
Plasmacytoid Dendritic Cells Enhance Mortality during ... · Plasmacytoid Dendritic Cells Enhance...
8
of July 16, 2018. This information is current as by Eliminating Virus-Specific CD8 T Cells Mortality during Lethal Influenza Infections Plasmacytoid Dendritic Cells Enhance Ryan A. Langlois and Kevin L. Legge http://www.jimmunol.org/content/184/8/4440 doi: 10.4049/jimmunol.0902984 March 2010; 2010; 184:4440-4446; Prepublished online 10 J Immunol Material Supplementary 4.DC1 http://www.jimmunol.org/content/suppl/2010/03/11/jimmunol.090298 References http://www.jimmunol.org/content/184/8/4440.full#ref-list-1 , 17 of which you can access for free at: cites 38 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2010 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on July 16, 2018 http://www.jimmunol.org/ Downloaded from by guest on July 16, 2018 http://www.jimmunol.org/ Downloaded from
Transcript of Plasmacytoid Dendritic Cells Enhance Mortality during ... · Plasmacytoid Dendritic Cells Enhance...

of July 16, 2018.This information is current as
by Eliminating Virus-Specific CD8 T CellsMortality during Lethal Influenza Infections Plasmacytoid Dendritic Cells Enhance
Ryan A. Langlois and Kevin L. Legge
http://www.jimmunol.org/content/184/8/4440doi: 10.4049/jimmunol.0902984March 2010;
2010; 184:4440-4446; Prepublished online 10J Immunol
MaterialSupplementary
4.DC1http://www.jimmunol.org/content/suppl/2010/03/11/jimmunol.090298
Referenceshttp://www.jimmunol.org/content/184/8/4440.full#ref-list-1
, 17 of which you can access for free at: cites 38 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2010 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 16, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Plasmacytoid Dendritic Cells Enhance Mortality duringLethal Influenza Infections by Eliminating Virus-Specific CD8T Cells
Ryan A. Langlois*,† and Kevin L. Legge*,†,‡
Previous studies have shown that the reduction in CD8 T cell immunity observed during high-dose influenza Avirus (IAV) infection
is mediated via lymph node (LN) dendritic cells (DCs) that express Fas ligand (FasL) and drive FasL-Fas (DC-T)–induced apoptosis.
However, the specific DC subset(s) within the LN and the additional factors required for DC-mediated elimination of IAV-specific
CD8 T cells remain unknown. In this paper, we demonstrate that plasmacytoid DCs (pDCs), which downregulate FasL during
sublethal, but not lethal, IAV infection, accumulate to greater numbers within the LNs of lethal dose-infected mice. Further our
findings show that pDCs from lethal, but not sublethal, dose IAV infections drive elimination of Fas+ CD8 T cells and that this
elimination occurs only in the absence of TCR recognition of IAV peptide-MHC class I complexes. Together, these results suggest
that pDCs play a heretofore unknown deleterious role during lethal dose IAV infections by limiting the CD8 T cell response. The
Journal of Immunology, 2010, 184: 4440–4446.
Clearance of primary influenza A virus (IAV) infections isgreatly enhanced by the generation of an effector CD8T cell response (1–3). Initially, these CD8 T cells are
primed by dendritic cells (DCs) within lung draining lymph nodes(LNs) (4–7) and then subsequently traffic to the lung, where theyeliminate virally infected cells via effector mechanisms, including;perforin, Fas ligand (FasL), and TRAIL (2, 8). Previously, we havedemonstrated a novel regulatory mechanism whereby lymph noderesident DCs (LNDCs) directly mediate apoptosis of IAV-specificCD8 T cells within the LN during lethal, but not sublethal, IAVinfections (9). This loss of IAV-specific CD8 T cells leads to de-creased numbers of CTLs that enter the lungs, resulting in a failureto clear the infection and ultimately the death of the host. Suchelimination of Fas+ IAV-specific CD8 T cells occurs subsequent tothe initial priming of CD8 T cells on days 3 and 4 postinfection(p.i.). In contrast to lethal dose IAV infections, during sublethalinfections, LNDCs downregulate FasL expression, allowing thedeveloping Fas+ IAV-specific CD8 T cells to escape apoptosis andtraffic into the lungs in sufficient numbers to clear the infection (9).At least six distinct populations of DCs have been described
within the lung-draining LN. There are four LNDC subsets that canbe identified phenotypically as CD4+ DCs, CD8a+ DCs and CD42
CD8a2 DCs, also known as double negative (DN) DCs (10–12).
Furthermore, in response to infection, plasmacytoid DCs (pDCs)are recruited into the LNs from the blood (13, 14). In addition tothese LNDCs, at least two respiratory DC (rDC) populations mi-grate from the lung into the draining LNs during infections (4–7).Once they enter the LN these migratory rDCs are thought to shareAgs with LNDCs, particularly with CD8a+ DCs, allowing thisLNDC subset to also participate in activation of naive CD8 T cells(7, 15). Interestingly, although Ags may be shared with all LNDCsubsets, not all LNDC subsets are able to present IAVAgs to naiveCD8 T cells as pDCs purified from the LNs of IAV-infected miceare unable to activate CD8 T cells directly ex vivo (4, 5, 7, 15). Insupport of the idea that pDCs do not participate in the activation ofnaive CD8 T cells, when pDCs are depleted in vivo during sublethalIAV infections there is no diminution of the CD8 T cell response (5,16). However, in contrast when pDCs are IAV-infected or pulsedwith IAV peptides in vitro they are now able to activate naive CD8T cells (17, 18). Together, these results suggest that although in-herently capable of presenting viral Ags to naive CD8 T cells, pDCmay not present IAV-Ags via MHC class I in the LN due to in-efficient processing of acquired viral proteins in vivo (5, 16).Although our previous work has demonstrated that LNDCs
eliminate IAV-specific CD8 T cells through a FasL-mediated path-way during lethal dose infections, it did not pinpoint the individualDC subset(s) involved, nor what role MHC class I presentation ofviral Ags by these DC subsets might play in the LNDC FasL-Fas–mediated elimination of IAV-specific CD8 T cells. This reportdemonstrates that whereas all LNDC subsets were FasL+ duringlethal dose IAV infection, only pDCs were able to eliminate acti-vated IAV-specific CD8 T cells. In contrast, pDCs isolated fromsublethal dose IAV-infected mice downregulated FasL and weretherefore unable to eliminate activated Fas+ CD8 T cells. Inter-estingly, our findings also demonstrate that the recruitment of pDCsinto the lung draining LNs was increased during lethal versus sub-lethal dose IAV infections, elevating the putative in vivo E:T ratio(i.e., pDC/T cell), and that transfer of FasL+ pDCs into mice de-ficient in functional FasL (i.e., gldmice)was sufficient to reverse thepreviously described protection of gld mice from lethal dose IAVinfections (9). Finally, this report demonstrates that lethal dose pDCelimination of IAV-specific CD8 T cells occurs in the absence of
*Department of Pathology, †Interdisciplinary Graduate Program in Immunology, and‡Department of Microbiology, University of Iowa, Iowa City, IA 52242
Received for publication September 9, 2009. Accepted for publication February 9,2010.
This work was supported by National Institutes of Health Grants AI-071085 and AI-076989 and by Department of Pathology Start-Up Funds (to K.L.L.).
Address correspondence and reprint requests to Dr. Kevin L. Legge, Department ofPathology, 1036ML, 200 Hawkins Drive, Iowa City, IA 52242. E-mail address:[email protected]
The online version of this article contains supplemental material.
Abbreviations used in this paper: 7-AAD, 7-aminoactinomycin D; DC, dendritic cell;DN, double negative; FasL, Fas ligand; IAV, influenza A virus; LN, lymph node;LNDC, lymph node resident dendritic cell; MFI, mean fluorescence intensity; n.s.,not significant; pDC, plasmacytoid dendritic cell; p.i., postinfection; rDC, respiratorydendritic cell; WT, wild-type.
Copyright� 2010 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/10/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0902984
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

direct viral-peptideMHC class I presentation. Taken together, thesedata suggest that pDCs are the cell type responsible for dampeningthe CD8 T cell response during lethal IAV infection in vivo andthat such elimination occurs in the absence of cognate IAV Agpresentation.
Materials and MethodsMice
Wild-type (WT) BALB/c mice were purchased from the National CancerInstitute (Frederick, MD). BALB/c CD90.1 congenic mice were kind giftsfrom Dr. Richard Enelow (Dartmouth College, Hanover, NH) and Dr. JohnT.Harty (University of Iowa, IowaCity, IA). Clone-4 (CL-4)TCR transgenicmice specific for the HA533/HA529 epitope of H1 and H2 IAV viruses, re-spectively were a kind gift from Dr. Linda Sherman (The Scripps ResearchInstitute, La Jolla, CA). BALB/c gld mice (CPt.C3-Tnfsf6gld/J) were ob-tained from The Jackson Laboratory (Bar Habor, ME). DUC18 TCR trans-genic mice specific for mutated tERK136–144 were kindly provided byDr. Paul Allen (Washington University, St. Louis, MO). All experimentswere performed in accordance with federal and institutional guidelines ap-proved by the University of Iowa Animal Care and Use Committee.
Virus infection
BALB/c mice 6–10 wk old were anesthetized by halothane or isofluoraneand infected intranasally with either a 10 LD50 or a 0.1 LD50 dose ofmouse-adapted A/JAPAN/305/57 in 50 ml Iscoves media. Viruses weregrown and stored as previously described (9).
MHC I tetramers
Tetramers HA204 (H-2K(d)/LYQNVGTYV), HA529 (H-2K(d)/IYATVAGSL),and NP147 (H2K(d)/TYQRTRALV) were obtained from the National Instituteof Allergy and InfectiousDiseaseMHCTetramer Core Facility (Atlanta, GA).
Flow cytometry
LN cells were stained with the following monoclonal Abs: rat anti-mouseCD8a (53-6.7), hamster anti-mouse CD11c (HL3), rat anti-mouse CD3ε(145-2C11), rat anti-mouse CD4 (CT-CD4), and rat anti-mouse CD43 (S7)purchased from BD Biosciences (San Jose, CA); mouse anti-mouse CD90.2(5a-8), rat anti-mouse CD45R (RA3-6B2), rat anti-mouse DX5, and rat anti-mouse CD19 (6D5) purchased from Caltag (Invitrogen, Carlsbad, CA).Anti-FasL CD95L (MFL3) was purchased from eBioscience (San Diego,CA). For FasL staining, cells were blocked with 1:100 rat serum, 1:100hamster serum, and 1:400 free streptavidin (Molecular Probes, Eugene, OR)on ice for 25 min. Cells were then washed twice and stained with 23 biotin-congugated anti-FasL (MFL3) followed by streptavidin-PE purchased fromBD Biosciences. For surface staining, isolated cells (106) were stained withAbs, and then fixed using BD FACS Lysing Solution (BD Biosciences). Allflow cytometry data were acquired on a BD FACS Calibur or BD FACSCanto II (BD Immunocytometry Systems/BD Biosciences) and analyzedusing FlowJo software (TreeStar, Ashland, OR).
CD8 CL-4 T cell purification and adoptive transfer
Spleens from CL-4 mice were removed and processed into single-cellsuspensions. Cells were labeled with anti-CD8a microbeads and purifiedaccording to the manufacturer’s instructions (Miltenyi Biotec, Auburn,CA). The purified CD90.2+ CL-4 cells (2 3 106) were then adoptivelytransferred i.v. into BALB/c CD90.1+ mice. At 24 h posttransfer, the hostmice were infected intranasally with a 0.1 LD50 IAV, as described above.For isolation of activated CL-4 CD8 T cells, lung draining LNs from IAV-infected CD90.1–CL-4 transferred host mice were removed on day 3 p.i.and digested, as described above. CD8 T cells were enriched using anti-CD8a beads according to the manufacturer’s instructions (Miltenyi Bio-tec). CD8a+ cells were then stained with Abs to CD90.2 and CD43, andactivated CD90.2+CD43+ CL-4 cells were sort purified using a FACS DiVa(BD Biosciences).
LNDC purification
Lung draining LNs (peribronchiolar and mediastinal) were removed anddigested with 4000 U type IV collagenase (Worthington Biochemical,Lakewood,NJ) and600UDNase1 (Sigma-Aldrich, StLouis,MO) in Iscovesmedia for 10min at room temperature. LNs were processed into a single-cellsuspension and RBCs lysed using NH4Cl-Tris. Cells were then stained withanti-CD3ε–PE and anti-CD19–PE mAb, followed by anti-PE microbeadsaccording to the manufacturer’s instructions (Miltenyi Biotec). Labeledcells were isolated using an autoMACS. The negative fraction was saved,stained with Abs to CD11c, CD45R (B220), CD8a, and CD4 and, witha FACS DiVa, sorted into CD11cmodCD45R+CD8+ cells (i.e., pDCs);CD11c+CD45R2CD8+ cells (i.e., CD8a+ DCs); CD11c+CD45R2CD82
CD42 cells (i.e., DN DCs); and CD11c+CD45R2CD82CD4+ cells (i.e.,CD4+ DCs).
Ex vivo LNDC killing assay
Purified LNDC subsets (104) were coincubated with 104 activated CL-4 cellsand 2.5 mg rmFas-human Fc (R&D Systems, Minneapolis, MN), 1 mMHA529 peptide (Bio-Synthesis, Lewisville, TX), or control media for 18 h at37˚C. To determine viability, cells were resuspended in 13 annexin bindingbuffer and stained with Annexin-V-APC and 7-aminoactinomycin D (7-AAD) (40% per BD Biosciences’s recommendation for 106 cells). Thedegree of DC killing of T cells was determined by measuring the fraction ofCD8 T cells that were nonapoptotic (i.e., Annexin-V2 7-AAD2, live cells)and normalizing this value to CD8 T cells cultured alone.
In vivo pDC transfer studies
Spleens from naive WT or gld mice were removed, and single-cell sus-pensions stained with anti–PDCA-1 microbeads and purified according tothe manufacturer’s instructions (Miltenyi Biotec). Then 23 106 WT or gldpDCs were adoptively transferred i.v. into gld mice 18 h post lethal IAVinfection. Some groups of pDCs were also pulsed with 1 mM HA529
peptide for 30 min at 37˚C. Mice were monitored daily for weight loss andmortality, or on day 4 p.i. lung draining LNs were removed to determineIAV-specific T cell responses in the LNs.
FIGURE 1. pDCs and CD8a+ DCs modulate FasL expression during IAV infection. Mice were infected with either a 10 LD50 (white portion of graph) or
a 0.1 LD50 (shaded portion of graph) of IAV, and on day 3 p.i. cells fromdrainingLNs (pooled) from each groupwere examined for FasL expression. The dotted line
represents staining with an isotype control mAb. A, FasL expression on CD11cmodCD45R+CD8+ cells (i.e., pDCs); 10 LD50 mean fluorescence intensity (MFI) =
60.5, and 0.1 LD50 MFI = 28.3. B, FasL expression on CD11c+CD45R2CD8+ cells (i.e., CD8a+ DCs); 10 LD50 MFI = 139, and 0.1 LD50 MFI = 118. C, FasL
expression on CD11c+CD45R2CD82 cells (i.e., CD8a2DCs); 10 LD50MFI = 618, and 0.1 LD50MFI = 496.2. TheMFI of staining with isotype control mAb has
been subtracted from the FasL MFI for the DC subsets, to yield the MFI reported above. Data are representative of five independent experiments.
The Journal of Immunology 4441
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
Statistical analysis
Statistical analysis between two data sets was performed using a one-tailedStudent t test. Differences were considered statistically significant at pvalues at or below 0.05. Statistical analysis for mortality experiments wasperformed using Kaplan–Meier survival analysis.
ResultsCD8a+ DCs and pDCs downregulate FasL expression duringlethal dose IAV infections
Our previous studies have demonstrated that reduction in IAV-specific CD8 T cell numbers in lethal dose IAV infections (Sup-plemental Fig. 1) is linked toFasL expression onDCswithin theLNs(9). This FasL expression on LNDCs decreases during sublethal,compared with lethal, IAV infection, therein allowing activatedeffector IAV-specific CD8 T cells to leave the LNs and traffic to thelungs to fight the infection. However, the decrease in FasL ex-pression on LNDCs during sublethal IAV infections was not uni-form, with some LNDCs maintaining high levels of FasL expres-sion. This finding suggested that a unique LNDC subset or subsetsmay be triggering the elimination of CD8 T cells during lethal doseIAV infection (9). To determine which LNDC subset or subsetsdifferentially modulate FasL expression during IAV infections,mice were infected with either a lethal or a sublethal dose of IAV,and the level of FasL on individual LNDC subsets was determinedon day 3 p.i., the time point where LNDC-mediated elimination ofIAV-specific CD8 T cells begins (9). Interestingly, although bothCD8a+ DCs and pDCs decreased FasL expression during sublethalcompared with lethal dose IAV infection, CD8a2 DCs, includingboth the CD4+ and DN DC subsets, did not modulate FasL ex-pression between the two IAV infection doses (Fig. 1). Further, therDC subsets, which have migrated from the lungs to the LNs duringIAV infection, remained FasL2 during both lethal and sublethalIAV infection (data not shown). Given that the pDC and CD8a+ DCsubsets are the only LNDC populations to downmodulate FasLexpression during sublethal dose IAV infections and that IAV-spe-cific CD8 T cell responses are rescued at this dose of infection(Supplemental Fig. 1) despite the remaining CD8a2 DC FasL
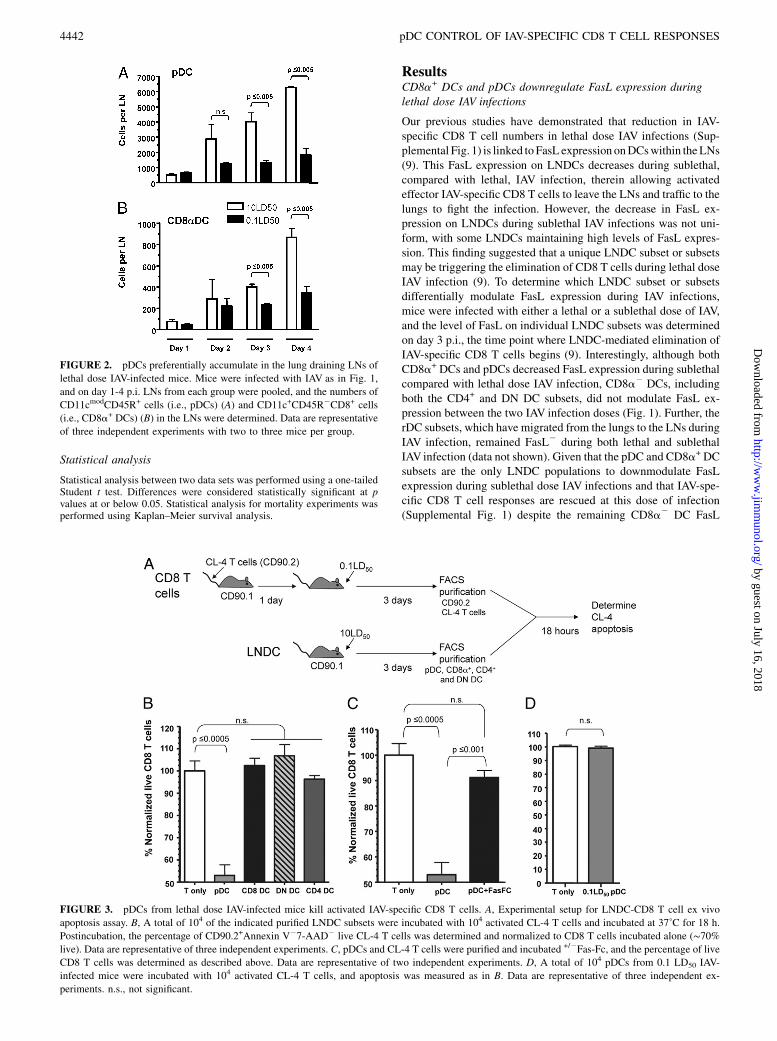
FIGURE 2. pDCs preferentially accumulate in the lung draining LNs of
lethal dose IAV-infected mice. Mice were infected with IAV as in Fig. 1,
and on day 1-4 p.i. LNs from each group were pooled, and the numbers of
CD11cmodCD45R+ cells (i.e., pDCs) (A) and CD11c+CD45R2CD8+ cells
(i.e., CD8a+ DCs) (B) in the LNs were determined. Data are representative
of three independent experiments with two to three mice per group.
FIGURE 3. pDCs from lethal dose IAV-infected mice kill activated IAV-specific CD8 T cells. A, Experimental setup for LNDC-CD8 T cell ex vivo
apoptosis assay. B, A total of 104 of the indicated purified LNDC subsets were incubated with 104 activated CL-4 T cells and incubated at 37˚C for 18 h.
Postincubation, the percentage of CD90.2+Annexin V27-AAD2 live CL-4 T cells was determined and normalized to CD8 T cells incubated alone (∼70%live). Data are representative of three independent experiments. C, pDCs and CL-4 T cells were purified and incubated +/2Fas-Fc, and the percentage of live
CD8 T cells was determined as described above. Data are representative of two independent experiments. D, A total of 104 pDCs from 0.1 LD50 IAV-
infected mice were incubated with 104 activated CL-4 T cells, and apoptosis was measured as in B. Data are representative of three independent ex-
periments. n.s., not significant.
4442 pDC CONTROL OF IAV-SPECIFIC CD8 T CELL RESPONSES
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

expression (Fig. 1) (9), the suggestion may be made that CD8a2
DCs are not the cells responsible for elimination of CD8 T cellsduring lethal dose IAV infections. Further, these results importantlyindicate that pDCs and/or CD8a+ DCs are likely responsible formediating FasL-driven CD8 T cell apoptosis during lethal dose IAVinfections.
pDCs accumulate in the lung draining LNs in greater numbersduring lethal dose IAV infections
Because our above results suggested the involvement of CD8a+
DCs and/or pDCs in the induction of CD8 T cell apoptosis duringlethal dose IAV infections, we next determined the kinetics andmagnitude of their recruitment/expansion within the lung drainingLNs during lethal and sublethal dose IAV infections. Our resultsshow that the number of both pDC and CD8a+ DC increases withinthe LNs between days 2 and 4 (Fig. 2); that is, the time point duringLNDC-mediated induction of CD8 T cell apoptosis occurs withinthe LNs (9). Specifically, the number of pDCs substantially in-creased in the LNs during lethal versus sublethal IAV infectionsbetween days 2 and 4, reaching significant differences by day 3 p.i.(Fig. 2A). Importantly, the number of pDCs present in the LNs wasapproximately six times greater than the number of CD8a+ DCspresent during lethal dose IAV infections. These results, togetherwith the fact that ∼6500 activated CD43+CD8 T cells were presentin the LNs on day 4 p.i. during lethal IAV infection (data notshown), indicate that a putative in vivo E:T (i.e., DC/activated Tcell) ratio for pDCs is conservatively∼1:1, compared with ∼1:6 forCD8a+ DCs (data not shown). Given that pDCs exhibited enhancedLN recruitment during lethal dose IAV infections (Fig. 2) and thatFasL expression on pDCs was dependent on the dose of IAV in-fection (Fig. 1), these data suggest that pDCs may be the pre-dominant LNDC population responsible for elimination of IAV-specific CD8 T cells during lethal IAV infections.
pDCs directly kill IAV-specific CD8 T cells during lethal doseIAV infections
To directly determine the LNDC subset responsible for drivingapoptosis of IAV-specific CD8 T cells during lethal dose IAVinfections, we used an ex vivo apoptosis assay. Briefly, transgenicCD90.2+ clone-4 T cells (Cl-4), which are specific for the HA529
epitope of IAV,were transferred into CD90.1 hosts, whichwere theninfected with a sublethal dose of IAV. Activated donor CD90.2+
CL-4 cells, which express higher levels of Fas (Supplemental Fig.2), were then purified on day 3 p.i. (i.e., the time directly before Tcell apoptosis occurs during lethal dose IAV infections) (9). Theseactivated CD90.2+ CL-4 T cells were then incubated with CD90.1+
LNDC subsets purified from day 3 lethal dose IAV-infected mice.Subsequent CL-4 CD8 T cell apoptosis was measured following18 h of coculture (Fig. 3). Interestingly, despite the fact that all lethaldose LNDC subsets express FasL at this time point (Fig. 1), onlypDCs induced statistically significant levels of apoptosis of the IAV-specific CL-4 CD8 T cells after coculture (Fig. 3B). The T cell ap-optosis induced by pDCs was FasL-Fas dependent as coculture in
FIGURE 5. pDC induction of apoptosis in IAV-specific
CD8 T cells is abrogated by cognate viral peptide-MHC I
presentation. A, CL-4 T cells and pDCs were purified as
described in Fig. 4 and cocultured +/21 mM HA529 pep-
tide, and then the percentage of live CD8 T cells was de-
termined as in Fig. 3. B, pDCs were purified as in Fig. 4,
pulsedwith1mMHA529 peptide, and transferred into IAV-
infectedmice, and CL-4 T cell numbers weremeasured as
in Fig. 4. Data are representative of two or three in-
dependent experiments. n.s., not significant.
FIGURE 4. Donnor WT pDCs reduce IAV-specific T cell numbers and
cause enhanced mortality in mice deficient in functional FasL. A, CD90.2
WT and gld mice received 2 3 106 CD90.1 CL-4 T cells and 24 h post-
transfer were infected with a 10 LD50 dose of IAV. At 24 h p.i., one group
of gld mice received 2 3 106 WT pDCs. On day 4 p.i., lung draining LNs
were removed, and CD3+CD8+CD90.1+ Cl-4 T cells were enumerated.
Data are pooled from two independent experiments with 8–10 mice per
group. B, gld or WT mice were infected with a 2.5 LD50 of IAV. At 24 h
p.i., pDCs were purified from naive WT or gld mice and then transferred
(2 3 106) into IAV-infected gld mice. Mortality was then monitored for
10 d p.i. p values are as follows: WT versus GLD = 0.0285, WT versus
(GLD + WT pDC) = 0.7013, GLD versus (GLD + WT pDC) = 0.0157,
(GLD + WT pDC) versus (GLD + GLD pDC) = 0.0103, and (GLD + GLD
pDC) versus GLD = 0.5722. Data are pooled from three independent ex-
periments with n values equaling WT = 15, GLD+WT pDC = 14, GLD+
GLD pDC = 9, and GLD = 18.
The Journal of Immunology 4443
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

the presence of Fas-Fc, which blocks FasL-mediated apoptosis andthereby abrogated the ability of pDCs to drive T cell apoptosis (Fig.3C). Given that pDCs during sublethal IAV infection downregulateFasL expression (Fig. 1),we next determined if pDCs from sublethaldose IAV-infected mice showed a similar ability to induce apoptosisof activated IAV-specific CD8 T cells. Consistent with their down-regulation of FasL during sublethal IAV infection, pDCs fromsublethal dose IAV-infectedmicewere unable to induce apoptosis ofIAV-specific CD8 T cells (Fig. 3D).
pDCs contribute to increased mortality during lethal IAV infection
Our previous studies have shown that gld mice (i.e., mice lackingfunctional FasL) exhibit increased IAV-specific CD8 T cell expan-sion and protection during lethal dose IAVinfections (9). Therefore,we used gld mice as hosts for pDCs to directly determine if WTpDCs (i.e., FasL+) could mediate a similar reduction in CD8 T cellresponses in vivo. Indeed, when WT pDCs were adoptively trans-ferred into gld mice, the number of CL-4 T cells within the LNs onday 4 p.i. was significantly reduced (∼65%) (Fig. 4A). In fact, thisreduction mirrored the number of IAV-specific T cells found in theLNs of lethal dose-infectedWTmice. Further. whenWTpDCsweretransferred into lethal dose IAV-infected gld mice, it resulted in anenhancedmortality thatwas statistically similar to that in lethal doseIAV-infected WT mice (Fig. 4B). This increase in disease severitywas FasL dependent, as adoptive transfer of gld pDCs into lethaldose IAV-infected gld mice did not increase the severity of disease.Importantly, in these latter experiments, the transferred donor pDCsare the only cells expressing FasL and therefore the only LNDCpopulation able to induce FasL-Fas–mediated apoptosis of IAV-specific CD8 T cells. Together, these data suggest that pDCs aresufficient to dampen in vivo the magnitude of the IAV-specific CD8T cell response, ultimately leading to an enhanced mortality.
pDCs eliminate IAV-specific CD8 T cells through FasL–Fasinteractions in the absence of cognate IAV Ag presentation
Given that both CD8a+ DCs and pDCs downregulate FasL ex-pression in sublethal dose IAV-infected mice, it was surprising thatonly the pDCs eliminated IAV-specific CD8 T cells directly ex vivo.Further, both CD4 and DN DCs expressed FasL during lethal doseIAV infections yet did not lead to any detectable apoptosis. Wetherefore next undertook experiments to determine the mechanismregulating this differential killing. Previous studies have demon-strated that pDCs isolated directly ex vivo from IAV-infected miceare unable to stimulate naive CD8 T cells, suggesting that pDCs donot present IAVAgs viaMHC class I in vivo (4, 5, 7, 15). In contrastto pDCs, CD8a+ DCs and CD8a2 DCs purified from the LNs ofIAV-infected mice are able to induce proliferation of naive CD8T cells directly ex vivo (4, 7, 15). Given this differential ability ofLNDC subsets to present IAVAg to CD8 T cells, we hypothesizedthat IAV peptide-MHC I presentation might rescue the IAV-specificCD8 T cells from FasL-mediated apoptosis. To test this hypothesis,activated CL-4 T cells were incubated with pDCs from the LNs ofday 3 lethal dose IAV-infected mice in the presence or absence ofexogenous IAV peptide, and the ability of pDCs to drive apoptosisof IAV-specific CD8 T cells was determined. Strikingly, culturingpDCs with IAV peptide abrogated their ability to eliminate IAV-specific CD8 T cells (Fig. 5A). To determine if Ag presentation bypDCs ablates their ability to eliminate IAV-specific CD8 T cellsin vivo, pDCs were pulsed with IAV peptide and transferred into gldmice; the number of transferred CL-4 cells was then measuredfollowing IAV infection on day 4 p.i. Unlike unpulsed pDCs (Figs.4A, 5B), pDCs pulsed with a cognate IAV peptide epitope did noteliminate IAV-specific CD8 T cells, leading to a similar T cell re-sponse to that observed in gld mice that did not receive pDCs (Fig.
5B). These data suggest that concomitant viral Ag peptide-MHC Ipresentation overrides the ability of pDCs to induce FasL-mediatedapoptosis in vitro and in vivo and suggest that pDC elimination ofIAV-specific CD8 T cells is critically tied to a lack of IAV Agpresentation. Therefore, the lack of apoptosis induction by CD8a+
and CD8a2 DCs may relate to their presentation of IAV Ags.Consistent with this idea, pDCs from lethal dose IAV-infectedmice were also able to mediate the apoptosis of in vitro activatedDUC18 (i.e., a non–IAV-specific T cell that displays an activationphenotype similar to that of in vivo activated CL-4 T cells; Fig. 6B,Supplemental Fig. 3), similar to their elimination of in vitro acti-vated CL-4 cells (Fig. 6A). Furthermore, although CD8a+ DCspurified from IAV-infected mice were unable to eliminate IAV-specific transgenic T cells (Figs. 3, 6A), they were capable ofeliminating non–IAV-specific transgenic T cells (Fig. 6B), sug-gesting that CD8a+ DCs can mediate elimination of activatedT cells in the absence of Ag presentation. Together these datasupport the idea that elimination of IAV-specific T cells duringlethal IAV infection does not require engagement of TCRs and infact TCR engagement may inhibit such apoptosis. Therefore, CD8T cell apoptosis may instead relate to the overall T cell activationstate and Fas expression.
DiscussionDC elimination of T cells through FasL–Fas interactions has beenpreviously described by multiple investigators (19–21). Suss andShortman (20) demonstrated that CD8a+ splenic DCs expressingFasL were able to eliminate CD4 T cells during an MLR. In ad-dition, multiple groups have used adoptive transfer of DCs trans-fected with FasL to control T cell numbers in a variety of diseasesettings, including autoimmunity, cancer, and viral infection (19,21, 22). In this paper, we have shown that LN resident pDCs canmediate the elimination of IAV-specific CD8 T cells during lethaldose IAV infections. Together, our results suggest that eliminationof activated Fas+ T cells by FasL+ pDCs may represent an integralmechanism for dampening T cell numbers.
FIGURE 6. pDCs from lethal dose IAV-infected mice eliminate IAV-
specific and non–IAV-specific activated CD8 T cells. CD8 T cells from the
spleens of naive CL-4 (A) and DUC18 (B) transgenic mice were purified
and cultured for 3 d in the presence of aCD3/CD28 Abs to activate the
T cells. Activated transgenic T cells (104) were incubated with pDCs or
CD8a+ DCs (104) from lethal dose IAV-infected mice at 37˚C for 18 h, as
described in Fig. 3. After incubation, the percentage of CD90.2+Annexin
V27-AAD2 live transgenic T cells was determined and normalized to CD8
T cells incubated alone. Data are representative of two independent ex-
periments. n.s. = p . 0.1.
4444 pDC CONTROL OF IAV-SPECIFIC CD8 T CELL RESPONSES
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Whereas DC-mediated reduction of the effector T cell responsein autoimmune reactions or at the conclusion of an immune responsewould be beneficial, the loss of effector CD8 T cells is clearly det-rimental to survival during a high-dose IAVinfection (9). Indeed, ourprevious studies have shown that in the absence of functional FasL,sufficient numbers of CD8 T cells develop to control the high-doseIAVinoculum (Fig. 4) (9). Thus, enhanced recruitment of pDCs intothe LNs observed during lethal dose IAVinfections (Fig. 2), coupledwith pDC-mediated elimination of the IAV-specific CD8 T cell re-sponse (Figs. 3, 4), allows the virus to escape adaptive immunecontrol, leading to death of the infected host. In this regard, recentstudies have shown that individuals infected with highly pathogenicavian (H5N1) IAV have dampened or reduced CD4 and CD8 T cellresponses (23). Similarly, mice and monkeys infected with highlypathogenic H5N1 IAV develop T cell lymphopenia (24–26), withloss of CD8 T cells in the lungs and lymph nodes, associated withenhanced levels of apoptosis (24). The exact pathway or pathwaysand cell type mediating the apoptosis responsible for T cell lym-phopenia remain poorly understood at this time. However, given ourresults, it will be important to determine what role FasL expressionby pDCs plays in the lymphopenia associated with highly patho-genic avian H5N1 IAV infections.Surprisingly, LNDCelimination of IAV-specificCD8T cells does
not require cognate MHC class I Ag presentation. In fact, the rescueof these T cells from FasL+ DC-induced apoptosis during recog-nition of viral peptide-MHC I complexes may be due in part toTCR-mediated upregulation of NK-kB, which also has been shownto protect T cells, macrophages, and B cells from Fas-mediatedapoptosis (27–30). Interestingly, signals through the BCR, whichupregulate NF-kB, result in transient (,24 h) protection from Fas-mediated apoptosis (28). In our studies, pDC-driven apoptosis oc-curs in the LNs at a time point concomitant with DC-mediated Agpresentation. Signaling through Fas on T cells during activation canact as a costimulatory molecule and has been demonstrated to in-crease T cell proliferation and activation (31–34). Together thesefindings suggest the possibility that during a narrow window im-mediately following the activation of naive T cells within the LNs,cognate MHC class I Ag–TCR interactions in the presence of FasL-Fas engagement may lead to enhanced T cell responses rather thanapoptosis. Conversely, FasL-Fas engagement alone or after egressfrom the draining LNs would mediate death. Consistent with ourresults showing that coculturing of pDCs and CL-4 T cells with IAVpeptide epitopes reverses the pDC-mediated loss of T cells (Fig.5A), a recent report has demonstrated that Ag-pulsed FasL-trans-fected DCs enhance Ag-specific CD8 T cell responses, rather thaninduce apoptosis. This finding suggests that the presentation of Agby these transfected DCs inhibits their ability to drive eliminationof cognate T cells (35). In contrast to this report, and our own re-sults, other groups have suggested that DC FasL-mediated elimi-nation of CD4 and CD8 T cells can occur in an Ag-dependentmanner (19, 21, 36). The reason for these differences is not clear atthis time; however, in contrast to our own studies, these latter ex-periments used conventional bone marrow-derived DCs or DC celllines, rather than pDCs obtained from the LNs. In addition, thesestudies used effector DCs transfected with FasL cDNA, resulting inconstitutively high levels of FasL.Our results suggest that pDC-mediated induction of apoptosis
in activated T cells during lethal dose IAV infections occurs in-dependently of IAVAg presentation (Fig. 5). In agreement with thisidea, recent reports have demonstrated that although LN residentpDCs contain IAV proteins following infection, they are unable tostimulate naive or memory CD8 T cells and may instead regulate Bcell responses (5). Unlike pDCs, LN resident CD8a+ DCs acquireIAV Ag (likely from migratory rDCs) and cross-present this Ag
during IAV infections (15). Interestingly, our results show that al-though CD8a+ DCs are also able to regulate FasL expression in anIAV dose-dependent manner (Fig. 1), they do not eliminate IAV-specific CD8 T cells during lethal dose IAV infections (Fig. 3). Thischaracteristic appears to be due to their presentation of viral pep-tides, as our preliminary results suggest that CD8a+ DCs from le-thally infected b2m
2/2 mice induce substantial apoptosis of IAV-specific T cells (data not shown). Together, these results, along withthose demonstrating that pDCs from lethal dose IAV-infected micekill both activated non–IAV-specific and IAV-specific CD8 T cellswith the same efficiency (Fig. 6A), indicate that pDC elimination ofeffector CD8 T cells during lethal dose IAV infections is in-dependent of TCR engagement. Rather, pDC-mediated eliminationof T cells is associated with the T cell activation state and Fas ex-pression. Thus, Fas-expressing activated or memory CD8 T cells ofany specificity might be susceptible to pDC-mediated apoptosisduring lethal dose IAV infections.In addition to differential FasL expression on pDCs from lethal
and sublethal dose IAV-infected mice, our results demonstrate anenhanced recruitment of pDCs into the LNs of lethal dose-infectedmice (Fig. 2). pDC recruitment into LNs through high endothelialvenules is thought to be mediated partly in response to CXCL12and CXCL9 expression (13, 14). Interestingly, CXCL9 is upregu-lated in response to IL-1 and IFN-g expression (37). Consistentwith these findings, our preliminary experiments have demon-strated increased secretion of IFN-g, IL-1a, and IL-1b from in vitrocultured LNs obtained from lethal compared with sublethal doseIAV-infected mice (data not shown). Importantly, as pDCs leave thehigh endothelial venules and enter the LNs, the increased re-cruitment of pDCs during lethal IAV infections will also raise thelocal in vivo E:T ratio. Even without factoring in a colocalization ofthese recruited pDCs into areas of the LNs that accumulate newlyactivated Fas+ T cells (38), our data suggest that in vivo there is ∼1pDC available for every activated CD8 T cell. In our ex vivoanalysis (Fig. 3), pDCs cultured at a 1:1 ratio with T cells were ableto reduce a static number of T cells by ∼45% in the span of 18 h.Therefore, the ∼80–95% reduction of endogenous T cell responseobserved in vivo on days 5 and 6 p.i. [(9) and Supplemental Fig. 1]may relate to the fact that elimination of activated T cells wouldalso reduce the subsequent burst size of the total response and/or toan increased local LNs in vivo E:T ratio.The data presented in this paper describe a novel role for pDCs
during lethal dose IAV infection—namely, the elimination of acti-vatedCD8T cells, leading to enhancedmortality. Given the emergingthreat of highly pathogenic pandemic IAV and the detrimental roleFasL+ pDCs can play during a lethal IAV infection, our findingssuggest that pDCs and FasLmay be strong candidates for therapeuticblockade during highly virulent IAV infections.
AcknowledgmentsWe thank Drs. J. Harty, J. Heusel, T. Waldschmidt, and S. Varga for critical
reading of this manuscript, the University of Iowa Flow Cytometry Facility
for expert assistance, and J. McGill and B. VanOosten-Anderson for tech-
nical assistance.
DisclosuresThe authors have no financial conflicts of interest.
References1. Eichelberger, M., W. Allan, M. Zijlstra, R. Jaenisch, and P. C. Doherty. 1991.
Clearance of influenza virus respiratory infection in mice lacking class I major
histocompatibility complex-restricted CD8+ T cells. J. Exp. Med. 174: 875–
880.
The Journal of Immunology 4445
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

2. Topham, D. J., R. A. Tripp, and P. C. Doherty. 1997. CD8+ T cells clear in-fluenza virus by perforin or Fas-dependent processes. J. Immunol. 159: 5197–5200.
3. Graham, M. B., and T. J. Braciale. 1997. Resistance to and recovery from lethalinfluenza virus infection in B lymphocyte-deficient mice. J. Exp. Med. 186:2063–2068.
4. Kim, T. S., and T. J. Braciale. 2009. Respiratory dendritic cell subsets differ intheir capacity to support the induction of virus-specific cytotoxic CD8+ T cellresponses. PLoS One 4: e4204.
5. GeurtsvanKessel, C. H., M. A. Willart, L. S. van Rijt, F. Muskens, M. Kool,C. Baas, K. Thielemans, C. Bennett, B. E. Clausen, H. C. Hoogsteden, et al.2008. Clearance of influenza virus from the lung depends on migratory langerin+CD11b- but not plasmacytoid dendritic cells. J. Exp. Med. 205: 1621–1634.
6. Legge, K. L., and T. J. Braciale. 2003. Accelerated migration of respiratorydendritic cells to the regional lymph nodes is limited to the early phase ofpulmonary infection. Immunity 18: 265–277.
7. Belz, G. T., C. M. Smith, L. Kleinert, P. Reading, A. Brooks, K. Shortman,F. R. Carbone, and W. R. Heath. 2004. Distinct migrating and nonmigratingdendritic cell populations are involved in MHC class I-restricted antigen pre-sentation after lung infection with virus. Proc. Natl. Acad. Sci. USA 101: 8670–8675.
8. Brincks, E. L., A. Katewa, T. A. Kucaba, T. S. Griffith, and K. L. Legge. 2008.CD8 T cells utilize TRAIL to control influenza virus infection. J. Immunol. 181:4918–4925.
9. Legge, K. L., and T. J. Braciale. 2005. Lymph node dendritic cells control CD8+T cell responses through regulated FasL expression. Immunity 23: 649–659.
10. Maldonado-Lopez, R., T. De Smedt, P. Michel, J. Godfroid, B. Pajak,C. Heirman, K. Thielemans, O. Leo, J. Urbain, and M. Moser. 1999. CD8alpha+and CD8alpha- subclasses of dendritic cells direct the development of distinctT helper cells in vivo. J. Exp. Med. 189: 587–592.
11. Maldonado-Lopez, R., T. De Smedt, B. Pajak, C. Heirman, K. Thielemans,O. Leo, J. Urbain, C. R. Maliszewski, and M. Moser. 1999. Role of CD8alpha+and CD8alpha- dendritic cells in the induction of primary immune responsesin vivo. J. Leukoc. Biol. 66: 242–246.
12. Henri, S., D. Vremec, A. Kamath, J. Waithman, S. Williams, C. Benoist,K. Burnham, S. Saeland, E. Handman, and K. Shortman. 2001. The dendritic cellpopulations of mouse lymph nodes. J. Immunol. 167: 741–748.
13. Yoneyama, H., K. Matsuno, Y. Zhang, T. Nishiwaki, M. Kitabatake, S. Ueha,S. Narumi, S. Morikawa, T. Ezaki, B. Lu, et al. 2004. Evidence for recruitmentof plasmacytoid dendritic cell precursors to inflamed lymph nodes through highendothelial venules. Int. Immunol. 16: 915–928.
14. Krug, A., R. Uppaluri, F. Facchetti, B. G. Dorner, K. C. Sheehan, R. D. Schreiber,M. Cella, and M. Colonna. 2002. IFN-producing cells respond to CXCR3 ligandsin the presence of CXCL12 and secrete inflammatory chemokines upon activation.J. Immunol. 169: 6079–6083.
15. Belz, G. T., S. Bedoui, F. Kupresanin, F. R. Carbone, and W. R. Heath. 2007.Minimal activation of memory CD8+ T cell by tissue-derived dendritic cellsfavors the stimulation of naive CD8+ T cells. Nat. Immunol. 8: 1060–1066.
16. Wolf, A. I., D. Buehler, S. E. Hensley, L. L. Cavanagh, E. J. Wherry, P. Kastner,S. Chan, and W. Weninger. 2009. Plasmacytoid dendritic cells are dispensableduring primary influenza virus infection. J. Immunol. 182: 871–879.
17. Fonteneau, J. F., M. Gilliet, M. Larsson, I. Dasilva, C. Munz, Y. J. Liu, andN. Bhardwaj. 2003. Activation of influenza virus-specific CD4+ and CD8+T cells: a new role for plasmacytoid dendritic cells in adaptive immunity. Blood101: 3520–3526.
18. Di Pucchio, T., B. Chatterjee, A. Smed-Sorensen, S. Clayton, A. Palazzo,M. Montes, Y. Xue, I. Mellman, J. Banchereau, and J. E. Connolly. 2008. Directproteasome-independent cross-presentation of viral antigen by plasmacytoid den-dritic cells on major histocompatibility complex class I. Nat. Immunol. 9: 551–557.
19. Matsue, H., K. Matsue, M. Walters, K. Okumura, H. Yagita, and A. Takashima.1999. Induction of antigen-specific immunosuppression by CD95L cDNA-transfected ‘killer’ dendritic cells. Nat. Med. 5: 930–937.
20. Suss, G., and K. Shortman. 1996. A subclass of dendritic cells kills CD4 T cellsvia Fas/Fas-ligand-induced apoptosis. J. Exp. Med. 183: 1789–1796.
21. Wolfe, T., C. Asseman, A. Hughes, H. Matsue, A. Takashima, and M. G. vonHerrath. 2002. Reduction of antiviral CD8 lymphocytes in vivo with dendriticcells expressing Fas ligand-increased survival of viral (lymphocytic choriome-ningitis virus) central nervous system infection. J. Immunol. 169: 4867–4872.
22. Kim, S. H., N. Bianco, R. Menon, E. R. Lechman, W. J. Shufesky, A. E. Morelli,and P. D. Robbins. 2006. Exosomes derived from genetically modified DC ex-pressing FasL are anti-inflammatory and immunosuppressive. Mol. Ther. 13:289–300.
23. de Jong, M. D., C. P. Simmons, T. T. Thanh, V. M. Hien, G. J. Smith, T. N. Chau,D. M. Hoang, N. V. Chau, T. H. Khanh, V. C. Dong, et al. 2006. Fatal outcome ofhuman influenza A (H5N1) is associated with high viral load and hyper-cytokinemia. Nat. Med. 12: 1203–1207.
24. Tumpey, T. M., X. Lu, T. Morken, S. R. Zaki, and J. M. Katz. 2000. Depletion oflymphocytes and diminished cytokine production in mice infected with a highlyvirulent influenza A (H5N1) virus isolated from humans. J. Virol. 74: 6105–6116.
25. Perrone, L. A., J. K. Plowden, A. Garcıa-Sastre, J. M. Katz, and T. M. Tumpey.2008. H5N1 and 1918 pandemic influenza virus infection results in early andexcessive infiltration of macrophages and neutrophils in the lungs of mice. PLoSPathog. 4: e1000115.
26. Baskin, C. R., H. Bielefeldt-Ohmann, T. M. Tumpey, P. J. Sabourin, J. P. Long,A. Garcıa-Sastre, A. E. Tolnay, R. Albrecht, J. A. Pyles, P. H. Olson, et al. 2009.Early and sustained innate immune response defines pathology and death innonhuman primates infected by highly pathogenic influenza virus. Proc. Natl.Acad. Sci. USA 106: 3455–3460.
27. Dudley, E., F. Hornung, L. Zheng, D. Scherer, D. Ballard, and M. Lenardo. 1999.NF-kappaB regulates Fas/APO-1/CD95- and TCR- mediated apoptosis of Tlymphocytes. Eur. J. Immunol. 29: 878–886.
28. Carey, G. B., D. Donjerkovic, C. M. Mueller, S. Liu, J. A. Hinshaw, L. Tonnetti,W. Davidson, and D. W. Scott. 2000. B-cell receptor and Fas-mediated signalsfor life and death. Immunol. Rev. 176: 105–115.
29. Lu, B., L. Wang, D. Medan, D. Toledo, C. Huang, F. Chen, X. Shi, andY. Rojanasakul. 2002. Regulation of Fas (CD95)-induced apoptosis by nuclearfactor-kappaB and tumor necrosis factor-alpha in macrophages. Am. J. Physiol.Cell Physiol. 283: C831–C838.
30. Schram, B. R., and T. L. Rothstein. 2003. NF-kappa B is required for surface Ig-induced Fas resistance in B cells. J. Immunol. 170: 3118–3124.
31. Chun, D. H., K. C. Jung, W. S. Park, I. S. Lee, W. J. Choi, C. J. Kim, S. H. Park,and Y. Bae. 2000. Costimulatory effect of Fas in mouse T lymphocytes. Mol.Cells 10: 642–646.
32. Maksimow, M., T. S. Soderstrom, S. Jalkanen, J. E. Eriksson, and A. Hanninen.2006. Fas costimulation of naive CD4 T cells is controlled by NF-kappaB sig-naling and caspase activity. J. Leukoc. Biol. 79: 369–377.
33. Maksimow, M., M. Santanen, S. Jalkanen, and A. Hanninen. 2003. Respondingnaive T cells differ in their sensitivity to Fas engagement: early death of manyT cells is compensated by costimulation of surviving T cells. Blood 101: 4022–4028.
34. Siegel, R. M., F. K. Chan, H. J. Chun, and M. J. Lenardo. 2000. The multifacetedrole of Fas signaling in immune cell homeostasis and autoimmunity. Nat. Im-munol. 1: 469–474.
35. Buonocore, S., N. O. Haddou, F. Moore, S. Florquin, F. Paulart, C. Heirman,K. Thielemans, M. Goldman, and V. Flamand. 2008. Neutrophil-dependent tu-mor rejection and priming of tumoricidal CD8+ T cell response induced bydendritic cells overexpressing CD95L. J. Leukoc. Biol. 84: 713–720.
36. Kusuhara, M., K. Matsue, D. Edelbaum, J. Loftus, A. Takashima, and H. Matsue.2002. Killing of naive T cells by CD95L-transfected dendritic cells (DC): in vivostudy using killer DC-DC hybrids and CD4(+) T cells from DO11.10 mice. Eur.J. Immunol. 32: 1035–1043.
37. Sanmiguel, J. C., F. Olaru, J. Li, E. Mohr, and L. E. Jensen. 2009. Interleukin-1regulates keratinocyte expression ofT cell targeting chemokines through interleukin-1 receptor associated kinase-1 (IRAK1) dependent and independent pathways. Cell.Signal. 21: 685–694.
38. Barchet, W., A. Blasius, M. Cella, and M. Colonna. 2005. Plasmacytoid dendriticcells: in search of their niche in immune responses. Immunol. Res. 32: 75–83.
4446 pDC CONTROL OF IAV-SPECIFIC CD8 T CELL RESPONSES
by guest on July 16, 2018http://w
ww
.jimm
unol.org/D
ownloaded from


















