Physiological mechanisms controlling anestrus and infertility in postpartum beef.pdf
20
R E Short, R A Bellows, R B Staigmiller, J G Berardinelli and E E Custer cattle. Physiological mechanisms controlling anestrus and infertility in postpartum beef 1990, 68:799-816. J ANIM SCI http://www.journalofanimalscience.org/content/68/3/799 the World Wide Web at: The online version of this article, along with updated information and services, is located on www.asas.org by guest on September 24, 2014 www.journalofanimalscience.org Downloaded from by guest on September 24, 2014 www.journalofanimalscience.org Downloaded from
Transcript of Physiological mechanisms controlling anestrus and infertility in postpartum beef.pdf

R E Short, R A Bellows, R B Staigmiller, J G Berardinelli and E E Custercattle.
Physiological mechanisms controlling anestrus and infertility in postpartum beef
1990, 68:799-816.J ANIM SCI
http://www.journalofanimalscience.org/content/68/3/799the World Wide Web at:
The online version of this article, along with updated information and services, is located on
www.asas.org
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

PHYSIOLOGICAL MECHANISMS CONTROLLING ANESTRUS AND INFERTILITY IN POSTPARTUM BEEF
R. E. Short3 R. A. Bellows3, R. B. Staigmiller3, J. G. Berardinelli4 and E. E. Custep
U. S. Department of Agriculture, Miles City, MT 59301 and Montana State University, Bozeman 59717
ABSTRACT
Postpartum infertility is caused by four factors: general infertility, lack of uterine involution, short estrous cycles and anestrus. The general infertility component is common to any estrous cycle and reduces potential fertility by 20 to 30%. Incomplete uterine involution prevents fertilization during the first 20 d after calving but is not related to anestrus. Short estrous cycles prevent fertility during the first 40 d after calving by causing the cow to return to estrus before pregnancy recognition occurs. Anestrus is the major component of postpartum infertility and is affected by several minor factors: season, breed, parity, dystocia, presence of a bull, uterine palpation and carryover effects from the previous pregnancy as well as two major factors: suckling and nuhition. These major factors have direct effects on anestrus but also interact with one or more other factors to control postpartum anestrus. Physiological mechanisms associated with anestrus involve blockage of the GnRH “pulse generator” in the hypothalamus, but other pathways also must be involved because bypassing the pulse generator is not an effective treatment for all cows. The primary cause of anestrus probably is different for different stages of anestrus. The mediating mechanisms for anestrus are not involved with prolactin, oxytocin, the adrenal or direct neural input from the mammary gland but are at least partially involved with blood glucose and the endogenous opioid peptide system. Management options to decrease the impact of anestrus and infertility include: 1) restrict breeding season to 545 d; 2) manage nutrition so body condition score is 5 to 7 before calving; 3) minimize effects of dystocia and stimulate estrous activity with a sterile bull and estrous synchronization; and 4) judicious use of complete, partial or short-term weaning. (Key Words: Cattle, Beef, Postpartum Interval, Infertility, Anestrus.)
J . Anim. Sci. 1990. 68:7’)9-816
‘This research is a contribution 10 Western Regional Re- search Project W-112, Reproductive Performance in Do- mestic Ruminants. Conuibution No. 2288 from Montana Agric. Exp. Sta. Presented ai a symposium titled “Postpar- tum Rebrdiiig in Cattle” at the ASAS Both Annu. Mtg.. New Brunswick, NJ.
2The authors express appreciation to former graduate students P. F. Clemente, D. L. Walters and G . L. Faltys for their contributions to this work and to The Upjohn Company. Kalamazoo, MI for financial support and generous contribu- tions of Lutalyse and to American Breeders Service, De- Forest, WI for semen used in some of these studies.
3USDA-ARS. Ft. Keogh Livest. and Range Res. Lab., Rt. 1 Box 2021, Miles City, MT.
4Dept. of Anim. and Range Sci., Montana State Univ., Bozeman.
Received January 3.1989. Accepted June 12,1989.
Introduction
Reproduction is the main factor limiting production efficiency of beef cattle (Dickerson, 1970; Dziuk and Bellows, 1983; Koch and Algeo, 1983). The largest loss of the potential calf crop occurs because cows fail to become pregnant (Wiltbank et al., 1961; Bellows et al., 1979). Postpartum anestrus or the interval from parturition to estrus (postpartum interval) largely determines the likelihood of cow’ becoming pregnant during the breeding season (Symington, 1969; Wiltbank, 1970). The pur- pose of this review is to enumerate factors that affect the postpartum interval of cows and to show how these factors affect the endocrine system.
799
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

800 SHORT ET AL.
TABLE 1. EARLY HISMRY OF POSTPARTUM RESEARCH
Referena Location commentsa
Hammmd (1927) Guilben and McDonald (1933) Clapp (1937)
Lasley and Bogan (1943) Casida and Wisnicky ( 1950) Jouben (1954) Wamick (1955) Bunis and =ode (1958) Wiltbank and Cook (1958) Foote et al. (1960a.b)
Biswal and Rao (1960) Wiltbank et al. (1962, 1963) Perkms and Kiddw (1 %3) Foote and Hunter (1964) BhaIla et al. (1966,1967) Oxenrcider ( 1968) Saiduddii et al. (1%8a) Casida (1%8); Graves et al.
England California Wisconsin
Missouri Wisconsin S. Africa Oregon USDA USDA Wisconsin
India USDA W. Virginia Nevada India Iowa Nevada Wisconsin
Herdsmen reported nurse cows had longer PPI. Distribution of PPI in beef cows. Four times milked or suckled dairy cows has longer PPI than those two times milked. PPI of Hereford range cows. Diethylstibcsterol (DES) did not affect PPI. Nutritional effects on calving interval, beef and daily. PPI for Angus and Hereford and its relationship to day of calving. Date of calving vs pregnancy rate. PPI of suckled and milked cows. Uterine involution not related to PPI; pretreatment did not affect uter- ine involution but delayed estrus. Weaning reduced PPI and effect decreased with increasing parity. Re- and post-calving nutritional effects on PPI in Hereford cows. Uterine involution did not affect conception rate, PPI more important. P4 and E2 treclunents reduced PPI, short estrous cycles. Parity and weaning decreased PPI. Suckling effects in beef cows. PPI shortened by steroid treatment. Most definitive study in postpartum cow.
(1968) Lauderdale et al. (1968); Riesen et al. 1968; Saiduddin et al. (1968b) Shocl et al. (1972) USDA Suckling effect independent of nutrition; mastectomy enhances wean-
ing effect.
*PPI = Interval from calving to estrus.
Postpartum infertility and anestrus were first recognized as problems over 60 yr ago (Hammond, 1927). Research on postpartum reproduction progressed slowly for a period of time (see Table 1 for a summary of the early work in this area). A tremendous increase in postpartum research occurred during the last 20 yr; the majority was reported in the last decade (see Figure l5 for summary of research activity during the last 60 yr). In spite of the research activity during the last decade, post- partum infertility is still a significant problem in the industry, but, as shown in Figure 1, research activity in this area is decreasing rapidly.
Over the years, several review papers summarizing reproduction and anestrus re- search in cattle have been published either as independent articles (Baker, 1969; Wiltbank,
5A computer database of postpartum cow literature with over 700 references was compiled by R. E. Short and B. W. Knapp and is available from the authors on computer disc or hard copy with any of the following fields: authors, year, title. journal, volume and page, keywords, author address and abstract.
1970; Tervit et al., 1977; Kilkenny, 1978; Lamming et al., 1981; Garcia, 1982; Peters, 1984a; Hanzen, 1986; Short and Adams, 1988) or as part of various books or proceedings of symposia (Ninth Animal Reproduction Sympo- sium: Casida, 1971; W. C. Foote, 1971; W. D. Foote, 1971; Tucker, 1971; Wagner and Oxenreider, 1971 - BARC Symposium on Animal Reproduction: Bellows et al., 1979; Inskeep and Lishman, 1979 - Fourteenth Animal Reproduction Symposium: Wette- mann, 1980; Kiracofe, 1980; Dunn and Kalten- bach, 1980 Edgerton, 1980; Christensen, 1980 - Diamond Jubilee Symposium for American Society of Animal Science: Koch and Algeo, 1983; Dziuk and Bellows, 1983 - Current Topics in Vet. Med. and Animal Science: Terqui, 1985; Schallenberger and Walters, 1985 - Reproductive Endocrinology of Do- mestic Ruminants Symposia: Nett, 1987). However, there are no recent reviews that are comprehensive and address the problem of postpartum infertility and anestrus in beef cattle from a practical point of view. There- fore, the emphasis of this review is to cover this subject for beef cattle with relevant literature included from both beef and dairy cattle because the difference between these two
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

POSTPARTUM ANESTRUS AND INFERTILITY 80 1
80
70 C n I fertility by 20 to 30% for any estrus regardless of whether it occurs after calving or at any other reproductive state. This component will not be discussed further, but it must be recognized as a factor in postpartum infertility. The other three factors are related specifically to the postpartum period and will be discussed. Shown in Figure 2 are the time relationships that several components have to infertility; the order of the four factors listed does not indicate relative importance.
25 35 45 55 65 75 85 95 YEAR (19--)
Figure 1. Numbers of citatim reviewed on postpartum cow research from 1927 to 1988 (673 citations, complter database; Short and Knapp5).
is due primarily to management and not to physiology. Literature that pertains mainly to the management aspects of postpartum dairy cattle is not included (e.g., use of milk progesterone for detecting ovarian cycles).
Factors Affecting Padparturn Infertility
After parturition, cows are infertile for a variable period of time. Several factors con- tribute to that infertility. Illustrated in Figure 2 are 1) the relationship between probability of a pregnancy occuning (infertility vs fertility) and time after calving and 2) the four main factors that contribute to infertility. A general infertility component exists that decreases
Uterine Involution
Kiracofe (1 980) thoroughly reviewed the literature on this subject and concluded that uterine involution has no relationship to length of the anestrous period. Nevertheless, uterine involution is a barrier to fertility during the early postpartum period. Graves et al. (1968) recovered ova or palpated for pregnancy in cows that were bred at the first postpartum estrus. They found that fertilization rates and pregnancy rates were very low when cows were bred <20 d after calving, but fertility returned to normal between 20 to 40 d after calving.
In several experiments, we bred cows that showed estrus very early after calving (Short et al., 1974). Half the cows were bred by normal AI procedures, and half were bred by surgi- cally &positing semen into the tip of the uterine horns. Ova were recovered 3 d after insemination to determine fertilization rates. Inseminations into the tip of the uterine horn reduced the interval to conception by increas-
Figure 2. Relationship of f d l i t y (probability of a pregnancy occurring) 10 time after calving. The factors contributing to infertility arc shown above the fmility probability line.
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

802 SHORT ET AL.
ing the proportion of cows with fertilized ova when inseminations were done before 20 d after calving. Fertilization rates after 20 d were similar for both groups. We concluded that infertility during the first 20 d after calving is caused by a physical barrier to sperm transport and not by any inherent defect in the ova or other physiolcgical mechanisms. We do not have data on whether an early postpartum uterus actually could maintain a pregnancy if a fertilized ovum were present. The nonin- voluted uterus may be a barrier not only to sperm transport but also to implantation.
From a practical point of view, uterine involution is not a problem for beef cattle because it does not affect anestrus. Very few cows would exhibit estrus early enough after calving for uterine involution to interfere with conception so long as disease conditions do not prevent or delay normal involution.
Short Estrous Cycles
Short estrous cycles also contribute to postpartum infertility during the first 30 to 40 d after calving (Figure 2). Most estrous cycles after 40 d have normal length, although some evidence exists that short estrous cycles will occur later (Lishman et al., 1979). The short estrous cycle phenomenon was one of the first abnormalities observed in the postpartum cow (Foote and Hunter, 1964; Morrow et al., 1968; Lauderdale et al., 1968; Casida, 1971; Short et al., 1972). Ovulations following esms des- tined to start a short cycle are normal with ova released that can be fertilized. However, no pregnancies were detected, apparently because the corpus luteum (CL) regressed before the ovary received a signal from the uterus that a pregnancy existed (Graves et al., 1968; Short et al., 1972, 1974; Odde et al., 1980; Ramirez- Godinez et al., 1982a,b).
Why is this first estrous cycle short? Is it because the CL is not capable of functioning normally, or is it because regression signals are being given prematurely? The CL formed during a short estrous cycle is smaller, secretes less progesterone (P4) and is less responsive to stimulation (Short et al., 1974; Lishman et al., 1979; Kesler et al., 1981; Rutter and Randel, 1984; Duby et al., 1985; Carmthers et al., 1986). Gonadotropic stimulation of the short- cycle CL appears to be normal and not to be a limitation to its development (Manns et al., 1983; Carmthers et al., 1986 Garverick et al.,
1988), although FSH concentrations may be low preceding this ovulation (Ramirez-Godi- nez et al., 1982b). The short-cycle CL is not hypersensitive to prostaglandin (Copelin et al., 1988), nor is there any evidence of a problem with receptors or enzymes (Smith et al., 1986). So far, this evidence doesn’t answer the question asked earlier; it merely reinforces the evidence that the short-cycle CL is function- ally deficient.
Prostaglandin F2a (PGF) appears to be the normal physiological signal whereby the uterus causes regression of the CL at the end of the estrous cycle. During the early postpartum period, high PGF concentrations probably prolong the interval to normal CL function because hysterectomy reduces PGF concentra- tions and maintains normal CL function (Edqvist et al., 1978; Lindell et al., 1982a,b; Madej et al., 1984; Copelin et al., 1987). The early postpartum uterus produces and metabo- lizes large amounts of PGF, and suppression or infusions of PGF further implicate a role of PGF in the control of CL function in the postpartum cow (Guilbault et al., 1984, 1987a,b). Therefore, evidence exists to answer the question raised earlier. The functional capabilities of the early-postpartum CL are normal, but the CL is caused to regress prematurely by abnormally high PGF concen- trations from the uterus. Regression occurs before the signal can be given that a pregnancy exists. These high FGF concentrations presum- ably are a part of the mechanisms involved in involution of the uterus. Uterine involution is involved with prevention of pregnancy, not only blocking fertilization as described earlier, but also by preventing normal CL function.
Normal CL function during an early post- partum estrous cycle can be obtained by pretreating with a progestin (Ramirez-Godinez et al., 1981; Pratt et al.. 1982; Sheffel et al., 1982; Rutter et al., 1985; Smith et al., 1987). The mechanism associated with the progestin effect involves a reduction in PGF (Troxel and Kesler, 1984) with the effect mediated via the ovary rather than the pituitary. Some evidence exists for a LH-follicle involvement (Williams et al., 1982b; Garcia-Winder et al., 1986b). Even ovarian or follicular involvements could be mediated indirectly through alteration in uterine endocrinology.
The short-estrous-cycle syndrome is a prac- tical beef cattle problem because some cows exhibit estrus and are bred during the period
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

POSTPARTUM ANESTRUS AND INFERTILITY 803
that these estrous cycles occur, especially in more intensive operations where postpartum intervals are short and estrous synchronization is used. For early postpartum cows, estrous synchronizing treatments that include a proges- tin may have an advantage (see review by Odde, 1990).
Postpartum Anestrus
In beef production systems, postpartum anestrus can affect infertility for a long time (Figure 2); therefore, anestrus is a more serious problem than either uterine involution or short estrous cycles. P o s t p m anestrus commonly is referred to as postpartum interval (PPI), the interval from pamuition to first estrus. Inter- vals to other endpoints such as to ovulation or to conception also are important, but from a practical point of view, estrus is the most logical measure of the commencement of potential fertility.
Ovulation may precede the first observed estrus and often is called a “silent” or “quiet” heat (Casida and Venzke, 1936; Labsetwar et al., 1963). The true incidence of this phenom- ena will be affected by how well we “listen” (Casida, 1971). but even under ideal estrous detection conditions, some ovulations are not accompanied by a detectable estrus. This condition is much more prevalent in milked dairy cows and when ovulation occurs very early after calving.
The opposite condition, estrus without ovulation, has been observed at puberty (non- puberal estrus, Nelsen et al., 1985; Rutter and Randel, 1986), but it has been reported only once during postpartum anestrus in cattle, and that was in diury canle (Casida, 1968). The appropriate experiments have not been con- ducted to determine the extent to which this condition exists in beef cattle.
Postpartum anestrus or PPI is a€fected by several minor factors, including season, breed, age or parity, dystocia, presence of a bull, uterine palpation and carryover effects from the previous pregnancy, as well as by the two major factors of suckling and nutrition. These factors all have direct effects on PPI, but each factor can interact with one or more of the other factors. Because of the many factors and interactions that affect PPI, control and man- agement of postpartum anestrus is complex. Minor Factors. Season was one of the first factors to be associated with postpartum
anestrus (Buch et al., 1955; Warnick, 1955). In the study reported by Warnick, season was included in the analysis as day-of-year calved with spring-calving cows. Cows calving later in the spring had shorter PPI. In this case, day- of-year effects could be true season effects associated with light and temperature differ- ences, or the effect could be confounded with nutritional changes as year progressed. Subse- quent work has confirmed that day-of-year decreases PPI as the spring calving season progresses and has shown that cows calving from late spring to early fall have shorter PPI than cows calving from late fall to early spring (Bellows and Short, 1978; Peters and Riley, 1982a,b; King and MacLeod, 1983; Smeaton et al., 1986). These seasonal effects are not due to nutritional and management differences (Boyd, 1977; Bellows and Short, 1978) but are true seasonal effects related to changes in light (Hansen and Hauser, 1984; Hauser, 1984; Garel et al., 1987). However, seasonal effects are modified by numtion and other factors such as genotype and suckling (Hansen and Hauser, 1983; Hansen, 1985; Montgomery et al., 1985). From recent evidence, it appears that seasonal effects are mediated via the pineal because injections of melatonin will increase PPI (Sharpe et al., 1986). Not much can be done through management to correct seasonal effects on PPI other than to adjust for seasonal effects.
Breed and genotype effects on postpartum anestrus can be seen by making comparisons between studies. Dairy breeds that are milked have shorter PPI than suckled beef breeds, but when dairy cows are suckled, they have longer PPI than beef cows. Comparisons also have been made within studies that have shown that, when managed comparably, dairy genotypes have longer PPI than beef genotypes and that these effects are more pronounced at first parity and at lower dietary intakes (Kropp et al.. 1973; Hansen et al., 1982). Differences also have been reported between beef breeds, with the effect being more pronounced at lower &etary intakes (Dunn et al., 1969; Bellows and Short, 1978). How genotype affects PPI is not known; effects may be due to true physiological differences among breeds, and(or) they may be due to confounding effects such as differences in amount of milk produced or appetite and feed intake. The effect of breed on PPI is an important factor, and even though breed usually is predeter-
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

804 SHORT ET AL.
mined, this effect must be considered when managing postpartum cows.
The effect of age and parity on PPI is due mostly to the difference between 2- and 3-yr-old dams or older, with young cows having longer PPI and lower reproductive potential. Dystocia also is associated with age and will increase PPI and delay rebreeding (Brinks et al., 1973; Laster et al., 1973; Bellows and Short, 1978; Doornbos et al., 1984; Izaike et al., 1984). The adverse effects of dystocia can be overcome at least partially by providing early obstemcal assistance (Bel- lows et al., 1988).
Presence of a bull decreases PPI (2alesky et al., 1984; Alberio et al., 1987; Berardinelli et al., 1987; Scott and Montgomery, 1987; Custer, 1988). The mechanism whereby pres- ence of a bull accelerates the physiological processes that initiate resumption of estrous cycles is not understood; the available evi- dence implicates no involvement with either P4 or LH (Custer, 1988). Putting sterile teaser bulls with cows during the early postpartum period could be a useful tool in managing postpartum anestrus in beef cattle.
Postpartum reproduction also may be af- fected by other factors such as rectal palpation, treatment with a synthetic prostaglandin (Tol- leson and Randel, 1988) and effects of the calf. The effects of the calf apparently are related to differences in growth rate and amount of milk consumed; fast-growing, larger calves and(or) calves consuming more milk will have dams with longer PPI (Rice et al., 1961; Reynolds et al., 1978; Bellows et al., 1982). These effects are more subtle or are fixed and are less likely to be manipulated by management. However, they should be considered when devising management strategies.
Major Factors. Suckling and nutrition are by far the most important factors determining the length of postpartum anestrus. Suckling probably has the most dramatic effect on PPI and was one of the first factors to be related to postpartum reproduction (Clapp, 1937; Wilt- bank and Cook, 1958). Cows that have their calves weaned at birth have shorter PPI than do cows that are suckled (Graves et al., 1968; Oxenreider, 1968; Short et al., 1972). If calves are weaned at some time after birth but before estrous cycles begin (usually between 20 and 40 d after calving), cows will return to estrus in a few days (Walters et al., 1982b). Regulation of the suckling and lactation
stimulus is a viable management option to decrease PPI. Postpartum intervals can be decreased by complete weaning, short-term weaning (48 h) or partial weaning (restricting suckling to short periods of time each day). However, response to weaning treatments will vary with other factors such as age, nutrition, genotype of the cow and age of the calf. The effects of suckling and lactation have been reviewed in this symposium as well as in an earlier symposium (Edgerton, 1980 Williams, 1990).
Nutritional effects were firmly established as a factor controlling postpartum anestrus by the classical work of Wiltbank (Wiltbank et al., 1%2, 1963; Dunn et al., 1969). Reviews on the effects of nutrition have detailed these effects (Dunn and Kaltenbach, 1980; Short and Adams, 1988; Randel, 1990). Nutritional ef- fects are elicited via a complex interplay among many variables such as quantity and quality of feed intake, nutrient reserves stored in the body and competition for nutrients from other physiological functions besides reproduc- tion. The effects of nutrition have been most commonly measured using energy as a varia- ble. Other nutrients also may affect reproduc- tion, but their effects are not well documented.
The allocation of nutrients to various body functions is commonly referred to as nutrient partitioning. A schematic representation of these relationships is shown in Figure 3 (Short and Adams, 1988). Illustrated in Figure 3 are several points about nutritional and production interrelationships. The first is that cattle (and other ruminants) have a unique niche in animal agriculture because they have the ability to convert lowquality forages to useful products. The second is that excess nutrients can be stored during excess periods to be retrieved at a later time to maintain production. However, extended periods of limited nutrient availabil- ity because of low quantity and(or) poor quality will decrease production. The third is that nutrients are partitioned by a priority to first maintain life of the cow and then to propagate the species. The approximate order of priority for partitioning of nutrients is as follows: 1) basal metabolism, 2) activity, 3) growth, 4) basic energy reserves, 5) pregnancy, 6) lactation, 7) additional energy reserves, 8) estrous cycles and initiation of pregnancy and 9) excess reserves. The relative priority of these functions can change depending on what functions are present and at what level.
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

POSTPARTUM ANESTRUS AND INFERTILITY 805
Figure 3. Partitioning of numents in a cow with nutrient intake varying in quantity and quality (from Shon and Adams, 1988).
Energy reserves can constitute almost 50% of an animal’s maximum possible weight (Short et al., unpublished data). One of the simplest ways to estimate nutrient reserves is the use of body condition scores (BCS; Lowman et al., 1973; Bellows et al., 1982; Richards et al., 1986). As shown in Figure 3, estrous cycles usually can be maintained if BCS is 4 or greater, although this may differ depending on other factors such as breed and whether an animal is entering into or coming out of anestrus (Hansen et al., 1982; Louw et al., 1988).
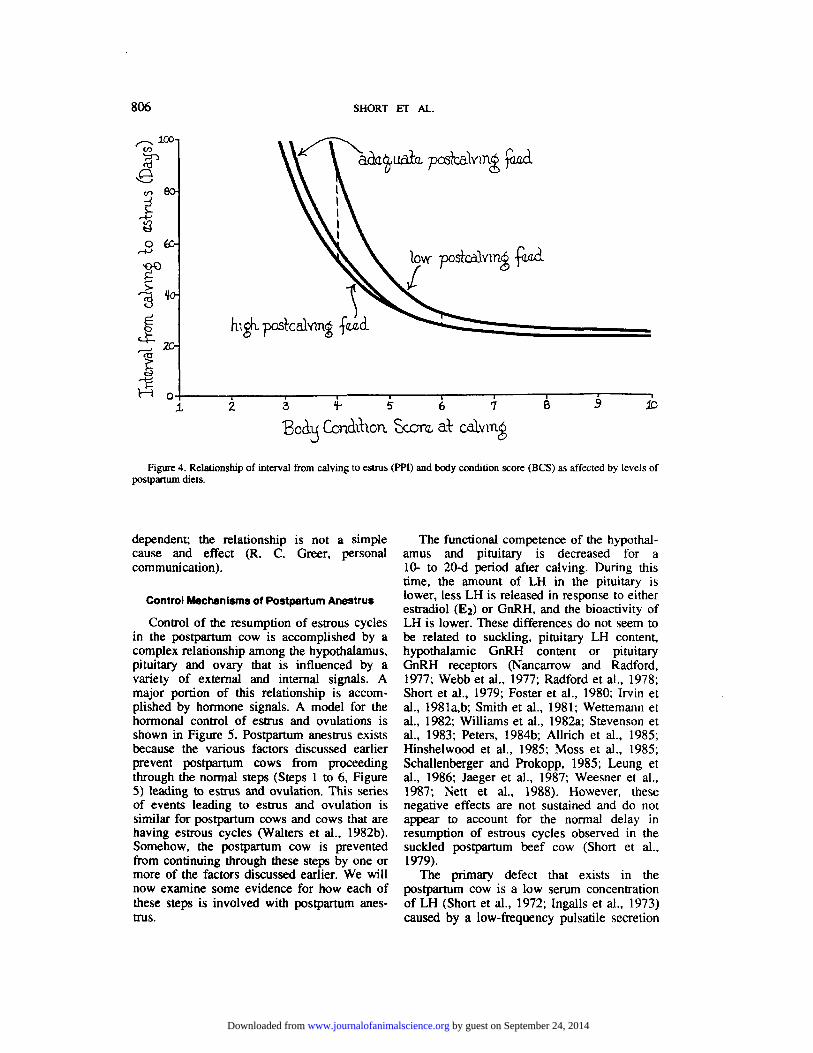
The effect of nutrition on postpartum reproduction depends somewhat on whether nutritional differences exist before or after calving. In general, the differences that exist at calving as estimated by precalving BCS are more important than those that exist after calving. These relationships are depicted in Figure 4. The effects of BCS at calving on PPI are nonlinear, with the effects being greatest at
very low scores (<4), becoming less as BCS increases, with very little effect as BCS increases above 7. Postcalving dietary intake can alter these relationships, but postcalving differences in diets have their main effect when BCS at calving is <6 and when the feed intake is low vs adequate (100% of NRC) rather than adequate vs high.
A degree of caution should be exercised when interpreting simple correlation and re- gression relationships among BCS and PPI. Both of these variables are physiological responses in a system in which the relation- ships may not be cause-effect but merely are cogenerated. This type of relationship has been shown between age at first estrus and weight at first estrus in heifers (Greer et al., 1983). Analyses of postpartum data using a path hagram approach and generalized least squares procedures to estimate parameters of a simultaneous equations model have shown that BCS at calving and PPI are recursively
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from
806 SHORT ET AL.
Figure 4. Relationship of interval from calving to estrus (PPI) and body condition score (BCS) as affected by levels of postpartum diets.
dependent; the relationship is not a simple cause and effect (R. C. Greer, personal communication).
Control Mechnnlrms of Postpartum Anestrus
Control of the resumption of estrous cycles in the postpartum cow is accomplished by a complex relationship among the hypothalamus, pituitary and ovary that is influenced by a variety of external and internal signals. A major portion of this relationship is accom- plished by hormone signals. A model for the hormonal control of estrus and ovulations is shown in Figure 5. Postpartum anestrus exists because the various factors discussed earlier prevent postpartum cows from proceeding through the normal steps (Steps 1 to 6, Figure 5) leading to estrus and ovulation. This series of events leading to estrus and ovulation is similar for postpartum cows and cows that are having estrous cycles (Walters et al., 1982b). Somehow, the postpartum cow is prevented from continuing through these steps by one or more of the factors discussed earlier. We will now examine some evidence for how each of these steps is involved with postpartum anes- trus.
The functional competence of the hypothal- amus and pituitary is decreased for a 10- to 204 period after calving. During this time, the amount of LH in the pituitary is lower, less LH is released in response to either estradiol (E2) or GnRH, and the bioactivity of LH is lower. These differences do not seem to be related to suckling, pituitary LH content, hypothalamic GnRH content or pituitary GnRH receptors (Nancarrow and Radford, 1977; Webb et al.. 1977; Radford et al., 1978; Short et al., 1979; Foster et al., 1980; Irvin et al., 1981a,b; Smith et al., 1981; Wettemann et al., 1982; Williams et al., 1982a; Stevenson et al., 1983; Peters, 1984b; Allrich et al., 1985; Hinshelwood et al., 1985; Moss et al., 1985; Schallenberger and Prokopp, 1985; Leung et al., 1986; Jaeger et al., 1987; Weesner et al., 1987; Nett et al., 1988). However, these negative effects are not sustained and do not appear to account for the normal delay in resumption of estrous cycles observed in the suckled postpartum beef cow (Short et al., 1979).
The primary defect that exists in the postpartum cow is a low serum concentration of LH (Short et al., 1972; Ingalls et al., 1973) caused by a low-frequency pulsatile secretion
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

POSTPARTUM ANESTRUS AND INFERTILITY
ANESTRUS and
807
Figure 5. A model depicting the hormonal control of esttu and ovulation in postpartum cows.
pattern of LH and, presumably, GnRH (Steps 2 and 3, Figure 5). During postpartum anestrus, the pulse pattern of LH typically has a very low frequency (<1 pulse/4 h). The frequency increases during the time preceding estrus to about one pulse every 1 to 2 h (Carruthers and Hafs, 1980 Carruthers et al., 1980; Walters et al., 1982a, Schallenberger. 1985). The GnRH “pulse generator“ somehow must be inhibited during the early postpartum period.
A pulsatile pattern of LH release can be reproduced by giving low-dose injections of GnRH every 1 to 2 h. Postpartum cows treated in such a manner have shorter postpartum intervals (Riley et al., 1981; Walters et al., 1982c; Jagger et al., 1987; Wildeus et al., 1987). However, all cows do not respond to such treatment, especially if they are early in the postpartum period or on a low dietary intake (Short et al., 1981; Edwards et al., 1983; Peters et al.. 1985; Jagger et al., 1987; Short and Adams, 1988). The inconsistency in the response to these treatments leads us to conclude that inhibition of the GnRH “pulse generator” is not the only cause of anestrus.
The mechanisms for control of anestrus may be different depending on the cause (i.e., nutrition vs suckling), or the primary defect may change as the postpartum cow progresses from “deep” anesnus to “shallow” anestrus with time after calving. The amount of time required to make this transition varies with the amount of input from each of the controlling factors.
A pulsatile pattern of GnRH release may be only the strategy evolved to maintain a certain level of secretion because continuous infusions of GnRH will elicit a response similar to pulse injections (Lofstedt et al., 1981; Jagger et al., 1987). Prolonged infusions of GnRH result in a decreased release of LH over time, but this decreased sensitivity to GnRH is not reflected by the endogenous pulsatile release of LH (Lamming and MacLeod, 1988).
At present, there is no evidence implicating FSH as a limiting factor in postpartum anestrus (Walters et al., 1982a; Moss et al., 1985).
The ovarian component of anestrus (Step 4, Figure 5 ) is highly dependent on the status of the preceding steps, but in general the ovary
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

808 SHORT ET AL.
per se does not Seem to be a limiting factor because it will respond to exogenous stimula- tion (Casida et al.. 1943). Follicular develop ment and function are normal, although there are some carryover effects of the CL of pregnancy. Follicles are selected from a pool of medium-sized follicles, and as follicles mature, they show an increase in number of LH receptors and switch from P4 to E2 production. Atresia is not related to gonadotro- pin binding (Braden et al., 1986; Spicer et al., 1986a,b,c; Inskeep et al., 1988). The main evidence for the ovary’s involvement in anestrus is that the pituitary-hypothalamus is hypersensitive to negative feedback of estrogen rather than an absence of the positive feedback mechanism discussed earlier (Acosta et al., 1983; Garcia-Winder et al., 1986a; Chang and Reeves, 1987).
This hypersensitivity to estrogen gradually is overcome to allow Steps 2 to 4 to proceed, resulting in the continuation through preovula- tory LH release, estrus and ovulation (Steps 5 and 6, Figure 5) . Then the cycle repeats itself or pregnancy occurs (Steps 7a or 7b).
The stimuli that are detected by control mechanisms to retard or block the normal progression through these steps in Figure 5 are not understood, but several possibilities have been investigated. A role of the adrenal was implicated by some circumstantial evidence, but careful experimentation has ruled this out (Wagner and Oxenreider, 1971; Dunlap et al., 1981a,bcc; Ellicot et al., 1981; Smith et al., 1974; Whisnant et al., 1985; Faltys et al., 1987; Spicer and Zinn, 1987). Prolactin also was explored as a possible mediator but was ruled out (Smith et al., 1974; Cummins et al., 1977; Clemente et al., 1978; Schallenberger et al., 1978; Holness and Jeffers, 1980 Williams and Ray, 1980; Webb and Lamming, 1981; Weiss at al., 1981; Montgomery, 1982; Whee- ler et al., 1982). Oxytocin was injected three times daily in an experiment, and we found no evidence that oxytocin was involved with anestrus (Short et al., 1972); this was con- fumed by Stewart and Stevenson (1987). A direct neural involvement from the udder does not seem to be present (Short et al., 1976; Williams, 1990).
The only positive evidence for a control pathway involves glucose (see review by Short and Adams, 1988) and the endogenous opioid peptide (EOP) system. Even though evidence exists that the EOP system is involved in the
control of LH release during the postpartum period (Gregg et al., 1986; Malven et al., 1986; Short et al., 1986; Whisnant et al., 1986a,b,c,d; Cross et al., 1987; Peck et al.. 1988; Trout and Malven, 1988), this system is only a part of the control system because prolonged treatment with an EOP antagonist only temporarily will release LH without subsequent ovulation (Short et al., 1986).
Conclusions and Management Considerations
Postpartum infertility and anestrus are com- plex phenomena controlled by many factors that act either individually or in concert to decrease the production potential of beef cattle. Many of the factors that affect postpartum reproduction cannot be eliminated, but they can be considered when one makes manage- ment decisions relative to other factors that can be manipulated. In making a management decision about postpartum anestrus, one must consider not only the potential benefits of a treatment on postpartum fertility but also the cost of implementing this treatment. For most production systems, maximum fertility will not be the most profitable. Optimum fertility rate for a given production unit is that which returns the greatest profit (or some other defined measure of success) over a period of time. Unfortunately, only limited data are available on the costs of most treatments that affect postpartum fertility, so that aspect of the decision making process is left to the individ- ual producer.
The first management decision that has a major impact on postpartum fertility is the length of the breeding season. Having a restricted breeding season has many advan- tages, such as a more uniform, older calf crop, but examination of Figure 6 reveals an even more important reason for limiting the breed- ing season to 45 d or less (Figure 6 is Figure 2 redrawn with the addition of variable lengths of breeding seasons). The three examples of breeding seasons shown (45, 60 and 82 d) illustrate stages of the postpartum period for cows when the next breeding season starts. This assumes a gestation period of 280 d and an interval of 365 d between the start of the two breeding seasons. If breeding seasons are 45 d or less, than all cows have a high probability for pregnancy at the beginning of the breeding season because they are beyond the effects of uterine involution and short
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

POSTPARTUM ANESTRUS AND INFERTILITY 809
py qftar Calnng
Figure 6. Relationship of tbe length of the breeding Season to fertility during the postpartum period.
estrous cycles and beyond the major effects of anestrus if proper management has been accomplished (see later points on this). Con- versely, as breeding seasons extend beyond 45 d, more and more cows will have a lower potential fertility at the beginning of the next breeding season. Cows that calve in the last 2 wk in a 60-d breeding season system or in the last 6 wk in a 90-d breeding system will have increasing problems with anestrus, short es- trous cycles and uterine involution. Any cow that is bred after 82 d in a long breeding season system will not have calved by the time the next breeding season starts. If producers presently are using a long breeding season (A5 d), it can be a slow but beneficial process to change to a shorter season. Because cows that presently are in a long breeding season program are difficult to convert to a short breeding season, it is best to start with the replacement heifers and gradually work the breeding season shorter as these heifers and each subsequent year’s replacements enter the herd. By using this method and culling nonpregnant and late-calving cows, the whole herd can be shifted to a breeding season of 45 d or less in a few years.
The second management decision to con- sider for decreasing postpartum infertility is nutrition. Nutritional levels and BCS have major impact on anestrus and potential fertili- ty. The best way to monitor a nutritional
program is by following body weight changes and BCS. Often it is not practical to measure body weight in many production settings, but in most situations a procedure can be devel- oped for monitoring BCS. For optimum reproduction, it is best to manage for a BCS at calving of 5 to 7 (on a 9- or 10-point scale). Condition scores higher than 7 are not necessary and probably waste valuable re- sources unless severe nutritional deficiencies are anticipated after calving. Condition scores lower than 5 will lower potential fertility unless there is a high availability of numents after calving. It is difficult and expensive to correct nutritional problems that exist at calving by increasing dietary intake after calving. When selecting a target condition score at calving for managing a specific group of cows, it is important to consider other factors unique to that herd such as breed, amount of milk produced and incidence of dystocia and then adjust the precalving BCS as necessary.
The third management decision to minimize postpartum infertility is to consider several options that include minimizing stress from dystocia, putting a sterile bull with cows during the early postpartum period before the breeding season starts and using estrous synchronizing treatments that include a proges- tin to increase estrous response and decrease the incidence of short estrous cycles. The
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

810 SHORT
deleterious effects of dystocia can be mini- mized by careful attention to the following: selection of replacement heifers, selection of sires to be used on replacement heifers, management of both cows and heifers before and after calving and providing appropriate obstetrical assistance at parturition. Putting a bull with cows during the postpartum period to reduce postpartum infertility is a new ap- proach. More information is needed concern- ing the optimum bukcow ratio, when the bulls should be turned in and whether other forms of social interaction will elicit the same response (e.g.. androgenized cows). However, the rela- tive cost of using bulls is low and should be considered by producers. If estrous synchroni- zation is a viable alternative in a production system, it can be used to shorten the effective breeding season and take advantage of possible stimulatory effects of a progestin.
The fourth and last management alternative to consider is decreasing the suckling stimulus. We view this alternative as a last resort because this alternative potentially has more serious economic and management conse- quences, such as an increased potential for calf diseases and for reduced calf weaning weights. All the other management alternatives should be considered and dealt with before resorting to this option. The suckling stimulus can be decreased by complete weaning, limiting the time during which suckling can occur to once or twice a day or weaning for a short period of time (48 h) associated with an estrous synchro- nizing treatment. In order for these treatments to be effective for reducing infertility, they must be accomplished at some point before the breeding season starts. However, in some cases of limited feed supplies later in lactation, weaning can be an effective tool for managing BCS going into the winter in order to minimize fertility problems in the subsequent year. In all cases of partial or complete weaning, the potential economic and manage- ment disadvantages should be carefully weighed against the potential advantages of increased fertility.
Implications
Postpartum infertility can be a serious problem that decreases production efficiency of beef production systems. Many factors contribute to infertility through a complex interaction of physiological and endocrine
ET AL.
mechanisms. Anestrus is the main component of postpartum infertility, but uterine involution and short estrous cycles as well as a general infertility component also are involved. Man- agement options to increase fertility during the postpartum period include shorter breeding seasons, using BCS to monitor nutritional management, minimizing dystocia, using a sterile teaser bull before breeding, synchroniz- ing esms and decreasing the suckling stimu- lus.
Literature Cited
Acosta, B., G. K. T X M V S ~ ~ , T. E. Platt, D. L. Hamernik. 1. L. Brown, H. M. Schoenemann and J. J. Reeves. 1983. Nursing enhances the negative effect of estrogen on LH release in the cow. J. Anim. Sci. 57:1530.
Alberio. R. H.. G. Schiersmann, N. Carou and 1. Mestre. 1987. Effect of a teaser bull on ovarian and behavioral activity of suckling beef cows. Anim. Reprod. Sci. 14: 263.
Allrich. R. D., J. R. Parfet. C. A. Marvin. G. E. Moss and P. V. Malven. 1985. In vitro release of LHRH from the bovine pituitary stalk-median eminence alterations during the postpartum period. Domest. Axurn. Endo- crinol. 2:191.
Baker, A. A. 1969. Post partum anoestrus in cattle. A u t . Vet. J. 45:180.
Bellows, R. A. and R. E. Short. 1978. Effects of precalving feed level on birth weight, calving difficulty and subsequent fertility. I. Anim. Sci. 46:1522.
Bellows, R. A., R. E. Short and G. V. Richardson. 1982. Effects of sire, age of dam and gestation feed level on dystocia and postpartum reproduction. J. Anim. Sci. 55:18.
Bellows, R. A,. R. E. Short and R. B. Staigmiller. 1979. Research areas in beef cattle reproduction. In: H. Hawk (Ed.) Proc. Beltsville Symp. on Animal Reproduction. No. 3. p 3. Allanheld, Osmun, Mont- clair.
Bellows, R. A,, R. E. Short, R. B. Staigmiller and W. L. Milmine. 1988. Effects of induced panwition and early obstetrical assistance in beef cattle. J. Anim. Sci. 66: 1073.
Berardinelli, J. G., M. S. Roberson and R. Adair. 1987. Shortening the anestrus period. Montana AgResearch 4(2):25.
Bhal1a.R. C.. DP.S. Sengar and B. K. Soni. 1967. Studies on post-parturn o e s t ~ ~ ~ in Murrah buffaloes and Sahiwal cows and factors affecting them. Indian J. Dairy Sci. 20: 189.
Bhalla, R. C.. B. K. Soni and D.P.S. Sengar. 1966. Studieson reproduction in Murrah buffaloes. 11. Involution of uterus. Indian Vet. 1. 433392.
Biswal, G. and A. M. Rao. 1960. Effect of weaning on Red- Si& cows. Part I. Lactational length, lactation yield, daily milk yield and days dry. Indian Vet. 1. 37:379.
Boyd, H. 1977. Anoestrus in cattle. Vet. Rec. 100:150. Braden, T. D., J. G. Manns. D. L. Cermak, T. M. Nett and G.
D. Niswender. 1986. Follicular development following pamuition and during the estrous cycle in beef cows. Theriogenology 25:833.
Brinks, J. S., J . E. Olson and E. J. Carroll. 1973. Calving
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

POSTPARTUM ANESTRUS AND INFERTILITY 81 1
difficulty and its association with subsequent prcduc- tivity in Herefords. J. h i m . Sci. 36:l l .
Buch, N. C.. W. 1. Tyler and L. E. Casida. 1955. Postpartum e s m and involution of the uterus in an experimental herd of Holstein-Friesian cows. J. Dairy Sci. 38:73.
Burris, M. J. and B. M. Priode. 1958. Effect of calving date on subsequent calving performance. J. h i m . Sci. 17: 527.
Carruthers, T. D.,E. M. Convey, J. S. Kesner, H. D. Hafs and K. W. Cheng. 1980. The hypothalamo-pituitary gonadotrophic axis of suckled and nonsuckled dairy cows postpartum. J. Anim. Sci. 51949.
Carruthers, T. D. and H. D. Hafs. 1980. Suckling and four- times daily milking: Influence on ovulation, esuus and serum luteinizing hormone, glucocorticoids and pro- lactin in postpartum Holsteins. J. Anim. Sci. 50:919.
Carrurhers, T. D., J. G. Manns and L. M. Rutter. 1986. Failure of human chorionic gonadotropin injections to sustain gonadotropin-releasing hormone-induced cor- pora lutea in postpartum beef cows. Biol. Reprod. 35: 846.
Casida, L. E. 1968. The postpartum cow-A resume. Univ. of Wisconsin Res. Bull. No. 270, pp 48-54, Madmn.
Casida, L. E. 1971. The postpartum interval and its relation to fertility in the cow, sow and ewe. J. Anim. Sci. 32(Suppl. 1):66.
Casida, L. E., R. K. Meyer, W. H. McShan and W. Wisnicky. 1943. Effects of pituitary gonadotropins on the ovaries and the induction of superfecundity in cattle. Am. J. Vet. Res. 4:76.
Casida. L. E. and W. G. Venzke. 1936. Observations on reproductive processes in dairy cattle and their relation to breedingeffciency. Roc. Am. Soc. Anun. Rod. 36: 221.
Casida, L. E. and W. Wisnicky. 1950. Effects of diethylstil- bestrol dipropionate upon postpartum changes in the cow. J. Anim. Sci. 9238.
Chaug. C.-F. and J. J. Reeves. 1987. Postpanum interval in beef cows shortened by enclomiphene. J. Anim. Sci. 65:217.
Christenson, R. K. 1980. Environmental influences on the postpartum animal. J. h m . Sci. 5l(Suppl.I1):53.
Clapp, H. 1937. A factor in breeding efficiency of dairy cattle. Roc. Am. Soc. Anim. Prod., p 259.
Clemente, P. F., R. E. Short, R. B. Staigmiller. R. A. Bellows, C. C. Kaltenbach and T. G. Dunn. 1978. Effect of precalving nutrition, early weaning, CB-154. and antiprolactin ueatment on postpartum interval lengih in beef cows. I. Anim. Sci. 47(Suppl. 1):351 (Ab=.).
Copelin, I. P., M. F. Smith, H. A. Garverick and R. S. Youngquist. 1987. Effect of the uterus on subnormal luteal function in anesuous beef cows. J. Anim. Sci. 64:1506.
Copelin, J. P., M. F. Smith, H. A. Garverick, R. S. Youngquist. W. R. McVey, Jr. and E. K. Inskeep. 1988. Responsiveness of bovine corpora lutea to prostaglandin F2a: Comparison of corpora lutea anticipated to have short or normal lifespans. J. h i m . Sci. 66:1236.
Cross, J. C., L. M. Rutter and J. G. Manns. 1987. Effects of progesterone and weaning on LH and FSH responses to naloxone in postpartum beef cows. Domest. h i m . Endocrinol. 4: 1 I 1.
Cummins, L. J., 1. K. Findlay and R.A.S. Lawson. 1977. An attempt at the use of CB-154 (ergocryptine) to reduce post-partum anoestrus in Hereford heifers. Thenogen-
ology 8:193. Custer, E. E. 1988. Postpartum interval to csms and patterns
of luteinizing hormone (LH) ConcenKations in first- calf suckled beef cows exposed to mature bulls. M. S. Thesis. Montana State Univ., Bozeman.
Dickerson, G. 1970. Efficiency of animal production-mold- ing the biological components. J. Anim. Sci. 30:849.
hrnbos, D. E., R. A. Bellows, P. J. Burfening and B. W. Knapp. 1984. Effects of dam age, prepartum nutrition and duration of labor on productivity and postpartum reproduction in beef females. J . h i m . Sci. 59:l.
Duby, R. T., T. Browning, D. Carey and D. L. Black. 1985. Progesterone synthesis and histology of postpartum bovine corpora lutea. Theriogenology 23:619.
Dunlap. S. E., T. E. Kiser, N. M. Cox, F. N. Thompson, G. B. Rampacek, L. L. Benyshek and R. R. Kraeling. 19Xla. Cortisol and luteinizing hormone after adrenocroticouopic hormone administration to post- partum beef cows. J. Anim. Sei. 52587.
Dunlap. S. E., T. E. Kiser, G. B. Rampacek, R. R. Kraelmg and F. N. Thompson. 1981b. Effect of suckling 011
cortisol, progesterone and luteinizing hormone in postpartum beef cows. Theriogenology 16: 185.
Dunlap. S. E.. T. E. Kiser, J. J. Vallner, G. B. Rampacek, R. R. Kraeling and L. L. Benyshek. 1981~. Clearance of serum cortisol in suckled and nonsuckled postpanum beef cows. 1. Anim. Sci. 53:1082.
Dunn. T. G., J. E. Ingalls, D. R. Zimmerman and J. N. Wiltbank. 1969. Reproductive performance of 2-year-old Hereford and Angus heifers as influenced by pre- and post-calving energy intake. J. Anim. Sei. 29:719.
Dunn, T. G. and C. C. Kaltenbach. 1980. Nutrition and the postpartum interval of the ewe, sow and cow. 1. Anim. Sci. 51(Suppl. II):2Y.
h i & , P. 1. and R. A. Bellows. 1983. Management of reproduction of beef cattle, sheep and pigs. J. Anini. Sci. 57(Suppl. 2):355.
Edgerton, L. A. 1980. Effect of lactation upon the postpartum interval. I. Anim. Sci. 51(Suppl. 11):40.
Edqvist, L.-E. , H. Kindahl and G. Stabenfeldt. I 9 7 X . Release of prostaglandin Fza during the bovine penpartal period. Prostaglandins 16: 11 1,
Edwards, S., J. F. Roche and G. D. Niswender. 1983. Response of sucklmg beef cows to multiple, low-dose injections of Gn-RH with or without progesterone pretreaunent. J. Reprod. Fertil. 6965.
Ellicott, A. R.. D. M. Heruicks, T. Gimenez and T. E. Kiser. 1981. Suckling induced cortisol secretion in young beef cows. Theriogenology 16:469.
Faltys, G. L.. E. M. Convey, R. E. Short, C. A. Keech and R. L. Fogwell. 1987. Relationship between weaning and secretion of luteinizing hormone, cortisol and transcor- tin in beef cows. I. Anirn. Sci. 64:1498.
Foote, W. C. 1971. Some influences of lactation and hormone treament on uterine changes in postpartum sheep. J. Anim. Sci. 32(Suppl. I):48.
Foote. W. D. 1971. Endocrine changes in the bovine during the postpartum period. J. Anim. Sci. 32(Suppl. 1):73.
Foote, W. D.. E. R. Hauser and L. E. Casida. 1960a. Some causes of variation in post-partun reproductive activity in Hereford cows. J. Anim. Sci. 19:238.
Foote, W. D.. E. R. Hauser and L. E. Casida. 1960b. Influence of progesterone treatment on post-pru7um reproductive activity in beef cattle. J. Anim. Sci. 19: 674.
Foote, W. D. and J. E. Hunter. 1964. Post-partum intervals of
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

812 SHORT
beef cows mated with progesterone and esuogen. J. Anim. Sci. 23517.
F o s l ~ . J. P.. G. E. Lamming and A. R. Petas. 1980. Short- ~ u m relationships between plasma LH. FSH and
and Ihe effect of Gn-RH injection. J. Reprod. Fenil. 5 9 321.
Garcia, M. 1982. Reproductive functions during the post p a n m period in the cow. Nord. Veterinaermed. 34: 264.
Garcia-Winder, M.. K. Imakawa. M. L. Day, D. D. Zalesky. R. J. Kittok and J. E. Kindcr. 1986a. Effccts of suckling and low dosts of estradiol on luteinizing hormone sccrctim during the postpartum period in beef cows. Domera Anim. Endocrinol. 3:79.
Garcia-Winder, M. P. E. Lewis. D. R. Deaver, V. G. Smith, G. S. Lewis and E. K. Ioskeep. 1986b. Endocrine profiles associated with life span of induced corpora lulea in postpartum beef cows. I. Anim. Sci. 621353.
Garel. J. P., D. Gauthier, M. &tit and J. 'Ibimonier. 1987. Influence of photoperiod on IIIC post-partum changes in live weight and ovarian function in suckled cow. Re@. Nutr. Dev. 27:3M.
Gmerick, H. A., J. R. Parfet, C. N. Lee. J. P. Copelin. R. S. Youngquist and M F. Smith. 1988. Relationship of p- and post-ovulatory gonadoaopin concentrations to subnormal luteal function in posipartum beef cattle. J. Anim. Sci. 66:104.
Graves, W. E.. J. W. Lauderdale. E. R. Hauser and L. E. Casida. 1968. Relation of postpartum interval to pituitary gonadotropins. ovarian follicular develop- ment and fertility in beef cows (Effect of suckling and intmal to breeding). Univ. of Wisconsin Res. Bull.
Grca, R. C., R. W. Whirman. R. B. Staigmiller and D. C. Anderson 1983. Estimating the impact of management decisions on the occurrence of puberty in beef heifers. J. Anim. Sci. 56:30.
Gregg. D. W., G. E. Moss. R. E. Hudgens and P. V. Malven. 1986. Eadogtnous opioid modulation of luteinizing hormone aad prolactin secretion in postpartum ewes and cows. J. Anim. Sci. 63838.
Guilbault, L. A., W. W. Thatcher, M. h o s t and G. K. Haibel. 1987a. Influence of a physiological infusion of prostaglandin F b into postpartum cows with partially suppnsscd endogenous production of prostaglandins. 1. Uterine and ovarian morphological responses. Theriogenology 27:93 1.
Guilbault. L. A., W. W. Thatcher, D. 8. Foster and D. Caton. 1984. Relationship of 15-keto- 13.14-dihydro-Rosta- glandin F-2-a concentrations in peripheral plasma with local uterhe production of Fseries prostaglandins and changes in uterine blood flow during the early postpartum mod of cattle. Biol. Reprod. 31:870.
Guilbault. L. A., W. W. Thatcher and C. J. Wilcox. 1987b. Influence of a physiological infusion of prostaglandin F2a into postptum cows with partially suppressed endogenous production of prostaglandins. 2. lnterrela- tionships of hormmal, ovarian and uterine responses. Theriogtnology 2 7 9 7 .
Guilbcn. H. R. and A. McDonald. 1933. Weight records of purtbred beef cattle during growth, gestation, and lactatim. togethcr with data on reproduction. Roc.
Hammond, J. 1927. The pzlysiology of Reproduction in the
Hansen, P. J. 1985. Scasonal modulation of puberty and the
progesterrnt c~acenvati~n~ in p~st-partum dairy COWS
NO. 270. pp 23-26.
Am. SIX. of h i m . Rod. pp 244-253.
Cow. University Res, Cambridge.
ET AL.
postpartum anestrus in cattle: A review. Livest. Prod. Sci. 12309.
Haascn. P. J.. D. 8. Baik, J. J. Rutledge and E. R. Hauser. 1982. Genotype x environmental interactions on reproductive traiu of bovine females. 11. Postpartum reproduction as influenced by genotype, dietary regimen, level of milk production and parity. J. A n i . Sci. 55:1458.
Hansen. P. J. and E. R. Hauser. 1983. Genotype x environmental interactions on reproductive traits of bovine females. HI. Seasonal variation in postpartum reproduction as influenced by genotype, suckling and dietary regimen. J. Anim. Sci. 56:1362.
Hansen, P. J. and E. R. Hauser. 1984. Photoperiodic alteration of postpartum reproductive function in suckled cows. Theriogenology 22: 1.
Hanzen. Ch. 1986. Endocrine regulation of postpartum ovarian activity in cattle: a review. Reprod. Nutr. Dev. 26:1219.
Hauser. E. R. 1984. Seasonal effects on female reproduction in the bovine (Bus u r u s ) (European breeds). Therio- genology 21:150.
Hinshelwood, M. M., D. J. Dierschke and E. R. Hauser. 1985. Effect of suckling on the hypothalamic-pituitary axis in postpartum beef cows, independent of ovarian secretions. Biol. Reprod. 32:290.
Holness, D. H. and C. Jeffers. 1980. Oestrus and ovarian activity in lactating beef cows treated with 2-bromo-aergocryptine. South Afr. J. Anim. Sci. 10: 77.
Ingalls, W. G.. E. M. Convey and H. D. Hafs. 1973. Bovine serum LH. GH, and prolactin during late pregnancy, parturition and early lactation. Proc. Soc. Exp. Biol. Med. 143:161.
Inskeep.E. K., T. D. Braden, P. E. Lewis. M. Garcia-Winder and G. D. Niswender. 1988. Receptors for luteinizing hormone and follicle-stimulating hormone in largest follicles of postpartum beef cows. Biol. Reprod. 38: 587.
postpartum anestrus in beef cattle. In: Harold Hawk (Ed.) Beltsville Symposium on Animal Reproduction, No. 3, p 277. Allanheld. Osmun. Montclair.
Irvin, H. J., V. M. Pllantz. R. E. Morrow, B. N. Day and H. A. Garverick. 1981a. GnRH induced LH release in suckled beef cows. II. The effects of exogenous corticoids and estradiol benzoate on luteinizing hormone release by GnRH. Theriogenology 16513.
Irvin, H. J., A. A. Zaied. B. N. Day and H. A. Garverick. 1981b. GnRH induced LH release in suckled beef cows. I. The effects of days postpartum and estradiol- 178 concentrations on the release of LH following administration of GnRH. Theriogenology 15443.
Izaike, Y., A. Okano, K. Shimada and T. Oishi. 1984. Relationship between postpartum reproductive perfor- mance and parity in beef cows. Jpn. J. Anim. Reprod. 30206.
Jaeger, J. R., H. A. Turner and F. Stormshak. 1987. Gondatropin releasing hormone-induced secretion of luteinizing hormone during the milk-ejection reflex in the postpamUn beef cow. J. Anim. Sci. 65543.
Jagger, J. P., A. R. Peters and G. E. Lamming. 1987. Hormone responses to low-dose GNRH in postpartum beef cows. J. Reprod. Fertil. 80263.
Joubert. D. M. 1954. The influence of winter nutritional depressions on the growth, reproduction and produc- tion of cattle. J. A&. Sci. 445.
~nslrecp, E. K. and A. w. Listunan. 1979. Fwtors arrwting
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

POSTPARTUM ANESTRUS AND INFERTILITY 813
Kesler. D. J.. P. J. Weaon, C. A. Pimental, T. R. Toxler. D. L. Vincent and J. E. Hixon. 1981. Diminution of the in vitro response to luteinizing hormone by corpora lutea induced by gonadotropin releasing hormone treatment of postpartum suckled beef cows. J. Anim. Sci. 53:749.
Kilkmay, J. B. Reproductive performance of beef cows. 1978. Wald Rev. Anim. Prod. 1465.
King, G. J. and G. A. M a c W . 1983. Reproductive function in beef COWS calving in the spring or fall. Anim. Reprod. Sci. 6:255.
Kiracofe, G. H. 1980. Uterine involution: Its role in regulating postpartum intervals. J. Anim. Sci. Sl(Supp1. II):l6.
Koch, R. M. and J. W. Alga. 1983. The beef canle industry: changes and challenges. J. Anim. Sci. 57(Suppl. 2):28.
Kropp, J. R., D. F. Stephens, J. W. Holloway, J. V. Whiteman, L. KnOn: and R. Totusek. 1973. Perfor- mance on range and in drylot of two-year-old Hereford, Hereford x Holstein and Holstein females as influenced by level of winter supplementation. 1. Anim. Sci. 37:1222.
Labhsetwar. A. P., W. J. Tyler and L. E. Casida. 1963. Genetic and environmental factors affecting quiet owlations in Holstein cattle. J. Dairy Sci. 46:843.
Lamming. G. E. and B. J. MacLeod. 1988. CorUinuous W o n of GnRH reduces the LH response to an intravcnous GnRH injedon but does not inhibit endogenous LH secretion in cows. J. Reprod. Fenil. 82237.
Lamming. G. E., D. C. Wathes and A. R. Peters. 1981. Endocrine patterns of the postparnun cow. J. Reprod. Fertil. Suppl. 30155.
Lasley, J. F. and R. Bogart. 1943. Some factors influencing reproductive efficiency of range cattle under artificial and natural breeding conditions. Missouri Agric. Exp. Sta. Res. Bull. 376.
Laster, D. B.. H. A. Glirnp. L. V. Cundiff and K. E. Gregory. 1973. Factors affecting dystocia and the effects of dystocia on subsequent reproduction in beef cattle. J. Anim. Sci. 36:695.
Lauderdale, J. W., W. E. Graves, E. R. Hauser and L. E. Casida. 1%8. Relation ofpostpartum interval to corpus luteum development, pituitary prolactin activity and uterine involution in beef cows (effect of suckling and interval to breeding). Univ. of Wisconsin Res. Bull. No. 270, pp 4248. Madison.
Leung. K., V. Padmanabhan, L. J. Spicer, H. A. Tucker, R. E. Short and E. M. Convey. 1986. Relationdup between pituitary GnRH-binding sites and pituitary release of gonadotropins in post-partum beef cows. 1. Reprod. Fertil. 7653.
Lmdell, J.-O., H. Kindahl. L.-E. Edqvist and G. Tufvesson. 1982a. Effect of hysterectomy on the postpartum prostaglandin levels in the cow. Acta Vet. Scand. 23: 144.
Lindell, J.-0.. H. Kindahl, L. lansson and L.-E. Edqvist. 1982b. Post-partum release of prostaglandin F2a and uterine involution in the cow. Theriogenology 17:237.
Lishman. A. W., S.M.J. Allison, R. L. Fogwell, R. L. Butcher and E. K. lnskeep. 1979. Follicular develop- ment and function of induced corpora lutea in underfed postpartum anestrous beef cows. J. Anim. Sci. 48:867.
Lofstedt, R. M.. J. G. Manns. B. D. Murphy. W. D. Humphrey and R. J. Mapletoft. 1981. Influence of GnRH infusion on endocrine parameters and duration of postpartum anesms in beef cows. Theriogenology 15:359.
Louw. B. P., C. R. 'Ilomas and A. W. Lishman. 1988. The influence of loss and gain of body mass on ovarian activity in beef cows. South Afi. J. h i m . Sci. 18:l.
Lowman, B. G., N. Scott and S. Somerville. 1W3. Condition scoring of cattle. E. of Scotland Coll. of Agric. Bull. No. 6.
Madej, A., H. Kindahl. W. Woyno, L.-E. Edqvist and R. Stupnicki. 1984. Blood levels of 15-keto-13, 14 dihydro-prostaglandin F2.a during the postpartum period in primiparous cows. Theriogenology 21:279.
Malven, P. V., J. R. Parfet, D. W. Gregg, R. D. Allrich and 0. E. Moss. 1986. Relationships among concentrations of four opioid neuropeptides and luteinizing hormone- releasing hormone in neural tissues of beef cows following early weaning. J. Anim. Sci. 62:723.
Manns, J. G.. W. D. Humphrey, P. F. Flood, R. J. Mapletoft, N. Rawlings and K. W. Cheng. 1983. Endocrine profiles and functional characteristics of corpora lutea following onset of postpartum ovarian activity in beef cows. Can. J. Anim. Sci. 63:331.
Montgomery, G. W. 1982. Influence of suckling frequency and bromccriptine treatment on the resumption of ovarian cycles in post-partun beef cattle. Theriogenol- ogy 17:551.
Montgomery. G. W., I. C. Scott and N. Hudson. 1985. An interaction between season of calving and nutrition on the resumption of ovarian cycles in post-partum beef cattle. J. Reprod. Fenil. 73:45.
Morrow, D. A,. S. J. Roberts and K. McEntee. 1968. Latent effects of pregnancy on postpartum estrous cycle length in dairy cattle. J. Anim. Sci. 27:1404.
Moss, G. E., J. R. Parfet, C. A. Marvin, R. D. Allnch and M. A. Diekman. 1985. Pituitary concentrations of gonado- tropins and receptors for GnRH in suckled beef cows at various intervals after calving. J. Anim. Sci. 60285.
Nancarrow, C. D., H. M. Radford, R. J. Scaramuzzi and T. B. Post. 1977. Responses to injected oestrogen in suckled cows. Theriogenology 8:192 (Abstr.).
Nelsen, T. C., R. E. Short, D. A. Phelps and R. B. Staigmiller. 1985. Nonpuberal estrus and mature cow influences on growth and puberty in heifers. J. Ann. Sci. 61:470.
Nett, T. M. 1987. Function of the hypothalamic-hypophysial axis during the post-partum period in ewes and cows. J. Reprod. Fertil. (Suppl. 34):201.
Nett, T. M., D. Cermak, T. Braden, J. Manns and G. Niswender. 1988. Pituitary receptors for GnRH and estradiol, and pituitary content of gonadotropins in beef cows. 11. Changes during the postpartum period. Domest. Anim. Endocrinol. 5:81.
Odde, K. G. 1990. A review of synchronization of estrus in postpartum cattle. J. Anim. Sci. 68:817.
Odde, K. G.. H. S. Ward, G. H. Kiracofe. R. M. McKee and R. J. Kittok. 1980. Short estrous cycles and associated serum progesterone levels in beef cows. Theriogenol- ogy 14:105.
Oxenreider. S. L. 1968. Effects of suckling and ovarian function on postpartum reproductive activity in beef cows. Am. J. Vet. Res. 29:2099.
Peck, D. D.. F. N. Thompson, A. Jernigan and T. E. Kiser. 1988. Effect of morphine on serum gonadotropin concentrations in postpartum beef cows. I. Anim. Sci. 662930.
Perkins, J. L. and H. E. Kidder. 1963. Relation of utcriiie involution and postpartum interval to reproductive efficiency in beef cattle. J. Anim. Sci. 2 3 1 3 .
Peters, A. R. 1984a. Reproductive activity of the cow in the
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

8 14 SHORT
post-partum period. I. Factors affecting the length of the post-panum acyclic period. Br. Vet. J. 140:76.
Peters, A. R. 1984b. Effect of exogenous oestradiol-17b on goaadmophin secretion in post-partum beef cows. J. Reprod. Fertil. 72:473.
Peters, A. R.. M. G. Pimentel and G. E. Lamming. 1985. Hormone responses to exogenous GnRH pulses in post-partum dairy cows. J. Reprod. Fertil. 75557.
Peters, A. R. and G. M. Riley. 1982a. Is the cow a seasonal breeder? Br. Vet. J. 138533.
Peters, A. R. and G. M. Riley. 1982b. Milk progesterone profiles and factors affecting post partum ovarian activity in k e f cows. h i m . Prod. 34:145.
Pratt, B. R.. 1. G. Berarh l l i , L. P. Stevens and E. K. Inskeep. 1982. Induced corpora lutea in the postpartum beef cow. I. Comparison of gonadotropin releasing hormone and human chorionic gonadotropin and effects of progestogen and estrogen. J. An i . Sci. 54: 822.
Radford, H. M.. C. D. Nancmow and P. E. Maimer. 1978. Ovarian function in suckling and non-suckling beef cows post partum. J. Reprod. Fenil. 54:49.
Ramirez-Godinez, J. A., G. H. Kiracofe, D. L. Carnahan, M. F. Spire, K. B. Beeman, J. S. Stevenson and R. R. Schalles. 1982a. Evidence for d a t i o n and fertiliza- tion in beef cows with short estrous cycles. Theriogen- ology 17:409.
RamirezGodinez, J. A.,G. H. Kiracofe, R. M. McKee, R. R. Schalles and R. J. Kittok. 1981. Reducing the incidence of shon estrous cycles in beef cows with norgestomet. Theriogenology 15613.
Ramirez-Godinez, J. A., G. H. Kiracofe, R. R. Schalles and G. D. Niswender. 1982b. Endocrine patterns in the postpartum beef cow associated with weaning: A comparison of the shon and subsequent normal cycles. J. Anim. Sci. 59153.
Randel, R. D. 1990. Nutrition and postpartum rebreeding in cattle. J. Anim. Sci. 68:853.
Reynolds, W. L., T. M. DeRouen and R. A. Bellows. 1978. Relationships of milk yield of dam to early & r o d rate of straightbrad and crossbred calves. J. Anim. Sci. 47: 584.
Rice, F. J., R. R. Woodward, J. R. Quesenberry and F. S. Willson. 1961. Fertility of beef cattle raised under range conditions. Montana Agric. Exp. Sta. Bull. 561.
Richards, M. W., J. C. Spitzer and M. B. Warner. 1986. Effect of varying levels of postpartum nuuition and body condition at calving on subsequent reproductive performance in beef cattle. J. Anim. Sci. 62:300.
Riesen, J. W., S. Saiduddin, W. J. Tyler and L. E. Casida. 1%8. Relatioa of postpartum interval to corpus luteum development, pituitary prolactin activity, and uterine involution in dairy cows (effect of suckling). Univ. of Wisconsin Res. Bull. No. 270:2741.
Riley, G. M.. A. R. Peters and G. E. Lamming. 1981. Induction of pulsatile LH release, FSH release and ovulation in post-panum acyclic beef cows by repeated small doses of Gn-RH. J. Reprod. Fertil. 63:559.
Runer, L. M.. T. D. Carruthers and I. G. Manns. 1985. The postpartum induced corpus luteum: Functional differ- ences from that of cycling cows and the effects of progesterone pretreatment. Biol. Reprod. 33560.
Rutter, L. M. and R. D. Randel. 1984. Luteal competency during the resumption of ovarian cyclicity in postpar- tum Brahman cows. Tbcriogenology 21:713.
Runer, L. M. and R. D. Randcl. 1986. Nonpuberal estrus in k e f heifers. J. Amm. Sci. 63:1049. '
ET AL..
Saiduddin. S. , M. M. Quevedo and W. D. Foote. 1968a. Response of beef cows to exogenous progesterone and estradiol at various stages postpartum. J. A n i . Sci. 27: 1015.
Saiduddin. S. , J. W. Riesen, W. J. Tyler and L. E. Casida. 1%8b. Relation of postpartum interval to pituitary gonadotropins, ovarian follicular development and fertility in dauy cows. (Effect of suckling, feeding level, breeding management and genetic level of milk production). Univ. Wisconsin Res. Bull. No. 270:
Schallenberger, E. 1985. Gonadotrophins and ovarian steroids in cattle. 3. Pulsatile changes of gonadotrophin concentrations in the jugular vein postpartum. Acta Endocrinol. 109:37.
Schallenberger, E. and S. Prokopp. 1985. Gonadotrophins and ovarian steroids in cattle. IV. Re-establishment of the stimulatory feedback action of oestradiol-17p on LH and FSH. Acta Endocrinol. 1W44.
Schallenberger. E., D. Schams and K. Zottmeier. 1978. Response of lutropin (LH) and follitropin (FSH) to the adminiseation of gonadoliberin (GnRH) in pregnant and post-partum cattle including experiments with prolactin suppression. Theriogenology 10:35.
Schallenberger. E. and D. L. Walters. 1985. Endocrine mechanisms contributing to postpartum anoestrus in dairy and beef cattle. In: F. Ellendorff and F. Elsasser (Ed.) Current Topics in Vet. Med. and Anim. Sci. Vol. 31. The Hague: Martinus Nijhoff.
Scott, I. C. and G. W. Montgomery. 1987. Introduction of bulls induces return of cyclic ovarian function in post- partum beef cows. N.Z. J. Agric. Res. 30:189.
Sharpe, P. H., D. R. Gifford, P. F. Flavel, M. G. Nottle and D. T. Armstrong. 1986. Effect of melatonin on postpar- tum anesms in beef cows. Theriogenology 26621.
Sheffel, C. E., B. R. Pran, W. L. Ferrell and E. K. Inskeep. 1982. Induced corpora lutea in the postpartum beef cow. 11. Effects of treatment with progestogen and gonadotropins. J. Anim. Sci. 54:830.
Short, R. E. and D. C. Adams. 1988. Nutritional and hormonal interrelationshps in beef cattle reproduc- tion. Can. J. Anim. Sci. 68:29.
Short, R. E., R. A. Bellows, E. L. Moody and B. E. Howland. 1972. Effects of suckling and mastectomy on bovine postpartum reproduction. J. Anim. Sci. 34:70.
Short, R. E.. E. M. Convey, R. B. Staigmiller and R. A. Bellows. 1981. Effects of intermittent smalldose injections of GnRH in anestrous postpartum beef cows. J. Anim. Sci. 53 (Supp1.1):366 (Abstr.)
Short, R. E., A. R. Peters, M. P. Easdon and G. E. Lamming. 1986. The effect of an opiate antagonist on LH secretion i n k f cows. Paper presented at Annu. Conf., Soc. for Study of Fertil.. Bristol, UK. p 17.
Short, R. E., R. D. Randel and R. A. Bellows. 1974. Factors affecting reproduction in the postpartum cow. J. Anim. Sci. 39226 (Abstr.).
Short, R. E., R. D. Randel, R. B. Staigmiller and R. A. Bellows. 1979. Factors affecting estrogen-induced LH release in the cow. Biol. Reprod. 21:683.
Short. R. E., R. B. Staigmiller. J. K. Baber, J. B. C m and R. A. Bellows. 1976. Effects of mammary denervation in postpamun cows. 1. Anim. Sci. 43:304 (Abstr).
Smeaton, D. C., D. G. McCall and J. B. Clayton. 1986. Calving date effects on beef cow productivity. Proc. N.Z. Soc. Anim. Prod. 46:149.
Smilh, J. F.. E. Payne, H. R. Twi t . L. T. McCowan, R. Fairclough and R. Kilgour. 1981. The effect of
15-22.
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

POSTPARTUM ANESTRUS AND INFERTILITY 815
suckling upon the endocrine changes associated with anestrus in identical twin dairy cows. J. Reprod. Fertil.
Smith, M. F., H. A. Gmetick, R. S. Youngquist and W. L. Zahler. 1986. Luteinizing hormone receptor concen- trations, adenylate cyclase activity and phosphodies- (aase activity of bovi i corpora lutea: Comparison of short and normal estrous cycles. Domest. Anim. Mocrinol. 3:127.
Smith, V. G.. T. W. k k . E. M. Convey and H. A. Tucker. 1974. Bovine scrum prolactin. growth hormone, cortisol and milk yield after ergocripline. Neuroendo- crinology 15:172.
Smirh, V. G.. J. R. Chenault, J. C. McAllister and J. W. Lauderdale. 1987. Responsc of postpartum beef cows to exogenous progestogens and gonadotropin releasing hormone. J. Anim. Sci. 64540
Spicer, L. J.. E. M. Convey. K. hung, R. E. Short and H. A. Tucker. 1986a. Anovulation in poslpanum suckled
cows. 11. Associations among binding of &-labckd goaaQtropins to granulosa and thecal cells , md conccntntions of steroids in scm and various sizcd ovarian follicles. J. Anim. Sci. 62:742.
Spicer, L. J., E. M. Convey, H. A. Tucker and S. E. Echtemkamp. 1986b. Effects of in ermittent injections of L m cm specific biding of 1 h - l a b e w g o d o - vopins to granulosa and theca. and concentrations of staoids in serum and ovarian follicles during postpar- tum anovulation in suckled beef cows. J. h i m . Sci. 62: 1324.
Spicer, L. J.. K. hung, E. M. Convey, J. Gunther. R. E. Shon and H. A. Tucker. 1986c. Anovulation in postpartum suckled beef cows. 1. Associations among size and numben of ovarian follicles, uterine involu- tion, and hormones in serum and follicular fluid J. Anim. Sci. 62:734
Spicer, L. J. and S. A. Z~M. 1987. Relationship between concentrations of cortisol in ovarian follicular fluid and various biochemical markers of follicular differen- tiation in cyclic and anovulatory cattle. J. Reprod. Fertil. 81:221.
Stevenson, J. S., M. F. Spire and J. H. Britt. 1983. Influence of the ovary on estradiol-induced luteinizing hormone release in postpartum milked and suckled Holstein cows. J. Anim. Sci. 57:692.
Stewart, R. E. and J. S. Stevenson. 1987. Hormonal, estrual,
following multiple daily injections of oxytocin. J. Anim. Sci. 65:1585.
Symington, R. B. 1%9. Factors affecting postpartum fertility in cattle with special emphasis on the hannonal aspects of the problem in ranch cows in Soulhcm Africa. Roc. South Afr. Soc. Anim. Rod. 8: 29.
Terqui. M. 1985. Rcproductve potential during the post panum period in cows. In: F. Ellendorff and F. Elsasser (Ed.). Current Topics in Veterinary Medicine and Animal Science 31:199-205. The Hague, Maninus Nijhoff.
Tervit, H. R.. J. F. Smith and C. C. Kaltenbach. 1977. Post- partum anoestrus in bed cattle: A review. Roc. N.Z. Soc. Anim. Prod. 37:109.
Tolleson, D. R. and R. D. Randel. 1988. Effects of alfaprostol and uterine palpation on postpartum interval and pregnancy rate to embryo transfer in Brahman influenced k e f cows. Theriogenology 2 9 555.
Suppl. 30:241.
ovulatory and milk traits in postpartum dairy cows
Trout, W. E. and P. V. Malven. 1988. Quantification of naloxone binding sites in brains from suckled beef cows during postpartum anestrus and resumption of m o u s cycles. J. Anim. Sci. 66:954.
Troxel. T. R. and D. J. Kesler. 1984. The effect of progestin and GnRH treatments a ovarian function and reproductive hormone secretions of anestrous postpar- tum suckled beef cows. Theriogenology 21599.
Tucker, H. A. 1971. Hormonal response to milking. J. Anim. Sci. 32(Suppl. 1):137.
Wagner, W. C. and S. L. Oxenreider. 1971. Endocrine physiology following parturition. J. Anim. Sci. 32(Suppl. 1):l.
Walters. D. L.. C. C. Kaltenbach, T. G. Dunn and R. E. Short. 1982a. Pituitary and ovarian function in postpartum beef cows. I. Effect of suckling on serum and follicular fluid hormones and follicular gonadotropin receptors. Biol. Reprod. 26 :W.
Walters. D. L.. R. E. Short. E. M. Convey. R. B. Staigmiller, T. G. DUM and C. C. Kaltenbach. 1982b. Pituitary and ovarian function in postpartum beef cows. 11. Endo- crine changes prior to ovulation in suckled and non- suckled postpartum cows compared to cycling cows. Biol. Reprod. 26:647.
Walters, D. L., R. E. Short, E. M. Convey, R. B. Staigmiller, T. G. Dunn and C. C. Kaltenbach. 1982~. Pituitary and ovarian function in postpartum beef cows. 111. Induction of estrus, ovulation, and luteal function with intermittent snall-dose injections of GnRH. Biol. Reprod. 26555.
Wamick, A. C. 1955. Factors associated with h e interval from parturition to fust estrus in beef cattle. J. A n i . Sci. 14:1003.
Webb, R. and G. E. Lamming. 1981. Patterns of plasma prolactin in post-partum suckled cows. J. Endocrinol. 90:391.
Webb, R., G. E. Lamming. N. B. Haynes. H. D. Hafs and J. G. Manns. 1977. Response of cyclic and post-partum suckled cows to injections of synthetic LH-RH. J. Reprod. Fenil. 50:203.
Weesner, G. D., T. A. Noms, D. W. Forrest and P. G. Harms. 1987. Biological activity of luteinizing hormone in the peripartum cow: Least activity at parturition with an increase throughout the postpartum interval. Biol. Reprod. 37:85 1.
Weiss, T. I., C. D. Nancarrow. D. T. Armstrong and J. B. Donnelly. 1981. Modulation of functional capacity of small ovarian follicles in the post-partum cow by prolactin. Aust. J. Biol. Sci. 34:479.
Weuemann. R. P. 1980. Postpartum endocrine function of cattle, sheep and swine. J. Anim. Sci. Sl(Supp1. 2):2.
Wettemann. R. P., T. W. Beck, E. 1. Turman and R. L. Hintz. 1982. Endocrine response of postpartum anestrous beef cows to GnRH or PMSG. Theriogenology 18599.
Wheeler, M. B., G. B. Anderson. C. J. Munro and G. H. Stabenfeldt. 1982. Prolactin response in beef cows and heifers suckling one or two calves. J. Reprod. Fertil. 64243.
Whimant, C. S., T. E. Kiser and F. N. Thompson. 1985. Effect of calf removal on serum luteinizing hormone and cortisol concentrations in postpartum cows. Theriogenology 2 4 119.
Whimant. C. S..T. E. Kiser. F. N. Thompson and C. R. Barb. 1986a. Influence of calf removal on the serum luteinizing hormone response to naloxone in the
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

816 SHORT
postpartum beef cow. J. Anim. Sci. 63561. Whimant. C. S., T. E. Kiser. F. N. Thompson and C. R. Barb.
1986b. Naloxone infusioo increases pulsatile luteiniz- ing hormoncrclease in postpartum beef cows. Domest. Anim. Endocrinol. 3:49.
Whisurnt, C. S.. T. E. Kiser, F. N. Thompson and C. R. Barb. 1986c. Opioid inhibitioo of luteinizing hormone sccretioo during tbe postpartum period in suckled beef cows. J. Anim. Sci. 63:1445.
Whimat. C. S.. F. N. Thompson, T. E. Kiser and C. R. Barb. 1986d. Effect of naloxone on serum luteinizing hormone, ca i so l and prolactin concentrations in anestrous beef cows. J. Anim. Sci. 62:1340.
Wildcus, S.. R. D. Randel and W. D. Humphrey. 1987. laflueace of repeated low doses of gonadotropin releasing hormone on postpartum interval and serum luteinizing hormone in Brahman cross cows. Therio- genology 27:7 1 1.
Williams. G. L. 1990. Suckling as a regulator of postpartum rcbrceding in canle: A review. J. Anim. Sci. 68:831.
Williams. G. L., J. Kofwica, W. D. Slanger. D. K. Olson. J. E. Tilton and L. 1. Johnson 1982a. Effect of suckling on pituitary rcspcmaveness to gonadotropin-releasing hormcme throughout tbe early postpartum period of beef cows. J. Anim. Sci. 54594.
Williams. G. L., B. J. Petersen and J. E. Tilton. 1982b. Pituitary and ovarian rtsponses of postpartum dairy cows to progesterone priming and single or double
ET fu.
injections of gonadotropin-releasing hormone. Therio- genology 18561.
Williams, G. L. and D. E. Ray. 1980. Hormonal and reproductive profiles of early postpartum beef heifers after prolactin suppression or steroid-induced luteal function. J. Anim. Sci. 50:906.
Wiltbank, J. N. 1970. Research needs in k f cattlc reproduction. J. Anim. Sci. 31:755.
Wiltbank, J. N. and A. C. Cook. 1958. The comparative reproductive performance of nursed cows and milked cows. J. h i m . Sci. 17540.
Wiltbank, J. N.. W. W. Rowden. J. E. Ingalls. K. E. Gregory and R. M. Koch. 1962. Effect of energy level on reproductive phenomena of mature Hereford cows. J. Anim. Sci. 21:219.
Wiltbank, J. N., W. W. Rowden, J. E. Ingalls and D. R. Zimmerman. 1963. Muence of post-partum energy level on reproductive performance of Hereford cows restricted in energy intake prior to calving. J. Anim. Sci. 23:1049.
Wiltbank, J. N.. E. J. Warwick, E. H. Vernon and B. M. Riode. 1%1. Factors affecting net calf crop in beef cattle. J. Anim. Sci. 20:409.
Zalesky, D. D., M. L. Day, M. Garcia-Winder, K. Imakawa. R. J. Kittok. M. J. D'Occhio and J. E. Kinder. 1984. Influence of exposure to bulls on resumption of estrous cycles following panurition in beef cows. J. h i m . Sci. 5 9 1135.
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from

Citationshttp://www.journalofanimalscience.org/content/68/3/799#otherarticlesThis article has been cited by 42 HighWire-hosted articles:
by guest on September 24, 2014www.journalofanimalscience.orgDownloaded from