Properties of DNA rosettes and their relevance to chromosome structure
8
Chromosoma (Berl) (1983) 88:307-314 CHROMOSOMA Springer-Verlag 1983 Properties of DNA rosettes and their relevance to chromosome structure Pedro Le6n and Gabriel Macaya Centro de Investigaci6n en Biologia Celular y Molecular, Universidad de Costa Rica, Ciudad Universitaria "Rodrigo Facio", San Jos~, Costa Rica Abstract. We have studied the spreading conditions that lead to the formation of rosettes in DNA and chromatin preparations from the amphibians Bufo marinus and Boli- toglossa subpalrnata and the bacterium Shigella. Both nucle- ar preparations and extensively deproteinized DNA pro- duced rosettes. The longest fibers and the most symmetric rosettes were observed in amphibian nuclear spreadings. In this procedure purified nuclei were submitted immediate- ly to Kleinschmidt spreading over various types of hypo- phase. Distilled-water hypophases were most conducive for rosette production or stability. Rosettes were observed with cytochrome C as the basic protein, but not with ribonucle- ase A and bovine serum albumin. We cannot prove that all rosettes are artifacts of the spreading procedure, but we believe that at least some result from the expansion of compact DNA doughnuts and other structures that are apparently formed in the presence of basic proteins in salt concentrations over 40 mM (Olins and Olins 1971; Man- ning 1979). The dilute hypophase requirement is explainable by the assumption that dilution and spreading effects un- fold a compact precursor. Occasionally we have detected structures that appear to be intermediates in the process of doughnut unfolding and that illustrate a procedure that may give rise to rosettes. Introduction DNA rosettes were first reported by Kleinschmidt and Zahn (1959) who observed them after spreading DNA from fish and bacterial ceils on an air-water film of denatured, basic protein. Kleinschmidt and collaborators (1959, 1968) considered rosettes to be artifacts of the procedure, since they disappeared after the molarity of the hypophase was raised. Several investigators (Inman 1967; Davis et al. 1971) have since agreed that the structure is an artifact induced by spreading DNA with cytochrome C over dilute hypo- phases, but others (Sonnenbichler 1969a, b; Comings and Okada 1976; Okada and Comings 1979) have proposed that rosettes represent packaging domains of the chromatin fiber and have presented evidence for the origin of rosettes from the intrinsic periodicity in the fiber. The presence of loops in dehistonized eucaryotic chro- mosomes was independently detected by sedimentation stu- dies using sucrose gradient centrifugation of dehistonized metaphase chromosomes and interphase nuclei in the pres- ence of ethidium bromide (Cook and Brazell 1975; Benya- jail and Worcel 1976; Adolph etal. 1977; Paulson and Laemmli 1977). As the concentration of ethidium bromide in the solution is increased, sedimentation behavior typical of covalently closed circular molecules is observed. This suggests that the chromosomal fiber is anchored at distant sites and prevented from rotation. Surface spreading of de- histonized metaphase chromosomes reveals 10- to 30-gm loops anchored to a proteinaceous scaffold; rosettes are not evident in these preparations (Paulson and Laemmli 1977; Hadlacsky et al. 1981 ; Mullinger and Johnson 1980). Loops in sedimentation studies are estimated to range in size from 30 to 90 kilobase pairs (kb) in most reported cases (Benyajati and Worcel 1976; Paulson and Laemmli 1977). Individual loops in rosettes are much smaller (1 5 kb) (Okada and Comings 1979; this paper). Several cytological studies with light and electron mi- croscopy reveal loops in whole-mount and sectioned chro- mosomes, from a wide variety of eucaryotes (Sorsa 1973; Comings and Okada 1973). The possibility that DNA ro- settes correspond on a cytological level to chromomeres was first proposed by Sonnenbichler (1969). We studied the structure of chromatin from the nuclei of amphibian red blood cells (RBCs) by centrifugation, elec- trophoresis, and spreading procedures for electron micros- copy in an attempt to clarify the origin of rosettes and other aggregates. We found that several types of aggregates could be produced, depending on the state and source of the DNA, the basic-protein requirements, and the type of hypophase used. From our results we conclude that at least some rosettes form during cytochrome C spreading of the DNA over dilute hypophases, as Kleinschmidt proposed. Materials and methods Organisms used. The toad Bufo marinus, collected in San Jos6, was kept in shallow open aquaria and the salamander Bolitoglossa subpalmata, from the Cerro de la Muerte, lo- cated along the Pan American Highway south of San Jos~, was kept at 18 ~ C in moss terraria. The toads were anesthe- tized with diethyl ether; the salamanders, with a dilute chloretone solution. Isolation of nuclei. RBCs obtained by heart puncture were lysed in ice-cold 0.1 x SSC (SSC is 0.15 M NaC1, 0.015 M sodium citrate, pH 6.8) for 30-60 min. Lysis was monitored by phase-contrast microscopy. Nuclei from lysed cells were pelleted by centrifugation (2,000 rpm for 15 min in a Hita-
-
Upload
pedro-leon -
Category
Documents
-
view
213 -
download
1
Transcript of Properties of DNA rosettes and their relevance to chromosome structure

Chromosoma (Berl) (1983) 88:307-314 CHROMOSOMA �9 Springer-Verlag 1983
Properties of D N A rosettes and their relevance to chromosome structure
Pedro Le6n and Gabriel Macaya Centro de Investigaci6n en Biologia Celular y Molecular, Universidad de Costa Rica, Ciudad Universitaria "Rodrigo Facio", San Jos~, Costa Rica
Abstract. We have studied the spreading conditions that lead to the formation of rosettes in DNA and chromatin preparations from the amphibians Bufo marinus and Boli- toglossa subpalrnata and the bacterium Shigella. Both nucle- ar preparations and extensively deproteinized DNA pro- duced rosettes. The longest fibers and the most symmetric rosettes were observed in amphibian nuclear spreadings. In this procedure purified nuclei were submitted immediate- ly to Kleinschmidt spreading over various types of hypo- phase. Distilled-water hypophases were most conducive for rosette production or stability. Rosettes were observed with cytochrome C as the basic protein, but not with ribonucle- ase A and bovine serum albumin. We cannot prove that all rosettes are artifacts of the spreading procedure, but we believe that at least some result from the expansion of compact DNA doughnuts and other structures that are apparently formed in the presence of basic proteins in salt concentrations over 40 mM (Olins and Olins 1971; Man- ning 1979). The dilute hypophase requirement is explainable by the assumption that dilution and spreading effects un- fold a compact precursor. Occasionally we have detected structures that appear to be intermediates in the process of doughnut unfolding and that illustrate a procedure that may give rise to rosettes.
Introduction
DNA rosettes were first reported by Kleinschmidt and Zahn (1959) who observed them after spreading DNA from fish and bacterial ceils on an air-water film of denatured, basic protein. Kleinschmidt and collaborators (1959, 1968) considered rosettes to be artifacts of the procedure, since they disappeared after the molarity of the hypophase was raised. Several investigators (Inman 1967; Davis et al. 1971) have since agreed that the structure is an artifact induced by spreading DNA with cytochrome C over dilute hypo- phases, but others (Sonnenbichler 1969a, b; Comings and Okada 1976; Okada and Comings 1979) have proposed that rosettes represent packaging domains of the chromatin fiber and have presented evidence for the origin of rosettes from the intrinsic periodicity in the fiber.
The presence of loops in dehistonized eucaryotic chro- mosomes was independently detected by sedimentation stu- dies using sucrose gradient centrifugation of dehistonized metaphase chromosomes and interphase nuclei in the pres- ence of ethidium bromide (Cook and Brazell 1975; Benya-
jail and Worcel 1976; Adolph etal. 1977; Paulson and Laemmli 1977). As the concentration of ethidium bromide in the solution is increased, sedimentation behavior typical of covalently closed circular molecules is observed. This suggests that the chromosomal fiber is anchored at distant sites and prevented from rotation. Surface spreading of de- histonized metaphase chromosomes reveals 10- to 30-gm loops anchored to a proteinaceous scaffold; rosettes are not evident in these preparations (Paulson and Laemmli 1977; Hadlacsky et al. 1981 ; Mullinger and Johnson 1980). Loops in sedimentation studies are estimated to range in size from 30 to 90 kilobase pairs (kb) in most reported cases (Benyajati and Worcel 1976; Paulson and Laemmli 1977). Individual loops in rosettes are much smaller (1 5 kb) (Okada and Comings 1979; this paper).
Several cytological studies with light and electron mi- croscopy reveal loops in whole-mount and sectioned chro- mosomes, from a wide variety of eucaryotes (Sorsa 1973; Comings and Okada 1973). The possibility that DNA ro- settes correspond on a cytological level to chromomeres was first proposed by Sonnenbichler (1969).
We studied the structure of chromatin from the nuclei of amphibian red blood cells (RBCs) by centrifugation, elec- trophoresis, and spreading procedures for electron micros- copy in an attempt to clarify the origin of rosettes and other aggregates. We found that several types of aggregates could be produced, depending on the state and source of the DNA, the basic-protein requirements, and the type of hypophase used. From our results we conclude that at least some rosettes form during cytochrome C spreading of the DNA over dilute hypophases, as Kleinschmidt proposed.
Materials and methods
Organisms used. The toad Bufo marinus, collected in San Jos6, was kept in shallow open aquaria and the salamander Bolitoglossa subpalmata, from the Cerro de la Muerte, lo- cated along the Pan American Highway south of San Jos~, was kept at 18 ~ C in moss terraria. The toads were anesthe- tized with diethyl ether; the salamanders, with a dilute chloretone solution.
Isolation of nuclei. RBCs obtained by heart puncture were lysed in ice-cold 0.1 x SSC (SSC is 0.15 M NaC1, 0.015 M sodium citrate, pH 6.8) for 30-60 min. Lysis was monitored by phase-contrast microscopy. Nuclei from lysed cells were pelleted by centrifugation (2,000 rpm for 15 min in a Hita-

308
chi 18PR-3 refrigerated centrifuge with an RP16 rotor, ap- proximately 400 g). When large volumes of lysis buffer are used (1 : 1000 dilution of whole blood), very efficient lysis (> 98%) can be produced.
Preparation ofchromatin. Nuclei were resuspended in small volumes of either: (a) Worcel's lysis medium (0.4% Nonidet NP-40, 0.9 M NaC1, 0.05 M Tris-HC1, pH 8; Benjayati and Worcel ]976); (b) 1 or 2 M NaC1, 0.05 M Tris-HC1 (pH 8), 0.1 M EDTA (ethyldiaminetetraacetic acid); or (c) 0.15 M NaC1, 0.05 M Tris-HC1 (pH 8.0), 0.1 M EDTA for immediate spreading with formamide and cytochrome C. Alternatively chromatin from nuclei resuspended in concen- trated salt solutions was precipitated with 2.5 volumes of cold ethanol, spooled onto glass rods, washed with 70% ethanol, and stored in 70% ethanol at - 2 0 ~ C. To obtain clean DNA the lysed nuclear suspension was digested for 2 h with self-digested Proteinase K (500 gg/ml, Merck), ex- tracted with redistilled phenol and chloroform :isoamyl al- cohol (25: I), and precipitated with ethanol as before, or dialyzed against the spreading buffer. The isolation of nu- cleic acids from Shigella was carried out on a strain of the bacterium kindly provided by Elena Pefiaranda, using procedures described elsewhere (Croza et al. 1977).
Enzyme treatment. Precipitated chromatin was dissolved in 0.1 M Tris-HC1, 0.05 M NaC1, 0.01 M MgC12 (pH 7.5) and treated with trypsin (100 ~tg/ml, Sigma Type III), pancreatic RNAse A (100 gg/ml, Sigma Type IIA) or TI RNAse (20 units/ml, Sigma Type V) for 2 h at 37 ~ C. To obtain nicked molecules, DNase I (Sigma Type I) was used at a concentra- tion of 0.5-1.0 gg/ml for 1 h at 4 ~ C before spreading at 50 gg/ml DNA concentration. Trypsin was self-digested briefly before use, and RNAse A was placed in a boiling waterbath for 5 min to denature contaminating DNAse. The restriction enzyme Eco RI was purchased from Miles Biochemicals and used under the digestion conditions re- commended by the supplier.
Shearing procedures. Resuspended nuclei were sheared at maximum speed for 4 min with a Sorvall Omnimixer, using the high-speed microadaptor with a 4-ml capacity in an ice bath. Then they were spread directly after the addition of an equal volume of 99% recrystallized formamide (J.T. Baker) or gently extracted with chloroform: isoamyl alcohol before spreading.
Formamide spreading. We followed the procedures of Davis et al. (1971) using collodion film coated with carbon, and acid-cleaned slides as ramps on plastic Petri dishes. All solu- tions were prepared with Millipore-filtered, glass-distilled water. Spreading was done after mixing equal volumes of recrystallized 99 % formamide with either (a) purified nuclei in Worcel's lysis medium; (b) chromatin resuspended in Worcel's lysis medium with ethidium bromide (50 gg/ml final concentration) 15 min before mixing with formamide; (c) enzyme digestion mixtures after the addition of the ap- propriate volume of a 10 x stock "spreading buffer" (Davis et al. 1971); or (d) samples from the aqueous phase after phenol or chloroform extraction, diluted at least 20 times.
After the nucleic acid was carefully mixed with form- amide, either bovine serum albumin (BSA; Sigma Fraction V powder), ribonuclease A (heat-treated, Sigma Type IIIA), or cytochrome C (Sigma Type VI from bovine heart) was
added to a final concentration of 0.1% and spread over a glass-distilled water or salt hypophase. The grids were stained with alcoholic uranyl acetate or rotary-shadowed with platinum:paladium (80:20) at a 7 ~ angle in a Hitachi HUS-5 high-vacuum evaporator.
Preparations were observed and photographed with a Hitachi HU12A electron microscope at 75 kV. Measure- ments were taken with an opisometer on enlarged photo- graphs and calibrated with respect to two circular plasmids of molecular weight 2.2 x 1 0 6 and 4.1 x 1 0 6 (Croza et al. 1977) present in the Shigella strain used.
Polyacrylamide gel electrophoresis and ultraviolet spectra. The proteins that remain associated with the extracted chro- matin, were separated in SDS (sodium dodecyl sulfate) polyacrylamide gels, following the method of Laemmli (1970). Nuclear preparations were suspended in a small vol- ume of "sample buffer" with 2% SDS, heated to 90~ for 5 rain, and separated in the Tris-glycine discontinous system, using 3% and 10% acrylamide for the stacking and separating gels, respectively (Laemmli 1970).
The ultraviolet absorption spectrum of the different chromatin preparations was determined with a Model 200-20 dual-beam Hitachi spectrophotometer in 0.15 M NaC1, 0.05 M Tris-HC1 (pH 8) buffer with the appropriate standards. The protein content of the same chromatin was assayed by the method of Lowry et al. (1951).
Results
The effect of several spreading conditions on the appear- ance of rosettes in chromatin and DNA of the amphibians Bufo marinus and Bolitoglossa subpalmata and several other organisms is presented in the following sections.
Hypophase effects
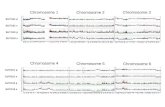
We found that the composition of the hypophase is crucial for rosette formation. Distilled-water hypophases favor the appearance of rosettes even when the composition of the spreading solution is altered. Spreading nuclear material from Worcel's lysis medium is in our hands the most reliable procedure for the production of large symmetric "typical rosettes". These have a single central nucleating region where loops converge, often with only two free ends, indi- cating that they are intramolecular aggregations (Fig. 1 a, b). Larger rosettes with several nucleating regions are not uncommon in the undigested nuclear preparations. Loops in these rosettes do not appear supercoiled under most spreading conditions (Fig. I a, b). Raising the molarity of the hypophase above 0.15 M ammonium aceate abolishes the "typical rosettes," although other structures are still found (Fig. I c, and section below).
Changes in the state and origin of the DNA
The state of the DNA at the time of spreading appears to affect its capacity to form rosettes on distilled-water hy- pophase. The largest rosettes were obtained with nuclei sus- pended in Worcel's lysis medium and spread with forma- mide and cytochrome C (Fig. 1 a), with minimal shearing.
Nuclei that have been extensively digested with protein- ase K or trypsin and spread immediately over water hypo- phase also produce large symmetric rosettes, which are

90
309
70
SO
~_ 30 o o o
70
70 _Q .C~
E E 2 50 2
30
10
0 1 2 3 L/am 0 1 2 3 4pro 5
Fig. 2. Size distribution of loops in rosettes measured from enlarged photographs of preparations from (a) Bufo marinus (n=439; 2= 0.93 _+ 0.26 gin) and b Shigella (n = 188 ; :~ = 0.54 _+ 0.50 gm) and two Shigella plasmids used for calibration. Length distribution of (e) the free ends (n = 234; 2 = 0.43 4-0.24 gin) and (d) the linear mole- cules (n = 342; ~ =0.63 • 0.49 gm) in Eco RI-digested DNA from Bolitoglossa subpalmata RBCs spread over distilled-water hypo- phase
sometimes par t of extremely long molecules ( > 2 0 0 gm). Large, symmetric rosettes are also obtained using undi- gested nuclei resuspended in lysis medium or in 2 M NaC1 and spread directly with cytochrome C.
"Typ ica l roset tes" have an average of 2 7 + 12 loops, which generally appear relaxed (Fig. 1 a, b). The size distri- but ion of loops in these rosettes is presented in the histo- gram of Figure 2 a. Chromat in from Bufo and Bolitoglossa RBCs produce indist inguishable rosettes. Treat ing nuclei with high concentrat ions of r ibonuclease does not affect the appearance o f " typ ica l rosettes."
Very long molecules are recovered if carefully handled nuclei are spread immediately. The fiber in large rosettes can be over 100 pm in length. "Typica l roset tes" have ap- proximate ly 30_+ 12 pm D N A , which corresponds to a mo- lecular weight of about 60 x 106 daltons. Adjacent rosettes are separated by fibers that measure from a few to over 25 gm in length (Fig. 3 a).
Shearing of the D N A from these animals decreases the size of the loops and rosettes recovered (Fig. 1 b). I t is also possible to observe rosettes in D N A that has been digested with proteinase K and repeatedly extracted with chloroform and phenol. These rosettes have fewer and smaller loops (Fig. 3 b), and often have several free ends.
Extensive shearing of the D N A with an Omnimixer pro- duces even smaller rosettes with many free ends, which we have called the "as ter isk type" . Digestion with Eco RI also gives rise to "as te r i sk - type" rosettes (Fig. 3c). Loops and free l inear molecules were measured in prepara t ions (Fig. 2c, d) that were extensively digested with Eco RI under condit ions in which it recognizes the hexanucleotide
Fig. 1. Spreading of DNA preparations from Bufo marinus under various conditions. DNA concentration approximately 5 gg/ml, a '~ Typical rosette" from Bufo marinus RBC nuclei with cytochrome C (50 pg/ml) and 50% formamide, spread directly over a distilled-water hypophase. Upper rosette with two nucleating sites, lower rosette with one. b Spread as in (a) but using deproteinized DNA. Many rosettes have two long free ends; this one has, in addition, two short free ends. (e) Spread of deproteinized DNA as in (a) but using a 0.2 M ammonium acetate hypophase. An area without doughnuts is shown. Bars represent 0.5 pm

310
Fig. 3. Bolitoglossa DNA (5 gg/ml) spread over a distilled-water hypophase, a Two adjacent rosettes observed in a nuclear spread. The interrosette DNA is 15 gm long. b Rosettes observed in sheared, deproteinized DNA; three doughnuts are also present, e DNA restricted with Eco RI shows many "asterisk-type" rosettes. Bars represent 0.5 gM
sequence 5 ' -GAATTC (Roberts 1980). Thus in a random sequence this restriction endonuclease cleaves approxi- mately every 4 kb. "Aster isk-type" rosettes contain an av- erage of 15 loops with 25 kb D N A and should be nicked at least six times (40% density of nicks), if composed of random sequences. The density of nicks gives the percentage of nicked loops out of the total number of loops. Table 1 shows that in a sample of 50 rosettes, most (96%) have a density of nicks lower than that (40%) expected for a random nucleotide sequence. The mean length of nicked
Table 1. Density of nicked loops expressed as the percentage of nicked to total loops. The number of nicked loops was determined by substracting 2 from the number of fi-ee ends (since unnicked loops usually have two free ends) and dividing the remainder by two. A total of 748 loops were scored in 50 rosettes for an average number of 15 loops per rosette
Density Percent of nicked loops of rosettes
0-5 24 5-10 10
10-15 10 15 20 18 20-25 16 2~30 8 30-35 4 35-40 6 4045 4 45-50 0
loops (Fig. 2c) suggests that most loops are nicked only once, since multiple nicks in many loops would lower this value to under half the length o f whole loops. Measure- ments of free linear molecules present alongside the nicked rosettes (Fig. 2d), reveal fibers up to 10 gm in length, as well as molecules much shorter than expected, including a size class of approximaely 0.08 gm, abundant in number but insignificant in mass. Presumably these results are due to the abundance of tandem repetitive sequences, most of them lacking the restriction sequence.
Nicking the D N A briefly with DNAse I, or by keeping it at room temperature for several days, gradually abolishes the appearance of loops and rosettes in all hypophases (Fig. 4 a). Spreading clean D N A under the partially dena- turing conditions described by Moreau et al. (1981), pro- duces small rosettes with double-stranded molecules. Long single-stranded molecules in the same preparations do not form loops or aggregates (Fig. 4b). These observations sug- gest that the duplex molecule is necessary for rosette forma- tion.
Rosettes are formed by D N A from many sources other than amphibian RBCs, including the bacteria Escherichia coli and Shigella (Fig. 5), rat liver, commercial calf thymus, and human lymphocytes. Loops in bacterial rosettes are slightly smaller in length (Fig. 2 b) than those from amphibi- an sources.
Some of the plasmid circles from Shigella appear re- laxed; others are supercoiled into structures resembling small rosettes without free ends. We assume that the relaxed forms are nicked molecules. No D N A concentration effects were detected at 0.5-5 gg/ml.

Fig. 4. a Deproteinized DNA from Bufo marinus was (a) briefly nicked with DNAse I and spread with cytochrome C over a dis- tilled-water hypophase, b DNA was spread under the denaturing conditions described by Moreau et al. (1981). The long fiber has the beaded appearance of single-stranded molecules typical for such preparations, whereas the double-stranded fibers are thinner and sometimes resemble rosettes. The consistent presence of com- pact double-stranded structures and extended single-stranded fibers is noteworthy. Bars represent 0.5 gm
Changes in the composition of the spreading solution
Formamide is not essential for rosette formation, since ro- settes appear with aqua-spreading procedures from 2 M ammonium acetate. The detergent Nonidet can also be omitted from the spreading solution without affecting ro- sette formation. When cytochrome C is replaced by pancre- atic ribonuclease A or bovine serum albumin (BSA), ro- settes are not found, although doughnuts and other com- pact structures are still present (see next section).
311
After ethidium bromide is added to the spreading solu- tion, distinct aggregates with highly supercoiled loops ap- pear, with either low or high salt hypophases. These aggre- gates differ from the typical rosettes, since they usually lack a central nucleating site where loops converge. Relaxed loops are sometimes evident among the supercoiled ones.
Other aggregates
Stems and doughnut-shaped objects, essentially identical to those described by Olins and Olins (1971), are found in many of the spreads with cytochrome C and BSA on all hypophases used (Figs. 3a, 6a). Stems are of variable length; doughnuts have an inner diameter of 0.08 0.16 gm and an outer diameter of 0 .1~0.40 gin. Both structures have been observed in D N A that was extensively deprotein- ized and centrifuged to equilibrium in CsC1. Doughnuts are less prevalent in spread nuclei. Occasionally we found partially unraveled stems and doughnuts, revealing an or- dered packing of single, long D N A molecules (Fig. 6b-d). When resuspended nuclei are used immediately, spreading in all hypophases produces "scaffold like" fragments, which are absent if the D N A is deproteinized. The loops of the scaffolds are considerably longer than in typical ro- settes. Although their size is difficult to measure, most of them have a circumference of well over 10 ~tm. Scaffolds were observed regardless of the hypophase utilized for spreading, as long as undigested nuclear preparations were usd. In dilute hypophases rosettes and doughnuts are some- times seen in association with scaffolds.
Protein analysis
High salt and chloroform extraction of the RBC nuclei from these amphibians does not remove all proteins from the DNA. Lowry protein determinations of ethanol-precipi- tated D N A or of D N A lyophilized after extensive dialysis gave positive reactions, and the UV spectrum was typical of nucleoprotein (Table 2). The results of disc gel electro- phoresis with SDS showed that salt and chloroform extrac- tion selectively removed proteins from the chromatin, leav- ing behind two or three polypeptides that range in molecu- lar weight f rom 68,000 to 80,000 daltons. Nuclei suspended in Worcel 's lysis medium retained a strong protein band that comigrates with histone H4 and a minor band corre- sponding to H3, in addition to the high-molecular-weight
Fig. 5. DNA from Shigella obtained from a CsCl gradient and spread with cytochrome C over a water hypophase. This material contains the plasmid circles that were measured and included in the histogram in Figure 2b. Bar represents 0.5 I~m

312
Table 2. Absorption spectra from 350 to 220 nm were taken in standard l-cm pathlength quartz cuvettes using appropriate blanks. From the spectra the ratios indicated were obtained, uncor- rected for light scattering. Data presented are for Bufo marinus RBC preparations, indicating the mean and the standard deviation for 8 separate measurements. For Lowry protein determinations the material was first precipitated with ethanol and spooled onto glass rods before resuspension in the appropriate buffer. Protein content is expressed as micrograms percent with respect to DNA
Absorption Whole Dehistonized Clean ratio nuclei chromatin DNA
260/230 1.07_+ 0 . 0 9 1.12_+0.09 1.90_+0.30 260/280 1.61_ 0.09 1.71 +0.09 1.80+_0.05 260/320 50 +38 125 -+33 200 +47
Protein content 50-65 3-5 < 0.005
components. Chromatin suspended in 2 M NaC1 loses all traces of histomes although the high-molecular-weight pro- teins are not removed.
Nuclei treated with proteinase K and repeatedly extract- ed with phenol did not give positive Lowry reactions, had a UV spectrum typical of pure DNA (Table 2), and pro- duced no definitive bands by SDS polyacrylimide gel elec- trophoresis.
Fig. 6. Occasionally, a single deproteinized DNA preparation from Bufo marinus spread over distilled water, showed DNA doughnuts in various states of unravelling (a-c) and well-spread rosettes (d). Bars represent 0.5 gm
Discussion
The packaging of the DNA molecule in eucaryotic chromo- somes and chromatin was considerably clarified by the dis- covery of the nucleosomal organization of histones and the detection of higher-order structures mediated' by histone H1 (for review see McGhee and Felsenfeld 1980; Thoma and Koller 1981). Is there a higher level of chromosomal packaging that is not mediated by the histones in eukar- yotes ? Several papers cited in the introduction suggest that removal of histones does not completely destroy long-range organization of chromosomes and chromatin. Presumably nonhistone proteins mediate such interaction, although the precise way in which this is accomplished is not understood. Scaffolds (Adolph etal. 1977), rosettes (Sonnenbichler 1969; Okada and Comings 1979), and other arrangements (Sorsa 1972; Mullinger and Johnson 1980) have been pro- posed for the dehistonized DNA fiber.
Rosettes formally represent an effective way of packag- ing long DNA fibers while maintaining their long-range uninemy. An attractive feature of this packaging proposal is the simple way it explains the organization of chromo- meres and, by rosette fusion, the C, G and Q bands in metaphase chromosomes (Okada and Comings 1979).
The evidence for and against the existence of rosettes in chromosomes is summarized by Okada and Comings (1979). Our results contribute to this debate in several ways.
As evidence for rosettes being real biological structures, Okada and Comings (1979) cite the nonrandom distribution of loops, rosettes, and interrosette length in DNA from Chinese hamster cells. Our data from Bufo and Bolitoglossa agree well with their measurements. Furthermore, bacterial and clean-DNA rosettes have fewer and smaller loops than those from spread nuclear material. The largest symmetrical rosettes are produced by immediately spreading nuclei

313
under conditions in which the long D N A molecules remain unsheared.
Our results contribute four lines of evidence that rosettes are artifacts. (1) Rosettes are found using the same proce- dures with D N A from bacterial sources. (2) Chromatin ex- tensively digested with proteinase K and repeatedly extract- ed with chloroform and phenol can produce rosettes. (3) Rosettes are not abundant unless dilute hypophases are used. (4) Albumin and ribonuclease A substitution of cyto- chrome C for spreading does not favor the appearance of rosettes, regardless of the hypophase. Furthermore, prelimi- nary work with chromatin using BAC spreading (Cole et al. 1978) and Dubochet adhesion (Dubochet et al. 1971) has not clearly revealed rosettes (Bonilla and Le6n in prepara- tion).
How are rosettes produced? We can envision at least three different general mechanisms: (1) Rosettes could be remnants of a native structure that emerges when chromatin is depleted of histones under conditons that do not remove all chromosomal proteins. In this case rosettes could, for instance, be morphologically related to chromomeres. (2) Rosettes could be formed during the procedure in specific regions of the fiber by complexes of bound proteins and cytochrome C. In this case rosettes would not necessarily be found in vivo, but their study could still reveal interesting properties of the bound chromosomal proteins. (3) Rosettes could be unspecific artifacts due to intrinsic properties of the basic pro te in-DNA complex.
Polyelectrolyte theory (Manning 1979; Olins and Olins 1971; Belmont and Nicolini 1981) suggests a mechanism that is consistent with most of our results, including the dilute hypophase requirement and the extensive occurrence of doughnuts in this preparation. Charge neutralizaton of the D N A in solution presumably induces the formation of compact structures that can be detected by various physi- co-chemical procedures. These compact structures, which appear in electron micrographs as doughnuts and strands (Olins and Olins 1971; Fisher and Williams 1979; this paper) do not occur below a narrow salt transition (20-40 mM). Thus, an expanded structure presumably emerges when the D N A is transferred to a dilute environ- ment below this salt transition. We suggest that at least some of the structures that we have observed result from the unfolding of compact aggregates formed in high salt in the presence of basic protein, as indicated by the observa- tion of intermediate forms between the compact doughnut and the open rosette. It is possible that we have observed several types of rosettes of different origin; however, we could not identify consistent differences in the nucleating centers that might be characteristic of different types of rosettes.
We have estimated that our " typ ica l" rosette obtained from nuclear spreadings contains 21 l~m D N A (42x 106 daltons) approximately 60 kb. We ignore the D N A content of the pachytene chromomeres in Bufo and Bolitoglossa, but judging from published data on salamander chromo- somes (Vlad and Macgregor 1975; Lima-de-Faria 1972) one would expect five to ten times more D N A in salamander pachytene chromomeres.
The analysis of the restricted rosettes indicates that they contain diverse nucleotide sequences. It also agrees with the finding of extensive interstitial repetition in the chromo- somes of these large-genome salamanders (Mizuno and Macgregor 1974; Macgregor et al. 1976). The abundance
of very small restricted molecules, contrasts with their virtu- al absence in mechanically sheared preparations (data not shown). It also indicates that digestion was complete.
Although we cannot conclude that rosettes have no bio- logical relevance, it remains to be proved that they represent biological structures instead of artifacts. We suggest that cytochrome C and conditions that suddenly change the ionic environment of the polynucleotide should be avoided in this type of study.
Acknowledgements. We would like to acknowledge the technical help of Cesar F. Bonilla and Oscar Chavarria. We also extend our gratitude to Reynaldo Pereira for help with the illustrations, to Flora Gonzfilez for typing the manuscript, and to the staff of the Electron Microscopy Unit. This work was financed with grants from CONICIT (Consejo Nacional de Investigaciones Cientificas y Tecnol6gicas) and the Vice-rectory of Research, University of Costa Rica.
References
Adolph KW, Cheng SM, Paulson JR, Laemmli UK (1977) Isola- tion of a protein scaffold from mitotic HeLa cell chromosomes. Proc Natl Acad Sci 74:4937-4941
Belmont A, Nicolini C (1981) Polyelectrolyte theory and chromatin - DNA quaternary structure. J Theor Biol 90:169-179
Benyajati C, Worcel A (1976) Isolation, characterization, and structure of the folded interphase genome of Drosophila mel- anogaster. Cell 9 : 393-407
Cole MD, Beer M, Koller Th, Strycharz WA, Nomura N (1978) Electron microscopic determination of the binding sites of ribo- somal proteins $4 and $8 on 16S RNA. Proc Natl Acad Sci 75 : 270-274
Comings DE, Okada TA (1973) Some aspects of chromosome structure in eukaryotes. Cold Spring Harbor Syrup Quant Biol 38:145-153
Comings DE, Okada TA (1976) Nuclear proteins. III. The fibrillar nature of the nuclear matrix. Exptl Cell Res 103:341-360
Cook PR, Brazell IA (1975) Supercoils in human DNA. J Cell Sci 19:261 279
Croza JH, Olarte J, Mata LJ, Lutthropp LK, Pefiaranda E (1977) Characterization of an R-plasmid associated with ampicillin resistance in Shigella dysenteriae Type 1 isolated from epidem- ics. Antimicrobial Agents and Chemotherapy 11:553-558
Davis RW, Simon MN, Davidson N (1971) Electron microscope heteroduplex methods for mapping regions of base sequence homology in nucleic acids. In: Grossman L, Moldave K (eds) Methods in Enzymology 21:413 428
Dubochet J, Docommun M, Zollinger M, Kellenberger E (1971) A new preparation method for dark-field electron microscopy of biomolecules. J Ultrastruct Res 35:147-167
Fisher HW, Williams RC (1979) Electron microscopic visualization of nucleic acids and their complexes with proteins. Ann Rev Biochem 48 : 649-679
Hadlacsky G, Sumner AT, Ross A (1981) Protein-depeleted chro- mosomes. I and II. Chromosoma 81 : 537-567
Inman RB (1967) Some factors affecting electron microscopic length of deoxyribonucleic acid. J Mol Biol 25:209-216
Kleinschmidt A (1968) Monolayer techniques in electron microsco- py of nucleic acid molecules. In: Grossman L, Moldave K (eds) Methods in Enzymology 12 B : Academic Press. New York and London, pp 361-377
Kleinschmidt A, Zahn RK (1959) Uber Desoxyribonucleinsfiure- molekulin in Protein-Mischfilmen. Naturforsch b 14:770-778
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-685
Lima-de-Faria A (1973) The molecular organization of the chro- momeres of Acheta involved in ribosomal DNA amplification. Cold Spring Harbor Syrup Quant Biol 38:559-571

314
Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the Folin phenol reagent. J Biol Chem 193 : 265
Macgregor HC, Mizuno S, Vlad M (1976) Chromosomes and DNA sequences in salamanders. Chromosomes Today 5:331-339
Manning GS (1979) Theory of Hi-mediated control of higher orders of structure in chromatin. Biopolymers 18:2929-2942
McGhee JD, Felsenfeld G (1980) Nucleosome structure. Ann Rev Biochem 49 : 1115-1156
Mizuno S, Macgregor HC (1974) Chromosomes, DNA sequences and evolution in salamanders of the genus Plethodon. Chromo- soma 48:239-296
Moreau J, Matyash-Smirniaguina L, Scherrer K (1981) Systematic punctuation of eucaryotic DNA by A + T-rich sequences. Proc Natl Acad Sci 78:1341-1345
Mullinger AM, Johnson RT (1980) Packing DNA into chromo- somes. J Cell Sci 46:61-86
Okada TA, Comings DE (1979) Higher-order structure of chromo- somes. Chromosoma 72:1-14
Olins DE, Olins AL (1971) Model nucleohistones: the interaction of F1 and F2al histones with native T7 DNA. J Mol Biol 57:437-455
Paulson JR, Laemmli UK (1977) The structure of histone-depleted metaphase chromosomes. Cell 12: 817-828
Roberts RJ (1980) Directory of restriction endonucleases. In: Grossman L, Moldave K (eds) Methods in Enzymology 65:1-15
Sonnenbichler J (1969a) Substructure of chromosomes. Nature 223 : 205-206
Sonnenbichler J (1969b) Nucleoprotein complexes: possible sub- units of chromosomes. Hoppe-Seyler's Z Physiol Chem 350:761-766
Sorsa V (1973) Organization of chromomeres. Cold Spring Harbor Symp Quant Biol 38:601-608
Thoma F, Koller Th (1981) Unravelled nucleosomes, nucleosome beads and higher-order structures of chromatin: Influence of non-histone components and histone H1. J Mol Biol 149: 709-733
Vlad M, Macgregor HC (1975) Chromomere number and its genet- ic significance in Lampbrush chromosomes. Chromosoma 50: 327-347
Received November 11, 1982 / in revised form April 20, 1983 Accepted by H.C. Macgregor