
Patogenicidad de Phytophthora spp. causantes de ... · generó una demanda de planta forestal a la...
17
Bol . San. Veg. Plagas, 30: 385-401, 2004 Patogenicidad de Phytophthora spp. causantes de podredumbre radical de Quercus ilex ssp. ballota en viveros forestales M. E. SÁNCHEZ, S. ANDICOBERRY, A. TRAPERO Como parte de una línea de investigación sobre calidad de planta para forestación de tierras agrarias, se ha estudiado la etiología de las podredumbres radicales que afectan a la encina {Quercus ilex ssp. ballota) en viveros forestales de Andalucía. La sintomatolo- gía mostrada por las plántulas afectadas consistió en marchitez foliar asociada a la necro- sis extensa de las raicillas absorbentes. A partir de las raíces necróticas se obtuvieron diversos aislados fúngicos que han sido identificados como Phytophthora cinnamomi, P. drechsleri y P. cryptogea. El estudio del crecimiento micelial de los aislados en función de la temperatura, ha mostrado que las tres especies de Phytophthora poseen curvas de crecimiento similares, con temperaturas óptimas alrededor de 24° C. Las inoculaciones efectuadas sobre plantones sanos han mostrado claramente el carácter patogénico de todos los aislados ensayados en Q. ilex ssp. ballota y Q. súber, sin que se apreciara especificidad de huésped. Las condiciones de exceso de agua en el sustrato de cultivo resultaron decisivas para el establecimiento de la infección y el desarrollo de la enferme- dad radical. Phytophthora cinnamomi ha resultado ser la especie más virulenta. También se ha establecido que Q. ilex ssp. ballota es más susceptible a la infección por P. cinna- momi que Q. súber. El presente trabajo constituye la primera descripción de P. cinnamo- mi, P. drechsleri y P. cryptogea como patógenos de plantones de encina en vivero. M. E. SÁNCHEZ, S. ANDICOBERRY, A. TRAPERO. Dpto. Agronomía. ETSIAM. Universidad de Córdoba. Apdo. 3048. 14080-Córdoba. Dirección de correo electrónico: [email protected] Palabras clave: Phytophthora cinnamomi, P. cryptogea, P. drechsleri, Quercus rotundifolia, Q. suber INTRODUCCIÓN Desde 1993 hasta 1997 fueron forestadas en Andalucía cerca de 120.000 ha dentro del Programa de Forestación de Tierras Agrarias de 1993 (GÓMEZ-JOVER y JIMÉNEZ, 1997). Este programa ofrecía ayudas a la repobla- ción con especies forestales en parcelas sometidas a un aprovechamiento agrícola previo. La puesta en marcha del Programa generó una demanda de planta forestal a la que respondieron los viveros andaluces con un aumento de la producción de las especies más solicitadas, pero sin fijar unos determi- nados criterios de calidad de planta. De todas las especies, las que experimentaron una mayor demanda fueron la encina (Quercus ilex spp. ballota) y el alcornoque (Q. súber), ocupando entre ambas alrededor del 70% de las tierras agrarias forestadas dentro de dicho Programa (GÓMEZ-JOVER y JIMÉNEZ, 1997). La calidad de la planta forestal producida por el viverista va a determinar, en gran medida, el éxito de la plantación, en cuanto al crecimiento y supervivencia de las espe- cies empleadas (NAVARRO et al., 1998). En las condiciones ambientales particulares del vivero se corre el riesgo de que las enferme- dades se extiendan e intensifiquen con rapi- dez, dando lugar a la aparición de problemas
Transcript of Patogenicidad de Phytophthora spp. causantes de ... · generó una demanda de planta forestal a la...

Bol . San. Veg. Plagas, 30: 385-401, 2004
Patogenicidad de Phytophthora spp. causantes de podredumbreradical de Quercus ilex ssp. ballota en viveros forestales
M. E. SÁNCHEZ, S. ANDICOBERRY, A. TRAPERO
Como parte de una línea de investigación sobre calidad de planta para forestación detierras agrarias, se ha estudiado la etiología de las podredumbres radicales que afectan ala encina {Quercus ilex ssp. ballota) en viveros forestales de Andalucía. La sintomatolo-gía mostrada por las plántulas afectadas consistió en marchitez foliar asociada a la necro-sis extensa de las raicillas absorbentes. A partir de las raíces necróticas se obtuvierondiversos aislados fúngicos que han sido identificados como Phytophthora cinnamomi, P.drechsleri y P. cryptogea. El estudio del crecimiento micelial de los aislados en funciónde la temperatura, ha mostrado que las tres especies de Phytophthora poseen curvas decrecimiento similares, con temperaturas óptimas alrededor de 24° C. Las inoculacionesefectuadas sobre plantones sanos han mostrado claramente el carácter patogénico detodos los aislados ensayados en Q. ilex ssp. ballota y Q. súber, sin que se apreciaraespecificidad de huésped. Las condiciones de exceso de agua en el sustrato de cultivoresultaron decisivas para el establecimiento de la infección y el desarrollo de la enferme-dad radical. Phytophthora cinnamomi ha resultado ser la especie más virulenta. Tambiénse ha establecido que Q. ilex ssp. ballota es más susceptible a la infección por P. cinna-momi que Q. súber. El presente trabajo constituye la primera descripción de P. cinnamo-mi, P. drechsleri y P. cryptogea como patógenos de plantones de encina en vivero.
M. E. SÁNCHEZ, S. ANDICOBERRY, A. TRAPERO. Dpto. Agronomía. ETSIAM. Universidadde Córdoba. Apdo. 3048. 14080-Córdoba. Dirección de correo electrónico:[email protected]
Palabras clave: Phytophthora cinnamomi, P. cryptogea, P. drechsleri, Quercusrotundifolia, Q. suber
INTRODUCCIÓN
Desde 1993 hasta 1997 fueron forestadasen Andalucía cerca de 120.000 ha dentro delPrograma de Forestación de Tierras Agrariasde 1993 (GÓMEZ-JOVER y JIMÉNEZ, 1997).Este programa ofrecía ayudas a la repobla-ción con especies forestales en parcelassometidas a un aprovechamiento agrícolaprevio. La puesta en marcha del Programageneró una demanda de planta forestal a laque respondieron los viveros andaluces conun aumento de la producción de las especiesmás solicitadas, pero sin fijar unos determi-nados criterios de calidad de planta. De todas
las especies, las que experimentaron unamayor demanda fueron la encina (Quercusilex spp. ballota) y el alcornoque (Q. súber),ocupando entre ambas alrededor del 70% delas tierras agrarias forestadas dentro de dichoPrograma (GÓMEZ-JOVER y JIMÉNEZ, 1997).
La calidad de la planta forestal producidapor el viverista va a determinar, en granmedida, el éxito de la plantación, en cuantoal crecimiento y supervivencia de las espe-cies empleadas (NAVARRO et al., 1998). Enlas condiciones ambientales particulares delvivero se corre el riesgo de que las enferme-dades se extiendan e intensifiquen con rapi-dez, dando lugar a la aparición de problemas

a corto plazo, en el mismo vivero, o a máslargo plazo, apareciendo los síntomas de laenfermedad una vez que los plantones infec-tados ya se han establecido en las parcelas derepoblación (SÁNCHEZ et al., 1999). El con-trol sanitario en los viveros es, pues, clavepara conseguir un buen desarrollo de los cul-tivos y producir una planta de calidad para larepoblación. Se han descrito más de 30 espe-cies fúngicas diferentes causantes de podre-dumbres radicales de plántulas en especiesforestales. Algunos de ellos son parásitoscon escasa capacidad saprofítica (Phytopht-hora y Pythium), aunque la mayoría sonsaprofitos facultativos que pueden actuarcomo patógenos cuando se ven favorecidospor las condiciones ambientales (BUTIN,1995). En los viveros españoles, la mayoríade los problemas de muerte de plántulas sondebidos a enfermedades causadas por hon-gos, especialmente los causantes de caída deplántulas (damping-off), y los derivados dela aparición de malas hierbas en los cultivos(PEÑUELAS y OCAÑA, 1996). En España, lapodredumbre de las raíces causada por Phy-tophthora rara vez ha sido identificada enviveros forestales y normalmente suelen atri-buirse las pérdidas a una mala aireación delsustrato (PEÑUELAS y OCAÑA, 1996). Por otraparte, en el género Quer cus las podredum-bres radicales de plantones en vivero no sonmuy frecuentes, y no está estudiada la inci-dencia de estas enfermedades en las especiesde Quercus que se producen en España.
El desconocimiento existente sobre lasprincipales enfermedades de vivero en elcontexto específico de las especies más utili-zadas en la forestación de tierras agrarias enAndalucía, hace necesario un estudio en pro-fundidad sobre su etiología, como primerpaso necesario para establecer un controlefectivo de las mismas (SÁNCHEZ et al.,1999).
Por ello, como parte de una línea de inves-tigación sobre calidad de planta en los vive-ros forestales andaluces, los objetivos delpresente trabajo han sido (i) caracterizar lasintomatologia de las principales micosisque afectan a los plantones de encina en los
viveros andaluces, (ii) identificar los hongosasociados con las podredumbres radicales dela encina en vivero y (iii) determinar la pato-genicidad sobre alcornoque y encina de loshongos consistentemente aislados de lasplantas enfermas.
MATERIAL Y MÉTODOS
Manejo de las muestrasLa toma de muestras se realizó en dos
viveros forestales de la provincia de Córdo-ba afectados de muerte de plantones. El pri-mero de ellos (Vivero I) era un vivero de pro-ducción propia para forestación de una fincaprivada ubicada en la sierra de Córdoba. Losplantones de encina afectados crecían enbolsas de polietileno negro de 1 1 de capaci-dad, utilizando como sustrato suelo naturalde monte. El otro vivero (Vivero II) estádedicado a la producción comercial intensi-va de planta en envase y utilizan como sus-trato turba rubia / negra (7/3 en volumen) enbandejas de 9 x 5 alvéolos de 18 cm de altu-ra y 300 cm^ de volumen. En los dos vive-ros, las plántulas de encina afectadas eran de1 savia y estaban próximas a su plantaciónen campo.
Tras consignar la sintomatologia aérea dela enfermedad, para el muestreo se tomaronseis plantones sintomáticos en cada vivero,desechando aquellos que aparecían comple-tamente marchitos. Ya en el laboratorio, trassacar los plantones de sus envases, se proce-dió a un lavado cuidadoso de las raíces bajoel chorro de agua. Las raíces con síntomas depodredumbre de cada uno de los plantonesse trataron separadamente. Se cortaron conescalpelo estéril en pequeños trozos (3-4 mmde longitud) bajo condiciones asépticas, y sesembraron en placas de Petri. Los medios decultivo empleados para los aislamientos fue-ron: APDA (medio genérico para el aisla-miento de hongos) (SÁNCHEZ et al., 2002c),PARP (específico para el aislamiento dehongos pitiáceos) y PARPH (específico parael aislamiento de especies del género Phy-tophthora) (JEFFERS y MARTIN, 1986). Sesembraron 6 placas de cada medio de cultivo

Cuadro 1. Aislamiento de Phytophthora spp. a partir de las raíces necróticas de plantones de encina afectados depodredumbre radical
ViveroI
n
Aislados purificados
PV1,PV2
PV3, PV4
PV5
Especie fúngica
Phytophthora cinnamomi A2
Phytophthora drechsleri A2
Phytophthora cryptogea Al
Aislamiento (% ) 8
32
17
34a Porcentaje expresado como n° de trocitos de raíz que dieron lugar a una colonia sobre el n° total de trochos de raíz
colocados en medio selectivo (PARP o PARPH).
por plantón. Las placas se incubaron a 22° Cen oscuridad durante 4 días. Al cabo de 2días, las placas que contenían el medioAPDA se colocaron bajo luz durante otros 4días (12 h/día). Las colonias obtenidas seagruparon en función de su morfología y setransfirieron aquellas que presentaron mayorfrecuencia de aislamiento, con el fin de obte-ner cultivos puros (aislados) de cada grupomorfológico. Se seleccionaron 1 ó 2 aisladosde cada grupo (Cuadro 1) para su caracteri-zación e identificación.
Caracterización e identificación de losaislados
Morfología de las coloniasPara caracterizar la morfología de la colo-
nia se utilizó el medio de zanahoria-agar(CA) (DHINGRA y SINCLAIR, 1995). Los ais-lados se incubaron en oscuridad y a 20° C yse observaron a los 4 días de crecimiento.Además, se realizaron observaciones delmicelio al microscopio óptico invertido.
Estructuras de reproducción asexualPara estimular la producción de esporan-
gios, se colocaron trozos de CA colonizadospor cada uno de los aislados en un mediolíquido de extracto de suelo (RIBEIRO, 1978;SÁNCHEZ et al, 2000; 2002b) a dos tempera-turas de incubación: 15 y 20° C, renovandoel extracto de suelo cada 24 h. Se realizaronobservaciones periódicas desde las 48 h deincubación hasta los 5 días con el microsco-pio invertido. Cuando se observó la produc-ción de esporangios, se realizaron montajesteñidos con fucsina ácida en lactofenol parasu observación en el microscopio óptico. Porcada aislado se observaron y midieron 30esporangios maduros.
Estructuras de reproducción sexualPara la producción de gametangios se reali-
zaron cultivos dobles en medio CA con aisla-dos de referencia de tipo de apareamiento Aly A2, ya que ninguno de los aislados produjoestructuras sexuales en cultivo simple. Para loscruzamientos se utilizaron como aislados dereferencia P. cinnamomi Al (proporcionadopor la Dra. Pérez Giménez, CIFA, Málaga), P.cinnamomi A2 (obtenido de la micoteca delDpto. de Agronomía de la ETSIAM, Córdoba)y P. drechsleri Al y A2 (proporcionados por elDr. Brasier, Forestry Commission ResearchAgency, UK). Se prepararon 3 repeticionespor combinación y las placas se incubaron a22° C en oscuridad. En las placas donde loscruzamientos tuvieron éxito se procedió a lapreparación de montajes microscópicos teñi-dos con fucsina ácida en lactofenol. Por cadaaislado se observaron y midieron 30 oogonios,30 anteridios y 30 oosporas maduras.
Tasa de crecimiento a distintas tempera-turas
Cada uno de los aislados se sembró en elmedio CA y las placas se incubaron a 5, 10,15,20, 25, 30 o 35° C. Se midió el diámetromedio de la colonia a los2 ,4 ,6 ,10y l5 díasde incubación a cada temperatura. Con losdatos de tres repeticiones por aislado y tem-peratura, se calculó la tasa máxima de creci-miento diario en función de la temperatura.Estos datos se ajustaron matemáticamente auna curva de regresión, mediante el progra-ma Statistix for Windows (Analytical Soft-ware, Tallahassee, USA), eligiendo el mode-lo que mejor se ajustaba para cada uno de losaislados. Utilizando la ecuación resultante,se calcularon las temperaturas cardinales decrecimiento.

Ensayos de patogenicidad en plantonesde encina y alcornoque
Experimento 1: Los dos aislados proce-dentes del Vivero I (PV1 y PV2) se inocula-ron en plantones sanos de encina y alcorno-que de 1 savia. Se ensayaron tres métodos deinoculación de los plantones: mediante laaplicación de inoculo producido en medioVAV8 (Vermiculita-avena-V8) (WILCOX yMIRCETICH, 1985), sustituyendo el jugo devegetales V8 por extracto de zanahoria (SÁN-CHEZ et a/., 2000; 2002b); en medio de mijo(MOREIRA et al., 1997) y en suspensiónacuosa (SÁNCHEZ et al., 2000; 2002b).
En todos los casos el inoculo se aplicó alcepellón de los plantones extraídos de susenvases originales, a razón de 100 mi porplanta para los medios de vermiculita y sus-
pensión acuosa y de 50 mi para el medio demijo (Figura la, Ib). El cepellón así tratadose introdujo en una maceta previamentedesinfestada con lejía y se añadió suelo hastacompletar un volumen de 2.5 1 por maceta.Este suelo era una mezcla de arena, limo yturba (1:1:1 en volumen) (TRAPERO y JIMÉ-NEZ, 1985). Se prepararon 6 repeticiones poraislado y tipo de inoculo. Las macetas secolocaron en bandejas, también previamentedesinfestadas con lejía, de forma que en cadabandeja sólo hubiera plantas inoculadas conel mismo aislado. Tras una semana de acon-dicionamiento en cámara de crecimiento, lasbandejas se llenaron de agua (Figura le),manteniéndose llenas durante todo el experi-mento, ya que éstas son las condiciones quefavorecen la infección radical de la encina y
Figura 1: Inoculación de plantones de encina y alcornoque de 1 savia (Experimento 1), a) aplicación al cepellón delinoculo en el medio VAV8 modificado, b) aplicación al cepellón del inoculo en el medio de mijo, c) encharcamiento
del sustrato mediante inundación de las bandejas.

el alcornoque por Phytophthora spp. (SÁN-CHEZ et al., 2000; 2002b). El experimentotranscurrió a 22 ± 2o C de temperatura y 14h de luz diaria. Los plantones testigo fuerontratados de la misma manera, salvo por laausencia de cualquier tipo de inoculo.
Experimento 2: Se ensayó la influenciadel contenido hídrico del suelo en la infec-ción. Para ello, los aislados PV2 y PV5 seinocularon en plantones sanos de encina yalcornoque de 1,5 savias. La inoculación serealizó con suspensiones acuosas en dosis de100 mi por planta. Todos los plantones,incluyendo los testigos, fueron sometidos atres regímenes hídricos distintos: saturacióncontinua (SÁNCHEZ et al, 2000; 2002b),saturación periódica corta (2 días por sema-na) y saturación periódica larga (2 días cada2 semanas). Para ello, todas las plantas fue-ron colocadas en bandejas de plástico deforma que en cada bandeja sólo hubieraplantas inoculadas con el mismo aislado ytras una semana de acondicionamiento encámara de cultivo a 22 ± 2o C, se procedió a
la inundación de las bandejas según las con-diciones hídricas que les correspondiesen. Elexperimento se llevó a cabo en umbráculo,con una temperatura máxima media de 31,5o
C y temperatura mínima media de 12,2° C.Se prepararon 6 repeticiones por aislado yrégimen hídrico.
Experimento 3: En este ensayo se inocula-ron plantones de encina y alcornoque de 9meses de edad con los 5 aislados procedentesde encina más un aislado de referencia de P.drechsleri procedente de plántulas de pinocarrasco (SÁNCHEZ et al., 2002a), conservadoen la micoteca del Dpto. de Agronomía de laUniversidad de Córdoba (aislado PPH). Parainfectar las plantas se eligió el método de ino-culación con suspensiones acuosas. Las plan-tas inoculadas y las testigo fueron sometidasa condiciones de encharcamiento periódicocorto (2 días por semana). El experimento sellevó a cabo en umbráculo, con temperaturamáxima media de 33,5° C y temperaturamínima media de 18,1° C. Se prepararon 6repeticiones por aislado inoculado.
Figura 2: Síntomas de la podredumbre radical en plantones de encina, a) síntomas foliares: marchitez en los márgenesde las hojas, b) marchitez foliar generalizada, c) necrosis de raicillas absorbentes, d) desprendimiento de la corteza
externa o síntoma "raíz pelada".

En todos los experimentos se realizó unseguimiento de los síntomas foliares, y sedieron por finalizados cuando las plantasinoculadas mostraron la sintomatologiaaérea de la enfermedad. En este momento sesacaron las plantas de sus macetas, se lava-ron a chorro de agua las raíces y se evalua-ron. Se utilizó la escala 0-4 para la evalua-ción de la severidad de las infecciones enparte aérea y raíz (0 = sin síntomas, 1 = 1-33%, 2 = 34-66%, 3 = 67-99%, 4 = 100% departe aérea o raíz sintomática) (SÁNCHEZ eta/., 2000; 2002b). A estos datos se les aplicóel análisis de la varianza y los valoresmedios fueron comparados mediante el testLSD (mínima diferencia significativa) prote-gido de Fisher al nivel de significación P =0,05 (STEEL y TORRIE, 1985). Tras la evalua-ción se realizaron aislamientos de las raici-llas en el medio PARPH para determinar elgrado de recuperación del patógeno.
RESULTADOS
Sintomatologia observada en viveroLos síntomas observados en los plantones
de encina fueron similares en los dos vive-ros. Las plántulas presentaban clorosis, dese-cación y marchitez foliar, que comenzabapor los márgenes e iba avanzando hacia elnervio principal hasta la completa marchitezde la hoja (Figura 2a). En algunos casos se
produjo defoliación, pero en general lashojas marchitas permanecían prendidas altallo (Figura 2b). Los sistemas radicales apa-recían reducidos y con necrosis extensas enlas raicillas absorbentes aún unidas a la raízprincipal leñosa, que sin embargo no presen-taba lesiones (Figura 2c). En estas raícesabsorbentes necrosadas frecuentemente seapreciaba un desprendimiento de la cortezaexterna, dando lugar al síntoma denominado"raíz pelada" (Figura 2d) (SÁNCHEZ et al,1998a; 2000). En los dos viveros el drenajede los envases era deficiente, por lo que seveían sometidas a encharcamientos periódi-cos. En el Vivero I, las plántulas estaban dis-puestas sobre láminas de polietileno negrosobre un suelo de hormigón. El riego se rea-lizaba diariamente con manguera y el excesode agua se acumulaba en el suelo, formandocharcos que llegaban a cubrir los orificios dedrenaje de las bolsas en las que crecían estosplantones. Todas las plántulas de encina deeste vivero mostraban en mayor o menorgrado síntomas de podredumbre radical. Enel Vivero II los problemas de drenaje estabanoriginados por la alta capacidad de retenciónhídrica del sustrato utilizado para el cultivo.En este vivero la enfermedad se manifestó enforma de rodales que se distribuían en tornoa los aspersores de riego.
De estos dos viveros se obtuvieron cuatrotipos de colonias fúngicas en el medio PARP
Figura 3: Morfología de las colonias de Phytophthora spp. a los 4 días de crecimiento a 20° C en el medio CA, a) coloniapetaloide con micelio aéreo algodonoso del aislado PV1, identificado como P. cinnamomi, b) colonia tipo roseta del
aislado PV5, identificado como P. cryptogea, c) colonia tipo crisantemo del aislado PV4, identificado como P. drechsleri.

Cuadro 2. Características de los esporangios producidos por los aislados de Phytophthora spp. en medio líquidode extracto de suelo
a Valores expresados en \\,m. Cada valor es la media de 30 observaciones ± desviación estándar.
y PARPH asociadas a la podredumbre radi-cal, de los cuales se conservaron y caracteri-zaron 5 aislados. Los porcentajes de aisla-miento aparecen reflejados en el Cuadro 1.Además, en el medio PARP se obtuvieronpuntualmente algunas colonias pertene-cientes al género Pythium, y en el medioAPDA se aisló esporádicamente alguna
colonia perteneciente a los géneros Cylin-drocarpon y Fusarium.
Caracterización e identificación de losaislados
Morfología de las coloniasA los 4 días de crecimiento a 20° C en el
medio CA, los aislados PV1 y PV2 formaron
Figura 4: Morfología de las estructuras de reproducción asexual de los aislados de Phytophthora spp., a) esporangioelipsoide de P. cinnamomi, b) ramificación simpodial de los esporangióforos de P. cinnamomi, c) esporangio
subesférico de P. drechsleri, d) esporangio obpiriforme de P. cryptogea.

392 M. E. SÁNCHEZ, S. ANDICOBERRY, A. TRAPERO
colonias de color blanco, con abundantemicelio aéreo algodonoso y crecimiento uni-forme o ligeramente petaloide (Figura 3a).Las colonias de los aislados PV3 y PV5 pre-sentaron formaciones miceliares blanqueci-nas a modo de pétalos, dando lugar a unacolonia tipo "roseta", con micelio aéreo esca-so (Figura 3b). Por último, el aislado PV4formó colonias blancas tipo "crisantemo",con pétalos pequeños e imbricados, micelioaéreo escaso y pulverulento (Figura 3c).
En las observaciones realizadas con elmicroscopio invertido, todos los aisladospresentaron un micelio cenocítico muy rami-ficado. Los aislados PV1, PV2 y PV5 pre-sentaron ramificaciones botriosas. Los aisla-dos PV1 y PV2 mostraron además hinchazo-nes hifales grandes, esféricas, terminales eintercalares, simples y en racimo.
Caracterización de estructuras de repro-ducción asexual
De las dos temperaturas ensayadas, la de20° C fue la más favorable para la formaciónde esporangios. A esta temperatura el tiempoque tardaron en formarse los esporangiososciló entre las 48 y las 72 h, mientras que a15° C osciló entre los 4 y 5 días y se produ-jeron en menor número.
En todos los aislados los esporangios for-mados fueron persistentes y no papilados. Suscaracterísticas se recogen en el Cuadro 2.
Los esporangios producidos por el aislado
PV1 fueron de forma ovoide-elipsoide, conun alto porcentaje de esporangios con ápicetruncado (Figura 4a). Los esporangióforosfueron en su gran mayoría simples y largos,aunque también los hubo ramificados simpo-dialmente (Figura 4b). Los esporangios pro-ducidos por PV2 fueron similares, aunque eneste caso el porcentaje que presentaban elápice truncado fue menor. PV3 produjoesporangios subesféricos (Figura 4c) y enmenor proporción ovoides. Sólo un pequeñoporcentaje de los esporangios presentó elápice truncado. Los esporangióforos fueronen su gran mayoría simples y largos, conproliferación externa, generalmente no rami-ficados. Los esporangios producidos por elaislado PV4 fueron de forma ovoide o elip-soide. Un alto porcentaje presentó ápicetruncado. Los esporangióforos fueron simi-lares a los de PV3. El aislado PV5 produjoesporangios obpiriformes (Figura 4d) y elip-soides con un 100% de ápices truncados yesporangióforos simples.
En todos los aislados se observó la forma-ción de hinchazones hifales en medio líquidode extracto de suelo, siendo más abundantesa 15° C que a 20° C. Éstas fueron en sumayoría esféricas, intercalares y en racimos,aunque también las hubo solitarias y termi-nales, siendo de mayor tamaño y mucho másabundantes las producidas por los aisladosPV1 y PV2.
b)
Figura 5: Morfología de las estructuras de reproducción sexual de los aislados de Phytophthora spp. obtenidosmediante cruzamientos con aislados de referencia: oosporas pleróticas y anteridios anfiginos de a) P. cinnamomi,
b) P, drechsleri, c) P. cryptogea.

Cuadro 3. Formación de estructuras de reproducción sexual de los aislados de Phytophthora spp. en cultivodoble con aislados de referencia de ambos tipos de apareamiento (Al y Â2)
Especie
P. cinnamomi
P. drechsleri
P. cryptogea
Aislado
PV1
PV2
PV3
PV4
PV5
P. cinnamomiAl
+
+
+
+
-
Aislado de referencia
P. cinnamomiA2
•
+
P. drechsleriAl
+
+
+
+
-
: P. drechsleriA2
+
Tipo deapareamiento
A2
A2
A2
A2
Al
+ = Formación de gametangios en cultivo doble.- = Ausencia de gametangios en cultivo doble.
Caracterización de estructuras de repro-ducción sexual
Los resultados de los cruzamientos de losaislados a identificar con los aislados dereferencia se recogen en el Cuadro 3. Casitodos los aislados produjeron gametangiosen cultivo doble con los aislados de referen-cia Al, por lo que su tipo de apareamiento esA2. La única excepción fue el aislado PV5,que resultó de tipo de apareamiento Al. Parala caracterización de las estructuras sexualesde los aislados con tipo de apareamiento A2,se utilizaron las producidas en los cruza-mientos realizados con P. cinnamomi Al .Para el aislado PV5, se observaron y midie-ron los gametangios obtenidos en los cruza-mientos realizados con P. drechsleri A2. Entodos estos casos se produjeron estructurassexuales en las interfases y en zonas de laplaca próximas a la pieza de agar de siembradel aislado a identificar. Los montajes se rea-lizaron con estructuras procedentes de estas
zonas, reduciéndose así la eventualidad deincluir gametangios producidos por los aisla-dos de referencia.
En todos los casos se produjeron oogo-nios esféricos, de pared lisa y con oosporaspleróticas (Figura 5). Los anteridios fueronen todos los casos anfiginos y unicelulares(Figura 5), salvo algunos pertenecientes a losaislados PV1 y PV2, que fueron bicelulares(Cuadro 4). Las dimensiones de los oogo-nios, grosor de la pared de las oosporas ytamaño de los anteridios aparecen en Cuadro4. La relación entre el diámetro de la oospo-ra y la longitud del anteridio (Do/La) paratodos los aislados osciló entre 2,3 y 2,6,excepto para el aislado PV5, que fue de 1,2(Cuadro 4).
Identificación de los aisladosTodos los aislados obtenidos se caracteri-
zaron por ser heterotálicos, con anteridiosanfiginos y oosporas pleróticas. Además, losesporangios aparecían en posición terminal
Cuadro 4. Características de las estructuras sexuales de los aislados de Phytophthora spp. producidos en cultivosdobles con aislados de referencia
' Valores expresados en \im. Cada valor es la media de 30 observaciones ± desviación estándar.

M. E. SÁNCHEZ, S. ANDICOBERRY, A. TRAPERO
Figura 6. Curvas de crecimiento en el medio de cultivo CA para cada una de las especies de Phytophthora causantesmuerte de plántulas de encina en vivero. Las flechas indican las temperaturas óptimas de crecimiento estimadas. Los
puntos representan los valores medios de tasa de crecimiento de los aislados de cada especie a las distintastemperaturas ensayadas.

Cuadro 5. Temperaturas mínimas y óptimas de crecimiento estimadas para cada especie de Phytophthora en elmedio de cultivo CA
Especie
P. cinnamomi
P. drechsleri
P. cryptogea
T" mínima8
7,3
4,2
1,4
T" óptimab
24,7
24,4
23,7a Las temperaturas mínimas fueron determinadas calculando los valores de T para y = 0 de las ecuaciones polinómicas
de regresión (Figura 6) ajustadas para cada especie.b Las temperaturas óptimas fueron estimadas a partir de la derivación de las ecuaciones polinómicas de regresión (Figura 6)
ajustadas para cada especie.
en el esporangióforo, eran no papilados ypersistentes. Por todo ello, pertenecen algénero Phytophthora, dentro del grupo VI deWATERHOUSE (1963). La caracterización desus estructuras vegetativas y de reproducciónsexual y asexual (Cuadros 2 y 4), han permi-tido su adscripción a diferentes especies mor-fológicas: PV1 y PV2 han sido identificadoscomo P. cinnamomi; PV3 y PV4 como P.drechsleri y PV5 como P. cryptogea, segúnlas descripciones de WATERHOUSE (1963),STAMPS et al. (1990) y ERWIN y RIBEIRO
(1996).Tasa de crecimiento a distintas tempera-
turasLos valores de tasa diaria máxima de cre-
cimiento en función de la temperatura paralos distintos aislados se ajustaron a unmodelo polinómico de 3er grado forzado en
el origen, cuya expresión general es:y=a*T3+b*T2+c*T, donde y representa latasa de crecimiento máximo diario, T es latemperatura de crecimiento y a, b y c son loscoeficientes de regresión. Las curvas de cre-cimiento para cada una de las especies iden-tificadas aparecen en la Figura 6. A partir delas ecuaciones obtenidas se estimaron lastemperaturas mínimas y óptimas de creci-miento (Cuadro 5). Las temperaturas ópti-mas de crecimiento difirieron poco entreespecies y estuvieron alrededor de 24° C. Porotra parte, se estimó la temperatura mínimade crecimiento para cada especie, que seajustó en todos los casos a las observacionesexperimentales, estimándose temperaturasmínimas superiores a 5o C para P. cinnamo-mi e inferiores a 5o C para P. drechsleri y P.cryptogea (Cuadro 5). Las temperaturas
Cuadro 6. Influencia del método de inoculación en la patogenicidad de aislados de P. cinnamomi sobre plantonesde encina y alcornoque (Experimento 1)
1 La escala utilizada para evaluar la severidad de los síntomas fue: 0 = sin síntomas, 1 = 1-33%, 2 = 34-66%, 3 = 67-99%, 4 = 100% del tejido afectado. Para cada columna, medias seguidas de una letra común, no difierensignificativamente (P = 0,05), según el test de la mínima diferencia significativa (LSD) protegido de Fisher (Steel yTome, 1985).
máximas de crecimiento estimadas para cadaespecie excedieron del intervalo de tempera-turas ensayado, por lo que no se consideraron.
Ensayos de patogenicidadEn todas las condiciones ensayadas, en
las dos especies de Quer cus, con las distintastécnicas de inoculación y aislados inocula-dos, se reprodujeron los síntomas de laenfermedad observados en vivero (Figura 7).En ningún caso se observó esta sintomatolo-gia en las plantas testigo utilizadas en losdiferentes ensayos, con lo cual, los resulta-dos que se exponen a continuación muestranclaramente el carácter patógeno de todos losaislados en las dos especies de Quercus ino-culadas. En todos los experimentos se recu-peró el hongo inoculado a partir de las raícesnecróticas de las plantas inoculadas, mien-tras que el porcentaje de recuperación dePhytophthora spp. en las raíces de las plan-tas testigo fue del 0% en todos los casos.
Experimento 1: Se dio por concluido alcabo de 4 semanas. La severidad de los sínto-mas aéreos y radicales correspondientes a lasdiferentes especies de planta, aislados y técni-cas de inoculación, se recogen en el Cuadro 6.También aparecen los análisis de la varianza ycomparaciones de medias individualizadospor especie de planta. El análisis global de lavarianza para la severidad de síntomas aéreosmostró diferencias significativas entre lasplantas inoculadas y las testigo sin inocular.También se apreciaron diferencias significati-vas en función de la especie de planta inocula-da, así como en la interacción aislado-planta.Sin embargo, no existieron diferencias en fun-ción de los distintos métodos de inoculaciónempleados, los aislados ensayados, ni en lainteracción entre ambas variables A nivel radi-cal sólo hubo diferencias significativas entreplantas testigo e inoculadas.
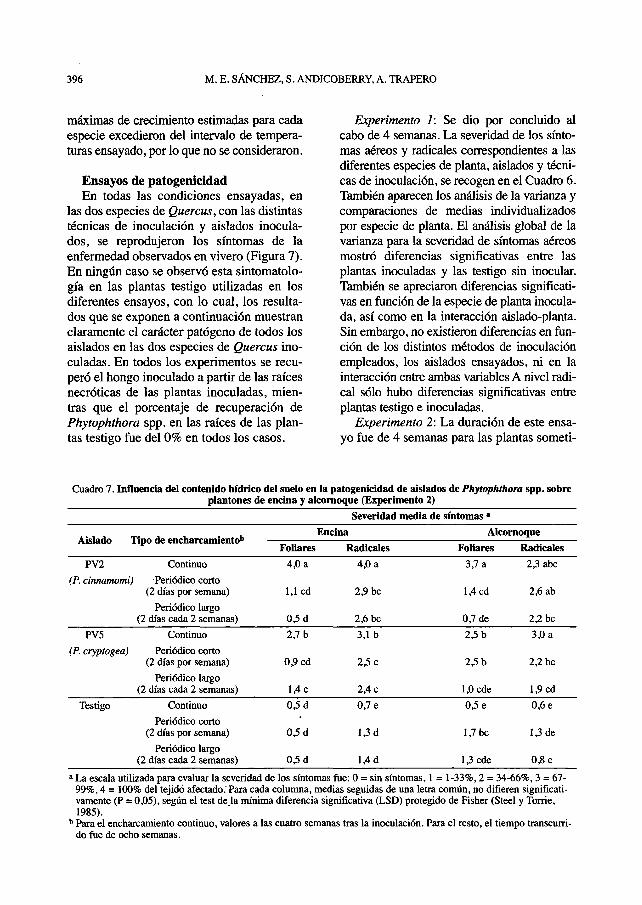
Experimento 2: La duración de este ensa-yo fue de 4 semanas para las plantas someti-
Cuadro 7. Influencia del contenido hídrico del suelo en la patogenicidad de aislados de Phytophthora spp. sobreplantones de encina y alcornoque (Experimento 2)
a La escala utilizada para evaluar la severidad de los síntomas fue: 0 = sin síntomas, 1 = 1-33%, 2 = 34-66%, 3 = 67-99%, 4 = 100% del tejido afectado .'Para cada columna, medias seguidas de una letra común, no difieren significati-vamente (P = 0,05), según el test de la mínima diferencia significativa (LSD) protegido de Fisher (Steel y Torne,1985).
b Para el encharcamiento continuo, valores a las cuatro semanas tras la inoculación. Para el resto, el tiempo transcurri-do fue de ocho semanas.
Continuo
Periódico corto(2 días por semana)
Periódico largo(2 días cada 2 semanas)
Continuo
Periódico corto(2 días por semana)
Periódico largo(2 días cada 2 semanas)
Continuo
Periódico corto(2 días por semana)
Periódico largo(2 días cada 2 semanas)

Figura 7: Síntomas expresados por los plantones inoculados en los experimentos de patogenicidad. Experimento 3,a) marchitez foliar y necrosis radical en plantón de encina inoculado con P. cinnamomi (PV1), a la izquierda plantón
testigo b) síntomas foliares y necrosis radical en plantones de alcornoque. De izquierda a derecha: plantón testigo,inoculado con P. cinnamomi (PV1), inoculado con P. cinnamomi (PV2), inoculado con P. drechderi (PV4).
das a encharcamiento continuo y de 8 sema-nas para las encharcadas periódicamente. Eneste experimento, el análisis global de lavarianza mostró diferencias significativas enla severidad de síntomas aéreos y radicalesen función de las condiciones de encharca-
miento, entre plantas inoculadas y testigo yentre las dos especies de plantas, así como enlas interacciones entre las condiciones deencharcamiento y cada una de las demásvariables. Así, los encharcamientos continuoy periódico corto resultaron los más favora-
Cuadro 8. Patogenicidad de los aislados de Phytophthora spp. sobre plantones de encina y alcornoque(Experimento 3)
a La escala utilizada para evaluar la severidad de los síntomas fue: 0 = sin síntomas, 1 = 1-33%, 2 = 34-66%, 3 = 67-99%, 4 = 100% del tejido afectado. Para cada columna, medias seguidas de una letra común, no difieren significativa-mente (P = 0,05), según el test de la mínima diferencia significativa (LSD) protegido de Fisher (Steel y Torne, 1985).
b PPH = Aislado de P. drechsleri causante de podredumbre radical en plántulas de Pinus halepensis (Sánchez et al.,2002a).

bles para el desarrollo de la enfermedad. Nose apreciaron diferencias significativas entreel aislado de P. cinnamomi (PV2) y el de P.cryptogea (PV5). Este análisis también mos-tró una mayor severidad de síntomas radicalesen encina, si bien los síntomas secundarios dela enfermedad, es decir, los síntomas aéreos,fueron significativamente más severos enalcornoque. En el Cuadro 7 se muestran losresultados del experimento y los análisis de lavarianza y comparaciones de medias indivi-dualizados por especie de planta.
Experimento 3: Se dio por concluido a las8 semanas. El análisis global de la varianzapara la severidad de síntomas aéreos y radi-cales y su comparación de medias permitióestablecer diferencias de patogenicidad entrelos distintos aislados ensayados, resultandolos aislados PV1 y PV2, identificados comoP. cinnamomi, los más patogénicos tanto enencina como en alcornoque (Cuadro 8). Elaislado de referencia de P. drechsleri (PPH)resultó igualmente muy patogénico en enci-na, aunque no así para el alcornoque. En esteexperimento no se apreciaron diferenciassignificativas a nivel aéreo entre encinas yalcornoques, aunque sí las hubo a nivel radi-cal (Figura 7), siendo de nuevo las encinaslas que mostraron síntomas más severos.
DISCUSIÓN
El síndrome de muerte de plantonesobservado en los dos viveros afectados secorresponde plenamente a la enfermedadconocida como "colapso tardío" o "dam-ping-off tardío", según la denominación deBUTIN (1995).
Los hongos asociados a la podredumbreradical pertenecen en todos los casos al géne-ro Phytophthora. De las tres especies identifi-cadas (P. cinnamomi, P. drechsleri y P. crypto-gea), sólo P. cinnamomi ha sido descrita cau-sando enfermedades radicales en pies adultosde Q. ilex ssp. ballota y Q. súber (BRASIER,1996; TUSET et al, 1996, MOREIRA et al,
1997; SÁNCHEZ et al, 2000; 2002b).Todos los aislados presentaron curvas de
crecimiento muy parecidas, con elevadas
tasas de crecimiento y temperaturas óptimasalrededor de 24° C. Estas temperaturas con-trastan con las temperaturas óptimas descri-tas para P. cinnamomi (30° C) procedentes deaislamientos realizados en Andalucía de raízde encinas adultas (SÁNCHEZ et al, 2000;2002b), sugiriendo una cierta adaptación delos aislados a las condiciones específicas delvivero.
Los experimentos de patogenicidad reali-zados han demostrado que todos los aisladosde Phytophthora son claramente patógenasde encina y alcornoque en vivero y reprodu-cen los síntomas de podredumbre radical ymarchitez foliar en todas las condicionesensayadas. Además, la patogenicidad de P.cinnamomi, P. drechsleri y P. cryptogeaviene determinada por el contenido hídricodel suelo, ya que la aparición de la enferme-dad en los viveros estudiados estuvo asocia-da a un estado de saturación hídrica periódi-ca de los sustratos en los que crecían losplantones. Estas condiciones de encharca-miento favorecen la aparición de este tipo deenfermedades (JUZWIK et al, 1998; PETTTTTet al, 1998; SÁNCHEZ et al, 1998b; 2000;2001; 2002b). El bajo nivel de síntomasmostrados por las plantas testigo en todos losexperimentos, significativamente menoresque los valores de severidad mostrados porlas plantas inoculadas, indica que el efectodel encharcamiento, en ausencia de patóge-no, no es el responsable de los síntomasobservados en las plantas inoculadas. Tal ycomo ya se demostró en el caso de la podre-dumbre radical del olivo causada por Phy-tophthora megasperma (SÁNCHEZ et al,1998b; 2001), los resultados obtenidossugieren que lo que se viene considerandocomo alta sensibilidad de los Quercus a laasfixia radical (PEÑUELAS y OCAÑA, 1996)por encharcamiento de los sustratos de culti-vo, se debe contemplar más apropiadamentecomo una muy elevada susceptibilidad aestos patógenos en las condiciones de satu-ración hídrica que favorecen la infección delas raíces.
Los aislados reprodujeron la enfermedaden las dos especies de Quercus inoculadas,

no pudiéndose apreciar especificidad dehuésped, tal y como cabía esperar de hongoscausantes de "damping-off" (MANION, 1991;BUTIN, 1995).
En el primer ensayo de patogenicidad(Experimento 1) los tres tipos de inoculodieron lugar a la infección radical de losplantones, sin que se apreciaran diferenciassignificativas en cuanto a la severidad de sín-tomas radicales obtenida. DHINGRA y SIN-CLAIR (1995) consideran más efectivos losmétodos de inoculación de raíces en los cua-les el inoculo se añade suspendido en unmedio nutritivo. En nuestro caso, la ausenciade nutrientes en el medio de inoculación(suspensión acuosa), resultó tan efectivocomo los otros dos tipos de inoculo, en loscuales el hongo se añadía colonizando gra-nos de avena y partículas de vermiculita(medio VAV8 modificado) o granos de mijo.Este hecho puede ser debido a la gran capa-cidad infectiva que muestran los aisladoscuando las condiciones de humedad y tem-peratura son favorables. Por este motivo, seeligió la suspensión acuosa para posterioresinoculaciones, por ser el método de inocula-ción más sencillo en cuanto a su preparacióny posterior manejo.
El Experimento 2 tuvo como objetivoconocer la influencia de la saturación hídricadel suelo en el desarrollo de la enfermedad.La mayoría de autores señalan las condicio-nes de encharcamiento periódico como lasmejores para el desarrollo de las infeccionesproducidas por Phytophthora en huéspedesleñosos (RIBEIRO, 1978; BROWNE y MIRCE-TICH, 1988). En nuestros ensayos, las condi-ciones de encharcamiento continuo resultaronlas más favorables, ya que dieron lugar ainfecciones más severas y además, la sinto-matologia de la enfermedad se desarrolló enun tiempo menor. Los plantones testigo some-tidos a encharcamiento continuo mostraronunos valores de necrosis radical bajísimos,incluso menores que los ya bajos valoresobtenidos para los plantones testigo encharca-dos periódicamente. No obstante, para lossiguientes experimentos se optó por someter alos plantones a encharcamiento periódico
corto, por ser éstas las condiciones que más seasemejan a las condiciones reales de vivero.
El Experimento 3 mostró diferencias depatogenicidad entre los distintos aisladosensayados. En este sentido, atendiendo a lasintomatologia primaria de la enfermedad, P.cinnamomi resultó ser la especie más viru-lenta para ambas especies de Quercus. Enencina, el aislado PPH procedente de pino,identificado como P. drechsleri (SÁNCHEZ etal., 2002a), dio lugar a una sintomatologiatan severa como los aislados de P. cinnamo-mi, en contraste con el resto de aislados de P.drechsleri, que dieron lugar a una severidadde síntomas menor, a pesar de haber sidoobtenidos de raíces enfermas de encina. Esimportante señalar que, en las condicionesde este experimento, los alcornoques presen-taron una menor severidad de síntomas folia-res que las encinas, aunque las diferencias nofueron tan marcadas a nivel radical. Lamayor susceptibilidad de la encina a la infec-ción radical por P. cinnamomi ya había sidopuesta de manifiesto en inoculaciones artifi-ciales sobre plantones (MOREIRA et al., 1997;SÁNCHEZ et al, 2000; 2002b).
En cuanto al posible origen de la enfer-medad en los viveros estudiados, hay quediferenciar la situación de cada uno de ellos.En el Vivero I, el suelo de monte utilizadocomo sustrato para el cultivo aparece comola fuente de inoculo más probable. El hechode que P. cinnamomi, patógeno extremada-mente virulento, haya sido aislado única-mente bajo estas condiciones especiales,hace pensar que no debe tratarse de un agen-te que habitualmente cause pérdidas en vive-ro. En cuanto al Vivero II, es más difícildeterminar el posible origen del inoculo, quepudo llegar al vivero con el sustrato de culti-vo o con el agua de riego. Hay que señalarque en este vivero se utiliza el mismo sustra-to y régimen de riegos para la producción deencinas y alcornoques y, sin embargo, sólolas primeras se vieron afectadas por la enfer-medad radical, a pesar de que nuestros expe-rimentos muestran que el alcornoque tam-bién es susceptible a Phytophthora spp. Estaaparente contradicción puede explicarse si

tenemos en cuenta las diferencias que exis-ten en el cultivo de ambas especies. La épocade siembra es más tardía para el alcornoque,mientras que el endurecimiento (preacondi-cionamiento hídrico) de esta especie ocurreantes, con objeto de evitar portes demasiadoelevados (NAVARRO et al., 1998). De estamanera, el tiempo que el alcornoque pasa enlas condiciones de saturación hídrica perió-dica que favorecen la enfermedad es sensi-blemente menor que en el caso de la encina.Este hecho, unido a la menor susceptibilidaddel alcornoque a la infección por Phytopht-hora, podría justificar la ausencia de sínto-mas en alcornoque en el vivero, al tiempoque no nos permite descartar el sustrato o elagua como el origen del inoculo. En cual-quier caso, no debe olvidarse la semillacomo otra posible vía de entrada al viverodel inoculo inicial.
Independientemente del origen del inocu-lo, la constatación de que el exceso de aguaes la condición fundamental que favorece lainfección de las raíces, implica que la mejormedida de control contra la enfermedad en elcontexto del vivero consistirá en evitar losencharcamientos periódicos que sufren losplantones (BLOOMBERG, 1985; ERWIN yRIBEIRO, 1996), bien por instalaciones inade-cuadas, excesos de riego, empleo de sustra-tos poco porosos y/o con gran capacidad deretención de agua, etc.
Hasta el momento, sólo una especie dePhytophthora (P. cinnamomi) ha sido des-crita como agente causal de podredumbresradicales en encinas y alcornoques (BRA-SIER, 1996; TUSET et al., 1996; MOREIRA etal, 1997; SÁNCHEZ et al, 2000; 2002b),aunque nunca en plantones de vivero. Estetrabajo constituye la primera descripción deP. cinnamomi, P. drechsleri y P. cryptogeacomo agentes de podredumbre radical tar-día de Q. ilex ssp. ballota en vivero. Tam-bién se ha demostrado que las tres especiesde Phytophthora constituyen un peligropotencial para la producción en vivero deQ. súber.
AGRADECIMIENTOS
Los autores quieren expresar su agradeci-miento a los Dres. Brasier (Forestry Commis-sion Research Agency, UK) y Pérez Giménez(CIFA, Málaga) por habernos proporcionadolos cultivos de referencia para los cruzamien-tos de Phytophthora spp. Este trabajo ha sidofinanciado por los Proyectos de InvestigaciónFO96-006 y AGL2002-00530, así como porel Convenio suscrito entre la Consejería deMedio Ambiente de la Junta de Andalucía y laUniversidad de Córdoba para el estudio de laSeca de Quercus en Andalucía. ME. Sánchezdisfruta de un contrato del Programa "Ramóny Cajal" del MCyT.
ABSTRACT
SÁNCHEZ M.E., S. ANDICOBERRY, A. TRAPERO. 2004. Pathogenicity of Phytophthoraspp. causing root rot of Quercus ilex ssp. ballota in forest nurseries. Bol. San. Veg. Pla-gas, 30: 385-401.
Within a research project on quality of plants for forestation of agricultural lands, westudied the etiology of a root rot affecting holm oak (Quercus ilex ssp. ballota) seedlingsin two forest nurseries in southern Spain. Major disease symptoms were foliar wiltingand necrosis of feeder roots. Three fungal species, Phytophthora cinnamomi, P. crypto-gea and P. drechsleri, were isolated from necrotic roots of holm oak. Isolates of the threespecies showed similar growth patterns at different temperatures with optimum tempera-tures near 24 °C. Selected isolates of the three Phytophthora species were pathogenic toQuercus ilex ssp. ballota and Q. suber seedlings in artificial inoculations. Soil floodingconditions were essential for infection and root rot development. There was not host spe-cificity among fungal species, being the isolates of P. cinnamomi the most virulent in allinoculated plants. In these inoculations, Q. ilex ssp. ballota plants were more susceptible

than those of Q. suber were. This work is the first report of P. cinnamomi, P. drechsleriand P. cryptogea affecting Q. ilex ssp. ballota in forest nurseries.
Key words: Phytophthora cinnamomi, P. cryptogea, P. drechsleri, Quercus rotundi-folia, Q. suber.
REFERENCIAS
BLOOMBERG WJ. 1985. The epidemiology of forest nur-sery diseases. Annu. Rev. Phytopathol., 23: 83-96.
BRASIER, CM. 1996. Phytophthora cinnamomi and oakdecline in southern Europe. Environmental cons-traints including climate change. Ann. ScilFor., 53:347-358.
BROWNE, G.T., MIRCETICH, S.M. 1988. Effects of floodduration of the development of Phytophthora rootand crown rots of apple. Phytopathology, 78: 846-851.
BUTIN, H. 1995. Tree diseases and disorders. OxfordUniv. Press, Oxford.
ERWIN, D.C., RIBEIRO, O.K. 1996. Phytophthora dise-ases worldwide. APS Press, St. Paul, Minnesota.
DHINGRA, O.D., SINCLAIR, J.B. 1995. Basic plant patho-logy methods. CRC Press, Boca Raton, Rórida.
GÓMEZ-JOVER, F., JIMÉNEZ, F. J. 1997. Forestación detierras agrícolas. MAPA-Universidad de Córdoba,Madrid.
JEFFERS, N.S., MARTIN, J.B. 1986. Comparison of twomedia selective for Phytophthora and Pythium spe-cies. Plant Dis., 70: 1038-1043.
JUZWIK, J., GUST, K.M., ALLMARAS, R.R. 1998. Influen-ce of cultural-practices on edaphic factors related toroot disease in Pinus nursery seedlings. Plant andSoi/,207: 195-208.
MANION, P.D. 1991. Tree diseases concepts. Prentice-Hall, London.
MOREIRA, A.C., FERRAZ, J.F.P., CLEGG, J.M. 1997. Sus-ceptibility of Quercus suber and Q. rotundifolia toinfection to Phytophthora cinnamomi. Proceedingsdel 10th Congress of the Mediterranean Phytopat-hological Union, Montpellier. pp: 427-431.
NAVARRO, R.M., CAMPO, A. D. DEL, ALEJANO, R., ALVA-REZ, L. 1998. Caracterización de calidad final deplanta de encina, alcornoque, algarrobo y acebucheen cinco viveros en Andalucía. Informaciones Téc-nicas 53/98. Junta de Andalucía, Sevilla.
PEÑUELAS, J. L., OCAÑA, L. 1996. Cultivo de plantasforestales en contenedor. MAPA. Mundi-Prensa,Madrid.
PETTnr, T.R., FINLAY, A.R., SCOTT, M.A., DAVIES, E.M.1998. Development of a system simulating com-mercial production conditions for assessing thepotential spread of Phytophthora cryptogea root rotof hardy nursery stock in recirculating irrigationwater. Ann. Appl. Biol., 132: 61-75.
RIBEIRO, O.K. 1978. A source book of the genus Phy-tophthora. J. Cramer, Vaduz.
SÁNCHEZ, M.E., PÉREZ DE ALGABA, A., BLANCO, M.A.,TRAPERO, A. 1998a. La "seca" de olivos jóvenes I:
Sintomatologia e incidencia de los agentes asocia-dos. Bol. San. Veg. Plagas, 24: 551- 572.
SANCHEZ, M.E., Ruiz, A., PÉREZ DE ALGABA, A., BLAN-CO, M.A., TRAPERO, A. 1998b. Occurrence andetiology of death of young olive trees in southenSpain. Eur. J. Plant Pathol, 104:347-357.
SÁNCHEZ, M.E., VARO, M.R., ANDICOBERRY; S., TRAPE-RO, A. 1999. Enfermedades que afectan a la pro-ducción de planta forestal en viveros andaluces.Agricultura, 809: 1030-1034.
SÁNCHEZ, M.E., CAETANO, P., FERRAZ, J., TRAPERO, A.,2000. El decaimiento y muerte de encinas en tresdehesas de la provincia de Huelva. Bol. San. Veg.Plagas, 26: 447-464.
SÁNCHEZ, M.E., MUÑOZ, M., BRASIER, CM., TRAPERO,A. 2001. Identity and pathogenicity of two Phy-tophthora taxa associated with a new root diseaseof olive trees. Plant Dis., 85: 411-416.
SÁNCHEZ, M.E., ANDICOBERRY, S., TRAPERO, A. 2002 a.Phytophthora root rot of Aleppo pine seedlings in aforest nursery in Spain. Plant Dis., 86: 563.
SÁNCHEZ, M.E., CAETANO, P., FERRAZ, J., TRAPERO, A.2002b. Phytophthora disease of Quercus ilex insouthwestern Spain. For. Path., 32: 5-18.
SÁNCHEZ, M.E., GUTIÉRREZ, J., TRAPERO, A. 2002c.Botryosphaeria canker of Cistus ladanifer. PlantPathol., 51: 364-372.
STAMPS, D.T., WATERHOUSE, G.M., NEWHOOK, FJ.,HALL, G.S. 1990. Revised tabular key to the speciesof Phytophthora. CAB International, Kew, Surrey.
STEEL, R.G.D., TORRIE, J.H. 1985. Bioestadística: Prin-cipios y procedimientos. McGraw-Hill, Bogotá.
TRAPERO, A., JIMÉNEZ, R.M. 1985. Fungal wilt and rootrot diseases of chickpea in Southern Spain. Phyto-pathology, 75: 1146-1151.
TUSET, J., HINAREJOS, C , MIRA, J.L., COBOS, J. 1996.Implicación de Phytophthora cinnamomi Rands enla enfermedad de la seca en encinas y alcornoques.Bol. San. Veg. Plagas, 22: 491-499.
WATERHOUSE, G.M. 1963. Key to the species of Phy-tophthora de Bary. CMI Mycological Papers 92,Kew.
Wnxox, W.F., MIRCETICH, S.M. 1985. Pathogenicityand relative virulence of seven Phytophthora spp.on Mahaleb and Mazzard cherry. Phytopathology,75: 221-226.
(Recepción: 28 julio 2003)(Aceptación: 19 abril 2004)


















