
PESTEPORCINECLASSIQUE:INFLUENCEDES … fileHALId: hal-00900934 Submittedon1Jan1977 HAL is a...
15
HAL Id: hal-00900934 https://hal.archives-ouvertes.fr/hal-00900934 Submitted on 1 Jan 1977 HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci- entific research documents, whether they are pub- lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers. L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés. PESTE PORCINE CLASSIQUE : INFLU ENCE DES ANTICORPS PASSIFS D’ORIGINE COLOSTRALE SUR LA REPONSE IMMUNITAIRE DU PORCELET CONSECUTIVE A LA VACCINATION AVEC L’AIDE DE LA SOUCHE LAPINISEE DITE ” CHINOISE ” (*) Maria Mierzejewska, S. Tereszczuk, G. Corthier, J.M. Aynaud To cite this version: Maria Mierzejewska, S. Tereszczuk, G. Corthier, J.M. Aynaud. PESTE PORCINE CLASSIQUE: IN- FLU ENCE DES ANTICORPS PASSIFS D’ORIGINE COLOSTRALE SUR LA REPONSE IMMU- NITAIRE DU PORCELET CONSECUTIVE A LA VACCINATION AVEC L’AIDE DE LA SOUCHE LAPINISEE DITE ” CHINOISE ” (*). Annales de Recherches Vétérinaires, INRA Editions, 1977, 8 (3), pp.227-240. hal-00900934
Transcript of PESTEPORCINECLASSIQUE:INFLUENCEDES … fileHALId: hal-00900934 Submittedon1Jan1977 HAL is a...

HAL Id: hal-00900934https://hal.archives-ouvertes.fr/hal-00900934
Submitted on 1 Jan 1977
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
PESTE PORCINE CLASSIQUE : INFLU ENCE DESANTICORPS PASSIFS D’ORIGINE COLOSTRALESUR LA REPONSE IMMUNITAIRE DU PORCELETCONSECUTIVE A LA VACCINATION AVEC L’AIDEDE LA SOUCHE LAPINISEE DITE ” CHINOISE ” (*)
Maria Mierzejewska, S. Tereszczuk, G. Corthier, J.M. Aynaud
To cite this version:Maria Mierzejewska, S. Tereszczuk, G. Corthier, J.M. Aynaud. PESTE PORCINE CLASSIQUE : IN-FLU ENCE DES ANTICORPS PASSIFS D’ORIGINE COLOSTRALE SUR LA REPONSE IMMU-NITAIRE DU PORCELET CONSECUTIVE A LA VACCINATION AVEC L’AIDE DE LA SOUCHELAPINISEE DITE ” CHINOISE ” (*). Annales de Recherches Vétérinaires, INRA Editions, 1977, 8(3), pp.227-240. �hal-00900934�

PESTE PORCINE CLASSIQUE : INFLU ENCE DES ANTICORPS PASSIFSD’ORIGINE COLOSTRALE SUR LA REPONSE IMMUNITAIRE
DU PORCELET CONSECUTIVE A LA VACCINATION AVEC L’AIDEDE LA SOUCHE LAPINISEE DITE « CHINOISE » (*)
Maria MIERZEJEWSKA, S. TERESZCZUK, G. CORTHIER J.M. AYNAUD
(1 J Institut Vétérinaire, Laboratoire de Pathologie Porcine, AI Partyzantow 57, PULAWY (Pologne)12J I.N.R.A., Laboratoire de Pathologie Porcine, 78850 THIVERVAL-GRIGNON (France)
Summary
HOG CHOLERA VIRUS : INFLUENCE OF COLOSTRAL PASSIVE ANTIBODY ON IMMUNERESPONSE OF PIG FOLLOWING VACCINATION WITH THE RABBIT ADAPTED CHINESESTRAIN. - Using the rabbit adapted chinese strain of Hog cholera, active immunizationof piglets having passive colostral antibodies was studied. 65 piglets born from 11 sowswere used. Concerning sows, vaccination was performed 5-6 months and 1 month beforeservice (3 sows), 30 days (2 sows) and 60 days (3 sows) after service. Divided in 5 lots,piglets were vaccinated at 4 different periods after birth (15, 30, 60 and 90 days).Hog cholera immunity was determined for each animal by means of kinetic of serum neutra-
lizing antibodies and resistance to virulent challenge performed 5 months after birth. Highlevels of neutralizing antibodies were observed in serum of each vaccinated sow at thetime of farrowing. In piglets having ingested low quantities of colostrum, vaccination in-duces a good antigenic stimulation characterized by a normal humoral immune responseand challenge resistance. But in piglets having ingested a normal quantities of colostrum,colostral passive antibodies have a partial or complete suppressive effect on primary im-mune response which is characterized by a delay in serum antibodies formation and by alow level at the time of challenge. According the conditions of sows vaccination, differen-ces were observed in the properties of colostral passive antibodies (intensity of suppressiveeffect on active immune response, in vitro &dquo; avidity &dquo; for Hog cholera virus, mean valueof half-life) present in piglets serum. On practical aspect, vaccination with the chinesestrain becomes fully effective in piglets having passive immunity when they are 30-60 daysold.
Introduction.
L’ingestion du colostrum (Porter 1975,Smith et ai. 1975), permet au porcelet d’ac-quérir passivement une immunité sériquecomparable à celle de sa mère par son in-
tensité, sa nature et sa spécificité. Les anti-
corps maternels présents dans le sérum du
jeune porcelet peuvent persister jusqu’à 14semaines selon la quantité initiale. Leur demi-vie est de 6-22 jours selon les classes d’im-
munoglobulines (Coggins 1964, Loan 1966,
(*) Ce travail expérimental est le résultat de la collaboration entre les deux laboratoires dans lecadre des échanges culturels franco-polonais.
:r

Curtis 1973, Launais 1976). On sait que l’im-munité passive exerce un effet inhibiteur surle développement de l’immunité active in-duite normalement à la suite de l’injectionde l’antigène (Uhr 1968, Lang 1969, Pierce
1969, Feldmann 1970, Liew 1972, Lieper 1975).Rouze et al. (1975) ont récemment étudiél’effet de l’immunité passive sur l’immunisa-tion du porcelet induite à l’aide d’un anti-
gène inerte : le lysozyme, Dans le cas del’influenza du porc, le problème a été abordépar Easterday 1972 et Renshaw 1975. Lamise en oeuvre de la vaccination contre la
peste porcine chez le jeune porcelet né demère immune, se heurte au problème del’immunité passive d’origine colostrale (Cog-gins 1962-1964, Aiken 1964). L’étude des con-ditions de l’immunisation du jeune porceletcontre la peste porcine en présence des
anticorps maternels a fait l’objet de nom-breux travaux : Lin 1969, Sasahara 1969 et
Hanaki 1972 ont examiné le problème dansle cas de la souche G.P.E. La vaccinationà l’aide de la souche « Thiverval » a étéétudiée par Corthier 1975-1976 et par Lau-nais 1975-1976. Enfin, le cas de la souche
lapinisée dite « chinoise a été abordé suc-cessivement par Zuffa 1967, Bran 1969, Olah1969, Precausta 1975, Goret 1976, Terpstra1976.Les travaux réalisés avec la souche G.P.E.
ou avec la souche a Thiverval » ont permisde préciser l’influence des anticorps passifsd’origine colostrale sur la cinétique d’appa-rition des anticorps actifs produits à lasuite de la vaccination. L’inhibition observée
dépendrait de l’intensité de l’immunité pas-sive présente chez le jeune au moment dela vaccination : ce paramètre serait lui-mêmeétroitement dépendant du niveau de l’immu-nité maternelle au moment de la mise
bas, de la quantité de colostrum ingéré et del’âge du porcelet au moment de la vaccina-tion.
En revanche, dans le cas de la souche« chinoise », seuls Precausta 1975 et Terpstra1976 ont abordé le problème dans cette opti-que. Les recherches entreprises par les au-tres auteurs (Zuffa 1967, Bran 1969, Olah1969) ont eu seulement pour objet de déter-miner l’âge à partir duquel on peut vaccinerefficacement le porcelet né de mère immune.
Les expériences présentées dans ce rap-port ont eu pour but d’étudier Ies conditionsde l’immunisation active des porcelets avecla souche « chinoise » sous le couvert d’uneimmunité maternelle d’intensité variable se-
lon les portées. L’immunité anti peste por-cine a été étudiée au cours de la réponseprimaire et au cours de la réponse secon-daire chez chacun des animaux par la ciné-
tique d’apparition des anticorps neutralisantssériques et par la résistance à l’infectionvirulente d’épreuve.
1. Matériel et méthodes.
1.1. Le vaccin anti peste porcine ;
Le vaccin a Lapest » préparé et produit àPulawy à partir de la souche « chinoise », a
été utilisé pour vacciner les truies et leurs
porcelets.
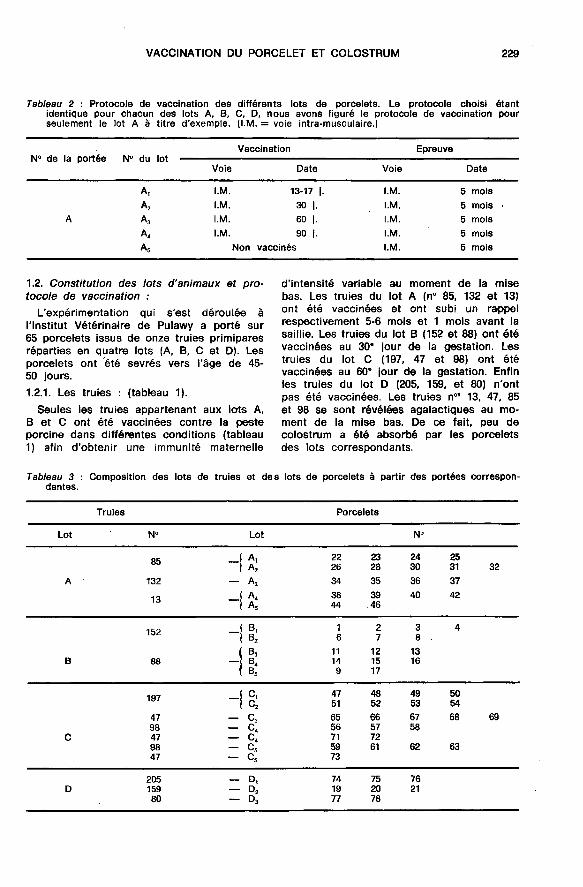
1.2. Constitution des lots d’animaux et pro-tocole de vaccination :
L’expérimentation qui s’est déroulée àl’Institut Vétérinaire de Pulawy a porté sur
65 porcelets issus de onze truies primiparesréparties en quatre lots (A, B, C et D). Lesporcelets ont ’été sevrés vers l’âge de 45-50 jours.1.2.1. Les truies : (tableau 1).
Seules les truies appartenant aux lots A,B et C ont été vaccinées contre la pesteporcine dans différentes conditions (tableau1) afin d’obtenir une immunité maternelle
d’intensité variable au moment de la misebas. Les truies du lot A (n° 85, 132 et 13)ont été vaccinées et ont subi un rappelrespectivement 5-6 mois et 1 mois avant lasaillie. Les truies du lot B (152 et 88) ont étévaccinées au 30e jour de la gestation. Lestruies du lot C (197, 47 et 98) ont étévaccinées au 60° jour de la gestation. Enfinles truies du lot D (205, 159, et 80) n’ont
pas été vaccinées. Les truies n’l 13, 47, 85et 98 se sont révélées agalactiques au mo-ment de la mise bas. De ce fait, peu decolostrum a été absorbé par les porceletsdes lots correspondants.
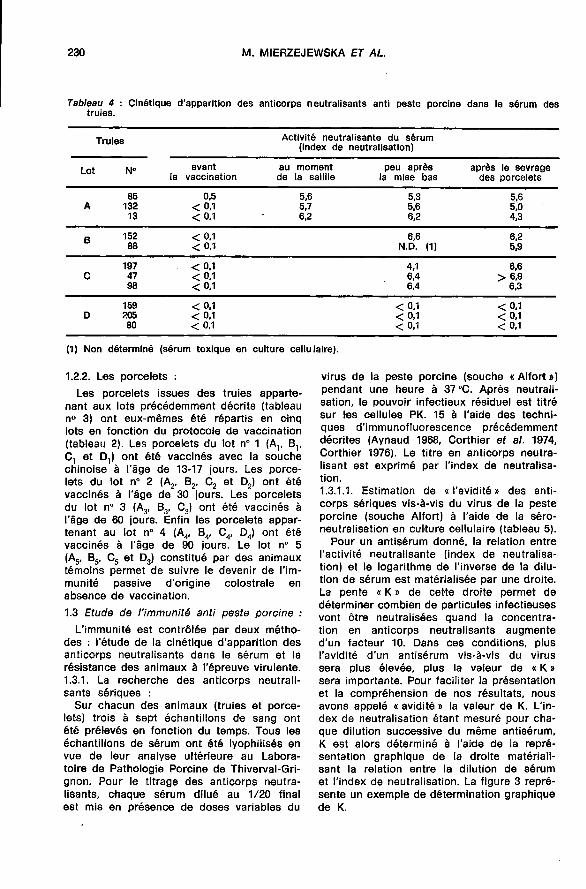
1.2.2. Les poroelets :Les porcelets issues des truies apparte-
nant aux lots précédemment décrits (tableaun° 3) ont eux-mêmes été répartis en cinqlots en fonction du protocole de vaccination(tableau 2). Les porcelets du lot n° 1 (AI, Bl’Ci et Dy ont été vaccinés avec la souchechinoise à l’âge de 13-17 jours. Les porce-lets du lot n° 2 (A2, B2, C2 et D2) ont étévaccinés à l’âge de 30 jours. Les porceletsdu lot n° 3 (A3, B3’ C3) ont été vaccinés àl’âge de 60 jours. Enfin les porcelets appar-tenant au lot n° 4 (A4, B4’ C,, D4) ont étévaccinés à l’âge de 90 jours. Le lot n° 5
(A5, B5, C5 et D3) constitué par des animauxtémoins permet de suivre le devenir de l’im-munité passive d’origine colostrale en
absence de vaccination.
1.3 Etude de l’immunité anti peste porcine :L’immunité est contrôlée par deux métho-
des : l’étude de la cinétique d’apparition desanticorps neutralisants dans le sérum et larésistance des animaux à l’épreuve virulente.1.3.1. La recherche des anticorps neutrali-sants sériques :Sur chacun des animaux (truies et porce-
lets) trois à sept échantillons de sang ont
été prélevés en fonction du temps. Tous leséchantillons de sérum ont été lyophilisés envue de leur analyse ultérieure au Labora-toire de Pathologie Porcine de Thiverval-Gri-gnon. Pour le titrage des anticorps neutra-
lisants, chaque sérum dilué au 1/20 finalest mis en présence de doses variables du
virus de la peste porcine (souche « Alfort »)pendant une heure à 37 °C. Après neutrali-
sation, le pouvoir infectieux résiduel est titrésur les cellules PK. 15 à l’aide des techni-
ques d’immunofluorescence précédemmentdécrites (Aynaud 1968, Corthier et al. 1974,Corthier 1976). Le titre en anticorps neutra-lisant est exprimé par l’index de neutralisa-tion.1.3.1.1. Estimation de « l’avidité des anti-
corps sériques vis-à-vis du virus de la pesteporcine (souche Alfort) à l’aide de la séro-neutralisation en culture cellulaire (tableau 5).Pour un antisérum donné, la relation entre
l’activité neutralisante (index de neutralisa-
tion) et le logarithme de l’inverse de la dilu-tion de sérum est matérialisée par une droite.La pente « K » de cette droite permet dedéterminer combien de particules infectieusesvont être neutralisées quand la concentra-
tion en anticorps neutralisants augmented’un facteur 10. Dans ces conditions, plusl’avidité d’un antisérum vis-à-vis du virussera plus élevée, plus la valeur de « K »
sera importante. Pour faciliter la présentationet la compréhension de nos résultats, nousavons appelé « avidité u la valeur de K. L’in-dex de neutralisation étant mesuré pour cha-
que dilution successive du même antisérum,K est alors déterminé à l’aide de la repré-sentation graphique de la droite matériali-sant la relation entre la dilution de sérumet l’index de neutralisation. La figure 3 repré-sente un exemple de détermination graphiquede K.
1.3.1.2. Détermination de la demi-vie des
anticorps passifs d’origine maternelle.Le temps au bout duquel la concentration
en anticorps maternels a diminué de 50 %(soit une variation du logarithme de la dilu-tion en anticorps (log. D) égale à 0,3) carac-térise la demi-vie.
Les valeurs successives de « K » présen-tées dans le tableau 3 permettent de prévoirles variations de neutralisation du sérumquand le logarithme de la dilution en anti-
corps (log. D) diminue de 1. La demie-viesera déterminée par le temps au bout duquella valeur de l’index de neutralisation aura
varié de K/3,3. Les valeurs de K (tableau 5)et les courbes (figure 1) ont permis d’éta!blirainsi la demi-vie moyenne des anticorpsmaternels pour chacun des différents lots deporcelets nés de mères immunes.
1.3.2. Résistance à l’infection virulente
d’épreuve :La souche pathogène « Washington » (sang
virul!ent) a été injectée par voie intramuscu-laire à la dose de 1 mi à chacun des 65 por-celets, âgés de cinq mois environ. Les
signes cliniques, l’hyperthermie, la mortalitéet les lésions sont relevés.
2. Résultats.
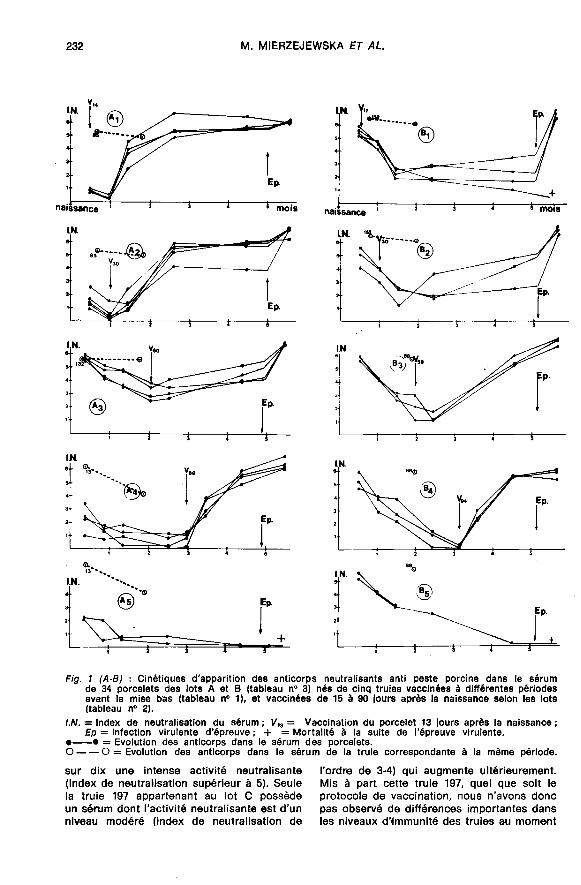
2.1. Porcelets nés de truies non immunisées(figure 2).Les lots D,, D2 et D3 étaient constitués
.,,
des porcelets nés de trubes non vaccinées(no 80, 159 et 205). Les sérums de ces truiesainsi que ceux de leurs porcelets avant la
vaccination sont totalement dépourvus d’ac-tivité neutralisante anti peste porcine. Dansces conditions, pratiquée à l’âge de 17 oude 32 jours, la vaccination induit rapidementune réponse immunitaire caractérisée parune intense production d’anticorps neutrali-sants sériques dont le taux maximum atteintau bout de six semaines, se maintient à unniveau constant jusqu’à cinq-six mois. A
cette date, soumis à une épreuve virulente,tous les animaux vaccinés résistent à lamaladie tandis que les animaux témoins nonvaccinés (lot D3), succombent dans les dé-lais habituels.
2.2. Porcelets nés de truies immunisées :
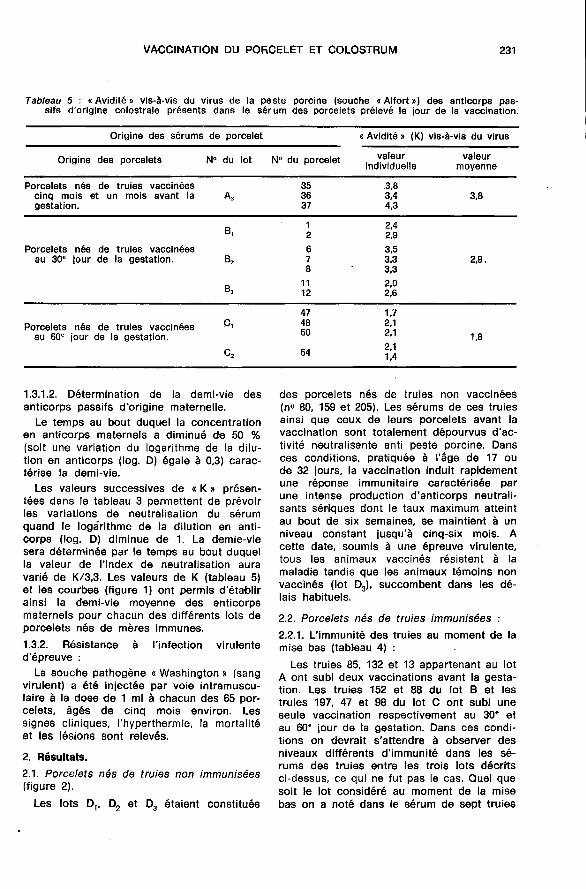
2.2.1. L’immunité des truies au moment de lamise bas (tableau 4) : -
Les truies 85, 132 et 13 appartenant au lotA ont subi deux vaccinations avant la gesta-tion. Les truies 152 et 88 du lot B et lestruies 197, 47 et 98 du lot C ont subi uneseule vaccination respectivement au 30’ et
au 60’ jour de la gestation. Dans ces condi-tions on devrait s’attendre à observer desniveaux différents d’immunité dans les sé-rums des truies entre les trois lots décritsci-dessus, ce qui ne fut pas le cas. Quel quesoit le lot considéré au moment de la misebas on a noté dans le sérum de sept truies
sur dix une intense activité neutralisante
(index de neutral,isation supérieur à 5). Seulela truie 197 appartenant au lot C possèdeun sérum dont l’activité neutralisante est d’unniveau modéré (index de neutralisation de
l’ordre de 3-4) qui augmente ultérieurement.Mis à part cette truie 197, quel que soit le
protocole de vaccination, nous n’avons doncpas observé de différences importantes dansles niveaux d’immunité des truies au moment

de la mise bas. Il faut noter enfin que l’im-munité post-vaccinale se maintenait de façonconstante à un niveau élevé tout au longde la période d’observation qui porte prèsde dix mois dans le cas du lot A par exem-ple.
2.2.2. L’immunisation active des porcelets nésde mères immunes :
L’activité neutralisante du sérum des por-celets lors du premier prélèvement effectuévers l’âge de 12-17 jours résultait de l’immu-nité passive d’origine colostrale. Si l’on con-sidère les niveaux respectifs de l’activité .neutralisante du sérum des truies et du sé-rum des porcelets dans les premiers joursde la vie, on constate les faits importantssuivants : les sérums des truies 88, 132,

152 et 197 présentaient à ce moment un titreélevé en anticorps (index de neutrali-sation :5-6) très voisin sinon identique à celui dusérum de leurs porcelets. En revanche, lessérums des porcelets nés des truies 13, 47,85 et 98 présentaient une faible activité neu-tralisante (index de neutralisation : 1-3) con-trastant avec la forte activité anticorps obser-vée dans le- sérum des mères respectives(index de neutralisation : 5-6). Dans ce der-nier cas la situation était anormale. Elle étaitla conséquence d’une ingestion insuffisantede colostrum par les porcelets et ceci enraison de l’agalaxie présentée par les quatretruies correspondantes.En raison des perturbations provoquées
dans nos résultats expérimentaux par l’aga-laxie chez quatre truies et en vue d’unemeilleure interprétation, nous avons jugé pré-férable de considérer surtout les animaux
ayant ingéré une quantité normale de colos-trum, c’est-à-dire tes porcelets des lots A.,BI’ B2, B3’ Ci et C2 issus des truies 88, 132,152 et 197. L’immunité passive d’origine co-lostrale diminuait lentement pour disparaîtrecomplètement chez les témoins non vaccinésà des dates variables selon le niveau d’anti-
corps ini2ialement observé.
2.2.2.1. Caractéristiques de la réponse pri-maire consécutive à la vaccination.- Chez les porcelets ayant reçu une quan-
tité insuffisante de colostrum (lots A,, A2,A4’ B4’ C3 et C4), pratiquée à un âge où les
anticorps maternels résiduels ont disparu, lavaccination se traduisait au niveau du sérum
par le développement d’une réponse immu-nitaire semblable à celle précédemmentobservée chez les porcelets nés de mèresnon immunes (4ots D).- Chez les porcelets ayant ingéré une
quantité normale de colostrum, l’immunité
passive d’origine maternelle, exerçai2 un effetinhibiteur sur la réponse primaire qui se
traduit par un retard dans l’apparition desanticorps et par un niveau plus faible au
moment de l’épreuve virulente (figure 1).Par ailleurs, l’immun!ité passive semblait
posséder des qualités différentes qui furent
mises en évidence au moyen de la compa-raison des sérums des porcelets des lots A,B et C et ceci grâce à l’étude des trois cri-tères suivants :- L’intensité de l’effet inhibiteur sur la
réponse primaire (mesurée par la productiondes anticorps sériques) à la suite de la vac-cination (figure 2).

- L’ « avidité » in vitro des anticorps pas-sifs vis-à-BAs du virus de la peste porcine(souche virulente Alfort) [tableau 5].- La demi-vie moyenne des anticorps
passifs.2.2.2.1.1. L’intensité de l’effet inhibiteur sur
la réponse primaire :Considérons les animaux à l’âge de 5 mois
environ appartenant respectivement aux lots
A3’ aux lots BI’ B2 et B3 et aux lots Ci et C2.L’immunité active (activité neutralisan2e du
sérum) observée chez ces animaux peut êtreconsidérée comme la résultante de l’influenoeinhi-bitrice des anticorps passifs sur la ré-
ponse primaire induite à la suite de la vacci-nation.
L’analyse statistique montre qu’il existeune étroite corrélation ( P< 0,001) entre l’in-dex de neutralisation du sérum au momentde la vaccination et celui observé chez cesmêmes animaux vers l’âge de 5 mois (figuren° 2). La droite obtenue peut être caractériséepar son intersection avec l’axe des ordon-nées (anticorps passifs) qui détermine ainsile niveau minimum d’anticorps passifs(LN.p.) capable d’inhiber totalement la pro-duction active d’anticorps à la suite de la
vaccination. La valeur de l’I.N.p. est faible
(LN:p. = 3,0) pour les porcelets nés de truiesvaccinées au 60. jour de la gestation (lotsci et C2). En revanche, LN.P. est élevé(LN.p. = 9,0) pour Ies porcelets nés de truiesdes lots A et B.
Nos résultats mettent en relief deux pointsintéressants :- L’intensité de la réponse primaire me-
surée vers l’âge de 5 mois est inversementproportionnelle au niveau des anticorps pas-sifs présents au moment de la vaccination.- Les anticorps passifs présents dans le
sérum des porcelets des lots Ci et C2 sont
plus inhibiteurs sur la réponse primaire queles anticorps passifs présents dans le sérumdes porcelets des lots A3’ BI, B2 et B,, End’autres termes, pratiquée 55 jours avant lamise bas, la vaccination avec la souche « chi-noise » induit chez la truie une immunité
qui après transmission passive par Ie colos-
trum, exerce une intense activité inhibitricesur le développement de la réponse primaireconsécutive à la vaccination précoce chezIe porcelet.Sur Ie plan pratique, nos résultats sug-
gèrent que l’intensité de l’effet inhibiteur desanticorps passifs sur la réponse primaire
serait liée dans une certaine mesure aux
conditions de la vaccination chez la truie.
2.2.2.1.2. L’ « avidité » in vitro des anticorpspassifs d’origine colostrale vis-à-vis du virus,étudiée à t’aide de la sProneutratisation enculture cellulaire :A une constante près « l’avidité » des anti-
corps passifs peut être estimée in vitro en
étudiant la relation existant entre l’index de
neutralisation et le logarithme de la dilutionde sérum, c’est-à-dire la concentration en
anticorps. La pente K de cette droite (figure3) qui est fonction de « l’avidité » des anti-
corps, exprime la quantité de particules in-fectieuses neutralisées quand la concentra-tion en anticorps dans le mélange réactionnelin vitro augmente de 10. Dans ces conditionsK est propor2ionnel à « l’avidité » des anti-
corps passifs. Pour le lot A3 d’une part, B,,B2’ B3 d’autre part et CI, C2 en dernier lieu,les valeurs moyennes de K sont respecti-vement de 3,8, 2,9 et 1,8 (tableau 5). Les
différences observées sont significativement

différentes. En d’autres termes, les conditionsde la vaccination en fonction du temps parrapport à la date de la mise bas exercent
une influence sur les qualités des immuno-globulines anti peste porcine transmises parle colostrum et en particulier leur « avidité »
vis-à-vis du virus.2.2.2.1.3. La demi-vie moyenne des anticorpspassifs :Chez les porcelets des lots B,, B2 et B3
d’une part et des lots Cl, C2 d’autre part,les anticorps passifs d’origine maternelle pré-sentent respectivement une demi-vie moyen-ne de 10 et 6 jours. Ces résultats suggèrentl’existence de classes différentes d’immuno-globulines dans ies sérums des lots consi-
dérés.
2.2.2.2. Caractéristiques de la réponse secon-daire consécutive à l’infection virulente
d’épreuve :Mis à part cinq animaux des lots Bl’ Ci
et C2, tous les poroelets vaccinés ont résistéà I’infection viruiente en présentant pourcertains (lots Ci, BI et B2) une productiond’anticorps précoce et intense typique d’uneréponse de type secondaire (tableau 6 et
figure 1). En revanche, tous les animaux nonvaccinés (lots Ay Bs et C,) ont succombédans les délais habituels.
3. Discussion.
3.1. Après la mise bas, le sérum des truiesvaccinées dans différentes conditions pré-sente une activité neutr,alisante élevée (in-dex de neutralisation compris entre 4 et 7).Peu après la naissance, on devrait donc s’at-tendre à observer dans le sang des porceletsdes valeurs voisines de celles des truies
correspondantes. C’est le cas seulement des
portées issues des truies nl’ 132, 197, 152
et 88. Chez les quatre autres portées (issues
des truies n’l 85, 13, 47 et 98) le niveau des
anticorps neutralisants est anormalement bascomparé à celui des truies correspondantes.Résultant de l’agalaxie, ce phénomène sou-ligne l’importance de l’ingestion d’une quan-tité normale de colostrum et également 1’,in-térêt de recourir à la mesure de l’activiténeutralisante des sérums pour rechercheravec précision les niveaux respectifs d’anti-corps anti peste porcine chez la truie et ses
porcelets.3.2. Chez les porcelets nés de mères immu-nes et possédant une forte immunité passive,on constate que l’activité neutralisante d’ori-gine maternelle persiste pendant 3-4 mois
après la naissance (lot B,). Ces résultatsconfirment ceux précédemment obtenus avecla .souehe « Thiverval » (Corthier 1975-1976,Launais 1975-1976) et soulignent à nouveau
l’intérêt de pratiquer l’épreuve virulente vers4-5 mois, c’est-à=dire à un âge où toute
l’activité neutralisante d’origine maternelle a
totalement disparu.3.3. L’activité neutralisante présente dans lesérum des porcelets peu après la naissance,est une fonction directe de l’immunité dela truie au moment de la mise bas. C’est
pourquoi les différences « d‘avidité quenous observons au cours de l’étude dessérums des porcelets nés des truies vacci-
nées 85 jours (lots B,, B2’ B3) ou 55 joursavant la mise bas (lots 1, et C2) peuventêtre interprétées comme résultant d’une dif-férence dans l’état immunitaire des truies.Eisen et Siskind 1964, Walker et Siskind
1968, et Karush 1970, ont démontré avec
d’autres systèmes (anticorps anti D.N.P.) queles anticorps produits en début d’immuni-sation étaient moins affins que ceux produitsen fin d’immunisation. De même, dans notre
système, nous observons que les anticorps

passifs présents dans les sérums des por-celets nés de truies en début d’immunisation(lots Bl’ B2’ B3) présentent une « avidité »moindre (K = 1,8) par rapport aux sérumsdes porcelets nés de truies vaccinées au
30e jour de la gestation (lots Cl et C2) pourlesquels « l’avidité » est plus élevée (K = 2,9).Il faut noter enfin que les anticorps passifsprésents dans le sérum des porcelets dulot A3 (truies vaccinées six mois avant la
gestation) possèdent une avidité plus élevée(K = 3,8).Nous observons des différences importan-
tes dans la valeur de la demi-vie moyennedes anticorps passifs d’origine maternelle
présents dans le sérum des porcelets appar-tenant d’une part aux lots B,, B2 et B3 ’(demi-vie moyenne : dix jours) et d’autre part auxlots Cl et C2 (demi-vie moyenne : six jours).Comme cela a été démontré dans d’autres
systèmes (Webster 1968), les différencesainsi observées dans les propriétés des anti-corps passifs pourraient refléter leur appar-tenance à des classes d’immunoglobulinesrespectivement différentes.
L’influence des différentes classes d’immu-noglobulines sur la régulation de la réponseprimaire semble variable selon les systèmesétudiés. Dans le cas des anticorps anti DNPil a été démontré que l’effet immunosuppres-seur des anticorps passifs est lié à l’intensitéde leur affinité pour l’a,n,tigène et à la classed’immunoglobuline à laquelle ils appartien-nent. Les anticorps prélevés en fin d’immu-nisation appartiennent surtout aux immuno-globulines de la classe G, et sont caractérisés
par une affinité plus élevée (Eisen et Siskind1964, Walker et Siskind 1968) et par un effetinhibiteur plus marqué sur la formation desanticorps actifs (Walker et Siskind 1968).Dans 4e cas du phage <& X 174 ou dans lecas du DNP, Finkelstein et Uhr (1964) et
Moller et Wigzell (1965) ont montré que les
anticorps passifs 7 S sont plus immunosup-pressifs que les anticorps passifs 19 S.Dans notre système, nous observons au
contraire que les anticorps passifs exercentun effet inhibiteur sur Ie développement dela réponse primaire consécutive à la valcci-nation, apparemment inversement proportion-nel à leur « avidité » pour le virus (tableau 7).Réalisés avec un antigène vivant dont la
multiplication dans l’organisme est néces-saire à l’induction de l’immunité, nos résul-tats sont apparemment en contradiction avecles travaux des auteurs précédemment cités.Mais dans les systèmes utilisés par ces der-niers, il faut noter que les antigènes étaientconstitués par des antigènes inertes tels quele DNP, les globules rouges de mouton ou le
bactériophage <& X 174, antigènes incapablesde ses multiplier chez 4e lapin, la souris oule cobaye. On sait que les caractères de la
réponse immunitaire sont différents selon
que l’antigène est inerte ou capable de répli-cation dans l’organisme. Dans ces conditions,on peut penser que la nature de l’antigèneutilisé doit vraisemblablement constituer unparamètre important dans l’induction de l’im-munisation active en présence d’anticorpspassifs. Ce qui pourrait expliquer les diffé-rences observées dans notre cas.

Sur un plan pratique, dans le cas de lasouche « chinoise », nos résultats expérimen-taux suggèrent que la vaccination de latruie peu de temps avant la mise bas pour-rait entraîner le transfert aux porcelets d’uneimmunité d’intensité modérée, mais en re-
vanche, capable d’inhiber plus fortement la
réponse primaire consécutive à la vaccina-tion.
3.4. Chez les porcelets nés de truies non
immunes, l’âge ne semble pas avoir d’in-fluence sur Ia réponse immunitaire dans nosconditions expérimentales. Vaccinés à l’âgede 15, 30, 60 ou 90 jours, les animaux déve-loppent une excellente réponse immunitaire.Concernant l’âge optimal à partir duquel ildevient possible de vacciner les porceletsnés de mères immunes, divers auteurs ontapporté des précisions. Dans une expérienceportant sur 103 porcelets âgés de 1 à 90
jours, Olah (1969), avait recherché l’influencede l’immunité passive sur le développementde l’immunisation active déterminée par larésistance à l’épreuve virulente. Cet auteuravait ainsi montré qu’il fallait attendre l’âgede deux mois pour pouvoir vacciner effica-cement les porcelets avec la souche chi-noise « Suvac ». Dans une expérience por-tant sur 143 porcelets nés de truies vacci-nées avec la souche x chinoise n au momentde la saillie, et se basant uniquement sur leniveau maximum des anticorps sériquesaprès la vaccination, Terpstra, 1976, montrequ’il faut attendre l’âge de 70-76 jours pourobtenir une vaccination efficace chez 80-90 %des animaux. Selon Precausta, 1975, en rai-son de la sensibilité de la souche chinoise
aux anticorps spécifiques, la vaccination des
porcelets issus de mères immunes ne peutintervenir dans le troisième mois de la viedu porcelet. Dans le cas de la souche « Thi-vervai », et se basant sur la résistance à
l’épreuve virulente, Launais et al. (1975, 1976)ont démontré qu’il était possible de vaccinerefficacement les porcelets à partir de l’âgede 35 jours. Les expériences présentéesdans ce rapport, ne nous permettent pas deconclure de façon précise dans ce domaine,en raison du faible nombre d’animaux et sur-tout de l’hétérogénéité des réponses chez
quatre portées de porcelet, compte tenu del’agalaxie présentée chez les quatre truies
correspondantes. Il semble cependant que lavaccination s’avère efficace à partir d’une
période située entre le 30e et le 60e jouraprès la naissance. De nouvelles expérimen-tations portant sur un plus grand nombrede portées seront nécessaires pour précisercette période.Quoi qu&dquo;il en soit, l’étude de la cinétique
d’apparition des anticorps chez chacun desporcelets a eu le grand intérêt de rendre
compte de façon précise dans le cas de la
souche « chinoise », des conditions de l’in-
fluence inhibitrice des anticorps colostrauxsur la réponse primaire au niveau humoral
(tableau 7).
Accepté pour publication en avril 1977.
Remerciements.
Nous remercions Chantai Galicher, Jac-
queline Gelfi et Elisabeth Petit pour leur effi-cace collaboration technique.
Résumé
Les expériences présentées dans ce travail ont eu pour but l’étude des conditions de l’im-munisation active des porcelets avec la souche lapinisée dite chinoise du virus de la
peste porcine classique en présence d’immunité passive d’origine colostrale d’intensitévariable selon les portées. L’expérimentation a porté sur 65 porcelets issus de onze truiesprimipares. Chez tes truies, la vaccination a été pratiquée dans les conditions suivantes :trois truies ont été vaccinées et ont subi un rappel respectivement 5-6 mois et un moisavant la saillie, deux et trois truies ont été vaccinées respectivement au 30’ et au 60e jourde la gestation. Enfin trois truies n’ont pas subi de vaccination. Les 65 porcelets issus deces onze truies sont répartis en cinq lots et ont été vaccinés à quatre périodes différentesaprès la naissance selon les lots : 13°-17° jours pour les lots n° 1, 301 jour pour les lotsno 2, 60’ jour pour les lots n° 3 et 90, jour pour les lots n° 4.L’immunité anti peste porcine a été étudiée au cours de la réponse primaire et au coursde la réponse secondaire chez chacun des animaux par la cinétique d’apparition des anti-corps neutralisants sériques et par la résistance à l’infection virulente d’épreuve réaliséechez les porcelets cinq mois après la naissance. Les résultats obtenus sont les suivants :
quelque soit le lot considéré on note au moment de la mise bas dans le sérum de sept

truies sur dix une intense activité neutralisante. Chez les poroelets ayant reçu une quan-tité faible de colostrum, la vaccination entraîne le développement d’une bonne réponseimmunitaire. Chez les porcelets ayant ingéré une quantité normale de colostrum les anti-
corps passifs d’origine colostrale exercent un effet inhibiteur sur la réponse primaire quise manifeste par un retard dans l’apparition des anticorps et par un niveau plus faible aumoment de l’épreuve virulente. Selon les conditions de vaccination des truies, on observedes différences dans les propriétés des anticorps passifs d’origine colostrale présents dansle sérum des porcelets des lots correspondants. Les différences observées se manifestentau niveau :
1. De l’intensité de l’effet inhibiteur sur la réponse primaire.2. « L’avidité » in vitro des anticorps passifs vis-à-vis du virus.
3. La demi-vie moyenne des anticorps passifs.Sur un plan pratique il apparaît que la vaccination s’avère efficaoe à partir d’une périodesituée entre le 30’ et le 60e jour après la naissance chez des porcelets possédant une
immunité passive d’origine colostrale.
References
AIKEN J.M., BLORE I.C., 1964, Immunology of new born pigs : response to lapinized Hog choleravirus in colostrum deprived and suckling pigs. - Am. J. Vet. Res. 25, 1134-1139.
AYNAUD J.M., 1968. Etude de la multiplication in vitro d’un clone du virus de la peste porcine. -- Rech. Vet. 1, 25-36.
BRAN L., MIHAITA S., POPA M., TOTORCEA N., ALBU T., TIGAERU N., 1969. Recherches concer-
nant l’immunisation des porcelets contre la peste porcine avec la souche lapinis6e « C » dansles grandes unites d’61evage. - Rev. Zoot. Med. 1, 43-49.
COGGINS L., SHEFFY B.E., BAKER J.A., 1962. Response of swine to Hog cholera vaccines. - Proc.U.S. Livestock. San. Ass. 66, 316-323.
COGGINS L., 1964. Study of Hog cholera colostral antibody and its effect on active Hog choleraimmunization. - Am. J. Vet. Res. 25, 613-616.
CORTHIER G., AYNAUD J.M., GALICHER C., GELFI J., 1974. Activite antigénique compar6e de deuxTogavirus : le virus de la peste porcine et le virus de la maladie des muqueuses. ― Ann. Rech.Vet. 5, 373-380,
CORTHIER G., 1975. Influence de l’immunit6 passive sur la r6ponse du porcelet aux infections virales.Application 6 la peste porcine classique. - Journées de la Recherche Porcine en France, Paris.347-354.
CORTHIER G., 1976. Comparaison de deux methodes in vitro de titrage des anticorps neutralisantle virus de la peste porcine classique. - Ann. Rech. Vet. 7, 349-360.
CORTHIER G., 1976. Swine fever : influence of passive immunity on pig immune response followingvaccination. - Ann. Rech. V6t. 7, 361-372.
CURTIS J., BOURNE F.J., 1973. Half-lives of immunoglobulins IgG, IgA and IgM in the serum of newborn pigs. - Immunology 24, 147-151.
EASTERDAY B.C., 1972. Immunologic considerations of swine influenza. - J. Am. Vet. Med. Ass.,160, 645-650.
EISEN H.N. and SISKIND G.W., 1964. Variations in affinities of antibodies during the immune res-
ponse. - Biochemistry 3, 996-1000. ·
FELDMANN M.. DIENER E., 1970. Antibody-mediated suppression of the immune response in vitro.I. Evidence for a central effect. - J. exp. Med. 131, 247-256.
FINKELSTEIN M.S. and UHR J.W., 1964. Specific inhibition of antibody formation by passively admi-nistered 19.S and 7.S antibody. - Science, 146, 67-69.
GORET P.. 1976. Vaccination contre la peste porcine classique 6 I’aide des virus modifies. - Rec.M6d, Vet. 152, 175-179.
HANAKI T., OGAWA N., NAKAGAWA H., SAWADA M., 1972. Neutralizing antibody titer of pigs ino-culated with the Hog cholera live virus vaccine prepared from G.P.E. strain. (Communicationpersonnelle.)
HANAKI T., OGAWA N., NAKAGAWA H., SAWADA M., SAZAWA H., 1972. Antibody response in
pigs holding maternal immunity and inoculated with the G.P.E. strain of attenuated Hog cho-lera virus. - Annual Report of the Nat. Vet. Assay Laboratory no 9, 93-96.
JONSSON A., 1973. Transfer of immunoglobulins from mother to offspring in the pig. - Acta. Vet.Scand., suppl. 43.

KARUSH F., 1970. Affinity and the immune resp onse. - Ann. New York Acad. Sci., 169, 56-64.
LANG W.. NASE S., REJEWSKY K., 1969 Inhibition of the immune response in vitro to sheep redblood cells by passive antibody. - Nature, 223, 949-951.
LAUNAIS M., AYNAUD J.M., CORTHIER G., 1972. Peste porcine classique, propri6t6s d’un clone (sou-che Thiverval) iso[6 en culture cellulaire a basse temperature. Application a la vaccination. -
Rev. M6d. Vet., 123, 1537-1554.LAUNAIS M., AYNAUD J.M.. CORTHIER G., LAUDE H., 1974. Peste porcine classique : caract6res
de I’immunité, innocult6 vis-a-vis des truies en gestation et stabilit6 g6n6tique de la souche« Thiverval ». - Rev. Med. Vet., 125, 175-194.
LAUNAIS M., AYNAUD J.M., 1976. Peste porcine classique : durée de l’immunit6 chez la truie avecla souche r Thiverval » dans les conditions de la pratique. - Journées de la Recherche Por-cine en France, Paris. 227-280.
LAUNAIS M., AYNAUD J.M., 1975. Vaccination du jeune porcelet contre la peste porcine a I’aidede la souche Thiverval en presence d’immunit6 maternelle dans les conditions du terrain. -
Journees de la Recherche Porcine en France, Paris. 355-362.
LAUNAIS M., AYNAUD J.M., CORTHIER G., 1977. Hog cholera virus : Active immunization of pigletswith the x Thiverval » strain in presence of colostral passive immunity. (To be published.)
LEIPER J.B. and SOLOMO J.B.. 1975. Role of maternal antibody causing immunosuppresive delay inthe onset of plaque-forming cell responses in rats and rabbits. - In : Maternofoetal trans-mission of immunoglobulins. Edited by Hemmings W.A., Cambridge Univ. Press.
LIEW F.Y., PARISH C.R., 1972. Regulation of the immune response by antibody. I. Suppression ofantibody formation and concomitant enhancement of cell-mediated immunity by passive antibody.- Cell. lmmunol., 4, 66-85.
LIN T.C.. SHIEH C.M.. CHEN Y.C.. CHENN C.C., LEE C.S.. LAI S.S., 1969. The distribution of colostralantibody titers of piglets in Taiwan and the relationship between colostral antibody and theimmunity acquired from Hog cholera live vaccine. Exp. Report of Taiwan Prov. - Res. Inst.Anim. Hlth, 6, 11-14.
LOAN R.W., RODABAUGH D.E., 1966. Serologic studies of Hog cholera immunization. - Am. J.Vet. Res., 27, 1333-1338.
MOLLER G., WIGZELL H., 1965. Antibody synthesis at the cellular level. Antibody induced suppres-sion of 19.S and 7.S antibody response. - J. Exp. Med., 121, 969-988.
OLAH P., 1969. Immunization of suckling pigs against Swine fever using SUVAC strain. - Mag.Allatorv. Lapja. 7. 371-372.
PIERCE C.W., 1969. Immune response in vitro : li. Suppression of the immune response in vitro byspecific antibody. - J. Exp. Med., 130, 365-379.
PORTER P., 1975. Intestinal absorption of colostral IgA anti-E. Coli antibodies by the neonatalpiglet and calf. - In : Maternofoetal transmission of immunoglobulins. Edited by HemmingsW.A., Cambridge Univ. Press.
PRECAUSTA P., BRUN A., KATO E., TERRE J. et MARCON Ch., 1975. Peste porcine classique. Etuded’un vaccin prepare a partir de la souche chinoise CL adaptee 6 la culture cellulaire. - Rev.M6d. V6t., 126, 969-981.
RENSHAW H.W., 1975. Influence of antibody mediated immune suppression on clinical viral and im-mune responses to swine influenza infection. - Ann. J. Veter., 36, 5-13.
’
ROUZE P., HOUDAYER M., METZGER J.J., 1975. La r6ponse immunitaire du porcelet. Effets de l’im-munit6 passive sur l’immunisation du porcelet induite a I’aide d’un antigene inerte : le lysozyme.- Journées de la Recherche Porcine en France, Paris. 363-370.
SASAHARA J., KUMAGAI T., SHIMIZU Y., FURUUCHI S., 1969. Field experiments of Hog choleralive vaccine prepared on guinea pig kidney cell culture. - Nat. lnst. Anim. Hlth. Quart., 9,83-91.
SMITH M.W., MUNN E.A., BURTON K.A. and HENRIQUE DE JESUS C., 1975. Functional changesin the brush border surface of the newborn pig intestine. - In : Maternofoetal transmission of
immunoglobulins. Edited by Hemmings Cambridge Univ. Press.
TERPSTRA C., TIELEN J.M., 1976. Antibody response against Swine fever following vaccination withC. strain. - Zbl. Vet. Med. B 23, 809-821.
UHR J.W., MOLLER G., 1968. Regulatory effect of antibody on the Immune response. - Adv. lmmu-nol. 8, 81.
’
WALKER J.G. and SISKIND G.W., 1968. Effect of antibody affinity upon its ability to suppress anti-
body formation. - Immunology, 14, 21-28.WEBSTER R.G., 1968. The immune response to influenza virus. Ill. Changes in the avidity and spe-
cificity of early [gM and IgG antibodies. - Immunology, 14, 39-52.
ZUFFA A., CERNEK K., VLCEK J., WAGNER E., 1967. Study on the properties of chinese LapinizedHog cholera virus. Ill. Immunogenicity of the virus for suckling piglets from immune sows. -Vet. A4ed. 12, 621-630.


















