
Population genetic structure of the red imported fire ant, … · Population genetic structure of...
12
Research article Population genetic structure of the red imported fire ant, Solenopsis invicta, in Taiwan C.C. Yang 1 , D. DeWayne Shoemaker 2 , W.J. Wu 1 and C.J. Shih 1, * 1 Department of Entomology, National Taiwan University, No. 1, Sec. 4, Roosevelt Road, Taipei 106, Taiwan, e-mail: [email protected], [email protected], [email protected] 2 USDA, ARS, Center for Medical, Agricultural & Veterinary Entomology, 1600/1700 SW 23rd Drive, Gainesville, FL, 32608, U.S.A., e-mail: [email protected] Received 18 June 2007; revised 11 October 2007; accepted 17 October 2007. Published Online First 10 November 2007 Abstract. We generated and analyzed microsatellite genotypic data and mtDNA sequence data from the fire ant Solenopsis invicta collected from two separate infested areas (Taoyuan and Chiayi) in Taiwan to infer the population and colony structure of these recently established populations. These genetic analyses revealed the following patterns: 1) Relatedness among worker nestmates was significantly greater than zero for both social forms from both populations; 2) No significant isolation by distance was found among nests within each social form from either population; 3) Significant mtDNA but no nuclear differentiation occurs between sympatric social forms in Taoyuan; 4) Molecular signa- tures of genetic bottlenecks associated with recent introductions are evident in both populations; and 5) The two sampled populations, Taoyuan and Chiayi, are highly genetically differentiated at both the nuclear or mtDNA genomes and most likely derive from two separate introductions into Taiwan. While results from these analyses generally were consistent with predictions based on the known biology of these ants and similar studies of S. invicta in the U.S.A. and South America, some patterns likely reflect the recent introduction and human-mediated inadvertent transport of ants in Taiwan. This is the first study to investigate the population and colony structure of fire ants in Taiwan and results from our study represent an important contribution to the ongoing efforts aimed at eradicating this invasive pest in Taiwan. Keywords: Solenopsis invicta, control strategy, fire ants, invasive species, population structure. Introduction Invasive species pose major threats to agricultural and natural environments and to the public health (Porter and Savignano, 1990; Williams and Porter, 1994; Lee, 2002; Sax et al., 2005). The negative impact of invasive species on agricultural systems is evident from the fact that a vast number of major pest insects in the U.S.A. are of exotic origin (Carruthers, 2003). Because many invasive species occupy human-disturbed habitats and are often found in areas of high commercial activity, the importance of invasive species is an issue of growing concern given increasing human activities, such as habitat destruction and international trade and transportation (Drake et al., 1989; Sax et al., 2005). Despite these negative impacts and increasing concerns, studies of invasive species offer unique opportunities to the study of evolutionary genetic processes as well as the genetic and evolutionary con- sequences of invasions (Ross et al., 1993; Tsutsui and Case, 2001; Giraud et al., 2002; Lee, 2002; JaquiȖry et al., 2005). For instance, because the history of many invasions are well documented and have occurred in recent times, inferences drawn from patterns of variation often are not confounded by historical processes. Further, the intro- duction of species into novel environments provides ideal scenarios for studying adaptation to novel environments, effects of bottlenecks on genetic diversity, and genetic changes occurring during invasion and spread (Sax et al. , 2005). Also, comparative studies across species or within * Author for correspondence, e-mail: [email protected] Insect. Soc. 55 (2008) 54 – 65 0020-1812/08/010054-12 DOI 10.1007/s00040-007-0969-y # BirkhȨuser Verlag, Basel, 2007 Insectes Sociaux
Transcript of Population genetic structure of the red imported fire ant, … · Population genetic structure of...

Research article
Population genetic structure of the red imported fire ant Solenopsis invictain Taiwan
CC Yang1 D DeWayne Shoemaker2 WJ Wu1 and CJ Shih1
1 Department of Entomology National Taiwan University No 1 Sec 4 Roosevelt Road Taipei 106 Taiwan e-mail d95632004ntuedutwwuwjntuedutw shihcjntuedutw
2 USDA ARS Center for Medical Agricultural amp Veterinary Entomology 16001700 SW 23rd Drive Gainesville FL 32608 USA e-maildewayneshoemakerarsusdagov
Received 18 June 2007 revised 11 October 2007 accepted 17 October 2007Published Online First 10 November 2007
Abstract We generated and analyzed microsatellitegenotypic data and mtDNA sequence data from the fireant Solenopsis invicta collected from two separateinfested areas (Taoyuan and Chiayi) in Taiwan to inferthe population and colony structure of these recentlyestablished populations These genetic analyses revealedthe following patterns 1) Relatedness among workernestmates was significantly greater than zero for bothsocial forms from both populations 2) No significantisolation by distance was found among nests within eachsocial form from either population 3) SignificantmtDNA but no nuclear differentiation occurs betweensympatric social forms in Taoyuan 4) Molecular signa-tures of genetic bottlenecks associated with recentintroductions are evident in both populations and 5)The two sampled populations Taoyuan and Chiayi arehighly genetically differentiated at both the nuclear ormtDNA genomes and most likely derive from twoseparate introductions into Taiwan While results fromthese analyses generally were consistent with predictionsbased on the known biology of these ants and similarstudies of S invicta in the USA and South Americasome patterns likely reflect the recent introduction andhuman-mediated inadvertent transport of ants in TaiwanThis is the first study to investigate the population andcolony structure of fire ants in Taiwan and results fromour study represent an important contribution to theongoing efforts aimed at eradicating this invasive pest inTaiwan
Keywords Solenopsis invicta control strategy fire antsinvasive species population structure
Introduction
Invasive species pose major threats to agricultural andnatural environments and to the public health (Porter andSavignano 1990 Williams and Porter 1994 Lee 2002Sax et al 2005) The negative impact of invasive specieson agricultural systems is evident from the fact that a vastnumber of major pest insects in the USA are of exoticorigin (Carruthers 2003) Because many invasive speciesoccupy human-disturbed habitats and are often found inareas of high commercial activity the importance ofinvasive species is an issue of growing concern givenincreasing human activities such as habitat destructionand international trade and transportation (Drake et al 1989 Sax et al 2005) Despite these negative impactsand increasing concerns studies of invasive species offerunique opportunities to the study of evolutionary geneticprocesses as well as the genetic and evolutionary con-sequences of invasions (Ross et al 1993 Tsutsui andCase 2001 Giraud et al 2002 Lee 2002 Jaquiry et al 2005) For instance because the history of many invasionsare well documented and have occurred in recent timesinferences drawn from patterns of variation often are notconfounded by historical processes Further the intro-duction of species into novel environments provides idealscenarios for studying adaptation to novel environmentseffects of bottlenecks on genetic diversity and geneticchanges occurring during invasion and spread (Sax et al 2005) Also comparative studies across species or within Author for correspondence e-mail shihcjntuedutw
Insect Soc 55 (2008) 54 ndash 650020-181208010054-12DOI 101007s00040-007-0969-y Birkhuser Verlag Basel 2007
Insectes Sociaux
species that have been introduced to more than onelocation can increase our understanding of what charac-teristics of these species are important for invasionsuccess (Lee 2002 May et al 2006)
Many social insects particularly ants are well knownas being important invasive pest species Indeed theinvasive potential and abilities of ants to colonize almostevery available habitat is such that they often are referredto as tramp species (Hccedillldobler and Wilson 1990Vander Meer et al 1990) One of the most notoriousinvasive ant species is the red imported fire ant (RIFA)Solenopsis invicta Buren (Lofgren 1986 Jouvenaz 1990Patterson 1994 Orr 1996) This exotic pest was inad-vertently introduced into the USA from South Americaabout 75 years ago Since that time it has spreadthroughout the southern states in the USA S invicta isof profound economic importance in the USA because1) it is an aggressive stinging insect causing massenvenomation incidents and hypersensitivity reactionsin humans 2) it occurs primarily in human-modifiedhabitats 3) it constructs large mounds that are unsightlyand capable of damaging farm machinery 4) it feeds onseveral important cultivated plants and tends homopter-ans that are also plant pests and 5) it negatively affectspopulations of native ants and other ground-dwellinganimals (Lofgren et al 1975 Lofgren 1986 Porter andSavignano 1990 Allen et al 1994 1998 2000 2001Vinson 1994 1997 Giuliano et al 1996 Carroll andHoffman 2000 Gotelli and Arnett 2000 Kaspari 2000Kemp et al 2000 Kopachena et al 2000 Eubanks2001 Forys et al 2001 Wojcik et al 2001 Morrison2002) These pest attributes of S invicta presumably areattributable largely to the fact that the introducedpopulations have almost none of the competitors andnatural enemies that normally act to suppress theirpopulations with the effect that population densities inthe USA are orders of magnitude greater than in thenative South American range (Porter et al 1992 1997Morrison 2000)
More recently S invicta has been inadvertentlyintroduced to other regions of the world including severalwestern states (eg New Mexico California) the Carri-bean Australia mainland China and Taiwan (MacKayand Fagerlund 1997 Buckley 1999 Davis et al 2001Nattrass and Vanderwoude 2001 McCubbin and Weiner2002 Huang et al 2004 Chen et al 2006) In all of theseareas S invicta is considered a significant pest such thatits negative agricultural ecological and public healthimpacts can only be expected to intensify in the nearfuture
While an extensive body of literature exists on studiesof S invicta from native South American and introducedpopulations in the USA few studies have been con-ducted on S invicta in these more recently invaded areas(Henshaw et al 2005 Chen et al 2006) Such studies areof interest not only for development of effective inte-grated management strategies for control and eradicationof fire ants in these areas but also provide the opportunity
for comparative studies of the genetic changes associatedwith or occurring during separate invasions (Ross et al 1993 Tsutsui and Case 2001 Giraud et al 2002Jaquiry et al 2005) To that end we present here theresults of a study exploring patterns of nuclear (micro-satellite) and of mtDNA variation in the fire ant S invictafrom two separate infested areas in Taiwan (Taoyuan andChiayi counties Fig 1) While the occurrence of S invictain Taiwan was first reported in 2003 analyses of the sizeand distribution of colonies suggest that this species likelyarrived some 3 ndash 5 years earlier (Huang et al 2004 Chenet al 2006)
Previous studies also have shown that both knownsocial forms of S invicta are present in both of theinfested areas in Taiwan (Huang et al 2004 Chen et al 2006) Two distinct types of colony social organizationoccur within this species (Glancey et al 1973) Coloniesof the monogyne (M) form possess a single egg-layingqueen whereas colonies of the polygyne (P) formpossess several to hundreds of these queens Associationstudies have shown that social form in this and other fireant species depends on the presence within a colony ofspecific coding region variants of the single gene Gp-9(reviewed in Gotzek and Ross 2007) Two classes ofvariants designated as B-like and b-like alleles occur inS invicta Monogyne colonies contain only the B-likeallele whereas polygyne colonies invariably contain theb-like allele as well as the B-like allele (Ross 1997 Rossand Keller 1998 Krieger and Ross 2002) The twosocial forms of S invicta differ not only in colony queennumber and Gp-9 genotypes but also in importantfeatures of their reproductive and dispersal behaviorswhich are expected to have a number of importanteffects on the distribution of genetic variation at variousspatial scales as well as management strategies em-ployed for eradication Thus knowledge of distributionof the two social forms is critical to constructing aneffective predictive model of their spread and expansionand assessing the potential of successful eradication of Sinvicta (Drees and Vinson 1990) Finally previousstudies have shown that the unique social and breedingbiology of each social form can act as strong constraintson particular routes of interform gene flow (Shoemakerand Ross 1996) Specifically the selective elimination ofqueens of the alternate social form by workers (Kellerand Ross 1998 Vander Meer and Porter 2001 Kriegerand Ross 2002 Ross and Keller 2002) and the highproportion of sterile diploid males in P nests inintroduced areas (Ross and Fletcher 1985a 1986Krieger et al 1999) severely restrict female-mediatedgene flow and limit male-mediated interform gene flowmostly to instances involving M males mating with Pfemales (Ross and Shoemaker 1993 1997 Shoemakerand Ross 1996 Shoemaker et al 2006) Therefore thepresence of both social forms in a novel environmentpresents the opportunity to test the generality of thesepatterns observed in earlier studies in the introducedrange in the USA
Insect Soc Vol 55 2008 Research article 55
This study has several objectives First we wished tolearn whether the current distribution of fire ants in thesetwo areas is the result of a single or separate introductionsinto Taiwan The finding of multiple introductions mayindicate that the island of Taiwan is at high risk ofexperiencing additional introductions Second we aimedto infer whether one social form was possibly derivedfrom local populations of the alternate social form as wellas whether substantial gene flow occurs between sym-patric populations of each social form Lastly we utilizedour genetic data to assess whether a genetic signaturecharacteristic of a recent population bottleneck is evidentin either of two areas infested by S invicta While dataexamining the alternative colony genetic structure andassociated genetic differences between native and intro-duced fire ant populations have been proposed for theperiod of several decades after their first introduction tothe USA (Ross and Fletcher 1985b Ross et al 1996Ross et al 1999) the more recent introduction of fireants into Taiwan provides a novel opportunity to study thegenetic changes characteristics of invasiveness of thisspecies over an extremely short time period particularlyin the initial 5 ndash10 years after the original introduction
Materials and methods
Source of ant samples
Ants representing both social forms were collected from two locationsTaoyuan county and Chiayi county in Taiwan (these two locations areseparated by ca 250 km) Initially the social form of S invicta wasdetermined by visual inspection of the average size of the workers theheight of the mounds and the number of queens (Greenberg et al1985) Based on the preliminary determinations of social form in thefield adult workers were collected from nests of each putative socialform separated by distances of 30 ndash5000 (polygyne) and 600ndash2000 m(monogyne) (Fig 1) In total we sampled 30 colonies from Taoyuancounty and 25 colonies from the Chiayi county Collected workers werestored in 95 alcohol and frozen at -20 8C
DNA extractions and determination of social form
Two separate sets of total genomic DNA were extracted fromindividuals sampled from each nest For the first set 10ndash15 workerswere pooled and extracted in bulk using the Viogene DNA extractionkit (Viogene Inc Taipei Taiwan) These bulk-extracted samples wereused in Gp-9 assays for social form determination (Valles and Porter2003) For the second set total genomic DNA was extracted from 10individual workers from each sampled nest These samples were used assource DNA for microsatellite genotyping and mtDNA sequencing(single individual per nest see below) Inspection of the distribution ofmultilocus genotypes from 10 individuals per sampled nest provides apowerful and direct approach for determining the social form of eachnest
Microsatellite analyses
We genotyped 10 workers from each colony representing 550individuals in total at six dinucleotide-repeat microsatellite loci(Table 1) previously developed for S invicta (Table 1 Krieger andKeller 1997) Amplification of the single locus Sol-20 was performed in
10-ml reactions with 30 ndash100 ng DNA 1 ml of 10X Super-Therm GoldPCR buffer 1 ml of 25 mM dNTPs 03 ml of a 10 mM FAM-labeledforward primer 03 ml of a 10 mM unlabeled reverse primer and 015 Uof Super-Therm Gold Hot-start Taq DNA polymerase (AppliedBiosystems) The remaining five loci were amplified in two separatemultiplex PCR reactions In the first reaction Sol-11 Sol-42 and Sol-49were amplified simultaneously in a single tube Reaction cocktails andconditions for Sol-11 Sol-42 and Sol-49 were identical to thosedescribed earlier (Ross et al 2003) except forward primers werelabeled with FAM In the second multiplex reaction Sol-6 and Sol-55were co-amplified in a single 20-ml reaction with 30ndash100 ng DNA 24 mlof 10X Super-Therm Gold PCR buffer 16 ml of 25 mM dNTPs 05 mlof a 10 mM FAM-labeled Sol-6 forward primer 05 ml of a 10 mMunlabeled Sol-6 reverse primer 06 ml of a 10 mM FAM-labeled Sol-55forward primer 06 ml of a 10 mM unlabeled Sol-55 reverse primer and1U of Super-Therm Gold Hot-start Taq DNA polymerase PCRamplifications were carried out on an ABI 9700 thermal cycler(Applied Biosystems) using the following profile initial denaturationat 95 8C for 10 min 30 cycles of 94 8C for 1 min 58 8C for 1 min and72 8C for 1 min a final extension of 72 8C for 30 min All PCR productswere visualized on an ABI 3100 genetic analyzer using laser detectionand scoring of genotypes was performed using the software GENES-CAN 312 (Applied Biosystems)
MtDNA sequencing
A 920-bp fragment of the mitochondrial genome that includes portionsof the cytochrome oxidase subunit I (COI) and subunit II (COII) geneswas amplified from a single individual per nest using the primers C1-J-2195 (Simon et al 1994) and DDS-COII-4 (Ross and Shoemaker1997) PCR reactions were carried out using identical reaction cocktailsand thermal cycling profiles described in Ahrens et al (2005) MtDNAamplicons were purified and used directly in standard fluorescent cycle-sequencing PCR reactions (ABI Prism Big Dye terminator chemistryApplied Biosystems) Sequencing reactions were subsequently purifiedand run on an ABI 3730 sequencer All mtDNA amplicons weresequenced in both directions and corresponding sequence pairs wereedited and combined into a single consensus sequence file Allsequences were then aligned by eye using previously published Sinvicta mtDNA sequence obtained from GenBank (AY950762)
Genetic analyses
The number of alleles allele frequencies observed heterozygosity andexpected heterozygosity at each microsatellite locus for both popula-tions were calculated using the program MSA 300 (Dieringer andSchlccediltterer 2002) Genotype proportions in the two populations weretested for conformity to Hardy-Weinberg expectations (HWE) usingexact tests implemented in the program GENEPOP 33 (Raymond andRousset 1995b) The program DAMBE (Xia and Xie 2001) was usedto determine haplotype counts from the mtDNA sequence dataGenetic data from only one individual per nest was used for the aboveanalyses
Relatedness of nestmate workers in each form from both pop-ulations was estimated from genotype distributions at the six micro-satellite loci using the program RELATEDNESS 508 (Goodnight andQueller 1999) Triploid individuals were discarded when performingthe relatedness calculations (see below) The reference population forrelatedness estimation within social forms was defined as either allmonogyne or polygyne nests from a given population For all related-ness calculations nests were weighted equally and standard errors andtheir derivative 95 confidence limits were obtained by jackknifingover nests
Population genetic differentiation was examined in several waysPairwise FST values based on microsatellite and mtDNA sequence datawere calculated between sympatric populations of each social formbetween the two separate geographic populations and between nestpairs within in each area using the program FSTAT (Goudet 2001)Significance of the genetic differentiation measured between pairs of
56 CC Yang et al Fire ants in Taiwan
populations was determined by means of exact tests implemented inGENEPOP 33 (Raymond and Rousset 1995a) Genetic distancevalues (Neis D) also were calculated to further assess divergenceamong populations (Nei 1972)
FST values were also estimated simultaneously at two levels (socialform geographic population) using the hierarchical analysis ofmolecular variance procedure in ARLEQUIN (Excoffier et al1992) Statistical significance of the differentiation at each level wasestablished by means of 20000 data permutations
We tested for correlations between pairwise FST values [FST(1ndashFST)] and the natural logarithm of pairwise geographic distances amongnests from each social form in the two areas except for monogynousindividuals from Chiayi at all six microsatellite loci using the Manteltest in GENEPOP 33 (Raymond and Rousset 1995b) to determine ifthere was a significant positive correlation between genetic differ-entiation and geographic distance indicative of isolation by distance
We also used an individual-based Bayesian method to statisticallyrecognize distinct genetic clusters of individuals and to infer levels ofpopulation admixture from individual multilocus genotypic data(Pritchard et al 2000 Mank and Avise 2004) The method assumesa model with K populations each with characteristic allele frequenciesthat are estimated while individuals are probabilistically assigned toeach population Prior information such as the geographic location ofsamples or colony social form also can be incorporated into the modelto assist in clustering The program STRUCTURE (Pritchard et al2000 URL httppritchbsduchicagoedu) was used to explore theparameter space of each model using Markov chain Monte Carlo(MCMC) algorithms with competing models evaluated on the basis ofthe posterior probabilities given the data model and prior informationSeparate runs of STRUCTURE were performed on samples from eachsingle geographic population only with different numbers of assumedpopulations ranging from one to five (K=1ndash5) and on samples fromboth populations combined into a single dataset (K=1ndash10) Separateruns were performed using the nuclear data only and the nuclear andmtDNA data combined All simulations used 100000 MCMC iter-ations in the burnin phase and 300000 iterations in the data collectionphase with four independent runs conducted on each set of data andparameter values No prior information regarding social form orpopulation of origin were included and all analyses were run assumingcorrelated allele frequencies We ensured accurate estimates of thesimulation values by checking that model parameters equilibratedbefore the end of the burnin phase and that the posterior probabilitieswere consistent across the four runs for each data and parameter setThe estimated membership coefficients (degree of admixture) ofindividuals and their averages for each of the populations wereobtained from the simulations yielding the preferred estimates ofnumbers of distinct clusters as determined by the method of Evannoet al (2005) Again for all of the analyses of genetic differentiation weutilized the multilocus genetic data from only a single individual persampled nest
One signature of a recent population bottleneck is reduction inallelic diversity but not observed heterozygosity simply because suchpopulation bottlenecks tend to reduce allelic number substantiallymore than genetic diversity at polymorphic loci (Luikart et al 1998)We used the program BOTTLENECK (Cornuet and Luikart 1996Piry et al 1999) to identify whether a mode-shift distortion of allelefrequency distributions characteristic of recent bottlenecks was evidentfor either of the two recently established fire ant populations in Taiwanusing the infinite allele model (IAM) and the two phase model (TPM)Significance was assessed by the sign standardized differences andWilcoxon tests Additionally we also compared NA and HE for thepopulations from South America (Ross et al 2007) the USA(Shoemaker et al 2006) and Taiwan and examined the bottleneckeffect in Taiwanese populations if results showed NA similar betweenUSA and Taiwan but considerably lower than those found in SouthAmerica
The proportions of triploid individuals from P and M colonies ofthe two infested areas in Taiwan were estimated from the proportion ofindividuals harboring three different alleles at one or more micro-satellite locus We repeated PCR amplification and genotyping of themicrosatellite loci for all individuals in which three alleles at any locus
was detected to confirm they indeed were triploid Triploid individualswere not used for any of the population genetic analyses above Insteadwe randomly selected one diploid individual from the same nest for allpopulation genetic analyses
Results
Distribution of the monogyne and polygyne social forms
All nests putatively identified as monogyne in the fieldwere identified as such by examining genotype distribu-tions for the 10 nestmate females surveyed at the sixmicrosatellites and by detecting only the Gp-9B alleleamong these workers Also all putative polygyne nestswere confirmed as such by inspection of genotypedistributions of 10 nestmate offspring females for depar-ture from those expected for offspring in monogyne nests(single female mated to a single male) surveyed at sixmicrosatellite loci andor by detecting individuals bearingthe Gp-9b allele In every case the two complementaryapproaches to determining social form were consistent inclassifying nests as either monogyne or polygyne Thepreliminary analyses show that the distribution of the twosocial forms is quite different in the two infested areas(Fig 1) In Taoyuan townships centrally located in thecounty (Fig 1b) harbor a higher proportion of P colonies(P 60 ) whereas surrounding townships (Fig 1b) areinfested more commonly by ants of the M social form (M765) In contrast the large infested area in Chiayiconsists mostly of P nests (92 of colonies collectedFig 1c)
Genetic diversity
From one to five alleles were found at each microsatellitelocus (23 and 26 alleles in the Taoyuan and Chiayipopulations respectively Table 1) A total of sevenprivate alleles (ie unique to a single population) acrossall six microsatellite loci were found Tests for conformityof genotype proportions to Hardy-Weinberg expectationsrevealed only one significant deviation involving Sol-11 inthe Taoyuan polygyne population (Table 1) Score testsreveal that this significant departure resulted from adeficiency of heterozygotes
Analyses of the mtDNA sequence data revealed onlythree unique variants occur in Taiwan (designatedhaplotypes H5 H22 and H36 see Shoemaker andRoss 1996) all of which are commonly observed in fireants from the USA Individuals from the Taoyuanpopulation harbored either the H36 or H5 mtDNAhaplotype whereas all individuals from the Chiayi pop-ulation bear mtDNA haplotype H22 (Fig 1)
Insect Soc Vol 55 2008 Research article 57
Figure 1 (a) Map of Taiwan showing the two areas (Taoyuan and Chiayi counties) infested by Solenopsis invicta (indicated by shading) Histogramsshow the proportion of nests harboring each mtDNA haplotype Pie diagrams in the enlarged map of Taoyuan county (b) show the proportion of themonogyne and polygyne nests found in the central townships (smaller dotted-line circle) and in the surrounding townships (areas between larger andsmaller dotted-line circles) A pie diagram in the enlarged map of Chiayi county (c) indicates the proportion of two social forms in the sampling area(solid-line circle) Social form of each nest was determined by using a Gp-9 allele-specific assay and by examining the distribution of genotype arraysat six microsatellite loci within nests (see text)
Table 1 Genetic diversity measures within each social form from the two study populations of S invicta in Taiwan n denotes total number ofindividuals (nests) for a given social form in each area NA denotes observed number of alleles found at each locus from each populationHE =expected heterozygosity HO = observed heterozygosity
Locus Taoyuan Chiayi
Monogyne(n= 11)
Polygyne( n=19)
Monogyne( n=2)
Polygyne(n=23)
NA HE HO NA HE HO NA HE HO NA HE HO
Sol-6 1 0000 0000 2 0193 0105 3 0833 0800 4 0599 0434
Sol-11 4 0731 0545 4 0755 0526 2 0500 0700 4 0657 0652
Sol-20 5 0814 0818 5 0734 0631 3 0833 0800 4 0679 0782
Sol-42 4 0610 0363 4 0613 0473 2 0500 0450 5 0656 0608
Sol-49 4 0697 0634 5 0768 0842 3 0833 0850 5 0735 0782
Sol-55 3 0634 0454 3 0624 0789 2 0667 0850 4 0486 0608
Mean 35 0581 0469 38 0614 0561 25 0603 0742 43 0635 0645
58 CC Yang et al Fire ants in Taiwan
Within-nest relatedness
The estimates of relatedness among workers withinmonogyne nests from each area were consistent witheach nest headed by single queen mated to a single male(Taoyuan R=071870036 Chiayi R=071970038Fig 2) Relatedness among workers within polygynenests was significantly greater than zero but significantlylower (Plt0001) than the values for monogyne nests(Taoyuan R=011220006 Chiayi R=014440007Fig 2)
Population genetic differentiation
Exact tests based on six microsatellite loci revealedsignificant differentiation between Taoyuan and Chiayipopulations (Plt002) No significant differentiation wasobserved between sympatric populations of the two socialforms in the two areas (Pgt013) Exact tests for differ-entiation using mtDNA sequence data revealed signifi-cant differentiation occurs between the two geographicpopulations (Plt00001) as well as between the two socialforms in Taoyuan (Plt004) but not between the twosocial forms in Chiayi (only one mtDNA variant occurshere)
Nuclear and mtDNA variation was partitioned byestimating FST simultaneously at both levels using theanalysis of molecular variance approach (Excoffier et al 1992) Results of these analyses for the nuclear locishowed that only the between-population variance com-ponent is significant (Plt0000001 Fig 3a) providingadditional support to the above analyses suggesting thesetwo populations are genetically distinct For analysesinvolving mtDNA data both the between-population andbetween-social form variance components are significant(both Plt005 Fig 3b)
Bayesian simulations using STRUCTURE revealedthat individuals from the Taoyuan population represent a
single distinct genetic cluster suggesting no differentia-tion between sympatric social forms exists whether usingnuclear data only or nuclear and mtDNA combinedIdentical results were obtained for individuals from theChiayi population In contrast simulations incorporatingall individuals from the two areas gave posterior proba-bilities that strongly support the existence of two distinctgenetic clusters using dataset from the nuclear loci orcombination of the two classes of markers (K=2 Fig 4)The two clusters hypothesized by the Bayesian methodcorrespond roughly to geographic populations based onaverage membership coefficientsgt05 for one of the twoposterior clusters in each of these regions The firstcluster (indicated by lighter gray) is comprised largely ofindividuals from the Taoyuan population whereas thesecond cluster (indicated by the darker gray) is comprisedmostly of the individuals from Chiayi Identical clusteringresults were obtained when simulations were run incor-porating both the microsatellite and mtDNA data(Fig 4)
Figure 2 Average relatedness (95 confidence interval) valuesamong workers within nests of the polygyne and monogyne social formsfrom Taoyuan and Chiayi counties Taiwan
Figure 3 Partitioning of genetic variance in S invicta at six micro-satellite loci (a) and mtDNA (b) estimated using analysis of molecularvariance This hierarchical approach partitioned the total variancebetween infested regions (Taoyuan and Chiayi counties) between thetwo social forms within each infested region and within the social formsin each region The percentage of the total variance partitioned at eachlevel is shown Asterisks indicate statistically significant differentiationat a given level (a=005)
Figure 4 Assignment of individuals representing Solenopsis invictafrom both study populations to two genetic clusters inferred fromSTRUCTURE simulations (average membership coefficient Q) Eachvertical bar represents the individual membership coefficients dividedinto parts proportional to the individuals proposed ancestry in eachcluster
Insect Soc Vol 55 2008 Research article 59
Isolation by distance and tests for genetic bottlenecks
No correlation exists between pairwise FST or FST(1ndash FST)values and geographic distance for individuals of thepolygyne social form in either population (TaoyuanPgt022 Chiayi Pgt048) Further no significant iso-lation by distance for the monogyne nests from Taoyuanpopulation was found (Pgt065) We did not perform theMantel tests for the Chiayi monogyne population sinceonly two nests were collected
All tests revealed a significant excess of heterozygos-ity in Taoyuan under both models except the sign test(Table 3) A significant excess of heterozygosity wasfound in Chiayi for the infinite allele model but not thetwo phase model (Table 3) Combined with a mode shiftof allele frequency distribution (Table 3 Fig 5a) theTaoyuan population appears to have undergone a recentbottleneck In contrast the most abundant alleles in theChiayi population belong to the low frequency class(lt01 Fig 5b) resulting in a normal L-shape distribu-
tion We also compared NA (number of observed allelesTable 2) and HE (expected heterozygosity Table 2)among the South American USA and Taiwanesepopulations to determine whether a genetic signaturecharacteristic of a bottleneck was evident for the latterpopulations In order to minimize the effect of populationsize we randomly selected 30 individuals from the USAand South American dataset (Shoemaker et al 2006Ross et al 2007) Results showed that the NA and HE inTaiwan and the USA populations were consistentlylower than in South American fire ant populations Twopopulations in Taiwan had slightly lower NA than inUSA but had similar HE values
Additional findings
Analyses of individual genotypes revealed that 108 ofthe 190 workers sampled from polygyne nests in Taoyuanand 11 of 230 individuals from polygyne nests in Chiayi
Table 2 Genetic diversity measures within the two study populations of S invicta in Taiwan NA denotes observed number of alleles found at eachlocus from each population HE =expected heterozygosity HO = observed heterozygosity Numbers in parentheses represent the number of privatealleles occurring in only one of the two Taiwanese populations Genetic diversity measures for two population from the USA and South Americaalso are shown (see text)
Source of collection Population Sample size Mean no of alleles NA HE HO
TaiwanTaoyuan county 30 38 23 (2) 0609 0527Chiayi county 25 48 26 (5) 0636 0660
USAMississippi 30 57 34 0651 0559western Louisiana 30 58 35 0628 0607
South AmericaFormosa Argentina 30 158 195 0835 0842Corrientes Argentina 30 158 95 0836 0826
Figure 5 Distribution of allele frequencies at six microsatellite loci for ants (single individual per nest) from the Taoyuan (a) and Chiayi populations(b)
60 CC Yang et al Fire ants in Taiwan
harbored three distinct alleles whereas all of the individ-uals sampled from monogyne nests harbored at most twodistinct alleles These results indicate that a substantialnumber of polygyne fire ants are triploid a findingconsistent with an earlier study of fire ants from theUSA (Krieger et al 1999)
Discussion
We studied the colony genetic and population structure ofthe invasive fire ant S invicta in Taiwan by collecting andanalyzing microsatellite genotypic data and mtDNAsequence data of ants collected from two separateinfested areas Our genetic analyses revealed severalpatterns which are discussed in more detail below andcan be summarized as follows 1) Estimates of within nestrelatedness were significantly greater than zero for bothsocial forms from both populations 2) No significantisolation by distance was found among nests within eachsocial form from either population 3) SignificantmtDNA but no nuclear differentiation occurs betweensympatric social forms in Taoyuan (no differentiationbetween forms at either genome is evident in Chiayi) 4)Molecular signatures of genetic bottlenecks associatedwith recent introductions are evident in both populations5) The two sampled populations Taoyuan and Chiayi arehighly genetically differentiated at both the nuclear ormtDNA genomes In addition to these general findingswe found numerous triploid individuals in ants of thepolygyne (P) but not the monogyne (M) form anddistinct geographic distribution of each of the two socialforms between the two populations
Colony social organization and patterns of intracolonyworker relatedness
The estimate of relatedness among workers withinmonogyne colonies was not significantly different from075 the predicted value for a nest headed by one singly-mated queen These data also are consistent with datafrom earlier studies of monogyne S invicta colonies inboth the USA (Ross and Fletcher 1985b) and SouthAmerica (Ross et al 1997) and suggest that the socialand mating biology of M queens is virtually invariantwith M nests representing closed societies headed by onesingly-mated reproductive queen
In contrast colony genetic structure of P social formdoes differ between native South America and intro-duced populations in the USA (Ross et al 1996) Pnests of S invicta in the native range contain relativelyfew queens which are close relatives and relatednessamong nestmate workers is significantly greater thanzero whereas P nests in the USA contain highnumbers of unrelated queens and relatedness amongnestmate workers is not significantly greater than zeroThese differences in queen number and relatedness
between the two ranges are consistent with predictionsbased on differences in the population densities of antsin these two areas (Ross et al 1996) which aresubstantially higher in the USA compared to SouthAmerica (Porter et al 1997) One consequence of thehigher population densities in the USA is that nestingsites are more often saturated such that opportunitiesfor independent founding by queens are more limitedin the introduced range (Ross et al 1996) Theseconstraints on independent founding lead to increasedselection pressure on queens to seek adoption intoexisting colonies and on resident workers of colonies toaccept such queens As queen number within thesenests increases relatedness among nestmate workers isexpected to decrease
Relatedness among workers within P nests in Taiwanis small yet significantly greater than zero and intermedi-ate to the values observed for native and USApopulations The social organization of P nests in Taiwanlikely reflects the fact that nesting sites are much lesssaturated compared to sites in the USA since thepopulation densities of ants in Taiwan currently are muchlower than those in the USA However while such adifference between the two introduced ranges are evidentnow it is unlikely to persist especially if S invictacontinues to spread in Taiwan so that habitats becomesaturated If S invicta continues to spread so that habitatsbecome more saturated future studies of the socialevolution of P populations in Taiwan could shed addi-tional light on whether such a change in habitat avail-ability does in fact lead to a shift in social organizationtowards a higher queen number within nests and lowerrelatedness among nestmates
Patterns of isolation by distance
The lack of isolation by distance among colonies of the Mform within Taoyuan was not unexpected given that mostM queens take part in aerial mating flights oftendispersing several kilometers or more before landingand attempting to found a new nest (Toom et al 1976Ross and Fletcher 1985b Ross et al 1999) Such long-distance dispersal is predicted to erase any signature ofnuclear differentiation among closely spaced nests
The lack of detectable isolation by distance within theP form at both studied sites was somewhat unexpectedconsidering the reproductive habits of P queens whichoften mate in or near their natal nests and form newcolonies by budding or by entering into nearby existingnests (Shoemaker and Ross 1996 Ross et al 1997 Rossand Shoemaker 1997) One explanation for the lack ofdetectable isolation by distance is simply that malesdisperse rather long distances to mate with P queensUnder this scenario one would predict that long distancemale-mediated gene flow would result in homogenizationof allele frequencies at nuclear loci yet significantmicrogeographic mtDNA structure would remain espe-
Insect Soc Vol 55 2008 Research article 61
cially if queen dispersal is limited However no suchpattern was found Significant isolation by distance wasabsent for both the nuclear and mtDNA markers Anadditional explanation for these results is that thesignature of isolation by distance predicted to emergesolely from the natural dispersal biology of these ants isconfounded by the fact that these ants are frequentlyinadvertently transported relatively great distances byhumans Such frequent inadvertent human transport maybe a common characteristic of invasive ants found in closeproximity to human habitats and thus play a role inshaping the genetic structure in introduced areas Finallyit is possible that the lack of a detectable signal of isolationby distance is due to the extremely recent introduction offire ants into Taiwan with insufficient time having passedfor populations to achieve migrationdrift equilibrium
Gene flow between sympatric social forms
The pronounced mtDNA differentiation between sym-patric social forms in Taoyuan is consistent with resultsfrom earlier studies in both South America and the USAshowing that interform gene flow mediated by queens israre or absent Queens of each social form rarely succeedin becoming reproductives in colonies of the alternateform as a result of behavioral decisions (attributed largelyto differences in genotypes of queens at the single geneGp-9) made by both queens and workers (Shoemaker andRoss 1996 Ross and Shoemaker 1997 Keller and Ross1998 DeHeer et al 1999 Goodisman et al 2000 Rossand Keller 2002) The contrasting weak or nonexistentinterform divergence at nuclear genes also is consistentwith studies of S invicta in the USA showing that queensof the P form predominantly mate with males of the Mform where the two forms co-occur (Ross and Shoe-maker 1993 Ross and Keller 1995 Shoemaker andRoss 1996) In the USA most males in P colonies aresterile diploid males which means that P queens relylargely on immigrant M males for mating opportunities(Ross and Fletcher 1985a Ross and Shoemaker 1993)Such interform matings act as a homogenizing force onlyfor nuclear markers The similar frequency of triploidworkers in P nests in Taiwan and the USA suggestsmany diploid males also occur in Taiwan as well (Rosset al 1993 1999 Krieger and Keller 1997 Krieger et al 1999) which means that P queens here must also matewith M males The finding of contrasting patterns ofdifferentiation between the two genomes parallels resultsfrom earlier studies involving S invicta in both the USAand South America (Shoemaker and Ross 1996 Rosset al 1997 Shoemaker et al 2006) One final caveathowever is that we cannot completely rule out thepossibility that the lack of detectable nuclear differ-entiation is due to insufficient time having passed fordivergence to occur despite restricted gene flow betweenthe two forms
Understanding the distributions of the two socialforms in the two infested areas in Taiwan each of whichexhibits rather distinct dispersal strategies is importantfor developing effective control strategies of fire ants inthese infested areas (Porter et al 1991) The distribu-tional pattern of social forms in Taoyuan suggests that theexpansion of this population has involved persistentoutward movement of ants from the core infestationcoupled with frequent long distance movement of Mreproductives into outlying areas (Fig 1b) Based onthese results we recommend that bait broadcasting andsurveillance efforts in Taoyuan should include not onlythe currently infested areas but also should include anarea extending well beyond the infested areas In contrastto the distributional pattern of social forms in Taoyuanthe large infested area in Chiayi consists mostly of P nests(92 of colonies collected Fig 1c) However despitethis difference we suggest a similar eradication strategyshould be employed This is because even thoughcomprehensive bait broadcasting within the entire areamay be more effective than for Taoyuan because of thelow queen vagility and relatively slow rate of reinvasionby budding of polygynous queens (Porter et al 1991)some M colonies do occur in Chiayi and there also ispotential for additional M colonies to arise locally fromthe P social form a possibility that is especially morelikely in areas of low density (Ross and Shoemaker 1997DeHeer and Tschinkel 1998 DeHeer et al 1999)
Genetic bottlenecks in Taoyuan and Chiayi populations
Simulations and statistical tests using the program BOT-TLENECK provide evidence of a significant geneticbottleneck associated with the introduction of fire antsinto Taoyuan with only one exception (Table 3) How-ever this single exception is based on a sign test whichhas relatively low statistical power when the number ofloci is below 20 such as our study (Cornuet and Luikart1996) In Chiayi significant excess in heterozygosity usingthe infinite alleles model (IAM) was observed for all testsbut not for the two-phase model (TPM Table 3) BecauseLuikart and Cornuet (1998) suggest that implementingthe test under the IAM model may be more reliable thanother models these results coupled with finding thatallelic frequencies fit a normal L-shaped distributionprovide evidence for a genetic bottleneck Interestinglyestimates of NA and HE (Table 2) are remarkably similarfor introduced populations from Taiwan and the USAand considerably lower than those from the native SouthAmerican range While the geographic origin of ants inTaiwan is unknown the similar genetic diversity measuresbetween the introduced ranges raises the issue that theintroduction of ants into Taiwan has left only a modestfootprint on patterns of diversity if these ants originatedfrom the USA
62 CC Yang et al Fire ants in Taiwan
Genetic differentiation between Taoyuan and Chiayipopulations
All analyses of population differentiation using themicrosatellite and mtDNA data indicate that the twogeographic populations are strongly differentiated andthus likely originated from separate invasions Whilemultiple lines of evidence support this notion thestrongest evidence is the confinement each of the threemtDNA variants to only one of the two geographicpopulations Interestingly all three variants found inTaiwan are the same three variants commonly found inants from the USA (Shoemaker et al 2006) China(GenBank accession numbers DQ831670-DQ831672)and Australia (CCY and DDS unpublished results) Theoccurrence of these three variants in all recently invadedareas suggests that ants from Taiwan could have origi-nated from any of these other areas We currently aregenerating the necessary baseline data for ants from all ofthese introduced regions as well as from populations intheir native range to infer the source population andinvasion history of ants in Taiwan and these other areas
From the perspective of managing fire ants in Taiwantreating each infested area as a separate management unitwould appear more reasonable and means that scientistsprobably should focus on separate plans for eliminationof two isolated populations (Hampton et al 2004) Onthe other hand because these two areas are at high risk ofconnectivity via long distance human-mediated dispersalcare must be taken to prevent reinvasion into these areaseven after successful eradication Stringent inter-countyquarantine measures such as movement control shouldbe established to monitor the risk of transportation of fireant-infested goods to prevent jump dispersal between thetwo areas and decrease probability of successful erad-ication (Drees and Gold 2003)
Acknowledgments
The authors would like to thank Chung-Chi Lin for providing most ofthe specimens for this study including the first identified samples inTaoyuan and Chiayi counties We are particularly grateful to Hwei-yuChang Hon-Tsen Yu and Chia-Hung Hsieh for helpful discussions andcomments on earlier versions of this manuscript Comments by twoanonymous reviewers greatly improved this manuscript This research
was financially supported by grants from Academia Sinica Taiwan Theuse of trade firm or corporation names in this publication is for theinformation and convenience of the reader Such use does notconstitute an official endorsement or approval by the United StatesDepartment of Agriculture or the Agricultural Research Service of anyproduct or service to the exclusion of others that may be suitable
References
Ahrens ME Ross KG and Shoemaker DD 2005 Phylogeographicstructure of the fire ant Solenopsis invicta in its native SouthAmerican range Roles of natural barriers and habitat connectivityEvolution 59 1733ndash1743
Allen CR Demarais S and Lutz RS 1994 Red imported fire antimpact on wildlife an overview Texas J Sci 46 51ndash59
Allen CR Lutz RS and Demarais S 1998 Ecological effects of theinvasive nonindigenous ant Solenopsis invicta on native verte-brates The wheels on the bus Transactions of the North AmericanWildlife and Natural Resources Conference 63 56 ndash65
Allen CR Willey RD Myers PE Horton PM and Buffa J 2000Impact of red imported fire ant infestation on northern bobwhitequail abundance trends in southeastern United States J AgrUrban Entomol 17 43ndash51
Allen CR Forys EA Rice KG and Wojcik DP 2001 Effects of fireants (Hymenoptera Formicidae) on hatching turtles and preva-lence of fire ants on sea turtle nesting beaches in Florida FloridaEntomol 84 250ndash253
Bouwma AM Ahrens ME DeHeer CJ and Shoemaker DD 2006Distribution and prevalence of Wolbachia in introduced popula-tions of the fire ant Solenopsis invicta Insect Mol Biol 15 89ndash93
Briano JA Calcaterra LA Vander Meer R Valles SM and LivoreJP 2006 New survey for the fire ant microsporidia Vairimorphainvictae and Thelohania solenopsae in southern South Americawith observations on their field persistence and prevalence of dualinfections Environ Entomol 35 1358ndash1365
Buckley A 1999 Fire ants in California Am Bee J 139 88Carroll CR and Hoffman CA 2000 The pervasive ecological effects
of invasive species Exotic and native fire ants In Invertebrates asWeb-Masters in Ecosystems (Coleman DC and Hendrix PF Eds)Wallingford Oxon UK pp 221ndash232
Carruthers RI 2003 Invasive species research in the United StatesDepartment of Agriculture ndash Agricultural Research Service PestManag Sci 59 827ndash834
Chen JSC Shen CH and Lee HJ 2006 Monogynous and polygynousred imported fire ants Solenopsis invicta Buren (HymenopteraFormicidae) in Taiwan Environ Entomol 35 167ndash172
Cornuet JM and Luikart G 1996 Description and power analysis oftwo tests for detecting recent population bottlenecks from allelefrequency data Genetics 144 2001ndash2014
Davis LR Vander Meer RK and Porter SD 2001 Red imported fireants expand their range across the West Indies Florida Entomol84 735ndash736
DeHeer CJ and Tschinkel WR 1998 The success of alternativereproductive tactics in monogyne Solenopsis invicta significancefor transitions in social organization Behav Ecol 9 130ndash135
DeHeer CJ Goodisman MAD and Ross KG 1999 Queen dispersalstrategies in the multiple-queen form of the fire ant Solenopsisinvicta Am Nat 153 660ndash675
Dieringer D and Schlccediltterer C 2002 Microsatellite analyser (MSA) aplatform independent analysis tool for large microsatellite datasets Mol Ecol Notes 3 167ndash169
Drake JA Mooney HA Castri F Groves RH Kruger MRejmnek M and Williamson M 1989 Biological Invasions AGlobal Perspective Chichester UK 550 pp
Drees BM and Vinson SB 1990 Comparison of the control ofmonogynous and polygynous forms of the red imported fire ant(Hymenoptera Formicidae) with a chloripyrifos mound drench JEntomol Sci 25 317ndash324
Table 3 Tests for genetic bottlenecks using the infinite allele model(IAM) and two-phase model (TPM 90 stepwise mutation model and10 multistep mutation model [Garza and Williamson 2001])implemented in the program BOTTLENECK (a=005)Significance at given statistic level
Population Simulatedmodel
Significancetest
Standardizeddifferences test
Wilcoxontest
Taoyuan IAM 0148 0003 0015TPM 0186 0035 0023
Chiayi IAM 0037 0022 0007TPM 0521 0293 0281
Insect Soc Vol 55 2008 Research article 63
Drees BM and Gold RE 2003 Development of integrated pestmanagement programs for the red imported fire ant (Hymeno-ACHTUNGTRENNUNGptera Formicidae) J Entomol Sci 38 170ndash180
Eubanks MD 2001 Estimates of the direct and indirect effects of redimported fire ants on biological control in field crops Biol Control21 35 ndash43
Evanno G Regnaut S and Goudet J 2005 Detecting the number ofclusters of individuals using the software STRUCTURE asimulation study Mol Ecol 14 2611ndash2620
Excoffier L Smouse PE and Quattro JM 1992 Analysis of molecularvariance inferred from metric distances among DNA haplotypesApplication to human mitochondrial DNA restriction dataGenetics 131 479ndash491
Forys EA Quistorff A and Allen CR 2001 Potential fire ant(Hymenoptera Formicidae) impact on the endangered Schausswallowtail (Lepidoptera Papilionidae) Florida Entomol 84 254ndash258
Garza JC and Williamson EG 2001 Detection of reduction inpopulation size using data from microsatellite loci Mol Ecol 10 305ndash318
Giraud T Pedersen JS and Keller L 2002 Evolution of supercoloniesthe Argentine ants of southern Europe Proc Natl Acad Sci U SA 99 6075ndash6079
Giuliano WM Allen CR Lutz RS and Demarais S 1996 Effects ofred imported fire ants on northern bobwhite chicks J WildlManage 60 309ndash313
Glancey BM Craig CH Stringer CE and Bishop PM 1973Multiple fertile queens in colonies of the imported fire antSolenopsis invicta J Georgia Entomol Soc 8 327ndash328
Goodisman MAD DeHeer CJ and Ross KG 2000 Mating behaviorof queens from multiple-queen colonies of Solenopsis invicta JInsect Behav 13 455ndash468
Goodnight KF and Queller DC 1999 Computer software forperforming likelihood tests of pedigree relationship using geneticmarkers Mol Ecol 8 1231ndash1234
Gotelli NJ and Arnett AE 2000 Biogeographic effects of red fire antinvasion Ecol Lett 3 257ndash261
Gotzek D and Ross KG 2007 Genetic regulation of colony socialorganization in fire ants An integrative overview Q Rev Biol 82 201ndash226
Goudet J 2001 FSTAT A Program to Estimate and Test GeneDiversities and Fixation Indices Version 293 http wwwunilch izeasoftwares fstathtml
Greenberg L Fletcher DJC and Vinson SB 1985 Differences inworker size and mound distribution in monogynous and polygy-nous colonies of the fire ant Solenopsis invicta Buren J KansasEntomol Soc 58 9ndash18
Hampton JO Spencer PBS Alpers DL Twigg LE WoolnoughAP Doust J Higgs T and Pluske J 2004 Molecular techniqueswildlife management and the importance of genetic populationstructure and dispersal a case study with feral pigs J Appl Ecol41 735ndash743
Henshaw MT Kunzmann N Vanderwoude C Sanetra M andCrozier RH 2005 Population genetics and history of theintroduced fire ant Solenopsis invicta Buren (HymenopteraFormicidae) in Australia Aus J Entomol 44 37ndash44
Hccedillldobler B and Wilson EO 1990 The Ants The Belknap Press ofHarvard University Press Cambridge Massachusetts 732 pp
Huang TC Chou YC and Chou HC 2004 The infestation andcontrol of the red imported fire ant in Taiwan In Proceedingsof the Symposium on the Control of the Red Imported Fire AntBureau of Animal and Plant Health Inspection and QuarantineCouncil of Agriculture Executive Yuan Taipei Taiwan pp1 ndash 13
Jaquiry J Vogel V and Keller L 2005 Multilevel genetic analyses oftwo European supercolonies of the Argentine ant Linepithemahumile Mol Ecol 14 589ndash598
Jeyaprakash A and Hoy MA 2000 Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropods Insect Mol Biol 9 393ndash405
Jouvenaz DP 1990 Approaches to biological control of fire ants in theUnited States In Applied Myrmecology A World Perspective(Vander Meer RK Jaffe K and Cedeno A Eds) Westview PressBoulder Colorado pp 620ndash627
Kaspari M 2000 Do imported fire ants impact canopy arthropodsEvidence from simple arboreal pitfall traps Southwestern Nat 45 118ndash122
Keller L and Ross KG 1998 Selfish genes a green beard in the red fireant Nature 394 573ndash575
Kemp SF deShazo RD Moffitt JE Williams DF and Buhner WA2000 Expanding habitat of the imported fire ant (Solenopsisinvicta) A public health concern J Allergy Clin Immunol 105 683ndash691
Kopachena JG Buckley AJ and Potts GA 2000 Effects of the redimported fire ant (Solenopsis invicta) on reproductive success ofbarn swallows (Hirundo rustica) in northeast Texas SouthwesternNat 45 477ndash482
Krieger MJB and Keller L 1997 Polymorphism at dinucleotidemicrosatellite loci in fire ant Solenopsis invicta populations MolEcol 6 997ndash999
Krieger MJB Ross KG Chang CWY and Keller L 1999Frequency and origin of triploidy in the fire ant Solenopsis invictaHeredity 82 142ndash150
Krieger MJB and Ross KG 2002 Identification of a major generegulating complex social behavior Science 295 328ndash332
Lee CE 2002 Evolutionary genetics of invasive species Trends EcolEvol 17 386ndash391
Lofgren CS Banks WA and Glancey BM 1975 Biology and controlof imported fire ants Annu Rev Entomol 20 1ndash30
Lofgren CS 1986 History of imported fire ants in the United States InFire Ants and Leaf Cutting Ants Biology and Management(Lofgren CS and Vander Meer RK Eds) Westview PressBoulder Colorado pp 36ndash49
Luikart G Allendorf FW Cornuet JM and Sherwin WB 1998Distortion of allele frequency distributions provides a test forrecent population bottlenecks J Hered 89 238ndash247
Luikart G and Cornuet JM 1998 Empirical evaluation of a test foridentifying recently bottlenecked populations from allele frequen-cy data Conserv Biol 12 228ndash237
MacKay WP and Fagerlund R 1997 Range expansion of the redimported fire ant Solenopsis invicta Buren (Hymenoptera For-micidae) into New Mexico and extreme western Texas ProcEntomol Soc Wash 99 757ndash758
Mank JE and Avise JC 2004 Individual organisms as units of analysisBayesian-clustering alternatives in population genetics Genet Res84 135ndash143
May GE Gelembiuk GW Panov VE Orlova MI and Lee CE2006 Molecular ecology of zebra mussel invasions Mol Ecol 15 1021ndash1031
McCubbin KI and Weiner JM 2002 Fire ants in Australia A newmedical and ecological hazard Med J Aus 176 518ndash519
Morrison LW 2000 Mechanisms of interspecific competition amongan invasive and two native fire ants Oikos 90 238ndash252
Morrison LW 2002 Long-term impacts of an arthropod-communityinvasion by the imported fire ant Solenopsis invicta Ecology 83 2337ndash2345
Nattrass R and Vanderwoude C 2001 A preliminary investigation ofthe ecological effects of red imported fire ants (Solenopsis invicta)in Brisbane Ecol Manag Resto 2 220ndash223
Nei M 1972 Genetic distance between populations Am Nat 106 283ndash292
Orr MR 1996 Host manipulation by Wolbachia is a neutral trait withinsingle populations Anim Behav 51 1183ndash1185
Patterson RS 1994 Biological control of introduced ant species InExotic Ants Biology Impact and Control of Introduced Species(Williams DF Ed) Westview Press Boulder Colorado pp 293ndash308
Piry S Luikart G and Cornuet JM 1999 BOTTLENECK acomputer program for detecting recent reductions in the effectivepopulation size using allele frequency data J Hered 90 502ndash503
64 CC Yang et al Fire ants in Taiwan
Porter SD and Savignano DA 1990 Invasion of polygyne fire antsdecimates native ants and disrupts arthropod community Ecology7 2095ndash2106
Porter SD Bhatkar A Mulder R Vinson SB and Clair DJ 1991Distribution and density of polygyne fire ants (HymenopteraFormicidae) in Texas J Econ Entomol 84 866ndash874
Porter SD Fowler HG and MacKay WP 1992 Fire ant mounddensities in the United States and Brazil (Hymenoptera Formi-cidae) J Econ Entomol 85 1154ndash1161
Porter SD Williams DF Patterson RS and Fowler HG 1997Intercontinental differences in the abundance of Solenopsis fireants (Hymenoptera Formicidae) Escape from natural enemiesEnviron Entomol 26 373ndash384
Pritchard JK Stephens M and Donnelly P 2000 Inference ofpopulation structure using multilocus genotype data Genetics155 945ndash959
Raymond M and Rousset F 1995a An exact test for populationdifferentiation Evolution 49 1280ndash1283
Raymond M and Rousset F 1995b GENEPOP (version 12)population genetics software for exact tests and ecumenicism JHered 86 248ndash249
Reuter M Pedersen JS and Keller L 2005 Loss of Wolbachiainfection during colonisation in the invasive Argentine ant Line-pithema humile Heredity 94 364ndash369
Ross KG and Fletcher DJC 1985a Genetic origin of male diploidy inthe fire ant Solenopsis invicta (Hymenoptera Formicidae) and itsevolutionary significance Evolution 39 888ndash903
Ross KG and Fletcher DJC 1985b Comparative study of genetic andsocial structure in two forms of the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 17 349ndash356
Ross KG and Fletcher DJC 1986 Diploid male productionndashasignificant colony mortality factor in the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 19 283ndash291
Ross KG and Shoemaker DD 1993 An unusual pattern of gene flowbetween the two social forms of the fire ant Solenopsis invictaEvolution 47 1595ndash1605
Ross KG Vargo EL Keller L and Trager JC 1993 Effect of afounder event on variation in the genetic sex-determining system ofthe fire ant Solenopsis invicta Genetics 135 843ndash854
Ross KG and Keller L 1995 Joint influence of gene flow and selectionon a reproductively important genetic polymorphism in the fire antSolenopsis invicta Am Nat 146 325ndash348
Ross KG Vargo EL and Keller L 1996 Social evolution in a newenvironment The case of introduced fire ants Proc Natl AcadSci U S A 93 3021ndash3025
Ross KG 1997 Multilocus evolution in fire ants Effects of selectiongene flow and recombination Genetics 145 961ndash974
Ross KG Krieger MJB Shoemaker DD Vargo EL and Keller L1997 Hierarchical analysis of genetic structure in native fire antpopulations Results from three classes of molecular markersGenetics 147 643ndash655
Ross KG and Shoemaker DD 1997 Nuclear and mitochondrialgenetic structure in two social forms of the fire ant Solenopsisinvicta Insights into transitions to an alternate social organizationHeredity 78 590ndash602
Ross KG and Keller L 1998 Genetic control of social organization inan ant Proc Natl Acad Sci U S A 95 14232ndash14237
Ross KG Shoemaker DD Krieger MJ DeHeer CJ and Keller L1999 Assessing genetic structure with multiple classes of molecularmarkers A case study involving the introduced fire ant Solenopsisinvicta Mol Biol Evol 16 525ndash543
Ross KG and Keller L 2002 Experimental conversion of colony socialorganization by manipulation of worker genotype composition infire ants (Solenopsis invicta) Behav Ecol Sociobiol 51 287ndash295
Ross KG Krieger MJB and Shoemaker DD 2003 Alternativegenetic foundations and evolutionary routes for a key socialpolymorphism in fire ants Genetics 165 1853ndash1867
Ross KG Krieger MJB Keller L and Shoemaker DD 2007Genetic variation and structure in native populations of the fire antSolenopsis invicta Evolutionary and demographic implicationsBiol J Linnean Soc 92 541ndash560
Rousset F 1997 Genetic differentiation and estimation of gene flowfrom F-statistics under isolation by distance Genetics 145 1219ndash1228
Sax DF Stachowicz JJ and Gaines SD 2005 Species InvasionsInsights into Ecology Evolution and Biogeography SinauerAssociates Sunderland Massachussetts 495 pp
Shoemaker DD and Ross KG 1996 Effects of social organization ongene flow in the fire ant Solenopsis invicta Nature 383 613ndash616
Shoemaker DD Ross KG Keller L Vargo EL and Werren JH2000 Wolbachia infections in native and introduced populations offire ants (Solenopsis spp) Insect Mol Biol 9 661ndash673
Shoemaker DD Ahrens M Sheill L Mescher M Keller L and RossKG 2003 Distribution and prevalence of Wolbachia infections innative populations of the fire ant Solenopsis invicta (HymenopteraFormicidae) Environ Entomol 32 1329ndash1336
Shoemaker DD DeHeer CJ Krieger MJB and Ross KG 2006Population genetics of the invasive fire ant Solenopsis invicta(Hymenoptera Formicidae) in the USA Ann Entomol Soc Am99 1213ndash1233
Simon C Frati F Beckenbach A Crespi B Liu H and Flook P 1994Evolution weighting and phylogenetic utility of mitochondrialgene sequences and a compilation of conserved polymerase chainreaction primers Ann Entomol Soc Am 87 651ndash701
Toom PM Cupp E Johnson CP and Griffin I 1976 Utilization ofbody reserves for minim brood development by queens of theimported fire ant Solenopsis invicta J Insect Physiol 22 217ndash220
Tsutsui ND and Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepethema humile) in its nativeand introduced ranges Evolution 55 976ndash985
Tsutsui ND Kauppinen SN Oyafuso AF and Grosberg RK 2003The distribution and evolutionary history of Wolbachia infection innative and introduced populations of the invasive argentine ant(Linepithema humile) Mol Ecol 12 3057ndash3068
Valles SM and Porter SD 2003 Identification of polygyne andmonogyne fire ant colonies (Solenopsis invicta) by multiplex PCRof Gp-9 alleles Insect Soc 50 199ndash200
Valles SM Oi DH Briano JA and Williams DF 2004 Simultaneousdetection of Vairimorpha invictae (Microsporidia Burenellidae)and Thelohania solenopsae (Microsporidia Thelohaniidae) in fireants by PCR Florida Entomol 87 85ndash87
Vander Meer R K and Porter SD 2001 Fate of newly-mated queensintroduced into monogyne and polygyne Solenopsis invicta (Hy-menoptera Formicidae) colonies Ann Entomol Soc Am 94 289ndash297
Vander Meer RK Jaffe K and Cedeno A 1990 Applied Myrmecol-ogy A World Perspective Westview Press Boulder CO 741 pp
Vinson SB 1994 Impact and invasion of Solenopsis invicta (Buren) onnative food webs In Exotic Ants Biology Impact and Control ofIntroduced Species (Williams DF Ed) Westview Press BoulderColorado pp 293ndash308
Vinson SB 1997 Invasion of the red imported fire ant (HymenopteraFormicidae) Spread biology and impact Am Entomol 43 23ndash29
Williams DF and Porter SD 1994 Fire ant control Science 264 1653ndash1653
Wojcik D Allen CR Brenner RJ Forys EA Jouvenaz D and LutzRS 2001 Red imported fire ants Impact on biodiversity AmEntomol 47 16ndash23
Xia X and Xie Z 2001 DAMBE Data analysis in molecular biologyand evolution J Hered 92 371ndash373
Insect Soc Vol 55 2008 Research article 65

species that have been introduced to more than onelocation can increase our understanding of what charac-teristics of these species are important for invasionsuccess (Lee 2002 May et al 2006)
Many social insects particularly ants are well knownas being important invasive pest species Indeed theinvasive potential and abilities of ants to colonize almostevery available habitat is such that they often are referredto as tramp species (Hccedillldobler and Wilson 1990Vander Meer et al 1990) One of the most notoriousinvasive ant species is the red imported fire ant (RIFA)Solenopsis invicta Buren (Lofgren 1986 Jouvenaz 1990Patterson 1994 Orr 1996) This exotic pest was inad-vertently introduced into the USA from South Americaabout 75 years ago Since that time it has spreadthroughout the southern states in the USA S invicta isof profound economic importance in the USA because1) it is an aggressive stinging insect causing massenvenomation incidents and hypersensitivity reactionsin humans 2) it occurs primarily in human-modifiedhabitats 3) it constructs large mounds that are unsightlyand capable of damaging farm machinery 4) it feeds onseveral important cultivated plants and tends homopter-ans that are also plant pests and 5) it negatively affectspopulations of native ants and other ground-dwellinganimals (Lofgren et al 1975 Lofgren 1986 Porter andSavignano 1990 Allen et al 1994 1998 2000 2001Vinson 1994 1997 Giuliano et al 1996 Carroll andHoffman 2000 Gotelli and Arnett 2000 Kaspari 2000Kemp et al 2000 Kopachena et al 2000 Eubanks2001 Forys et al 2001 Wojcik et al 2001 Morrison2002) These pest attributes of S invicta presumably areattributable largely to the fact that the introducedpopulations have almost none of the competitors andnatural enemies that normally act to suppress theirpopulations with the effect that population densities inthe USA are orders of magnitude greater than in thenative South American range (Porter et al 1992 1997Morrison 2000)
More recently S invicta has been inadvertentlyintroduced to other regions of the world including severalwestern states (eg New Mexico California) the Carri-bean Australia mainland China and Taiwan (MacKayand Fagerlund 1997 Buckley 1999 Davis et al 2001Nattrass and Vanderwoude 2001 McCubbin and Weiner2002 Huang et al 2004 Chen et al 2006) In all of theseareas S invicta is considered a significant pest such thatits negative agricultural ecological and public healthimpacts can only be expected to intensify in the nearfuture
While an extensive body of literature exists on studiesof S invicta from native South American and introducedpopulations in the USA few studies have been con-ducted on S invicta in these more recently invaded areas(Henshaw et al 2005 Chen et al 2006) Such studies areof interest not only for development of effective inte-grated management strategies for control and eradicationof fire ants in these areas but also provide the opportunity
for comparative studies of the genetic changes associatedwith or occurring during separate invasions (Ross et al 1993 Tsutsui and Case 2001 Giraud et al 2002Jaquiry et al 2005) To that end we present here theresults of a study exploring patterns of nuclear (micro-satellite) and of mtDNA variation in the fire ant S invictafrom two separate infested areas in Taiwan (Taoyuan andChiayi counties Fig 1) While the occurrence of S invictain Taiwan was first reported in 2003 analyses of the sizeand distribution of colonies suggest that this species likelyarrived some 3 ndash 5 years earlier (Huang et al 2004 Chenet al 2006)
Previous studies also have shown that both knownsocial forms of S invicta are present in both of theinfested areas in Taiwan (Huang et al 2004 Chen et al 2006) Two distinct types of colony social organizationoccur within this species (Glancey et al 1973) Coloniesof the monogyne (M) form possess a single egg-layingqueen whereas colonies of the polygyne (P) formpossess several to hundreds of these queens Associationstudies have shown that social form in this and other fireant species depends on the presence within a colony ofspecific coding region variants of the single gene Gp-9(reviewed in Gotzek and Ross 2007) Two classes ofvariants designated as B-like and b-like alleles occur inS invicta Monogyne colonies contain only the B-likeallele whereas polygyne colonies invariably contain theb-like allele as well as the B-like allele (Ross 1997 Rossand Keller 1998 Krieger and Ross 2002) The twosocial forms of S invicta differ not only in colony queennumber and Gp-9 genotypes but also in importantfeatures of their reproductive and dispersal behaviorswhich are expected to have a number of importanteffects on the distribution of genetic variation at variousspatial scales as well as management strategies em-ployed for eradication Thus knowledge of distributionof the two social forms is critical to constructing aneffective predictive model of their spread and expansionand assessing the potential of successful eradication of Sinvicta (Drees and Vinson 1990) Finally previousstudies have shown that the unique social and breedingbiology of each social form can act as strong constraintson particular routes of interform gene flow (Shoemakerand Ross 1996) Specifically the selective elimination ofqueens of the alternate social form by workers (Kellerand Ross 1998 Vander Meer and Porter 2001 Kriegerand Ross 2002 Ross and Keller 2002) and the highproportion of sterile diploid males in P nests inintroduced areas (Ross and Fletcher 1985a 1986Krieger et al 1999) severely restrict female-mediatedgene flow and limit male-mediated interform gene flowmostly to instances involving M males mating with Pfemales (Ross and Shoemaker 1993 1997 Shoemakerand Ross 1996 Shoemaker et al 2006) Therefore thepresence of both social forms in a novel environmentpresents the opportunity to test the generality of thesepatterns observed in earlier studies in the introducedrange in the USA
Insect Soc Vol 55 2008 Research article 55
This study has several objectives First we wished tolearn whether the current distribution of fire ants in thesetwo areas is the result of a single or separate introductionsinto Taiwan The finding of multiple introductions mayindicate that the island of Taiwan is at high risk ofexperiencing additional introductions Second we aimedto infer whether one social form was possibly derivedfrom local populations of the alternate social form as wellas whether substantial gene flow occurs between sym-patric populations of each social form Lastly we utilizedour genetic data to assess whether a genetic signaturecharacteristic of a recent population bottleneck is evidentin either of two areas infested by S invicta While dataexamining the alternative colony genetic structure andassociated genetic differences between native and intro-duced fire ant populations have been proposed for theperiod of several decades after their first introduction tothe USA (Ross and Fletcher 1985b Ross et al 1996Ross et al 1999) the more recent introduction of fireants into Taiwan provides a novel opportunity to study thegenetic changes characteristics of invasiveness of thisspecies over an extremely short time period particularlyin the initial 5 ndash10 years after the original introduction
Materials and methods
Source of ant samples
Ants representing both social forms were collected from two locationsTaoyuan county and Chiayi county in Taiwan (these two locations areseparated by ca 250 km) Initially the social form of S invicta wasdetermined by visual inspection of the average size of the workers theheight of the mounds and the number of queens (Greenberg et al1985) Based on the preliminary determinations of social form in thefield adult workers were collected from nests of each putative socialform separated by distances of 30 ndash5000 (polygyne) and 600ndash2000 m(monogyne) (Fig 1) In total we sampled 30 colonies from Taoyuancounty and 25 colonies from the Chiayi county Collected workers werestored in 95 alcohol and frozen at -20 8C
DNA extractions and determination of social form
Two separate sets of total genomic DNA were extracted fromindividuals sampled from each nest For the first set 10ndash15 workerswere pooled and extracted in bulk using the Viogene DNA extractionkit (Viogene Inc Taipei Taiwan) These bulk-extracted samples wereused in Gp-9 assays for social form determination (Valles and Porter2003) For the second set total genomic DNA was extracted from 10individual workers from each sampled nest These samples were used assource DNA for microsatellite genotyping and mtDNA sequencing(single individual per nest see below) Inspection of the distribution ofmultilocus genotypes from 10 individuals per sampled nest provides apowerful and direct approach for determining the social form of eachnest
Microsatellite analyses
We genotyped 10 workers from each colony representing 550individuals in total at six dinucleotide-repeat microsatellite loci(Table 1) previously developed for S invicta (Table 1 Krieger andKeller 1997) Amplification of the single locus Sol-20 was performed in
10-ml reactions with 30 ndash100 ng DNA 1 ml of 10X Super-Therm GoldPCR buffer 1 ml of 25 mM dNTPs 03 ml of a 10 mM FAM-labeledforward primer 03 ml of a 10 mM unlabeled reverse primer and 015 Uof Super-Therm Gold Hot-start Taq DNA polymerase (AppliedBiosystems) The remaining five loci were amplified in two separatemultiplex PCR reactions In the first reaction Sol-11 Sol-42 and Sol-49were amplified simultaneously in a single tube Reaction cocktails andconditions for Sol-11 Sol-42 and Sol-49 were identical to thosedescribed earlier (Ross et al 2003) except forward primers werelabeled with FAM In the second multiplex reaction Sol-6 and Sol-55were co-amplified in a single 20-ml reaction with 30ndash100 ng DNA 24 mlof 10X Super-Therm Gold PCR buffer 16 ml of 25 mM dNTPs 05 mlof a 10 mM FAM-labeled Sol-6 forward primer 05 ml of a 10 mMunlabeled Sol-6 reverse primer 06 ml of a 10 mM FAM-labeled Sol-55forward primer 06 ml of a 10 mM unlabeled Sol-55 reverse primer and1U of Super-Therm Gold Hot-start Taq DNA polymerase PCRamplifications were carried out on an ABI 9700 thermal cycler(Applied Biosystems) using the following profile initial denaturationat 95 8C for 10 min 30 cycles of 94 8C for 1 min 58 8C for 1 min and72 8C for 1 min a final extension of 72 8C for 30 min All PCR productswere visualized on an ABI 3100 genetic analyzer using laser detectionand scoring of genotypes was performed using the software GENES-CAN 312 (Applied Biosystems)
MtDNA sequencing
A 920-bp fragment of the mitochondrial genome that includes portionsof the cytochrome oxidase subunit I (COI) and subunit II (COII) geneswas amplified from a single individual per nest using the primers C1-J-2195 (Simon et al 1994) and DDS-COII-4 (Ross and Shoemaker1997) PCR reactions were carried out using identical reaction cocktailsand thermal cycling profiles described in Ahrens et al (2005) MtDNAamplicons were purified and used directly in standard fluorescent cycle-sequencing PCR reactions (ABI Prism Big Dye terminator chemistryApplied Biosystems) Sequencing reactions were subsequently purifiedand run on an ABI 3730 sequencer All mtDNA amplicons weresequenced in both directions and corresponding sequence pairs wereedited and combined into a single consensus sequence file Allsequences were then aligned by eye using previously published Sinvicta mtDNA sequence obtained from GenBank (AY950762)
Genetic analyses
The number of alleles allele frequencies observed heterozygosity andexpected heterozygosity at each microsatellite locus for both popula-tions were calculated using the program MSA 300 (Dieringer andSchlccediltterer 2002) Genotype proportions in the two populations weretested for conformity to Hardy-Weinberg expectations (HWE) usingexact tests implemented in the program GENEPOP 33 (Raymond andRousset 1995b) The program DAMBE (Xia and Xie 2001) was usedto determine haplotype counts from the mtDNA sequence dataGenetic data from only one individual per nest was used for the aboveanalyses
Relatedness of nestmate workers in each form from both pop-ulations was estimated from genotype distributions at the six micro-satellite loci using the program RELATEDNESS 508 (Goodnight andQueller 1999) Triploid individuals were discarded when performingthe relatedness calculations (see below) The reference population forrelatedness estimation within social forms was defined as either allmonogyne or polygyne nests from a given population For all related-ness calculations nests were weighted equally and standard errors andtheir derivative 95 confidence limits were obtained by jackknifingover nests
Population genetic differentiation was examined in several waysPairwise FST values based on microsatellite and mtDNA sequence datawere calculated between sympatric populations of each social formbetween the two separate geographic populations and between nestpairs within in each area using the program FSTAT (Goudet 2001)Significance of the genetic differentiation measured between pairs of
56 CC Yang et al Fire ants in Taiwan
populations was determined by means of exact tests implemented inGENEPOP 33 (Raymond and Rousset 1995a) Genetic distancevalues (Neis D) also were calculated to further assess divergenceamong populations (Nei 1972)
FST values were also estimated simultaneously at two levels (socialform geographic population) using the hierarchical analysis ofmolecular variance procedure in ARLEQUIN (Excoffier et al1992) Statistical significance of the differentiation at each level wasestablished by means of 20000 data permutations
We tested for correlations between pairwise FST values [FST(1ndashFST)] and the natural logarithm of pairwise geographic distances amongnests from each social form in the two areas except for monogynousindividuals from Chiayi at all six microsatellite loci using the Manteltest in GENEPOP 33 (Raymond and Rousset 1995b) to determine ifthere was a significant positive correlation between genetic differ-entiation and geographic distance indicative of isolation by distance
We also used an individual-based Bayesian method to statisticallyrecognize distinct genetic clusters of individuals and to infer levels ofpopulation admixture from individual multilocus genotypic data(Pritchard et al 2000 Mank and Avise 2004) The method assumesa model with K populations each with characteristic allele frequenciesthat are estimated while individuals are probabilistically assigned toeach population Prior information such as the geographic location ofsamples or colony social form also can be incorporated into the modelto assist in clustering The program STRUCTURE (Pritchard et al2000 URL httppritchbsduchicagoedu) was used to explore theparameter space of each model using Markov chain Monte Carlo(MCMC) algorithms with competing models evaluated on the basis ofthe posterior probabilities given the data model and prior informationSeparate runs of STRUCTURE were performed on samples from eachsingle geographic population only with different numbers of assumedpopulations ranging from one to five (K=1ndash5) and on samples fromboth populations combined into a single dataset (K=1ndash10) Separateruns were performed using the nuclear data only and the nuclear andmtDNA data combined All simulations used 100000 MCMC iter-ations in the burnin phase and 300000 iterations in the data collectionphase with four independent runs conducted on each set of data andparameter values No prior information regarding social form orpopulation of origin were included and all analyses were run assumingcorrelated allele frequencies We ensured accurate estimates of thesimulation values by checking that model parameters equilibratedbefore the end of the burnin phase and that the posterior probabilitieswere consistent across the four runs for each data and parameter setThe estimated membership coefficients (degree of admixture) ofindividuals and their averages for each of the populations wereobtained from the simulations yielding the preferred estimates ofnumbers of distinct clusters as determined by the method of Evannoet al (2005) Again for all of the analyses of genetic differentiation weutilized the multilocus genetic data from only a single individual persampled nest
One signature of a recent population bottleneck is reduction inallelic diversity but not observed heterozygosity simply because suchpopulation bottlenecks tend to reduce allelic number substantiallymore than genetic diversity at polymorphic loci (Luikart et al 1998)We used the program BOTTLENECK (Cornuet and Luikart 1996Piry et al 1999) to identify whether a mode-shift distortion of allelefrequency distributions characteristic of recent bottlenecks was evidentfor either of the two recently established fire ant populations in Taiwanusing the infinite allele model (IAM) and the two phase model (TPM)Significance was assessed by the sign standardized differences andWilcoxon tests Additionally we also compared NA and HE for thepopulations from South America (Ross et al 2007) the USA(Shoemaker et al 2006) and Taiwan and examined the bottleneckeffect in Taiwanese populations if results showed NA similar betweenUSA and Taiwan but considerably lower than those found in SouthAmerica
The proportions of triploid individuals from P and M colonies ofthe two infested areas in Taiwan were estimated from the proportion ofindividuals harboring three different alleles at one or more micro-satellite locus We repeated PCR amplification and genotyping of themicrosatellite loci for all individuals in which three alleles at any locus
was detected to confirm they indeed were triploid Triploid individualswere not used for any of the population genetic analyses above Insteadwe randomly selected one diploid individual from the same nest for allpopulation genetic analyses
Results
Distribution of the monogyne and polygyne social forms
All nests putatively identified as monogyne in the fieldwere identified as such by examining genotype distribu-tions for the 10 nestmate females surveyed at the sixmicrosatellites and by detecting only the Gp-9B alleleamong these workers Also all putative polygyne nestswere confirmed as such by inspection of genotypedistributions of 10 nestmate offspring females for depar-ture from those expected for offspring in monogyne nests(single female mated to a single male) surveyed at sixmicrosatellite loci andor by detecting individuals bearingthe Gp-9b allele In every case the two complementaryapproaches to determining social form were consistent inclassifying nests as either monogyne or polygyne Thepreliminary analyses show that the distribution of the twosocial forms is quite different in the two infested areas(Fig 1) In Taoyuan townships centrally located in thecounty (Fig 1b) harbor a higher proportion of P colonies(P 60 ) whereas surrounding townships (Fig 1b) areinfested more commonly by ants of the M social form (M765) In contrast the large infested area in Chiayiconsists mostly of P nests (92 of colonies collectedFig 1c)
Genetic diversity
From one to five alleles were found at each microsatellitelocus (23 and 26 alleles in the Taoyuan and Chiayipopulations respectively Table 1) A total of sevenprivate alleles (ie unique to a single population) acrossall six microsatellite loci were found Tests for conformityof genotype proportions to Hardy-Weinberg expectationsrevealed only one significant deviation involving Sol-11 inthe Taoyuan polygyne population (Table 1) Score testsreveal that this significant departure resulted from adeficiency of heterozygotes
Analyses of the mtDNA sequence data revealed onlythree unique variants occur in Taiwan (designatedhaplotypes H5 H22 and H36 see Shoemaker andRoss 1996) all of which are commonly observed in fireants from the USA Individuals from the Taoyuanpopulation harbored either the H36 or H5 mtDNAhaplotype whereas all individuals from the Chiayi pop-ulation bear mtDNA haplotype H22 (Fig 1)
Insect Soc Vol 55 2008 Research article 57
Figure 1 (a) Map of Taiwan showing the two areas (Taoyuan and Chiayi counties) infested by Solenopsis invicta (indicated by shading) Histogramsshow the proportion of nests harboring each mtDNA haplotype Pie diagrams in the enlarged map of Taoyuan county (b) show the proportion of themonogyne and polygyne nests found in the central townships (smaller dotted-line circle) and in the surrounding townships (areas between larger andsmaller dotted-line circles) A pie diagram in the enlarged map of Chiayi county (c) indicates the proportion of two social forms in the sampling area(solid-line circle) Social form of each nest was determined by using a Gp-9 allele-specific assay and by examining the distribution of genotype arraysat six microsatellite loci within nests (see text)
Table 1 Genetic diversity measures within each social form from the two study populations of S invicta in Taiwan n denotes total number ofindividuals (nests) for a given social form in each area NA denotes observed number of alleles found at each locus from each populationHE =expected heterozygosity HO = observed heterozygosity
Locus Taoyuan Chiayi
Monogyne(n= 11)
Polygyne( n=19)
Monogyne( n=2)
Polygyne(n=23)
NA HE HO NA HE HO NA HE HO NA HE HO
Sol-6 1 0000 0000 2 0193 0105 3 0833 0800 4 0599 0434
Sol-11 4 0731 0545 4 0755 0526 2 0500 0700 4 0657 0652
Sol-20 5 0814 0818 5 0734 0631 3 0833 0800 4 0679 0782
Sol-42 4 0610 0363 4 0613 0473 2 0500 0450 5 0656 0608
Sol-49 4 0697 0634 5 0768 0842 3 0833 0850 5 0735 0782
Sol-55 3 0634 0454 3 0624 0789 2 0667 0850 4 0486 0608
Mean 35 0581 0469 38 0614 0561 25 0603 0742 43 0635 0645
58 CC Yang et al Fire ants in Taiwan
Within-nest relatedness
The estimates of relatedness among workers withinmonogyne nests from each area were consistent witheach nest headed by single queen mated to a single male(Taoyuan R=071870036 Chiayi R=071970038Fig 2) Relatedness among workers within polygynenests was significantly greater than zero but significantlylower (Plt0001) than the values for monogyne nests(Taoyuan R=011220006 Chiayi R=014440007Fig 2)
Population genetic differentiation
Exact tests based on six microsatellite loci revealedsignificant differentiation between Taoyuan and Chiayipopulations (Plt002) No significant differentiation wasobserved between sympatric populations of the two socialforms in the two areas (Pgt013) Exact tests for differ-entiation using mtDNA sequence data revealed signifi-cant differentiation occurs between the two geographicpopulations (Plt00001) as well as between the two socialforms in Taoyuan (Plt004) but not between the twosocial forms in Chiayi (only one mtDNA variant occurshere)
Nuclear and mtDNA variation was partitioned byestimating FST simultaneously at both levels using theanalysis of molecular variance approach (Excoffier et al 1992) Results of these analyses for the nuclear locishowed that only the between-population variance com-ponent is significant (Plt0000001 Fig 3a) providingadditional support to the above analyses suggesting thesetwo populations are genetically distinct For analysesinvolving mtDNA data both the between-population andbetween-social form variance components are significant(both Plt005 Fig 3b)
Bayesian simulations using STRUCTURE revealedthat individuals from the Taoyuan population represent a
single distinct genetic cluster suggesting no differentia-tion between sympatric social forms exists whether usingnuclear data only or nuclear and mtDNA combinedIdentical results were obtained for individuals from theChiayi population In contrast simulations incorporatingall individuals from the two areas gave posterior proba-bilities that strongly support the existence of two distinctgenetic clusters using dataset from the nuclear loci orcombination of the two classes of markers (K=2 Fig 4)The two clusters hypothesized by the Bayesian methodcorrespond roughly to geographic populations based onaverage membership coefficientsgt05 for one of the twoposterior clusters in each of these regions The firstcluster (indicated by lighter gray) is comprised largely ofindividuals from the Taoyuan population whereas thesecond cluster (indicated by the darker gray) is comprisedmostly of the individuals from Chiayi Identical clusteringresults were obtained when simulations were run incor-porating both the microsatellite and mtDNA data(Fig 4)
Figure 2 Average relatedness (95 confidence interval) valuesamong workers within nests of the polygyne and monogyne social formsfrom Taoyuan and Chiayi counties Taiwan
Figure 3 Partitioning of genetic variance in S invicta at six micro-satellite loci (a) and mtDNA (b) estimated using analysis of molecularvariance This hierarchical approach partitioned the total variancebetween infested regions (Taoyuan and Chiayi counties) between thetwo social forms within each infested region and within the social formsin each region The percentage of the total variance partitioned at eachlevel is shown Asterisks indicate statistically significant differentiationat a given level (a=005)
Figure 4 Assignment of individuals representing Solenopsis invictafrom both study populations to two genetic clusters inferred fromSTRUCTURE simulations (average membership coefficient Q) Eachvertical bar represents the individual membership coefficients dividedinto parts proportional to the individuals proposed ancestry in eachcluster
Insect Soc Vol 55 2008 Research article 59
Isolation by distance and tests for genetic bottlenecks
No correlation exists between pairwise FST or FST(1ndash FST)values and geographic distance for individuals of thepolygyne social form in either population (TaoyuanPgt022 Chiayi Pgt048) Further no significant iso-lation by distance for the monogyne nests from Taoyuanpopulation was found (Pgt065) We did not perform theMantel tests for the Chiayi monogyne population sinceonly two nests were collected
All tests revealed a significant excess of heterozygos-ity in Taoyuan under both models except the sign test(Table 3) A significant excess of heterozygosity wasfound in Chiayi for the infinite allele model but not thetwo phase model (Table 3) Combined with a mode shiftof allele frequency distribution (Table 3 Fig 5a) theTaoyuan population appears to have undergone a recentbottleneck In contrast the most abundant alleles in theChiayi population belong to the low frequency class(lt01 Fig 5b) resulting in a normal L-shape distribu-
tion We also compared NA (number of observed allelesTable 2) and HE (expected heterozygosity Table 2)among the South American USA and Taiwanesepopulations to determine whether a genetic signaturecharacteristic of a bottleneck was evident for the latterpopulations In order to minimize the effect of populationsize we randomly selected 30 individuals from the USAand South American dataset (Shoemaker et al 2006Ross et al 2007) Results showed that the NA and HE inTaiwan and the USA populations were consistentlylower than in South American fire ant populations Twopopulations in Taiwan had slightly lower NA than inUSA but had similar HE values
Additional findings
Analyses of individual genotypes revealed that 108 ofthe 190 workers sampled from polygyne nests in Taoyuanand 11 of 230 individuals from polygyne nests in Chiayi
Table 2 Genetic diversity measures within the two study populations of S invicta in Taiwan NA denotes observed number of alleles found at eachlocus from each population HE =expected heterozygosity HO = observed heterozygosity Numbers in parentheses represent the number of privatealleles occurring in only one of the two Taiwanese populations Genetic diversity measures for two population from the USA and South Americaalso are shown (see text)
Source of collection Population Sample size Mean no of alleles NA HE HO
TaiwanTaoyuan county 30 38 23 (2) 0609 0527Chiayi county 25 48 26 (5) 0636 0660
USAMississippi 30 57 34 0651 0559western Louisiana 30 58 35 0628 0607
South AmericaFormosa Argentina 30 158 195 0835 0842Corrientes Argentina 30 158 95 0836 0826
Figure 5 Distribution of allele frequencies at six microsatellite loci for ants (single individual per nest) from the Taoyuan (a) and Chiayi populations(b)
60 CC Yang et al Fire ants in Taiwan
harbored three distinct alleles whereas all of the individ-uals sampled from monogyne nests harbored at most twodistinct alleles These results indicate that a substantialnumber of polygyne fire ants are triploid a findingconsistent with an earlier study of fire ants from theUSA (Krieger et al 1999)
Discussion
We studied the colony genetic and population structure ofthe invasive fire ant S invicta in Taiwan by collecting andanalyzing microsatellite genotypic data and mtDNAsequence data of ants collected from two separateinfested areas Our genetic analyses revealed severalpatterns which are discussed in more detail below andcan be summarized as follows 1) Estimates of within nestrelatedness were significantly greater than zero for bothsocial forms from both populations 2) No significantisolation by distance was found among nests within eachsocial form from either population 3) SignificantmtDNA but no nuclear differentiation occurs betweensympatric social forms in Taoyuan (no differentiationbetween forms at either genome is evident in Chiayi) 4)Molecular signatures of genetic bottlenecks associatedwith recent introductions are evident in both populations5) The two sampled populations Taoyuan and Chiayi arehighly genetically differentiated at both the nuclear ormtDNA genomes In addition to these general findingswe found numerous triploid individuals in ants of thepolygyne (P) but not the monogyne (M) form anddistinct geographic distribution of each of the two socialforms between the two populations
Colony social organization and patterns of intracolonyworker relatedness
The estimate of relatedness among workers withinmonogyne colonies was not significantly different from075 the predicted value for a nest headed by one singly-mated queen These data also are consistent with datafrom earlier studies of monogyne S invicta colonies inboth the USA (Ross and Fletcher 1985b) and SouthAmerica (Ross et al 1997) and suggest that the socialand mating biology of M queens is virtually invariantwith M nests representing closed societies headed by onesingly-mated reproductive queen
In contrast colony genetic structure of P social formdoes differ between native South America and intro-duced populations in the USA (Ross et al 1996) Pnests of S invicta in the native range contain relativelyfew queens which are close relatives and relatednessamong nestmate workers is significantly greater thanzero whereas P nests in the USA contain highnumbers of unrelated queens and relatedness amongnestmate workers is not significantly greater than zeroThese differences in queen number and relatedness
between the two ranges are consistent with predictionsbased on differences in the population densities of antsin these two areas (Ross et al 1996) which aresubstantially higher in the USA compared to SouthAmerica (Porter et al 1997) One consequence of thehigher population densities in the USA is that nestingsites are more often saturated such that opportunitiesfor independent founding by queens are more limitedin the introduced range (Ross et al 1996) Theseconstraints on independent founding lead to increasedselection pressure on queens to seek adoption intoexisting colonies and on resident workers of colonies toaccept such queens As queen number within thesenests increases relatedness among nestmate workers isexpected to decrease
Relatedness among workers within P nests in Taiwanis small yet significantly greater than zero and intermedi-ate to the values observed for native and USApopulations The social organization of P nests in Taiwanlikely reflects the fact that nesting sites are much lesssaturated compared to sites in the USA since thepopulation densities of ants in Taiwan currently are muchlower than those in the USA However while such adifference between the two introduced ranges are evidentnow it is unlikely to persist especially if S invictacontinues to spread in Taiwan so that habitats becomesaturated If S invicta continues to spread so that habitatsbecome more saturated future studies of the socialevolution of P populations in Taiwan could shed addi-tional light on whether such a change in habitat avail-ability does in fact lead to a shift in social organizationtowards a higher queen number within nests and lowerrelatedness among nestmates
Patterns of isolation by distance
The lack of isolation by distance among colonies of the Mform within Taoyuan was not unexpected given that mostM queens take part in aerial mating flights oftendispersing several kilometers or more before landingand attempting to found a new nest (Toom et al 1976Ross and Fletcher 1985b Ross et al 1999) Such long-distance dispersal is predicted to erase any signature ofnuclear differentiation among closely spaced nests
The lack of detectable isolation by distance within theP form at both studied sites was somewhat unexpectedconsidering the reproductive habits of P queens whichoften mate in or near their natal nests and form newcolonies by budding or by entering into nearby existingnests (Shoemaker and Ross 1996 Ross et al 1997 Rossand Shoemaker 1997) One explanation for the lack ofdetectable isolation by distance is simply that malesdisperse rather long distances to mate with P queensUnder this scenario one would predict that long distancemale-mediated gene flow would result in homogenizationof allele frequencies at nuclear loci yet significantmicrogeographic mtDNA structure would remain espe-
Insect Soc Vol 55 2008 Research article 61
cially if queen dispersal is limited However no suchpattern was found Significant isolation by distance wasabsent for both the nuclear and mtDNA markers Anadditional explanation for these results is that thesignature of isolation by distance predicted to emergesolely from the natural dispersal biology of these ants isconfounded by the fact that these ants are frequentlyinadvertently transported relatively great distances byhumans Such frequent inadvertent human transport maybe a common characteristic of invasive ants found in closeproximity to human habitats and thus play a role inshaping the genetic structure in introduced areas Finallyit is possible that the lack of a detectable signal of isolationby distance is due to the extremely recent introduction offire ants into Taiwan with insufficient time having passedfor populations to achieve migrationdrift equilibrium
Gene flow between sympatric social forms
The pronounced mtDNA differentiation between sym-patric social forms in Taoyuan is consistent with resultsfrom earlier studies in both South America and the USAshowing that interform gene flow mediated by queens israre or absent Queens of each social form rarely succeedin becoming reproductives in colonies of the alternateform as a result of behavioral decisions (attributed largelyto differences in genotypes of queens at the single geneGp-9) made by both queens and workers (Shoemaker andRoss 1996 Ross and Shoemaker 1997 Keller and Ross1998 DeHeer et al 1999 Goodisman et al 2000 Rossand Keller 2002) The contrasting weak or nonexistentinterform divergence at nuclear genes also is consistentwith studies of S invicta in the USA showing that queensof the P form predominantly mate with males of the Mform where the two forms co-occur (Ross and Shoe-maker 1993 Ross and Keller 1995 Shoemaker andRoss 1996) In the USA most males in P colonies aresterile diploid males which means that P queens relylargely on immigrant M males for mating opportunities(Ross and Fletcher 1985a Ross and Shoemaker 1993)Such interform matings act as a homogenizing force onlyfor nuclear markers The similar frequency of triploidworkers in P nests in Taiwan and the USA suggestsmany diploid males also occur in Taiwan as well (Rosset al 1993 1999 Krieger and Keller 1997 Krieger et al 1999) which means that P queens here must also matewith M males The finding of contrasting patterns ofdifferentiation between the two genomes parallels resultsfrom earlier studies involving S invicta in both the USAand South America (Shoemaker and Ross 1996 Rosset al 1997 Shoemaker et al 2006) One final caveathowever is that we cannot completely rule out thepossibility that the lack of detectable nuclear differ-entiation is due to insufficient time having passed fordivergence to occur despite restricted gene flow betweenthe two forms
Understanding the distributions of the two socialforms in the two infested areas in Taiwan each of whichexhibits rather distinct dispersal strategies is importantfor developing effective control strategies of fire ants inthese infested areas (Porter et al 1991) The distribu-tional pattern of social forms in Taoyuan suggests that theexpansion of this population has involved persistentoutward movement of ants from the core infestationcoupled with frequent long distance movement of Mreproductives into outlying areas (Fig 1b) Based onthese results we recommend that bait broadcasting andsurveillance efforts in Taoyuan should include not onlythe currently infested areas but also should include anarea extending well beyond the infested areas In contrastto the distributional pattern of social forms in Taoyuanthe large infested area in Chiayi consists mostly of P nests(92 of colonies collected Fig 1c) However despitethis difference we suggest a similar eradication strategyshould be employed This is because even thoughcomprehensive bait broadcasting within the entire areamay be more effective than for Taoyuan because of thelow queen vagility and relatively slow rate of reinvasionby budding of polygynous queens (Porter et al 1991)some M colonies do occur in Chiayi and there also ispotential for additional M colonies to arise locally fromthe P social form a possibility that is especially morelikely in areas of low density (Ross and Shoemaker 1997DeHeer and Tschinkel 1998 DeHeer et al 1999)
Genetic bottlenecks in Taoyuan and Chiayi populations
Simulations and statistical tests using the program BOT-TLENECK provide evidence of a significant geneticbottleneck associated with the introduction of fire antsinto Taoyuan with only one exception (Table 3) How-ever this single exception is based on a sign test whichhas relatively low statistical power when the number ofloci is below 20 such as our study (Cornuet and Luikart1996) In Chiayi significant excess in heterozygosity usingthe infinite alleles model (IAM) was observed for all testsbut not for the two-phase model (TPM Table 3) BecauseLuikart and Cornuet (1998) suggest that implementingthe test under the IAM model may be more reliable thanother models these results coupled with finding thatallelic frequencies fit a normal L-shaped distributionprovide evidence for a genetic bottleneck Interestinglyestimates of NA and HE (Table 2) are remarkably similarfor introduced populations from Taiwan and the USAand considerably lower than those from the native SouthAmerican range While the geographic origin of ants inTaiwan is unknown the similar genetic diversity measuresbetween the introduced ranges raises the issue that theintroduction of ants into Taiwan has left only a modestfootprint on patterns of diversity if these ants originatedfrom the USA
62 CC Yang et al Fire ants in Taiwan
Genetic differentiation between Taoyuan and Chiayipopulations
All analyses of population differentiation using themicrosatellite and mtDNA data indicate that the twogeographic populations are strongly differentiated andthus likely originated from separate invasions Whilemultiple lines of evidence support this notion thestrongest evidence is the confinement each of the threemtDNA variants to only one of the two geographicpopulations Interestingly all three variants found inTaiwan are the same three variants commonly found inants from the USA (Shoemaker et al 2006) China(GenBank accession numbers DQ831670-DQ831672)and Australia (CCY and DDS unpublished results) Theoccurrence of these three variants in all recently invadedareas suggests that ants from Taiwan could have origi-nated from any of these other areas We currently aregenerating the necessary baseline data for ants from all ofthese introduced regions as well as from populations intheir native range to infer the source population andinvasion history of ants in Taiwan and these other areas
From the perspective of managing fire ants in Taiwantreating each infested area as a separate management unitwould appear more reasonable and means that scientistsprobably should focus on separate plans for eliminationof two isolated populations (Hampton et al 2004) Onthe other hand because these two areas are at high risk ofconnectivity via long distance human-mediated dispersalcare must be taken to prevent reinvasion into these areaseven after successful eradication Stringent inter-countyquarantine measures such as movement control shouldbe established to monitor the risk of transportation of fireant-infested goods to prevent jump dispersal between thetwo areas and decrease probability of successful erad-ication (Drees and Gold 2003)
Acknowledgments
The authors would like to thank Chung-Chi Lin for providing most ofthe specimens for this study including the first identified samples inTaoyuan and Chiayi counties We are particularly grateful to Hwei-yuChang Hon-Tsen Yu and Chia-Hung Hsieh for helpful discussions andcomments on earlier versions of this manuscript Comments by twoanonymous reviewers greatly improved this manuscript This research
was financially supported by grants from Academia Sinica Taiwan Theuse of trade firm or corporation names in this publication is for theinformation and convenience of the reader Such use does notconstitute an official endorsement or approval by the United StatesDepartment of Agriculture or the Agricultural Research Service of anyproduct or service to the exclusion of others that may be suitable
References
Ahrens ME Ross KG and Shoemaker DD 2005 Phylogeographicstructure of the fire ant Solenopsis invicta in its native SouthAmerican range Roles of natural barriers and habitat connectivityEvolution 59 1733ndash1743
Allen CR Demarais S and Lutz RS 1994 Red imported fire antimpact on wildlife an overview Texas J Sci 46 51ndash59
Allen CR Lutz RS and Demarais S 1998 Ecological effects of theinvasive nonindigenous ant Solenopsis invicta on native verte-brates The wheels on the bus Transactions of the North AmericanWildlife and Natural Resources Conference 63 56 ndash65
Allen CR Willey RD Myers PE Horton PM and Buffa J 2000Impact of red imported fire ant infestation on northern bobwhitequail abundance trends in southeastern United States J AgrUrban Entomol 17 43ndash51
Allen CR Forys EA Rice KG and Wojcik DP 2001 Effects of fireants (Hymenoptera Formicidae) on hatching turtles and preva-lence of fire ants on sea turtle nesting beaches in Florida FloridaEntomol 84 250ndash253
Bouwma AM Ahrens ME DeHeer CJ and Shoemaker DD 2006Distribution and prevalence of Wolbachia in introduced popula-tions of the fire ant Solenopsis invicta Insect Mol Biol 15 89ndash93
Briano JA Calcaterra LA Vander Meer R Valles SM and LivoreJP 2006 New survey for the fire ant microsporidia Vairimorphainvictae and Thelohania solenopsae in southern South Americawith observations on their field persistence and prevalence of dualinfections Environ Entomol 35 1358ndash1365
Buckley A 1999 Fire ants in California Am Bee J 139 88Carroll CR and Hoffman CA 2000 The pervasive ecological effects
of invasive species Exotic and native fire ants In Invertebrates asWeb-Masters in Ecosystems (Coleman DC and Hendrix PF Eds)Wallingford Oxon UK pp 221ndash232
Carruthers RI 2003 Invasive species research in the United StatesDepartment of Agriculture ndash Agricultural Research Service PestManag Sci 59 827ndash834
Chen JSC Shen CH and Lee HJ 2006 Monogynous and polygynousred imported fire ants Solenopsis invicta Buren (HymenopteraFormicidae) in Taiwan Environ Entomol 35 167ndash172
Cornuet JM and Luikart G 1996 Description and power analysis oftwo tests for detecting recent population bottlenecks from allelefrequency data Genetics 144 2001ndash2014
Davis LR Vander Meer RK and Porter SD 2001 Red imported fireants expand their range across the West Indies Florida Entomol84 735ndash736
DeHeer CJ and Tschinkel WR 1998 The success of alternativereproductive tactics in monogyne Solenopsis invicta significancefor transitions in social organization Behav Ecol 9 130ndash135
DeHeer CJ Goodisman MAD and Ross KG 1999 Queen dispersalstrategies in the multiple-queen form of the fire ant Solenopsisinvicta Am Nat 153 660ndash675
Dieringer D and Schlccediltterer C 2002 Microsatellite analyser (MSA) aplatform independent analysis tool for large microsatellite datasets Mol Ecol Notes 3 167ndash169
Drake JA Mooney HA Castri F Groves RH Kruger MRejmnek M and Williamson M 1989 Biological Invasions AGlobal Perspective Chichester UK 550 pp
Drees BM and Vinson SB 1990 Comparison of the control ofmonogynous and polygynous forms of the red imported fire ant(Hymenoptera Formicidae) with a chloripyrifos mound drench JEntomol Sci 25 317ndash324
Table 3 Tests for genetic bottlenecks using the infinite allele model(IAM) and two-phase model (TPM 90 stepwise mutation model and10 multistep mutation model [Garza and Williamson 2001])implemented in the program BOTTLENECK (a=005)Significance at given statistic level
Population Simulatedmodel
Significancetest
Standardizeddifferences test
Wilcoxontest
Taoyuan IAM 0148 0003 0015TPM 0186 0035 0023
Chiayi IAM 0037 0022 0007TPM 0521 0293 0281
Insect Soc Vol 55 2008 Research article 63
Drees BM and Gold RE 2003 Development of integrated pestmanagement programs for the red imported fire ant (Hymeno-ACHTUNGTRENNUNGptera Formicidae) J Entomol Sci 38 170ndash180
Eubanks MD 2001 Estimates of the direct and indirect effects of redimported fire ants on biological control in field crops Biol Control21 35 ndash43
Evanno G Regnaut S and Goudet J 2005 Detecting the number ofclusters of individuals using the software STRUCTURE asimulation study Mol Ecol 14 2611ndash2620
Excoffier L Smouse PE and Quattro JM 1992 Analysis of molecularvariance inferred from metric distances among DNA haplotypesApplication to human mitochondrial DNA restriction dataGenetics 131 479ndash491
Forys EA Quistorff A and Allen CR 2001 Potential fire ant(Hymenoptera Formicidae) impact on the endangered Schausswallowtail (Lepidoptera Papilionidae) Florida Entomol 84 254ndash258
Garza JC and Williamson EG 2001 Detection of reduction inpopulation size using data from microsatellite loci Mol Ecol 10 305ndash318
Giraud T Pedersen JS and Keller L 2002 Evolution of supercoloniesthe Argentine ants of southern Europe Proc Natl Acad Sci U SA 99 6075ndash6079
Giuliano WM Allen CR Lutz RS and Demarais S 1996 Effects ofred imported fire ants on northern bobwhite chicks J WildlManage 60 309ndash313
Glancey BM Craig CH Stringer CE and Bishop PM 1973Multiple fertile queens in colonies of the imported fire antSolenopsis invicta J Georgia Entomol Soc 8 327ndash328
Goodisman MAD DeHeer CJ and Ross KG 2000 Mating behaviorof queens from multiple-queen colonies of Solenopsis invicta JInsect Behav 13 455ndash468
Goodnight KF and Queller DC 1999 Computer software forperforming likelihood tests of pedigree relationship using geneticmarkers Mol Ecol 8 1231ndash1234
Gotelli NJ and Arnett AE 2000 Biogeographic effects of red fire antinvasion Ecol Lett 3 257ndash261
Gotzek D and Ross KG 2007 Genetic regulation of colony socialorganization in fire ants An integrative overview Q Rev Biol 82 201ndash226
Goudet J 2001 FSTAT A Program to Estimate and Test GeneDiversities and Fixation Indices Version 293 http wwwunilch izeasoftwares fstathtml
Greenberg L Fletcher DJC and Vinson SB 1985 Differences inworker size and mound distribution in monogynous and polygy-nous colonies of the fire ant Solenopsis invicta Buren J KansasEntomol Soc 58 9ndash18
Hampton JO Spencer PBS Alpers DL Twigg LE WoolnoughAP Doust J Higgs T and Pluske J 2004 Molecular techniqueswildlife management and the importance of genetic populationstructure and dispersal a case study with feral pigs J Appl Ecol41 735ndash743
Henshaw MT Kunzmann N Vanderwoude C Sanetra M andCrozier RH 2005 Population genetics and history of theintroduced fire ant Solenopsis invicta Buren (HymenopteraFormicidae) in Australia Aus J Entomol 44 37ndash44
Hccedillldobler B and Wilson EO 1990 The Ants The Belknap Press ofHarvard University Press Cambridge Massachusetts 732 pp
Huang TC Chou YC and Chou HC 2004 The infestation andcontrol of the red imported fire ant in Taiwan In Proceedingsof the Symposium on the Control of the Red Imported Fire AntBureau of Animal and Plant Health Inspection and QuarantineCouncil of Agriculture Executive Yuan Taipei Taiwan pp1 ndash 13
Jaquiry J Vogel V and Keller L 2005 Multilevel genetic analyses oftwo European supercolonies of the Argentine ant Linepithemahumile Mol Ecol 14 589ndash598
Jeyaprakash A and Hoy MA 2000 Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropods Insect Mol Biol 9 393ndash405
Jouvenaz DP 1990 Approaches to biological control of fire ants in theUnited States In Applied Myrmecology A World Perspective(Vander Meer RK Jaffe K and Cedeno A Eds) Westview PressBoulder Colorado pp 620ndash627
Kaspari M 2000 Do imported fire ants impact canopy arthropodsEvidence from simple arboreal pitfall traps Southwestern Nat 45 118ndash122
Keller L and Ross KG 1998 Selfish genes a green beard in the red fireant Nature 394 573ndash575
Kemp SF deShazo RD Moffitt JE Williams DF and Buhner WA2000 Expanding habitat of the imported fire ant (Solenopsisinvicta) A public health concern J Allergy Clin Immunol 105 683ndash691
Kopachena JG Buckley AJ and Potts GA 2000 Effects of the redimported fire ant (Solenopsis invicta) on reproductive success ofbarn swallows (Hirundo rustica) in northeast Texas SouthwesternNat 45 477ndash482
Krieger MJB and Keller L 1997 Polymorphism at dinucleotidemicrosatellite loci in fire ant Solenopsis invicta populations MolEcol 6 997ndash999
Krieger MJB Ross KG Chang CWY and Keller L 1999Frequency and origin of triploidy in the fire ant Solenopsis invictaHeredity 82 142ndash150
Krieger MJB and Ross KG 2002 Identification of a major generegulating complex social behavior Science 295 328ndash332
Lee CE 2002 Evolutionary genetics of invasive species Trends EcolEvol 17 386ndash391
Lofgren CS Banks WA and Glancey BM 1975 Biology and controlof imported fire ants Annu Rev Entomol 20 1ndash30
Lofgren CS 1986 History of imported fire ants in the United States InFire Ants and Leaf Cutting Ants Biology and Management(Lofgren CS and Vander Meer RK Eds) Westview PressBoulder Colorado pp 36ndash49
Luikart G Allendorf FW Cornuet JM and Sherwin WB 1998Distortion of allele frequency distributions provides a test forrecent population bottlenecks J Hered 89 238ndash247
Luikart G and Cornuet JM 1998 Empirical evaluation of a test foridentifying recently bottlenecked populations from allele frequen-cy data Conserv Biol 12 228ndash237
MacKay WP and Fagerlund R 1997 Range expansion of the redimported fire ant Solenopsis invicta Buren (Hymenoptera For-micidae) into New Mexico and extreme western Texas ProcEntomol Soc Wash 99 757ndash758
Mank JE and Avise JC 2004 Individual organisms as units of analysisBayesian-clustering alternatives in population genetics Genet Res84 135ndash143
May GE Gelembiuk GW Panov VE Orlova MI and Lee CE2006 Molecular ecology of zebra mussel invasions Mol Ecol 15 1021ndash1031
McCubbin KI and Weiner JM 2002 Fire ants in Australia A newmedical and ecological hazard Med J Aus 176 518ndash519
Morrison LW 2000 Mechanisms of interspecific competition amongan invasive and two native fire ants Oikos 90 238ndash252
Morrison LW 2002 Long-term impacts of an arthropod-communityinvasion by the imported fire ant Solenopsis invicta Ecology 83 2337ndash2345
Nattrass R and Vanderwoude C 2001 A preliminary investigation ofthe ecological effects of red imported fire ants (Solenopsis invicta)in Brisbane Ecol Manag Resto 2 220ndash223
Nei M 1972 Genetic distance between populations Am Nat 106 283ndash292
Orr MR 1996 Host manipulation by Wolbachia is a neutral trait withinsingle populations Anim Behav 51 1183ndash1185
Patterson RS 1994 Biological control of introduced ant species InExotic Ants Biology Impact and Control of Introduced Species(Williams DF Ed) Westview Press Boulder Colorado pp 293ndash308
Piry S Luikart G and Cornuet JM 1999 BOTTLENECK acomputer program for detecting recent reductions in the effectivepopulation size using allele frequency data J Hered 90 502ndash503
64 CC Yang et al Fire ants in Taiwan
Porter SD and Savignano DA 1990 Invasion of polygyne fire antsdecimates native ants and disrupts arthropod community Ecology7 2095ndash2106
Porter SD Bhatkar A Mulder R Vinson SB and Clair DJ 1991Distribution and density of polygyne fire ants (HymenopteraFormicidae) in Texas J Econ Entomol 84 866ndash874
Porter SD Fowler HG and MacKay WP 1992 Fire ant mounddensities in the United States and Brazil (Hymenoptera Formi-cidae) J Econ Entomol 85 1154ndash1161
Porter SD Williams DF Patterson RS and Fowler HG 1997Intercontinental differences in the abundance of Solenopsis fireants (Hymenoptera Formicidae) Escape from natural enemiesEnviron Entomol 26 373ndash384
Pritchard JK Stephens M and Donnelly P 2000 Inference ofpopulation structure using multilocus genotype data Genetics155 945ndash959
Raymond M and Rousset F 1995a An exact test for populationdifferentiation Evolution 49 1280ndash1283
Raymond M and Rousset F 1995b GENEPOP (version 12)population genetics software for exact tests and ecumenicism JHered 86 248ndash249
Reuter M Pedersen JS and Keller L 2005 Loss of Wolbachiainfection during colonisation in the invasive Argentine ant Line-pithema humile Heredity 94 364ndash369
Ross KG and Fletcher DJC 1985a Genetic origin of male diploidy inthe fire ant Solenopsis invicta (Hymenoptera Formicidae) and itsevolutionary significance Evolution 39 888ndash903
Ross KG and Fletcher DJC 1985b Comparative study of genetic andsocial structure in two forms of the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 17 349ndash356
Ross KG and Fletcher DJC 1986 Diploid male productionndashasignificant colony mortality factor in the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 19 283ndash291
Ross KG and Shoemaker DD 1993 An unusual pattern of gene flowbetween the two social forms of the fire ant Solenopsis invictaEvolution 47 1595ndash1605
Ross KG Vargo EL Keller L and Trager JC 1993 Effect of afounder event on variation in the genetic sex-determining system ofthe fire ant Solenopsis invicta Genetics 135 843ndash854
Ross KG and Keller L 1995 Joint influence of gene flow and selectionon a reproductively important genetic polymorphism in the fire antSolenopsis invicta Am Nat 146 325ndash348
Ross KG Vargo EL and Keller L 1996 Social evolution in a newenvironment The case of introduced fire ants Proc Natl AcadSci U S A 93 3021ndash3025
Ross KG 1997 Multilocus evolution in fire ants Effects of selectiongene flow and recombination Genetics 145 961ndash974
Ross KG Krieger MJB Shoemaker DD Vargo EL and Keller L1997 Hierarchical analysis of genetic structure in native fire antpopulations Results from three classes of molecular markersGenetics 147 643ndash655
Ross KG and Shoemaker DD 1997 Nuclear and mitochondrialgenetic structure in two social forms of the fire ant Solenopsisinvicta Insights into transitions to an alternate social organizationHeredity 78 590ndash602
Ross KG and Keller L 1998 Genetic control of social organization inan ant Proc Natl Acad Sci U S A 95 14232ndash14237
Ross KG Shoemaker DD Krieger MJ DeHeer CJ and Keller L1999 Assessing genetic structure with multiple classes of molecularmarkers A case study involving the introduced fire ant Solenopsisinvicta Mol Biol Evol 16 525ndash543
Ross KG and Keller L 2002 Experimental conversion of colony socialorganization by manipulation of worker genotype composition infire ants (Solenopsis invicta) Behav Ecol Sociobiol 51 287ndash295
Ross KG Krieger MJB and Shoemaker DD 2003 Alternativegenetic foundations and evolutionary routes for a key socialpolymorphism in fire ants Genetics 165 1853ndash1867
Ross KG Krieger MJB Keller L and Shoemaker DD 2007Genetic variation and structure in native populations of the fire antSolenopsis invicta Evolutionary and demographic implicationsBiol J Linnean Soc 92 541ndash560
Rousset F 1997 Genetic differentiation and estimation of gene flowfrom F-statistics under isolation by distance Genetics 145 1219ndash1228
Sax DF Stachowicz JJ and Gaines SD 2005 Species InvasionsInsights into Ecology Evolution and Biogeography SinauerAssociates Sunderland Massachussetts 495 pp
Shoemaker DD and Ross KG 1996 Effects of social organization ongene flow in the fire ant Solenopsis invicta Nature 383 613ndash616
Shoemaker DD Ross KG Keller L Vargo EL and Werren JH2000 Wolbachia infections in native and introduced populations offire ants (Solenopsis spp) Insect Mol Biol 9 661ndash673
Shoemaker DD Ahrens M Sheill L Mescher M Keller L and RossKG 2003 Distribution and prevalence of Wolbachia infections innative populations of the fire ant Solenopsis invicta (HymenopteraFormicidae) Environ Entomol 32 1329ndash1336
Shoemaker DD DeHeer CJ Krieger MJB and Ross KG 2006Population genetics of the invasive fire ant Solenopsis invicta(Hymenoptera Formicidae) in the USA Ann Entomol Soc Am99 1213ndash1233
Simon C Frati F Beckenbach A Crespi B Liu H and Flook P 1994Evolution weighting and phylogenetic utility of mitochondrialgene sequences and a compilation of conserved polymerase chainreaction primers Ann Entomol Soc Am 87 651ndash701
Toom PM Cupp E Johnson CP and Griffin I 1976 Utilization ofbody reserves for minim brood development by queens of theimported fire ant Solenopsis invicta J Insect Physiol 22 217ndash220
Tsutsui ND and Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepethema humile) in its nativeand introduced ranges Evolution 55 976ndash985
Tsutsui ND Kauppinen SN Oyafuso AF and Grosberg RK 2003The distribution and evolutionary history of Wolbachia infection innative and introduced populations of the invasive argentine ant(Linepithema humile) Mol Ecol 12 3057ndash3068
Valles SM and Porter SD 2003 Identification of polygyne andmonogyne fire ant colonies (Solenopsis invicta) by multiplex PCRof Gp-9 alleles Insect Soc 50 199ndash200
Valles SM Oi DH Briano JA and Williams DF 2004 Simultaneousdetection of Vairimorpha invictae (Microsporidia Burenellidae)and Thelohania solenopsae (Microsporidia Thelohaniidae) in fireants by PCR Florida Entomol 87 85ndash87
Vander Meer R K and Porter SD 2001 Fate of newly-mated queensintroduced into monogyne and polygyne Solenopsis invicta (Hy-menoptera Formicidae) colonies Ann Entomol Soc Am 94 289ndash297
Vander Meer RK Jaffe K and Cedeno A 1990 Applied Myrmecol-ogy A World Perspective Westview Press Boulder CO 741 pp
Vinson SB 1994 Impact and invasion of Solenopsis invicta (Buren) onnative food webs In Exotic Ants Biology Impact and Control ofIntroduced Species (Williams DF Ed) Westview Press BoulderColorado pp 293ndash308
Vinson SB 1997 Invasion of the red imported fire ant (HymenopteraFormicidae) Spread biology and impact Am Entomol 43 23ndash29
Williams DF and Porter SD 1994 Fire ant control Science 264 1653ndash1653
Wojcik D Allen CR Brenner RJ Forys EA Jouvenaz D and LutzRS 2001 Red imported fire ants Impact on biodiversity AmEntomol 47 16ndash23
Xia X and Xie Z 2001 DAMBE Data analysis in molecular biologyand evolution J Hered 92 371ndash373
Insect Soc Vol 55 2008 Research article 65

This study has several objectives First we wished tolearn whether the current distribution of fire ants in thesetwo areas is the result of a single or separate introductionsinto Taiwan The finding of multiple introductions mayindicate that the island of Taiwan is at high risk ofexperiencing additional introductions Second we aimedto infer whether one social form was possibly derivedfrom local populations of the alternate social form as wellas whether substantial gene flow occurs between sym-patric populations of each social form Lastly we utilizedour genetic data to assess whether a genetic signaturecharacteristic of a recent population bottleneck is evidentin either of two areas infested by S invicta While dataexamining the alternative colony genetic structure andassociated genetic differences between native and intro-duced fire ant populations have been proposed for theperiod of several decades after their first introduction tothe USA (Ross and Fletcher 1985b Ross et al 1996Ross et al 1999) the more recent introduction of fireants into Taiwan provides a novel opportunity to study thegenetic changes characteristics of invasiveness of thisspecies over an extremely short time period particularlyin the initial 5 ndash10 years after the original introduction
Materials and methods
Source of ant samples
Ants representing both social forms were collected from two locationsTaoyuan county and Chiayi county in Taiwan (these two locations areseparated by ca 250 km) Initially the social form of S invicta wasdetermined by visual inspection of the average size of the workers theheight of the mounds and the number of queens (Greenberg et al1985) Based on the preliminary determinations of social form in thefield adult workers were collected from nests of each putative socialform separated by distances of 30 ndash5000 (polygyne) and 600ndash2000 m(monogyne) (Fig 1) In total we sampled 30 colonies from Taoyuancounty and 25 colonies from the Chiayi county Collected workers werestored in 95 alcohol and frozen at -20 8C
DNA extractions and determination of social form
Two separate sets of total genomic DNA were extracted fromindividuals sampled from each nest For the first set 10ndash15 workerswere pooled and extracted in bulk using the Viogene DNA extractionkit (Viogene Inc Taipei Taiwan) These bulk-extracted samples wereused in Gp-9 assays for social form determination (Valles and Porter2003) For the second set total genomic DNA was extracted from 10individual workers from each sampled nest These samples were used assource DNA for microsatellite genotyping and mtDNA sequencing(single individual per nest see below) Inspection of the distribution ofmultilocus genotypes from 10 individuals per sampled nest provides apowerful and direct approach for determining the social form of eachnest
Microsatellite analyses
We genotyped 10 workers from each colony representing 550individuals in total at six dinucleotide-repeat microsatellite loci(Table 1) previously developed for S invicta (Table 1 Krieger andKeller 1997) Amplification of the single locus Sol-20 was performed in
10-ml reactions with 30 ndash100 ng DNA 1 ml of 10X Super-Therm GoldPCR buffer 1 ml of 25 mM dNTPs 03 ml of a 10 mM FAM-labeledforward primer 03 ml of a 10 mM unlabeled reverse primer and 015 Uof Super-Therm Gold Hot-start Taq DNA polymerase (AppliedBiosystems) The remaining five loci were amplified in two separatemultiplex PCR reactions In the first reaction Sol-11 Sol-42 and Sol-49were amplified simultaneously in a single tube Reaction cocktails andconditions for Sol-11 Sol-42 and Sol-49 were identical to thosedescribed earlier (Ross et al 2003) except forward primers werelabeled with FAM In the second multiplex reaction Sol-6 and Sol-55were co-amplified in a single 20-ml reaction with 30ndash100 ng DNA 24 mlof 10X Super-Therm Gold PCR buffer 16 ml of 25 mM dNTPs 05 mlof a 10 mM FAM-labeled Sol-6 forward primer 05 ml of a 10 mMunlabeled Sol-6 reverse primer 06 ml of a 10 mM FAM-labeled Sol-55forward primer 06 ml of a 10 mM unlabeled Sol-55 reverse primer and1U of Super-Therm Gold Hot-start Taq DNA polymerase PCRamplifications were carried out on an ABI 9700 thermal cycler(Applied Biosystems) using the following profile initial denaturationat 95 8C for 10 min 30 cycles of 94 8C for 1 min 58 8C for 1 min and72 8C for 1 min a final extension of 72 8C for 30 min All PCR productswere visualized on an ABI 3100 genetic analyzer using laser detectionand scoring of genotypes was performed using the software GENES-CAN 312 (Applied Biosystems)
MtDNA sequencing
A 920-bp fragment of the mitochondrial genome that includes portionsof the cytochrome oxidase subunit I (COI) and subunit II (COII) geneswas amplified from a single individual per nest using the primers C1-J-2195 (Simon et al 1994) and DDS-COII-4 (Ross and Shoemaker1997) PCR reactions were carried out using identical reaction cocktailsand thermal cycling profiles described in Ahrens et al (2005) MtDNAamplicons were purified and used directly in standard fluorescent cycle-sequencing PCR reactions (ABI Prism Big Dye terminator chemistryApplied Biosystems) Sequencing reactions were subsequently purifiedand run on an ABI 3730 sequencer All mtDNA amplicons weresequenced in both directions and corresponding sequence pairs wereedited and combined into a single consensus sequence file Allsequences were then aligned by eye using previously published Sinvicta mtDNA sequence obtained from GenBank (AY950762)
Genetic analyses
The number of alleles allele frequencies observed heterozygosity andexpected heterozygosity at each microsatellite locus for both popula-tions were calculated using the program MSA 300 (Dieringer andSchlccediltterer 2002) Genotype proportions in the two populations weretested for conformity to Hardy-Weinberg expectations (HWE) usingexact tests implemented in the program GENEPOP 33 (Raymond andRousset 1995b) The program DAMBE (Xia and Xie 2001) was usedto determine haplotype counts from the mtDNA sequence dataGenetic data from only one individual per nest was used for the aboveanalyses
Relatedness of nestmate workers in each form from both pop-ulations was estimated from genotype distributions at the six micro-satellite loci using the program RELATEDNESS 508 (Goodnight andQueller 1999) Triploid individuals were discarded when performingthe relatedness calculations (see below) The reference population forrelatedness estimation within social forms was defined as either allmonogyne or polygyne nests from a given population For all related-ness calculations nests were weighted equally and standard errors andtheir derivative 95 confidence limits were obtained by jackknifingover nests
Population genetic differentiation was examined in several waysPairwise FST values based on microsatellite and mtDNA sequence datawere calculated between sympatric populations of each social formbetween the two separate geographic populations and between nestpairs within in each area using the program FSTAT (Goudet 2001)Significance of the genetic differentiation measured between pairs of
56 CC Yang et al Fire ants in Taiwan
populations was determined by means of exact tests implemented inGENEPOP 33 (Raymond and Rousset 1995a) Genetic distancevalues (Neis D) also were calculated to further assess divergenceamong populations (Nei 1972)
FST values were also estimated simultaneously at two levels (socialform geographic population) using the hierarchical analysis ofmolecular variance procedure in ARLEQUIN (Excoffier et al1992) Statistical significance of the differentiation at each level wasestablished by means of 20000 data permutations
We tested for correlations between pairwise FST values [FST(1ndashFST)] and the natural logarithm of pairwise geographic distances amongnests from each social form in the two areas except for monogynousindividuals from Chiayi at all six microsatellite loci using the Manteltest in GENEPOP 33 (Raymond and Rousset 1995b) to determine ifthere was a significant positive correlation between genetic differ-entiation and geographic distance indicative of isolation by distance
We also used an individual-based Bayesian method to statisticallyrecognize distinct genetic clusters of individuals and to infer levels ofpopulation admixture from individual multilocus genotypic data(Pritchard et al 2000 Mank and Avise 2004) The method assumesa model with K populations each with characteristic allele frequenciesthat are estimated while individuals are probabilistically assigned toeach population Prior information such as the geographic location ofsamples or colony social form also can be incorporated into the modelto assist in clustering The program STRUCTURE (Pritchard et al2000 URL httppritchbsduchicagoedu) was used to explore theparameter space of each model using Markov chain Monte Carlo(MCMC) algorithms with competing models evaluated on the basis ofthe posterior probabilities given the data model and prior informationSeparate runs of STRUCTURE were performed on samples from eachsingle geographic population only with different numbers of assumedpopulations ranging from one to five (K=1ndash5) and on samples fromboth populations combined into a single dataset (K=1ndash10) Separateruns were performed using the nuclear data only and the nuclear andmtDNA data combined All simulations used 100000 MCMC iter-ations in the burnin phase and 300000 iterations in the data collectionphase with four independent runs conducted on each set of data andparameter values No prior information regarding social form orpopulation of origin were included and all analyses were run assumingcorrelated allele frequencies We ensured accurate estimates of thesimulation values by checking that model parameters equilibratedbefore the end of the burnin phase and that the posterior probabilitieswere consistent across the four runs for each data and parameter setThe estimated membership coefficients (degree of admixture) ofindividuals and their averages for each of the populations wereobtained from the simulations yielding the preferred estimates ofnumbers of distinct clusters as determined by the method of Evannoet al (2005) Again for all of the analyses of genetic differentiation weutilized the multilocus genetic data from only a single individual persampled nest
One signature of a recent population bottleneck is reduction inallelic diversity but not observed heterozygosity simply because suchpopulation bottlenecks tend to reduce allelic number substantiallymore than genetic diversity at polymorphic loci (Luikart et al 1998)We used the program BOTTLENECK (Cornuet and Luikart 1996Piry et al 1999) to identify whether a mode-shift distortion of allelefrequency distributions characteristic of recent bottlenecks was evidentfor either of the two recently established fire ant populations in Taiwanusing the infinite allele model (IAM) and the two phase model (TPM)Significance was assessed by the sign standardized differences andWilcoxon tests Additionally we also compared NA and HE for thepopulations from South America (Ross et al 2007) the USA(Shoemaker et al 2006) and Taiwan and examined the bottleneckeffect in Taiwanese populations if results showed NA similar betweenUSA and Taiwan but considerably lower than those found in SouthAmerica
The proportions of triploid individuals from P and M colonies ofthe two infested areas in Taiwan were estimated from the proportion ofindividuals harboring three different alleles at one or more micro-satellite locus We repeated PCR amplification and genotyping of themicrosatellite loci for all individuals in which three alleles at any locus
was detected to confirm they indeed were triploid Triploid individualswere not used for any of the population genetic analyses above Insteadwe randomly selected one diploid individual from the same nest for allpopulation genetic analyses
Results
Distribution of the monogyne and polygyne social forms
All nests putatively identified as monogyne in the fieldwere identified as such by examining genotype distribu-tions for the 10 nestmate females surveyed at the sixmicrosatellites and by detecting only the Gp-9B alleleamong these workers Also all putative polygyne nestswere confirmed as such by inspection of genotypedistributions of 10 nestmate offspring females for depar-ture from those expected for offspring in monogyne nests(single female mated to a single male) surveyed at sixmicrosatellite loci andor by detecting individuals bearingthe Gp-9b allele In every case the two complementaryapproaches to determining social form were consistent inclassifying nests as either monogyne or polygyne Thepreliminary analyses show that the distribution of the twosocial forms is quite different in the two infested areas(Fig 1) In Taoyuan townships centrally located in thecounty (Fig 1b) harbor a higher proportion of P colonies(P 60 ) whereas surrounding townships (Fig 1b) areinfested more commonly by ants of the M social form (M765) In contrast the large infested area in Chiayiconsists mostly of P nests (92 of colonies collectedFig 1c)
Genetic diversity
From one to five alleles were found at each microsatellitelocus (23 and 26 alleles in the Taoyuan and Chiayipopulations respectively Table 1) A total of sevenprivate alleles (ie unique to a single population) acrossall six microsatellite loci were found Tests for conformityof genotype proportions to Hardy-Weinberg expectationsrevealed only one significant deviation involving Sol-11 inthe Taoyuan polygyne population (Table 1) Score testsreveal that this significant departure resulted from adeficiency of heterozygotes
Analyses of the mtDNA sequence data revealed onlythree unique variants occur in Taiwan (designatedhaplotypes H5 H22 and H36 see Shoemaker andRoss 1996) all of which are commonly observed in fireants from the USA Individuals from the Taoyuanpopulation harbored either the H36 or H5 mtDNAhaplotype whereas all individuals from the Chiayi pop-ulation bear mtDNA haplotype H22 (Fig 1)
Insect Soc Vol 55 2008 Research article 57
Figure 1 (a) Map of Taiwan showing the two areas (Taoyuan and Chiayi counties) infested by Solenopsis invicta (indicated by shading) Histogramsshow the proportion of nests harboring each mtDNA haplotype Pie diagrams in the enlarged map of Taoyuan county (b) show the proportion of themonogyne and polygyne nests found in the central townships (smaller dotted-line circle) and in the surrounding townships (areas between larger andsmaller dotted-line circles) A pie diagram in the enlarged map of Chiayi county (c) indicates the proportion of two social forms in the sampling area(solid-line circle) Social form of each nest was determined by using a Gp-9 allele-specific assay and by examining the distribution of genotype arraysat six microsatellite loci within nests (see text)
Table 1 Genetic diversity measures within each social form from the two study populations of S invicta in Taiwan n denotes total number ofindividuals (nests) for a given social form in each area NA denotes observed number of alleles found at each locus from each populationHE =expected heterozygosity HO = observed heterozygosity
Locus Taoyuan Chiayi
Monogyne(n= 11)
Polygyne( n=19)
Monogyne( n=2)
Polygyne(n=23)
NA HE HO NA HE HO NA HE HO NA HE HO
Sol-6 1 0000 0000 2 0193 0105 3 0833 0800 4 0599 0434
Sol-11 4 0731 0545 4 0755 0526 2 0500 0700 4 0657 0652
Sol-20 5 0814 0818 5 0734 0631 3 0833 0800 4 0679 0782
Sol-42 4 0610 0363 4 0613 0473 2 0500 0450 5 0656 0608
Sol-49 4 0697 0634 5 0768 0842 3 0833 0850 5 0735 0782
Sol-55 3 0634 0454 3 0624 0789 2 0667 0850 4 0486 0608
Mean 35 0581 0469 38 0614 0561 25 0603 0742 43 0635 0645
58 CC Yang et al Fire ants in Taiwan
Within-nest relatedness
The estimates of relatedness among workers withinmonogyne nests from each area were consistent witheach nest headed by single queen mated to a single male(Taoyuan R=071870036 Chiayi R=071970038Fig 2) Relatedness among workers within polygynenests was significantly greater than zero but significantlylower (Plt0001) than the values for monogyne nests(Taoyuan R=011220006 Chiayi R=014440007Fig 2)
Population genetic differentiation
Exact tests based on six microsatellite loci revealedsignificant differentiation between Taoyuan and Chiayipopulations (Plt002) No significant differentiation wasobserved between sympatric populations of the two socialforms in the two areas (Pgt013) Exact tests for differ-entiation using mtDNA sequence data revealed signifi-cant differentiation occurs between the two geographicpopulations (Plt00001) as well as between the two socialforms in Taoyuan (Plt004) but not between the twosocial forms in Chiayi (only one mtDNA variant occurshere)
Nuclear and mtDNA variation was partitioned byestimating FST simultaneously at both levels using theanalysis of molecular variance approach (Excoffier et al 1992) Results of these analyses for the nuclear locishowed that only the between-population variance com-ponent is significant (Plt0000001 Fig 3a) providingadditional support to the above analyses suggesting thesetwo populations are genetically distinct For analysesinvolving mtDNA data both the between-population andbetween-social form variance components are significant(both Plt005 Fig 3b)
Bayesian simulations using STRUCTURE revealedthat individuals from the Taoyuan population represent a
single distinct genetic cluster suggesting no differentia-tion between sympatric social forms exists whether usingnuclear data only or nuclear and mtDNA combinedIdentical results were obtained for individuals from theChiayi population In contrast simulations incorporatingall individuals from the two areas gave posterior proba-bilities that strongly support the existence of two distinctgenetic clusters using dataset from the nuclear loci orcombination of the two classes of markers (K=2 Fig 4)The two clusters hypothesized by the Bayesian methodcorrespond roughly to geographic populations based onaverage membership coefficientsgt05 for one of the twoposterior clusters in each of these regions The firstcluster (indicated by lighter gray) is comprised largely ofindividuals from the Taoyuan population whereas thesecond cluster (indicated by the darker gray) is comprisedmostly of the individuals from Chiayi Identical clusteringresults were obtained when simulations were run incor-porating both the microsatellite and mtDNA data(Fig 4)
Figure 2 Average relatedness (95 confidence interval) valuesamong workers within nests of the polygyne and monogyne social formsfrom Taoyuan and Chiayi counties Taiwan
Figure 3 Partitioning of genetic variance in S invicta at six micro-satellite loci (a) and mtDNA (b) estimated using analysis of molecularvariance This hierarchical approach partitioned the total variancebetween infested regions (Taoyuan and Chiayi counties) between thetwo social forms within each infested region and within the social formsin each region The percentage of the total variance partitioned at eachlevel is shown Asterisks indicate statistically significant differentiationat a given level (a=005)
Figure 4 Assignment of individuals representing Solenopsis invictafrom both study populations to two genetic clusters inferred fromSTRUCTURE simulations (average membership coefficient Q) Eachvertical bar represents the individual membership coefficients dividedinto parts proportional to the individuals proposed ancestry in eachcluster
Insect Soc Vol 55 2008 Research article 59
Isolation by distance and tests for genetic bottlenecks
No correlation exists between pairwise FST or FST(1ndash FST)values and geographic distance for individuals of thepolygyne social form in either population (TaoyuanPgt022 Chiayi Pgt048) Further no significant iso-lation by distance for the monogyne nests from Taoyuanpopulation was found (Pgt065) We did not perform theMantel tests for the Chiayi monogyne population sinceonly two nests were collected
All tests revealed a significant excess of heterozygos-ity in Taoyuan under both models except the sign test(Table 3) A significant excess of heterozygosity wasfound in Chiayi for the infinite allele model but not thetwo phase model (Table 3) Combined with a mode shiftof allele frequency distribution (Table 3 Fig 5a) theTaoyuan population appears to have undergone a recentbottleneck In contrast the most abundant alleles in theChiayi population belong to the low frequency class(lt01 Fig 5b) resulting in a normal L-shape distribu-
tion We also compared NA (number of observed allelesTable 2) and HE (expected heterozygosity Table 2)among the South American USA and Taiwanesepopulations to determine whether a genetic signaturecharacteristic of a bottleneck was evident for the latterpopulations In order to minimize the effect of populationsize we randomly selected 30 individuals from the USAand South American dataset (Shoemaker et al 2006Ross et al 2007) Results showed that the NA and HE inTaiwan and the USA populations were consistentlylower than in South American fire ant populations Twopopulations in Taiwan had slightly lower NA than inUSA but had similar HE values
Additional findings
Analyses of individual genotypes revealed that 108 ofthe 190 workers sampled from polygyne nests in Taoyuanand 11 of 230 individuals from polygyne nests in Chiayi
Table 2 Genetic diversity measures within the two study populations of S invicta in Taiwan NA denotes observed number of alleles found at eachlocus from each population HE =expected heterozygosity HO = observed heterozygosity Numbers in parentheses represent the number of privatealleles occurring in only one of the two Taiwanese populations Genetic diversity measures for two population from the USA and South Americaalso are shown (see text)
Source of collection Population Sample size Mean no of alleles NA HE HO
TaiwanTaoyuan county 30 38 23 (2) 0609 0527Chiayi county 25 48 26 (5) 0636 0660
USAMississippi 30 57 34 0651 0559western Louisiana 30 58 35 0628 0607
South AmericaFormosa Argentina 30 158 195 0835 0842Corrientes Argentina 30 158 95 0836 0826
Figure 5 Distribution of allele frequencies at six microsatellite loci for ants (single individual per nest) from the Taoyuan (a) and Chiayi populations(b)
60 CC Yang et al Fire ants in Taiwan
harbored three distinct alleles whereas all of the individ-uals sampled from monogyne nests harbored at most twodistinct alleles These results indicate that a substantialnumber of polygyne fire ants are triploid a findingconsistent with an earlier study of fire ants from theUSA (Krieger et al 1999)
Discussion
We studied the colony genetic and population structure ofthe invasive fire ant S invicta in Taiwan by collecting andanalyzing microsatellite genotypic data and mtDNAsequence data of ants collected from two separateinfested areas Our genetic analyses revealed severalpatterns which are discussed in more detail below andcan be summarized as follows 1) Estimates of within nestrelatedness were significantly greater than zero for bothsocial forms from both populations 2) No significantisolation by distance was found among nests within eachsocial form from either population 3) SignificantmtDNA but no nuclear differentiation occurs betweensympatric social forms in Taoyuan (no differentiationbetween forms at either genome is evident in Chiayi) 4)Molecular signatures of genetic bottlenecks associatedwith recent introductions are evident in both populations5) The two sampled populations Taoyuan and Chiayi arehighly genetically differentiated at both the nuclear ormtDNA genomes In addition to these general findingswe found numerous triploid individuals in ants of thepolygyne (P) but not the monogyne (M) form anddistinct geographic distribution of each of the two socialforms between the two populations
Colony social organization and patterns of intracolonyworker relatedness
The estimate of relatedness among workers withinmonogyne colonies was not significantly different from075 the predicted value for a nest headed by one singly-mated queen These data also are consistent with datafrom earlier studies of monogyne S invicta colonies inboth the USA (Ross and Fletcher 1985b) and SouthAmerica (Ross et al 1997) and suggest that the socialand mating biology of M queens is virtually invariantwith M nests representing closed societies headed by onesingly-mated reproductive queen
In contrast colony genetic structure of P social formdoes differ between native South America and intro-duced populations in the USA (Ross et al 1996) Pnests of S invicta in the native range contain relativelyfew queens which are close relatives and relatednessamong nestmate workers is significantly greater thanzero whereas P nests in the USA contain highnumbers of unrelated queens and relatedness amongnestmate workers is not significantly greater than zeroThese differences in queen number and relatedness
between the two ranges are consistent with predictionsbased on differences in the population densities of antsin these two areas (Ross et al 1996) which aresubstantially higher in the USA compared to SouthAmerica (Porter et al 1997) One consequence of thehigher population densities in the USA is that nestingsites are more often saturated such that opportunitiesfor independent founding by queens are more limitedin the introduced range (Ross et al 1996) Theseconstraints on independent founding lead to increasedselection pressure on queens to seek adoption intoexisting colonies and on resident workers of colonies toaccept such queens As queen number within thesenests increases relatedness among nestmate workers isexpected to decrease
Relatedness among workers within P nests in Taiwanis small yet significantly greater than zero and intermedi-ate to the values observed for native and USApopulations The social organization of P nests in Taiwanlikely reflects the fact that nesting sites are much lesssaturated compared to sites in the USA since thepopulation densities of ants in Taiwan currently are muchlower than those in the USA However while such adifference between the two introduced ranges are evidentnow it is unlikely to persist especially if S invictacontinues to spread in Taiwan so that habitats becomesaturated If S invicta continues to spread so that habitatsbecome more saturated future studies of the socialevolution of P populations in Taiwan could shed addi-tional light on whether such a change in habitat avail-ability does in fact lead to a shift in social organizationtowards a higher queen number within nests and lowerrelatedness among nestmates
Patterns of isolation by distance
The lack of isolation by distance among colonies of the Mform within Taoyuan was not unexpected given that mostM queens take part in aerial mating flights oftendispersing several kilometers or more before landingand attempting to found a new nest (Toom et al 1976Ross and Fletcher 1985b Ross et al 1999) Such long-distance dispersal is predicted to erase any signature ofnuclear differentiation among closely spaced nests
The lack of detectable isolation by distance within theP form at both studied sites was somewhat unexpectedconsidering the reproductive habits of P queens whichoften mate in or near their natal nests and form newcolonies by budding or by entering into nearby existingnests (Shoemaker and Ross 1996 Ross et al 1997 Rossand Shoemaker 1997) One explanation for the lack ofdetectable isolation by distance is simply that malesdisperse rather long distances to mate with P queensUnder this scenario one would predict that long distancemale-mediated gene flow would result in homogenizationof allele frequencies at nuclear loci yet significantmicrogeographic mtDNA structure would remain espe-
Insect Soc Vol 55 2008 Research article 61
cially if queen dispersal is limited However no suchpattern was found Significant isolation by distance wasabsent for both the nuclear and mtDNA markers Anadditional explanation for these results is that thesignature of isolation by distance predicted to emergesolely from the natural dispersal biology of these ants isconfounded by the fact that these ants are frequentlyinadvertently transported relatively great distances byhumans Such frequent inadvertent human transport maybe a common characteristic of invasive ants found in closeproximity to human habitats and thus play a role inshaping the genetic structure in introduced areas Finallyit is possible that the lack of a detectable signal of isolationby distance is due to the extremely recent introduction offire ants into Taiwan with insufficient time having passedfor populations to achieve migrationdrift equilibrium
Gene flow between sympatric social forms
The pronounced mtDNA differentiation between sym-patric social forms in Taoyuan is consistent with resultsfrom earlier studies in both South America and the USAshowing that interform gene flow mediated by queens israre or absent Queens of each social form rarely succeedin becoming reproductives in colonies of the alternateform as a result of behavioral decisions (attributed largelyto differences in genotypes of queens at the single geneGp-9) made by both queens and workers (Shoemaker andRoss 1996 Ross and Shoemaker 1997 Keller and Ross1998 DeHeer et al 1999 Goodisman et al 2000 Rossand Keller 2002) The contrasting weak or nonexistentinterform divergence at nuclear genes also is consistentwith studies of S invicta in the USA showing that queensof the P form predominantly mate with males of the Mform where the two forms co-occur (Ross and Shoe-maker 1993 Ross and Keller 1995 Shoemaker andRoss 1996) In the USA most males in P colonies aresterile diploid males which means that P queens relylargely on immigrant M males for mating opportunities(Ross and Fletcher 1985a Ross and Shoemaker 1993)Such interform matings act as a homogenizing force onlyfor nuclear markers The similar frequency of triploidworkers in P nests in Taiwan and the USA suggestsmany diploid males also occur in Taiwan as well (Rosset al 1993 1999 Krieger and Keller 1997 Krieger et al 1999) which means that P queens here must also matewith M males The finding of contrasting patterns ofdifferentiation between the two genomes parallels resultsfrom earlier studies involving S invicta in both the USAand South America (Shoemaker and Ross 1996 Rosset al 1997 Shoemaker et al 2006) One final caveathowever is that we cannot completely rule out thepossibility that the lack of detectable nuclear differ-entiation is due to insufficient time having passed fordivergence to occur despite restricted gene flow betweenthe two forms
Understanding the distributions of the two socialforms in the two infested areas in Taiwan each of whichexhibits rather distinct dispersal strategies is importantfor developing effective control strategies of fire ants inthese infested areas (Porter et al 1991) The distribu-tional pattern of social forms in Taoyuan suggests that theexpansion of this population has involved persistentoutward movement of ants from the core infestationcoupled with frequent long distance movement of Mreproductives into outlying areas (Fig 1b) Based onthese results we recommend that bait broadcasting andsurveillance efforts in Taoyuan should include not onlythe currently infested areas but also should include anarea extending well beyond the infested areas In contrastto the distributional pattern of social forms in Taoyuanthe large infested area in Chiayi consists mostly of P nests(92 of colonies collected Fig 1c) However despitethis difference we suggest a similar eradication strategyshould be employed This is because even thoughcomprehensive bait broadcasting within the entire areamay be more effective than for Taoyuan because of thelow queen vagility and relatively slow rate of reinvasionby budding of polygynous queens (Porter et al 1991)some M colonies do occur in Chiayi and there also ispotential for additional M colonies to arise locally fromthe P social form a possibility that is especially morelikely in areas of low density (Ross and Shoemaker 1997DeHeer and Tschinkel 1998 DeHeer et al 1999)
Genetic bottlenecks in Taoyuan and Chiayi populations
Simulations and statistical tests using the program BOT-TLENECK provide evidence of a significant geneticbottleneck associated with the introduction of fire antsinto Taoyuan with only one exception (Table 3) How-ever this single exception is based on a sign test whichhas relatively low statistical power when the number ofloci is below 20 such as our study (Cornuet and Luikart1996) In Chiayi significant excess in heterozygosity usingthe infinite alleles model (IAM) was observed for all testsbut not for the two-phase model (TPM Table 3) BecauseLuikart and Cornuet (1998) suggest that implementingthe test under the IAM model may be more reliable thanother models these results coupled with finding thatallelic frequencies fit a normal L-shaped distributionprovide evidence for a genetic bottleneck Interestinglyestimates of NA and HE (Table 2) are remarkably similarfor introduced populations from Taiwan and the USAand considerably lower than those from the native SouthAmerican range While the geographic origin of ants inTaiwan is unknown the similar genetic diversity measuresbetween the introduced ranges raises the issue that theintroduction of ants into Taiwan has left only a modestfootprint on patterns of diversity if these ants originatedfrom the USA
62 CC Yang et al Fire ants in Taiwan
Genetic differentiation between Taoyuan and Chiayipopulations
All analyses of population differentiation using themicrosatellite and mtDNA data indicate that the twogeographic populations are strongly differentiated andthus likely originated from separate invasions Whilemultiple lines of evidence support this notion thestrongest evidence is the confinement each of the threemtDNA variants to only one of the two geographicpopulations Interestingly all three variants found inTaiwan are the same three variants commonly found inants from the USA (Shoemaker et al 2006) China(GenBank accession numbers DQ831670-DQ831672)and Australia (CCY and DDS unpublished results) Theoccurrence of these three variants in all recently invadedareas suggests that ants from Taiwan could have origi-nated from any of these other areas We currently aregenerating the necessary baseline data for ants from all ofthese introduced regions as well as from populations intheir native range to infer the source population andinvasion history of ants in Taiwan and these other areas
From the perspective of managing fire ants in Taiwantreating each infested area as a separate management unitwould appear more reasonable and means that scientistsprobably should focus on separate plans for eliminationof two isolated populations (Hampton et al 2004) Onthe other hand because these two areas are at high risk ofconnectivity via long distance human-mediated dispersalcare must be taken to prevent reinvasion into these areaseven after successful eradication Stringent inter-countyquarantine measures such as movement control shouldbe established to monitor the risk of transportation of fireant-infested goods to prevent jump dispersal between thetwo areas and decrease probability of successful erad-ication (Drees and Gold 2003)
Acknowledgments
The authors would like to thank Chung-Chi Lin for providing most ofthe specimens for this study including the first identified samples inTaoyuan and Chiayi counties We are particularly grateful to Hwei-yuChang Hon-Tsen Yu and Chia-Hung Hsieh for helpful discussions andcomments on earlier versions of this manuscript Comments by twoanonymous reviewers greatly improved this manuscript This research
was financially supported by grants from Academia Sinica Taiwan Theuse of trade firm or corporation names in this publication is for theinformation and convenience of the reader Such use does notconstitute an official endorsement or approval by the United StatesDepartment of Agriculture or the Agricultural Research Service of anyproduct or service to the exclusion of others that may be suitable
References
Ahrens ME Ross KG and Shoemaker DD 2005 Phylogeographicstructure of the fire ant Solenopsis invicta in its native SouthAmerican range Roles of natural barriers and habitat connectivityEvolution 59 1733ndash1743
Allen CR Demarais S and Lutz RS 1994 Red imported fire antimpact on wildlife an overview Texas J Sci 46 51ndash59
Allen CR Lutz RS and Demarais S 1998 Ecological effects of theinvasive nonindigenous ant Solenopsis invicta on native verte-brates The wheels on the bus Transactions of the North AmericanWildlife and Natural Resources Conference 63 56 ndash65
Allen CR Willey RD Myers PE Horton PM and Buffa J 2000Impact of red imported fire ant infestation on northern bobwhitequail abundance trends in southeastern United States J AgrUrban Entomol 17 43ndash51
Allen CR Forys EA Rice KG and Wojcik DP 2001 Effects of fireants (Hymenoptera Formicidae) on hatching turtles and preva-lence of fire ants on sea turtle nesting beaches in Florida FloridaEntomol 84 250ndash253
Bouwma AM Ahrens ME DeHeer CJ and Shoemaker DD 2006Distribution and prevalence of Wolbachia in introduced popula-tions of the fire ant Solenopsis invicta Insect Mol Biol 15 89ndash93
Briano JA Calcaterra LA Vander Meer R Valles SM and LivoreJP 2006 New survey for the fire ant microsporidia Vairimorphainvictae and Thelohania solenopsae in southern South Americawith observations on their field persistence and prevalence of dualinfections Environ Entomol 35 1358ndash1365
Buckley A 1999 Fire ants in California Am Bee J 139 88Carroll CR and Hoffman CA 2000 The pervasive ecological effects
of invasive species Exotic and native fire ants In Invertebrates asWeb-Masters in Ecosystems (Coleman DC and Hendrix PF Eds)Wallingford Oxon UK pp 221ndash232
Carruthers RI 2003 Invasive species research in the United StatesDepartment of Agriculture ndash Agricultural Research Service PestManag Sci 59 827ndash834
Chen JSC Shen CH and Lee HJ 2006 Monogynous and polygynousred imported fire ants Solenopsis invicta Buren (HymenopteraFormicidae) in Taiwan Environ Entomol 35 167ndash172
Cornuet JM and Luikart G 1996 Description and power analysis oftwo tests for detecting recent population bottlenecks from allelefrequency data Genetics 144 2001ndash2014
Davis LR Vander Meer RK and Porter SD 2001 Red imported fireants expand their range across the West Indies Florida Entomol84 735ndash736
DeHeer CJ and Tschinkel WR 1998 The success of alternativereproductive tactics in monogyne Solenopsis invicta significancefor transitions in social organization Behav Ecol 9 130ndash135
DeHeer CJ Goodisman MAD and Ross KG 1999 Queen dispersalstrategies in the multiple-queen form of the fire ant Solenopsisinvicta Am Nat 153 660ndash675
Dieringer D and Schlccediltterer C 2002 Microsatellite analyser (MSA) aplatform independent analysis tool for large microsatellite datasets Mol Ecol Notes 3 167ndash169
Drake JA Mooney HA Castri F Groves RH Kruger MRejmnek M and Williamson M 1989 Biological Invasions AGlobal Perspective Chichester UK 550 pp
Drees BM and Vinson SB 1990 Comparison of the control ofmonogynous and polygynous forms of the red imported fire ant(Hymenoptera Formicidae) with a chloripyrifos mound drench JEntomol Sci 25 317ndash324
Table 3 Tests for genetic bottlenecks using the infinite allele model(IAM) and two-phase model (TPM 90 stepwise mutation model and10 multistep mutation model [Garza and Williamson 2001])implemented in the program BOTTLENECK (a=005)Significance at given statistic level
Population Simulatedmodel
Significancetest
Standardizeddifferences test
Wilcoxontest
Taoyuan IAM 0148 0003 0015TPM 0186 0035 0023
Chiayi IAM 0037 0022 0007TPM 0521 0293 0281
Insect Soc Vol 55 2008 Research article 63
Drees BM and Gold RE 2003 Development of integrated pestmanagement programs for the red imported fire ant (Hymeno-ACHTUNGTRENNUNGptera Formicidae) J Entomol Sci 38 170ndash180
Eubanks MD 2001 Estimates of the direct and indirect effects of redimported fire ants on biological control in field crops Biol Control21 35 ndash43
Evanno G Regnaut S and Goudet J 2005 Detecting the number ofclusters of individuals using the software STRUCTURE asimulation study Mol Ecol 14 2611ndash2620
Excoffier L Smouse PE and Quattro JM 1992 Analysis of molecularvariance inferred from metric distances among DNA haplotypesApplication to human mitochondrial DNA restriction dataGenetics 131 479ndash491
Forys EA Quistorff A and Allen CR 2001 Potential fire ant(Hymenoptera Formicidae) impact on the endangered Schausswallowtail (Lepidoptera Papilionidae) Florida Entomol 84 254ndash258
Garza JC and Williamson EG 2001 Detection of reduction inpopulation size using data from microsatellite loci Mol Ecol 10 305ndash318
Giraud T Pedersen JS and Keller L 2002 Evolution of supercoloniesthe Argentine ants of southern Europe Proc Natl Acad Sci U SA 99 6075ndash6079
Giuliano WM Allen CR Lutz RS and Demarais S 1996 Effects ofred imported fire ants on northern bobwhite chicks J WildlManage 60 309ndash313
Glancey BM Craig CH Stringer CE and Bishop PM 1973Multiple fertile queens in colonies of the imported fire antSolenopsis invicta J Georgia Entomol Soc 8 327ndash328
Goodisman MAD DeHeer CJ and Ross KG 2000 Mating behaviorof queens from multiple-queen colonies of Solenopsis invicta JInsect Behav 13 455ndash468
Goodnight KF and Queller DC 1999 Computer software forperforming likelihood tests of pedigree relationship using geneticmarkers Mol Ecol 8 1231ndash1234
Gotelli NJ and Arnett AE 2000 Biogeographic effects of red fire antinvasion Ecol Lett 3 257ndash261
Gotzek D and Ross KG 2007 Genetic regulation of colony socialorganization in fire ants An integrative overview Q Rev Biol 82 201ndash226
Goudet J 2001 FSTAT A Program to Estimate and Test GeneDiversities and Fixation Indices Version 293 http wwwunilch izeasoftwares fstathtml
Greenberg L Fletcher DJC and Vinson SB 1985 Differences inworker size and mound distribution in monogynous and polygy-nous colonies of the fire ant Solenopsis invicta Buren J KansasEntomol Soc 58 9ndash18
Hampton JO Spencer PBS Alpers DL Twigg LE WoolnoughAP Doust J Higgs T and Pluske J 2004 Molecular techniqueswildlife management and the importance of genetic populationstructure and dispersal a case study with feral pigs J Appl Ecol41 735ndash743
Henshaw MT Kunzmann N Vanderwoude C Sanetra M andCrozier RH 2005 Population genetics and history of theintroduced fire ant Solenopsis invicta Buren (HymenopteraFormicidae) in Australia Aus J Entomol 44 37ndash44
Hccedillldobler B and Wilson EO 1990 The Ants The Belknap Press ofHarvard University Press Cambridge Massachusetts 732 pp
Huang TC Chou YC and Chou HC 2004 The infestation andcontrol of the red imported fire ant in Taiwan In Proceedingsof the Symposium on the Control of the Red Imported Fire AntBureau of Animal and Plant Health Inspection and QuarantineCouncil of Agriculture Executive Yuan Taipei Taiwan pp1 ndash 13
Jaquiry J Vogel V and Keller L 2005 Multilevel genetic analyses oftwo European supercolonies of the Argentine ant Linepithemahumile Mol Ecol 14 589ndash598
Jeyaprakash A and Hoy MA 2000 Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropods Insect Mol Biol 9 393ndash405
Jouvenaz DP 1990 Approaches to biological control of fire ants in theUnited States In Applied Myrmecology A World Perspective(Vander Meer RK Jaffe K and Cedeno A Eds) Westview PressBoulder Colorado pp 620ndash627
Kaspari M 2000 Do imported fire ants impact canopy arthropodsEvidence from simple arboreal pitfall traps Southwestern Nat 45 118ndash122
Keller L and Ross KG 1998 Selfish genes a green beard in the red fireant Nature 394 573ndash575
Kemp SF deShazo RD Moffitt JE Williams DF and Buhner WA2000 Expanding habitat of the imported fire ant (Solenopsisinvicta) A public health concern J Allergy Clin Immunol 105 683ndash691
Kopachena JG Buckley AJ and Potts GA 2000 Effects of the redimported fire ant (Solenopsis invicta) on reproductive success ofbarn swallows (Hirundo rustica) in northeast Texas SouthwesternNat 45 477ndash482
Krieger MJB and Keller L 1997 Polymorphism at dinucleotidemicrosatellite loci in fire ant Solenopsis invicta populations MolEcol 6 997ndash999
Krieger MJB Ross KG Chang CWY and Keller L 1999Frequency and origin of triploidy in the fire ant Solenopsis invictaHeredity 82 142ndash150
Krieger MJB and Ross KG 2002 Identification of a major generegulating complex social behavior Science 295 328ndash332
Lee CE 2002 Evolutionary genetics of invasive species Trends EcolEvol 17 386ndash391
Lofgren CS Banks WA and Glancey BM 1975 Biology and controlof imported fire ants Annu Rev Entomol 20 1ndash30
Lofgren CS 1986 History of imported fire ants in the United States InFire Ants and Leaf Cutting Ants Biology and Management(Lofgren CS and Vander Meer RK Eds) Westview PressBoulder Colorado pp 36ndash49
Luikart G Allendorf FW Cornuet JM and Sherwin WB 1998Distortion of allele frequency distributions provides a test forrecent population bottlenecks J Hered 89 238ndash247
Luikart G and Cornuet JM 1998 Empirical evaluation of a test foridentifying recently bottlenecked populations from allele frequen-cy data Conserv Biol 12 228ndash237
MacKay WP and Fagerlund R 1997 Range expansion of the redimported fire ant Solenopsis invicta Buren (Hymenoptera For-micidae) into New Mexico and extreme western Texas ProcEntomol Soc Wash 99 757ndash758
Mank JE and Avise JC 2004 Individual organisms as units of analysisBayesian-clustering alternatives in population genetics Genet Res84 135ndash143
May GE Gelembiuk GW Panov VE Orlova MI and Lee CE2006 Molecular ecology of zebra mussel invasions Mol Ecol 15 1021ndash1031
McCubbin KI and Weiner JM 2002 Fire ants in Australia A newmedical and ecological hazard Med J Aus 176 518ndash519
Morrison LW 2000 Mechanisms of interspecific competition amongan invasive and two native fire ants Oikos 90 238ndash252
Morrison LW 2002 Long-term impacts of an arthropod-communityinvasion by the imported fire ant Solenopsis invicta Ecology 83 2337ndash2345
Nattrass R and Vanderwoude C 2001 A preliminary investigation ofthe ecological effects of red imported fire ants (Solenopsis invicta)in Brisbane Ecol Manag Resto 2 220ndash223
Nei M 1972 Genetic distance between populations Am Nat 106 283ndash292
Orr MR 1996 Host manipulation by Wolbachia is a neutral trait withinsingle populations Anim Behav 51 1183ndash1185
Patterson RS 1994 Biological control of introduced ant species InExotic Ants Biology Impact and Control of Introduced Species(Williams DF Ed) Westview Press Boulder Colorado pp 293ndash308
Piry S Luikart G and Cornuet JM 1999 BOTTLENECK acomputer program for detecting recent reductions in the effectivepopulation size using allele frequency data J Hered 90 502ndash503
64 CC Yang et al Fire ants in Taiwan
Porter SD and Savignano DA 1990 Invasion of polygyne fire antsdecimates native ants and disrupts arthropod community Ecology7 2095ndash2106
Porter SD Bhatkar A Mulder R Vinson SB and Clair DJ 1991Distribution and density of polygyne fire ants (HymenopteraFormicidae) in Texas J Econ Entomol 84 866ndash874
Porter SD Fowler HG and MacKay WP 1992 Fire ant mounddensities in the United States and Brazil (Hymenoptera Formi-cidae) J Econ Entomol 85 1154ndash1161
Porter SD Williams DF Patterson RS and Fowler HG 1997Intercontinental differences in the abundance of Solenopsis fireants (Hymenoptera Formicidae) Escape from natural enemiesEnviron Entomol 26 373ndash384
Pritchard JK Stephens M and Donnelly P 2000 Inference ofpopulation structure using multilocus genotype data Genetics155 945ndash959
Raymond M and Rousset F 1995a An exact test for populationdifferentiation Evolution 49 1280ndash1283
Raymond M and Rousset F 1995b GENEPOP (version 12)population genetics software for exact tests and ecumenicism JHered 86 248ndash249
Reuter M Pedersen JS and Keller L 2005 Loss of Wolbachiainfection during colonisation in the invasive Argentine ant Line-pithema humile Heredity 94 364ndash369
Ross KG and Fletcher DJC 1985a Genetic origin of male diploidy inthe fire ant Solenopsis invicta (Hymenoptera Formicidae) and itsevolutionary significance Evolution 39 888ndash903
Ross KG and Fletcher DJC 1985b Comparative study of genetic andsocial structure in two forms of the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 17 349ndash356
Ross KG and Fletcher DJC 1986 Diploid male productionndashasignificant colony mortality factor in the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 19 283ndash291
Ross KG and Shoemaker DD 1993 An unusual pattern of gene flowbetween the two social forms of the fire ant Solenopsis invictaEvolution 47 1595ndash1605
Ross KG Vargo EL Keller L and Trager JC 1993 Effect of afounder event on variation in the genetic sex-determining system ofthe fire ant Solenopsis invicta Genetics 135 843ndash854
Ross KG and Keller L 1995 Joint influence of gene flow and selectionon a reproductively important genetic polymorphism in the fire antSolenopsis invicta Am Nat 146 325ndash348
Ross KG Vargo EL and Keller L 1996 Social evolution in a newenvironment The case of introduced fire ants Proc Natl AcadSci U S A 93 3021ndash3025
Ross KG 1997 Multilocus evolution in fire ants Effects of selectiongene flow and recombination Genetics 145 961ndash974
Ross KG Krieger MJB Shoemaker DD Vargo EL and Keller L1997 Hierarchical analysis of genetic structure in native fire antpopulations Results from three classes of molecular markersGenetics 147 643ndash655
Ross KG and Shoemaker DD 1997 Nuclear and mitochondrialgenetic structure in two social forms of the fire ant Solenopsisinvicta Insights into transitions to an alternate social organizationHeredity 78 590ndash602
Ross KG and Keller L 1998 Genetic control of social organization inan ant Proc Natl Acad Sci U S A 95 14232ndash14237
Ross KG Shoemaker DD Krieger MJ DeHeer CJ and Keller L1999 Assessing genetic structure with multiple classes of molecularmarkers A case study involving the introduced fire ant Solenopsisinvicta Mol Biol Evol 16 525ndash543
Ross KG and Keller L 2002 Experimental conversion of colony socialorganization by manipulation of worker genotype composition infire ants (Solenopsis invicta) Behav Ecol Sociobiol 51 287ndash295
Ross KG Krieger MJB and Shoemaker DD 2003 Alternativegenetic foundations and evolutionary routes for a key socialpolymorphism in fire ants Genetics 165 1853ndash1867
Ross KG Krieger MJB Keller L and Shoemaker DD 2007Genetic variation and structure in native populations of the fire antSolenopsis invicta Evolutionary and demographic implicationsBiol J Linnean Soc 92 541ndash560
Rousset F 1997 Genetic differentiation and estimation of gene flowfrom F-statistics under isolation by distance Genetics 145 1219ndash1228
Sax DF Stachowicz JJ and Gaines SD 2005 Species InvasionsInsights into Ecology Evolution and Biogeography SinauerAssociates Sunderland Massachussetts 495 pp
Shoemaker DD and Ross KG 1996 Effects of social organization ongene flow in the fire ant Solenopsis invicta Nature 383 613ndash616
Shoemaker DD Ross KG Keller L Vargo EL and Werren JH2000 Wolbachia infections in native and introduced populations offire ants (Solenopsis spp) Insect Mol Biol 9 661ndash673
Shoemaker DD Ahrens M Sheill L Mescher M Keller L and RossKG 2003 Distribution and prevalence of Wolbachia infections innative populations of the fire ant Solenopsis invicta (HymenopteraFormicidae) Environ Entomol 32 1329ndash1336
Shoemaker DD DeHeer CJ Krieger MJB and Ross KG 2006Population genetics of the invasive fire ant Solenopsis invicta(Hymenoptera Formicidae) in the USA Ann Entomol Soc Am99 1213ndash1233
Simon C Frati F Beckenbach A Crespi B Liu H and Flook P 1994Evolution weighting and phylogenetic utility of mitochondrialgene sequences and a compilation of conserved polymerase chainreaction primers Ann Entomol Soc Am 87 651ndash701
Toom PM Cupp E Johnson CP and Griffin I 1976 Utilization ofbody reserves for minim brood development by queens of theimported fire ant Solenopsis invicta J Insect Physiol 22 217ndash220
Tsutsui ND and Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepethema humile) in its nativeand introduced ranges Evolution 55 976ndash985
Tsutsui ND Kauppinen SN Oyafuso AF and Grosberg RK 2003The distribution and evolutionary history of Wolbachia infection innative and introduced populations of the invasive argentine ant(Linepithema humile) Mol Ecol 12 3057ndash3068
Valles SM and Porter SD 2003 Identification of polygyne andmonogyne fire ant colonies (Solenopsis invicta) by multiplex PCRof Gp-9 alleles Insect Soc 50 199ndash200
Valles SM Oi DH Briano JA and Williams DF 2004 Simultaneousdetection of Vairimorpha invictae (Microsporidia Burenellidae)and Thelohania solenopsae (Microsporidia Thelohaniidae) in fireants by PCR Florida Entomol 87 85ndash87
Vander Meer R K and Porter SD 2001 Fate of newly-mated queensintroduced into monogyne and polygyne Solenopsis invicta (Hy-menoptera Formicidae) colonies Ann Entomol Soc Am 94 289ndash297
Vander Meer RK Jaffe K and Cedeno A 1990 Applied Myrmecol-ogy A World Perspective Westview Press Boulder CO 741 pp
Vinson SB 1994 Impact and invasion of Solenopsis invicta (Buren) onnative food webs In Exotic Ants Biology Impact and Control ofIntroduced Species (Williams DF Ed) Westview Press BoulderColorado pp 293ndash308
Vinson SB 1997 Invasion of the red imported fire ant (HymenopteraFormicidae) Spread biology and impact Am Entomol 43 23ndash29
Williams DF and Porter SD 1994 Fire ant control Science 264 1653ndash1653
Wojcik D Allen CR Brenner RJ Forys EA Jouvenaz D and LutzRS 2001 Red imported fire ants Impact on biodiversity AmEntomol 47 16ndash23
Xia X and Xie Z 2001 DAMBE Data analysis in molecular biologyand evolution J Hered 92 371ndash373
Insect Soc Vol 55 2008 Research article 65

populations was determined by means of exact tests implemented inGENEPOP 33 (Raymond and Rousset 1995a) Genetic distancevalues (Neis D) also were calculated to further assess divergenceamong populations (Nei 1972)
FST values were also estimated simultaneously at two levels (socialform geographic population) using the hierarchical analysis ofmolecular variance procedure in ARLEQUIN (Excoffier et al1992) Statistical significance of the differentiation at each level wasestablished by means of 20000 data permutations
We tested for correlations between pairwise FST values [FST(1ndashFST)] and the natural logarithm of pairwise geographic distances amongnests from each social form in the two areas except for monogynousindividuals from Chiayi at all six microsatellite loci using the Manteltest in GENEPOP 33 (Raymond and Rousset 1995b) to determine ifthere was a significant positive correlation between genetic differ-entiation and geographic distance indicative of isolation by distance
We also used an individual-based Bayesian method to statisticallyrecognize distinct genetic clusters of individuals and to infer levels ofpopulation admixture from individual multilocus genotypic data(Pritchard et al 2000 Mank and Avise 2004) The method assumesa model with K populations each with characteristic allele frequenciesthat are estimated while individuals are probabilistically assigned toeach population Prior information such as the geographic location ofsamples or colony social form also can be incorporated into the modelto assist in clustering The program STRUCTURE (Pritchard et al2000 URL httppritchbsduchicagoedu) was used to explore theparameter space of each model using Markov chain Monte Carlo(MCMC) algorithms with competing models evaluated on the basis ofthe posterior probabilities given the data model and prior informationSeparate runs of STRUCTURE were performed on samples from eachsingle geographic population only with different numbers of assumedpopulations ranging from one to five (K=1ndash5) and on samples fromboth populations combined into a single dataset (K=1ndash10) Separateruns were performed using the nuclear data only and the nuclear andmtDNA data combined All simulations used 100000 MCMC iter-ations in the burnin phase and 300000 iterations in the data collectionphase with four independent runs conducted on each set of data andparameter values No prior information regarding social form orpopulation of origin were included and all analyses were run assumingcorrelated allele frequencies We ensured accurate estimates of thesimulation values by checking that model parameters equilibratedbefore the end of the burnin phase and that the posterior probabilitieswere consistent across the four runs for each data and parameter setThe estimated membership coefficients (degree of admixture) ofindividuals and their averages for each of the populations wereobtained from the simulations yielding the preferred estimates ofnumbers of distinct clusters as determined by the method of Evannoet al (2005) Again for all of the analyses of genetic differentiation weutilized the multilocus genetic data from only a single individual persampled nest
One signature of a recent population bottleneck is reduction inallelic diversity but not observed heterozygosity simply because suchpopulation bottlenecks tend to reduce allelic number substantiallymore than genetic diversity at polymorphic loci (Luikart et al 1998)We used the program BOTTLENECK (Cornuet and Luikart 1996Piry et al 1999) to identify whether a mode-shift distortion of allelefrequency distributions characteristic of recent bottlenecks was evidentfor either of the two recently established fire ant populations in Taiwanusing the infinite allele model (IAM) and the two phase model (TPM)Significance was assessed by the sign standardized differences andWilcoxon tests Additionally we also compared NA and HE for thepopulations from South America (Ross et al 2007) the USA(Shoemaker et al 2006) and Taiwan and examined the bottleneckeffect in Taiwanese populations if results showed NA similar betweenUSA and Taiwan but considerably lower than those found in SouthAmerica
The proportions of triploid individuals from P and M colonies ofthe two infested areas in Taiwan were estimated from the proportion ofindividuals harboring three different alleles at one or more micro-satellite locus We repeated PCR amplification and genotyping of themicrosatellite loci for all individuals in which three alleles at any locus
was detected to confirm they indeed were triploid Triploid individualswere not used for any of the population genetic analyses above Insteadwe randomly selected one diploid individual from the same nest for allpopulation genetic analyses
Results
Distribution of the monogyne and polygyne social forms
All nests putatively identified as monogyne in the fieldwere identified as such by examining genotype distribu-tions for the 10 nestmate females surveyed at the sixmicrosatellites and by detecting only the Gp-9B alleleamong these workers Also all putative polygyne nestswere confirmed as such by inspection of genotypedistributions of 10 nestmate offspring females for depar-ture from those expected for offspring in monogyne nests(single female mated to a single male) surveyed at sixmicrosatellite loci andor by detecting individuals bearingthe Gp-9b allele In every case the two complementaryapproaches to determining social form were consistent inclassifying nests as either monogyne or polygyne Thepreliminary analyses show that the distribution of the twosocial forms is quite different in the two infested areas(Fig 1) In Taoyuan townships centrally located in thecounty (Fig 1b) harbor a higher proportion of P colonies(P 60 ) whereas surrounding townships (Fig 1b) areinfested more commonly by ants of the M social form (M765) In contrast the large infested area in Chiayiconsists mostly of P nests (92 of colonies collectedFig 1c)
Genetic diversity
From one to five alleles were found at each microsatellitelocus (23 and 26 alleles in the Taoyuan and Chiayipopulations respectively Table 1) A total of sevenprivate alleles (ie unique to a single population) acrossall six microsatellite loci were found Tests for conformityof genotype proportions to Hardy-Weinberg expectationsrevealed only one significant deviation involving Sol-11 inthe Taoyuan polygyne population (Table 1) Score testsreveal that this significant departure resulted from adeficiency of heterozygotes
Analyses of the mtDNA sequence data revealed onlythree unique variants occur in Taiwan (designatedhaplotypes H5 H22 and H36 see Shoemaker andRoss 1996) all of which are commonly observed in fireants from the USA Individuals from the Taoyuanpopulation harbored either the H36 or H5 mtDNAhaplotype whereas all individuals from the Chiayi pop-ulation bear mtDNA haplotype H22 (Fig 1)
Insect Soc Vol 55 2008 Research article 57
Figure 1 (a) Map of Taiwan showing the two areas (Taoyuan and Chiayi counties) infested by Solenopsis invicta (indicated by shading) Histogramsshow the proportion of nests harboring each mtDNA haplotype Pie diagrams in the enlarged map of Taoyuan county (b) show the proportion of themonogyne and polygyne nests found in the central townships (smaller dotted-line circle) and in the surrounding townships (areas between larger andsmaller dotted-line circles) A pie diagram in the enlarged map of Chiayi county (c) indicates the proportion of two social forms in the sampling area(solid-line circle) Social form of each nest was determined by using a Gp-9 allele-specific assay and by examining the distribution of genotype arraysat six microsatellite loci within nests (see text)
Table 1 Genetic diversity measures within each social form from the two study populations of S invicta in Taiwan n denotes total number ofindividuals (nests) for a given social form in each area NA denotes observed number of alleles found at each locus from each populationHE =expected heterozygosity HO = observed heterozygosity
Locus Taoyuan Chiayi
Monogyne(n= 11)
Polygyne( n=19)
Monogyne( n=2)
Polygyne(n=23)
NA HE HO NA HE HO NA HE HO NA HE HO
Sol-6 1 0000 0000 2 0193 0105 3 0833 0800 4 0599 0434
Sol-11 4 0731 0545 4 0755 0526 2 0500 0700 4 0657 0652
Sol-20 5 0814 0818 5 0734 0631 3 0833 0800 4 0679 0782
Sol-42 4 0610 0363 4 0613 0473 2 0500 0450 5 0656 0608
Sol-49 4 0697 0634 5 0768 0842 3 0833 0850 5 0735 0782
Sol-55 3 0634 0454 3 0624 0789 2 0667 0850 4 0486 0608
Mean 35 0581 0469 38 0614 0561 25 0603 0742 43 0635 0645
58 CC Yang et al Fire ants in Taiwan
Within-nest relatedness
The estimates of relatedness among workers withinmonogyne nests from each area were consistent witheach nest headed by single queen mated to a single male(Taoyuan R=071870036 Chiayi R=071970038Fig 2) Relatedness among workers within polygynenests was significantly greater than zero but significantlylower (Plt0001) than the values for monogyne nests(Taoyuan R=011220006 Chiayi R=014440007Fig 2)
Population genetic differentiation
Exact tests based on six microsatellite loci revealedsignificant differentiation between Taoyuan and Chiayipopulations (Plt002) No significant differentiation wasobserved between sympatric populations of the two socialforms in the two areas (Pgt013) Exact tests for differ-entiation using mtDNA sequence data revealed signifi-cant differentiation occurs between the two geographicpopulations (Plt00001) as well as between the two socialforms in Taoyuan (Plt004) but not between the twosocial forms in Chiayi (only one mtDNA variant occurshere)
Nuclear and mtDNA variation was partitioned byestimating FST simultaneously at both levels using theanalysis of molecular variance approach (Excoffier et al 1992) Results of these analyses for the nuclear locishowed that only the between-population variance com-ponent is significant (Plt0000001 Fig 3a) providingadditional support to the above analyses suggesting thesetwo populations are genetically distinct For analysesinvolving mtDNA data both the between-population andbetween-social form variance components are significant(both Plt005 Fig 3b)
Bayesian simulations using STRUCTURE revealedthat individuals from the Taoyuan population represent a
single distinct genetic cluster suggesting no differentia-tion between sympatric social forms exists whether usingnuclear data only or nuclear and mtDNA combinedIdentical results were obtained for individuals from theChiayi population In contrast simulations incorporatingall individuals from the two areas gave posterior proba-bilities that strongly support the existence of two distinctgenetic clusters using dataset from the nuclear loci orcombination of the two classes of markers (K=2 Fig 4)The two clusters hypothesized by the Bayesian methodcorrespond roughly to geographic populations based onaverage membership coefficientsgt05 for one of the twoposterior clusters in each of these regions The firstcluster (indicated by lighter gray) is comprised largely ofindividuals from the Taoyuan population whereas thesecond cluster (indicated by the darker gray) is comprisedmostly of the individuals from Chiayi Identical clusteringresults were obtained when simulations were run incor-porating both the microsatellite and mtDNA data(Fig 4)
Figure 2 Average relatedness (95 confidence interval) valuesamong workers within nests of the polygyne and monogyne social formsfrom Taoyuan and Chiayi counties Taiwan
Figure 3 Partitioning of genetic variance in S invicta at six micro-satellite loci (a) and mtDNA (b) estimated using analysis of molecularvariance This hierarchical approach partitioned the total variancebetween infested regions (Taoyuan and Chiayi counties) between thetwo social forms within each infested region and within the social formsin each region The percentage of the total variance partitioned at eachlevel is shown Asterisks indicate statistically significant differentiationat a given level (a=005)
Figure 4 Assignment of individuals representing Solenopsis invictafrom both study populations to two genetic clusters inferred fromSTRUCTURE simulations (average membership coefficient Q) Eachvertical bar represents the individual membership coefficients dividedinto parts proportional to the individuals proposed ancestry in eachcluster
Insect Soc Vol 55 2008 Research article 59
Isolation by distance and tests for genetic bottlenecks
No correlation exists between pairwise FST or FST(1ndash FST)values and geographic distance for individuals of thepolygyne social form in either population (TaoyuanPgt022 Chiayi Pgt048) Further no significant iso-lation by distance for the monogyne nests from Taoyuanpopulation was found (Pgt065) We did not perform theMantel tests for the Chiayi monogyne population sinceonly two nests were collected
All tests revealed a significant excess of heterozygos-ity in Taoyuan under both models except the sign test(Table 3) A significant excess of heterozygosity wasfound in Chiayi for the infinite allele model but not thetwo phase model (Table 3) Combined with a mode shiftof allele frequency distribution (Table 3 Fig 5a) theTaoyuan population appears to have undergone a recentbottleneck In contrast the most abundant alleles in theChiayi population belong to the low frequency class(lt01 Fig 5b) resulting in a normal L-shape distribu-
tion We also compared NA (number of observed allelesTable 2) and HE (expected heterozygosity Table 2)among the South American USA and Taiwanesepopulations to determine whether a genetic signaturecharacteristic of a bottleneck was evident for the latterpopulations In order to minimize the effect of populationsize we randomly selected 30 individuals from the USAand South American dataset (Shoemaker et al 2006Ross et al 2007) Results showed that the NA and HE inTaiwan and the USA populations were consistentlylower than in South American fire ant populations Twopopulations in Taiwan had slightly lower NA than inUSA but had similar HE values
Additional findings
Analyses of individual genotypes revealed that 108 ofthe 190 workers sampled from polygyne nests in Taoyuanand 11 of 230 individuals from polygyne nests in Chiayi
Table 2 Genetic diversity measures within the two study populations of S invicta in Taiwan NA denotes observed number of alleles found at eachlocus from each population HE =expected heterozygosity HO = observed heterozygosity Numbers in parentheses represent the number of privatealleles occurring in only one of the two Taiwanese populations Genetic diversity measures for two population from the USA and South Americaalso are shown (see text)
Source of collection Population Sample size Mean no of alleles NA HE HO
TaiwanTaoyuan county 30 38 23 (2) 0609 0527Chiayi county 25 48 26 (5) 0636 0660
USAMississippi 30 57 34 0651 0559western Louisiana 30 58 35 0628 0607
South AmericaFormosa Argentina 30 158 195 0835 0842Corrientes Argentina 30 158 95 0836 0826
Figure 5 Distribution of allele frequencies at six microsatellite loci for ants (single individual per nest) from the Taoyuan (a) and Chiayi populations(b)
60 CC Yang et al Fire ants in Taiwan
harbored three distinct alleles whereas all of the individ-uals sampled from monogyne nests harbored at most twodistinct alleles These results indicate that a substantialnumber of polygyne fire ants are triploid a findingconsistent with an earlier study of fire ants from theUSA (Krieger et al 1999)
Discussion
We studied the colony genetic and population structure ofthe invasive fire ant S invicta in Taiwan by collecting andanalyzing microsatellite genotypic data and mtDNAsequence data of ants collected from two separateinfested areas Our genetic analyses revealed severalpatterns which are discussed in more detail below andcan be summarized as follows 1) Estimates of within nestrelatedness were significantly greater than zero for bothsocial forms from both populations 2) No significantisolation by distance was found among nests within eachsocial form from either population 3) SignificantmtDNA but no nuclear differentiation occurs betweensympatric social forms in Taoyuan (no differentiationbetween forms at either genome is evident in Chiayi) 4)Molecular signatures of genetic bottlenecks associatedwith recent introductions are evident in both populations5) The two sampled populations Taoyuan and Chiayi arehighly genetically differentiated at both the nuclear ormtDNA genomes In addition to these general findingswe found numerous triploid individuals in ants of thepolygyne (P) but not the monogyne (M) form anddistinct geographic distribution of each of the two socialforms between the two populations
Colony social organization and patterns of intracolonyworker relatedness
The estimate of relatedness among workers withinmonogyne colonies was not significantly different from075 the predicted value for a nest headed by one singly-mated queen These data also are consistent with datafrom earlier studies of monogyne S invicta colonies inboth the USA (Ross and Fletcher 1985b) and SouthAmerica (Ross et al 1997) and suggest that the socialand mating biology of M queens is virtually invariantwith M nests representing closed societies headed by onesingly-mated reproductive queen
In contrast colony genetic structure of P social formdoes differ between native South America and intro-duced populations in the USA (Ross et al 1996) Pnests of S invicta in the native range contain relativelyfew queens which are close relatives and relatednessamong nestmate workers is significantly greater thanzero whereas P nests in the USA contain highnumbers of unrelated queens and relatedness amongnestmate workers is not significantly greater than zeroThese differences in queen number and relatedness
between the two ranges are consistent with predictionsbased on differences in the population densities of antsin these two areas (Ross et al 1996) which aresubstantially higher in the USA compared to SouthAmerica (Porter et al 1997) One consequence of thehigher population densities in the USA is that nestingsites are more often saturated such that opportunitiesfor independent founding by queens are more limitedin the introduced range (Ross et al 1996) Theseconstraints on independent founding lead to increasedselection pressure on queens to seek adoption intoexisting colonies and on resident workers of colonies toaccept such queens As queen number within thesenests increases relatedness among nestmate workers isexpected to decrease
Relatedness among workers within P nests in Taiwanis small yet significantly greater than zero and intermedi-ate to the values observed for native and USApopulations The social organization of P nests in Taiwanlikely reflects the fact that nesting sites are much lesssaturated compared to sites in the USA since thepopulation densities of ants in Taiwan currently are muchlower than those in the USA However while such adifference between the two introduced ranges are evidentnow it is unlikely to persist especially if S invictacontinues to spread in Taiwan so that habitats becomesaturated If S invicta continues to spread so that habitatsbecome more saturated future studies of the socialevolution of P populations in Taiwan could shed addi-tional light on whether such a change in habitat avail-ability does in fact lead to a shift in social organizationtowards a higher queen number within nests and lowerrelatedness among nestmates
Patterns of isolation by distance
The lack of isolation by distance among colonies of the Mform within Taoyuan was not unexpected given that mostM queens take part in aerial mating flights oftendispersing several kilometers or more before landingand attempting to found a new nest (Toom et al 1976Ross and Fletcher 1985b Ross et al 1999) Such long-distance dispersal is predicted to erase any signature ofnuclear differentiation among closely spaced nests
The lack of detectable isolation by distance within theP form at both studied sites was somewhat unexpectedconsidering the reproductive habits of P queens whichoften mate in or near their natal nests and form newcolonies by budding or by entering into nearby existingnests (Shoemaker and Ross 1996 Ross et al 1997 Rossand Shoemaker 1997) One explanation for the lack ofdetectable isolation by distance is simply that malesdisperse rather long distances to mate with P queensUnder this scenario one would predict that long distancemale-mediated gene flow would result in homogenizationof allele frequencies at nuclear loci yet significantmicrogeographic mtDNA structure would remain espe-
Insect Soc Vol 55 2008 Research article 61
cially if queen dispersal is limited However no suchpattern was found Significant isolation by distance wasabsent for both the nuclear and mtDNA markers Anadditional explanation for these results is that thesignature of isolation by distance predicted to emergesolely from the natural dispersal biology of these ants isconfounded by the fact that these ants are frequentlyinadvertently transported relatively great distances byhumans Such frequent inadvertent human transport maybe a common characteristic of invasive ants found in closeproximity to human habitats and thus play a role inshaping the genetic structure in introduced areas Finallyit is possible that the lack of a detectable signal of isolationby distance is due to the extremely recent introduction offire ants into Taiwan with insufficient time having passedfor populations to achieve migrationdrift equilibrium
Gene flow between sympatric social forms
The pronounced mtDNA differentiation between sym-patric social forms in Taoyuan is consistent with resultsfrom earlier studies in both South America and the USAshowing that interform gene flow mediated by queens israre or absent Queens of each social form rarely succeedin becoming reproductives in colonies of the alternateform as a result of behavioral decisions (attributed largelyto differences in genotypes of queens at the single geneGp-9) made by both queens and workers (Shoemaker andRoss 1996 Ross and Shoemaker 1997 Keller and Ross1998 DeHeer et al 1999 Goodisman et al 2000 Rossand Keller 2002) The contrasting weak or nonexistentinterform divergence at nuclear genes also is consistentwith studies of S invicta in the USA showing that queensof the P form predominantly mate with males of the Mform where the two forms co-occur (Ross and Shoe-maker 1993 Ross and Keller 1995 Shoemaker andRoss 1996) In the USA most males in P colonies aresterile diploid males which means that P queens relylargely on immigrant M males for mating opportunities(Ross and Fletcher 1985a Ross and Shoemaker 1993)Such interform matings act as a homogenizing force onlyfor nuclear markers The similar frequency of triploidworkers in P nests in Taiwan and the USA suggestsmany diploid males also occur in Taiwan as well (Rosset al 1993 1999 Krieger and Keller 1997 Krieger et al 1999) which means that P queens here must also matewith M males The finding of contrasting patterns ofdifferentiation between the two genomes parallels resultsfrom earlier studies involving S invicta in both the USAand South America (Shoemaker and Ross 1996 Rosset al 1997 Shoemaker et al 2006) One final caveathowever is that we cannot completely rule out thepossibility that the lack of detectable nuclear differ-entiation is due to insufficient time having passed fordivergence to occur despite restricted gene flow betweenthe two forms
Understanding the distributions of the two socialforms in the two infested areas in Taiwan each of whichexhibits rather distinct dispersal strategies is importantfor developing effective control strategies of fire ants inthese infested areas (Porter et al 1991) The distribu-tional pattern of social forms in Taoyuan suggests that theexpansion of this population has involved persistentoutward movement of ants from the core infestationcoupled with frequent long distance movement of Mreproductives into outlying areas (Fig 1b) Based onthese results we recommend that bait broadcasting andsurveillance efforts in Taoyuan should include not onlythe currently infested areas but also should include anarea extending well beyond the infested areas In contrastto the distributional pattern of social forms in Taoyuanthe large infested area in Chiayi consists mostly of P nests(92 of colonies collected Fig 1c) However despitethis difference we suggest a similar eradication strategyshould be employed This is because even thoughcomprehensive bait broadcasting within the entire areamay be more effective than for Taoyuan because of thelow queen vagility and relatively slow rate of reinvasionby budding of polygynous queens (Porter et al 1991)some M colonies do occur in Chiayi and there also ispotential for additional M colonies to arise locally fromthe P social form a possibility that is especially morelikely in areas of low density (Ross and Shoemaker 1997DeHeer and Tschinkel 1998 DeHeer et al 1999)
Genetic bottlenecks in Taoyuan and Chiayi populations
Simulations and statistical tests using the program BOT-TLENECK provide evidence of a significant geneticbottleneck associated with the introduction of fire antsinto Taoyuan with only one exception (Table 3) How-ever this single exception is based on a sign test whichhas relatively low statistical power when the number ofloci is below 20 such as our study (Cornuet and Luikart1996) In Chiayi significant excess in heterozygosity usingthe infinite alleles model (IAM) was observed for all testsbut not for the two-phase model (TPM Table 3) BecauseLuikart and Cornuet (1998) suggest that implementingthe test under the IAM model may be more reliable thanother models these results coupled with finding thatallelic frequencies fit a normal L-shaped distributionprovide evidence for a genetic bottleneck Interestinglyestimates of NA and HE (Table 2) are remarkably similarfor introduced populations from Taiwan and the USAand considerably lower than those from the native SouthAmerican range While the geographic origin of ants inTaiwan is unknown the similar genetic diversity measuresbetween the introduced ranges raises the issue that theintroduction of ants into Taiwan has left only a modestfootprint on patterns of diversity if these ants originatedfrom the USA
62 CC Yang et al Fire ants in Taiwan
Genetic differentiation between Taoyuan and Chiayipopulations
All analyses of population differentiation using themicrosatellite and mtDNA data indicate that the twogeographic populations are strongly differentiated andthus likely originated from separate invasions Whilemultiple lines of evidence support this notion thestrongest evidence is the confinement each of the threemtDNA variants to only one of the two geographicpopulations Interestingly all three variants found inTaiwan are the same three variants commonly found inants from the USA (Shoemaker et al 2006) China(GenBank accession numbers DQ831670-DQ831672)and Australia (CCY and DDS unpublished results) Theoccurrence of these three variants in all recently invadedareas suggests that ants from Taiwan could have origi-nated from any of these other areas We currently aregenerating the necessary baseline data for ants from all ofthese introduced regions as well as from populations intheir native range to infer the source population andinvasion history of ants in Taiwan and these other areas
From the perspective of managing fire ants in Taiwantreating each infested area as a separate management unitwould appear more reasonable and means that scientistsprobably should focus on separate plans for eliminationof two isolated populations (Hampton et al 2004) Onthe other hand because these two areas are at high risk ofconnectivity via long distance human-mediated dispersalcare must be taken to prevent reinvasion into these areaseven after successful eradication Stringent inter-countyquarantine measures such as movement control shouldbe established to monitor the risk of transportation of fireant-infested goods to prevent jump dispersal between thetwo areas and decrease probability of successful erad-ication (Drees and Gold 2003)
Acknowledgments
The authors would like to thank Chung-Chi Lin for providing most ofthe specimens for this study including the first identified samples inTaoyuan and Chiayi counties We are particularly grateful to Hwei-yuChang Hon-Tsen Yu and Chia-Hung Hsieh for helpful discussions andcomments on earlier versions of this manuscript Comments by twoanonymous reviewers greatly improved this manuscript This research
was financially supported by grants from Academia Sinica Taiwan Theuse of trade firm or corporation names in this publication is for theinformation and convenience of the reader Such use does notconstitute an official endorsement or approval by the United StatesDepartment of Agriculture or the Agricultural Research Service of anyproduct or service to the exclusion of others that may be suitable
References
Ahrens ME Ross KG and Shoemaker DD 2005 Phylogeographicstructure of the fire ant Solenopsis invicta in its native SouthAmerican range Roles of natural barriers and habitat connectivityEvolution 59 1733ndash1743
Allen CR Demarais S and Lutz RS 1994 Red imported fire antimpact on wildlife an overview Texas J Sci 46 51ndash59
Allen CR Lutz RS and Demarais S 1998 Ecological effects of theinvasive nonindigenous ant Solenopsis invicta on native verte-brates The wheels on the bus Transactions of the North AmericanWildlife and Natural Resources Conference 63 56 ndash65
Allen CR Willey RD Myers PE Horton PM and Buffa J 2000Impact of red imported fire ant infestation on northern bobwhitequail abundance trends in southeastern United States J AgrUrban Entomol 17 43ndash51
Allen CR Forys EA Rice KG and Wojcik DP 2001 Effects of fireants (Hymenoptera Formicidae) on hatching turtles and preva-lence of fire ants on sea turtle nesting beaches in Florida FloridaEntomol 84 250ndash253
Bouwma AM Ahrens ME DeHeer CJ and Shoemaker DD 2006Distribution and prevalence of Wolbachia in introduced popula-tions of the fire ant Solenopsis invicta Insect Mol Biol 15 89ndash93
Briano JA Calcaterra LA Vander Meer R Valles SM and LivoreJP 2006 New survey for the fire ant microsporidia Vairimorphainvictae and Thelohania solenopsae in southern South Americawith observations on their field persistence and prevalence of dualinfections Environ Entomol 35 1358ndash1365
Buckley A 1999 Fire ants in California Am Bee J 139 88Carroll CR and Hoffman CA 2000 The pervasive ecological effects
of invasive species Exotic and native fire ants In Invertebrates asWeb-Masters in Ecosystems (Coleman DC and Hendrix PF Eds)Wallingford Oxon UK pp 221ndash232
Carruthers RI 2003 Invasive species research in the United StatesDepartment of Agriculture ndash Agricultural Research Service PestManag Sci 59 827ndash834
Chen JSC Shen CH and Lee HJ 2006 Monogynous and polygynousred imported fire ants Solenopsis invicta Buren (HymenopteraFormicidae) in Taiwan Environ Entomol 35 167ndash172
Cornuet JM and Luikart G 1996 Description and power analysis oftwo tests for detecting recent population bottlenecks from allelefrequency data Genetics 144 2001ndash2014
Davis LR Vander Meer RK and Porter SD 2001 Red imported fireants expand their range across the West Indies Florida Entomol84 735ndash736
DeHeer CJ and Tschinkel WR 1998 The success of alternativereproductive tactics in monogyne Solenopsis invicta significancefor transitions in social organization Behav Ecol 9 130ndash135
DeHeer CJ Goodisman MAD and Ross KG 1999 Queen dispersalstrategies in the multiple-queen form of the fire ant Solenopsisinvicta Am Nat 153 660ndash675
Dieringer D and Schlccediltterer C 2002 Microsatellite analyser (MSA) aplatform independent analysis tool for large microsatellite datasets Mol Ecol Notes 3 167ndash169
Drake JA Mooney HA Castri F Groves RH Kruger MRejmnek M and Williamson M 1989 Biological Invasions AGlobal Perspective Chichester UK 550 pp
Drees BM and Vinson SB 1990 Comparison of the control ofmonogynous and polygynous forms of the red imported fire ant(Hymenoptera Formicidae) with a chloripyrifos mound drench JEntomol Sci 25 317ndash324
Table 3 Tests for genetic bottlenecks using the infinite allele model(IAM) and two-phase model (TPM 90 stepwise mutation model and10 multistep mutation model [Garza and Williamson 2001])implemented in the program BOTTLENECK (a=005)Significance at given statistic level
Population Simulatedmodel
Significancetest
Standardizeddifferences test
Wilcoxontest
Taoyuan IAM 0148 0003 0015TPM 0186 0035 0023
Chiayi IAM 0037 0022 0007TPM 0521 0293 0281
Insect Soc Vol 55 2008 Research article 63
Drees BM and Gold RE 2003 Development of integrated pestmanagement programs for the red imported fire ant (Hymeno-ACHTUNGTRENNUNGptera Formicidae) J Entomol Sci 38 170ndash180
Eubanks MD 2001 Estimates of the direct and indirect effects of redimported fire ants on biological control in field crops Biol Control21 35 ndash43
Evanno G Regnaut S and Goudet J 2005 Detecting the number ofclusters of individuals using the software STRUCTURE asimulation study Mol Ecol 14 2611ndash2620
Excoffier L Smouse PE and Quattro JM 1992 Analysis of molecularvariance inferred from metric distances among DNA haplotypesApplication to human mitochondrial DNA restriction dataGenetics 131 479ndash491
Forys EA Quistorff A and Allen CR 2001 Potential fire ant(Hymenoptera Formicidae) impact on the endangered Schausswallowtail (Lepidoptera Papilionidae) Florida Entomol 84 254ndash258
Garza JC and Williamson EG 2001 Detection of reduction inpopulation size using data from microsatellite loci Mol Ecol 10 305ndash318
Giraud T Pedersen JS and Keller L 2002 Evolution of supercoloniesthe Argentine ants of southern Europe Proc Natl Acad Sci U SA 99 6075ndash6079
Giuliano WM Allen CR Lutz RS and Demarais S 1996 Effects ofred imported fire ants on northern bobwhite chicks J WildlManage 60 309ndash313
Glancey BM Craig CH Stringer CE and Bishop PM 1973Multiple fertile queens in colonies of the imported fire antSolenopsis invicta J Georgia Entomol Soc 8 327ndash328
Goodisman MAD DeHeer CJ and Ross KG 2000 Mating behaviorof queens from multiple-queen colonies of Solenopsis invicta JInsect Behav 13 455ndash468
Goodnight KF and Queller DC 1999 Computer software forperforming likelihood tests of pedigree relationship using geneticmarkers Mol Ecol 8 1231ndash1234
Gotelli NJ and Arnett AE 2000 Biogeographic effects of red fire antinvasion Ecol Lett 3 257ndash261
Gotzek D and Ross KG 2007 Genetic regulation of colony socialorganization in fire ants An integrative overview Q Rev Biol 82 201ndash226
Goudet J 2001 FSTAT A Program to Estimate and Test GeneDiversities and Fixation Indices Version 293 http wwwunilch izeasoftwares fstathtml
Greenberg L Fletcher DJC and Vinson SB 1985 Differences inworker size and mound distribution in monogynous and polygy-nous colonies of the fire ant Solenopsis invicta Buren J KansasEntomol Soc 58 9ndash18
Hampton JO Spencer PBS Alpers DL Twigg LE WoolnoughAP Doust J Higgs T and Pluske J 2004 Molecular techniqueswildlife management and the importance of genetic populationstructure and dispersal a case study with feral pigs J Appl Ecol41 735ndash743
Henshaw MT Kunzmann N Vanderwoude C Sanetra M andCrozier RH 2005 Population genetics and history of theintroduced fire ant Solenopsis invicta Buren (HymenopteraFormicidae) in Australia Aus J Entomol 44 37ndash44
Hccedillldobler B and Wilson EO 1990 The Ants The Belknap Press ofHarvard University Press Cambridge Massachusetts 732 pp
Huang TC Chou YC and Chou HC 2004 The infestation andcontrol of the red imported fire ant in Taiwan In Proceedingsof the Symposium on the Control of the Red Imported Fire AntBureau of Animal and Plant Health Inspection and QuarantineCouncil of Agriculture Executive Yuan Taipei Taiwan pp1 ndash 13
Jaquiry J Vogel V and Keller L 2005 Multilevel genetic analyses oftwo European supercolonies of the Argentine ant Linepithemahumile Mol Ecol 14 589ndash598
Jeyaprakash A and Hoy MA 2000 Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropods Insect Mol Biol 9 393ndash405
Jouvenaz DP 1990 Approaches to biological control of fire ants in theUnited States In Applied Myrmecology A World Perspective(Vander Meer RK Jaffe K and Cedeno A Eds) Westview PressBoulder Colorado pp 620ndash627
Kaspari M 2000 Do imported fire ants impact canopy arthropodsEvidence from simple arboreal pitfall traps Southwestern Nat 45 118ndash122
Keller L and Ross KG 1998 Selfish genes a green beard in the red fireant Nature 394 573ndash575
Kemp SF deShazo RD Moffitt JE Williams DF and Buhner WA2000 Expanding habitat of the imported fire ant (Solenopsisinvicta) A public health concern J Allergy Clin Immunol 105 683ndash691
Kopachena JG Buckley AJ and Potts GA 2000 Effects of the redimported fire ant (Solenopsis invicta) on reproductive success ofbarn swallows (Hirundo rustica) in northeast Texas SouthwesternNat 45 477ndash482
Krieger MJB and Keller L 1997 Polymorphism at dinucleotidemicrosatellite loci in fire ant Solenopsis invicta populations MolEcol 6 997ndash999
Krieger MJB Ross KG Chang CWY and Keller L 1999Frequency and origin of triploidy in the fire ant Solenopsis invictaHeredity 82 142ndash150
Krieger MJB and Ross KG 2002 Identification of a major generegulating complex social behavior Science 295 328ndash332
Lee CE 2002 Evolutionary genetics of invasive species Trends EcolEvol 17 386ndash391
Lofgren CS Banks WA and Glancey BM 1975 Biology and controlof imported fire ants Annu Rev Entomol 20 1ndash30
Lofgren CS 1986 History of imported fire ants in the United States InFire Ants and Leaf Cutting Ants Biology and Management(Lofgren CS and Vander Meer RK Eds) Westview PressBoulder Colorado pp 36ndash49
Luikart G Allendorf FW Cornuet JM and Sherwin WB 1998Distortion of allele frequency distributions provides a test forrecent population bottlenecks J Hered 89 238ndash247
Luikart G and Cornuet JM 1998 Empirical evaluation of a test foridentifying recently bottlenecked populations from allele frequen-cy data Conserv Biol 12 228ndash237
MacKay WP and Fagerlund R 1997 Range expansion of the redimported fire ant Solenopsis invicta Buren (Hymenoptera For-micidae) into New Mexico and extreme western Texas ProcEntomol Soc Wash 99 757ndash758
Mank JE and Avise JC 2004 Individual organisms as units of analysisBayesian-clustering alternatives in population genetics Genet Res84 135ndash143
May GE Gelembiuk GW Panov VE Orlova MI and Lee CE2006 Molecular ecology of zebra mussel invasions Mol Ecol 15 1021ndash1031
McCubbin KI and Weiner JM 2002 Fire ants in Australia A newmedical and ecological hazard Med J Aus 176 518ndash519
Morrison LW 2000 Mechanisms of interspecific competition amongan invasive and two native fire ants Oikos 90 238ndash252
Morrison LW 2002 Long-term impacts of an arthropod-communityinvasion by the imported fire ant Solenopsis invicta Ecology 83 2337ndash2345
Nattrass R and Vanderwoude C 2001 A preliminary investigation ofthe ecological effects of red imported fire ants (Solenopsis invicta)in Brisbane Ecol Manag Resto 2 220ndash223
Nei M 1972 Genetic distance between populations Am Nat 106 283ndash292
Orr MR 1996 Host manipulation by Wolbachia is a neutral trait withinsingle populations Anim Behav 51 1183ndash1185
Patterson RS 1994 Biological control of introduced ant species InExotic Ants Biology Impact and Control of Introduced Species(Williams DF Ed) Westview Press Boulder Colorado pp 293ndash308
Piry S Luikart G and Cornuet JM 1999 BOTTLENECK acomputer program for detecting recent reductions in the effectivepopulation size using allele frequency data J Hered 90 502ndash503
64 CC Yang et al Fire ants in Taiwan
Porter SD and Savignano DA 1990 Invasion of polygyne fire antsdecimates native ants and disrupts arthropod community Ecology7 2095ndash2106
Porter SD Bhatkar A Mulder R Vinson SB and Clair DJ 1991Distribution and density of polygyne fire ants (HymenopteraFormicidae) in Texas J Econ Entomol 84 866ndash874
Porter SD Fowler HG and MacKay WP 1992 Fire ant mounddensities in the United States and Brazil (Hymenoptera Formi-cidae) J Econ Entomol 85 1154ndash1161
Porter SD Williams DF Patterson RS and Fowler HG 1997Intercontinental differences in the abundance of Solenopsis fireants (Hymenoptera Formicidae) Escape from natural enemiesEnviron Entomol 26 373ndash384
Pritchard JK Stephens M and Donnelly P 2000 Inference ofpopulation structure using multilocus genotype data Genetics155 945ndash959
Raymond M and Rousset F 1995a An exact test for populationdifferentiation Evolution 49 1280ndash1283
Raymond M and Rousset F 1995b GENEPOP (version 12)population genetics software for exact tests and ecumenicism JHered 86 248ndash249
Reuter M Pedersen JS and Keller L 2005 Loss of Wolbachiainfection during colonisation in the invasive Argentine ant Line-pithema humile Heredity 94 364ndash369
Ross KG and Fletcher DJC 1985a Genetic origin of male diploidy inthe fire ant Solenopsis invicta (Hymenoptera Formicidae) and itsevolutionary significance Evolution 39 888ndash903
Ross KG and Fletcher DJC 1985b Comparative study of genetic andsocial structure in two forms of the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 17 349ndash356
Ross KG and Fletcher DJC 1986 Diploid male productionndashasignificant colony mortality factor in the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 19 283ndash291
Ross KG and Shoemaker DD 1993 An unusual pattern of gene flowbetween the two social forms of the fire ant Solenopsis invictaEvolution 47 1595ndash1605
Ross KG Vargo EL Keller L and Trager JC 1993 Effect of afounder event on variation in the genetic sex-determining system ofthe fire ant Solenopsis invicta Genetics 135 843ndash854
Ross KG and Keller L 1995 Joint influence of gene flow and selectionon a reproductively important genetic polymorphism in the fire antSolenopsis invicta Am Nat 146 325ndash348
Ross KG Vargo EL and Keller L 1996 Social evolution in a newenvironment The case of introduced fire ants Proc Natl AcadSci U S A 93 3021ndash3025
Ross KG 1997 Multilocus evolution in fire ants Effects of selectiongene flow and recombination Genetics 145 961ndash974
Ross KG Krieger MJB Shoemaker DD Vargo EL and Keller L1997 Hierarchical analysis of genetic structure in native fire antpopulations Results from three classes of molecular markersGenetics 147 643ndash655
Ross KG and Shoemaker DD 1997 Nuclear and mitochondrialgenetic structure in two social forms of the fire ant Solenopsisinvicta Insights into transitions to an alternate social organizationHeredity 78 590ndash602
Ross KG and Keller L 1998 Genetic control of social organization inan ant Proc Natl Acad Sci U S A 95 14232ndash14237
Ross KG Shoemaker DD Krieger MJ DeHeer CJ and Keller L1999 Assessing genetic structure with multiple classes of molecularmarkers A case study involving the introduced fire ant Solenopsisinvicta Mol Biol Evol 16 525ndash543
Ross KG and Keller L 2002 Experimental conversion of colony socialorganization by manipulation of worker genotype composition infire ants (Solenopsis invicta) Behav Ecol Sociobiol 51 287ndash295
Ross KG Krieger MJB and Shoemaker DD 2003 Alternativegenetic foundations and evolutionary routes for a key socialpolymorphism in fire ants Genetics 165 1853ndash1867
Ross KG Krieger MJB Keller L and Shoemaker DD 2007Genetic variation and structure in native populations of the fire antSolenopsis invicta Evolutionary and demographic implicationsBiol J Linnean Soc 92 541ndash560
Rousset F 1997 Genetic differentiation and estimation of gene flowfrom F-statistics under isolation by distance Genetics 145 1219ndash1228
Sax DF Stachowicz JJ and Gaines SD 2005 Species InvasionsInsights into Ecology Evolution and Biogeography SinauerAssociates Sunderland Massachussetts 495 pp
Shoemaker DD and Ross KG 1996 Effects of social organization ongene flow in the fire ant Solenopsis invicta Nature 383 613ndash616
Shoemaker DD Ross KG Keller L Vargo EL and Werren JH2000 Wolbachia infections in native and introduced populations offire ants (Solenopsis spp) Insect Mol Biol 9 661ndash673
Shoemaker DD Ahrens M Sheill L Mescher M Keller L and RossKG 2003 Distribution and prevalence of Wolbachia infections innative populations of the fire ant Solenopsis invicta (HymenopteraFormicidae) Environ Entomol 32 1329ndash1336
Shoemaker DD DeHeer CJ Krieger MJB and Ross KG 2006Population genetics of the invasive fire ant Solenopsis invicta(Hymenoptera Formicidae) in the USA Ann Entomol Soc Am99 1213ndash1233
Simon C Frati F Beckenbach A Crespi B Liu H and Flook P 1994Evolution weighting and phylogenetic utility of mitochondrialgene sequences and a compilation of conserved polymerase chainreaction primers Ann Entomol Soc Am 87 651ndash701
Toom PM Cupp E Johnson CP and Griffin I 1976 Utilization ofbody reserves for minim brood development by queens of theimported fire ant Solenopsis invicta J Insect Physiol 22 217ndash220
Tsutsui ND and Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepethema humile) in its nativeand introduced ranges Evolution 55 976ndash985
Tsutsui ND Kauppinen SN Oyafuso AF and Grosberg RK 2003The distribution and evolutionary history of Wolbachia infection innative and introduced populations of the invasive argentine ant(Linepithema humile) Mol Ecol 12 3057ndash3068
Valles SM and Porter SD 2003 Identification of polygyne andmonogyne fire ant colonies (Solenopsis invicta) by multiplex PCRof Gp-9 alleles Insect Soc 50 199ndash200
Valles SM Oi DH Briano JA and Williams DF 2004 Simultaneousdetection of Vairimorpha invictae (Microsporidia Burenellidae)and Thelohania solenopsae (Microsporidia Thelohaniidae) in fireants by PCR Florida Entomol 87 85ndash87
Vander Meer R K and Porter SD 2001 Fate of newly-mated queensintroduced into monogyne and polygyne Solenopsis invicta (Hy-menoptera Formicidae) colonies Ann Entomol Soc Am 94 289ndash297
Vander Meer RK Jaffe K and Cedeno A 1990 Applied Myrmecol-ogy A World Perspective Westview Press Boulder CO 741 pp
Vinson SB 1994 Impact and invasion of Solenopsis invicta (Buren) onnative food webs In Exotic Ants Biology Impact and Control ofIntroduced Species (Williams DF Ed) Westview Press BoulderColorado pp 293ndash308
Vinson SB 1997 Invasion of the red imported fire ant (HymenopteraFormicidae) Spread biology and impact Am Entomol 43 23ndash29
Williams DF and Porter SD 1994 Fire ant control Science 264 1653ndash1653
Wojcik D Allen CR Brenner RJ Forys EA Jouvenaz D and LutzRS 2001 Red imported fire ants Impact on biodiversity AmEntomol 47 16ndash23
Xia X and Xie Z 2001 DAMBE Data analysis in molecular biologyand evolution J Hered 92 371ndash373
Insect Soc Vol 55 2008 Research article 65
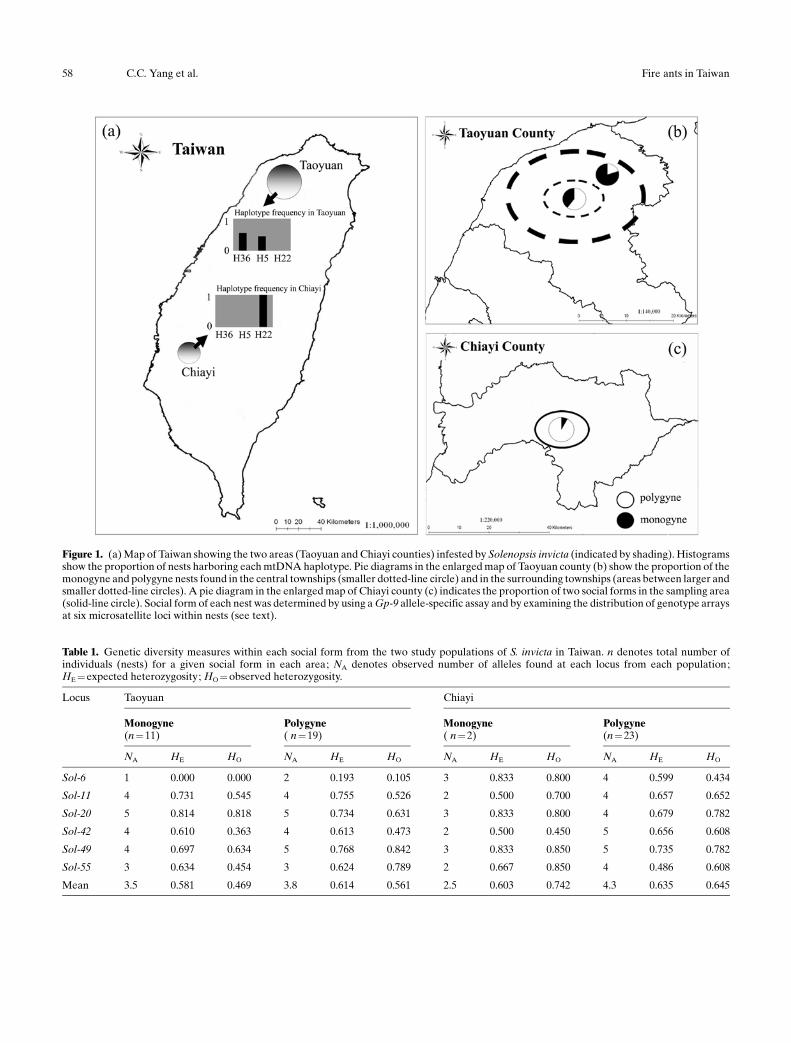
Figure 1 (a) Map of Taiwan showing the two areas (Taoyuan and Chiayi counties) infested by Solenopsis invicta (indicated by shading) Histogramsshow the proportion of nests harboring each mtDNA haplotype Pie diagrams in the enlarged map of Taoyuan county (b) show the proportion of themonogyne and polygyne nests found in the central townships (smaller dotted-line circle) and in the surrounding townships (areas between larger andsmaller dotted-line circles) A pie diagram in the enlarged map of Chiayi county (c) indicates the proportion of two social forms in the sampling area(solid-line circle) Social form of each nest was determined by using a Gp-9 allele-specific assay and by examining the distribution of genotype arraysat six microsatellite loci within nests (see text)
Table 1 Genetic diversity measures within each social form from the two study populations of S invicta in Taiwan n denotes total number ofindividuals (nests) for a given social form in each area NA denotes observed number of alleles found at each locus from each populationHE =expected heterozygosity HO = observed heterozygosity
Locus Taoyuan Chiayi
Monogyne(n= 11)
Polygyne( n=19)
Monogyne( n=2)
Polygyne(n=23)
NA HE HO NA HE HO NA HE HO NA HE HO
Sol-6 1 0000 0000 2 0193 0105 3 0833 0800 4 0599 0434
Sol-11 4 0731 0545 4 0755 0526 2 0500 0700 4 0657 0652
Sol-20 5 0814 0818 5 0734 0631 3 0833 0800 4 0679 0782
Sol-42 4 0610 0363 4 0613 0473 2 0500 0450 5 0656 0608
Sol-49 4 0697 0634 5 0768 0842 3 0833 0850 5 0735 0782
Sol-55 3 0634 0454 3 0624 0789 2 0667 0850 4 0486 0608
Mean 35 0581 0469 38 0614 0561 25 0603 0742 43 0635 0645
58 CC Yang et al Fire ants in Taiwan
Within-nest relatedness
The estimates of relatedness among workers withinmonogyne nests from each area were consistent witheach nest headed by single queen mated to a single male(Taoyuan R=071870036 Chiayi R=071970038Fig 2) Relatedness among workers within polygynenests was significantly greater than zero but significantlylower (Plt0001) than the values for monogyne nests(Taoyuan R=011220006 Chiayi R=014440007Fig 2)
Population genetic differentiation
Exact tests based on six microsatellite loci revealedsignificant differentiation between Taoyuan and Chiayipopulations (Plt002) No significant differentiation wasobserved between sympatric populations of the two socialforms in the two areas (Pgt013) Exact tests for differ-entiation using mtDNA sequence data revealed signifi-cant differentiation occurs between the two geographicpopulations (Plt00001) as well as between the two socialforms in Taoyuan (Plt004) but not between the twosocial forms in Chiayi (only one mtDNA variant occurshere)
Nuclear and mtDNA variation was partitioned byestimating FST simultaneously at both levels using theanalysis of molecular variance approach (Excoffier et al 1992) Results of these analyses for the nuclear locishowed that only the between-population variance com-ponent is significant (Plt0000001 Fig 3a) providingadditional support to the above analyses suggesting thesetwo populations are genetically distinct For analysesinvolving mtDNA data both the between-population andbetween-social form variance components are significant(both Plt005 Fig 3b)
Bayesian simulations using STRUCTURE revealedthat individuals from the Taoyuan population represent a
single distinct genetic cluster suggesting no differentia-tion between sympatric social forms exists whether usingnuclear data only or nuclear and mtDNA combinedIdentical results were obtained for individuals from theChiayi population In contrast simulations incorporatingall individuals from the two areas gave posterior proba-bilities that strongly support the existence of two distinctgenetic clusters using dataset from the nuclear loci orcombination of the two classes of markers (K=2 Fig 4)The two clusters hypothesized by the Bayesian methodcorrespond roughly to geographic populations based onaverage membership coefficientsgt05 for one of the twoposterior clusters in each of these regions The firstcluster (indicated by lighter gray) is comprised largely ofindividuals from the Taoyuan population whereas thesecond cluster (indicated by the darker gray) is comprisedmostly of the individuals from Chiayi Identical clusteringresults were obtained when simulations were run incor-porating both the microsatellite and mtDNA data(Fig 4)
Figure 2 Average relatedness (95 confidence interval) valuesamong workers within nests of the polygyne and monogyne social formsfrom Taoyuan and Chiayi counties Taiwan
Figure 3 Partitioning of genetic variance in S invicta at six micro-satellite loci (a) and mtDNA (b) estimated using analysis of molecularvariance This hierarchical approach partitioned the total variancebetween infested regions (Taoyuan and Chiayi counties) between thetwo social forms within each infested region and within the social formsin each region The percentage of the total variance partitioned at eachlevel is shown Asterisks indicate statistically significant differentiationat a given level (a=005)
Figure 4 Assignment of individuals representing Solenopsis invictafrom both study populations to two genetic clusters inferred fromSTRUCTURE simulations (average membership coefficient Q) Eachvertical bar represents the individual membership coefficients dividedinto parts proportional to the individuals proposed ancestry in eachcluster
Insect Soc Vol 55 2008 Research article 59
Isolation by distance and tests for genetic bottlenecks
No correlation exists between pairwise FST or FST(1ndash FST)values and geographic distance for individuals of thepolygyne social form in either population (TaoyuanPgt022 Chiayi Pgt048) Further no significant iso-lation by distance for the monogyne nests from Taoyuanpopulation was found (Pgt065) We did not perform theMantel tests for the Chiayi monogyne population sinceonly two nests were collected
All tests revealed a significant excess of heterozygos-ity in Taoyuan under both models except the sign test(Table 3) A significant excess of heterozygosity wasfound in Chiayi for the infinite allele model but not thetwo phase model (Table 3) Combined with a mode shiftof allele frequency distribution (Table 3 Fig 5a) theTaoyuan population appears to have undergone a recentbottleneck In contrast the most abundant alleles in theChiayi population belong to the low frequency class(lt01 Fig 5b) resulting in a normal L-shape distribu-
tion We also compared NA (number of observed allelesTable 2) and HE (expected heterozygosity Table 2)among the South American USA and Taiwanesepopulations to determine whether a genetic signaturecharacteristic of a bottleneck was evident for the latterpopulations In order to minimize the effect of populationsize we randomly selected 30 individuals from the USAand South American dataset (Shoemaker et al 2006Ross et al 2007) Results showed that the NA and HE inTaiwan and the USA populations were consistentlylower than in South American fire ant populations Twopopulations in Taiwan had slightly lower NA than inUSA but had similar HE values
Additional findings
Analyses of individual genotypes revealed that 108 ofthe 190 workers sampled from polygyne nests in Taoyuanand 11 of 230 individuals from polygyne nests in Chiayi
Table 2 Genetic diversity measures within the two study populations of S invicta in Taiwan NA denotes observed number of alleles found at eachlocus from each population HE =expected heterozygosity HO = observed heterozygosity Numbers in parentheses represent the number of privatealleles occurring in only one of the two Taiwanese populations Genetic diversity measures for two population from the USA and South Americaalso are shown (see text)
Source of collection Population Sample size Mean no of alleles NA HE HO
TaiwanTaoyuan county 30 38 23 (2) 0609 0527Chiayi county 25 48 26 (5) 0636 0660
USAMississippi 30 57 34 0651 0559western Louisiana 30 58 35 0628 0607
South AmericaFormosa Argentina 30 158 195 0835 0842Corrientes Argentina 30 158 95 0836 0826
Figure 5 Distribution of allele frequencies at six microsatellite loci for ants (single individual per nest) from the Taoyuan (a) and Chiayi populations(b)
60 CC Yang et al Fire ants in Taiwan
harbored three distinct alleles whereas all of the individ-uals sampled from monogyne nests harbored at most twodistinct alleles These results indicate that a substantialnumber of polygyne fire ants are triploid a findingconsistent with an earlier study of fire ants from theUSA (Krieger et al 1999)
Discussion
We studied the colony genetic and population structure ofthe invasive fire ant S invicta in Taiwan by collecting andanalyzing microsatellite genotypic data and mtDNAsequence data of ants collected from two separateinfested areas Our genetic analyses revealed severalpatterns which are discussed in more detail below andcan be summarized as follows 1) Estimates of within nestrelatedness were significantly greater than zero for bothsocial forms from both populations 2) No significantisolation by distance was found among nests within eachsocial form from either population 3) SignificantmtDNA but no nuclear differentiation occurs betweensympatric social forms in Taoyuan (no differentiationbetween forms at either genome is evident in Chiayi) 4)Molecular signatures of genetic bottlenecks associatedwith recent introductions are evident in both populations5) The two sampled populations Taoyuan and Chiayi arehighly genetically differentiated at both the nuclear ormtDNA genomes In addition to these general findingswe found numerous triploid individuals in ants of thepolygyne (P) but not the monogyne (M) form anddistinct geographic distribution of each of the two socialforms between the two populations
Colony social organization and patterns of intracolonyworker relatedness
The estimate of relatedness among workers withinmonogyne colonies was not significantly different from075 the predicted value for a nest headed by one singly-mated queen These data also are consistent with datafrom earlier studies of monogyne S invicta colonies inboth the USA (Ross and Fletcher 1985b) and SouthAmerica (Ross et al 1997) and suggest that the socialand mating biology of M queens is virtually invariantwith M nests representing closed societies headed by onesingly-mated reproductive queen
In contrast colony genetic structure of P social formdoes differ between native South America and intro-duced populations in the USA (Ross et al 1996) Pnests of S invicta in the native range contain relativelyfew queens which are close relatives and relatednessamong nestmate workers is significantly greater thanzero whereas P nests in the USA contain highnumbers of unrelated queens and relatedness amongnestmate workers is not significantly greater than zeroThese differences in queen number and relatedness
between the two ranges are consistent with predictionsbased on differences in the population densities of antsin these two areas (Ross et al 1996) which aresubstantially higher in the USA compared to SouthAmerica (Porter et al 1997) One consequence of thehigher population densities in the USA is that nestingsites are more often saturated such that opportunitiesfor independent founding by queens are more limitedin the introduced range (Ross et al 1996) Theseconstraints on independent founding lead to increasedselection pressure on queens to seek adoption intoexisting colonies and on resident workers of colonies toaccept such queens As queen number within thesenests increases relatedness among nestmate workers isexpected to decrease
Relatedness among workers within P nests in Taiwanis small yet significantly greater than zero and intermedi-ate to the values observed for native and USApopulations The social organization of P nests in Taiwanlikely reflects the fact that nesting sites are much lesssaturated compared to sites in the USA since thepopulation densities of ants in Taiwan currently are muchlower than those in the USA However while such adifference between the two introduced ranges are evidentnow it is unlikely to persist especially if S invictacontinues to spread in Taiwan so that habitats becomesaturated If S invicta continues to spread so that habitatsbecome more saturated future studies of the socialevolution of P populations in Taiwan could shed addi-tional light on whether such a change in habitat avail-ability does in fact lead to a shift in social organizationtowards a higher queen number within nests and lowerrelatedness among nestmates
Patterns of isolation by distance
The lack of isolation by distance among colonies of the Mform within Taoyuan was not unexpected given that mostM queens take part in aerial mating flights oftendispersing several kilometers or more before landingand attempting to found a new nest (Toom et al 1976Ross and Fletcher 1985b Ross et al 1999) Such long-distance dispersal is predicted to erase any signature ofnuclear differentiation among closely spaced nests
The lack of detectable isolation by distance within theP form at both studied sites was somewhat unexpectedconsidering the reproductive habits of P queens whichoften mate in or near their natal nests and form newcolonies by budding or by entering into nearby existingnests (Shoemaker and Ross 1996 Ross et al 1997 Rossand Shoemaker 1997) One explanation for the lack ofdetectable isolation by distance is simply that malesdisperse rather long distances to mate with P queensUnder this scenario one would predict that long distancemale-mediated gene flow would result in homogenizationof allele frequencies at nuclear loci yet significantmicrogeographic mtDNA structure would remain espe-
Insect Soc Vol 55 2008 Research article 61
cially if queen dispersal is limited However no suchpattern was found Significant isolation by distance wasabsent for both the nuclear and mtDNA markers Anadditional explanation for these results is that thesignature of isolation by distance predicted to emergesolely from the natural dispersal biology of these ants isconfounded by the fact that these ants are frequentlyinadvertently transported relatively great distances byhumans Such frequent inadvertent human transport maybe a common characteristic of invasive ants found in closeproximity to human habitats and thus play a role inshaping the genetic structure in introduced areas Finallyit is possible that the lack of a detectable signal of isolationby distance is due to the extremely recent introduction offire ants into Taiwan with insufficient time having passedfor populations to achieve migrationdrift equilibrium
Gene flow between sympatric social forms
The pronounced mtDNA differentiation between sym-patric social forms in Taoyuan is consistent with resultsfrom earlier studies in both South America and the USAshowing that interform gene flow mediated by queens israre or absent Queens of each social form rarely succeedin becoming reproductives in colonies of the alternateform as a result of behavioral decisions (attributed largelyto differences in genotypes of queens at the single geneGp-9) made by both queens and workers (Shoemaker andRoss 1996 Ross and Shoemaker 1997 Keller and Ross1998 DeHeer et al 1999 Goodisman et al 2000 Rossand Keller 2002) The contrasting weak or nonexistentinterform divergence at nuclear genes also is consistentwith studies of S invicta in the USA showing that queensof the P form predominantly mate with males of the Mform where the two forms co-occur (Ross and Shoe-maker 1993 Ross and Keller 1995 Shoemaker andRoss 1996) In the USA most males in P colonies aresterile diploid males which means that P queens relylargely on immigrant M males for mating opportunities(Ross and Fletcher 1985a Ross and Shoemaker 1993)Such interform matings act as a homogenizing force onlyfor nuclear markers The similar frequency of triploidworkers in P nests in Taiwan and the USA suggestsmany diploid males also occur in Taiwan as well (Rosset al 1993 1999 Krieger and Keller 1997 Krieger et al 1999) which means that P queens here must also matewith M males The finding of contrasting patterns ofdifferentiation between the two genomes parallels resultsfrom earlier studies involving S invicta in both the USAand South America (Shoemaker and Ross 1996 Rosset al 1997 Shoemaker et al 2006) One final caveathowever is that we cannot completely rule out thepossibility that the lack of detectable nuclear differ-entiation is due to insufficient time having passed fordivergence to occur despite restricted gene flow betweenthe two forms
Understanding the distributions of the two socialforms in the two infested areas in Taiwan each of whichexhibits rather distinct dispersal strategies is importantfor developing effective control strategies of fire ants inthese infested areas (Porter et al 1991) The distribu-tional pattern of social forms in Taoyuan suggests that theexpansion of this population has involved persistentoutward movement of ants from the core infestationcoupled with frequent long distance movement of Mreproductives into outlying areas (Fig 1b) Based onthese results we recommend that bait broadcasting andsurveillance efforts in Taoyuan should include not onlythe currently infested areas but also should include anarea extending well beyond the infested areas In contrastto the distributional pattern of social forms in Taoyuanthe large infested area in Chiayi consists mostly of P nests(92 of colonies collected Fig 1c) However despitethis difference we suggest a similar eradication strategyshould be employed This is because even thoughcomprehensive bait broadcasting within the entire areamay be more effective than for Taoyuan because of thelow queen vagility and relatively slow rate of reinvasionby budding of polygynous queens (Porter et al 1991)some M colonies do occur in Chiayi and there also ispotential for additional M colonies to arise locally fromthe P social form a possibility that is especially morelikely in areas of low density (Ross and Shoemaker 1997DeHeer and Tschinkel 1998 DeHeer et al 1999)
Genetic bottlenecks in Taoyuan and Chiayi populations
Simulations and statistical tests using the program BOT-TLENECK provide evidence of a significant geneticbottleneck associated with the introduction of fire antsinto Taoyuan with only one exception (Table 3) How-ever this single exception is based on a sign test whichhas relatively low statistical power when the number ofloci is below 20 such as our study (Cornuet and Luikart1996) In Chiayi significant excess in heterozygosity usingthe infinite alleles model (IAM) was observed for all testsbut not for the two-phase model (TPM Table 3) BecauseLuikart and Cornuet (1998) suggest that implementingthe test under the IAM model may be more reliable thanother models these results coupled with finding thatallelic frequencies fit a normal L-shaped distributionprovide evidence for a genetic bottleneck Interestinglyestimates of NA and HE (Table 2) are remarkably similarfor introduced populations from Taiwan and the USAand considerably lower than those from the native SouthAmerican range While the geographic origin of ants inTaiwan is unknown the similar genetic diversity measuresbetween the introduced ranges raises the issue that theintroduction of ants into Taiwan has left only a modestfootprint on patterns of diversity if these ants originatedfrom the USA
62 CC Yang et al Fire ants in Taiwan
Genetic differentiation between Taoyuan and Chiayipopulations
All analyses of population differentiation using themicrosatellite and mtDNA data indicate that the twogeographic populations are strongly differentiated andthus likely originated from separate invasions Whilemultiple lines of evidence support this notion thestrongest evidence is the confinement each of the threemtDNA variants to only one of the two geographicpopulations Interestingly all three variants found inTaiwan are the same three variants commonly found inants from the USA (Shoemaker et al 2006) China(GenBank accession numbers DQ831670-DQ831672)and Australia (CCY and DDS unpublished results) Theoccurrence of these three variants in all recently invadedareas suggests that ants from Taiwan could have origi-nated from any of these other areas We currently aregenerating the necessary baseline data for ants from all ofthese introduced regions as well as from populations intheir native range to infer the source population andinvasion history of ants in Taiwan and these other areas
From the perspective of managing fire ants in Taiwantreating each infested area as a separate management unitwould appear more reasonable and means that scientistsprobably should focus on separate plans for eliminationof two isolated populations (Hampton et al 2004) Onthe other hand because these two areas are at high risk ofconnectivity via long distance human-mediated dispersalcare must be taken to prevent reinvasion into these areaseven after successful eradication Stringent inter-countyquarantine measures such as movement control shouldbe established to monitor the risk of transportation of fireant-infested goods to prevent jump dispersal between thetwo areas and decrease probability of successful erad-ication (Drees and Gold 2003)
Acknowledgments
The authors would like to thank Chung-Chi Lin for providing most ofthe specimens for this study including the first identified samples inTaoyuan and Chiayi counties We are particularly grateful to Hwei-yuChang Hon-Tsen Yu and Chia-Hung Hsieh for helpful discussions andcomments on earlier versions of this manuscript Comments by twoanonymous reviewers greatly improved this manuscript This research
was financially supported by grants from Academia Sinica Taiwan Theuse of trade firm or corporation names in this publication is for theinformation and convenience of the reader Such use does notconstitute an official endorsement or approval by the United StatesDepartment of Agriculture or the Agricultural Research Service of anyproduct or service to the exclusion of others that may be suitable
References
Ahrens ME Ross KG and Shoemaker DD 2005 Phylogeographicstructure of the fire ant Solenopsis invicta in its native SouthAmerican range Roles of natural barriers and habitat connectivityEvolution 59 1733ndash1743
Allen CR Demarais S and Lutz RS 1994 Red imported fire antimpact on wildlife an overview Texas J Sci 46 51ndash59
Allen CR Lutz RS and Demarais S 1998 Ecological effects of theinvasive nonindigenous ant Solenopsis invicta on native verte-brates The wheels on the bus Transactions of the North AmericanWildlife and Natural Resources Conference 63 56 ndash65
Allen CR Willey RD Myers PE Horton PM and Buffa J 2000Impact of red imported fire ant infestation on northern bobwhitequail abundance trends in southeastern United States J AgrUrban Entomol 17 43ndash51
Allen CR Forys EA Rice KG and Wojcik DP 2001 Effects of fireants (Hymenoptera Formicidae) on hatching turtles and preva-lence of fire ants on sea turtle nesting beaches in Florida FloridaEntomol 84 250ndash253
Bouwma AM Ahrens ME DeHeer CJ and Shoemaker DD 2006Distribution and prevalence of Wolbachia in introduced popula-tions of the fire ant Solenopsis invicta Insect Mol Biol 15 89ndash93
Briano JA Calcaterra LA Vander Meer R Valles SM and LivoreJP 2006 New survey for the fire ant microsporidia Vairimorphainvictae and Thelohania solenopsae in southern South Americawith observations on their field persistence and prevalence of dualinfections Environ Entomol 35 1358ndash1365
Buckley A 1999 Fire ants in California Am Bee J 139 88Carroll CR and Hoffman CA 2000 The pervasive ecological effects
of invasive species Exotic and native fire ants In Invertebrates asWeb-Masters in Ecosystems (Coleman DC and Hendrix PF Eds)Wallingford Oxon UK pp 221ndash232
Carruthers RI 2003 Invasive species research in the United StatesDepartment of Agriculture ndash Agricultural Research Service PestManag Sci 59 827ndash834
Chen JSC Shen CH and Lee HJ 2006 Monogynous and polygynousred imported fire ants Solenopsis invicta Buren (HymenopteraFormicidae) in Taiwan Environ Entomol 35 167ndash172
Cornuet JM and Luikart G 1996 Description and power analysis oftwo tests for detecting recent population bottlenecks from allelefrequency data Genetics 144 2001ndash2014
Davis LR Vander Meer RK and Porter SD 2001 Red imported fireants expand their range across the West Indies Florida Entomol84 735ndash736
DeHeer CJ and Tschinkel WR 1998 The success of alternativereproductive tactics in monogyne Solenopsis invicta significancefor transitions in social organization Behav Ecol 9 130ndash135
DeHeer CJ Goodisman MAD and Ross KG 1999 Queen dispersalstrategies in the multiple-queen form of the fire ant Solenopsisinvicta Am Nat 153 660ndash675
Dieringer D and Schlccediltterer C 2002 Microsatellite analyser (MSA) aplatform independent analysis tool for large microsatellite datasets Mol Ecol Notes 3 167ndash169
Drake JA Mooney HA Castri F Groves RH Kruger MRejmnek M and Williamson M 1989 Biological Invasions AGlobal Perspective Chichester UK 550 pp
Drees BM and Vinson SB 1990 Comparison of the control ofmonogynous and polygynous forms of the red imported fire ant(Hymenoptera Formicidae) with a chloripyrifos mound drench JEntomol Sci 25 317ndash324
Table 3 Tests for genetic bottlenecks using the infinite allele model(IAM) and two-phase model (TPM 90 stepwise mutation model and10 multistep mutation model [Garza and Williamson 2001])implemented in the program BOTTLENECK (a=005)Significance at given statistic level
Population Simulatedmodel
Significancetest
Standardizeddifferences test
Wilcoxontest
Taoyuan IAM 0148 0003 0015TPM 0186 0035 0023
Chiayi IAM 0037 0022 0007TPM 0521 0293 0281
Insect Soc Vol 55 2008 Research article 63
Drees BM and Gold RE 2003 Development of integrated pestmanagement programs for the red imported fire ant (Hymeno-ACHTUNGTRENNUNGptera Formicidae) J Entomol Sci 38 170ndash180
Eubanks MD 2001 Estimates of the direct and indirect effects of redimported fire ants on biological control in field crops Biol Control21 35 ndash43
Evanno G Regnaut S and Goudet J 2005 Detecting the number ofclusters of individuals using the software STRUCTURE asimulation study Mol Ecol 14 2611ndash2620
Excoffier L Smouse PE and Quattro JM 1992 Analysis of molecularvariance inferred from metric distances among DNA haplotypesApplication to human mitochondrial DNA restriction dataGenetics 131 479ndash491
Forys EA Quistorff A and Allen CR 2001 Potential fire ant(Hymenoptera Formicidae) impact on the endangered Schausswallowtail (Lepidoptera Papilionidae) Florida Entomol 84 254ndash258
Garza JC and Williamson EG 2001 Detection of reduction inpopulation size using data from microsatellite loci Mol Ecol 10 305ndash318
Giraud T Pedersen JS and Keller L 2002 Evolution of supercoloniesthe Argentine ants of southern Europe Proc Natl Acad Sci U SA 99 6075ndash6079
Giuliano WM Allen CR Lutz RS and Demarais S 1996 Effects ofred imported fire ants on northern bobwhite chicks J WildlManage 60 309ndash313
Glancey BM Craig CH Stringer CE and Bishop PM 1973Multiple fertile queens in colonies of the imported fire antSolenopsis invicta J Georgia Entomol Soc 8 327ndash328
Goodisman MAD DeHeer CJ and Ross KG 2000 Mating behaviorof queens from multiple-queen colonies of Solenopsis invicta JInsect Behav 13 455ndash468
Goodnight KF and Queller DC 1999 Computer software forperforming likelihood tests of pedigree relationship using geneticmarkers Mol Ecol 8 1231ndash1234
Gotelli NJ and Arnett AE 2000 Biogeographic effects of red fire antinvasion Ecol Lett 3 257ndash261
Gotzek D and Ross KG 2007 Genetic regulation of colony socialorganization in fire ants An integrative overview Q Rev Biol 82 201ndash226
Goudet J 2001 FSTAT A Program to Estimate and Test GeneDiversities and Fixation Indices Version 293 http wwwunilch izeasoftwares fstathtml
Greenberg L Fletcher DJC and Vinson SB 1985 Differences inworker size and mound distribution in monogynous and polygy-nous colonies of the fire ant Solenopsis invicta Buren J KansasEntomol Soc 58 9ndash18
Hampton JO Spencer PBS Alpers DL Twigg LE WoolnoughAP Doust J Higgs T and Pluske J 2004 Molecular techniqueswildlife management and the importance of genetic populationstructure and dispersal a case study with feral pigs J Appl Ecol41 735ndash743
Henshaw MT Kunzmann N Vanderwoude C Sanetra M andCrozier RH 2005 Population genetics and history of theintroduced fire ant Solenopsis invicta Buren (HymenopteraFormicidae) in Australia Aus J Entomol 44 37ndash44
Hccedillldobler B and Wilson EO 1990 The Ants The Belknap Press ofHarvard University Press Cambridge Massachusetts 732 pp
Huang TC Chou YC and Chou HC 2004 The infestation andcontrol of the red imported fire ant in Taiwan In Proceedingsof the Symposium on the Control of the Red Imported Fire AntBureau of Animal and Plant Health Inspection and QuarantineCouncil of Agriculture Executive Yuan Taipei Taiwan pp1 ndash 13
Jaquiry J Vogel V and Keller L 2005 Multilevel genetic analyses oftwo European supercolonies of the Argentine ant Linepithemahumile Mol Ecol 14 589ndash598
Jeyaprakash A and Hoy MA 2000 Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropods Insect Mol Biol 9 393ndash405
Jouvenaz DP 1990 Approaches to biological control of fire ants in theUnited States In Applied Myrmecology A World Perspective(Vander Meer RK Jaffe K and Cedeno A Eds) Westview PressBoulder Colorado pp 620ndash627
Kaspari M 2000 Do imported fire ants impact canopy arthropodsEvidence from simple arboreal pitfall traps Southwestern Nat 45 118ndash122
Keller L and Ross KG 1998 Selfish genes a green beard in the red fireant Nature 394 573ndash575
Kemp SF deShazo RD Moffitt JE Williams DF and Buhner WA2000 Expanding habitat of the imported fire ant (Solenopsisinvicta) A public health concern J Allergy Clin Immunol 105 683ndash691
Kopachena JG Buckley AJ and Potts GA 2000 Effects of the redimported fire ant (Solenopsis invicta) on reproductive success ofbarn swallows (Hirundo rustica) in northeast Texas SouthwesternNat 45 477ndash482
Krieger MJB and Keller L 1997 Polymorphism at dinucleotidemicrosatellite loci in fire ant Solenopsis invicta populations MolEcol 6 997ndash999
Krieger MJB Ross KG Chang CWY and Keller L 1999Frequency and origin of triploidy in the fire ant Solenopsis invictaHeredity 82 142ndash150
Krieger MJB and Ross KG 2002 Identification of a major generegulating complex social behavior Science 295 328ndash332
Lee CE 2002 Evolutionary genetics of invasive species Trends EcolEvol 17 386ndash391
Lofgren CS Banks WA and Glancey BM 1975 Biology and controlof imported fire ants Annu Rev Entomol 20 1ndash30
Lofgren CS 1986 History of imported fire ants in the United States InFire Ants and Leaf Cutting Ants Biology and Management(Lofgren CS and Vander Meer RK Eds) Westview PressBoulder Colorado pp 36ndash49
Luikart G Allendorf FW Cornuet JM and Sherwin WB 1998Distortion of allele frequency distributions provides a test forrecent population bottlenecks J Hered 89 238ndash247
Luikart G and Cornuet JM 1998 Empirical evaluation of a test foridentifying recently bottlenecked populations from allele frequen-cy data Conserv Biol 12 228ndash237
MacKay WP and Fagerlund R 1997 Range expansion of the redimported fire ant Solenopsis invicta Buren (Hymenoptera For-micidae) into New Mexico and extreme western Texas ProcEntomol Soc Wash 99 757ndash758
Mank JE and Avise JC 2004 Individual organisms as units of analysisBayesian-clustering alternatives in population genetics Genet Res84 135ndash143
May GE Gelembiuk GW Panov VE Orlova MI and Lee CE2006 Molecular ecology of zebra mussel invasions Mol Ecol 15 1021ndash1031
McCubbin KI and Weiner JM 2002 Fire ants in Australia A newmedical and ecological hazard Med J Aus 176 518ndash519
Morrison LW 2000 Mechanisms of interspecific competition amongan invasive and two native fire ants Oikos 90 238ndash252
Morrison LW 2002 Long-term impacts of an arthropod-communityinvasion by the imported fire ant Solenopsis invicta Ecology 83 2337ndash2345
Nattrass R and Vanderwoude C 2001 A preliminary investigation ofthe ecological effects of red imported fire ants (Solenopsis invicta)in Brisbane Ecol Manag Resto 2 220ndash223
Nei M 1972 Genetic distance between populations Am Nat 106 283ndash292
Orr MR 1996 Host manipulation by Wolbachia is a neutral trait withinsingle populations Anim Behav 51 1183ndash1185
Patterson RS 1994 Biological control of introduced ant species InExotic Ants Biology Impact and Control of Introduced Species(Williams DF Ed) Westview Press Boulder Colorado pp 293ndash308
Piry S Luikart G and Cornuet JM 1999 BOTTLENECK acomputer program for detecting recent reductions in the effectivepopulation size using allele frequency data J Hered 90 502ndash503
64 CC Yang et al Fire ants in Taiwan
Porter SD and Savignano DA 1990 Invasion of polygyne fire antsdecimates native ants and disrupts arthropod community Ecology7 2095ndash2106
Porter SD Bhatkar A Mulder R Vinson SB and Clair DJ 1991Distribution and density of polygyne fire ants (HymenopteraFormicidae) in Texas J Econ Entomol 84 866ndash874
Porter SD Fowler HG and MacKay WP 1992 Fire ant mounddensities in the United States and Brazil (Hymenoptera Formi-cidae) J Econ Entomol 85 1154ndash1161
Porter SD Williams DF Patterson RS and Fowler HG 1997Intercontinental differences in the abundance of Solenopsis fireants (Hymenoptera Formicidae) Escape from natural enemiesEnviron Entomol 26 373ndash384
Pritchard JK Stephens M and Donnelly P 2000 Inference ofpopulation structure using multilocus genotype data Genetics155 945ndash959
Raymond M and Rousset F 1995a An exact test for populationdifferentiation Evolution 49 1280ndash1283
Raymond M and Rousset F 1995b GENEPOP (version 12)population genetics software for exact tests and ecumenicism JHered 86 248ndash249
Reuter M Pedersen JS and Keller L 2005 Loss of Wolbachiainfection during colonisation in the invasive Argentine ant Line-pithema humile Heredity 94 364ndash369
Ross KG and Fletcher DJC 1985a Genetic origin of male diploidy inthe fire ant Solenopsis invicta (Hymenoptera Formicidae) and itsevolutionary significance Evolution 39 888ndash903
Ross KG and Fletcher DJC 1985b Comparative study of genetic andsocial structure in two forms of the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 17 349ndash356
Ross KG and Fletcher DJC 1986 Diploid male productionndashasignificant colony mortality factor in the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 19 283ndash291
Ross KG and Shoemaker DD 1993 An unusual pattern of gene flowbetween the two social forms of the fire ant Solenopsis invictaEvolution 47 1595ndash1605
Ross KG Vargo EL Keller L and Trager JC 1993 Effect of afounder event on variation in the genetic sex-determining system ofthe fire ant Solenopsis invicta Genetics 135 843ndash854
Ross KG and Keller L 1995 Joint influence of gene flow and selectionon a reproductively important genetic polymorphism in the fire antSolenopsis invicta Am Nat 146 325ndash348
Ross KG Vargo EL and Keller L 1996 Social evolution in a newenvironment The case of introduced fire ants Proc Natl AcadSci U S A 93 3021ndash3025
Ross KG 1997 Multilocus evolution in fire ants Effects of selectiongene flow and recombination Genetics 145 961ndash974
Ross KG Krieger MJB Shoemaker DD Vargo EL and Keller L1997 Hierarchical analysis of genetic structure in native fire antpopulations Results from three classes of molecular markersGenetics 147 643ndash655
Ross KG and Shoemaker DD 1997 Nuclear and mitochondrialgenetic structure in two social forms of the fire ant Solenopsisinvicta Insights into transitions to an alternate social organizationHeredity 78 590ndash602
Ross KG and Keller L 1998 Genetic control of social organization inan ant Proc Natl Acad Sci U S A 95 14232ndash14237
Ross KG Shoemaker DD Krieger MJ DeHeer CJ and Keller L1999 Assessing genetic structure with multiple classes of molecularmarkers A case study involving the introduced fire ant Solenopsisinvicta Mol Biol Evol 16 525ndash543
Ross KG and Keller L 2002 Experimental conversion of colony socialorganization by manipulation of worker genotype composition infire ants (Solenopsis invicta) Behav Ecol Sociobiol 51 287ndash295
Ross KG Krieger MJB and Shoemaker DD 2003 Alternativegenetic foundations and evolutionary routes for a key socialpolymorphism in fire ants Genetics 165 1853ndash1867
Ross KG Krieger MJB Keller L and Shoemaker DD 2007Genetic variation and structure in native populations of the fire antSolenopsis invicta Evolutionary and demographic implicationsBiol J Linnean Soc 92 541ndash560
Rousset F 1997 Genetic differentiation and estimation of gene flowfrom F-statistics under isolation by distance Genetics 145 1219ndash1228
Sax DF Stachowicz JJ and Gaines SD 2005 Species InvasionsInsights into Ecology Evolution and Biogeography SinauerAssociates Sunderland Massachussetts 495 pp
Shoemaker DD and Ross KG 1996 Effects of social organization ongene flow in the fire ant Solenopsis invicta Nature 383 613ndash616
Shoemaker DD Ross KG Keller L Vargo EL and Werren JH2000 Wolbachia infections in native and introduced populations offire ants (Solenopsis spp) Insect Mol Biol 9 661ndash673
Shoemaker DD Ahrens M Sheill L Mescher M Keller L and RossKG 2003 Distribution and prevalence of Wolbachia infections innative populations of the fire ant Solenopsis invicta (HymenopteraFormicidae) Environ Entomol 32 1329ndash1336
Shoemaker DD DeHeer CJ Krieger MJB and Ross KG 2006Population genetics of the invasive fire ant Solenopsis invicta(Hymenoptera Formicidae) in the USA Ann Entomol Soc Am99 1213ndash1233
Simon C Frati F Beckenbach A Crespi B Liu H and Flook P 1994Evolution weighting and phylogenetic utility of mitochondrialgene sequences and a compilation of conserved polymerase chainreaction primers Ann Entomol Soc Am 87 651ndash701
Toom PM Cupp E Johnson CP and Griffin I 1976 Utilization ofbody reserves for minim brood development by queens of theimported fire ant Solenopsis invicta J Insect Physiol 22 217ndash220
Tsutsui ND and Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepethema humile) in its nativeand introduced ranges Evolution 55 976ndash985
Tsutsui ND Kauppinen SN Oyafuso AF and Grosberg RK 2003The distribution and evolutionary history of Wolbachia infection innative and introduced populations of the invasive argentine ant(Linepithema humile) Mol Ecol 12 3057ndash3068
Valles SM and Porter SD 2003 Identification of polygyne andmonogyne fire ant colonies (Solenopsis invicta) by multiplex PCRof Gp-9 alleles Insect Soc 50 199ndash200
Valles SM Oi DH Briano JA and Williams DF 2004 Simultaneousdetection of Vairimorpha invictae (Microsporidia Burenellidae)and Thelohania solenopsae (Microsporidia Thelohaniidae) in fireants by PCR Florida Entomol 87 85ndash87
Vander Meer R K and Porter SD 2001 Fate of newly-mated queensintroduced into monogyne and polygyne Solenopsis invicta (Hy-menoptera Formicidae) colonies Ann Entomol Soc Am 94 289ndash297
Vander Meer RK Jaffe K and Cedeno A 1990 Applied Myrmecol-ogy A World Perspective Westview Press Boulder CO 741 pp
Vinson SB 1994 Impact and invasion of Solenopsis invicta (Buren) onnative food webs In Exotic Ants Biology Impact and Control ofIntroduced Species (Williams DF Ed) Westview Press BoulderColorado pp 293ndash308
Vinson SB 1997 Invasion of the red imported fire ant (HymenopteraFormicidae) Spread biology and impact Am Entomol 43 23ndash29
Williams DF and Porter SD 1994 Fire ant control Science 264 1653ndash1653
Wojcik D Allen CR Brenner RJ Forys EA Jouvenaz D and LutzRS 2001 Red imported fire ants Impact on biodiversity AmEntomol 47 16ndash23
Xia X and Xie Z 2001 DAMBE Data analysis in molecular biologyand evolution J Hered 92 371ndash373
Insect Soc Vol 55 2008 Research article 65

Within-nest relatedness
The estimates of relatedness among workers withinmonogyne nests from each area were consistent witheach nest headed by single queen mated to a single male(Taoyuan R=071870036 Chiayi R=071970038Fig 2) Relatedness among workers within polygynenests was significantly greater than zero but significantlylower (Plt0001) than the values for monogyne nests(Taoyuan R=011220006 Chiayi R=014440007Fig 2)
Population genetic differentiation
Exact tests based on six microsatellite loci revealedsignificant differentiation between Taoyuan and Chiayipopulations (Plt002) No significant differentiation wasobserved between sympatric populations of the two socialforms in the two areas (Pgt013) Exact tests for differ-entiation using mtDNA sequence data revealed signifi-cant differentiation occurs between the two geographicpopulations (Plt00001) as well as between the two socialforms in Taoyuan (Plt004) but not between the twosocial forms in Chiayi (only one mtDNA variant occurshere)
Nuclear and mtDNA variation was partitioned byestimating FST simultaneously at both levels using theanalysis of molecular variance approach (Excoffier et al 1992) Results of these analyses for the nuclear locishowed that only the between-population variance com-ponent is significant (Plt0000001 Fig 3a) providingadditional support to the above analyses suggesting thesetwo populations are genetically distinct For analysesinvolving mtDNA data both the between-population andbetween-social form variance components are significant(both Plt005 Fig 3b)
Bayesian simulations using STRUCTURE revealedthat individuals from the Taoyuan population represent a
single distinct genetic cluster suggesting no differentia-tion between sympatric social forms exists whether usingnuclear data only or nuclear and mtDNA combinedIdentical results were obtained for individuals from theChiayi population In contrast simulations incorporatingall individuals from the two areas gave posterior proba-bilities that strongly support the existence of two distinctgenetic clusters using dataset from the nuclear loci orcombination of the two classes of markers (K=2 Fig 4)The two clusters hypothesized by the Bayesian methodcorrespond roughly to geographic populations based onaverage membership coefficientsgt05 for one of the twoposterior clusters in each of these regions The firstcluster (indicated by lighter gray) is comprised largely ofindividuals from the Taoyuan population whereas thesecond cluster (indicated by the darker gray) is comprisedmostly of the individuals from Chiayi Identical clusteringresults were obtained when simulations were run incor-porating both the microsatellite and mtDNA data(Fig 4)
Figure 2 Average relatedness (95 confidence interval) valuesamong workers within nests of the polygyne and monogyne social formsfrom Taoyuan and Chiayi counties Taiwan
Figure 3 Partitioning of genetic variance in S invicta at six micro-satellite loci (a) and mtDNA (b) estimated using analysis of molecularvariance This hierarchical approach partitioned the total variancebetween infested regions (Taoyuan and Chiayi counties) between thetwo social forms within each infested region and within the social formsin each region The percentage of the total variance partitioned at eachlevel is shown Asterisks indicate statistically significant differentiationat a given level (a=005)
Figure 4 Assignment of individuals representing Solenopsis invictafrom both study populations to two genetic clusters inferred fromSTRUCTURE simulations (average membership coefficient Q) Eachvertical bar represents the individual membership coefficients dividedinto parts proportional to the individuals proposed ancestry in eachcluster
Insect Soc Vol 55 2008 Research article 59
Isolation by distance and tests for genetic bottlenecks
No correlation exists between pairwise FST or FST(1ndash FST)values and geographic distance for individuals of thepolygyne social form in either population (TaoyuanPgt022 Chiayi Pgt048) Further no significant iso-lation by distance for the monogyne nests from Taoyuanpopulation was found (Pgt065) We did not perform theMantel tests for the Chiayi monogyne population sinceonly two nests were collected
All tests revealed a significant excess of heterozygos-ity in Taoyuan under both models except the sign test(Table 3) A significant excess of heterozygosity wasfound in Chiayi for the infinite allele model but not thetwo phase model (Table 3) Combined with a mode shiftof allele frequency distribution (Table 3 Fig 5a) theTaoyuan population appears to have undergone a recentbottleneck In contrast the most abundant alleles in theChiayi population belong to the low frequency class(lt01 Fig 5b) resulting in a normal L-shape distribu-
tion We also compared NA (number of observed allelesTable 2) and HE (expected heterozygosity Table 2)among the South American USA and Taiwanesepopulations to determine whether a genetic signaturecharacteristic of a bottleneck was evident for the latterpopulations In order to minimize the effect of populationsize we randomly selected 30 individuals from the USAand South American dataset (Shoemaker et al 2006Ross et al 2007) Results showed that the NA and HE inTaiwan and the USA populations were consistentlylower than in South American fire ant populations Twopopulations in Taiwan had slightly lower NA than inUSA but had similar HE values
Additional findings
Analyses of individual genotypes revealed that 108 ofthe 190 workers sampled from polygyne nests in Taoyuanand 11 of 230 individuals from polygyne nests in Chiayi
Table 2 Genetic diversity measures within the two study populations of S invicta in Taiwan NA denotes observed number of alleles found at eachlocus from each population HE =expected heterozygosity HO = observed heterozygosity Numbers in parentheses represent the number of privatealleles occurring in only one of the two Taiwanese populations Genetic diversity measures for two population from the USA and South Americaalso are shown (see text)
Source of collection Population Sample size Mean no of alleles NA HE HO
TaiwanTaoyuan county 30 38 23 (2) 0609 0527Chiayi county 25 48 26 (5) 0636 0660
USAMississippi 30 57 34 0651 0559western Louisiana 30 58 35 0628 0607
South AmericaFormosa Argentina 30 158 195 0835 0842Corrientes Argentina 30 158 95 0836 0826
Figure 5 Distribution of allele frequencies at six microsatellite loci for ants (single individual per nest) from the Taoyuan (a) and Chiayi populations(b)
60 CC Yang et al Fire ants in Taiwan
harbored three distinct alleles whereas all of the individ-uals sampled from monogyne nests harbored at most twodistinct alleles These results indicate that a substantialnumber of polygyne fire ants are triploid a findingconsistent with an earlier study of fire ants from theUSA (Krieger et al 1999)
Discussion
We studied the colony genetic and population structure ofthe invasive fire ant S invicta in Taiwan by collecting andanalyzing microsatellite genotypic data and mtDNAsequence data of ants collected from two separateinfested areas Our genetic analyses revealed severalpatterns which are discussed in more detail below andcan be summarized as follows 1) Estimates of within nestrelatedness were significantly greater than zero for bothsocial forms from both populations 2) No significantisolation by distance was found among nests within eachsocial form from either population 3) SignificantmtDNA but no nuclear differentiation occurs betweensympatric social forms in Taoyuan (no differentiationbetween forms at either genome is evident in Chiayi) 4)Molecular signatures of genetic bottlenecks associatedwith recent introductions are evident in both populations5) The two sampled populations Taoyuan and Chiayi arehighly genetically differentiated at both the nuclear ormtDNA genomes In addition to these general findingswe found numerous triploid individuals in ants of thepolygyne (P) but not the monogyne (M) form anddistinct geographic distribution of each of the two socialforms between the two populations
Colony social organization and patterns of intracolonyworker relatedness
The estimate of relatedness among workers withinmonogyne colonies was not significantly different from075 the predicted value for a nest headed by one singly-mated queen These data also are consistent with datafrom earlier studies of monogyne S invicta colonies inboth the USA (Ross and Fletcher 1985b) and SouthAmerica (Ross et al 1997) and suggest that the socialand mating biology of M queens is virtually invariantwith M nests representing closed societies headed by onesingly-mated reproductive queen
In contrast colony genetic structure of P social formdoes differ between native South America and intro-duced populations in the USA (Ross et al 1996) Pnests of S invicta in the native range contain relativelyfew queens which are close relatives and relatednessamong nestmate workers is significantly greater thanzero whereas P nests in the USA contain highnumbers of unrelated queens and relatedness amongnestmate workers is not significantly greater than zeroThese differences in queen number and relatedness
between the two ranges are consistent with predictionsbased on differences in the population densities of antsin these two areas (Ross et al 1996) which aresubstantially higher in the USA compared to SouthAmerica (Porter et al 1997) One consequence of thehigher population densities in the USA is that nestingsites are more often saturated such that opportunitiesfor independent founding by queens are more limitedin the introduced range (Ross et al 1996) Theseconstraints on independent founding lead to increasedselection pressure on queens to seek adoption intoexisting colonies and on resident workers of colonies toaccept such queens As queen number within thesenests increases relatedness among nestmate workers isexpected to decrease
Relatedness among workers within P nests in Taiwanis small yet significantly greater than zero and intermedi-ate to the values observed for native and USApopulations The social organization of P nests in Taiwanlikely reflects the fact that nesting sites are much lesssaturated compared to sites in the USA since thepopulation densities of ants in Taiwan currently are muchlower than those in the USA However while such adifference between the two introduced ranges are evidentnow it is unlikely to persist especially if S invictacontinues to spread in Taiwan so that habitats becomesaturated If S invicta continues to spread so that habitatsbecome more saturated future studies of the socialevolution of P populations in Taiwan could shed addi-tional light on whether such a change in habitat avail-ability does in fact lead to a shift in social organizationtowards a higher queen number within nests and lowerrelatedness among nestmates
Patterns of isolation by distance
The lack of isolation by distance among colonies of the Mform within Taoyuan was not unexpected given that mostM queens take part in aerial mating flights oftendispersing several kilometers or more before landingand attempting to found a new nest (Toom et al 1976Ross and Fletcher 1985b Ross et al 1999) Such long-distance dispersal is predicted to erase any signature ofnuclear differentiation among closely spaced nests
The lack of detectable isolation by distance within theP form at both studied sites was somewhat unexpectedconsidering the reproductive habits of P queens whichoften mate in or near their natal nests and form newcolonies by budding or by entering into nearby existingnests (Shoemaker and Ross 1996 Ross et al 1997 Rossand Shoemaker 1997) One explanation for the lack ofdetectable isolation by distance is simply that malesdisperse rather long distances to mate with P queensUnder this scenario one would predict that long distancemale-mediated gene flow would result in homogenizationof allele frequencies at nuclear loci yet significantmicrogeographic mtDNA structure would remain espe-
Insect Soc Vol 55 2008 Research article 61
cially if queen dispersal is limited However no suchpattern was found Significant isolation by distance wasabsent for both the nuclear and mtDNA markers Anadditional explanation for these results is that thesignature of isolation by distance predicted to emergesolely from the natural dispersal biology of these ants isconfounded by the fact that these ants are frequentlyinadvertently transported relatively great distances byhumans Such frequent inadvertent human transport maybe a common characteristic of invasive ants found in closeproximity to human habitats and thus play a role inshaping the genetic structure in introduced areas Finallyit is possible that the lack of a detectable signal of isolationby distance is due to the extremely recent introduction offire ants into Taiwan with insufficient time having passedfor populations to achieve migrationdrift equilibrium
Gene flow between sympatric social forms
The pronounced mtDNA differentiation between sym-patric social forms in Taoyuan is consistent with resultsfrom earlier studies in both South America and the USAshowing that interform gene flow mediated by queens israre or absent Queens of each social form rarely succeedin becoming reproductives in colonies of the alternateform as a result of behavioral decisions (attributed largelyto differences in genotypes of queens at the single geneGp-9) made by both queens and workers (Shoemaker andRoss 1996 Ross and Shoemaker 1997 Keller and Ross1998 DeHeer et al 1999 Goodisman et al 2000 Rossand Keller 2002) The contrasting weak or nonexistentinterform divergence at nuclear genes also is consistentwith studies of S invicta in the USA showing that queensof the P form predominantly mate with males of the Mform where the two forms co-occur (Ross and Shoe-maker 1993 Ross and Keller 1995 Shoemaker andRoss 1996) In the USA most males in P colonies aresterile diploid males which means that P queens relylargely on immigrant M males for mating opportunities(Ross and Fletcher 1985a Ross and Shoemaker 1993)Such interform matings act as a homogenizing force onlyfor nuclear markers The similar frequency of triploidworkers in P nests in Taiwan and the USA suggestsmany diploid males also occur in Taiwan as well (Rosset al 1993 1999 Krieger and Keller 1997 Krieger et al 1999) which means that P queens here must also matewith M males The finding of contrasting patterns ofdifferentiation between the two genomes parallels resultsfrom earlier studies involving S invicta in both the USAand South America (Shoemaker and Ross 1996 Rosset al 1997 Shoemaker et al 2006) One final caveathowever is that we cannot completely rule out thepossibility that the lack of detectable nuclear differ-entiation is due to insufficient time having passed fordivergence to occur despite restricted gene flow betweenthe two forms
Understanding the distributions of the two socialforms in the two infested areas in Taiwan each of whichexhibits rather distinct dispersal strategies is importantfor developing effective control strategies of fire ants inthese infested areas (Porter et al 1991) The distribu-tional pattern of social forms in Taoyuan suggests that theexpansion of this population has involved persistentoutward movement of ants from the core infestationcoupled with frequent long distance movement of Mreproductives into outlying areas (Fig 1b) Based onthese results we recommend that bait broadcasting andsurveillance efforts in Taoyuan should include not onlythe currently infested areas but also should include anarea extending well beyond the infested areas In contrastto the distributional pattern of social forms in Taoyuanthe large infested area in Chiayi consists mostly of P nests(92 of colonies collected Fig 1c) However despitethis difference we suggest a similar eradication strategyshould be employed This is because even thoughcomprehensive bait broadcasting within the entire areamay be more effective than for Taoyuan because of thelow queen vagility and relatively slow rate of reinvasionby budding of polygynous queens (Porter et al 1991)some M colonies do occur in Chiayi and there also ispotential for additional M colonies to arise locally fromthe P social form a possibility that is especially morelikely in areas of low density (Ross and Shoemaker 1997DeHeer and Tschinkel 1998 DeHeer et al 1999)
Genetic bottlenecks in Taoyuan and Chiayi populations
Simulations and statistical tests using the program BOT-TLENECK provide evidence of a significant geneticbottleneck associated with the introduction of fire antsinto Taoyuan with only one exception (Table 3) How-ever this single exception is based on a sign test whichhas relatively low statistical power when the number ofloci is below 20 such as our study (Cornuet and Luikart1996) In Chiayi significant excess in heterozygosity usingthe infinite alleles model (IAM) was observed for all testsbut not for the two-phase model (TPM Table 3) BecauseLuikart and Cornuet (1998) suggest that implementingthe test under the IAM model may be more reliable thanother models these results coupled with finding thatallelic frequencies fit a normal L-shaped distributionprovide evidence for a genetic bottleneck Interestinglyestimates of NA and HE (Table 2) are remarkably similarfor introduced populations from Taiwan and the USAand considerably lower than those from the native SouthAmerican range While the geographic origin of ants inTaiwan is unknown the similar genetic diversity measuresbetween the introduced ranges raises the issue that theintroduction of ants into Taiwan has left only a modestfootprint on patterns of diversity if these ants originatedfrom the USA
62 CC Yang et al Fire ants in Taiwan
Genetic differentiation between Taoyuan and Chiayipopulations
All analyses of population differentiation using themicrosatellite and mtDNA data indicate that the twogeographic populations are strongly differentiated andthus likely originated from separate invasions Whilemultiple lines of evidence support this notion thestrongest evidence is the confinement each of the threemtDNA variants to only one of the two geographicpopulations Interestingly all three variants found inTaiwan are the same three variants commonly found inants from the USA (Shoemaker et al 2006) China(GenBank accession numbers DQ831670-DQ831672)and Australia (CCY and DDS unpublished results) Theoccurrence of these three variants in all recently invadedareas suggests that ants from Taiwan could have origi-nated from any of these other areas We currently aregenerating the necessary baseline data for ants from all ofthese introduced regions as well as from populations intheir native range to infer the source population andinvasion history of ants in Taiwan and these other areas
From the perspective of managing fire ants in Taiwantreating each infested area as a separate management unitwould appear more reasonable and means that scientistsprobably should focus on separate plans for eliminationof two isolated populations (Hampton et al 2004) Onthe other hand because these two areas are at high risk ofconnectivity via long distance human-mediated dispersalcare must be taken to prevent reinvasion into these areaseven after successful eradication Stringent inter-countyquarantine measures such as movement control shouldbe established to monitor the risk of transportation of fireant-infested goods to prevent jump dispersal between thetwo areas and decrease probability of successful erad-ication (Drees and Gold 2003)
Acknowledgments
The authors would like to thank Chung-Chi Lin for providing most ofthe specimens for this study including the first identified samples inTaoyuan and Chiayi counties We are particularly grateful to Hwei-yuChang Hon-Tsen Yu and Chia-Hung Hsieh for helpful discussions andcomments on earlier versions of this manuscript Comments by twoanonymous reviewers greatly improved this manuscript This research
was financially supported by grants from Academia Sinica Taiwan Theuse of trade firm or corporation names in this publication is for theinformation and convenience of the reader Such use does notconstitute an official endorsement or approval by the United StatesDepartment of Agriculture or the Agricultural Research Service of anyproduct or service to the exclusion of others that may be suitable
References
Ahrens ME Ross KG and Shoemaker DD 2005 Phylogeographicstructure of the fire ant Solenopsis invicta in its native SouthAmerican range Roles of natural barriers and habitat connectivityEvolution 59 1733ndash1743
Allen CR Demarais S and Lutz RS 1994 Red imported fire antimpact on wildlife an overview Texas J Sci 46 51ndash59
Allen CR Lutz RS and Demarais S 1998 Ecological effects of theinvasive nonindigenous ant Solenopsis invicta on native verte-brates The wheels on the bus Transactions of the North AmericanWildlife and Natural Resources Conference 63 56 ndash65
Allen CR Willey RD Myers PE Horton PM and Buffa J 2000Impact of red imported fire ant infestation on northern bobwhitequail abundance trends in southeastern United States J AgrUrban Entomol 17 43ndash51
Allen CR Forys EA Rice KG and Wojcik DP 2001 Effects of fireants (Hymenoptera Formicidae) on hatching turtles and preva-lence of fire ants on sea turtle nesting beaches in Florida FloridaEntomol 84 250ndash253
Bouwma AM Ahrens ME DeHeer CJ and Shoemaker DD 2006Distribution and prevalence of Wolbachia in introduced popula-tions of the fire ant Solenopsis invicta Insect Mol Biol 15 89ndash93
Briano JA Calcaterra LA Vander Meer R Valles SM and LivoreJP 2006 New survey for the fire ant microsporidia Vairimorphainvictae and Thelohania solenopsae in southern South Americawith observations on their field persistence and prevalence of dualinfections Environ Entomol 35 1358ndash1365
Buckley A 1999 Fire ants in California Am Bee J 139 88Carroll CR and Hoffman CA 2000 The pervasive ecological effects
of invasive species Exotic and native fire ants In Invertebrates asWeb-Masters in Ecosystems (Coleman DC and Hendrix PF Eds)Wallingford Oxon UK pp 221ndash232
Carruthers RI 2003 Invasive species research in the United StatesDepartment of Agriculture ndash Agricultural Research Service PestManag Sci 59 827ndash834
Chen JSC Shen CH and Lee HJ 2006 Monogynous and polygynousred imported fire ants Solenopsis invicta Buren (HymenopteraFormicidae) in Taiwan Environ Entomol 35 167ndash172
Cornuet JM and Luikart G 1996 Description and power analysis oftwo tests for detecting recent population bottlenecks from allelefrequency data Genetics 144 2001ndash2014
Davis LR Vander Meer RK and Porter SD 2001 Red imported fireants expand their range across the West Indies Florida Entomol84 735ndash736
DeHeer CJ and Tschinkel WR 1998 The success of alternativereproductive tactics in monogyne Solenopsis invicta significancefor transitions in social organization Behav Ecol 9 130ndash135
DeHeer CJ Goodisman MAD and Ross KG 1999 Queen dispersalstrategies in the multiple-queen form of the fire ant Solenopsisinvicta Am Nat 153 660ndash675
Dieringer D and Schlccediltterer C 2002 Microsatellite analyser (MSA) aplatform independent analysis tool for large microsatellite datasets Mol Ecol Notes 3 167ndash169
Drake JA Mooney HA Castri F Groves RH Kruger MRejmnek M and Williamson M 1989 Biological Invasions AGlobal Perspective Chichester UK 550 pp
Drees BM and Vinson SB 1990 Comparison of the control ofmonogynous and polygynous forms of the red imported fire ant(Hymenoptera Formicidae) with a chloripyrifos mound drench JEntomol Sci 25 317ndash324
Table 3 Tests for genetic bottlenecks using the infinite allele model(IAM) and two-phase model (TPM 90 stepwise mutation model and10 multistep mutation model [Garza and Williamson 2001])implemented in the program BOTTLENECK (a=005)Significance at given statistic level
Population Simulatedmodel
Significancetest
Standardizeddifferences test
Wilcoxontest
Taoyuan IAM 0148 0003 0015TPM 0186 0035 0023
Chiayi IAM 0037 0022 0007TPM 0521 0293 0281
Insect Soc Vol 55 2008 Research article 63
Drees BM and Gold RE 2003 Development of integrated pestmanagement programs for the red imported fire ant (Hymeno-ACHTUNGTRENNUNGptera Formicidae) J Entomol Sci 38 170ndash180
Eubanks MD 2001 Estimates of the direct and indirect effects of redimported fire ants on biological control in field crops Biol Control21 35 ndash43
Evanno G Regnaut S and Goudet J 2005 Detecting the number ofclusters of individuals using the software STRUCTURE asimulation study Mol Ecol 14 2611ndash2620
Excoffier L Smouse PE and Quattro JM 1992 Analysis of molecularvariance inferred from metric distances among DNA haplotypesApplication to human mitochondrial DNA restriction dataGenetics 131 479ndash491
Forys EA Quistorff A and Allen CR 2001 Potential fire ant(Hymenoptera Formicidae) impact on the endangered Schausswallowtail (Lepidoptera Papilionidae) Florida Entomol 84 254ndash258
Garza JC and Williamson EG 2001 Detection of reduction inpopulation size using data from microsatellite loci Mol Ecol 10 305ndash318
Giraud T Pedersen JS and Keller L 2002 Evolution of supercoloniesthe Argentine ants of southern Europe Proc Natl Acad Sci U SA 99 6075ndash6079
Giuliano WM Allen CR Lutz RS and Demarais S 1996 Effects ofred imported fire ants on northern bobwhite chicks J WildlManage 60 309ndash313
Glancey BM Craig CH Stringer CE and Bishop PM 1973Multiple fertile queens in colonies of the imported fire antSolenopsis invicta J Georgia Entomol Soc 8 327ndash328
Goodisman MAD DeHeer CJ and Ross KG 2000 Mating behaviorof queens from multiple-queen colonies of Solenopsis invicta JInsect Behav 13 455ndash468
Goodnight KF and Queller DC 1999 Computer software forperforming likelihood tests of pedigree relationship using geneticmarkers Mol Ecol 8 1231ndash1234
Gotelli NJ and Arnett AE 2000 Biogeographic effects of red fire antinvasion Ecol Lett 3 257ndash261
Gotzek D and Ross KG 2007 Genetic regulation of colony socialorganization in fire ants An integrative overview Q Rev Biol 82 201ndash226
Goudet J 2001 FSTAT A Program to Estimate and Test GeneDiversities and Fixation Indices Version 293 http wwwunilch izeasoftwares fstathtml
Greenberg L Fletcher DJC and Vinson SB 1985 Differences inworker size and mound distribution in monogynous and polygy-nous colonies of the fire ant Solenopsis invicta Buren J KansasEntomol Soc 58 9ndash18
Hampton JO Spencer PBS Alpers DL Twigg LE WoolnoughAP Doust J Higgs T and Pluske J 2004 Molecular techniqueswildlife management and the importance of genetic populationstructure and dispersal a case study with feral pigs J Appl Ecol41 735ndash743
Henshaw MT Kunzmann N Vanderwoude C Sanetra M andCrozier RH 2005 Population genetics and history of theintroduced fire ant Solenopsis invicta Buren (HymenopteraFormicidae) in Australia Aus J Entomol 44 37ndash44
Hccedillldobler B and Wilson EO 1990 The Ants The Belknap Press ofHarvard University Press Cambridge Massachusetts 732 pp
Huang TC Chou YC and Chou HC 2004 The infestation andcontrol of the red imported fire ant in Taiwan In Proceedingsof the Symposium on the Control of the Red Imported Fire AntBureau of Animal and Plant Health Inspection and QuarantineCouncil of Agriculture Executive Yuan Taipei Taiwan pp1 ndash 13
Jaquiry J Vogel V and Keller L 2005 Multilevel genetic analyses oftwo European supercolonies of the Argentine ant Linepithemahumile Mol Ecol 14 589ndash598
Jeyaprakash A and Hoy MA 2000 Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropods Insect Mol Biol 9 393ndash405
Jouvenaz DP 1990 Approaches to biological control of fire ants in theUnited States In Applied Myrmecology A World Perspective(Vander Meer RK Jaffe K and Cedeno A Eds) Westview PressBoulder Colorado pp 620ndash627
Kaspari M 2000 Do imported fire ants impact canopy arthropodsEvidence from simple arboreal pitfall traps Southwestern Nat 45 118ndash122
Keller L and Ross KG 1998 Selfish genes a green beard in the red fireant Nature 394 573ndash575
Kemp SF deShazo RD Moffitt JE Williams DF and Buhner WA2000 Expanding habitat of the imported fire ant (Solenopsisinvicta) A public health concern J Allergy Clin Immunol 105 683ndash691
Kopachena JG Buckley AJ and Potts GA 2000 Effects of the redimported fire ant (Solenopsis invicta) on reproductive success ofbarn swallows (Hirundo rustica) in northeast Texas SouthwesternNat 45 477ndash482
Krieger MJB and Keller L 1997 Polymorphism at dinucleotidemicrosatellite loci in fire ant Solenopsis invicta populations MolEcol 6 997ndash999
Krieger MJB Ross KG Chang CWY and Keller L 1999Frequency and origin of triploidy in the fire ant Solenopsis invictaHeredity 82 142ndash150
Krieger MJB and Ross KG 2002 Identification of a major generegulating complex social behavior Science 295 328ndash332
Lee CE 2002 Evolutionary genetics of invasive species Trends EcolEvol 17 386ndash391
Lofgren CS Banks WA and Glancey BM 1975 Biology and controlof imported fire ants Annu Rev Entomol 20 1ndash30
Lofgren CS 1986 History of imported fire ants in the United States InFire Ants and Leaf Cutting Ants Biology and Management(Lofgren CS and Vander Meer RK Eds) Westview PressBoulder Colorado pp 36ndash49
Luikart G Allendorf FW Cornuet JM and Sherwin WB 1998Distortion of allele frequency distributions provides a test forrecent population bottlenecks J Hered 89 238ndash247
Luikart G and Cornuet JM 1998 Empirical evaluation of a test foridentifying recently bottlenecked populations from allele frequen-cy data Conserv Biol 12 228ndash237
MacKay WP and Fagerlund R 1997 Range expansion of the redimported fire ant Solenopsis invicta Buren (Hymenoptera For-micidae) into New Mexico and extreme western Texas ProcEntomol Soc Wash 99 757ndash758
Mank JE and Avise JC 2004 Individual organisms as units of analysisBayesian-clustering alternatives in population genetics Genet Res84 135ndash143
May GE Gelembiuk GW Panov VE Orlova MI and Lee CE2006 Molecular ecology of zebra mussel invasions Mol Ecol 15 1021ndash1031
McCubbin KI and Weiner JM 2002 Fire ants in Australia A newmedical and ecological hazard Med J Aus 176 518ndash519
Morrison LW 2000 Mechanisms of interspecific competition amongan invasive and two native fire ants Oikos 90 238ndash252
Morrison LW 2002 Long-term impacts of an arthropod-communityinvasion by the imported fire ant Solenopsis invicta Ecology 83 2337ndash2345
Nattrass R and Vanderwoude C 2001 A preliminary investigation ofthe ecological effects of red imported fire ants (Solenopsis invicta)in Brisbane Ecol Manag Resto 2 220ndash223
Nei M 1972 Genetic distance between populations Am Nat 106 283ndash292
Orr MR 1996 Host manipulation by Wolbachia is a neutral trait withinsingle populations Anim Behav 51 1183ndash1185
Patterson RS 1994 Biological control of introduced ant species InExotic Ants Biology Impact and Control of Introduced Species(Williams DF Ed) Westview Press Boulder Colorado pp 293ndash308
Piry S Luikart G and Cornuet JM 1999 BOTTLENECK acomputer program for detecting recent reductions in the effectivepopulation size using allele frequency data J Hered 90 502ndash503
64 CC Yang et al Fire ants in Taiwan
Porter SD and Savignano DA 1990 Invasion of polygyne fire antsdecimates native ants and disrupts arthropod community Ecology7 2095ndash2106
Porter SD Bhatkar A Mulder R Vinson SB and Clair DJ 1991Distribution and density of polygyne fire ants (HymenopteraFormicidae) in Texas J Econ Entomol 84 866ndash874
Porter SD Fowler HG and MacKay WP 1992 Fire ant mounddensities in the United States and Brazil (Hymenoptera Formi-cidae) J Econ Entomol 85 1154ndash1161
Porter SD Williams DF Patterson RS and Fowler HG 1997Intercontinental differences in the abundance of Solenopsis fireants (Hymenoptera Formicidae) Escape from natural enemiesEnviron Entomol 26 373ndash384
Pritchard JK Stephens M and Donnelly P 2000 Inference ofpopulation structure using multilocus genotype data Genetics155 945ndash959
Raymond M and Rousset F 1995a An exact test for populationdifferentiation Evolution 49 1280ndash1283
Raymond M and Rousset F 1995b GENEPOP (version 12)population genetics software for exact tests and ecumenicism JHered 86 248ndash249
Reuter M Pedersen JS and Keller L 2005 Loss of Wolbachiainfection during colonisation in the invasive Argentine ant Line-pithema humile Heredity 94 364ndash369
Ross KG and Fletcher DJC 1985a Genetic origin of male diploidy inthe fire ant Solenopsis invicta (Hymenoptera Formicidae) and itsevolutionary significance Evolution 39 888ndash903
Ross KG and Fletcher DJC 1985b Comparative study of genetic andsocial structure in two forms of the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 17 349ndash356
Ross KG and Fletcher DJC 1986 Diploid male productionndashasignificant colony mortality factor in the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 19 283ndash291
Ross KG and Shoemaker DD 1993 An unusual pattern of gene flowbetween the two social forms of the fire ant Solenopsis invictaEvolution 47 1595ndash1605
Ross KG Vargo EL Keller L and Trager JC 1993 Effect of afounder event on variation in the genetic sex-determining system ofthe fire ant Solenopsis invicta Genetics 135 843ndash854
Ross KG and Keller L 1995 Joint influence of gene flow and selectionon a reproductively important genetic polymorphism in the fire antSolenopsis invicta Am Nat 146 325ndash348
Ross KG Vargo EL and Keller L 1996 Social evolution in a newenvironment The case of introduced fire ants Proc Natl AcadSci U S A 93 3021ndash3025
Ross KG 1997 Multilocus evolution in fire ants Effects of selectiongene flow and recombination Genetics 145 961ndash974
Ross KG Krieger MJB Shoemaker DD Vargo EL and Keller L1997 Hierarchical analysis of genetic structure in native fire antpopulations Results from three classes of molecular markersGenetics 147 643ndash655
Ross KG and Shoemaker DD 1997 Nuclear and mitochondrialgenetic structure in two social forms of the fire ant Solenopsisinvicta Insights into transitions to an alternate social organizationHeredity 78 590ndash602
Ross KG and Keller L 1998 Genetic control of social organization inan ant Proc Natl Acad Sci U S A 95 14232ndash14237
Ross KG Shoemaker DD Krieger MJ DeHeer CJ and Keller L1999 Assessing genetic structure with multiple classes of molecularmarkers A case study involving the introduced fire ant Solenopsisinvicta Mol Biol Evol 16 525ndash543
Ross KG and Keller L 2002 Experimental conversion of colony socialorganization by manipulation of worker genotype composition infire ants (Solenopsis invicta) Behav Ecol Sociobiol 51 287ndash295
Ross KG Krieger MJB and Shoemaker DD 2003 Alternativegenetic foundations and evolutionary routes for a key socialpolymorphism in fire ants Genetics 165 1853ndash1867
Ross KG Krieger MJB Keller L and Shoemaker DD 2007Genetic variation and structure in native populations of the fire antSolenopsis invicta Evolutionary and demographic implicationsBiol J Linnean Soc 92 541ndash560
Rousset F 1997 Genetic differentiation and estimation of gene flowfrom F-statistics under isolation by distance Genetics 145 1219ndash1228
Sax DF Stachowicz JJ and Gaines SD 2005 Species InvasionsInsights into Ecology Evolution and Biogeography SinauerAssociates Sunderland Massachussetts 495 pp
Shoemaker DD and Ross KG 1996 Effects of social organization ongene flow in the fire ant Solenopsis invicta Nature 383 613ndash616
Shoemaker DD Ross KG Keller L Vargo EL and Werren JH2000 Wolbachia infections in native and introduced populations offire ants (Solenopsis spp) Insect Mol Biol 9 661ndash673
Shoemaker DD Ahrens M Sheill L Mescher M Keller L and RossKG 2003 Distribution and prevalence of Wolbachia infections innative populations of the fire ant Solenopsis invicta (HymenopteraFormicidae) Environ Entomol 32 1329ndash1336
Shoemaker DD DeHeer CJ Krieger MJB and Ross KG 2006Population genetics of the invasive fire ant Solenopsis invicta(Hymenoptera Formicidae) in the USA Ann Entomol Soc Am99 1213ndash1233
Simon C Frati F Beckenbach A Crespi B Liu H and Flook P 1994Evolution weighting and phylogenetic utility of mitochondrialgene sequences and a compilation of conserved polymerase chainreaction primers Ann Entomol Soc Am 87 651ndash701
Toom PM Cupp E Johnson CP and Griffin I 1976 Utilization ofbody reserves for minim brood development by queens of theimported fire ant Solenopsis invicta J Insect Physiol 22 217ndash220
Tsutsui ND and Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepethema humile) in its nativeand introduced ranges Evolution 55 976ndash985
Tsutsui ND Kauppinen SN Oyafuso AF and Grosberg RK 2003The distribution and evolutionary history of Wolbachia infection innative and introduced populations of the invasive argentine ant(Linepithema humile) Mol Ecol 12 3057ndash3068
Valles SM and Porter SD 2003 Identification of polygyne andmonogyne fire ant colonies (Solenopsis invicta) by multiplex PCRof Gp-9 alleles Insect Soc 50 199ndash200
Valles SM Oi DH Briano JA and Williams DF 2004 Simultaneousdetection of Vairimorpha invictae (Microsporidia Burenellidae)and Thelohania solenopsae (Microsporidia Thelohaniidae) in fireants by PCR Florida Entomol 87 85ndash87
Vander Meer R K and Porter SD 2001 Fate of newly-mated queensintroduced into monogyne and polygyne Solenopsis invicta (Hy-menoptera Formicidae) colonies Ann Entomol Soc Am 94 289ndash297
Vander Meer RK Jaffe K and Cedeno A 1990 Applied Myrmecol-ogy A World Perspective Westview Press Boulder CO 741 pp
Vinson SB 1994 Impact and invasion of Solenopsis invicta (Buren) onnative food webs In Exotic Ants Biology Impact and Control ofIntroduced Species (Williams DF Ed) Westview Press BoulderColorado pp 293ndash308
Vinson SB 1997 Invasion of the red imported fire ant (HymenopteraFormicidae) Spread biology and impact Am Entomol 43 23ndash29
Williams DF and Porter SD 1994 Fire ant control Science 264 1653ndash1653
Wojcik D Allen CR Brenner RJ Forys EA Jouvenaz D and LutzRS 2001 Red imported fire ants Impact on biodiversity AmEntomol 47 16ndash23
Xia X and Xie Z 2001 DAMBE Data analysis in molecular biologyand evolution J Hered 92 371ndash373
Insect Soc Vol 55 2008 Research article 65

Isolation by distance and tests for genetic bottlenecks
No correlation exists between pairwise FST or FST(1ndash FST)values and geographic distance for individuals of thepolygyne social form in either population (TaoyuanPgt022 Chiayi Pgt048) Further no significant iso-lation by distance for the monogyne nests from Taoyuanpopulation was found (Pgt065) We did not perform theMantel tests for the Chiayi monogyne population sinceonly two nests were collected
All tests revealed a significant excess of heterozygos-ity in Taoyuan under both models except the sign test(Table 3) A significant excess of heterozygosity wasfound in Chiayi for the infinite allele model but not thetwo phase model (Table 3) Combined with a mode shiftof allele frequency distribution (Table 3 Fig 5a) theTaoyuan population appears to have undergone a recentbottleneck In contrast the most abundant alleles in theChiayi population belong to the low frequency class(lt01 Fig 5b) resulting in a normal L-shape distribu-
tion We also compared NA (number of observed allelesTable 2) and HE (expected heterozygosity Table 2)among the South American USA and Taiwanesepopulations to determine whether a genetic signaturecharacteristic of a bottleneck was evident for the latterpopulations In order to minimize the effect of populationsize we randomly selected 30 individuals from the USAand South American dataset (Shoemaker et al 2006Ross et al 2007) Results showed that the NA and HE inTaiwan and the USA populations were consistentlylower than in South American fire ant populations Twopopulations in Taiwan had slightly lower NA than inUSA but had similar HE values
Additional findings
Analyses of individual genotypes revealed that 108 ofthe 190 workers sampled from polygyne nests in Taoyuanand 11 of 230 individuals from polygyne nests in Chiayi
Table 2 Genetic diversity measures within the two study populations of S invicta in Taiwan NA denotes observed number of alleles found at eachlocus from each population HE =expected heterozygosity HO = observed heterozygosity Numbers in parentheses represent the number of privatealleles occurring in only one of the two Taiwanese populations Genetic diversity measures for two population from the USA and South Americaalso are shown (see text)
Source of collection Population Sample size Mean no of alleles NA HE HO
TaiwanTaoyuan county 30 38 23 (2) 0609 0527Chiayi county 25 48 26 (5) 0636 0660
USAMississippi 30 57 34 0651 0559western Louisiana 30 58 35 0628 0607
South AmericaFormosa Argentina 30 158 195 0835 0842Corrientes Argentina 30 158 95 0836 0826
Figure 5 Distribution of allele frequencies at six microsatellite loci for ants (single individual per nest) from the Taoyuan (a) and Chiayi populations(b)
60 CC Yang et al Fire ants in Taiwan
harbored three distinct alleles whereas all of the individ-uals sampled from monogyne nests harbored at most twodistinct alleles These results indicate that a substantialnumber of polygyne fire ants are triploid a findingconsistent with an earlier study of fire ants from theUSA (Krieger et al 1999)
Discussion
We studied the colony genetic and population structure ofthe invasive fire ant S invicta in Taiwan by collecting andanalyzing microsatellite genotypic data and mtDNAsequence data of ants collected from two separateinfested areas Our genetic analyses revealed severalpatterns which are discussed in more detail below andcan be summarized as follows 1) Estimates of within nestrelatedness were significantly greater than zero for bothsocial forms from both populations 2) No significantisolation by distance was found among nests within eachsocial form from either population 3) SignificantmtDNA but no nuclear differentiation occurs betweensympatric social forms in Taoyuan (no differentiationbetween forms at either genome is evident in Chiayi) 4)Molecular signatures of genetic bottlenecks associatedwith recent introductions are evident in both populations5) The two sampled populations Taoyuan and Chiayi arehighly genetically differentiated at both the nuclear ormtDNA genomes In addition to these general findingswe found numerous triploid individuals in ants of thepolygyne (P) but not the monogyne (M) form anddistinct geographic distribution of each of the two socialforms between the two populations
Colony social organization and patterns of intracolonyworker relatedness
The estimate of relatedness among workers withinmonogyne colonies was not significantly different from075 the predicted value for a nest headed by one singly-mated queen These data also are consistent with datafrom earlier studies of monogyne S invicta colonies inboth the USA (Ross and Fletcher 1985b) and SouthAmerica (Ross et al 1997) and suggest that the socialand mating biology of M queens is virtually invariantwith M nests representing closed societies headed by onesingly-mated reproductive queen
In contrast colony genetic structure of P social formdoes differ between native South America and intro-duced populations in the USA (Ross et al 1996) Pnests of S invicta in the native range contain relativelyfew queens which are close relatives and relatednessamong nestmate workers is significantly greater thanzero whereas P nests in the USA contain highnumbers of unrelated queens and relatedness amongnestmate workers is not significantly greater than zeroThese differences in queen number and relatedness
between the two ranges are consistent with predictionsbased on differences in the population densities of antsin these two areas (Ross et al 1996) which aresubstantially higher in the USA compared to SouthAmerica (Porter et al 1997) One consequence of thehigher population densities in the USA is that nestingsites are more often saturated such that opportunitiesfor independent founding by queens are more limitedin the introduced range (Ross et al 1996) Theseconstraints on independent founding lead to increasedselection pressure on queens to seek adoption intoexisting colonies and on resident workers of colonies toaccept such queens As queen number within thesenests increases relatedness among nestmate workers isexpected to decrease
Relatedness among workers within P nests in Taiwanis small yet significantly greater than zero and intermedi-ate to the values observed for native and USApopulations The social organization of P nests in Taiwanlikely reflects the fact that nesting sites are much lesssaturated compared to sites in the USA since thepopulation densities of ants in Taiwan currently are muchlower than those in the USA However while such adifference between the two introduced ranges are evidentnow it is unlikely to persist especially if S invictacontinues to spread in Taiwan so that habitats becomesaturated If S invicta continues to spread so that habitatsbecome more saturated future studies of the socialevolution of P populations in Taiwan could shed addi-tional light on whether such a change in habitat avail-ability does in fact lead to a shift in social organizationtowards a higher queen number within nests and lowerrelatedness among nestmates
Patterns of isolation by distance
The lack of isolation by distance among colonies of the Mform within Taoyuan was not unexpected given that mostM queens take part in aerial mating flights oftendispersing several kilometers or more before landingand attempting to found a new nest (Toom et al 1976Ross and Fletcher 1985b Ross et al 1999) Such long-distance dispersal is predicted to erase any signature ofnuclear differentiation among closely spaced nests
The lack of detectable isolation by distance within theP form at both studied sites was somewhat unexpectedconsidering the reproductive habits of P queens whichoften mate in or near their natal nests and form newcolonies by budding or by entering into nearby existingnests (Shoemaker and Ross 1996 Ross et al 1997 Rossand Shoemaker 1997) One explanation for the lack ofdetectable isolation by distance is simply that malesdisperse rather long distances to mate with P queensUnder this scenario one would predict that long distancemale-mediated gene flow would result in homogenizationof allele frequencies at nuclear loci yet significantmicrogeographic mtDNA structure would remain espe-
Insect Soc Vol 55 2008 Research article 61
cially if queen dispersal is limited However no suchpattern was found Significant isolation by distance wasabsent for both the nuclear and mtDNA markers Anadditional explanation for these results is that thesignature of isolation by distance predicted to emergesolely from the natural dispersal biology of these ants isconfounded by the fact that these ants are frequentlyinadvertently transported relatively great distances byhumans Such frequent inadvertent human transport maybe a common characteristic of invasive ants found in closeproximity to human habitats and thus play a role inshaping the genetic structure in introduced areas Finallyit is possible that the lack of a detectable signal of isolationby distance is due to the extremely recent introduction offire ants into Taiwan with insufficient time having passedfor populations to achieve migrationdrift equilibrium
Gene flow between sympatric social forms
The pronounced mtDNA differentiation between sym-patric social forms in Taoyuan is consistent with resultsfrom earlier studies in both South America and the USAshowing that interform gene flow mediated by queens israre or absent Queens of each social form rarely succeedin becoming reproductives in colonies of the alternateform as a result of behavioral decisions (attributed largelyto differences in genotypes of queens at the single geneGp-9) made by both queens and workers (Shoemaker andRoss 1996 Ross and Shoemaker 1997 Keller and Ross1998 DeHeer et al 1999 Goodisman et al 2000 Rossand Keller 2002) The contrasting weak or nonexistentinterform divergence at nuclear genes also is consistentwith studies of S invicta in the USA showing that queensof the P form predominantly mate with males of the Mform where the two forms co-occur (Ross and Shoe-maker 1993 Ross and Keller 1995 Shoemaker andRoss 1996) In the USA most males in P colonies aresterile diploid males which means that P queens relylargely on immigrant M males for mating opportunities(Ross and Fletcher 1985a Ross and Shoemaker 1993)Such interform matings act as a homogenizing force onlyfor nuclear markers The similar frequency of triploidworkers in P nests in Taiwan and the USA suggestsmany diploid males also occur in Taiwan as well (Rosset al 1993 1999 Krieger and Keller 1997 Krieger et al 1999) which means that P queens here must also matewith M males The finding of contrasting patterns ofdifferentiation between the two genomes parallels resultsfrom earlier studies involving S invicta in both the USAand South America (Shoemaker and Ross 1996 Rosset al 1997 Shoemaker et al 2006) One final caveathowever is that we cannot completely rule out thepossibility that the lack of detectable nuclear differ-entiation is due to insufficient time having passed fordivergence to occur despite restricted gene flow betweenthe two forms
Understanding the distributions of the two socialforms in the two infested areas in Taiwan each of whichexhibits rather distinct dispersal strategies is importantfor developing effective control strategies of fire ants inthese infested areas (Porter et al 1991) The distribu-tional pattern of social forms in Taoyuan suggests that theexpansion of this population has involved persistentoutward movement of ants from the core infestationcoupled with frequent long distance movement of Mreproductives into outlying areas (Fig 1b) Based onthese results we recommend that bait broadcasting andsurveillance efforts in Taoyuan should include not onlythe currently infested areas but also should include anarea extending well beyond the infested areas In contrastto the distributional pattern of social forms in Taoyuanthe large infested area in Chiayi consists mostly of P nests(92 of colonies collected Fig 1c) However despitethis difference we suggest a similar eradication strategyshould be employed This is because even thoughcomprehensive bait broadcasting within the entire areamay be more effective than for Taoyuan because of thelow queen vagility and relatively slow rate of reinvasionby budding of polygynous queens (Porter et al 1991)some M colonies do occur in Chiayi and there also ispotential for additional M colonies to arise locally fromthe P social form a possibility that is especially morelikely in areas of low density (Ross and Shoemaker 1997DeHeer and Tschinkel 1998 DeHeer et al 1999)
Genetic bottlenecks in Taoyuan and Chiayi populations
Simulations and statistical tests using the program BOT-TLENECK provide evidence of a significant geneticbottleneck associated with the introduction of fire antsinto Taoyuan with only one exception (Table 3) How-ever this single exception is based on a sign test whichhas relatively low statistical power when the number ofloci is below 20 such as our study (Cornuet and Luikart1996) In Chiayi significant excess in heterozygosity usingthe infinite alleles model (IAM) was observed for all testsbut not for the two-phase model (TPM Table 3) BecauseLuikart and Cornuet (1998) suggest that implementingthe test under the IAM model may be more reliable thanother models these results coupled with finding thatallelic frequencies fit a normal L-shaped distributionprovide evidence for a genetic bottleneck Interestinglyestimates of NA and HE (Table 2) are remarkably similarfor introduced populations from Taiwan and the USAand considerably lower than those from the native SouthAmerican range While the geographic origin of ants inTaiwan is unknown the similar genetic diversity measuresbetween the introduced ranges raises the issue that theintroduction of ants into Taiwan has left only a modestfootprint on patterns of diversity if these ants originatedfrom the USA
62 CC Yang et al Fire ants in Taiwan
Genetic differentiation between Taoyuan and Chiayipopulations
All analyses of population differentiation using themicrosatellite and mtDNA data indicate that the twogeographic populations are strongly differentiated andthus likely originated from separate invasions Whilemultiple lines of evidence support this notion thestrongest evidence is the confinement each of the threemtDNA variants to only one of the two geographicpopulations Interestingly all three variants found inTaiwan are the same three variants commonly found inants from the USA (Shoemaker et al 2006) China(GenBank accession numbers DQ831670-DQ831672)and Australia (CCY and DDS unpublished results) Theoccurrence of these three variants in all recently invadedareas suggests that ants from Taiwan could have origi-nated from any of these other areas We currently aregenerating the necessary baseline data for ants from all ofthese introduced regions as well as from populations intheir native range to infer the source population andinvasion history of ants in Taiwan and these other areas
From the perspective of managing fire ants in Taiwantreating each infested area as a separate management unitwould appear more reasonable and means that scientistsprobably should focus on separate plans for eliminationof two isolated populations (Hampton et al 2004) Onthe other hand because these two areas are at high risk ofconnectivity via long distance human-mediated dispersalcare must be taken to prevent reinvasion into these areaseven after successful eradication Stringent inter-countyquarantine measures such as movement control shouldbe established to monitor the risk of transportation of fireant-infested goods to prevent jump dispersal between thetwo areas and decrease probability of successful erad-ication (Drees and Gold 2003)
Acknowledgments
The authors would like to thank Chung-Chi Lin for providing most ofthe specimens for this study including the first identified samples inTaoyuan and Chiayi counties We are particularly grateful to Hwei-yuChang Hon-Tsen Yu and Chia-Hung Hsieh for helpful discussions andcomments on earlier versions of this manuscript Comments by twoanonymous reviewers greatly improved this manuscript This research
was financially supported by grants from Academia Sinica Taiwan Theuse of trade firm or corporation names in this publication is for theinformation and convenience of the reader Such use does notconstitute an official endorsement or approval by the United StatesDepartment of Agriculture or the Agricultural Research Service of anyproduct or service to the exclusion of others that may be suitable
References
Ahrens ME Ross KG and Shoemaker DD 2005 Phylogeographicstructure of the fire ant Solenopsis invicta in its native SouthAmerican range Roles of natural barriers and habitat connectivityEvolution 59 1733ndash1743
Allen CR Demarais S and Lutz RS 1994 Red imported fire antimpact on wildlife an overview Texas J Sci 46 51ndash59
Allen CR Lutz RS and Demarais S 1998 Ecological effects of theinvasive nonindigenous ant Solenopsis invicta on native verte-brates The wheels on the bus Transactions of the North AmericanWildlife and Natural Resources Conference 63 56 ndash65
Allen CR Willey RD Myers PE Horton PM and Buffa J 2000Impact of red imported fire ant infestation on northern bobwhitequail abundance trends in southeastern United States J AgrUrban Entomol 17 43ndash51
Allen CR Forys EA Rice KG and Wojcik DP 2001 Effects of fireants (Hymenoptera Formicidae) on hatching turtles and preva-lence of fire ants on sea turtle nesting beaches in Florida FloridaEntomol 84 250ndash253
Bouwma AM Ahrens ME DeHeer CJ and Shoemaker DD 2006Distribution and prevalence of Wolbachia in introduced popula-tions of the fire ant Solenopsis invicta Insect Mol Biol 15 89ndash93
Briano JA Calcaterra LA Vander Meer R Valles SM and LivoreJP 2006 New survey for the fire ant microsporidia Vairimorphainvictae and Thelohania solenopsae in southern South Americawith observations on their field persistence and prevalence of dualinfections Environ Entomol 35 1358ndash1365
Buckley A 1999 Fire ants in California Am Bee J 139 88Carroll CR and Hoffman CA 2000 The pervasive ecological effects
of invasive species Exotic and native fire ants In Invertebrates asWeb-Masters in Ecosystems (Coleman DC and Hendrix PF Eds)Wallingford Oxon UK pp 221ndash232
Carruthers RI 2003 Invasive species research in the United StatesDepartment of Agriculture ndash Agricultural Research Service PestManag Sci 59 827ndash834
Chen JSC Shen CH and Lee HJ 2006 Monogynous and polygynousred imported fire ants Solenopsis invicta Buren (HymenopteraFormicidae) in Taiwan Environ Entomol 35 167ndash172
Cornuet JM and Luikart G 1996 Description and power analysis oftwo tests for detecting recent population bottlenecks from allelefrequency data Genetics 144 2001ndash2014
Davis LR Vander Meer RK and Porter SD 2001 Red imported fireants expand their range across the West Indies Florida Entomol84 735ndash736
DeHeer CJ and Tschinkel WR 1998 The success of alternativereproductive tactics in monogyne Solenopsis invicta significancefor transitions in social organization Behav Ecol 9 130ndash135
DeHeer CJ Goodisman MAD and Ross KG 1999 Queen dispersalstrategies in the multiple-queen form of the fire ant Solenopsisinvicta Am Nat 153 660ndash675
Dieringer D and Schlccediltterer C 2002 Microsatellite analyser (MSA) aplatform independent analysis tool for large microsatellite datasets Mol Ecol Notes 3 167ndash169
Drake JA Mooney HA Castri F Groves RH Kruger MRejmnek M and Williamson M 1989 Biological Invasions AGlobal Perspective Chichester UK 550 pp
Drees BM and Vinson SB 1990 Comparison of the control ofmonogynous and polygynous forms of the red imported fire ant(Hymenoptera Formicidae) with a chloripyrifos mound drench JEntomol Sci 25 317ndash324
Table 3 Tests for genetic bottlenecks using the infinite allele model(IAM) and two-phase model (TPM 90 stepwise mutation model and10 multistep mutation model [Garza and Williamson 2001])implemented in the program BOTTLENECK (a=005)Significance at given statistic level
Population Simulatedmodel
Significancetest
Standardizeddifferences test
Wilcoxontest
Taoyuan IAM 0148 0003 0015TPM 0186 0035 0023
Chiayi IAM 0037 0022 0007TPM 0521 0293 0281
Insect Soc Vol 55 2008 Research article 63
Drees BM and Gold RE 2003 Development of integrated pestmanagement programs for the red imported fire ant (Hymeno-ACHTUNGTRENNUNGptera Formicidae) J Entomol Sci 38 170ndash180
Eubanks MD 2001 Estimates of the direct and indirect effects of redimported fire ants on biological control in field crops Biol Control21 35 ndash43
Evanno G Regnaut S and Goudet J 2005 Detecting the number ofclusters of individuals using the software STRUCTURE asimulation study Mol Ecol 14 2611ndash2620
Excoffier L Smouse PE and Quattro JM 1992 Analysis of molecularvariance inferred from metric distances among DNA haplotypesApplication to human mitochondrial DNA restriction dataGenetics 131 479ndash491
Forys EA Quistorff A and Allen CR 2001 Potential fire ant(Hymenoptera Formicidae) impact on the endangered Schausswallowtail (Lepidoptera Papilionidae) Florida Entomol 84 254ndash258
Garza JC and Williamson EG 2001 Detection of reduction inpopulation size using data from microsatellite loci Mol Ecol 10 305ndash318
Giraud T Pedersen JS and Keller L 2002 Evolution of supercoloniesthe Argentine ants of southern Europe Proc Natl Acad Sci U SA 99 6075ndash6079
Giuliano WM Allen CR Lutz RS and Demarais S 1996 Effects ofred imported fire ants on northern bobwhite chicks J WildlManage 60 309ndash313
Glancey BM Craig CH Stringer CE and Bishop PM 1973Multiple fertile queens in colonies of the imported fire antSolenopsis invicta J Georgia Entomol Soc 8 327ndash328
Goodisman MAD DeHeer CJ and Ross KG 2000 Mating behaviorof queens from multiple-queen colonies of Solenopsis invicta JInsect Behav 13 455ndash468
Goodnight KF and Queller DC 1999 Computer software forperforming likelihood tests of pedigree relationship using geneticmarkers Mol Ecol 8 1231ndash1234
Gotelli NJ and Arnett AE 2000 Biogeographic effects of red fire antinvasion Ecol Lett 3 257ndash261
Gotzek D and Ross KG 2007 Genetic regulation of colony socialorganization in fire ants An integrative overview Q Rev Biol 82 201ndash226
Goudet J 2001 FSTAT A Program to Estimate and Test GeneDiversities and Fixation Indices Version 293 http wwwunilch izeasoftwares fstathtml
Greenberg L Fletcher DJC and Vinson SB 1985 Differences inworker size and mound distribution in monogynous and polygy-nous colonies of the fire ant Solenopsis invicta Buren J KansasEntomol Soc 58 9ndash18
Hampton JO Spencer PBS Alpers DL Twigg LE WoolnoughAP Doust J Higgs T and Pluske J 2004 Molecular techniqueswildlife management and the importance of genetic populationstructure and dispersal a case study with feral pigs J Appl Ecol41 735ndash743
Henshaw MT Kunzmann N Vanderwoude C Sanetra M andCrozier RH 2005 Population genetics and history of theintroduced fire ant Solenopsis invicta Buren (HymenopteraFormicidae) in Australia Aus J Entomol 44 37ndash44
Hccedillldobler B and Wilson EO 1990 The Ants The Belknap Press ofHarvard University Press Cambridge Massachusetts 732 pp
Huang TC Chou YC and Chou HC 2004 The infestation andcontrol of the red imported fire ant in Taiwan In Proceedingsof the Symposium on the Control of the Red Imported Fire AntBureau of Animal and Plant Health Inspection and QuarantineCouncil of Agriculture Executive Yuan Taipei Taiwan pp1 ndash 13
Jaquiry J Vogel V and Keller L 2005 Multilevel genetic analyses oftwo European supercolonies of the Argentine ant Linepithemahumile Mol Ecol 14 589ndash598
Jeyaprakash A and Hoy MA 2000 Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropods Insect Mol Biol 9 393ndash405
Jouvenaz DP 1990 Approaches to biological control of fire ants in theUnited States In Applied Myrmecology A World Perspective(Vander Meer RK Jaffe K and Cedeno A Eds) Westview PressBoulder Colorado pp 620ndash627
Kaspari M 2000 Do imported fire ants impact canopy arthropodsEvidence from simple arboreal pitfall traps Southwestern Nat 45 118ndash122
Keller L and Ross KG 1998 Selfish genes a green beard in the red fireant Nature 394 573ndash575
Kemp SF deShazo RD Moffitt JE Williams DF and Buhner WA2000 Expanding habitat of the imported fire ant (Solenopsisinvicta) A public health concern J Allergy Clin Immunol 105 683ndash691
Kopachena JG Buckley AJ and Potts GA 2000 Effects of the redimported fire ant (Solenopsis invicta) on reproductive success ofbarn swallows (Hirundo rustica) in northeast Texas SouthwesternNat 45 477ndash482
Krieger MJB and Keller L 1997 Polymorphism at dinucleotidemicrosatellite loci in fire ant Solenopsis invicta populations MolEcol 6 997ndash999
Krieger MJB Ross KG Chang CWY and Keller L 1999Frequency and origin of triploidy in the fire ant Solenopsis invictaHeredity 82 142ndash150
Krieger MJB and Ross KG 2002 Identification of a major generegulating complex social behavior Science 295 328ndash332
Lee CE 2002 Evolutionary genetics of invasive species Trends EcolEvol 17 386ndash391
Lofgren CS Banks WA and Glancey BM 1975 Biology and controlof imported fire ants Annu Rev Entomol 20 1ndash30
Lofgren CS 1986 History of imported fire ants in the United States InFire Ants and Leaf Cutting Ants Biology and Management(Lofgren CS and Vander Meer RK Eds) Westview PressBoulder Colorado pp 36ndash49
Luikart G Allendorf FW Cornuet JM and Sherwin WB 1998Distortion of allele frequency distributions provides a test forrecent population bottlenecks J Hered 89 238ndash247
Luikart G and Cornuet JM 1998 Empirical evaluation of a test foridentifying recently bottlenecked populations from allele frequen-cy data Conserv Biol 12 228ndash237
MacKay WP and Fagerlund R 1997 Range expansion of the redimported fire ant Solenopsis invicta Buren (Hymenoptera For-micidae) into New Mexico and extreme western Texas ProcEntomol Soc Wash 99 757ndash758
Mank JE and Avise JC 2004 Individual organisms as units of analysisBayesian-clustering alternatives in population genetics Genet Res84 135ndash143
May GE Gelembiuk GW Panov VE Orlova MI and Lee CE2006 Molecular ecology of zebra mussel invasions Mol Ecol 15 1021ndash1031
McCubbin KI and Weiner JM 2002 Fire ants in Australia A newmedical and ecological hazard Med J Aus 176 518ndash519
Morrison LW 2000 Mechanisms of interspecific competition amongan invasive and two native fire ants Oikos 90 238ndash252
Morrison LW 2002 Long-term impacts of an arthropod-communityinvasion by the imported fire ant Solenopsis invicta Ecology 83 2337ndash2345
Nattrass R and Vanderwoude C 2001 A preliminary investigation ofthe ecological effects of red imported fire ants (Solenopsis invicta)in Brisbane Ecol Manag Resto 2 220ndash223
Nei M 1972 Genetic distance between populations Am Nat 106 283ndash292
Orr MR 1996 Host manipulation by Wolbachia is a neutral trait withinsingle populations Anim Behav 51 1183ndash1185
Patterson RS 1994 Biological control of introduced ant species InExotic Ants Biology Impact and Control of Introduced Species(Williams DF Ed) Westview Press Boulder Colorado pp 293ndash308
Piry S Luikart G and Cornuet JM 1999 BOTTLENECK acomputer program for detecting recent reductions in the effectivepopulation size using allele frequency data J Hered 90 502ndash503
64 CC Yang et al Fire ants in Taiwan
Porter SD and Savignano DA 1990 Invasion of polygyne fire antsdecimates native ants and disrupts arthropod community Ecology7 2095ndash2106
Porter SD Bhatkar A Mulder R Vinson SB and Clair DJ 1991Distribution and density of polygyne fire ants (HymenopteraFormicidae) in Texas J Econ Entomol 84 866ndash874
Porter SD Fowler HG and MacKay WP 1992 Fire ant mounddensities in the United States and Brazil (Hymenoptera Formi-cidae) J Econ Entomol 85 1154ndash1161
Porter SD Williams DF Patterson RS and Fowler HG 1997Intercontinental differences in the abundance of Solenopsis fireants (Hymenoptera Formicidae) Escape from natural enemiesEnviron Entomol 26 373ndash384
Pritchard JK Stephens M and Donnelly P 2000 Inference ofpopulation structure using multilocus genotype data Genetics155 945ndash959
Raymond M and Rousset F 1995a An exact test for populationdifferentiation Evolution 49 1280ndash1283
Raymond M and Rousset F 1995b GENEPOP (version 12)population genetics software for exact tests and ecumenicism JHered 86 248ndash249
Reuter M Pedersen JS and Keller L 2005 Loss of Wolbachiainfection during colonisation in the invasive Argentine ant Line-pithema humile Heredity 94 364ndash369
Ross KG and Fletcher DJC 1985a Genetic origin of male diploidy inthe fire ant Solenopsis invicta (Hymenoptera Formicidae) and itsevolutionary significance Evolution 39 888ndash903
Ross KG and Fletcher DJC 1985b Comparative study of genetic andsocial structure in two forms of the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 17 349ndash356
Ross KG and Fletcher DJC 1986 Diploid male productionndashasignificant colony mortality factor in the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 19 283ndash291
Ross KG and Shoemaker DD 1993 An unusual pattern of gene flowbetween the two social forms of the fire ant Solenopsis invictaEvolution 47 1595ndash1605
Ross KG Vargo EL Keller L and Trager JC 1993 Effect of afounder event on variation in the genetic sex-determining system ofthe fire ant Solenopsis invicta Genetics 135 843ndash854
Ross KG and Keller L 1995 Joint influence of gene flow and selectionon a reproductively important genetic polymorphism in the fire antSolenopsis invicta Am Nat 146 325ndash348
Ross KG Vargo EL and Keller L 1996 Social evolution in a newenvironment The case of introduced fire ants Proc Natl AcadSci U S A 93 3021ndash3025
Ross KG 1997 Multilocus evolution in fire ants Effects of selectiongene flow and recombination Genetics 145 961ndash974
Ross KG Krieger MJB Shoemaker DD Vargo EL and Keller L1997 Hierarchical analysis of genetic structure in native fire antpopulations Results from three classes of molecular markersGenetics 147 643ndash655
Ross KG and Shoemaker DD 1997 Nuclear and mitochondrialgenetic structure in two social forms of the fire ant Solenopsisinvicta Insights into transitions to an alternate social organizationHeredity 78 590ndash602
Ross KG and Keller L 1998 Genetic control of social organization inan ant Proc Natl Acad Sci U S A 95 14232ndash14237
Ross KG Shoemaker DD Krieger MJ DeHeer CJ and Keller L1999 Assessing genetic structure with multiple classes of molecularmarkers A case study involving the introduced fire ant Solenopsisinvicta Mol Biol Evol 16 525ndash543
Ross KG and Keller L 2002 Experimental conversion of colony socialorganization by manipulation of worker genotype composition infire ants (Solenopsis invicta) Behav Ecol Sociobiol 51 287ndash295
Ross KG Krieger MJB and Shoemaker DD 2003 Alternativegenetic foundations and evolutionary routes for a key socialpolymorphism in fire ants Genetics 165 1853ndash1867
Ross KG Krieger MJB Keller L and Shoemaker DD 2007Genetic variation and structure in native populations of the fire antSolenopsis invicta Evolutionary and demographic implicationsBiol J Linnean Soc 92 541ndash560
Rousset F 1997 Genetic differentiation and estimation of gene flowfrom F-statistics under isolation by distance Genetics 145 1219ndash1228
Sax DF Stachowicz JJ and Gaines SD 2005 Species InvasionsInsights into Ecology Evolution and Biogeography SinauerAssociates Sunderland Massachussetts 495 pp
Shoemaker DD and Ross KG 1996 Effects of social organization ongene flow in the fire ant Solenopsis invicta Nature 383 613ndash616
Shoemaker DD Ross KG Keller L Vargo EL and Werren JH2000 Wolbachia infections in native and introduced populations offire ants (Solenopsis spp) Insect Mol Biol 9 661ndash673
Shoemaker DD Ahrens M Sheill L Mescher M Keller L and RossKG 2003 Distribution and prevalence of Wolbachia infections innative populations of the fire ant Solenopsis invicta (HymenopteraFormicidae) Environ Entomol 32 1329ndash1336
Shoemaker DD DeHeer CJ Krieger MJB and Ross KG 2006Population genetics of the invasive fire ant Solenopsis invicta(Hymenoptera Formicidae) in the USA Ann Entomol Soc Am99 1213ndash1233
Simon C Frati F Beckenbach A Crespi B Liu H and Flook P 1994Evolution weighting and phylogenetic utility of mitochondrialgene sequences and a compilation of conserved polymerase chainreaction primers Ann Entomol Soc Am 87 651ndash701
Toom PM Cupp E Johnson CP and Griffin I 1976 Utilization ofbody reserves for minim brood development by queens of theimported fire ant Solenopsis invicta J Insect Physiol 22 217ndash220
Tsutsui ND and Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepethema humile) in its nativeand introduced ranges Evolution 55 976ndash985
Tsutsui ND Kauppinen SN Oyafuso AF and Grosberg RK 2003The distribution and evolutionary history of Wolbachia infection innative and introduced populations of the invasive argentine ant(Linepithema humile) Mol Ecol 12 3057ndash3068
Valles SM and Porter SD 2003 Identification of polygyne andmonogyne fire ant colonies (Solenopsis invicta) by multiplex PCRof Gp-9 alleles Insect Soc 50 199ndash200
Valles SM Oi DH Briano JA and Williams DF 2004 Simultaneousdetection of Vairimorpha invictae (Microsporidia Burenellidae)and Thelohania solenopsae (Microsporidia Thelohaniidae) in fireants by PCR Florida Entomol 87 85ndash87
Vander Meer R K and Porter SD 2001 Fate of newly-mated queensintroduced into monogyne and polygyne Solenopsis invicta (Hy-menoptera Formicidae) colonies Ann Entomol Soc Am 94 289ndash297
Vander Meer RK Jaffe K and Cedeno A 1990 Applied Myrmecol-ogy A World Perspective Westview Press Boulder CO 741 pp
Vinson SB 1994 Impact and invasion of Solenopsis invicta (Buren) onnative food webs In Exotic Ants Biology Impact and Control ofIntroduced Species (Williams DF Ed) Westview Press BoulderColorado pp 293ndash308
Vinson SB 1997 Invasion of the red imported fire ant (HymenopteraFormicidae) Spread biology and impact Am Entomol 43 23ndash29
Williams DF and Porter SD 1994 Fire ant control Science 264 1653ndash1653
Wojcik D Allen CR Brenner RJ Forys EA Jouvenaz D and LutzRS 2001 Red imported fire ants Impact on biodiversity AmEntomol 47 16ndash23
Xia X and Xie Z 2001 DAMBE Data analysis in molecular biologyand evolution J Hered 92 371ndash373
Insect Soc Vol 55 2008 Research article 65

harbored three distinct alleles whereas all of the individ-uals sampled from monogyne nests harbored at most twodistinct alleles These results indicate that a substantialnumber of polygyne fire ants are triploid a findingconsistent with an earlier study of fire ants from theUSA (Krieger et al 1999)
Discussion
We studied the colony genetic and population structure ofthe invasive fire ant S invicta in Taiwan by collecting andanalyzing microsatellite genotypic data and mtDNAsequence data of ants collected from two separateinfested areas Our genetic analyses revealed severalpatterns which are discussed in more detail below andcan be summarized as follows 1) Estimates of within nestrelatedness were significantly greater than zero for bothsocial forms from both populations 2) No significantisolation by distance was found among nests within eachsocial form from either population 3) SignificantmtDNA but no nuclear differentiation occurs betweensympatric social forms in Taoyuan (no differentiationbetween forms at either genome is evident in Chiayi) 4)Molecular signatures of genetic bottlenecks associatedwith recent introductions are evident in both populations5) The two sampled populations Taoyuan and Chiayi arehighly genetically differentiated at both the nuclear ormtDNA genomes In addition to these general findingswe found numerous triploid individuals in ants of thepolygyne (P) but not the monogyne (M) form anddistinct geographic distribution of each of the two socialforms between the two populations
Colony social organization and patterns of intracolonyworker relatedness
The estimate of relatedness among workers withinmonogyne colonies was not significantly different from075 the predicted value for a nest headed by one singly-mated queen These data also are consistent with datafrom earlier studies of monogyne S invicta colonies inboth the USA (Ross and Fletcher 1985b) and SouthAmerica (Ross et al 1997) and suggest that the socialand mating biology of M queens is virtually invariantwith M nests representing closed societies headed by onesingly-mated reproductive queen
In contrast colony genetic structure of P social formdoes differ between native South America and intro-duced populations in the USA (Ross et al 1996) Pnests of S invicta in the native range contain relativelyfew queens which are close relatives and relatednessamong nestmate workers is significantly greater thanzero whereas P nests in the USA contain highnumbers of unrelated queens and relatedness amongnestmate workers is not significantly greater than zeroThese differences in queen number and relatedness
between the two ranges are consistent with predictionsbased on differences in the population densities of antsin these two areas (Ross et al 1996) which aresubstantially higher in the USA compared to SouthAmerica (Porter et al 1997) One consequence of thehigher population densities in the USA is that nestingsites are more often saturated such that opportunitiesfor independent founding by queens are more limitedin the introduced range (Ross et al 1996) Theseconstraints on independent founding lead to increasedselection pressure on queens to seek adoption intoexisting colonies and on resident workers of colonies toaccept such queens As queen number within thesenests increases relatedness among nestmate workers isexpected to decrease
Relatedness among workers within P nests in Taiwanis small yet significantly greater than zero and intermedi-ate to the values observed for native and USApopulations The social organization of P nests in Taiwanlikely reflects the fact that nesting sites are much lesssaturated compared to sites in the USA since thepopulation densities of ants in Taiwan currently are muchlower than those in the USA However while such adifference between the two introduced ranges are evidentnow it is unlikely to persist especially if S invictacontinues to spread in Taiwan so that habitats becomesaturated If S invicta continues to spread so that habitatsbecome more saturated future studies of the socialevolution of P populations in Taiwan could shed addi-tional light on whether such a change in habitat avail-ability does in fact lead to a shift in social organizationtowards a higher queen number within nests and lowerrelatedness among nestmates
Patterns of isolation by distance
The lack of isolation by distance among colonies of the Mform within Taoyuan was not unexpected given that mostM queens take part in aerial mating flights oftendispersing several kilometers or more before landingand attempting to found a new nest (Toom et al 1976Ross and Fletcher 1985b Ross et al 1999) Such long-distance dispersal is predicted to erase any signature ofnuclear differentiation among closely spaced nests
The lack of detectable isolation by distance within theP form at both studied sites was somewhat unexpectedconsidering the reproductive habits of P queens whichoften mate in or near their natal nests and form newcolonies by budding or by entering into nearby existingnests (Shoemaker and Ross 1996 Ross et al 1997 Rossand Shoemaker 1997) One explanation for the lack ofdetectable isolation by distance is simply that malesdisperse rather long distances to mate with P queensUnder this scenario one would predict that long distancemale-mediated gene flow would result in homogenizationof allele frequencies at nuclear loci yet significantmicrogeographic mtDNA structure would remain espe-
Insect Soc Vol 55 2008 Research article 61
cially if queen dispersal is limited However no suchpattern was found Significant isolation by distance wasabsent for both the nuclear and mtDNA markers Anadditional explanation for these results is that thesignature of isolation by distance predicted to emergesolely from the natural dispersal biology of these ants isconfounded by the fact that these ants are frequentlyinadvertently transported relatively great distances byhumans Such frequent inadvertent human transport maybe a common characteristic of invasive ants found in closeproximity to human habitats and thus play a role inshaping the genetic structure in introduced areas Finallyit is possible that the lack of a detectable signal of isolationby distance is due to the extremely recent introduction offire ants into Taiwan with insufficient time having passedfor populations to achieve migrationdrift equilibrium
Gene flow between sympatric social forms
The pronounced mtDNA differentiation between sym-patric social forms in Taoyuan is consistent with resultsfrom earlier studies in both South America and the USAshowing that interform gene flow mediated by queens israre or absent Queens of each social form rarely succeedin becoming reproductives in colonies of the alternateform as a result of behavioral decisions (attributed largelyto differences in genotypes of queens at the single geneGp-9) made by both queens and workers (Shoemaker andRoss 1996 Ross and Shoemaker 1997 Keller and Ross1998 DeHeer et al 1999 Goodisman et al 2000 Rossand Keller 2002) The contrasting weak or nonexistentinterform divergence at nuclear genes also is consistentwith studies of S invicta in the USA showing that queensof the P form predominantly mate with males of the Mform where the two forms co-occur (Ross and Shoe-maker 1993 Ross and Keller 1995 Shoemaker andRoss 1996) In the USA most males in P colonies aresterile diploid males which means that P queens relylargely on immigrant M males for mating opportunities(Ross and Fletcher 1985a Ross and Shoemaker 1993)Such interform matings act as a homogenizing force onlyfor nuclear markers The similar frequency of triploidworkers in P nests in Taiwan and the USA suggestsmany diploid males also occur in Taiwan as well (Rosset al 1993 1999 Krieger and Keller 1997 Krieger et al 1999) which means that P queens here must also matewith M males The finding of contrasting patterns ofdifferentiation between the two genomes parallels resultsfrom earlier studies involving S invicta in both the USAand South America (Shoemaker and Ross 1996 Rosset al 1997 Shoemaker et al 2006) One final caveathowever is that we cannot completely rule out thepossibility that the lack of detectable nuclear differ-entiation is due to insufficient time having passed fordivergence to occur despite restricted gene flow betweenthe two forms
Understanding the distributions of the two socialforms in the two infested areas in Taiwan each of whichexhibits rather distinct dispersal strategies is importantfor developing effective control strategies of fire ants inthese infested areas (Porter et al 1991) The distribu-tional pattern of social forms in Taoyuan suggests that theexpansion of this population has involved persistentoutward movement of ants from the core infestationcoupled with frequent long distance movement of Mreproductives into outlying areas (Fig 1b) Based onthese results we recommend that bait broadcasting andsurveillance efforts in Taoyuan should include not onlythe currently infested areas but also should include anarea extending well beyond the infested areas In contrastto the distributional pattern of social forms in Taoyuanthe large infested area in Chiayi consists mostly of P nests(92 of colonies collected Fig 1c) However despitethis difference we suggest a similar eradication strategyshould be employed This is because even thoughcomprehensive bait broadcasting within the entire areamay be more effective than for Taoyuan because of thelow queen vagility and relatively slow rate of reinvasionby budding of polygynous queens (Porter et al 1991)some M colonies do occur in Chiayi and there also ispotential for additional M colonies to arise locally fromthe P social form a possibility that is especially morelikely in areas of low density (Ross and Shoemaker 1997DeHeer and Tschinkel 1998 DeHeer et al 1999)
Genetic bottlenecks in Taoyuan and Chiayi populations
Simulations and statistical tests using the program BOT-TLENECK provide evidence of a significant geneticbottleneck associated with the introduction of fire antsinto Taoyuan with only one exception (Table 3) How-ever this single exception is based on a sign test whichhas relatively low statistical power when the number ofloci is below 20 such as our study (Cornuet and Luikart1996) In Chiayi significant excess in heterozygosity usingthe infinite alleles model (IAM) was observed for all testsbut not for the two-phase model (TPM Table 3) BecauseLuikart and Cornuet (1998) suggest that implementingthe test under the IAM model may be more reliable thanother models these results coupled with finding thatallelic frequencies fit a normal L-shaped distributionprovide evidence for a genetic bottleneck Interestinglyestimates of NA and HE (Table 2) are remarkably similarfor introduced populations from Taiwan and the USAand considerably lower than those from the native SouthAmerican range While the geographic origin of ants inTaiwan is unknown the similar genetic diversity measuresbetween the introduced ranges raises the issue that theintroduction of ants into Taiwan has left only a modestfootprint on patterns of diversity if these ants originatedfrom the USA
62 CC Yang et al Fire ants in Taiwan
Genetic differentiation between Taoyuan and Chiayipopulations
All analyses of population differentiation using themicrosatellite and mtDNA data indicate that the twogeographic populations are strongly differentiated andthus likely originated from separate invasions Whilemultiple lines of evidence support this notion thestrongest evidence is the confinement each of the threemtDNA variants to only one of the two geographicpopulations Interestingly all three variants found inTaiwan are the same three variants commonly found inants from the USA (Shoemaker et al 2006) China(GenBank accession numbers DQ831670-DQ831672)and Australia (CCY and DDS unpublished results) Theoccurrence of these three variants in all recently invadedareas suggests that ants from Taiwan could have origi-nated from any of these other areas We currently aregenerating the necessary baseline data for ants from all ofthese introduced regions as well as from populations intheir native range to infer the source population andinvasion history of ants in Taiwan and these other areas
From the perspective of managing fire ants in Taiwantreating each infested area as a separate management unitwould appear more reasonable and means that scientistsprobably should focus on separate plans for eliminationof two isolated populations (Hampton et al 2004) Onthe other hand because these two areas are at high risk ofconnectivity via long distance human-mediated dispersalcare must be taken to prevent reinvasion into these areaseven after successful eradication Stringent inter-countyquarantine measures such as movement control shouldbe established to monitor the risk of transportation of fireant-infested goods to prevent jump dispersal between thetwo areas and decrease probability of successful erad-ication (Drees and Gold 2003)
Acknowledgments
The authors would like to thank Chung-Chi Lin for providing most ofthe specimens for this study including the first identified samples inTaoyuan and Chiayi counties We are particularly grateful to Hwei-yuChang Hon-Tsen Yu and Chia-Hung Hsieh for helpful discussions andcomments on earlier versions of this manuscript Comments by twoanonymous reviewers greatly improved this manuscript This research
was financially supported by grants from Academia Sinica Taiwan Theuse of trade firm or corporation names in this publication is for theinformation and convenience of the reader Such use does notconstitute an official endorsement or approval by the United StatesDepartment of Agriculture or the Agricultural Research Service of anyproduct or service to the exclusion of others that may be suitable
References
Ahrens ME Ross KG and Shoemaker DD 2005 Phylogeographicstructure of the fire ant Solenopsis invicta in its native SouthAmerican range Roles of natural barriers and habitat connectivityEvolution 59 1733ndash1743
Allen CR Demarais S and Lutz RS 1994 Red imported fire antimpact on wildlife an overview Texas J Sci 46 51ndash59
Allen CR Lutz RS and Demarais S 1998 Ecological effects of theinvasive nonindigenous ant Solenopsis invicta on native verte-brates The wheels on the bus Transactions of the North AmericanWildlife and Natural Resources Conference 63 56 ndash65
Allen CR Willey RD Myers PE Horton PM and Buffa J 2000Impact of red imported fire ant infestation on northern bobwhitequail abundance trends in southeastern United States J AgrUrban Entomol 17 43ndash51
Allen CR Forys EA Rice KG and Wojcik DP 2001 Effects of fireants (Hymenoptera Formicidae) on hatching turtles and preva-lence of fire ants on sea turtle nesting beaches in Florida FloridaEntomol 84 250ndash253
Bouwma AM Ahrens ME DeHeer CJ and Shoemaker DD 2006Distribution and prevalence of Wolbachia in introduced popula-tions of the fire ant Solenopsis invicta Insect Mol Biol 15 89ndash93
Briano JA Calcaterra LA Vander Meer R Valles SM and LivoreJP 2006 New survey for the fire ant microsporidia Vairimorphainvictae and Thelohania solenopsae in southern South Americawith observations on their field persistence and prevalence of dualinfections Environ Entomol 35 1358ndash1365
Buckley A 1999 Fire ants in California Am Bee J 139 88Carroll CR and Hoffman CA 2000 The pervasive ecological effects
of invasive species Exotic and native fire ants In Invertebrates asWeb-Masters in Ecosystems (Coleman DC and Hendrix PF Eds)Wallingford Oxon UK pp 221ndash232
Carruthers RI 2003 Invasive species research in the United StatesDepartment of Agriculture ndash Agricultural Research Service PestManag Sci 59 827ndash834
Chen JSC Shen CH and Lee HJ 2006 Monogynous and polygynousred imported fire ants Solenopsis invicta Buren (HymenopteraFormicidae) in Taiwan Environ Entomol 35 167ndash172
Cornuet JM and Luikart G 1996 Description and power analysis oftwo tests for detecting recent population bottlenecks from allelefrequency data Genetics 144 2001ndash2014
Davis LR Vander Meer RK and Porter SD 2001 Red imported fireants expand their range across the West Indies Florida Entomol84 735ndash736
DeHeer CJ and Tschinkel WR 1998 The success of alternativereproductive tactics in monogyne Solenopsis invicta significancefor transitions in social organization Behav Ecol 9 130ndash135
DeHeer CJ Goodisman MAD and Ross KG 1999 Queen dispersalstrategies in the multiple-queen form of the fire ant Solenopsisinvicta Am Nat 153 660ndash675
Dieringer D and Schlccediltterer C 2002 Microsatellite analyser (MSA) aplatform independent analysis tool for large microsatellite datasets Mol Ecol Notes 3 167ndash169
Drake JA Mooney HA Castri F Groves RH Kruger MRejmnek M and Williamson M 1989 Biological Invasions AGlobal Perspective Chichester UK 550 pp
Drees BM and Vinson SB 1990 Comparison of the control ofmonogynous and polygynous forms of the red imported fire ant(Hymenoptera Formicidae) with a chloripyrifos mound drench JEntomol Sci 25 317ndash324
Table 3 Tests for genetic bottlenecks using the infinite allele model(IAM) and two-phase model (TPM 90 stepwise mutation model and10 multistep mutation model [Garza and Williamson 2001])implemented in the program BOTTLENECK (a=005)Significance at given statistic level
Population Simulatedmodel
Significancetest
Standardizeddifferences test
Wilcoxontest
Taoyuan IAM 0148 0003 0015TPM 0186 0035 0023
Chiayi IAM 0037 0022 0007TPM 0521 0293 0281
Insect Soc Vol 55 2008 Research article 63
Drees BM and Gold RE 2003 Development of integrated pestmanagement programs for the red imported fire ant (Hymeno-ACHTUNGTRENNUNGptera Formicidae) J Entomol Sci 38 170ndash180
Eubanks MD 2001 Estimates of the direct and indirect effects of redimported fire ants on biological control in field crops Biol Control21 35 ndash43
Evanno G Regnaut S and Goudet J 2005 Detecting the number ofclusters of individuals using the software STRUCTURE asimulation study Mol Ecol 14 2611ndash2620
Excoffier L Smouse PE and Quattro JM 1992 Analysis of molecularvariance inferred from metric distances among DNA haplotypesApplication to human mitochondrial DNA restriction dataGenetics 131 479ndash491
Forys EA Quistorff A and Allen CR 2001 Potential fire ant(Hymenoptera Formicidae) impact on the endangered Schausswallowtail (Lepidoptera Papilionidae) Florida Entomol 84 254ndash258
Garza JC and Williamson EG 2001 Detection of reduction inpopulation size using data from microsatellite loci Mol Ecol 10 305ndash318
Giraud T Pedersen JS and Keller L 2002 Evolution of supercoloniesthe Argentine ants of southern Europe Proc Natl Acad Sci U SA 99 6075ndash6079
Giuliano WM Allen CR Lutz RS and Demarais S 1996 Effects ofred imported fire ants on northern bobwhite chicks J WildlManage 60 309ndash313
Glancey BM Craig CH Stringer CE and Bishop PM 1973Multiple fertile queens in colonies of the imported fire antSolenopsis invicta J Georgia Entomol Soc 8 327ndash328
Goodisman MAD DeHeer CJ and Ross KG 2000 Mating behaviorof queens from multiple-queen colonies of Solenopsis invicta JInsect Behav 13 455ndash468
Goodnight KF and Queller DC 1999 Computer software forperforming likelihood tests of pedigree relationship using geneticmarkers Mol Ecol 8 1231ndash1234
Gotelli NJ and Arnett AE 2000 Biogeographic effects of red fire antinvasion Ecol Lett 3 257ndash261
Gotzek D and Ross KG 2007 Genetic regulation of colony socialorganization in fire ants An integrative overview Q Rev Biol 82 201ndash226
Goudet J 2001 FSTAT A Program to Estimate and Test GeneDiversities and Fixation Indices Version 293 http wwwunilch izeasoftwares fstathtml
Greenberg L Fletcher DJC and Vinson SB 1985 Differences inworker size and mound distribution in monogynous and polygy-nous colonies of the fire ant Solenopsis invicta Buren J KansasEntomol Soc 58 9ndash18
Hampton JO Spencer PBS Alpers DL Twigg LE WoolnoughAP Doust J Higgs T and Pluske J 2004 Molecular techniqueswildlife management and the importance of genetic populationstructure and dispersal a case study with feral pigs J Appl Ecol41 735ndash743
Henshaw MT Kunzmann N Vanderwoude C Sanetra M andCrozier RH 2005 Population genetics and history of theintroduced fire ant Solenopsis invicta Buren (HymenopteraFormicidae) in Australia Aus J Entomol 44 37ndash44
Hccedillldobler B and Wilson EO 1990 The Ants The Belknap Press ofHarvard University Press Cambridge Massachusetts 732 pp
Huang TC Chou YC and Chou HC 2004 The infestation andcontrol of the red imported fire ant in Taiwan In Proceedingsof the Symposium on the Control of the Red Imported Fire AntBureau of Animal and Plant Health Inspection and QuarantineCouncil of Agriculture Executive Yuan Taipei Taiwan pp1 ndash 13
Jaquiry J Vogel V and Keller L 2005 Multilevel genetic analyses oftwo European supercolonies of the Argentine ant Linepithemahumile Mol Ecol 14 589ndash598
Jeyaprakash A and Hoy MA 2000 Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropods Insect Mol Biol 9 393ndash405
Jouvenaz DP 1990 Approaches to biological control of fire ants in theUnited States In Applied Myrmecology A World Perspective(Vander Meer RK Jaffe K and Cedeno A Eds) Westview PressBoulder Colorado pp 620ndash627
Kaspari M 2000 Do imported fire ants impact canopy arthropodsEvidence from simple arboreal pitfall traps Southwestern Nat 45 118ndash122
Keller L and Ross KG 1998 Selfish genes a green beard in the red fireant Nature 394 573ndash575
Kemp SF deShazo RD Moffitt JE Williams DF and Buhner WA2000 Expanding habitat of the imported fire ant (Solenopsisinvicta) A public health concern J Allergy Clin Immunol 105 683ndash691
Kopachena JG Buckley AJ and Potts GA 2000 Effects of the redimported fire ant (Solenopsis invicta) on reproductive success ofbarn swallows (Hirundo rustica) in northeast Texas SouthwesternNat 45 477ndash482
Krieger MJB and Keller L 1997 Polymorphism at dinucleotidemicrosatellite loci in fire ant Solenopsis invicta populations MolEcol 6 997ndash999
Krieger MJB Ross KG Chang CWY and Keller L 1999Frequency and origin of triploidy in the fire ant Solenopsis invictaHeredity 82 142ndash150
Krieger MJB and Ross KG 2002 Identification of a major generegulating complex social behavior Science 295 328ndash332
Lee CE 2002 Evolutionary genetics of invasive species Trends EcolEvol 17 386ndash391
Lofgren CS Banks WA and Glancey BM 1975 Biology and controlof imported fire ants Annu Rev Entomol 20 1ndash30
Lofgren CS 1986 History of imported fire ants in the United States InFire Ants and Leaf Cutting Ants Biology and Management(Lofgren CS and Vander Meer RK Eds) Westview PressBoulder Colorado pp 36ndash49
Luikart G Allendorf FW Cornuet JM and Sherwin WB 1998Distortion of allele frequency distributions provides a test forrecent population bottlenecks J Hered 89 238ndash247
Luikart G and Cornuet JM 1998 Empirical evaluation of a test foridentifying recently bottlenecked populations from allele frequen-cy data Conserv Biol 12 228ndash237
MacKay WP and Fagerlund R 1997 Range expansion of the redimported fire ant Solenopsis invicta Buren (Hymenoptera For-micidae) into New Mexico and extreme western Texas ProcEntomol Soc Wash 99 757ndash758
Mank JE and Avise JC 2004 Individual organisms as units of analysisBayesian-clustering alternatives in population genetics Genet Res84 135ndash143
May GE Gelembiuk GW Panov VE Orlova MI and Lee CE2006 Molecular ecology of zebra mussel invasions Mol Ecol 15 1021ndash1031
McCubbin KI and Weiner JM 2002 Fire ants in Australia A newmedical and ecological hazard Med J Aus 176 518ndash519
Morrison LW 2000 Mechanisms of interspecific competition amongan invasive and two native fire ants Oikos 90 238ndash252
Morrison LW 2002 Long-term impacts of an arthropod-communityinvasion by the imported fire ant Solenopsis invicta Ecology 83 2337ndash2345
Nattrass R and Vanderwoude C 2001 A preliminary investigation ofthe ecological effects of red imported fire ants (Solenopsis invicta)in Brisbane Ecol Manag Resto 2 220ndash223
Nei M 1972 Genetic distance between populations Am Nat 106 283ndash292
Orr MR 1996 Host manipulation by Wolbachia is a neutral trait withinsingle populations Anim Behav 51 1183ndash1185
Patterson RS 1994 Biological control of introduced ant species InExotic Ants Biology Impact and Control of Introduced Species(Williams DF Ed) Westview Press Boulder Colorado pp 293ndash308
Piry S Luikart G and Cornuet JM 1999 BOTTLENECK acomputer program for detecting recent reductions in the effectivepopulation size using allele frequency data J Hered 90 502ndash503
64 CC Yang et al Fire ants in Taiwan
Porter SD and Savignano DA 1990 Invasion of polygyne fire antsdecimates native ants and disrupts arthropod community Ecology7 2095ndash2106
Porter SD Bhatkar A Mulder R Vinson SB and Clair DJ 1991Distribution and density of polygyne fire ants (HymenopteraFormicidae) in Texas J Econ Entomol 84 866ndash874
Porter SD Fowler HG and MacKay WP 1992 Fire ant mounddensities in the United States and Brazil (Hymenoptera Formi-cidae) J Econ Entomol 85 1154ndash1161
Porter SD Williams DF Patterson RS and Fowler HG 1997Intercontinental differences in the abundance of Solenopsis fireants (Hymenoptera Formicidae) Escape from natural enemiesEnviron Entomol 26 373ndash384
Pritchard JK Stephens M and Donnelly P 2000 Inference ofpopulation structure using multilocus genotype data Genetics155 945ndash959
Raymond M and Rousset F 1995a An exact test for populationdifferentiation Evolution 49 1280ndash1283
Raymond M and Rousset F 1995b GENEPOP (version 12)population genetics software for exact tests and ecumenicism JHered 86 248ndash249
Reuter M Pedersen JS and Keller L 2005 Loss of Wolbachiainfection during colonisation in the invasive Argentine ant Line-pithema humile Heredity 94 364ndash369
Ross KG and Fletcher DJC 1985a Genetic origin of male diploidy inthe fire ant Solenopsis invicta (Hymenoptera Formicidae) and itsevolutionary significance Evolution 39 888ndash903
Ross KG and Fletcher DJC 1985b Comparative study of genetic andsocial structure in two forms of the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 17 349ndash356
Ross KG and Fletcher DJC 1986 Diploid male productionndashasignificant colony mortality factor in the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 19 283ndash291
Ross KG and Shoemaker DD 1993 An unusual pattern of gene flowbetween the two social forms of the fire ant Solenopsis invictaEvolution 47 1595ndash1605
Ross KG Vargo EL Keller L and Trager JC 1993 Effect of afounder event on variation in the genetic sex-determining system ofthe fire ant Solenopsis invicta Genetics 135 843ndash854
Ross KG and Keller L 1995 Joint influence of gene flow and selectionon a reproductively important genetic polymorphism in the fire antSolenopsis invicta Am Nat 146 325ndash348
Ross KG Vargo EL and Keller L 1996 Social evolution in a newenvironment The case of introduced fire ants Proc Natl AcadSci U S A 93 3021ndash3025
Ross KG 1997 Multilocus evolution in fire ants Effects of selectiongene flow and recombination Genetics 145 961ndash974
Ross KG Krieger MJB Shoemaker DD Vargo EL and Keller L1997 Hierarchical analysis of genetic structure in native fire antpopulations Results from three classes of molecular markersGenetics 147 643ndash655
Ross KG and Shoemaker DD 1997 Nuclear and mitochondrialgenetic structure in two social forms of the fire ant Solenopsisinvicta Insights into transitions to an alternate social organizationHeredity 78 590ndash602
Ross KG and Keller L 1998 Genetic control of social organization inan ant Proc Natl Acad Sci U S A 95 14232ndash14237
Ross KG Shoemaker DD Krieger MJ DeHeer CJ and Keller L1999 Assessing genetic structure with multiple classes of molecularmarkers A case study involving the introduced fire ant Solenopsisinvicta Mol Biol Evol 16 525ndash543
Ross KG and Keller L 2002 Experimental conversion of colony socialorganization by manipulation of worker genotype composition infire ants (Solenopsis invicta) Behav Ecol Sociobiol 51 287ndash295
Ross KG Krieger MJB and Shoemaker DD 2003 Alternativegenetic foundations and evolutionary routes for a key socialpolymorphism in fire ants Genetics 165 1853ndash1867
Ross KG Krieger MJB Keller L and Shoemaker DD 2007Genetic variation and structure in native populations of the fire antSolenopsis invicta Evolutionary and demographic implicationsBiol J Linnean Soc 92 541ndash560
Rousset F 1997 Genetic differentiation and estimation of gene flowfrom F-statistics under isolation by distance Genetics 145 1219ndash1228
Sax DF Stachowicz JJ and Gaines SD 2005 Species InvasionsInsights into Ecology Evolution and Biogeography SinauerAssociates Sunderland Massachussetts 495 pp
Shoemaker DD and Ross KG 1996 Effects of social organization ongene flow in the fire ant Solenopsis invicta Nature 383 613ndash616
Shoemaker DD Ross KG Keller L Vargo EL and Werren JH2000 Wolbachia infections in native and introduced populations offire ants (Solenopsis spp) Insect Mol Biol 9 661ndash673
Shoemaker DD Ahrens M Sheill L Mescher M Keller L and RossKG 2003 Distribution and prevalence of Wolbachia infections innative populations of the fire ant Solenopsis invicta (HymenopteraFormicidae) Environ Entomol 32 1329ndash1336
Shoemaker DD DeHeer CJ Krieger MJB and Ross KG 2006Population genetics of the invasive fire ant Solenopsis invicta(Hymenoptera Formicidae) in the USA Ann Entomol Soc Am99 1213ndash1233
Simon C Frati F Beckenbach A Crespi B Liu H and Flook P 1994Evolution weighting and phylogenetic utility of mitochondrialgene sequences and a compilation of conserved polymerase chainreaction primers Ann Entomol Soc Am 87 651ndash701
Toom PM Cupp E Johnson CP and Griffin I 1976 Utilization ofbody reserves for minim brood development by queens of theimported fire ant Solenopsis invicta J Insect Physiol 22 217ndash220
Tsutsui ND and Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepethema humile) in its nativeand introduced ranges Evolution 55 976ndash985
Tsutsui ND Kauppinen SN Oyafuso AF and Grosberg RK 2003The distribution and evolutionary history of Wolbachia infection innative and introduced populations of the invasive argentine ant(Linepithema humile) Mol Ecol 12 3057ndash3068
Valles SM and Porter SD 2003 Identification of polygyne andmonogyne fire ant colonies (Solenopsis invicta) by multiplex PCRof Gp-9 alleles Insect Soc 50 199ndash200
Valles SM Oi DH Briano JA and Williams DF 2004 Simultaneousdetection of Vairimorpha invictae (Microsporidia Burenellidae)and Thelohania solenopsae (Microsporidia Thelohaniidae) in fireants by PCR Florida Entomol 87 85ndash87
Vander Meer R K and Porter SD 2001 Fate of newly-mated queensintroduced into monogyne and polygyne Solenopsis invicta (Hy-menoptera Formicidae) colonies Ann Entomol Soc Am 94 289ndash297
Vander Meer RK Jaffe K and Cedeno A 1990 Applied Myrmecol-ogy A World Perspective Westview Press Boulder CO 741 pp
Vinson SB 1994 Impact and invasion of Solenopsis invicta (Buren) onnative food webs In Exotic Ants Biology Impact and Control ofIntroduced Species (Williams DF Ed) Westview Press BoulderColorado pp 293ndash308
Vinson SB 1997 Invasion of the red imported fire ant (HymenopteraFormicidae) Spread biology and impact Am Entomol 43 23ndash29
Williams DF and Porter SD 1994 Fire ant control Science 264 1653ndash1653
Wojcik D Allen CR Brenner RJ Forys EA Jouvenaz D and LutzRS 2001 Red imported fire ants Impact on biodiversity AmEntomol 47 16ndash23
Xia X and Xie Z 2001 DAMBE Data analysis in molecular biologyand evolution J Hered 92 371ndash373
Insect Soc Vol 55 2008 Research article 65

cially if queen dispersal is limited However no suchpattern was found Significant isolation by distance wasabsent for both the nuclear and mtDNA markers Anadditional explanation for these results is that thesignature of isolation by distance predicted to emergesolely from the natural dispersal biology of these ants isconfounded by the fact that these ants are frequentlyinadvertently transported relatively great distances byhumans Such frequent inadvertent human transport maybe a common characteristic of invasive ants found in closeproximity to human habitats and thus play a role inshaping the genetic structure in introduced areas Finallyit is possible that the lack of a detectable signal of isolationby distance is due to the extremely recent introduction offire ants into Taiwan with insufficient time having passedfor populations to achieve migrationdrift equilibrium
Gene flow between sympatric social forms
The pronounced mtDNA differentiation between sym-patric social forms in Taoyuan is consistent with resultsfrom earlier studies in both South America and the USAshowing that interform gene flow mediated by queens israre or absent Queens of each social form rarely succeedin becoming reproductives in colonies of the alternateform as a result of behavioral decisions (attributed largelyto differences in genotypes of queens at the single geneGp-9) made by both queens and workers (Shoemaker andRoss 1996 Ross and Shoemaker 1997 Keller and Ross1998 DeHeer et al 1999 Goodisman et al 2000 Rossand Keller 2002) The contrasting weak or nonexistentinterform divergence at nuclear genes also is consistentwith studies of S invicta in the USA showing that queensof the P form predominantly mate with males of the Mform where the two forms co-occur (Ross and Shoe-maker 1993 Ross and Keller 1995 Shoemaker andRoss 1996) In the USA most males in P colonies aresterile diploid males which means that P queens relylargely on immigrant M males for mating opportunities(Ross and Fletcher 1985a Ross and Shoemaker 1993)Such interform matings act as a homogenizing force onlyfor nuclear markers The similar frequency of triploidworkers in P nests in Taiwan and the USA suggestsmany diploid males also occur in Taiwan as well (Rosset al 1993 1999 Krieger and Keller 1997 Krieger et al 1999) which means that P queens here must also matewith M males The finding of contrasting patterns ofdifferentiation between the two genomes parallels resultsfrom earlier studies involving S invicta in both the USAand South America (Shoemaker and Ross 1996 Rosset al 1997 Shoemaker et al 2006) One final caveathowever is that we cannot completely rule out thepossibility that the lack of detectable nuclear differ-entiation is due to insufficient time having passed fordivergence to occur despite restricted gene flow betweenthe two forms
Understanding the distributions of the two socialforms in the two infested areas in Taiwan each of whichexhibits rather distinct dispersal strategies is importantfor developing effective control strategies of fire ants inthese infested areas (Porter et al 1991) The distribu-tional pattern of social forms in Taoyuan suggests that theexpansion of this population has involved persistentoutward movement of ants from the core infestationcoupled with frequent long distance movement of Mreproductives into outlying areas (Fig 1b) Based onthese results we recommend that bait broadcasting andsurveillance efforts in Taoyuan should include not onlythe currently infested areas but also should include anarea extending well beyond the infested areas In contrastto the distributional pattern of social forms in Taoyuanthe large infested area in Chiayi consists mostly of P nests(92 of colonies collected Fig 1c) However despitethis difference we suggest a similar eradication strategyshould be employed This is because even thoughcomprehensive bait broadcasting within the entire areamay be more effective than for Taoyuan because of thelow queen vagility and relatively slow rate of reinvasionby budding of polygynous queens (Porter et al 1991)some M colonies do occur in Chiayi and there also ispotential for additional M colonies to arise locally fromthe P social form a possibility that is especially morelikely in areas of low density (Ross and Shoemaker 1997DeHeer and Tschinkel 1998 DeHeer et al 1999)
Genetic bottlenecks in Taoyuan and Chiayi populations
Simulations and statistical tests using the program BOT-TLENECK provide evidence of a significant geneticbottleneck associated with the introduction of fire antsinto Taoyuan with only one exception (Table 3) How-ever this single exception is based on a sign test whichhas relatively low statistical power when the number ofloci is below 20 such as our study (Cornuet and Luikart1996) In Chiayi significant excess in heterozygosity usingthe infinite alleles model (IAM) was observed for all testsbut not for the two-phase model (TPM Table 3) BecauseLuikart and Cornuet (1998) suggest that implementingthe test under the IAM model may be more reliable thanother models these results coupled with finding thatallelic frequencies fit a normal L-shaped distributionprovide evidence for a genetic bottleneck Interestinglyestimates of NA and HE (Table 2) are remarkably similarfor introduced populations from Taiwan and the USAand considerably lower than those from the native SouthAmerican range While the geographic origin of ants inTaiwan is unknown the similar genetic diversity measuresbetween the introduced ranges raises the issue that theintroduction of ants into Taiwan has left only a modestfootprint on patterns of diversity if these ants originatedfrom the USA
62 CC Yang et al Fire ants in Taiwan
Genetic differentiation between Taoyuan and Chiayipopulations
All analyses of population differentiation using themicrosatellite and mtDNA data indicate that the twogeographic populations are strongly differentiated andthus likely originated from separate invasions Whilemultiple lines of evidence support this notion thestrongest evidence is the confinement each of the threemtDNA variants to only one of the two geographicpopulations Interestingly all three variants found inTaiwan are the same three variants commonly found inants from the USA (Shoemaker et al 2006) China(GenBank accession numbers DQ831670-DQ831672)and Australia (CCY and DDS unpublished results) Theoccurrence of these three variants in all recently invadedareas suggests that ants from Taiwan could have origi-nated from any of these other areas We currently aregenerating the necessary baseline data for ants from all ofthese introduced regions as well as from populations intheir native range to infer the source population andinvasion history of ants in Taiwan and these other areas
From the perspective of managing fire ants in Taiwantreating each infested area as a separate management unitwould appear more reasonable and means that scientistsprobably should focus on separate plans for eliminationof two isolated populations (Hampton et al 2004) Onthe other hand because these two areas are at high risk ofconnectivity via long distance human-mediated dispersalcare must be taken to prevent reinvasion into these areaseven after successful eradication Stringent inter-countyquarantine measures such as movement control shouldbe established to monitor the risk of transportation of fireant-infested goods to prevent jump dispersal between thetwo areas and decrease probability of successful erad-ication (Drees and Gold 2003)
Acknowledgments
The authors would like to thank Chung-Chi Lin for providing most ofthe specimens for this study including the first identified samples inTaoyuan and Chiayi counties We are particularly grateful to Hwei-yuChang Hon-Tsen Yu and Chia-Hung Hsieh for helpful discussions andcomments on earlier versions of this manuscript Comments by twoanonymous reviewers greatly improved this manuscript This research
was financially supported by grants from Academia Sinica Taiwan Theuse of trade firm or corporation names in this publication is for theinformation and convenience of the reader Such use does notconstitute an official endorsement or approval by the United StatesDepartment of Agriculture or the Agricultural Research Service of anyproduct or service to the exclusion of others that may be suitable
References
Ahrens ME Ross KG and Shoemaker DD 2005 Phylogeographicstructure of the fire ant Solenopsis invicta in its native SouthAmerican range Roles of natural barriers and habitat connectivityEvolution 59 1733ndash1743
Allen CR Demarais S and Lutz RS 1994 Red imported fire antimpact on wildlife an overview Texas J Sci 46 51ndash59
Allen CR Lutz RS and Demarais S 1998 Ecological effects of theinvasive nonindigenous ant Solenopsis invicta on native verte-brates The wheels on the bus Transactions of the North AmericanWildlife and Natural Resources Conference 63 56 ndash65
Allen CR Willey RD Myers PE Horton PM and Buffa J 2000Impact of red imported fire ant infestation on northern bobwhitequail abundance trends in southeastern United States J AgrUrban Entomol 17 43ndash51
Allen CR Forys EA Rice KG and Wojcik DP 2001 Effects of fireants (Hymenoptera Formicidae) on hatching turtles and preva-lence of fire ants on sea turtle nesting beaches in Florida FloridaEntomol 84 250ndash253
Bouwma AM Ahrens ME DeHeer CJ and Shoemaker DD 2006Distribution and prevalence of Wolbachia in introduced popula-tions of the fire ant Solenopsis invicta Insect Mol Biol 15 89ndash93
Briano JA Calcaterra LA Vander Meer R Valles SM and LivoreJP 2006 New survey for the fire ant microsporidia Vairimorphainvictae and Thelohania solenopsae in southern South Americawith observations on their field persistence and prevalence of dualinfections Environ Entomol 35 1358ndash1365
Buckley A 1999 Fire ants in California Am Bee J 139 88Carroll CR and Hoffman CA 2000 The pervasive ecological effects
of invasive species Exotic and native fire ants In Invertebrates asWeb-Masters in Ecosystems (Coleman DC and Hendrix PF Eds)Wallingford Oxon UK pp 221ndash232
Carruthers RI 2003 Invasive species research in the United StatesDepartment of Agriculture ndash Agricultural Research Service PestManag Sci 59 827ndash834
Chen JSC Shen CH and Lee HJ 2006 Monogynous and polygynousred imported fire ants Solenopsis invicta Buren (HymenopteraFormicidae) in Taiwan Environ Entomol 35 167ndash172
Cornuet JM and Luikart G 1996 Description and power analysis oftwo tests for detecting recent population bottlenecks from allelefrequency data Genetics 144 2001ndash2014
Davis LR Vander Meer RK and Porter SD 2001 Red imported fireants expand their range across the West Indies Florida Entomol84 735ndash736
DeHeer CJ and Tschinkel WR 1998 The success of alternativereproductive tactics in monogyne Solenopsis invicta significancefor transitions in social organization Behav Ecol 9 130ndash135
DeHeer CJ Goodisman MAD and Ross KG 1999 Queen dispersalstrategies in the multiple-queen form of the fire ant Solenopsisinvicta Am Nat 153 660ndash675
Dieringer D and Schlccediltterer C 2002 Microsatellite analyser (MSA) aplatform independent analysis tool for large microsatellite datasets Mol Ecol Notes 3 167ndash169
Drake JA Mooney HA Castri F Groves RH Kruger MRejmnek M and Williamson M 1989 Biological Invasions AGlobal Perspective Chichester UK 550 pp
Drees BM and Vinson SB 1990 Comparison of the control ofmonogynous and polygynous forms of the red imported fire ant(Hymenoptera Formicidae) with a chloripyrifos mound drench JEntomol Sci 25 317ndash324
Table 3 Tests for genetic bottlenecks using the infinite allele model(IAM) and two-phase model (TPM 90 stepwise mutation model and10 multistep mutation model [Garza and Williamson 2001])implemented in the program BOTTLENECK (a=005)Significance at given statistic level
Population Simulatedmodel
Significancetest
Standardizeddifferences test
Wilcoxontest
Taoyuan IAM 0148 0003 0015TPM 0186 0035 0023
Chiayi IAM 0037 0022 0007TPM 0521 0293 0281
Insect Soc Vol 55 2008 Research article 63
Drees BM and Gold RE 2003 Development of integrated pestmanagement programs for the red imported fire ant (Hymeno-ACHTUNGTRENNUNGptera Formicidae) J Entomol Sci 38 170ndash180
Eubanks MD 2001 Estimates of the direct and indirect effects of redimported fire ants on biological control in field crops Biol Control21 35 ndash43
Evanno G Regnaut S and Goudet J 2005 Detecting the number ofclusters of individuals using the software STRUCTURE asimulation study Mol Ecol 14 2611ndash2620
Excoffier L Smouse PE and Quattro JM 1992 Analysis of molecularvariance inferred from metric distances among DNA haplotypesApplication to human mitochondrial DNA restriction dataGenetics 131 479ndash491
Forys EA Quistorff A and Allen CR 2001 Potential fire ant(Hymenoptera Formicidae) impact on the endangered Schausswallowtail (Lepidoptera Papilionidae) Florida Entomol 84 254ndash258
Garza JC and Williamson EG 2001 Detection of reduction inpopulation size using data from microsatellite loci Mol Ecol 10 305ndash318
Giraud T Pedersen JS and Keller L 2002 Evolution of supercoloniesthe Argentine ants of southern Europe Proc Natl Acad Sci U SA 99 6075ndash6079
Giuliano WM Allen CR Lutz RS and Demarais S 1996 Effects ofred imported fire ants on northern bobwhite chicks J WildlManage 60 309ndash313
Glancey BM Craig CH Stringer CE and Bishop PM 1973Multiple fertile queens in colonies of the imported fire antSolenopsis invicta J Georgia Entomol Soc 8 327ndash328
Goodisman MAD DeHeer CJ and Ross KG 2000 Mating behaviorof queens from multiple-queen colonies of Solenopsis invicta JInsect Behav 13 455ndash468
Goodnight KF and Queller DC 1999 Computer software forperforming likelihood tests of pedigree relationship using geneticmarkers Mol Ecol 8 1231ndash1234
Gotelli NJ and Arnett AE 2000 Biogeographic effects of red fire antinvasion Ecol Lett 3 257ndash261
Gotzek D and Ross KG 2007 Genetic regulation of colony socialorganization in fire ants An integrative overview Q Rev Biol 82 201ndash226
Goudet J 2001 FSTAT A Program to Estimate and Test GeneDiversities and Fixation Indices Version 293 http wwwunilch izeasoftwares fstathtml
Greenberg L Fletcher DJC and Vinson SB 1985 Differences inworker size and mound distribution in monogynous and polygy-nous colonies of the fire ant Solenopsis invicta Buren J KansasEntomol Soc 58 9ndash18
Hampton JO Spencer PBS Alpers DL Twigg LE WoolnoughAP Doust J Higgs T and Pluske J 2004 Molecular techniqueswildlife management and the importance of genetic populationstructure and dispersal a case study with feral pigs J Appl Ecol41 735ndash743
Henshaw MT Kunzmann N Vanderwoude C Sanetra M andCrozier RH 2005 Population genetics and history of theintroduced fire ant Solenopsis invicta Buren (HymenopteraFormicidae) in Australia Aus J Entomol 44 37ndash44
Hccedillldobler B and Wilson EO 1990 The Ants The Belknap Press ofHarvard University Press Cambridge Massachusetts 732 pp
Huang TC Chou YC and Chou HC 2004 The infestation andcontrol of the red imported fire ant in Taiwan In Proceedingsof the Symposium on the Control of the Red Imported Fire AntBureau of Animal and Plant Health Inspection and QuarantineCouncil of Agriculture Executive Yuan Taipei Taiwan pp1 ndash 13
Jaquiry J Vogel V and Keller L 2005 Multilevel genetic analyses oftwo European supercolonies of the Argentine ant Linepithemahumile Mol Ecol 14 589ndash598
Jeyaprakash A and Hoy MA 2000 Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropods Insect Mol Biol 9 393ndash405
Jouvenaz DP 1990 Approaches to biological control of fire ants in theUnited States In Applied Myrmecology A World Perspective(Vander Meer RK Jaffe K and Cedeno A Eds) Westview PressBoulder Colorado pp 620ndash627
Kaspari M 2000 Do imported fire ants impact canopy arthropodsEvidence from simple arboreal pitfall traps Southwestern Nat 45 118ndash122
Keller L and Ross KG 1998 Selfish genes a green beard in the red fireant Nature 394 573ndash575
Kemp SF deShazo RD Moffitt JE Williams DF and Buhner WA2000 Expanding habitat of the imported fire ant (Solenopsisinvicta) A public health concern J Allergy Clin Immunol 105 683ndash691
Kopachena JG Buckley AJ and Potts GA 2000 Effects of the redimported fire ant (Solenopsis invicta) on reproductive success ofbarn swallows (Hirundo rustica) in northeast Texas SouthwesternNat 45 477ndash482
Krieger MJB and Keller L 1997 Polymorphism at dinucleotidemicrosatellite loci in fire ant Solenopsis invicta populations MolEcol 6 997ndash999
Krieger MJB Ross KG Chang CWY and Keller L 1999Frequency and origin of triploidy in the fire ant Solenopsis invictaHeredity 82 142ndash150
Krieger MJB and Ross KG 2002 Identification of a major generegulating complex social behavior Science 295 328ndash332
Lee CE 2002 Evolutionary genetics of invasive species Trends EcolEvol 17 386ndash391
Lofgren CS Banks WA and Glancey BM 1975 Biology and controlof imported fire ants Annu Rev Entomol 20 1ndash30
Lofgren CS 1986 History of imported fire ants in the United States InFire Ants and Leaf Cutting Ants Biology and Management(Lofgren CS and Vander Meer RK Eds) Westview PressBoulder Colorado pp 36ndash49
Luikart G Allendorf FW Cornuet JM and Sherwin WB 1998Distortion of allele frequency distributions provides a test forrecent population bottlenecks J Hered 89 238ndash247
Luikart G and Cornuet JM 1998 Empirical evaluation of a test foridentifying recently bottlenecked populations from allele frequen-cy data Conserv Biol 12 228ndash237
MacKay WP and Fagerlund R 1997 Range expansion of the redimported fire ant Solenopsis invicta Buren (Hymenoptera For-micidae) into New Mexico and extreme western Texas ProcEntomol Soc Wash 99 757ndash758
Mank JE and Avise JC 2004 Individual organisms as units of analysisBayesian-clustering alternatives in population genetics Genet Res84 135ndash143
May GE Gelembiuk GW Panov VE Orlova MI and Lee CE2006 Molecular ecology of zebra mussel invasions Mol Ecol 15 1021ndash1031
McCubbin KI and Weiner JM 2002 Fire ants in Australia A newmedical and ecological hazard Med J Aus 176 518ndash519
Morrison LW 2000 Mechanisms of interspecific competition amongan invasive and two native fire ants Oikos 90 238ndash252
Morrison LW 2002 Long-term impacts of an arthropod-communityinvasion by the imported fire ant Solenopsis invicta Ecology 83 2337ndash2345
Nattrass R and Vanderwoude C 2001 A preliminary investigation ofthe ecological effects of red imported fire ants (Solenopsis invicta)in Brisbane Ecol Manag Resto 2 220ndash223
Nei M 1972 Genetic distance between populations Am Nat 106 283ndash292
Orr MR 1996 Host manipulation by Wolbachia is a neutral trait withinsingle populations Anim Behav 51 1183ndash1185
Patterson RS 1994 Biological control of introduced ant species InExotic Ants Biology Impact and Control of Introduced Species(Williams DF Ed) Westview Press Boulder Colorado pp 293ndash308
Piry S Luikart G and Cornuet JM 1999 BOTTLENECK acomputer program for detecting recent reductions in the effectivepopulation size using allele frequency data J Hered 90 502ndash503
64 CC Yang et al Fire ants in Taiwan
Porter SD and Savignano DA 1990 Invasion of polygyne fire antsdecimates native ants and disrupts arthropod community Ecology7 2095ndash2106
Porter SD Bhatkar A Mulder R Vinson SB and Clair DJ 1991Distribution and density of polygyne fire ants (HymenopteraFormicidae) in Texas J Econ Entomol 84 866ndash874
Porter SD Fowler HG and MacKay WP 1992 Fire ant mounddensities in the United States and Brazil (Hymenoptera Formi-cidae) J Econ Entomol 85 1154ndash1161
Porter SD Williams DF Patterson RS and Fowler HG 1997Intercontinental differences in the abundance of Solenopsis fireants (Hymenoptera Formicidae) Escape from natural enemiesEnviron Entomol 26 373ndash384
Pritchard JK Stephens M and Donnelly P 2000 Inference ofpopulation structure using multilocus genotype data Genetics155 945ndash959
Raymond M and Rousset F 1995a An exact test for populationdifferentiation Evolution 49 1280ndash1283
Raymond M and Rousset F 1995b GENEPOP (version 12)population genetics software for exact tests and ecumenicism JHered 86 248ndash249
Reuter M Pedersen JS and Keller L 2005 Loss of Wolbachiainfection during colonisation in the invasive Argentine ant Line-pithema humile Heredity 94 364ndash369
Ross KG and Fletcher DJC 1985a Genetic origin of male diploidy inthe fire ant Solenopsis invicta (Hymenoptera Formicidae) and itsevolutionary significance Evolution 39 888ndash903
Ross KG and Fletcher DJC 1985b Comparative study of genetic andsocial structure in two forms of the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 17 349ndash356
Ross KG and Fletcher DJC 1986 Diploid male productionndashasignificant colony mortality factor in the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 19 283ndash291
Ross KG and Shoemaker DD 1993 An unusual pattern of gene flowbetween the two social forms of the fire ant Solenopsis invictaEvolution 47 1595ndash1605
Ross KG Vargo EL Keller L and Trager JC 1993 Effect of afounder event on variation in the genetic sex-determining system ofthe fire ant Solenopsis invicta Genetics 135 843ndash854
Ross KG and Keller L 1995 Joint influence of gene flow and selectionon a reproductively important genetic polymorphism in the fire antSolenopsis invicta Am Nat 146 325ndash348
Ross KG Vargo EL and Keller L 1996 Social evolution in a newenvironment The case of introduced fire ants Proc Natl AcadSci U S A 93 3021ndash3025
Ross KG 1997 Multilocus evolution in fire ants Effects of selectiongene flow and recombination Genetics 145 961ndash974
Ross KG Krieger MJB Shoemaker DD Vargo EL and Keller L1997 Hierarchical analysis of genetic structure in native fire antpopulations Results from three classes of molecular markersGenetics 147 643ndash655
Ross KG and Shoemaker DD 1997 Nuclear and mitochondrialgenetic structure in two social forms of the fire ant Solenopsisinvicta Insights into transitions to an alternate social organizationHeredity 78 590ndash602
Ross KG and Keller L 1998 Genetic control of social organization inan ant Proc Natl Acad Sci U S A 95 14232ndash14237
Ross KG Shoemaker DD Krieger MJ DeHeer CJ and Keller L1999 Assessing genetic structure with multiple classes of molecularmarkers A case study involving the introduced fire ant Solenopsisinvicta Mol Biol Evol 16 525ndash543
Ross KG and Keller L 2002 Experimental conversion of colony socialorganization by manipulation of worker genotype composition infire ants (Solenopsis invicta) Behav Ecol Sociobiol 51 287ndash295
Ross KG Krieger MJB and Shoemaker DD 2003 Alternativegenetic foundations and evolutionary routes for a key socialpolymorphism in fire ants Genetics 165 1853ndash1867
Ross KG Krieger MJB Keller L and Shoemaker DD 2007Genetic variation and structure in native populations of the fire antSolenopsis invicta Evolutionary and demographic implicationsBiol J Linnean Soc 92 541ndash560
Rousset F 1997 Genetic differentiation and estimation of gene flowfrom F-statistics under isolation by distance Genetics 145 1219ndash1228
Sax DF Stachowicz JJ and Gaines SD 2005 Species InvasionsInsights into Ecology Evolution and Biogeography SinauerAssociates Sunderland Massachussetts 495 pp
Shoemaker DD and Ross KG 1996 Effects of social organization ongene flow in the fire ant Solenopsis invicta Nature 383 613ndash616
Shoemaker DD Ross KG Keller L Vargo EL and Werren JH2000 Wolbachia infections in native and introduced populations offire ants (Solenopsis spp) Insect Mol Biol 9 661ndash673
Shoemaker DD Ahrens M Sheill L Mescher M Keller L and RossKG 2003 Distribution and prevalence of Wolbachia infections innative populations of the fire ant Solenopsis invicta (HymenopteraFormicidae) Environ Entomol 32 1329ndash1336
Shoemaker DD DeHeer CJ Krieger MJB and Ross KG 2006Population genetics of the invasive fire ant Solenopsis invicta(Hymenoptera Formicidae) in the USA Ann Entomol Soc Am99 1213ndash1233
Simon C Frati F Beckenbach A Crespi B Liu H and Flook P 1994Evolution weighting and phylogenetic utility of mitochondrialgene sequences and a compilation of conserved polymerase chainreaction primers Ann Entomol Soc Am 87 651ndash701
Toom PM Cupp E Johnson CP and Griffin I 1976 Utilization ofbody reserves for minim brood development by queens of theimported fire ant Solenopsis invicta J Insect Physiol 22 217ndash220
Tsutsui ND and Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepethema humile) in its nativeand introduced ranges Evolution 55 976ndash985
Tsutsui ND Kauppinen SN Oyafuso AF and Grosberg RK 2003The distribution and evolutionary history of Wolbachia infection innative and introduced populations of the invasive argentine ant(Linepithema humile) Mol Ecol 12 3057ndash3068
Valles SM and Porter SD 2003 Identification of polygyne andmonogyne fire ant colonies (Solenopsis invicta) by multiplex PCRof Gp-9 alleles Insect Soc 50 199ndash200
Valles SM Oi DH Briano JA and Williams DF 2004 Simultaneousdetection of Vairimorpha invictae (Microsporidia Burenellidae)and Thelohania solenopsae (Microsporidia Thelohaniidae) in fireants by PCR Florida Entomol 87 85ndash87
Vander Meer R K and Porter SD 2001 Fate of newly-mated queensintroduced into monogyne and polygyne Solenopsis invicta (Hy-menoptera Formicidae) colonies Ann Entomol Soc Am 94 289ndash297
Vander Meer RK Jaffe K and Cedeno A 1990 Applied Myrmecol-ogy A World Perspective Westview Press Boulder CO 741 pp
Vinson SB 1994 Impact and invasion of Solenopsis invicta (Buren) onnative food webs In Exotic Ants Biology Impact and Control ofIntroduced Species (Williams DF Ed) Westview Press BoulderColorado pp 293ndash308
Vinson SB 1997 Invasion of the red imported fire ant (HymenopteraFormicidae) Spread biology and impact Am Entomol 43 23ndash29
Williams DF and Porter SD 1994 Fire ant control Science 264 1653ndash1653
Wojcik D Allen CR Brenner RJ Forys EA Jouvenaz D and LutzRS 2001 Red imported fire ants Impact on biodiversity AmEntomol 47 16ndash23
Xia X and Xie Z 2001 DAMBE Data analysis in molecular biologyand evolution J Hered 92 371ndash373
Insect Soc Vol 55 2008 Research article 65

Genetic differentiation between Taoyuan and Chiayipopulations
All analyses of population differentiation using themicrosatellite and mtDNA data indicate that the twogeographic populations are strongly differentiated andthus likely originated from separate invasions Whilemultiple lines of evidence support this notion thestrongest evidence is the confinement each of the threemtDNA variants to only one of the two geographicpopulations Interestingly all three variants found inTaiwan are the same three variants commonly found inants from the USA (Shoemaker et al 2006) China(GenBank accession numbers DQ831670-DQ831672)and Australia (CCY and DDS unpublished results) Theoccurrence of these three variants in all recently invadedareas suggests that ants from Taiwan could have origi-nated from any of these other areas We currently aregenerating the necessary baseline data for ants from all ofthese introduced regions as well as from populations intheir native range to infer the source population andinvasion history of ants in Taiwan and these other areas
From the perspective of managing fire ants in Taiwantreating each infested area as a separate management unitwould appear more reasonable and means that scientistsprobably should focus on separate plans for eliminationof two isolated populations (Hampton et al 2004) Onthe other hand because these two areas are at high risk ofconnectivity via long distance human-mediated dispersalcare must be taken to prevent reinvasion into these areaseven after successful eradication Stringent inter-countyquarantine measures such as movement control shouldbe established to monitor the risk of transportation of fireant-infested goods to prevent jump dispersal between thetwo areas and decrease probability of successful erad-ication (Drees and Gold 2003)
Acknowledgments
The authors would like to thank Chung-Chi Lin for providing most ofthe specimens for this study including the first identified samples inTaoyuan and Chiayi counties We are particularly grateful to Hwei-yuChang Hon-Tsen Yu and Chia-Hung Hsieh for helpful discussions andcomments on earlier versions of this manuscript Comments by twoanonymous reviewers greatly improved this manuscript This research
was financially supported by grants from Academia Sinica Taiwan Theuse of trade firm or corporation names in this publication is for theinformation and convenience of the reader Such use does notconstitute an official endorsement or approval by the United StatesDepartment of Agriculture or the Agricultural Research Service of anyproduct or service to the exclusion of others that may be suitable
References
Ahrens ME Ross KG and Shoemaker DD 2005 Phylogeographicstructure of the fire ant Solenopsis invicta in its native SouthAmerican range Roles of natural barriers and habitat connectivityEvolution 59 1733ndash1743
Allen CR Demarais S and Lutz RS 1994 Red imported fire antimpact on wildlife an overview Texas J Sci 46 51ndash59
Allen CR Lutz RS and Demarais S 1998 Ecological effects of theinvasive nonindigenous ant Solenopsis invicta on native verte-brates The wheels on the bus Transactions of the North AmericanWildlife and Natural Resources Conference 63 56 ndash65
Allen CR Willey RD Myers PE Horton PM and Buffa J 2000Impact of red imported fire ant infestation on northern bobwhitequail abundance trends in southeastern United States J AgrUrban Entomol 17 43ndash51
Allen CR Forys EA Rice KG and Wojcik DP 2001 Effects of fireants (Hymenoptera Formicidae) on hatching turtles and preva-lence of fire ants on sea turtle nesting beaches in Florida FloridaEntomol 84 250ndash253
Bouwma AM Ahrens ME DeHeer CJ and Shoemaker DD 2006Distribution and prevalence of Wolbachia in introduced popula-tions of the fire ant Solenopsis invicta Insect Mol Biol 15 89ndash93
Briano JA Calcaterra LA Vander Meer R Valles SM and LivoreJP 2006 New survey for the fire ant microsporidia Vairimorphainvictae and Thelohania solenopsae in southern South Americawith observations on their field persistence and prevalence of dualinfections Environ Entomol 35 1358ndash1365
Buckley A 1999 Fire ants in California Am Bee J 139 88Carroll CR and Hoffman CA 2000 The pervasive ecological effects
of invasive species Exotic and native fire ants In Invertebrates asWeb-Masters in Ecosystems (Coleman DC and Hendrix PF Eds)Wallingford Oxon UK pp 221ndash232
Carruthers RI 2003 Invasive species research in the United StatesDepartment of Agriculture ndash Agricultural Research Service PestManag Sci 59 827ndash834
Chen JSC Shen CH and Lee HJ 2006 Monogynous and polygynousred imported fire ants Solenopsis invicta Buren (HymenopteraFormicidae) in Taiwan Environ Entomol 35 167ndash172
Cornuet JM and Luikart G 1996 Description and power analysis oftwo tests for detecting recent population bottlenecks from allelefrequency data Genetics 144 2001ndash2014
Davis LR Vander Meer RK and Porter SD 2001 Red imported fireants expand their range across the West Indies Florida Entomol84 735ndash736
DeHeer CJ and Tschinkel WR 1998 The success of alternativereproductive tactics in monogyne Solenopsis invicta significancefor transitions in social organization Behav Ecol 9 130ndash135
DeHeer CJ Goodisman MAD and Ross KG 1999 Queen dispersalstrategies in the multiple-queen form of the fire ant Solenopsisinvicta Am Nat 153 660ndash675
Dieringer D and Schlccediltterer C 2002 Microsatellite analyser (MSA) aplatform independent analysis tool for large microsatellite datasets Mol Ecol Notes 3 167ndash169
Drake JA Mooney HA Castri F Groves RH Kruger MRejmnek M and Williamson M 1989 Biological Invasions AGlobal Perspective Chichester UK 550 pp
Drees BM and Vinson SB 1990 Comparison of the control ofmonogynous and polygynous forms of the red imported fire ant(Hymenoptera Formicidae) with a chloripyrifos mound drench JEntomol Sci 25 317ndash324
Table 3 Tests for genetic bottlenecks using the infinite allele model(IAM) and two-phase model (TPM 90 stepwise mutation model and10 multistep mutation model [Garza and Williamson 2001])implemented in the program BOTTLENECK (a=005)Significance at given statistic level
Population Simulatedmodel
Significancetest
Standardizeddifferences test
Wilcoxontest
Taoyuan IAM 0148 0003 0015TPM 0186 0035 0023
Chiayi IAM 0037 0022 0007TPM 0521 0293 0281
Insect Soc Vol 55 2008 Research article 63
Drees BM and Gold RE 2003 Development of integrated pestmanagement programs for the red imported fire ant (Hymeno-ACHTUNGTRENNUNGptera Formicidae) J Entomol Sci 38 170ndash180
Eubanks MD 2001 Estimates of the direct and indirect effects of redimported fire ants on biological control in field crops Biol Control21 35 ndash43
Evanno G Regnaut S and Goudet J 2005 Detecting the number ofclusters of individuals using the software STRUCTURE asimulation study Mol Ecol 14 2611ndash2620
Excoffier L Smouse PE and Quattro JM 1992 Analysis of molecularvariance inferred from metric distances among DNA haplotypesApplication to human mitochondrial DNA restriction dataGenetics 131 479ndash491
Forys EA Quistorff A and Allen CR 2001 Potential fire ant(Hymenoptera Formicidae) impact on the endangered Schausswallowtail (Lepidoptera Papilionidae) Florida Entomol 84 254ndash258
Garza JC and Williamson EG 2001 Detection of reduction inpopulation size using data from microsatellite loci Mol Ecol 10 305ndash318
Giraud T Pedersen JS and Keller L 2002 Evolution of supercoloniesthe Argentine ants of southern Europe Proc Natl Acad Sci U SA 99 6075ndash6079
Giuliano WM Allen CR Lutz RS and Demarais S 1996 Effects ofred imported fire ants on northern bobwhite chicks J WildlManage 60 309ndash313
Glancey BM Craig CH Stringer CE and Bishop PM 1973Multiple fertile queens in colonies of the imported fire antSolenopsis invicta J Georgia Entomol Soc 8 327ndash328
Goodisman MAD DeHeer CJ and Ross KG 2000 Mating behaviorof queens from multiple-queen colonies of Solenopsis invicta JInsect Behav 13 455ndash468
Goodnight KF and Queller DC 1999 Computer software forperforming likelihood tests of pedigree relationship using geneticmarkers Mol Ecol 8 1231ndash1234
Gotelli NJ and Arnett AE 2000 Biogeographic effects of red fire antinvasion Ecol Lett 3 257ndash261
Gotzek D and Ross KG 2007 Genetic regulation of colony socialorganization in fire ants An integrative overview Q Rev Biol 82 201ndash226
Goudet J 2001 FSTAT A Program to Estimate and Test GeneDiversities and Fixation Indices Version 293 http wwwunilch izeasoftwares fstathtml
Greenberg L Fletcher DJC and Vinson SB 1985 Differences inworker size and mound distribution in monogynous and polygy-nous colonies of the fire ant Solenopsis invicta Buren J KansasEntomol Soc 58 9ndash18
Hampton JO Spencer PBS Alpers DL Twigg LE WoolnoughAP Doust J Higgs T and Pluske J 2004 Molecular techniqueswildlife management and the importance of genetic populationstructure and dispersal a case study with feral pigs J Appl Ecol41 735ndash743
Henshaw MT Kunzmann N Vanderwoude C Sanetra M andCrozier RH 2005 Population genetics and history of theintroduced fire ant Solenopsis invicta Buren (HymenopteraFormicidae) in Australia Aus J Entomol 44 37ndash44
Hccedillldobler B and Wilson EO 1990 The Ants The Belknap Press ofHarvard University Press Cambridge Massachusetts 732 pp
Huang TC Chou YC and Chou HC 2004 The infestation andcontrol of the red imported fire ant in Taiwan In Proceedingsof the Symposium on the Control of the Red Imported Fire AntBureau of Animal and Plant Health Inspection and QuarantineCouncil of Agriculture Executive Yuan Taipei Taiwan pp1 ndash 13
Jaquiry J Vogel V and Keller L 2005 Multilevel genetic analyses oftwo European supercolonies of the Argentine ant Linepithemahumile Mol Ecol 14 589ndash598
Jeyaprakash A and Hoy MA 2000 Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropods Insect Mol Biol 9 393ndash405
Jouvenaz DP 1990 Approaches to biological control of fire ants in theUnited States In Applied Myrmecology A World Perspective(Vander Meer RK Jaffe K and Cedeno A Eds) Westview PressBoulder Colorado pp 620ndash627
Kaspari M 2000 Do imported fire ants impact canopy arthropodsEvidence from simple arboreal pitfall traps Southwestern Nat 45 118ndash122
Keller L and Ross KG 1998 Selfish genes a green beard in the red fireant Nature 394 573ndash575
Kemp SF deShazo RD Moffitt JE Williams DF and Buhner WA2000 Expanding habitat of the imported fire ant (Solenopsisinvicta) A public health concern J Allergy Clin Immunol 105 683ndash691
Kopachena JG Buckley AJ and Potts GA 2000 Effects of the redimported fire ant (Solenopsis invicta) on reproductive success ofbarn swallows (Hirundo rustica) in northeast Texas SouthwesternNat 45 477ndash482
Krieger MJB and Keller L 1997 Polymorphism at dinucleotidemicrosatellite loci in fire ant Solenopsis invicta populations MolEcol 6 997ndash999
Krieger MJB Ross KG Chang CWY and Keller L 1999Frequency and origin of triploidy in the fire ant Solenopsis invictaHeredity 82 142ndash150
Krieger MJB and Ross KG 2002 Identification of a major generegulating complex social behavior Science 295 328ndash332
Lee CE 2002 Evolutionary genetics of invasive species Trends EcolEvol 17 386ndash391
Lofgren CS Banks WA and Glancey BM 1975 Biology and controlof imported fire ants Annu Rev Entomol 20 1ndash30
Lofgren CS 1986 History of imported fire ants in the United States InFire Ants and Leaf Cutting Ants Biology and Management(Lofgren CS and Vander Meer RK Eds) Westview PressBoulder Colorado pp 36ndash49
Luikart G Allendorf FW Cornuet JM and Sherwin WB 1998Distortion of allele frequency distributions provides a test forrecent population bottlenecks J Hered 89 238ndash247
Luikart G and Cornuet JM 1998 Empirical evaluation of a test foridentifying recently bottlenecked populations from allele frequen-cy data Conserv Biol 12 228ndash237
MacKay WP and Fagerlund R 1997 Range expansion of the redimported fire ant Solenopsis invicta Buren (Hymenoptera For-micidae) into New Mexico and extreme western Texas ProcEntomol Soc Wash 99 757ndash758
Mank JE and Avise JC 2004 Individual organisms as units of analysisBayesian-clustering alternatives in population genetics Genet Res84 135ndash143
May GE Gelembiuk GW Panov VE Orlova MI and Lee CE2006 Molecular ecology of zebra mussel invasions Mol Ecol 15 1021ndash1031
McCubbin KI and Weiner JM 2002 Fire ants in Australia A newmedical and ecological hazard Med J Aus 176 518ndash519
Morrison LW 2000 Mechanisms of interspecific competition amongan invasive and two native fire ants Oikos 90 238ndash252
Morrison LW 2002 Long-term impacts of an arthropod-communityinvasion by the imported fire ant Solenopsis invicta Ecology 83 2337ndash2345
Nattrass R and Vanderwoude C 2001 A preliminary investigation ofthe ecological effects of red imported fire ants (Solenopsis invicta)in Brisbane Ecol Manag Resto 2 220ndash223
Nei M 1972 Genetic distance between populations Am Nat 106 283ndash292
Orr MR 1996 Host manipulation by Wolbachia is a neutral trait withinsingle populations Anim Behav 51 1183ndash1185
Patterson RS 1994 Biological control of introduced ant species InExotic Ants Biology Impact and Control of Introduced Species(Williams DF Ed) Westview Press Boulder Colorado pp 293ndash308
Piry S Luikart G and Cornuet JM 1999 BOTTLENECK acomputer program for detecting recent reductions in the effectivepopulation size using allele frequency data J Hered 90 502ndash503
64 CC Yang et al Fire ants in Taiwan
Porter SD and Savignano DA 1990 Invasion of polygyne fire antsdecimates native ants and disrupts arthropod community Ecology7 2095ndash2106
Porter SD Bhatkar A Mulder R Vinson SB and Clair DJ 1991Distribution and density of polygyne fire ants (HymenopteraFormicidae) in Texas J Econ Entomol 84 866ndash874
Porter SD Fowler HG and MacKay WP 1992 Fire ant mounddensities in the United States and Brazil (Hymenoptera Formi-cidae) J Econ Entomol 85 1154ndash1161
Porter SD Williams DF Patterson RS and Fowler HG 1997Intercontinental differences in the abundance of Solenopsis fireants (Hymenoptera Formicidae) Escape from natural enemiesEnviron Entomol 26 373ndash384
Pritchard JK Stephens M and Donnelly P 2000 Inference ofpopulation structure using multilocus genotype data Genetics155 945ndash959
Raymond M and Rousset F 1995a An exact test for populationdifferentiation Evolution 49 1280ndash1283
Raymond M and Rousset F 1995b GENEPOP (version 12)population genetics software for exact tests and ecumenicism JHered 86 248ndash249
Reuter M Pedersen JS and Keller L 2005 Loss of Wolbachiainfection during colonisation in the invasive Argentine ant Line-pithema humile Heredity 94 364ndash369
Ross KG and Fletcher DJC 1985a Genetic origin of male diploidy inthe fire ant Solenopsis invicta (Hymenoptera Formicidae) and itsevolutionary significance Evolution 39 888ndash903
Ross KG and Fletcher DJC 1985b Comparative study of genetic andsocial structure in two forms of the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 17 349ndash356
Ross KG and Fletcher DJC 1986 Diploid male productionndashasignificant colony mortality factor in the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 19 283ndash291
Ross KG and Shoemaker DD 1993 An unusual pattern of gene flowbetween the two social forms of the fire ant Solenopsis invictaEvolution 47 1595ndash1605
Ross KG Vargo EL Keller L and Trager JC 1993 Effect of afounder event on variation in the genetic sex-determining system ofthe fire ant Solenopsis invicta Genetics 135 843ndash854
Ross KG and Keller L 1995 Joint influence of gene flow and selectionon a reproductively important genetic polymorphism in the fire antSolenopsis invicta Am Nat 146 325ndash348
Ross KG Vargo EL and Keller L 1996 Social evolution in a newenvironment The case of introduced fire ants Proc Natl AcadSci U S A 93 3021ndash3025
Ross KG 1997 Multilocus evolution in fire ants Effects of selectiongene flow and recombination Genetics 145 961ndash974
Ross KG Krieger MJB Shoemaker DD Vargo EL and Keller L1997 Hierarchical analysis of genetic structure in native fire antpopulations Results from three classes of molecular markersGenetics 147 643ndash655
Ross KG and Shoemaker DD 1997 Nuclear and mitochondrialgenetic structure in two social forms of the fire ant Solenopsisinvicta Insights into transitions to an alternate social organizationHeredity 78 590ndash602
Ross KG and Keller L 1998 Genetic control of social organization inan ant Proc Natl Acad Sci U S A 95 14232ndash14237
Ross KG Shoemaker DD Krieger MJ DeHeer CJ and Keller L1999 Assessing genetic structure with multiple classes of molecularmarkers A case study involving the introduced fire ant Solenopsisinvicta Mol Biol Evol 16 525ndash543
Ross KG and Keller L 2002 Experimental conversion of colony socialorganization by manipulation of worker genotype composition infire ants (Solenopsis invicta) Behav Ecol Sociobiol 51 287ndash295
Ross KG Krieger MJB and Shoemaker DD 2003 Alternativegenetic foundations and evolutionary routes for a key socialpolymorphism in fire ants Genetics 165 1853ndash1867
Ross KG Krieger MJB Keller L and Shoemaker DD 2007Genetic variation and structure in native populations of the fire antSolenopsis invicta Evolutionary and demographic implicationsBiol J Linnean Soc 92 541ndash560
Rousset F 1997 Genetic differentiation and estimation of gene flowfrom F-statistics under isolation by distance Genetics 145 1219ndash1228
Sax DF Stachowicz JJ and Gaines SD 2005 Species InvasionsInsights into Ecology Evolution and Biogeography SinauerAssociates Sunderland Massachussetts 495 pp
Shoemaker DD and Ross KG 1996 Effects of social organization ongene flow in the fire ant Solenopsis invicta Nature 383 613ndash616
Shoemaker DD Ross KG Keller L Vargo EL and Werren JH2000 Wolbachia infections in native and introduced populations offire ants (Solenopsis spp) Insect Mol Biol 9 661ndash673
Shoemaker DD Ahrens M Sheill L Mescher M Keller L and RossKG 2003 Distribution and prevalence of Wolbachia infections innative populations of the fire ant Solenopsis invicta (HymenopteraFormicidae) Environ Entomol 32 1329ndash1336
Shoemaker DD DeHeer CJ Krieger MJB and Ross KG 2006Population genetics of the invasive fire ant Solenopsis invicta(Hymenoptera Formicidae) in the USA Ann Entomol Soc Am99 1213ndash1233
Simon C Frati F Beckenbach A Crespi B Liu H and Flook P 1994Evolution weighting and phylogenetic utility of mitochondrialgene sequences and a compilation of conserved polymerase chainreaction primers Ann Entomol Soc Am 87 651ndash701
Toom PM Cupp E Johnson CP and Griffin I 1976 Utilization ofbody reserves for minim brood development by queens of theimported fire ant Solenopsis invicta J Insect Physiol 22 217ndash220
Tsutsui ND and Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepethema humile) in its nativeand introduced ranges Evolution 55 976ndash985
Tsutsui ND Kauppinen SN Oyafuso AF and Grosberg RK 2003The distribution and evolutionary history of Wolbachia infection innative and introduced populations of the invasive argentine ant(Linepithema humile) Mol Ecol 12 3057ndash3068
Valles SM and Porter SD 2003 Identification of polygyne andmonogyne fire ant colonies (Solenopsis invicta) by multiplex PCRof Gp-9 alleles Insect Soc 50 199ndash200
Valles SM Oi DH Briano JA and Williams DF 2004 Simultaneousdetection of Vairimorpha invictae (Microsporidia Burenellidae)and Thelohania solenopsae (Microsporidia Thelohaniidae) in fireants by PCR Florida Entomol 87 85ndash87
Vander Meer R K and Porter SD 2001 Fate of newly-mated queensintroduced into monogyne and polygyne Solenopsis invicta (Hy-menoptera Formicidae) colonies Ann Entomol Soc Am 94 289ndash297
Vander Meer RK Jaffe K and Cedeno A 1990 Applied Myrmecol-ogy A World Perspective Westview Press Boulder CO 741 pp
Vinson SB 1994 Impact and invasion of Solenopsis invicta (Buren) onnative food webs In Exotic Ants Biology Impact and Control ofIntroduced Species (Williams DF Ed) Westview Press BoulderColorado pp 293ndash308
Vinson SB 1997 Invasion of the red imported fire ant (HymenopteraFormicidae) Spread biology and impact Am Entomol 43 23ndash29
Williams DF and Porter SD 1994 Fire ant control Science 264 1653ndash1653
Wojcik D Allen CR Brenner RJ Forys EA Jouvenaz D and LutzRS 2001 Red imported fire ants Impact on biodiversity AmEntomol 47 16ndash23
Xia X and Xie Z 2001 DAMBE Data analysis in molecular biologyand evolution J Hered 92 371ndash373
Insect Soc Vol 55 2008 Research article 65

Drees BM and Gold RE 2003 Development of integrated pestmanagement programs for the red imported fire ant (Hymeno-ACHTUNGTRENNUNGptera Formicidae) J Entomol Sci 38 170ndash180
Eubanks MD 2001 Estimates of the direct and indirect effects of redimported fire ants on biological control in field crops Biol Control21 35 ndash43
Evanno G Regnaut S and Goudet J 2005 Detecting the number ofclusters of individuals using the software STRUCTURE asimulation study Mol Ecol 14 2611ndash2620
Excoffier L Smouse PE and Quattro JM 1992 Analysis of molecularvariance inferred from metric distances among DNA haplotypesApplication to human mitochondrial DNA restriction dataGenetics 131 479ndash491
Forys EA Quistorff A and Allen CR 2001 Potential fire ant(Hymenoptera Formicidae) impact on the endangered Schausswallowtail (Lepidoptera Papilionidae) Florida Entomol 84 254ndash258
Garza JC and Williamson EG 2001 Detection of reduction inpopulation size using data from microsatellite loci Mol Ecol 10 305ndash318
Giraud T Pedersen JS and Keller L 2002 Evolution of supercoloniesthe Argentine ants of southern Europe Proc Natl Acad Sci U SA 99 6075ndash6079
Giuliano WM Allen CR Lutz RS and Demarais S 1996 Effects ofred imported fire ants on northern bobwhite chicks J WildlManage 60 309ndash313
Glancey BM Craig CH Stringer CE and Bishop PM 1973Multiple fertile queens in colonies of the imported fire antSolenopsis invicta J Georgia Entomol Soc 8 327ndash328
Goodisman MAD DeHeer CJ and Ross KG 2000 Mating behaviorof queens from multiple-queen colonies of Solenopsis invicta JInsect Behav 13 455ndash468
Goodnight KF and Queller DC 1999 Computer software forperforming likelihood tests of pedigree relationship using geneticmarkers Mol Ecol 8 1231ndash1234
Gotelli NJ and Arnett AE 2000 Biogeographic effects of red fire antinvasion Ecol Lett 3 257ndash261
Gotzek D and Ross KG 2007 Genetic regulation of colony socialorganization in fire ants An integrative overview Q Rev Biol 82 201ndash226
Goudet J 2001 FSTAT A Program to Estimate and Test GeneDiversities and Fixation Indices Version 293 http wwwunilch izeasoftwares fstathtml
Greenberg L Fletcher DJC and Vinson SB 1985 Differences inworker size and mound distribution in monogynous and polygy-nous colonies of the fire ant Solenopsis invicta Buren J KansasEntomol Soc 58 9ndash18
Hampton JO Spencer PBS Alpers DL Twigg LE WoolnoughAP Doust J Higgs T and Pluske J 2004 Molecular techniqueswildlife management and the importance of genetic populationstructure and dispersal a case study with feral pigs J Appl Ecol41 735ndash743
Henshaw MT Kunzmann N Vanderwoude C Sanetra M andCrozier RH 2005 Population genetics and history of theintroduced fire ant Solenopsis invicta Buren (HymenopteraFormicidae) in Australia Aus J Entomol 44 37ndash44
Hccedillldobler B and Wilson EO 1990 The Ants The Belknap Press ofHarvard University Press Cambridge Massachusetts 732 pp
Huang TC Chou YC and Chou HC 2004 The infestation andcontrol of the red imported fire ant in Taiwan In Proceedingsof the Symposium on the Control of the Red Imported Fire AntBureau of Animal and Plant Health Inspection and QuarantineCouncil of Agriculture Executive Yuan Taipei Taiwan pp1 ndash 13
Jaquiry J Vogel V and Keller L 2005 Multilevel genetic analyses oftwo European supercolonies of the Argentine ant Linepithemahumile Mol Ecol 14 589ndash598
Jeyaprakash A and Hoy MA 2000 Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropods Insect Mol Biol 9 393ndash405
Jouvenaz DP 1990 Approaches to biological control of fire ants in theUnited States In Applied Myrmecology A World Perspective(Vander Meer RK Jaffe K and Cedeno A Eds) Westview PressBoulder Colorado pp 620ndash627
Kaspari M 2000 Do imported fire ants impact canopy arthropodsEvidence from simple arboreal pitfall traps Southwestern Nat 45 118ndash122
Keller L and Ross KG 1998 Selfish genes a green beard in the red fireant Nature 394 573ndash575
Kemp SF deShazo RD Moffitt JE Williams DF and Buhner WA2000 Expanding habitat of the imported fire ant (Solenopsisinvicta) A public health concern J Allergy Clin Immunol 105 683ndash691
Kopachena JG Buckley AJ and Potts GA 2000 Effects of the redimported fire ant (Solenopsis invicta) on reproductive success ofbarn swallows (Hirundo rustica) in northeast Texas SouthwesternNat 45 477ndash482
Krieger MJB and Keller L 1997 Polymorphism at dinucleotidemicrosatellite loci in fire ant Solenopsis invicta populations MolEcol 6 997ndash999
Krieger MJB Ross KG Chang CWY and Keller L 1999Frequency and origin of triploidy in the fire ant Solenopsis invictaHeredity 82 142ndash150
Krieger MJB and Ross KG 2002 Identification of a major generegulating complex social behavior Science 295 328ndash332
Lee CE 2002 Evolutionary genetics of invasive species Trends EcolEvol 17 386ndash391
Lofgren CS Banks WA and Glancey BM 1975 Biology and controlof imported fire ants Annu Rev Entomol 20 1ndash30
Lofgren CS 1986 History of imported fire ants in the United States InFire Ants and Leaf Cutting Ants Biology and Management(Lofgren CS and Vander Meer RK Eds) Westview PressBoulder Colorado pp 36ndash49
Luikart G Allendorf FW Cornuet JM and Sherwin WB 1998Distortion of allele frequency distributions provides a test forrecent population bottlenecks J Hered 89 238ndash247
Luikart G and Cornuet JM 1998 Empirical evaluation of a test foridentifying recently bottlenecked populations from allele frequen-cy data Conserv Biol 12 228ndash237
MacKay WP and Fagerlund R 1997 Range expansion of the redimported fire ant Solenopsis invicta Buren (Hymenoptera For-micidae) into New Mexico and extreme western Texas ProcEntomol Soc Wash 99 757ndash758
Mank JE and Avise JC 2004 Individual organisms as units of analysisBayesian-clustering alternatives in population genetics Genet Res84 135ndash143
May GE Gelembiuk GW Panov VE Orlova MI and Lee CE2006 Molecular ecology of zebra mussel invasions Mol Ecol 15 1021ndash1031
McCubbin KI and Weiner JM 2002 Fire ants in Australia A newmedical and ecological hazard Med J Aus 176 518ndash519
Morrison LW 2000 Mechanisms of interspecific competition amongan invasive and two native fire ants Oikos 90 238ndash252
Morrison LW 2002 Long-term impacts of an arthropod-communityinvasion by the imported fire ant Solenopsis invicta Ecology 83 2337ndash2345
Nattrass R and Vanderwoude C 2001 A preliminary investigation ofthe ecological effects of red imported fire ants (Solenopsis invicta)in Brisbane Ecol Manag Resto 2 220ndash223
Nei M 1972 Genetic distance between populations Am Nat 106 283ndash292
Orr MR 1996 Host manipulation by Wolbachia is a neutral trait withinsingle populations Anim Behav 51 1183ndash1185
Patterson RS 1994 Biological control of introduced ant species InExotic Ants Biology Impact and Control of Introduced Species(Williams DF Ed) Westview Press Boulder Colorado pp 293ndash308
Piry S Luikart G and Cornuet JM 1999 BOTTLENECK acomputer program for detecting recent reductions in the effectivepopulation size using allele frequency data J Hered 90 502ndash503
64 CC Yang et al Fire ants in Taiwan
Porter SD and Savignano DA 1990 Invasion of polygyne fire antsdecimates native ants and disrupts arthropod community Ecology7 2095ndash2106
Porter SD Bhatkar A Mulder R Vinson SB and Clair DJ 1991Distribution and density of polygyne fire ants (HymenopteraFormicidae) in Texas J Econ Entomol 84 866ndash874
Porter SD Fowler HG and MacKay WP 1992 Fire ant mounddensities in the United States and Brazil (Hymenoptera Formi-cidae) J Econ Entomol 85 1154ndash1161
Porter SD Williams DF Patterson RS and Fowler HG 1997Intercontinental differences in the abundance of Solenopsis fireants (Hymenoptera Formicidae) Escape from natural enemiesEnviron Entomol 26 373ndash384
Pritchard JK Stephens M and Donnelly P 2000 Inference ofpopulation structure using multilocus genotype data Genetics155 945ndash959
Raymond M and Rousset F 1995a An exact test for populationdifferentiation Evolution 49 1280ndash1283
Raymond M and Rousset F 1995b GENEPOP (version 12)population genetics software for exact tests and ecumenicism JHered 86 248ndash249
Reuter M Pedersen JS and Keller L 2005 Loss of Wolbachiainfection during colonisation in the invasive Argentine ant Line-pithema humile Heredity 94 364ndash369
Ross KG and Fletcher DJC 1985a Genetic origin of male diploidy inthe fire ant Solenopsis invicta (Hymenoptera Formicidae) and itsevolutionary significance Evolution 39 888ndash903
Ross KG and Fletcher DJC 1985b Comparative study of genetic andsocial structure in two forms of the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 17 349ndash356
Ross KG and Fletcher DJC 1986 Diploid male productionndashasignificant colony mortality factor in the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 19 283ndash291
Ross KG and Shoemaker DD 1993 An unusual pattern of gene flowbetween the two social forms of the fire ant Solenopsis invictaEvolution 47 1595ndash1605
Ross KG Vargo EL Keller L and Trager JC 1993 Effect of afounder event on variation in the genetic sex-determining system ofthe fire ant Solenopsis invicta Genetics 135 843ndash854
Ross KG and Keller L 1995 Joint influence of gene flow and selectionon a reproductively important genetic polymorphism in the fire antSolenopsis invicta Am Nat 146 325ndash348
Ross KG Vargo EL and Keller L 1996 Social evolution in a newenvironment The case of introduced fire ants Proc Natl AcadSci U S A 93 3021ndash3025
Ross KG 1997 Multilocus evolution in fire ants Effects of selectiongene flow and recombination Genetics 145 961ndash974
Ross KG Krieger MJB Shoemaker DD Vargo EL and Keller L1997 Hierarchical analysis of genetic structure in native fire antpopulations Results from three classes of molecular markersGenetics 147 643ndash655
Ross KG and Shoemaker DD 1997 Nuclear and mitochondrialgenetic structure in two social forms of the fire ant Solenopsisinvicta Insights into transitions to an alternate social organizationHeredity 78 590ndash602
Ross KG and Keller L 1998 Genetic control of social organization inan ant Proc Natl Acad Sci U S A 95 14232ndash14237
Ross KG Shoemaker DD Krieger MJ DeHeer CJ and Keller L1999 Assessing genetic structure with multiple classes of molecularmarkers A case study involving the introduced fire ant Solenopsisinvicta Mol Biol Evol 16 525ndash543
Ross KG and Keller L 2002 Experimental conversion of colony socialorganization by manipulation of worker genotype composition infire ants (Solenopsis invicta) Behav Ecol Sociobiol 51 287ndash295
Ross KG Krieger MJB and Shoemaker DD 2003 Alternativegenetic foundations and evolutionary routes for a key socialpolymorphism in fire ants Genetics 165 1853ndash1867
Ross KG Krieger MJB Keller L and Shoemaker DD 2007Genetic variation and structure in native populations of the fire antSolenopsis invicta Evolutionary and demographic implicationsBiol J Linnean Soc 92 541ndash560
Rousset F 1997 Genetic differentiation and estimation of gene flowfrom F-statistics under isolation by distance Genetics 145 1219ndash1228
Sax DF Stachowicz JJ and Gaines SD 2005 Species InvasionsInsights into Ecology Evolution and Biogeography SinauerAssociates Sunderland Massachussetts 495 pp
Shoemaker DD and Ross KG 1996 Effects of social organization ongene flow in the fire ant Solenopsis invicta Nature 383 613ndash616
Shoemaker DD Ross KG Keller L Vargo EL and Werren JH2000 Wolbachia infections in native and introduced populations offire ants (Solenopsis spp) Insect Mol Biol 9 661ndash673
Shoemaker DD Ahrens M Sheill L Mescher M Keller L and RossKG 2003 Distribution and prevalence of Wolbachia infections innative populations of the fire ant Solenopsis invicta (HymenopteraFormicidae) Environ Entomol 32 1329ndash1336
Shoemaker DD DeHeer CJ Krieger MJB and Ross KG 2006Population genetics of the invasive fire ant Solenopsis invicta(Hymenoptera Formicidae) in the USA Ann Entomol Soc Am99 1213ndash1233
Simon C Frati F Beckenbach A Crespi B Liu H and Flook P 1994Evolution weighting and phylogenetic utility of mitochondrialgene sequences and a compilation of conserved polymerase chainreaction primers Ann Entomol Soc Am 87 651ndash701
Toom PM Cupp E Johnson CP and Griffin I 1976 Utilization ofbody reserves for minim brood development by queens of theimported fire ant Solenopsis invicta J Insect Physiol 22 217ndash220
Tsutsui ND and Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepethema humile) in its nativeand introduced ranges Evolution 55 976ndash985
Tsutsui ND Kauppinen SN Oyafuso AF and Grosberg RK 2003The distribution and evolutionary history of Wolbachia infection innative and introduced populations of the invasive argentine ant(Linepithema humile) Mol Ecol 12 3057ndash3068
Valles SM and Porter SD 2003 Identification of polygyne andmonogyne fire ant colonies (Solenopsis invicta) by multiplex PCRof Gp-9 alleles Insect Soc 50 199ndash200
Valles SM Oi DH Briano JA and Williams DF 2004 Simultaneousdetection of Vairimorpha invictae (Microsporidia Burenellidae)and Thelohania solenopsae (Microsporidia Thelohaniidae) in fireants by PCR Florida Entomol 87 85ndash87
Vander Meer R K and Porter SD 2001 Fate of newly-mated queensintroduced into monogyne and polygyne Solenopsis invicta (Hy-menoptera Formicidae) colonies Ann Entomol Soc Am 94 289ndash297
Vander Meer RK Jaffe K and Cedeno A 1990 Applied Myrmecol-ogy A World Perspective Westview Press Boulder CO 741 pp
Vinson SB 1994 Impact and invasion of Solenopsis invicta (Buren) onnative food webs In Exotic Ants Biology Impact and Control ofIntroduced Species (Williams DF Ed) Westview Press BoulderColorado pp 293ndash308
Vinson SB 1997 Invasion of the red imported fire ant (HymenopteraFormicidae) Spread biology and impact Am Entomol 43 23ndash29
Williams DF and Porter SD 1994 Fire ant control Science 264 1653ndash1653
Wojcik D Allen CR Brenner RJ Forys EA Jouvenaz D and LutzRS 2001 Red imported fire ants Impact on biodiversity AmEntomol 47 16ndash23
Xia X and Xie Z 2001 DAMBE Data analysis in molecular biologyand evolution J Hered 92 371ndash373
Insect Soc Vol 55 2008 Research article 65

Porter SD and Savignano DA 1990 Invasion of polygyne fire antsdecimates native ants and disrupts arthropod community Ecology7 2095ndash2106
Porter SD Bhatkar A Mulder R Vinson SB and Clair DJ 1991Distribution and density of polygyne fire ants (HymenopteraFormicidae) in Texas J Econ Entomol 84 866ndash874
Porter SD Fowler HG and MacKay WP 1992 Fire ant mounddensities in the United States and Brazil (Hymenoptera Formi-cidae) J Econ Entomol 85 1154ndash1161
Porter SD Williams DF Patterson RS and Fowler HG 1997Intercontinental differences in the abundance of Solenopsis fireants (Hymenoptera Formicidae) Escape from natural enemiesEnviron Entomol 26 373ndash384
Pritchard JK Stephens M and Donnelly P 2000 Inference ofpopulation structure using multilocus genotype data Genetics155 945ndash959
Raymond M and Rousset F 1995a An exact test for populationdifferentiation Evolution 49 1280ndash1283
Raymond M and Rousset F 1995b GENEPOP (version 12)population genetics software for exact tests and ecumenicism JHered 86 248ndash249
Reuter M Pedersen JS and Keller L 2005 Loss of Wolbachiainfection during colonisation in the invasive Argentine ant Line-pithema humile Heredity 94 364ndash369
Ross KG and Fletcher DJC 1985a Genetic origin of male diploidy inthe fire ant Solenopsis invicta (Hymenoptera Formicidae) and itsevolutionary significance Evolution 39 888ndash903
Ross KG and Fletcher DJC 1985b Comparative study of genetic andsocial structure in two forms of the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 17 349ndash356
Ross KG and Fletcher DJC 1986 Diploid male productionndashasignificant colony mortality factor in the fire ant Solenopsis invicta(Hymenoptera Formicidae) Behav Ecol Sociobiol 19 283ndash291
Ross KG and Shoemaker DD 1993 An unusual pattern of gene flowbetween the two social forms of the fire ant Solenopsis invictaEvolution 47 1595ndash1605
Ross KG Vargo EL Keller L and Trager JC 1993 Effect of afounder event on variation in the genetic sex-determining system ofthe fire ant Solenopsis invicta Genetics 135 843ndash854
Ross KG and Keller L 1995 Joint influence of gene flow and selectionon a reproductively important genetic polymorphism in the fire antSolenopsis invicta Am Nat 146 325ndash348
Ross KG Vargo EL and Keller L 1996 Social evolution in a newenvironment The case of introduced fire ants Proc Natl AcadSci U S A 93 3021ndash3025
Ross KG 1997 Multilocus evolution in fire ants Effects of selectiongene flow and recombination Genetics 145 961ndash974
Ross KG Krieger MJB Shoemaker DD Vargo EL and Keller L1997 Hierarchical analysis of genetic structure in native fire antpopulations Results from three classes of molecular markersGenetics 147 643ndash655
Ross KG and Shoemaker DD 1997 Nuclear and mitochondrialgenetic structure in two social forms of the fire ant Solenopsisinvicta Insights into transitions to an alternate social organizationHeredity 78 590ndash602
Ross KG and Keller L 1998 Genetic control of social organization inan ant Proc Natl Acad Sci U S A 95 14232ndash14237
Ross KG Shoemaker DD Krieger MJ DeHeer CJ and Keller L1999 Assessing genetic structure with multiple classes of molecularmarkers A case study involving the introduced fire ant Solenopsisinvicta Mol Biol Evol 16 525ndash543
Ross KG and Keller L 2002 Experimental conversion of colony socialorganization by manipulation of worker genotype composition infire ants (Solenopsis invicta) Behav Ecol Sociobiol 51 287ndash295
Ross KG Krieger MJB and Shoemaker DD 2003 Alternativegenetic foundations and evolutionary routes for a key socialpolymorphism in fire ants Genetics 165 1853ndash1867
Ross KG Krieger MJB Keller L and Shoemaker DD 2007Genetic variation and structure in native populations of the fire antSolenopsis invicta Evolutionary and demographic implicationsBiol J Linnean Soc 92 541ndash560
Rousset F 1997 Genetic differentiation and estimation of gene flowfrom F-statistics under isolation by distance Genetics 145 1219ndash1228
Sax DF Stachowicz JJ and Gaines SD 2005 Species InvasionsInsights into Ecology Evolution and Biogeography SinauerAssociates Sunderland Massachussetts 495 pp
Shoemaker DD and Ross KG 1996 Effects of social organization ongene flow in the fire ant Solenopsis invicta Nature 383 613ndash616
Shoemaker DD Ross KG Keller L Vargo EL and Werren JH2000 Wolbachia infections in native and introduced populations offire ants (Solenopsis spp) Insect Mol Biol 9 661ndash673
Shoemaker DD Ahrens M Sheill L Mescher M Keller L and RossKG 2003 Distribution and prevalence of Wolbachia infections innative populations of the fire ant Solenopsis invicta (HymenopteraFormicidae) Environ Entomol 32 1329ndash1336
Shoemaker DD DeHeer CJ Krieger MJB and Ross KG 2006Population genetics of the invasive fire ant Solenopsis invicta(Hymenoptera Formicidae) in the USA Ann Entomol Soc Am99 1213ndash1233
Simon C Frati F Beckenbach A Crespi B Liu H and Flook P 1994Evolution weighting and phylogenetic utility of mitochondrialgene sequences and a compilation of conserved polymerase chainreaction primers Ann Entomol Soc Am 87 651ndash701
Toom PM Cupp E Johnson CP and Griffin I 1976 Utilization ofbody reserves for minim brood development by queens of theimported fire ant Solenopsis invicta J Insect Physiol 22 217ndash220
Tsutsui ND and Case TJ 2001 Population genetics and colonystructure of the Argentine ant (Linepethema humile) in its nativeand introduced ranges Evolution 55 976ndash985
Tsutsui ND Kauppinen SN Oyafuso AF and Grosberg RK 2003The distribution and evolutionary history of Wolbachia infection innative and introduced populations of the invasive argentine ant(Linepithema humile) Mol Ecol 12 3057ndash3068
Valles SM and Porter SD 2003 Identification of polygyne andmonogyne fire ant colonies (Solenopsis invicta) by multiplex PCRof Gp-9 alleles Insect Soc 50 199ndash200
Valles SM Oi DH Briano JA and Williams DF 2004 Simultaneousdetection of Vairimorpha invictae (Microsporidia Burenellidae)and Thelohania solenopsae (Microsporidia Thelohaniidae) in fireants by PCR Florida Entomol 87 85ndash87
Vander Meer R K and Porter SD 2001 Fate of newly-mated queensintroduced into monogyne and polygyne Solenopsis invicta (Hy-menoptera Formicidae) colonies Ann Entomol Soc Am 94 289ndash297
Vander Meer RK Jaffe K and Cedeno A 1990 Applied Myrmecol-ogy A World Perspective Westview Press Boulder CO 741 pp
Vinson SB 1994 Impact and invasion of Solenopsis invicta (Buren) onnative food webs In Exotic Ants Biology Impact and Control ofIntroduced Species (Williams DF Ed) Westview Press BoulderColorado pp 293ndash308
Vinson SB 1997 Invasion of the red imported fire ant (HymenopteraFormicidae) Spread biology and impact Am Entomol 43 23ndash29
Williams DF and Porter SD 1994 Fire ant control Science 264 1653ndash1653
Wojcik D Allen CR Brenner RJ Forys EA Jouvenaz D and LutzRS 2001 Red imported fire ants Impact on biodiversity AmEntomol 47 16ndash23
Xia X and Xie Z 2001 DAMBE Data analysis in molecular biologyand evolution J Hered 92 371ndash373
Insect Soc Vol 55 2008 Research article 65


















