
Phylogeography of the ectomycorrhizal Pisolithus species as inferred from nuclear ribosomal DNA ITS...
13
© New Phytologist (2002) 153 : 345 – 357 www.newphytologist.com 345 Research Blackwell Science Ltd Phylogeography of the ectomycorrhizal Pisolithus species as inferred from nuclear ribosomal DNA ITS sequences Francis Martin 1,3 , Jesús Díez 1,3 , Bernard Dell 2 and Christine Delaruelle 1 1 UMR INRA–UHP ‘Interactions Arbres/Micro-Organismes’, Centre INRA de Nancy, F-54280 Champenoux, France; 2 School of Biological Sciences and Biotechnology, Murdoch University, Perth WA 6150, Australia Summary • The fungal genus Pisolithus is cosmopolitan in warm temperate regions and forms ectomycorrhizal associations with a wide range of woody plants. To delimit phylo- genetic Pisolithus species and identify their geographical distribution, 102 collec- tions were made worldwide and their rDNA internal transcribed spacer (ITS) was sequenced. • Phylogenetic analyses of these sequences, together with 46 additional GenBank accessions, identified 11 species. • A strong phylogeographical pattern was observed related to the native geograph- ical origin of the host plants. In the Holarctic, P. tinctorius was widely distributed, associated with Pinus and Quercus . It has been co-introduced with pines to other biogeographical regions. Several Pisolithus lineages, including P. aurantioscabrosus , occurred in restricted biogeographical regions associated with endemic plants, such as Afzelia in eastern Africa. Pisolithus albus , P. marmoratus and P. microcarpus were associated with Australasian hosts ( Eucalyptus , Acacia ) and were distributed with their hosts worldwide. By contrast, two additional unnamed species were restricted to Australia. • The present study shows that evolutionary lineages within Pisolithus are related to the biogeographical origin of the hosts. In addition, regional floras and endemic plants could act as hosts of endemic species of Pisolithus . Key words: Acacia , ectomycorrhizal fungus, Eucalyptus , internal transcribed spacers, Pisolithus , phylogenetic species, phylogeography, ribosomal DNA. © New Phytologist (2002) 153 : 345–357 Author for correspondence: Francis Martin Tel: +33 383 39 40 80 Fax: +33 383 39 40 69 Email: [email protected] Received: 22 June 2001 Accepted: 3 October 2001 Introduction The fungal genus Pisolithus is cosmopolitan in warm temperate regions of the world and forms ectomycorrhizal associations with a wide range of woody plants (Marx, 1977; Chambers & Cairney, 1999) including members of the Pinaceae, Myrtaceae, Fagaceae, Mimosaceae, Dipterocarpaceae and Cistaceae. Although considerable heterogeneity exists in terms of sporocarp, spore and isolate culture morphology, taxa within the genus Pisolithus have been widely regarded as conspecific and grouped as Pisolithus tinctorius (Chambers & Cairney, 1999). Pisolithus collections from throughout the world differ in their ecology, morphological and molecular characteristics, symbiotic efficiency, physiology and geographical distribution (Lamhamedi et al. , 1990; Burgess et al. , 1994, 1995; Anderson et al. , 1998, 2001; Martin et al. , 1998; Gomes et al. , 2000; Díez et al. , 2001). Based on mating tests, Kope & Fortin (1990) proposed that the genus Pisolithus comprises several biological species. DNA-based analysis further supported the occurrence of several phylogenetic species of Pisolithus . So far, four major clades were identified within two main phylogenetic lineages (Anderson et al. , 1998, 2001; Martin et al. , 1998; Gomes et al. , 2000; Díez et al. , 2001). Several morphological species of Pisolithus are common in forests and woodlands in Australia ( P. microcarpus (Cke & Mass.) Cunn.) (Cunningham, 1942; Bougher & Syme, 1998), in tropical rainforests of southeast Asia, [ P. kisslingi Fisch. and 3 These authors contributed equally to this work.
-
Upload
francis-martin -
Category
Documents
-
view
213 -
download
0
Transcript of Phylogeography of the ectomycorrhizal Pisolithus species as inferred from nuclear ribosomal DNA ITS...

©
New Phytologist
(2002)
153
: 345–357
www.newphytologist.com
345
Research
Blackwell Science Ltd
Phylogeography of the ectomycorrhizal
Pisolithus
species
as inferred from nuclear ribosomal DNA ITS sequences
Francis Martin
1,3
, Jesús Díez
1,3
, Bernard Dell
2
and Christine Delaruelle
1
1
UMR INRA–UHP ‘Interactions Arbres/Micro-Organismes’, Centre INRA de Nancy, F-54280 Champenoux, France;
2
School of Biological Sciences and
Biotechnology, Murdoch University, Perth WA 6150, Australia
Summary
• The fungal genus
Pisolithus
is cosmopolitan in warm temperate regions and formsectomycorrhizal associations with a wide range of woody plants. To delimit phylo-genetic
Pisolithus
species and identify their geographical distribution, 102 collec-tions were made worldwide and their rDNA internal transcribed spacer (ITS) wassequenced.• Phylogenetic analyses of these sequences, together with 46 additional GenBankaccessions, identified 11 species.• A strong phylogeographical pattern was observed related to the native geograph-ical origin of the host plants. In the Holarctic,
P. tinctorius
was widely distributed,associated with
Pinus
and
Quercus
. It has been co-introduced with pines to otherbiogeographical regions. Several
Pisolithus
lineages, including
P. aurantioscabrosus
,occurred in restricted biogeographical regions associated with endemic plants, suchas
Afzelia
in eastern Africa.
Pisolithus albus
,
P. marmoratus
and
P. microcarpus
wereassociated with Australasian hosts (
Eucalyptus
,
Acacia
) and were distributed withtheir hosts worldwide. By contrast, two additional unnamed species were restrictedto Australia.• The present study shows that evolutionary lineages within
Pisolithus
are relatedto the biogeographical origin of the hosts. In addition, regional floras and endemicplants could act as hosts of endemic species of
Pisolithus
.
Key words:
Acacia
, ectomycorrhizal fungus,
Eucalyptus
, internal transcribed spacers,
Pisolithus
, phylogenetic species, phylogeography, ribosomal DNA.
©
New Phytologist
(2002)
153
: 345–357
Author for correspondence:
Francis Martin Tel: +33 383 39 40 80 Fax: +33 383 39 40 69 Email: [email protected]
Received:
22 June 2001
Accepted:
3 October 2001
Introduction
The fungal genus
Pisolithus
is cosmopolitan in warm temperateregions of the world and forms ectomycorrhizal associationswith a wide range of woody plants (Marx, 1977; Chambers& Cairney, 1999) including members of the Pinaceae,Myrtaceae, Fagaceae, Mimosaceae, Dipterocarpaceae andCistaceae. Although considerable heterogeneity exists in termsof sporocarp, spore and isolate culture morphology, taxa withinthe genus
Pisolithus
have been widely regarded as conspecificand grouped as
Pisolithus tinctorius
(Chambers & Cairney,1999).
Pisolithus
collections from throughout the world differ
in their ecology, morphological and molecular characteristics,symbiotic efficiency, physiology and geographical distribution(Lamhamedi
et al.
, 1990; Burgess
et al.
, 1994, 1995; Anderson
et al.
, 1998, 2001; Martin
et al.
, 1998; Gomes
et al.
, 2000;Díez
et al.
, 2001). Based on mating tests, Kope & Fortin(1990) proposed that the genus
Pisolithus
comprises severalbiological species. DNA-based analysis further supportedthe occurrence of several phylogenetic species of
Pisolithus
.So far, four major clades were identified within two mainphylogenetic lineages (Anderson
et al.
, 1998, 2001; Martin
et al.
, 1998; Gomes
et al.
, 2000; Díez
et al.
, 2001). Severalmorphological species of
Pisolithus
are common in forestsand woodlands in Australia (
P. microcarpus
(Cke & Mass.)Cunn.) (Cunningham, 1942; Bougher & Syme, 1998), intropical rainforests of southeast Asia, [
P. kisslingi
Fisch. and
3
These authors contributed equally to this work.
NPH_313.fm Page 345 Wednesday, January 9, 2002 1:37 PM

www.newphytologist.com
©
New Phytologist
(2002)
153
: 345–357
Research346
P. aurantioscabrosus
Wat. (Watling
et al.
, 1995, 1999)], and inNew Caledonia (
P. pusillum
Pat.). Two additional Australianspecies have recently been provisionally described:
P. albus
(Cooke & Mass.) M. J. Priest,
nom. prov
., and
P. marmoratus
(Berk.) Priest,
nom. prov
. (Bougher & Syme, 1998). Thetotal number of identified
Pisolithus
species may be as manyas 10 (Sims
et al.
, 1999; Anderson
et al.
, 2001). There isthus growing consensus that the genus
Pisolithus
is moregenetically diverse than was previously assumed and thattaxonomic revision on a global scale is required (Bougher &Syme, 1998; Martin
et al.
, 1998; Chambers & Cairney,1999). The accurate delimitation and naming of those speciesis crucial because a wealth of biological information is linkedwith the current names.
The dispersal of ectomycorrhizal trees, such as eucalyptsand pines, outside their natural range, has resulted in theintroduction of a narrow range of exotic ectomycorrhizal fungiinto forest plantations in many countries of Europe, SouthAmerica and Africa (Marx, 1977; Garbaye
et al.
, 1988; Martin
et al.
, 1998; Gomes
et al.
, 2000; Díez
et al.
, 2001). Whether thedistribution of these fungi extend beyond these areas dependson the host range of the fungi and their ability to competewith indigenous species.
Studies that are based on DNA sequences serve to re-evaluate earlier classifications and provide more accuratespecies delimitation (Taylor
et al.
, 2000). Studies of phylogenyand its implications for understanding the biogeography ofectomycorrhizal fungi showing cryptic speciation have beenprogressing significantly in recent years as a result of DNAsequencing (Kretzer
et al.
, 1996; Jarosh & Bresinsky, 1999;Wu
et al.
, 2000). To date, the phylogeny of
Pisolithus
hasbeen assessed using a single locus, the internal transcribedspacer (ITS) region of the nuclear ribosomal DNA (rDNA),on a limited number of isolates from a restricted number ofgeographical regions: New South Wales (NSW) (easternAustralia) (Anderson
et al.
, 1998), Kenya (Martin
et al.
, 1998),Brazil (Gomes
et al.
, 2000) and the western Mediterraneanregion (Díez
et al.
, 2001).The objectives of this study were first to determine the phylo-
genetic relationships among
Pisolithus
species and second toextend the understanding of their host specificity and bio-geographical patterns.
Materials and Methods
Isolates
The 102 isolates of
Pisolithu
s analysed in this study were col-lected from a variety of geographical localities (i.e. ecosystems)throughout the world (Table 1) and deposited at the Collectionsof Ectomycorrhizal Fungi of the Trees–Microorganisms Inter-actions Unit (INRA-Nancy, France), School of BiologicalSciences and Biotechnology (Murdoch University, WA, Australia),and CSIRO Division of Forestry and Forest Products (Wembley,
WA, Australia). Morphological descriptions of representativespecimens were reported in Burgess
et al
. (1995), Watling
et al
. (1995, 1999), Anderson
et al
. (1998, 2001), Martin
et al
. (1998), Cairney
et al
. (1999), and Díez
et al
. (2001).Samples collected by the authors were excised from the centraltissues of the basidiome stipe to avoid contamination by otherfungi. Cultures obtained from basidiome were maintainedon modified Melin Norkrans’s (MMN) medium agar (Marx,1981) or Pachlewski medium (Martin
et al.
, 1998) at 25
°
Cwith subculturing every 2–3 months.
DNA manipulations
Samples for DNA extraction were excised from fresh basidiomata,basidiomata preserved in glycerol /ethanol/water (30/30/40)or the outer part of mycelial agar cultures. DNA was isolatedfrom each isolate with the DNAeasy Kit (Qiagen, Dusseldorf,Germany). The ITS of nuclear rDNA were amplified as describedby Martin
et al
. (1998) with the ITS1 and ITS4 primers (White
et al.
, 1990). Amplifications were performed on a GeneAmp9600 PCR system (Perkin Elmer, Norwalk, CT, USA). Thetwo ITS alleles amplified from the fruiting body of isolatePasoh01 from
P. aurantioscabrosus
were subcloned in the plas-mid pCR4-TOPO (Invitrogen, Groningen, The Netherlands)before sequencing. DNA amplification products were purifiedwith BIO 101 GeneClean III Kit (BIO 101, Inc., Carlsbad,CA, USA) and sequenced using the
Taq
Big Dye TerminatorCycle Sequencing Kit (Applied Biosystems Inc., Norwalk, CT,USA), following the manufacturer’s instructions. Sequenceswere generated for both strands using the ITS1 and ITS4primers. Reactions were then electrophoresed on either an ABI377 (at Murdoch University, WA, Australia) or ABI Genotyper310 (at INRA-Nancy) automated sequencer (Applied BiosystemsInc.). Gels were tracked using the ABI Prism Sequencingprogramme and raw sequence data were edited using theSequence Analysis 3.3 (Applied Biosystems Inc.) programmeand assembled with Sequencher 3.1.1 (Gene Codes Corporation,Ann Arbor, MI, USA) programme. About 20% of the ampli-fied ITS were sequenced on both machines to check forbase calling reproducibility. Sequences were deposited inthe National Center for Biotechnology Information (NCBI)GenBank (accession n
°
AF374622 to AF374720, AF415226& AF415227) (Table 1).
Sequence and data analyses
Search for sequence identity in the GenBank DNA database wasconducted by Gapped BlastN (NCBI) (Altschul
et al.
, 1997).The 102 sequences from the present survey and 46 additional
Pisolithus
sequences retrieved from GenBank were alignedusing the programme MultAlin (http://www.toulouse.inra.fr/multalin.html) (Corpet, 1988). The resulting multiple align-ments were optimized visually. Only unambiguous alignmentswere used in the phylogenetic and distance analyses. The
NPH_313.fm Page 346 Wednesday, January 9, 2002 1:37 PM

©
New Phytologist
(2002)
153
: 345–357
www.newphytologist.com
Research 347
Table 1
Pisolithus
isolates studied, with information concerning their host, geographical origin, code used in the cladogramme, and GenBank accession number
Isolate no. Host
1
Locality Code GenBank accession
Afzelia
(1) 5105
Afzelia quanzensis
Arabuko-Sokoke Forest Reserve, Kenya
5105_Afz_Ara_Ken AF003915
3
(2) K915
A. quanzensis
Arabuko-Sokoke Forest Reserve, Kenya
K915_Afz_Ara_Ken AF228653
5
Acacia
(3) COI 24
Acacia holosericea
Sinthian Malène, Senegal COI24_Aca_Sin_Sen AF374622(4) COI 07
A. mangium
Casamance, Senegal COI07_Aca_Cas_Sen AF374623(5) CSH4461
Acacia
sp. Townsville, Queensland CSH4461_Aca_Two_Qld AF374624(6) Gemas
A. mangium
Gemas, Malaysia Gemas_Aca_Gem_Mal AF374638
Dipterocarps
(7) Pasoh01
Shorea macroprera
Forest Service, Pasoh, Malaysia Pasoh01_Sho_Pas_Mal AF415226 & AF415227
Pinus, Quercus
or
Cistus
(8) Pt301
Pinus
sp. Georgia, USA Pt301_Pin_Geo_USA AF143233
4
(9) MH208
P. pinaster
unknown, Portugal MH208_Pin_unk_Por AF374626(10) MP9812
Pinus
sp. Voeltjiesdord, South Africa MP9812_Pin_Voe_SAf AF374627(CSH557)(11) MP9813
P. radiata
Grabouw, South Africa MH9813_Pin_Grb_SAf AF374628(CSH558)(12) PtJap
P. pumila/Betula ermanii
Mt Iou, Shiretoko Peninsula, PtJapan_Pin_Iou_Jap AF374629(MH175) Hokkaido, Japan(13) Hel1
P. pinaster/Quercus suber
unknown, Portugal Hel1_Pin/Que_unk_Por AF374630(MH177)(14) MSN
P. kesiya
Mae Sa Nam, Chiangmai MSN_Pin_Mae_Tha AF374625(MH181) Province, Northern Thailand(15) THAI 1
P. kesiya
Chiangmai Province, THAI1_Pin_Chi_Tha AF374631(MH140) Northern Thailand(16) MARX270
P. elliotii
Georgia, USA MARX270_Pin_Geo_USA AF374632(17) 5111
P. caribaea
Arabuko, Kenya 5111_Pin_Ara_Ken AF003916
3
(18) esp07
P. caribaea
Arabuko, Kenya esp07_Pin_Ara_Ken AF228647(19) NIC (S382)
P. caribaea
Puerto Cabezas, Nicaragua NIC(S382)_Pin_Pue_Nic AF374633(20) OAK
Pinus
sp. Georgia, USA OAK_Que_Geo_USA AF374634(21) ATH
Pinus
sp. Georgia, USA ATH_Que_Geo_USA AF374635(22) Pt92JG
P. pinaster/Q. suber
Alcácer du Sal, Portugal Pt92JG_Pin/Que_Alc_Por AF374636(MH171)cab01
Cistus ladanifer
Cabañeros National Park, Ciudad Real, Spain
cab01_Cis_Cab_Spa AF228644
5
(23) ch01
Q. ilex/C. ladanifer
Chapiñeria, Madrid, Spain ch01_Que_Cha_Spa AF228645
5
(25) ch02
Q. ilex/C. ladanifer
Chapiñeria, Madrid, Spain ch02_Que_Cha_Spa AF228646
5
(26) cr01
C. ladanifer
Abenojar, Ciudad Real, Spain cr01_Cis_Abe_Spa AF228641
5
(27) cr02
C. ladanifer
Abenojar,Ciudad Real, Spain cr02_Cis_Abe_Spa AF2286425
(28) cr04 C. ladanifer Abenojar, Ciudad Real, Spain cr04_Cis_Abe_Spa AF2286435
(29) gr13 P. halepensis/Q. coccifera Granada, Spain gr13_Pin/Que_Gra_Spa AF2286505
(30) m14 P. halepensis Moratalla, Murcia, Spain m14_Pin_Mor_Spa AF2286525
(31) pt03 Q. ilex/Q. coccifera Valencia, Spain pt03_Que_Val_Spa AF2286485
(32) pt04 Q. ilex Fuentidueña, Spain pt04_Que_Fue_Spa AF2286495
(33) pt05 Q. ilex Fuentidueña, Spain pt05_Que_Fue_Spa AF2286515
(34) MH143 P. yunnanensis Kunming, China MH143_Pin_Kun_Chi AF374717(35) H529 Pinus sp. unknown, USA H529_Pin_unk_USA AF374709(36) F22 P. pinaster Lac d’Aureilhan, Mimizan, France F22_Pin_Mim_Fra AF374707(37) H614 P. elliotii Kaiping, China H614_Pin_Kai_Chi AF374710(38) H613 Pinus/Eucalyptus Kaiping, China H613_Pin/Euc_Kai_Chi AF374711(39) H237 Pinus sp. unknown, France H237_Pin_unk_Fra AF374712
Eucalyptus plantations and native forests in Australia(40) MU98/20 E. calophylla Karridale, Western Australia MU98/20_Euc_Kar_WA AF374637(41) MU98/15 E. calophylla North Augusta, Western Australia MU98/15_Euc_Aug_WA AF374639(42) WM01 Eucalyptus sp. Westmead, New South Wales WM01_Euc_Wes_NSW AF0047342
(43) CS01 Eucalyptus sp. unknown, New South Wales CS01_Euc_unk_NSW AF0047322
(44) LJ07 Eucalyptus sp. N. Turramurra, New South Wales LJ07_Euc_Tur_NSW AF0047332
(45) R01 Eucalyptus sp. unknown, New South Wales R01_Euc_unk_NSW AF0047352
(46) MU98/1 E. marginata Augusta, Western Australia MU98/1_Euc_Aug_WA AF374640(47) MU98/2 E. marginata Augusta, Western Australia MU98/2_Euc_Aug_WA AF374641
NPH_313.fm Page 347 Wednesday, January 9, 2002 1:37 PM

www.newphytologist.com © New Phytologist (2002) 153: 345–357
Research348
(48) MU98/3 E. marginata Augusta, Western Australia MU98/3_Euc_Aug_WA AF374642(49) MU98/4 E. marginata N. Augusta, Western Australia MU98/4_Euc_Aug_WA AF374643(50) MU98/5A E. globulus Kudardup, Western Australia MU98/5A_Euc_Kud_WA AF374644(51) MU98/5B E. globulus Kudardup, Western Australia MU98/5B_Euc_Kud_WA AF374645(52) MU98/6 E. globulus Scott R, Western Australia MU98/6_Euc_Sco_WA AF374646(53) MU98/7 E. globulus Scott R, Western Australia MU98/7_Euc_Sco_WA AF374647(54) MU98/8 E. globulus Scott R, Western Australia MU98/8_Euc_Sco_WA AF374648(55) MU98/9 E. globulus Scott R, Western Australia MU98/9_Euc_Sco_WA AF374649(56) MU98/10 E. marginata W. Manjimup, Western Australia MU98/10_Euc_Man_WA AF374650(57) MU98/11 E. marginata E. Manjimup, Western Australia MU98/11_Euc_Man_WA AF374651(58) MU98/12 Eucalyptus spp. Manjimup, Western Australia MU98/12_Euc_Man_WA AF374652(59) MU98/13 Eucalyptus spp. Manjimup, Western Australia MU98/13_Euc_Man_WA AF374653(60) MU98/14 Eucalyptus spp. South-west, Western Australia MU98/14_Euc_Sou_WA AF374654(61) MU98/16 E. globulus Kudardup, Western Australia MU98/16_Euc_Kud_WA AF374655(62) MU98/17 E. globulus Kudardup, Western Australia MU98/17_Euc_Kud_WA AF374656(63) MU98/18 E. globulus Kudardup, Western Australia MU98/18_Euc_Kud_WA AF374657(64) MU98/19 E. globulus Kudardup, Western Australia MU98/19_Euc_Kud_WA AF374658(65) MU98/21 E. globulus Albany, Western Australia MU98/21_Euc_Alb_WA AF374659(66) MU98/22 E. globulus Albany, Western Australia MU98/22_Euc_Alb_WA AF374660(67) MU98/101 E. camaldulensis S. Eneabba, Western Australia MU98/101_Euc_Ene_WA AF374661(68) MU98/102 Eucalyptus/Acacia Eneabba, Western Australia MU98/102_Euc/Aca_Ene_WA AF374662(69) MU98/103 Eucalyptus/Acacia Eneabba, Western Australia MU98/103_Euc/Aca_Ene_WA AF374663(70) MU98/104 E. globulus N. Albany, Western Australia MU98/104_Euc_Alb_WA AF374664(71) MU98/105 E. patens Kalamunda, Western Australia MU98/105_Euc_Kal_WA AF374665(72) CSH4338 Eucalyptus sp., Atherton, Queensland CSH4338_Euc_Ath_Qld AF374666
Melaleuca sp.(73) CSH4742 E. grandis Mt Spec, Queensland CSH4742_Euc_MtS_Qld AF374667(74) CSH5822 Eucalyptus sp. New England National Park,
New South WalesCSH5822_Euc_New_NSW AF374668
(75) MH19 (CSH445)
E. marginata Jarrahdale, Western Australia MH19_Euc_Jar_WA AF374669
(76) H4937 (MH132)
E. tereticornis, E. tesselaris Fanning River, Queensland H4937_Euc_FR_Qld AF374670
(77) MH155 (CSH1101)
E. globulus Northcliffe, Western Australia MH155_Euc_Nor_WA AF374671
(78) MH156 (CSH4334)
Eucalyptus sp. Atherton Tableland, Quensland MH156_Euc_Ath_WA AF374672
(79) MH158 (CSH6386)
E. urophylla Guangdong, China MH158_Euc_Gua_Chi AF374673
(80) MH730 (CSH4111)
Eucalyptus sp. Beerwah, Queensland MH730_Euc_Bee_Qld AF374674
(81) CSH4339 (MH731)
Eucalyptus sp. Atherton Tableland, Queensland MH731_Euc_Ath_Qld AF374675
(82) MH732 (CSH6291)
Eucalyptus sp. Cardwell, Queensland MH732_Euc_Car_Qld AF374676
(83) MH97 Eucalyptus sp., N. Morawa, Western Australia MH97_Euc_Mow_WA AF374713Melaleuca sp.
(84) MH115 Eucalyptus/Acacia Wittenoom, Western Australia MH115_Euc/Aca_Wit_WA AF374714(85) MH22 Eucalyptus/Acacia Wubin, Western Australia MH22_Euc/Aca _Wub_WA AF374715(86) H4111 Eucalyptus sp. unknown, Queensland H4111_Euc_unk_Qld AF374718(87) MH56 Eucalyptus/Acacia Kalbarri, Western Australia MH56_Euc/Aca _Klb_WA AF374708(88) SA01 Eucalyptus sp. Uraidla, Adelaide Hills, South
AustraliaSA01_Euc_Ura_SA AF2707876
(89) QLD05 Eucalyptus sp. Mareeba, North Queensland QLD05_Euc_Mar_Qld AF2707866
(90) QLD01 Eucalyptus sp. Wangetti, North Queensland QLD01_Euc_Wan_Qld AF2707856
(91) NSW5 Eucalyptus sp. Ballina, New South Wales NSW5_Euc_Bal_NSW AF2707846
(92) QLD02 Eucalyptus sp. Wangetti, North Queensland QLD02_Euc_Wan_Qld AF2707836
(93) CA02 Eucalyptus sp. Canberra, Australia Capital Territory
CA02_Euc_Can_ACT AF2707826
(94) VIC01 Eucalyptus sp. Warrandyte, Victoria VIC01_Euc_War_Vic AF2707816
(95) WM02 Eucalyptus sp. Westmead, New South Wales WM02_Euc_Wes_NSW AF2707806
(96) W34 Eucalyptus sp. N. Wilberforce, New South Wales W34_Euc_Wil_NSW AF2707796
(97) R08 Eucalyptus sp. Wattamolla, New South Wales R08_Euc_Wat_NSW AF2707786
(98) LJ30 Eucalyptus sp. N. Turramurra, New South Wales LJ30_Euc_Tur_NSW AF2707746
Isolate no. Host1 Locality Code GenBank accession
Table 1 continued
NPH_313.fm Page 348 Wednesday, January 9, 2002 1:37 PM

© New Phytologist (2002) 153: 345–357 www.newphytologist.com
Research 349
(99) BW02 Eucalyptus sp. Patonga, New South Wales BW02_Euc_Pat_NSW AF2707736
(100) BW01 Eucalyptus sp. Patonga, New South Wales BW01_Euc_Pat_NSW AF2707726
(101) BTH1 Eucalyptus sp. Bathurst, New south Wales BTH1_Euc_Bat_NSW AF2707716
(102) MH2 E. saligna Perth, Western Australia MH2_Euc_Per_WA AF374716(103) KS781 E. calophylla Denmark, Western Australia KS781_Euc_Den_WA AF374719
Eucalyptus plantations outside Australia(104) mam17 Eucalyptus sp. Mammora Forest, Morocco mam17_Euc_Mam_Mor AF2286575
(105) mar01 Eucalyptus sp. Mammora Forest, Morocco mar01_Euc_Mam_Mor AF2286545
(106) mar02 Eucalyptus sp. Mammora Forest, Morocco mar02_Euc_Mam_Mor AF2286555
(107) ast05 Eucalyptus sp. The Miravete Pass, Cáceres, Spain ast05_Euc_Mir_Spa AF2286565
(108) 5110 E. camaldulensis Gede Forest Station, Kenya 5110_Euc_Ged_Ken AF0039143
(109) 441 E. citriodora Sao Paulo, Brazil 441_Euc_Sao_Bra U626661
(110) Pt90A Eucalyptus sp. Vizcosa, Brazil Pt90A_Euc_Viz_Bra AF1405474
(111) MH210 E. globulus unknown, Portugal MH210_Euc_unk_Por AF374677(112) MH727 (CSH705)
E. urophylla Zenhai, China MH727_Euc_Zen_Chi AF374678
(113) MH728 (CSH708)
Eucalyptus sp. Yanxi, China MH728_Euc_Yan_Chin AF374679
(114) MH729 E. grandis Yanxi, China H729_Euc_Yan_Chi AF374680(CSH709)(115) MH745 (CSH710)
E. urophylla Guangdong, China MH745_Euc_Gua_Chi AF374681
(116) MP9811 Eucalyptus sp. Wiesonhof, South Africa MP9811_Euc_Wie_SAf AF374682(CSH556)(117) MP9814 E. tereticornus Tamil Nadiu, India MP9814_Euc_Tam_Chi AF374683(CSH753)(118) HCX1 E. maidenii Chuxiong, China HCX1_Euc_Chu_Chi AF374684(119) COI03 E. camaldulensis Bandia, Senegal COI03_Euc_Ban_Sen AF374685(120) COI05 E. camaldulensis Djibelor, Senegal COI05_Euc_Dji_Sen AF374686(121) COI12 E. camaldulensis Kabrousse, Senegal COI12_Euc_Kab_Sen AF374687(122) COI15 E. camaldulensis Casamance, Senegal COI15_Euc_Cas_Sen AF374688(123) COI16 E. camaldulensis Casamance, Senegal COI16_Euc_Cas_Sen AF374689(124) COI22 E. camaldulensis Casamance, Senegal COI22_Euc_Cas_Sen AF374690(125) COI25 E. camaldulensis Casamance, Senegal COI25_Euc_Cas_Sen AF374691(126) COI26 E. camaldulensis Casamance, Senegal COI26_Euc_Cas_Sen AF374692(127) COI27 E. camaldulensis Casamance, Senegal COI27_Euc_Cas_Sen AF374693(128) COI30 E. camaldulensis Ngan, Senegal COI30_Euc_Nga_Sen AF374694(129) R15 E. globulus Castelo Branco, Portugal R15_Euc_Cst_Por AF374695(130) UFSC-Pt22 Eucalyptus sp. Três Barras, SC, Brazil UFSC22_Euc_Trê_Bra AF374696(131) UFSC-Pt23 Eucalyptus sp. UFSC, Florianópolis, SC, Brazil UFSC23_Euc_Flo_Bra AF374697(132) UFSC-Pt25 Eucalyptus sp. UFSC, Florianópolis, SC, Brazil UFSC25_Euc_Flo_Bra AF374698(133) UFSC-Pt26 Eucalyptus sp. UFSC, Florianópolis, SC, Brazil UFSC26_Euc_Flo_Bra AF374699(134) UFSC-Pt27 Eucalyptus sp. UFSC, Florianópolis, SC, Brazil UFSC27_Euc_Flo_Bra AF374700(135) UFSC-Pt44 Eucalyptus sp. UFSC, Florianópolis, SC, Brazil UFSC44_Euc_Flo_Bra AF374701(136) UFSC-Pt49 Eucalyptus sp. UFSC, Florianópolis, SC, Brazil UFSC49_Euc_Flo_Bra AF374702(137) UFSC-116 E. dunni Três Barras, SC, Brazil UFSC116_Euc_Trê_Bra AF374703(138) UFSC-132 E. dunni Três Barras, SC, Brazil UFSC132_Euc_Trê_Bra AF374704(139) UFSC-145 Eucalyptus sp. UFSC, Florianópolis, SC, Brazil UFSC145_Euc_Flo_Bra AF374705(140) ITA6 Eucalyptus sp. Itamarandiba, MG, Brazil ITA6_Euc_Ita_Bra AF374706(141) RS26 Eucalyptus sp. Vizcosa, Brazil RS26_Euc_Viz_Bra AF1429914
Unknown (host)(142) PF Unknown unknown, France PF_unk_unk_Fra AF1432344
(143) DSM4271 Unknown unknown DSM4271_unk_unk AF0969766
(144) W15 Unknown N. Wilberforce, New South Wales W15_unk_Wil_NSW AF0047362
(145) W16 Unknown N. Wilberforce, New South Wales W16_unk_Wil_NSW AF0047372
(146) SU1 Unknown North Sumatra SU1_unk_unk_NSu AF2707776
(147) PH1 Unknown Philippines PH1_unk_unk_Phi AF2707766
(148) NA1 Unknown USA NA1_unk_unk_USA AF2707756
(149) H714 Unknown unknown, Western Australia H714_unk_unk_WA AF374720
*Ectomycorrhizal plants in the vicinity of the collected fruit bodies were listed as hosts. Sequences obtained from GenBank: 1Carnero-Díaz et al. (1997); 2Anderson et al. (1998); 3Martin et al. (1998); 4Gomes et al. (2000); 5Díez et al. (2001); 6Anderson et al. (2001); and 7GenBank, unpublished.
Isolate no. Host1 Locality Code GenBank accession
Table 1 continued
NPH_313.fm Page 349 Wednesday, January 9, 2002 1:37 PM

www.newphytologist.com © New Phytologist (2002) 153: 345–357
Research350
aligned sequences were exported to a NEXUS file andanalysed using PAUP*4.08b (PPC/Altivec) (Swofford, 1999).The final alignment of the ITS1/5.8S/ITS2 region had a totalof 706 nonambiguously aligned sites: 133 characters were con-stant, 92 variable characters were parsimony-uninformativeand 481 characters were parsimony-informative. The finalalignment of Pisolithus sequences is available on request.
Maximum parsimony (MP) was the primary method toinfer phylogenetic relationships between the Pisolithus isolates,and all final results were compared with neighbour joining(NJ) and maximum likelihood (ML) (Swofford, 1999) to detectany algorithm-specific results. MP analysis was carried outwith data sets that included and excluded gaps, to detect anycharacter type-specific results. MP trees were identified usingheuristic searches based on 1000 random sequence additions,retaining clades compatible with the 50% majority-rule inthe bootstrap consensus tree, using the Nearest-NeighbourInterchange (NNI) branch-swapping option and saving nomore than 100 trees with > 1000 length for each replicationand MULPART in effect (Swofford, 1999). All characterswere treated as unordered and with equal weight. Trees weredrawn with TreeView (Page, 1996). ITS sequences of Paxillusinvolutus (GenBank accession n° AF167700) and Suillus luteus(GenBank accession n° L54110) were used as outgroup taxa.Pairwise distance between isolate sequences were calculatedaccording to the Jukes–Cantor model ( Jukes & Cantor, 1969).The criteria used to define a phylogenetic species were two-fold: the existence of a monophyletic terminal clade and a ITSsequence divergence among isolates within the clade ≤ 12%.
Results
The phylogeny of Pisolithus
We obtained the same overall topology for well-resolvedphylogenetic trees regardless of the methods MP, NJ or ML(data not shown). The placement of single sequences wasalso constant in parsimony and distance-based trees (data notshown). The 50% majority-rule consensus tree, generated bybootstrap heuristic search (Swofford, 1999), resulted in threedistinct Pisolithus lineages, A, B and C with strong bootstrapsupport (Fig. 1). These lineages comprised 11 major terminalclades (referred to as phylogenetic species). Lineage C onlycomprised P. aurantioscabrosus, which formed a basal clade.P. tinctorius and P. marmoratus grouped in lineage A, whereasP. albus and P. microcarpus belonged to lineage B.
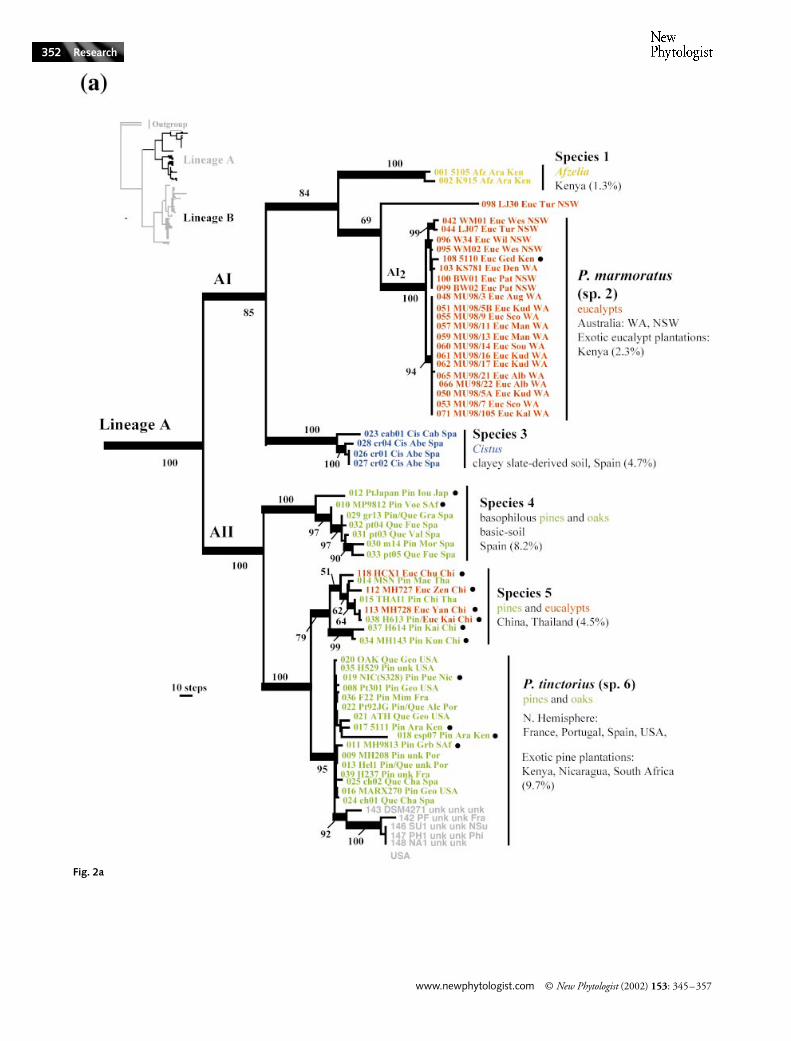
Lineage A branched into two well-supported sublineages:AI and AII (Fig. 2a). The sublineage AI terminated in threewell supported clades (species 1, 3 and P. marmoratus) and anadditional isolate of New South Wales (NSW), LJ30. Isolatesof species 1 were collected under an ectomycorrhizal Caes-alpinioideae of the coastal Miombo-like woodlands of Kenya,Afzelia quanzensis (Martin et al., 1998). Isolates of species 2were found in eucalypt plantations in the coastal lowlands of
Kenya (Martin et al., 1998), in native sclerophyllous forests ofNSW (Anderson et al., 1998, 2001) and in natural forests andplantations in WA. Species 2 is P. marmoratus as the sequenceof the voucher specimen KS781 (WA Herbarium), which wasused for the published description in Bougher & Syme (1998),falls within this clade. Species 3 included isolates that werefound in association with Cistus in Spain (Díez et al., 2001).
Sublineage AII branched into three terminal clades withstrong bootstrap support (species 4, 5 and P. tinctorius) (Fig. 2a).Species 4 included isolates found in association with pinesand oaks on basic soils in Spain (Díez et al., 2001). It alsocomprised two isolates collected under pines in Japan andSouth Africa. Isolates of species 5 were collected under variouspine and eucalypt species in Thailand and China. They do notcorrespond with the south-east Asian taxon, P. aurantioscabrosus(Watling et al., 1995, 1999) (Fig. 1), but to a species closelyrelated to P. tinctorius. Isolates of P. tinctorius were found inassociation with pine and oak species in North America andEurope, and in pine plantations in Nicaragua, Kenya andSouth Africa.
Lineage B included isolates associated with eucalyptsand acacias. The sublineage BI (Fig. 2b) ramified into severalmoderately-to-well supported branches and terminal clades.The topology of this sublineage was similar regardless of themethods (MP, NJ and ML). It identified three main groups(species 7–9). Isolates of species 7 showed the morphologicalfeatures of P. albus sensu Bougher & Syme (1998). The sequencesof isolates H4339 and H4937 were 100% identical to the voucherspecimen T25070 (Oregon State University Herbarium)deposited as P. albus. They were found in plantations of euca-lypts and acacias and in forests in various regions of Australia(ACT, NSW, Qld, Southern Australia, Victoria and WA) and inplantations of eucalypts and acacias in China, India, Malaysia,Morocco, Spain and Senegal. The ITS sequence variationwithin this group was significant (= 10.2%). Species 8 onlyincluded isolates collected under acacias and eucalypts in WA.Isolates of species 9 exhibited the morphological features ofP. microcarpus (Cunningham, 1942) and the sequenced isolatesUFSCPt132 and UFSCPt145 (Fig. 2b) correspond to twovoucher specimens (Herbarium of the Universidad Federal deSanta Catarina, Brazil) deposited as P. microcarpus (Giachiniet al., 2000). P. microcarpus isolates were found in plantationsof eucalypts and acacias and forests in Australia (WA, NSWand Qld) and in plantations throughout the world (e.g. Brazil,China, Morocco, Portugal, Senegal, South Africa).
Isolates of species 10 (lineage BII) were collected undereucalypts in WA and are probably a new species. It showed12% genetic distance with the acacia-associated isolate CSH4461from Queensland, which might represent a further species.
Evolutionary distance between and within species
Percentage pairwise sequence divergence across the entire ITSsequence ranged from 7% (between species 5 and P. tinctorius)
NPH_313.fm Page 350 Wednesday, January 9, 2002 1:37 PM

© New Phytologist (2002) 153: 345–357 www.newphytologist.com
Research 351
to (36%) (species 1 and 10). Within-species divergence rangedfrom 0.1% (P. marmoratus) to 9.7% (P. tinctorius). MaximumITS divergence between species associated with eucalypts andacacias was 32% (P. marmoratus vs species 10), whereas thehighest divergence within Pisolithus associated with Holartichosts was 23% (species 3 vs species 5) (Table 2).
Discussion
Within Pisolithus, biological species that are difficult or evenimpossible to distinguish based on morphological features wererevealed using mating tests (Kope & Fortin, 1990). DNA-basedphylogenetic analyses of a restricted set of isolates suggested
the occurrence of several phylogenetic species within Pisolithus(Anderson et al., 1998, 2001; Martin et al., 1998; Cairneyet al., 1999; Díez et al., 2001). In the present survey, we haveextended these previous phylogenetic studies to delimit Pisolithustaxa among 148 collections from natural vegetation and forestplantations throughout the world. There are significant sim-ilarities among tree topologies generated with the MP, NJ andML methods in this study, strengthening the evolutionaryrelationships inferred. The bootstrap value of clades used todefine phylogenetic species was moderate (78% for species 5)to high (100%) (Fig. 1). The evolutionary distance betweenterminal clades indicated the likelihood that they representat least 11 distinct phylogenetic species. Whether additional
Fig. 1 Phylogenetic relationships among Pisolithus species based on the rDNA 5.8S/ITS2 sequence. The 50% majority rule consensus tree shown here is based on a parsimony analysis. Bootstrap heuristic search (1000 replicates) was conducted using PAUP 4.0b8. Tree length, 994 steps; consistency index (CI), 0.639, retention index (RI), 0.846; rescaled consistency index (RC), 0.540. Tree was rooted using Paxillus and Suillus sequences as outgroup. Branch lengths are proportional to the number of nucleotides changes. Significant bootstrap frequencies (> 50%) are indicated. Outlines indicate terminal clades (species) and major lineages. Strain code: (isolate name) (host) (locality) (country). Hosts: Aca, Acacia spp.; Afz, Afzelia quanzensis; Cis, Cistus spp.; Euc, Eucalyptus spp.; Pin, Pinus spp.; Que, Quercus spp.; Sho, Shorea macroprera; unk, unknown.Locality: Ara, Arabuko; Abe, Abenojar (Ciudad Real); Cab, Cabañeros National Park (Ciudad Real); Can, Canberra; Cha, Chapinería (Madrid); Den, Denmark; Ene, Eneabba (RGC mine); Gem, Gemas; Geo, Georgia, USA; Iou, Mt Iou, Shiretoko Peninsula, Hokkaido; Kai, Kaiping; Kal, Kalamunda; Klb, Kalbarri; Mea, Mea Sa Nam; Mow, Morawa; Sao, Sao Paulo; Sin, Sinthian Malène; Sco, Scott; Tow, Townsville; Tre, Três Barras; Tur, Turramurra; unk, unknown; Val, Valencia; Voe, Voeltjiesdord; Wil, Wilberforce; Yan, Yanxi.Country: ACT, Australian Capital Territory; Bra, Brazil; Chi, China; Jap, Japan; Ken, Kenya; Mal, Malaysia; NSW, New South Wales (Australia); Qld, Queensland (Australia); Sen, Senegal; SAf, South Africa; Spa, Spain; Tha, Thailand; WA, Western Australia.
NPH_313.fm Page 351 Wednesday, January 9, 2002 1:37 PM
www.newphytologist.com © New Phytologist (2002) 153: 345–357
Research352
Fig. 2a
NPH_313.fm Page 352 Wednesday, January 9, 2002 1:37 PM

© New Phytologist (2002) 153: 345–357 www.newphytologist.com
Research 353
Fig. 2b. Caption on following page.
NPH_313.fm Page 353 Wednesday, January 9, 2002 1:37 PM

www.newphytologist.com © New Phytologist (2002) 153: 345–357
Research354
terminal clades (e.g. isolates LJ30, CS01, and CSH4461) corres-pond to new phylogenetic species will await additional samplingand sequencing.
Pisolithus tinctorius as a lineage associated with Holartic hosts
It has been suggested (Bougher & Syme, 1998; Díez et al.,2001) that P. tinctorius is confined to Holartic hosts. In thepresent analysis, P. tinctorius associated with pines and oaks(e.g. MH208) in the northern hemisphere. P. tinctorius alsooccurred in pine plantations in Nicaragua (NIC(S382)), SouthAfrica (MH9813) and Kenya (5111, Martin et al., 1998, esp07,
Díez et al., 2001), in northern Sumatra (SU1) and the Philippines(PH1). Thus, the present study supports the hypothesis thatthis Pisolithus species was introduced to the southern hemi-sphere with pines (Dunstan et al., 1998; Díez et al., 2001).Two isolates from pine plantations in South Africa (MP9812)and Japan (PtJap) did not belong to P. tinctorius, but they areclosely related to species 4 occurring in basophilous woodlandsin Spain (Díez et al., 2001), suggesting that they have beenintroduced from the Mediterranean region into South Africaand Asia along with pines.
In this survey, P. tinctorius isolates were not found inAustralia, even though extensive basidiocarp surveys have beenundertaken in eucalypt forests and plantations. By contrast to
Table 2 Pairwise comparisons of ITS1, 5.8S and ITS2 sequences between species of Pisolithus. The percentage of divergence was estimated according to the Jukes & Cantor model
Species 1 2 3 4 5 6 7 8 9 10 11
Lineage A1 Species 1 –2 P. marmoratus (sp. 2) 22 –3 Species 3 25 16 –4 Species 4 25 17 19 –5 Species 5 27 20 23 14 –6 P. tinctorius (sp. 6) 26 20 21 14 7 –
Lineage B7 P. albus (sp. 7) 33 28 31 32 9 29 –8 Species 8 32 29 32 31 29 29 7 –9 P. microcarpus 34 29 32 33 31 30 8 10 –10 Species 10 36 32 33 36 33 33 23 5 21 –
Lineage C*11 P. aurantioscabrosus (sp. 11) 37 38 40 41 40 39 42 41 41 42 –
Comparison for species 11 (P. aurantioscabrosus) was based on the 5.8S and ITS2 sequences. The percentage of divergence between the two ITS alleles of P. auriantoscabrosus was 4%. Isolates used in the analysis: 1, 001_5105; 2, 055_MU98/9; 3, 026_cr01; 4, 031_pt03; 5, 014_MH181; 6, 016_MARX270; 7, 119_COI3; 8, 068_MU98/102; 9, 109_441; 10, 052_MU98/6; 11, 007_Pasoh01_allele1.
Fig. 2 Phylogenetic relationships among Pisolithus collected worldwide based on rDNA 5.8S, ITS1 and ITS2 sequences. The 50% majority rule consensus tree shown here is based on parsimony analysis using sequence data from 148 Pisolithus isolates. Bootstrap heuristic search (1000 replicates) was conducted using PAUP 4.0b8. Tree length, 2678 steps; consistency index (CI), 0.539; retention index (RI), 0.943; rescaled consistency index (RC), 0.508. The tree was rooted using Suillus and Paxillus sequences as outgroup. Branch lengths are proportional to the number of nucleotides changes. Significant bootstrap frequencies (> 50%) are indicated. Outlines indicate terminal clades (species). Within-clade genetic distances are indicated in brackets (Jukes–Cantor model). (a) Detail, lineage A. Yellow lines, Afzelia; blue lines, Cistus; red lines, Eucalyptus; green lines, Pinus and Quercus; grey lines, unknown; circles, exotic plantation. (b) Detail, lineage B. Purple lines, Acacia; red lines, Eucalyptus; grey lines, unknown; circles, exotic plantation. Strain code (first three letters): (isolate name) (host) (locality) (country). Locality: Alc, Alcácer du Sal; Ara, Arabuko; Aben, Abenojar (Ciudad Real); Ath, Atherton Tableland; Aug, Augusta; Bal, Ballina; Ban, Bandia; Bee, Beerwah; Cab, Cabañeros National Park (Ciudad Real); Can, Canberra; Car, Cardwell; Cas, Casamance; Cha, Chapinería (Madrid); Chi, Chiang Mai; Chu, Chuxiong; Cst, Castelo do Branco; Den, Denmark; Dji, Djiblor; Ene, Eneabba (RGC mine); Flo, Florianopolis, SC (UFSC); FR, Fanning River; Fue, Fuentidueña; Ged, Gede Forest Station; Geo, Georgia, USA; Gem, Gemas; Gra, Granada; Grb, Grabouw; Gua, Guangdong; Hud, Kudardup; Ita, Itamarandiba; Iou, Mt Iou, Shiretoko Peninsula, Hokkaido; Jar, Jarrahdale; Kab, Kabrouse; Kai, Kaiping; Kal, Kalamunda; Kar, Karridale; Klb, Kalbarri; kud, Kudardup; Kun, Kunming; Mar, Mareeba; Mam, Mammora Forest; Man, Manjimup; Mae, Mae Sa Nam; Mim, Mimizan (Lac d’Aureilhan); Mir, The Miravete Pass (Cáceres); Mor, Moratalla (Murcia); Mow, Morawa; Mts, Mount Spec, Queensland; New, New England National Park, NSW; Nga, Ngan; Nor, Northcliffe; Pas, Forest Service, Pasoh; Per, Perth; Pue, Puerto Cabezas; Sao, Sao Paulo; Sin, Sinthian Malène; Sco, Scott; Sou, South-west; Tam, Tamil Madiu; Tow, Townsville; Trê, Três Barras; Tur, Turramurra; unk, unknown; Ura, Uraidla; Val, Valencia; Viz, Vizcosa; Voe, Voeltjiesdord; Wan, Wangetti; Wat, Wattamolla; Wes, Westmead; Wie, Wiesonhof; Wil, Wilberforce; Wit, Wittenoom; Wub, Wubin; Yan, Yanxi; Zen, Zenhai.Country: ACT, Australian Capital Territory; Bra, Brazil; Chi, China; Fra, France; Ind, India; Jap, Japan; Ken, Kenya; Mal, Malaysia; Mor, Morocco; NSu, North Sumatra; Nic, Nicaragua; NSW, New South Wales (Australia); Phi, Philippines; Por, Portugal; Qld, Queensland (Australia); SA, South Australia; SAf, South Africa; Sen, Senegal; Spa, Spain; Tha, Thailand; unk, unknown; Vic, Victoria, Australia; WA, Western Australia.
NPH_313.fm Page 354 Wednesday, January 9, 2002 1:37 PM

© New Phytologist (2002) 153: 345–357 www.newphytologist.com
Research 355
America and Africa, there is no reliable record of P. tinctoriusfrom pine plantations in Australia and New Zealand (Dunstanet al., 1998). P. tinctorius does not occur in Australia, at leastnot in natural ecosystems (Bougher & Syme, 1998; Cairneyet al., 1999). P. tinctorius probably was not among the Holarticectomycorrhizal fungi brought by early settlers into Australiawith soil or pine seedlings. Later, stringent quarantine regula-tions on the importation of soils and plants probably precludedthe introduction of P. tinctorius.
Pisolithus lineages and regional floras
Several species identified in the present analysis consisted ofisolates limited to specific geographical regions (species 1, 3,4, 5), most of them confined to endemic plants (species 1, 3and 4). Species 1 is composed of isolates associated with anectomycorrhizal Caesalpinioideae of the coastal Miombo-likewoodlands of Kenya, Afzelia quanzensis Welw. (Martin et al.,1998). Species 3 consisted of isolates associated with Cistusladanifer L. (Cistaceae) in closed shrublands on clayey slate-derived soils in the western Mediterranean region (e.g. cr01;Díez et al., 2001). Isolates of species 4 associated with basophilouspines and sclerophyllous oaks in the Mediterranean region(e.g. m14 and pt03; Díez et al., 2001).
Isolates collected in pine plantations in China and Thailandand in eucalypt plantations in China did not belong toP. tinctorius, but to a related species. The success of plantationsof pine and eucalyptus in southeast Asia was probably facilit-ated by the occurrence of such compatible indigenous strainsof Pisolithus, rather than facilitated by the co-introductionof exotic ectomycorrhizal fungi on seedlings. The isolates ofthe so-called ‘southeast Asia’-type (species 5) were collectedin P. kesiya plantations in Thailand (e.g. MSN, THAI1), inP. yunnanensis plantations in China (e.g. MH143), and ineucalypt plantations in China (e.g. MH727). Specimens withsimilar sporocarp and spore morphologies have been observedin natural stands of P. kesiya and P. merkusii in Thailand,Indonesia and the Philippines, and P. yunnanensis in China(B. Dell, unpublished). We hypothesized that this speciescould correspond to P. aurantioscabrosus, a taxon found in nativeforests of Malaysia (Watling et al., 1995, 1999). We thusanalysed a sample collected under the dipterocarp Shoreamacroprera in the locality where the holotype was collectedand morphologically identified as P. aurantioscabrosus (LSSee, Forest Service, Pasoh, Malaysia). The phylogeneticanalysis (Fig. 1) showed that species 5 does not correspondto P. aurantioscabrosus.
Pisolithus lineages in Australia and their recent dispersal worldwide
Australian material from native forests accounts for all oflineage B and part of lineage A. The wide divergence of theirITS sequences (≤ 42%) (Table 2) suggests that the centre of
diversification of the genus Pisolithus may well be Australasia.The ancestral Pisolithus was probably a generalist mycorrhizalsymbiont. Allopatric speciation probably led to the radiationof this genus after the break up of Pangaea into the northernand southern landmasses of Laurasia and Gondwana in theTriasic, as suggested for other mycorrhizal genera (e.g. Tylopilluscf. in Halling, 2001).
Because eucalypts have evolved in isolation from the ecto-mycorrhizal fungal flora associated with Pinus and Quercus inthe northern hemisphere, Pisolithus tinctorius is probably notable to associate with eucalypts in silva (Bougher & Syme,1998; Cairney et al., 1999). P. marmoratus is considered thesouthern hemisphere counterpart of P. tinctorius. They aremorphologically very similar (Bougher & Syme, 1998), butP. marmoratus is associated with eucalypts, while P. tinctoriusis not. The species P. marmoratus and P. albus are very com-mon in WA (Bougher & Syme, 1998). The isolates found insclerophyllous NSW forests (LJ07 and WW01; Andersonet al., 1998, 2001) correspond to P. marmoratus (Bougher &Syme, 1998), together with 13 isolates collected in nativewoodlands and open forests of E. marginata/E. calophylla, andin plantations of E. globulus in WA.
The lineage B is composed of four major terminal clades,which represent species specifically associated with Australasianacacias (e.g. Acacia mangium) and eucalypts. The sublineage BIincludes three species (P. albus, 8 and P. microcarpus), whereasthe sublineage BII comprises species 10. The evolutionaryrelationships within the sublineage BI were not well resolved(Fig. 2b). The ITS region did not have enough parsimonyinformative characters to fully resolve the topology of this partof the tree, in terms of both branch length and bootstrapsupport. However, good support in terminal clades allowed usto delineate species boundaries. We considered species 7 to beP. albus based on morphological features. Isolates of thesespecies were collected throughout the world (e.g. India, Senegal,Spain) in eucalypt plantations. Species 8 and 10 were onlyfound in WA and might represent endemic species in con-cordance with other Pisolithus taxa, such as species 3, 4 and 5.Species 9 corresponds to the morphological species P. microcarpus,illustrated in Bougher and Syme (1998). Although nativeto Australia, isolates of this species were collected in exoticplantations worldwide (e.g. Brazil, China, Portugal, Senegal,South Africa).
The lack of significant ITS divergence among isolates fromdifferent continents within P. albus or P. microcarpus (Fig. 2b),and the significant divergence among isolates belonging tothese different species from the same geographical region (e.g.WA) (Fig. 2b; Table 2) are consistent with recent cosmopolitandissemination of these species with eucalypts through humanactivity. Australian eucalypts were introduced to southeastAsia, India, Africa, South America, South Africa and southernEurope in the last 200 yr as seeds and saplings and this facilit-ated the dispersal of Australian Pisolithus. Similarly, it has beensuggested that dissemination of the basidiomycetous yeast
NPH_313.fm Page 355 Wednesday, January 9, 2002 1:37 PM

www.newphytologist.com © New Phytologist (2002) 153: 345–357
Research356
Cryptococcus neoformans, a human pathogen, associated withEucalyptus leaves in southern California (Casadevall & Perfect,1998) and India (Chakrabarti et al., 1997), resulted from thetransplantation of Eucalyptus from Australia. The evidence heredoes not indicate the precise geographical origin of Eucalyptusdisseminated through the world, although their likely placeof origin within Australia is the east coast (Victoria, NSW,Queensland) where many commercial species of plantationeucalypts emanated. To trace the geographical origin ofPisolithus found in plantations worldwide, more isolates ofeach species from numerous geographical regions in Australiamust be analysed using multilocus genetic approaches (e.g.microsatellites) or mitochondrial DNA. The mechanisms ofdispersion facilitated by eucalypt plantations worldwide couldplace certain isolates of divergent lineages (e.g. eucalypt-associated- and Afzelia-associated Pisolithus) in close proximity,and subsequent mating among them could generate hybrids.Recent hybridization between fungal species has been reportedbetween the poplar rusts Melampsora medusae and M. occidentalis(Brasier, 2000), and between major lineages of Cryptococcusneoformans (Xu et al., 2000). However, no genetic exchange hasyet been found in silva between pine-, eucalypt- and Afzelia-associated Pisolithus in woodlands of Kenya (Martin et al., 1998)or in the Mediterranean region where natural and introducedPisolithus coexist (Díez et al., 2001).
Conclusions
The present study clearly shows that the evolutionary lineageswithin Pisolithus are related to biogeographical origin of thehosts. Each phylogenetic species of Pisolithus was confinedto hosts from one single biogeographical realm. Living insymbiotic association for a long evolutionary time in the samebiogeographical area could have lead to the acquisition of asignificant degree of host-fungus specificity. This specificitymight be strengthened by ecological factors (Molina et al., 1992),which probably accounts for the strict host–fungus associationpatterns observed in silva. The present phylogeographicalsurvey also showed that regional floras and endemic plants(e.g. Afzelia and Cistus) could act as hosts of endemic speciesof Pisolithus. We included in the present study specimensdeposited in herbaria and identified by classical taxonomy inorder to assign morphological species to the obtained phylo-genetic species. Mating tests are needed to delimit biologicalspecies, in particular which of the phylogenetic species arereproductively isolated.
Acknowledgements
We wish to thank the many field collectors and colleagues whoprovided isolates of Pisolithus : M. Abourouh, A. Ba, T. Burgess,A. Casares, C. Chamberlain, M. Duccousso, W. Dunstan,R. Duponnois, J. Garbaye, A. Hernandes, M. Honrubia,M. Ivory, L. Khabar, F. Lapeyrie, L. S. See, H. Machado,
N. Malajczuk, D. Marx, V. Lopes de Oliveira, U. Sangwanit,D. Sylvia and J. Trappe. We are grateful to I. Alexander,I. Anderson, B. Anta, N. Bougher, J. Cairney, M. Honrubia,M. Ivory, F. Lapeyrie, F. Le Tacon, J. L. Manjón, N. Malajczuk,G. Moreno, Romero de la Osa, and J. Trappe for their helpfuldiscussion over the years. Additional technical support atMU was provided by R. Tothill and M. Farbey. This work wassupported by an EU postdoctoral fellowship (Contract HPMF-CT-1999–00174) and a ‘Ramon y Cajal’ contract from theMCYT (Spain) to JD, research grants from the INRA andBureau des Ressources Génétiques to FM, and MU SpecialResearch Grants to BD. The present study is a joint projectbetween the School of Biological Sciences and Biotechnology(Murdoch University, Perth, WA, Australia) and the UMRIaM (INRA-Nancy, France).
References
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Research 25: 3389–3402.
Anderson IC, Chambers SM, Cairney JWG. 1998. Molecular determination of genetic variation in Pisolithus isolates from a defined region in New South Wales, Australia. New Phytologist 138: 151–162.
Anderson IC, Chambers SM, Cairney JWG. 2001. ITS-RFLP and ITS sequence diversity in Pisolithus from central and eastern Australia sclerophyll forests. Mycological Research 105: 1304–1312.
Bougher NL, Syme K. 1998. Fungi of Southeastern Australia. Perth, Australia: University of Western Australia Press.
Brasier C. 2000. Plant pathology. The rise of the hybrid fungi. Nature 405: 134–135.
Burgess T, Dell B, Malajczuk N. 1994. Variation in mycorrhizal development and growth stimulation by 20 Pisolithus isolates inoculated onto Eucalyptus grandis W. Hill ex Maiden. New Phytologist 127: 731–739.
Burgess T, Malajczuk N, Dell B. 1995. Variation in Pisolithus based on basidiome and basidiospore morphology, culture characteristics and analysis of polypeptides using 1D-SDS-PAGE. Mycological Research 99: 1–13.
Cairney JWG, Chambers SM, Anderson IC. 1999. Pisolithus systematic – molecular methods provide fresh insights. Mycologist 13: 31–35.
Carnero-Díaz E, Tagu D, Martin F. 1997. Ribosomal DNA internal transcribed spacers to estimate the proportion of Pisolithus tinctorius and Eucalyptus globulus RNAs in ectomycorrhiza. Applied and Environmental Microbiology 63: 840–843.
Casadevall A, Perfect JR. 1998. Cryptococcus Neoformans. Washington DC, USA: American Society for Microbiology Press.
Chakrabarti A, Jatana M, Kumar P, Chatha L, Kaushal A, Padhye AA. 1997. Isolation of Cryptococcus neoformans var. gattii from Eucalyptus camaldulensis. Indiain. Journal of Clinical Microbiology 35: 3340–3342.
Chambers SM, Cairney JWG. 1999. Pisolithus. In: Cairney JWG, Chambers SM, eds. Ectomycorrhizal fungi. Key genera in profile. Berlin, Germany: Springer-Verlag, 1–31.
Corpet F. 1988. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Research 16: 10881–10890.
Cunningham GH. 1942. The gasteromycetes of Australia and New Zealand. Dunedin, New Zealand: John McIndoe.
Díez J, Anta B, Manjón JL, Honrubia M. 2001. Genetic variability of Pisolithus isolates associated with native hosts and exotic eucalyptus in the western Mediterranean region. New Phytologist 149: 577–587.
Dunstan WA, Dell B, Malajczuk N. 1998. The diversity of ectomycorrhizal fungi associated with introduced Pinus spp. in the southern hemisphere, with particular reference to Western Australia. Mycorrhiza 8: 71–79.
NPH_313.fm Page 356 Wednesday, January 9, 2002 1:37 PM

© New Phytologist (2002) 153: 345–357 www.newphytologist.com
Research 357
Garbaye J, Delwaulle JC, Diangana D. 1988. Growth response of eucalyptus in the Congo to ectomycorrhizal inoculation. Forest Ecology and Management 24: 151–157.
Giachini AJ, Oliveira VL, Castellano MA, Trappe JM. 2000. Ectomycorrhizal fungi in Eucalyptus and Pinus plantations in southern Brazil. Mycologia 92: 1166–1177.
Gomes EA, Abreu LM, Borges AC, Araujo EF. 2000. ITS sequences and mitochondrial DNA polymorphism in Pisolithus isolates. Mycological Research 104: 911–918.
Halling RE. 2001. Co-evolution, significance and biogeography. Annals of the Missouri Botanical Garden 88: 5–13.
Jarosh M, Bresinsky A. 1999. Speciation and phylogenetic distance within Paxillus s. str. (Basidiomycetes, Boletales). Plant Biology 1: 701–706.
Jukes TH, Cantor CR. 1969. Evolution of protein molecules. In: Munro HN, ed. Mammalian protein metabolism. New York, USA: Academic Press, 21–132.
Kope HH, Fortin JA. 1990. Germination and comparative morphology of basidiospores of Pisolithus arhizus. Mycologia 82: 350–357.
Kretzer A, Li Y, Szaro T, Bruns TD. 1996. Internal transcribed spacer sequences from 38 recognized species of Suillus sensu lato: phylogenetic and taxonomic implications. Mycologia 88: 776–785.
Lamhamedi MS, Fortin JA, Kope HH, Kropp BR. 1990. Genetic variation in ectomycorrhiza formation by Pisolithus arhizus on Pinus pinaster and Pinus banksiana. New Phytologist 115: 689–697.
Martin F, Delaruelle C, Ivory M. 1998. Genetic variability in intergenic spacers of ribosomal DNA in Pisolithus isolates associated with pine, eucalyptus and Afzelia in lowland Kenyan forests. New Phytologist 139: 341–352.
Marx DH. 1977. Tree host range and world distribution of the ectomycorrhizal fungus Pisolithus tinctorius. Canadian Journal of Microbiology 23: 217–223.
Marx DH. 1981. Variability in ectomycorrhizal development and growth among isolates of Pisolithus tinctorius as affected by source, age and reisolation. Canadian Journal of Forest Research 11: 168–174.
Molina RH, Massicotte H, Trappe J. 1992. Specificity phenomena in mycorrhizal symbiosis: Community-ecological consequences and practical implications. In: Allen MF, ed. Mycorrhizal functioning. London, UK: Chapman & Hall, 357–423.
Page RDM. 1996. TREEVIEW: An application to display phylogenetic trees on personal computers. Computer Applications in the Biosciences 12: 357–358.
Sims KP, Sen R, Watling R, Jeffries P. 1999. Species and population structures of Pisolithus and Scleroderma identified by combined phenotypic and genomic marker analysis. Mycological Research 103: 449–458.
Swofford DL. 1999. PAUP*. Phylogenetic analysis using parsimony (*and other methods), Version 4.08b8. Sunderland, MA, USA: Sinauer Associates.
Taylor JW, Jacobson DJ, Kroken S, Kasuga T, Geiser DM, Hibbett DS, Fisher MC. 2000. Phylogenetic species recognition and species concepts in fungi. Fungal Genetics and Biology 31: 21–32.
Watling R, Taylor A, See LS, Sims K, Alexander IJ. 1995. A rainforest Pisolithus: its taxonomy and ecology. Nova Hedwigia 61: 417–419.
Watling R, Turnbull E, See LS. 1999. Pisolithus aurantioscabrosus Watl. (Pisolithaceae; Basidiomycota) – an expanded view. Nova Hedwigia 69: 433–437.
White TJ, Bruns T, Lee SS, Taylor J. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ, eds. PCR protocols: a guide to methods and applications. New York, USA: Academic Press, 315–322.
Wu Q-X, Mueller GM, Lutzoni FM, Huang Y-Q, Guo S-Y. 2000. Phylogenetic and biogeographical relationships of eastern Asian and eastern North America disjunct Suillus species (Fungi) as inferred from nuclear ribosomal RNA ITS sequences. Molecular Phylogenetics and Evolution 17: 37–47.
Xu JP, Vilgalys R, Mitchell TG. 2000. Multiple gene genealogies reveal recent dispersion and hybridization in the human pathogenic fungus Cryptococcus neoformans. Molecular Ecology 9: 1471–1481.
About New Phytologist
• New Phytologist is owned by a non-profit-making charitable trust dedicated to the promotion of plant science. Regular papers,Letters, Research reviews, Rapid reports and Methods papers are encouraged. Complete information is available atwww.newphytologist.com
• All the following are free – essential colour costs, 100 offprints for each article, online summaries and ToC alerts (go to thewebsite and click on Synergy)
• You can take out a personal subscription to the journal for a fraction of the institutional price. Rates start at £83 in Europe/$133in the USA & Canada for the online edition (go to the website and click on Subscriptions)
• If you have any questions, do get in touch with Central Office ([email protected]; tel +44 1524 594691) or, for a localcontact in North America, the USA Office ([email protected]; tel 865 576 5251)
NPH_313.fm Page 357 Wednesday, January 9, 2002 1:37 PM


















