Partial molar heat capacities and volumes of transfer of some amino acids and peptides from water to...
7
J. Phys. Chem. 1985,89, 1099-1 105 1099 is related to the emission energy with no ion pairing, E‘,,, and the ion-pairing energies in the excited (Utx) and ground states (AEx) by E,, = I ? ‘ , , + (AE’x - AEx). The internal energy changes associated with the emission energies are negative quantities, a fact which is incorporated into this expression. The ionic charge is the same in the ground and excited states leading, to a first approximation, to a cancellation of anion-cation Cou- lombic terms in AEx and AE’X. However, a significant anion- dipole term does exist for the excited state of the form, AEJx 0: -elpl/R2, where e is the unit electron charge, 1p1 the magnitude of the dipole moment, and R is the separation distance between the anion and the center of the dipole. There is no anion-dipole term in AEx because Ipl = 0 given the D3 symmetry of the ground state. The net effect is that the excited and ground states are expected to be stabilized about equally by Coulombic interactions with the surrounding anions but a further stabilization of the excited state occurs based on the anion-dipole interaction. The analysis based on the thermodynamic cycle suggests that E,, should vary as -l/(a + rx-)2 where R = a + r,, rx- is the radius of the ion, and a is determined by the properties of the excited-state dipole. There is insufficient information about the excited-state dipole and insufficient experimental data to test this prediction. However, it is evident from the data in Table I that E, does decrease as rx- decreases. In addition, although the sense of the changes in E,, with rx- are the same for Os(4,4’- Ph2phen)32+, they are much less profound, consistent with the larger size of the cation and the concomittant increase in R. The fact that correlations exist between the experimental emission energies and the anionic radii is significant in supporting an electrostatic origin for the variations in E,, and through E,,, k,. It is important to note that correlations do not exist with other properties. In particular, a significant contribution to stabilizing the excited state and decreasing the energy gap could have been the extent of mixing of the outer sphere charge transfer (OSCT) excited state, O~”(phen)~+, X, into the MLCT state. However, if such an effect exists it must be of significantly less importance than the simple electrostatic effect identified here, since there is no obvious correlation of E,, with ionization energies or redox potentials for the oxidation of the anions, X- - X + e-. Final Comments The results obtained here show that it is possible to account for the observation of specific anion effects based on the energy gap law and that the effects can be rationalized based on elec- trostatics. Even though the effects are small, the experimental observations provide a further experimental verification of the energy gap law and provide an understanding of the factors which determine excited-state lifetimes for MLCT excited states. With the acquisition of sufficient data it may be possible to turn the argument around and use excited-state lifetime measurements to obtain information concerning the surrounding medium. It is important to realize that the experiments described here were carried out in fluid solution where environmental redistri- bution is rapid on the lifetime of the excited state. In media where translational and orientational correlation times are restricted, as in polymeric films, glasses, or solids, counterion effects may lead to some distinctively different features since the environment is constrained to be that of the ground state. If, for example, there is an anionic asymmetry, it may contribute to the trapping of the excited electron predominantly onto a single ligand even in com- plexes like O ~ ( p h e n ) ~ ~ + which have D3 intramolecular symmetry. Acknowledgment. This work was supported by the Department of Energy under Grant No. DE-AS05-78ER06034. Registry No. [O~(phen),](PF,)~, 75441-76-8; [O~(phen),](ClO~)~, 14586-79-9; [O~(phen)~]Cl~, 73466-62-3;[Os(phen),]Br2, 82313-96-0; [O~(phen),l(SCN)~, 82313-97-1; [Os(phen),](BPh,),, 88545-38-4; [0~(4,4’-Ph~phen),](PF~)~, 94598-19-3; [0~(4,4/-Ph~phen)~](ClO~)~, 94598-20-6;[Os(4,4’-Ph2phen),]C12, 94598-21-7;[0~(4,4’-Ph~phen)~]- Br2, 94598-22-8; [0~(4,4’-Ph~phen),](sCN)~, 94598-23-9. Partial Molar Heat Capacities and Volumes of Transfer of Some Amino Acids and Peptides from Water to Aqueous Sodium Chloride Solutions at 298.15 K Rajiv Bhat and Jagdish C. Ahluwalia* Department of Chemistry, Indian Institute of Technology, Hauz Khas, New Delhi- 110016, India (Received: July I I, 1984; In Final Form: November 16, 1984) Partial molar heat capacities (Cpo) and volumes (P) of some amino acids and peptides have been determined in water and aqueous sodium chloride solutions at 298.15 K with a Picker flow microcalorimeter and a vibrating-tube digital density meter. These data have been utilized to calculate the respective heat capacities and volumes of transfer for the amino acids and peptides from water to various molalities of sodium chloride solutions. The results show that heat capacities and volumes of transfer for the amino acids, peptides, and the peptide group are positive and may be regarded to result from the dominant interactions of sodium chloride ions with the charged centers of the amino acids and peptides. The results have been rationalized by a cosphere overlap model. Introduction of has proved quite challenging and still remains a subject of extensive investigationS,1 o w i n g to the complexities arising from a direct thermodynamic study on proteins, investigations of the behavior of model compounds of proteins like amino acids and peptides in aqueous and mixed aqueous solvents make it easy to understand the factors governing the stability of the bi0polymers.2~ Salt solutions have large effects on the structure and properties of proteins including their solubility, denaturation, dissociation into subunits, and the activity of enzymes? In the literature there are some reports’ about the effect of various neutral salts on the (2) Rialdi, G.; Biltonen, R. In ‘International Review of Science”, Physical Chemistry”, Series 2; Skinner, H. A., Ed.; Butterworth: London, 1975; Vol. 10, pp 147-189. (3) Kauzmann, W. Adv. Prorein Chem. 1959, 14, 1. (4) (a) Von Hippel, P. H,; Schleich, T. In “Structure and Stability of Biological Macromolecules”; Vol. 2, Timasheff, S. N., Fasman, G. D., Eds.; Marcel Dekker: New York, 1969; p 417. (b) Jencks, W. P. In ‘Catalysis in Chemistry and Enzymology”; Mc-Graw Hill: New York, 1969; p 351. (c) Von Hippel, P. H.; Schleich, T. Acc. Chem. Res. 1969, 2, 257. The study of the thermodynamic stability of the native (1) Timasheff, S. N., Fasman, G. D., Eds. “Structure and Stability of Biological Macromolecules”, Marcel Dekker: New York, 1969; Vol. 2, (a) Chapter 2, p 65; (b) Chapter 3, p 213. 0022-3654/8S/2089-1099$01.~0/0 0 1985 American Chemical Society
Transcript of Partial molar heat capacities and volumes of transfer of some amino acids and peptides from water to...

J . Phys. Chem. 1985,89, 1099-1 105 1099
is related to the emission energy with no ion pairing, E‘,,, and the ion-pairing energies in the excited (Utx) and ground states (AEx) by E,, = I?‘,, + (AE’x - AEx). The internal energy changes associated with the emission energies are negative quantities, a fact which is incorporated into this expression. The ionic charge is the same in the ground and excited states leading, to a first approximation, to a cancellation of anion-cation Cou- lombic terms in AEx and AE’X. However, a significant anion- dipole term does exist for the excited state of the form, AEJx 0: -elpl/R2, where e is the unit electron charge, 1p1 the magnitude of the dipole moment, and R is the separation distance between the anion and the center of the dipole. There is no anion-dipole term in AEx because Ipl = 0 given the D3 symmetry of the ground state. The net effect is that the excited and ground states are expected to be stabilized about equally by Coulombic interactions with the surrounding anions but a further stabilization of the excited state occurs based on the anion-dipole interaction.
The analysis based on the thermodynamic cycle suggests that E,, should vary as - l / ( a + rx-)2 where R = a + r,, rx- is the radius of the ion, and a is determined by the properties of the excited-state dipole. There is insufficient information about the excited-state dipole and insufficient experimental data to test this prediction. However, it is evident from the data in Table I that E , does decrease as rx- decreases. In addition, although the sense of the changes in E,, with rx- are the same for Os(4,4’- Ph2phen)32+, they are much less profound, consistent with the larger size of the cation and the concomittant increase in R .
The fact that correlations exist between the experimental emission energies and the anionic radii is significant in supporting an electrostatic origin for the variations in E,, and through E,,, k,. It is important to note that correlations do not exist with other properties. In particular, a significant contribution to stabilizing the excited state and decreasing the energy gap could have been the extent of mixing of the outer sphere charge transfer (OSCT) excited state, O~”(phen)~+, X, into the MLCT state. However,
if such an effect exists it must be of significantly less importance than the simple electrostatic effect identified here, since there is no obvious correlation of E,, with ionization energies or redox potentials for the oxidation of the anions, X- - X + e-.
Final Comments
The results obtained here show that it is possible to account for the observation of specific anion effects based on the energy gap law and that the effects can be rationalized based on elec- trostatics. Even though the effects are small, the experimental observations provide a further experimental verification of the energy gap law and provide an understanding of the factors which determine excited-state lifetimes for MLCT excited states. With the acquisition of sufficient data it may be possible to turn the argument around and use excited-state lifetime measurements to obtain information concerning the surrounding medium.
It is important to realize that the experiments described here were carried out in fluid solution where environmental redistri- bution is rapid on the lifetime of the excited state. In media where translational and orientational correlation times are restricted, as in polymeric films, glasses, or solids, counterion effects may lead to some distinctively different features since the environment is constrained to be that of the ground state. If, for example, there is an anionic asymmetry, it may contribute to the trapping of the excited electron predominantly onto a single ligand even in com- plexes like O ~ ( p h e n ) ~ ~ + which have D3 intramolecular symmetry.
Acknowledgment. This work was supported by the Department of Energy under Grant No. DE-AS05-78ER06034.
Registry No. [O~(phen),](PF,)~, 75441-76-8; [O~(phen),](ClO~)~, 14586-79-9; [O~(phen)~]Cl~, 73466-62-3; [Os(phen),]Br2, 823 13-96-0; [O~(phen),l(SCN)~, 82313-97-1; [Os(phen),](BPh,),, 88545-38-4; [0~(4,4’-Ph~phen),](PF~)~, 94598-19-3; [0~(4,4/-Ph~phen)~](ClO~)~, 94598-20-6; [Os(4,4’-Ph2phen),]C12, 94598-21-7; [0~(4,4’-Ph~phen)~]- Br2, 94598-22-8; [0~(4,4’-Ph~phen),](sCN)~, 94598-23-9.
Partial Molar Heat Capacities and Volumes of Transfer of Some Amino Acids and Peptides from Water to Aqueous Sodium Chloride Solutions at 298.15 K
Rajiv Bhat and Jagdish C. Ahluwalia* Department of Chemistry, Indian Institute of Technology, Hauz Khas, New Delhi- 110016, India (Received: July I I , 1984; In Final Form: November 16, 1984)
Partial molar heat capacities (Cpo) and volumes (P) of some amino acids and peptides have been determined in water and aqueous sodium chloride solutions at 298.15 K with a Picker flow microcalorimeter and a vibrating-tube digital density meter. These data have been utilized to calculate the respective heat capacities and volumes of transfer for the amino acids and peptides from water to various molalities of sodium chloride solutions. The results show that heat capacities and volumes of transfer for the amino acids, peptides, and the peptide group are positive and may be regarded to result from the dominant interactions of sodium chloride ions with the charged centers of the amino acids and peptides. The results have been rationalized by a cosphere overlap model.
Introduction
of has proved quite challenging and still remains a subject of extensive investigationS,1 owing to the complexities arising from a direct thermodynamic study on proteins, investigations of the behavior of model compounds of proteins like amino acids and peptides in aqueous and mixed aqueous solvents make it easy to understand the factors governing the stability of the bi0polymers.2~
Salt solutions have large effects on the structure and properties of proteins including their solubility, denaturation, dissociation into subunits, and the activity of enzymes? In the literature there are some reports’ about the effect of various neutral salts on the
(2) Rialdi, G.; Biltonen, R. In ‘International Review of Science”, Physical Chemistry”, Series 2; Skinner, H. A., Ed.; Butterworth: London, 1975; Vol. 10, pp 147-189.
(3) Kauzmann, W. Adv. Prorein Chem. 1959, 14, 1. (4) (a) Von Hippel, P. H,; Schleich, T. In “Structure and Stability of
Biological Macromolecules”; Vol. 2, Timasheff, S. N., Fasman, G. D., Eds.; Marcel Dekker: New York, 1969; p 417. (b) Jencks, W. P. In ‘Catalysis in Chemistry and Enzymology”; Mc-Graw Hill: New York, 1969; p 351. (c) Von Hippel, P. H.; Schleich, T. Acc. Chem. Res. 1969, 2, 257.
The study of the thermodynamic stability of the native
(1) Timasheff, S. N., Fasman, G. D., Eds. “Structure and Stability of Biological Macromolecules”, Marcel Dekker: New York, 1969; Vol. 2, (a) Chapter 2, p 65; (b) Chapter 3, p 213.
0022-3654/8S/2089-1099$01.~0/0 0 1985 American Chemical Society

1100 The Journal of Physical Chemistry, Vol. 89, No. 7, 1985 Bhat and Ahluwalia
TABLE II: Limiting Partial Molar Heat Capacities (J K-' mol-') of Some Amino Acids and Peptides in Water and Aqueous Sodium Chloride Solutions alone with Their Correspondhe Transfer Parameters at 298.15 K'
compd c;
water 1 mNaCl 2mNaCl 3mNaCl 4mNaCl glycine diglycine triglycine DL-ahine DL-a-amino-n-butyric acid L-valine L-leucine
c;,, W + l m W + 2 m W - 3 m W - 4 m
NaCl NaCl NaCl NaCl 42.9 (1.1) 84.7 (1.0) 108.4 (1.9) 116.3 (0.9) 126.9 (2.0) 41.7 (1.5) 65.4 (2.2) 73.3 (1.4) 83.9 (2.3) 99.3 (1.0) 173.0 (2.1) 206.5 (1.6) 231.6 (1.3) 245.8 (1.5) 73.7 (2.3) 107.2 (1.9) 132.3 (1.6) 146.5 (1.8)
185.9b (0.9) 284.9 (3.7) 308.8 (3.1) 99.0 (3.8) 122.9 (3.2) 146.OC (5.0) 165.4 (2.0) 191.7 (1.6) 19.4 (5.4) 45.7 (5.2) 234.1d (1.0) 252.3 (3.4) 270.5 (3.8) 18.2 (3.5) 36.4 (3.9) 3O7Sd (1.0) 330.0 (2.8) 339.0 (6.9) 22.5 (3.0) 32.5 (6.9) 402.4d (3.0) 420.4 (2.6) 418.2 (5.2) 18.0 (4.0) 14.6 (6.0)
57.1 (6.6) 46.6 (4.7)
50.0 (6.5) 40.6 (4.7) 24.1 (3.5)
CReference 16. dReference 15.
glycyl-DL-alanine 219.7 (6.5) 276.8 (1.4) glycyl-DL-a-amino-n-butyric 306.2 (3.6) 352.8 (3.0)
glycyl-r-valine 352.6 (5.3) 402.6 (3.7) glycyl-L-leucine 473.3 (0.8) 513.9 (4.6) L-leucylgl ycine 486.2 (0.3) 510.3 (3.5)
acid
'Entries in parentheses are the standard deviations. bReference 31.
transition temperatures of proteins and enzymes. The effectiveness of various neutral salts toward the destabilizing tendency of proteins has been classified in the form of a series known as the Hofmeister ~ e r i e s . ~ The peptide group is strongly salted in or stabilized by NaI, NaC104, NaSCN, NaC1,CCOO; it is salted in less strongly by KF, LiC1, NaCl, KCl, CsCl, and NaBr; and (CH3)4NBr and Na2S04 have negligible salting in effects.
Harrington and Herskovid in a paper on the subunit structure and dissociation of Lumbricus terrestris hemoglobin by salts have pointed out that various neutral salts including sodium chloride tend to dissociate the duodecameric structure of hemoglobin into its subunits. Nagy and Jencks' have pointed out that electrolytes induce dissociation in the protein F-actin without causing any conformational change or denaturation. They have suggested that salts interact directly with the peptide groups of the protein and bring about its dissociation. Therefore, in order to understand the behavior of proteins in aqueous salt solutions, we have studied the thermodynamic parameters, partial molar heat capacities and volumes, for some amino acids and peptides in aqueous sodium chloride solutions.
Experimental Section The amino acids and peptides selected for the present study
were glycine, alanine, DL-a-amino-n-butyric acid, valine, leucine, diglycine, glycylalanine, glycyl-DL-a-amino-n-butyric acid, gly- cylvaline, glycylleucine, leucylglycine, and triglycine. All of them were procured from Sigma except for glycine (pro analysi grade) and valine, which were obtained from Merck, and diglycine, which was procured from BDH. Analytical reagent grade sodium chloride from Sarabhai Chemicals (India) was used. All the amino acids and peptides were used as such after drying them under vacuum over P205 in a vacuum desiccator. Distilled water was deionized by passing it through a Barnstead mixed-bed ion-ex- change resin column. The deionized water was further purified by distilling it from alkaline KMn04 and it was used for all the heat capacity and density measurements. All the solutions were made by weight.
Heat capacities of the solutions were measured with a Picker dynamic flow microcalorimeter coupled in series with an Anton Paar vibrating-tube digital density meter (Model DMA 60/602). The principle and operational procedure for the flow microcalo- rimeters and density meter9 has been described elsewhere. The precision of the microcalorimeter is 0.5% and the accuracy of the density meter is f 1.5 X 10" g cm-3. The temperature around the density meter cell was maintained by circulating water from
( 5 ) Hofmeister, F. Arch. Exp. Pathol. Pharmakol. 1888, 24, 247. (6) Harrington, J. P.; Herskovits, T. T. Biochemistry 1975, 14, 4972. (7) Nagy, B.; Jencks, W. P. J . Am. Chem. SOC. 1965.87, 2480. (8) Picker, P.; Leduc, P. A.; Philip, P. R.; Desnoyers, J. E. J . Chem.
(9) Picker, P.; Tremblay, E.; Jolicoeur, C. J. Solution Chem. 1974, 3, 377. Thermodyn. 1971, 3, 63 1.
a constant-temperature bath through the metal block of the density meter at a flow rate of ca. 3 dm3 m i d . The temperature of the bath was controlled by using a PTC-40 proportional temperature controller and an MK-70 ultracryostat. Thermal stability of the bath was better than f l X
The Picker flow microcalorimeter and the density meter were calibrated with sodium chloride solutions. A correction of 4% in power was required in the heat capacity results after comparing our data with those of Picker et ai.* Density results showed excellent agreement with the l i t e r a t ~ r e . ~ The density meter was calibrated with water and dry air everyday. The density of water a t 298.15 K was taken as 0.997 047 g cm-3 from Kell's data.1° Reproducibility of the density measurements was better than 3 PPm.
Results The apparent molar heat capacity (4,) and the apparent molar
volume (4,) were calculated from the specific heat capacity and the solution density results with the following equations.
(1)
(2)
where M is the molar mass of the solute and Cpo and Cp are the specific heat capacities of solvent and solution, respectively. In studies where aqueous sodium chloride was used as a solvent, Cpo is the specific heat capacity of the aqueous sodium chloride solution and C, is the specific heat capacity of the ternary system of amino acid (or peptide), sodium chloride, and water; m is the molality of the solute in water or aqueous sodium chloride solution. d and do are the densities of the solute in water (or aqueous sodium chloride solution) and the density of water (or aqueous sodium chloride solution), respectively. Densities and specific heat ca- pacities of aqueous sodium chloride solutions were measured separately for each set of experiments.
@, and 4, for the amino acids and peptides vary linearly with their concentration in sodium chloride solutions and are presented in Table I (Table I is available as supplementary material; see paragraph at the end of text regarding supplementary material). These results were fitted by the least-squares method to the equations:
K.
@, = MC, - 1000(Cpo - C p ) / m
@v = M / d - 1000(d - do) /mddo
@c = @cO + Scm
@v = @"O 4- Svm
(3)
(4)
where dCo and 9," are the infinite dilution apparent molar heat capacities and volumes, respectively, and S, and S, are the cor- responding experimental slopes of the @, and @" vs. m plots. In those cases where there was no dependence with m, @co and were calculated by taking an average of all the data points.
(10) Kell, G. S. J. Chem. Eng. Data 1967, 12, 66.
cpo and ro of Amino Acids and Peptides
7
Sodium chloride [mol k g - l )
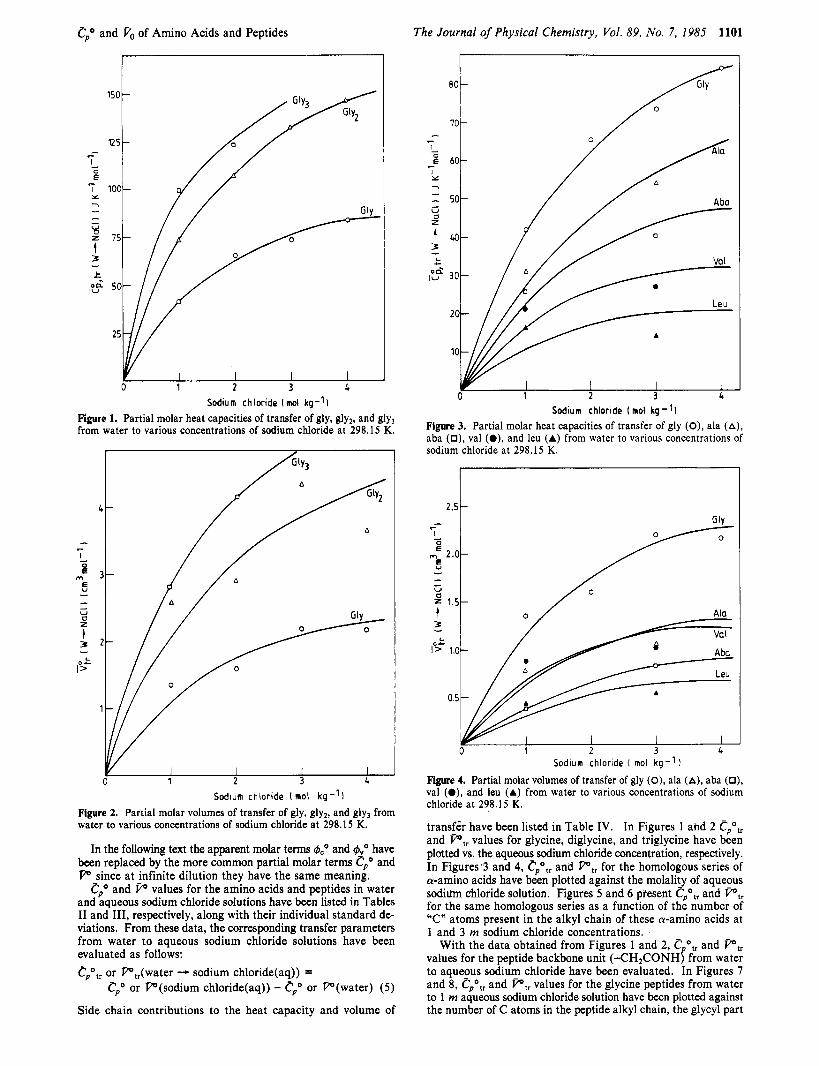
Figure 1. Partial molar heat capacities of transfer of gly, gly2, and gly3 from water to various concentrations of sodium chloride at 298.15 K.
/ / A
Sodium chloride [ mol kg - ' I Figure 2. Partial molar volumes of transfer of gly, gly,, and gly, from water to various concentrations of sodium chloride at 298.15 K.
In the following text the apparent molar terms &O and 4vo have been replaced by the more common partial molar terms Cpo and P since at infinite dilution they have the same meaning.
Cpo and i" values for the amino acids and peptides in water and aqueous sodium chloride solutions have been listed in Tables I1 and 111, respectively, along with their individual standard de- viations. From these data, the corresponding transfer parameters from water to aqueous sodium chloride solutions have been evaluated as follows: cpo,, or Ptr(water - sodium chloride(aq)) =
Cpo or P(sodium chloride(aq)) - Cpo or P(water ) ( 5 )
Side chain contributions to the heat capacity and volume of
The Journal of Physical Chemistry, Vol. 89, No. 7, 1985 1101
Sodium chloride [mol kg-1) Figure 3. Partial molar heat capacities of transfer of gly (0), ala (A), aba (0), Val (O), and leu (A) from water to various concentrations of sodium chloride at 298.15 K.
2.5 L c: - I
Sodium chloride [ mol kg-'
Figure 4. Partial molar volumes of transfer of gly (0), ala (A), aba (0), val (O), and leu (A) from water to various concentrations of sodium chloride at 298.15 K.
transfer have been listed in Table IV. In Figures 1 ahd 2 cpo,, and P,, values for glycine, diglycine, and triglycine have been plotted vs. the aqueous sodium chloride concentration, respectively. In Figures 3 and 4, cpo,, and i",, for the homologous series of a-amino acids have been plotted against the molality of aqueous sodium chloride solution. Figures 5 and 6 present Cpotr and PI, for the same homologous series as a function of the number of "C" atom$ present in the alkyl chain of these a-amino acids at 1 and 3 m sodium chloride concentrations.
With the data obtained from Figures 1 and 2, cpotr and P,, values for the peptide backbone unit (-CH,CONH) from water to aqueous sodium chloride have been evaluated. In Figures 7 and 8, cpo,, and P,, values for the glycine peptides from water to 1 m aqueous sodium chloride solution have been plotted against the number of C atoms in the peptide alkyl chain, the glycyl part
1102
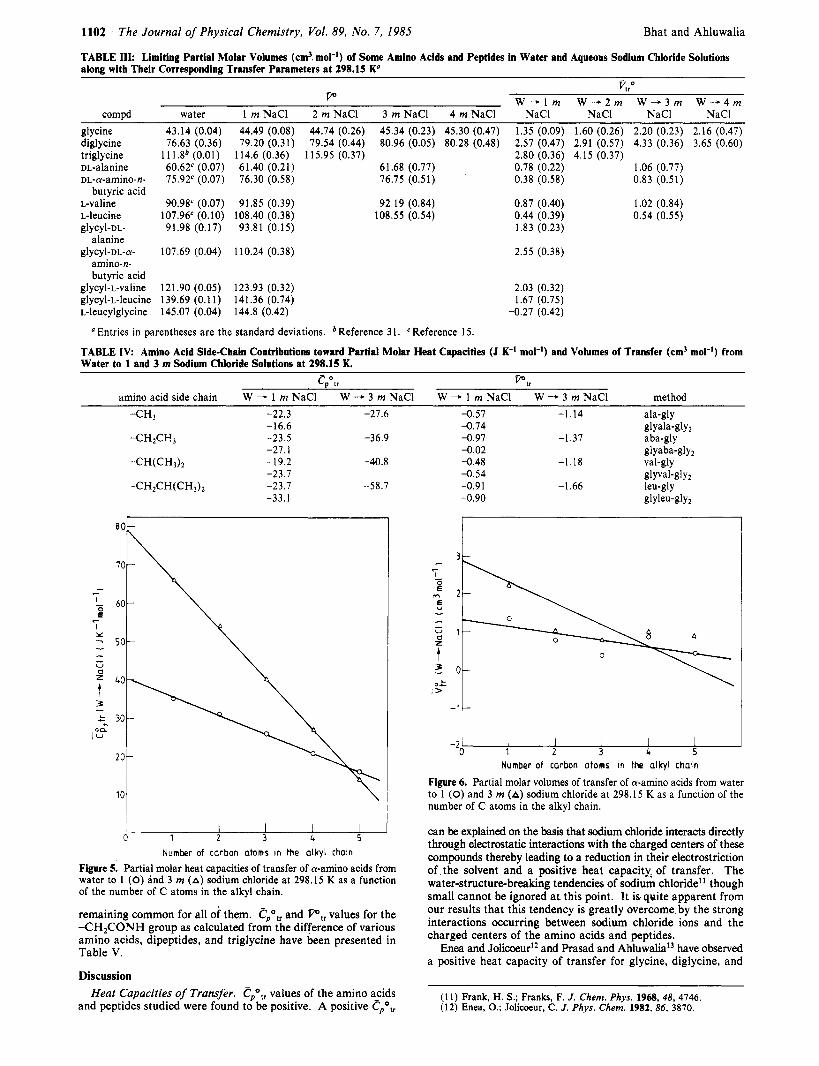
TABLE III: Limiting Partial Molar Volumes (cm3 mol-') of Some Amino Acids and Peptides in Water and Aqueous Sodium Chloride Solutions along with Their Corresponding Transfer Parameters at 298.15 K'
The Journal of Physical Chemistry, Vol. 89, No. 7, 1985 Bhat and Ahluwalia
V,," W - l m W - 2 m W - 3 m W - 4 m r"
compd water 1 mNaCl 2 m N a C l 3 m N a C l 4mNaCl NaCl NaCl NaCl NaCl g 1 y c i n e diglycine triglycine DL-alanine DL-a-amino-n-
butyric acid L-valine L-leucine
alanine
amino-n- butyric acid
glycyl-L-valine glycyl-L-leucine L-leucylglycine
glyCyl-DL-
glycyl-DL-a-
43.14 (0.04) 44.49 (0.08) 44.74 (0.26) 76.63 (0.36) 79.20 (0.31) 79.54 (0.44)
11 l.gb (0.01) 114.6 (0.36) 115.95 (0.37) 60.62c (0.07) 61.40 (0.21) 75.92c (0.07) 76.30 (0.58)
90.98' (0.07) 91.85 (0.39) 107.96c (0.10) 108.40 (0.38) 91.98 (0.17) 93.81 (0.15)
107.69 (0.04) 110.24 (0.38)
121.90 (0.05) 123.93 (0.32) 139.69 (0.11) 141.36 (0.74) 145.07 (0.04) 144.8 (0.42)
45.34 (0.23) 45.30 (0.47) 1.35 (0.09) 1.60 (0.26) 2.20 (0.23) 2.16 (0.47) 80.96 (0.05) 80.28 (0.48) 2.57 (0.47) 2.91 (0.57) 4.33 (0.36) 3.65 (0.60)
61.68 (0.77) 0.78 (0.22) 1.06 (0.77) 76.75 (0.51) 0.38 (0.58) 0.83 (0.51)
92.19 (0.84) 0.87 (0.40) 1.02 (0.84) 108.55 (0.54) 0.44 (0.39) 0.54 (0.55)
2.80 (0.36) 4.15 (0.37)
1.83 (0.23)
2.55 (0.38)
2.03 (0.32) 1.67 (0.75)
-0.27 (0.42)
' Entries in parentheses are the standard deviations. Reference 3 1. e Reference 15.
TABLE IV: Amino Acid Side-Chain Contributions toward Partial Molar Heat Capacities (J K-I mol-') and Volumes of Transfer (cm' mol-') from Water to 1 and 3 m sodium Chloride Solutions at 298.15 K.
c p o tr r" tr
amino acid side chain W - 1 m NaCl W - 3 m NaCl W - 1 m NaCl W - 3 m NaCl method -22.3 -27.6 -16.6 -23.5 -36.9 -27.1 -19.2 -23.7
-40.8
-CH,CH(CH3)2 -23.7 -58.7 -33.1
\
0 1 2 3 4 5 Number of carbon atoms in the alkyl chain
Figure 5. Partial molar heat capacities of transfer of a-amino acids from water to 1 (0) and 3 m (A) sodium chloride at 298.15 K as a function of the number of C atoms in the alkyl chain.
remaining common for all of them. cpo,, and P,, values for the -CH,CONH group as calculated from the difference of various amino acids, dipeptides, and triglycine have been presented in Table V.
Discussion Heat Capacities of Transfer. cpo,, values of the amino acids
and peptides studied were found to be positive. A positive cpot,
-0.57 -0.74 -0.97 -0.02 -0.48 -0.54 -0.91 -0.90
-1.14 ala-gly
-1.37 aba-gl y
-1.18 Val-gly
-1.66 leu-gly
glyala-gly 2
glYaba-glY2
glyvaWy2
glYleu-glY2
-1L
\
1 I I I I 1 2 3 4 5
Number of carbon atoms in the alkyl chain
Figure 6. Partial molar volumes of transfer of a-amino acids from water to 1 (0) and 3 m (A) sodium chloride at 298.15 K as a function of the number of C atoms in the alkyl chain.
can be explained on the basis that sodium chloride interacts directly through electrostatic interactions with the charged centers of these compounds thereby leading to a reduction in their electrostriction of the solvent and a positive heat capacity. of transfer. The water-structurebreakng tendencies of sodium chloride' though small cannot be ignored at this point. It i s quite apparent from our results that this tendency is greatly overcome by the strong interactions occurring between sodium chloride ions and the charged centers of the amino acids and peptides.
Enea and Jolicoeur12 and Prasad and Ahluwalia13 have observed a positive heat capacity of transfer for glycine, diglycine, and
(11) Frank, H. S.; Franks, F. J . Chem. Phys. 1968, 48, 4746. (12) Enea, 0.; Jolicoeur, C. J . Phys. Chem. 1982, 86, 3870.

cpo and Vo of Amino Acids and Peptides The Journal of Physical Chemistry, Vol. 89, No. 7, 1985 1103
TABLE V Peptide Group Contributions toward Partial Molar Heat Capacities (J K-' mol-') and Volumes of Transfer (cm' mol-') from Water to Various Molalities of Sodium Chloride at 298.15 K.
Cpo,(CH2CONH) ~ , , (CH,CONH) C,OdCONH) P,,(CONH)
W - l m W 4 2 m W - 3 m W - 4 m W - l m W - 2 m W - 3 m W - 4 m W - l m W - 3 m W - l m W - 3 m NaCl NaCI NaCl NaCl NaCl NaCl NaCl NaCl NaCl NaCl NaCl NaCl 32.0' 41.8" 59.0" 62.6' 1.22" 1.31" 2.13" 1.49" 54.38 86.68 1.799 3.278 25.3b 1 5.7b 0.23b 1 .24b 38.9h 1.45* 37.7c 1 .05c 24.1' 1.68'
27.S 1.16e 28.4d 2.17d 32.0' 1.59
22 .6 1.23f
gly2-gly. bgly,-gly2. Cglyala-ala. dglyaba-aba. e glyval-Val. fglyleu-leu. ggly,-ala. glyala-aba. 'glyaba-Val. jglyval-leu.
80 c
~ 1 2 3 4 5
Number o f carbon atoms in the alkyl chain OO
Figure 7. Partial molar heat capacities of transfer of glycine peptides from water to 1 m sodium chloride at 298.15 K as a function of the number of C atoms in their alkyl chain.
m 5 3 - -
L p.-'
m l
' 5 3 - -
L p.-'
't 0 1 2 3 4 5
Number o f carbon atoms in the alkyl chain
Figure 8. Partial molar volumes of transfer of glycine peptides from water to 1 m sodium chloride at 298.15 K as a function of the number of C atoms in their alkyl chain.
triglycine from water to aqueous urea solutions. They have ex- plained their results on the basis of interactions occurring between urea molecules and the peptide groups. In Figure 1 we observe that a t a particular molality of sodium chloride, triglycine has a higher cpo, value than diglycine which in turn has a higher cpo, value than glycine. This can be explained on the basis that the peptide group being hydrophilic interacts with sodium chloride
(13) Prasad, K. P.; Ahluwalia, J. C. Biopolymers 1980, 19, 273.
ions through the overlap of their hydration cospheres leading to a positive excess heat capacity of transfer in the order triglycine > diglycine > glycine.
From Figure 1 we have calculated the heat capacity of transfer of a peptide backbone unit (-CH,CONH) from water to 1, 2, 3, and 4 m aqueous sodium chloride solutions as follows:
cpot,(-CH2CONH) = cpo,,(triglycine) - Cpo,,(diglycine) (6)
or
cpo,,(-CH2CONH) = Cpo,,(diglycine) - CPo,,(glycine) (7)
The -CH2CONH group contributions to cpot, at various sodium chloride concentrations as evaluated from the difference betwekn the glycine peptides and the corresponding amino acids are positive (Table V).
With increasing concentration of sodium chloride (see Figure l ) , we get higher cpotr values showing that salt-amino acid (or peptide) interactions increase with increasing salt concentration. In the ternary system of amino acid (or peptide), sodium chloride, and water the following interactions may be occurring: (a) ion-ion interactions between Na+ ions and the COO- group and between C1- ions and the NH3+ group; (b) ion-peptide group interactions; (c) ion-nonpolar group interactions. If we follow the cosphere overlap modelI4 then ion-ion interactions and ion-peptide group interactions would lead to a positive Cpotr, but on the contrary ion-nonpolar group interactions will lead to a negative Cpotr. The overall cpou obtained experimentally can be viewed as comprising of these contributions. Since we observe positive cPo!, for all the amino acids and peptides, we conclude that ion-ion interactions and ion-hydrophilic group interactions are stronger than ion- nonpolar group interactions. It also explains the higher cpot, values observed at higher molalities of sodium chloride for these com- pounds.
In Figure 5 we have plotted cpo,, values for the homologous series of a-amino acids vs. the number of C atoms in their alkyl chains. data for DL-a-amino-n-butyric acid, valine, and leucine in water have been taken from Di Paola and Belleau.I5 The cpo value for alanine in water has been taken from Spink and Wadso.I6 Since these data were obtained from different authors, a linear regression analysis was carried out for these points as a function of the number of C atoms in the alkyl chain of a-amino acids. A linear regression analysis was also carried out for the Cpo values obtained by us in 1 and 3 m sodium chloride. cpo,, values for the amino acids were evaluated by taking the difference between individual points on the best-fitted lines obtained in each case.
Di Paola and BelleauI5 have measured cpou and P"tr for glycine from water to 1 m aqueous sodium chloride. Our values of cpotr (41.7 J K-' mol-') and P,, (1.35 cm3 mol-') for glycine agree very well with their values which are 46.7 J K-' mol-' and 1.85 cm3 mol-', respectively. In Figure 5 we observe that at 1 m sodium chloride concentration cpot, decreases by ca. 4 J K-I mol-' as the
(14) Friedman, H. L.; Krishnan, C. V. In "Water, A Comprehensive Treatise"; Franks, F., Ed. Plenum Press: New York, 1973; Vol. 3, Chapter 1.
(1 5 ) Di Paola, G.; Belleau, B. Can. J . Chem. 1978, 56, 1827. (16) Spink, C. H.; Wadso, I. J. Chem. Thermodn. 1975, 7, 561.

1104
number of CHI groups increases. This decrease can be attributed to enhanced ion-nonpolar group interactions as the CH2 content of these amino acids increases. At 3 m sodium chloride con- centration we again observe a decrease in cpotr_ But as the number of CH2 groups increases the difference in C,:,, values for the individual amino acids at 1 and 3 m sodium chlonde concentrations keeps on decreasing. For leucine we observe almost no difference in cpo, at 1 and 3 m concentrations of sodium chloride. In leucine because of the presence of five CH2 groups, ion-nonpolar group interactions reduce cpo,, to such an extent that it almost exactly compensates for the increase in Cpotr due to ion-ion interactions as a result of increasing the molality of sodium chloride. In Figure 7 we again find a decrease of ca. 7 J K-' mol-' for the transfer of a CH2 group from water to 1 m aqueous sodium chloride solution, showing that the scheme of obtaining CH2 group con- tribution to the cpo,, holds equally well in case of dipeptides as well. However, this decrease in NaCl solution is much less as compared to a decrease of ca. 32 J K-' mol-' for the transfer of a CH2 group from water to 6 M aqueous urea s o l ~ t i o n . ' ~
We have already found that the heat capacity of transfer for the peptide backbone unit is positive. Combining this with the negative contribution of the CHI group to cpotr, we conclude that heat capacity of transfer for a peptide group (-CONH) should be slightly more positive than a -CH,CONH group. Sodium chloride interacts with -CONH groups through the overlap of hydration cospheres and as a result of this overlap there is a reduction in the overall water-structure-breaking tendencies of sodium chloride. This leads to a positive heat capacity of transfer for the interaction of sodium chloride with the peptide group.
Lilley et al.17-22 have done a large number of studies on the interaction of various salts including sodium chloride with some amino acids and peptides and have evaluated the free energy and enthalpy of interaction parameters based on a McMillan-Mayer approach. They have concluded that interaction of the ions of sodium chloride with the a-amino acids is localized at the zwit- terionic head group of the amino acids and that attractive in- teractions occur between ions and the peptide group in peptides while repulsive interactions occur between the ions and the non- polar groups of amino acids. These conclusions do support our f .dings from heat capacities and volumes of transfer studies.
Volumes of Transfer. In Figure 2 we find that P,, values from water to aqueous sodium chloride increase with increasing con- centration of sodium chloride. A similar trend was observed in the heat capacity of transfer data too. i", for a peptide backbone unit is also positive like its cpot, value. The interaction of sodium chloride with the charged centers of these compounds reduces the effect of electrostriction of solvent thereby giving rise to a positive volume of transfer from water to aqueous sodium chloride.
Franks et al.23 have pointed out that the partial molar volume of a nonelectrolyte is a combination of two factors, viz., the intrinsic volume of the solute and the volume changes due to its interactions with the solvent. The intrinsic volume has been considered to be made up of two types of contribution^^^^^^
The Journal of Physical Chemistry, Vol. 89, No. 7, 1985 Bhat and Ahluwalia
(17) Lilley, T. H.; Tasker, I. R. J . Chem. Soc., Faraday trans. 1 1982, 78,
(18) Kelley, B. P.; Lilley, T. H. J. Chem. Thermodyn. 1979, 11, 513. (19) Lilley, T. H.; Moses, E.; Tasker, I. R. J . Chem. Soc., Faraday Trans.
1 1980, 76, 906. (20) Kelley, B. P.; Lilley, T. H. J. Chem. Thermodyn. 1978, 10, 703. (21) Kel! i , B. P.; Lilley, T. H. J . Chem. Soc., Faraday Trans. 1 1978,
(22) Kelley, B. P.; Lilley, T. H. J . Chem. Soc., Faraday Trans. 1 1978,
(23) Franks, F.; Quickenden, M. A,; Reid, D. S.; Watson, B. Trans.
(24) Shahidi, F.; Farrell, P. G . ; Edwards, J. T. J . Solution Chem. 1976,
(25) Terasawa, S.; Itsuki, H.; Arakawa, S. J . Phys. Chem. 1975,79,2345.
1 .
74, 2771.
74, 2779.
Faraday SOC. 1970 66, 582.
5, 807.
where Vvw is van der Waals volume26327 and Vvoid is the volume associated with the voids or empty spaces present thereinS2*
Shahidi et al.24 have modified the above equation in order to evaluate the contribution of a solute molecule toward its partial molar volume as
P = Vvw + Vvoid - nu,
where us is the shrinkage in volume produced by the interaction of hydrogen bonding groups present in the solute with water molecules and n is the potential number of hydrogen bonding sites in the molecule. Finally, t" of an amino acid can be evaluated as
= vvW + Vvoid - Vshrinkage
If we assume that Vvw and Vvold remain of the same magnitude in water and sodium chloride, the positive volumes of transfer for the amino acids can be explained to result from the decrease in volume of shrinkage in presence of sodium chloride molecules in aqueous solutions. Because of stronger interactions occurring between NH3+ and C1- and COO- and Na+ centers of glycine and other amino acids and peptides, it is obvious that the electro- striction of the neighboring water molecules due to these charged centers will be reduced in the presence of sodium chloride. This will lead to a reduction in the shrinkage volume. Furthermore, due to these interactions the structure-breaking tendency of sodium chloride will be reduced. This will also lead to a positive volume of transfer.29
The results shown in Figure 2 on the P,, of glycine, diglycine, and triglycine indicate that P,, for the peptide backbone unit CH2CONH is positive at all molalities of sodium chloride. P,, of a homologous series of compounds of amino acids and glycine peptides from water to sodium chloride solution which are plotted as a function of number of C atoms in their alkyl chain in Figures 6 and 8 show that the P, for CH, is negative. It may therefore be concluded that the positive contribution of P,, for CONH is more than the negative contribution of P,, of CH2. The positive P,, of CONH can be rationalized in terms of a reduction in its H-bonding interaction with water as a result of its interaction with Na+ and C1- ions in aqueous sodium chloride solution.
Another interesting observation concerns the difference in Cpotr and P,, values of glycylleucine and leucylglycine from water to 1 m sodium chloride. In water leucylglycine has a higher heat capacity and volume as compared to glycylleucine. This may be due to a greater decrease in electrostriction of NH3+ when it is near the alkyl group (as in leucylglycine) as compared to when COO- is near the alkyl group (as in g lycyl le~c ine) .~~ Upon transferring these compounds to 1 m sodium chloride, electro- striction of the COO- in glycylleucine is reduced further, giving rise to an increase in the heat capacities andavolume, whereas in leucylglycine electrostriction of the NH3+ group due to the alkyl group being near it is already reduced so much that the addition of sodium chloride does not reduce it further as is evident from no increase in P,, and a slight increase in Cpotr as we go from water to sodium chloride. This result further ascertains the validity of the cosphere overlap model in explaining the heat capacities and volumes of transfer in case of amino acids and peptides.
However, it has been pointed out by a referee that the im- portance of the electrostatic contribution to Cp3. or p, is perhaps underestimated in this work and the suggestion has been made for a study of derivatives of amino acids to elucidate the relative importance of the cosphere overlap effect and purely electrostatic effects. Nevertheless, it should be worthwhile to have such data for amino acids and their derivatives in aqueous solutions of various
(26) Bondi, A. J . Phys. Chem. 1964, 68, 441. (27) Bondi, A. In "Physical Properties of Molecular Crystals, Liquids, and
(28) Bondi, A. J . Phys. Chem. 1959, 58, 929. (29) Fortier, J. L.; Philip, P. R.; Desnoyers, J . E. J . Solution Chem. 1974,
(30) Zana, R. J . Phys. Chem. 1977, 81, 1817. (31) Jolicoeur, C.; Boileau, J. Can. J . Chem. 1978, 56, 2707.
Glasses"; New York, 1968; Chapter 14.
3. 523.

J . Phys. Chem. 1985,89, 1105-1 107 1105
salts to understand better the role of various effects that govern the conformational stability of a protein in aqueous neutral salt solutions.
Registry No. NaCI, 7647-14-5; glycine, 56-40-6; diglycine, 142-73-4; triglycine, 139-1 3-9; DL-alanine, 302-72-7; DL-a-aminobutyric acid, 2835-81-6; L-valine, 72-1 8-4; L-leucine, 61-90-5; glycyl-DL-alanine, 926- 77-2; glycyl-DL-a-aminobutyric acid, 7369-76-8; glycyl-L-valine, 1963-
21-9; glycyl-L-leucine, 869- 19-2; L-leucylglycine, 686-50-0.
Supplementary Material Available: Data on apparent molar volumes (4") and apparent molar heat capacities (&) for some amino acids and peptides a t 298.15 K in aqueous solutions of Sodium chloride as a function of concentration of sodium chloride given in Table I (7 pages). Ordering information is available on any current masthead page.
A Reexamination of the CH,' -t HCN Association Reaction Including the CH,+/CD,+ Isotope Effect
Paul R. Kemper, Lewis M. Bass,+ and Michael T. Bowers* Department of Chemistry, University of California, Santa Barbara, California 931 06 (Received: August 1, 1984; In Final Form: October 22, 1984)
The association reaction CH3+ + HCN - [CH3-HCN]+ has been reexamined. Previous work suggested this reaction occurred with a bimolecular rate constant of - 1 X lo-'' cm3/s due to radiative association at temperatures near 300 K. New work reported here suggests an upper limit of 5 X lo-'' cm3/s for radiative association at 300 K. Third-order association rate constants are reported for He, Ne, Ar, and HCN third bodies. Relative stabilization efficiencies of 0.11, 0.23, 0.25, and 1 .OO, respectively, are obtained. The effect of substituting CD3+ for CH3+ was also studied. The third-order rate constants increased by approximately a factor of 2, in excellent agreement with phase space theory calculations. The reaction CD3+ + HCN - CD2H+ + DCN was observed to occur at approximately 15% of the collision rate, indicating a barrier to isomerization exists with a height comparable to the [CD3-HCN]+ dissociation energy.
Introduction Several years ago we reported' an investigation, both experi-
mental and theoretical, of the CH3+ + H C N association reaction
The experiments were done using a drift ion cyclotron resonance spectrometer (drift ICR). Experimental evidence indicated that the stable product was produced both by radiative and collisional stabilization of an excited association complex. Two experimental findings indicated that radiative stabilization occurred: first, the observed second-order reaction rate coefficient did not extrapolate to zero at zero pressure. This indicated that stable product was formed in the absence of collisions. Second, ion cyclotron dou- ble-resonance experiments indicated that the CH3+ reactant and the [CH3-HCN]+ product were chemically coupled at low pressure.
In this paper we present new measurements of both the CH3+ + H C N and the CD3+ + H C N systems. The experiments were done using both drift and tandem ICR spectrometers. We now conclude that radiative stabilization does not occur to a significant extent in these systems. We do, however, find very significant differences between the third-order association rates in the CH3+ and CD3+ systems. These differences are well reproduced in theoretical calculations using statistical theory.
Experimental Section Both drift ICR and tandem ICR (TICR) spectrometers were
used in the experiments reported here. Both spectrometers have been described previou~ly.~-~ The drift ICR experiments can be briefly described as follows (see also ref 1). The CH3+ reactant was formed by low-energy electron impact on CHJ. The resulting CH31+ (>90% of the ionization) was ejected throughout the cell by using ICR double resonance. This was necessary since the CH,I+ ion also reacted with H C N to give a mass 42+ product (the product of interest)
Present address: Division of Natural Sciences, University of South Carolina at Aiken, Aiken, SC 29801.
0022-365418512089-1105$01 SO10
CH31+ + H C N - [CH3-HCN]+ + I (2) In the previous study, we obtained the observed second-order rate from the relative CH3+ and M42+ intensities knowing the reaction time, H C N pressure, and bath gas pressure. The same experi- mental conditions were used in the present experiments; however, the drift ICR was used only for low-pressure, thermal energy ICR double-resonance experiments and no rate constants were mea- sured. In these doubleresonance experiments, the M42' product ion was monitored in the second region of the cell while the reactant ion of interest was irradiated at its resonant cyclotron frequency in the first region of the cell. The resulting increase in reaction kinetic energy allowed determination of the effect of kinetic energy in the reaction, as well as the different reaction pathways which lead to the M42+ product.
The buik of the experiments presented here were done using the TICR spectrometer. In the TICR the ion formation and reaction regions are separated and interfering reactions (such as the CH31+ + H C N reaction) do not occur. Ions are formed in an ion source by electron impact. Both CH4 and CH31 were used as sources of CH3+ ions; no difference in the results was found. After formation the ions were drifted out of the source, accelerated to -3 KeV, mass selected in a 180' Dempster mass filter, de- celerated to near zero kinetic energy, and injected into the ICR reaction cell where the reaction with HCN occurred. The HCN used was purified by freeze-pump-thaw cycles. The source and reaction cell pressures were measured directly with a capacitance manometer. Reaction drift times were measured by using a trapping pulse method.3 The CH3+ kinetic energy upon injection into the reaction cell is about 0.5 eV (lab).3 This excess energy could have a significant effect on the association reaction. The effect-of-this injection energy was investigated by adding non- reactive collisional buffer gases to the reaction cell. This point
(1) L. M. Bass, P. R. Kemper, V. G . Anicich, and M. T. Bowers, J . Am.
(2) A. G . Wren, P. Gilbert, and M. T. Bowers, Rea Sci. Inrtrum., 49, 531 Chem. Soc., 103, 5283 (1981).
(1978). '
52, l(1983). (3jP. R. Kemper and M. T. Bowers, Int . J . Mass. Spectrom. Ion Phys.,
0 1985 American Chemical Society