
Post-Transcriptional Regulation Biochemistry – Third Edition by Garrett & Grisham Chapter 29 –...
59
Post- Transcriptional Regulation • Biochemistry – Third Edition by Garrett & Grisham • Chapter 29 – Pages 974-981
-
Upload
violet-russell -
Category
Documents
-
view
219 -
download
0
Transcript of Post-Transcriptional Regulation Biochemistry – Third Edition by Garrett & Grisham Chapter 29 –...

Post-Transcriptional Regulation
• Biochemistry – Third Edition by Garrett & Grisham
• Chapter 29 – Pages 974-981

Post-transcriptional processing of mRNA in eukaryotes
Translation closely follows transcription in prokaryote – no nucleus
In eukaryotes, these processes are separated - transcription occurs in the nucleus, translation occurs in the cytoplasm
On the way from nucleus to cytoplasm, the mRNA is converted from "primary transcript" (or pre-mRNA) to "mature mRNA"

What is post-transcriptional regulation ?
• Transcription: The processes of RNA synthesis.
• Post-transcriptional regulation: The regulation AFTER transcription, but not translation.

mRNA degradation mRNA localization
post-transcriptional regulation

Comparison between procaryotic and eucaryotic mRNA
polycistronic
monocistronic

• Introns intervene between exons doesn’t code for a protein
• Exon: protein coding region; intron: noncoding region
• Exon size is much smaller than intron size
• Examples: actin gene has 309-bp intron separates first three amino acids and the other 350 or so
• Chicken pro-alpha-2 collagen gene is 40-kbp (40,000 bp) long, with 51 exons of only 5 kbp total; the exons range in size from 45 to 249 bases
• Most introns (lengths vary from 60-10,000 bps) have stop codons in all 3 reading frames, thus they are untranslatable; they need to be removed - splicing
• Mechanism by which introns are excised and exons are spliced together is complex and must be precise
Eukaryotic genes are splitBecause contain introns and exons

The organization of split eucaryotic genes
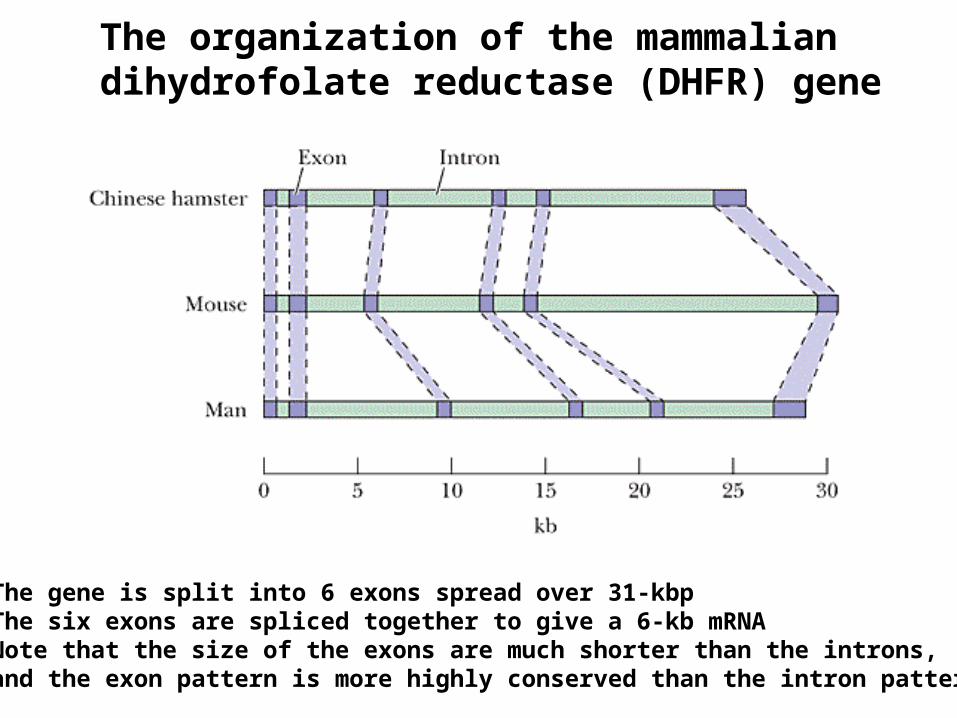
The organization of the mammalian dihydrofolate reductase (DHFR) gene
The gene is split into 6 exons spread over 31-kbpThe six exons are spliced together to give a 6-kb mRNANote that the size of the exons are much shorter than the introns, and the exon pattern is more highly conserved than the intron pattern

• Primary transcripts (pre-mRNAs) are “capped” as soon as they are transcribed by RNA polymerase II
• The reaction is catalyzed by guanylyl transferase using GTP as a substrate
• Capped G residue is methylated at N7-position
• Additional methylation occurs at C2'-O positions of next two residues and at 6-amino group of the first adenine (if A is the initial nucleotide)
Capping and Methylation
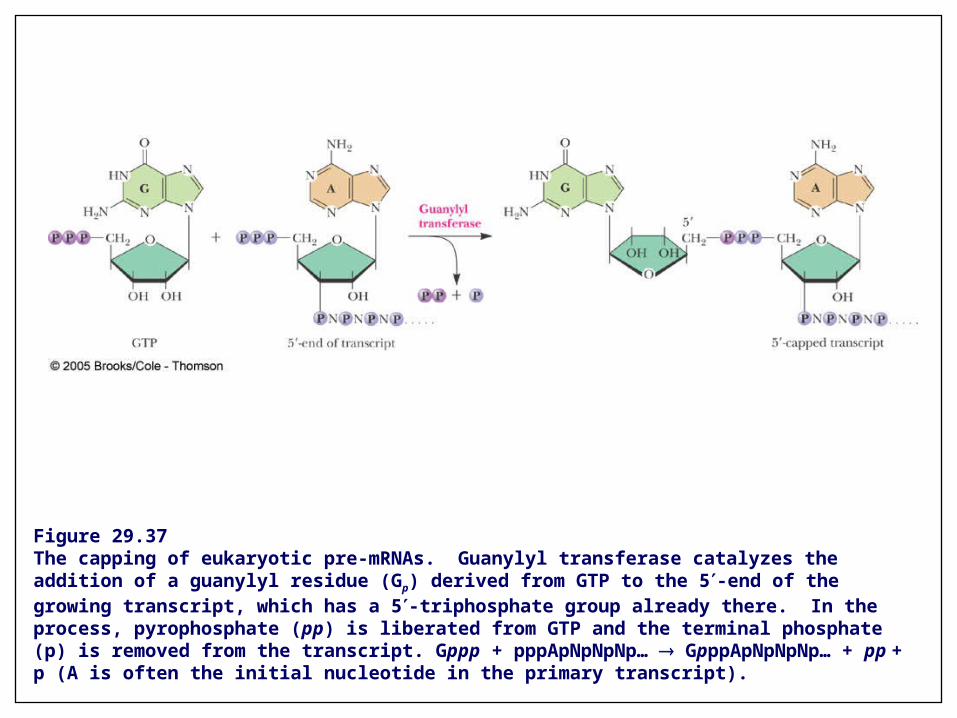
Figure 29.37The capping of eukaryotic pre-mRNAs. Guanylyl transferase catalyzes the addition of a guanylyl residue (Gp) derived from GTP to the 5-end of the growing transcript, which has a 5-triphosphate group already there. In the process, pyrophosphate (pp) is liberated from GTP and the terminal phosphate (p) is removed from the transcript. Gppp + pppApNpNpNp… GpppApNpNpNp… + pp + p (A is often the initial nucleotide in the primary transcript).

Figure 29.38Methylation of several specific sites located at the 5-end of eukaryotic pre-mRNAs is an essential step in mRNA maturation. A cap bearing only a single CH3 on the guanyl is termed cap 0. This methylation occurs in all eukaryotic mRNAs. If a methyl is also added to the 2-O position of the first nucleoside after the cap, a cap 1 structure is generated. This is the predominant cap form in all multicellular eukaryotes. Some species add a third CH3 to the 2-O position of the second nucleoside after the cap, giving a cap 2 structure. Also, if the first base after the cap is an adenine, it may be methylated on its 6-NH2. In addition, approximately 0.1% of the adenine bases throughout the mRNA of higher eukaryotes carry methylation on their 6-NH2 groups.

Phosphatase
Guanylyl transferase
Guanine 7-Methyl transferase
2’ O-Methyl transferase
Enzymes involved in the 5’ Capping

Why do cells need to cap their mRNA?
- cap is recognized by cap-binding proteins
- cap distinguishes mRNAs from other types of RNA molecules (RNA pol I and III produce uncapped RNAs)
- mRNAs need a cap (and poly A tail) for export from the nucleus
- cap is necessary for translation
- cap stabilizes mRNA in the cytoplasm

1. Requires C-terminal domain (CTD) of the largest subunit of RNA polymerase II
2. CTD has repeats (52 copies) of heptapeptide ‘YSPTSPS’. 5 of these 7 have –OH; which are differentially phosphorylated during elongation
3. Phosphorylation of CTD provides binding site for factors involved in capping, splicing, and 3’ end formation
4. These mRNA processing factors are transferred to the nascent RNA at the appropriate time
5. Dephosphorylation of CTD occurs at the end of transcription; necessary for re-initiation
CTD
RNA factory - mRNA processing is coupled to transcription
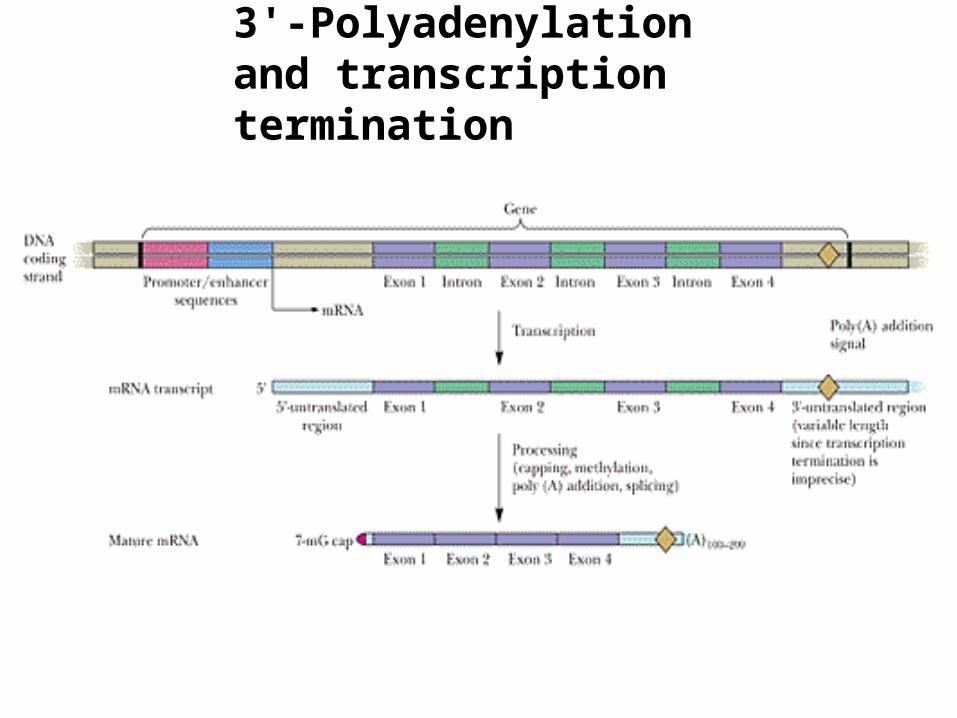
3'-Polyadenylation and transcription termination

• Termination of transcription occurs only after RNA polymerase has transcribed a consensus AAUAAA
sequence - the poly(A) signal
• 10-30 nucleotides after this site [the poly(A) signal], the mRNA is cleaved and a string of ~200 adenine residues is added to the mRNA transcript - the poly(A) tail
• poly(A) polymerase adds these A residues
• poly(A) tail bound by PABP: • stimulates translation and • governs the stability of mRNA
3'-Polyadenylation
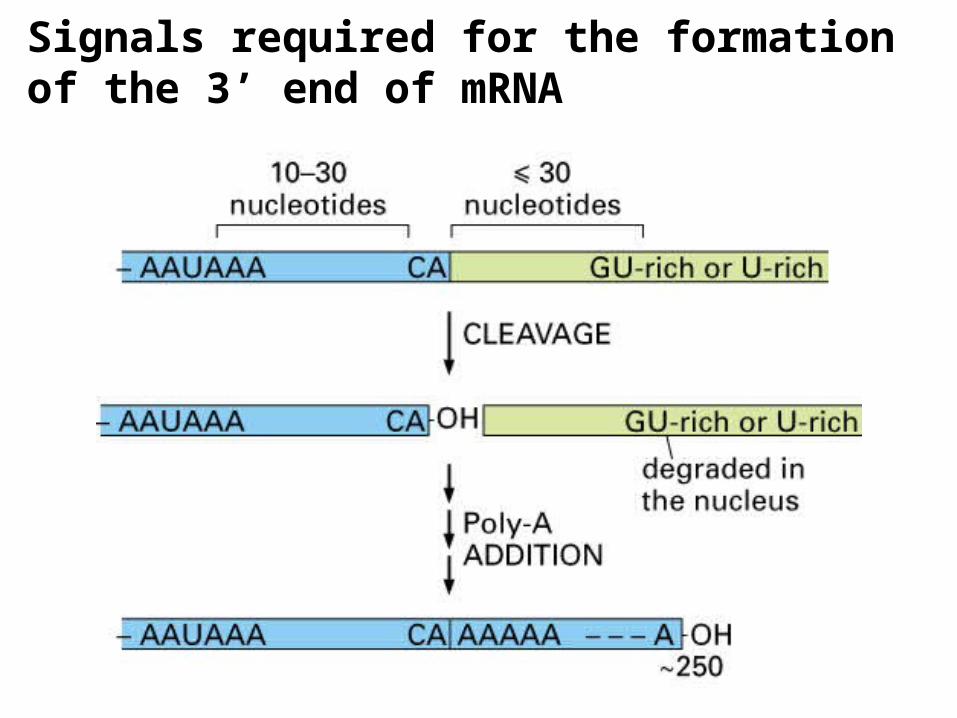
Signals required for the formation of the 3’ end of mRNA

Mammalian pre-mRNA 3’ end processing complex
recognizes the poly(A) signal
recognizes the downstream GU-rich sequences
CPSF- cleavage, polyadenylation specificity factor
CstF- cleavage stimulation factor
CF- cleavage factor

Figure 29.39Poly(A) addition to the 3-ends of transcripts occurs 10 to 35 nucleotides downstream from a consensus AAUAAA sequence, defined as the polyadenylylation signal. CPSF (cleavage and polyadenylylation specificity factor) binds to this signal sequence and mediates looping of the 3-end of the transcript through interactions with a G/U-rich sequence even further downstream. Cleavage factors (CFs) then bind and bring about the endonucleolytic cleavage of the transcript to create a new 3-end 10 to 35 nucleotides downstream from the polyadenylylation signal. Poly(A) polymerase (PAP) then successively adds 200 to 250 adenylyl residues to the new 3-end. (RNA polymerase II is also a significant part of the polyadenylylation complex at the 3-end of the transcript, but for simplicity in illustration, its presence is not shown in the lower part of the figure.)

Mechanism of poly A tail formation - coupled to transcription

Polyadenylation of mRNA
A. Where is the template? - does not require a template - the poly(A) tail is not encoded in the genome
B. What’s the function of the polyA tail? - by interaction with poly(A) binding protein (PABP),
it is necessary for efficient translation and protection from mRNA degradation (2 purposes)

Pre-mRNA splicing
splicing
translation

• Within the nucleus, pre-mRNAs form ribonucleoprotein particles (RNPs) through association with a characteristic set of nuclear proteins
• In "splicing", the introns are excised and the exons are sewn together to form mature mRNA
• The substrate for splicing is the capped primary transcript emerging from the RNA polymerase II in the form of RNP complex
• Splicing occurs only in the nucleus; mature mRNA is then exported to the cytoplasm
• Consensus sequences defining the exon/intron junctions are derived from analysis of the splice sites in vertebrate genes
• The 5'-end of an intron (5’ splice site) in higher eukaryotes is always GU and the 3'-end (3’ splice site) is always AG
• All introns have a "branch site" 18 to 40 nucleotides upstream from 3'-splice site
• 5’ and 3’ splice sites and branch site are essential for splicing
Splicing of pre-mRNA

5’ splice site
3’ splice site
Branch site (usually closer to 3’ss)
What makes an intron?
R: Purine A or GY: Pyrimidine U or C

• Branch site is usually YNYRAY, where Y = pyrimidine (C or U), R = purine (A or G) and N is anything
• The intron is excised as a lariat structure after the completion of splicing
• The "lariat” a covalently closed loop of RNA, is formed by attachment of the 5'-P
of the intron's invariant 5'-G to the 2'-OH at the branch A site
• The lariat is excised when the 3’-OH of the consensus G at the 3’ end of the 5’ exon joins with the 5’-P at the 5’ end of the 3’ exon
• The lariat product is unstable; the 2'-5' phosphodiester is quickly cleaved and the linear intron is degraded in the nucleus
• The reactions that occur are transesterification reactions where an OH group reacts with a phosphoester bond, displacing an OH to form a new phosphoester link
• Because the reactions lead to no net change in the number of phosphodiester linkages, no energy input is needed
The branch site and lariat

Figure 29.41Splicing of mRNA precursors. A representative precursor mRNA is depicted. Exon 1 and Exon 2 indicate two exons separated by an intervening sequence (or intron) with consensus 5, 3, and branch sites. The fate of the phosphates at the 5- and 3’-splice sties can be followed by tracing the fate of the respective ps. The products of the splicing reaction, the lariat form of the excised intron and the united exons, are shown at the bottom of the figure. The lariat intermediate is generated when the invariant G at the 5-end of the intron attaches
via its 5-phosphate to the 2-OH of the invariant A within the branch site. The consensus guanosine residue at the 3-end of Exon 1 (the 5-splice site) then reacts with the 5-phosphate at the 3-splice site (the 5-end of Exon 2), ligating the two exons and releasing the lariat structure. Although the reaction is shown here in a stepwise fashion, 5-cleavage, lariat formation, and exon ligation/lariat excision are believed to occur in a concerted fashion. (Adapted from Figure 1 in Sharp,P.A., 1987. Splicing of messenger RNA precursors. Science 235:766.)

The spliceosome - RNA/protein complex
Composition:
Small nuclear RNAs (snRNAs):
5 snRNAs (U1, U2, U4, U5, U6), called U because it is uridine rich, range in size from 106-189 nucleotide long (small).
Proteins:
10 identified associated with each of the snRNAs to form the small nuclear ribonucleoproteins (snRNPs).
non-snRNP splicing factors: SR-family proteins and other splicing factors
(current estimate of total proteins is >50 different)

The importance of snRNP
• Splicing depends on small nuclear ribonucleoprotein particles - snRNPs, pronounced "snurps"
• A snRNP consists of a small nuclear RNA (100-200 nucleotides long) and about 10 different proteins
• Some of the 10 proteins form a core set common to all snRNPs, whereas others are unique to a specific snRNP


Assembly of the spliceosome
• snRNPs and pre-mRNA form a multicomponent complex called spliceosome
• Spliceosome is about the size of ribosome and its assembly requires ATP
• U1 snRNP binds at the 5'- splice site and U2 snRNP binds at the branch site
• Interactions between the snRNPs brings 5'- and 3'- splice sites together so lariat can form and exon ligation can occur
• In addition to the snRNPs, a number of proteins with RNA-annealing functions as well as ATP-dependent RNA-unwinding activity participate in spliceosome function
• The transesterification reactions that join the exons may in fact be catalyzed by snRNAs themselves, but not by snRNP proteins

Figure 29.42Mammalian U1 snRNA can be arranged in a secondary structure where its 5-end is single-stranded and can base-pair with the consensus 5-splice site of the intron. (Adapted from Figure 1 in Rosbash, M., and Seraphin, B., 1991. Who’s on first? The U1 snRNP-5- splice site interaction and aplicing. Trends in Biochemical Sciences 16:187.)

Figure 29.43Events in spliceosome assembly. U1 snRNP binds at the 5-splice site, followed by the association of U2 snRNP with the UACUAA*C branch-point sequence. The triple U4/U6-U5 snRNP complex replaces U1 at the 5-splice site and directs the juxtaposition of the branch-point sequence with the 5-splice site, whereupon U4 snRNP is released. Lariat formation occurs, freeing the 3-end of the 5-exon to join with the 5-end of the 3-exon, followed by exon ligation. U2, U5, and U6 snRNPs dissociate from the lariat following exon ligation. Spliceosome assembly, rearrangement, and disassembly require ATP as well as various RNA-binding proteins (not shown). (Adapted form Figure 2 in Staley,J.P., and Guthrie,C., 1998. Mechanical devices of the spliceosome: Motors, clocks, springs, and things. Cell 92:315-326.)
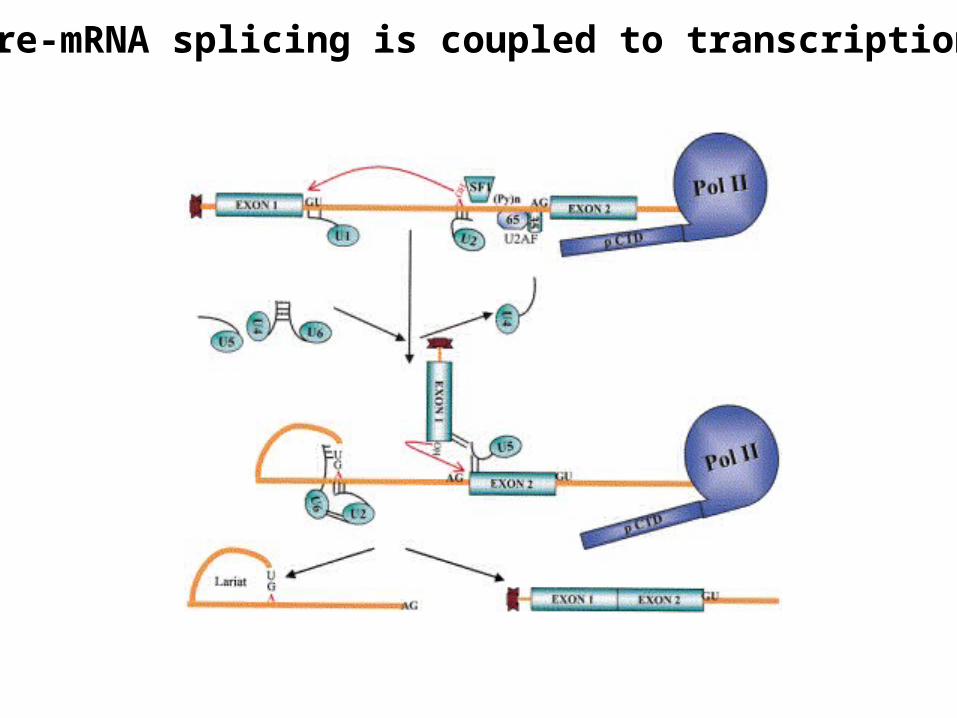
Pre-mRNA splicing is coupled to transcription


Constitutive pre-mRNA Splicing
Every intron is removed and every exon is incorporated into mature RNA without exception – results in a single form of mature mRNA
I II III
I II III
A cell
I II III
I II III
B cell

Alternative pre-mRNA splicing
Pre-mRNA can be spliced in different ways – gives rise to multiple forms of mature mRNAs and increases the coding capacity of the genome
A single gene makes possible a set of related polypeptides, protein isoforms
Occurs because there is intron sequence ambiguity; different choices are made by chance on different transcripts
Several versions of the protein encoded by the gene are made in all cells -constitutive alternative splicing
Regulated alternative splicing - many cases
Switch from production of a nonfunctional protein to the production of a functional one
Cell-type specific splicing - generate different versions of a protein in different cell types
RNA splicing can be regulated negatively, which prevents access to a particular splice site, or positively, which activates an otherwise overlooked splice site.
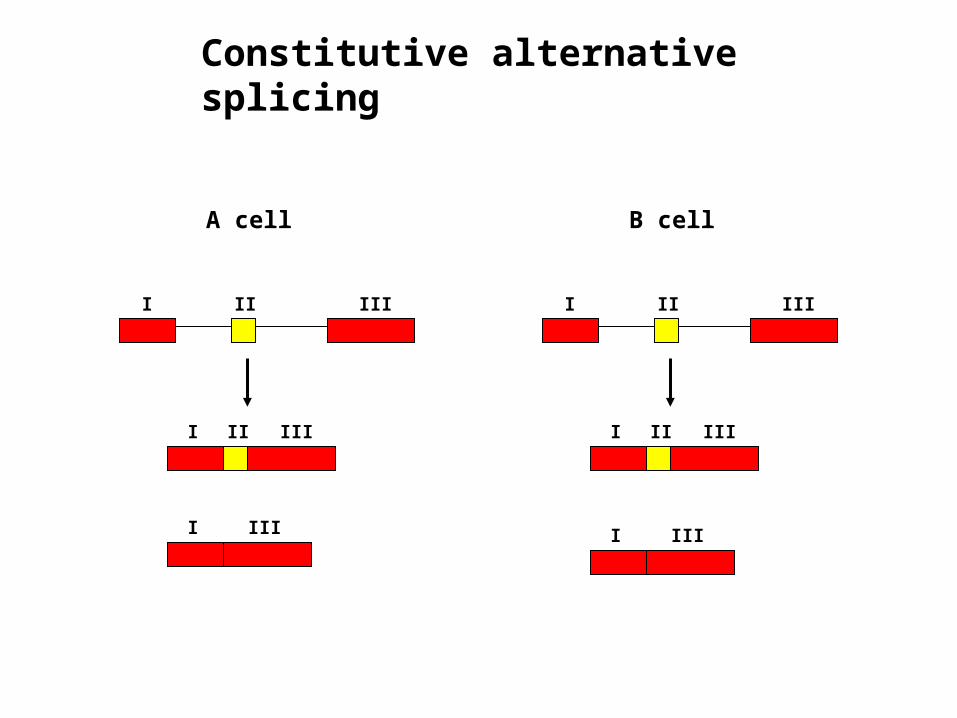
Constitutive alternative splicing
I II III
I II III
A cell
I II III
I II III
B cell
I III I III

Regulated alternative splicing
I II III
I II III
A cell
I II III
I II III
B cell
I III
I II III
C cell
I III

It contains 18 exons
Eleven constitutive exons (exon 1-3, 9-15, and 18) are found in all mature mRNAs (always there)
Five exons (4-8) are combinatorial; (none, 1,2, etc or different combinations) individually included or excluded; 32 possible combinations
Two exons (16 and 17) are mutually exclusive; one or the other is always present never together. Only 2 possibilities
Each mRNA includes all constitutive exons, one of the 32 possible combinations from exon 4-8, and exon 16 or 17.
32 X 2 = 64 possible mature mRNAs can be generated from the primary transcript
Alternative splicing expands the coding potential of the genome
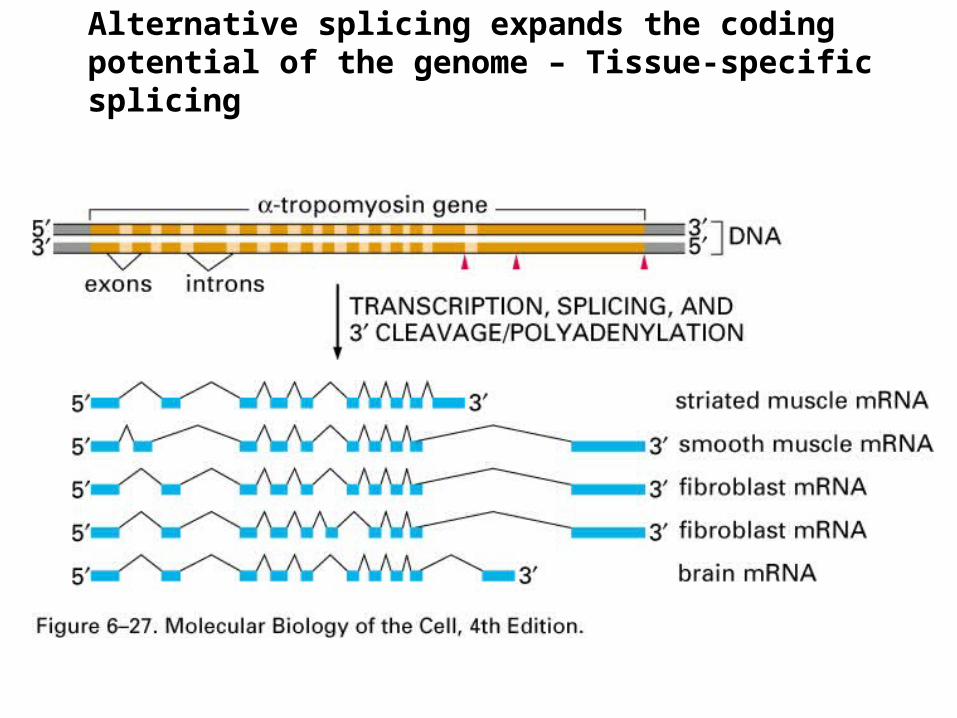
Alternative splicing expands the coding potential of the genome – Tissue-specific splicing

Negative and positive control of splicing

7mGppp AAAAAA
mRNA exportmRNA Quality Control:
Nuclear Retention and Degradation
mRNA processing
mRNA QUALITY CONTROL
7mGppp
7mGppp
7mGppp
AAAAAAlong
normal abnormal

mRNA transport through the nuclear pore complex
phenylalanine-glycine repeats
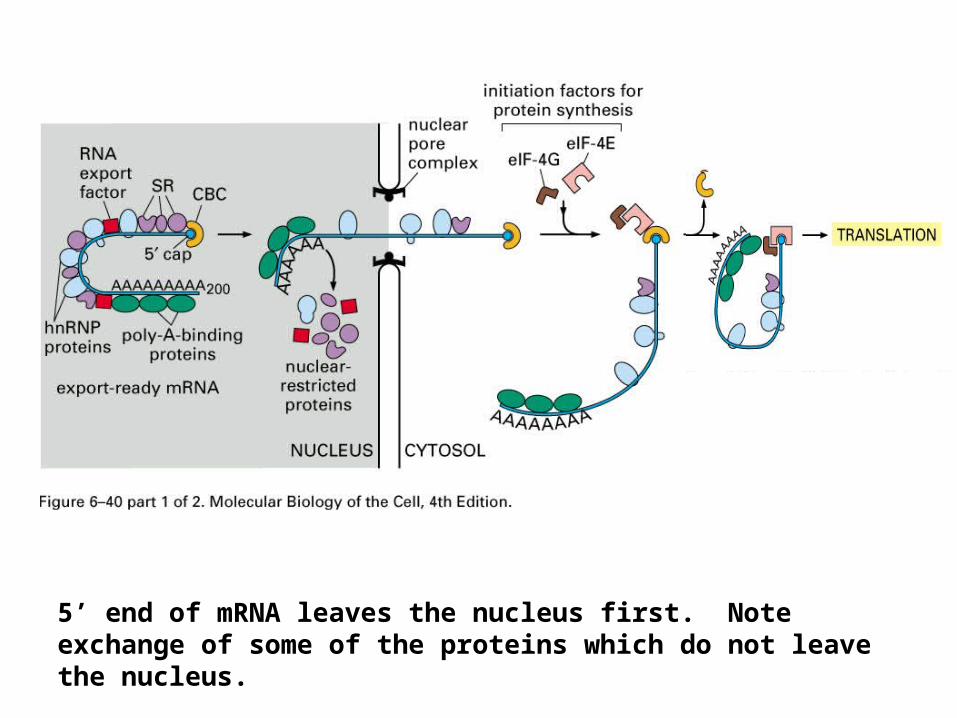
5’ end of mRNA leaves the nucleus first. Note exchange of some of the proteins which do not leave the nucleus.

1. Splicing-dependent recruitment
2. Transcription-dependent recruitment
Two mechanisms by which mRNA export factors are recruited onto mRNAs : two mechanisms

Splicing-dependent recruitment of mRNA export factors
UAP56: a spliceosomal component; deposited onto mRNAs during splicing
ALY forms a complex with UAP56 and is recruited onto the mRNA
TAP/p15 binds ALY and releases UAP56
The mRNP is exported from the nucleus

Transcription-dependent recruitment of mRNA export factors
THO complex: interacts withRNAPII and functions in transcription elongation.
THO complex associates with Sub2p and Yra1p to form TREX (Transcription/EXport) complex.

Fates of mRNAs in the cytoplasm
• Translation
• Degradation
• Localization → translation

mRNA stability
Procaryotes
- Bacterial mRNAs are very unstable- They are synthesized and degraded rapidly- Bacteria can adapt quickly to environmental changes
Eucaryotes
- Most eucaryotic mRNAs are more stable; half-life > 10 hr- Some mRNAs, however, have half-life = 30 min or less; encode regulatory proteins whose production rates need to be changed rapidly in order to respond to environmental changes- Why some mRNAs are unstable?- How mRNAs are degraded in eucaryotic cells?

AAAAAm7G
AAAAAm7G AAAAAm7GAAAAAm7G
Very stable Stable Unstable
-Globin mRNA Most mRNAs Cytokine and protooncogene mRNAs
Stabilizer Destabilizer
Determinants of mRNA stability

Two mechanisms of eucaryotic mRNA decay
General decay pathway: most mRNAs Certain mRNAs: requires specific sequences and specific proteins and specific endonucleases

mRNA localization
Generally mRNAs are translated in the cytoplasm by free ribosome; their products may be directed to other sites in the cell
mRNA encoding secreted or membrane-bound proteins are directed to endoplasmic reticulum (ER) by a signal at the amino terminus of the protein
Some mRNAs are directed to specific intracellular locations before translation begins, i.e. translated at the site where the protein functions
RNA localization may be an ancient mechanism for producing cytoplasmic asymmetry
Basic features of mRNA localization include cis-acting elements within the mRNA that targets that message to a subcellular region, a protein-RNA complex that effects localization, and the cytoskeleton that acts as a “road” for RNA movement
Most localization signal sequences appear to be in the 3'UTR (untranslated region) of mRNAs

The importance of 3’ UTR in mRNA localization
mRNA encoding hairy protein is normally localized to the apical site of nuclei
Injected hairy RNA containing the 3’ UTR
Injected hairy RNA lacking the 3’ UTR

RNA Editing
RNA Editing is the change in RNA sequence after transcription by processes other than splicing
mRNAs in the mitochondria of Trypanosome: one or more U are inserted (or, less frequently, removed); so extensively that over half of the nucleotides in the mature mRNA are U that were inserted
mRNAs in the mitochondria of chloroplast: changes from C to U, without nucleotide insertions or deletions
Much more limited kind occurs in mammals

RNA editing – A to I editing in mammals
Carried out by ADAR- adenosine deaminase acting on RNA, which recognizes a double-stranded RNA region
Such regions form when an exon region containing the A to be edited base pairs with a complementary base sequences in an intron known as the editing site complementarysequence
Occurred in transcripts encoding glutamate receptors; changes a glutamine codon (CAG) to an arginine codon (CGG); altering the conductance properties of the receptor
A U
I C
---
------
---
1. Altering amino acid codon
2. Changing splice site
3. Introducing premature stop codons
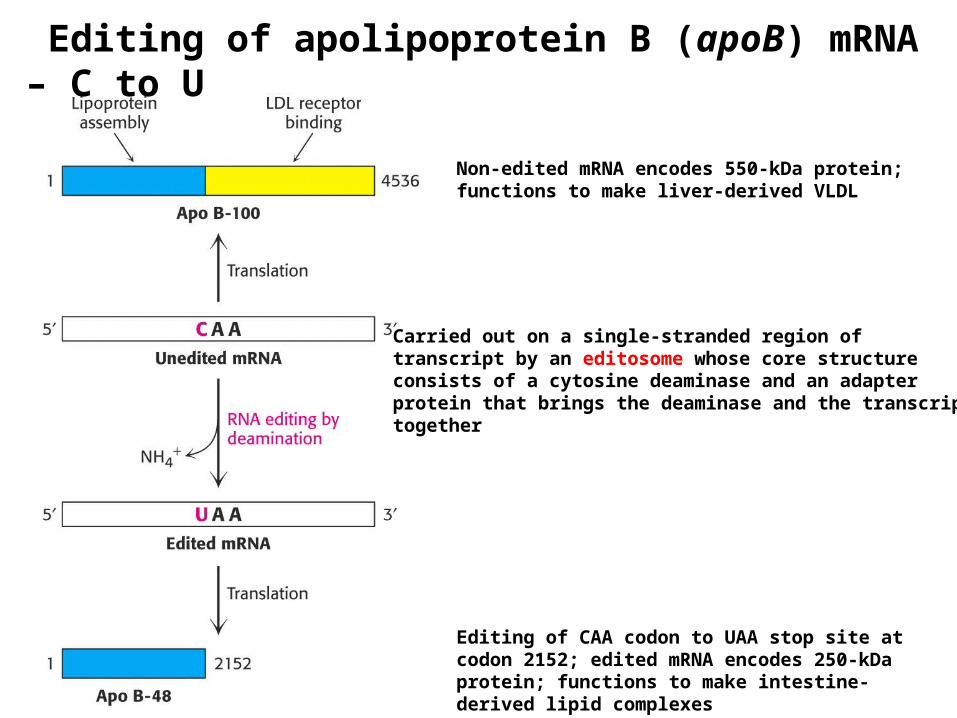
Editing of apolipoprotein B (apoB) mRNA – C to U
Non-edited mRNA encodes 550-kDa protein; functions to make liver-derived VLDL
Editing of CAA codon to UAA stop site at codon 2152; edited mRNA encodes 250-kDa protein; functions to make intestine-derived lipid complexes
Carried out on a single-stranded region oftranscript by an editosome whose core structure consists of a cytosine deaminase and an adapter protein that brings the deaminase and the transcript together

Stages: transcription → transcript processing → mRNA export → translation
Traditionally they have been presented as a linear series of events (a pathway of discrete and independent steps) – each going to completionbefore the next begins
Now it is clear that each stage is part of a continuous process with physical and functional connections between the transcriptional and processing machineries.
Capping, RNA splicing, 3’ end formation and polyadenylation, and nuclear export are coupled to transcriptional machinery
Regulation occurs at multiple levels in this continuous processin a coordinated fashion
Eucaryotic cells have elaborate mRNA surveillance systems to destroy any messages containing errors
Unified theory of gene expression

Traditional View of Gene Expression
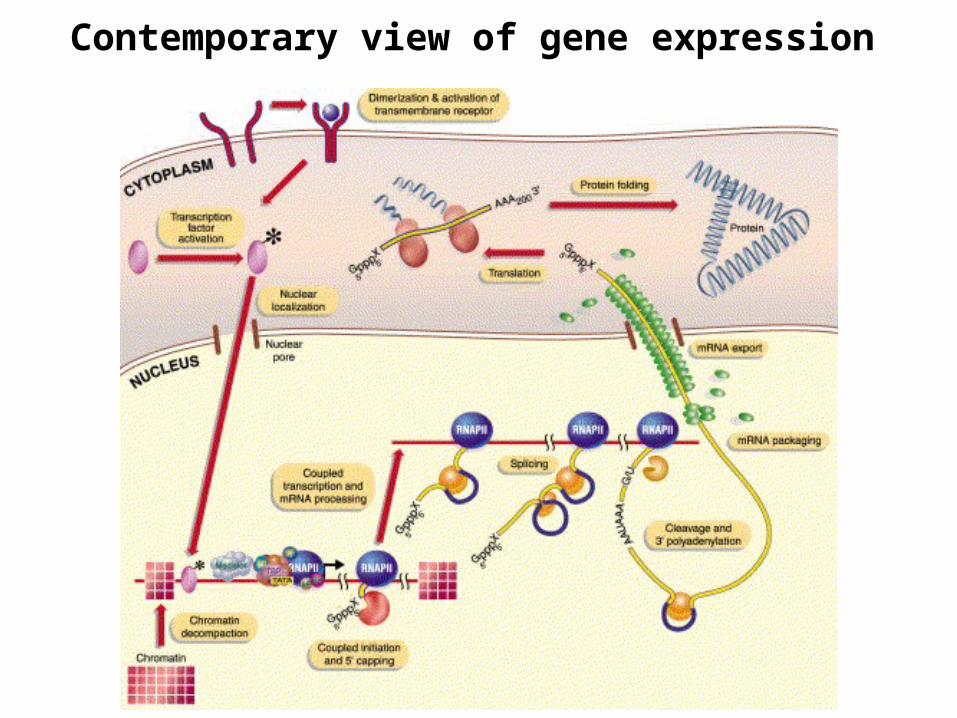
Contemporary view of gene expression


















