
Physical mapping of 18S rDNA and heterochromatin in ... · Genetics and Molecular Research 13 (1):...
14
©FUNPEC-RP www.funpecrp.com.br Genetics and Molecular Research 13 (1): 2186-2199 (2014) Physical mapping of 18S rDNA and heterochromatin in species of family Lygaeidae (Hemiptera: Heteroptera) V.B. Bardella 1,2 , T.R. Sampaio 1 , N.B. Venturelli 1 , A.L. Dias 1 , L. Giuliano-Caetano 1 , J.A.M. Fernandes 3 and R. da Rosa 1 1 Laboratório de Citogenética Animal, Departamento de Biologia Geral, Centro de Ciências Biológicas, Universidade Estadual de Londrina, Londrina, PR, Brasil 2 Instituto de Biociências, Letras e Ciências Exatas, Departamento de Biologia, Universidade Estadual Paulista, São José do Rio Preto, SP, Brasil 3 Instituto de Ciências Biológicas, Universidade Federal do Pará, Belém, PA, Brasil Corresponding author: R. da Rosa E-mail: [email protected] Genet. Mol. Res. 13 (1): 2186-2199 (2014) Received June 17, 2013 Accepted December 5, 2013 Published March 26, 2014 DOI http://dx.doi.org/10.4238/2014.March.26.7 ABSTRACT. Analyses conducted using repetitive DNAs have contributed to better understanding the chromosome structure and evolution of several species of insects. There are few data on the organization, localization, and evolutionary behavior of repetitive DNA in the family Lygaeidae, especially in Brazilian species. To elucidate the physical mapping and evolutionary events that involve these sequences, we cytogenetically analyzed three species of Lygaeidae and found 2n (♂) = 18 (16 + XY) for Oncopeltus femoralis; 2n (♂) = 14 (12 + XY) for Ochrimnus sagax; and 2n (♂) = 12 (10 + XY) for Lygaeus peruvianus. Each species showed different quantities of heterochromatin, which also showed variation in their molecular composition by fluorochrome
Transcript of Physical mapping of 18S rDNA and heterochromatin in ... · Genetics and Molecular Research 13 (1):...

©FUNPEC-RP www.funpecrp.com.brGenetics and Molecular Research 13 (1): 2186-2199 (2014)
Physical mapping of 18S rDNA and heterochromatin in species of family Lygaeidae (Hemiptera: Heteroptera)
V.B. Bardella1,2, T.R. Sampaio1, N.B. Venturelli1, A.L. Dias1, L. Giuliano-Caetano1, J.A.M. Fernandes3 and R. da Rosa1
1Laboratório de Citogenética Animal, Departamento de Biologia Geral, Centro de Ciências Biológicas, Universidade Estadual de Londrina, Londrina, PR, Brasil2Instituto de Biociências, Letras e Ciências Exatas, Departamento de Biologia, Universidade Estadual Paulista, São José do Rio Preto, SP, Brasil3Instituto de Ciências Biológicas, Universidade Federal do Pará, Belém, PA, Brasil
Corresponding author: R. da RosaE-mail: [email protected]
Genet. Mol. Res. 13 (1): 2186-2199 (2014)Received June 17, 2013Accepted December 5, 2013Published March 26, 2014DOI http://dx.doi.org/10.4238/2014.March.26.7
ABSTRACT. Analyses conducted using repetitive DNAs have contributed to better understanding the chromosome structure and evolution of several species of insects. There are few data on the organization, localization, and evolutionary behavior of repetitive DNA in the family Lygaeidae, especially in Brazilian species. To elucidate the physical mapping and evolutionary events that involve these sequences, we cytogenetically analyzed three species of Lygaeidae and found 2n (♂) = 18 (16 + XY) for Oncopeltus femoralis; 2n (♂) = 14 (12 + XY) for Ochrimnus sagax; and 2n (♂) = 12 (10 + XY) for Lygaeus peruvianus. Each species showed different quantities of heterochromatin, which also showed variation in their molecular composition by fluorochrome

2187
©FUNPEC-RP www.funpecrp.com.brGenetics and Molecular Research 13 (1): 2186-2199 (2014)
Physical mapping in Lygaeidae
staining. Amplification of the 18S rDNA generated a fragment of approximately 787 bp. The alignment of the consensus sequence with sequences from other species of Heteroptera deposited in the GenBank revealed a similarity of 98% with small differences. Fluorescent in situ hybridization with the 18S rDNA fragment revealed that this ribosomal gene was located in 1 autosomal pair at different positions in the three species. No cytogenetic data are available for these Brazilian species. The basal number and the possible chromosomal changes that occurred among the different species, as well as the evolution of these DNA sequences, are discussed.
Key words: Chromosome structure; Fluorescent in situ hybridization; Heterochromatin; Ribosomal genes
INTRODUCTION
The ribosomal DNA has been used in many molecular analyses in insects, especially those involving clarifications about phylogenetic relationships. In Hemiptera, these studies have revealed important findings compared to those obtained in other groups of insects (Caterino et al., 2000). Besides 18S rDNA, other classes of repetitive DNA with heterochromatic sequences have also been widely used as cytogenetic markers for comparisons across different groups of insects such as in Orthoptera (Cabral-de-Mello et al., 2011), Coleoptera (Almeida et al., 2010), Diptera (Rafael et al., 2006), and Heteroptera (Papeschi and Bressa, 2006; Bardella et al., 2010; Grozeva et al., 2010; Panzera et al., 2012). Some types of repetitive DNA can be chromosome-specific, such as those mostly found in Drosophila melanogaster (Bonaccorsi and Lohe, 1991) and those found on the sex chromosomes in species such as Megoura (Bizzaro et al., 1996).
Few studies have investigated the identification and location of repetitive DNA, par-ticularly in Heteroptera. These studies revealed the distribution of 18S rDNA sites in auto-somal chromosomes, such as those observed in Pachylis argentinus Berg, 1879, and Nezara viridula Linnaeus, 1758 (Papeschi et al., 2003), as well as in sex chromosomes such as those of the families Belostomatidae (Papeschi and Bressa, 2006), Cimicidae (Grozeva et al., 2010), and Reduviidae (Bardella et al., 2010). Although studies investigating the location of het-erochromatin are more common in Heteroptera, most of them are restricted to conventional analyses such as C-banding technique (Papeschi and Bressa, 2006). Studies involving the mo-lecular characterization of these repetitive DNAs in insects are scarce, compared to the great diversity of the group.
In Lygaeidae, cytogenetic studies shown a wide karyotypic diversity; however, most of them are restricted to conventional analysis. Thus far, chromosomal data are available for 13 subfamilies (Ueshima, 1979; Ueshima and Ashlock, 1980; Grozeva and Kuznetsova, 1993; Kaur et al., 2010), which suggest the existence of a chromosomal variation from 2n = 11 in Cryphula nitens Barber 1955 (Ueshima, 1979) to 2n = 30 in different species of the subfamily Cyminae. In addition, different sex chromosome systems, as well as the presence/absence of m-chromosomes, were observed (Grozeva and Kuznetsova, 1993).
Although the karyotype variability within the family Lygaeidae has been well docu-mented, the distribution of rDNA and different classes of heterochromatin and the evolution-

2188
©FUNPEC-RP www.funpecrp.com.brGenetics and Molecular Research 13 (1): 2186-2199 (2014)
V.B. Bardella et al.
ary events that occurred in this group are not well elucidated. Thus, in this study, we performed both conventional and molecular cytogenetic analyses of three species of the family Lygaeidae and determined the possible events that led to karyotype differentiation of this interesting group of insects.
MATERIAL AND METHODS
Samples and collection sites
Three species of family Lygaeidae [Oncopeltus femoralis Stål, 1874 (Figure 1a), Ochrimnus sagax Brailovsky, 1982 (Figure 1b), and Lygaeus peruvianus Brailovsky, 1978 (Figure 1c)] were collected from southern and southeastern Brazil (Table 1). Individuals from each species were identified and deposited at Universidade Federal do Pará (UFPA).
Figure 1. Lygaeidae species from Brazil southeast: a. Oncopeltus femoralis; b. Lygaeus peruvianus; and c. Ochrimnus sagax. Bar = 0,5 cm.
Specie Male Female Locality (City/State) Geographic coordinates
Oncopeltus femoralis 17 12 Londrina/Paraná 23°21'47.06''S and 51°06'28.95''WOchrimnus sagax 4 8 Assis/São Paulo 22°28'39.80''S and 50°20'55.73''WLygaeus peruvianus 6 6 Londrina/Paraná 23°21'47.06''S and 51°06'28.95''W
Table 1. Collection sites of Lygaeidae species.
Chromosome preparations and conventional staining
The gonads of adult specimens were dissected in physiological solution for insects (7.5 g NaCl, 2.38 g Na2HPO4, and 2.72 g KH2PO4 in 1 L distilled water). The testicles were treated with a hypotonic solution (tap water) for 3 min and fixed in methanol/acetic acid (3:1) for 30 min. Chromosome preparations were obtained by generating cellular suspension by maceration in 1 drop of 60% acetic acid; each gonad was previously treated with 45% acetic acid. These preparations were submitted to conventional staining with 3% Giemsa and chro-mosome banding techniques. Chromosome measurements were performed using the com-puter application MicroMeasure version 3.2 (Reeves and Tear, 2000).

2189
©FUNPEC-RP www.funpecrp.com.brGenetics and Molecular Research 13 (1): 2186-2199 (2014)
Physical mapping in Lygaeidae
Chromosome banding
The distribution of heterochromatin was analyzed by Giemsa C-banding after treatment with 0.2 M HCl, Ba(OH)2, and 2X saline sodium citrate (SSC; Sumner, 1972). The GC- and AT-rich bands were detected using chromomycin A3 (CMA3) and 4'-6-diamino-2-phenylindole (DAPI), respectively. The slides were stained with 0.5 mg/mL CMA3 for 1.5 h, washed in distilled water, and sequentially stained with 2 µg/mL DAPI for 30 min. Slides were mounted with a medium composed of glycerol/McIlvaine’s buffer (pH 7.0; 1:1), plus 2.5 mM MgCl2.
DNA isolation, polymerase chain reaction amplification, and genomic sequencing
In addition to karyotype studies, genomic DNA from a male of Oncopeltus femoralis was obtained from the muscle tissue sample. After the sample was digested with proteinase K for 3 h, phenol/Tris-HCl, pH 8.0 was added, followed by centrifugation and washing with phenol/Tris-HCl, pH 8.0 and chloroform-isoamyl alcohol. After an additional centrifugation, chloroform-isoamyl alcohol was added, and the DNA was precipitated with absolute and cold ethanol for 12 h at -20°C and eluted in Tris-EDTA (TE; 1:10) + RNAse.
Unlabeled 18S rDNA probes were generated by polymerase chain reaction (PCR) by using universal arthropod primers: forward 5'-CCTGAGAAACGGCTACCACATC-3' and re-verse 5'-GAGTCTCGTTCGTTATCGGA-3' (Whiting, 2002). The fragment obtained was puri-fied and sequenced using an automatic sequencer (Applied Biosystems® 3500xL Genetic Ana-lyzers) equipped with 50 cm capillaries and POP6 polymer (Applied Biosystems®). The DNA sequence obtained was submitted to nucleotide BLAST (Zhang et al., 2000), in which homolo-gous sequences of 18S rDNA from other species of Heteroptera were found, which, together with the sequence obtained from Oncopeltus femoralis, were submitted to nucleotide alignment by using MEGA version 5 (Tamura et al., 2011) and Multalin (Corpet, 1998) programs. Se-quence data were deposited in the GenBank sequence database under accession No. JX110161.
Fluorescence in situ hybridization
The 18S rDNA probe isolated using PCR was labeled with biotin-14-dATP by PCR and nick translation. Slides were dehydrated in alcohol series and washed in 15% formamide/0.2X SSC, pre-treated with DNAse-free RNAse (40 µg/mL in 2X SSC) at 37°C for 1 h and pepsin (0.005% in 10 mM HCl) at 37°C for 30 min, fixed with 4% fresh paraformaldehyde, dehydrated in an alcohol series, and air dried. The chromosomes were then denatured in 70% formamide/2X SSC at 70°C for 5 min. Subsequently, the slides were treated with 30 µL hybridization mixture containing 100 ng labeled probe (4 µL), 50% formamide (15 µL), 50% polyethylene glycol (6 µL), 20X SSC (3 µL), 100 ng calf thymus DNA (1 µL), and 10% SDS (1 µL). The material was denatured at 90°C for 10 min, and hybridization was performed overnight at 37°C in a humidified chamber. Post-hybridization washes were carried out at 42°C in 2X SSC, 20% formamide in 0.1X SSC, 0.1X SSC, and 4X SSC/0.2% Tween 20. The probe was detected using a solution of 5% bovine serum albumin (BSA) and fluorescein isothiocyanate (FITC)-conjugated avidin (50:0.5, v:v). The post-detection washes were performed in 4X SSC/0.2% Tween 20 at room temperature. Slides were mounted with 25 µL of a medium composed of 23 L 1,4-diazabicyclo[2.2.2]-octane (DABCO) solution [(2.3%), 20 mM Tris-HCl, pH 8.0, (2%), and glycerol (90%) in distilled water], 1 µL 2 µg/mL DAPI, and 1 µL 50 mM MgCl2.

2190
©FUNPEC-RP www.funpecrp.com.brGenetics and Molecular Research 13 (1): 2186-2199 (2014)
V.B. Bardella et al.
All the images were acquired using a Leica DM 4500 B microscope equipped with a DFC 300FX camera and Leica IM50 4.0 software and optimized for best contrast and bright-ness by using the iGrafx Image software.
RESULTS
Chromosome arrangements in the insect species
Oncopeltus femoralis
The chromosome number for all samples of Oncopeltus femoralis was 2n (♂) = 16 + XY (Figure 2). All specimens showed holokinetic chromosomes with homogeneous size among all chromosomes (Figure 2). It was possible to observe in diplotene/diakinesis the occurrence of up to 2 chiasmata in the longest bivalent (Figure 2a-c). The results of meio-cyte analyses in metaphase I showed a small difference in size between chromosomes X and Y (Figure 2d-f) and, at this stage, all cells showed the bivalents arranged in a ring and XY chromosomes positioned within the ring. Furthermore, the bivalents were confirmed to divide reductionally, whereas sex chromosomes divided equationally.
Figure 2. Meiotic and mitotic stages in males of Oncopeltus femoralis (2n = 16+XY) with Giemsa (a. d. g.), DAPI (b. e. h.), and CMA3 (c. f. i.) staining. a. b. c. Diplotene; d. e. f. metaphase I; g. h. i. gonial mitotic metaphase. Arrows show the GC-rich blocks in one autosomal pair. Arrowheads show the sex chromosomes. Bars = 5 µm.

2191
©FUNPEC-RP www.funpecrp.com.brGenetics and Molecular Research 13 (1): 2186-2199 (2014)
Physical mapping in Lygaeidae
C/CMA3/DAPI-banding showed 2 heterochromatic blocks, which were heteromor-phic (Figure 2i). The distribution and heteromorphism of the heterochromatic blocks were confirmed in diplotene/diakinesis (Figure 2b and c), and their location in an autosomal biva-lent was confirmed in metaphase I (Figure 2f). The analysis of spermatogonial cells in mitotic metaphase showed interstitial distribution of the heterochromatic block and CMA3
+/DAPI- block in a large-sized chromosome pair and confirmed the telokinetic pairing (Figure 2f).
Ochrimnus sagax
Ochrimnus sagax showed 2n (♂) = 12 + XY (Figure 3), with a karyotype consisting of 6 large, 7 medium, and 1 small chromosome (Y), all holokinetic. It was possible to observe in diplotene/diakinesis the occurrence of 1 or 2 terminal chiasmata per bivalent; the sex chromosomes were achiasmatic and behaved as univalent (Figure 3a). Observing the pairing of autosomal chromosomes around chromosomes X and Y was possible in metaphase II (Figure 3d-f). The 2 longest autosomal chromosomes showed a nucleolar constriction in mitotic metaphase (Figure 3g-i).
Figure 3. Male meiosis and mitosis in Ochrimnus sagax with Giemsa (a. d. g.), DAPI (b. e. h.), and CMA3 (c. f. i.) staining. a. b. c. Diplotene; d. e. f. metaphase II; g. h. i. mitotic metaphase. Arrowheads show the sex chromosomes. Arrows indicate the CMA3
+/DAPI- blocks in the nucleolar constriction. Note the presence of these sites in an autosome pair and in the Y chromosome. Bars = 5 µm.
The heterochromatic CMA3+/DAPI- blocks were evidenced in the larger autosomal
pair, coincident with nucleolar constriction. Besides this block, the Y chromosome showed an interstitial CMA3
+/DAPI- block (Figure 3i). The location of these blocks could also be con-

2192
©FUNPEC-RP www.funpecrp.com.brGenetics and Molecular Research 13 (1): 2186-2199 (2014)
V.B. Bardella et al.
firmed by the analyses performed on the meiotic chromosomes in diplotene/diakinesis (Figure 3b and c) and metaphase II (Figure 3h and i).
Lygaeus peruvianus
All samples of L. peruvianus showed 2n (♂) = 10 + XY, with a karyotype consisting of 2 large, 9 medium, and 1 small chromosome, all holokinetic (Figure 4a-c). Analyses of mei-ocytes in metaphase I and II showed the arrangement in which the autosomal chromosomes formed a ring around the sex chromosomes. Chromosome pairings revealed the occurrence of the XY sex chromosome system, and the analysis of cells in mitosis enabled the determination of the difference in size between them (Figure 4d-i).
Figure 4. Meiotic and mitotic cells from Lygaeus peruvianus with Giemsa (a. d. g.), DAPI (b. e. h.), and CMA3 (c. f. i.) staining. a. b. c. Diplotene; d. e. f. metaphase II; g. h. i. mitotic metaphase. Arrowheads show the Y chromosome. Arrows indicate the CMA3
+/DAPI- blocks. Note the presence of CMA30/DAPI+ terminal sites in all
autosome pairs and in the Y chromosome. Bars = 5 µm.
C-banding and CMA3/DAPI staining revealed the localization of GC- and AT-rich blocks. In diplotene/diakinesis, a wide distribution of AT-rich terminal blocks in most biva-lents were possible; however, in metaphase II, only the AT-rich block in Y chromosome was observed, probably due to the condensation of autosomes. It was possible to observe in a bivalent a GC-rich block in synteny with an AT-rich region (Figure 4a-f). The distribution of AT-rich blocks was also observed in mitotic cells, where the largest autosomal pair showed a DAPI+ marking placed in the terminal position (Figure 4h). In this stage, it was observed the

2193
©FUNPEC-RP www.funpecrp.com.brGenetics and Molecular Research 13 (1): 2186-2199 (2014)
Physical mapping in Lygaeidae
autosomal pair bearer of GC/AT-rich syntenic blocks. In addition, 7 chromosomes showed DAPI+ blocks in both terminal regions, and 3 chromosomes, including the X chromosomes, showed this sequence only in 1 terminal region. The Y chromosome showed DAPI+ marking throughout both in mitotic cells and in metaphase II (Figure 4h and i).
Sequencing and fluorescent in situ hybridization
Amplification of the 18S rDNA region of Oncopeltus femoralis with the specific primers generated a fragment of approximately 787 bp. The 719-bp consensus sequence of Oncopeltus femoralis generated through the alignment of the sequences consisted of 23.6% A, 23.9% C, 24.8% T, and 27.7% G bases (Figure 5a). The alignment of the consensus sequence with the 18S sequences from other species of Heteroptera deposited in the GenBank (Figure 5b) revealed a maximum similarity of 98% (Table 2) with small differences.
Figure 5. a. Partial sequence of 18S rDNA gene of Oncopeltus femoralis (GenBank accession No. JX110161); b. alignment of partial sequence of 18S gene of O. femoralis, with 18S sequences from other species of Heteroptera obtained from the GenBank (Graptostethus quadrisignatus accession AY627325, Spilostethus hospes accession AY627319, Notobitus meleagris accession AY627321, Leptocorisa sp accession AY627320, Pachygrontha antennata accession AY324852) showing highly conserved regions among the sequences analyzed. Dots indicate sequence identity and letters denote absence of homology.
a
b
2194
©FUNPEC-RP www.funpecrp.com.brGenetics and Molecular Research 13 (1): 2186-2199 (2014)
V.B. Bardella et al.
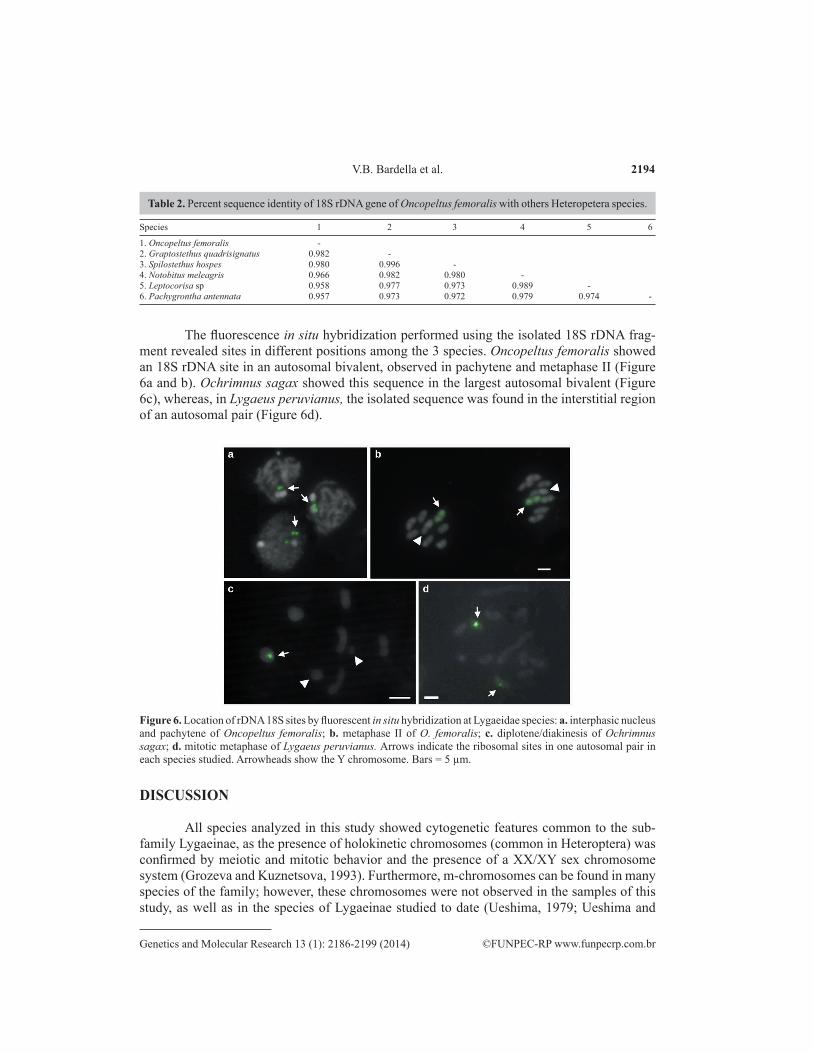
The fluorescence in situ hybridization performed using the isolated 18S rDNA frag-ment revealed sites in different positions among the 3 species. Oncopeltus femoralis showed an 18S rDNA site in an autosomal bivalent, observed in pachytene and metaphase II (Figure 6a and b). Ochrimnus sagax showed this sequence in the largest autosomal bivalent (Figure 6c), whereas, in Lygaeus peruvianus, the isolated sequence was found in the interstitial region of an autosomal pair (Figure 6d).
Species 1 2 3 4 5 6
1. Oncopeltus femoralis - 2. Graptostethus quadrisignatus 0.982 - 3. Spilostethus hospes 0.980 0.996 - 4. Notobitus meleagris 0.966 0.982 0.980 - 5. Leptocorisa sp 0.958 0.977 0.973 0.989 - 6. Pachygrontha antennata 0.957 0.973 0.972 0.979 0.974 -
Table 2. Percent sequence identity of 18S rDNA gene of Oncopeltus femoralis with others Heteropetera species.
Figure 6. Location of rDNA 18S sites by fluorescent in situ hybridization at Lygaeidae species: a. interphasic nucleus and pachytene of Oncopeltus femoralis; b. metaphase II of O. femoralis; c. diplotene/diakinesis of Ochrimnus sagax; d. mitotic metaphase of Lygaeus peruvianus. Arrows indicate the ribosomal sites in one autosomal pair in each species studied. Arrowheads show the Y chromosome. Bars = 5 µm.
DISCUSSION
All species analyzed in this study showed cytogenetic features common to the sub-family Lygaeinae, as the presence of holokinetic chromosomes (common in Heteroptera) was confirmed by meiotic and mitotic behavior and the presence of a XX/XY sex chromosome system (Grozeva and Kuznetsova, 1993). Furthermore, m-chromosomes can be found in many species of the family; however, these chromosomes were not observed in the samples of this study, as well as in the species of Lygaeinae studied to date (Ueshima, 1979; Ueshima and

2195
©FUNPEC-RP www.funpecrp.com.brGenetics and Molecular Research 13 (1): 2186-2199 (2014)
Physical mapping in Lygaeidae
Ashlock, 1980; Grozeva and Kuznetsova, 1993).In the specimens of the family Lygaeidae analyzed herein, a change in the diploid
number was observed, which ranged from 2n (♂) = 18 in Oncopeltus femoralis to 2n (♂) = 14 in Ochrimnus sagax, while Lygaeus peruvianus showed 2n (♂) = 12. Besides the variation in the diploid number, in O. femoralis and O. sagax, chromosome numbers decreased homo-geneously in size, i.e., there was not a great difference in size between the chromosome pairs (Figures 2 and 3). The same was not true for L. peruvianus, where the first chromosome pair had a different size compared to that of the other chromosomes (Figure 4). Another interesting finding was the difference in size of sex chromosomes in the 3 species. The X and Y chromo-somes in O. femoralis are of similar size, while, in O. sagax, the X chromosome is consider-ably longer than the Y chromosome, as observed in L. peruvianus. This increase/reduction in diploid number as well as the difference in size between chromosomes might be related to agmatoploidy/simploidy events, previously found in other groups of Heteroptera (Bressa et al., 2009; Bardella et al., 2010). This variation in the diploid number between specimens was also found in different species of the family Lygaeidae. The greatest variation in diploid number occurred in the subfamilies Rhyparochrominae (2n = 11 to 20) and Cyminae (2n = 16 to 30), although there are reports of such events in most of the other subfamilies (Ueshima and Ashlock, 1980; Grozeva and Kuznetsova, 1993).
With regard to heterochromatin, the distribution in heteropterans ranges from absence to presence of terminal heterochromatic blocks in most chromosomes, which, in some species, were characterized as AT-rich (Bressa et al., 2005; Papeschi and Bressa, 2007). In many cases, only 1 GC-rich heterochromatic block (interstitial or terminal) is recognized, which is often associated with the nucleolus organizer region (NOR; Camacho et al., 1985; Papeschi et al., 2003; Grozeva et al., 2004). Reports of interstitial heterochromatic blocks scattered through-out most autosomes are scarce and restricted to the family Curiae (Holhymenia rubiginosa and Spartocera batatas) (Fabricius, 1798) (Franco et al., 2006; Bressa et al., 2008). In the family Lygaeidae, studies on the heterochromatic location are limited to the genus Dieuches (Eyles, 1973), in which the 3 species analyzed showed AT-rich heterochromatic blocks already described in Heteroptera (Kaur et al., 2010).
Among all species studied, Oncopeltus femoralis had the lowest amount of hetero-chromatin on only 1 chromosome pair (the largest of the complement) with interstitial CMA3
+/DAPI-, which also showed size heteromorphism (Figures 2i and 7). This heteromorphism has already been reported in other species of Heteroptera, including Holhymenia rubiginosa (Bressa et al., 2008). In Ochrimnus sagax, although the CMA3
+/DAPI- blocks have also been observed on the first pair of autosomal chromosomes, they were different from those found in Oncopeltus femoralis, since their positioning was coincident with the nucleolar constriction and in a region closer to the telomere. Furthermore, this species showed an interstitial CMA3
+/DAPI- block in the Y chromosome (Figures 3i and 7).
Even though both species showed differences in the location of GC-rich repetitive DNA, they did not show any AT-rich blocks. In contrast, in L. peruvianus, although 2 autoso-mal chromosomes showed interstitial blocks (Figures 4i and 7), large DAPI+ heterochromatic terminal blocks were observed on all autosomes and on the X chromosome, while the Y chro-mosome appeared entirely DAPI+ (Figures 4h and 7). The large amount of DAPI+ heterochro-matin in L. peruvianus can be explained by the equilocal distribution (Heitz, 1957).

2196
©FUNPEC-RP www.funpecrp.com.brGenetics and Molecular Research 13 (1): 2186-2199 (2014)
V.B. Bardella et al.
The isolation, characterization, and localization of 18S rDNA sequences also proved to be interesting in the cytogenetic differentiation of species of Lygaeidae. Comparison of sequences from the GenBank by using the BLAST tool revealed that the 18S rDNA sequence obtained showed a similarity of 98% to those of other species of the family (Table 2). Small differences could be observed through the alignments, but this similarity confirms the nature of the isolated sequence. Almeida et al. (2010) analyzed the 18S rDNA isolated sequence of Omophoita octoguttata (Fabricius, 1875) (Coleoptera) and also found these differences in these kinds of repetitive DNA. Those authors attributed such differences to the result of transi-tion and transversion base changes. Analysis of the alignments of the 18S rDNA isolated in this study suggested that both base changes occurred with the same frequency. Thus, these changes could maintain the homogeneity of DNA, thereby preserving the structure and func-tion of the ribosome. However, the molecular process of concerted evolution might also be responsible for the changes in this sequence, suggesting that this family of repetitive DNA did not evolve independently; this has been proposed for the evolution of ribosomal genes in eukaryotic organisms (Elder and Turner, 1995; Liao, 1999).
The fluorescent in situ hybridization with the isolated 18S rDNA showed different
Figure 7. Idiograms and chromosomal physical mapping of Lygaeidae species studied.

2197
©FUNPEC-RP www.funpecrp.com.brGenetics and Molecular Research 13 (1): 2186-2199 (2014)
Physical mapping in Lygaeidae
location patterns among species of Lygaeidae. Oncopeltus femoralis showed an 18S rDNA interstitial site in an autosomal bivalent; Ochrimnus sagax showed this sequence in the largest autosomal bivalent in the interstitial terminal region, whereas, in Lygaeus peruvianus, the rDNA cistrons were found in the interstitial region of an autosomal pair. In Heteroptera, studies focusing on the identification and localization of 18S rDNA on chromosomes are restricted to a few species of some families and the results have shown that the 18S rRNA gene might be located in both the autosomes and the sex chromosomes (Papeschi et al., 2003; Bardella et al., 2010; Grozeva et al., 2010; Poggio et al., 2011; Panzera et al., 2012). However, most of the studies revealed that these cistrons at terminal position of the chromosomes were different from those found in the species analyzed in this study.
The isolated 18S rDNA sequence, along with the localization and identification of the base composition of heterochromatic blocks, constituted an important tool for the physical mapping of such chromosomes and differentiation among species. In many organisms, the GC-rich sites have been associated with the location of NORs (da Rosa et al., 2009; Bardella et al., 2010). In the 3 species of Lygaeidae, the 18S rDNA site varied with regard to the chro-mosomal location and was co-located with the CMA3
+ heterochromatic blocks in all the 3 spe-cies. Moreover, the hybridization of the probe revealed that the CMA3
+ block present on the Y chromosome of Ochrimnus sagax did not show any 18S rDNA sequences, and that this block corresponded to another type of GC-rich heterochromatin (Figure 7).
Our data showed a great diversity in the distribution of heterochromatic blocks and 18S rDNA in the individuals of the family Lygaeidae; similar findings have been reported in Coreidae (Bressa et al., 2005) and Triatiominae (Bardella et al., 2010). The differences in the sequence of 18S rDNA can be associated with some transitions/transversions of bases among other species of Heteroptera, and events of concerted evolution might have been involved in the evolution of this sequence. Considering the proposition made by Ueshima (1979) that the karyotype 2n = 14 (12 + XY) is the most common for the subfamily Lygaeinae, we can suggest that Ochrimnus sagax presents a plesiomorphic karyotype similar to that found in Oncopeltus femoralis and L. peruvianus; the latter would be species with derived karyotypes resulting from simploidy and agmatoploidy events, respectively. Due to the lack of chromosomal and phy-logenetic studies in the subfamily Lygaeinae and the little knowledge of the biogeographical distribution of these species, identifying the ancestral karyotype of this group is not possible.
ACKNOWLEDGMENTS
The authors are very grateful to Anna Maria Santos Silva and Paulo Roberto Maxi-miano da Silva for sample collection and to the Brazilian agencies Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Fundação Araucária and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for their financial support. The researchers received permission from Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA) to collect insect specimens.
REFERENCES
Almeida MC, Goll LG, Artoni RF, Nogaroto V, et al. (2010). Physical mapping of 18S rDNA cistron in species of the Omophoita genus (Coleoptera, Alticinae) using fluorescent in situ hybridization. Micron 41: 729-734.
Bardella VB, Gaeta ML, Vanzela ALL and Azeredo-Oliveira MTV (2010). Chromosomal location of heterochromatin

2198
©FUNPEC-RP www.funpecrp.com.brGenetics and Molecular Research 13 (1): 2186-2199 (2014)
V.B. Bardella et al.
and 45S rDNA sites in four South American triatomines (Heteroptera: Reduviidae). Comp. Cytogenet 4: 141-149.Bizzaro D, Manicardi GC and Bianchi U (1996). Chromosomal localization of a highly repeated EcoRI DNA fragment
in Megoura viciae (Homoptera, Aphididae) by nick translation and fluorescence in situ hybridization. Chromosome Res. 4: 392-396.
Bonaccorsi S and Lohe A (1991). Fine mapping of satellite DNA sequences along the Y chromosome of Drosophila melanogaster: relationships between satellite sequences and fertility factors. Genetics 129: 177-189.
Bressa MJ, Larramendy ML and Papeschi AG (2005). Heterochromatin characterization in five species of Heteroptera. Genetica 124: 307-317.
Bressa MJ, Franco MJ, Toscani MA and Papeschi AG (2008). Heterochromatin heteromorphism in Holhymenia rubiginosa (Heteroptera: Coreidae). Eur. J. Entomol. 105: 65-72.
Bressa MJ, Papeschi AG, Vitková M, Kubícková S, et al. (2009). Sex chromosome evolution in cotton stainers of the genus Dysdercus (Heteroptera: Pyrrhocoridae). Cytogenet. Genome Res. 125: 292-305.
Cabral-de-Mello DC, Martins C, Souza MJ and Moura RC (2011). Cytogenetic mapping of 5S and 18S rRNAs and H3 histone genes in 4 ancient Proscopiidae grasshopper species: contribution to understanding the evolutionary dynamics of multigene families. Cytogenet. Genome Res. 132: 89-93.
Camacho JPM, Belda J and Cabero J (1985). Meiotic behavior of the holocentric chromosomes of Nezara viridula (Insecta, Heteroptera) analysed by C-banding and silver impregnation. Can. J. Genet. Cytol. 27: 490-497.
Caterino MS, Cho S and Sperling FA (2000). The current state of insect molecular systematics: a thriving Tower of Babel. Annu. Rev. Entomol. 45: 1-54.
Corpet F (1988). Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 16: 10881-10890.da Rosa R, Laforga Vanzela AL, Rubert M, Martins-Santos IC, et al. (2009). Differentiation of Y chromosome in the
X(1)X(1)X(2)X(2)/X(1)X(2)Y sex chromosome system of Hoplias malabaricus (Characiformes, Erythrinidae). Cytogenet. Genome Res. 127: 54-60.
Elder JF Jr and Turner BJ (1995). Concerted evolution of repetitive DNA sequences in eukaryotes. Q. Rev. Biol. 70: 297-320.
Franco JM, Bressa MJ and Papeschi AG (2006). Karyotype and male meiosis in Spartocera batatas (Fabricius) and meiotic behavior of multiple sex chromosome in Coreidae, Heteroptera. Eur. J. Entomol. 103: 9-16.
Grozeva SM and Kuznetsova V (1993). Notes on the karyotype of some lygaeid bugs (Heteroptera, Pentatomomorpha, Lygaeidae). Folia Biol. 41: 65-75.
Grozeva S, Kuznetsova VG and Nokkala S (2004). Patterns of chromosome banding in four nabid species (Heteroptera, Cimicomorpha, Nabidae) with high chromosome number karyotypes. Hereditas 140: 99-104.
Grozeva S, Kuznetsova V and Anokhin B (2010). Bed bug cytogenetics: Karyotype, sex chromosome system, FISH mapping of 18S rDNA and male meiosis in Cimex lectularius Linnaeus, 1758 (Heteroptera: Cimicidae). Comp. Cytogenet 4: 151-160.
Heitz E (1957). Die Chromosomenstruktur im Kern Während der Kernteilung und der Entwicklung des Organismus. Conference on Chromosomes WEJ, Zwolle, 1-22.
Kaur H, Suman V and Kaur R (2010). First report on C-banding and fluorescent banding in species of Dieuches (Rhyparochrominae: Lygaeidae: Heteroptera). Entomol. Res. 40: 1-7.
Liao D (1999). Concerted evolution: molecular mechanism and biological implications. Am. J. Hum. Genet. 64: 24-30.Panzera Y, Pita S, Ferreiro MJ, Ferrandis I, et al. (2012). High dynamics of rDNA cluster location in kissing bug holocentric
chromosomes (Triatominae, Heteroptera). Cytogenet. Genome Res. 138: 56-67.Papeschi AG and Bressa MJ (2006). Evolutionary cytogenetics in Heteroptera. J. Biol. Res. 5: 3-21.Papeschi AG and Bressa MJ (2007). Classical and Molecular Cytogenetics in Heteroptera. In: Research Advances in
Entomology (Mohan RM, ed.). Kerala, 1-9.Papeschi AG, Mola LM, Bressa MJ, Greizerstein EJ, et al. (2003). Behaviour of ring bivalents in holokinetic systems:
alternative sites of spindle attachment in Pachylis argentinus and Nezara viridula (Heteroptera). Chromosome Res. 11: 725-733.
Poggio MG, Bressa MJ and Papeschi AG (2011). Male meiosis, heterochromatin characterization and chromosomal location of rDNA in Microtomus lunifer (Berg, 1900) (Hemiptera: Reduviidae: Hammacerinae). Comp. Cytogenet 5: 1-22.
Rafael MS, Santos IP Jr, Tadei WP, Carvalho KA, et al. (2006). Cytogenetic study of Anopheles albitarsis (Diptera: Culicidae) by C-banding and in situ hybridization. Hereditas 143: 62-67.
Reeves A and Tear J (2000). MicroMeasure for Windows. Version 3.3. Free Program Distributed by the Authors Over the Internet. Available at [http://www.colostate.edu/Depts/Biology/MicroMeasure]. Accessed February, 2013.
Sumner AT (1972). A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 75: 304-306.

2199
©FUNPEC-RP www.funpecrp.com.brGenetics and Molecular Research 13 (1): 2186-2199 (2014)
Physical mapping in Lygaeidae
Tamura K, Peterson D, Peterson N, Stecher G, et al. (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28: 2731-2739.
Ueshima N (1979). Insecta 6. Hemiptera II. Heteroptera. In: Animal Cytogenetics (John B, ed.). Gebruder Borntraeger, Stuttgart, 117.
Ueshima N and Ashlock PD (1980). Cytotaxonomy of the Lygaeidae (Heteroptera). Univ. Kans. Sci. Bull. 51: 71-801.Zhang Z, Schwartz S, Wagner L and Miller W (2000). A greedy algorithm for aligning DNA sequences. J. Comput. Biol.
7: 203-214.


















