
PATCH-CLAMP ANALYSIS OF THE EFFECTS OF …jeb.biologists.org/content/jexbio/163/1/65.full.pdf ·...
20
J. exp. Biol. 163, 65-84 (1992) 65 Printed in Great Britain © The Company of Biologists Limited 1992 PATCH-CLAMP ANALYSIS OF THE EFFECTS OF THE INSECTICIDE DELTAMETHRIN ON INSECT NEURONES BY M. AMAR, Y. PICHON* Departement de Biophysique, Laboratoire de Neurobiologie Cellulaire et Moleculaire du CNRS, F-91198, Gif sur Yvette Cedex, France AND I. INOUE Institute for Enzyme Research, Tokushima University, Tokushima, 770, Japan Accepted 4 September 1991 Summary 1. The mode of action of the pyrethroid insecticide deltamethrin on inexcitable embryonic cultured cockroach neurones has been investigated using the patch- clamp technique. 2. Whole-cell recordings of the current induced by step depolarizations of the cell membrane showed that concentrations of deltamethrin ranging from 10~ 8 to 5xlO~ 6 moll~' induced a small tetrodotoxin (TTX)-sensitive inward current that peaked at around -KlOmV and reversed at around +60 mV. The activation and inactivation kinetics of this current were much slower than those of the axonal sodium current in this same species and were relatively insensitive to membrane potential. Steady-state inactivation was almost absent. 3. Single-channel activity associated with the action of the insecticide was analyzed using the cell-attached configuration. Three distinct patterns of activity were found: (1) discrete single-channel events of relatively short duration, (2) long events of comparatively small amplitude and (3) complex bursts made up of a succession of openings and closings to several levels. These three patterns were analyzed quantitatively using specially designed programs. 4. The first pattern of activity could be seen in most patches. It consisted of short (l-10ms) rectangular events of comparatively small amplitude (1.5pA at rest) and very low open time probability (around 0.001). The current-voltage relationship of these small events was linear over the voltage range studied and the (extrapolated) reversal potential approximated £ Na . 5. The second pattern of activity was observed less frequently. The channels could stay open for very long periods (up to several seconds) and occasionally flickered between two or more levels. 6. The third pattern of activity was observed in many patches. During the burst, which could last from a few milliseconds to a few hundred milliseconds, the single- channel current jumped almost continuously between several levels (up to 7 or 8). *To whom reprint requests should be addressed Key words: sodium channels, patch-clamp, cultured neurones, insect, deltamethrin, Periplaneta americana.
Transcript of PATCH-CLAMP ANALYSIS OF THE EFFECTS OF …jeb.biologists.org/content/jexbio/163/1/65.full.pdf ·...

J. exp. Biol. 163, 65-84 (1992) 6 5Printed in Great Britain © The Company of Biologists Limited 1992
PATCH-CLAMP ANALYSIS OF THE EFFECTS OF THEINSECTICIDE DELTAMETHRIN ON INSECT NEURONES
BY M. AMAR, Y. PICHON*
Departement de Biophysique, Laboratoire de Neurobiologie Cellulaire etMoleculaire du CNRS, F-91198, Gif sur Yvette Cedex, France
AND I. INOUE
Institute for Enzyme Research, Tokushima University, Tokushima, 770, Japan
Accepted 4 September 1991
Summary
1. The mode of action of the pyrethroid insecticide deltamethrin on inexcitableembryonic cultured cockroach neurones has been investigated using the patch-clamp technique.
2. Whole-cell recordings of the current induced by step depolarizations of thecell membrane showed that concentrations of deltamethrin ranging from 10~8 to5xlO~6moll~' induced a small tetrodotoxin (TTX)-sensitive inward current thatpeaked at around -KlOmV and reversed at around +60 mV. The activation andinactivation kinetics of this current were much slower than those of the axonalsodium current in this same species and were relatively insensitive to membranepotential. Steady-state inactivation was almost absent.
3. Single-channel activity associated with the action of the insecticide wasanalyzed using the cell-attached configuration. Three distinct patterns of activitywere found: (1) discrete single-channel events of relatively short duration, (2) longevents of comparatively small amplitude and (3) complex bursts made up of asuccession of openings and closings to several levels. These three patterns wereanalyzed quantitatively using specially designed programs.
4. The first pattern of activity could be seen in most patches. It consisted ofshort (l-10ms) rectangular events of comparatively small amplitude (1.5pA atrest) and very low open time probability (around 0.001). The current-voltagerelationship of these small events was linear over the voltage range studied and the(extrapolated) reversal potential approximated £Na.
5. The second pattern of activity was observed less frequently. The channelscould stay open for very long periods (up to several seconds) and occasionallyflickered between two or more levels.
6. The third pattern of activity was observed in many patches. During the burst,which could last from a few milliseconds to a few hundred milliseconds, the single-channel current jumped almost continuously between several levels (up to 7 or 8).
*To whom reprint requests should be addressed
Key words: sodium channels, patch-clamp, cultured neurones, insect, deltamethrin, Periplanetaamericana.

66 M. AMAR, Y. PICHON AND I. INOUE
7. The size of the different (single-channel) current levels was the same for thethree patterns of activity. Furthermore, these levels corresponded to multiples ofthe same minimum level (around 1 pA at the resting potential).
8. These results suggest that deltamethrin activates several states of one orseveral populations of sodium channels.
Introduction
Agents that modify the properties of membrane ionic channels are useful toolsfor deciphering channel structure and function. Several molecules are known toactivate the sodium conductance and are often referred to as agonists by analogywith receptor-gated channels; they include toxins such as grayanotoxin andbatrachotoxin, alkaloids such as aconitine, veratridine and cevadine and insecti-cides such as pyrethroids and DDT. Veratridine, pyrethroid insecticides and DDThave very similar effects on the membrane: they induce repetitive firing,depolarizing afterpotentials and/or maintained depolarizations (see Ulbricht,1969; reviewed by Hille et al. 1987). This resemblance is suggestive of a commonmode of action. Thus, in his paper on the node of Ranvier, Hille (1968) suggestedthat DDT and veratridine act similarly in maintaining the sodium channels in the'open' position. A few years later, Vijverberg et al. (1972) proposed thatpyrethroids and DDT have a similar mode of action on sodium channel gating inmyelinated nerves. The same conclusions were reached more recently byLeibowitz et al. (1987) for frog skeletal muscle treated with several insecticide andalkaloid agonists of the sodium channel. Investigations with single-channel andwhole-cell voltage-clamp recordings of N18 neuroblastoma cells also support thehypothesis that veratridine modifies open channels (Barnes and Hille, 1988).
In contrast with the preceding conclusion, early voltage-clamp experiments onisolated cockroach axons revealed that micromolar concentrations of veratridineand cevadine (the two major components of veratrine sulphate) exerted theireffects through a slow voltage-dependent sodium channel activation independentof the normal fast sodium activation and inactivation processes responsible forspike production (Pichon and Boistel, 1969; Pichon, 1974). Under similarexperimental conditions, the main effect of DDT was to maintain in the openposition a certain proportion of the sodium channels (Pichon and Boistel, 1969), asin the node of Ranvier (Hille, 1968) and lobster axons (Narahashi and Haas, 1968).More recent experiments on the same preparation confirmed the previousobservations with DDT and showed that the mode of action of several pyrethroidmolecules was different, resembling that of veratridine (Laufer et al. 1984; Pichonand Pelhate, 1985; Pichon et al. 1985, 1987). In a recent study of the kineticproperties of sodium channels of rat neuroblastoma cells modified by thepyrethroid fenvalerate, Holloway et al. (1989) reached a similar conclusion. Ourrecent observation that micromolar concentrations of veratridine induce a TTX-sensitive inward current underlain by a complex single-channel activity ininexcitable cultured embryonic cockroach neurones strengthens this hypothesis(AmsLietal. 1991).

Deltamethrin effects on insect neurones 67
Part of the observed discrepancy could be due to species differences and also, asdiscussed by Amar et al. (1991), to the use, in the former experiments, of highconcentrations of agonists dissolved in most cases in strong solvents such asdimethylsulphoxide (DMSO). Since it is known that, in insects, as in most otherpreparations, the major effect of deltamethrin is to depolarize the nervemembrane slowly at submicromolar concentrations, even in the absence of nervestimulation (Heilig, 1984; Laufer et al. 1984; Roche et al. 1985; Guillet et al. 1986;Pichon et al. 1986, 1987), it was important to study in detail the effects of suchconcentrations on an insect preparation. The present paper describes the effects ofthe pyrethroid molecule deltamethrin on embryonic cockroach neurones inculture. The goals of this study were (1) to determine whether deltamethrin couldactivate a sodium conductance in otherwise inexcitable neurones and to character-ize this conductance and (2) to analyze the single-channel activity that underliesmembrane depolarization in deltamethrin-treated neurones.
A preliminary report of some of these results has been published elsewhere(Amar and Pichon, 1989).
Materials and methods
Cultures
Neuronal cultures were prepared from the brains of 21-day-old embryos ofPeriplaneta americana (L.) as described by Dewhurst and Beadle (1985). Briefly,the egg cases (which contained up to 16 embryos) were sterilized with absoluteethanol for lOmin and opened and the heads of the embryos removed and storedin Schneider's revised Drosophila medium. The brains were then extracted fromthe head capsules and the cells dissociated by gentle trituration with a Pasteurpipette. The cultures were initiated in a second medium made up of five parts ofSchneider's medium and four parts of Eagle's basal medium containinglOOi.u.ml"1 penicillin and lOO/^gml"1 streptomycin: three drops of the cellsuspension were placed in the middle of a 50 mm Falcon light-seal plastic Petri dishand covered with a sterile glass coverslip. After 1 h, the culture dishes were turnedupside down, to separate the live cells from the debris, and stored in an incubatorat 29°C. After 7 days, the culture medium was replaced by a third medium madeup of equal parts of Leibovitz's L-15 and Yunker's modified Grace mediumcontaining penicillin and streptomycin supplemented with foetal calf serum (7 %) .In the second medium, the vast majority of the glial cells did not survive and littlegrowth was seen. The third medium enabled the fast development of neuriticprocesses into a dense network. All culture media were obtained from Gibco Ltd(Life Technologies), Paisley, Scotland.
Experimental procedure
Immediately before an experiment, the culture medium was replaced by astandard saline containing 210mmoir1 NaCl, S.lmmoll"1 KC1, lOmmolP1
CaCl2 and lOmmolT1 Hepes buffer, adjusted to pH7.2 using NaOH. The dish

68 M. AMAR, Y. PICHON AND I. INOUE
was placed directly onto the stage of a Nikon TMS inverted microscope and thecells were observed under phase contrast. All experiments were performed atroom temperature (24-26 °C).
For the experiments using the whole-cell configuration of the patch-clamptechnique (Hamill et al. 1981), the electrodes were filled with a solution containing140 mmol P 1 potassium gluconate (normal-K+ internal solution) or caesiumaspartate (K+-free internal solution), lOmmoir1 EGTA, Immoll"3 CaCl2,10 mmol I"1 MgCl2 and 10 mmol P 1 Hepes buffer, adjusted to pH 7.2 using NaOH.Whenever needed, tetraethylammonium (TEA+) chloride (100 mmol I"1) and/or4-aminopyridine (4-AP) (10 mmol I"1) was added to the solution to block thepotassium current and tetrodotoxin (TTX) (3,umolPJ) was used to block thesodium current. Deltamethrin was first dissolved in a small volume of ethanol.Appropriate volumes of this stock solution were added to the saline contained inthe patch pipette (internal solution). Deltamethrin ( 1 R , 3 R , O S ) was a gift fromProcida (Roussel UCLAF, Marseilles); the other compounds were purchasedfrom Sigma.
Electrophysiological techniques
Electrodes were pulled from 1.5 mm borosilicate microhaematocrit tubing,using a Narishige micropipette puller, and fire polished. Their resistance rangedfrom 2 to 5MQ. The electrode was advanced towards the cell soma (10-15 t̂m indiameter) until gentle contact was established. A small negative pressure was thenusually sufficient to create a seal of 1—10GQ. For whole-cell recordings, themembrane patch was subsequently destroyed by applying a short (0.1-0.5 ms)1.5 V pulse across the membrane.
The patch electrode was connected to the input stage of an Axon Instruments(Axopatch 1C) patch amplifier. The outputs of the amplifier were connected to aTektronix oscilloscope, to the input stages of a modified Sony digital audioprocessor (PCM 701) connected to a Sony Betamax video recorder (Lamb, 1985)and to the input amplifier of a laboratory-made data-acquisition and stimulationsystem (DAISY-12C) connected to a Hewlett Packard series 9000 model 310microcomputer.
For the whole-cell clamp experiments, the command voltage steps weresupplied by the Hewlett-Packard-based system and the resulting currents weredigitized, after amplification and partial analogue compensation of the capacita-tive artefacts, and stored onto 3.5 inch floppy disks. For cell-attached experiments,the membrane potential was changed using the holding potential command of thepatch amplifier.
Data analysis
Whole-cell currents were analyzed on the Hewlett Packard microcomputer: theleak current corresponding to a 20 mV hyperpolarizing voltage pulse was sub-

Deltamethrin effects on insect neurones 69
tracted, after scaling, from the stored current traces and the resulting currentdisplayed on the screen and plotted on a digital plotter. The corrected recordswere stored for further analysis.
Single-channel data were analyzed on an IBM-AT-compatible microcomputer.Twenty-second-long recordings were selected from the video tape, digitized usinga data translation (DT 2801A) A/D converter and stored directly onto the harddisk. The resulting 12-bit binary files were then analyzed. In the early experiments,the program called IPROC-2, originally developed by Sachs et al. (1982), was used(see Beadle et al. 1989). Later analyses were performed using an original programcalled PAT2, a program derived from ANALYSIS (Pichon and Pichon, 1989) anddesigned specifically for fast analysis and display of long recordings with complexchannel behaviour. The outputs of both programs consisted of a series of files thatwere subsequently used, in connection with a statistical library, to build histo-grams.
All results are given as mean±s.D. unless stated otherwise.
ResultsWhole-cell clamp experiments
lAOmmoir1 K+ internal solution
Under these experimental conditions, embryonic cockroach neurones in culturehad a relatively low resting potential of -45±6.9mV (N=25) and were electricallyinexcitable. Under most conditions, however, strong voltage depolarizations inthe current-clamp mode gave rise to small (less than 15 mV) biphasic spikesoriginating from the axonal processes, indicating that, as in most adult insectneurones, the cell bodies were inexcitable and lacked fast voltage-sensitive sodiumchannels although normal sodium spikes were present in the axons (see Pichon,1974). Under voltage-clamp conditions, depolarizing voltage steps failed to elicitany significant inward current but gave rise to a large outward current. Varying theholding potential between —100 mV and — 40 mV or using long-duration(200-400 ms) hyperpolarizing conditioning prepulses failed to de-inactivate apotentially inactivated sodium conductance.
The addition of low concentrations of deltamethrin (l-S^moll"1) to thesolution contained in the patch pipette had a slight depolarizing effect on theresting potential (—39±7.5mV, N=9 for SjumolP1 deltamethrin) and induced asmall voltage-dependent inward current. This current turned on soon aftermembrane depolarization, reached a peak value of 50-100pA (mean -81±26pA,7V=7) during the first few milliseconds, then decreased and was replaced by a largeoutward potassium current (Fig. 1A). The maximum current was obtained forcommand steps to a membrane potential of about +10mV and the apparentreversal potential (calculated by extrapolation from the linear portion of the I-Vcurve) was around +60 mV. This inward current was sensitive to the addition oftetrodotoxin to the bath.

70 M. AMAR, Y. PICHON AND I. INOUE
2.5 ms 20 msD
-150
0-
-50-
-100-
15(1-
A
X«
\
is
i /
A
- 6 0 - 3 0 0 30 - 6 0 - 3 0
Membrane potential (mV)
30 60
Fig. 1. Ionic currents recorded from the soma of cultured cockroach neurones in thewhole-cell clamp configuration in the presence of 10~6moll~' deltamethrin.(A) Family of membrane currents corresponding to step membrane depolarizationsfrom a holding potentials of — 60 mV to membrane potentials between —50 and+60mV in 10 mV increments (internal solution: potassium gluconate). (B) Family ofmembrane currents corresponding to step membrane depolarizations from a holdingpotential of -60 mV to membrane potentials between -50 and OmV in 10 mVincrements (internal solution: caesium aspartate). (C) Current-voltage relationshipfor the peak sodium current recorded before (A) and after a 3min perfusion with3^molP' TTX (A); holding potential — 60mV. (D) Current-voltage relationship forthe peak sodium current corresponding to step depolarizations to the indicatedpotentials from three different holding potentials: -50mV (V), -60mV (A) and-70mV(A). InB, C and D, the outward current was blocked with lOmmoir' TEA"1"andO.lmmoir1 4-AP.

Deltamethrin effects on insect neurones 71
K+-free internal solution containing caesium
To analyze the time course of the deltamethrin-induced sodium current, most ofthe outward current was removed by replacing potassium gluconate in the internalsolution with caesium aspartate. The mean resting potential under these con-ditions was -47.7±2.6mV (N=3) in the absence and -42±14.4mV (N=6) in thepresence of potassium channel blockers in the bathing solution. The current-voltage relationship of the deltamethrin-induced peak inward current, measuredunder voltage-clamp conditions for neurones in which the turning on of the currentwas graded with potential, was basically the same as with potassium gluconate; themaximum current was larger, as was to be expected from the inhibition of theoutward current (maximum inward current around -120pA). Here again, thepeak current was obtained for a membrane potential of about +10mV and themean (extrapolated) equilibrium potential was around +60 mV (Fig. 1C). Thiscurrent was partly inhibited by bath application of TTX (3/imolP1) (Fig. 1C).
Kinetics of the deltamethrin-induced current
In nearly 70% of the 20 neurones internally perfused with caesium aspartate,the turning on of the deltamethrin-induced inward current was not graded andsome current traces exhibited notches, both features reflecting poor control of themembrane potential of the cell. These cells were discarded and the analysis wasperformed on the remaining 30 %. In these remaining cells, the time course of thesodium current was characterized by a comparatively fast rising phase followed bya slow inactivation (Fig. IB). For membrane potentials between -40mV and+20 mV (values for which the inward current was large and the outward currentwas negligible), the current traces were fitted with a combination of an exponen-tially rising phase, an exponentially falling phase and a plateau. In most cases, as inthe experiments illustrated in Fig. IB, the time constants exhibited little, if any,voltage sensitivity between —30 and OmV (mean time constant of activation3.6±0.64ms, mean time constant of inactivation 46±20ms for the cell illustratedin Fig. IB).
Steady-state inactivation
Changing the holding potential of the cells from -80mV to -40mV was foundto have little effect on the time course and the intensity of the deltamethrin-induced current. This feature is illustrated in Fig. ID (for the intensity). Long-lasting changes in the holding potential (several minutes) were, however, found tomodulate the size of the current in the direction expected for long-terminactivation; thus, the peak current was consistently smaller for a holding potentialof —60 mV if the membrane had been held for some time at - 40 mV. Conversely,the peak current was larger than the control if the membrane had previously beenheld at -80 mV.
Single-channel experiments
Single-channel activity induced by deltamethrin was studied using the cell-

72 M. AMAR, Y. PICHON AND I. INOUE
5pA
v •
40 ms
Fig. 2. Various patterns of single-channel activity induced by applying 0.5(Umoll 'deltamethrin onto the membrane of a cockroach neurone. Cell-attached configurationof the patch-clamp technique. Patch held at H90mV (i.e. hyperpolarized by 90 mVcompared to rest). Outward current blocked with TEA+ (lOmmolF1) and 4-AP(lmmolF1). Low-pass filter set at 3 kHz; sampling frequency 10 kHz. Note thepresence in the same recording of rectangular events (V), of long openings (•) and ofcomplex bursts of activity (•)
attached configuration of the patch-clamp technique. Deltamethrin was added tothe solution contained in the patch pipette at concentrations ranging from 0.01 tolOjitmoir1.
Under normal conditions (i.e. in the absence of any agonist in the bathingmedium or in the patch pipette and with lOmmolP1 Ca2+ in the externalsolution), the single-channel activity observed in these neurones was outwardlydirected and corresponded to that of voltage-dependent and/or calcium-depen-dent potassium channels (Christensen et al. 1988). It was blocked in the presentexperiments using high concentrations of TEA+ and 4-AP so that no detectableactivity could be recorded.
Addition of deltamethrin to the pipette solution induced characteristic, butcomplex, single-channel activity (Fig. 2) that was totally abolished in the presenceof TTX. Three distinct patterns of activity could be observed: relatively simplerectangular openings of variable duration, long openings of variable amplitudeand bursts of complex events. The three patterns were often mixed in the samerecording and were not characteristic of any given patch. The effects of membranepotential on the first pattern of activity (which is easier to analyze) are described inthe following section. The properties of the other two patterns are brieflydescribed in the last two sections.

Deltamethrin effects on insect neurones 73
Effects of membrane potential
The effects of deltamethrin on single-channel activity were investigated for awide range of potentials ranging from 150 mV in the hyperpolarizing direction to100 mV in the depolarizing direction (approximately —190 mV to +60 mVabsolute membrane potential, based on an estimated resting potential of-40mV). However, for depolarized membrane potentials, reliable estimates ofthe amplitude and kinetics of the single-channel events were seriously hamperedby two factors: the comparatively large background noise (1-1.5 pA peak to peak)at 3 kHz and the presence of a small population of (outward) potassium channelsthat were not totally blocked by TEA+ and 4-AP. The quantitative analysis was,therefore, limited to potentials ranging from 150 to — 20mV (i.e. approximately-190mV to -20mV absolute membrane potential).
Effect of membrane potential on the mean single-channel current and on single-channel amplitude distribution. As in the experiment illustrated in Fig. 3, the
H20mV
H30mV
H4QmV
H50mV
H60mV
5pA
50 ms
Fig. 3. Effect of membrane potential on single-channel activity induced by theaddition of O.l^moll"1 deltamethrin to the solution contained in the patch pipette.Cell-attached configuration of the patch-clamp technique. Outward current blockedwith TEA+ (lOmmoir1) and 4-AP (Immoir1). The activity was recorded at fivedifferent potential levels, as indicated; hyperpolarized by 20-60 mV compared withrest. Low-pass filter set at 3 kHz, sampling frequency 10 kHz. In this recording, onlytwo categories of events could be distinguished: rectangular openings and shortunresolved events. These two categories of event, which could be superimposed, asshown for H40 mV, increased in size but not in duration with membrane hyperpolariz-ation.
74 M. AMAR, Y. PICHON AND I. INOUE
<a.
1-
0-
- 1 -
- 2 -
- 3 -
- V
A _
/ • *A
D
i
•
\ <£5
-150 -100 -50 0 50
Pipette potential (mV)
100 150
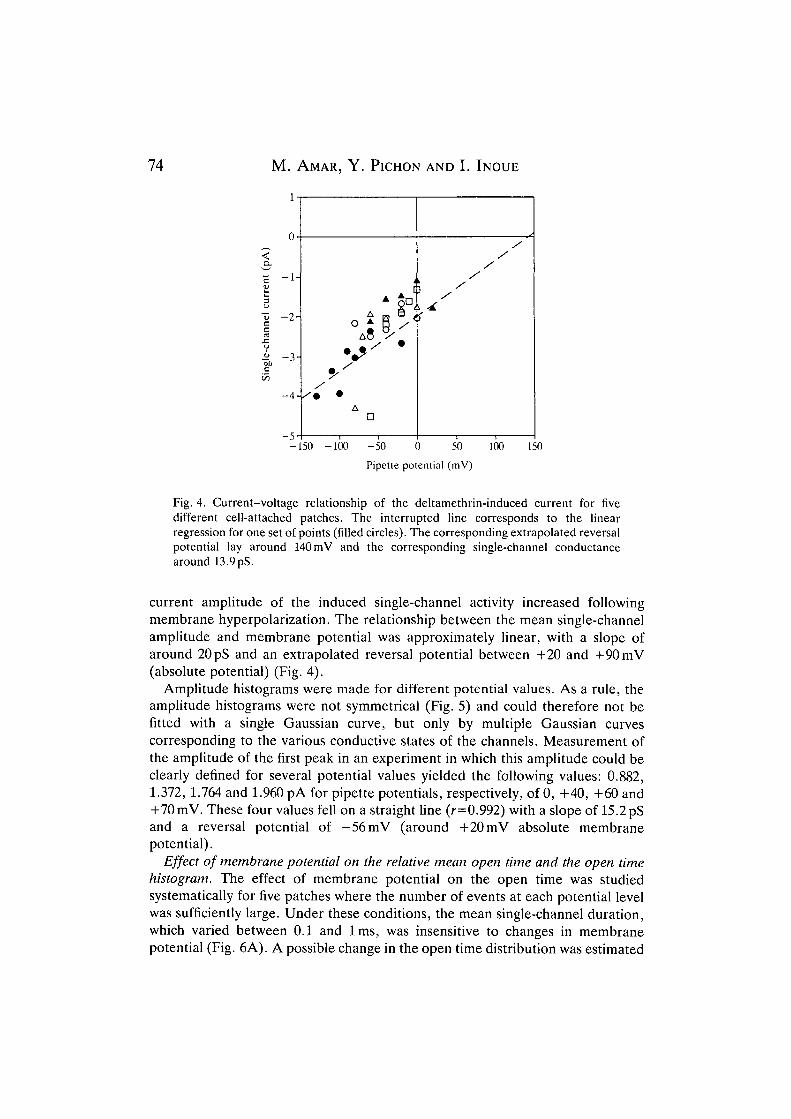
Fig. 4. Current-voltage relationship of the deltamethrin-induced current for fivedifferent cell-attached patches. The interrupted line corresponds to the linearregression for one set of points (filled circles). The corresponding extrapolated reversalpotential lay around 140 mV and the corresponding single-channel conductancearound 13.9 pS.
current amplitude of the induced single-channel activity increased followingmembrane hyperpolarization. The relationship between the mean single-channelamplitude and membrane potential was approximately linear, with a slope ofaround 20 pS and an extrapolated reversal potential between +20 and +90 mV(absolute potential) (Fig. 4).
Amplitude histograms were made for different potential values. As a rule, theamplitude histograms were not symmetrical (Fig. 5) and could therefore not befitted with a single Gaussian curve, but only by multiple Gaussian curvescorresponding to the various conductive states of the channels. Measurement ofthe amplitude of the first peak in an experiment in which this amplitude could beclearly defined for several potential values yielded the following values: 0.882,1.372, 1.764 and 1.960 pA for pipette potentials, respectively, of 0, +40, +60 and+70mV. These four values fell on a straight line (r=0.992) with a slope of 15.2 pSand a reversal potential of -56mV (around +20 mV absolute membranepotential).
Effect of membrane potential on the relative mean open time and the open timehistogram. The effect of membrane potential on the open time was studiedsystematically for five patches where the number of events at each potential levelwas sufficiently large. Under these conditions, the mean single-channel duration,which varied between 0.1 and lms, was insensitive to changes in membranepotential (Fig. 6A). A possible change in the open time distribution was estimated

Deltamethrin effects on insect neurones 75
200 T
1 5 0 - •
e
100--
50- -
•+•2 3
Amplitude (pA)
Fig. 5. Amplitude histogram of the 'single-channel' activity induced bydeltamethrin. Holding potential H70mV (70mV hyperpolarized compared with rest).The histogram shows a main peak at approximately 1.3 pA but cannot be fitted with asingle Gaussian curve, reflecting the existence of multiple conducting states.
10.00
I l.OOt-c
0.10
0.01-100 - 6 0 - 2 0 20
Pipette potential (mV)
1O.
10
• U "
~21
- 3 :
- 4 .
- 5 -
B
A
•
A
•O
A
-f—
O
A
DO A
A ^
A
—1
O
—1
10
io-4i•
-100 - 6 0 - 2 0 20
Pipette potential (mV)
60
Fig. 6. Effects of membrane potential on the relative mean open time (A) and therelative open time probability (B) of the deltamethrin-induced single-channel activityfrom different cell-attached patches. Despite a large scatter, the same general trendscan be observed. (A) The mean open time was in the millisecond range and showedlittle voltage-dependency. (B) The relative open time probability was very low (0.001)at the resting potential level and increased significantly with membrane hyperpolariz-ation. The interrupted line in B corresponds to an exponential fit of the pooled data(N=46) and had a correlation coefficient of 0.627.

76 M. AMAR, Y. PICHON AND I. INOUE
by comparing the duration histograms of the same patch held at five differentholding potentials. It was found that there was no significant change in thedistribution between 0 and 80mV (not shown).
Effects of membrane potential on the relative open time probability. The effect ofmembrane potential on the relative open time probability was studied for fivemembrane patches as above and the results are summarized in Fig. 6B. Therelative open time probability, which was very low (around 0.001) at the restinglevel, increased significantly to approximately 0.01 when the membrane washyperpolarized by 100 mV.
Long-duration channel openings
Long-duration channel openings were studied on selected records. The sam-pling frequency was reduced fivefold to 2 kHz and the data were filtered at 1 kHz.Selected records of some of these events are illustrated in Fig. 7A. Their open timeand amplitude varied widely even in the absence of any change in the holdingpotential (here 60 mV). The amplitude histogram illustrated in Fig. 7B is typical: itis asymmetrical and exhibits several peaks. As illustrated in Fig. 7C for the samedata, the open time histogram of these channels resembled that of normal eventson a 50 times expanded time scale.
Bursting channels
The properties of the bursts were studied on selected records filtered at 1 kHzand acquired at 10 kHz. During a burst, two categories of events could bedistinguished: very short triangular events (i.e. events that were not adequatelyresolved owing to the limited bandwidth of the recording system), during whichthe trace jumped directly from zero to one level or another and jumped backalmost immediately to zero, and longer complex events during which the tracemoved almost continuously from one level to another (Fig. 8A). Analysis of suchbursts gave amplitude histogram distributions very similar to those observednormally but included many more events. The histogram is again asymmetrical,sometimes consisting of a succession of reasonably well-resolved peaks. In theanalysis illustrated in Fig. 8B, the peaks were located at 0.976, 1.954, 2.93 and4.1 pA. These values, which are multiples of 1 pA, corresponded almost exactly tothe levels reached by the brief events. The duration histogram (Fig. 8C) could befitted by two exponentials with time constants almost identical to those of normalevents.
Discussion
The data presented here clearly indicate that low concentrations of deltamethrinactivate a small TTX-sensitive sodium conductance in cultured embryonicneurones of the cockroach.
Whole-cell recordings
The kinetics of the current are much slower and less sensitive to changes in

Deltamethrin effects on insect neurones 11
"T^ftv^as^^
20
2 15
° 10
E3
z 5
UnM n lln rm
^ ^ ^
150
SJ 100
I- C
\^i^^^
o5E3
z50
5pA
1 2 3 4Amplitude (pA)
250Duration (ms)
500
200 ms
Fig. 7. Long-duration single-channel activity induced by 0.5 jumol 1 l deltamethrin in acell-attached patch held hyperpolarized by 60 mV compared with rest. Outwardcurrent was blocked with TEA+ (lOmmoll"1) and 4-AP (lmmolT1). (A) Selectednon-consecutive traces of the single-channel activity. Low-pass filter set at 1kHz,sampling frequency 2 kHz. The interrupted line indicates zero current. Note that thesize and the duration of the single-channel events were variable. (B, C) Amplitude (B)and duration (C) histograms of the events illustrated in A analyzed using the PAT2program with a minimum open time of 2 ms and a minimum closed time of 25 ms. Asexpected from A, the amplitude of the single-channel activity could not be fitted by asimple Gaussian function. The duration histogram was best fitted by two exponentialfunctions with a fast time constant of 4.3 ms and a slow time constant of 86 ms(r=0.9979) (interrupted line).
membrane potential than the normal sodium currents. In some respects, thekinetics of the deltamethrin-induced current are reminiscent of those of thecalcium current in adult cockroach neurones (M. Amar and Y. Pichon, inpreparation). A further important difference between the deltamethrin-inducedcurrent and the fast axonal sodium current is the absence of steady-stateinactivation.
As expected from our previous experiments with pyrethroid insecticides, thecurrent-voltage relationship differs significantly from that of the normal axonal

78 M. AMAR, Y. PICHON AND I. INOUE
B150 T
a; 100-
"o
00
400 C
10Amplitude (pA)
5pA
2 300c
>
"° 200
|Z 100
25 ms
10
Duration (ms)
Fig. 8. Bursting activity induced by 5^moll~' deltamethrin in a cell-attached patchheld hyperpolarized by 60 mV compared with rest. Outward current was blocked withTEA+ (lOmmoir1) and 4-AP (Immoir1). Low-pass filter set at 1kHz, samplingfrequency 10 kHz. (A) Selected non-consecutive tracings showing the single-channelactivity during a burst. For each recording, the upper interrupted line corresponds tozero current and the four others to the main peaks of the amplitude histogramillustrated in B. As in Fig. 2, the events that constituted the bursts could be divided intotwo categories: simple short unresolved events and complex events during which thecurrent jumped between up to six or seven distinct levels, probably corresponding tosubstates of the sodium channel. (B,C) Amplitude (B) and duration (C) histograms ofthe events illustrated in A analyzed using the PAT2 program with both minimum openand minimum closed time set to 0. Here again, the amplitude histogram wasasymmetrical with a main peak at 0.976 pA and a succession of smaller peaks at 1.954,2.93 and 4.10 pA (V). The duration histogram was fitted by a double exponentialfunction with a fast time constant of 0.73 ms and a slow time constant of 4.4 ms(r=0.999) (interrupted line).
(Pichon and Boistel, 1967; Pichon, 1968, 1974) or neuronal (Lapied et al. 1990;M. Amar and Y. Pichon, in preparation) sodium currents. The peak current isdisplaced by 20-30 mV towards more depolarized potentials. As illustrated inFig. 9, the voltage-dependency of the deltamethrin-induced sodium current is

Deltamethrin effects on insect neurones 79
-40 0 40
Membrane potential (mV)
80
Fig. 9. Relationship between membrane potential and the pyrethroid-induced sodiumconductance. The open triangles correspond to the tail current induced byS-bioallethrin in cockroach axons (modified from Pichon and Pelhate, 1985), the filledtriangles correspond to the whole-cell current induced by 1/^molP1 deltamethrin incultured cockroach neurones. The interrupted line corresponds to the best fit of theaxonal data of equation 1 of Hodgkin and Huxley (1952) (y=l/[l+e(l/"t/m)/7])) thedotted line represents the voltage-dependency of the fast sodium current of cockroachaxons (Pichon, 1974).
almost identical to that of the tail current recorded after application ofS-bioallethrin (Pichon and Pelhate, 1985) or methanotetramethrin (Laufer etal.1984; Pichon et al. 1987) to the cockroach axonal preparation. The current density(around lOOjuAcirT2 assuming a cell diameter of lOjum) is very close to thatreported for similar concentrations of insecticide molecules on the axon (Pichonetal. 1987). These results are in full agreement with our proposed hypothesisconcerning the mode of action of pyrethroid insecticides.
In most, but not all, respects the effects of deltamethrin were reminiscent ofthose of similar concentrations of veratridine on the same preparation (Amar et al.1991). The origin of the induced conductance remains speculative. The simplestinterpretation of our data, which fits in well with experiments by other authors onnon-excitable tissues or cultured cells (Romey etal. 1979), would be that theinsecticide unveils 'silent' channels (i.e. channels that are present in the membranebut are not functional). The sensitivity to TTX supports this interpretation.Another interpretation would be that functional sodium channels are present inthe membrane but their opening time is so short that they contribute very little tothe whole-cell current and they are not detectable under patch-clamp conditionsbecause of the limited bandwidth of the patch amplifiers. The effect of theinsecticide molecule would be to lengthen the open time of these channels so thatthe net current increases and the single-channel activity becomes detectable.
The shift from the current-voltage relationship of the deltamethrin-inducedsodium current to that of the fast axonal or neuronal sodium currents observed in

80 M. AMAR, Y. PICHON AND I. INOUE
the same species is reminiscent, as for veratridine (Amar et al. 1991), of thatobserved by Goldin et al. (1990) for the sodium current induced by rat brain IIAcvsubunit of the voltage-gated sodium channel injected into Xenopus oocytes. It isalso suggestive of a modification of the (voltage-sensitive) S4 segment of domain IIof the channel protein.
Single-channel recordings
Concentrations of deltamethrin ranging from 10~8 to 5x lO~ 6 moi r ' inducedcomplex but characteristic single-channel activity. Despite this complexity, thecurrent-voltage relationship of the mean and peak currents could usually be fittedwith a straight line with an extrapolated reversal potential between 30 and 90 mV(i.e. not very different from E^a). The corresponding mean single-channelconductance (20 pS) is very similar, although slightly lower, than that observedwith similar concentrations of veratridine on the same preparation (25.6 pS, Amaret al. 1991) and is of the same order of magnitude as that reported for normalsodium channels in neuroblastoma cells (Quandt and Narahashi, 1982; Nagy et al.1983; Yamamoto et al. 1984; Chinn and Narahashi, 1986,1990; Nagy, 1987; Barnesand Hille, 1988), for vertebrate muscle sodium channels (Sigel, 1987a), forneuronal sodium channels from wild-type Drosophila (Stiihmer et al. 1987) and formutant Drosophila neuronal sodium channels (Stiihmer et al. 1989) expressed inXenopus oocytes. It is slightly lower than the 21-25 pS reported for chick neuronalsodium channels expressed in Xenopus oocytes (Sigel, 19876).
Another similarity between our single-channel data and those obtained on otherpreparations with or without treatment with sodium channel 'agonists', such asbatrachotoxin, veratridine or pyrethroid insecticides, is the presence of channelswith more than one conducting state. The conclusion that the recorded activitycorresponds to multiple states of the same channel rather than to the superimpo-sition of a number of independent channels is based on several considerations. Thefirst is that, if one considers the extremely low open time probability of the single-channel events under our experimental conditions (around 1%, Fig. 6), thenumber of superimposed events greatly exceeds that predicted from the randomsuperimposition of independent channels. The second argument is the observationthat the number of transitions exceeding one or two unitary levels is several ordersof magnitude larger than predicted on the basis of independent channels. Thethird argument is based on the statistical distribution within the bursts of thevarious time courses of 'complex' events (see Colquhoun and Hawkes, 1983). Thiswas particularly obvious in the case of 'bursting' channels, where up to seven oreight 'levels' could be clearly separated, the lowest level having a calculatedconductance of 6pS, i.e. about one-third of the mean conductance value. Quiteunexpectedly, and at variance with previous reports and our own observations onveratridine-treated neurones, the levels appeared to be exact multiples of thesmallest unitary conductance. This observation is consistent with the hypothesisproposed by Sigel (19876) that the small conductance could represent the realunitary conductance of the sodium channel and that the normally observed

Deltamethrin effects on insect neurones 81
4 6
Size of transitions
10 4 6
Conductance level
Fig. 10. Example of the distribution of the transitions and the conductance levelsduring a burst of activity induced by 10"6moll~1 deltamethrin. (A) Number ofoccurrences of each transition category plotted against its size. (B) Number ofoccurrences of each conductance level plotted against its size. The interrupted linescorrespond to fits of the experimental data with one exponential, the dotted linecorresponds to a fit of levels 3-10 with one exponential. The transition size was takenas the size of an abrupt change in current from one level to another (size 2 would, forexample, correspond to a direct transition from level 0 to level 2 or to a direct transitionfrom level 3 to level 1). The level size was taken as the value of the current after eachstep. Both sizes were expressed as multiples of the unitary current, which was taken asone. For explanations, see text. Membrane potential hyperpolarized by 60 mV.
10
conductances reflect the cooperative opening of several elementary channels(corresponding, for example, to three of the four a-subunits of the sodium channelprotein).
The large number of substates observed in our experiments could reflect thecomplexity of a primitive sodium channel and the fact that our experiments wereperformed at a comparatively high temperature (24-26°C). Thus, in recentexperiments on deltamethrin-treated neuroblastoma cells, Chinn and Narahashi(1990) observed that the number of subconducting states was greater at roomtemperature than at 11 °C. To test these hypotheses, it would be of interest torepeat our experiments on dissociated adult cockroach neurones that expressfunctional sodium channels and to repeat the experiments at a lower temperature.
One interpretation of our data could be that the observed activity results fromthe transient aggregation of a variable number of (possibly identical) subunits(Pichon and Amar, 1991). The amount of cooperativity between the elementarychannels can be estimated from the number of occurrences of each subconduc-tance level and the distribution of sizes of the transitions. Thus, from the recordingpartly illustrated in Fig. 8, the distribution of the number of transitions of eachcategory plotted against their size can be fitted with a single decaying exponentialwith a slope of 0.59±0.03 (Fig. 10A). The corresponding distribution of thenumber of occurrences of each conductance level against its size can be fitted by aplateau (between levels 1 and 3) followed by a decaying exponential (betweenlevels 3 and 10) with a slope of 0.76±0.03 (Fig. 10B). These results suggest (1) that

82 M. AMAR, Y. PICHON AND I. INOUE
the cooperativity between each individual opening is the same for all subunits, (2)that the probability of recording conductance levels 1, 2 and 3 is comparativelyhigher than that of recording higher conductance levels. It would be of interest toestablish the correlation between these results and the structure of the putativesodium channel in embryonic cockroach neurones.
Mode of action of deltamethrin
Our experiments suggest that deltamethrin activates 'silent' sodium channels.The origin of these channels remains unknown. It is interesting to note, however,that, in the central nervous system of arthropods and particularly of insects, cellbodies are generally inexcitable (Hoyle, 1970; Pichon and Ashcroft, 1985), whichagrees with our findings in cultured neurones, but may become excitable and givefairly normal overshooting sodium spikes following colchicine treatment (Pitman,1975) or dissociation (Wu etal. 1983; O'Dowd and Aldrich, 1988; Gundel et al.1989; M. Amar, unpublished observation). This indicates that, at least in theseneurones, fast sodium channels may exist in the membrane in a non-functionalconfiguration and may be transformed into functional channels under certainconditions. Insect neurones may prove to be very useful in the study of theexpression of functional sodium channels during development and followingvarious physical and/or chemical treatments.
The authors are indebted to Dr R. Kado for lending them his patch pipettepuller. M.A. was supported by a grant from the Fondation Roussel, Paris.
ReferencesAMAR, M. AND PICHON, Y. (1989). Effects of two sodium channel 'agonists': veratridine and
deltamethrin on single channel activity in inexcitable embryonic cockroach neurones.Biophys. J. 55, 315a.
AMAR, M., PICHON, Y. AND INOUE\ I. (1991). Micromolar concentrations of veratridine activatesodium channels in embryonic cockroach neurones in culture. Pfliigers Arch. 417, 500-508.
BARNES, S. AND HILLE, B. (1988). Veratridine modifies open sodium channels. J. gen. Physiol.91, 421-443.
BEADLE, D. J., HORSEMAN, G., PICHON, Y., AMAR, M. AND SHIMAHARA, T. (1989).Acetylcholine-activated ion channels in embryonic cockroach neurones growing in culture.J. exp. Biol. 142, 337-355.
CHINN, K. AND NARAHASHI, T. (1986). Stabilization of sodium channel states by deltamethrin inmouse neuroblastoma cells. J. Physiol., Lond. 380, 191-207.
CHINN, K. AND NARAHASHI, T. (1990). Temperature-dependent subconducting states andkinetics of deltamethrin-modified sodium channels of neuroblastoma cells. Pfliigers Arch.413, 571-579.
CHRISTENSEN, B. N., LARMET, Y., SHIMAHARA, T., BEADLE, D. J. AND PICHON, Y. (1988). Ioniccurrents in neurones cultured from embryonic cockroach (Periplaneta americana) brains.J. exp. Biol. 135, 193-214.
COLQUHOUN, D. AND HAWKES, A. G. (1983). The principles of stochastic interpretation of ionchannel mechanisms. In Single Channel Recording fed. B. Sakmann and E. Neher), pp.135-175. New York: Plenum Press.
DEWHURST, S. AND BEADLE, D. J. (1985). Cell and tissue culture from the insect nervous system.In Neurochemical Techniques in Insect Research (ed. H. Breer ant T. A. Miller), pp. 207-222.New York: Springer Verlag.

Deltamethrin effects on insect neurones 83
GOLDIN, A. L., AULD, V. J., HEBERT, T., KRAFTE, D. S., CATTERALL, W. A., LESTER, H. A.,DAVIDSON, N. AND DUNN, R. J. (1990). Structural and functional analysis of the voltage gatedsodium channel. Biophys. J. 57, 210a.
GUILLET, J. C , ROCHE, M. AND PICHON, Y. (1986). Effects of topical and bath applications ofthe insecticide deltamethrin on electrical activity in a leg mechanoreceptor of the cockroach,Periplaneta americana. Pest. Biochem. Physiol. 26, 183-192.
GUNDEL, J., WICHER, C , TENNIGKEIT, M. AND MATTHIES, H. (1989). Investigations of
excitability of isolated and non-isolated neurons from the terminal ganglion of Periplanetaamericana by current/voltage clamp and intracellular perfusion. Gen. Physiol. Biophys. 8,579-587.
HAMILL, O. P., MARTY, A., NEHER, E., SAKMANN, B. AND SIGWORTH, F. (1981). Improved patchclamp techniques for high resolution current recording from cells and cell-free membranepatches. Pflugers Arch. 391, 85-100.
HEILIG, U. (1984). Etude de la neurotoxicit6 de trois pyrdthrinoides chez deux especes de blattesPeriplaneta americana et Blabera craniifer Biirm. Toxicit6 globale et effets sur l'activite-electrique du systeme nerveux. These de troisieme Cycle, Paris, pp. 1-106.
HILLE, B. (1968). Pharmacological modifications of the sodium channels of frog nerve. J. gen.Physiol. 51, 199-219.
HILLE, B., LEIBOWITZ, M. D., SUTRO, J. B., SCHWARZ, J. R. AND HOLAN, G. (1987). In Proteins
of Excitable Membranes (ed. B. Hille and D. M. Fambrough), pp. 109-124. New York: JohnWhiley and Sons.
HODGKIN, A. L. AND HUXLEY, A. F. (1952). The dual effect of membrane potential on sodiumconductance in the giant axon of Loligo. J. Physiol, Lond. 116, 497-506.
HOLLOWAY, S. F., SALGADO, V. L., Wu, C. H. AND NARAHASHI, T. (1989). Kinetic properties ofsingle sodium channels modified by fenvalerate in mouse neuroblastoma cells. Pfliigers Arch.414, 613-621.
HOYLE, G. (1970). Cellular mechanisms underlying behavior: neuroethology. Adv. InsectPhysiol. 7, 349-444.
LAMB, T. D. (1985). An inexpensive digital tape recorder suitable for neurophysiological signals.J. Neurosci. Meth. 15, 1-13.
LAPIED, B., MALECOT, C. O. ANDPELHATE, M. (1990). Patch-clamp study of the properties of thesodium current in cockroach single isolated adult aminergic neurones. J. exp. Biol. 151,387-403.
LAUFER, J., ROCHE, M., PELHATE, M., ELLIOTT, M., JANES, N. F. AND SATTELLE, D. B. (1984).
Pyrethroid insecticides: actions of deltamethrin and related compounds on insect axonalsodium channels. J. Insect Physiol. 30, 341-349.
LEIBOWITZ, M. D., SCHWARZ, J. R., HOLAN, G. AND HILLE, B. (1987). Electrophysiological
comparison of insecticide and alkaloid agonists of Na channels. J. gen. Physiol. 90, 75-93.NAGY, K. (1987). Subconductance states of single sodium channels modified by chloramine-T
and sea anemone toxin in neuroblastoma cells. Eur. biophys. J. 15,129-132.NAGY, K., KISS, T. AND HOF, D. (1983). Single Na channels in mouse neuroblastoma cell
membrane. Indications for two open states. Pfliigers Arch. 399, 302-308.NARAHASHI, T. AND HAAS, H. G. (1968). Interaction of DDT with the components of lobster
nerve membrane conductance. J. gen. Physiol. 51, 177-198.O ' D O W D , D. K. AND ALDRICH, R. W. (1988). Voltage-clamp analysis of sodium channels in
wild-type and mutant Drosophila neurons. J. Neurosci. 8, 3633-3643.PICHON, C. AND PICHON, Y. (1989). A program for automatic analysis of action potentials in
squid axons. J. Physiol., Lond. 415, 4P.PICHON, Y. (1968). Nature des courants membranaires dans une fibre nerveuse d'insecte:
l'axone g6ant de Periplaneta americana. C.r. Seanc. Soc. Biol. 162, 2233-2240.PICHON, Y. (1974). Axonal conduction in insects. In Insect Neurobiology (ed. J. E. Treherne),
pp. 73—117. Amsterdam: North Holland.PICHON, Y. AND AMAR, M. (1991). 'Agonist' induced sodium channels in insect neurones: a key
to evolution of the sodium conductance? Biophys. J. (Abstr.) (in press).PICHON, Y. AND ASHCROFT, F. M. (1985). Nerve and muscle: electrical activity. In
Comprehensive Insect Physiology, Biochemistry and Pharmacology, vol. V (ed. G. A. Kerkutand L. I. Gilbert), pp. 85-113. Oxford: Pergamon Press.

84 M. AMAR, Y. PICHON AND I. INOUE
PICHON, Y. AND BOISTEL, J. (1967). Current-voltage relations in the isolated giant axons of thecockroach under voltage-clamp conditions. J. exp. Biol. 47, 343-356.
PICHON, Y. AND BOISTEL, J. (1969). Effets compares du DDT et de la veratrine sur laperm£abilite ionique de la membrane nerveuse d'insecte. J. Physiol., Paris 61, 373-374.
PICHON, Y., GUILLET, J. C.,HEILIG, U. AND PELHATE, M. (1985). Recent studies on the effects ofDDT and pyrethroid insecticides on nervous activity in cockroaches. Pesticide Sci. 16,627-640.
PICHON, Y. AND PELHATE, M. (1985). Electrical activity in insect axons: its modifications byDDT and pyrethroid insecticides. In Frontiers in Membrane Research in Agriculture: BeltsvilleSymposium 9 (ed. J. B. St John, E. Berlin and P. E. Jackson), pp. 421-438. Totowa: Rowmanand Allanheld.
PICHON, Y., PELHATE, M. AND HEILIG, U. (1986). Mode of action of pyrethroid insecticides onthe insect nerve membrane. Proceedings of the Sixth International Congress on PesticideChemistry, Ottawa 3A-22 (Abstract).
PICHON, Y., PELHATE, M. AND HEILIG, U. (1987). Sodium channels: primary targets forinsecticide action. In Sites of Action for Neurotoxic Pesticides (ed. R. M. Hollingworth andM. B. Green), ACS Symposium Series 356, pp. 212-225. Washington: ACS.
PITMAN, R. M. (1975). The ionic dependence of action potentials induced by colchicine in aninsect motorneurone cell body. J. Physiol., Lond. 247', 511-520.
QUANDT, F. N. AND NARAHASHI, T. (1982). Modification of single Na+ channels bybatrachotoxin. Proc. natn. Acad. Sci. U.S.A. 79, 6732-6736.
ROCHE, M., FRELIN, C , BRUNEAU, P. AND MEINARD, C. (1985). Interaction of tralomethrin,tralocythrin, and related pyrethroids in Na+ channels of insect and mammalian neuronal cells.Pest. Biochem. Physiol. 24, 306-316.
ROMEY, G., JACQUES, Y., SCHWEITZ, H., FOSSET, M. AND LAZDUNSKI, M. (1979). The sodiumchannel in non-impulsive cells. Interaction with specific neurotoxins. Biochim. biophys. Ada556, 344-353.
SACHS, F., NEIL, J. AND BARKAKATI, N. (1982). The automated analysis of data from single ionicchannels. Pflugers Arch. 395, 331-340.
SIGEL, E. (1987a). Properties of single sodium channels translated by Xenopus oocytes afterinjection with messenger ribonucleic acid. /. Physiol., Lond. 386, 73-90.
SIGEL, E. (19876). Effects of veratridine on single neuronal sodium channels expressed inXenopus oocytes. Pflugers Arch. 410, 112-120.
STUHMER, W., CONTI, F., SUZUKI, H., WANG, X., NODA, M., YAHAGI, N., KUBO, H. AND NUMA,S. (1989). Structural parts involved in activation and inactivation of the sodium channel.Nature 339, 597-603.
STUHMER, W., METHFESSEL, C , SAKMANN, B., NODA, B. AND NUMA, S. (1987). Patch-clampcharacterization of sodium channels expressed from brain cDNA. Eur. biophys. J. 14,131-138.
ULBRICHT, W. (1969). The effect of veratridine on excitable membranes of nerve and muscle.Ergeb. Physiol. 61, 18-71.
VUVERBERG, H. P. M., VAN DER ZALM, J. M. AND VAN DEN BERCKEN, J. (1972). Similar mode ofaction of pyrethroids and DDT on sodium channel gating in myelinated nerve. Nature 295,601-603.
Wu, C. F., SUZUKI, N. AND POO, M. M. (1983). Dissociated neurons from normal and mutantDrosophila larval central nervous system in cell culture. J. Neurosci. 3, 1888-1899.
YAMAMOTO, D., YEH, J. Z. AND NARAHASHI, T. (1984). Voltage-dependent calcium block ofnormal and tetramethrin-modified single sodium channels. Biophys. J. 45, 337-344.


















