
Plasmodium falciparum: Biochemical characterization of the cysteine protease falcipain-2′
6
Experimental Parasitology 112 (2006) 187–192 www.elsevier.com/locate/yexpr 0014-4894/$ - see front matter 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.exppara.2005.10.007 Research brief Plasmodium falciparum: Biochemical characterization of the cysteine protease falcipain-2 Naresh Singh, Puran S. Sijwali, Kailash C. Pandey, Philip J. Rosenthal ¤ Department of Medicine, San Francisco General Hospital, University of California, San Francisco, CA 94143-0811, USA Received 27 June 2005; received in revised form 24 October 2005; accepted 27 October 2005 Available online 7 December 2005 Abstract The Plasmodium falciparum cysteine proteases falcipain-2 and falcipain-3 are hemoglobinases and potential antimalarial drug targets. The falcipain-2 gene was identiWed recently and is nearly identical in sequence to falcipain-2. The product of this gene has not been stud- ied previously. The mature protease domain of falcipain-2 was expressed in Escherichia coli, puriWed, and refolded to active enzyme. Functional analysis revealed similar biochemical properties to those of falcipain-2, including pH optima (pH 5.5–7.0), reducing require- ments, and substrate preference. Studies with cysteine protease inhibitors showed similar inhibition of falcipain-2 and falcipain-2, although speciWcities were not identical. Considering activity against the presumed biological substrate, both enzymes readily hydrolyzed hemoglobin. Our results conWrm that falcipain-2 is an active hemoglobinase and suggest that falcipain-2 and falcipain-2 play similar roles in erythrocytic parasites but that, for promising cysteine protease inhibitors, it will be important to conWrm activity against this additional target. 2005 Elsevier Inc. All rights reserved. Index Descriptors and Abbreviations: Plasmodium falciparum; Malaria; Cysteine protease; Proteinase; Hemoglobin; Falcipain; DTT, dithiothreitol; GSH, reduced glutathione; AMC, 7-amino-4-methyl coumarin; E-64, trans-epoxysuccinyl-L-leucylamido-(4-guanidino)butane; Mu-Leu-Hph-VSPh, morpholine urea-leucine-homophenylalanine-phenyl vinyl sulfone; Z-Leu-Hph-CH 2 F, benzyloxycarbonyl-leucine-homophenylalanine-Xuoromethyl ketone; DMSO, dimethyl sulfoxide The control of malaria, in particular that caused by Plasmodium falciparum, is undermined by resistance to most available drugs (White, 2004). New antimalarial drugs, ideally directed against new targets, are urgently needed (Rosenthal, 2003). Among promising targets are P. falciparum cysteine proteases (Rosenthal, 2004). In par- ticular, falcipain-2 and falcipain-3 appear to play key roles in hemoglobin hydrolysis by erythrocytic parasites and to be the targets of eVective antimalarial cysteine protease inhibitors (Shenai et al., 2000; Sijwali et al., 2004). This report describes falcipain-2, an enzyme that is very similar in sequence to falcipain-2, but whose existence was not appreciated until elaboration of the P. falciparum genome sequence. Characterization of falcipain-2 is necessary to fully appreciate the action of antimalarial cysteine protease inhibitors. Malaria parasites digest host hemoglobin in an acidic food vacuole to obtain free amino acids and maintain osmotic stability (Francis et al., 1997; Lew et al., 2003). Cys- teine protease inhibitors block hemoglobin hydrolysis (Rosenthal et al., 1988, 1991), indicating that cysteine prote- ases are required for this process. The Wrst characterized P. falciparum cysteine protease, falcipain-1, was hypothesized to mediate hemoglobin hydrolysis (Rosenthal and Nelson, 1992), but recent studies showed that disruption of the falci- pain-1 gene has no obvious eVect on erythrocytic parasites (Eksi et al., 2004; Sijwali et al., 2004). Therefore, although the function of falcipain-1 remains uncertain, it does not appear to be essential for hemoglobin hydrolysis by erythro- The nucleotide sequence reported in this paper has been submitted to the GenBank/NCBI Data Bank with Accession No. AY966007 (FP2). * Corresponding author. Fax: +1 415 648 8425. E-mail address: [email protected] (P.J. Rosenthal).
-
Upload
naresh-singh -
Category
Documents
-
view
212 -
download
0
Transcript of Plasmodium falciparum: Biochemical characterization of the cysteine protease falcipain-2′

Experimental Parasitology 112 (2006) 187–192
www.elsevier.com/locate/yexpr
Research brief
Plasmodium falciparum: Biochemical characterization of the cysteine protease falcipain-2� �
Naresh Singh, Puran S. Sijwali, Kailash C. Pandey, Philip J. Rosenthal ¤
Department of Medicine, San Francisco General Hospital, University of California, San Francisco, CA 94143-0811, USA
Received 27 June 2005; received in revised form 24 October 2005; accepted 27 October 2005Available online 7 December 2005
Abstract
The Plasmodium falciparum cysteine proteases falcipain-2 and falcipain-3 are hemoglobinases and potential antimalarial drug targets.The falcipain-2� gene was identiWed recently and is nearly identical in sequence to falcipain-2. The product of this gene has not been stud-ied previously. The mature protease domain of falcipain-2� was expressed in Escherichia coli, puriWed, and refolded to active enzyme.Functional analysis revealed similar biochemical properties to those of falcipain-2, including pH optima (pH 5.5–7.0), reducing require-ments, and substrate preference. Studies with cysteine protease inhibitors showed similar inhibition of falcipain-2 and falcipain-2�,although speciWcities were not identical. Considering activity against the presumed biological substrate, both enzymes readily hydrolyzedhemoglobin. Our results conWrm that falcipain-2� is an active hemoglobinase and suggest that falcipain-2 and falcipain-2� play similarroles in erythrocytic parasites but that, for promising cysteine protease inhibitors, it will be important to conWrm activity against thisadditional target. 2005 Elsevier Inc. All rights reserved.
Index Descriptors and Abbreviations: Plasmodium falciparum; Malaria; Cysteine protease; Proteinase; Hemoglobin; Falcipain; DTT, dithiothreitol; GSH,reduced glutathione; AMC, 7-amino-4-methyl coumarin; E-64, trans-epoxysuccinyl-L-leucylamido-(4-guanidino)butane; Mu-Leu-Hph-VSPh, morpholineurea-leucine-homophenylalanine-phenyl vinyl sulfone; Z-Leu-Hph-CH2F, benzyloxycarbonyl-leucine-homophenylalanine-Xuoromethyl ketone; DMSO,dimethyl sulfoxide
The control of malaria, in particular that caused byPlasmodium falciparum, is undermined by resistance tomost available drugs (White, 2004). New antimalarialdrugs, ideally directed against new targets, are urgentlyneeded (Rosenthal, 2003). Among promising targets areP. falciparum cysteine proteases (Rosenthal, 2004). In par-ticular, falcipain-2 and falcipain-3 appear to play key rolesin hemoglobin hydrolysis by erythrocytic parasites and tobe the targets of eVective antimalarial cysteine proteaseinhibitors (Shenai et al., 2000; Sijwali et al., 2004). Thisreport describes falcipain-2�, an enzyme that is very similarin sequence to falcipain-2, but whose existence was not
� The nucleotide sequence reported in this paper has been submitted tothe GenBank/NCBI Data Bank with Accession No. AY966007 (FP2�).
* Corresponding author. Fax: +1 415 648 8425.E-mail address: [email protected] (P.J. Rosenthal).
0014-4894/$ - see front matter 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.exppara.2005.10.007
appreciated until elaboration of the P. falciparum genomesequence. Characterization of falcipain-2� is necessary tofully appreciate the action of antimalarial cysteine proteaseinhibitors.
Malaria parasites digest host hemoglobin in an acidicfood vacuole to obtain free amino acids and maintainosmotic stability (Francis et al., 1997; Lew et al., 2003). Cys-teine protease inhibitors block hemoglobin hydrolysis(Rosenthal et al., 1988, 1991), indicating that cysteine prote-ases are required for this process. The Wrst characterized P.falciparum cysteine protease, falcipain-1, was hypothesizedto mediate hemoglobin hydrolysis (Rosenthal and Nelson,1992), but recent studies showed that disruption of the falci-pain-1 gene has no obvious eVect on erythrocytic parasites(Eksi et al., 2004; Sijwali et al., 2004). Therefore, althoughthe function of falcipain-1 remains uncertain, it does notappear to be essential for hemoglobin hydrolysis by erythro-

188 N. Singh et al. / Experimental Parasitology 112 (2006) 187–192
cytic parasites. Falcipain-2 and falcipain-3 were character-ized more recently (Shenai et al., 2000; Sijwali et al.,2001a,b). They are fairly typical papain-family cysteine pro-teases, although they have some unusual features for thisfamily. In particular, they contain unusual amino-terminalextensions that mediate folding of the mature proteasesindependent of the prodomain (Pandey et al., 2004; Sijwaliet al., 2002) and an insertion near the carboxy terminus thatmediates interaction with hemoglobin (Pandey et al., 2005).Falcipain-2 and falcipain-3 are expressed sequentially byerythrocytic parasites (Dahl and Rosenthal, 2005), and itappears that they both play roles in hemoglobin hydrolysis.Strongly supporting this hypothesis, disruption of the falci-pain-2 gene caused trophozoites to accumulate undegradedhemoglobin in the parasite food vacuole (Sijwali andRosenthal, 2004). This disruption was not lethal, probablybecause of increased expression of falcipain-3 and falcipain-2� (Sijwali and Rosenthal, 2004). Disruption of the falcipain-3 gene using similar methods has not been successful(Sijwali, unpublished observation), suggesting that this dis-ruption is lethal for erythrocytic parasites.
Supporting the consideration of falcipains as therapeutictargets, cysteine protease inhibitors that block the activityof these enzymes prevent the development of cultured para-sites (Rosenthal et al., 1991; Shenai et al., 2003) and curemalaria in infected mice (Olson et al., 1999; Rosenthal et al.,1993). Optimization of cysteine protease inhibitors as anti-malarials is facilitated by an appreciation of the speciWcactions of these compounds. Recombinant falcipain-2 andfalcipain-3 are available, and analyses of inhibition of theseenzymes is routinely performed as part of drug discoveryeVorts (Shenai et al., 2003). However, biochemicalcharacterization of the most recently identiWed falcipain,falcipain-2�, has not previously been reported.
The genes encoding falcipain-2, falcipain-3, and falci-pain-2� are nearly contiguous, and located within a »12 kbstretch of chromosome 11. The falcipain-1 gene is encodedon a diVerent chromosome (chromosome 14). All falcipainsare expressed by erythrocytic parasites (Sijwali et al., 2004).Falcipain-2 and falcipain-3 are quite similar in sequence(68% identical in the catalytic domains), but falcipain-2 andfalcipain-2� are much more similar, diVering by only 3amino acids (FP2� ! FP2, I150V, P157A, E171Q) in theircatalytic domains (the catalytic domain begins at residue 18of the mature protease) (Fig. 1A). It was predicted that fal-cipain-2� is very similar biochemically to falcipain-2 andthat the enzymes share the function of hemoglobin hydro-lysis. Nonetheless, it was important to test this hypothesis,as diVerences between falcipain-2 and falcipain-2� mighthave important implications for chemotherapy with cys-teine protease inhibitors. We therefore set out to expressand biochemically characterize falcipain-2�.
The sequences of the falcipain-2 and falcipain-2� genesdiVer »10% within portions encoding the prodomains,allowing selective ampliWcation of their genes. To expressfalcipain-2�, a gene segment (1691bp) was ampliWed fromP. falciparum 3D7 genomic DNA with forward primer
FP2.1UPS (5�-CTATAAATTATATATTTCAATGAATTGATTATATAAG-3�) and reverse primer FP2.1RH (5�-TTAaagcttATTTATGATAGTCCCCCTTTTAAAATACTACTC-3�, HindIII site is shown in small letters) using ventDNA polymerase (New England Bio Labs). The ampliWedgene was cloned into the pGEM-T cloning vector (Promega)and transformed into Escherichia coli DH5� (Invitrogen).DNA puriWed from these bacteria was cleaved with BglIIand HindIII to yield a transcript encoding mature falcipain-2� (813bp, including 42 nucleotides upstream of the predictedmature enzyme cleavage site). This fragment was subclonedinto the pRSET-B expression plasmid (Invitrogen) and usedto transform E. coli BL(21)DE3pLysS (Novagen) for proteinexpression. Falcipain-2� was expressed after induction withisopropyl-1-thio-�-D-galactopyranoside using techniques asdescribed previously for falcipain-2 (Sijwali et al., 2001a,b).The enzyme was then puriWed under denaturing conditionsusing nickel–nitrilotriacetic acid chromatography, refoldedto active protein at alkaline pH, and further puriWed by Q-Sepharose anion exchange chromatography (AmershamBiosciences), all as previously described for falcipain-2(Sijwali et al., 2001a,b). For comparative studies, falcipain-2was expressed and puriWed as previously described (Sijwaliet al., 2001a,b). PuriWed enzymes were stored at ¡20 °C inequal volumes of glycerol. The nucleotide sequence of thefalcipain-2� transformation clone and the N-terminalsequence of recombinant falcipain-2� were conWrmed bystandard methods.
Recombinant falcipain-2� cross-reacted with antiserumgenerated against falcipain-2 (Sijwali et al., 2001a,b), anexpected result considering their similarity in sequence(Fig. 1B). Falcipain-2� was biochemically active, as docu-mented by gelatin substrate PAGE (Fig. 1B). Both enzymesrequired reductants for appreciable activity; activity wasmarkedly stimulated by both dithiothreitol (DTT) and thephysiological reductant glutathione (GSH) (Shenai andRosenthal, 2002). Both enzymes also underwent eYcientautohydrolysis during refolding at alkaline pH, a featurethat distinguishes them from recombinant falcipain-3,which undergoes minimal processing until transfer to acidicpH (Sijwali et al., 2001a,b). Additional biochemical proper-ties of falcipain-2 and falcipain-2� were assessed in parallel.When studied against the peptide Xuorogenic substratebenzyloxycarbonyl-Leu-Arg-7-amino-4-methyl coumarin(Z-Leu-Arg-AMC), the two enzymes had identical pHoptima of 6.0–6.5, consistent with action in the acidic foodvacuole (Fig. 1C). Falcipain-2� was somewhat less stableupon incubation at 37 °C, but both enzymes retained mostof their activity after a 2 h incubation (Fig. 1D).
Kinetic properties of the recombinant enzymes againstWve diVerent peptide substrates were determined as previ-ously described (Shenai et al., 2000). Relative and absoluteactivities against the tested substrates were similar. Nota-bly, as described previously for falcipain-2 (Shenai et al.,2000) and falcipain-3 (Sijwali et al., 2001a,b), falcipain-2�also preferred substrates with leucine at the P2 position.However, some diVerences were seen. Notably, falcipain-2�
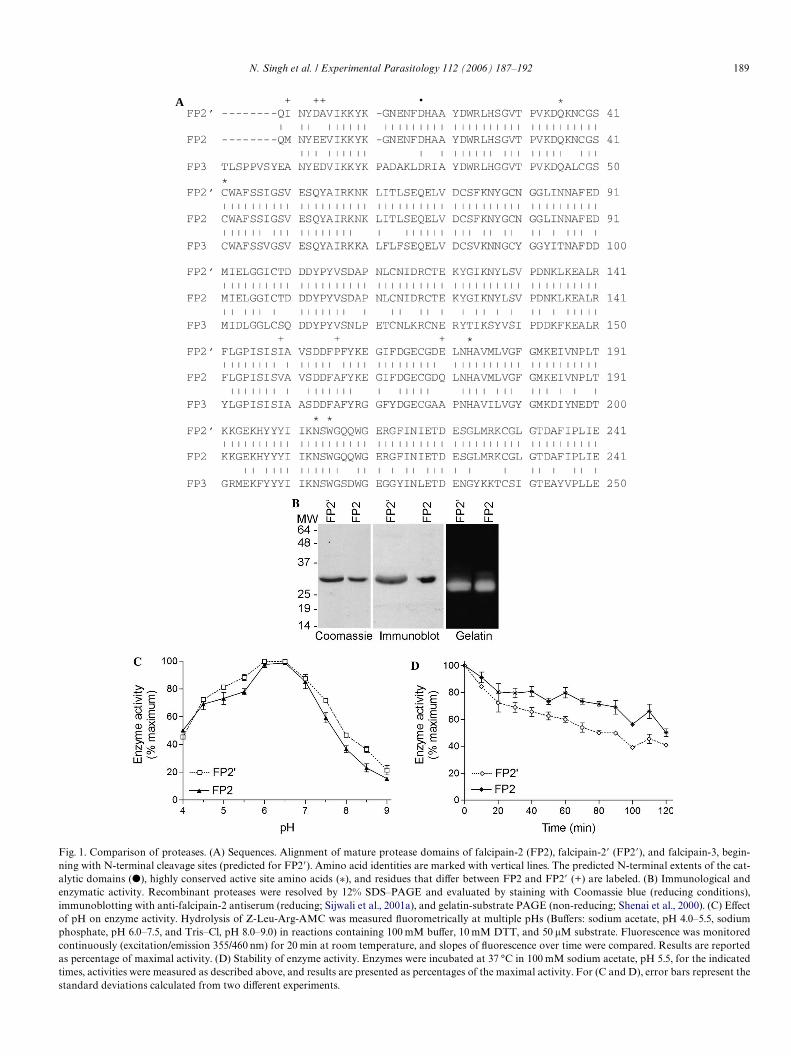
N. Singh et al. / Experimental Parasitology 112 (2006) 187–192 189
Fig. 1. Comparison of proteases. (A) Sequences. Alignment of mature protease domains of falcipain-2 (FP2), falcipain-2� (FP2�), and falcipain-3, begin-ning with N-terminal cleavage sites (predicted for FP2�). Amino acid identities are marked with vertical lines. The predicted N-terminal extents of the cat-alytic domains (�), highly conserved active site amino acids (¤), and residues that diVer between FP2 and FP2� (+) are labeled. (B) Immunological andenzymatic activity. Recombinant proteases were resolved by 12% SDS–PAGE and evaluated by staining with Coomassie blue (reducing conditions),immunoblotting with anti-falcipain-2 antiserum (reducing; Sijwali et al., 2001a), and gelatin-substrate PAGE (non-reducing; Shenai et al., 2000). (C) EVectof pH on enzyme activity. Hydrolysis of Z-Leu-Arg-AMC was measured Xuorometrically at multiple pHs (BuVers: sodium acetate, pH 4.0–5.5, sodiumphosphate, pH 6.0–7.5, and Tris–Cl, pH 8.0–9.0) in reactions containing 100 mM buVer, 10 mM DTT, and 50 �M substrate. Fluorescence was monitoredcontinuously (excitation/emission 355/460 nm) for 20 min at room temperature, and slopes of Xuorescence over time were compared. Results are reportedas percentage of maximal activity. (D) Stability of enzyme activity. Enzymes were incubated at 37 °C in 100 mM sodium acetate, pH 5.5, for the indicatedtimes, activities were measured as described above, and results are presented as percentages of the maximal activity. For (C and D), error bars represent thestandard deviations calculated from two diVerent experiments.
A
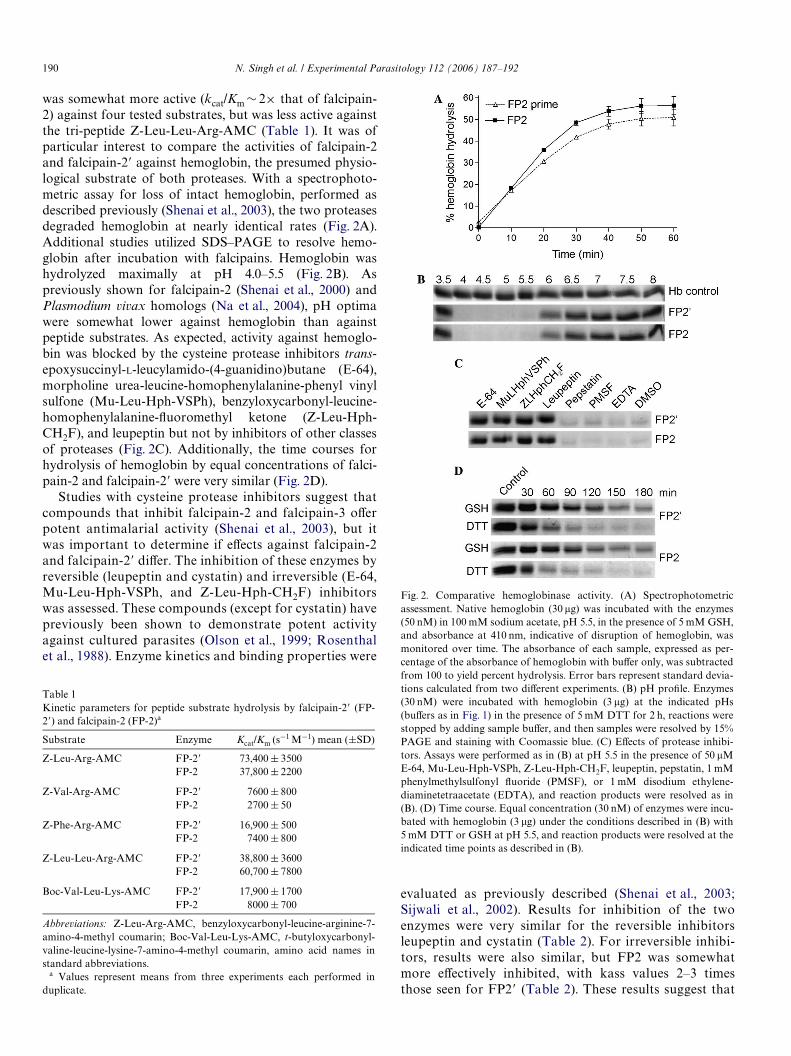
190 N. Singh et al. / Experimental Parasitology 112 (2006) 187–192
was somewhat more active (kcat/Km » 2£ that of falcipain-2) against four tested substrates, but was less active againstthe tri-peptide Z-Leu-Leu-Arg-AMC (Table 1). It was ofparticular interest to compare the activities of falcipain-2and falcipain-2� against hemoglobin, the presumed physio-logical substrate of both proteases. With a spectrophoto-metric assay for loss of intact hemoglobin, performed asdescribed previously (Shenai et al., 2003), the two proteasesdegraded hemoglobin at nearly identical rates (Fig. 2A).Additional studies utilized SDS–PAGE to resolve hemo-globin after incubation with falcipains. Hemoglobin washydrolyzed maximally at pH 4.0–5.5 (Fig. 2B). Aspreviously shown for falcipain-2 (Shenai et al., 2000) andPlasmodium vivax homologs (Na et al., 2004), pH optimawere somewhat lower against hemoglobin than againstpeptide substrates. As expected, activity against hemoglo-bin was blocked by the cysteine protease inhibitors trans-epoxysuccinyl-L-leucylamido-(4-guanidino)butane (E-64),morpholine urea-leucine-homophenylalanine-phenyl vinylsulfone (Mu-Leu-Hph-VSPh), benzyloxycarbonyl-leucine-homophenylalanine-Xuoromethyl ketone (Z-Leu-Hph-CH2F), and leupeptin but not by inhibitors of other classesof proteases (Fig. 2C). Additionally, the time courses forhydrolysis of hemoglobin by equal concentrations of falci-pain-2 and falcipain-2� were very similar (Fig. 2D).
Studies with cysteine protease inhibitors suggest thatcompounds that inhibit falcipain-2 and falcipain-3 oVerpotent antimalarial activity (Shenai et al., 2003), but itwas important to determine if eVects against falcipain-2and falcipain-2� diVer. The inhibition of these enzymes byreversible (leupeptin and cystatin) and irreversible (E-64,Mu-Leu-Hph-VSPh, and Z-Leu-Hph-CH2F) inhibitorswas assessed. These compounds (except for cystatin) havepreviously been shown to demonstrate potent activityagainst cultured parasites (Olson et al., 1999; Rosenthalet al., 1988). Enzyme kinetics and binding properties were
Table 1Kinetic parameters for peptide substrate hydrolysis by falcipain-2� (FP-2�) and falcipain-2 (FP-2)a
Abbreviations: Z-Leu-Arg-AMC, benzyloxycarbonyl-leucine-arginine-7-amino-4-methyl coumarin; Boc-Val-Leu-Lys-AMC, t-butyloxycarbonyl-valine-leucine-lysine-7-amino-4-methyl coumarin, amino acid names instandard abbreviations.
a Values represent means from three experiments each performed induplicate.
Substrate Enzyme Kcat/Km (s¡1 M¡1) mean (§SD)
Z-Leu-Arg-AMC FP-2� 73,400 § 3500FP-2 37,800 § 2200
Z-Val-Arg-AMC FP-2� 7600 § 800FP-2 2700 § 50
Z-Phe-Arg-AMC FP-2� 16,900 § 500FP-2 7400 § 800
Z-Leu-Leu-Arg-AMC FP-2� 38,800 § 3600FP-2 60,700 § 7800
Boc-Val-Leu-Lys-AMC FP-2� 17,900 § 1700FP-2 8000 § 700
evaluated as previously described (Shenai et al., 2003;Sijwali et al., 2002). Results for inhibition of the twoenzymes were very similar for the reversible inhibitorsleupeptin and cystatin (Table 2). For irreversible inhibi-tors, results were also similar, but FP2 was somewhatmore eVectively inhibited, with kass values 2–3 timesthose seen for FP2� (Table 2). These results suggest that
Fig. 2. Comparative hemoglobinase activity. (A) Spectrophotometricassessment. Native hemoglobin (30 �g) was incubated with the enzymes(50 nM) in 100 mM sodium acetate, pH 5.5, in the presence of 5 mM GSH,and absorbance at 410 nm, indicative of disruption of hemoglobin, wasmonitored over time. The absorbance of each sample, expressed as per-centage of the absorbance of hemoglobin with buVer only, was subtractedfrom 100 to yield percent hydrolysis. Error bars represent standard devia-tions calculated from two diVerent experiments. (B) pH proWle. Enzymes(30 nM) were incubated with hemoglobin (3 �g) at the indicated pHs(buVers as in Fig. 1) in the presence of 5 mM DTT for 2 h, reactions werestopped by adding sample buVer, and then samples were resolved by 15%PAGE and staining with Coomassie blue. (C) EVects of protease inhibi-tors. Assays were performed as in (B) at pH 5.5 in the presence of 50 �ME-64, Mu-Leu-Hph-VSPh, Z-Leu-Hph-CH2F, leupeptin, pepstatin, 1 mMphenylmethylsulfonyl Xuoride (PMSF), or 1 mM disodium ethylene-diaminetetraacetate (EDTA), and reaction products were resolved as in(B). (D) Time course. Equal concentration (30 nM) of enzymes were incu-bated with hemoglobin (3 �g) under the conditions described in (B) with5 mM DTT or GSH at pH 5.5, and reaction products were resolved at theindicated time points as described in (B).

N. Singh et al. / Experimental Parasitology 112 (2006) 187–192 191
inhibitors that act against falcipain-2 will show similaractivity against falcipain-2� but that for lead compoundsit will be prudent to assess activities against bothenzymes.
We have characterized for the Wrst time recombinantfalcipain-2�, an enzyme that is expressed by erythrocyticparasites and is very similar in sequence to falcipain-2. Inall assessments, including hydrolysis of the presumedphysiological substrate hemoglobin and inhibition bypotent inhibitors, the activities of falcipain-2 and falci-pain-2� were similar. Thus, it is anticipated that speciWcinhibitors that target falcipain-2 will also inhibit falci-pain-2�. Indeed, a broader approach is often successful, asmany inhibitors have blocked the activities of both falci-pain-2 and falcipain-3 and exhibited potent antimalarialactivity (Shenai et al., 2003), suggesting that eVectiveinhibitors that target all three related plasmodial hemo-globinases can be generated. Considering these results, itis appropriate to consider whether the functions of falci-pain-2 and falcipain-2� are fully redundant. In fact, theyare not, as the enzymes are expressed somewhat diVer-ently across the erythrocytic cycle, and disruption of thefalcipain-2 gene was not fully complemented by over-expression of falcipain-2� (Sijwali and Rosenthal, 2004).DiVerences in expression or other biological featuresmight be mediated by sequence diVerences between theprodomains and amino-terminal extensions of falcipain-2and falcipain-2�, which diVer more markedly than do thecatalytic domains, and/or by sequence diVerences in non-coding regulatory regions. These diVerences might lead todiVerences in biological functions, including actionagainst substrates in addition to hemoglobin (e.g., eryth-rocyte cytoskeletal proteins to facilitate rupture bymature parasites (Dua et al., 2001)). In any event, our bio-chemical data oVer promise that potent inhibitors of falci-pain-2 will act similarly against the related proteasefalcipain-2�.
Acknowledgments
We thank Jiri Gut and Anthony Lau for their technicalsupport. This work was supported by grants from theNational Institutes of Health (AI35800 and AI35707) andthe Medicines for Malaria Venture. P.J.R. is a Doris DukeCharitable Foundation Distinguished Clinical Scientist.
Table 2Inhibition of falcipain-2� and falcipain-2a
a Values (§SD) represent means of two experiments, each performed induplicate.
Inhibitor FP2� FP2
kass (M¡1 s¡1) E-64 5100 § 1000 11,600 § 1200Mu-Leu-Hph-VSPh 36,800 § 4500 95,100 § 13,400Z-Leu-Hph-CH2F 17,700 § 2300 30,000 § 2000
Ki (nM) Leupeptin 5.3 § 0.6 4.7 § 0.5Cystatin 1.4 § 0.1 2.4 § 0.1
References
Dahl, E.L., Rosenthal, P.J., 2005. Biosynthesis, localization, and processingof falcipain cysteine proteases of Plasmodium falciparum. Mol. Bio-chem. Parasitol. 139, 205–212.
Dua, M., Raphael, P., Sijwali, P.S., Rosenthal, P.J., Hanspal, M., 2001.Recombinant falcipain-2 cleaves erythrocyte membrane ankyrin andprotein 4.1. Mol. Biochem. Parasitol. 116, 95–99.
Eksi, S., Czesny, B., Greenbaum, D.C., Bogyo, M., Williamson, K.C., 2004.Targeted disruption of Plasmodium falciparum cysteine protease, falci-pain 1, reduces oocyst production, not erythrocytic stage growth. Mol.Microbiol. 53, 243–250.
Francis, S.E., Sullivan Jr., D.J., Goldberg, D.E., 1997. Hemoglobin metabo-lism in the malaria parasite Plasmodium falciparum. Annu. Rev. Micro-biol. 51, 97–123.
Lew, V.L., TiVert, T., Ginsburg, H., 2003. Excess hemoglobin digestion andthe osmotic stability of Plasmodium falciparum-infected red blood cells.Blood 101, 4189–4194.
Na, B.K., Shenai, B.R., Sijwali, P.S., Choe, Y., Pandey, K.C., Singh, A.,Craik, C.S., Rosenthal, P.J., 2004. IdentiWcation and biochemical char-acterization of vivapains, cysteine proteases of the malaria parasitePlasmodium vivax. Biochem. J. 378, 529–538.
Olson, J.E., Lee, G.K., Semenov, A., Rosenthal, P.J., 1999. AntimalarialeVects in mice of orally administered peptidyl cysteine protease inhibi-tors. Bioorg. Med. Chem. 7, 633–638.
Pandey, K.C., Sijwali, P.S., Singh, A., Na, B.K., Rosenthal, P.J., 2004. Inde-pendent intramolecular mediators of folding, activity, and inhibitionfor the Plasmodium falciparum cysteine protease falcipain-2. J. Biol.Chem. 279, 3484–3491.
Pandey, K.C., Wang, S.X., Sijwali, P.S., Lau, A.L., McKerrow, J.H., Rosen-thal, P.J., 2005. The Plasmodium falciparum cysteine protease falcipain-2 captures its substrate, hemoglobin, via a unique motif. Proc. Natl.Acad. Sci. USA 102, 9138–9143.
Rosenthal, P.J., 2003. Antimalarial drug discovery: old and newapproaches. J. Exp. Biol. 206, 3735–3744.
Rosenthal, P.J., 2004. Cysteine proteases of malaria parasites. Int. J. Paras-itol. 34, 1489–1499.
Rosenthal, P.J., Lee, G.K., Smith, R.E., 1993. Inhibition of a Plasmodium vinc-kei cysteine proteinase cures murine malaria. J. Clin. Invest. 91, 1052–1056.
Rosenthal, P.J., McKerrow, J.H., Aikawa, M., Nagasawa, H., Leech, J.H.,1988. A malarial cysteine proteinase is necessary for hemoglobin deg-radation by Plasmodium falciparum. J. Clin. Invest. 82, 1560–1566.
Rosenthal, P.J., Nelson, R.G., 1992. Isolation and characterization of acysteine proteinase gene of Plasmodium falciparum. Mol. Biochem.Parasitol. 51, 143–152.
Rosenthal, P.J., Wollish, W.S., Palmer, J.T., Rasnick, D., 1991. Antimalar-ial eVects of peptide inhibitors of a Plasmodium falciparum cysteineproteinase. J. Clin. Invest. 88, 1467–1472.
Shenai, B.R., Lee, B.J., Alvarez-Hernandez, A., Chong, P.Y., Emal, C.D.,Neitz, R.J., Roush, W.R., Rosenthal, P.J., 2003. Structure–activity rela-tionships for inhibition of cysteine protease activity and developmentof Plasmodium falciparum by peptidyl vinyl sulfones. Antimicrob.Agents Chemother. 47, 154–160.
Shenai, B.R., Rosenthal, P.J., 2002. Reducing requirements for hemoglobinhydrolysis by Plasmodium falciparum cysteine proteases. Mol. Bio-chem. Parasitol. 122, 99–104.
Shenai, B.R., Sijwali, P.S., Singh, A., Rosenthal, P.J., 2000. Characteriza-tion of native and recombinant falcipain-2, a principal trophozoite cys-teine protease and essential hemoglobinase of Plasmodium falciparum.J. Biol. Chem. 275, 29000–29010.
Sijwali, P.S., Brinen, L.S., Rosenthal, P.J., 2001a. Systematic optimizationof expression and refolding of the Plasmodium falciparum cysteine pro-tease falcipain-2. Protein Expr. Purif. 22, 128–134.
Sijwali, P.S., Shenai, B.R., Gut, J., Singh, A., Rosenthal, P.J., 2001b. Expres-sion and characterization of the Plasmodium falciparum haemoglobin-ase falcipain-3. Biochem. J. 360, 481–489.
Sijwali, P.S., Kato, K., Seydel, K.B., Gut, J., Lehman, J., Klemba, M., Gold-berg, D.E., Miller, L.H., Rosenthal, P.J., 2004. Plasmodium falciparum

192 N. Singh et al. / Experimental Parasitology 112 (2006) 187–192
cysteine protease falcipain-1 is not essential in erythrocytic stagemalaria parasites. Proc. Natl. Acad. Sci. USA 101, 8721–8726.
Sijwali, P.S., Rosenthal, P.J., 2004. Gene disruption conWrms a critical rolefor the cysteine protease falcipain-2 in hemoglobin hydrolysis by Plas-modium falciparum. Proc. Natl. Acad. Sci. USA 101, 4384–4389.
Sijwali, P.S., Shenai, B.R., Rosenthal, P.J., 2002. Folding of the Plasmo-dium falciparum cysteine protease falcipain-2 is mediated by achaperone-like peptide and not the prodomain. J. Biol. Chem. 277,14910–14915.
White, N.J., 2004. Antimalarial drug resistance. J. Clin. Invest. 113, 1084–1092.
![Op Access M acteriza Plasmodium falciparum tifolat ... · Plasmodium falciparum and Plasmodium vivax has been reported as early as t[2]ulfado–pyrimeth - ()olate pathway inhibitor](https://static.fdocuments.in/doc/165x107/5fa82a31367407357973068f/op-access-m-acteriza-plasmodium-falciparum-tifolat-plasmodium-falciparum-and.jpg)

















