
Part I Channel structure and gating 1912 Julius Bernstein ... Membrane bilayers – act as...
72
Ion channels 7, 10, 12, & 14 Feb 14 Dan Minor, Ph.D. Cardiovascular Research Institute University of California, San Francisco [email protected] Part I Channel structure and gating Thursday, 6 February 14
Transcript of Part I Channel structure and gating 1912 Julius Bernstein ... Membrane bilayers – act as...

Ion channels 7, 10, 12, & 14 Feb 14
Dan Minor, Ph.D.Cardiovascular Research InstituteUniversity of California, San [email protected]
Part I Channel structure and gating
Thursday, 6 February 14

Ion Channels - Structures and gating mechanisms
- Voltage-gated channels - Idea of a gate - potassium channel example- Voltage-sensors : how to open the gate- Voltage-gated channels -inactivation- Ligand-gated ion channels : a different sort of gate and gating
mechanism
- Permeation - Chemical recognition at high speed- Toxins and channels- Engineering light driven channels
- Mechanosensation- MALS
Thursday, 6 February 14

Gating – channel’s response to a stimulus, opening or closing –100 µs
Selective permeability – restricted passage of ion type – 25 ns ion-1
Fastest enzyme (Carbonic anhydrase) ~1 µs Helix-coil transitions 10 nsBackbone motions 1 ns
Ion channels are FAST!Ion - ‘Greek for that which goes’ (Faraday 1834)
Thursday, 6 February 14

Brief history of electrical signals in cellular systems
1881 – 1887 Sidney Ringer – perfused frog hearts require salts of sodium, potassium, and calcium in definite proportion for the heart to beat
1888 Walther Nernst – electrolyte diffusion theory inspires ideas about bioelectric potentials1902, 1912 Julius Bernstein – Proposes correctly that excitable cells are surrounded by a membrane with selective potassium permeability and that during excitation this permeability changes for other ions
1930s-1950s Cole, Curtis, Hodgkin, Huxley – Squid giant axon, action potential theory1960s-1970s – Pores or carriers?
1980s – First channels cloned1998 – First high resolution crystal structure of a channel protein
Thursday, 6 February 14
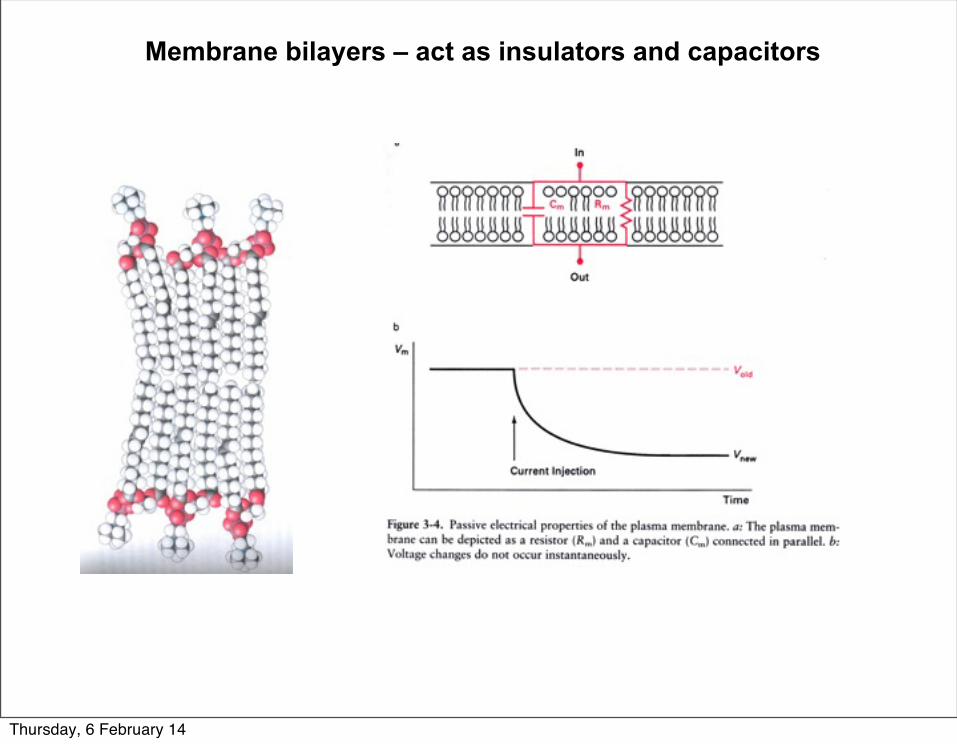
Membrane bilayers – act as insulators and capacitors
Thursday, 6 February 14
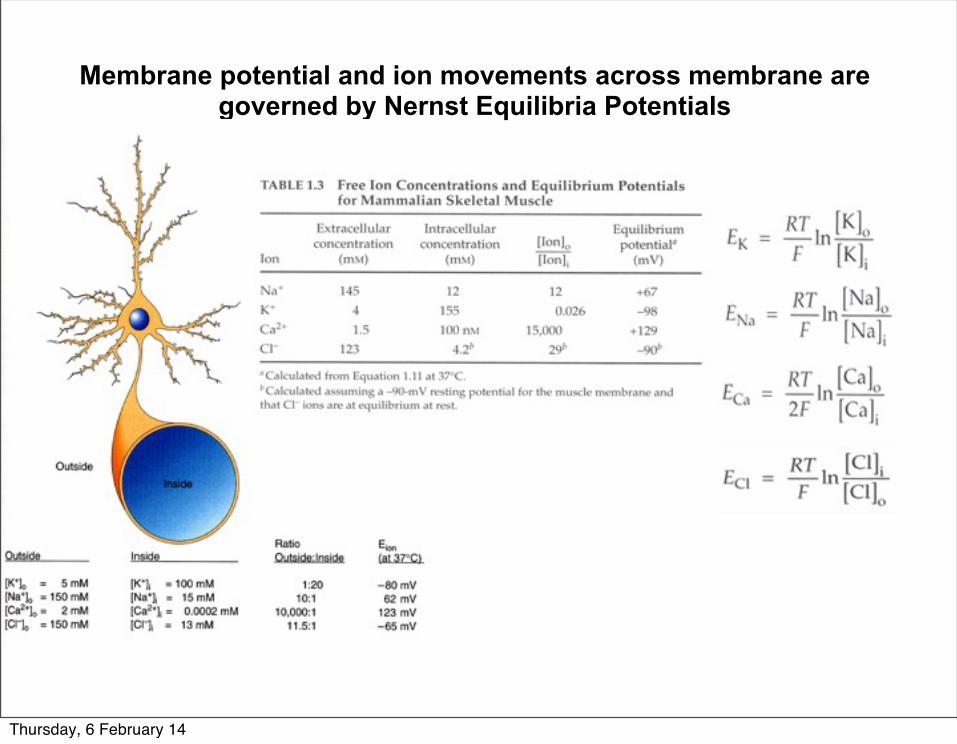
Membrane potential and ion movements across membrane are governed by Nernst Equilibria Potentials
Thursday, 6 February 14
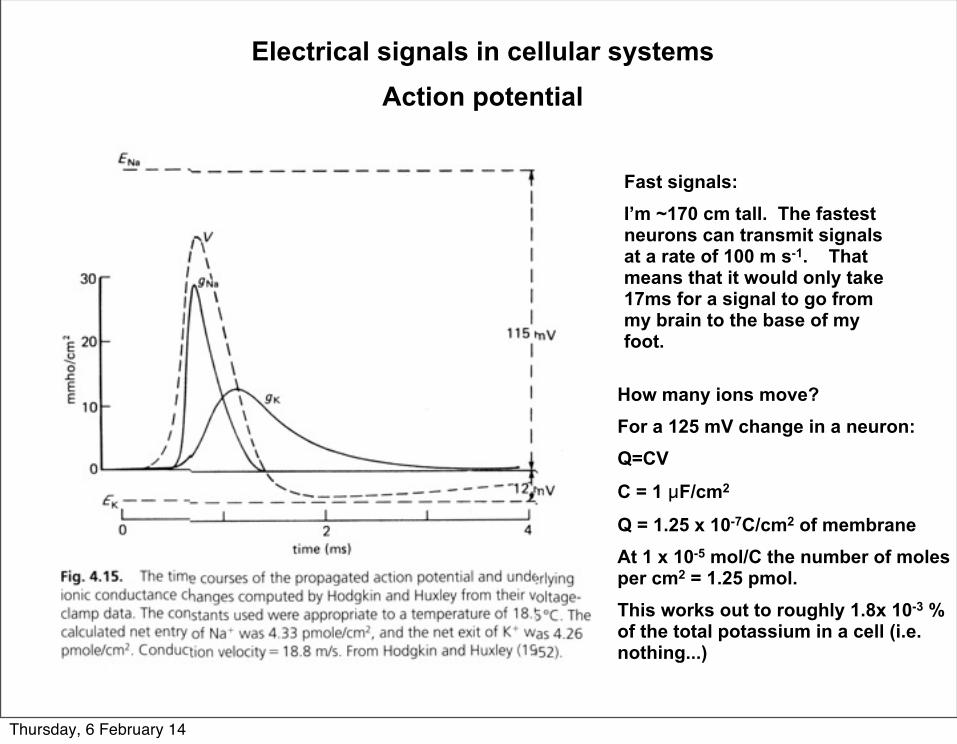
Electrical signals in cellular systemsAction potential
Fast signals:I’m ~170 cm tall. The fastest neurons can transmit signals at a rate of 100 m s-1. That means that it would only take 17ms for a signal to go from my brain to the base of my foot.
How many ions move?For a 125 mV change in a neuron:Q=CV
C = 1 µF/cm2
Q = 1.25 x 10-7C/cm2 of membrane At 1 x 10-5 mol/C the number of moles per cm2 = 1.25 pmol.This works out to roughly 1.8x 10-3 % of the total potassium in a cell (i.e. nothing...)
Thursday, 6 February 14
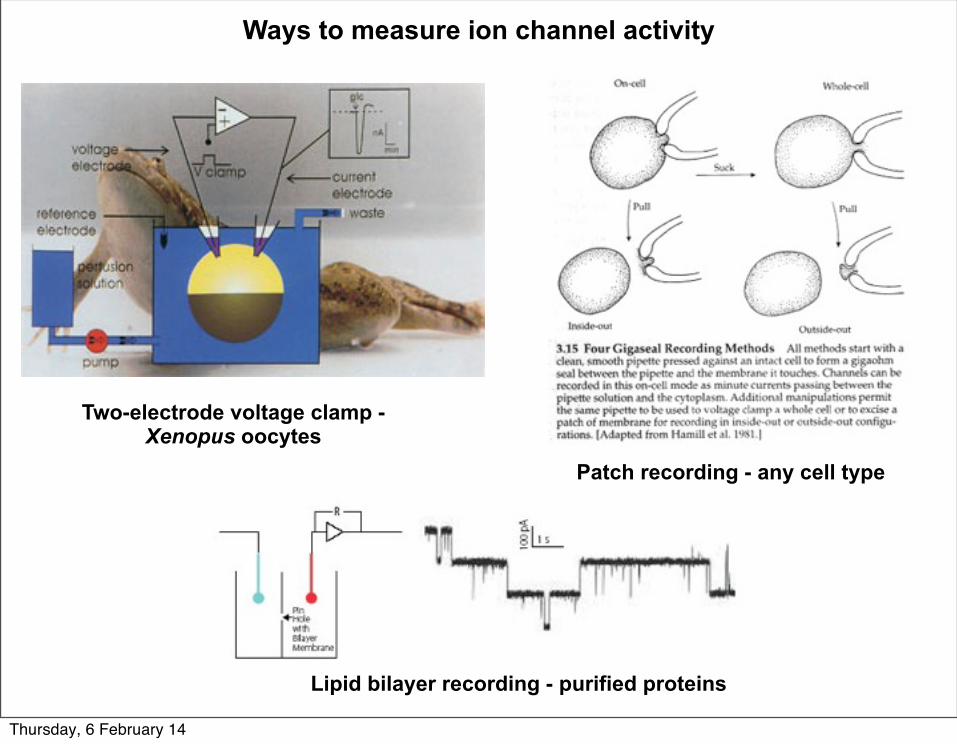
Ways to measure ion channel activity
Two-electrode voltage clamp - Xenopus oocytes
Patch recording - any cell type
Lipid bilayer recording - purified proteins
Thursday, 6 February 14
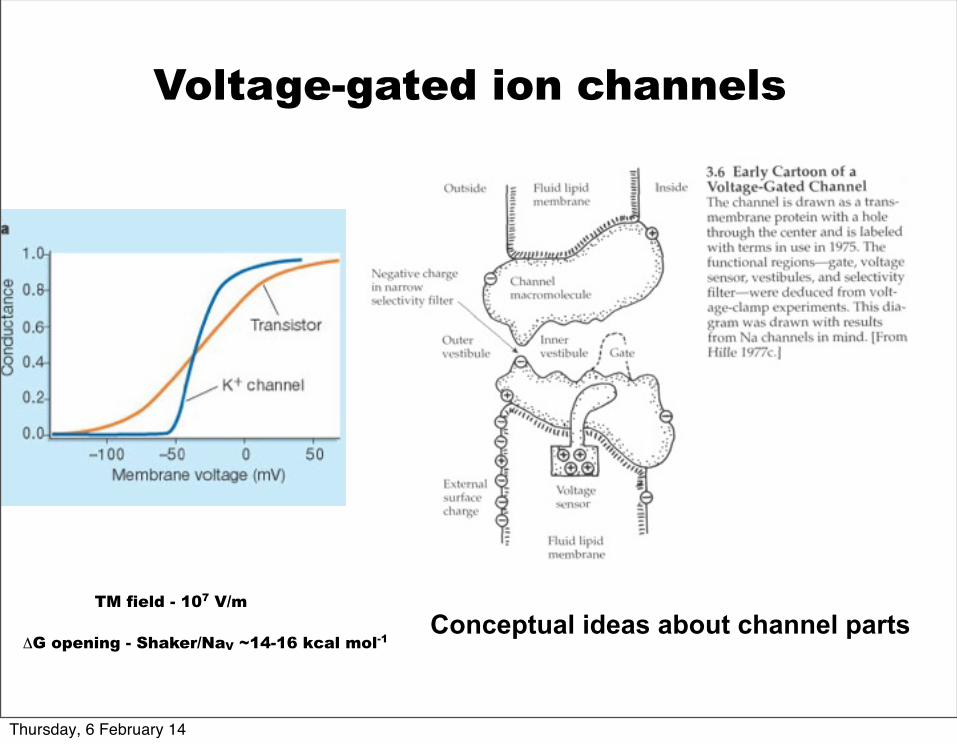
Conceptual ideas about channel parts
Voltage-gated ion channels
TM field - 107 V/m
ΔG opening - Shaker/NaV ~14-16 kcal mol-1
Thursday, 6 February 14
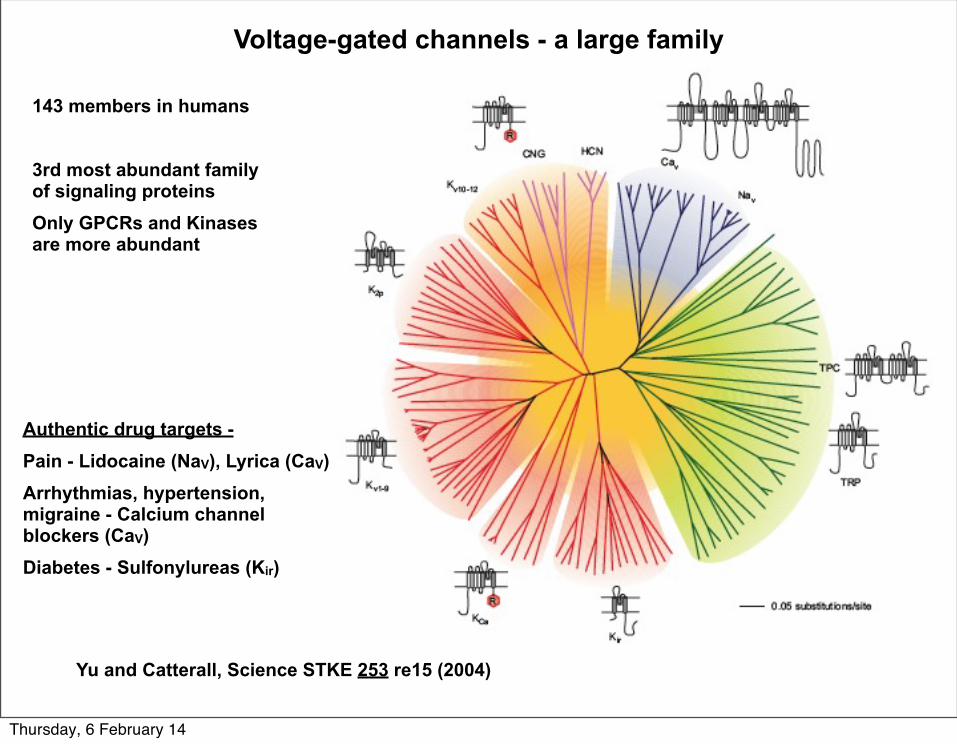
Voltage-gated channels - a large family
Yu and Catterall, Science STKE 253 re15 (2004)
143 members in humans
3rd most abundant family of signaling proteins Only GPCRs and Kinases are more abundant
Authentic drug targets -Pain - Lidocaine (NaV), Lyrica (CaV)Arrhythmias, hypertension, migraine - Calcium channel blockers (CaV)Diabetes - Sulfonylureas (Kir)
Thursday, 6 February 14
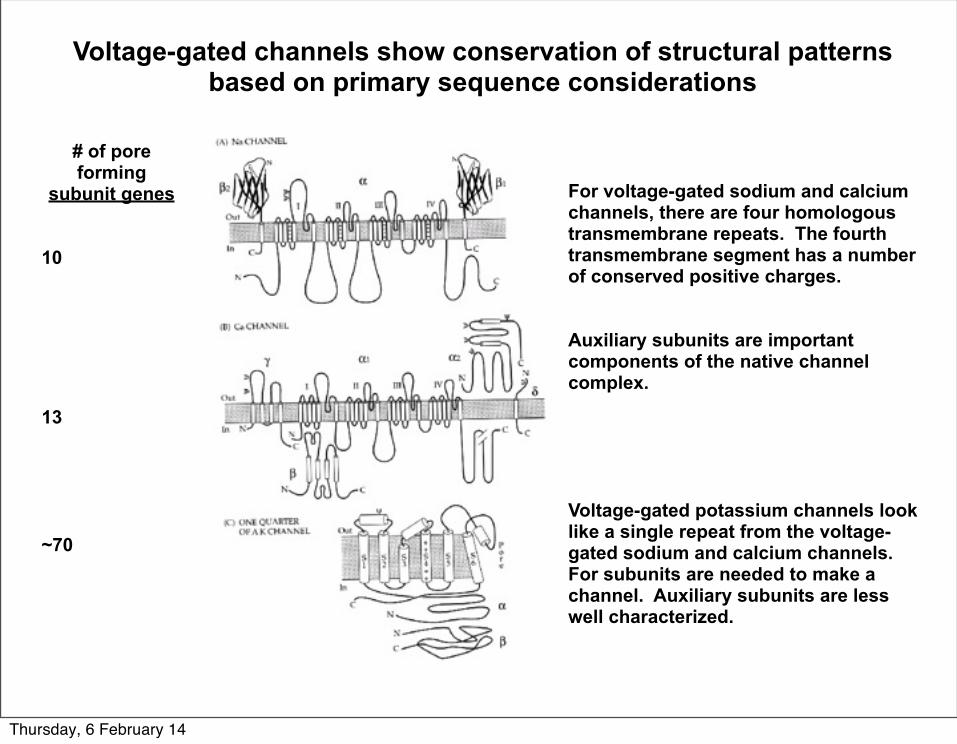
Voltage-gated channels show conservation of structural patterns based on primary sequence considerations
For voltage-gated sodium and calcium channels, there are four homologous transmembrane repeats. The fourth transmembrane segment has a number of conserved positive charges.
Auxiliary subunits are important components of the native channel complex.
Voltage-gated potassium channels look like a single repeat from the voltage-gated sodium and calcium channels. For subunits are needed to make a channel. Auxiliary subunits are less well characterized.
# of pore forming
subunit genes
10
13
~70
Thursday, 6 February 14
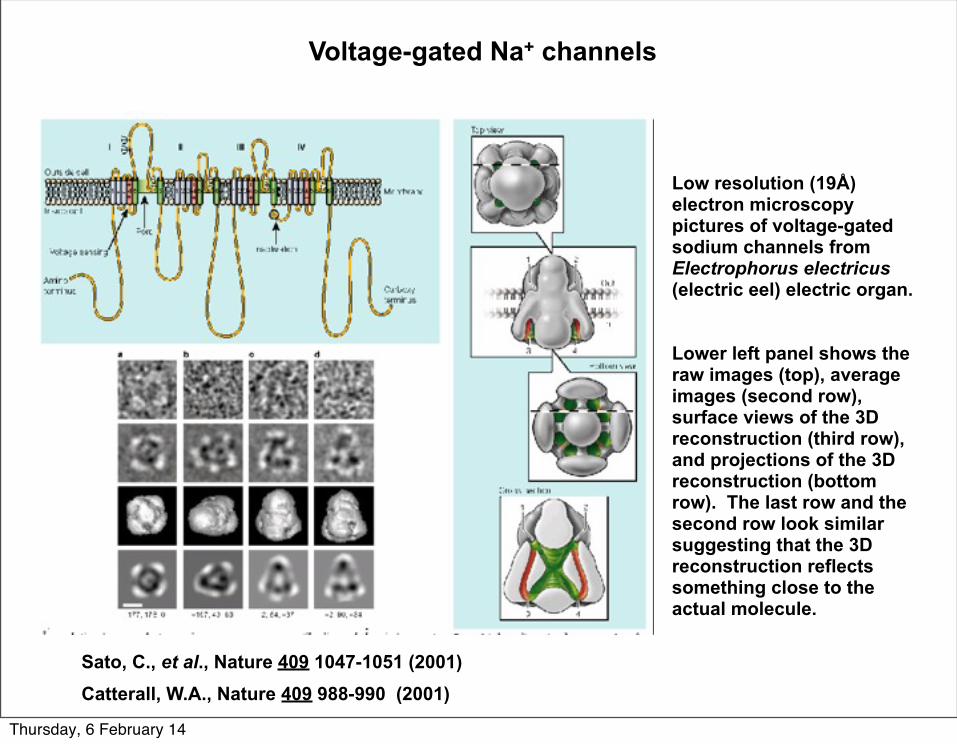
Sato, C., et al., Nature 409 1047-1051 (2001)Catterall, W.A., Nature 409 988-990 (2001)
Low resolution (19Å) electron microscopy pictures of voltage-gated sodium channels from Electrophorus electricus (electric eel) electric organ.
Lower left panel shows the raw images (top), average images (second row), surface views of the 3D reconstruction (third row), and projections of the 3D reconstruction (bottom row). The last row and the second row look similar suggesting that the 3D reconstruction reflects something close to the actual molecule.
Voltage-gated Na+ channels
Thursday, 6 February 14

Voltage drop across the membrane--most of the drop happens in the hydrophobic layer
Voltage-gated channels have intrinsic voltage sensors (charges/dipoles) in the membrane
The S4 segments with multiple basic residues correspond to the intrinsic voltage sensor
Voltage-gating - how do charge movements in the channel open
the pore?
Thursday, 6 February 14

Voltage sensor positive charges are conserved across a wide range of voltage-sensitive channels
Early models of voltage sensor motions postulated a helical-screw motion where the voltage-sensor moves towards the extracellular space upon depolarization.
Hille, B., Ion Channels of Excitable Membranes, 3rd edition, Sinauer (Sunderland, MA) 2001Thursday, 6 February 14

Hille, B., Ion Channels of Excitable Membranes, 3rd edition, Sinauer (Sunderland, MA) 2001
Cysteine has a unique reactivity among the twenty natural amino acids, a thiol (-SH) group.
Experiments that measure changes in cysteine reactivity can reveal conformational changes, There are many caveats…..
Thursday, 6 February 14
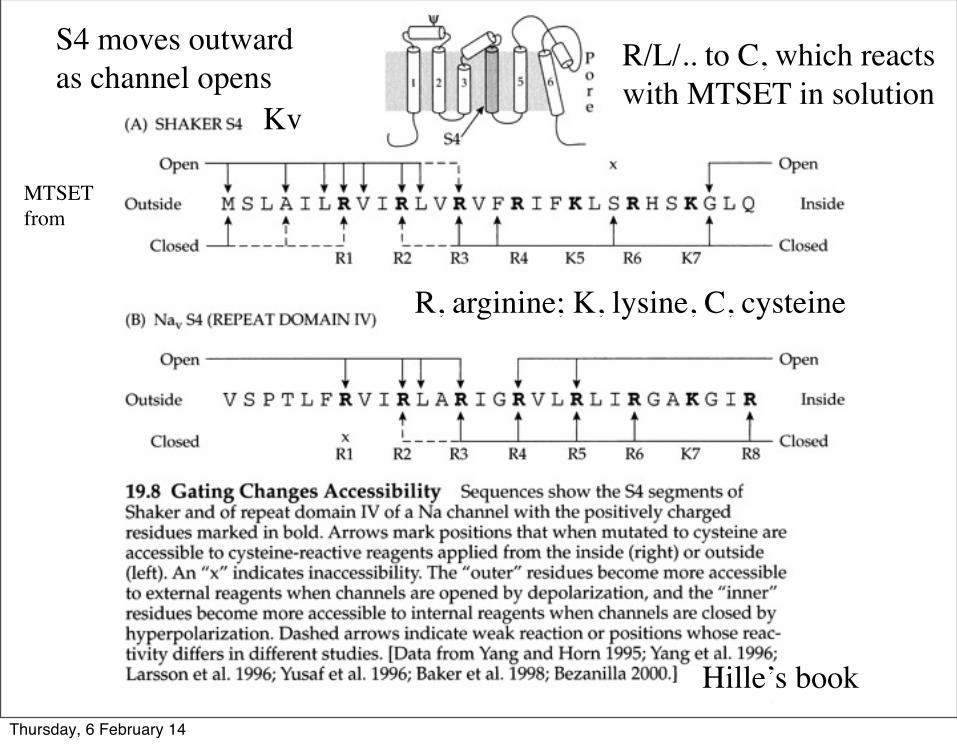
R/L/.. to C, which reacts with MTSET in solution
R, arginine; K, lysine, C, cysteine
Kv
Hille’s book
MTSETfrom
S4 moves outwardas channel opens
Thursday, 6 February 14
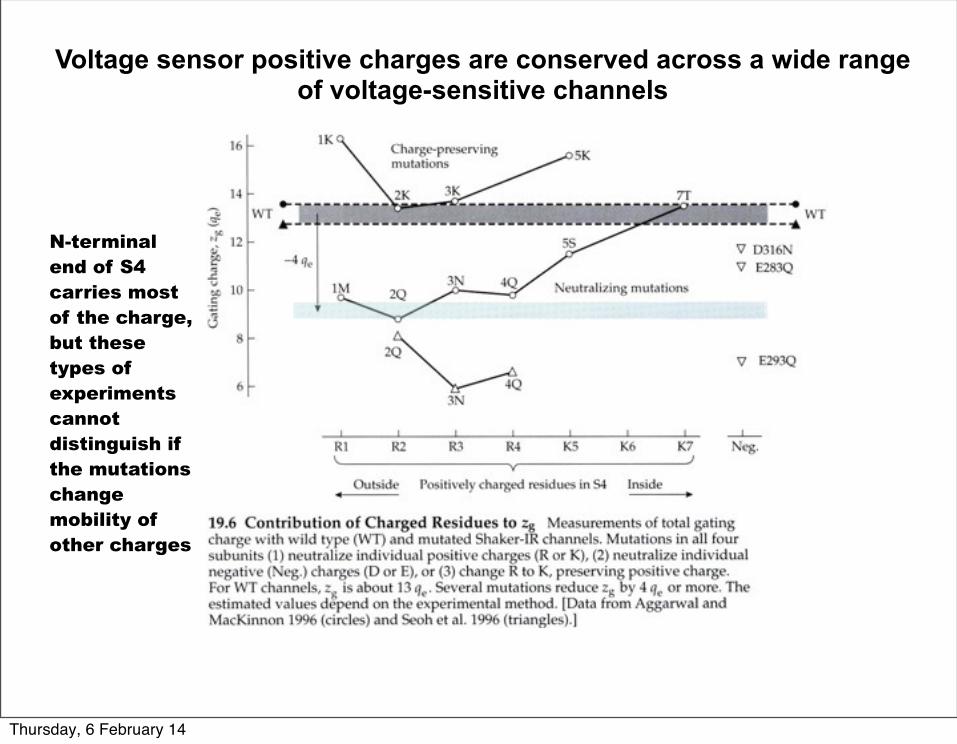
Voltage sensor positive charges are conserved across a wide range of voltage-sensitive channels
N-terminal end of S4 carries most of the charge,but these types of experiments cannot distinguish if the mutations change mobility of other charges
Thursday, 6 February 14
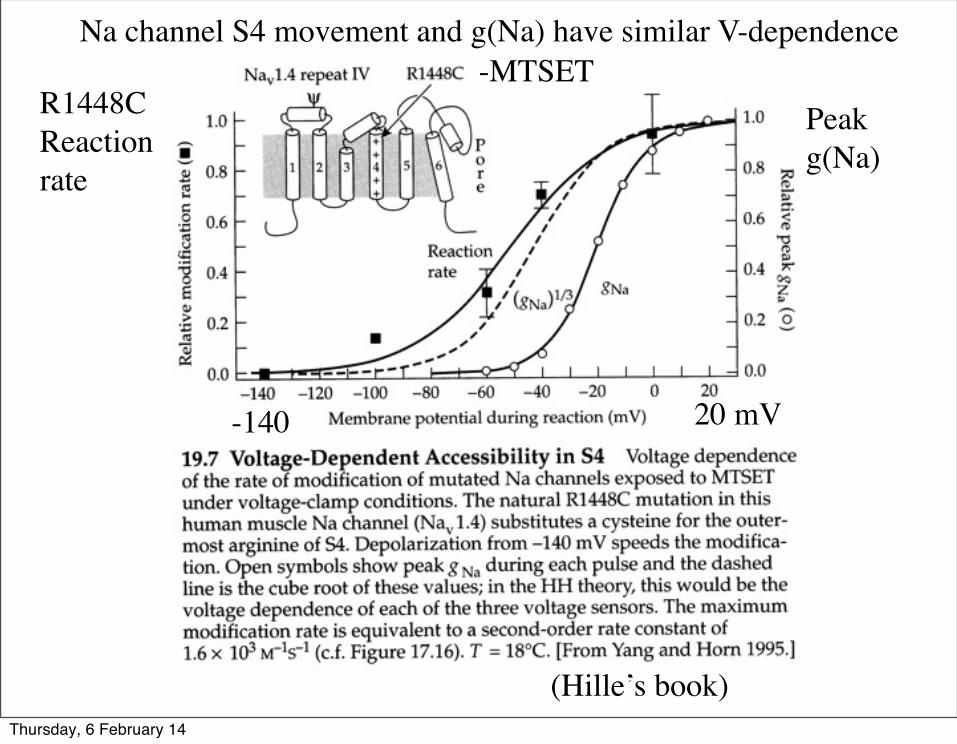
-MTSET
(Hille’s book)
Na channel S4 movement and g(Na) have similar V-dependence
Peak g(Na)
R1448C Reactionrate
-140 20 mV
Thursday, 6 February 14
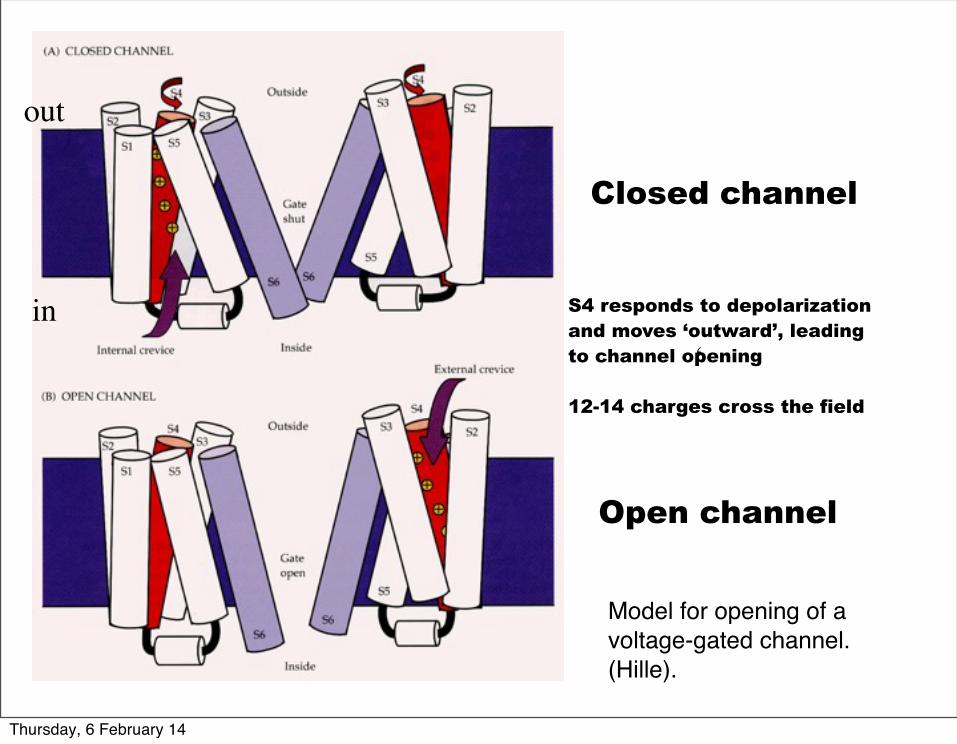
Model for opening of a voltage-gated channel. (Hille).
Closed channel
Open channel
S4 responds to depolarizationand moves ‘outward’, leading to channel opening
12-14 charges cross the field
‘
out
in
Thursday, 6 February 14
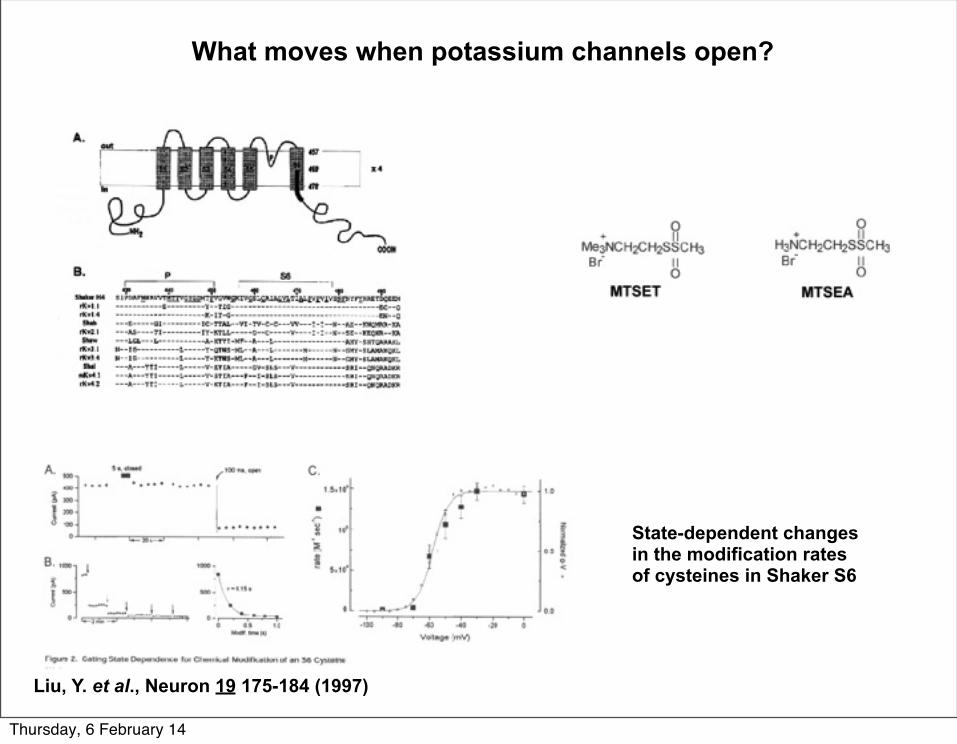
What moves when potassium channels open?
State-dependent changes in the modification rates of cysteines in Shaker S6
Liu, Y. et al., Neuron 19 175-184 (1997)
Thursday, 6 February 14
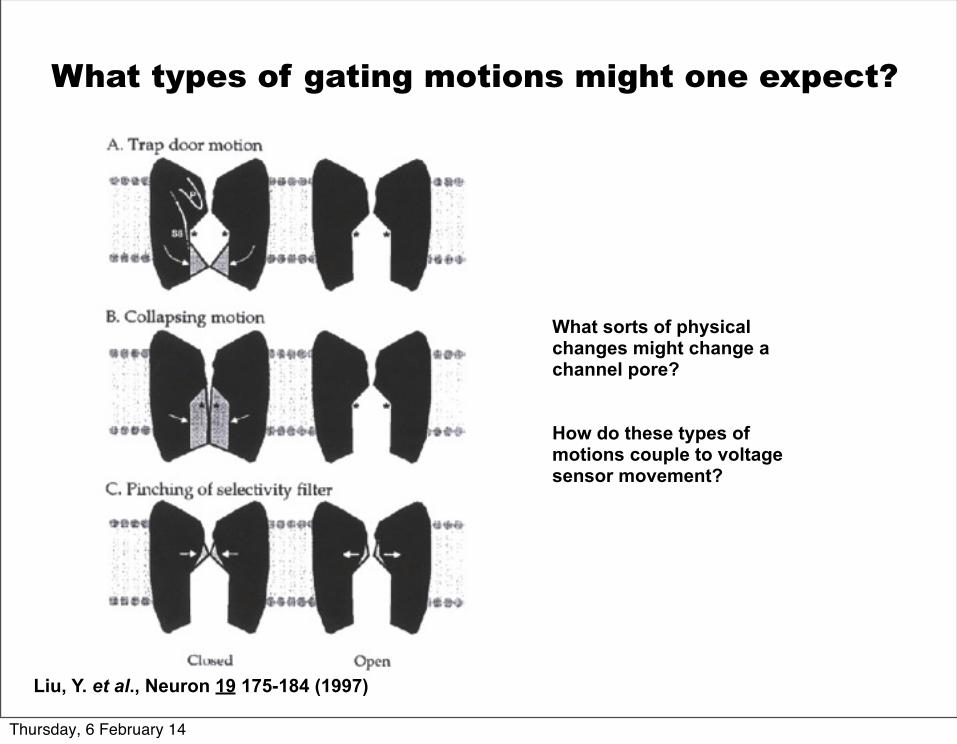
What sorts of physical changes might change a channel pore?
How do these types of motions couple to voltage sensor movement?
Liu, Y. et al., Neuron 19 175-184 (1997)
What types of gating motions might one expect?
Thursday, 6 February 14
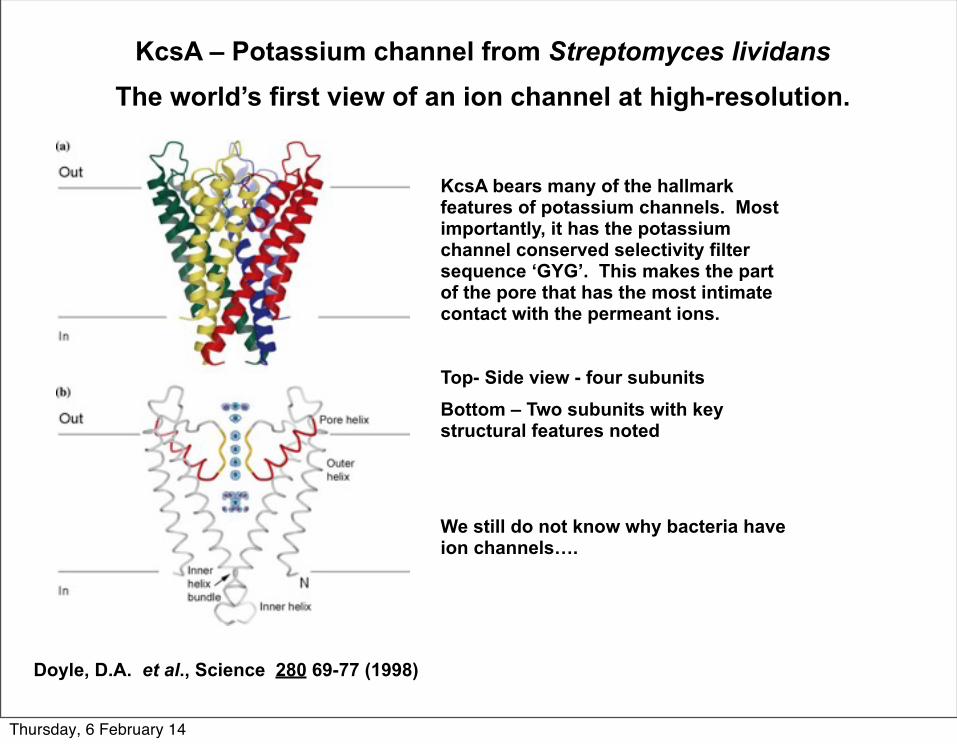
KcsA – Potassium channel from Streptomyces lividansThe world’s first view of an ion channel at high-resolution.
KcsA bears many of the hallmark features of potassium channels. Most importantly, it has the potassium channel conserved selectivity filter sequence ‘GYG’. This makes the part of the pore that has the most intimate contact with the permeant ions.
Top- Side view - four subunitsBottom – Two subunits with key structural features noted
We still do not know why bacteria have ion channels….
Doyle, D.A. et al., Science 280 69-77 (1998)
Thursday, 6 February 14
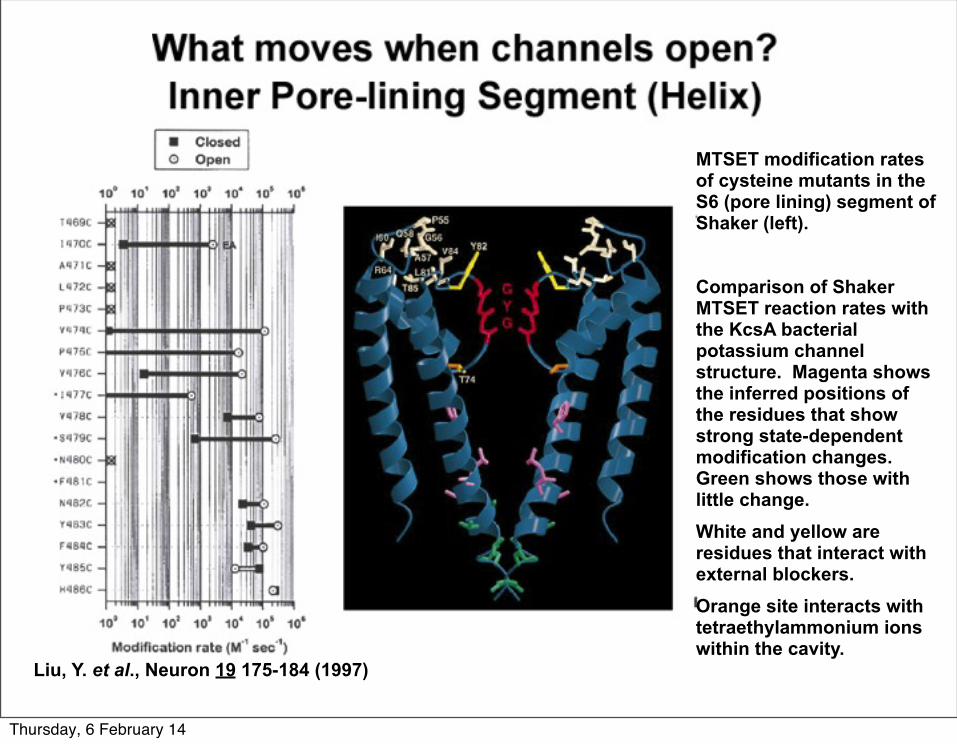
Liu, Y. et al., Neuron 19 175-184 (1997)
MTSET modification rates of cysteine mutants in the S6 (pore lining) segment of Shaker (left).
Comparison of Shaker MTSET reaction rates with the KcsA bacterial potassium channel structure. Magenta shows the inferred positions of the residues that show strong state-dependent modification changes. Green shows those with little change. White and yellow are residues that interact with external blockers.Orange site interacts with tetraethylammonium ions within the cavity.
Thursday, 6 February 14

Comparison of transmembrane sequences shows conservation of key residues in the voltage-sensor, S4, residues that interact with the charges in the voltage sensro (S2 & S3), and in the potassium selectivity filter (all shown in red).
Ruta, V., et al., Nature 422 180-185 (2003)
Voltage-gated channelsKvAP (Aeropyrum pernix, archea, thermal vent)
Thursday, 6 February 14
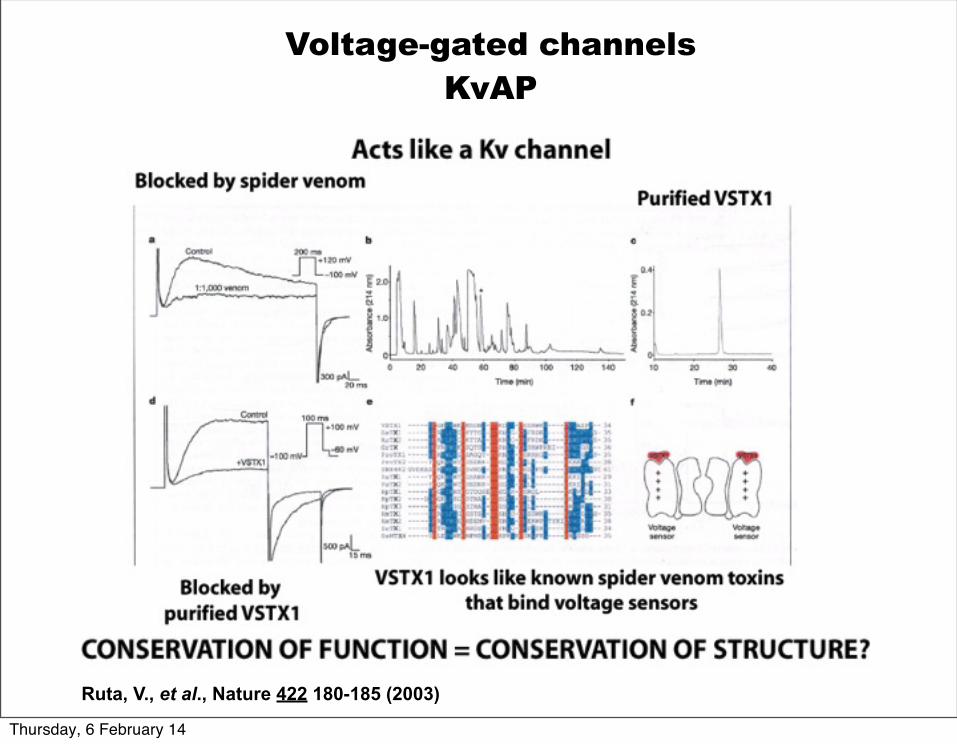
Ruta, V., et al., Nature 422 180-185 (2003)
Voltage-gated channelsKvAP
Thursday, 6 February 14

Jiang, Y., et al., Nature 423 33-41 (2003)
View from the cytoplasmic side of the KvAP structure. Each subunit is colored differently. The green molecules are conformational specific monoclonal Fab’s that bind to the voltage sensor.
Voltage-gated channelsKvAP
3.2 Å
Thursday, 6 February 14
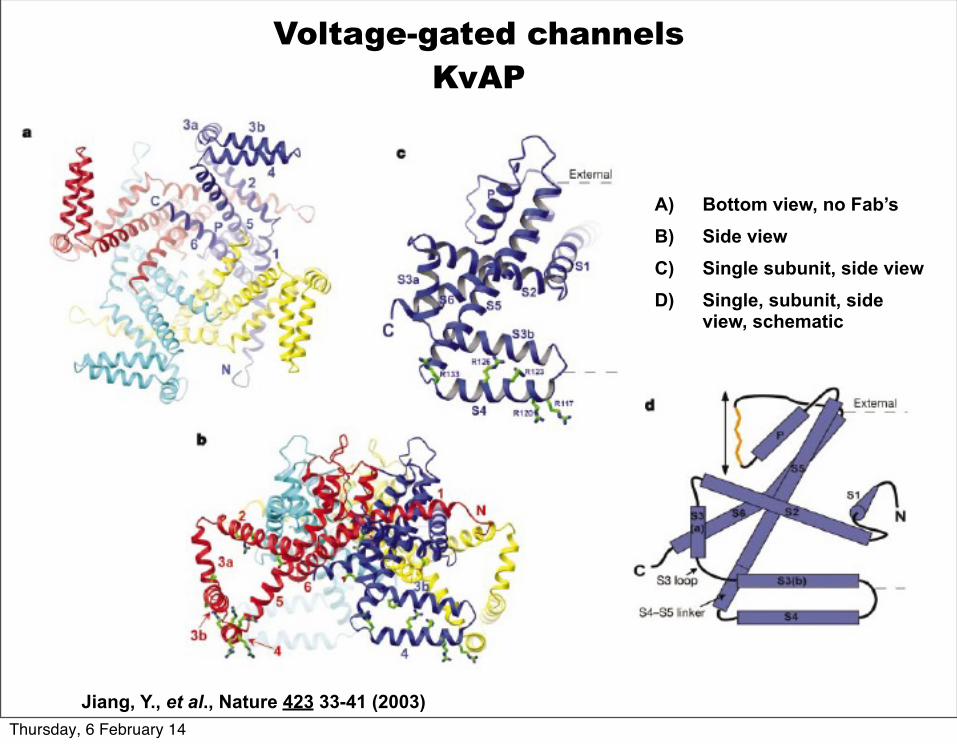
A) Bottom view, no Fab’sB) Side viewC) Single subunit, side viewD) Single, subunit, side
view, schematic
Jiang, Y., et al., Nature 423 33-41 (2003)
Voltage-gated channelsKvAP
Thursday, 6 February 14

Voltage-gated channelsKvAP - The paddle model
Jiang, Y., et al., Nature 423 42-48(2003)
An accessibility experiment to test the paddle model
Thursday, 6 February 14
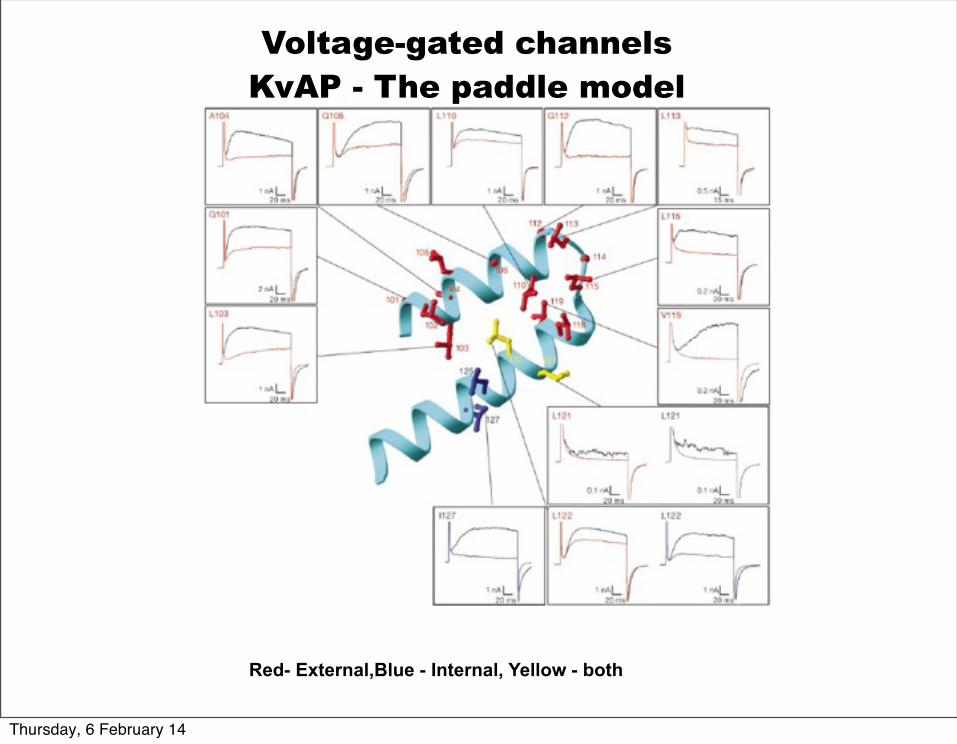
Voltage-gated channelsKvAP - The paddle model
Red- External,Blue - Internal, Yellow - both
Thursday, 6 February 14

Jiang, Y., et al., Nature 423 42-48(2003)
Voltage-gated channelsKvAP - The paddle model
Thursday, 6 February 14

Jiang, Y., et al., Nature 423 33-41 (2003)
Voltage-gated channelsKvAP Paddles vs. Pistons
Thursday, 6 February 14

Ruta, V., et al., Cell 123 463-475 (2005)
Voltage-gated channelsKvAP - The paddle model - further tests
Thursday, 6 February 14

Red- External, Blue - InternalYellow - both, Black - Neither
White - Zone of inaccessibility
Voltage-gated channelsKvAP - The paddle model - further tests
Ruta, V., et al., Cell 123 463-475 (2005)Thursday, 6 February 14
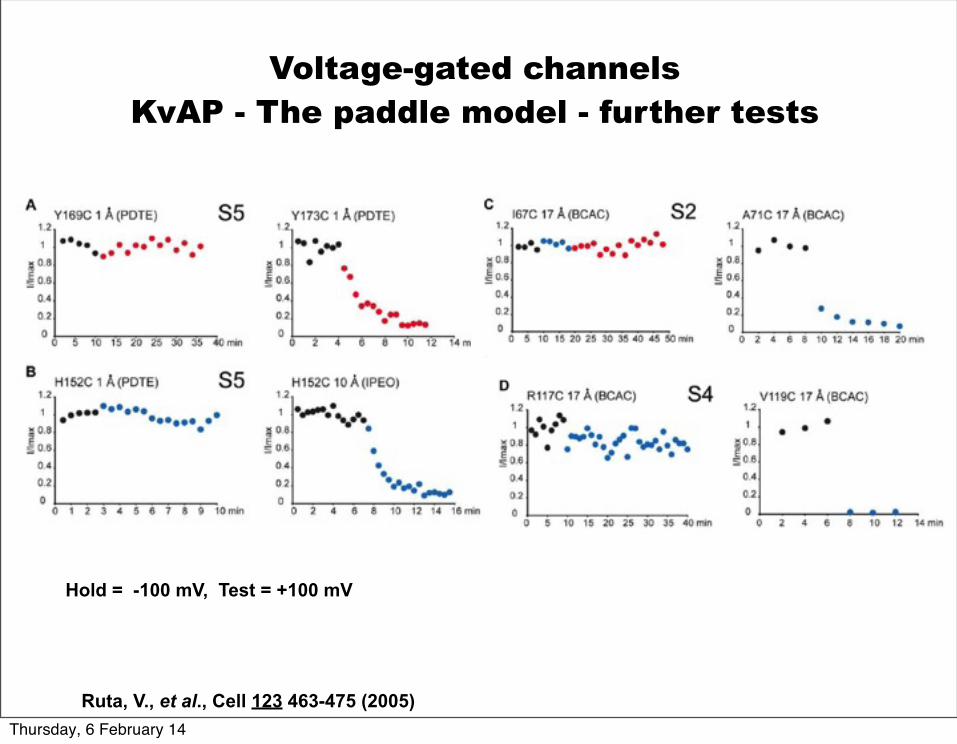
Ruta, V., et al., Cell 123 463-475 (2005)
Voltage-gated channelsKvAP - The paddle model - further tests
Hold = -100 mV, Test = +100 mV
Thursday, 6 February 14

Ruta, V., et al., Cell 123 463-475 (2005)
Red- External, Blue - InternalYellow - both, Black - Neither
White - Zone of inaccessibility
Voltage-gated channelsKvAP - The paddle model - further tests S1/S2
Thursday, 6 February 14

Red- External, Blue - InternalYellow - both, Black - Neither
White - Zone of inaccessibility
Voltage-gated channelsKvAP - The paddle model - further tests S3/S4
Thursday, 6 February 14
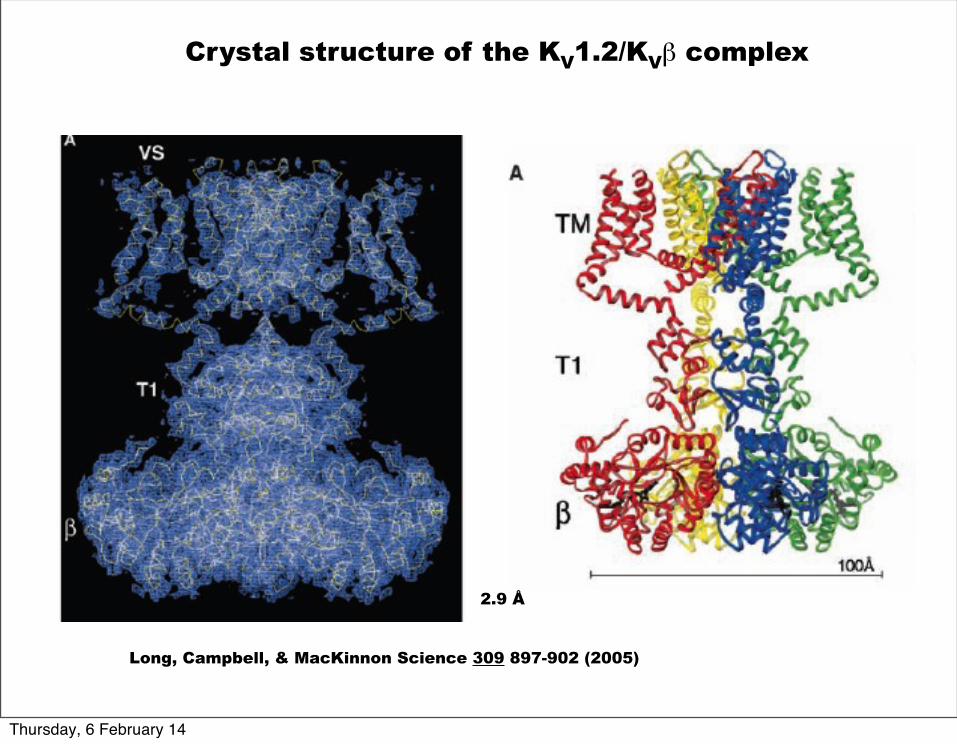
Long, Campbell, & MacKinnon Science 309 897-902 (2005)
Crystal structure of the KV1.2/KVβ complex
2.9 Å
Thursday, 6 February 14

Long, Campbell, & MacKinnon Science 309 897-902 (2005)
Crystal structure of the KV1.2/KVβ complex
Voltage sensors (S1-S4) are packed against the pore (S5-S6) of the adjacent subunit.
S1-S4
S5-S6
Thursday, 6 February 14
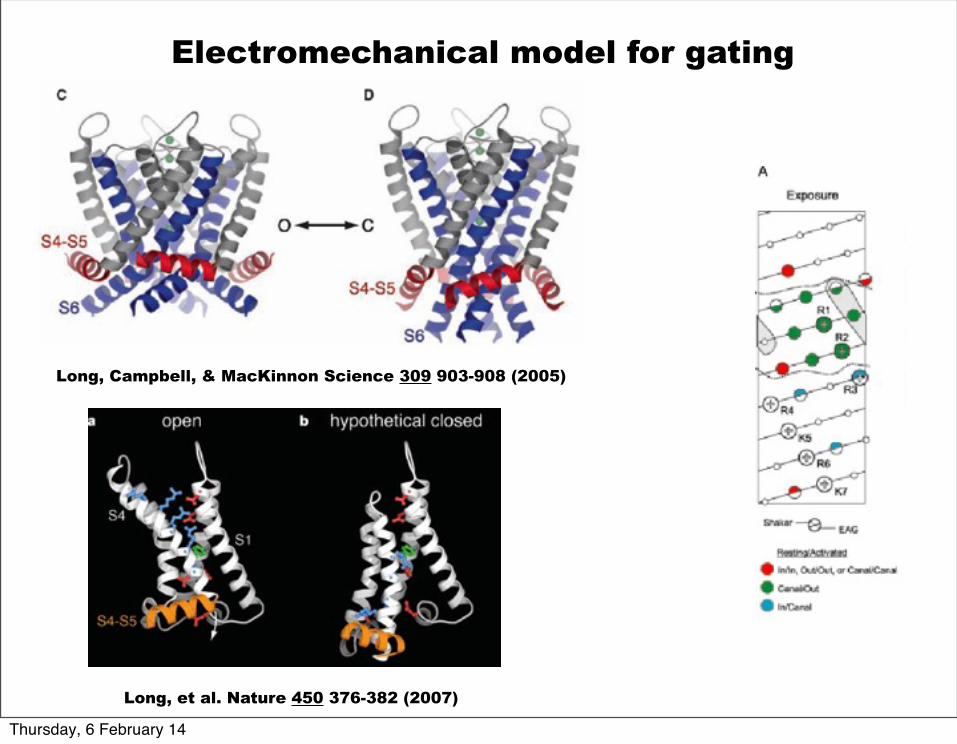
Long, Campbell, & MacKinnon Science 309 903-908 (2005)
Electromechanical model for gating
Long, et al. Nature 450 376-382 (2007)
Thursday, 6 February 14

Pathak et al. Neuron 56 124-140 (2007)
Models of how voltage sensing works
Thursday, 6 February 14
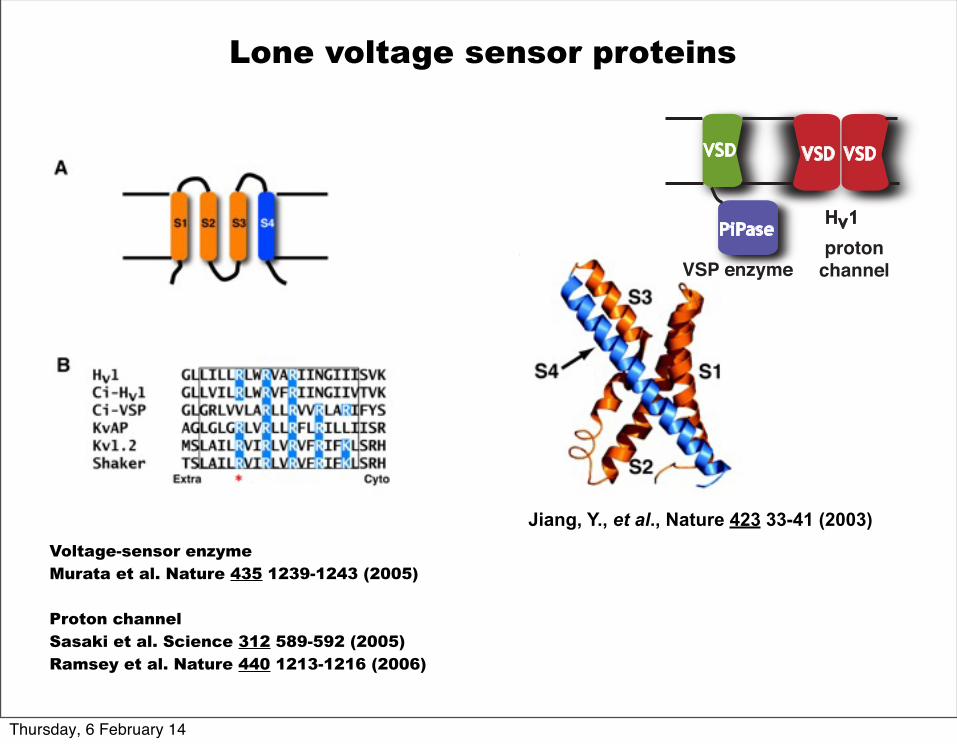
Lone voltage sensor proteins
Voltage-sensor enzymeMurata et al. Nature 435 1239-1243 (2005)
Proton channelSasaki et al. Science 312 589-592 (2005)Ramsey et al. Nature 440 1213-1216 (2006)
Jiang, Y., et al., Nature 423 33-41 (2003)
PiPaseHv1
VSP enzymeproton
channel
VSD VSDVSD
Thursday, 6 February 14
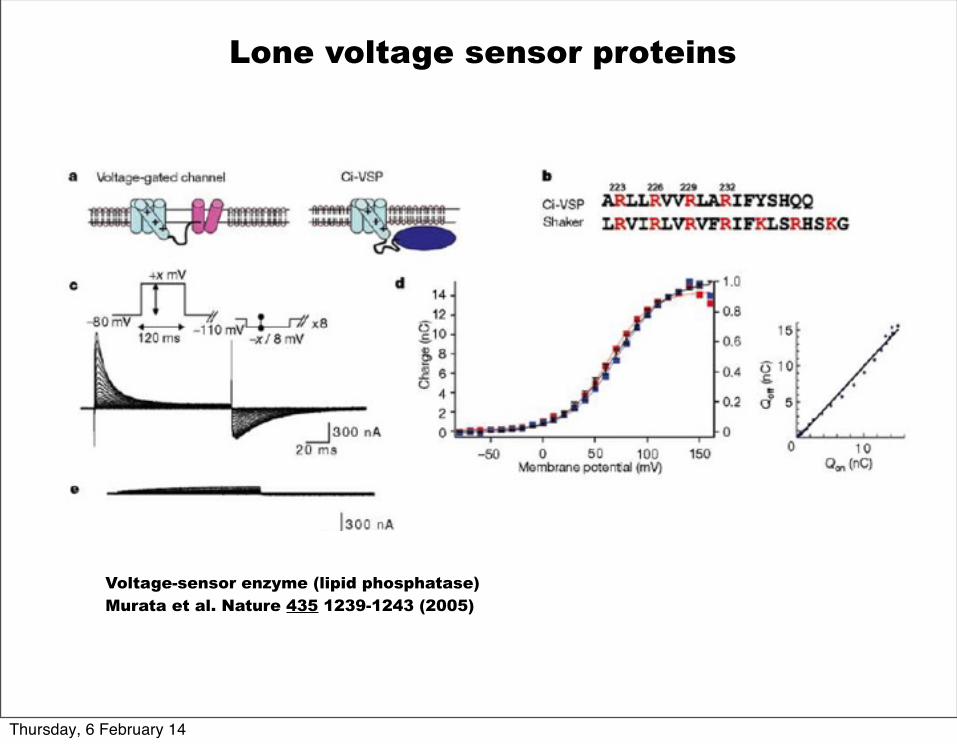
Voltage-sensor enzyme (lipid phosphatase)Murata et al. Nature 435 1239-1243 (2005)
Lone voltage sensor proteins
Thursday, 6 February 14
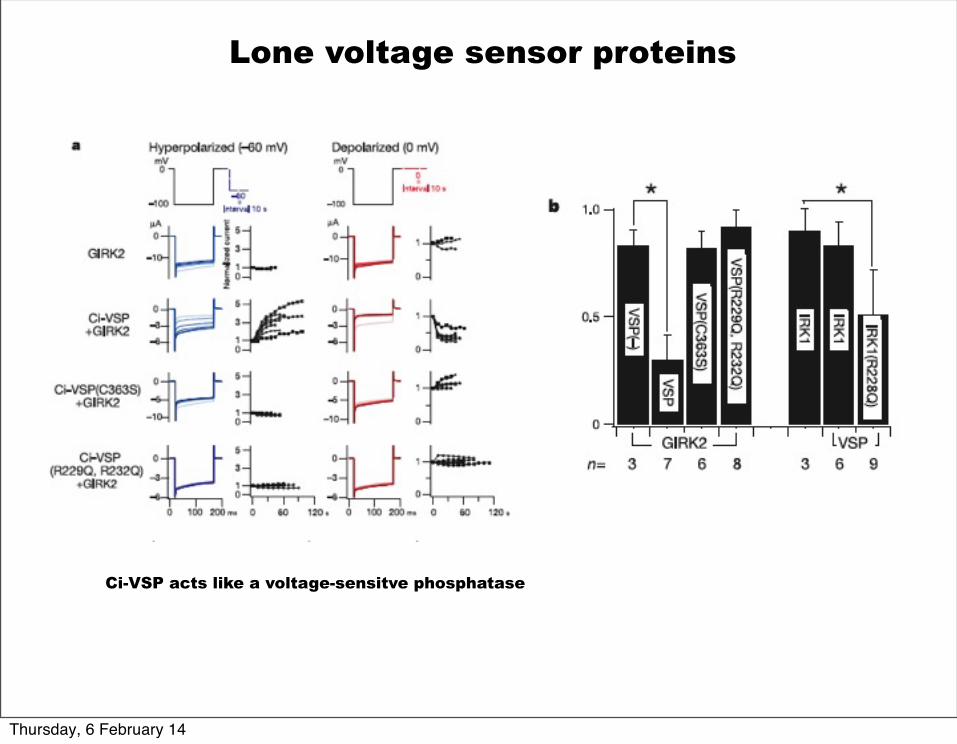
Lone voltage sensor proteins
Ci-VSP acts like a voltage-sensitve phosphatase
Thursday, 6 February 14
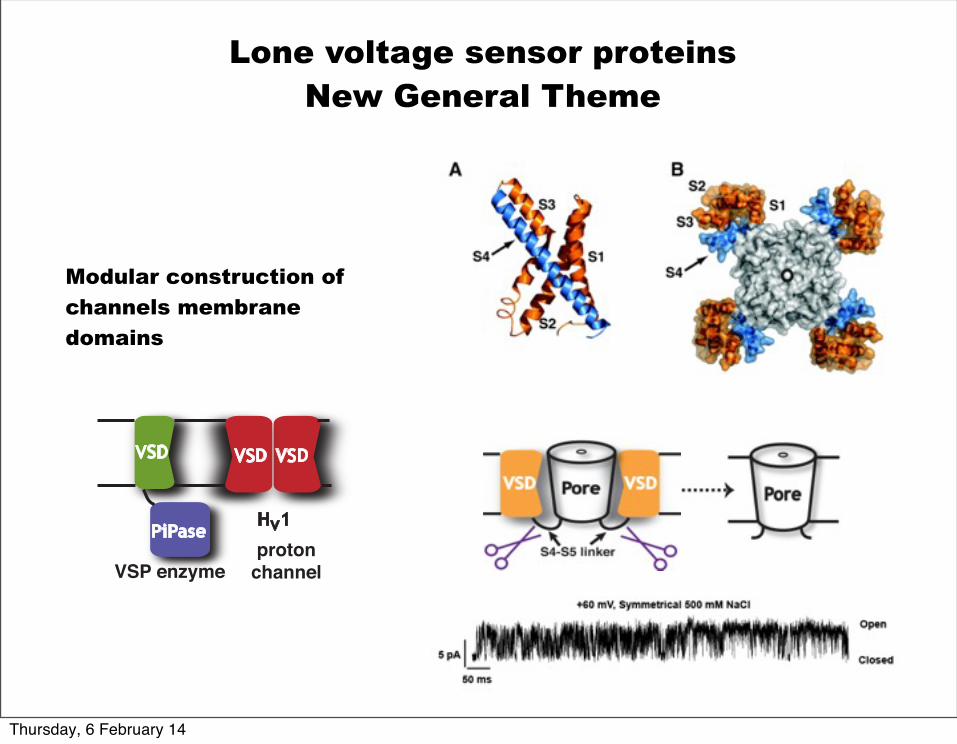
Lone voltage sensor proteins New General Theme
Modular construction of channels membrane domains
PiPaseHv1
VSP enzymeproton
channel
VSD VSDVSD
Thursday, 6 February 14
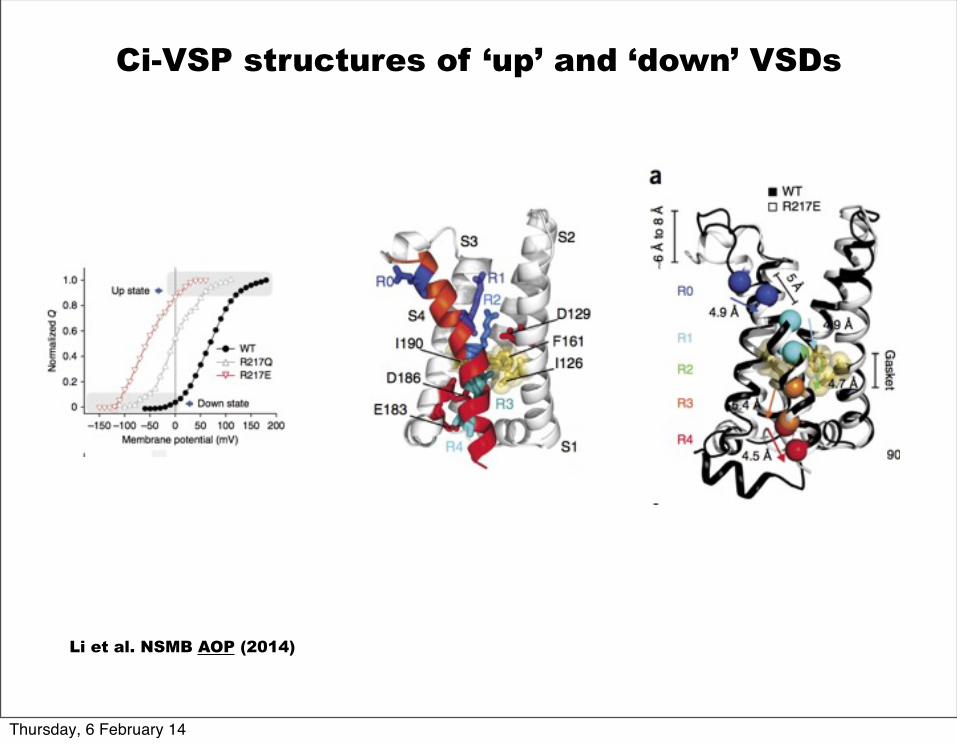
Ci-VSP structures of ‘up’ and ‘down’ VSDs
Li et al. NSMB AOP (2014)
Thursday, 6 February 14
46
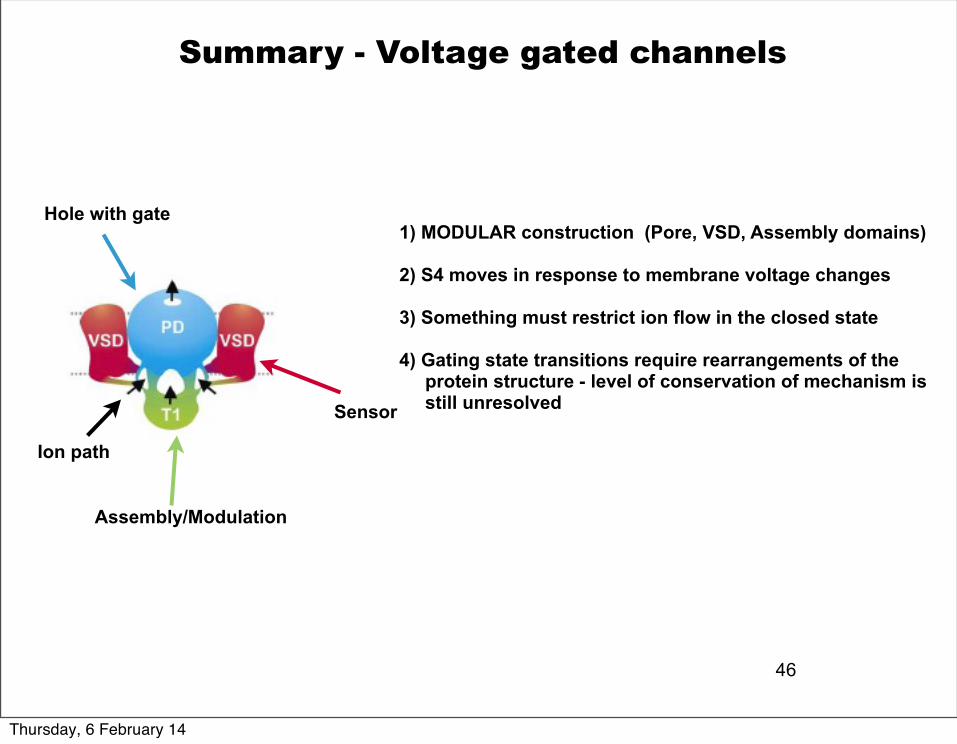
Summary - Voltage gated channels
1) MODULAR construction (Pore, VSD, Assembly domains)
2) S4 moves in response to membrane voltage changes
3) Something must restrict ion flow in the closed state
4) Gating state transitions require rearrangements of the protein structure - level of conservation of mechanism is still unresolved
Hole with gate
Sensor
Assembly/Modulation
Ion path
Thursday, 6 February 14

Ligand-gated ion channels
Acetylcholine receptor is a pentamer and is related to serotonin receptor (5HT3R), glycine receptor and GABAA receptor
Thursday, 6 February 14
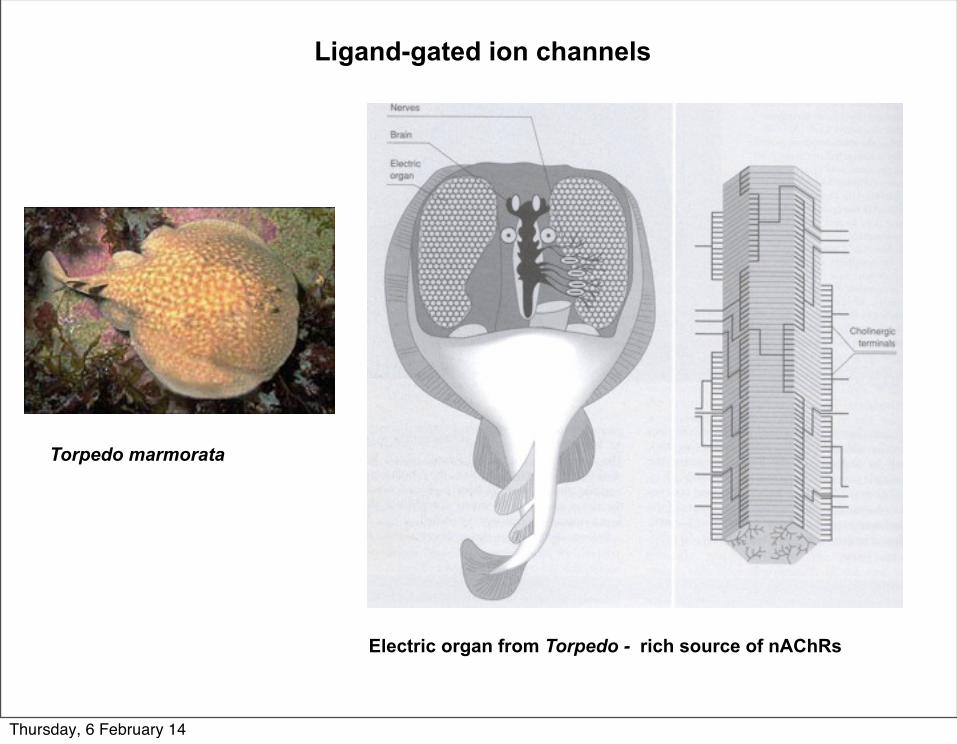
Ligand-gated ion channels
Torpedo marmorata
Electric organ from Torpedo - rich source of nAChRs
Thursday, 6 February 14
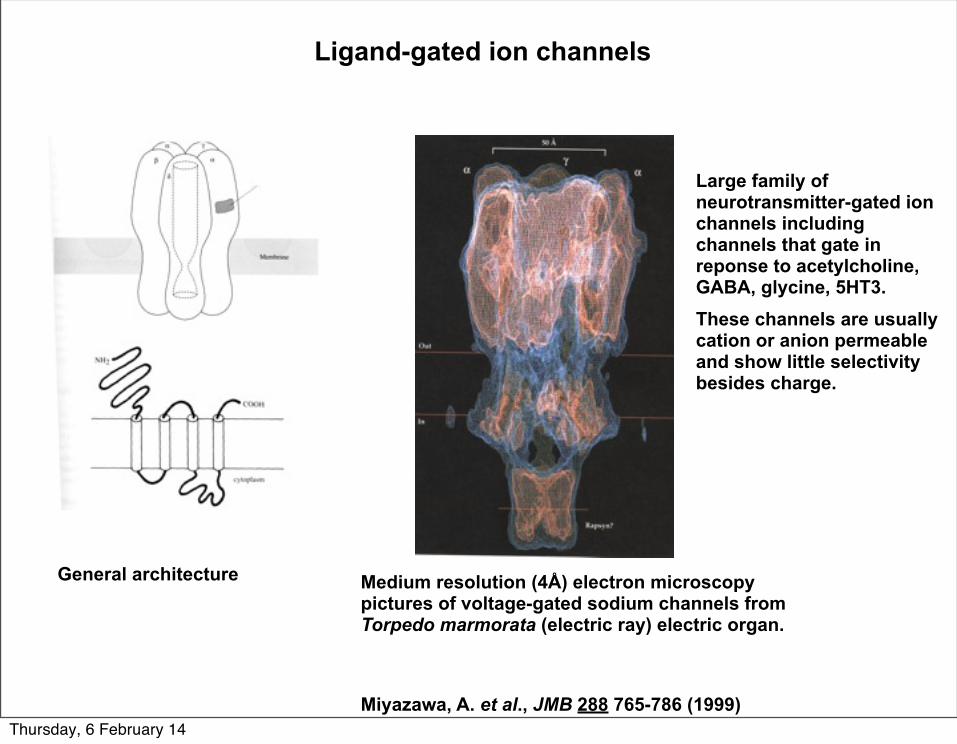
Ligand-gated ion channels
Large family of neurotransmitter-gated ion channels including channels that gate in reponse to acetylcholine, GABA, glycine, 5HT3.These channels are usually cation or anion permeable and show little selectivity besides charge.
General architecture Medium resolution (4Å) electron microscopy pictures of voltage-gated sodium channels from Torpedo marmorata (electric ray) electric organ.
Miyazawa, A. et al., JMB 288 765-786 (1999)Thursday, 6 February 14
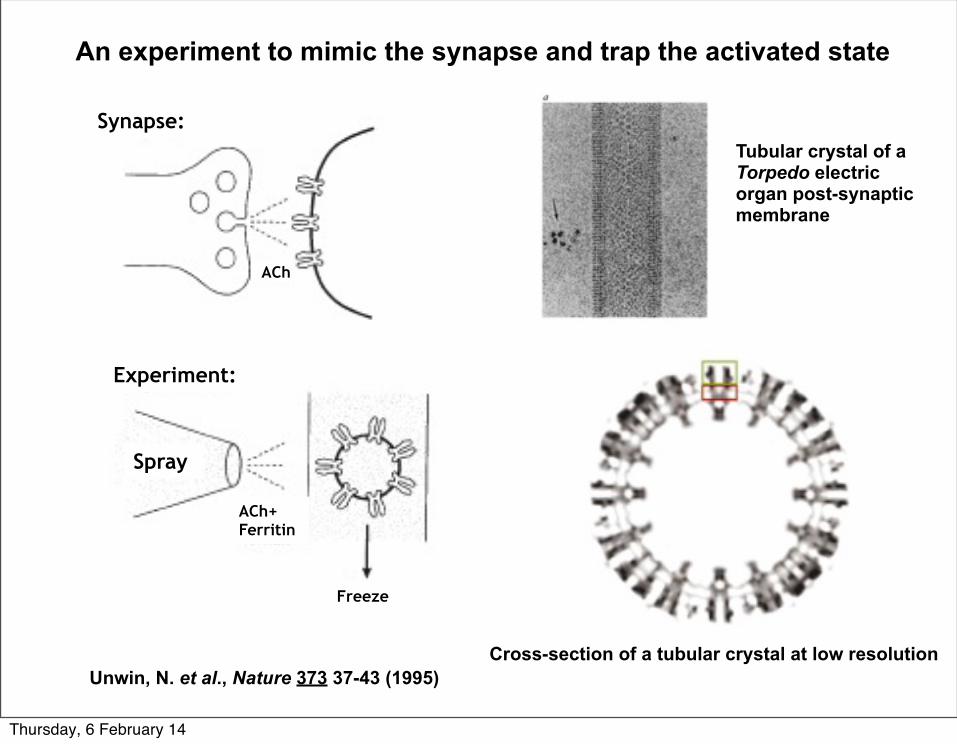
Spray
Experiment:
Synapse:
ACh+ Ferritin
Freeze
An experiment to mimic the synapse and trap the activated state
Unwin, N. et al., Nature 373 37-43 (1995)Cross-section of a tubular crystal at low resolution
Tubular crystal of a Torpedo electric organ post-synaptic membrane
ACh
Thursday, 6 February 14
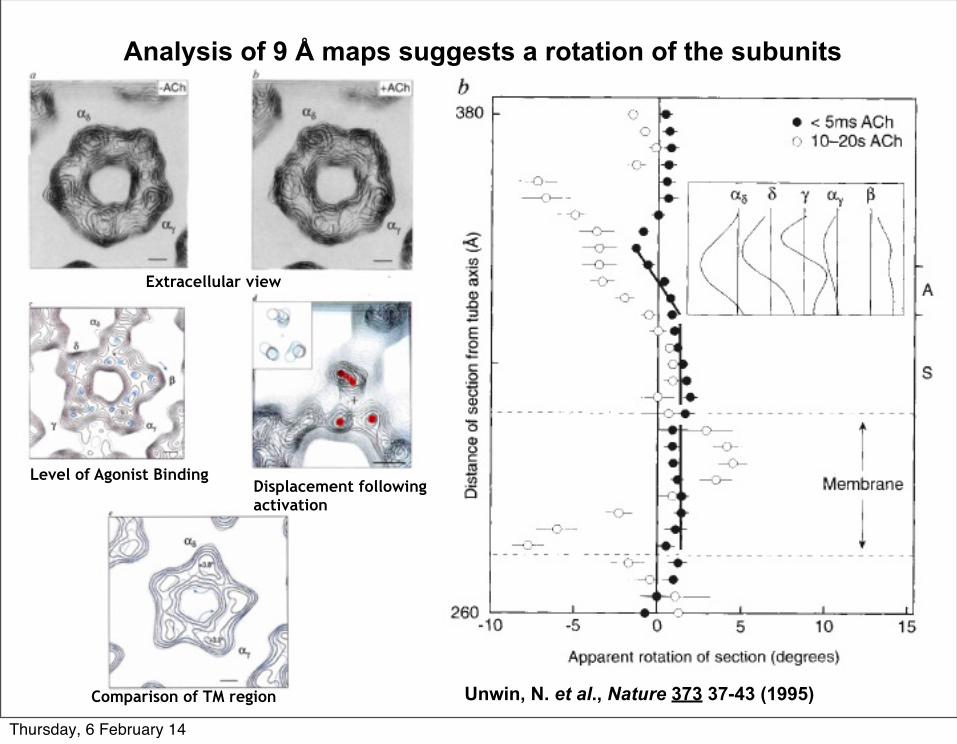
Analysis of 9 Å maps suggests a rotation of the subunits
Unwin, N. et al., Nature 373 37-43 (1995)
Extracellular view
Level of Agonist BindingDisplacement following activation
Comparison of TM region
Thursday, 6 February 14
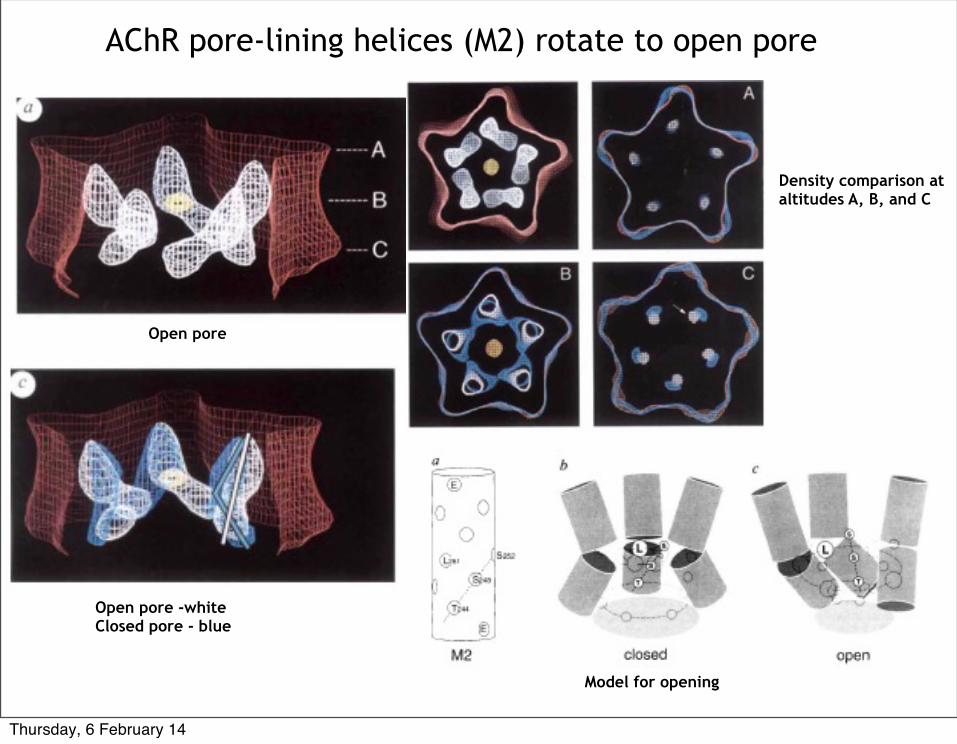
AChR pore-lining helices (M2) rotate to open pore
Open pore
Open pore -whiteClosed pore - blue
Density comparison at altitudes A, B, and C
Model for opening
Thursday, 6 February 14
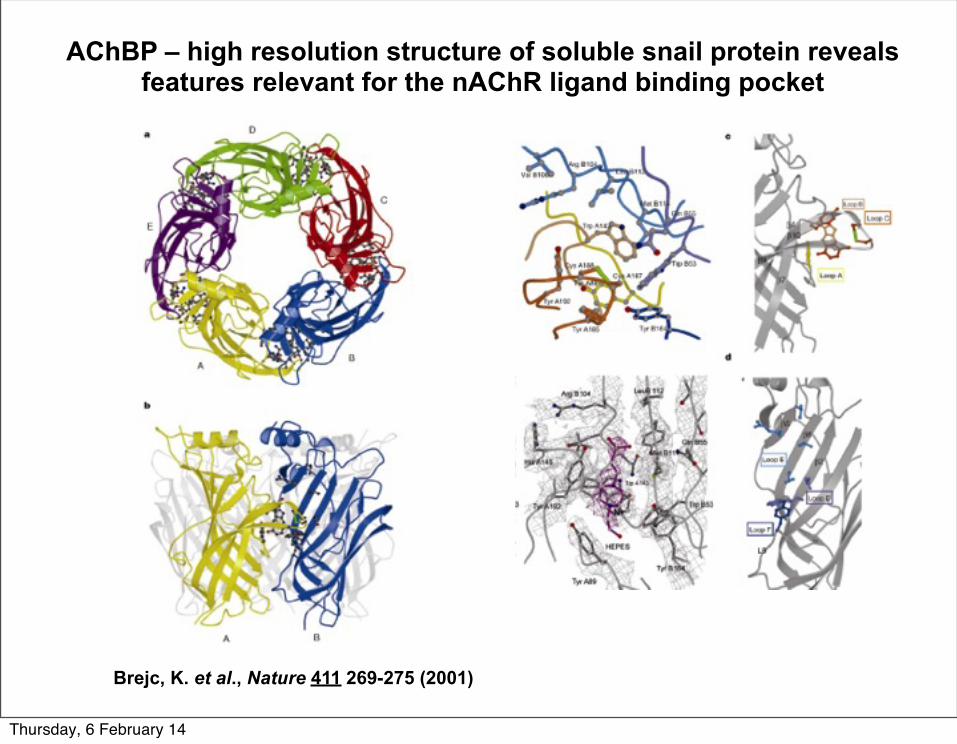
AChBP – high resolution structure of soluble snail protein reveals features relevant for the nAChR ligand binding pocket
Brejc, K. et al., Nature 411 269-275 (2001)
Thursday, 6 February 14
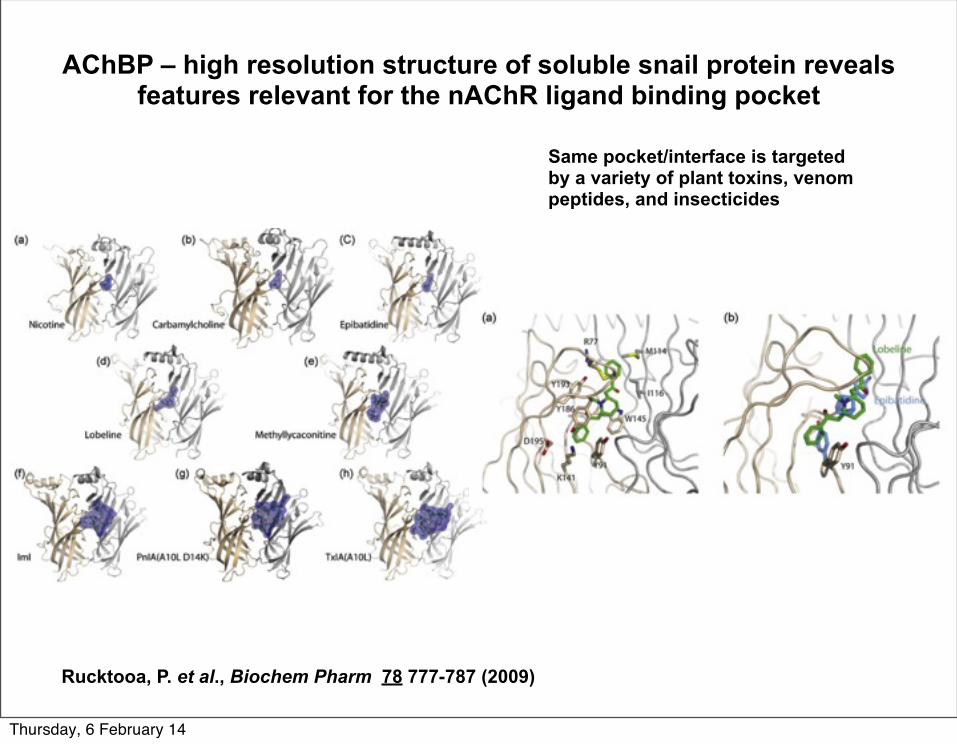
AChBP – high resolution structure of soluble snail protein reveals features relevant for the nAChR ligand binding pocket
Rucktooa, P. et al., Biochem Pharm 78 777-787 (2009)
Same pocket/interface is targeted by a variety of plant toxins, venom peptides, and insecticides
Thursday, 6 February 14
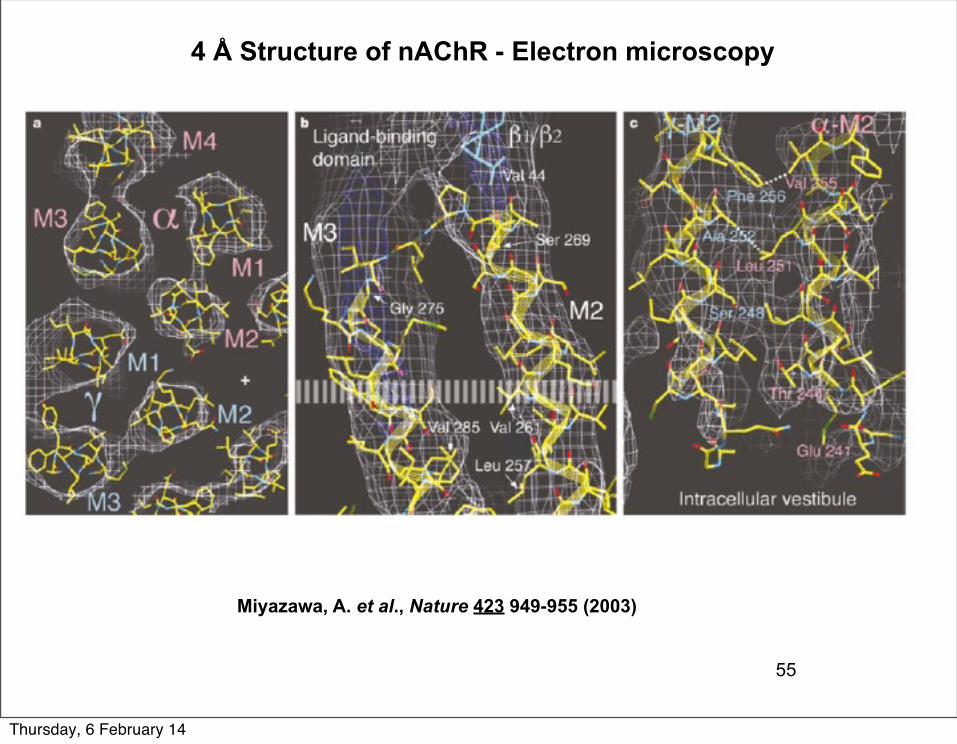
55
Miyazawa, A. et al., Nature 423 949-955 (2003)
4 Å Structure of nAChR - Electron microscopy
Thursday, 6 February 14
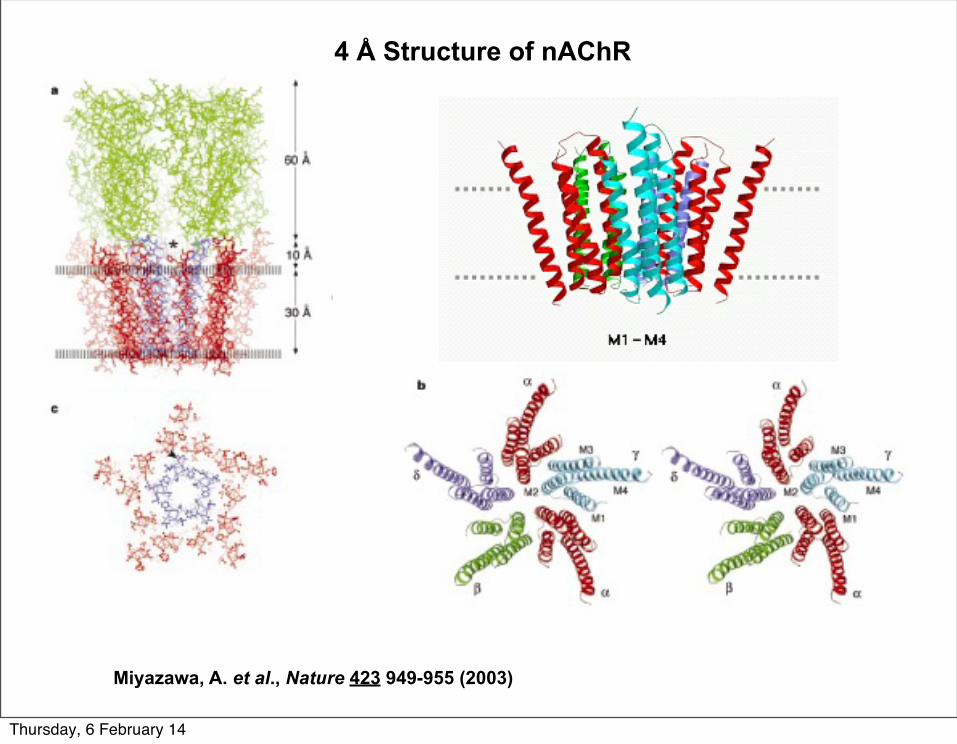
4 Å Structure of nAChR
Miyazawa, A. et al., Nature 423 949-955 (2003)
Thursday, 6 February 14
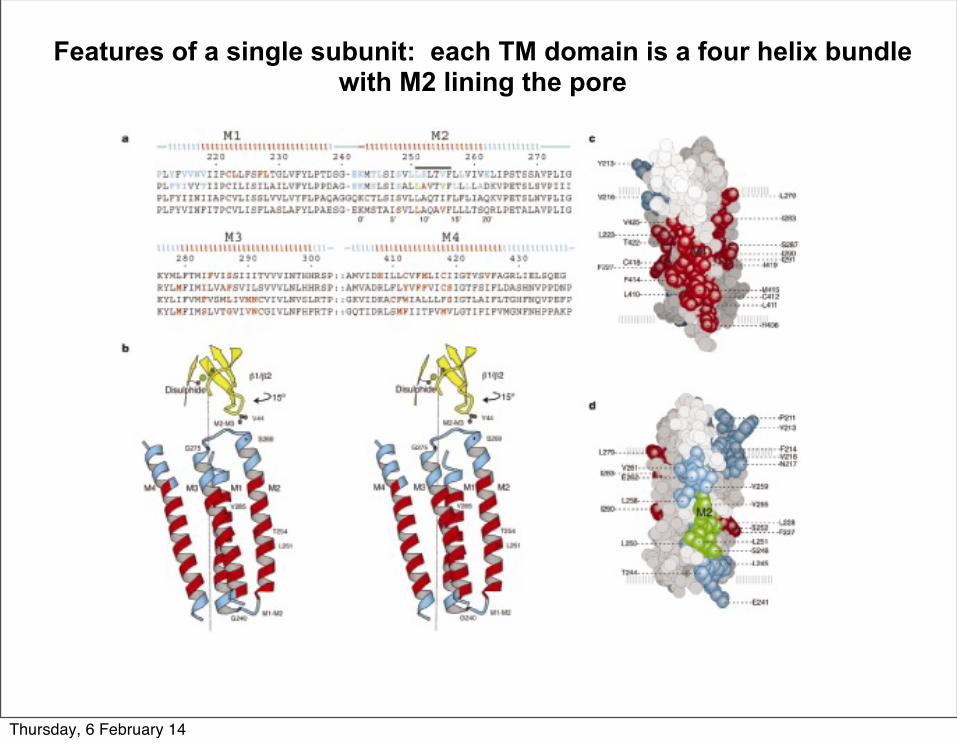
Features of a single subunit: each TM domain is a four helix bundle with M2 lining the pore
Thursday, 6 February 14
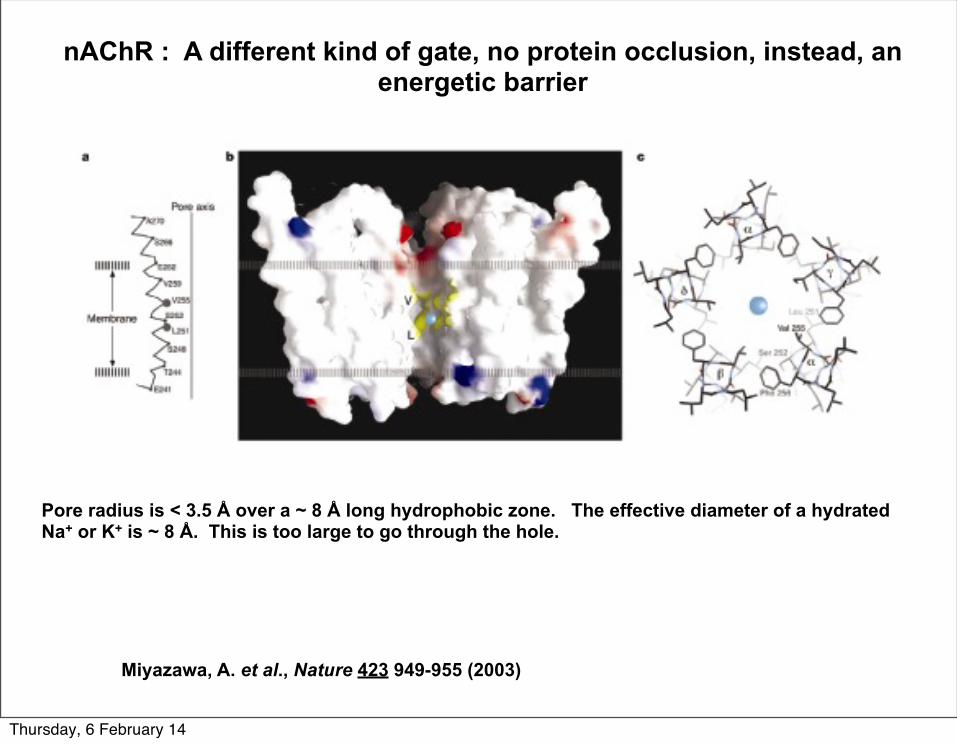
nAChR : A different kind of gate, no protein occlusion, instead, an energetic barrier
Miyazawa, A. et al., Nature 423 949-955 (2003)
Pore radius is < 3.5 Å over a ~ 8 Å long hydrophobic zone. The effective diameter of a hydrated Na+ or K+ is ~ 8 Å. This is too large to go through the hole.
Thursday, 6 February 14
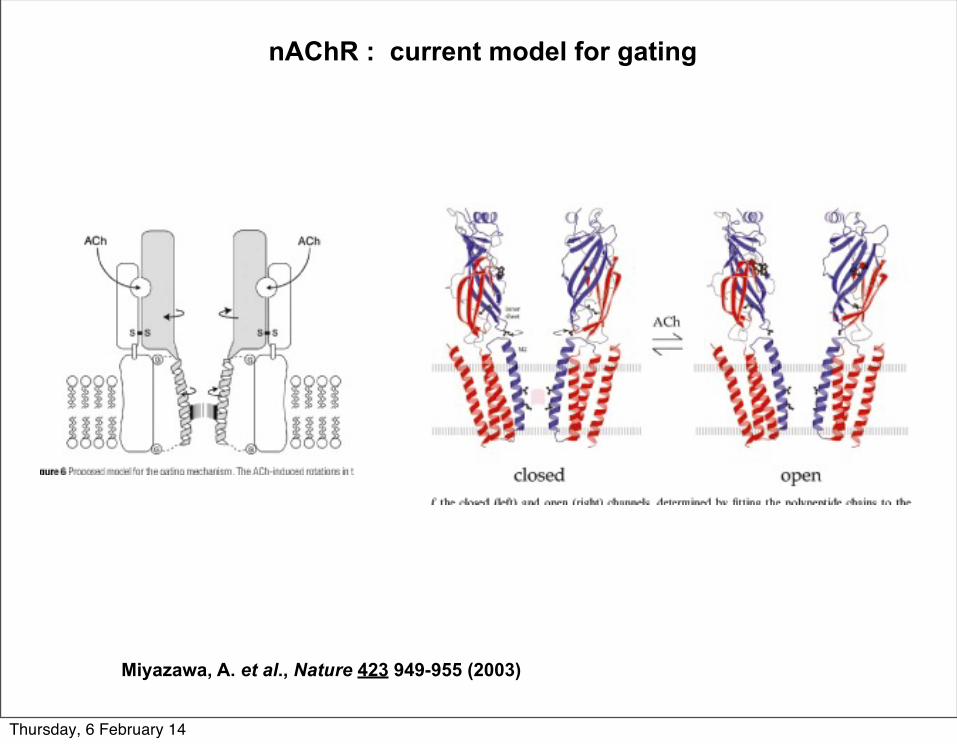
nAChR : current model for gating
Miyazawa, A. et al., Nature 423 949-955 (2003)
Thursday, 6 February 14
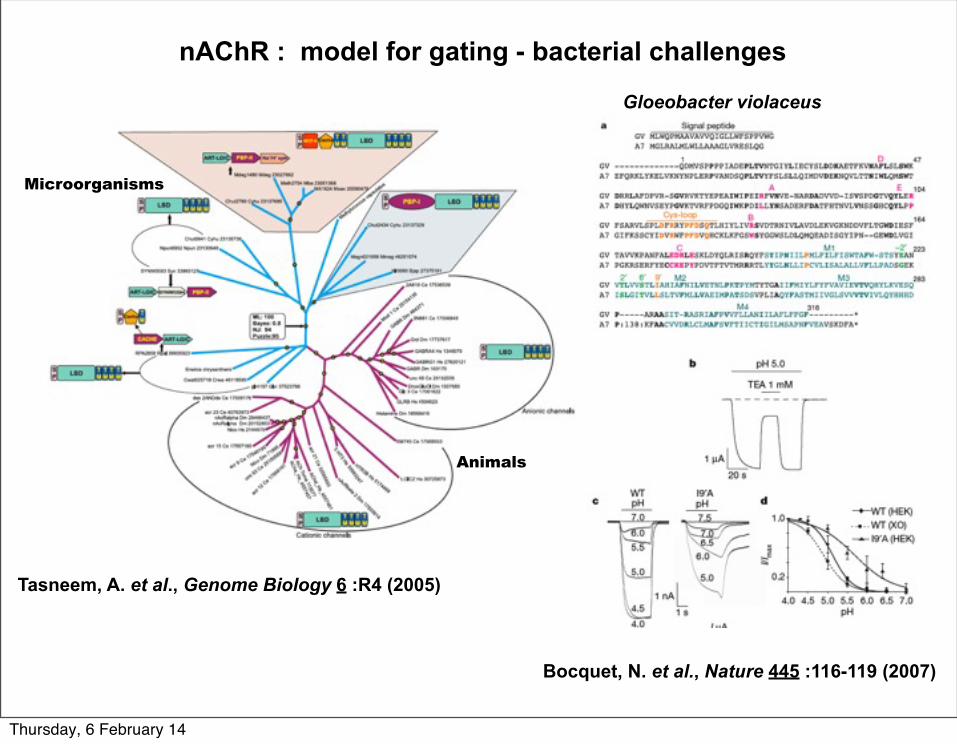
nAChR : model for gating - bacterial challenges
Tasneem, A. et al., Genome Biology 6 :R4 (2005)
Microorganisms
Animals
Bocquet, N. et al., Nature 445 :116-119 (2007)
Gloeobacter violaceus
Thursday, 6 February 14
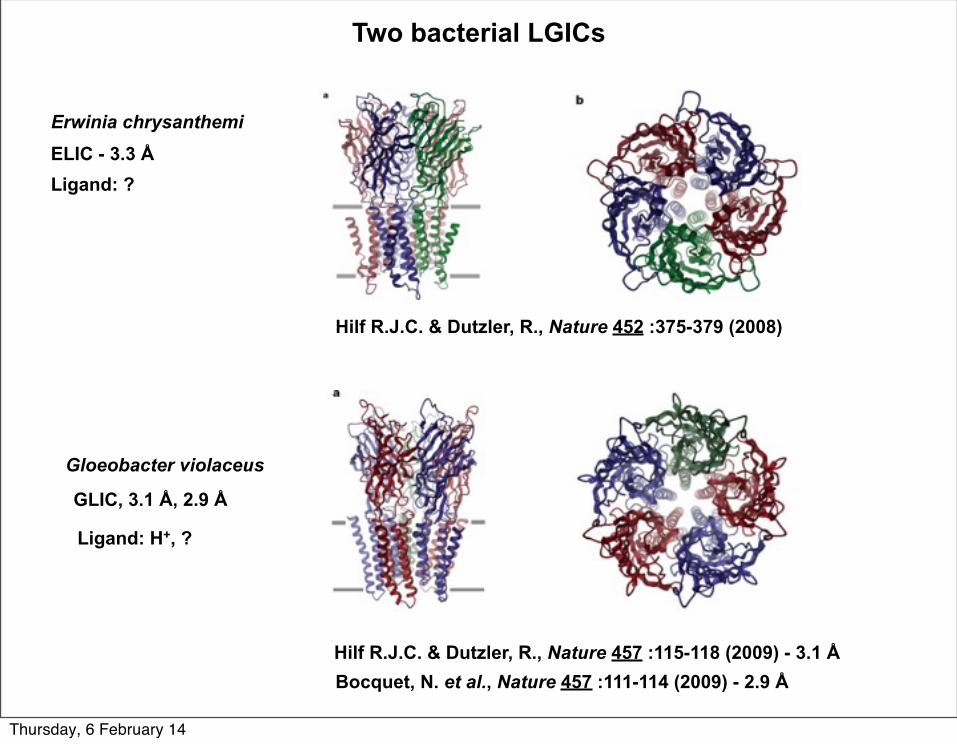
Hilf R.J.C. & Dutzler, R., Nature 457 :115-118 (2009) - 3.1 Å
Two bacterial LGICs
ELIC - 3.3 Å
GLIC, 3.1 Å, 2.9 Å
Ligand: ?
Ligand: H+, ?
Bocquet, N. et al., Nature 457 :111-114 (2009) - 2.9 Å
Hilf R.J.C. & Dutzler, R., Nature 452 :375-379 (2008)
Erwinia chrysanthemi
Gloeobacter violaceus
Thursday, 6 February 14
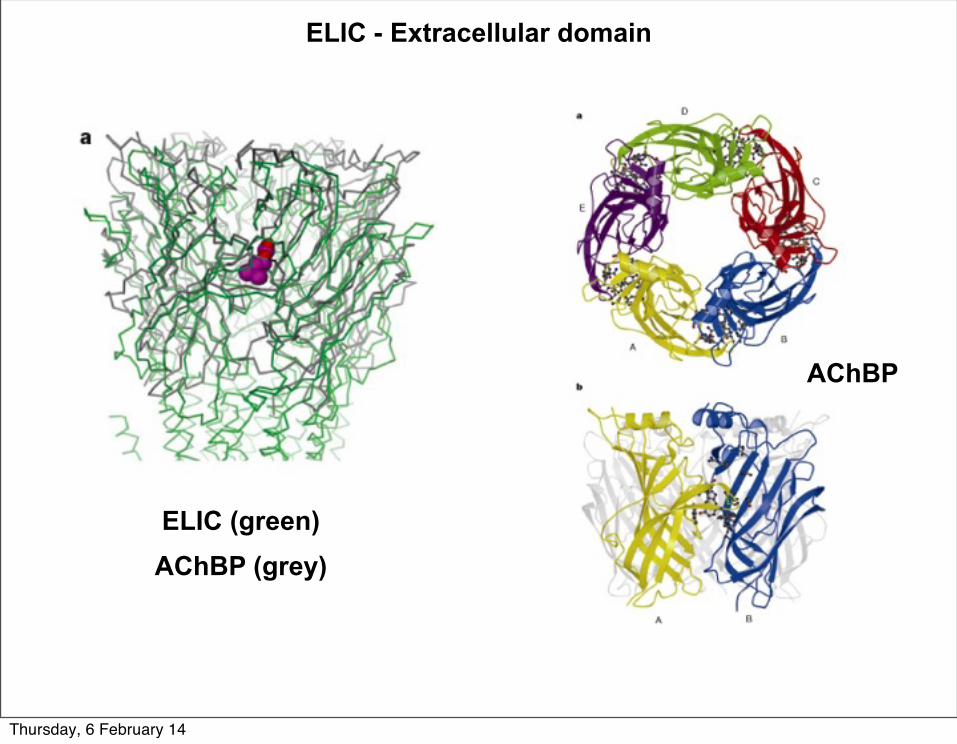
AChBP
ELIC (green)AChBP (grey)
ELIC - Extracellular domain
Thursday, 6 February 14
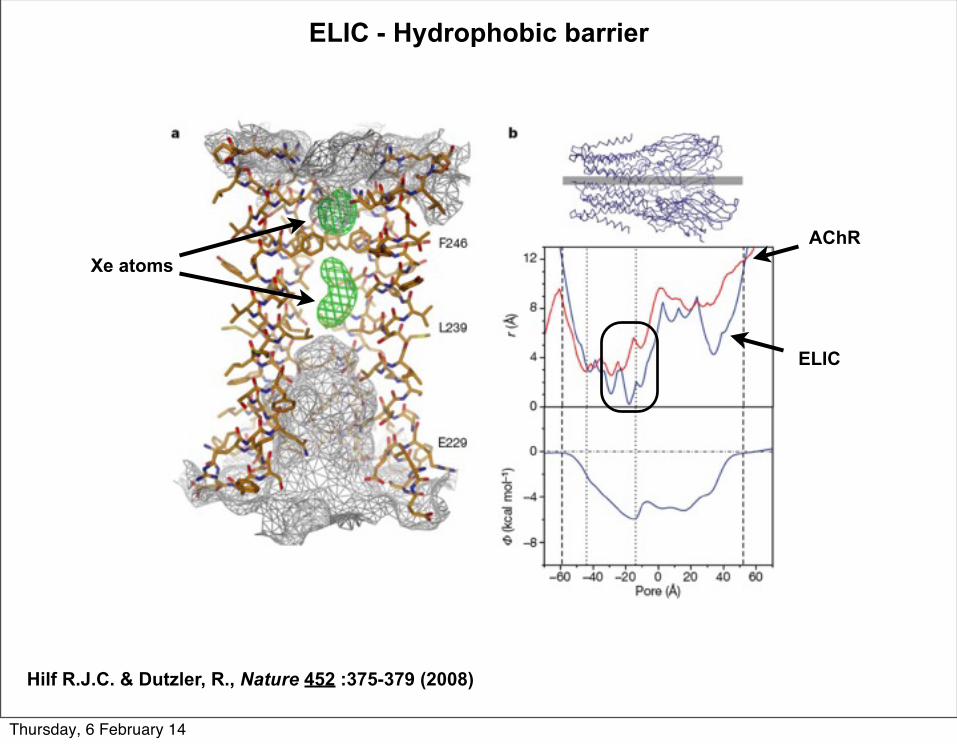
Xe atoms
ELIC
AChR
ELIC - Hydrophobic barrier
Hilf R.J.C. & Dutzler, R., Nature 452 :375-379 (2008)
Thursday, 6 February 14
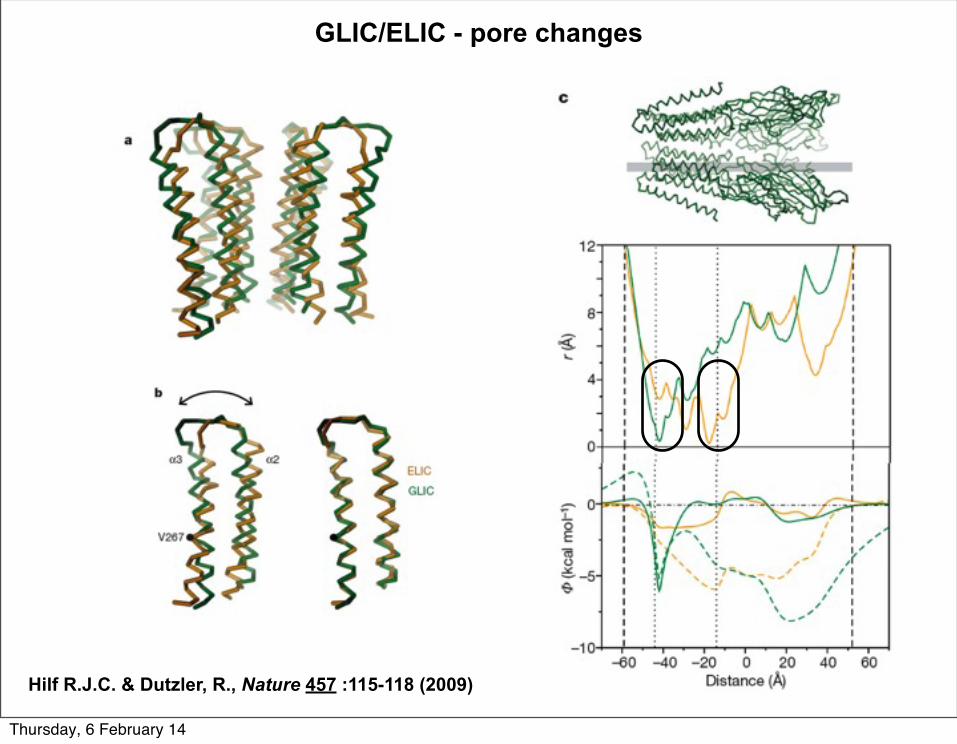
GLIC/ELIC - pore changes
Hilf R.J.C. & Dutzler, R., Nature 457 :115-118 (2009)
Thursday, 6 February 14
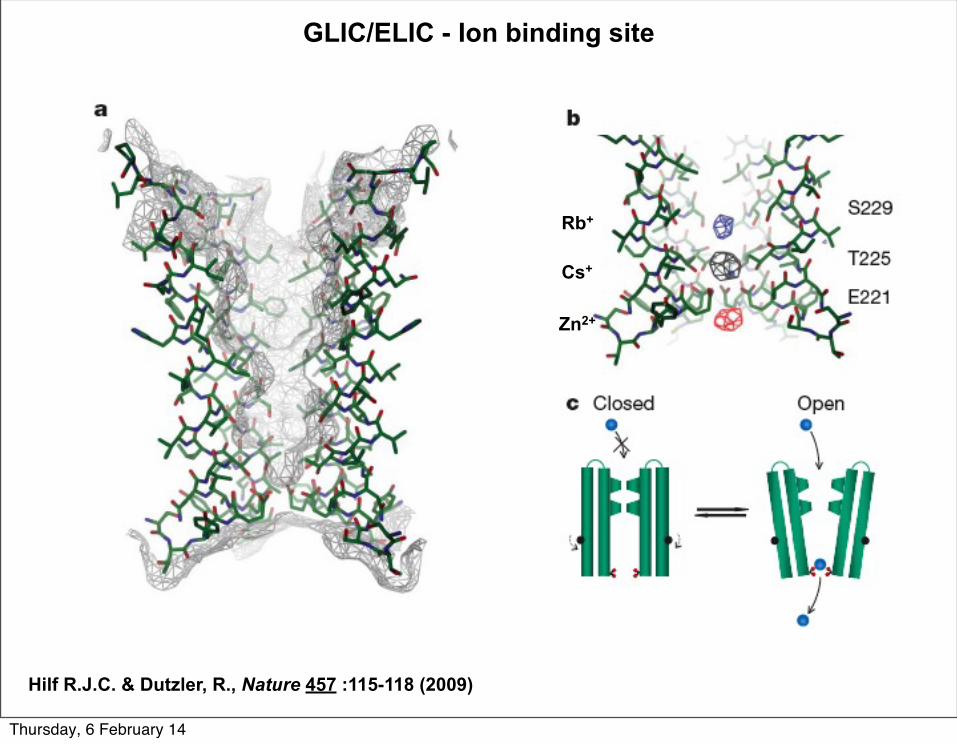
GLIC/ELIC - Ion binding site
Hilf R.J.C. & Dutzler, R., Nature 457 :115-118 (2009)
Rb+
Zn2+
Cs+
Thursday, 6 February 14
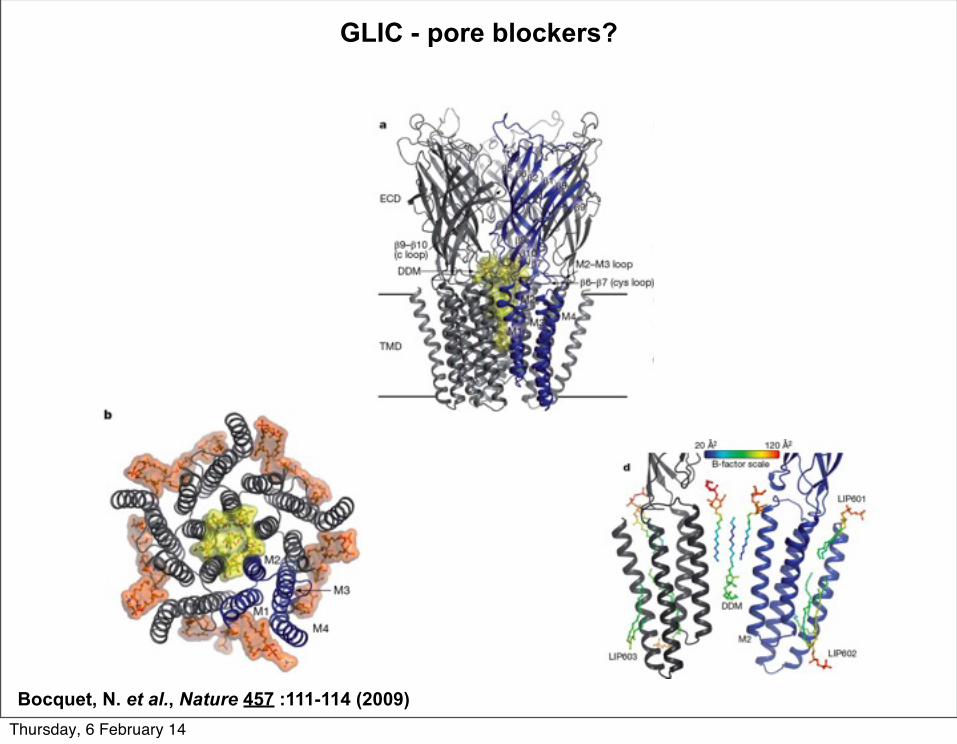
GLIC - pore blockers?
Bocquet, N. et al., Nature 457 :111-114 (2009)Thursday, 6 February 14
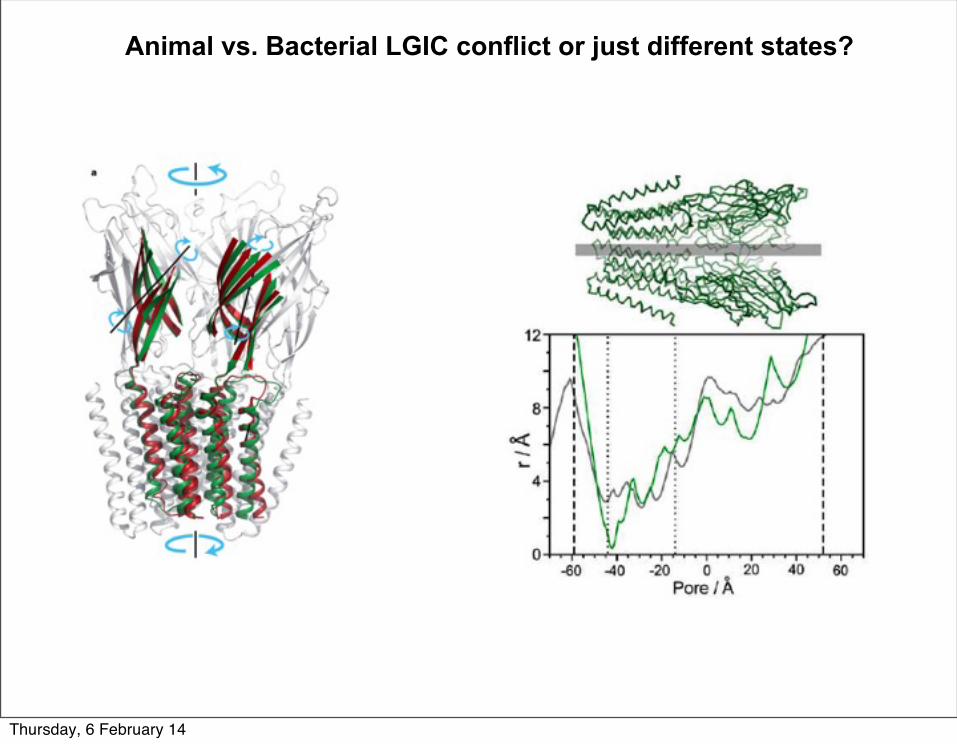
Animal vs. Bacterial LGIC conflict or just different states?
Thursday, 6 February 14
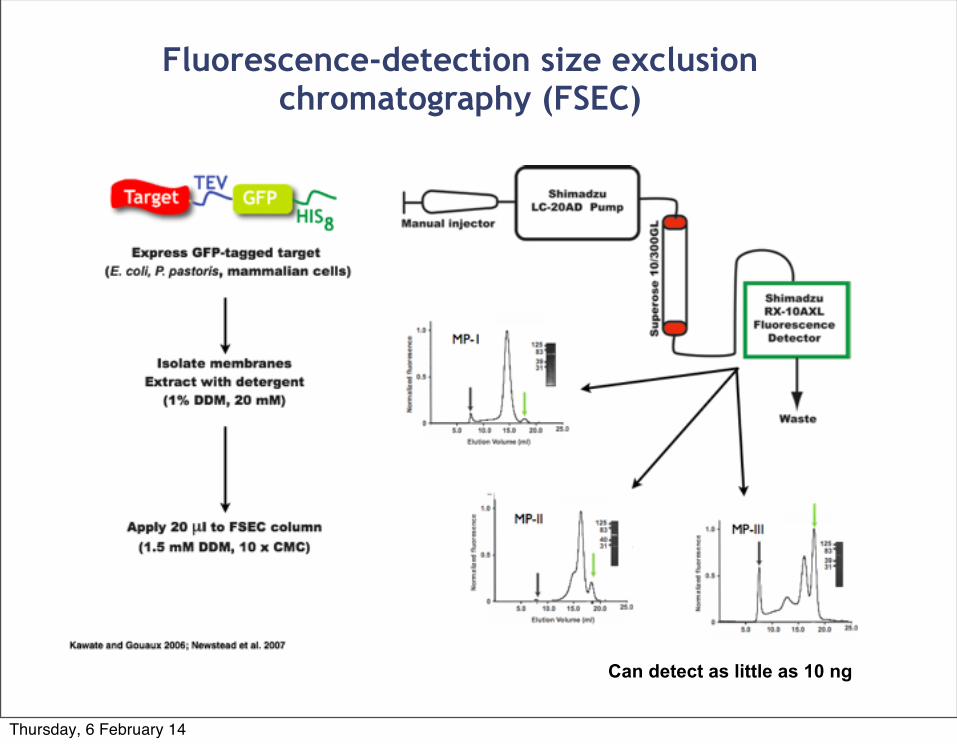
Fluorescence-detection size exclusion chromatography (FSEC)
Can detect as little as 10 ng
Thursday, 6 February 14
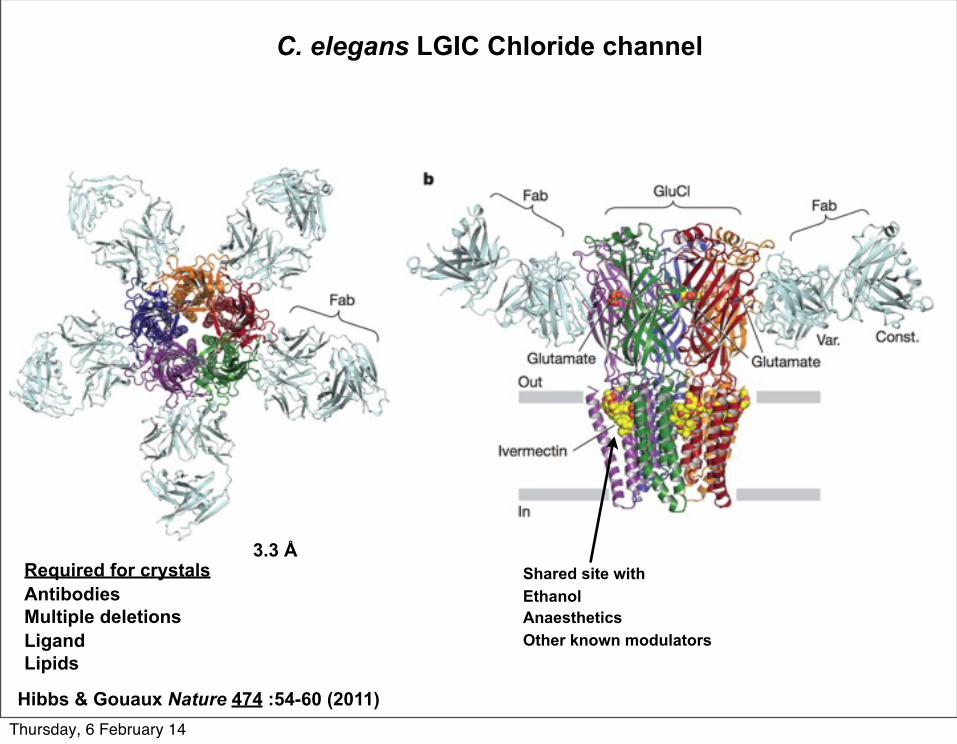
C. elegans LGIC Chloride channel
Required for crystalsAntibodiesMultiple deletionsLigandLipids
Hibbs & Gouaux Nature 474 :54-60 (2011)
3.3 ÅShared site withEthanolAnaestheticsOther known modulators
Thursday, 6 February 14
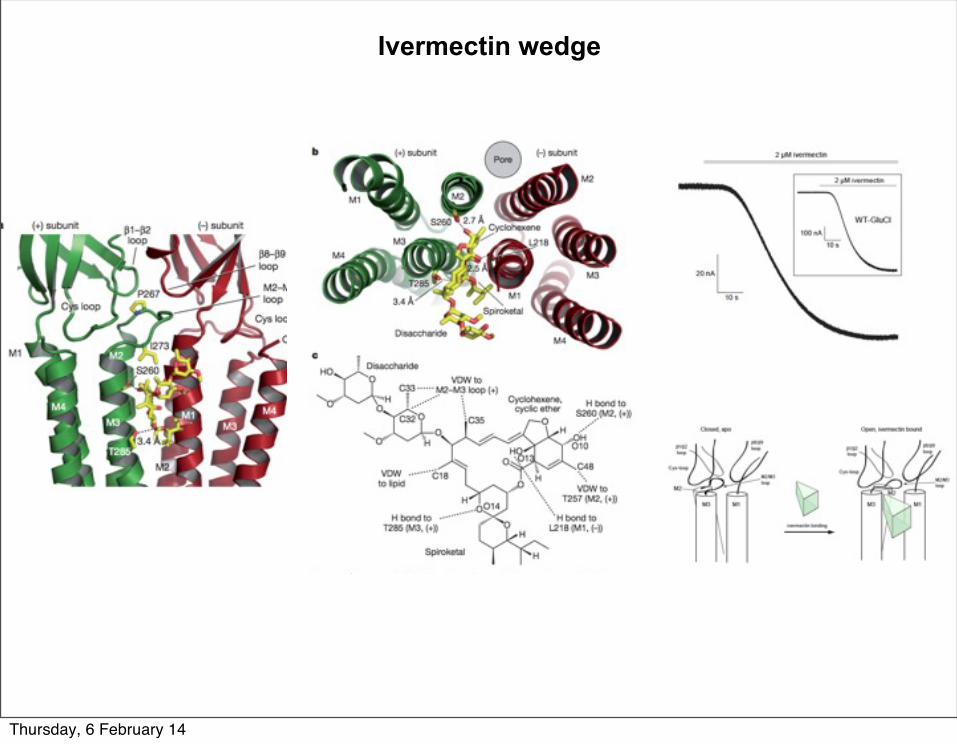
Ivermectin wedge
Thursday, 6 February 14
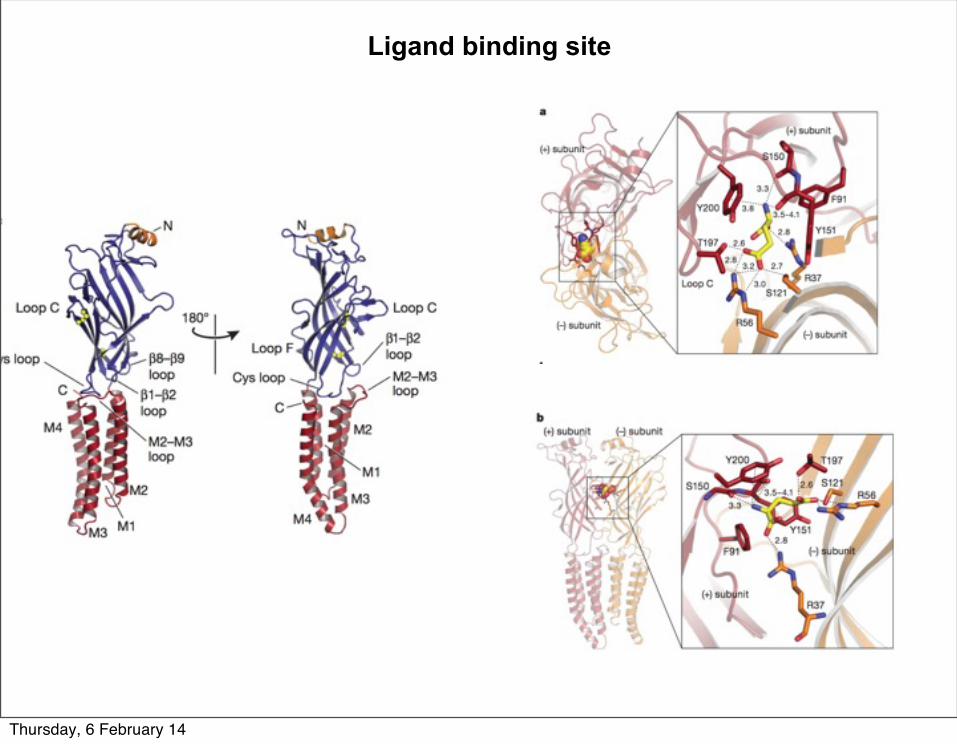
Ligand binding site
Thursday, 6 February 14
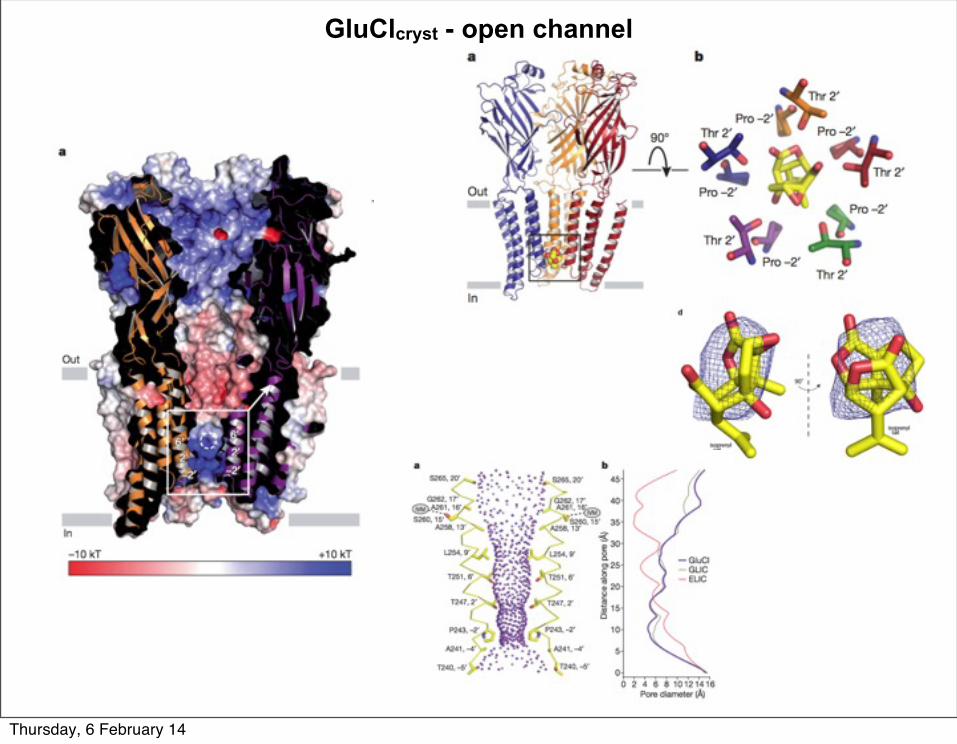
GluClcryst - open channel
Thursday, 6 February 14


















