
Population Structure, Mobility and Habitat Preferences of the Violet Copper Lycaena Helle...
10
Population structure, mobility and habitat preferences of the violet copper Lycaena helle (Lepidoptera: Lycaenidae) in Western Germany: implications for conservation Klaus Fischer, Burkhard Beinlich and Harald Plachter* University of Marburg, Faculty of Biology, Institute for Nature Conservation, Lahnberge, D-35032 Marburg, Germany Received: 23 March 1998; accepted: 10 August 1998 We describe population structure, mobility and habitat preferences of the butterfly Lycaena helle using a survey of the popula- tions in the Westerwald area (Western Germany) and mark-recapture techniques. Three out of 83 recorded populations were intensively studied in 1995. In all, 1596 individuals (537 females) were uniquely marked. The average adult residence time was 8 days, the maximum 34 days. L. helle is extremely sedentary. Average distances between first and subsequent captures were 37 and 61 meters for males and females respectively. Population-specific differences are interpreted as being caused by the influence of barriers and corridors. The potential for colonization is regarded as poor, and isolated populations seem to be under a higher risk of extinction than other ones. Preferred habitats were abandoned moist meadows with rich aspects of Polygonum bistorta on sheltered, warm, and humid stands. Owing to the destruction of natural habitats (moorland) L. helle now colonizes almost exclusively anthropogenic ephemeral habitats (abandoned moist meadows) in Central Europe. This results in serious conservation problems, as the biology of the species does not seem to be adapted to a high turn-over rate of habitats. Keywords: butterfly; metapopulation; dispersal; sedentariness; habitat turn-over Introduction Recent studies on butterfly ecology have contributed considerable additional information in the fields of population structure, dispersal, and conservation (e.g. Thomas and Harrison, 1992; Hanski et al., 1995; Pullin, 1995; Lewis et al., 1997). In particular, metapopulation structures and the types and amount of dispersal seem to be major factors in the long-term survival of popula- tions. As declining patch occupancy and an increasing threat to survival is caused by progressive habitat frag- mentation (e.g. Hanski et al. 1994, 1995; Hill et al., 1996), the persistence of a species often relies on networks of suitable habitats, sufficiently close to allow dispersal between them (Thomas et al., 1992; Dennis and Eales, 1997). However, knowledge about the role of habitat corridors in facilitating dispersal and hence maintain- ing viable populations as well as the knowledge about the distances allowing colonization of empty habitat patches is still fairly poor. Both aspects are of impor- tance to species survival in fragmented landscapes as is the case in Central Europe (e.g. Settele et al., 1996). In contrast to previous studies we chose a butterfly species, the Violet Copper, Lycaena helle Denis and Schifferm ¨ uller, 1775, which in Central Europe has prob- ably already been living in more or less fragmented habitat islands for several thousands of years. Being a postglacial relict, the species has always been restricted to a few localities only (Bernardi and De Lesse, 1951; Meyer, 1982 a,b). The strong spatial restriction led to the assumption that dispersal ability might be decid- edly poor. The hygrophilous butterfly colonizes moor- land and, if lacking such natural habitats, prefers abandoned moist meadows (R´ eal, 1962 b; Meineke, 1982). An important reason for the decline of this spe- cies seems to be the loss of habitats due to the deteri- oration of wetlands (e.g. Hasselbach, 1985; Meyer, 1980). Nowadays, L. helle is one of the rarest butterfly species in Central Europe and is mentioned in the ‘Red Data Lists’ of Germany (Pretscher, 1984; ‘endangered’) and Europe (Munguira et al., 1993; ‘vulnerable’). Unfor- tunately, precise information on the butterfly’s ecology which would be crucial for conservation management is still lacking. Taking into account the above men- tioned deficiencies and the specific situation in Central Europe, we focus on the following questions essential * To whom correspondence should be addressed. Journal of Insect Conservation, 3 , 43–52 (1999) 1366–638X © 1999 Kluwer Academic Publishers
-
Upload
klaus-fischer -
Category
Documents
-
view
222 -
download
4
Transcript of Population Structure, Mobility and Habitat Preferences of the Violet Copper Lycaena Helle...

Population structure, mobility and habitat preferences of the violetcopper Lycaena helle (Lepidoptera: Lycaenidae) in WesternGermany: implications for conservation
Klaus Fischer, Burkhard Beinlich and Harald Plachter*
University of Marburg, Faculty of Biology, Institute for Nature Conservation, Lahnberge, D-35032Marburg, Germany
Received: 23 March 1998; accepted: 10 August 1998
We describe population structure, mobility and habitat preferences of the butterfly Lycaena helle using a survey of the popula-tions in the Westerwald area (Western Germany) and mark-recapture techniques. Three out of 83 recorded populations wereintensively studied in 1995. In all, 1596 individuals (537 females) were uniquely marked. The average adult residence time was8 days, the maximum 34 days. L. helle is extremely sedentary. Average distances between first and subsequent captures were 37and 61 meters for males and females respectively. Population-specific differences are interpreted as being caused by theinfluence of barriers and corridors. The potential for colonization is regarded as poor, and isolated populations seem to beunder a higher risk of extinction than other ones. Preferred habitats were abandoned moist meadows with rich aspects ofPolygonum bistorta on sheltered, warm, and humid stands. Owing to the destruction of natural habitats (moorland) L. helle nowcolonizes almost exclusively anthropogenic ephemeral habitats (abandoned moist meadows) in Central Europe. This results inserious conservation problems, as the biology of the species does not seem to be adapted to a high turn-over rate ofhabitats.
Keywords: butterfly; metapopulation; dispersal; sedentariness; habitat turn-over
J o u r n a l o f I n s e c t C o n s e r v a t i o n , 3 , 4 3 – 5 2 ( 1 9 9 9 )
Introduction
Recent studies on butterfly ecology have contributedconsiderable additional information in the fields ofpopulation structure, dispersal, and conservation (e.g.Thomas and Harrison, 1992; Hanski et al., 1995; Pullin,1995; Lewis et al., 1997). In particular, metapopulationstructures and the types and amount of dispersal seemto be major factors in the long-term survival of popula-tions. As declining patch occupancy and an increasingthreat to survival is caused by progressive habitat frag-mentation (e.g. Hanski et al. 1994, 1995; Hill et al., 1996),the persistence of a species often relies on networks ofsuitable habitats, sufficiently close to allow dispersalbetween them (Thomas et al., 1992; Dennis and Eales,1997). However, knowledge about the role of habitatcorridors in facilitating dispersal and hence maintain-ing viable populations as well as the knowledge aboutthe distances allowing colonization of empty habitatpatches is still fairly poor. Both aspects are of impor-tance to species survival in fragmented landscapes as isthe case in Central Europe (e.g. Settele et al., 1996).
In contrast to previous studies we chose a butterfly
* To whom correspondence should be addressed.
1366–638X © 1999 Kluwer Academic Publishers
species, the Violet Copper, Lycaena helle Denis andSchiffermuller, 1775, which in Central Europe has prob-ably already been living in more or less fragmentedhabitat islands for several thousands of years. Being apostglacial relict, the species has always been restrictedto a few localities only (Bernardi and De Lesse, 1951;Meyer, 1982 a,b). The strong spatial restriction led tothe assumption that dispersal ability might be decid-edly poor. The hygrophilous butterfly colonizes moor-land and, if lacking such natural habitats, prefersabandoned moist meadows (Real, 1962 b; Meineke,1982). An important reason for the decline of this spe-cies seems to be the loss of habitats due to the deteri-oration of wetlands (e.g. Hasselbach, 1985; Meyer,1980). Nowadays, L. helle is one of the rarest butterflyspecies in Central Europe and is mentioned in the ‘RedData Lists’ of Germany (Pretscher, 1984; ‘endangered’)and Europe (Munguira et al., 1993; ‘vulnerable’). Unfor-tunately, precise information on the butterfly’s ecologywhich would be crucial for conservation managementis still lacking. Taking into account the above men-tioned deficiencies and the specific situation in CentralEurope, we focus on the following questions essential

K . F i s c h e r e t a l .
for the conservation of this organism: 1. Is L. helle infact a sedentary species and over what distance cancolonization occur regularly? 2. Do barriers and corri-dors influence dispersal? 3. What essential resourcescomprise a habitat for L. helle? 4. What appropriateland use could maintain these habitats?
Study sites and methods
The study was carried out in the Westerwald area (Rhi-neland-Palatinate, Western Germany) at elevationsfrom 500 to 600 meters (Fig. 1). The climate is sub-oce-anic, moderately cold, and humid (precipitation .
1000 mm/year, average temperature/year 1 6 °C;Sabel and Fischer, 1992). In May and June 1994 popula-tions of L. helle in the Westerwald area were system-atically mapped. Therefore, we searched for adultbutterflies in all suitable habitat patches in an area ofabout 250 km2, covering the whole (known) distribu-tion of the species in the Westerwald area. As the malesprefer sheltered locations in the vicinity of the larvalhostplant (Polygonum bistorta), they are usually easy tofind (cf. Thomas and Harrison, 1992). Each patch wasvisited at least twice during favourable weather condi-tions. In no case did the second search reveal L. hellewhere they had not been found on the first search. Foreach occupied patch, we determined patch area (range0.2–20 ha), distance to the nearest populated patch(between patch edges; range 200–2100 m), vegetationtype and current land use.
Out of 83 sites supporting populations of this butter-
Figure 1. Location of the survey area and the three study sites for mGermany.
4
fly species, three with sufficient population size anddifferent habitat configurations were chosen fordetailed studies in 1995 using mark-release-recapturetechniques (Fig. 2). The patchy habitat distributionwithin these study sites was suitable for studying theexchange of individuals between sub-populationsunder various conditions: fallow wetland with someintervening spruce forests (site B), light bog-woodlandwith clearings (site L), and afforestated wetland, whereL. helle is present in fens and along some ditches (siteH). The populations were sampled on every favourableday between 1st May and 10th July 1995 (except at siteH with a lower number of sampling days). When cap-tured for the first time the butterflies were individuallymarked (permanent-ink, felt-tip pen) and released atthe point of capture, after keeping them for a short timein a cool-box to avoid a ‘mark-release trauma’ (cf. Wattet al., 1977). For each capture study site, the locationwithin a grid system (each cell 20 3 20 m), date, num-ber of mark, sex, distance to the nearest shrubs/trees,percentage of ground covered by P. bistorta, behaviour,and – if required – nectar source plants were recorded.Mobility was estimated by the straight distancesbetween the places of capture and (next) recapture.This, of course, does not reflect the real flight pattern ofthe butterflies but it roughly indicates dispersal abilityand site fidelity. Those estimations as well as the fol-lowing one can only give minimum values. The indi-vidual home ranges were estimated by the minimumarea method (Southwood, 1976), using data gathered
ark-recapture studies in the north-eastern Rhineland-Palatinate region,
4

T h e C o p p e r B u t t e r f l y L y c a e n a h e l l e i n W e s t e r n G e r m a n y
from individuals observed at least four times. Fordescription of habitats, 15 plant sociological inventorieswere carried out according to the method of Braun-Blanquet regarding the plant species and their abun-dance (cf. Muhlenberg, 1989). The phytocoenosisobtained from the plant sociological inventories wasanalysed using ‘indicator values’ of vascular plants forecological parameters (Ellenberg et al., 1992).
Results
Capture results
In total, 1596 individuals of L. helle were marked dur-ing the mark-recapture studies, resulting in 866 recap-tures (total observations 2462). The overall recapturerate (in % of marked individuals) was 30.0%, showingconsiderable differences between the three study sites(Table 1). The average residence time was 7.7 6 6.8days (median: 6, n 5 471). This did not vary accordingto sex (males 7.7; females 7.8). The maximum residencetimes were 34 days for males and 33 days forfemales.
Figure 2. Map of the distribution of Lycaena helle in the Westerwaldextended habitat patches the points give only the patch centres. The tH refer to the three study sites for detailed population studies in 19generalized to avoid the exact localisation and exploitation of popula
4
Mobility and isolation of habitat patches
The majority of individuals were recaptured in the nearvicinity of previous captures. 63.5% of the linear dis-tances were less than 40 meters, 89.0% less than 100meters. Males were found to be significantly more sta-tionary than females (average 37.2 6 60.7 m comparedto 60.8 6 67.0 m; Mann & Whitney U-test, p , 0.0001,U 5 38163.5, n1 5 682, n2 5 165). Spearman’s rankcorrelation indicated a significant, but weak relation-ship between the distance covered and the time periodsbetween capture and recapture (r 5 0.17; p , 0.0001,n 5 824), thus ruling out serious method-relatederrors. The flight distances recorded for males weresignificantly different between the three study sites(Kruskal-Wallis-test, p , 0.0001, H 5 24.2, df 5 2, n5 682), whereas for females these site differences werenot significant (Kruskal-Wallis-test, p 5 0.073, H 5
5.2, df 5 2, n 5 165; tab. 2). If, in the case of multiplerecaptures, the respective distances covered by eachindividual are totalled, the average distances increaseto 67.5 6 86.1 m (males) and 97.6 6 107.6 m (females)
area in 1994. A survey identified 83 populations (black points). Forriangles mark the position of extinct populations, the letters L, B and95. Unoccupied patches have not been recorded. The plot is heavilytions of the protected species.
5
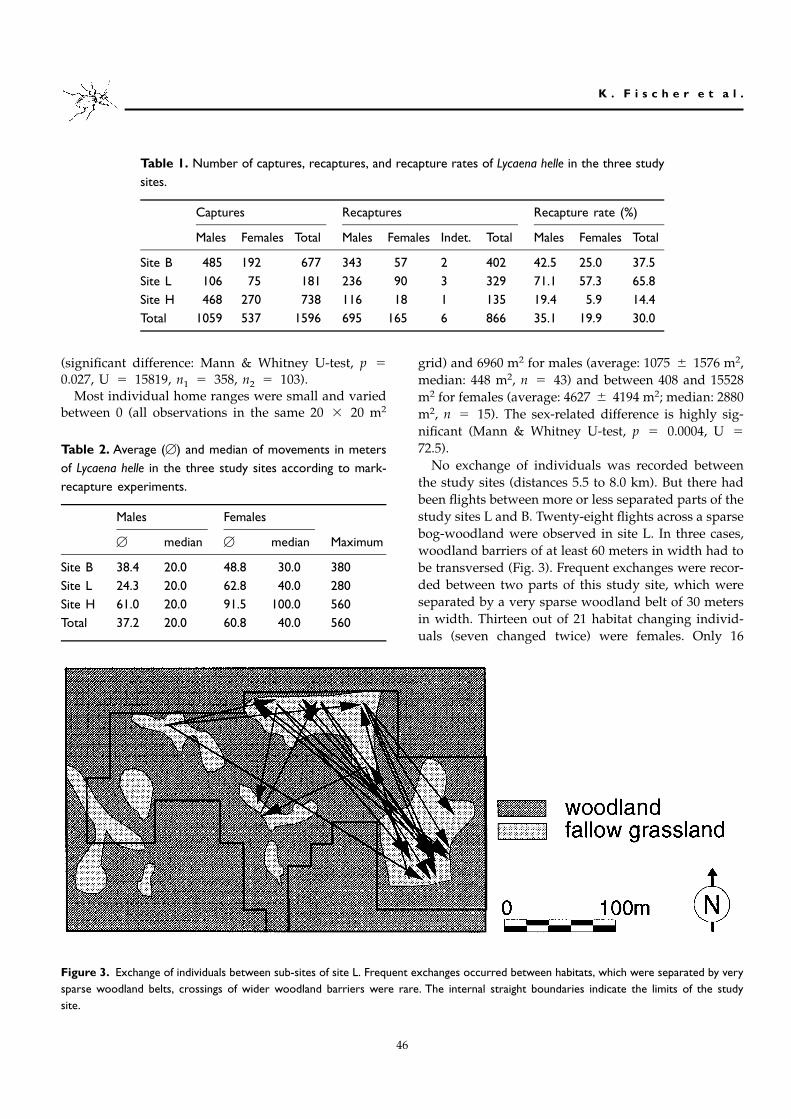
Table 1. Number of captures, recaptures, and recapture rates of Lycaena helle in the three studysites.
Captures Recaptures Recapture rate (%)
Males Females Total Males Females Indet. Total Males Females Total
Site B 485 192 677 343 57 2 402 42.5 25.0 37.5Site L 106 75 181 236 90 3 329 71.1 57.3 65.8Site H 468 270 738 116 18 1 135 19.4 5.9 14.4Total 1059 537 1596 695 165 6 866 35.1 19.9 30.0
K . F i s c h e r e t a l .
(significant difference: Mann & Whitney U-test, p 5
0.027, U 5 15819, n1 5 358, n2 5 103).Most individual home ranges were small and varied
between 0 (all observations in the same 20 3 20 m2
Table 2. Average ([) and median of movements in metersof Lycaena helle in the three study sites according to mark-recapture experiments.
Males Females
[ median [ median Maximum
Site B 38.4 20.0 48.8 30.0 380Site L 24.3 20.0 62.8 40.0 280Site H 61.0 20.0 91.5 100.0 560Total 37.2 20.0 60.8 40.0 560
Figure 3. Exchange of individuals between sub-sites of site L. Frequensparse woodland belts, crossings of wider woodland barriers were rsite.
4
grid) and 6960 m2 for males (average: 1075 6 1576 m2,median: 448 m2, n 5 43) and between 408 and 15528m2 for females (average: 4627 6 4194 m2; median: 2880m2, n 5 15). The sex-related difference is highly sig-nificant (Mann & Whitney U-test, p 5 0.0004, U 5
72.5).No exchange of individuals was recorded between
the study sites (distances 5.5 to 8.0 km). But there hadbeen flights between more or less separated parts of thestudy sites L and B. Twenty-eight flights across a sparsebog-woodland were observed in site L. In three cases,woodland barriers of at least 60 meters in width had tobe transversed (Fig. 3). Frequent exchanges were recor-ded between two parts of this study site, which wereseparated by a very sparse woodland belt of 30 metersin width. Thirteen out of 21 habitat changing individ-uals (seven changed twice) were females. Only 16
t exchanges occurred between habitats, which were separated by veryare. The internal straight boundaries indicate the limits of the study
6

T h e C o p p e r B u t t e r f l y L y c a e n a h e l l e i n W e s t e r n G e r m a n y
exchanges between sub-sites were recorded for site B.Here, suitable habitats were separated by woodlandbarriers as well as distance (120 and 280 m; cf. fig. 4).The habitats were connected by a path through thespruce forests. Two direct observations showed thatthis path is used by butterflies to fly from one habitat tothe other. Two other butterflies were caught by a gustof wind and crossed the forest this way. Twelve out of16 habitat changing individuals in site B were males.Numerous direct observations proved that L. hellemoves by using linear structures such as forest edges.In contrast, the species was never seen flying over openfields.
Based on the survey in 1994, isolation by distancebetween occupied habitat patches was analysed. Theaverage distance between occupied habitats next toeach other was 598 6 405 m (median 5 400 m, n 5
83). Forty-five (54.2%) patches were not more than 500m, 73 (88.0%) not further than 1000 m away from thenext population. Twelve formerly known populations(own unpubl. data) were found to be extinct. The aver-age distance of those to the next still existing popula-tion (1533 6 1087 m; median 1250) was significantlyhigher compared to the distances between occupiedpatches (Mann & Whitney U-test, p 5 0.0002, U 5
Figure 4. Exchange of individuals between sub-sites of site B. Habitatsinternal straight boundaries indicate the limits of the study site.
4
161.5, n1 5 83, n2 5 12). The habitat patch sizes of the83 recorded populations were roughly estimated lead-ing to an average size of 1.98 6 2.9 ha (median 5 1ha). Thirty-four (41.0%) patches were smaller than 1 ha,with the smallest occupied patch being only 0.2 ha inextent (1–2 ha: 33 patches; 3–10 ha: 13; . 10 ha: 3). Thesize of the largest patch was about 20 ha.
Behavioural aspects
According to the behaviour at the moment of encoun-ter, males ‘flew’ more frequently than females (51.1% to37.8%) and ‘fed’ (25.3% to 33.3%) and ‘rested’ (23.6% to28.9%) less frequently. These sex-specific differences inbehavioural patterns are highly significant (chi-square-test, p , 0.0001, x2 5 35.2, df 5 2, n 5 2450). Highnumbers of observations (up to 130) in some grids weredue to concentrations of males, taking up perches closeto each other. From these perches, they rapidly startedto chase any larger flying insect (such as bumble-bees,dragonflies, butterflies) that approached. In case ofnon-conspecific intruders the males immediatelyreturned to their perches. Conspecifics were pursued(usually in a typical co-rotating circle flight) until theyalighted or flew away. Afterwards, one butterfly
are separated not only by woodland barriers, but also by distance. The
7

K . F i s c h e r e t a l .
returned to the perch. Frequently males were capturedseveral times at exactly the same perch (up to tentimes). Males were never seen exhibiting patrollingbehaviour.
Habitat characteristics
In the study area moist meadows with abundant knot-grass (P. bistorta) were clearly preferred. The colonizedmeadows in particular belong to the Deschampsia cespi-tosa-Polygonum bistorta-association, often includingplant species indicative of nutrient-poor grassland (e.g.Potentilla erecta, Hypericum maculatum) and of fenland(e.g. Carex fusca, Comarum palustre, Eriophorum angusti-folium, Viola palustre). This plant community must beregarded as a fallow stage of unimproved moist grass-land (Sabel and Fischer, 1992). Indicator values for eco-logical parameters (Ellenberg et al., 1992) characterizethe habitats of L. helle as humid, moderately acidic, andfairly nutrient-poor (average indicator values of the 15plant sociological inventories: humidity 7.1, soil reac-tion 4.9, nutrient ratio 4.5). Those findings are suppor-ted by the results obtained from the survey of L. hellepopulations in the Westerwald area. The Deschampsiacespitosa-Polygonum bistorta-association occurred at 78out of 83 (94.0%) sites supporting populations, and at43 (51.8%) sites it was the only colonized plant commu-nity within the habitat patch. Only in five habitats,which consisted of low and transitional moor, this spe-cific association was not found at all. In the other casesa mixture of plant communities including theDeschampsia cespitosa-Polygonum bistorta-association,fenland (Caricetum fuscae, 20 sites), meadow-sweet fal-low land (Filipendulion ulmariae, 14 sites) and oligo-trophic grassland (Polygalo-Nardetum, 3) was found.As already indicated by the above mentioned plantcommunities, most of the habitats (79 out of 83; 95.2 %)underlie no current land use. Only four populationspersisted under extensive (2) or sporadic (2) grazingregimes. Within the moist meadows the butterflieswere restricted to the vicinity of woodland and shrubs.For all observations the average distance to the nearestshrubs/trees was only 3.6 6 2.7 meters (median: 3.0m, n 5 2461).
The occurrence of P. bistorta in the habitats is essen-tial, for egg-laying as well as feeding of larvae in Cen-tral Europe is restricted to this plant species (e.g.Weidemann, 1995). Females significantly prefer denserground coverage of this plant than males (25.4 6 16.2to 21.5 6 13.1%; Mann & Whitney U-test, p , 0.0001,U 5 540493.5, n1 5 702, n2 5 1753). Adults fed onflowers of 17 different plant species. Cardamine pratensis
4
and P. bistorta were most frequently visited (65.8% offeeding records). Other important nectare source plantswere Cardamine amara, Ranunculus repens and R. acris.
Discussion
Our data on mobility as well as the long resident timesstrongly suggest that L. helle can be an extremely seden-tary species. The maximum residence time of 34 days isone of the highest ever found in field studies for non-wintering butterflies in Europe. For British butterflies,the longest residence time, given by Shreeve (1992), is28 days (Maniola jurtina). The average residence timesof L. helle are also relatively long compared to field dataon other European Lycaenid species (cf. Kockelke et al.1994; Pauler et al. 1995; Vaisanen et al. 1994). The recor-ded recapture rates can be regarded as typical for sed-entary species (cf. Pauler et al., 1995; Scott and Opler,1975). The considerable differences between the studysites (14.4 to 65.8%) confirm that recapture rates are notonly due to species-specific differences, but also to theconfiguration of the study sites, size and seclusion ofpopulations, as well as study intensity.
The behaviour patterns of males and females differdistinctly, which can be related to the mating system(e.g. Rutowski, 1991). The mate-location behaviour ofL. helle is obviously aggressive perching (c.f. Corderoand Soberon, 1990). The higher flight activity of malesis due to the resulting inspection flights, characteristicfor many Lycaenid species (e.g. Scott, 1974; Scott andOpler, 1975; Schurian and Fiedler, 1996). That femaleswere more frequently observed ‘resting’ in the vegeta-tion, does not contradict their higher mobility. Due tothe territorial behaviour of males, they usually returnto their perches after flights resulting in site fidelity(e.g. Schurian and Fiedler, 1996; Pullin, 1997). Females,however, disperse for longer (but still short) distancesin search for oviposition sites (cf. Elligsen et al., 1997;Shreeve, 1992). Additionally, this active search doesexplain the stronger association of females with P. bis-torta, the larval hostplant.
Activity data differ considerably between the threestudy sites (highly significant for males), which is mostlikely related to the habitat configuration (influence ofbarriers and corridors). Linear corridors are probablyused by L. helle to travel considerable distancesbetween adjacent habitats, as is likely for ditches in siteH and paths in site B. The fact that the more sedentarymales changed habitats more often in site B is probablydue to passive wind-assisted drift, whereas for thewind sheltered site L, active movements can be
8

T h e C o p p e r B u t t e r f l y L y c a e n a h e l l e i n W e s t e r n G e r m a n y
assumed. Owing to frequent inspection flights (up to100 in two hours; Scott, 1974), males should be morefrequently affected by gusts than females. But in gen-eral, the arrangement and the structure of barriersbetween occupied habitats seems to be most importantfor the connectivity of adjacent populations. As thelevel of activity of L. helle is highly correlated with thesolar radiation (Meyer and Helminger, 1994; Real,1962a), it is extremely unlikely that individuals willcross dense spruce forests. On the other hand openfields probably act as barriers due to the restriction towind-sheltered stands, at least for active dispersal.Thus, L. helle seems to be characterized by a rather poorindividual ability to move between neighbouring hab-itats, if barriers are present. Furthermore, our data(infrequent dispersal between adjacent populations, cf.fig. 4) as well as the occurrence in distinct habitatpatches suggest that L. helle possibly exists in a meta-population structure, which seems to be a major factorin the persistence of many butterfly populations (e.g.Brunzel and Reich, 1996; Hanski et al., 1994; Hanski etal., 1995; Thomas and Harrison, 1992; Lewis et al., 1997).Therefore, the basic question is at what scale (and dis-tances) colonization of vacant habitats does occur.
Regarding habitat requirements L. helle, like otherbutterfly species, is restricted to sheltered locations inthe vicinity of shrubs or trees (c.f. Dover et al., 1997;Elligsen et al., 1997). The dependence on shelteredstands is apparently so strong, that more open habitatscannot be colonized, even if all other requirements aremet (Real, 1962d). Together with published data (Han-nemann, 1928; Real, 1962 a-d; Weidemann, 1995), nec-tar feeding of adults is confirmed for 33 plant species todate. Thus, L. helle can be regarded as fairly opportu-nistic concerning the use of nectar source plants.According to our observations and the bibliographicalreferences, the basic habitat requirements of L. helle inCentral Europe can be described as follows: (1) moistmeadows with abundant P. bistorta (larval hostplant),in particular the Deschampsia cespitosa-Polygonum bis-torta-association, (2) sheltered, warm and humid standsin the vicinity of shrubs, forest edges etc., (3) nectarsource plants. Neither these findings nor the existenceeven in fairly small habitat patches explain the rarity ofthe species in Central Europe (e.g. Meyer, 1980), forthese requirements should be met in many Europeanlandscapes. Indeed, the butterfly does not occur at allplaces, where it could be expected (SBN, 1987). We con-clude that its sedentary behaviour and a resulting lowdispersal ability might be the decisive factor. Of course,one has to take into consideration that mark-recapture
49
methods tend to underestimate the real dispersal abil-ity due to methodological constraints (cf. Dennis andBardell, 1996; Dennis and Shreeve, 1997; Settele et al.,1996). Nevertheless, studies using large scale surveysshow that at least for small sedentary species andunder normal conditions (for the possible impact ofextreme weather events see Dennis and Bardell, 1996)distances of 1 km or more between suitable habitatscan act as major barriers to dispersal and lead to a fail-ure of colonization (Thomas and Harrison, 1992; Lewiset al., 1997), whereas for other species with stronglydeveloped dispersal behaviour barriers seem to be nonexistent (Brunzel and Reich, 1996; Brunzel, 1996).
Some further evidence was found to support thisidea. The existing spatially restricted colonies in Cen-tral Europe are still concentrated close to the edges ofthe last glaciation (Meyer, 1982a). Additionally, numer-ous described local forms (even though they are notvalid as subspecies in a taxonomic sense) indicate avery low level of long-distance dispersal and exchange(cf. Bernardi and De Lesse, 1951; Meyer, 1982 a,b).Under natural conditions the butterfly is strictly linkedto ecosystems which are relatively constant over longperiods of time, especially bogs and swamps (cf. Real,1962 a-d). To survive there, efficient colonization strate-gies are not needed. Moreover, such moorland biotopeshave always been rare and often isolated in the moun-tain regions of Central Europe, thus resulting in a lowprobability of reaching a suitable and free habitat.There should therefore be a strong selection againstdispersal. Thus, L. helle seems to be in need of continu-ity of habitats in space and time.
Implications for conservation
As a consequence of the destruction and deteriorationof nearly all natural moorland in Central Europe, L.helle currently colonizes almost exclusively anthropo-genic habitats, particularly abandoned moist meadows.This results in serious problems for the conservation ofthis species, as these habitats are ephemeral over timeand the biology of L. helle does not seem to be adaptedto a high turn-over rate of habitats. Clearly, this butter-fly is able to colonize (neighbouring) habitat patchesclose enough to be reached, as shown by our mark-recapture experiments and the occurrence in secondaryhabitats. However, colonization seems only to happenat a strongly restricted spatial scale. Thus, besides theexistence of suitable habitat patches (cf. Dennis andEales, 1997), the distances allowing colonization ofempty habitat patches seem to be most important. The

K . F i s c h e r e t a l .
maximum single step colonization distances of butter-flies is species-specific (Thomas et al., 1992). For L. hellea frequent and successful colonization within a fewyears seems probable only for distances of a few hun-dred meters. Our data on the occurrence of L. helle inthe Westerwald area show that most of the existingpopulations were not more than 500 m distant from thenext occupied habitat patch. Furthermore, isolatedpopulations generally seem to have a higher extinctionrisk than connected ones (cf. Hanski et al., 1995; Hill etal., 1996). Thus, L. helle is likely to be extremely suscep-tible to habitat change in isolated habitat patches. Thismight have led to the extinction of the butterfly in mostof the formerly occupied locations (cf. Hasselbach,1985; Meyer, 1980). Conservation measures for this spe-cies must not only take into account how to manage thehabitats to support large populations, but the spatialarrangement and connectivity of suitable habitats aswell (cf. Thomas et al., 1992).
Because of the instability of the secondary habitats,some kind of management is unavoidable. Otherwise P.bistorta as well as other substantial habitat elementswill vanish in the course of succession. The regenera-tion and re-establishment of moorland will take longperiods of time and is impossible in most of the regionswhere L. helle currently occurs in Central Europe. Thereis obviously no alternative (apart from introductions)to the protection of the existing colonies within theirsecondary habitats, at least none, which would meetthe requirements of this species in time. Because mostcolonies exist on recently abandoned moist meadows(e.g. Meineke, 1982), mowing seems to be most appro-priate. Taking into account the sudden habitat deteri-oration caused by every cut, rotational mowing ofsingle habitat patches in autumn in cycles of severalyears might be most favourable. The mowing shouldfollow the pupation and leave out areas with shrubs asan essential habitat requirement. However, such amowing system is highly artificial, expensive and faraway from the economic requirements of the localfarmers. In general, the protection of specific land usepatterns in cultural landscapes by management meas-ures without real economic returns is a doubtful con-servation strategy (Plachter, 1995, 1996). Grazing mightbe a better alternative. Indeed, in the course of map-ping, four colonies of L. helle were found in rough cattlepastures, making such attempts promising. This wouldpresumably require pasturing on a fairly large spatialscale with irregularly grazed patches. Such a large-scale pasturing by herdsmen supports other butterflyspecies, too (cf. Elligsen et al., 1997).
5
Acknowledgements
The authors wish to thank the Ministry of Environmentand Forestry of the State of Rhineland-Palatinate forsupporting the study by grants from the scholarship‘Arten- und Biotopschutz Rheinland-Pfalz’, theKoblenz and Siegen-Wittgenstein district governmentsfor granting permission to pursue this study, KonradFiedler (University of Bayreuth) and two unknown ref-erees for critical and helpful comments on the manu-script, Angelika Fischer-Munsch (Westerburg), PeterZofel (University of Marburg), and Jason Audsley(Marburg) for assistance in data analysis and the lin-guistic improvement of the manuscript, respectively.Peter Fasel, Biological Station Rothaargebirge, andErnst Brockmann, Lepidopterological Study Group ofHesse, contributed unpublished local data on thebutterfly.
References
Bernardi, G. and De Lesse, H. (1952) La variabilite geogra-phique de Lycaena helle Denis et Schiff. (Lep. Lycaenidae).Rev. Franc. Lepidopt. 13/14, 203–214.
Brunzel, S. (1996) Populationsokologische Untersuchung amSchwalbenschwanz (Papilio machaon L. 1758). Z. Okologieu. Naturschutz 5, 37–46.
Brunzel, S. and Reich, M. (1996) Zur Metapopulationsstruk-tur des Roten Scheckenfalters (Melitaea didyma Esper1779) auf der Schwabischen Alb. Z. Okologie u. Natur-schutz 5, 243–253.
Cordero, C.R. and Soberon, J. (1990) Non-resource basedterritoriality in males of the butterfly Xamia xami (Lep-idoptera: Lycaenidae). J. Ins. Behav. 3, 719–732.
Dennis, R.L.H. and Bardell, P. (1996) The impact of extremeweather events on local populations of Hipparchia semele(L.) (Nymphalidae) and Plebejus argus (L.) (Lycaenidae):hindsight, inference and lost opportunities. Entomologist’sGaz. 47, 211–225.
Dennis, R.L.H. and Eales, T. (1997) Patch occupancy in Coeno-nympha tullia (Muller, 1764) (Lepidoptera: Satyrinae): hab-itat quality matters as much as patch size and isolation. J.Insect Conserv. 1, 167–176.
Dennis, R.L.H. and Shreeve, T.G. (1997): Diversity of butter-fly species on British islands: ecological influences under-lying the roles of area, isolation and faunal source. Biol. J.Linn. Soc. 60, 257–275.
Dover, J.W., Sparks, T.H. and Greatorex-Davies, J.N. (1997)The importance of shelter for butterflies in open land-scapes. J. Insect Conserv. 1, 89–97.
Ellenberg, H., Weber, H.E., Dull, R., Wirth, V., Werner, W. and
0

T h e C o p p e r B u t t e r f l y L y c a e n a h e l l e i n W e s t e r n G e r m a n y
Paulissen, D. (1992) Zeigerwerte von Pflanzen in Mitte-leuropa, 2. edition. Scripta Geobotanica 18, 1–258.
Elligsen, H., Beinlich, B. and Plachter, H. (1997) Effects oflarge scale cattle grazing on populations of Coenonymphaglycerion and Lasiommata megera (Lepidoptera: Satyridae).J. Insect Conserv. 1, 13–23.
Hannemann, J. (1928) Etwas uber Chrysophanus amphidamasEsp. Int. Entomol. Zeitschrift 22, 209–210.
Hanski, I., Kuussaari, M. and Nieminen, M. (1994) Metapopu-lation and migration in the butterfly Melitaea cinxia. Ecol-ogy 75 (3), 747–762.
Hanski, I., Pakkala, T., Kuussaari, M. and Lei, G. (1995) Met-apopulation persistence of an endangered butterfly in afragmented landscape. Oikos 72, 21–28.
Hasselbach, W. (1985) Lycaena helle – die Zucht einer in derBundesrepublik Deutschland vom Aussterben bedrohtenArt (Lep.: Lycaenidae). Entomol. Zeitschrift 95, 70–76.
Hill, J.K., Thomas, C.D. and Lewis, O.T. (1996) Effects of hab-itat patch size and isolation on dispersal by Hesperiacomma butterflies: implications for metapopulation struc-ture. J. Anim. Ecol. 65, 725–735.
Kockelke, K., Hermann, G., Kaule, G., Verhaagh, M. and Set-tele, J. (1994) Zur Autokologie und Verbreitung des Kreu-zenzian-Ameisenblaulings, Maculinea rebeli. Carolinea 52,93–109.
Meineke, J.U. (1982) Einige Aspekte des Moor-Biotopschut-zes fur Großschmetterlinge am Beispiel moorbewohnen-der Großschmetterlingsarten in Sudwestdeutschland.Telma 12, 85–98.
Meyer, M. (1980) Die Verbreitung von Lycaena helle in derBundesrepublik Deutschland (Lep.: Lycaenidae). Entomol.Zeitschrift 90, 217–224.
Meyer, M. (1982a) Revision systematique, chorologique etecologique de Lycaena helle Denis & Schiffermuller, 1775(Lycaenidae). Linneana Belgica 8, 238–260, 345–358,451–466.
Meyer, M. (1982b) Les races europeennes de Lycaena helleDenis & Schiffermuller, 1775, et leurs biotopes (Lepidop-tera, Lycaenidae). Proc. 3rd Congr. Europ. Lepid., Cam-bridge 1982, 125–137.
Meyer, M. and Helminger, T. (1994) Untersuchungen zu einerPopulation von Lycaena helle arduinnae Meyer, 1980 imnordwestlichen Osling (Lepidoptera, Lycaenidae). Bull.Soc. Nat. Luxemb. 95, 315–326.
Muhlenberg, M. (1989) Freilandokologie. Heidelberg, Wiesba-den: Quelle & Meyer.
Munguira, M.L., Martin, J. and Balletto, E. (1993) Conserva-tion biology of Lycaenidae: A European overview. In Con-servation biology of Lycaenidae (butterflies) (T.R. New, ed.),pp. 23–34. Oxford: Information Press.
Lewis, O.T., Thomas, C.D., Hill, J.K., Brookes, M.I., Crane,T.P.R., Graneau, Y.A., Mallet, J.L.B. and Rose, O.C. (1997)
51
Three ways of assessing metapopulation structure in thebutterfly Plebejus argus. Ecol. Entomol. 22, 283–293.
Pauler, R., Kaule, G., Verhaagh, M. and Settele, J. (1995)Untersuchung zur Autokologie des SchwarzgeflecktenAmeisenblaulings, Maculinea arion (Linnaeus 1758) (Lep-idoptera: Lycaenidae), in Sudwestdeutschland. Nachr.entomol. Verein Apollo, N.F. 16, 147–186.
Plachter, H. (1995) Functional criteria for the assessment ofcultural landscapes. In Cultural landscapes of universal value(Von Droste, B., Plachter, H. and Rossler, M., eds.), pp.393–404. Jena: G. Fischer.
Plachter, H. (1996) Central European approach for the pro-tection of biodiversity. In Nature conservation outside pro-tected areas (D. Ogrin, ed.), pp. 91–108. Conf. Proc.Ministry of Environment and Physical Planning, Ljubl-jana.
Pretscher, P. (1984) Rote Liste der Großschmetterlinge. InRote Liste der gefahrdeten Tiere und Pflanzen in der Bundes-republik Deutschland (Blab, J., Nowak, E., Trautmann, W.and Sukopp, H., eds.), 4th edition. Greven: Kilda.
Pullin, A.S. (ed., 1995) Ecology and conservation of butterflies.London: Chapman & Hall.
Pullin, A.S. (1997) Habitat requirements of Lycaena disparbatavus and implications for re-establishment in England.J. Insect Conserv. 1, 177–185.
Real, P. (1962a) Les Pyrenees Orientales, refuge entomolo-gique. Ann. sci. Univ. Besancon, 2e ser. Zool. 17, 97–110.
Real, P. (1962b) Les stations de Lycaena helle D. et Schiff. dansle Doubs. Note de faunistique jurassienne no 1. Ann. sci.Univ. Besançon, 2e ser. Zool. 17, 111–118.
Real, P. (1962c) Quelques remarques sur Lycaena helle D. etSchiff. dans les Monts de la Madeleine. Ann. sci. Univ.Besançon, 2e ser. Zool. 17, 119–120.
Real, P. (1962d) Lycaena helle D. et Schiff. (ex Heodes amphi-damas Esp.) dans le Massif de Sancy. Ann. sci. Univ.Besançon, 2e ser. Zool. 17, 121–134.
Rutowski, R.L. (1991) The evolution of male mate-locationbehaviour in butterflies. Amer. Naturalist 138,1121–1139.
Sabel, K.-J. and Fischer, E. (1992) Boden- und vegetations-geographische Untersuchungen im Westerwald. Frank-furter geowiss. Arbeiten, D 7, 1–268.
SBN (Schweizerischer Bund fur Naturschutz, ed.) (1987) Tag-falter und ihre Lebensraume. Arten-Gefahrdung-Schutz. –2nd edition. Basel: SBN.
Schurian, K.G. and Fiedler, K. (1996) Adult behaviour andearly stages of Lycaena ochimus (Herrich-Schafer [1851])(Lepidoptera: Lycaenidae). Nachr. entomol. Verein Apollo,N.F. 16, 329–343.
Scott, J.A. (1974) Population biology and adult behaviour ofLycaena arota (Lycaenidae). J. Lepid. Society 28, 64–75.

K . F i s c h e r e t a l .
Scott, J.A. and Opler, P.A. (1975) Population biology andadult behavior of Lycaena xanthoides (Lycaenidae). J. Lepid.Society 29, 63–66.
Settele, J., Henle, K. and Bender, C. (1996) Metapopulationenund Biotopverbund: Theorie und Praxis am Beispiel vonSchmetterlingen und Reptilien. Z. Okologie u. Naturschutz:5, 187–206.
Settele, J., Margules, C., Poschlod, P. and Henle, K. (eds.,1996) Species dispersal in fragmented landscapes. Dor-drecht, Kluwer.
Shreeve, T.G. (1992) Adult behaviour. In The ecology of butter-flies in Britain (R.L.H. Dennis, ed.), pp. 22 ff. Oxford: Uni-versity Press.
Southwood, T.R.E. (1976) Ecological methods, 2nd edition.London, New York: Chapman and Hall.
Thomas, C.D. and Harrison, S. (1992) Spatial dynamics of a
52
patchily distributed butterfly species. J. Animal Ecol. 61,437–446.
Thomas, C.D., Thomas, J.A. and Warren, M.S. (1992) Dis-tributions of occupied and vacant butterfly habitats infragmented landscapes. Oecologia 92, 563–567.
Vaisanen, R., Kuussaari, M. Nieminen, M. and Somerma, P.(1994) Biology and conservation of Pseudophilotes batonin Finland (Lepidoptera, Lycaenidae). Ann. Zool. Fennici 31,145–156.
Watt, W.B., Chew, F.S., Snyder, C.R.G., Watt, A.G. and Roths-child, D.E. (1977) Population structure of pierid butter-flies I. Numbers and movements of some Colias species.Oecologia 27, 1–22.
Weidemann, H.J. (1995) Tagfalter beobachten, bestimmen, 2ndedition. Augsburg: Naturbuch.


















