group to ornithine
The product - citruilline
Enzyme: ornithine carbamoyltransferase Reaction takes place in the mitochondrial matrix
Citrulline leaves the matrix and passes to the cytosol

In the cytosol citrulline in the presence of ATP reacts with aspartate to form argininosuccinate
Enzyme: argininosuccinate synthetase

Argininosuccinate is cleaved to free arginine and fumarateEnzyme: argininosuccinate lyase
The fumarate enters the tricarboxylic acid cycle

Arginine is hydrolyzed to generate urea and ornithineEnzyme: arginase (present only in liver of ureotelic animals)
Ornithine is transported back into the mitochondrion to begin another cycle
Urea is excreted (about 40 g per day)

The urea cycle

The Linkage between Urea Cycle, Citric Acid Cycle and Transamination of Oxaloacetate
Fumarate formed in urea cycle enters citric acid cycle and is converted to oxaloacetate.
Fates of oxaloacetate: (1) transamination to aspartate, (2) conversion into glucose,(3) condensation with acetyl CoA to form citrate,(4) conversion into pyruvate.










