
Projections of the lateral reticular nucleus to the cochlear
Upload
charles-watsonCategory
view
212download
0
ORIGINAL ARTICLE
Projections from the lateral vestibular nucleus to the spinal cordin the mouse
Huazheng Liang • Timea Bacskai • Charles Watson •
George Paxinos
Received: 19 December 2012 / Accepted: 27 February 2013
� Springer-Verlag Berlin Heidelberg 2013
Abstract The present study investigated the projections
from the lateral vestibular nucleus (LVe) to the spinal cord
using retrograde and anterograde tracers. Retrogradely
labeled neurons were found after fluoro-gold injections into
both the cervical and lumbar cord, with a smaller number
of labeled neurons seen after lumbar cord injections.
Labeled neurons in the LVe were found in clusters at
caudal levels of the nucleus, and a small gap separated
these clusters from labeled neurons in the spinal vestibular
nucleus (SpVe). In the anterograde study, BDA-labeled
fiber tracts were found in both the ventral and ventrolateral
funiculi on the ipsilateral side. These fibers terminated in
laminae 6–9. Some fibers were continuous with boutons in
contact with motor neurons in both the medial and lateral
motor neuron columns. In the lumbar and sacral segments,
some collaterals from the ipsilateral vestibulospinal tracts
were found on the contralateral side, and these fibers
mainly terminated in laminae 6–8. The present study
reveals for the first time the fiber terminations of the lateral
vestibular nucleus in the mouse spinal cord and therefore
enhances future functional studies of the vestibulospinal
system.
Keywords Hindbrain � Vestibulospinal tract �Spinal cord � Motor neurons � Vestibular system
Abbreviations
2SpL Lamina 2 of the spinal gray, lateral part
3Sp Lamina 3 of the spinal gray
4Sp Lamina 4 of the spinal gray
4V 4th ventricle
5SpM Lamina 5 of the spinal gray, medial part
5SpL Lamina 5 of the spinal gray, lateral part
6Sp Lamina 5 of the spinal gray
7n Facial nerve
7N Facial nucleus
7Sp Lamina 7 of the spinal gray
8Sp Lamina 8 of the spinal gray
10Sp Area 10 of the spinal gray
Ax9 Axial muscle motoneurons of lamina 9
Bi9 Biceps motoneurons of lamina 9
C4 4th cervical segment
C5 5th cervical segment
C6 6th cervical segment
CC Central canal
contra Contralateral
cu Cuneate fasciculus
DAB 3,30-Diaminobenzidine
DC Dorsal cochlear nucleus
De9 Deltoid muscle motoneurons of lamina 9
dcs Dorsal corticospinal tract
dr Dorsal root
Fl Flocculus
GAD Glutamic acid decarboxylase
Gi Gigantocellular reticular nucleus
icp Inferior cerebellar peduncle
IMM Intermediomedial column
IntA Interposed cerebellar nucleus, anterior part
H. Liang and T. Bacskai made an equal contribution to this work.
H. Liang � T. Bacskai � C. Watson � G. Paxinos (&)
Neuroscience Research Australia, Cnr Barker Street
and Hospital Road, Randwick, NSW 2031, Australia
e-mail: [email protected]
C. Watson
Faculty of Health Sciences, Curtin University,
Perth, WA 6845, Australia
G. Paxinos
School of Medical Sciences, The University of New South
Wales, Sydney, NSW 2052, Australia
123
Brain Struct Funct
DOI 10.1007/s00429-013-0536-4

IO Inferior olive
ipsi Ipsilateral
L3 3rd lumbar segment
L4 4th lumbar segment
L6 6th lumbar segment
Lat Lateral cerebellar nucleus
lf Lateral funiculus
LSp Lateral spinal nucleus
LVe Lateral vestibular nucleus
Med Medial cerebellar nucleus
mlf Medial longitudinal fasciculus
MVeMC Medial vestibular nucleus, magnocellular part
MVePC Medial vestibular nucleus, parvicellular part
PFl Paraflocculus
Pr Prepositus nucleus
py Pyramidal tract
rs Rubrospinal tract
scp Superior cerebellar peduncle
SI9 Supraspinatus and infraspinatus motoneurons
of lamina 9
sp5 Spinal trigeminal tract
Sr9 Serratus anterior motoneurons in lamina 9
SuVe Superior vestibular nucleus
T7 7th thoracic segment
VCPO Ventral cochlear nucleus, posterior part, octopus
cell area
vGluT2 Vesicular glutamate transporter 2
vmf Ventromedial fissure
vr Ventral root
vwc Ventral white commissure
Introduction
It has long been known that the lateral vestibular nucleus
(LVe) is involved in locomotion and postural control
through its connections with the spinal cord (Zemlan et al.
1979; Hayes and Rustioni 1981; Leong et al. 1984; Rose
et al. 1992; Masson et al. 1991; Wada et al. 1993; Auclair
et al. 1993; Kudo et al. 1993; de Boer-van Huizen and ten
Donkelaar 1999; Rose et al. 1999; Rice et al. 2010). The
lateral vestibular nucleus makes excitatory monosynaptic
connections with motor neurons of the spinal cord (Wilson
and Yoshida 1969; Grillner et al. 1970; Wilson et al. 1970;
Kasumacic et al. 2010). These vestibulospinal fibers also
have collaterals in the cervical and lower segments to
different degrees (Abzug et al. 1974; Huisman et al. 1984;
Shinoda et al. 1989; Boyle 2000). Boyle (2000) claimed
that certain vestibulospinal fibers projecting to the lumbo-
sacral cord of the squirrel monkey serve as a private
pathway to the motor circuits of the lower limb and tail
with few collaterals in the cervical cord. Other researchers
claim that descending fibers from the lateral vestibular
nucleus have ample collaterals in different segments as
demonstrated with tracer injections (Huisman et al. 1984;
Shinoda et al. 1989; Kuze et al. 1999). In the rat and cat, it
has been shown that fibers from the lateral vestibular
nucleus travel in the ventromedial funiculus (Huisman
et al. 1984; Shen et al. 1990; Rose et al. 1992, 1999;
Bacskai et al. 2002; Matesz et al. 2002) and ventrolateral
funiculus (Rose et al. 1992, 1999; Bacskai et al. 2002;
Matesz et al. 2002). These fibers mainly terminate in the
ventral horn especially in laminae 6–9 (Isu et al. 1988;
Rose et al. 1992, 1999; Bacskai et al. 2002; Matesz et al.
2002). In the mouse, spinal cord projections from the
lateral vestibular nucleus have been demonstrated with
injections of retrograde tracers into the spinal cord
(VanderHorst and Ulfhake 2006; Liang et al. 2011). How-
ever, the detailed fiber distribution of the lateral vestibular
nucleus in the mouse spinal cord is still unknown. We
examined the projections of the lateral vestibular nucleus to
the spinal cord with retrograde and anterograde tracing
techniques. We found that the lateral vestibular nucleus
projects to the gray matter at all levels in spinal cord with
fibers following both the ventral and lateral funiculi, and
terminals present in laminae 6–9. The present study
provides anatomical support for future vestibulospinal
research.
Materials and methods
Animals
Twenty-six C57/BL6 mice of 12 weeks of age, weighing
25–30 g were used. These mice were obtained from the
Animal Resource Centre in Western Australia. All exper-
imental procedures were approved by the Animal Care and
Ethics Committee of The University of New South Wales
(11/75A).
Retrograde tracing
Mice were anaesthetized with an intraperitoneal injection
of ketamine (80 mg/kg) and xylazine (5 mg/kg) and placed
in a mouse stereotaxic head holder (Kopf Instruments,
Tujunga, CA, USA). The ear bars were carefully tightened
and the mouse was stabilized in the stereotaxic holder.
After shaving the fur and sterilizing the skin, spinal cord
segments were exposed by laminectomy at C2 and T12.
The dura mater on the right side of the spinal cord was
penetrated with the tip of a 29-gauge insulin injection
needle and then the needle of a 5-ll Hamilton syringe
(Hamilton Company, Reno, NV, USA; the outer diameter
is 0.711 mm) was driven through this opening. 20–40 nl of
Brain Struct Funct
123

fluoro-gold (FG) (Fluorochrome, Denver, Co, USA; diluted
to 5 % in distilled water) solution was injected through the
needle into the right side of the spinal cord. The needle of
the Hamilton syringe was left in place for 10 min after the
injections. In the present study, ten mice were injected with
fluoro-gold into the upper cervical and upper lumbar seg-
ments (5 mice in each group). In the control group, two
mice received normal saline injections into the spinal cord
and another two mice received fluoro-gold injections into
the cisterna magna. After fluoro-gold injections, the soft
tissue and the skin were sutured and an antibiotic—tetra-
cycline (Pfizer)—was applied topically over the incision.
Buprenorphine (Temgesic, Reckitt Benckiser) solution was
injected to relieve pain.
Anterograde tracing
Mice were anaesthetized as above and positioned in the
stereotaxic instrument. After exposing the skull surface, a
hole was made dorsal to the lateral vestibular nucleus (drill
from Fine Science Tools, North Vancouver, BC, Canada).
10–20 nl of biotinylated dextran amine (BDA) solution
(10,000 MW, Invitrogen) (Bregma -5.88 to -6.24 mm,
midline ?1.25 to ?1.75 mm, surface -2.85 to -3.25 mm)
was injected into the lateral vestibular nucleus of six mice
using the Hamilton syringe described above. In the control
group, two mice received BDA injections into the cisterna
magna and another four into the adjacent vestibular nuclei
and cerebellum. The needle of the Hamilton syringe was left
in place for 10 min after BDA injections and then the skin
was sutured, buprenorphine was injected subcutaneously,
and topical tetracycline was sprayed over the incision.
Tissue preparation
Mice were anesthetized with a lethal dose of pentobarbital
solution (0.1 ml, 200 mg/ml) after 1 week (fluoro-gold
experiments) or 6 weeks (dextran experiments). They were
then perfused through the left ventricle with 60 ml of
0.9 % normal saline containing heparin (150 IU/mouse;
Sigma) using a pump. This was followed by 80 ml of 4 %
paraformaldehyde (Sigma) (in 0.1 M phosphate buffer) and
80 ml of 10 % sucrose solution (in 0.1 M phosphate buf-
fer). Mouse brain and spinal cord were removed and
postfixed in 4 % paraformaldehyde for 2 h at 4 �C, and
then transferred to 30 % sucrose (in 0.1 M PB solution).
After 48 h, brains and spinal cords were cut into 40 lm
thick sections using a Leica CM 1950 cryostat.
Immunohistochemistry
Brain and spinal cord sections from FG injected mice were
washed in 0.1 M PB and transferred to 1 % H2O2 in 50 %
ethanol at room temperature. After 30 min, the sections
were rinsed in 0.1 M PB and treated with 5 % goat serum
in 0.1 M PB to block the non-specific binding sites. The
sections were then incubated with the primary anti-FG
antibody (Chemicon, 1:5,000; raised in rabbit) overnight.
After three rinses in 0.1 M PB, the sections were incubated
with the secondary antibody (biotinylated goat anti-rabbit
IgG; Sigma, 1:200) for 2 h at room temperature. The sec-
tions were washed in 0.1 M PB and then transferred to the
extravidin peroxidase solution (Sigma, 1:1,000). After 2 h,
the sections were rinsed in 0.1 M PB and transferred to the
3,30-diaminobenzidine (DAB) reaction complex (Vector
lab, Burlingame, CA, USA) until the optimal color devel-
oped. The reaction was stopped by transferring the sections
into 0.1 M PB solution, and the sections were mounted
onto gelatinized slides and coverslipped after graded
dehydration and clearing. Sections from the dextran-
injected mouse brain and spinal cord were incubated in
1 % H2O2 in 50 % ethanol for 30 min at room temperature
and then transferred to 5 % goat serum in 0.1 M PB. These
sections were then incubated in the extravidin peroxidase
solution (Sigma, 1:1,000) for 2 h, followed by the DAB
reaction complex, before being mounted and coverslipped
as above.
Data analysis
Mouse brain sections after after visualization of fluoro-gold
injections into the spinal cord were scanned with an Aperio
scanner (ScanScope XT) under 209 magnification. Scan-
ned images were opened with Imagescope software and
images of 49 magnification were extracted and opened
with Adobe Illustrator 5. A new layer was created and then
the profile of sections was drawn. DAB-stained neurons
were mapped with purple dots. Cell numbers were calcu-
lated by counting the dots in every second section through
the entire lateral vestibular nucleus (3 series of sections
from upper cervical and upper lumbar cord injections,
respectively). For display purposes, one dot represents 3–5
labeled neurons in the diagrams. The Imagescope software
was also used to measure the nuclear diameter of labeled
neurons. The estimated cell number of lateral vestibular
neurons was calculated with the corrected Abercrombie
(1946) formula: P ¼ A MMþL. P is the estimated number of
cells, A is the actual count, M is the section thickness, and
L is the nuclear length (L was measured in every second
section). Images of 109 magnification were also extracted
from Imagescope to show the labeled neurons in the areas
of interest.
Mouse brain and spinal cord sections from dextran
injected mice were also scanned with the Aperio scanner.
Images of these sections were then extracted and overlaid
Brain Struct Funct
123

on a spinal cord diagram from the atlas of the mouse spinal
cord (Watson et al. 2009). Adjacent Nissl sections or DAB
stained sections counterstained with Nissl were referred to
in mapping. Images of 49, 59, and 109 were also
extracted to show injection sites and labeled fibers in the
brain and spinal cord.
Results
FG-labeled neurons in the lateral vestibular nucleus
After cervical cord injections of FG, labeled neurons were
first found in the vestibular complex at the level of the
facial genu. These neurons were dorsolateral to the facial
genu and ventral to the caudal part of the locus coeruleus,
occupying the ventral part of the magnocellular part of the
medial vestibular nucleus (MVe) in the mouse brain atlas
(Franklin and Paxinos 2008) (Fig. 1a, b). They were not a
homogenous group, but a mixture of large and small neu-
rons. In more caudal coronal sections, labeled neurons
were found in a wider area mediolaterally and dorsoven-
trally. They were medial to the spinal trigeminal tract and
the inferior cerebellar peduncle, leaving a narrow strip
between these two fiber bundles and the labeled neurons.
Furthermore, there was a small band devoid of labeled
neurons ventral to the superior cerebellar peduncle.
Labeled neurons formed two clusters: one large cluster in
the dorsolateral part of the vestibular complex and another
smaller cluster in the ventromedial part (Fig. 1c–e). The
majority of these neurons were large (30.3 ± 4.0 9
20.6 ± 2.2 lm) and triangular or fusiform. Medial to the
ventromedial cluster, there were some labeled neurons
scattered in the medial vestibular nucleus (MVe). They
were also large neurons. In the caudal pole of the lateral
vestibular nucleus, labeled neurons seemed to form only
one cluster and other labeled neurons were sparsely dis-
tributed in the ventral part of the lateral vestibular nucleus,
Fig. 1 Retrogradely labeled neurons after FG injections into the
cervical cord. a–f Diagrams showing FG-labeled neurons in the
vestibular complex. a0–f0 Photographs showing FG-labeled neurons in
the vestibular complex. From the rostral pole of the lateral vestibular
nucleus to the caudal pole, the number of labeled neurons increases
(a–d, a0–d0) and then wanes (e–f, e0–f0), with a dense cluster of
neurons in the dorsal part of the nucleus and a less dense cluster in the
ventral part. Some labeled neurons are diffusely distributed in the
caudoventral part of the vestibular complex, which might correspond
to the ventral part of the lateral vestibular nucleus. Note that a small
number of labeled neurons are present in the medial part of the
vestibular complex, which corresponds to the medial vestibular
nucleus. The green dashed lines indicate the SuVe boundary, the lightblue dashed lines indicate the MVe boundary and the red dashed linesindicate the LVe boundary. The photomicrograph in a shows the
injection site in the mouse cervical cord. The scale bar for all
photographs is 200 lm
Brain Struct Funct
123
which may correspond to the ventral part of the lateral
vestibular nucleus (Fig. 1f). In total, there were 660 ± 32
FG-labeled neurons.
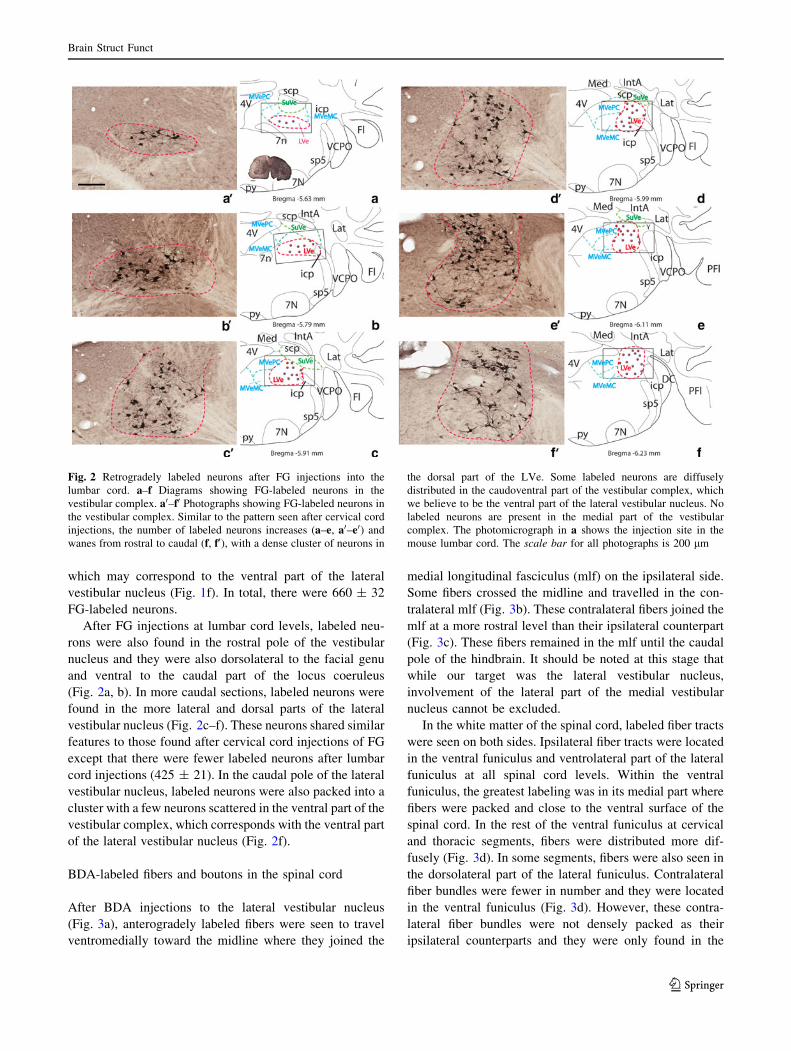
After FG injections at lumbar cord levels, labeled neu-
rons were also found in the rostral pole of the vestibular
nucleus and they were also dorsolateral to the facial genu
and ventral to the caudal part of the locus coeruleus
(Fig. 2a, b). In more caudal sections, labeled neurons were
found in the more lateral and dorsal parts of the lateral
vestibular nucleus (Fig. 2c–f). These neurons shared similar
features to those found after cervical cord injections of FG
except that there were fewer labeled neurons after lumbar
cord injections (425 ± 21). In the caudal pole of the lateral
vestibular nucleus, labeled neurons were also packed into a
cluster with a few neurons scattered in the ventral part of the
vestibular complex, which corresponds with the ventral part
of the lateral vestibular nucleus (Fig. 2f).
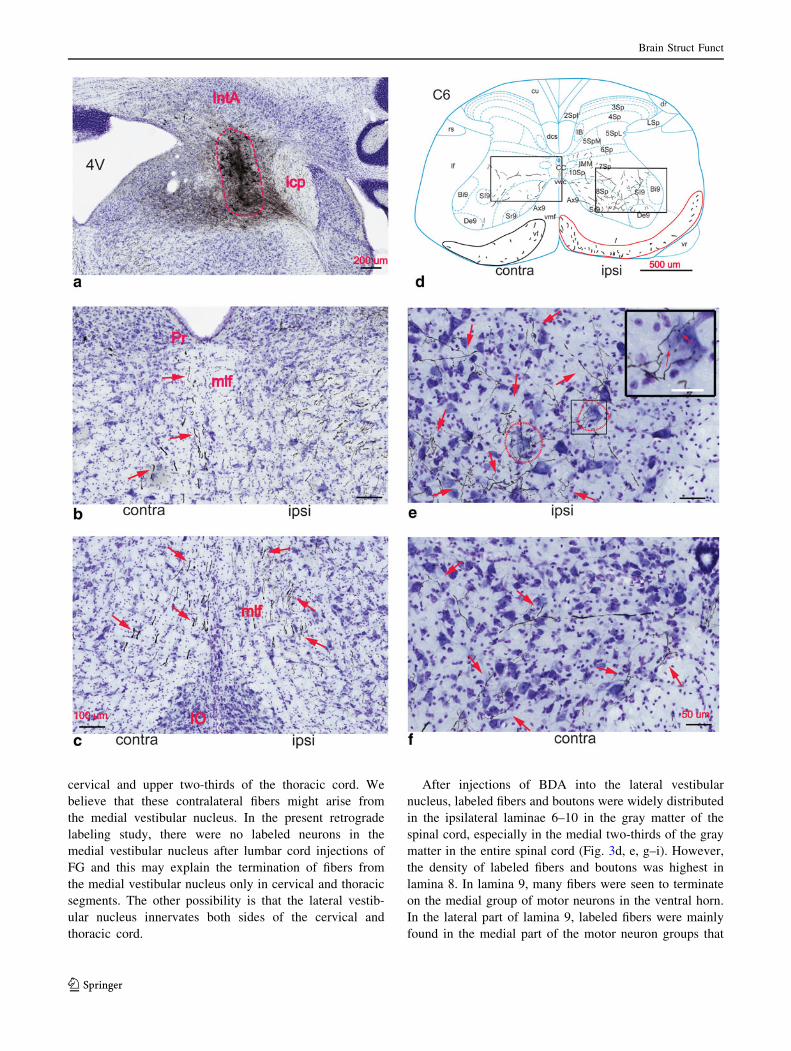
BDA-labeled fibers and boutons in the spinal cord
After BDA injections to the lateral vestibular nucleus
(Fig. 3a), anterogradely labeled fibers were seen to travel
ventromedially toward the midline where they joined the
medial longitudinal fasciculus (mlf) on the ipsilateral side.
Some fibers crossed the midline and travelled in the con-
tralateral mlf (Fig. 3b). These contralateral fibers joined the
mlf at a more rostral level than their ipsilateral counterpart
(Fig. 3c). These fibers remained in the mlf until the caudal
pole of the hindbrain. It should be noted at this stage that
while our target was the lateral vestibular nucleus,
involvement of the lateral part of the medial vestibular
nucleus cannot be excluded.
In the white matter of the spinal cord, labeled fiber tracts
were seen on both sides. Ipsilateral fiber tracts were located
in the ventral funiculus and ventrolateral part of the lateral
funiculus at all spinal cord levels. Within the ventral
funiculus, the greatest labeling was in its medial part where
fibers were packed and close to the ventral surface of the
spinal cord. In the rest of the ventral funiculus at cervical
and thoracic segments, fibers were distributed more dif-
fusely (Fig. 3d). In some segments, fibers were also seen in
the dorsolateral part of the lateral funiculus. Contralateral
fiber bundles were fewer in number and they were located
in the ventral funiculus (Fig. 3d). However, these contra-
lateral fiber bundles were not densely packed as their
ipsilateral counterparts and they were only found in the
Fig. 2 Retrogradely labeled neurons after FG injections into the
lumbar cord. a–f Diagrams showing FG-labeled neurons in the
vestibular complex. a0–f0 Photographs showing FG-labeled neurons in
the vestibular complex. Similar to the pattern seen after cervical cord
injections, the number of labeled neurons increases (a–e, a0–e0) and
wanes from rostral to caudal (f, f0), with a dense cluster of neurons in
the dorsal part of the LVe. Some labeled neurons are diffusely
distributed in the caudoventral part of the vestibular complex, which
we believe to be the ventral part of the lateral vestibular nucleus. No
labeled neurons are present in the medial part of the vestibular
complex. The photomicrograph in a shows the injection site in the
mouse lumbar cord. The scale bar for all photographs is 200 lm
Brain Struct Funct
123
cervical and upper two-thirds of the thoracic cord. We
believe that these contralateral fibers might arise from
the medial vestibular nucleus. In the present retrograde
labeling study, there were no labeled neurons in the
medial vestibular nucleus after lumbar cord injections of
FG and this may explain the termination of fibers from
the medial vestibular nucleus only in cervical and thoracic
segments. The other possibility is that the lateral vestib-
ular nucleus innervates both sides of the cervical and
thoracic cord.
After injections of BDA into the lateral vestibular
nucleus, labeled fibers and boutons were widely distributed
in the ipsilateral laminae 6–10 in the gray matter of the
spinal cord, especially in the medial two-thirds of the gray
matter in the entire spinal cord (Fig. 3d, e, g–i). However,
the density of labeled fibers and boutons was highest in
lamina 8. In lamina 9, many fibers were seen to terminate
on the medial group of motor neurons in the ventral horn.
In the lateral part of lamina 9, labeled fibers were mainly
found in the medial part of the motor neuron groups that
Brain Struct Funct
123

innervate the distal limb muscles (Fig. 3e, g, i). In both the
medial and lateral motor neuron groups, many swellings
presumed to be boutons were seen adjacent to motor neu-
rons (Fig. 3e). In lamina 10, fibers and boutons were pre-
dominantly seen in the area ventral and lateral to the
central canal (CC). Labeled fibers in the dorsal horn were
occasionally seen. In the contralateral gray matter of the
spinal cord, there were fewer labeled fibers and they were
distributed in the gray matter in a pattern similar to that of
ipsilateral fibers, but with a lower density (Fig. 3d, f, g).
These contralateral fiber tracts were only seen in cervical
and upper two-thirds of the thoracic spinal cord. Some
contralateral fibers were also seen in lower lumbar seg-
ments (Fig. 3h). These fibers were sparsely distributed in
laminae 6–8. We believe that they must be the collaterals
of the ipsilateral vestibulospinal fiber tract that crossed the
midline, because there were no labeled fiber bundles on the
contralateral side at these levels.
When BDA was injected into the cisterna magna or
cerebellar cortex, no labeled fibers were found in the spinal
cord. Small injections to the dorsal part of the lateral
vestibular nucleus mainly labeled fibers in the dorsolateral
funiculus (Fig. 4a–c). Large injections (Fig. 4d), which
involved the lateral part of the medial vestibular nucleus
and the superior vestibular nucleus as well as the lateral
nucleus, revealed a pattern of fiber distribution similar to
Fig. 3 continued
Fig. 3 Anterogradely labeled fibers after BDA injections into the
vestibular complex. a The injection site in the vestibular complex,
predominantly in the lateral vestibular nucleus. The dashed-linecircled area is the injection site. b BDA-labeled fiber tracts in the
contralateral medial longitudinal fasciculus at the level of the caudal
pole of the facial nucleus (arrows). c BDA-labeled fibers in bilateral
medial longitudinal fasciculus at the level of the rostral pole of the
hypoglossal nucleus (arrows). d A diagram of BDA-labeled fibers in
the sixth cervical segment. On the ipsilateral side, labeled fiber tracts
are present in the ventral and ventrolateral funiculi (red circled area).
Fibers and boutons are found in laminae 6–10 with predominance in
lamina 8. On the contralateral side, labeled fiber tracts are present in
the ventral funiculus only (black circled area) and they are diffusely
distributed. Labeled fibers and boutons are mainly present in laminae
7 and 8, but the density of these labeled fibers is lower than that of the
ipsilateral fibers. The majority of the fibers are in laminae 7 and 8. e A
higher magnification image of the right rectangular area in
d. Labeled fibers and boutons are present in laminae 8 and 9
(arrows). Some boutons are adjacent to motor neurons (dashed linecircled neurons) in the medial motor neuron group. Note the larger
magnification of the small rectangular area, which shows the boutons
terminating on the cellular area of a motor neuron at the upper rightcorner (arrow). f A higher magnification image of the left rectangular
area in d. Labeled fibers and boutons are present in laminae 7–9
(arrows). g A photograph showing BDA-labeled fiber tracts and
terminals in the seventh thoracic cord segment. Ipsilaterally labeled
fibers are present in both the ventral and ventrolateral funiculi (the redcircled area). The contralaterally labeled fibers are present in the
ventral funiculus only, with an even lower density than their
counterparts in the cervical cord. In the gray matter, fibers and
boutons are distributed similarly as those in the cervical segments
(arrows). h A photograph showing BDA-labeled fiber tracts and
terminals in the sixth lumbar segment. Fiber tracts are only present in
the ipsilateral ventral and ventrolateral funiculi (red circled area).
Fibers and boutons are present in laminae 6–9 with predominance in
lamina 8. A higher magnification image of the rectangular area is
shown in i (arrows). Note that a few fibers are present in the
contralateral gray matter. We believe that they arise from the
ipsilateral tracts. The scale bar for a, g, and h is 200 lm; the scalebar for b, c, and i is 200 lm; the scale bar for d is 500 lm; the scalebar for e and f is 50 lm. The scale bar for the photomicrograph in
d is 25 lm
b
Brain Struct Funct
123
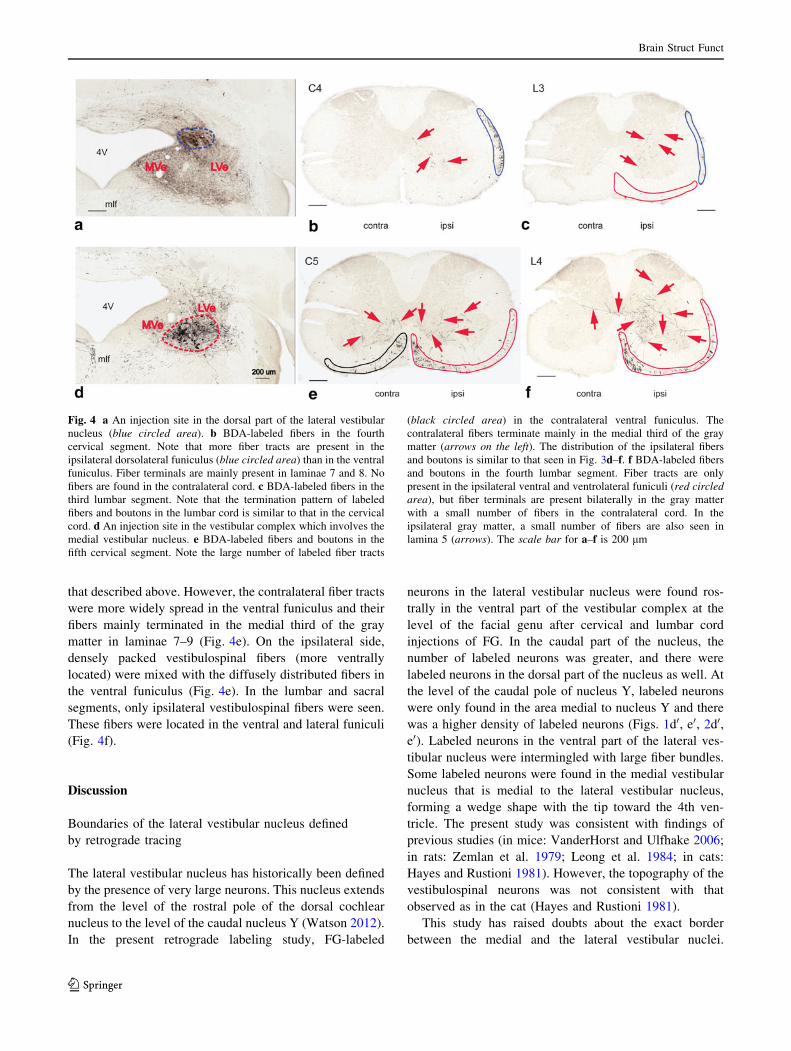
that described above. However, the contralateral fiber tracts
were more widely spread in the ventral funiculus and their
fibers mainly terminated in the medial third of the gray
matter in laminae 7–9 (Fig. 4e). On the ipsilateral side,
densely packed vestibulospinal fibers (more ventrally
located) were mixed with the diffusely distributed fibers in
the ventral funiculus (Fig. 4e). In the lumbar and sacral
segments, only ipsilateral vestibulospinal fibers were seen.
These fibers were located in the ventral and lateral funiculi
(Fig. 4f).
Discussion
Boundaries of the lateral vestibular nucleus defined
by retrograde tracing
The lateral vestibular nucleus has historically been defined
by the presence of very large neurons. This nucleus extends
from the level of the rostral pole of the dorsal cochlear
nucleus to the level of the caudal nucleus Y (Watson 2012).
In the present retrograde labeling study, FG-labeled
neurons in the lateral vestibular nucleus were found ros-
trally in the ventral part of the vestibular complex at the
level of the facial genu after cervical and lumbar cord
injections of FG. In the caudal part of the nucleus, the
number of labeled neurons was greater, and there were
labeled neurons in the dorsal part of the nucleus as well. At
the level of the caudal pole of nucleus Y, labeled neurons
were only found in the area medial to nucleus Y and there
was a higher density of labeled neurons (Figs. 1d0, e0, 2d0,e0). Labeled neurons in the ventral part of the lateral ves-
tibular nucleus were intermingled with large fiber bundles.
Some labeled neurons were found in the medial vestibular
nucleus that is medial to the lateral vestibular nucleus,
forming a wedge shape with the tip toward the 4th ven-
tricle. The present study was consistent with findings of
previous studies (in mice: VanderHorst and Ulfhake 2006;
in rats: Zemlan et al. 1979; Leong et al. 1984; in cats:
Hayes and Rustioni 1981). However, the topography of the
vestibulospinal neurons was not consistent with that
observed as in the cat (Hayes and Rustioni 1981).
This study has raised doubts about the exact border
between the medial and the lateral vestibular nuclei.
Fig. 4 a An injection site in the dorsal part of the lateral vestibular
nucleus (blue circled area). b BDA-labeled fibers in the fourth
cervical segment. Note that more fiber tracts are present in the
ipsilateral dorsolateral funiculus (blue circled area) than in the ventral
funiculus. Fiber terminals are mainly present in laminae 7 and 8. No
fibers are found in the contralateral cord. c BDA-labeled fibers in the
third lumbar segment. Note that the termination pattern of labeled
fibers and boutons in the lumbar cord is similar to that in the cervical
cord. d An injection site in the vestibular complex which involves the
medial vestibular nucleus. e BDA-labeled fibers and boutons in the
fifth cervical segment. Note the large number of labeled fiber tracts
(black circled area) in the contralateral ventral funiculus. The
contralateral fibers terminate mainly in the medial third of the gray
matter (arrows on the left). The distribution of the ipsilateral fibers
and boutons is similar to that seen in Fig. 3d–f. f BDA-labeled fibers
and boutons in the fourth lumbar segment. Fiber tracts are only
present in the ipsilateral ventral and ventrolateral funiculi (red circledarea), but fiber terminals are present bilaterally in the gray matter
with a small number of fibers in the contralateral cord. In the
ipsilateral gray matter, a small number of fibers are also seen in
lamina 5 (arrows). The scale bar for a–f is 200 lm
Brain Struct Funct
123

Paxinos and Watson (1982) in their rat brain atlas defined a
larger lateral vestibular nucleus at the expense of the
medial vestibular nucleus. In the 2008 edition of the mouse
brain atlas (Franklin and Paxinos 2008), as well as in the
more recent edition of the mouse brain atlas (Paxinos and
Franklin 2013), the medial vestibular nucleus was made
larger at the expense of the lateral vestibular nucleus.
Because immunohistochemical stains (Watson and Paxinos
2010) and in situ data (i.e., Allen Brain Atlas,
http://mouse.brain-map.org) do not reveal a distinct
boundary between the two nuclei, it is hard to know where
the true borders might lie. Some have argued that the medial
vestibular nucleus does not project below the thoracic cord
(Wilson et al. 1967). By that criterion, all that is currently
delineated in the atlas of the medial vestibular nucleus
should be transferred to the lateral vestibular nucleus (col-
ored dashed lines in Fig. 1) since these neurons project to
the lumbar spinal cord. Certainly the magnocellular part of
the medial vestibular nucleus has larger neurons than the
parvicellular part of the medial vestibular nucleus, but the
neurons of the magnocellular part of the medial vestibular
nucleus are not as large as those in the adjacent area which
the atlases label the lateral vestibular nucleus.
As noted above, the currently available data from
immunohistochemistry, in situ hybridization, and Nissl
staining do not adequately define the precise boundary
between the lateral vestibular nucleus and the medial
vestibular nucleus. We believe that the present tracing
study might be used to define a new boundary between
these nuclei, and the validity of this boundary could be
tested in future electrophysiological studies (Fig. 5).
It has been reported that some vestibulospinal neurons in
the lateral vestibular nucleus are glutamic acid decarbox-
ylase (GAD) positive (Valla et al. 2003). Whereas in
another study, 97 % of the vestibulospinal fibers inner-
vating the lumbar cord have been found to be vesicular
glutamate transporter 2 (vGluT2) positive (Du Beau et al.
2012), indicating that these vestibulospinal fibers are
excitatory. In the mouse, neurotransmitters used by the
vestibulospinal neurons have not been investigated.
Fig. 5 A summary diagram showing the origin, course and termina-
tion of the descending fibers from the lateral vestibular nucleus to the
spinal cord. Vestibulospinal fibers originate in the lateral vestibular
nucleus and descend in the hindbrain either within the ipsilateral
medial longitudinal fasciculus or lateral to it. These descending fibers
travel in the ipsilateral ventral and lateral funiculi in the white matter
of the spinal cord and terminate in laminae 6–10, with predominance
in laminae 8 of the ipsilateral side. A small number of fibers were
occasionally found in ipsilateral lamina 5. Contralateral collaterals
were present in the lumbosacral cord. It was uncertain whether
collaterals were also present in the cervical and thoracic segments.
Therefore, only collaterals in the lumbar cord were represented in this
diagram
b
Brain Struct Funct
123

Fiber termination of the lateral vestibular nucleus
in the spinal cord
We believe that the present study is the first to describe
the fiber termination of the lateral vestibular nucleus in the
spinal cord of the mouse. The fibers projecting from the
lateral vestibular nucleus travelled in the ipsilateral ventral
and lateral funiculi and terminated in the ventromedial gray
matter of the ipsilateral spinal cord. In some lumbar and
sacral cord sections, collaterals of vestibulospinal fibers
were found in the contralateral cord. In the cervical and
upper thoracic cord, some fiber tracts and terminals were
also found in the contralateral cord. We think it is likely that
these fibers have arisen from the medial vestibular nucleus.
In the rat, it has been reported that vestibulospinal tracts
travel in the ipsilateral ventral funiculus, and to a lesser
extent in the ipsilateral lateral funiculus. Their fibers
mainly terminate in the ventral horn, especially in laminae
7 and 8, and to a lesser extent in the dorsal horn. Fur-
thermore, a considerable number of collaterals are also
present in the cervical and lumbosacral segments (Bacskai
et al. 2002; Matesz et al. 2002). Our results are consistent
with the findings in the rat.
In the cat (Rose et al. 1999), fibers from the lateral
vestibular nucleus have been traced to the upper cervical
cord where they are bilaterally distributed in the ventro-
medial funiculus, but only ipsilaterally in the ventrolateral
and lateral funiculi. This finding is similar to ours in
relation to the distribution of vestibulospinal fibers in the
cervical cord. However, these authors did not investigate
the fiber termination in lower spinal segments. Another
study in the cat (Kuze et al. 1999) revealed the lateral
vestibular nucleus fiber distribution in the entire spinal cord
by injecting PHA-L into the lateral vestibular nucleus.
Vestibulospinal fibers were seen to travel in the ventral
funiculus bilaterally in the cervical cord, but only in the
ipsilateral ventral funiculus in lower segments. This result
is consistent with our findings.
In the squirrel monkey (Boyle 2000), vestibulospinal
fibers from the lateral vestibular nucleus were seen to reach
the lumbar cord with few collaterals in the cervical cord.
Boyle concluded that these lumbar projecting fibers may
serve as a ‘private pathway’ connecting the lateral vestib-
ular nucleus neurons and the lumbar cord.
In the present study, some fibers were also found in the
contralateral lumbar and sacral cord, but no contralateral
tract was observed. These contralateral fibers must there-
fore originate from the ipsilateral vestibulospinal tract.
This phenomenon was also observed in an electrophysio-
logical study, which showed that the lumbar cord was
innervated by the lateral vestibulospinal tract only, and that
there are bifurcating fibers innervating the contralateral
spinal cord (Kasumacic et al. 2010).
In this study, a few fibers were found in the dorsolateral
funiculus after injecting BDA to the dorsal part of the
lateral vestibular nucleus. These fibers are apparently dif-
ferent from those in the ventral funiculus. Based on their
location in the vestibular complex and their discrete char-
acter, we propose that they might arise from the superior
vestibular nucleus.
Functional significance of spinal projections
from the lateral vestibular nucleus
The present study has clearly shown that fibers from the
lateral vestibular nucleus innervate not only the axial motor
neurons, but also the motor neurons in the lateral motor
neuron group and interneurons in laminae 7 and 8. This is
consistent with previous findings that the lateral vestibular
nucleus is involved in both posture and balance control. The
contralateral extensor motor neurons and bilateral flexor
motor neurons might be activated and inhibited, respec-
tively, through commissural interneurons as found in other
mammals (Wilson and Yoshida 1969; Pompeiano 1972).
It has been shown that somatosensory and vestibular
inputs converge on interneurons in the spinal cord (Brink
et al. 1980). These interneurons might mediate various
responses to external stimuli through their connections
with other neurons in the spinal cord. For example, inter-
neurons in laminae 7 and 8 of the spinal cord may partic-
ipate in locomotion through connections with the central
pattern generator (Kjaerulff et al. 1994; Bracci et al. 1996;
Tresch and Kiehn 1999; Cina and Hochman 2000; Dai
et al. 2005). It is highly possible that vestibulospinal fiber
terminals on the interneurons in laminae 7 and 8 are indi-
rectly engaged in locomotion.
Conclusion
The present study revealed in the spinal cord the distribu-
tion pattern of fibers from the lateral vestibular nucleus and
proposed the boundary between the lateral and the medial
vestibular nuclei based on retrograde tracing results. This
can provide an anatomical foundation for further vestibular
research on the mouse.
References
Abercrombie M (1946) Estimation of nuclear population from
microtome sections. Anat Rec 94:239–247
Abzug C, Maeda M, Peterson BW, Wilson VJ (1974) Cervical
branching of lumbar vestibulospinal axons. J Physiol 243:
499–522
Auclair F, Belanger MC, Marchand R (1993) Ontogenetic study of
early brain stem projections to the spinal cord in the rat. Brain
Res Bull 30:281–289
Brain Struct Funct
123

Bacskai T, Szekely G, Matesz C (2002) Ascending and descending
projections of the lateral vestibular nucleus in the rat. Acta Biol
Hung 53:7–21
Boyle R (2000) Morphology of lumbar-projecting lateral vestibulo-
spinal neurons in the brainstem and cervical spinal cord in the
squirrel monkey. Arch Ital Biol 138:107–122
Bracci E, Ballerini L, Nistri A (1996) Localization of rhythmogenic
networks responsible for spontaneous bursts induced by strych-
nine and bicuculline in the rat isolated spinal cord. J Neurosci
16:7063–7076
Brink EE, Hirai N, Wilson VJ (1980) Influence of neck afferents on
vestibulospinal neurons. Exp Brain Res 38:285–292
Cina C, Hochman S (2000) Diffuse distribution of sulforhodamine
labeled neurons during serotonin-evoked locomotion in the
neonatal rat thoracolumbar spinal cord. J Comp Neurol 423:590–
602
Dai X, Noga BR, Douglas JR, Jordan LM (2005) Localization of
spinal neurons activated during locomotion using the c-fos
immunohistochemical method. J Neurophysiol 93:3442–3452
de Boer-van Huizen RT, ten Donkelaar HJ (1999) Early development
of descending supraspinal pathways: a tracing study in fixed and
isolated rat embryos. Anat Embryol 199:539–547
Du Beau A, Shakya Shrestha S, Bannatyne BA, Jalicy SM, Linnen S,
Maxwell DJ (2012) Neurotransmitter phenotypes of descending
systems in the rat lumbar spinal cord. Neuroscience 227:67–79
Franklin KBJ, Paxinos G (2008) The mouse brain in stereotaxic
coordinates, 3rd edn. Elsevier Academic Press, San Diego
Grillner S, Hongo T, Lund S (1970) The vestibulospinal tract. Effects
on alpha-motoneurones in the lumbosacral spinal cord in the cat.
Exp Brain Res 10:94–120
Hayes NL, Rustioni A (1981) Descending projections from brainstem
and sensorimotor cortex to spinal enlargements in the cat. Single
and double retrograde tracer studies. Exp Brain Res 41:89–107
Huisman AM, Ververs B, Cavada C, Kuypers HG (1984) Collater-
alization of brainstem pathways in the spinal ventral horn in rat
as demonstrated with the retrograde fluorescent double-labeling
technique. Brain Res 300:362–367
Isu N, Uchino Y, Nakashima H, Satoh S, Ichikawa T, Watanabe S
(1988) Axonal trajectories of posterior canal-activated secondary
vestibular neurons and their coactivation of extraocular and neck
flexor motoneurons in the cat. Exp Brain Res 70:181–191
Kasumacic N, Glover JC, Perreault MC (2010) Segmental patterns of
vestibular-mediated synaptic inputs to axial and limb motoneu-
rons in the neonatal mouse assessed by optical recording.
J Physiol 588:4905–4925
Kjaerulff O, Barajon I, Kiehn O (1994) Sulphorhodamine-labeled
cells in the neonatal rat spinal cord following chemically induced
locomotor activity in vitro. J Physiol 478:265–273
Kudo N, Furukawa F, Okado N (1993) Development of descending
fibers to the rat embryonic spinalcord. Neurosci Res 16:131–141
Kuze B, Matsuyama K, Matsui T, Miyata H, Mori S (1999) Segment-
specific branching patterns of single vestibulospinal tract axons
arising from the lateral vestibular nucleus in the cat: a PHA-L
tracing study. J Comp Neurol. 414:80–96
Leong SK, Shieh JY, Wong WC (1984) Localizing spinal-cord—
projecting neurons in adult albino rats. J Comp Neurol 228:1–17
Liang H, Paxinos G, Watson C (2011) Projections from the brain to
the spinal cord in the mouse. Brain Struct Funct 215:159–186
Masson RL, Sparkes ML, Ritz LA (1991) Descending projections to
the rat sacrocaudal spinal cord. J Comp Neurol 302:120–130
Matesz C, Bacskai T, Nagy E, Halasi G, Kulik A (2002) Efferent
connections of the vestibular nuclei in the rat: a neuromorpho-
logical study using PHA-L. Brain Res Bull 57:313–315
Paxinos G, Franklin KBJ (2013) The Mouse brain in stereotaxic
coordinates, 4th edn. Elsevier Academic Press, San Diego
Paxinos G, Watson C (1982) The Rat brain in stereotaxic coordinates.
Academic Press, Sydney
Pompeiano O (1972) Vestibulospinal relations: vestibular influences
on gamma motoneurons and primary afferents. Prog Brain Res
37:263–296
Rice CD, Weber SA, Waggoner AL, Jessell ME, Yates BJ (2010)
Mapping of neural pathways that influence diaphragm activity
and project to the lumbar spinal cord in cats. Exp Brain Res
203:205–211
Rose PK, Wainwright K, Neuber-Hess M (1992) Connections from
the lateral vestibular nucleus to the upper cervical spinal cord of
the cat: a study with the anterograde tracer PHA-L. J Comp
Neurol 321:312–324
Rose PK, Ely S, Norkum V, Neuber-Hess M (1999) Projections from
the lateral vestibular nucleus to the upper cervical spinal cord of
the cat: a correlative light and electron microscopic study of
axon terminals stained with PHA-L. J Comp Neurol 410:571–
585
Shen P, Arnold AP, Micevych PE (1990) Supraspinal projections to
the ventromedial lumbar spinal cord in adult male rats. J Comp
Neurol 300:263–272
Shinoda Y, Ohgaki T, Sugiuchi Y, Futami T (1989) Comparison of
the branching patterns of lateral and medial vestibulospinal tract
axons in the cervical spinal cord. Prog Brain Res 80:137–147
Tresch MC, Kiehn O (1999) Coding of locomotor phase in
populations of neurons in rostral and caudal segments of the
neonatal rat lumbar spinal cord. J Neurophysiol 82:3563–3574
Valla J, Delfini C, Diagne M, Pinganaud G, Buisseret P, Buisseret-
Delmas C (2003) Vestibulotrigeminal and vestibulospinal pro-
jections in rats: retrograde tracing coupled to glutamic acid
decarboxylase immunoreactivity. Neurosci Lett 340:225–228
VanderHorst VG, Ulfhake B (2006) The organization of the
brainstem and spinal cord of the mouse: relationships between
monoaminergic, cholinergic, and spinal projection systems.
J Chem Neuroanat 31:2–36
Wada N, Sugita S, Jouzaki A, Tokuriki M (1993) Descending
projections to coccygeal spinal segments in the cat. J Anat
182:259–265
Watson C (2012) Hindbrain. In: Watson C, Paxinos G, Puelles L (eds)
The Mouse Nervous System, chap. 12. Elsevier Academic Press,
San Diego, pp 408–409
Watson C, Paxinos G (2010) Chemoarchitectonic Atlas of the Mouse
Brain. Elsevier Academic Press, San Diego
Watson C, Paxinos G, Kayalioglu G, Heise C (2009) Atlas of the
mouse spinal cord. In: Watson C, Paxinos G, Kayalioglu G (eds)
The Spinal cord. Elsevier Academic Press, San Diego,
pp 308–379
Wilson VJ, Yoshida M (1969) Comparison of effects of stimulation
of Deiters’ nucleus and medial longitudinal fasciculus on
neck, forelimb, and hindlimb motoneurons. J Neurophysiol 32:
743–758
Wilson VJ, Wylie RM, Marco LA (1967) Projection to the spinal cord
from the medial and descending vestibular nuclei of the cat.
Nature 215:429–430
Wilson VJ, Yoshida M, Schor RH (1970) Supraspinal monosynaptic
excitation and inhibition of thoracic back montoneurons. Exp
Brain Res 11:282–295
Zemlan FP, Kow LM, Morrell JI, Pfaff DW (1979) Descending tracts
of the lateral columns of the rat spinal cord: a study using the
horseradish peroxidase and silver impregnation techniques.
J Anat 128:489–512
Brain Struct Funct
123









![Electrical Vestibular Stimulation after Vestibular ......electrical stimulation of the vestibular system to one ear [4,5,9]. However studies have also reported vestibular responses](https://static.fdocuments.in/doc/165x107/60f6b0762ca1b41e91018b73/electrical-vestibular-stimulation-after-vestibular-electrical-stimulation.jpg)








