
Prevalence of Localized Rearrangements Vs. Transpositions among ...
18
Copyright 0 1995 by the Genetics Society of America Prevalence of Localized Rearrangements us. Transpositions Among Events Induced by Drosophila P Element Transposase on a P Transgene Marion Delattre, Dominique Anxolabeh6re and Dario Coen Dipartement Dynamique du Ge’nome et Evolution, Institut Jacques Monod, F-75251 Paris Cedex 05, France and Universiti Pierre et Marie Curie (Paris 6), F-75252 Paris Cedex 05, France Manuscript received December 19, 1994 Accepted for publication August 18, 1995 ABSTRACT We have studied P transposase-induced events on a P[ w] transgene, P[ wd’], harboring the whole white gene with a 3.44kb direct duplication ofits 5’ regulatory sequences (containing the ZESTE-binding region, ZBR). We have recovered mutations leading to an increase or a decrease of zeste’ repression, generally as the consequence of modifications of the number of ZBR in close physical proximity and/ or jumps to other sites. We describe mutants displaying deletions of the original duplicated sequence or increases in the number of repeats from two to three or four. Internal deletions are more frequent than amplifications. Both require the integrity of Pelement ends. We havealso observed a high frequency of double Pelements localized at the original P[ 4’1 insertion site. These double Pelements are arranged in nonrandom configurations. We discuss the frequencies and the possible mechanisms leading to the various types of derivatives,in light of the current models for P excision and transposition. We propose that the P transposase induces mainly localized events. Some of these could result from frequent changes of template during gap-repair DNA synthesis, and/or from abortive transposition. P elements are a familyof transposable elements responsible for the phenomenon of P-M hybrid dysgenesis in Drosophila mlanogaster (reviewed by ENG ELS 1989). Complete P elements are 2907 bp in length and have 31-bp terminal inverted repeats (TIRs) (0’- and RUBIN 1983). They are autonomously functional for transposition (SPRADLING and RUBIN 1982). The complete P element contains four exons and encodes at least two overlapping genes, one for a 87-kD transposase protein and the other for a 66kD protein (Rro et al. 1986). The latter is a repressor of P transposition (ROBERTSON and ENGELS 1989; COEN 1990; MISRA and Rro 1990; LEMAITRE and COEN 1991). Shorter P elements usually derive from the 2.9-kb ele- ment by internal deletions (0’- and RUBIN 1983). Defective P elements are unable to transpose autono- mously but can be mobilized in trans by active elements (SPRADLING and RUBIN 1982; ENGELS 1984). Pelements generate an 8-bp duplication of genomic DNA at the site of insertion. P elements are thought to transpose by a nonreplica- tive “cut-and-paste” mechanism (ENGELS et al. 1990; KAUFMAN and Rro 1992) starting by an excision of the element that results in a double-strand gap at the donor site. This gap is subsequently repaired by a process simi- lar to gene conversion, using either the sister chroma- tid, the homologue or any homologous sequences in- serted in the genome, as well as oligonucleotides, as a Corresponding author: Dario Coen, Dtpartement Dynamique du GB nome et Evolution, Institut Jacques Monod, Tour 43, 2 place Jussieu, 75251 Paris Cedex 05, France. E-mail: [email protected] Genetics 141: 1407-1424 (December, 1995) template for repair (ENGELS et al. 1990; BANGA and BOYD 1992; JOHNSON-SCHLITZ and ENGELS 1993; NASSIF et al. 1994). The repair process has been used for a gene replacement procedure, using the white gene of D. melanogaster and involving templates with single-base substitutions, deletions and insertions (GLOOR et al. 1991; JOHNSON-SCHLITZ and ENGELS 1993; NASSIF et al. 1994). Complex conversion events have led NASSIF et al. (1994) to postulate that the two broken ends con- duct an independent genome search for a template and may use two different templates. They have proposed a synthesis-dependent strand annealing (SDSA) model for the repair of the break. According to their model, each terminus of the break invades a template indepen- dently and serves as a primer for DNA synthesis, with the newly synthesized DNA being immediately dis- placed from the template. These single strands then pair in a region of overlap, and synthesis is completed by each using the other as a template. We have studied P transposase-induced events on a P transgene, P[ wd’9.3](19DE).This transgene harbors a complete white’ gene with 3.44 kb of its 5’ regulatory sequences duplicated directly in tandem (C. ZUCKER, personal communication; COEN 1990).The white+ gene expression is repressed by the zeste’ mutation. This ef- fect requires the presence of two or more copies of white+ regulatory sequences brought into close proxim- ity by homologous chromosome pairing or by tandem duplication (transvection phenomenon, reviewed by PIRROTTA 1991). The regulatory sequences that lie 1185-1454 bp upstream of the transcription start site of the white gene contain the eye enhancer element
Transcript of Prevalence of Localized Rearrangements Vs. Transpositions among ...

Copyright 0 1995 by the Genetics Society of America
Prevalence of Localized Rearrangements us. Transpositions Among Events Induced by Drosophila P Element Transposase on a P Transgene
Marion Delattre, Dominique Anxolabeh6re and Dario Coen
Dipartement Dynamique du Ge’nome et Evolution, Institut Jacques Monod, F-75251 Paris Cedex 05, France and Universiti Pierre et Marie Curie (Paris 6), F-75252 Paris Cedex 05, France
Manuscript received December 19, 1994 Accepted for publication August 18, 1995
ABSTRACT We have studied P transposase-induced events on a P[ w] transgene, P[ wd’], harboring the whole white
gene with a 3.44kb direct duplication of its 5’ regulatory sequences (containing the ZESTE-binding region, ZBR). We have recovered mutations leading to an increase or a decrease of zeste’ repression, generally as the consequence of modifications of the number of ZBR in close physical proximity and/ or jumps to other sites. We describe mutants displaying deletions of the original duplicated sequence or increases in the number of repeats from two to three or four. Internal deletions are more frequent than amplifications. Both require the integrity of Pelement ends. We have also observed a high frequency of double Pelements localized at the original P[ 4’1 insertion site. These double Pelements are arranged in nonrandom configurations. We discuss the frequencies and the possible mechanisms leading to the various types of derivatives, in light of the current models for P excision and transposition. We propose that the P transposase induces mainly localized events. Some of these could result from frequent changes of template during gap-repair DNA synthesis, and/or from abortive transposition.
P elements are a family of transposable elements responsible for the phenomenon of P-M hybrid
dysgenesis in Drosophila mlanogaster (reviewed by ENG ELS 1989). Complete P elements are 2907 bp in length and have 31-bp terminal inverted repeats (TIRs) (0’- and RUBIN 1983). They are autonomously functional for transposition (SPRADLING and RUBIN 1982). The complete P element contains four exons and encodes at least two overlapping genes, one for a 87-kD transposase protein and the other for a 66kD protein (Rro et al. 1986). The latter is a repressor of P transposition (ROBERTSON and ENGELS 1989; COEN 1990; MISRA and Rro 1990; LEMAITRE and COEN 1991). Shorter P elements usually derive from the 2.9-kb ele- ment by internal deletions (0’- and RUBIN 1983). Defective P elements are unable to transpose autono- mously but can be mobilized in trans by active elements (SPRADLING and RUBIN 1982; ENGELS 1984). Pelements generate an 8-bp duplication of genomic DNA at the site of insertion.
P elements are thought to transpose by a nonreplica- tive “cut-and-paste” mechanism (ENGELS et al. 1990; KAUFMAN and Rro 1992) starting by an excision of the element that results in a double-strand gap at the donor site. This gap is subsequently repaired by a process simi- lar to gene conversion, using either the sister chroma- tid, the homologue or any homologous sequences in- serted in the genome, as well as oligonucleotides, as a
Corresponding author: Dario Coen, Dtpartement Dynamique du GB nome et Evolution, Institut Jacques Monod, Tour 43, 2 place Jussieu, 75251 Paris Cedex 05, France. E-mail: [email protected]
Genetics 141: 1407-1424 (December, 1995)
template for repair (ENGELS et al. 1990; BANGA and BOYD 1992; JOHNSON-SCHLITZ and ENGELS 1993; NASSIF et al. 1994). The repair process has been used for a gene replacement procedure, using the white gene of D. melanogaster and involving templates with single-base substitutions, deletions and insertions (GLOOR et al. 1991; JOHNSON-SCHLITZ and ENGELS 1993; NASSIF et al. 1994). Complex conversion events have led NASSIF et al. (1994) to postulate that the two broken ends con- duct an independent genome search for a template and may use two different templates. They have proposed a synthesis-dependent strand annealing (SDSA) model for the repair of the break. According to their model, each terminus of the break invades a template indepen- dently and serves as a primer for DNA synthesis, with the newly synthesized DNA being immediately dis- placed from the template. These single strands then pair in a region of overlap, and synthesis is completed by each using the other as a template.
We have studied P transposase-induced events on a P transgene, P [ wd’9.3](19DE). This transgene harbors a complete white’ gene with 3.44 kb of its 5’ regulatory sequences duplicated directly in tandem (C. ZUCKER, personal communication; COEN 1990). The white+ gene expression is repressed by the zeste’ mutation. This ef- fect requires the presence of two or more copies of white+ regulatory sequences brought into close proxim- ity by homologous chromosome pairing or by tandem duplication (transvection phenomenon, reviewed by PIRROTTA 1991). The regulatory sequences that lie 1185-1454 bp upstream of the transcription start site of the white gene contain the eye enhancer element

1408 M. Delattre, D. Anxolab&hSre and D. Cohen
and five binding sites for the ZESTE protein (Qw et al. 1992). This ZESTE binding region (ZBR) is neces- sary for the transvection phenomenon to occur. It is included in the 5' white duplicated sequence in the P[wd'9.3] transgene (Figure 1 ) . In a previous report, we have shown that the expression of the white gene carried by this transgene is repressed by the P-encoded repressor(s) (COEN 1990). This P repression occurs only in the presence of the zeste' allele and is strongly dependent on the insertion site of this transgene: this effect is very rarely recovered at other sites in the ge- nome (COEN 1990).
In the present work, we describe the recovery and analysis of mutations induced on this transgene by the P transposase. We have used the level of expression of the white gene, influenced by P and zeste repression, to monitor the modifications induced by the P transposase on the P[wd'9.3] transgene. Our prediction is that a darker phenotype ( i . e . , less repressed by zeste') would result in most cases, either from the loss of ZBR at the original site or from a jump to another site, where it is insensitive to P repression, and that a lighter phenotype ( i e . , more repressed by zestd) would result, in most cases, from an increase of the number of ZBR in close proximity. These predictions were mainly confirmed. Our results show that the P transposase induces dele- tions and amplifications of the original duplicated se- quence and that these two types of events occur at very different rates. We also describe many cases of double P elements at the original site. We observe that these double P elements are arranged in nonrandom config- urations. The relative frequencies of all types of events observed, including transposition to other chromo- somes, are discussed. Our results show that the P trans- posase induces preponderantly localized events on the P[wd'9.3] insertion. The implications of these results for the understanding of P element transposition are examined.
MATERIALS AND METHODS
The P[wd'] transgene: This transgene (C. ZUCKER and D. THIERRY-MIEG, personal communication) carries an EcoRI- KpnI fragment containing the whole white+ gene. This frag- ment includes 5' and 3' nontranscribed sequences sufficient for correct temporal and spatial expression of white (HA- ZELRIGG et al. 1984; LEVIS et UL. 1985b; PIRROTTA et U L . 1985). In addition, P[d'] (Figure 1) harbors a direct tandem dupli- cation of the 3.44kb 5' EcoRI-Hind111 fragment. This fragment contains the 5' regulatory sequences and the first exon (PIR- R O ~ A and BROCKL 1984), including the sequences responsi- ble for the interaction with the ZESTE protein (PIRROTTA 1991) (Figure 1).
Drosophila stocks: y z1 w1118: M stock. w11f8 is a viable partial deletion of the white locus (HAZELRIGG et al. 1984).
tion of the p[dl] transposon (Figure 1) at the cytological position 19DE (C. ZUCKER and D. THIERRY-MIEG, personal communication; RUBIN et al. 1985; COEN 1990). The inserted transgene and the strain harboring it will be abbreviated 9.3 and z w 9.3, respectively.
zl w1118 cuP[d19.3] (19DE): This M strain contains an inser-
Muller-5 designated M5: an M line with the Muller-5 balancer X chromosome (Basc chromosome marked with Bar w") (LINDSLEY and ZIMM 1992).
HS2-20 and 40102: These two lines were derived from the Gruta M stock by germline transformation with P element DNA (ANXOLABEHERE et al. 1987).
Harwich: A standard P reference stock (KIDWELL et al. 1977).
C(I)DX, y w f (M)/w u 1(1)44/Y: An M stock used to collect virgin females. 1(1)44 is a thermosensitive X-linked lethal mu- tation (BUSSON et al. 1983). This stock will be abbreviated C(I)DX (M) .
C(l)DX, y w f ( P ) / w u 1(1)44(P)/Y;Harwich: The Xchromo- somes of this stock were contaminated with P elements as described by ENGELS (1985). All the autosomes derive from Harwich. This stock will be designated as C(1)DX (P).
rJ"4. Sb P[ly+ A2-3](99B)/TM6: This strain contains an essentially immobile P element that produces a high rate of transposase somatically and germinally (ROBERTSON et al. 1988). It is designated hereafter as A2-3(99B).
All genetic symbols not described here are in LINDSLEY and ZIMM (1992).
Mobilization of P[wd'9.3](19DE) and P[d6](19DE), an X- l i e d P-red derivative of 9.3: All crosses were performed as described in Figures 2 and 3, respectively, at 25" unless other- wise mentioned.
DNA extraction and Southern analysis: DNA extraction from adult flies was performed as described by JUNAKOVIC et al. (1984). All standard molecular techniques, such as restriction digestion, agarose gel electrophoresis, Southern blotting and hybridization, were performed as described in MANIATIS et al. (1982).
PCR procedure and sequencing: Genomic DNA was treated with RNase A, proteinase K, phenol-chloroform, pre- cipitated with ethanol and resuspended in H20. All PCR am- plifications were performed in a 50-pl mixture covered with mineral oil, using the following conditions: 50 mM KC1, 10 mM Tris-HC1 pH 8.4, 2.0 mM MgC12, 0.2 mM of each dNTP, 100 ng of DNA, 2.5 U of Taq DNA polymerase (Perkin Elmer Cetus) and 50 pmol of each primer. Amplification was per- formed for 30 cycles on a Biometra Trio-Thermoblock as fol- lows: 1 min, 94"; 1 min, 64"; 1 min, 72".
The P element-specific primers used in PCR experiments were as follows (coordinates as in the P element sequence) (O'HARE and RUBIN 1983):
primer P1: P108-P89 5'CGTCCGCACACAACCTTTCC3' primer P2: P414P433 5'GGCTATACCAGTGGGAGTAC3' primer P3: P2767-P2748 5'CCTTAGCATGTCCGTGGGGT3' primer P 4 P2785-P2804 5'TCGCTGTCTCACTCAGACTC3'
A white specific primer, localized at position 13642-13661 (in white coordinates, Genbank accession number X02974) (O'HARE et al. 1984), was also used:
primer W 5'TCTCCCACCCGCTCTCCGCA3'
Direct sequencing of the PCR products was performed ac- cording to the procedure described by SACRAMENTO et al.
Inverse PCR procedure: This procedure was used to deter- mine the sequence of DNA flanking the 9.3 insert and its derivatives. Genomic DNA was digested with the appropriate restriction enzyme for 6 hr at 37" then treated with RNAse A for 15 min. After ethanol precipitation, 2 pg of digested DNA was incubated for 16 hr at 15" in ligation buffer (Tris-HC1 66 mM pH 7.5, MgC12 5 mM, DTT 1 mM, ATP 1 mM) with 5 U of T4 DNA ligase in a final volume of 50 pl. The ligated sample was then treated as described in the PCR procedure.
(1991).

P-Induced Localized Rearrangements 1409
A white +
wild type RNA
I~ Bg on B g B h X X b II I I I s s I s K
B PS1 Ps3
1 kb
P l g W P3 P4 4 - %
FIGURE 1,"Structure of the white gene and of the p[wd'] transgene. (A) Genomic and transcription map of the white+ locus (adapted from HAZELRICC 1987). Solid boxes, white exons; white ovals, ZBR. The sequences that have been duplicated in R w d l ] transgene are indicated under the white+ map (diagonally hatched bar). Spotted bars (A-D), white sequence probes used in Southern blot analysis. The A probe is identical to the sequence duplicated in P[ wd']. (B) The p[d'] transgene (C. ZUCKER, personal communication). The two Pelement termini are indicated by large black arrowheads at the extremities of the construct. Diagonally striped and open boxes represent, respectively, the duplicated and unique white sequences. Spotted bars represent P element specific probes, PS1 and PS3, used in Southern blot analysis (see MATERIALS AND METHODS). Small arrows above the construct (Pl, P2, P3, P4 and W) represent the position and direction of primers used for PCR (see MATERIALS AND METHODS). Restriction enzyme sites: B, BumHI; Bg, BgaI; E, EcoRI; H, HindIII; K, KpnI; S, Sua; X, XhoI.
Probes: Four plasmids containing white' sequences that to +878) and the Sua (+2411 to +2883) fragments of the cover almost the whole gene were used as probes (see Figure Pelement, were used as 5' and 3' specific Pelement probes. 1 for their localization on the white gene map). The pPSl The 2.2-kb flanking sequence of P[wd'9.3] (19DE) original and pPS3 plasmids, containing respectively the Hind111 (+40 insertion site was amplified by the inverse PCR procedure
L1 score for non-brown 6
or non-white $?
+ individually [Sbl F 1 d x C(1)DXy w f o 99
I [Sb+] Fz d
sensitivity to P cytotype?
+ establish line
FIGURE 2.-Genetic screen used to detect local modifications and transpositions of 9.3. (A) Use of Pstrains as sources of transposase. Dysgenic F, males harboring the z w 9.3 X chromosome were individually crossed to compound-X females of the P cytotype. F2 males with eye color that differs from the P- specific brown phenotype were retained for further analysis. (B) Use of A2-3(99B) as a source of transposase. Dysgenic F1 males were individually crossed to C(1)DX (M) virgin fe- males. Fz (Sb+) males with either normal red eyes or various nonred phenotypes were then crossed individually to C(1)DX (P) females to test the eye phenotype of their progeny in a P background. After 3 days, the same males were crossed to C(1)DX (M) females, to estab- lish an M stock. Males giving in the first cross (P) progeny not composed exclusively of brown males and white females were retained for further analysis. The same crossing scheme was used following injection of the P helper into z w 9.3 embryos, replacing F1 dysgenic males by Go males. Go females were crossed to M5 males, and their GI male prog- eny was treated as Go males.
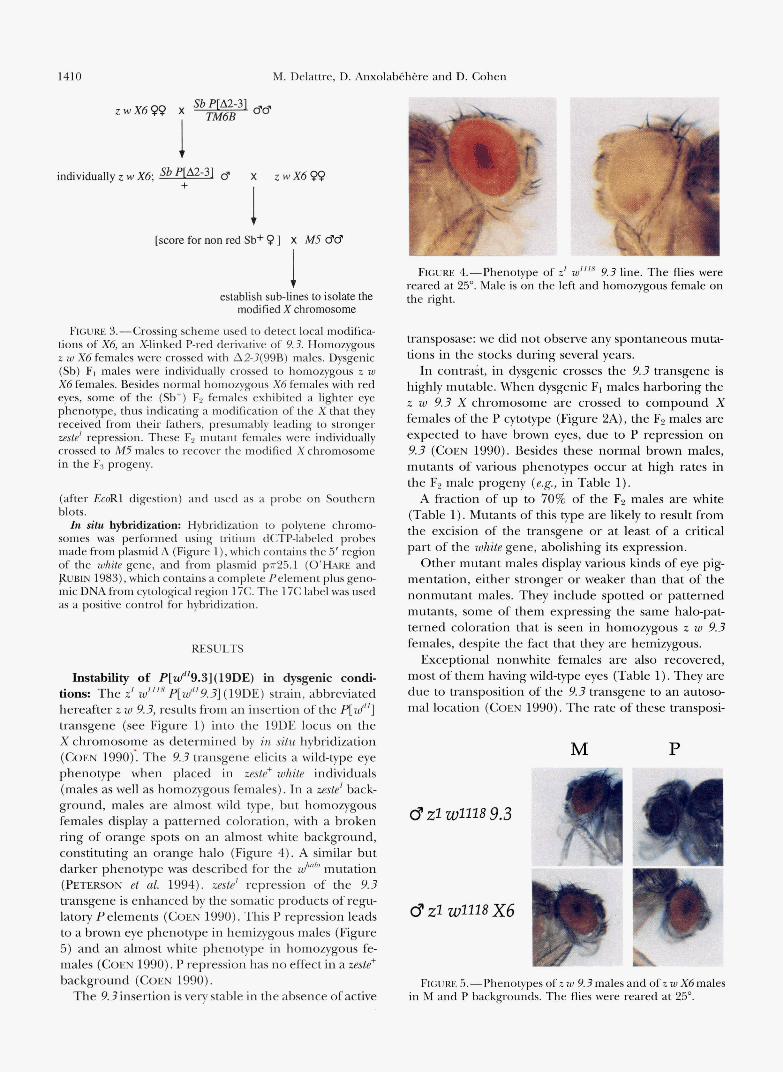
1410 M. Delattre, D. A n x c
zwX699 x 7-346~ Sb P[A2-31 @e
1 individuallyzwX6; - 6' X z w X 6 - +
1 [score for non red Sb+ 9 ] x M5 CPCF
I establish sub-lines to isolate the
modified X chromosome
FIGURE 3.-Crossing scheme used to detect local modifica- tions of X6, an X-linked P-red derivative of 9.3. Homozygous z w X6 females were crossed with A2-3(99B) males. Dysgenic (Sb) F, males were individually crossed to homozygous z w X6 females. Besides normal homozygous X6 females with red eyes, some of the (Sb+) FP females exhibited a lighter eye phenotype, thus indicating a modification of the X that they received from their fathers, presumably leading to stronger zestd repression. These F2 mutant females were individually crossed to M5 males to recover the modified X chromosome in the Fs progeny.
(after EcoRl digestion) and used as a probe on Southern blots. In situ hybridization: Hybridization to polytene chromo-
somes was performed using tritium dCTP-labeled probes made from plasmid A (Figure l), which contains the 5' region of the white gene, and from plasmid pn25.1 (0'- and RUBIN 1983), which contains a complete Pelement plus geno- mic DNA from cytological region 17C. The 17C label was used as a positive control for hybridization.
RESULTS
Instability of P[d19.3](19DE) in dysgenic condi- tions: The z' wZ1l8 p[wd19.3] (19DE) strain, abbreviated hereafter z w 9.3, results from an insertion of the P[ wd'] transgene (see Figure 1) into the 19DE locus on the X chromosome as determined by in situ hybridization (COEN 1990): The 9.3 transgene elicits a wild-type eye phenotype when placed in zeste+ white individuals (males as well as homozygous females). In a zeste' back- ground, males are almost wild type, but homozygous females display a patterned coloration, with a broken ring of orange spots on an almost white background, constituting an orange halo (Figure 4). A similar but darker phenotype was described for the Wh"b mutation (PETERSON et al. 1994). zeste' repression of the 9.3 transgene is enhanced by the somatic products of regu- latory P elements (COEN 1990). This P repression leads to a brown eye phenotype in hemizygous males (Figure 5 ) and an almost white phenotype in homozygous fe- males ( COEN 1990). P repression has no effect in a zeste' background (COEN 1990).
The 9.3 insertion is very stable in the absence of active
olab&h&e and D. &hen
FIGURE 4.-Phenotype of z1 w1'18 9.3 line. The flies were reared at 25". Male is on the left and homozygous female on the right.
transposase: we did not observe any spontaneous muta- tions in the stocks during several years.
In contrGt, in dysgenic crosses the 9.3 transgene is highly mutable. When dysgenic F, males harboring the z w 9.3 X chromosome are crossed to compound X females of the P cytotype (Figure 2A), the F2 males are expected to have brown eyes, due to P repression on 9.3 (COEN 1990). Besides these normal brown males, mutants of various phenotypes occur at high rates in the F2 male progeny (e.g., in Table 1).
A fraction of up to 70% of the F2 males are white (Table 1). Mutants of this type are likely to result from the excision of the transgene or at least of a critical part of the white gene, abolishing its expression.
Other mutant males display various kinds of eye pig- mentation, either stronger or weaker than that of the nonmutant males. They include spotted or patterned mutants, some of them expressing the same halo-pat- terned coloration that is seen in homozygous z w 9.3 females, despite the fact that they are hemizygous.
Exceptional nonwhite females are also recovered, most of them having wild-type eyes (Table 1). They are due to transposition of the 9.3 transgene to an autoso- mal location (COEN 1990). The rate of these transposi-
M P
0 21 w1118 9.3
0 ~ 1 ~ 1 1 1 8 X6
FIGURE 5.-Phenotypes of z w 9.3males and of z w X6males in M and P backgrounds. The flies were reared at 25".

P-Induced Localized Rearrangements
TABLE 1
Instability of P[wd‘9.3] in dysgenic conditions
141 1
F2 males F2 females
Transposase source“ Brown Red Lightb White Total White Wild type Other“ Totald
Harwich 64.0 20.4 2.9 12.7 2042 98.5 1.5 0.0 1035
40102 95.2 4.2 0.1 0.5 2882 99.8 0.2 0.0 2082 A2-?(99B) 6.7 15.0 7.7 70.6 963 94.7 5.3 0.0 607
HS2-20 60.5 14.3 5.8 19.4 6604 96.1 3.8 0.1 4820
a Crosses were performed according to the schemes depicted in Figure 2, using a P strain or A2-?(99B) as source of transposase.
*This class comprises individuals showing a lighter eye coloration as compared to the normal 9.3 phenotype. This includes
“This class comprises all individuals with a non-wild-type eye color.
The percentages of the different phenotypes are given as well as the total of F2 flies scored.
phenotypes ranging from orange to almost white and also spotted or patterned colorations.
Some of the F2 crosses were performed with C(1)DXyJP) females, thus preventing the detection of autosomal transposition among the female progeny. This explains the unequal sex ratio.
tion events is always lower than the rates of events o b served among F2 males.
P transposase induces frequent phenotypic variants displaying decreased zeste’ repression. These mutants display a red eye color despite being obtained in a P cytotype (Table 1). When Hanvich autosomes are used as the source of transposase, this class of mutants repre- sents as much as 20.4% of the total F2 male progeny (Table 1). This phenotype will be called “P-red”, in contrast to the normal “P-brown” phenotype of z w 9.3 (P) males.
P-red mutants may also be obtained in an M cytotype. This is achieved by using A2-3(99B) (ROBERTSON et al. 1988) as a source of transposase (Figure 2B) or by in-
jecting the helper P element p7r25 .7~~ (KARESS and RUBIN 1984) into z w 9.3 embryos. F2 males with red eyes are selected in the progeny of the dysgenic individuals obtained. They are then crossed individually to C(1)DX (P) females to test the eye phenotype of their progeny in a P background. After 3 days, the same males are crossed to C(I)DX (M) females to establish an M stock. Males giving progeny in the first cross (P) composed exclusively of brown males and white females are dis- carded as nonmutants. Males giving progeny containing at least a fraction of males with red eyes are retained (see below).
Genetic and molecular analysis of the P-red mu- tants Genetic analysis of 119 independent mutants of the P-red phenotype, induced by various sources of transposase (P strains, 42-3(99B) or injection of the helper) allowed grouping them into three classes: ( 1 ) 97 are X-linked; (2) eight are due to transposition to an autosomal location, the original 9.3 insert showing no phenotypically detectable modification; and (3) 14 result from transposition to an autosomal location ac- companied by a modification (e.g., excision) of the orig- inal insert. The first class (Xlinked mutants) is by far the most frequent with a relative frequency of 81.5%. The frequencies of the two other classes (6.7 and 11.7%) are not significantly different from each other.
The X6 derivative was obtained by injection of the helper Pelement into z w 9.3 embryos. X6males display the same eye coloration in an M or a P background (Figure 5). This showed that this mutant has indeed become insensitive to P repression. The homozygous X6 females display the same red eye color as hemizy- gous males (data not shown). Southern analysis of X6 genomic DNA revealed that X6 results from an excision of one of the two copies of the 3.44kb white5’ sequence duplication (Figure 6). This deletion appears to be precise at this level of analysis: the digestion pattern showed the conservation of all the sites tested except those included in the originally duplicated sequence. This Southern analysis showed no difference in the flanking sequences, indicating that the internal dele- tion event is not accompanied by transposition. PCR amplification and sequencing of the 5‘ flanking DNA confirm that X6 is at the same site as the original 9.3 insert (Figure 7A).
Southern analysis of genomic DNA was performed on 36 other X-linked P-red mutants. This analysis (results displayed in Figure 6) showed that 31 mutants among 36 are due to the same type of event as X6: an in situ excision of one of the two copies of the 3.44kb white 5‘ sequence duplication, appearing precise at the level of resolution of Southern analysis. Two derivatives are due to the same type of deletion accompanied by a modification of the flanking sequences (X45 and X42). The last three harbor larger internal deletions (X28, X37 and X43) (Figure 6).
All of the X-linked P-red derivatives analyzed are due to modifications involving at least the deletion of one copy of the original 3.44kb duplicated sequence. Therefore, P transposase appears to induce deletions of an original tandem repeat internal to P ends at a high rate. P transposase induces phenotypic variants displaying
increased zeste’ repression: Mutants with a lighter eye pigmentation than the original 9.3 phenotype repre- sent up to 7.7% of the total F2 progeny depending on

1412 M. Delattrr, D. Anxolah(.hi.re and D. Cohcn
P[wd'9.3] a bcdefgh i h' i' j lbmopqrst u v w x
a bcdefgh i h' khmopqrs t u v w x
x45 1 - " ~ : : " : : : 1 ~ ~ -
a gh i h' 0 U
X42 -m::Cl::IFT-- f gh i h' 0 U
S k b gh i h' - U
the transposase source (Table 1). In all cases, they occur less frequently than P-red mutants.
These mutants exhibit phenotypes ranging from brown to almost white ( ~ . g . , in Table 2). They include spotted or halo-patterned colorations. Twenty-nine in- dependent mutants of this class, displaying various phe- notypes, were genetically analyzed as described for the P-red mutants. Two classes were observed: 26 (out of 29) are X-linked and three result from a transposition to an autosomal location accompanied by an excision of the original insert in the same gamete. As with the P- red derivatives, the great majority (89.7%) of the lighter derivatives involve ?(-linked events.
Further analysis of 17 out of the 26 X-linked lines was undertaken. Their phenotypes are described in Table 2. One exceptional mutant line, named X80, displays the same eye phenotvpe in hemizygous males as well as in homozygous females. This phenotype is identical in an M and a P background (data not shown). For the other 16, the phenotype of homozygous females is lighter than that of hemizygous males, revealing that they remain sensitive to transvection (data not shown). Additionally, unlike X80, all of them remained sensitive to P repression (data not shown).
Genomic DNA from these 17 X-linked mutants was analyzed by Southern blotting: se\wal restriction diges- tions were performed and the blots were sequentially hybridized with probes covering the I'frcr"'] transgene and the 5' flanking sequence of 9.3 (see Figure 1 and MATERIALS ASD IMICTHODS). Southern blot analysis of X80 revealed the presence of one complete I'frcf"] ele- ment not inserted at the original 9.3 site, but at least 8 kb from it (data not shown). However, in situ hybridiza-
FI(;[.RE 6.--Molccular structure of P-rcd deriv- atives. Thr P[af''] clement insertion i n 19DE (9 .3) with the restriction map of flanking sc- quenccs tletermined b y Southern blot ;In;llysis. A l l the P-red derivatives arc X-linked. cxcrpr 7'20. which results from an autosomal insertion (sym- bolized by a zig-zag line). Thr 32 Y.3 tlrrivativcs resulting from ;In cxcision of one of the two cop irs of the original tandem rrpcat are itlentic;d according t o Sourhern blot anillysis. Twenty-four out of 32 were gcneraretl by P strains o r injcction ofa helper Pclrmcnt; the remaining eight cleriv;1- tivcs wcrc inducrtl by A2-3(99R). Thr fivr linrs (X45, S42, SJ3. S28 and 537) displaying more complex deletion events were obtainrd with AT- 3(998). Dashed lines antl dashed hoxrs, tlclcrcrl regions; gray boxes, uncerrainty on the limits of t h c tlrlerion. Refer to Figure I legends for all other symhols. For more legibility, the rcstriction sites have been named scqwntially by letters in alphabetic order. Restriction enzyme sites: a. HnmHI: I), i l indlll; c, Ntol; d and e , i \ 7 m l ; f, flnrrtl-ll; g. ifintllll; I1 antl h', EroRI; i and i', R n t r t I - I I ; j , EroRI; k. Az~ol; I, .Yhol; m, ,4vnI; 11, Mtol; o, NnmHI; p, Snn; q. c \~ml; r. Snn; s, Avnl; t. Snn; 11, polylinkcr (NnvtHI and Hindlll); v, .'\nd; I\.,
,Y/tol; x, BntnHI.
tion shows that the X80 element lies in 19DE like 9.3 (data not shown). Therefore X80appcars to result from a local transposition event a short distance from the starting site, associated with an apparent excision of the donor element. For the 16 other 9.3 derivatives displaying a lighter phenotvpe, molecular analysis has shown no modification of flanking sequences, revealing that the inserts are localized at the original 9.3 insertion site. They can he grouped into two general classes: inter- nal amplifications and double Pelements.
Atnp1;firnlion of t h p originnl dtrplirnfd stvpwnrc Three 9.3 derivatives are due t o an increase in the number o f copies of the previously duplicated 3.44-kb seqnencc. X i resrllts from an amplification from two to three rc- peats, everything else being unmodified (Figure 8). X13 and X8iare the consequence of an amplification from t w o to four repeats (Figure 8). A s for the deletions, the amplifications appear to be precise at this level of analysis. The Ptransposase can therefore induce ampli- fications of a tandem repeat internal to a Pelement, although at a lower frequency than deletions.
I lo trhlp P plrtnPn/.s: The 13 other mutants displaying an increased w.std repression harbor a second element close t o the original insert. The molecular characteris- tics of the different kinds of double P elements are shown in Figme 8. Double Pelements can be classified into three categories. (1) The two elements are com- plete and one element is inside the other. X93 harbors a second P [ r c f " ] element in the 3' region of the original insert, in reverse orientation. The hvo elements arc' \In- modified antl have kept the original duplicated 3.44- kb sequence (Figure 8). PCR amplification antl direct sequencing of the DNA flanking the internal element

P-Induced Localized Rearrangements
A Left
1413
1 31
9.3 g a t c m G G G C G G G A T A G T A C A C A W ? X A E A A A T A A C A T A A
1 31
X6 gatcTTTGGGCGGGATAGTACACAW4AXAXAAATAACATAAGGmTCCCGTCGATA
B Right
X6 TGCCGACGGGACCACCTTATGITATGTTATITCATCATGOTACAGAAGAGAGCGC. . . 31 1
X1 3.1 TGCCGACGGGACCACCCATGATGAAATAACATATATGTTATTTCATCATCATG~TACA~GATACAGAAGAGAGCGC . . 31 % 17 1
1- 17
FIGURE 7,“Genomic DNA sequences flanking the 9.3, X 6 and X13.1 insertion sites, and TIRs sequence of the elements. The sequences were amplified by the inverse PCR procedure, and the product of the reaction was directly sequenced (see MATERIALS AND METHODS). Bold face characters represent the 8-bp duplicated target site. Sequences underlined or topped with an arrow belong to the 31-bp TIRs of the P element. (A) Sequences at the left side of the elements (ie., the 5’ side of the 9.3 and X 6 elements according to their orientation and the 3‘ side of the X13.I element that is in reverse orientation). The 3’ 31-bp TIR of the X13.1 element contains an additional 26 bp of directly duplicated sequence. Small caps indicate the restriction site of NdeII (gatc), which was used in the inverse PCR procedure on the left side. (B) Sequences at the right side of the elements. The 3’ side of the 9.3 insert has not been sequenced. The 5’ 31-bp TIR of X13.1 contains a duplication of the first 17 bp of P sequence in a palindromic structure, associated with the deletion of base pair number 18.
in X 9 3 was performed. This showed that the internal element is inserted at the position 14182 (in white se- quence coordinates) (0’- et al. 1984) at a distance of 1326 bp from the 3’ end of the external element. The intact 5‘ and 3‘ ends of the internal element are flanked by direct duplication of 8 bp of its insertion site in the primary element, as expected from a normal transposition. (2) The two elements are contiguous and one of them is rearranged. Eight independent mutants were shown to result from the presence of two contigu- ous elements in the original 9.3 insertion site, associ- ated with a rearrangement (deletion or inversion) of one of the two elements.
XS, X 8 8 and X 9 8 contain a 5’ element missing its 3‘ region and a second complete P[wd’] element arranged in the same orientation (Figure 8). In the case of X98, the two elements harbor an amplification to three c o p ies of the original 3.44kb tandem repeat.
In the other mutants (X84 , X86 , X96 , X79 and X 7 1 ) , one of the two elements contains an inversion, and the second one is unrearranged (Figure 8). In all cases both elements have kept the original 3.44kb duplication of P[wd’]. The inversion involves either the 3’ extremity of the 5’-most element ( X 8 4 , X 8 6 and X96) or the 5’ extremity of the 3’-most element ( X 7 1 ) . One last mu-
tant, X79, harbors two contiguous elements associated with complex rearrangements that have not been fully elucidated.
The junction between the two elements present in X5 was sequenced after PCR amplification. The 5’-most element ends at position 14137 in white coordinates (0’- et al. 1984): 1371 bp of the 3’ end of p[wd’] are missing. The 5’-most element is directly joined to the 5’ end of the 3’-most p[wd’] element (data not shown),
The same analysis was performed on X 8 6 genomic DNA. A fragment containing almost all the inverted sequence from the 5’-most element and the junction with the 3‘-most element was PCR amplified. Direct sequencing of this junction reveals that the intact 5’ P end of the 3’-most element is joined to position 14088 (in whitecoordinates) (0’- et al. 1984), the inverted sequence therefore consisting of 1420 bp from the 3’ end of P[ wd‘] . As we did not sequence the other junc- tion (5’-most element 3’ P end of the inverted se- quence) we do not know if the X 8 6 inversion is precise or if some nucleotides are missing or duplicated.
(3) The two elements are arranged in a palindromic structure. After Southern blot analysis, X64 , X65 , X67 and X 7 5 appeared identical. They display a second ele-

1414
P[wd 1 9.31
x7
X 13, X87
x93
X5, X88
X98
X84, X86, X96
X71
M. Delattre, D. Anxolab6htke and D. Cohen
II II I I I I a bc def gh i h' i' jklmnopqrs t u v wx , 5kb ,
a bcdef gh i h' i' h' i'jklmnopqrs t u v wx
II II I I I I a bc def g h i h' i' h' i' h' i' jldmno pqrs t u v w x
a II II lr I II II I II II I I I
a bc def gh i h' i' jklmno pqrs tu t srqp onmlk/ i' h' i h g u v wx
a bc def g h i h' i' jktnnopqrstgh i h' i' jkbnno Wrs t u v wx
I II I I ;Ill IIII I+ I I I
I II I d T F W n I IIn F T F R n 11 I 11 n I t I I I
I I I I I ! P 4 Y w l II I I I II I FfyYl I I I I II II 11 1 I I
a bcdef gh i h' i' h' i' j l d m pqrs tgh i h' i' h' i' j l d m pqrs t u v wx
a b c d e f g h i h' i'jklrmo pqrs u tgh i h' i'jklrmo pqrs t u v wx
a bcdef gh i h' i' jklrmopqrs t ui' h' i h g j k l m pqrs t u v wx
X64, X65, X67, X75 1 1 1 a bc def u t srqp omkj i hggh i jkhno pqrs t u v wx
FIGURE 8.-Molecular structure of P transposase-induced 9.3 lighter derivatives. All these 9.3 derivatives were generated by the use of A2-3(99B) as a source of transposase, except X7, X13 and A%, which were induced by injection of the helper P element into z w 9.3 embryos. The phenotypes corresponding to the different X-linked lighter derivatives are given in Table 2. Legends are as in Figure 6. X 1 3 and X87 are strictly identical (by Southern blot analysis). The structures of X5 and X88 are similar but differ only by the size of the 3' missing sequence: 1371 bp in x5 and =2 kb in X88. The two elements of X98, although harboring an amplification to three copies of the original tandem repeat, are otherwise arranged as in X5. X84, X86 and X96 display a similar double P structure, with a 5'-most element containing an inversion of its 3' extremity and a second unmodified element. The size of the inverted sequence is -1.4 kb in X86 and X96, and 1.0 kb in X84. The 3'-most element in X71 contains a Gkb inversion of its 5' extremity, the other element is unmodified. X64, X65, X67 and X75 are strictly identical according to Southern blot results.
ment in a reverse orientation, localized very close to the 5' end of the original 9.3 insertion. The two ele- ments are 5' to 5' and display the same deletion of one of the two copies of the originally duplicated 3.44kb sequence (Figure 8). From Southern blot data, the two elements appear contiguous: the preciseness of this method leads us to conclude that there is <50 bp be- tween the two elements. So far our attempts to clone, PCR amplify and sequence the junctions between the elements have been unsuccessful, presumably because of their palindromic structures.
In conclusion, all the 9.3 derivatives more sensitive to zeste' repression are due either to an internal ampli- fication of the number of ZBR or to the integration of a second element within or very close to the starting insert.
Analysis of autosomal inserts resulting from the trans- position of 9.3: Eighty-two independent autosomal in-
sertions generated by the transposition of 9.3 (77 de- scribed in COEN 1990; five in this study) were examined. Most of them display a wild-type eye coloration. Only two are sensitive to P repression when in a zeste' back- ground. One, T37, is localized at cytological position 36AB (as determined by in situ hybridization, not shown). The other line, z w; CyO, C4/T(2;3)apxa was recovered in a search for insertions located on the Cy0 balancer chromosome.
Molecular analysis of 29 of these autosomal inser- tions, including the two that proved to have a cytotype- dependent expression, was performed by genomic Southern blotting (data not shown).
In 18 cases, the internal structure of the P[wd'] ele- ment appeared unchanged after transposition. These cases include T37, one of the autosomal insertions sen- sitive to P repression. In nine cases, the insert appeared to have undergone a precise deletion of the 3.44kb

P-Induced Localized Rearrangements 1415
duplication, leading to a structure analogous to that of X6. One of these lines contains two insertions of the precisely deleted transgene. In one case, an imprecise deletion had occurred (see T20 in Figure 6). Finally the C4 insertion displays an amplification of the duplicated sequences to four copies, leading to a structure similar to X 1 3 or X 8 7 (data not shown).
Therefore, 38% of the new insertions to autosomal locations are associated with modifications of the trans- posed insert (internal deletions or amplifications) re- sembling those recovered at the original insertion site.
Reversion of X6 insensitivity to P repression: X6, which results from a precise deletion of one copy of the original tandem repeat internal to 9.3, is insensitive to P repression (Figure 5). Presuming that this insensi- tivity was due to a lower sensitivity to zeste' repression, we decided to look for derivatives more sensitive to zeste' repression. To do so, the z w X 6 line was put in dysgenic conditions according to the crossing scheme depicted in Figure 3, and we looked for F2 (Sb+) females exhib- iting an eye color lighter than the normal red of homo- zygous X 6 females. They occurred at a rate of 7.8%. The modified X chromosomes, which they received from their dysgenic fathers, were recovered and 17 lines were established.
Males from these various lines display eye phenotypes ranging from almost wild type to orange, including spotted colorations. Their phenotypes are described in Table 2. All but one (X6.4) show a reversion to sensitiv- ity to P repression: they display a darker eye color in an M background than in a P background (data not shown).
Hemizygous males and homozygous females of the exceptional X6.4 mutant line display the same spotted orange phenotype, in M and P backgrounds. Molecular analysis of X6.4 reveals that the mutation results from a 1.4kb internal deletion (Figure 9) that removes the eye-specific enhancer and the ZBR (@AN et aZ. 1992). The 16 other X6 derivatives are double P elements lo- cated at the same site as the original X6 insert.
Southern analysis of X6.23 reveals that it contains two nested elements in opposite orientation (Figure 9). This structure is very similar to that of X93 (a 9.3 derivative, see Figure 8). They differ only by the number of the 3.44kb 5' regulatory regions of white gene and by the insertion site of the second element within the first (insertion at position 14182 in white sequence coor- dinates in the external element for X93, and 4300 bp downstream of that position for X6.23) . No nested ele- ment in direct orientation was recovered.
All the 15 remaining X 6 derivatives analyzed contain two very close elements in reverse orientation. Most of them (13) display, at the original site, the same double Xktype element in a 5' reverse configuration (see X6.20 and others in Figure 9). They are identical to one cate- gory of 9 . 3 derivatives described above (X64, X65, X67 and X75, see Figure 8) at the level of Southern analysis.
X6.9 differs from the 13 previous ones only by the fact that both elements contain the same small deletion of -500 bp of white sequences (Figure 9).
X6.7 is the only mutant displaying a structure not already seen with 9.3 derivatives. X6.7 harbors a double X6like element, at the original X6site, the two elements being in reverse orientation (Figure 9). However the two elements are arranged 3' to 3' instead of 5' to 5', like all the other double P elements that we have described before. In conclusion, all the X6 derivatives more sensitive to zeste' repression are due to the pres- ence of two nested or very close elements in reverse orientation.
A derivative of x13 is unable to transpose or to ex- cise: In a test for mobilization of X 1 3 (a 9.3 derivative harboring four copies of the original duplicated se- quence) (see Figure 8), a variant appeared that was selected because of its peculiar phenotype. Instead of the X 1 3 light halo patterned coloration, this derivative, named XI?. I , exhibits a red crescent-shaped pattern at the posterior part of the eye (data not shown) reminis- cent of the w"""' allele phenotype (PETERSON et al. 1994). X13.1 remains sensitive to P repression (data not shown). However, it seems insensitive to P transposase: it neither transposes nor excises nor gives derivatives with an altered phenotype. A stock harboring the XI?. I element together with the A2-3(99B) element was maintained during more than one year with no detect- able modification. In situ hybridization on polytene chromosomes showed that the X13.1 insert was at the same site as X13 (19DE, data not shown). This was con- firmed by Southern blot analysis. However, X13.1 ap- peared to be in the opposite orientation, as compared to X 1 3 or 9.3. Besides, the internal structure of the element appeared unmodified: it had kept the four copies of the 5' regulatory sequences present in XI?.
Analysis of the ends and flanking sequences of the X13.1 element was undertaken by inverse PCR and se- quencing (Figure 7). It revealed that the flanking se- quences on both sides are exactly the same as those of the 9.3 insert, the 8-bp duplicated target sequence remaining unchanged, and confirmed the reverse ori- entation of X l 3 . I (Figure 7). In addition, it showed that the 31-bp TIRs have been modified at both XI?. I ends. On the 3' side, the 31-bp TIR contains a direct duplica- tion of 26 bp (Figure 7A). At the 5' end, the first 17 bp of P sequence have been duplicated in a reverse orientation, giving a palindromic structure, and base pair number 18 (following the palindrome) is deleted (Figure 7B).
DISCUSSION
The results presented here show that P transposase induces preponderantly localized events on the p[zd'9.?] insertion. The types of X-linked events that we have recovered are summarized in Table 2 with their esti-

M. Delattre, D. Anxolabehere and D. Cohen 1416
TABLE 2
Types and frequencies of 9.3 and X6 X-linked derivatives ~ ~~ ~ ~ ~ ~~ ~ ~~ ~~ ~
ZBR Percentage Name Phenotype" number Molecular structure' Total' of events"
9.3 derivatives Internal rearrangements
Amplification of the 3.44 kb white duplicated sequence
x 7
X 1 3 and X 8 7
X 9 8
Dark orange halo
Yellow halo
Quasi white
3
4
2 x 3
1
1
2 x 1
1
2 x 1
2 x 2 2 x 2
2 x 2
2 x 3
2 x 2
2 x 2
2 x 2
2 x 2 1 x 2
0
2 x 1
One element containing precise amplification to three copies
One element containing precise amplification to four copies
Two contiguous elements in direct orientation (see below) containing precise amplification to three copies
One element with a precise deletion of 3.44kb repeat
One element with a precise deletion of 3.44kb repeat and a modification of the flanking sequences.
configuration (see below) containing a precise deletion
Unprecise deletion encompassing one ZBR or undetermined deletion
Two elements in reverse 5'
Both elements are complete
No double insertion in reverse 3' configuration
Both elements are complete One element with a 1.4kb deletion
of its 3' end/one complete element
One element with a 2.0-kb deletion of its 3' end/one complete element
of its 3' end/one complete element
One element with a 1.4-kb deletion
One complete element/one element with a 6.0-kb inversion of its 5' end
One element with a 1.0-kb inversion of its 3' end/oue complete element
One element with a 1.4kb inversion of i t s 3' end/one complete element
Complex rearrangements Excision of the original element and
insertion of an unmodified element at 2 8 kb (but still in IYDE)
One element containing a 1.4kb internal deletion that removes the ZBR
Two elements in reverse 5' configuration (see below) containing the same internal deletion of -500 bp
4
38
3'
4
0
1 7
1 1
2
1.6
12.8
0.9
1.6
0
0.4 2.8
0.4 0.4
0.9
Precise deletion of the duplicated sequence
X 6 and 31 others
X 4 2 and X45,
P-red
P-red
X64 , X65 , X67 and X 7 5
Brown spotted
Imprecise deletion X 2 8 , X 3 7 and X4?, P-red
Double P elements Separated by few bp
Reverse 5' configuration
Reverse 3' configuration
Nested Reverse orientation Direct orientation
(see discussion)
X64 , X65 , X67 and X 7 5
Brown spotted
Orange halo Yellow halo
Yellow halo X 8 8
X98 Pale yellow
x 7 1 Brown halo
X 8 4 Orange halo
Orange halo X 8 6 . X 9 6
Unknown orientation Insertion at a distance
x79 X 8 0
Brown Brown spots on
an almost white background
X 6 derivatives: Internal rearrangements
Imprecise deletion X 6 . 4
X 6 . 9
Orange spotted
Brown spotted

P-Induced Localized Rearrangements 1417
TABLE 2
Continued ~ ~ ~ ~
ZBR ~~
Percentage Name Phenotype" number Molecular structureb Totalc of eventsd
Double P elements Separated by few bp
Reverse 5' configuration X6.20 and 12 others Brown spotted 2 X 1 X6.9 Brown spotted 2 X 1
Reverse 3' configuration X6.7 Quasi-red 2 x 1
Reverse orientation X6. 23 Red 2 x 1 Direct orientation
Nested
Both elements are complete 14 6.4 Both element have the same
internal deletion of -500 bp Both elements are complete 1 0.5
Both elements are complete 1 0.5 (No double element in direct 0 0
orientation)
" Eye phenotype of hemizygous males at 25". *All these elements are localized at the original 9.3 insertion site except X80. 'Some double P derivatives have been considered twice: once as a double P element and once as an internal rearrangement
Estimated percentage of each type of event among the gametes of 9.3or X6males bearing A2-3(99B) as source of transposase (X98, X64, X65, X67, X75 and X6.9).
(based on the data from Table 1 and text). 'See Figure 6 for an example.
mated frequencies. The frequencies of the different events vary depending on the sources of transposase (Table 1 ) . As most of the derivatives studied were ob- tained with A2-?(99B), their frequencies of occurrence will be referred to in the following discussion and used to estimate the relative frequencies of each type of deriv- atives (Table 2). We observe high rates of deletions or amplifications of internal repeats and/or occurrence of double P elements in various configurations. Recently,
other authors have reported results similar to ours: transposition at close proximity (EGGLESTON 1990; DAN- IELS and CHOWICK 1993; TOWER et al. 1993; Z W G and SPRADLING 1993; DORER and HENIKOFF 1994) and internal amplification or deletion of repeated se- quences (PAQUES and WECNEZ 1993; KURKULOS et al. 1994). Our experimental design allows us to recover X- linked events as well as transposition to other chromo- somes in the progeny of the same cross and therefore
X6 I II II I I I I
a bc def gh i jkhno pqrs t u v wx
a bcdef gh jkhnopqrs t u v wx
X6.23 II I II II lr I II II I I I ~ I I I
a bc def gh i jklmno pqrs tu t srqp onmllQ i hg u v wx
X6.20 + 72 others I I I I IIII I I I1
a bc def u t srqp mlkj i hggh i j W m pqrs t u v wx
a bc def u t srqp ormlkj i hggh i jkhno pqrs t u v wx
X6.7 U I I IIII I I I I IIII Ill I I I a bcddgh i j k l m pqrs t u u t srqpomJkj I hg v wx
1 S k b
FIGURE 9.-Molecular structure of P transposase-induced X6 derivatives. Mutants were recovered according to the mating scheme shown in Figure 3. Dashed boxes, deleted regions. Other legends are as in Figure 6. X6.4contains an internal deletion that removes the ZBR and the eye-specific enhancer. The 13 X6derivatives that display double Pelement in 5' reverse configuration are identical at the level of Southern blot analysis. X6.9 differs from them only by a small deletion of -500 bp, contained in both elements. This deletion appears included in the first large intron of the gene (see Figure 1) and therefore does not dramatically influence the white expression.

1418 M. Delattre, D. Anxolabkhere and D. Cohen
to assess their relative frequencies. Localized events ap- pear to occur at a high rate (18.7%) in the progeny of dysgenic 9.3 males. The first conclusion that can be drawn is that their frequency is by far greater than the frequency of transposition to autosomal sites (4%). If all localized events are the consequence of the repair of the break caused by the excision of the target P element (ENGELS et al. 1990), one should conclude that all excisions do not lead to an insertion event as ex- pected from a conservative model for transposition. Ei- ther most of the excised elements are lost after excision (abortive transposition) or most of the events do not intrinsically give rise to a transposition intermediate (see below). Alternatively, it could be possible that most of the transposed inserts have lost the whitef gene ex- pression and are therefore undetectable. This could be due either to internal rearrangements or to position effect associated with the new insertion site (e.g. , hetero- chromatic sites). P transposase induces deletions and amplifications
of directly repeated sequences at high frequency: In all cases, the X-linked P-red derivatives result from the loss of one of the ZBR. Most of them have undergone a precise (at this level of resolution) deletion of one copy of the original tandem repeat contained in P[ w‘”] [34/37 (91.8%), including the two cases where it was accompanied by another event]. This type of event is also recovered among double P elements localized at the original site ( X 6 4 , X65, X67 and X75) and among insertions to autosomal sites (9/29). The deletion of one copy of the original tandem repeat is therefore present in up to 14% of all the gametes of dysgenic 9.3 males.
In contrast, some mutants that display a higher sensi- tivity to zeste’ repression are due to an increase to three or four copies of the 3.44kb tandemly arranged repeat in the P[wn’] transgene. Internal amplification occurs at the original site within simple (X7, X13, X87) or double insertions ( X 9 8 ) and is also found within a trans- posed insert. This type of event represents 1.7% of all the gametes of dysgenic 9.3 males.
The kind of rearrangements that we have observed here have also been reported by other authors. PAQUES and WEGNEL (1993) have described frequent deletions and amplifications of tandemly repeated ribosomal 5s genes located within a Pelement. KURKULOS et al. (1994) have observed 2% deletions between direct repeats sepa- rated by several kilobases of unique sequences. More recently, THOMPSON-STEWART et al. (1994) have de- scribed transposase-induced deletions and amplifica- tions of repeated sequences flanking a P element. Hence, it appears that P transposase frequently induces deletions and amplifications between direct repeats con- tained within a P transgene or outside it.
Internal deletions and amplifications require the in- tegrity of P element ends: The exceptional derivative X13.1 is insensitive to any transposase-induced re-
arrangement: in dysgenic conditions, the X13.1 insert appears unable to excise, to transpose or to undergo internal deletion or amplification, although it contains four tandem repeats of the 3.44kb white 5‘ sequence. Sequence analysis reveals that both X13.1 TIRs are mod- ified (Figure 7). The addition of 26 bp directly repeated at one Pend has already been described and has been shown not to prevent transposition (TAKASU-ISHIKAWA et al. 1992). Thus the immobility of X13.1 is probably not due to the 26-bp direct duplication at the 3‘ TIR but rather to the presence of the palindromic repeat of 17 bp associated with a 1-bp deletion at the other terminus (Figure 7). This is in agreement with the study of MULLINS et al. (1989) that showed that the integrity of the 31-bp inverted repeat is absolutely required for transposition.
This result clearly shows that, as with transposition, internal deletions and amplifications of direct repeats require, at least, an intact 5’ TIR. They are therefore a direct or indirect consequence of the process of P transposase-induced excision and/or transposition.
Mechanism for excisions and amplifications of direct repeats: According to the SDSA (synthesis-dependent strand annealing) model (NASSIF et al. 1994), Pelement excision results in a double-strand break (Figure 10A) that can be extended by degradation of the ends. Each 3‘ end then invades independently a homologous se- quence and acts as a primer for the initiation of DNA repair synthesis (Figure 10B). The growing single strands are released while the replication proceeds, dis- placing only a local loop of DNA (bubble migration). When overlapping sequences have been replicated, i.e., when the growing strands contain homologous se- quences, the two extended termini can anneal. The process is then terminated as in the single strand an- nealing (SSA) model (LIN et al. 1984; HABER 1992; Su- GAWARA and HABER 1992) by removal of any nonhomol- ogous sequences (that may have been synthesized beyond the overlap), DNA repair synthesis (each strand serving as a template for the other), and ligation.
The SSA model has been proposed to explain dele- tions between direct repeats in yeast (HABER 1992; SU- GAWARA and HABER 1992). KURKULOS et al. (1994) have used this model to interpret the occurrence of deletions between internal direct repeats included in a Pelement: when homologous sequences are encountered on both growing strands, annealing between them is possible leading therefore to premature termination of the re- pair process. To explain amplifications from two to four repeats, one needs to add some features to this model.
1. The invading strand could change template during the repair process, making a “copy-choice” possible (Figure 10). The work of GONZY-TR&BOUL et al. (1995) strongly supports the hypothesis of template change: they described chimerical elements that contain se- quences from two P elements used successively as tem- plates for the repair process. When repeats are encoun-

P-llltlrlccd I.oc;dizc.tl R(~;~r r ;~~lg(~~l~c .~ l~s 1.1 I!,
=LA A
G dj =I3 a
r,
- 0 0 __
1 - I 3 f "F c
0 - Restoration ( ~ f t h c tluplicatiorl 0 1 dc-lction
;3 D
- I klction
0 0 -3 &F 'A E
0 - Restoration o f thc duplication 0 1 dc-letion
. 3 F
- Restoration o f thc duplication or dcletion or tri- or quadriplication
F K ; ~ T W lO.-Mrcl1;mism of gcmralion o f drlc-tions ;Ind amplifications. Thin c r ~ n c d arrows inrlic;ttc* the position ;lnd clirc-c-tion o f t l w j m p hrtwvrc-n tcmlpl;ltcs ( s w t c ~ ) . Otlwr synllwls arc- ;IS in Figrwr I . Although t h c rcpcats ;~rr idrntic;d, lor t h c sitkc of clarity tllr diagonally harrhrtl p;Ittc*rns s y - holizirlg t h r n l ; ~ r c t l i f i i w - n t . (A) ' l o p : tloul)lr-stI-;mtl DSh I)rmk gcncmrrtl b y Pcwision. Rot tom: t l w r l n n l o t l i l i c t l sistrr cI1rom;ltitl. ( I ) ) Invasion of tlw rc-rnplatr by ~ h r I,rokrn I)Sh cntls. ;trcortling t o t h r S D S A motlcl (SASSIF ef crl. I ! W ) . ((1- F) Resolution of t h r rcp;Iir procrss: 1l1c monlcnt j u s t brforr t h r ;Irlnc-aling I)ctwecn t l ~ c Ilonlologorls scqucnccs prcscvlt o n both n;lsrrnt strands is rrpr(wntrd. V;wious possil)ilitics arc shown, tlcyxwling OII wllc~hrr o r n o t t l l r in\.;ding strand wi l l rhangr tcw1pl;llc.s ( D - F or (I), whrthcr t h c . , j t l m p of t h v inwding c m t l l ion1 o11c rrpcat t o t h r o t h c r o r r ~ ~ r s forward (I)) o r t)arkw;wtl (I< ;mtl F ) ; lnd wlwthrr o r 1101 tlw rrpliciltion procrctls ;~ltc*r I l l ( . c*nc'o~~ntrr of homologorw srqwnrrs o n b o t h growing strands ( F o r E). I n t h r c;~srs whrrc* mort Illill1
onr rc.pc;~t I1;n hrrn copic-tl o n onr of r h r strands. I h r rc~solu- tion wi l l tlepcntl o n tllc. honlologorls scqurncrs t h a t w i l l an- ncal i n thr last s ~ r p of the rrpair (not rrprrsrntctl) ( s w text for rxpl;\nation).
tercd, this capacity for changing tcmpl;ltt,s wil l a l l o w jumping backward or fonmrtl hctwccn repeats, leading t o amplifications o r dc+-tions.
2. The repair DNA synthesis should n o t ;11ways l w terminated as soon 21s homologous regions arc copied on both strands [ like in thc hornolo~-mor~itoring modcl proposctl by K[XMI.OS c/ nl. ( I W ~ ) ] . A s alrcittly post~~lated (Nnsstr- c/ nl. 1994), in some cases, thc repair
DNI\ syntlwsis could proccc~l lilrtl1cr, allowillg ill1lplili-
cations t o o c c ~ ~ r (scc I~clow ;Ind Figllrc IO). In o 1 1 r rc- s ~ ~ l t s , t h c ratio o f ' tlclction 79.v. ;~tnplific;rtion is highly 1,i;lsctl toward tlc+*tions ( 14% tlclctions conlp;~rctl t o
1.7% atnpli1ic;ttions in thc whole. progct~y o f ' 9.3:A2- 3(99R) milles). Jumping I)ctwccw rcpc;tts should occur a t similar frc-qucncics l im\xrrl and hackw~rtl, Iwtlillg t o ;Ipproximatcly c y ; d frcqllcwcics of dclctions ant1
;~mplific;~tions. This high csccss of clc*lctions can I)<* cs- p1;lincd i f \oc considcr that the growing singlc strat~tls arc in constant sc*;~rch for honlology. Thcrc*lim-, i n !nost ~ I I I not in ;dl c;~scs, t l l c rc-pair process 1vo111cI he tmni-
rcrc*tl. 111 thc / 7 r d " ] transgcnc, thc sizc o f 11w /' 3' end
+ t w o drlplicatc~tl regions (7470 111)) is ; ~ p p ~ o ~ i ~ ~ l ; ~ t ~ * l ~ cquivalcnt t o the sizc of the utliqw rcgioll o f r c & i / ~ + I' 3 f cntl (7481 bp). Tllcrcforc. if tllc c.xtcnsion pr(b ccwls at a sitnil;\r spcccl from both ?if cntls. they wi l l rc>l\cll holnologorls sc*qwnccs ;lpprosimatcIy it1 the mitldlc of thc / 7 7 d " ] c.lcmc.nt (Figure IO(:). I ' l lc* right- sick strand w i l l hc a h l c t o pair w i t h the Ic4i-sitlv strand, with cqtlal frcquctlcics, t o rc-pcat 2 (restoring thc- clrlpli- cation) or I (1c;ltling t o a dclction). If fon\.;u.tl.jltmpillg occurs (Figure 101)). the rc*srdt w i l l he a rlrlc*tion in most i f not ;dl c;~scs. If hsck\vard jumping O C C I I ~ S , in most C;ISCS only t w o rc.pcats woultl h v c hecn copictl on the Icft-sitlc strand \ohen the right-side strand wi l l rc;wh thc rcpc;~tcd rcgion (Figurc IOE). This w i l l rcntlt again in clclction o r restoration of thc tl~tp1ic;Itio~l with c * q l d prohahilitics. dcpcnrling on which rcpcat the right-siclc strand w i l l ant1c;d to (1 o r 2, rcspccti\rly). Thc I - C Y Y ) V C I ~
of an ;Implilic;ltiotl will r q u i r c that the repair p r o c c ~ is n o t intcrrrlptrd ;IS soon ;IS homology is (wcoImtcrcd. I n this case (Figure IOF). i t is possihlc t o obtain ;I tlclc- tion. restoration of the dllplication, o r tri- o r quxlripli- cation, tlc.pending on which rcyxat of thc Icft-sitlr str;Ind ( I , 2 o r 3) will anneal t o which repcat o f thr right-side strand ( 1 or 2). Thc overall rcsult will I ) c ;I
strong cxccss of dclctions o r restorations of the drtplica- tion 7t.v. amplifications. Onc prediction of this modcl is that thc ratio o f t h c tlifii.rc*nt types ofevents wi l l stro~lgly dcpcnd o n thc sizc and position of rrpc;lts h r l t ;~lso 0 1 1
their scyucncc. which co111tl haw a11 dYcct on the spcctl of thc rcplication and its stalling a t sonic positions (we (:I I ~ I X fg/ crl. 1994 and rcfcw*nccs thcwin).
I n thc PcIcmcnts ;wsayctl hy KIXW.I.OS P/ rrl. (19!)4), c.ilcll r c p ~ a t is close t o o11c cntl . allowing ol l ly (01. mostly) dclctions. This wi l l hold also for tlw 1;1rgc* csccss of extcrnal tlclctions 7,s. amplifications ohsc*~~ctl by TI l o ~ ~ l ~ s o s - ~ l . ~ \ ~ . . \ ~ r c/ crl. ( 1904). Thc. clcnlcnt assayrd by P..ic>l-w ;Ind \~'I(;sE% (1993) contains c4ght short rcpe;\ts (3.50 hp) loc;~tcd at one end and ;I rmiquc rc- gion (the YO.$!' gcnc, 7.2 kh) milch longer t h a n t l w r c p c ~ c c l region a s a whole (2.8 kh). Thcrcforc*. l,c.forr both growing str;lnrls rc;~ch an homologous sc.qwllcc. thcrc w i l l I>c cnough tinw for multiplc occurrcncc of
tliltcd ;Is soot1 ;Is llolllologorls Scq""1c"s !Yere cIlcollll-

1420 M. Delattre, D. Anxolab6hi.re and D. Cohen
jumps between repeats, by changing template. This will lead to an almost equal rate of deletions and amplifica- tions of various number of repeats.
Double P elements occur at high frequency: Besides internal amplifications, the great majority of 9.3 deriva- tives (13 out of 16) and all X 6 derivatives (16) that display a higher sensitivity to zeste' repression are due to the presence of a second element in close association with the donor element. These double insertion events occur at up to 5.2% in the progeny of dysgenic 9.3 males and at up to 7.4% in the progeny of dysgenic X 6 males. In all the cases of double insertions analyzed (29), the second element is adjacent to or integrated into the original insert.
A number of previous reports have described mutant alleles containing two or more closely integrated P ele- ments (NITASAKA et al. 1987; HAWLEY et al. 1988; ROIHA et al. 1988; EGGLESTON 1990; O'HARE et al. 1992; DAN- IELS and CHOVNICK 1993; TOWER et al. 1993; ZHANC and SPRADLINC 1993; DORER and HENIKOFF 1994). There- fore, the high frequency of multiple insertions appears to be a general property of the P element. This has been interpreted as resulting from a great tendency of the P element to transpose in the proximity of a preexisting element (EGGLESTON 1990).
Nested Pelements: Among the 29 double Pelements described in our study, two events can be clearly inter- preted as insertions inside the starting element: one out of 13 9.3 double P derivatives (X93) and one out of 16 X 6 double P derivatives ( X 6 . 2 3 ) . In both cases the two elements are in opposite orientation. The fact that sequences flanking the internal element in X93 harbor an 8-bp duplication, strongly argues in favor of a classi- cal P insertion.
In addition, eight contiguous double Pelements were observed among 9.3 derivatives, but none among X 6 derivatives. All of them are associated with rearrange- ments (inversion or deletion) modifymg the extremity of one of the elements. One interpretation for the ori- gin of this type of elements could be that they result from multiple nested insertions followed by excision and gap-repair events, according to the model of ENG ELS et al. (1990) (see above). Deletion and inversion would result from the choice of the homologous se- quence used as a template for repair on the sister chro- matid. Mechanisms for the origin of nested rearranged double Pelements that we have observed are proposed in Figure 11.
Regardless of the orientation, the second element seems to integrate frequently into the 3' region of the white transgene rather than into its 5' side. Eight out of the nine nested P elements that we have fully analyzed can be interpreted as resulting from a second insertion into the 3' region of white transgene. Five out of eight appear to be clustered in a region between 14088 and 14182 of white coordinates (O'HARE et al. 1984), as de- duced from the three sequencedjunctions of X5, X 8 6
A: x5. x88. X98
4
N
B: X71
J
k I
c: X84. X86. X96
I
FIGURE 11.-Model for the origin of nested rearranged double P elements. Nested insertions (---) are followed by excision of the internal element(s). This leaves a gap that is repaired by the invasion of the unmodified sister chromatid by the broken 3' ends (half-arrows) . We have represented the choice of templates that accounts for the observed structures. Refer to Figure 1 for all other symbols. (A) The structure of X 5 (similar to X 8 8 and X 9 8 ) could result from three successive events: insertion of a second element within the 3' side of 9.3 in direct orientation, excision of the internal element, and repair of the gap. The homologous template chosen for this repair would be the internal 3' end, leading to the loss of one of the duplicated 3' extremities in the resulting element. The same types of events, starting with two new insertions within the 5' side or 3' side of 9.3, would lead respectively to X 7 1 (B) , and X 8 4 , X 8 6 and X 9 6 ( C ) .

P-Induced Localized Rearrangements 1421
and X 9 3 (Southern analysis of two other derivatives, X 9 6 and X98, gave similar results). This region is close to, but does not include, the previously described hot spot for P insertions (RUBIN et al. 1982) that lies at position 13221 of whitecoordinates (0’- and RUBIN 1983). The three other cases of insertion into the white 3’ region ( X 8 4 , X 8 8 and X6.23) are dispersed in sites 500-1000 bp from the cluster region. Only X6.23 could result from an insertion in the conventional white hot spot, according to Southern blot results. This suggests that the specificity of P element insertion does not de- pend only on the sequences immediately flanking the insertion site. It can be influenced by sequences located at least 1 kb away from the insertion point in the white gene, as this specificity is modified in the P [ d * ] envi- ronment.
However it should be noted that all the nested ele- ments arranged in direct orientation share two features. (1) In all cases, both elements are identical in the sense that all have either retained the duplication (23, X88, X71, X79, X84, X86 and X96) or have undergone a triplication of the direct repeats ( X 9 8 ) . (2) In all of them, one element has one rearranged end. This does not happen when the two nested elements are in oppo- site orientation ( X 9 3 and X6.23): in this case, the two elements are both complete and unrearranged (besides the insertion of one element inside the other). Conse- quently, as discussed further, the possibility remains that the observed structures do not result from succes- sive and independent actual events of transposition, excision and gap-repair, but rather from the involve- ment of one single element in an aberrant and complex gap-repair event, leading to its partial or rearranged duplication.
Double P elements in close proximity: All of the re- maining (not nested) double insertions (19/29) con- tain two very close copies of the Pelement. These results are consistent with those of ECGLESTON (1990) who found that the vast majority of secondary insertions at the yellow and singed loci occurred within 50 bp of the primary element or into the inverted repeats. Of the 13 double-insertion alleles described by DANIELS and CHOVNICK (1993), eight contained a second element integrated into or immediately adjacent to the starting element. Such very closely integrated P elements have also been described by many other authors (HAWLEY et al. 1988; ROIHA et al. 1988; TOWER et al. 1993; ZHANC and SPRADLING 1993).
However, all these authors have also described dou- ble elements separated by intermediate or long dis- tances (0.2-200 kb). ZHANC and SPRADLINC (1993) showed that this type of event is more frequently ob- served in the progeny of females than those of males. However, they observed in males that 30% (4/13) of new insertions occurred at >6 kb from the donor ele- ment. In our study we have not recovered this type of double element structure. Such events would have been
detected as they would have predominantly resulted in a P-red phenotype like the autosomal insertions. This observation suggests that in our study the nature of the target element and/or of the flanking sequences (position effect) may have strongly influenced the type of events resulting from transposase action, in favor of transpositions at very short range.
Double Pelements have been thought to result from the tendency of Pelements to transpose inside or near other P elements (EGGLESTON 1990). This implies an affinity between the Pintermediate of transposition and inserted P elements. P elements containing a fragment of engrailed (HAMA et al. 1990; KASSIS et al. 1992) or polyhomeotic (FAUVARQUE and DURA 1993) are subjected to transposition to specific targets (homing). The same authors have proposed that this homing may result from an affinity between proteins bound to the se- quences included in the P element and to the target site. Double P elements may result from the same type of homing: for instance, the transposase that has affinity for P elements may remain attached to the excised P element and target it to other Pelements inserted any- where in the genome.
However, this mechanism cannot account for the very strong bias that we observed in favor of insertions in the 5‘ side rather than in the 3’ side of an inserted element (18 out of 19 not nested double Pelements). This strong preference for the 5‘ side was also observed by EGCLESTON (1990), TOWER et al. (1993) and ZHANC and SPRADLING (1993).
Besides the 5‘ side preference, we observed a very strong preference for reverse orientation. All the dou- ble P elements in close proximity that we have found (19) are arranged in reverse orientation (opposite ori- entation relative to the donor insert). Most (18) are 5’ to 5’, and in only one (X6.7) the two elements are 3’ to 3’. The same observation can be made for the multiple insertions described by other authors (HAWLEY et al. 1988; ROIHA et al. 1988; ECCLESTON 1990; DANIELS and CHOVNICK 1993; TOWER et al. 1993; ZHANC and SPRADLINC 1993; DORER and HENIKOFF 1994).
To account for this strong bias, ZHANG and SPRAD- LING (1993) and TOWER et al. (1993) have postulated that proteins would maintain the excised element in a fixed orientation and tightly associated to the donor site. This would lead the excised element to transpose preferentially at close proximity, with the orientation preference observed.
However, if double P elements result from transpo- sition of an excised element near another one, it is expected that the two elements may differ in certain occasions, given the high frequency of internal modifi- cations that we observe. All the double P elements in close proximity that we have observed contain two iden- tical elements. The two elements are unmodified (X6.20 and 12 others), have undergone the same pre- cise deletion of one copy of the originally duplicated

1422 M. Delattre, D. AnxolabChere and D. Cohen
A
B
C
D
E
J
I
FIGURE 12.-Model for the origin of double Pelement in reverse orientation. Small arrows, endonucleolytic cuts; dashed arrows, progression of the repair DNA synthesis. Other symbols are as in Figure 1. (A) Staggered cuts at the 5' end of P element. (B) Intramolecular ligation of the 5' and 3' broken ends to form a hairpin structure at the P 5' end. This hairpin structure will be resolved by a single-strand cut at the 3' end of the Pelement. On the other side, the 3' free end could invade the sister-chromatid and begin the repair process, regenerating an apparently unmodified flank- ing sequence (as suggested by Southern blot data). Alterna- tively, the free ends of this side could also be ligated to form a second hairpin structure. Resolution on this side would then occur by a cut in the single-stranded region of the hairpin. (C) Repair synthesis begins from the 3' ends. (D) The free ends are joined by ligation on the left side (curved line) and repair DNA synthesis proceeds through the hairpin. (E) This results in two closely integrated Pelements in 5' reverse con- figuration. The same process starting with staggered cuts at the 3' end of the P element will lead to a double P element in 3' reverse configuration.
sequence (X64, X65, X67, X75) or display the same little internal deletion (X6. 9).
To explain the identity of both members of a double P element in reverse orientation, we propose that, in most cases, they do not result from a local transposition event. They may rather result from a reverse duplication of the original element, subsequent to an abortive trans- position, according to the model described in Figure 12. This model shares some features with the models proposed for the excision of plant transposable ele- ments (SAFDLER and NEVERS 1985; COEN and CARPEN- TER 1988).
First, the P transposase would induce staggered cuts at only one P element end. It has recently been shown (HANIFORD and KLECKNER 1994) that cleavages at the two ends of the bacterial transposable element TnIO can occur at observably distinct times before transposi-
tion. Similarly, the two P termini may be not simultane- ously cleaved by the transposase. Then, ligation of the free ends would occur to form a hairpin structure. For the DNA molecule containing the P element, the hair- pin structure may be resolved by a single-strand cut at the 3' end of the element (Figure 12). The two DNA molecules are then ligated and repaired. Replication would proceed through the hairpin, giving rise to an inverted duplication of the starting Pelement.
The nature of the junction sequence between the two elements would be of utmost interest for an understand- ing of the mechanism generating these structures. Un- fortunately, our attempts to clone, PCR amplify or se- quence these regions have so far been unsuccessful, as for other authors (HAWLEY et al. 1988; ROIHA el al. 1988; EGGLESTON 1990).
The preponderance of 5' to 5' over 3' to 3' double Pelements could be due to the preferential occurrence of the first transposase cut at the 5' end of the P ele- ment. DANIELS and CHOVNICK (1993) observed small deletions (50-100 bp) in the adjacent 5' genomic DNA of several derivatives, but never in the 3' flanking DNA. These deletions may result from the extension by exo- nucleases of the first cut made by the transposase, pref- erentially at the 5' side of P elements.
In conclusion, at least three mechanisms may be in- volved in Pelement "transposition" for very short, me- dium and long distances. First, the model in Figure 12 could explain the production of double inserts in re- verse orientation integrated into the same target site. Second, the recovery of insertions at nearby sites or into the original P element without additional re- arrangements could imply an intermediate of transposi- tion that has a tendency to stay nearby the donor site. Finally, jumps to distant sites could arise by disassocia- tion of the excised P element from the donor site. The potential existence of different mechanisms for local- ized rearrangements and transposition to unlinked sites is reminiscent of the finding that intermolecular and intramolecular events of the IS911 transposable ele- ment are mutually exclusive (POLARD et nl. 1992). As with recombination mechanisms, the initial break(s) made by the P transposase may be resolved by using different pathways, most of them failing to engender genuine transposition.
P repression on 9.3 and its derivatives necessitates at least two ZBR in close proximity: P repression on 9.3 acts through the enhancement of the zestr' repression on this transgene and is strongly position dependent
Here we show that 9.3 derivatives insensitive to P repression are due to in situ modifications of the origi- nal insert (X-linked P-red mutants) or, less frequently, to transposition to a new genomic position. All the in situ modified P-red mutants have lost one copy of the originally duplicated 5' regulatory sequences.
This shows that the presence of at least two ZBR is a
(COEN 1990).

Hnduced Localized Rearrangements 1423
necessary condition for P repression. It confirms our previous conclusions showing that P repression involves zeste' repression on 9.3 and acts by enhancing it (COEN 1990). Amplifications leading to three or four copies of the regulatory regions in a direct tandem array do not prevent P repression and may even enhance it (data not shown). The same is true for secondary insertion at a short distance. The insensitivity to P repression due to the loss of one ZBR can even be reverted by the insertion of the same element at a short distance (e.g., X 6 and its revertants).
The two autosomal insertions that are sensitive to P repression have either retained the original duplication of whitf regulatory sequences or harbor an amplifica- tion to four copies of these sequences. Among the 27 autosomal insertions insensitive to P repression that have been molecularly analyzed, 10 have been modified upon transposition and lost one of the ZBR. The re- maining 17 appear to have retained the duplication of the regulatory sequences and thus two ZBR. They are nevertheless insensitive to P repression. This is also the case for X80, which has transposed at a nearby site. This confirms that, besides the presence of two ZBR, the flanking chromosomal sequences are important for the occurrence of the P repression that appears extremely sensitive to position effects, like the zeste-white interac- tion (LEVIS et al. 1985a).
Our allelic series of mutants, harboring various num- bers of regulatory regions of the same gene inserted exactly at the same localization, will be useful to study these interactions and the phenomenon of transvection as it alleviates the problem of position effects that often complicates these kinds of studies.
We thank D. N o u r l u for her excellent technical help and advices. R. LEVIS and L. R \ R I N O W kindly provided all the clones used to synthe- size the white probes. Special thanks to C. Z U C ~ R and D. TIIIERRY- MIK: who provided the original z' r ~ " ' ~ P [ d ' 9 . 3 ] (19DE) stock and shared unpublished informations on its structure. We are very grate- ful to A. PI.KSSIS, .J. DELTSCH and D. POINSO?' for fruitful discussion and their comments on the manuscript. We are particularly grateful to .J. HABER for stimulating discussions and his comments on the manuscript. This research was supported by the Centre National de la Recherche Scientifique and by a grant from the Groupement de Recherche et d'Etude sur les Genomes. M.D. was supported by fellow- ships from the Minist2re de I'Enseignement Supkrieur et de la Re- cherche a d from the Ligue Nationale Contre Le Cancer.
LITERATURE CITED
ANXOIAB~HER~.:, D., H. BENES, D. NOUAUD and G. PERIQUE^‘, 1987 Evolutionary steps and transposable elements in Drosophila mela- nogastrr: the missing RP type obtained by genetic transformation. Evolution 41: 846-853.
BANGA, S. S., and J. B. BOW, 1992 Oligonucleotidedirected site- specific mutagenesis in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 89: 1735- 1739.
BUSSON, D., M. G m s , K. KOMITOPOCJI.OU and M. MASSON, 1983 Ge- netic analysis of three dominant female-sterile mutations located on the X chromosomes of Drosophila mdanogaster. Genetics 105:
CHEDIN, F., E. DERWN, R. DERWN, S. D. EHRIKH and P. NOIROT, 1994 Frequency of deletion formation decreases exponentially
309-325,
with distance between short direct repeats. Mol. Microbiol. 12(4): 561-569.
COEN, D., 1990 Pelement regulatory products enhance zeste' repres- sion of P ( white'L''~'""'d) transgene in Drosophila melanogaster. Ge- netics 126: 949-960.
COEN, E. S., and R. CARPENTER, 1988 A semidominant allele, niv- 525, acts in trans to inhibit expression of its wild-type homologue in Antirrhinum majus. EMBO J. 7: 877-883.
DANIEIS, S. B., and A. CHOVNICK, 1993 P element transposition in Drosophila mlanogastm: an analysis of sister-chromatid pairs and the formation of intragenic secondary insertions during meiosis. Genetics 133: 623-636.
DORER, D. R., and S. HENIKOFF, 1994 Expansions of transgene re- peats cause heterochromatin formation and gene silencing in Drosophila. Cell 77: 993-1002.
EG(;I.ESTON, W. B., 1990 P element transposition and excision in Drosophila: interactions between elements. Ph.D. thesis, Univer- sity of Wisconsin, Madison.
ENGEIS, W. R., 1984 A trans-acting product needed for P factor transposition in Drosophila. Science 226: 1194- 1196.
ENGEIS, W. R., 1985 A set of P-cytotype balancer chromosomes. Drosophila Inform. Sew. 61: 70-71.
EN(;EI.s, W. R., 1989 Pelements in Drosophila melanogaster, pp. 437- 484 in Mobib DNA, edited by D. E. BERC, and M. M. Howx. American Society for Microbiology, Washington, DC.
ENGEIS, W. R., D. M.JoHNsoN-S(:1-11,17z, W. B. ECXI.ESTON and J. SVEI), 1990 High-frequency P element loss in Drosophila is homolog dependent. Cell 62: 515-525.
FALIVARQLIE, M.-O., and J.". DIIR4, 1993 polyhomeotic regulatory se- quences induce devlopmental regulatordependant variegation and targeted P-element insertions in Drosophila. Gene Dev. 7: 1508-1520.
GONZY-TR~BOUI., G., J. A. LEPESANT and J . DEUTSCH, 1995 En- hancer-trap targeting at the Broad-Complex locus of Drosophzla me- lanogastrr. Gene Dev. 9: 1137-1148.
GI.OOR, G. B., N. A. NASSIF, D. M. JOWNSONSCHIJTZ, C. R. PRESTON and W. R. ENGEI.S, 1991 Targeted gene replacement in Dro- sophila via P element-induced gap repair. Science 253: l l 10- 1117.
HABKR,.~. E., 1992 Exploring the pathways of homologous recombi- nation. Curr. Opin. Cell. Biol. 4: 401-412.
HAMA, C., Z. AI.I and T. B. KORNBERC,, 1990 Region-specific recom- bination and expression are directed by portions of the Drosoph- ila rngrailpd promoter. Gene Dev. 4 1079-1093.
HANIFORD, D., and N. KLECKNER, 1994 TnlU transposition in vivo: temporal separation of cleavages at the two transposon ends and roles of terminal basepairs subsequent to interaction of ends.
HAWI.EY, R. S., R. A. STEIJRER, C. H. MARCUS, R. SOHN, D. M. BARONAS et a/., 1988 Molecular analysis of an unstable P element inser- tion at the singed locus of Drosophila melanogastm: evidence for intracistronic transposition of a Pelement. Genetics 119: 85-94.
HAZEI.RI(;(;, T., 1987 The Drosophila whitr gene: a molecular u p date. Trends Genet. 3: 43-47.
HAZEI.RIG(:, T., R. LXVIS and G. M. RLJBIN, 1984 Transformation of white locus DNA in Drosophila: dosage compensation, ze.rtr interaction and position effects. Cell 36: 469-481.
JOHNSON-S(:HI.ITZ, D. M., and W. R. EN(;EI.s, 1993 P-element-in- duced interallelic gene conversion o f insertions and deletions in Drosophila mlanogastrr. Mol. Cell. Biol. 13: 7006-7018.
JUNAKOVIC, N., R. CANFVA and P. BAl.l.ARl0, 1984 Genomic distribu- tion of copia-like elements in laboratory stocks of Drosophila mela- nogaster, Chromosoma 90: 378-382.
KAREss, R. E., and G. M. RUBIN, 1984 Analysis of P transposabk element functions in Drosophila. Cell 38: 135-146.
KASSIS, J. A,, E. NOIL, E. P. VANSIC:KI.E and W. F. ODENWAI.D, 1992 Altering the insertional specificity of a Drosophila transposable element. Proc. Natl. Acad. Sci. USA 89: 1919-1923.
KAUFMAN, P. D., and D. C. RIO, 1992 P element transposition in vitro by a cut-and-paste mechanism and uses GTP as a cofactor. Cell 69: 27-39.
K m w w . I , , M. G.,.J. F. KIDWEI.I. and J. A. SWD, 1977 Hybrid dysgenesis in lhsophila mlanogmter: a syndrome of aberant trait . including
KURKLJI.OS, M.,J. M. WEINRF.RG, D. ROY and S. M. MOUNT, 1994 P mutation, sterility, and male recombination. Cenetics 86 813-833.
element-mediated in vivo deletion analysis of white-appncott dele-
EMBO J. 13: 3401 -341 1.

1424 M. Delattre, D. Anxolab6hkre and D. Cohen
tions between direct repeats are strongly favored. Genetics 136: 1001-1011.
LEMAITRE, B., and D. COEN, 1991 P regulatory products repress in
Acad. Sci. USA 88: 4419-4423. vivo the P promoter activity in P-hcZ fusion genes. Proc. Natl.
LEVIS, R., T. HAZELRIGC: and G. M. RUBIN, 1985a Effects of genomic position on the expression of transduced copies of the whitegene of Drosophila. Science 229: 558-561.
LEVIS, R., T. HAZELRIGG and G. M. RUBIN, 1985b Separable &-acting control elements for expression of the white gene of Drosophila. EMBO J. 4 3489-3499.
LIN, F.-L., K. SPERLE and N. STERNBERG, 1984 Model for homolc- gous recombination during transfer of DNA into mouse L cells: role for DNA ends in the recombination process. Mol. Cell. Biol. 4: 1020-1034.
LINDSIXY, D. I,., and G. G. ZIMM, 1992 The Genome of Drosophila melanogaster. Academic Press, San Diego.
MANIATIS, T., E. F. FRITSCH and J. SAMBROOK, 1982 Molecular Chn- ing: A Laboratmy Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
MISR&, S., and D. C. RIO, 1990 Cytotype control of Drosophila P element uansposition: the 66 kd protein is a repressor of trans- posase activity. Cell 62: 269-284.
MUILINS, M. C., D. C. RIO and G. M. RURIN, 1989 Cisacting DNA sequence requirements for P-element transposition. Gene Dev. 3: 729-738.
NASSIF, N., J. PENNEY, S. PAL, W. R. ENCELS and G. B. GLOOR, 1994 Efficient copying of nonhomologous sequences from ectopic sites via P-element-induced gap repair. Mol. Cell. Biol. 1 4 1613- 1625.
NITASAIW, E., T. MUKAI and T. YAMAZAKI, 1987 Repressor of P ele- ment in Drosophila melanogaster: cytotype determination by a de- fective P element with only open reading frames 0 through 2. Proc. Natl. Acad. Sci. USA 8 4 7605-7608.
O'HARE, K., and G. M. RURIN, 1983 Structure of P transposable elements and their sites of insertion and excision in the Drosophila melanogaster genome. Cell 34: 25-35.
O'HARE, K., C. MURPHY, R. LEVIS and G. M. RUBIN, 1984 DNA se- quence of the white locus of Drosophila melanogaster. J. Mol. Biol. 180: 437-455.
O'HARE, K., A. DRIVER, S. MCGRATH and D. M. JOHNSON-SCHLITZ, 1992 Distribution and structure of P elements from the Drosoph- ila melanoguster P strain ~ 2 . Genet. Res. 60: 33-41.
PAQLJFS, F., and M. WEGNEZ, 1993 Deletions and amplifications of tandemly arranged ribosomal 5s genes internal to a P element occur at a high rate in a dysgenic context. Genetics 135: 469-476.
PETERSON, K. M., P. S. DAVIS and B. H. JUDD, 1994 The determined state of white expression in the Drosophila eye is modified by zeste' in the dm family of mutants. Mol. Gen. Genet. 242: 717-726.
PIRROTTA, V., 1991 The genetics and molecular biology of reste in Drosophila melunoguster. Adv. Genet. 29: 301 -348.
PIRROTTA, V., and C. BR~CKL, 1984 Transcription of the Drosophila white locus and some of its mutants. EMBO J. 3: 563-568.
PIRROTTA, V., H. STEILER and M. P. BOZETTI, 1985 Multiple u p
stream regulatory elements control the expression o f the Drosoph- ila white gene. EMBO J. 4 3501 -3508.
POLARD, P., M. F. PRERE, 0. FAYET and M. CHANDLER, 1992 Transpos- aseinduced excision and circularization of the bacterial inser- tion sequence IS911. EMBO J. 11: 5079-5090.
QIAN, S., B. VARJAVANII and V. PIRROTTA, 1992 Molecular analysis of the rpste-whiteinteraction reveals a promoter-proximal element essential for distant enhancer-promoter communication. Genet- ics 131: 79-90.
RIO, D. C., F. A. LASKI and G. M. RUBIN, 1986 Identification and immunochemical analysis of biologically active Drmophilu P ele- ment transposase. Cell 44: 21-32.
ROBERTSON, H. M., and W. R. ENGELS, 1989 Modified Pelements that mimic the P cytotype in Dro.rophilu mehogaster. Genetics
ROBERTSON, H. M., C. R. PRESTON, R. W. PHIILIS, D. JOHNSON- SCHIJTZ, W. K. BENZ et al., 1988 A stable genomic source of P element transposase in Drosophila melanogaster. Genetics 118: 461 -470.
ROIHA, H., G. M. RUBIN and K. O'HARE, 1988 Pelement insertions and rearrangements at the singed locus of Drosophila melnnoguskr. Genetics 119: 75-83.
RUBIN, G. M., M. G. KIDW.I.I. and P. M. BINGHAM, 1982 The molecu- lar basis of P-M hybrid dysgenesis: the nature of induced muta- tions. Cell 29: 987-994.
RUBIN, G. M., T. HA%EI.RIGG, R. E. KARESS, F. A. LASKI, T. LAVERIY d uL, 1985 Germline specificity of P element transposition and some novel patterns of expression of transduced copies of the whitegene. Cold Spring Harbor Symp. Quant. Biol. 50: 329-335.
SACRAMENTO, D., H. BOURHY and N. TORDO, 1991 PCR technique as an alternative method for diagnosis and molecular epidemiol- o g y of rabies virus [published erratum appears in Mol. Cell Probes 1991 Oct;5(5):397]. Mol Cell Probes. 5: 229-240.
SAEDI.ER, H., and P. NEVER?, 1985 Transposition in plants: a molecu- lar model. EMBOJ. 4 585-590.
SPRADIJNG, A. C., and G. M. RUBIN, 1982 Transposition of cloned Pelements into Ihvsophilu germline chromosomes. Science 218: 341-347.
SUC~AWARA, N., and J . E. HABER, 1992 Characterization of double- strand break-induced recombination: homology requirements and single-stranded DNA formation. Mol. Cell. Bid. 12: 563-575.
TAKASU-ISHIKAWA, E., M. YOSHIHARA and Y. HOTTA, 1992 Extra se- quences found at P element excision sites in Dro.wphila melanogus- ter. Mol. Gen. Genet. 232: 17-23.
THOMPSON-STEWART, D., G. H. WEN and A. C . SPRADI.IN(:, 1994 A transposable element can drive the concerted evolution of tandemly repetitious DNA. Proc. Natl. Acad. Sci. USA 91: 9042- 9046.
TOWER, J., G. H. KARPEN, N. CKAIG and A. C. SPRADI.ING, 1993 Prefer- ential transposition of Drosophila Pelements to nearby chromo- somal sites. Genetics 188: 347-359.
ZHANG, P., and A. C. SPRADIJNG, 1993 Efficient and dispersed local Pelement transposition from Drosophila females. Genetics 188: 361-375.
123: 815-824.
Communicating editor: M. J. SIMMONS
















![[3,3]-Sigmatropic rearrangements - Massey Universitygjrowlan/stereo2/lecture11.pdf · 123.702 Organic Chemistry Claisen rearrangements • One of the most useful sigmatropic rearrangements](https://static.fdocuments.in/doc/165x107/5adcada77f8b9a213e8bd8b0/33-sigmatropic-rearrangements-massey-gjrowlanstereo2lecture11pdf123702.jpg)

