Single molecule detection of PARP1 and PARP2 interaction with ...

Cancer Therapeutics Insights
PARP1 Is Overexpressed in Nasopharyngeal Carcinoma andIts Inhibition Enhances Radiotherapy
Jeremy P.H. Chow1, Wing Yu Man1, Mao Mao1, Han Chen1, Florence Cheung2, John Nicholls3,Sai Wah Tsao4, Maria Li Lung5, and Randy Y.C. Poon1
AbstractNasopharyngeal carcinoma is a rare but highly invasive cancer. As options of agents for effective
combination chemoradiotherapy for advanced nasopharyngeal carcinoma are limited, novel therapeutic
approaches are desperately needed. The ubiquitin ligase CHFR is known to target PARP1 for degradation
and is epigenetically inactivated in nasopharyngeal carcinoma. We present evidence that PARP1 protein is
indeed overexpressed in nasopharyngeal carcinoma cells in comparison with immortalized normal nasopha-
ryngeal epithelial cells. Tissuemicroarray analysis also indicated that PARP1protein is significantly elevated in
primary nasopharyngeal carcinoma tissues, with strong correlation with all stages of nasopharyngeal
carcinoma development. We found that the PARP inhibitor AZD2281 (olaparib) increased DNA damage,
cell-cycle arrest, and apoptosis in nasopharyngeal carcinoma cells challenged with ionizing radiation or
temozolomide. Isobologram analysis confirmed that the cytotoxicity triggered by AZD2281 and DNA-
damaging agents was synergistic. Finally, AZD2281 also enhanced the tumor-inhibitory effects of ionizing
radiation in animal xenograft models. These observations implicate that PARP1 overexpression is an early
event in nasopharyngeal carcinoma development and provide a molecular basis of using PARP inhibitors to
potentiate treatment of nasopharyngeal carcinoma with radio- and chemotherapy. Mol Cancer Ther; 12(11);
2517–28. �2013 AACR.
IntroductionNasopharyngeal carcinoma is a highly invasive can-
cer. Although nasopharyngeal carcinoma is relativelyrare in most parts of the world (annual incidence rategenerally less than 1 per 100,000 population), high inci-dence rates are found in southern China and SoutheastAsia (1). For example, nasopharyngeal carcinoma amongwomen and men in Hong Kong is 7 and 18 per 100,000,respectively (2). The prevailing view is that infection byEpstein–Barr virus (EBV) is a significant etiologic factorfor nasopharyngeal carcinoma (reviewed in ref. 3). Epi-demiologic evidence also links the consumption ofdietary
nitrosamines (such as salted fish) to nasopharyngeal car-cinoma (reviewed in ref. 4).
Radiotherapyhas beenusedas theprimary treatment ofnasopharyngeal carcinoma (5). This reflects both the inac-cessible location of the nasopharynx as well as the radio-sensitivity of nasopharyngeal carcinoma cells. Morerecent advances in radiotherapy such as three-dimension-al conformal radiotherapy and intension-modulatedradiotherapy have helped to achieve good prognosis, inparticular for early-stage nasopharyngeal carcinoma (6,7). For advanced nasopharyngeal carcinoma, however,treatment outcomes have been poor due to distant metas-tases and recurrence (8). Chemotherapy is used in com-bination with radiotherapy to control advanced nasopha-ryngeal carcinoma. Commonly used chemotherapeuticagents are restricted to standard compounds such as theDNA-damaging agent cisplatin and the thymidylatesynthase inhibitor 5-flurouracil. Effective agents for com-bination chemoradiotherapy for nasopharyngeal carcino-ma are still to be established.
One of the earliest responses to DNA damage is thebinding of PARP to damaged sites. PARP catalyzes thetransfer and polymerization of ADP ribose units fromNADþ to form branched polymers of ADP ribose cova-lently linked to heterologous acceptor proteins or PARPsthemselves (reviewed in ref. 9). PARP-catalyzed poly(ADP-ribosyl)ation plays pivotal roles in the processingand resolution of DNA breaks. The binding of PARP to
Authors' Affiliations: 1Division of Life Science, Center for CancerResearch, and State Key Laboratory of Molecular Neuroscience, The HongKongUniversity of Science andTechnology,ClearWaterBay; 2Departmentof Pathology, Pamela Youde Nethersole Eastern Hospital; Departments of3Pathology, 4Anatomy, and 5Clinical Oncology and Center for CancerResearch, The University of Hong Kong, Hong Kong
Note: Supplementary data for this article are available at Molecular CancerTherapeutics Online (http://mct.aacrjournals.org/).
J.P.H. Chow and W.Y. Man contributed equally to this work.
Corresponding Author: Randy Y.C. Poon, Division of Life Science, HongKong University of Science and Technology, Clear Water Bay, Hong Kong.Phone: 852-23588703; Fax: 852-23581552; E-mail: [email protected]
doi: 10.1158/1535-7163.MCT-13-0010
�2013 American Association for Cancer Research.
MolecularCancer
Therapeutics
www.aacrjournals.org 2517
on July 2, 2018. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 26, 2013; DOI: 10.1158/1535-7163.MCT-13-0010

the site of DNA damage activates its catalytic activityand promotes local poly(ADP-ribose)–dependent recruit-ment of DNA repair enzymes. Poly(ADP-ribosyl)ation ofhistones and other proteins may also mediate the decon-densation of chromatin structure required for efficientDNA repair. PARP1 is the most abundant and activeenzyme of the PARP family, but roles of other membersincluding PARP2 and PARP3 in DNA damage responsesare emerging (10).
PARP inhibitors have been evaluated in clinical trialseither as single agents or in combination with DNA-damaging agents (reviewed in ref. 11). As stand-aloneagents, PARP inhibitors have been used to induce syn-thetic lethality in homologous repair–deficient tumors.For example, PARP inhibitors are effective in targetingBRCA1- or BRCA2-deficient breast and ovarian cancer.The rationale of the approach is that inhibition of PARPblocks the repair of spontaneous single-strand lesions,leading to the formation of double-strand breaks throughmechanisms such as stalled replication forks. These dou-ble-strand breaks become lethal in cells that are deficientin double-strand break repair (such as those with muta-tions in BRCA1 or BRCA2). Synthetic lethality of PARPinhibitors is not limited to BRCA1 or BRCA2 deficiency.Recently, it was discovered that PARP inhibitors marked-ly sensitize Ewing’s sarcoma cells harboring the EWS-FLI1 gene translocation (12). More importantly, PARPinhibitors couldpotentially beusedas agents that enhancechemo- or radiotherapy-induced DNA damage inpatients without defined gene mutations. This remainsan emerging fieldwithmanyparameters to be deciphered(reviewed in ref. 11).
Although PARP1 has been implicated as a therapeutictarget in several types of cancer, its role has not beenrigorously investigated in nasopharyngeal carcinoma.Here, we present evidence that PARP1 is overexpressedin nasopharyngeal carcinoma in comparison with normalnasopharyngeal cells. Importantly, nasopharyngeal car-cinoma cell growthwas inhibited synergistically byDNA-damaging agents and an inhibitor of PARP.
Materials and MethodsMaterials
Olaparib (AZD2281; Selleck Chemicals) and temozolo-mide(Sigma-Aldrich) were obtained from the indicatedsuppliers.
Cell cultureIMR90 (normal fibroblasts) were obtained from the
American Type Culture Collection. Nasopharyngeal car-cinoma cell lines C666-1 (13), CNE2 (14), HNE1 (15), andHONE1 (15) were obtained from nasopharyngeal carci-noma AoE Cell Line Repository and were propagated inRPMI-1640 (for C666-1) or Dulbecco’s Modified EagleMedium (DMEM; for other nasopharyngeal carcinomacells) supplemented with 10% (v/v) FBS (Life Technolo-gies) and 50 U/mL penicillin–streptomycin (Life Tech-
nologies). No authentication was carried out by theauthors. Telomerase-immortalized nasopharyngeal epi-thelial cell lines NP361, NP460, and NP550 (16) werepropagated in keratinocyte serum-free medium supple-mented (LifeTechnologies)with 50%v/vEpiLife (Sigma).Cells were grown in humidified incubators at 37�C in 5%CO2.
Trypan blue analysis was conducted as described pre-viously (17). For clonogenic survival assays, 500 cellswereseeded onto 60-mm dishes. After 16 hours, the cells wereirradiated with different doses of ionizing radiation andeither mock-treated or exposed to the different concentra-tions of AZD2281 for another 24 hours. After 10 days,colonies were fixed with methanol/acetic acid (2:1 v:v)and visualized by staining with 2% (w/v) crystal violet in20% methanol.
Flow cytometryFlow cytometry analysis after propidium iodide stain-
ing was conducted as described previously (17).
Ionizing radiationIonizing radiation was delivered with a caesium137
source from a MDS Nordion Gammacell 1000 Elite Irra-diator. Unless stated otherwise, cells were irradiated witha dose of 15 Gy.
g-H2AX stainingCells grown on poly-L-lysine–treated coverslips were
fixed by ice-cold methanol for 10 minutes. The cells werethen washed twice with PBS, 5 minutes each, beforepermeabilized and blocked with 3% bovine serum albu-min (BSA) and0.2%Tween-20 inPBS at room temperaturefor 30 minutes. The cells were washed twice with washbuffer (0.2% Tween-20 in PBS) before incubated withphosphor-histone H2AXSer139 antibody (Bethyl Laborato-ry; diluted in 3% BSA, 0.2% Tween-20with PBS) at 4�C for16 hours. The cells were thenwashedwithwash buffer forfour times, 5 minutes each, and incubated in dark withAlexa Fluor 594 goat anti-rabbit immunoglobulin G (IgG)secondary antibodies (Life Technologies) at 25�C for 2hours. After washing four times with wash buffer for 5minutes each, the cellswere stainedwithHoechst 33342 (5mg/mL in wash buffer) for 5 minutes, washed three timeswith wash buffer, before mounted with 100 mmol/L N-propyl-gallate in 9:1 v/v glycerol:PBS.
Isobologram analysisThe combination effects of two treatments were ana-
lyzed according to themedian-effect method of Chou andTalalay (18). Each dose–response curve was used to cal-culate the isobologram, the value of median-effect dose(Dm), the sigmoidicity of thedose–effect curve (m), and thelinear correlation coefficient of the median-effect plot (r).The combination index (CI) at different effectivedose (ED)ED50, ED75, and ED90 was calculated according to Chouand Talalay (18) using CalcuSyn Version 2.1 (Biosoft). CIvalue < 1 indicates synergistic effect (0.1–0.5 strong
Chow et al.
Mol Cancer Ther; 12(11) November 2013 Molecular Cancer Therapeutics2518
on July 2, 2018. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 26, 2013; DOI: 10.1158/1535-7163.MCT-13-0010

synergism; <0.1 very strong synergism); CI value of 1indicates additive effect; and CI value > 1 indicates antag-onistic effect.
Real-time PCRTotal RNA was extracted from approximately 3 � 106
cells using Nucleospin RNA (Macherey-Nagel). The totalRNA (1 mg) was used to synthesize cDNA using theHigh-Capacity cDNA Reverse Transcription Kit (LifeTechnologies) according to the manufacturer’s instruc-tion. Semiquantitative real-time PCR was conducted bymixing the cDNA with SYBR Green Real-Time PCRMaster Mixes in a 7500 Fast Real-Time PCR System (LifeTechnologies) using the following primers for actin (50-GGGAAATCGTGCGTGACATT-30; 50-GGAACCGCTC-ATTGCCAAT-30), CHFR (50-GGCAACCAGAGGTTTG-ACAT-30; 50-AGTCAGGACGGGATGTTACG-30), andPARP1 (50-GATGGGTTCTCTGAGCTTCG-30; 50-ACAC-CCCTTGCACGTACTTC-30).
Antibodies and immunoblottingAntibodies against b-actin (Sigma-Aldrich), phosphor-
histone H2AXSer139 (Bethyl Laboratory), cleaved PARP1(Asp214; BD Biosciences), and PARP1 (sc-7150; SantaCruz Biotechnology) were obtained from the indicatedsuppliers. Rabbit antibodies against CHFR were raisedagainst a bacterially expressed GST-CHFR(CD427) pro-tein. Immunoblotting was carried out as described pre-viously (19).
Nasopharyngeal carcinoma tissue microarrayanalysisA tissue microarray containing normal nasopharyn-
geal epithelium and nasopharyngeal carcinoma speci-mens was constructed. The nasopharyngeal carcinomaspecimens were obtained from patients with naso-pharyngeal carcinoma before treatment at QueenMary Hospital (Hong Kong) and Pamela Youde Neth-ersole Eastern Hospital (Hong Kong). Constructionof the nasopharyngeal carcinoma tissue microarrayand immunohistochemical staining using the standardstreptavidin–biotin–peroxidase complex method wassimilar to that previously reported (20). Slides wereincubated with antibodies against PARP1 at 1:200 dilu-tion and counterstained with hematoxylin. The immu-nohistochemically stained tissue microarray wasreviewed by two independent pathologists. For eval-uation of the staining, the nonmalignant and malignanttissues were scored by assessing the PARP1 staining inthe nucleus. A random four views were counted under�400 microscopy. The intensity of staining was gradedby an arbitrary scale that ranged from 0 to 3, represent-ing negative ("0"), weak ("1"), moderate ("2"), andstrong ("3") staining, respectively. Staining values of0 and 1 were classified as low expression, whereas 2and 3 were classified as high expression. The Pearsonx2 test was used to assess the association betweenPARP1 expression and several clinicopathologic vari-
ables. A P value less than 0.05 was regarded as statis-tically significant.
Tumor xenograftsThe experimental protocol was evaluated and
approved by the Animal Care Committee, Hong KongUniversity of Science and Technology (Clear Water Bay,Hong Kong). HONE1 cells (2 � 107) were injected sub-cutaneously into both sides of thedorsa of 4- to 6-week-oldfemale BALB/cnudemice. Four animals (n¼ 8) per groupwere used in control and combination treatment and 3animals were used in the single treatments. Tumors wereregularly measured using a Vernier caliper. Volume wascalculated according to the formula: p/6 � length �width2. Day ¼ 0 was designated as when the tumorvolume was approximately 100 mm3. AZD2281 (50mg/kg) was administrated with intraperitoneal injectioninto the mice daily from day 1 to 5. Ionizing radiation (2Gy) was delivered at day 1 and 4. In the combinationtreatment, AZD2281was administrated 30minutes beforeionizing radiation delivery. Mice were killed when thetumors in the control group reached 1,000 mm3.
ResultsOverexpression of PARP1 protein in nasopharyngealcarcinoma cell lines
Frequent alterations of gene expression have beenreported in nasopharyngeal carcinoma. However, theirimplication in tumorigenesis or treatment is not alwaysimmediately apparent. Among these, transcription of theubiquitin ligase CHFR was found to be epigeneticallyinactivated in nasopharyngeal carcinoma through pro-moter methylation (21). To study whether CHFR proteinis altered in nasopharyngeal carcinoma cells, antibodiesagainst CHFR were generated. Lysates from differentnasopharyngeal carcinoma cell lines (C666-1, CNE2,HNE1, and HONE1) were prepared and analyzed withimmunoblotting using antibodies specific for CHFR.Nor-mal nasopharyngeal epithelial cells that were immortal-ized with telomerase were used as a comparison. CHFRwas found to be downregulated in all the nasopharyngealcarcinoma cell lines (Fig. 1A). In agreement with this,CHFR was also present at a relatively high level in threedifferent immortalized nasopharyngeal epithelial celllines (NP361, NP460, and NP550; Fig. 1B). As a control,the normal human fibroblasts strain (IMR90) also con-tained a high abundance of CHFR.
Recently, CHFR was reported to be able to regulatePARP1 by targeting PARP1 for ubiquitination and deg-radation (22).We also found that PARP1 expression couldbe reduced by overexpression of CHFR and increasedby knockdown of CHFR (our unpublished data). Thisprompted us to examine the expression of PARP1 innasopharyngeal carcinoma cells. Significantly, PARP1displayed an inverse relationship with CHFR, expressingat high levels in all the nasopharyngeal carcinoma celllines and at low levels in the nasopharyngeal epithelial
PARP in Nasopharyngeal Carcinoma
www.aacrjournals.org Mol Cancer Ther; 12(11) November 2013 2519
on July 2, 2018. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 26, 2013; DOI: 10.1158/1535-7163.MCT-13-0010
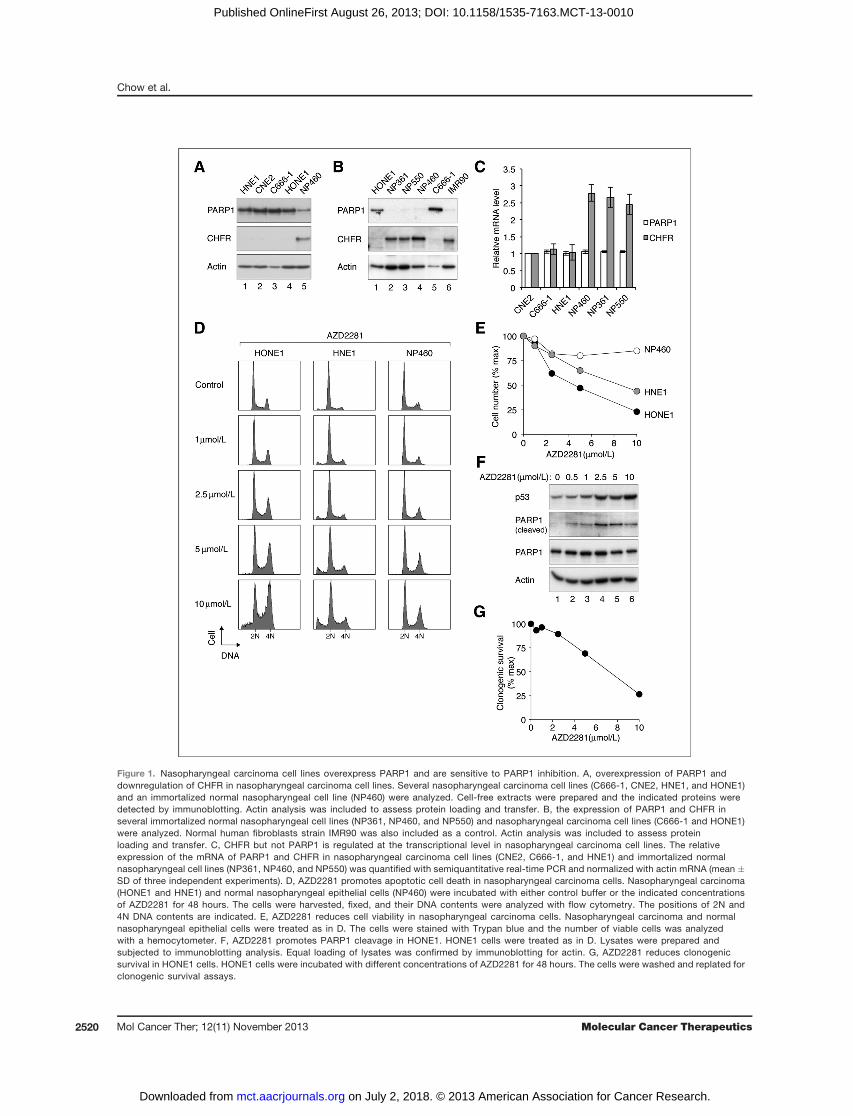
Figure 1. Nasopharyngeal carcinoma cell lines overexpress PARP1 and are sensitive to PARP1 inhibition. A, overexpression of PARP1 anddownregulation of CHFR in nasopharyngeal carcinoma cell lines. Several nasopharyngeal carcinoma cell lines (C666-1, CNE2, HNE1, and HONE1)and an immortalized normal nasopharyngeal cell line (NP460) were analyzed. Cell-free extracts were prepared and the indicated proteins weredetected by immunoblotting. Actin analysis was included to assess protein loading and transfer. B, the expression of PARP1 and CHFR inseveral immortalized normal nasopharyngeal cell lines (NP361, NP460, and NP550) and nasopharyngeal carcinoma cell lines (C666-1 and HONE1)were analyzed. Normal human fibroblasts strain IMR90 was also included as a control. Actin analysis was included to assess proteinloading and transfer. C, CHFR but not PARP1 is regulated at the transcriptional level in nasopharyngeal carcinoma cell lines. The relativeexpression of the mRNA of PARP1 and CHFR in nasopharyngeal carcinoma cell lines (CNE2, C666-1, and HNE1) and immortalized normalnasopharyngeal cell lines (NP361, NP460, and NP550) was quantified with semiquantitative real-time PCR and normalized with actin mRNA (mean �SD of three independent experiments). D, AZD2281 promotes apoptotic cell death in nasopharyngeal carcinoma cells. Nasopharyngeal carcinoma(HONE1 and HNE1) and normal nasopharyngeal epithelial cells (NP460) were incubated with either control buffer or the indicated concentrationsof AZD2281 for 48 hours. The cells were harvested, fixed, and their DNA contents were analyzed with flow cytometry. The positions of 2N and4N DNA contents are indicated. E, AZD2281 reduces cell viability in nasopharyngeal carcinoma cells. Nasopharyngeal carcinoma and normalnasopharyngeal epithelial cells were treated as in D. The cells were stained with Trypan blue and the number of viable cells was analyzedwith a hemocytometer. F, AZD2281 promotes PARP1 cleavage in HONE1. HONE1 cells were treated as in D. Lysates were prepared andsubjected to immunoblotting analysis. Equal loading of lysates was confirmed by immunoblotting for actin. G, AZD2281 reduces clonogenicsurvival in HONE1 cells. HONE1 cells were incubated with different concentrations of AZD2281 for 48 hours. The cells were washed and replated forclonogenic survival assays.
Chow et al.
Mol Cancer Ther; 12(11) November 2013 Molecular Cancer Therapeutics2520
on July 2, 2018. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 26, 2013; DOI: 10.1158/1535-7163.MCT-13-0010

cell lines (Fig. 1A and B). As expected, CHFR mRNAwaspresent at a lower level in nasopharyngeal carcinomacell lines than in the nasopharyngeal epithelial cell lines(Fig. 1C). Consistent with regulation by posttranslationalmechanisms,we found that PARP1mRNAwas expressedat similar levels between nasopharyngeal carcinoma andnasopharyngeal cell lines. Taken together, these resultsindicate that PARP1 is present at a significantly higherlevel in nasopharyngeal carcinoma cell lines than in nor-mal nasopharyngeal epithelial cells.
InhibitionofPARPenhances the cytotoxicity inducedby DNA-damaging agents in nasopharyngealcarcinoma cellsGiven that PARP1 is overexpressed in nasopharyngeal
carcinoma cells, we next addressed whether this trans-lates into alterations in the sensitivity to PARP inhibitors.Olaparib (AZD2281; ref. 23) is an orally active PARPinhibitor undergoing phase I and II clinical trials (11).Incubation of nasopharyngeal carcinoma and normalnasopharyngeal epithelial cells with AZD2281 aloneinduced a stress response. In the nasopharyngeal carci-noma cells HONE1 and HNE1, a relatively high concen-tration of AZD2281 (10 mmol/L) induced apoptotic celldeath (as indicated by the appearance of sub-G1 cells; Fig.1D). An increase in 4N DNA contents was also apparent,in particular forHONE1, suggesting a robust activation ofthe G2 DNA damage checkpoint. In contrast, no sub-G1
cells were detected when normal nasopharyngeal epithe-lial cells were challenged with the same concentrations ofAZD2281. In agreement with this, Trypan blue analysisindicated that a dose-dependent reduction of cell numberfor nasopharyngeal carcinoma cells (Fig. 1E). In contrast,only amarginal decrease in the number of viable cells wasobserved after normal nasopharyngeal epithelial cellswere incubated with AZD2281. Consistent with anincrease in apoptotic cell death in nasopharyngeal carci-noma cells, we found that AZD2281 stimulated an accu-mulation of cleaved PARP1 and p53 in a dose-dependentmanner (Fig. 1F). The long-term clonogenic survival ofHONE1 cells was likewise reduced by AZD2281 treat-ment (Fig. 1G).We next examined whether inhibition of PARP could
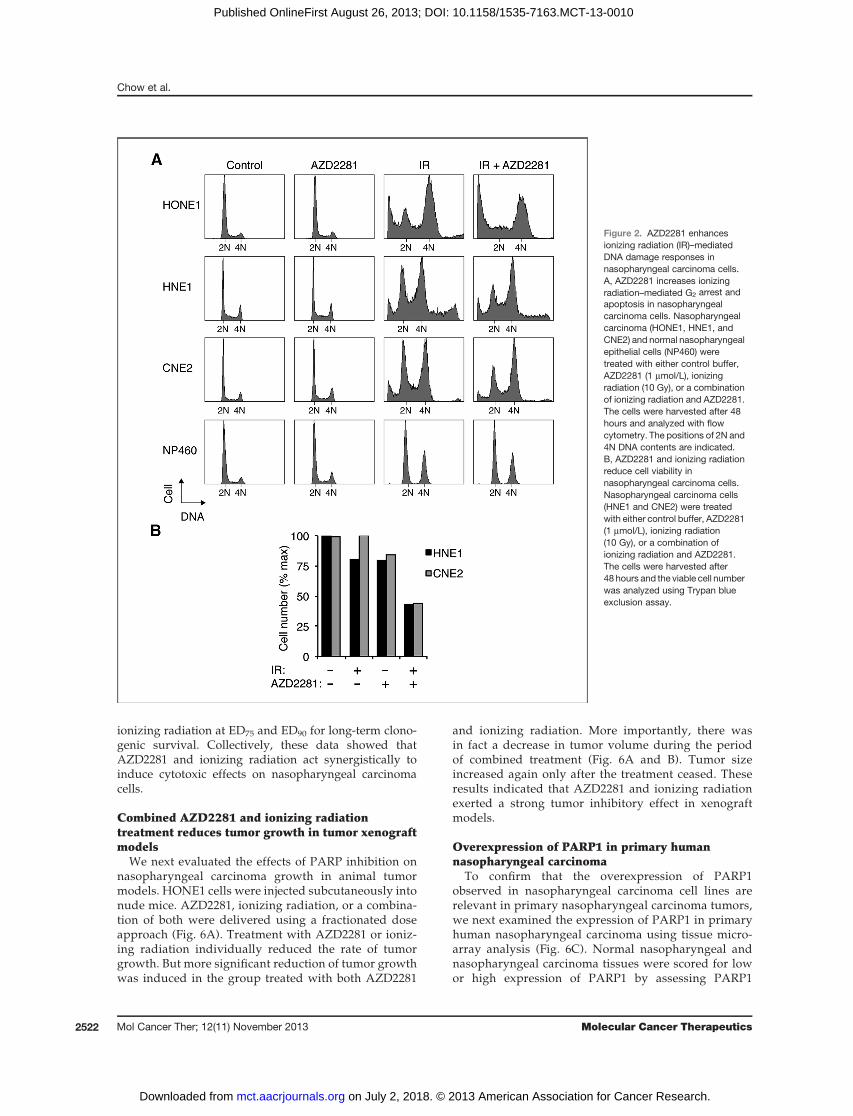
potentiate the cytotoxic effects of DNA-damaging agents.Apartial G2 arrestwas stimulated in both nasopharyngealcarcinoma and nasopharyngeal epithelial cells after irra-diated with 10 Gy of ionizing radiation (Fig. 2A). Signif-icantly, more nasopharyngeal carcinoma cells werearrested in G2 when they were irradiated in the presenceof 1 mmol/L of AZD2281. In marked contrast, normalnasopharyngeal epithelial cells seemed to be imperviousto AZD2281. As a control, exposure of nasopharyngealcarcinoma cells to AZD2281 alone did not affect normalcell-cycle progression (see also Fig. 1D). Moreover, anextensive sub-G1 population was induced when HONE1were treated with both ionizing radiation and AZD2281.Although sub-G1 population was not apparent for HNE1andCNE2 cells, Trypanblue exclusion analysis confirmed
that substantial cell death was triggered when the cellswere exposed to both ionizing radiation and AZD2281(Fig. 2B).
To ensure that the stimulation of nasopharyngealcarcinoma cytotoxicity by AZD2281 was not limitedto ionizing radiation, cells were also treated with theDNA-methylating agent temozolomide. Incubationwith either AZD2281 or temozolomide (125 mmol/L)alone did not significantly stimulate apoptosis. Massiveapoptosis was triggered when nasopharyngeal car-cinoma cells were incubated with both temozolomideand AZD2281 together (Fig. 3A). Although normalnasopharyngeal epithelial cells also responded to thecombined AZD2281 and temozolomide treatment bydisplaying more G2-arrested cells, there was no increasein sub-G1 population. Consistent with the results fromthe flow cytometry, high level of g-H2AX was inducedafter incubation with AZD2281 and temozolomide inHONE1, but not in NP460 (Fig. 3B).
To see whether the increase in DNAdamage–mediatedarrest and cell death in AZD2281-treated cells was causedby defective DNA repair, we next examined g-H2AXstaining in the presence or absence of AZD2281 (Fig.4A). As expected, irradiation with 5 Gy of ionizing radi-ation induced g-H2AX foci formation in both nasopha-ryngeal carcinoma (HONE1) and nasopharyngeal epithe-lial (NP460) cells. The g-H2AX foci staining returned to abackground level after 24 hours, suggesting that thedamaged DNA was repaired in both nasopharyngealcarcinoma and nasopharyngeal cells (Fig. 4B). Signi-ficantly, efficient DNA repair in HONE1 cells was atten-uated in the presence of AZD2281. In contrast, the DNArepair in NP460 cells was resistant to AZD2281.
Collectively, these results indicate that inhibition ofPARP enhances ionizing radiation- or temozolomide-mediatedDNAdamage and apoptosis in nasopharyngealcarcinoma cells but not in normal nasopharyngeal epi-thelial cells.
PARP inhibition and ionizing radiation actsynergistically to promote cytotoxicity
To investigate more rigorously whether the effects ofPARP inhibition and DNA damage on nasopharyngealcarcinoma cells represent synergism, isobologram analy-sis was carried out according to themedian-effectmethodof Chou and Talalay (18). Using Trypan blue exclusionassays to monitor cytotoxicity, we found that AZD2281and ionizing radiation acted synergistically on nasopha-ryngeal carcinoma cells (HONE1 and HNE1; Fig. 5Aand Table 1). The CI was less than 1, which indicatedsynergism, over a range of effective dose (ED50, ED75, andED90). In contrast, synergism between AZD2281 and ion-izing radiation were less effective on nasopharyngealepithelial cells (NP460), with the CI values actuallyexceeding 1 at ED90 (indicating antagonism). We alsoexamined the long-term survival of HONE1 cells usingclonogenic survival assays (Fig. 5B and Table 2). Similaranalysis indicated a synergism between AZD2281 and
PARP in Nasopharyngeal Carcinoma
www.aacrjournals.org Mol Cancer Ther; 12(11) November 2013 2521
on July 2, 2018. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 26, 2013; DOI: 10.1158/1535-7163.MCT-13-0010
ionizing radiation at ED75 and ED90 for long-term clono-genic survival. Collectively, these data showed thatAZD2281 and ionizing radiation act synergistically toinduce cytotoxic effects on nasopharyngeal carcinomacells.
Combined AZD2281 and ionizing radiationtreatment reduces tumor growth in tumor xenograftmodels
We next evaluated the effects of PARP inhibition onnasopharyngeal carcinoma growth in animal tumormodels. HONE1 cells were injected subcutaneously intonude mice. AZD2281, ionizing radiation, or a combina-tion of both were delivered using a fractionated doseapproach (Fig. 6A). Treatment with AZD2281 or ioniz-ing radiation individually reduced the rate of tumorgrowth. But more significant reduction of tumor growthwas induced in the group treated with both AZD2281
and ionizing radiation. More importantly, there wasin fact a decrease in tumor volume during the periodof combined treatment (Fig. 6A and B). Tumor sizeincreased again only after the treatment ceased. Theseresults indicated that AZD2281 and ionizing radiationexerted a strong tumor inhibitory effect in xenograftmodels.
Overexpression of PARP1 in primary humannasopharyngeal carcinoma
To confirm that the overexpression of PARP1observed in nasopharyngeal carcinoma cell lines arerelevant in primary nasopharyngeal carcinoma tumors,we next examined the expression of PARP1 in primaryhuman nasopharyngeal carcinoma using tissue micro-array analysis (Fig. 6C). Normal nasopharyngeal andnasopharyngeal carcinoma tissues were scored for lowor high expression of PARP1 by assessing PARP1
Figure 2. AZD2281 enhancesionizing radiation (IR)–mediatedDNA damage responses innasopharyngeal carcinoma cells.A, AZD2281 increases ionizingradiation–mediated G2 arrest andapoptosis in nasopharyngealcarcinoma cells. Nasopharyngealcarcinoma (HONE1, HNE1, andCNE2) and normal nasopharyngealepithelial cells (NP460) weretreated with either control buffer,AZD2281 (1 mmol/L), ionizingradiation (10 Gy), or a combinationof ionizing radiation and AZD2281.The cells were harvested after 48hours and analyzed with flowcytometry. The positions of 2N and4N DNA contents are indicated.B, AZD2281 and ionizing radiationreduce cell viability innasopharyngeal carcinoma cells.Nasopharyngeal carcinoma cells(HNE1 and CNE2) were treatedwith either control buffer, AZD2281(1 mmol/L), ionizing radiation(10 Gy), or a combination ofionizing radiation and AZD2281.The cells were harvested after48 hours and the viable cell numberwas analyzed using Trypan blueexclusion assay.
Chow et al.
Mol Cancer Ther; 12(11) November 2013 Molecular Cancer Therapeutics2522
on July 2, 2018. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 26, 2013; DOI: 10.1158/1535-7163.MCT-13-0010

staining in the nucleus (Table 3). High expression ofPARP1 was found in nasopharyngeal carcinoma tissues.Correlation was observed between PARP1 expressionand several clinicopathologic parameters, includingpatient age and survival. Importantly, an increase inPARP1 expression could be observed in all nasopha-ryngeal carcinoma stages, suggesting that the alterationof PARP1 expression is an early event in nasopharyn-geal carcinoma development.
DiscussionIn this study, we found that PARP1 protein is highly
expressed in nasopharyngeal carcinoma cell lines andprimary nasopharyngeal carcinoma tissues in compari-
sonwith nasopharyngeal epithelial cells. The PARP inhib-itor AZD2281 increased DNA damage, cell-cycle arrest,and apoptosis in nasopharyngeal carcinoma cell linestreated with ionizing radiation or temozolomide. Finally,AZD2281 also enhanced the tumor-inhibitory effects ofionizing radiation in mouse xenograft models.
A major focus of the clinical development of PARPinhibitors is to determine whether they can potentiateradio-/chemotherapy-mediated DNA damage, espe-cially for tumors that lack intrinsic defects in DNAdamage repair (reviewed in ref. 11). For nasopharyngealcarcinoma, we found that PARP inhibition was indeedeffective in potentiating the cytotoxicity of DNA-dam-aging agents using both cell line (Figs. 2 and 3) andanimal models (Fig. 6). Isobologram analysis indicated
Figure 3. AZD2281 enhancestemozolomide (TMZ)-mediatedDNA damage responses innasopharyngeal carcinomacells. A, AZD2281 increasestemozolomide-mediated G2 arrestand apoptosis in nasopharyngealcarcinoma cells. Nasopharyngealcarcinoma (HONE1 and C666-1)and normal nasopharyngealepithelial cells (NP460 and NP361)were incubated with either controlbuffer, AZD2281 (1 mmol/L),temozolomide (125 mmol/L), or acombination of temozolomide andAZD2281. Thecellswereharvestedafter 48 hours and analyzed withflowcytometry. The positions of 2Nand 4N DNA contents areindicated. B, AZD2281 increasestemozolomide-mediated DNAdamage in nasopharyngealcarcinoma cells. Nasopharyngealcarcinoma cells (HNE1 and CNE2)were incubated with either controlbuffer, AZD2281 (1 mmol/L),temozolomide (125 mmol/L), or acombination of temozolomide andAZD2281 for 48 hours. Lysateswere prepared and analyzed withimmunoblotting. Equal loading oflysates was confirmed byimmunoblotting for actin.
PARP in Nasopharyngeal Carcinoma
www.aacrjournals.org Mol Cancer Ther; 12(11) November 2013 2523
on July 2, 2018. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 26, 2013; DOI: 10.1158/1535-7163.MCT-13-0010

that at least for the cell line models, the effects ofAZD2281 and ionizing radiation were synergistic ratherthan simply additive (Fig. 5). AZD2281 and ionizingradiation displayed synergism on nasopharyngeal car-cinoma cells over the range of effective doses in bothshort-term growth assay (Fig. 5A and Table 1) and long-term cell survival (Fig. 5B and Table 2). Results from thexenograft studies further indicated that combinationaltreatment of AZD2281 and ionizing radiation couldfurther lead to shrinkage of tumor size (Fig. 6). How-ever, the treatment also has synergistic effects on nor-mal nasopharyngeal cells (for ED50 and ED75; Fig. 5Aand Table 1). Moreover, synergism was found at alleffective doses for long-term nasopharyngeal carcinomacell growth (Fig. 5B and Table 2). Furthermore, thecombined AZD2281 and ionizing radiation treatmentresulted in a approximately 15% weight loss of theanimals (Supplementary Fig. S1), suggesting potentialhigh toxicity to normal cells.
Hence, although our findings provide a rationale tosupport testing of PARP inhibitors and DNA-damagingagents on nasopharyngeal carcinoma, further research isrequired to uncover the precise clinical responses of thesetreatments.
Conceptually, PARP inhibition should only enhanceradio-/chemotherapy if DNA damage is selectivelyincreased in tumor compared with normal tissues.Tumors that already harbor DNA repair defects such asmutations in BRCA1 or BRCA2 genes serve as classicexamples. So far, there is no evidence of impairment ofthe components of the DNA repair pathways such asBRCA2 in nasopharyngeal carcinoma (24, 25). As impli-cated by our study, however, the abundance of PARP1 intumors may also play a role in selectively sensitizing thecells to PARP inhibitors. Nasopharyngeal carcinoma celllines contained significantly more PARP1 than nasopha-ryngeal epithelial cells (Fig. 1A and B) and were moresensitive to AZD2281 alone (Fig. 1D and E) or whenAZD2281 was added together with DNA-damagingagents (Figs. 2 and 3).
The observations that PARP1 was overexpressed evenat early stages of nasopharyngeal carcinomadevelopment(Table 3) imply that alteration of PARP1 could be a driverof tumorigenesis. Consistent with our observations, over-expression of PARP1 has also been reported in othertumors, including colorectal cancer (26), endometrial can-cer (27), hepatocellular carcinoma (28), and melanomas(27). Several studies have reported the association ofPARP1 protein expression with a worse prognosis. Forexample, PARP1 overexpression is associated with over-all poor prognosis, higher tumor grade, BRCA1-mutatedand triple-negative phenotype breast cancers (29–31). Innasopharyngeal carcinoma, PARP1 expression was cor-related with increase in patient age and poorer survival(Table 3).
What are the mechanisms that lead to PARP1 over-expression in nasopharyngeal carcinoma and othercancers? Transcription of the PARP1 gene relies on
Figure 4. Inhibition of PARP disrupts DNA damage repair innasopharyngeal carcinoma cells. A, AZD2281 inhibits repair of ionizingradiation (IR)–mediated DNA damage in nasopharyngeal carcinoma butnot in normal nasopharyngeal epithelial cells.Nasopharyngeal carcinoma(HONE1) andnasopharyngeal epithelial cells (NP460)were irradiatedwithionizing radiation (5 Gy), either in the absence or presence of AZD2281(1 mmol/L). The cells were fixed at either 1 or 24 hours after irradiation(cells treated with AZD2281 alone were also harvested after 24 hours).DNA damage and recovery were assessed by immunostainingwith an antibody against g-H2AX. Nuclei were identified by staining withHoechst 33342 dye. Representative images are shown of the varioustreatments. B, quantification of g-H2AX foci in the presence or absenceof AZD2281. Cells were treated as described in A. The percentageof cells positive for g-H2AXwas quantified (n¼ 75). Cells containingmorethan five g-H2AX foci in the nucleus were considered as positive. Mean�SD of three independent experiments.
Chow et al.
Mol Cancer Ther; 12(11) November 2013 Molecular Cancer Therapeutics2524
on July 2, 2018. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 26, 2013; DOI: 10.1158/1535-7163.MCT-13-0010

transcription factors such as Sp1, Sp3 (32), and NF1(33). However, the increase in PARP1 protein in naso-pharyngeal carcinoma was unlikely to be due to anincrease in transcription because no significant variationin PARP1 mRNA between nasopharyngeal carcinomaand nasopharyngeal epithelial cell lines was observed
(Fig. 1C). This is also consistent with the lack of reporton an alteration ofPARP1mRNAexpression fromvariousmicroarray studies. Instead, we believe that the proteinlevel of PARP1 is usually kept at a low level by CHFRin normal nasopharyngeal cells (22). The epigenetic in-activation of CHFR in nasopharyngeal carcinoma (21)provides a molecular basis of how PARP1 is upregulatedat the protein level (see Fig. 6D for a model). In thisconnection, it is possible that methylation of the CHFRpromoter can be used as a marker for PAPR1 expression,which in turn can be used as a prediction for sensitivityto PARP inhibitors.
An interesting question is why overexpression ofPARP1 would lead to an increase in sensitivity to PARPinhibition. We believe that "PARP1 addiction" may play arole on the reliance of nasopharyngeal carcinoma cells onPARP1 for DNA damage responses. As the DNA repaircircuitry gets rewired by the constitutive overexpressionof PARP1, the cell may become exquisitely dependent onPARP1 activity. Nasopharyngeal carcinoma cells wereclearly more dependent on PARP activity for effectiverepair than nasopharyngeal epithelial cells (Fig. 4). ThePARP1 addiction may be analogous to the addiction tomany oncogenes in cancer cells. Another possible reasonwhy PARP1-overexpressing cells were sensitive to PARPinhibitor is the recent finding that the cytotoxicity of PARP
Figure 5. AZD2281 and ionizingradiation (IR) act synergistically toinduce cytotoxicity innasopharyngeal carcinoma cells.A, AZD2281 and ionizing radiationact synergistically onnasopharyngeal carcinoma cellsbut not NP460 in reducing cellnumber. Nasopharyngealcarcinoma (HONE1 and HNE1) andnasopharyngeal epithelial cells(NP460) were treated with ionizingradiation and AZD2281 for 48hours and analyzed with Trypanblue analysis. Isobologram plots ofED90 are shown. Summary of theCIis shown in Table 1. B, AZD2281and ionizing radiation actsynergistically on clonogenicsurvival of nasopharyngealcarcinoma cells. HONE1 cells weretreated with ionizing radiation andAZD2281 and analyzed withclonogenic survival assays.Isobologramplots of effective doseED50, ED75, and ED90 are shown.Summary of the CI is shownin Table 2.
Table 1. AZD2281 and ionizing radiation actsynergistically to induce cytotoxicity innasopharyngeal carcinoma cells
CI
ED50 ED75 ED90 Dm m r
HONE1 0.510 0.633 0.786 1.037 0.940 0.998HNE1 0.199 0.181 0.165 0.707 0.764 0.961NP460 0.468 0.740 1.372 0.860 0.538 0.943
NOTE: Nasopharyngeal carcinoma (HONE1 and HNE1) andnasopharyngeal epithelial cells (NP460) were treated withionizing radiation and AZD2281 for 48 hours and analyzedwith Trypan blue analysis. The CI at ED50, ED75, and ED90
was calculated. The value of Dm, m, and r are indicated. ACI < 1 indicates synergism.
PARP in Nasopharyngeal Carcinoma
www.aacrjournals.org Mol Cancer Ther; 12(11) November 2013 2525
on July 2, 2018. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 26, 2013; DOI: 10.1158/1535-7163.MCT-13-0010

inhibitors including AZD2281 is in part caused the trap-ping of PARP1 at damaged DNA complexes (34). Hence,higher expression of PARP1 may allow larger number ofPARP1–DNA complexes to form.
Results from this study introduced further questionsabout PARP1 and nasopharyngeal carcinoma. PARP inhi-bitors in development mimic the nicotinamide moiety ofNADþ and bind PARP’s catalytic domain. In addition toAZD2281, several PARP inhibitors are in thedevelopmentpipeline. It would be interesting to see whether theyoffer better efficacy when combined with ionizing radia-tion in treatment of nasopharyngeal carcinoma. AlthoughPARP1 is believed to be the most abundant isoform of thePARP family, it is possible that other familymembersmayalso play a role in nasopharyngeal carcinoma. Inhibitorsthat target specific isoforms may be useful. Eventually,randomized studies will be needed to validate PARPinhibitors as therapeutic agents for patients with naso-pharyngeal carcinoma. Another unresolved issue isalthough we know that PARP1 is overexpressed in naso-pharyngeal carcinoma and other cancers, little is knownabout its activity. It would be interesting to assay PARP1activity in cell lines and tumor specimens from patients
Figure 6. AZD2281 and ionizing radiation (IR) inhibit tumor growth in mouse xenograft models. A, HONE1 cells were injected subcutaneously into nude mice.AZD2281 (closed arrowhead) and ionizing radiation (open arrowhead) were delivered at the indicated time points as described in Materials and Methods.The volume of the tumor was measured on different days (control, white square, n ¼ 8; AZD2281, white circle, n ¼ 6; ionizing radiation, gray circle, n ¼ 6;ionizing radiation and AZD2281, dark circle, n ¼ 8; mean � SD). B, representative mice with different treatments at day 28 are shown. C, PARP1 isoverexpressed in human primary nasopharyngeal carcinoma (NPC). The expression of PARP1 in normal nasopharyngeal and nasopharyngealcarcinoma tissueswas analyzed using tissuemicroarray and immunohistochemistry. Representative images of PARP1 and hematoxylin and eosin staining innormal nasopharyngeal and nasopharyngeal carcinoma tissues are shown. D, model of the roles of CHFR and PARP1 in nasopharyngeal carcinoma. Innasopharyngeal carcinoma, CHFR is epigenetically inactivated by promoter hypermethylation. The decrease in CHFR protein leads to a stabilization ofPARP1protein. As PARP1 is increased in early stages of nasopharyngeal carcinoma, the sustained increase inPARP1may contribute to tumor progression ofthis cancer. Moreover, PARP1 overexpression may lead to "PARP1 addiction," in which DNA repair becomes more reliant on PARP1 than in normal cells.
Table 2. AZD2281 and ionizing radiation actsynergistically on clonogenic survival ofnasopharyngeal carcinoma cells
CI
ED50 ED75 ED90 Dm m r
HONE1 1.142 0.830 0.606 1.072 3.940 1.000
NOTE: HONE1 cells were treated with ionizing radiation andAZD2281 and analyzedwith clonogenic survival assays. TheCI at ED50, ED75, and ED90 was calculated. A CI < 1 indicatessynergism.
Chow et al.
Mol Cancer Ther; 12(11) November 2013 Molecular Cancer Therapeutics2526
on July 2, 2018. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 26, 2013; DOI: 10.1158/1535-7163.MCT-13-0010

treated with PARP inhibitors. In addition to DNA repair,PARP family also participates in other processes, includ-
ing remodeling of chromatin and the regulation of tran-scription (reviewed in ref. 35). It is conceivable that thesensitization of nasopharyngeal carcinoma cells withPARP inhibition was not just due to disruption of DNArepair.
In conclusion, PARP1 overexpression is an early eventin nasopharyngeal carcinoma development. Cell line andanimal studies revealed that PARP inhibitors might beable to potentiate radiotherapy and chemotherapy fornasopharyngeal carcinoma.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: J.P.H. Chow, H. Chen, S.W. Tsao, R.Y.C. PoonDevelopment of methodology: J.P.H. Chow, M. Mao, H. ChenAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): J.P.H. Chow,W.Y.Man,M.Mao, F. Cheung,M.L.Lung, R.Y.C. PoonAnalysis and interpretation of data (e.g., statistical analysis, biostatis-tics, computational analysis): J.P.H. Chow, W.Y. Man, M. Mao, H. Chen,F. Cheung, J. Nicholls, R.Y.C. PoonWriting, review, and/or revision of the manuscript: J.P.H. Chow, W.Y.Man, M. Mao, M.L. Lung, R.Y.C. PoonAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): W.Y. Man, H. ChenStudy supervision: R.Y.C. Poon
AcknowledgmentsThe authors thank Anita Lau and Kenji Nishiura for technical assis-
tance, and Junyi Zhang for help in generating the CHFR antibodies.
Grant SupportThis work was supported in part by the Research Grants Council grant
AOE-MG/M-08/06 (R.Y.C. Poon).The costs of publication of this article were defrayed in part by the
payment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received January 7, 2013; revised August 7, 2013; accepted August 14,2013; published OnlineFirst August 26, 2013.
References1. JemalA,BrayF,CenterMM,Ferlay J,WardE, FormanD.Global cancer
statistics. CA Cancer J Clin 2011;61:69–90.2. Curado MP, Edwards B, Shin HR, Storm H, Ferlay J, Heanue M, et al.
Cancer incidence in five continents. Lyon, France: International Agen-cy for Research on Cancer; 2007. p. 896.
3. Dawson CW, Port RJ, Young LS. The role of the EBV-encoded latentmembrane proteins LMP1 and LMP2 in the pathogenesis of naso-pharyngeal carcinoma (NPC). Semin Cancer Biol 2012;22:144–53.
4. Chang ET, Adami HO. The enigmatic epidemiology of nasopharyngealcarcinoma. Cancer Epidemiol Biomarkers Prev 2006;15:1765–77.
5. Chan AT, Felip E. Nasopharyngeal cancer: ESMO clinical recommen-dations for diagnosis, treatment and follow-up. Ann Oncol 2009;20(Suppl 4):123–5.
6. Waldron J, Tin MM, Keller A, Lum C, Japp B, Sellmann S, et al.Limitation of conventional two dimensional radiation therapy planningin nasopharyngeal carcinoma. Radiother Oncol 2003;68:153–61.
7. Cheng JC, Chao KS, Low D. Comparison of intensity modulatedradiation therapy (IMRT) treatment techniques for nasopharyngealcarcinoma. Int J Cancer 2001;96:126–31.
8. Lee AW, Poon YF, Foo W, Law SC, Cheung FK, Chan DK, et al.Retrospective analysis of 5037 patients with nasopharyngeal carci-noma treated during 1976–1985: overall survival and patterns offailure. Int J Radiat Oncol Biol Phys 1992;23:261–70.
9. Luo X, Kraus WL. On PAR with PARP: cellular stress signalingthrough poly(ADP-ribose) and PARP-1. Genes Dev 2012;26:417–32.
10. De Vos M, Schreiber V, Dantzer F. The diverse roles and clinicalrelevance of PARPs in DNA damage repair: current state of the art.Biochem Pharmacol 2012;84:137–46.
11. Kummar S, Chen A, Parchment RE, Kinders RJ, Ji J, Tomaszewski JE,et al. Advances in using PARP inhibitors to treat cancer. BMC Med2012;10:25.
12. Garnett MJ, Edelman EJ, Heidorn SJ, Greenman CD, Dastur A, LauKW, et al. Systematic identification of genomic markers of drugsensitivity in cancer cells. Nature 2012;483:570–5.
13. Cheung ST, Huang DP, Hui AB, Lo KW, Ko CW, Tsang YS, et al.Nasopharyngeal carcinoma cell line (C666-1) consistently harbouringEpstein–Barr virus. Int J Cancer 1999;83:121–6.
Table 3. PARP1 is overexpressed in humanprimary nasopharyngeal carcinoma
PARP1expression
Low High
Variable n (%) n (%) Total P
Age�50 (median) 29 (58) 21 (42) 50<50 11 (24) 34 (76) 45 0.003a
SexMale 29 (43) 38 (57) 67Female 13 (42) 18 (58) 31 0.900
SurvivalAlive 19 (54) 16 (46) 35Dead 15 (29) 36 (71) 51 0.020a
Nasopharyngeal carcinoma stageNormal 10 (91) 1 (9) 11I 1 (20) 4 (80) 5 0.005a
II 12 (43) 16 (57) 28 0.006a
III 7 (41) 10 (59) 17 0.008a
IV 13 (39) 20 (61) 33 0.003a
M 3 (38) 5 (62) 8 0.013a
I, II, II, IV, M 36 (40) 55 (60) 91 0.001a
NOTE: The expression of PARP1 in normal nasopharyngealand nasopharyngeal carcinoma tissues was analyzed usingtissuemicroarray and immunohistochemistry.Nonmalignantandmalignant tissueswere scored for lowor high expressionof PARP1 by assessing PARP1 staining in the nucleus. ThePearsonx2 test was used to assess the association betweenPARP1 expression and several clinicopathologic variables.aVariables with statistical significance (P < 0.05) by Pearsonx2 test.
PARP in Nasopharyngeal Carcinoma
www.aacrjournals.org Mol Cancer Ther; 12(11) November 2013 2527
on July 2, 2018. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 26, 2013; DOI: 10.1158/1535-7163.MCT-13-0010

14. SizhongZ,XiukungG,Yi Z.Cytogenetic studieson anepithelial cell linederived from poorly differentiated nasopharyngeal carcinoma. Int JCancer 1983;31:587–90.
15. Glaser R, Zhang HY, Yao KT, Zhu HC, Wang FX, Li GY, et al. Twoepithelial tumor cell lines (HNE-1 and HONE-1) latently infected withEpstein–Barr virus that were derived from nasopharyngeal carcino-mas. Proc Natl Acad Sci U S A 1989;86:9524–8.
16. Li HM, Man C, Jin Y, Deng W, Yip YL, Feng HC, et al. Molecular andcytogenetic changes involved in the immortalization of nasopharyn-geal epithelial cells by telomerase. Int J Cancer 2006;119:1567–76.
17. Siu WY, Arooz T, Poon RY. Differential responses of proliferatingversus quiescent cells to adriamycin. Exp Cell Res 1999;250:131–41.
18. Chou TC, Talalay P. Quantitative analysis of dose–effect relationships:the combined effects of multiple drugs or enzyme inhibitors. AdvEnzyme Regul 1984;22:27–55.
19. Poon RY, Toyoshima H, Hunter T. Redistribution of the CDK inhibitorp27 between different cyclin. CDK complexes in the mouse fibroblastcell cycle and in cells arrested with lovastatin or ultraviolet irradiation.Mol Biol Cell 1995;6:1197–213.
20. Xie D, Sham JS, Zeng WF, Lin HL, Che LH, Wu HX, et al. Heteroge-neous expression and association of beta-catenin, p16 and c-myc inmultistage colorectal tumorigenesis and progression detected bytissue microarray. Int J Cancer 2003;107:896–902.
21. Cheung HW, Ching YP, Nicholls JM, Ling MT, Wong YC, Hui N, et al.Epigenetic inactivation ofCHFR in nasopharyngeal carcinoma throughpromoter methylation. Mol Carcinog 2005;43:237–45.
22. Kashima L, Idogawa M, Mita H, Shitashige M, Yamada T, Ogi K,et al. CHFR protein regulates mitotic checkpoint by targeting PARP-1 protein for ubiquitination and degradation. J Biol Chem 2012;287:12975–84.
23. Menear KA, Adcock C, Boulter R, Cockcroft XL, Copsey L, CranstonA, et al. 4-[3-(4-Cyclopropanecarbonylpiperazine-1-carbonyl)-4-fluorobenzyl]-2H-phthalazin-1-one: a novel bioavailable inhibitor ofpoly(ADP-ribose) polymerase-1. J Med Chem 2008;51:6581–91.
24. Qin HD, Shugart YY, Bei JX, Pan QH, Chen L, Feng QS, et al.Comprehensive pathway-based association study of DNA repair genevariants and the risk of nasopharyngeal carcinoma. Cancer Res 2011;71:3000–8.
25. Cheng Y, Lung HL, Wong PS, Hao DC, Man CS, Stanbridge EJ,et al. Chromosome 13q12 region critical for the viability and growth
of nasopharyngeal carcinoma hybrids. Int J Cancer 2004;109:357–62.
26. Idogawa M, Yamada T, Honda K, Sato S, Imai K, Hirohashi S. Poly(ADP-ribose) polymerase-1 is a component of the oncogenicT-cell factor-4/beta-catenin complex. Gastroenterology 2005;128:1919–36.
27. Ghabreau L, Roux JP, Frappart PO,Mathevet P, Patricot LM,MokniM,et al. Poly(ADP-ribose) polymerase-1, a novel partner of progesteronereceptors in endometrial cancer and its precursors. Int J Cancer 2004;109:317–21.
28. ShimizuS, Nomura F, Tomonaga T, SunagaM,NodaM, EbaraM, et al.Expression of poly(ADP-ribose) polymerase in human hepatocellularcarcinoma and analysis of biopsy specimens obtained under sono-graphic guidance. Oncol Rep 2004;12:821–5.
29. Rojo F, Garcia-Parra J, Zazo S, Tusquets I, Ferrer-Lozano J,Menendez S, et al. Nuclear PARP-1 protein overexpression isassociated with poor overall survival in early breast cancer. AnnOncol 2012;23:1156–64.
30. Domagala P, Huzarski T, Lubinski J, Gugala K, Domagala W. PARP-1expression in breast cancer including BRCA1-associated, triple neg-ative and basal-like tumors: possible implications for PARP-1 inhibitortherapy. Breast Cancer Res Treat 2011;127:861–9.
31. Ossovskaya V, Koo IC, Kaldjian EP, Alvares C, Sherman BM. Upre-gulation of Poly (ADP-Ribose) Polymerase-1 (PARP1) in triple-negativebreast cancer and other primary human tumor types. Genes Cancer2010;1:812–21.
32. Zaniolo K, Rufiange A, Leclerc S, Desnoyers S, Guerin SL. Regulationof the poly(ADP-ribose) polymerase-1 gene expression by the tran-scription factors Sp1 and Sp3 is under the influence of cell density inprimary cultured cells. Biochem J 2005;389:423–33.
33. Laniel MA, Bergeron MJ, Poirier GG, Guerin SL. A nuclear factorother than Sp1 binds the GC-rich promoter of the gene encoding ratpoly(ADP-ribose) polymerase in vitro. Biochem Cell Biol 1997;75:427–34.
34. Murai J, Huang SY, Das BB, Renaud A, Zhang Y, Doroshow JH, et al.TrappingofPARP1andPARP2byclinical PARP inhibitors.CancerRes2012;72:5588–99.
35. Krishnakumar R, Kraus WL. The PARP side of the nucleus: molecularactions, physiological outcomes, and clinical targets. Mol Cell 2010;39:8–24.
Chow et al.
Mol Cancer Ther; 12(11) November 2013 Molecular Cancer Therapeutics2528
on July 2, 2018. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 26, 2013; DOI: 10.1158/1535-7163.MCT-13-0010

2013;12:2517-2528. Published OnlineFirst August 26, 2013.Mol Cancer Ther Jeremy P.H. Chow, Wing Yu Man, Mao Mao, et al. Inhibition Enhances RadiotherapyPARP1 Is Overexpressed in Nasopharyngeal Carcinoma and Its
Updated version
10.1158/1535-7163.MCT-13-0010doi:
Access the most recent version of this article at:
Material
Supplementary
http://mct.aacrjournals.org/content/suppl/2013/08/27/1535-7163.MCT-13-0010.DC1
Access the most recent supplemental material at:
Cited articles
http://mct.aacrjournals.org/content/12/11/2517.full#ref-list-1
This article cites 34 articles, 7 of which you can access for free at:
Citing articles
http://mct.aacrjournals.org/content/12/11/2517.full#related-urls
This article has been cited by 3 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mct.aacrjournals.org/content/12/11/2517To request permission to re-use all or part of this article, use this link
on July 2, 2018. © 2013 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst August 26, 2013; DOI: 10.1158/1535-7163.MCT-13-0010