
PHOTOREACTIONS ASSOCIATED WITH IN VITRO HEMOLYSIS IN ERYTHROPOIETIC PROTOPORPHYRIA
11
Phufurhemisfry ondPhofobiolngv. 197 I. Vol. 13. pp. 67-77. Pergamon Press. Printed in Great Britain PHOTOREACTIONS ASSOCIATED WITH IN VZTRO HEMOLYSIS IN ERYTHROPOIETIC PROTOPORPHYRIA” JOSEPH HSU. BERNARD D. GOLDSTEIN and LEONARD C. HARBER Departments of Dermatology and Medicine, New York University School of Medicine, New York, N.Y. 10016, U.S.A.; and the Rockefeller University, New York, N.Y. 10021, U.S.A. (Received 2 April 1970; in revised form 3 August 1970) Abstract- Photohemolysis of erythrocytes from patients with erythropoietic protoporphyria results from damage to the cell membrane following photoexcitation of the protoporphyrin molecule by 400 nm radiation. Photochemical reactions were assessed for their roles in initiating cell destruction in an in vim model in which erythrocytes from patients with erythropoietic protoporphyria were irradiated. Electron transfer properties were demonstrated using 2,3‘,6- trichloroindophenol as an electron acceptor. Photohemolysis of these abnormal cells is due to cell membrane damage initiating colloid osmotic hemolysis. Additional data are offered demon- strating that this type of photohemolysis is oxygen-dependent, associated with peroxide forma- tion, and preceded by increased osmotic fragility. An hypothesis explaining the interrelation- ship of the photochemistry of protoporphyrin with the photobiologic observation of hemolysis is offered. INTRODUCTION IN 196 1 Magnus et (I/. [ 11 defined an inherited metabolic disorder, erythropoietic protoporphyria (EPP). This condition, associated with marked cutaneous photosen- sitivity, has to date been found to occur spontaneously only in man. It has been possible, however, to photosensitize guinea pigs by intraperitoneal injections of protoporphyrin [2] and also to induce protoporphyria in other animals with specific drugs[3,4]. The naturally-occurring disease is associated with elevated protoporphyrin levels in both red blood cells (rbc) and feces. The protoporphyrin concentration in the rbc ranges from 10 to 100- fold that of normal individuals. In uitro photohemolysis of rbc obtained from these patients occurs on exposure to 400 nm radiation[5-71. This corresponds closely to the action spectrum eliciting photosensitivity in man [ 11. Previous studies have demonstrated that photohemolysis of EPP cells is associated with damage to the cell membrane following photoactivation of the protoporphyrin molecule [8- 1 11. This observation is based primarily on two measurable phenomena: [ 13 Cellular potassium (K+) loss precedes visible hemolysis and is not initially associated with hemoglobin loss [8], and (2) photohemolysis can be inhibited in spite of K+ loss when non-membrane penetrating solutes such as sucrose are present in the medium external to the rbc in place of the normal electrolytes[7,8]. Photohemolysis occurring in the dark after being initiated by 400 nm radiation closely conforms to mechanisms for other types of photohemolysis described pre- viously by Wilbrandt, Leu, and others[l2-141 as being due to “colloid osmotic hemo- lysis” in which a “hole” on the cell membrane precedes cell destruction. The size of the photochemically induced hole in EPP photohemolysis has been estimated to be 10 A or less [8, 151. *This study was supported by Grants No. ES-00288 and HD-043 I3 from the U.S. Public Health Service and by Grant No. U-1761 of the Health Research Council of the City of New York. 67
-
Upload
joseph-hsu -
Category
Documents
-
view
212 -
download
0
Transcript of PHOTOREACTIONS ASSOCIATED WITH IN VITRO HEMOLYSIS IN ERYTHROPOIETIC PROTOPORPHYRIA

Phufurhemisfry ondPhofobiolngv. 197 I . Vol. 13. pp. 67-77. Pergamon Press. Printed in Great Britain
PHOTOREACTIONS ASSOCIATED WITH IN VZTRO HEMOLYSIS IN ERYTHROPOIETIC
PROTOPORPHYRIA” JOSEPH HSU. BERNARD D. GOLDSTEIN and LEONARD C. HARBER
Departments of Dermatology and Medicine, New York University School of Medicine, New York, N.Y. 10016, U.S.A.; and the Rockefeller University, New York, N.Y. 10021, U.S.A.
(Received 2 April 1970; in revised form 3 August 1970)
Abstract- Photohemolysis of erythrocytes from patients with erythropoietic protoporphyria results from damage to the cell membrane following photoexcitation of the protoporphyrin molecule by 400 nm radiation. Photochemical reactions were assessed for their roles in initiating cell destruction in an in v i m model in which erythrocytes from patients with erythropoietic protoporphyria were irradiated. Electron transfer properties were demonstrated using 2,3‘,6- trichloroindophenol as an electron acceptor. Photohemolysis of these abnormal cells is due to cell membrane damage initiating colloid osmotic hemolysis. Additional data are offered demon- strating that this type of photohemolysis is oxygen-dependent, associated with peroxide forma- tion, and preceded by increased osmotic fragility. An hypothesis explaining the interrelation- ship of the photochemistry of protoporphyrin with the photobiologic observation of hemolysis is offered.
INTRODUCTION
IN 196 1 Magnus et (I/. [ 11 defined an inherited metabolic disorder, erythropoietic protoporphyria (EPP). This condition, associated with marked cutaneous photosen- sitivity, has to date been found to occur spontaneously only in man. It has been possible, however, to photosensitize guinea pigs by intraperitoneal injections of protoporphyrin [2] and also to induce protoporphyria in other animals with specific drugs[3,4]. The naturally-occurring disease is associated with elevated protoporphyrin levels in both red blood cells (rbc) and feces. The protoporphyrin concentration in the rbc ranges from 10 to 100- fold that of normal individuals. In uitro photohemolysis of rbc obtained from these patients occurs on exposure to 400 nm radiation[5-71. This corresponds closely to the action spectrum eliciting photosensitivity in man [ 11. Previous studies have demonstrated that photohemolysis of EPP cells is associated with damage to the cell membrane following photoactivation of the protoporphyrin molecule [8- 1 11. This observation is based primarily on two measurable phenomena: [ 13 Cellular potassium (K+) loss precedes visible hemolysis and is not initially associated with hemoglobin loss [8], and (2) photohemolysis can be inhibited in spite of K+ loss when non-membrane penetrating solutes such as sucrose are present in the medium external to the rbc in place of the normal electrolytes[7,8].
Photohemolysis occurring in the dark after being initiated by 400 nm radiation closely conforms to mechanisms for other types of photohemolysis described pre- viously by Wilbrandt, Leu, and others[l2-141 as being due to “colloid osmotic hemo- lysis” in which a “hole” on the cell membrane precedes cell destruction. The size of the photochemically induced hole in EPP photohemolysis has been estimated to be 10 A or less [8, 151.
*This study was supported by Grants No. ES-00288 and HD-043 I3 from the U.S. Public Health Service and by Grant No. U-1761 of the Health Research Council of the City of New York.
67

68 J . HSU. B. D. GOLDSTEIN and L. C. HARBER
Data reported in this paper concern the photoprocesses initiating or associated with both protoporphyrin excitation and photohemolysis of EPP rbc. The particular phases investigated included : (a) electron transfer properties of irradiated proto- porphyrin; (b) oxygen requirements for photohernolysis; (c) hydrogen peroxide forma- tion following irradiation, and (d) alterations in osmotic fragility preceding photohemolysis.
MATERIALS A N D METHODS
(a) Detection of electron transfer from protoporphyrin using 2,3',6-trichlorophenol indophenol. Electron transfer from protoporphyrin IX was detected using 2,3',6- trichloroindophenol (TCIP) as an electron acceptor [ 161. A protoporphyrin solution (2.85 ml of a 2.84 X rnM) was added to 0.15 ml of a ImM TCIP solution. The solu- tion was adjusted to pH 9-00 with NH,OH. Samples to be irradiated were placed in Beckman 3-ml quartz cuvettes (optical path Icm) and similar solutions were kept in the dark. Control solutions of the dye alone in N H 4 0 H solution and protoporphyrin in NH,OH solution were also prepared for the study in the presence and absence of light. The photon source used for irradiation consisted of a Sylvania SG-8 Sun Gun placed at a target distance of 30-0 cm. The Sun Gun emits a continuous spectrum from U.V. to i.r. Three filters were used in order to obtain the desired wavelength : a Corning heat-resistent filter No. 4-97 (5.0 mm thick) was used to cut off irradiation wavelengths below 320 nm; a Schott filter BG-I 2 (1-0 nm thick) having a transmission range from 290 nm to 650nm with a peak of 87 per cent transmission at 400nm; a I-gallon Pyrex museum jar filled with distilled water (optical path 13-8 cm) was used to absorb heat and to cut off irradiation wavelengths below 320 nrn. The Sylvania Sun Gun irradiation intensity (measured with an Eppley thermopile) was 1-55 X 104pW/cm2 at a target distance of 30.0 cm as viewed through the 3 filters specified above. The major portion of the transmitted light was in the 400-460 nm region. During the period of continuous irradiation, optical density readings at 640 nm were obtained at intervals. lrradiation was continued for 5 hr, with data taken at 30-min intervals during the first 5 hr and then 18 hr later. Appropriate controls were simultaneously irradiated.
(b) Measurement of oxygen dependence for photohemolysis of rbc from EPP patients. Erythrocytes used in this study were obtained from a 12-yr old male with cutaneous photosensitivity who was found to have a free erythrocyte protoporphyrin value of 1.2 mgll00 ml of packed rbc as determined by the method of Wranne[ 171. The rbc of an unaffected volunteer with a protoporphyrin level of 27pg/100ml of packed rbc served as acontrol. Both types of rbc were washed three times and suspended in 1 : 1000 dilution in 0.85 per cent phosphate-buffered saline, pH 7.4[8]. Fifty ml of each suspension were placed in separate gas-collecting tubes.
A 100% nitrogen atmosphere in the collecting tube containing the cell suspension was established by using the following procedure : commercial nitrogen gas (99.5%) was purified by introducing it into a two-stage 125-ml Pyrex gas-collecting tube con- taining 50 ml of alkaline pyrogallol solution to remove the remaining oxygen [ 181. I t was then passed through high-pressure Cenco rubber tubing into another 125-ml Pyrex gas-collecting tube containing 50 ml of the buffered rbc suspension obtained from the EPP patient. Rotating samples were then irradiated with a bank of 4 General Elec- tric 20-W blacklight fluorescent tubes having an irradiation intensity of 1.14 x 10JpW/ cm2 at 25.4 cm (target distance). Percentage hemolysis was determined at periodic

Photoreactions associated with in oitro hemolysis 69
time intervals by duplicate direct cell count using a hemacytometer. Photohemolysis was also assayed in two other gas environments : 20% 0, (air) and 99.5% 0,. All irra- diated samples were compared with identically treated samples not exposed to light.
(c) Measurement of hydrogen peroxide formation. The rates of hydrogen peroxidase formation in rbc from patients with EPP were measured utilizing horseradish peroxide (HRP) in a modified procedure of the technique described by Bjorksten[l9] and Fox [20]. Stock solutions of HRP ( I mg/ml in distilled water) and hydrogen peroxide ( 1 ml of 30% Hz02/99 ml distilled water) were used as standards for all assays. The rate of formation of hydrogen peroxide in this test model was established by using 1 ml of the stock H,O, diluted to 100 ml with 0.01 M phosphate buffered saline at pH 6-0. A 1% 0-dianisidine solution in methyl alcohol (0.05 ml) was added to 6.0 ml of the sub- strate from which 2.9 ml was transferred to a 1 cm cuvette. The remainder was placed in a control cuvette. At zero time, 0.1 ml of the diluted enzyme was introduced in the control cuvette and absorbance at 460 nm was recorded with a Beckman DB spectro- photometer at 15-sec intervals for 1-2 min. The rate of change per minute was then determined.
Using 6.0 ml of a 1 : 1000 suspension of rbc as a substrate, the previous model was then used to assay H,O, formation of rbc in the presence of radiation. All irradiation of cells from EPP patients and normal volunteers was performed with a bank of 4 fluorescent blacklight lamps previously described. Both types of cells received identical doses of incident light (17 1 ergs/mm2-146 X 106 ergs/mm2). Red blood cell counts were performed on aliquots of the rbc suspension undergoing photohemolysis simul- taneously with the recording of absorbance. This furnished a means of correction for the rbc contribution to the absorbance of the photohemolysis suspension of both rbc and o-dianisidine, which were also assessed in a similar fashion.
(d) Measurement of effect of light on osmotic fragility. Venous blood in ethylene- diamine tetraacetate disodium (EDTA) anticoagulant was obtained from two patients with EPP ( I -2 mg/ 100 ml of packed rbc) and from hematologically normal volunteers (30pg/100 ml of packed rbc). The rbc’s were washed three times in an isotonic phos- phate-buffered saline solution, pH 7.4, prepared according to the method of Cartwright [21]. They were then suspended in isotonic buffer at a concentration of 1 : 400. Suitable aliquots were either irradiated or kept in the dark, the latter serving as controls. All studies were performed at 24 k 1°C. The radiation used was similar to that previously described. Rbc osmotic fragility was determined [2 11 after illumination by adding aliquots of the rbc suspension to tubes containing phosphate-buffered saline at concen- trations ranging from 0.85% to 0.30% saline and also to a tube containing distilled water. The tubes were centrifuged 30 min later at 2,000 rpm in an International PR-J centrifuge and the supernatant hemoglobin measured at 540nm in a Beckman DB spectro- photometer. Percentage hemolysis was calculated by comparing the supernatant hemo- globin concentration at each saline concentration with that of the distilled water tube in which complete hemolysis occurred.
Erythrocytes from a hematologically normal volunteer were obtained in EDTA anticoagulant in order to prepare pseudo-EPP cells. The normal rbc were washed 3 times in phosphate-buffered saline, 7.4, and suspended to a concentration of I : 100 in buffer to which 100pg/IOOml protoporphyrin 1X had been added. Red blood cell absorption of protoporphyrin 1X rapidly reached a peak concentration of 800 pg/ I00 ml of packed rbc in 5 min. Part of the suspension was irradiated and the remainder

70 J . HSU, 9. D. GOLDSTEIN and L. C. HARRER
0
was kept in the dark as a control sample. Erythrocyte osmotic fragility was then deter- mined in a manner identical to that of the cells from patients with EPP.
A' P P + NH, OH
4 P P + NH, OH I
RESULTS
(a) Determination of electron transfer from protoporphyrin using 2,3 ' ,6-tri- chloroindophenol (TCIP). Electron transfer from protoporphyrin IX following illumina- tion was demonstrated using TCIP as an electron acceptor (Fig. 1). This delocalization of the .rr-orbitals from the tetrapyrrole ring is manifested in the shift of optical density noted in Curve C (unilluminated). Both curves were obtained with a solution of proto- porphyrin IX at pH 9.0 to which TCIP had been added. Illumination was the sole variable. Using protoporphyrin in an initial concentration of 2.8 X M and the radiation factors noted, the maximum effect was seen during the first 5 hr with little alteration noted during the next 18 hr. N o variations following illumination were noted when either protoporphyrin or dye was removed from the test model (Curve A, A', B, B').
(b) Measurement of oxygen dependence in photohemolysis of. cells from EPP patient. Figure 2 represents the percentage hemolysis as a function of time following irradiation. Complete photohemolysis was observed within 2 hr in both an air and a
NH, OH + DYE I .oo
E
0 (0 t
E
c s W
0 . O O
0 . 6 0
0.40
0 . 2 0 t
........ * ......................
P P + NH, OH +DYE 'r

Photoreactions associated with in oirro hemolysis 71
m A CONTROL ' -
I I I I I I I 2 4 6 8 10 12 14 16 18 20
TIME ( H O U R S )
Fig. 2. Percentage hemolysis of EPP rbc as a function of time in hours; Graph demonstrates obligatory requirement of oxygen for photohemolysis of EPP rbc (xx and . . .). Cells from normal volunteers irradiated in air (a a) as well as EPP rbc irradiated in a nitrogen environment (A A) showed negligible hemolysis. Rbc from normal volunteers irradiated in 100% nitrogen
environment also showed minimal hemolysis.
99.5% oxygen environment using rbc from a patient with EPP. Less than 15 per cent photohemolysis was noted for normal or EPP rbc when irradiated in a nitrogen environ- ment for 20 hr. These results indicate that photohemolysis is oxygen dependent and that oxygen participates in the reaction which is subsequent to the light absorption process.
(c) Measurement of hydrogen peroxide formation. Figure 3, Curve E, demonstrates hydrogen peroxide formation in the rbc obtained from a patient with EPP following illumination. This curve represents the recorded absorbance in Curve B corrected for the absorbance due to rbc concentration obtained in Curve A (Curve B-Curve A). Less amounts of H,O, formation were noted with irradiated normal rbc (Curve D), which were also corrected for the absorbance due to rbc concentration (Curve C), (Curve D-CurveC). The level of protoporphyrin IX was 27-35 pg/ I00 ml of packed rbc for normal rbc as compared to 1-2 mg/lOO ml in EPP rbc.
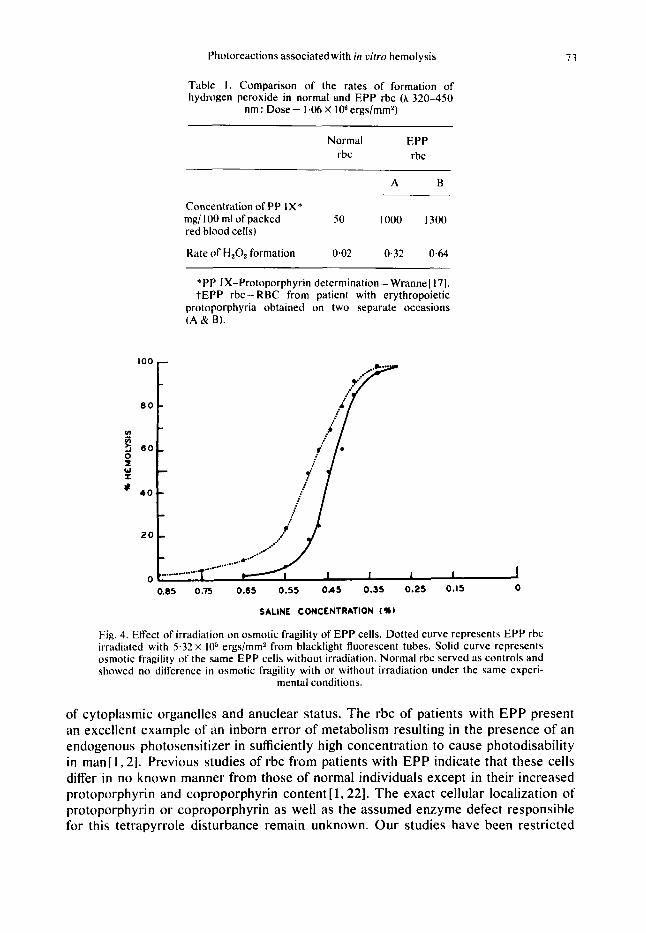
Table I represents a comparison of the rates of hydrogen peroxide formation in normal and EPP rbc. The rate of hydrogen peroxide formation of normal rbc was 0.02 nmole min, whereas those obtained from patients with EPP were 0.32-0-64 nmol min.
(d) Measurement of the effect of light on osmotic fragility. Irradiated rbc obtained from patients with EPP demonstrated an increased osmotic fragility as compared to the non-irradiated controls (Fig. 4 and Table 2). Continued exposure to light produced

72 J . HSU, B. D. GOLDSTEIN and L. C. HARBER
0.4
Om3[ 0.2 0 . I /- 0.3 - 0.4 - 0.5
0 .6
0.7
0.8
- - - -
I I I I I I I I l l 1 0.5 I I .5 2 2.5
TIME ( H O U R S 1
Fig. 3. Rates of hydrogen peroxide formation in EPP. Calculations for the rate of hydrogen peroxide were obtained by subtraction of the absorbance at 400 nm due to the presence of rbc from the total absorbance. The latter represents both the contributions of the dianisidine and rbc. Curve A represents decreasing absorbance due solely to hemolysis of EPP cells follow- ing illumination; Curve B represents the sum of hemolysis of EPP cells and hydrogen peroxide formation; Curve E indicates hydrogen peroxide formation from the EPP rbc receiving radia- tion. I t has been corrected for the loss of absorbance due to rbc undergoing photohemolysis (Curve B-CurveA); Curve C shows no hemolysis for normal rbc under blacklight illumination (1.06 X lo6 ergs/mm*); Curve D shows some hydrogen peroxide formation for normal rbc
under the same dosage of illumination.
further increases in osmotic fragility and eventually hemolysis as demonstrated in Table 2. Similar results were also obtained in a study of rbc from normal volunteers incubated in 100 pg/ 100 ml protoporphyrin IX (final concentration 800 pgl100 ml of packed rbc). These pseudo-EPP cells, when irradiated, also demonstrated an increase in osmotic fragility that was directly related to the dose of illumination (Fig. 5 ) . This was again shown to precede hemolysis. Erythrocytes from normal volunteers to which protoporphyrin 1X had not been added did not develop an increase in osmotic fragility after a 2 hr exposure.
DISCUSSION
Erythrocytes have afforded a unique model for studying the kinetics and localiza- tion of primary and secondary photobiologic reactions involving the cell membrane. This is primarily due to their ease of obtainment, stability in synthetic media, absence
Photoreactions associated with in uitro hemolysis 7 3
*PP IX-Protoporphyrin determination- Wranne[ 171. t E P P rbc - RBC from patient with erythropoietic
protoporphyria obtained on two separate occasions (A & B).
100
8 0
Y 2 6 0 0 I w I
* 40
2 0
0
Table I . Comparison of the rates of formation of hydrogen peroxide in normal and EPP rbc (A 320-450
nm : Dose - 1.06 X lo6 ergs/mm2)
Normal EPP rbc rbc
A B
Concentration of PP IX* mg/ 100 ml of packed 50 1000 1300 red blood cells)
Kate of H 2 0 2 formation 0.02 0.32 0.64
SALINE CONCENTRATION ( lb )
Fig. 4. Effect of irradiation on osmotic fragility of EPP cells. Dotted curve represents EPP rbc irradiated with 5.32 X lo5 ergs/mm2 from blacklight fluorescent tubes. Solid curve represents osmotic fragility of the same EPP cells without irradiation. Normal rbc served as controls and showed no difference in osmotic fragility with or without irradiation under the same experi-
mental conditions.
of cytoplasmic organelles and anuclear status. The rbc of patients with EPP present an excellent example of an inborn error of metabolism resulting in the presence of an endogenous photosensitizer in sufficiently high concentration to cause photodisability in man[l, 21. Previous studies of rbc from patients with EPP indicate that these cells differ in no known manner from those of normal individuals except in their increased protoporphyrin and coproporphyrin content [ 1,221. The exact cellular localization of protoporphyrin or coproporphyrin as well as the assumed enzyme defect responsible for this tetrapyrrole disturbance remain unknown. Our studies have been restricted

74 J. HSU, B. D. GOLDSTEIN and L. C. HARBER
Table 2. Effect of irradiation'! on hemolysis of EPP erythrocytes in
phosphate-buffered saline, pH 7.4
Percentage hemo- lysis at buffered
Radiation dose saline concentration ergs/mm* (0435%) (0.45%)
0 1 35 3.5 x 105 2 55
~ 3 . 2 8 x 105 22 82
Patient:W.S. 1.2 mg/l00 ml Protoporphyrin IX.
*Rbc from normal volunteers showed less than 1 per cent hemolysis at 8.28 X 105 ergs/mm*.
tG. E. Blacklight ( A 320-450nm; peak = 365 nm); Full width half maximum = 50 nm.
100 - I
8 0 - -
- 2 0 -
0.85 0.75 0.65 0.55 0.45 0.35
SALINE CONCENTRATION ( % I
Fig. 5 . Demonstration of the dose dependency of osmotic fragility in pseudo-EPP rbc. Curve A represents the average of 3 control readings of non-irradiated EPP rbc. Curves B, C and D represent the osmotic fragilities of the pseudo-EPP rbc irradiated with I .18X lo5, 2 . 3 6 ~ lo3
and 4.72 x lo5 ergs/mm2 respectively.
to protoporphyrin photokinetics primarily because previous studies of rbc with markedly elevated coproporphyrin levels, such as found in lead poisoning, have shown no evidence of in uitro photohemolysis [7,23] or clinical photosensitivity.
The data obtained following 400 nm irradiation of protoporphyrin or of cells from

Photoreactions associated with in uirro hemolysis 75
patients with EPP indicate that electron transfer occurs using the reducing dye, 2,3’,6- trichlorophenol indophenol, as the electron acceptor. Similar findings have been reported with hematoporphyrin [24] using glutathione as an electron donor and ferre- doxin as an electron acceptor.
The obligatory oxygen requirement for the photohemolysis of EPP cells (Fig. 2) indicates that the process is photodynamic. The results further show that the role of the excited oxygen is to participate in the reaction which is subsequent to the light absorption process; i.e., an active intermediary or intermediaries in the process of photodynamic hemol ysis. These findings reinforce earlier studies of normal rbc by Cook and Blum[l8] using a broad spectrum of ultraviolet radiation as an irradiation source. Their data indicated variations in oxygen dependence which were related to the spectral character of the ultraviolet radiation used. Accordingly, at least two chromo- pores were postulated to be related to photohemolysis of normal rbc; i.e., ( I ) a chromo- pore associated with non-oxygen dependent hemolysis having an action spectrum lower than 320nm and probably due to protein, and (2) a chromopore associated with an oxygen-dependent hemolysis pattern having an action spectrum in the 400 nm range, perhaps due to small quantities of protoporphyrin IX or possibly riboflavin. Cook and Blum [ 181 further noted that exceedingly high dosages of radiation were required for the oxygen-dependent process of normal rbc photohemolysis. Using lower irradiation factors in our studies, the negligible photohemolysis we observed in normal rbc is thus understandable. Our finding of marked photohemolysis occurring relatively rapidly in EPP cells whose protoporphyrin content was 1.2 mg/100 ml of packed rbc, as com- pared with negligible destruction of normal rbc, whose protoporphyrin content average 27pg/100ml of packed rbc, conforms well with the postulations of these authors of a “dose-squared/hemolysis-time relationship”[ 18,251 in a cell with a high concen- tration of chromopore.
The results of determination of hydrogen peroxide formation in the photodynamic hemolysis in EPP are in accord with those of Kasha[26,27]. His studies indicated that the singlet excited state of oxygen is sufficiently long-lived to allow time for it to enter into biologically significant chemical reactions, such as the formation of hydrogen peroxide (Fig. 3). Similar reasoning would predict that in aqueous media the presence of cell membrane lipids would also be associated with the formation of protoporphyrin peroxide and lipid peroxides. Studies presented by Ludwig et a1.[6] show that lipid peroxide formation can also be detected in EPP rbc undergoing photohemolysis. Accordingly, at least three peroxides could potentially participate in the production of lytic cell membrane lesions. Studies regarding protoporphyrin peroxide formation have not been reported to date.
The studies of irradiated rbc from patients with EPP and with normal rbc incubated in protoporphyrin IX (Figs. 4, 5 and Table 2) demonstrated an increased osmotic fragility preceding hemolysis. The results are consistent with colloid osmotic hemolysis [ 12,281 and are also compatible with membrane deformation. The alteration in osmotic fragility could be attributed to at least two mechanisms : (1) interference with active transport of cations (ATP “pump”)[29] or (2) damage to the membrane per se, pro- ducing an increased leak rate in the ce11[30]. Both of these types of primary damage would lead to intracellular sodium accumulation associated with swelling of the cell and loss of osmotic resistance. Another theoretical cause of an increased osmotic fragility is the loss of cell membrane with the resultant production of a smaller spherocytic cell

76 J . HSU, B. D. GOLDSTEIN and L. C. HARBER
which is more sensitive to osmotic stress due to its inability to swell in a hypotonic solution[3 11. This does not conform with the visual observations made[ 141. Previous studies with normal rbc undergoing photohemolysis after short-wave U.V. irradiation (320 nm) via a colloid osmotic hemolysis mechanism have also demonstrated a decreased resistance to external osmotic pressures [ 141.
I t is well appreciated that there are numerous additional facets concerning photo- hemolysis, such as enzyme destruction, repair processes and others, that have not been adequately studied to date. The diagram, Fig. 6, is offered as a current functional hypothesis to explain, in a unified fashion, the interrelationship of the photoprocesses and biologic reactions resulting in photohernolysis of EPP cells following protoporphyrin IX excitation.
0 2 PP* I" I I I I I I I !
PP PEROXIDE
LIPID PEROXIDE
NON-DEMON - STRATABLE n201
I I
I- Na' , K' ACTIVE CELL MEMBRANE TRANSPORT PUMP
-
CELLULAR K'LOSS CELLULAR Na' AND l$O INFLUX
I INCREASED OSMOTIC FRAGILITY PLUS
CELLULAR SWELLING I 1
COLLOID OSMOTIC HEMOLYSIS
Fig. 6. Functional hypothesis for biologic reactions following protoporphyrin 1X (PP) illumina- tion eventuating in cell destruction mediated through colloid osmotic hemolysis. (Key : PP* :
excited protoporphyrin; O* : excited oxygen molecule).

Photoreactions associated with in vitru hemolysis 77
R E F E R E N C E S I . I . A. Magnus, A. Jarrett, T. A. J. Prankard and C. Rimington, Lancet 2,448 ( 196 I ). 2. R. L. Baer, W. J . Miles, H. Rorsman and L. C. Harber, Dermafologica 135.5 ( I 967). 3. I. A. Magnus. D. A. Roeand L. K. Bhutani,J. Invest. Derm. 53,400( 1969). 4. S. Granick and G . Urata ,J . B i d . Chem. 238.82 I (1963). 5. L. C. Harber, A. S. Fleischer and R. L. Baer ,J .Am. Med. Assoc. 189, 191 (1964). 6. G. D. I.udwig. D. Bilheimer and L. Iverson, Clin. Res.Abstr. 15,284t1967). 7. E. Peterka. W. Runge and R. Fusaro,Archs Derm. 94,282 (1966). 8. A. S. Fleischer, I-. C. Harber. J. S. Cook and R. L. Baer,J. Invest. Derm. 46 ,505 ( 1966). 9. N. Kaplowitz, N . Javitt and L. C. Harber, New Engl. J. Med. 278, 1077 (1968).
10. C . Rimington, I . A. Magnus. E. A. Ryan and D. J. Cripps, Q. Jl. Med. 36.29 (1967). I 1. A. C. Allison. I . A. Magnus and M. R. Young, Nature 209,874 ( 1 966). 12. W. Wilbrandt,Arch.~es.Physiol.245,22(1941). 13. J . Leu. W. Wilbrandt and A. Liechti,Strahlentherap:.e 71.487 ( I 942). 14. J. S. Cook.J . Cell. Comp. Physiol. 47.55 (1,956). IS. E. M. Renkin,J. Gen. Physiol. 38,225 (1954). 16. M. Avron and A. T . Jagendorf. Archs Biochern. Biophys. 65,475 (lY56). 17. L. Wranne,Acta Pnecliut. 49 (suppl. 124). I(1960). 18. J. S. Cook and H . F. Blum. J . Cell. Comp. Physiol. 53,41 ( 1959). 19. F . Bjorksten.Actu Chem. Scand. 20. 1438 (1966). 20. L. R. Fox. W. K. Purres and H. I . Nakada, Biochern. 4,2754 ( I 965). 21. C. E. Cartwright, Diagnostic Laboratory Hematology. 4th Edn., p. 289. Crune& Stratton. N e w York
22. H. Langhof. H. Muller and L. Rietsche1,Arch. Klin. Exp. Derm., 212,506 (1961). 2 3 . R. G. Haining, T . E. Hulse and R. F. Labbe. Pruc. Soc. Exp. Biol. Med. 132,625 ( I 969). 24. K. K. Eisenstein and J. H. Wang.J. Biol. Chem. 244, (7). I720 ( 1969). 25. H. F. Blum, Photodynumic Action and Diseases Caused by Light. Reinhold, N e w York ( 1941). 26. M. Kasha, Health Phys. 15, 163 (1968). 27. M. Kasha and H. R. Rawls. Photochem. Photobiol. 7,561 (1968). 28. J. H. Jandl, Blood. 26.367 ( I 965). 29. T. A. Borgese and J. W. (3reen.J. Cell. Comp. Physiol. 59.215 (1962). 30. D. C. Tosteson, Regulation of cell volume by sodium and potassium transport In The Cellular Functions
Membrane Transport (Edited by J . H. Hoffman), pp. 3-22. Prentice-Hall, N e w York (1964). 3 I . R. I. Weed and C. F. Reed, Am. J. Med. 41,68 I ( I 966).
(196X).







![1. TITLE PAGE - vfa 20101236 - Observational... · Date: 23 May 2013 Page 8 of 48 Methoxy polyethylene glycol-epoetin beta (peg-EPO [Mircera ® ]) is another long-acting erythropoietic](https://static.fdocuments.in/doc/165x107/5cc1db8b88c9933e3a8cf5da/1-title-page-vfa-20101236-observational-date-23-may-2013-page-8-of.jpg)










