
Papéis Avulsos de Zoologia - SciELO · Papéis Avulsos de Zoologia ISSN 0031-1049 Museu de...
25
Volume 43(3):31-54, 2003 www.scielo.br/paz.htm Papéis Avulsos de Zoologia ISSN 0031-1049 Museu de Zoologia da Universidade de São Paulo COMPOSIÇÃO DE BANDOS MISTOS DE AVES EM FRAGMENTOS DE MATA ATLÂNTICA NO SUDESTE DO BRASIL MARCOS MALDONADO-COELHO 1,2 MIGUEL ANGELO MARINI 1,3 ABSTRACT Habitat fragmentation has been one of the most important subjects for conservation biology during the last years. However, few studies have evaluated the effects of forest fragmentation on mixed-species bird flocks. These flocks are associations of two or more species, which cohesion is related to networks of interactions between flock members. Decreased predation and increased foraging efficiency are two main selective advantages thought to explain the evolution of mixed-species flocking behavior. The aim of this work was to analyse mixed-species flock composition in relation to the size of Atlantic Forest fragments during rainy and dry seasons, at Zona da Mata region, Southeastern of Minas Gerais State, Brazil. Three distinct kinds of mixed-species flocks were identified at the study area: heterogeneous flocks, understory flocks and canopy flocks. The first one was observed in all forest fragments, although understory flocks were observed in only three. Canopy flocks were rare in all fragments. Some species were presente in two different kinds of flocks. The Black-goggled Tanager (Trichothraupis melanops) and the Golden-crowned Warbler (Basileuterus culicivorus) were nuclear species of heterogeneous flocks. The Red-crowned Ant-tanager (Habia rubica) was the most important species in understory flock formation and cohesion. In canopy flocks none of the species had similar characteristics. Here we analyzed the effect of season and forest fragment area on the composition of understory and heterogeneous mixed-flock based on two approaches: 1) – frequency of species and 2) – ecological traits of flocking species. In relation to species’ frequencies, seasonality affected heterogeneous flocks composition more strongly than fragment area. Only two forest fragments differed slightly in heterogeneous flocks’ composition. Both area and seasonality influenced understory flock composition. Excluding the category of canopy species of heterogeneous flocks, which decreased with fragment area in the rainy season, the overall proportion of species in the categories forest-dependence, foraging strata and foraging characteristics did not change in relation to forest fragment area or season, neither for heterogeneous nor for understory flocks. KEYWORDS: Atlantic Forest, Brazil, forest fragmentation, seasonal effects, mixed-species flock composition 1 Departamento de Biologia Geral, ICB, C.P. 486, Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brasil, 30161-970. 2 Endereço atual: Department of Biology and International Center for Tropical Ecology, University of Missouri, 8001 Natural Bridge Road, St. Louis, Missouri, 63121, EUA. 3 Endereço atual: Departamento de Zoologia, IB, Universidade de Brasília, Brasília, DF, Brasil, 70910-900.
Transcript of Papéis Avulsos de Zoologia - SciELO · Papéis Avulsos de Zoologia ISSN 0031-1049 Museu de...

Volume 43(3):31-54, 2003 www.scielo.br/paz.htm
Papéis Avulsos de Zoologia
ISSN 0031-1049
Museu de Zoologia da Universidade de São Paulo
COMPOSIÇÃO DE BANDOS MISTOS DE AVES EM FRAGMENTOS DEMATA ATLÂNTICA NO SUDESTE DO BRASIL
MARCOS MALDONADO-COELHO1,2
MIGUEL ANGELO MARINI1,3
ABSTRACT
Habitat fragmentation has been one of the most important subjects for conservation biology during the lastyears. However, few studies have evaluated the effects of forest fragmentation on mixed-species bird flocks.These flocks are associations of two or more species, which cohesion is related to networks of interactionsbetween flock members. Decreased predation and increased foraging efficiency are two main selective advantagesthought to explain the evolution of mixed-species flocking behavior. The aim of this work was to analysemixed-species flock composition in relation to the size of Atlantic Forest fragments during rainy and dryseasons, at Zona da Mata region, Southeastern of Minas Gerais State, Brazil. Three distinct kinds ofmixed-species flocks were identified at the study area: heterogeneous flocks, understory flocks and canopyflocks. The first one was observed in all forest fragments, although understory flocks were observed in onlythree. Canopy flocks were rare in all fragments. Some species were presente in two different kinds of flocks.The Black-goggled Tanager (Trichothraupis melanops) and the Golden-crowned Warbler (Basileuterusculicivorus) were nuclear species of heterogeneous flocks. The Red-crowned Ant-tanager (Habia rubica)was the most important species in understory flock formation and cohesion. In canopy flocks none of thespecies had similar characteristics. Here we analyzed the effect of season and forest fragment area on thecomposition of understory and heterogeneous mixed-flock based on two approaches: 1) – frequency ofspecies and 2) – ecological traits of flocking species. In relation to species’ frequencies, seasonality affectedheterogeneous flocks composition more strongly than fragment area. Only two forest fragments differedslightly in heterogeneous flocks’ composition. Both area and seasonality influenced understory flock composition.Excluding the category of canopy species of heterogeneous flocks, which decreased with fragment area inthe rainy season, the overall proportion of species in the categories forest-dependence, foraging strata andforaging characteristics did not change in relation to forest fragment area or season, neither for heterogeneousnor for understory flocks.
KEYWORDS: Atlantic Forest, Brazil, forest fragmentation, seasonal effects, mixed-speciesflock composition
1 Departamento de Biologia Geral, ICB, C.P. 486, Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brasil, 30161-970.2 Endereço atual: Department of Biology and International Center for Tropical Ecology, University of Missouri, 8001 Natural Bridge
Road, St. Louis, Missouri, 63121, EUA.3 Endereço atual: Departamento de Zoologia, IB, Universidade de Brasília, Brasília, DF, Brasil, 70910-900.

32 MALDONADO-COELHO & MARINI: BANDOS MISTOS DE AVES
INTRODUÇÃO
Investigações recentes têm se esforçado paraelucidar os padrões de composição de espécies de avesem fragmentos florestais e determinar quais proces-sos originam tais padrões (Willson et al. 1994, Stouffer& Bierregaard 1995, Sieving & Karr 1997, Warburton1997). As espécies de aves variam em suas respostasem relação à fragmentação na medida em que as po-pulações de algumas aumentam, algumas permanecemnão afetadas, e outras declinam ou desaparecem emfragmentos (Lynch & Whigham 1984, Blake & Karr1987, Stouffer & Bierregaard 1995). Uma grande vari-edade de características ecológicas e de histórias devida têm sido associadas com a propensão à extinçãolocal em populações isoladas de aves em florestas tro-picais, incluindo raridade natural (Christiansen & Pitter1997, Sieving & Karr 1997), grande tamanho corporal(Leck 1979), baixa fecundidade (Willis 1974, Sieving& Karr 1997), tipo e localização do ninho (Willson etal. 1994, Warburton 1997), dieta (Willis 1979, Stouffer& Bierregaard 1995), reduzida tolerância à matriz dehabitats modificados circundando os remanescentesflorestais (Lovejoy et al. 1986, Stouffer & Bierregaard1995, Bierregaard & Stouffer 1997, Warburton 1997)e endemismo (Willson et al. 1994, mas veja Marini2001).
Além disso, a fragmentação florestal gera possí-veis efeitos na cadeia trófica e nas interaçõesinterespecíficas que podem, em última análise, acarre-tar na diminuição da sobrevivência dos organismos.Um dos possíveis tipos de interações ecológicas quepodem ser alteradas com a fragmentação seriam osbandos mistos de aves (Stouffer & Bierregaard 1995,Maldonado-Coelho & Marini 2000). Bandos mistos sãoassociações multiespecíficas que ocorrem em todos osambientes terrestres, cuja coesão depende da respostamútua entre os membros integrantes do bando (Munn& Terborgh 1979, Powell 1985, Jullien & Thiollay 1998).Muitos estudos têm tentado explicar as funçõesadaptativas dos bandos mistos, sendo as mais impor-tantes o aumento na eficiência de forrageamento e odeclínio no risco de predação (Buskirk 1976, Morse1977, Powell 1985). Sendo assim, alterações na estru-tura e composição dos bandos podem ter conseqüên-cias negativas sobre a sobrevivência dos indivíduos.
Bandos mistos de aves podem ser caracterizadoscom base na dieta das espécies dos membros integran-tes e sua posição no estrato vertical. Assim, podem-sereconhecer nitidamente bandos de copa e bandos desub-bosque na Floresta Amazônica (Munn & Terborgh1979, Munn 1985), ao passo que esta separação é me-
nos evidente na Floresta Atlântica (Stotz 1993, Aleixo1997, Develey 1997). Os bandos mistos de copa sãoconstituídos principalmente por espécies que se ali-mentam tanto de insetos como de frutos e tendem aser instáveis, enquanto que os bandos de sub-bosquesão compostos em sua grande maioria por espéciescom dieta basicamente insetívora, possuindo elevadaestabilidade e diversidade de espécies (Munn &Terborgh 1979, Powell 1979, Gradwohl & Greenberg1980, Munn 1985, Jullien & Thiollay 1998).
Uma das principais variáveis influenciando o ta-manho e a composição dos bandos mistos de aves é asazonalidade. As variações sazonais na composição eestrutura dos bandos se devem, em grande parte, àsvariações na distribuição temporal e espacial dos re-cursos alimentares, juntamente com as atividadesreprodutivas (Davis 1946, Powell 1979, Alves &Cavalcanti 1996, Machado 1999, Develey & Peres2000). Adicionalmente, outro fator que pode influen-ciar sazonalmente a composição e tamanho dos ban-dos mistos é a presença de espécies migratórias(Morrison et al. 1987, Hutto 1994, Gram 1998).
Este trabalho tem como objetivo avaliar a influ-ência da área dos fragmentos e da sazonalidade na com-posição dos bandos mistos de aves em fragmentos flo-restais de Mata Atlântica no Sudeste de Minas Gerais.
MÉTODOS
Área de estudo
Este estudo foi conduzido nos municípios dePaula Cândido e Viçosa, localizados na Zona da Mata,sudeste do Estado de Minas Gerais (20°42’30” a20°50’00”S e 42°48’45” a 42°56’15”W). A vegetaçãooriginal da região é classificada como FlorestaEstacional Semidecidual Montana (Veloso et al. 1991),encontrando-se atualmente inteiramente fragmentada(Ribon 1998). Os fragmentos remanescentes são cons-tituídos por matas secundárias e localizam-se, quaseque exclusivamente, nas encostas e topos de morros,sendo muito raros aqueles encobrindo grotas e baixa-das (Valverde 1958). Os remanescentes florestais en-contram-se inseridos em uma matriz de vegetação cons-tituída exclusivamente por plantações e pastagens paraa criação de gado (Ribon 1998).
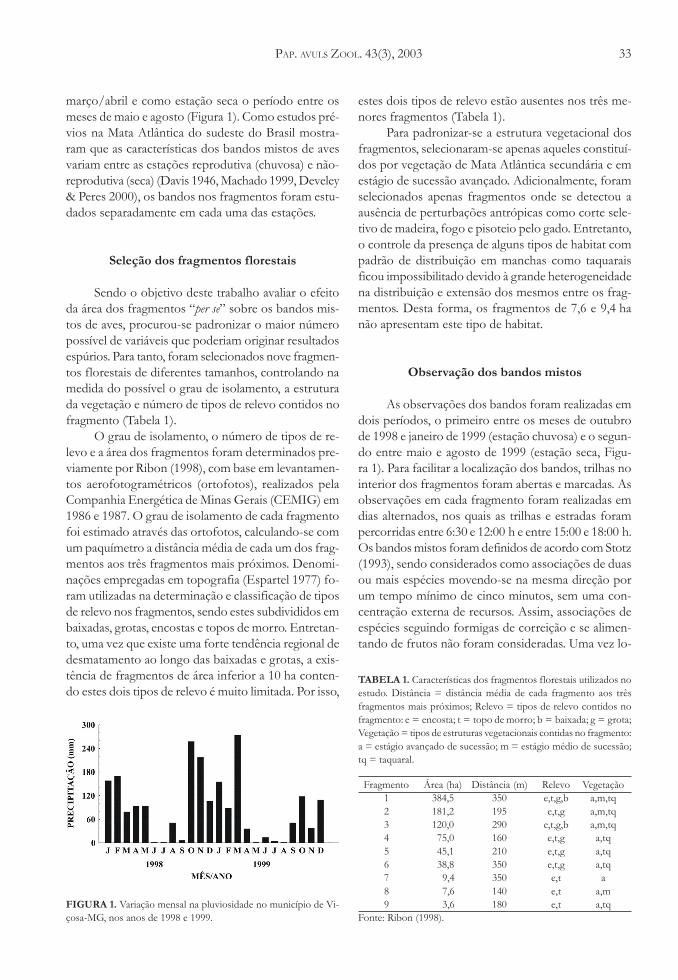
A região apresenta estações chuvosa e seca bemdefinidas e com reduzida variação anual em suas dura-ções. Desta forma, pode-se definir, com base na preci-pitação mensal, estação chuvosa como o período com-preendido entre os meses de setembro/outubro e
PAP. AVULS ZOOL. 43(3), 2003 33
março/abril e como estação seca o período entre osmeses de maio e agosto (Figura 1). Como estudos pré-vios na Mata Atlântica do sudeste do Brasil mostra-ram que as características dos bandos mistos de avesvariam entre as estações reprodutiva (chuvosa) e não-reprodutiva (seca) (Davis 1946, Machado 1999, Develey& Peres 2000), os bandos nos fragmentos foram estu-dados separadamente em cada uma das estações.
Seleção dos fragmentos florestais
Sendo o objetivo deste trabalho avaliar o efeitoda área dos fragmentos “per se” sobre os bandos mis-tos de aves, procurou-se padronizar o maior númeropossível de variáveis que poderiam originar resultadosespúrios. Para tanto, foram selecionados nove fragmen-tos florestais de diferentes tamanhos, controlando namedida do possível o grau de isolamento, a estruturada vegetação e número de tipos de relevo contidos nofragmento (Tabela 1).
O grau de isolamento, o número de tipos de re-levo e a área dos fragmentos foram determinados pre-viamente por Ribon (1998), com base em levantamen-tos aerofotogramétricos (ortofotos), realizados pelaCompanhia Energética de Minas Gerais (CEMIG) em1986 e 1987. O grau de isolamento de cada fragmentofoi estimado através das ortofotos, calculando-se comum paquímetro a distância média de cada um dos frag-mentos aos três fragmentos mais próximos. Denomi-nações empregadas em topografia (Espartel 1977) fo-ram utilizadas na determinação e classificação de tiposde relevo nos fragmentos, sendo estes subdivididos embaixadas, grotas, encostas e topos de morro. Entretan-to, uma vez que existe uma forte tendência regional dedesmatamento ao longo das baixadas e grotas, a exis-tência de fragmentos de área inferior a 10 ha conten-do estes dois tipos de relevo é muito limitada. Por isso,
estes dois tipos de relevo estão ausentes nos três me-nores fragmentos (Tabela 1).
Para padronizar-se a estrutura vegetacional dosfragmentos, selecionaram-se apenas aqueles constituí-dos por vegetação de Mata Atlântica secundária e emestágio de sucessão avançado. Adicionalmente, foramselecionados apenas fragmentos onde se detectou aausência de perturbações antrópicas como corte sele-tivo de madeira, fogo e pisoteio pelo gado. Entretanto,o controle da presença de alguns tipos de habitat compadrão de distribuição em manchas como taquaraisficou impossibilitado devido à grande heterogeneidadena distribuição e extensão dos mesmos entre os frag-mentos. Desta forma, os fragmentos de 7,6 e 9,4 hanão apresentam este tipo de habitat.
Observação dos bandos mistos
As observações dos bandos foram realizadas emdois períodos, o primeiro entre os meses de outubrode 1998 e janeiro de 1999 (estação chuvosa) e o segun-do entre maio e agosto de 1999 (estação seca, Figu-ra 1). Para facilitar a localização dos bandos, trilhas nointerior dos fragmentos foram abertas e marcadas. Asobservações em cada fragmento foram realizadas emdias alternados, nos quais as trilhas e estradas forampercorridas entre 6:30 e 12:00 h e entre 15:00 e 18:00 h.Os bandos mistos foram definidos de acordo com Stotz(1993), sendo considerados como associações de duasou mais espécies movendo-se na mesma direção porum tempo mínimo de cinco minutos, sem uma con-centração externa de recursos. Assim, associações deespécies seguindo formigas de correição e se alimen-tando de frutos não foram consideradas. Uma vez lo-
FIGURA 1. Variação mensal na pluviosidade no município de Vi-çosa-MG, nos anos de 1998 e 1999.
TABELA 1. Características dos fragmentos florestais utilizados noestudo. Distância = distância média de cada fragmento aos trêsfragmentos mais próximos; Relevo = tipos de relevo contidos nofragmento: e = encosta; t = topo de morro; b = baixada; g = grota;Vegetação = tipos de estruturas vegetacionais contidas no fragmento:a = estágio avançado de sucessão; m = estágio médio de sucessão;tq = taquaral.
Fragmento Área (ha) Distância (m) Relevo Vegetação1 384,5 350 e,t,g,b a,m,tq2 181,2 195 e,t,g a,m,tq3 120,0 290 e,t,g,b a,m,tq4 75,0 160 e,t,g a,tq5 45,1 210 e,t,g a,tq6 38,8 350 e,t,g a,tq7 9,4 350 e,t a8 7,6 140 e,t a,m9 3,6 180 e,t a,tq
Fonte: Ribon (1998).

34 MALDONADO-COELHO & MARINI: BANDOS MISTOS DE AVES
calizado, cada bando foi seguido por no máximo umahora, ou até ser perdido de vista, o que muitas vezesocorreu em conseqüência da elevada inclinação do ter-reno e do sub-bosque bastante denso. As observaçõesforam conduzidas com um binóculo 7-15 x 35 mm,sendo que para cada bando, quantificou-se o númerode espécies e de indivíduos de cada espécie.
Função das espécies nos bandos
As espécies foram classificadas de acordo comsua função nos bandos. Consideraram-se como espé-cies nucleares aquelas que contribuíram significativa-mente para a coesão e estabilidade dos bandos e apre-sentaram características comportamentais como asdescritas por Moynihan (1962): alta freqüência devocalizações e movimentos conspícuos. Uma espéciefoi considerada líder quando era a primeira espécie aatravessar um espaço aberto, sendo seguida pelas de-mais espécies, e como sentinela quando emitia gritosde alarme na presença de um possível predador.
Categorias ecológicas das espéciesassociadas aos bandos
As espécies foram categorizadas de acordo como grau de dependência florestal (dependentes,semidependentes e independentes; segundo Silva 1995)e com o comportamento de forrageamento (altura ecaracterísticas de forrageamento). Em relação à análi-se de altura de forrageamento, o estrato florestal foidividido em três níveis: sub-bosque (< 2 m), médioestrato (2-8 m) e copa (> 8 m). Para a categorizaçãoquanto às características de forrageamento, as espéci-es foram agrupadas em guildas, sendo classificadas deacordo com a dieta (insetívoro, onívoro e frugívoro),manobras e substrato de forrageamento. As guildasforam definidas principalmente baseadas em observa-ções pessoais das espécies na área de estudo(Maldonado-Coelho, dados não publicados) ecomplementariamente de acordo com Willis (1979) eSick (1997). Para as espécies insetívoras, quatro classesforam definidas: 1) generalistas (quando o substratode forrageamento inclui ar, folhas e ramos), 2)insetívoras que utilizam folhas como principal substratode captura da presa (insetívoras de folha), 3) insetívorasde tronco e 4) insetívoras aéreas. As espécies foramconsideradas onívoras ou frugívoras quando apresen-tam elevada proporção de frutos na dieta quando nãoassociadas aos bandos mistos.
Análises dos dados
As freqüências de participação de cada espécienos bandos para cada um dos fragmentos, durante asestações chuvosa e seca, foram estimadas por meio darazão entre o número de vezes em que cada espéciefoi registrada associada aos bandos de um determina-do fragmento pelo número total de bandos observa-dos neste fragmento. Estes cálculos foram realizadosseparadamente para os diferentes tipos de bandos mis-tos registrados nos fragmentos, e distintamente paraas estações chuvosa e seca.
Através das observações de campo, foram iden-tificados três tipos distintos de bandos mistos na áreade estudo, com base na estrutura e nos padrões deassociação entre as espécies participantes. Entretanto,para se confirmar se os diferentes tipos de bandosmistos diferiam na composição de espécies, foi reali-zada uma análise de correspondência utilizando-se oprograma MVSP (Kovac 1986). Como os três tipos deassociação foram observados no fragmento de maiorárea (384,5 ha), esta análise foi realizada utilizando-sesomente os bandos deste fragmento, nas estações chu-vosa e seca, separadamente. O método escolhido foi aanálise de correspondência destendenciada (DCA), queutiliza as similaridades nas associações de espécies paraordenar os bandos (Hill & Gauch 1980, Ludwig &Reynolds 1988). A DCA é derivada da análise de cor-respondência, porém é mais refinada que esta por re-duzir a compressão no primeiro eixo, e a distorção nosegundo e terceiros eixos (Hill & Gauch 1980). Paraesta análise, foi construída uma matriz de presença eausência das espécies, onde cada bando misto foi con-siderado como uma amostra (linhas), com a presençaou ausência (0 ou 1) das espécies listadas (colunas).Com a finalidade de evitar que espécies raramente de-tectadas nos bandos influenciassem a análise, apenasas espécies presentes em mais de 10% dos bandos emcada uma das duas estações foram consideradas. Osbandos foram agrupados de acordo com as similarida-des na composição específica, e para cada tipo de ban-do foram conduzidas análises distintas.
Análises de correspondência destendenciadas(DCA) foram utilizadas para ordenar os fragmentos,objetivando-se testar o efeito das variáveis estação e áreados fragmentos na composição de espécies dos bandos.Nesta análise utilizaram-se as freqüências de associaçãodas espécies em cada fragmento, nas estações chuvosa eseca separadamente. Os bandos de sub-bosque e hete-rogêneos (veja resultados) foram considerados distinta-mente nesta análise. Foram consideradas apenas as es-pécies com freqüência de associação igual ou superior a

PAP. AVULS ZOOL. 43(3), 2003 35
10% em cada tipo de bando, em pelo menos um dosfragmentos nos quais estes bandos foram registrados.
Variações nas proporções de espécies em diferen-tes categorias ecológicas (classes de dependência flores-tal, estrato e guildas alimentares) em bandos heterogê-neos em relação à área dos fragmentos foram analisa-das para cada estação separadamente, através de corre-lações não-paramétricas de Spearman. Com a finalida-de de se evitar a inflação da probabilidade de se rejeitara hipótese nula quando esta é verdadeira em decorrên-cia do elevado número de análises com dados relacio-nados (erro do Tipo I; Sokal & Rohlf 1998), foram uti-lizadas correções seqüenciais de Bonferroni (α = 0,05).A correção seqüencial de Bonferroni é mais adequadado que a correção padrão de Bonferroni por corrigir aprobabilidade de se cometer erro do Tipo I sem au-mentar a probabilidade de se cometer erro do Tipo II,ou seja, sem diminuir o poder do teste (Rice 1989). Es-tas análises foram conduzidas utilizando-se o programaestatístico SPSS (1998) versão 9.0 para Windows.
Em relação aos bandos de sub-bosque, diferençasnas proporções de espécies em cada uma das três cate-gorias ecológicas entre os fragmentos de 384,5, 75 e38,8 ha foram analisadas utilizando-se o teste G deheterogeneidade (Sokal & Rohlf 1998). Neste caso, acorreção seqüencial de Bonferroni não foi utilizada, poispara cada uma das três hipóteses nulas (cada categoriaecológica), somente uma análise estatística foi realizada.
RESULTADOS
Foram registradas 78 espécies em 356 bandosmistos, ao longo de 967 horas de observação nas esta-ções chuvosa e seca (Anexo 1). Na Tabela 2 encon-tram-se o número de dias e horas de observação nasestações chuvosa e seca em cada fragmento.
Um maior número de espécies foi observadodurante a estação seca (n = 71) em relação à estaçãochuvosa (n = 58). Passeriformes e não-passeriformesforam representados nos bandos por 72 e 6 espéciesrespectivamente (Anexo 1). As famílias mais represen-tativas nos bandos foram Tyrannidae (n = 26 espéci-es), Emberizidae (n = 16), Formicariidae (n = 8),Furnariidae (n = 7), Dendrocolaptidae (n = 6) e Picidae(n = 5) (Anexo 1).
Padrões de associação entre as espécies
Durante a estação chuvosa, 30 espécies foramincluídas na análise de correspondência destendenciada
(DCA) utilizada para se identificar os padrões de asso-ciação entre as espécies no fragmento de 384,5 ha (Fi-gura 2, Tabela 3). Quanto maior a proximidade de umbando ao outro no gráfico, maior a similaridade nacomposição específica entre eles. Por meio desta aná-lise verifica-se que as espécies formam três agrupa-mentos distintos. Na Figura 2A, três agrupamentosforam determinados pelo primeiro eixo, sendo: 1) as-
TABELA 2. Número total de dias e horas de observaçãodespendidas em cada um dos fragmentos florestais estudados, nasestações chuvosa e seca.
Fragmento Número de horas Número de dias(ha) de observação de observação
Chuvosa Seca Chuvosa Seca384,5 59 64 8 10181,2 61 46 10 6120,0 52 42 7 675,0 65 53 8 745,1 60 40 8 538,8 74 72 10 99,4 46 45 9 67,6 54 48 8 83,6 56 30 9 5
Total 527 440 77 62
FIGURA 2. Análise de correspondência destendenciada (DCA),avaliando-se os padrões de associação das espécies registradas embandos mistos no fragmento florestal de 384,5 ha, durante a esta-ção chuvosa (A) e a estação seca (B), utilizando-se em ambos oscasos o eixo I vs. o eixo II. Cada símbolo corresponde a um bando.

36 MALDONADO-COELHO & MARINI: BANDOS MISTOS DE AVES
sociações compostas principalmente por espécies desub-bosque como Habia rubica, Lepidocolaptes fuscus,Anabazenops fuscus , Automolus leucophthalmus,Campylorhamphus falcularius, Pyriglena leucoptera,Dysithamnus mentalis e Xiphocolaptes albicollis, e algumasespécies do estrato médio como Basileuterus culicivorus,Trichothraupis melanops e Leptopogon amaurocephalus deno-minada bandos de sub-bosque; 2) associações com-postas principalmente por espécies que ocupam osestratos superior, médio e inferior da floresta comoHylophilus amaurocephalus, Lepidocolaptes squamatus,Picumnus cirratus, Pachyramphus polychopterus, Trichothraupismelanops, Basileuterus culicivorus, Leptopogon amaurocephalus,Conirostrum speciosum, Myiopagis viridicata e Tolmomyiassulphurescens, sendo estas denominadas bandos hetero-gêneos; 3) associações compostas por espécies de copacomo Hemithraupis ruficapilla, C. speciosum e Tangaracyanoventris, denominadas bandos de copa. Desta for-ma, o eixo I mostra diferenças na composição dos ban-dos mistos de acordo com a posição que as espéciesocupam no estrato vertical da floresta durante a esta-ção chuvosa.
Durante a estação seca foram incluídas 40 espé-cies com freqüência de associação aos bandos superi-or a 10% no fragmento de 384,5 ha (Anexo 1). A aná-lise de correspondência mostra que, similarmente àestação chuvosa, os mesmos três tipos de bandos po-dem ser identificados quanto à composição de espéci-es (Figura 2B). As variâncias e autovalores também nãodiferiram entre as estações (Tabela 3). O eixo I sepa-rou os bandos mistos de sub-bosque dos demais ban-dos, revelando que H. rubica, L. fuscus, C. falcularius,A. leucophthalmus, A. fuscus e X. albicollis formaram, si-milarmente à estação chuvosa, associações distintas(Figura 2B). Este mesmo eixo mostrou que outras es-pécies como T. melanops, T. sulphurescens, Thamnophiluscaerulescens, Piaya cayana, P. cirratus, L. amaurocephalus,Synallaxis ruficapilla, C. speciosum, H. ruficapilla eH. amaurocephalus formaram associações distintas dosbandos de sub-bosque (bandos heterogêneos). Os ban-dos de copa compostos por H. ruficapilla, T. cyanoventris,Dacnis cayana e C. speciosum não apresentaramsobreposição com os demais bandos ao longo do eixoI, e desta forma puderam ser igualmente distinguidos.
Entretanto, apesar de algumas espécies se asso-ciarem preferencialmente a um tipo de bando misto,muitas espécies foram registradas freqüentemente emdois ou mais tipos de bando, em ambas as estações(Anexo 1). Desta forma, espécies preferencialmente dedossel (e.g. T. cyanoventris, D. cayana) foram observadasem bandos compostos de espécies de estrato médio einferior da floresta (bandos heterogêneos), e espéciesde estrato médio (e.g. B. culicivorus, T. melanops) foramregistradas em bandos de sub-bosque.
Os bandos mistos de sub-bosque foramregistrados em apenas três fragmentos florestais (384,5,75 e 38,8 ha) tanto na estação chuvosa (n = 42 ban-dos) quanto seca (n = 50 bandos), ao passo que osbandos heterogêneos foram observados em todos osnove fragmentos florestais estudados, tanto na esta-ção chuvosa (n = 105) como na seca (n = 146). Osregistros dos bandos de copa foram infreqüentes nasestações chuvosa (n = 7) e seca (n = 6), em todos osfragmentos estudados.
Uma vez que a análise de correspondênciadestendenciada (DCA) revelou que os três tipos debandos mistos formam associações discretas em ambasestações (Figuras 2A e 2B), juntamente com o fato deestes bandos possuírem estruturas distintas (veja emfunção das espécies nos bandos), optou-se por realizaranálises separadamente para cada tipo de bando.
Bandos heterogêneos
A ordenação dos bandos heterogêneos atravésda análise de correspondência destendenciada (DCA)baseada na composição de espécies produziu três ei-xos com variâncias reduzidas, indicando que as dife-renças entre os fragmentos e entre as duas estaçõesforam pequenas (Tabela 4). Entretanto, três grupospodem ser definidos pelo eixo I (autovalor = 0,16, Fi-gura 3): o primeiro constituído pelos bandos de todosos fragmentos durante a estação chuvosa (valores maisbaixos no eixo I), o segundo composto pelos bandosda maioria dos fragmentos durante a estação seca (va-lores intermediários no eixo I), e um terceiro constitu-ído especificamente pelos bandos do fragmento de
TABELA 3. Autovalores e variâncias para a análise de correspondência destendenciada (DCA) entre os bandos mistos de sub-bosque,heterogêneos e de copa registrados no fragmento de 384,5 ha, durante as estações chuvosa e seca.
Eixo 1 2 3Estação Chuvosa Seca Chuvosa Seca Chuvosa SecaAutovalor 0,59 0,59 0,30 0,25 0,16 0,19% da variância 18,7 19,1 9,3 8,1 5,1 6,3% da variância acumulada 18,7 19,1 28,0 27,2 33,1 33,5

PAP. AVULS ZOOL. 43(3), 2003 37
9,4 ha durante a estação seca (valores mais altos noeixo I). O eixo II (autovalor = 0,10) separa os bandosdo fragmento de 3,6 ha na estação seca (valores maisaltos no eixo II) dos demais (valores mais baixos noeixo II, Figura 3). Assim, os eixos I e II em conjuntomostram que os bandos nos fragmentos de 9,4 e 3,6 hanas estações seca e chuvosa, respectivamente, foramdistintos do padrão encontrado, uma vez que são osfragmentos que mais diferiram deles mesmos em dife-rentes estações, e dos demais fragmentos durante umamesma estação.
A separação dos fragmentos por estação no pri-meiro eixo indica que esta é a variável mais importantepara explicar as diferenças na composição dos bandosmistos heterogêneos. Isto está relacionado ao fato deque fragmentos de diferentes áreas em uma mesmaestação formam agrupamentos mais discretos do queum mesmo fragmento em diferentes estações (Figu-ra 3). O aparecimento e/ou aumento na freqüência deassociação de determinadas espécies aos bandos hete-rogêneos nos fragmentos durante a estação seca foi ofator responsável pela diferença sazonal na composi-ção dos bandos determinada pelo eixo I (Figura 3,
Anexo 1). Dentre as espécies que aumentaram a fre-qüência de associação aos bandos heterogêneos nosfragmentos estudados durante a estação seca pode-secitar: T. sulphurescens, S. ruficapilla, C. speciosum, Cyclarhisgujanensis, P. cayana, Platyrinchus mystaceus, T. cyanoventris,D. cayana e Todirostrum poliocephalum (Figura 3, Anexo 1).Adicionalmente, três espécies migratórias (Vireoolivaceus, Pachyramphus polychopterus e M. viridicata), queforam registradas nos bandos com maior freqüênciadurante a estação chuvosa (as duas primeiras) ou esta-vam ausentes durante a estação seca (M. viridicata), con-tribuíram para esta diferença sazonal.
Durante a estação chuvosa, o efeito de área nacomposição dos bandos heterogêneos foi maior nofragmento de 3,6 ha. Esta diferença, indicada pelo eixo2, se deve a um acentuado grau de associação deA. fuscus (87,5%), juntamente com a ausência de váriasespécies residentes como L. amaurocephalus,T. sulphurescens, Xenops rutilans, S. ruficapilla, C. gujanensise outras migratórias de verão como P. polychopterus eV. olivaceus nos bandos deste fragmento nesta estação(Anexo 1).
A associação das espécies Lepidocolaptesangustirostris e Contopus cinereus apenas aos bandos hete-rogêneos no fragmento de 9,4 ha durante a estaçãoseca explica a segregação deste fragmento em relaçãoaos demais ao longo do primeiro eixo (Anexo 1).
Bandos de sub-bosque
Nós excluímos Habia rubica da análise de DCA,uma vez que a sua presença foi utilizada para definiros bandos de sub-bosque (veja abaixo). A Tabela 5mostra que a proporção da variação total explicadapelos três primeiros eixos foi relativamente baixa(49,7%), indicando que a composição específica dosbandos de sub-bosque entre os três fragmentos (384,5,75 e 38,8 ha) e entre as estações diferiu pouco (Figu-ra 4). Entretanto, as diferenças entre fragmentos nacomposição dos bandos de sub-bosque foram maisimportantes do que a sazonalidade, devido ao fato deque a variação na composição dos bandos entre os frag-
TABELA 4. Autovalores e variâncias para a análise de correspon-dência destendenciada (DCA) comparando a composição dos ban-dos mistos heterogêneos entre os fragmentos e entre as estaçõeschuvosa e seca.
Eixo 1 2 3Autovalor 0,16 0,10 0,04% da variância 18,6 9,6 4,4% da variância acumulada 18,6 28,2 32,6
FIGURA 3. Ordenação dos bandos mistos heterogêneos por frag-mento e estação, eixos I vs. II. As estações do ano estão indicadaspor: c = estação chuvosa, s = estação seca.
TABELA 5. Autovalores e variâncias para a análise de correspon-dência destendenciada (DCA) comparando a composição dos ban-dos mistos de sub-bosque entre os fragmentos e entre as estaçõeschuvosa e seca.
Eixo 1 2 3Autovalor 0,10 0,04 0,01% da variância 34,4 13,0 2,3% da variância acumulada 34,4 47,4 49,7

38 MALDONADO-COELHO & MARINI: BANDOS MISTOS DE AVES
Bandos de copa
A raridade dos bandos de copa impossibilitou arealização de análises envolvendo a composição des-tes em relação à área dos fragmentos e à estação doano. Entretanto, puderam ser detectadas algumas dife-renças entre estações, uma vez que H. ruficapilla,C. speciosum e T. cyanoventris se associaram aos bandoscom maior freqüência durante a estação seca (Ane-xo 1). Adicionalmente, algumas espécies observadas aolongo da estação seca não foram registradas durante aestação chuvosa nestes bandos, como Camptostomaobsoletum, V. olivaceus, D. cayana e Nemosia pileata.
Análises das categorias ecológicas
A maioria das espécies nos bandos heterogêneose de sub-bosque nos fragmentos florestais, nas esta-ções chuvosa e seca, são espécies insetívoras de folha,dependentes de floresta e que forrageiam no médioestrato e sub-bosque (Tabelas 6 e 7). As análises en-volvendo as categorias ecológicas das espécies associ-adas aos bandos heterogêneos, revelaram que a pro-porção de espécies de copa aumentou significativamen-te com a área dos fragmentos durante a estação chu-vosa (r = 0,803; P = 0,005), não sendo observadas va-riações para as demais categorias nesta estação (Tabe-las 6 e 8). Durante a estação seca, nenhuma variaçãona proporção de espécies foi observada para quais-quer das categorias.
Em relação aos bandos de sub-bosque, não fo-ram observadas diferenças nas proporções para ne-nhuma das três categorias de dependência florestal(GH = 2,243; g.l. = 4; P > 0,05 para a estação chuvo-sa; GH = 0,864; g.l. = 4; P > 0,05 para a estação seca),estrato de forrageamento (GH = 0,188; g.l. = 4;P > 0,05 para a estação chuvosa; GH = 0,119; g.l. = 4;P > 0,05 para a estação seca) e característica deforrageamento (GH = 2,393; g.l. = 10; P > 0,05 paraa estação chuvosa; GH = 0,706; g.l. = 10; P > 0,05 paraa estação seca) entre os fragmentos de 384,5, 75 e38,8 ha, em ambas as estações (Tabela 7).
Função das espécies nos bandos
Dentre as espécies registradas nos bandos mis-tos heterogêneos nos fragmentos florestais estudados,T. melanops e B. culicivorus, além de serem as que commaior freqüência se associaram a estes bandos (Ane-xo 1), foram consideradas as espécies com maior im-
mentos foi maior que a variação em um mesmo frag-mento entre as duas estações.
A separação dos fragmentos ao longo do primei-ro eixo (autovalor = 0,10) está relacionada principal-mente a diferenças na composição dos bandos do frag-mento de 75 ha, em ambas as estações, em relação aosfragmentos de 384,5 e 38,8 ha. Adicionalmente, nesteeixo podem ser visualizadas pequenas diferenças sa-zonais na composição dos bandos nos fragmentos de75 e 384,5 ha (Figura 4). O eixo II (autovalor = 0,04),similarmente, parece mostrar primariamente diferen-ças na composição dos bandos de sub-bosque entreos fragmentos, separando o fragmento de 38,8 ha dosdois outros fragmentos maiores. Pode-se observar tam-bém um efeito sazonal para o fragmento de 38,8 ha(Figura 4).
A segregação do fragmento de 75 ha em relaçãoaos outros dois observada no primeiro eixo, pode seratribuída a uma reduzida freqüência de associação deA. fuscus e A. leucophthalmus, juntamente com o aumen-to na participação de L. amaurocephalus, nos bandosdeste fragmento. As variações sazonais na composi-ção dos bandos deste fragmento se devem a um au-mento na freqüência de associação de L. amaurocephalus,C. falcularius e X. rutilans durante a estação seca (Ane-xo 1). A variação sazonal observada para o fragmentode 384,5 ha se deve a um aumento na associação deA. leucophthalmus (38,8%) e Veniliornis maculifrons (33%)durante a estação seca (Figura 4, Anexo 1). A diferen-ça na composição dos bandos do fragmento de 38,8 haem relação aos demais se deve a uma redução na asso-ciação de Dysithamnus mentalis, X. albicollis e P. leucopteranos bandos deste fragmento (Anexo 1), enquanto quea variação sazonal neste fragmento se deve principal-mente a um aumento na freqüência de A. fuscus duran-te a estação seca (Anexo 1).
FIGURA 4. Ordenação dos bandos mistos de sub-bosque por frag-mento e estação, eixos I vs. II. As estações do ano estão indicadaspor: c = estação chuvosa, s = estação seca.

PAP. AVULS ZOOL. 43(3), 2003 39TA
BELA
6. N
úmer
o (%
) de
espé
cies
obs
erva
das
nos
band
os m
istos
het
erog
êneo
s pa
ra c
ada
uma
das
cate
goria
s de
pend
ênci
a flo
rest
al, e
stra
to d
e fo
rrag
eam
ento
e c
arac
terís
ticas
de
forr
agea
men
to n
osno
ve fr
agm
ento
s flo
rest
ais, d
uran
te a
s est
açõe
s chu
vosa
(C) e
seca
(S).
Cate
goria
Áre
a do
Fra
gmen
to (h
a)3,
67,
69,
438
,845
,175
120
181,
238
4,5
CS
CS
CS
CS
CS
CS
CS
CS
CS
Dep
endê
ncia
Flor
estal
Dep
ende
nte
821
1627
1619
1627
2022
2428
2428
2732
2332
(88,
9)(6
3,6)
(94,
1)(6
9,2)
(69,
6)(6
5,6)
(80,
0)(7
5,0)
(87,
0)(7
3,3)
(85,
7)(6
8,3)
(85,
7)(6
5,1)
(87,
1)(7
4,4)
(79,
3)(6
9,6)
Sem
idep
ende
nte
19
111
46
49
36
410
413
49
512
(11,
1)(2
7,3)
(5,9
)(2
8,2)
(17,
4)(2
0,7)
(20,
0)(2
5,0)
(13,
0)(2
0,0)
(14,
3)(2
4,4)
(14,
3)(3
0,2)
(12,
9)(2
0,9)
(17,
3)(2
6,1)
Inde
pend
ente
03
01
34
00
02
03
02
02
12
(9,1
)(2
,6)
(13,
0)(1
3,7)
(6,7
)(7
,3)
(4,7
)(4
,7)
(3,4
)(4
,3)
Estr
ato
de fo
rragea
ment
o
Copa
113
010
39
48
37
713
412
711
713
(11,
1)(3
9,4)
(25,
6)(1
3,0)
(31,
0)(2
0,0)
(22,
2)(1
3,0)
(23,
3)(2
5,0)
(31,
7)(1
4,3)
(27,
9)(2
2,6)
(25,
6)(2
4,1)
(28,
3)
Méd
io e
stra
to5
1310
1414
1510
1711
1516
2016
1714
1514
18(5
5,6)
(39,
4)(5
8,8)
(35,
9)(6
0,9)
(51,
8)(5
0,0)
(47,
2)(4
7,9)
(50,
0)(5
7,1)
(48,
8)(5
7,1)
(39,
5)(4
5,2)
(34,
9)(4
8,3)
(39,
1)
Sub-
bosq
ue3
77
156
56
119
85
88
1410
178
15(3
3,3)
(21,
2)(4
1,2)
(38,
5)(2
6,1)
(17,
2)(3
0,0)
(30,
6)(3
9,1)
(26,
7)(1
7,9)
(19,
5)(2
8,6)
(32,
6)(3
2,2)
(39,
5)(2
7,6)
(32,
6)
Cara
cterís
ticas
de f
orra
geame
nto
Inse
tívor
o ge
nera
lista
15
44
24
13
13
35
34
33
39
(11,
1)(1
5,1)
(23,
6)(1
0,2)
(8,8
)(1
3,8)
(5,0
)(8
,3)
(4,3
)(1
0,0)
(10,
7)(1
2,2)
(10,
7)(9
,4)
(9,7
)(7
,0)
(10,
3)(1
9,6)
Inse
tívor
o fo
lha
417
822
1113
1221
1517
1616
1723
1724
1523
(44,
4)(5
1,6)
(47,
0)(5
6,4)
(47,
8)(4
4,8)
(60,
0)(5
8,4)
(65,
2)(5
6,7)
(57,
0)(3
9,0)
(60,
7)(5
3,5)
(54,
8)(5
5,8)
(51,
8)(5
0,0)
Inse
tívor
o tro
nco
24
33
76
58
55
69
66
78
57
(22,
3)(1
2,1)
(17,
6)(7
,7)
(30,
4)(2
0,7)
(25,
0)(2
2,2)
(21,
8)(1
6,7)
(21,
4)(2
2,0)
(21,
4)(1
3,9)
(22,
6)(1
8,6)
(17,
2)(1
5,2)
Inse
tívor
o ar
00
00
01
00
01
01
01
00
00
(3,4
)(3
,3)
(2,4
)(2
,3)
Oní
voro
16
28
35
13
24
29
17
37
46
(11,
1)(1
8,2)
(11,
8)(2
0,6)
(13,
0)(1
7,3)
(5,0
)(8
,3)
(8,7
)(1
3,3)
(7,2
)(2
2,0)
(3,6
)(1
6,3)
(9,7
)(1
6,3)
(13,
8)(1
3,0)
Frug
ívor
o1
10
20
01
10
01
11
21
12
1(1
1,1)
(3,0
)(5
,1)
(5,0
)(2
,8)
(3,2
)(2
,4)
(3,6
)(4
,6)
(3,2
)(2
,3)
(6,9
)(2
,2)

40 MALDONADO-COELHO & MARINI: BANDOS MISTOS DE AVES
portância para sua coesão, sendo as únicas considera-das nucleares. Trichothraupis melanops e B. culicivorus pos-suem vários atributos em comum com outras espéciesnucleares (Moynihan 1962, Powell 1985), como eleva-da freqüência de vocalizações (para ambas) e movi-mentos conspícuos dentro dos bandos (neste caso paraT. melanops). Para maiores detalhes sobre o comporta-mento destas espécies nos bandos mistos em fragmen-tos florestais no Sudeste de Minas Gerais vejaMaldonado-Coelho & Marini (2000).
Para os bandos de sub-bosque, H. rubica foi aespécie com maior freqüência de associação (100% nosfragmentos de 384,5, 75 e 38,8 ha em ambas estações)e a mais importante, preenchendo as funções de espé-cie líder, nuclear e sentinela. A ocorrência de bandosde sub-bosque em um fragmento florestal estava con-dicionada unicamente à presença desta espécie, sendoque a ausência de H. rubica em um fragmento resulta-va no desaparecimento deste tipo de associação(Maldonado-Coelho & Marini, em preparação). Estaespécie foi a mais conspícua nestes bandos, apresen-tando alta freqüência de vocalizações, comportamen-
TABELA 7. Número (%) de espécies observadas nos bandos mistos de sub-bosque para cada uma das categorias dependência florestal,estrato de forrageamento e características de forrageamento nos fragmentos de 384,5, 75 e 38,8 ha, durante as estações chuvosa (C) eseca (S).
Categoria Área do Fragmento (ha)384,5 75 38,8
C S C S C SDependência Florestal
Dependente 19 18 14 17 11 13(100) (94,7) (93,3) (94,4) (100) (86,7)
Semidependente 0 1 1 1 0 2(5,3) (6,7) (5,6) (13,3)
Independente 0 0 0 0 0 0
Estrato de forrageamentoCopa 0 0 0 0 0 0Médio estrato 10 9 8 9 5 8
(52,6) (47,4) (53,3) (50) (45,4) (53,3)Sub-bosque 9 10 7 9 6 7
(47,4) (52,6) (46,7) (50) (54,6) (46,7)
Características de forrageamentoInsetívoro generalista 1 1 1 1 1 1
(5,3) (5,3) (6,7) (5,6) (9,1) (6,7)Insetívoro folha 8 10 8 9 5 7
(42,1) (52,6) (53,3) (50) (45,4) (46,7)Insetívoro tronco 5 5 4 6 3 4
(26,3) (26,3) (26,7) (33,3) (27,3) (26,6)Insetívoro ar 0 0 0 0 0 0Onívoro 4 3 2 2 2 3
(21,0) (15,8) (13,3) (11,1) (18,2) (20,0)Frugívoro 1 0 0 0 0 0
(5,3)
TABELA 8. Coeficientes de correlação de Spearman (r) entre aárea dos fragmentos e a proporção de espécies nos bandos hetero-gêneos para as categorias dependência florestal, estrato deforrageamento e características de forrageamento, nas estações chu-vosa e seca. As correlações significativas (após a correção seqüencialde Bonferroni) estão indicadas por um asterisco.
Categoria Chuva Secar P r P
Dependência FlorestalDependente -0,367 0,332 0,367 0,332Semidependente 0,293 0,444 -0,100 0,798Independente 0,137 0,725 -0,259 0,500
Estrato de forrageamentoCopa 0,803 0,009* -0,151 0,699Médio estrato -0,527 0,145 -0,233 0,546Sub-bosque -0,417 0,265 0,418 0,262
Características de forrageamentoInsetívoro generalista -0,259 0,500 -0,200 0,606Insetívoro folha 0,500 0,170 -0150 0,700Insetívoro tronco -0,343 0,366 0,233 0,546Insetívoro ar – – -0,037 0,926Onívoro -0,117 0,765 -0,427 0,252Frugívoro 0,119 0,761 -0,318 0,404

PAP. AVULS ZOOL. 43(3), 2003 41
to que presumivelmente tem funções de coesão e for-mação dos bandos de sub-bosque. Em todos estesbandos H. rubica foi invariavelmente a espécie líder,sendo prontamente seguida pelas demais espécies as-sociadas aos bandos.
A capacidade de H. rubica como espécie líder dosbandos pôde ser demonstrada em algumas ocasiões(n = 5), quando, uma vez dispersadas as demais espé-cies do grupo, os indivíduos de H. rubica começavam avocalizar intensamente, atraindo e sendo seguidas ime-diatamente pelas espécies associadas ao bando.
Em relação aos bandos de copa, nenhuma espé-cie com os requisitos de espécie nuclear, sentinela oulíder foi identificada.
DISCUSSÃO
A ocorrência de três tipos de bandos mistos nosfragmentos florestais na região de Viçosa é similar aopadrão encontrado por Aleixo (1997) e Develey (1997)em duas outras localidades de Mata Atlântica do su-deste do Brasil, onde igualmente registraram bandoscompostos de espécies de copa, bandos constituídosde espécies dos estratos superior-médio-inferior e ban-dos compostos por espécies de sub-bosque.
Bandos heterogêneos
Análise de correspondênciaA área dos fragmentos influenciou pouco a com-
posição dos bandos heterogêneos, sendo a estação doano uma variável mais importante para diferenciá-los.O reduzido efeito da área dos fragmentos na compo-sição dos bandos heterogêneos pode ser explicado pelaassociação de várias espécies a estes bandos em todosos fragmentos estudados (Anexo 1). Isto sugere queparte das espécies que se associam a bandos mistos naárea de estudo apresenta uma baixa sensibilidade à frag-mentação e que estas podem se movimentar entre osfragmentos (Maldonado-Coelho & Marini, em prepa-ração).
A separação dos fragmentos quanto à composi-ção dos bandos pela estação possivelmente está relaci-onada simultaneamente à variação temporal na dispo-nibilidade de alimento e às atividades reprodutivas.Diversos trabalhos mostram que uma maior abundân-cia de insetos ocorre nos períodos de maiorpluviosidade (Davis 1945, Poulin et al. 1992, Develey& Peres 2000). Como a disponibilidade de alimento éum fator crucial governando as atividades reprodutivas
das aves, as espécies insetívoras ajustam o seu períodoreprodutivo com aquele de maior abundância deartrópodes, ou seja, durante o período chuvoso (Poulinet al. 1992, Young 1994, Develey & Peres 2000).
Em Viçosa, o período reprodutivo das aves co-incide com o período de maior pluviosidade (Figura 1),quando muitas espécies foram registradas em ativida-des associadas à nidificação na área de estudo, comocópula, construção de ninhos, incubação e cuidadoscom a prole (obs. pess.). Estas atividades tendem a res-tringir as atividades das espécies apenas às proximida-des dos ninhos, dificultando a associação destas aosbandos mistos e conseqüentemente diminuindo o nú-mero de espécies participantes durante este período(Powell 1979, 1985, Develey & Peres 2000). Estudosdesenvolvidos na Mata Atlântica do Sudeste do Brasil(Davis 1946, Develey & Peres 2000) e na AméricaCentral (Powell 1979) igualmente verificaram uma di-minuição no número de espécies dos bandos mistosdurante a estação reprodutiva. Desta forma, uma dascausas para a variação sazonal na composição dos ban-dos estaria possivelmente relacionada com as ativida-des reprodutivas das espécies.
A maioria das espécies participantes dos bandosmistos heterogêneos nos fragmentos são predominan-temente insetívoras. Devido a este fato, e uma vez quea associação de um indivíduo a um bando misto tendea maximizar seu forrageio (Sullivan 1984, Powell 1985,Hino 1998), seria de se esperar que as espécies se asso-ciariam aos bandos mistos com maior freqüência du-rante épocas de baixa disponibilidade de insetos. Sen-do assim, isto também explicaria em parte a variaçãosazonal encontrada na composição dos bandos hete-rogêneos em Viçosa, com um incremento no númerode espécies e na freqüência de participação destas du-rante a estação seca.
Adicionalmente, a presença das espécies visitan-tes de verão nos bandos mistos heterogêneos durantea estação chuvosa, contribuindo para a diferença nacomposição destes bandos entre as estações, pode sercorroborada por outros estudos realizados no Sudestedo Brasil, que registraram estas espécies na região du-rante este período (Willis 1979, Machado 1997, Perei-ra et al. 1998). Entretanto, as populações de V. olivaceuse P. polychopterus aparentam ser apenas parcialmentemigratórias na região, uma vez que alguns indivíduosforam registrados (com menor freqüência) associadosaos bandos durante os meses secos.
Durante a estação chuvosa, a diferença na com-posição dos bandos apresentada pelo fragmento de3,6 ha se deve possivelmente à reduzida área deste frag-mento para as espécies residentes e migratórias de ve-

42 MALDONADO-COELHO & MARINI: BANDOS MISTOS DE AVES
rão sustentarem territórios reprodutivos, similarmen-te ao verificado por Maldonado-Coelho & Marini(2000) para bandos mistos observados em um frag-mento florestal de 1,7 ha, em outra localidade no Su-deste de Minas Gerais. A ocorrência de A. fuscus nosbandos deste fragmento pode ser explicada pela pre-sença de uma mancha de taquaras (Merostachys sp.) nesteremanescente florestal. Ribon (1998) registrou um in-divíduo de A. fuscus em um fragmento de 6,6 ha emapenas uma ocasião na mesma área de estudo, e suge-riu que este estaria em trânsito no fragmento, ou seriaum indivíduo que teria sido isolado durante o proces-so de fragmentação. No presente estudo é impossívelsaber se os indivíduos de A. fuscus estariam utilizandomais de um fragmento, mas a existência desta manchade taquaral no fragmento de 3,6 ha pode explicar aprovável persistência de A. fuscus. A espécie é altamen-te especializada em forragear neste micro-habitat(Rodrigues et al. 1994), e em todos fragmentos ondefoi observada em Viçosa, a espécie mostrou uma gran-de associação com taquarais (obs. pess.). Os indivídu-os de A. fuscus observados no fragmento de 3,6 hapodem ser remanescentes de uma população originalou residentes temporários que imigraram de outrosfragmentos maiores, possivelmente não se reproduzin-do, uma vez que locais propícios para a nidificação(cavidades e barrancos) não estão presentes (obs. pess.).Adicionalmente, estes resultados mostram que a pre-sença de determinados habitats é crucial para a persis-tência de espécies habitat-especialistas em fragmentosflorestais, podendo ser este fator, em algumas situa-ções, mais importante do que a área dos fragmentos.Marini (2001) observou que as espécies de aves de-pendentes de floresta da região do cerrado foram sen-sitivas à área dos fragmentos, mas que as espécies de-pendentes de floresta endêmicas do cerrado, não fo-ram sensitivas à área, mas à presença de florestas inun-dadas típicas deste bioma.
Em relação aos bandos dos demais fragmentos,ainda que os mesmos tenham apresentado elevada si-milaridade na composição, foi observada uma dimi-nuição na freqüência de associação (e.g. D. mentalis,T. melanops) ou na ausência (e.g. L. fuscus, Herpsilochmusrufimarginatus) de algumas espécies nos três fragmen-tos menores, quando comparados aos seis maiores.Exemplos similares considerando a perda de espéciesem bandos mistos observados em fragmentos flores-tais foram detectados em outros estudos na regiãoneotropical (Rappole & Morton 1985, Stotz 1993,Maldonado-Coelho & Marini 2000). Ainda que algunsestudos tenham observado o desaparecimento de ban-dos mistos em fragmentos de 10 ha após o isolamento
destes da floresta contínua (Bierregaard & Lovejoy1989, Stotz 1993), alguns fatores que poderiam expli-car a existência destas associações nos fragmentos pe-quenos, seriam as proximidades entre os fragmentos, adisponibilidade de recursos e a presença das espéciesnucleares nestes (Maldonado-Coelho 2000,Maldonado-Coelho & Marini 2000).
A diferença na composição dos bandos do frag-mento de 9,4 ha, devido à presença de C. cinereus eL. angustirostris, pode ser explicada, no caso de C. cinereuspela chegada de indivíduos visitantes de inverno naregião. Segundo Willis (1979), esta espécie inverna emlocais baixos, passando os meses de verão em áreasmais altas no Sudeste do Brasil. Porém, as observa-ções indicam que alguns indivíduos desta espécie resi-dem também nesta área durante o verão (fragmentode 384,5 ha, estação chuvosa, Anexo 1), sendo que oaumento de sua freqüência nos bandos no fragmentode 9,4 ha no inverno possivelmente se deve à chegadade outros indivíduos provenientes de locais mais altosna região, como por exemplo, o Parque Estadual daSerra do Brigadeiro.
A presença de L. angustirostris nos bandos do frag-mento de 9,4 ha constitui um caso a parte. Esta espé-cie possui seu centro de distribuição no bioma cerra-do (Silva 1995), sendo uma das espécies mais comunsem bandos mistos no Brasil Central (Alves & Cavalcanti1996). Provavelmente colonizou a área recentemente,uma vez que este é o primeiro registro paraL. angustirostris na região (Ribon, comun. pess.), ondeestaria provavelmente acompanhando o desmatamentoda Mata Atlântica. Atribui-se ao mesmo processo ainvasão de outros taxa de cerrado em regiões de MataAtlântica no estado de São Paulo (Alvarenga 1990,Willis 1991).
Pode-se sugerir aqui que a composição dos ban-dos heterogêneos estaria refletindo o conjunto de espé-cies presente nos fragmentos, como mostrado por Latta& Wunderle (1996), ao encontrarem uma correlaçãoentre a composição de espécies em pontos de escuta e acomposição específica dos bandos. Adicionalmente,Hutto (1994) e Aleixo (1997) mostraram que a compo-sição dos bandos era um reflexo da abundância das es-pécies na comunidade, ou seja, a freqüência de ocorrên-cia de uma espécie no bando refletiu sua abundânciarelativa no habitat. Entretanto, esta relação deve ser vis-ta com certa cautela neste estudo, pois, ainda que a fre-qüência de algumas espécies nos bandos mistos possaestar correlacionada com a sua abundância nos frag-mentos, é possível que a freqüência de outras espéciesseja explicada, por exemplo, por atividades reprodutivasou pela disponibilidade local de recursos.

PAP. AVULS ZOOL. 43(3), 2003 43
O fato da composição dos bandos entre frag-mentos de diferentes tamanhos ter sido similar emambas as estações parece corroborar a hipótese de queas comunidades em pequenos fragmentos não sãoamostras aleatórias de fragmentos maiores (Blake 1991,Newmark 1991, Andrén 1994, Warburton 1997), su-gerindo que parte das espécies participantes dos ban-dos heterogêneos possivelmente é menos afetada pelafragmentação.
Análises das categorias ecológicasO padrão encontrado para a análise envolvendo
a proporção de espécies dependentes de floresta asso-ciadas aos bandos heterogêneos não corrobora o ob-servado por outros estudos conduzidos em fragmen-tos florestais no Estado de Minas Gerais, que detecta-ram um aumento no número de espécies independen-tes (Christiansen & Pitter 1997) e semidependentes(Marini 2001) de floresta com a redução na área dosfragmentos. Como a razão entre a área de interior e aárea de borda diminui com a redução na área dos frag-mentos, seria de se esperar uma diminuição na pro-porção de espécies dependentes de floresta nos ban-dos heterogêneos observados nos pequenos fragmen-tos. O padrão encontrado neste estudo está relaciona-do ao fato de que espécies independentes esemidependentes de floresta se associam aos bandosheterogêneos em todos os fragmentos. Além disso,poucas espécies dependentes e semidependentes defloresta estiveram restritas aos bandos nos três frag-mentos menores (e.g. Myiophobus fasciatus, Lepidocolaptesangustirostris e Phacellodomus rufifrons).
O incremento na proporção de espécies de copanos bandos heterogêneos com o aumento na área dosfragmentos na estação chuvosa pode estar relaciona-do a vários fatores. Como exemplo, pode-se sugerir aocorrência de um maior número de espécies perten-centes a esta categoria ou uma maior disponibilidadede recursos nestes fragmentos. Para as demais catego-rias de estrato de forrageamento (espécies de médioestrato e sub-bosque), ainda que a riqueza destas espé-cies seja maior nos grandes fragmentos (Maldonado-Coelho, obs. pess.), isto não estaria afetando a compo-sição dos bandos heterogêneos.
Alguns estudos realizados na Mata Atlânticamostraram que espécies de aves de sub-bosque,insetívoras de folha, insetívoras de tronco e frugívorassão afetadas pela fragmentação (Willis 1979, Aleixo &Vielliard 1995, Anjos & Boçon 1999). Entretanto, aausência de variação na proporção das espécies asso-ciadas aos bandos heterogêneos nestas categorias coma área dos fragmentos não implicam que estas guildas
são pouco afetadas pela fragmentação na área de estu-do. Por exemplo, ainda que não tenham sido detecta-das variações significativas na guilda de insetívoros detronco, duas espécies de arapaçus (L. fuscus e X. albicollis)não foram observadas nos três menores fragmentosnem dentro nem fora dos bandos mistos. Estas duasespécies também não foram detectadas em pequenosfragmentos florestais naturais no estado do Paraná(Anjos & Boçon 1999).
Bandos de sub-bosque
Análise de correspondênciaApesar do objetivo principal deste trabalho ser
avaliar o efeito da área dos fragmentos na composiçãodos bandos, o fato dos bandos de sub-bosque teremsido observados em apenas três fragmentos impedeque os resultados para este tipo de bando sejamextrapolados para todos os fragmentos da região, de-vendo os mesmos serem analisados com cautela. Den-tre os possíveis fatores que poderiam influenciar a com-posição dos bandos de sub-bosque, pode-se citar a di-ferença na área dos fragmentos, que estaria afetando aabundância das espécies. Por exemplo, a abundânciapossivelmente maior de A. leucophthalmus no fragmen-to de 384,5 ha seria responsável por sua diferenciaçãoem relação aos fragmentos de 75 e 38,8 ha. Algunstrabalhos realizados na Mata Atlântica mostraram, porexemplo, uma diferença na abundância desta espécieentre fragmentos de tamanhos distintos (Willis 1979,Christiansen & Pitter 1997). Outros estudos em flo-restas tropicais (Bierregaard & Lovejoy 1989, Willsonet al. 1994, Stouffer & Bierregaard 1995, Ribon 1998)e temperadas (Lynch & Whigan 1984, Blake & Karr1987) igualmente registraram variações na abundânciae distribuição das espécies entre fragmentos de dife-rentes áreas. Entretanto, outras características não re-lacionadas à área dos fragmentos que explicariam estadiferença na composição dos bandos de sub-bosquepoderiam ser atribuídas por exemplo à disponibilida-de de recursos e dispersão dos indivíduos dentro e entreos fragmentos.
Algumas explicações para a variação sazonal emum mesmo fragmento poderiam estar relacionadas,novamente, com a disponibilidade de recursos (e.g. in-setos), juntamente com atividades reprodutivas dasespécies (Powell 1979, Develey & Peres 2000).
A função de H. rubica como espécie nuclear e lí-der dos bandos mistos tem sido documentada tam-bém em outros estudos da Mata Atlântica (Bencke1996, Develey 1997). Sua importância para a forma-

44 MALDONADO-COELHO & MARINI: BANDOS MISTOS DE AVES
ção e coesão dos bandos de sub-bosque pode serfortalecida pelo fato de que a presença destas associa-ções nos fragmentos está condicionada à ocorrênciadesta espécie (Maldonado-Coelho & Marini, em pre-paração).
Bandos mistos de composição similar (associa-ções entre H. rubica, L. fuscus e outras espécies) ocor-rem em várias localidades na Mata Atlântica, comoSanta Catarina e Rio Grande do Sul (Bencke 1996),São Paulo (Aleixo 1997, Develey 1997), Espírito Santo(Stotz 1993, obs. pess.) e em outras localidades no es-tado de Minas Gerais (obs. pess.), indicando que a as-sociação entre estas espécies é bastante comum nestebioma. A associação destas espécies nos bandos emlocalidades geográficas distintas pode estar relaciona-da a características comportamentais relacionadas aomimetismo (Willis 1989, Aleixo 1997, Develey 1997)e/ou à função coesiva e de sentinela de H. rubica.
Análises das categorias ecológicasNeste estudo, os bandos de sub-bosque foram
observados estritamente em áreas afastadas da borda(> 50 m) nos três fragmentos florestais, o que certa-mente reflete na predominância de espécies de interi-or de f lorestas associadas aos mesmos e nahomogeneidade na composição para a categoria de-pendência florestal. A preferência dos bandos de sub-bosque por áreas de interior de floresta reflete o com-portamento da espécie líder e nuclear destes bandos(H. rubica), que evita as proximidades da borda dos frag-mentos para forragear (Maldonado-Coelho 2000).Willis (1960) também observou o mesmo padrão decomportamento para esta espécie na América Central.
A similaridade nas proporções de espécies emdiferentes classes nas categorias estrato deforrageamento e características de forrageamento, nãoindica que os bandos de sub-bosque foram altamenteestáveis quanto à composição. O que estes resultadosmostram é que as proporções de espécies nas catego-rias ecológicas não se alteram entre os três fragmen-tos, não sendo o mesmo verdadeiro quando os mes-mos são comparados quanto à freqüência das espéci-es, como previamente demonstrado. Isto revela queparâmetros distintos em análises de composição debandos mistos em fragmentos florestais devem serconsiderados.
Bandos de copa
A raridade dos bandos mistos de copa nos frag-mentos na área de estudo não está relacionada ao efei-
to da fragmentação, mas sim à grande propensão dasespécies deste estrato a se associarem aos bandos he-terogêneos, tornando infreqüentes desta forma, asso-ciações compostas unicamente por espécies de copa.Stotz (1993) e Aleixo (1997), também observaram quebandos de copa bem definidos não ocorrem no Espí-rito Santo e em São Paulo. Este fato pode ser explica-do pela baixa estatura da vegetação, o que permitiriaque espécies de copa e dos estratos médio-inferior seassociassem, impedindo uma diferenciação mais acen-tuada, como ocorre na Amazônia (Munn 1985).
A composição dos bandos de copa aparentemen-te não é afetada pela área dos fragmentos, uma vezque as espécies que participam com maior freqüênciadeste tipo de associação (e.g. H. ruficapilla, C. speciosum,D. cayana e T. cyanoventris) foram regularmente obser-vadas em todos os fragmentos estudados. Estas espé-cies utilizam com freqüência a matriz de vegetaçãocircundante aos fragmentos, e desta forma a área dosfragmentos deve ter pouca influência nas suas distri-buições na paisagem, como sugerido por outros estu-dos (Willis 1979, Bierregaard & Stouffer 1997).
Similarmente aos bandos heterogêneos, a com-posição dos bandos de copa aparentemente é mais in-fluenciada pela estação do ano do que pela área dosfragmentos. Uma vez que grande parte destas espéciesapresenta uma grande proporção de frutos em sua di-eta (Rodrigues 1995, Sick 1997), e, como na região deViçosa, o pico de produtividade de frutos ocorre du-rante o final da estação seca e começo da estação chu-vosa (L.C. Marangon, com. pess.), uma das possíveisexplicações para a variação na composição dos ban-dos de copa seria uma mudança sazonal na dieta des-tas espécies, que se alimentariam principalmente deinsetos na estação seca, quando se associam aos ban-dos para maximizar o forrageio.
CONCLUSÕES
Os resultados do presente trabalho mostram quea área dos fragmentos exerce pouca influência sobre acomposição dos bandos heterogêneos, sendo a esta-ção do ano uma variável mais importante. O reduzidonúmero de fragmentos nos quais os bandos de sub-bosque foram observados impossibilita que diferen-ças na composição destas associações sejam avaliadassatisfatoriamente em relação à área dos fragmentos.Entretanto, a sazonalidade influencia a composiçãodesta associação de forma menos intensa, mostrandoque os bandos de sub-bosque e heterogêneos respon-dem independentemente a diferentes variáveis.

PAP. AVULS ZOOL. 43(3), 2003 45
Algumas espécies como H. rubica, L. fuscus,A. fuscus, T. melanops e B. culicivorus, apresentam alta fre-qüência de associação aos bandos mistos na área deestudo. Todas estas espécies são dependentes de áreasflorestadas e podem passar a maior parte da vida asso-ciadas a bandos mistos, sendo sua sobrevivência pos-sivelmente relacionada à existência destas associaçõesmulti-específicas. Como a região se encontra inteira-mente fragmentada, impossibilitando a conservação deáreas florestais contínuas, sugere-se que se mantenhaa maior área possível de cobertura vegetal dos frag-mentos juntamente com uma reduzida distância entreestes. Isto permitiria a existência de uma maior rique-za de espécies e facilitaria a dispersão dos organismosentre os fragmentos, favorecendo a manutenção deinterações biológicas como bandos mistos de aves. Esteestudo demonstra como a compreensão da ecologiados bandos mistos pode ajudar tanto a aumentar oconhecimento da biologia e comportamento das es-pécies participantes como a fornecer informações im-portantes em estratégias de conservação em paisagensfragmentadas.
AGRADECIMENTOS
M.M.C. recebeu financiamento do Fundo Mundi-al para a Natureza (WWF) e bolsa de estudos do Conse-lho Nacional de Desenvolvimento Científico eTecnológico (CNPq). M.A.M possuía uma Bolsa de Pro-dutividade em Pesquisa do CNPq. M.A.S. Alves, M.Rodrigues, R. Durães e dois revisores anônimos realiza-ram valiosas sugestões ao manuscrito. R. Durães e R.Ribon auxiliaram em várias etapas deste trabalho. Osautores são gratos aos proprietários rurais pela autori-zação em conduzir este estudo em suas propriedades.
RESUMO
A fragmentação de habitats tem sido um dos assuntosmais discutidos na biologia da conservação nos últimos anos.Entretanto, poucos estudos tem avaliado os efeitos da fragmen-tação de florestas em bandos mistos de aves. Esses bandos sãoassociações de duas ou mais espécies, cuja coesão está relacionadaà cadeia de interações entre os membros dos bandos. As duasvantagens seletivas invocadas para explicar a evolução do com-portamento de viver em bandos mistos de espécies são o declíniono risco de predação e aumento na eficiência no forrageamento.O objetivo do presente estudo foi analisar a composição de ban-dos mistos em relação ao tamanho dos fragmentos de Mata Atlân-tica durante as estações seca e chuvosa, na região da Zona da
Mata, sudoeste do Estado de Minas Gerais, Brasil. Foramidentificados três tipos distintos de bandos mistos na área estu-dada: bandos heterogêneos, bandos do sub-bosque e bandos decopa. O primeiro foi observado em todos os fragmentos florestais,enquanto bandos do sub-bosque foram observados em apenastrês fragmentos. Bandos de copa são raros em todos os fragmen-tos. Algumas espécies participaram de dois tipos de bandos.Trichothraupis melanops e Basileurus culicivorus foramas espécies nucleares dos bandos heterogêneos. Habia rubica foia espécie mais importante na formação e coesão de bandos dosub-bosque. Em bandos de copa, nenhuma das espécies apresen-tou características similares. Analisamos aqui o efeito da esta-ção e da área do fragmento florestal na composição de bandosheterogêneos e de sub-bosque, baseados em dois enfoques: 1) -freqüência de espécies e 2) - características ecológicas das espéciesdo bando. Em relação à freqüência das espécies, a sazonalidadeafetou a composição de bandos heterogêneos mais fortemente quea área do fragmento. Apenas dois fragmentos florestais diferi-ram levemente na composição de bandos heterogêneos. Tanto aárea quanto a sazonalidade influenciaram a composição de ban-dos do sub-bosque. Excluindo a categoria de espécies de dosseldos bandos heterogêneos, que decresceu com a área do fragmentona estação chuvosa, a proporção geral de espécies nas categoriasdependentes da floresta, estrato de forrageamento e característi-cas de forrageamento, não se alterou em relação à área do frag-mento florestal ou estação, tanto para bandos heterogêneos quan-to para bandos do sub-bosque.
PALAVRAS-CHAVE: Floresta Atlântica, Brasil,fragmentação florestal, efeitos sazonais, composiçãode bandos mistos.
REFERÊNCIAS
Aleixo, A. 1997. Composition of mixed-species bird flocks andabundance of flocking species in a semideciduous forest ofsoutheastern Brazil. Ararajuba, 5:11-18.
Alvarenga, H.M.F. 1990. Novos registros e expansões geográficasde aves no leste do estado de São Paulo. Ararajuba, 1:115-117.
Alves, M.A.S. & Cavalcanti, R.B. 1996. Sentinel behavior, seasonality,and the structure of bird flocks in Brazilian savanna. OrnitologiaNeotropical, 7:43-51.
Andrén, H. 1994. Effects of habitat fragmentation on birds andmammals in landscapes with different proportions of suitablehabitat: a review. Oikos, 71:355-366.
Anjos, L. & Boçon, R. 1999. Bird communities in natural forestpatches in southern Brazil. Wilson Bulletin, 111:397-414.
Bencke, G.A. 1996. New records and habitats of the White-browedfoliage Gleaner (Anabacerthia amaurotis) (Passeriformes:Furnariidae) in Rio Grande do Sul, southern Brazil. Ararajuba,4:24-28.
Bierregaard Jr., R.O. & Lovejoy, T.E. 1989. Effects of forestfragmentation on Amazonian understory bird communities.Acta Amazonica, 19:215-241.

46 MALDONADO-COELHO & MARINI: BANDOS MISTOS DE AVES
Bierregaard Jr., R.O. & Stouffer, P.C. 1997. Understory birds anddynamic habitat mosaics in Amazonian rainforests. In: Laurance,W.F. & Bierregaard Jr., R.O. (eds.). Tropical forest remnants: ecology,management and conservation of fragmented communities. Chicago, TheUniversity of Chicago Press. p.138-155.
Blake, J.G. & Karr, J.R. 1987. Breeding birds of isolated woodlots:area and habitat relationships. Ecology, 68:1724-1734.
Blake, J.G. 1991. Nested subsets and the distribution of birds onisolated woodlots. Conservation Biology, 5:58-66.
Buskirk, W.H. 1976. Social systems in a tropical forest avifauna.American Naturalist, 110:293-310.
Christiansen, M.C. & Pitter, E. 1997. Species loss in a forest birdcommunity near Lagoa Santa in southeastern Brazil. BiologicalConservation, 80:23-32.
Davis, D.E. 1945. The anual cycle of plants, mosquitoes, birds, andmammals in two Brazilian forests. Ecological Monographs,15:243-295.
Davis, D.E. 1946. A seasonal analysis of mixed flocks of birds inBrazil. Ecology, 27:168-181.
Develey, P.F. 1997. Ecologia de bandos mistos de aves de Mata Atlânticana Estação Ecológica da Juréia-Itatins. São Paulo, Instituto deBiociências Universidade de São Paulo. 75p. (Tese deMestrado)
Develey, P.F. & Peres, C.A. 2000. Resource seasonality and thestructure of mixed species bird flocks in coastal Atlantic Forestof southeastern Brazil. Journal of Tropical Ecology, 16:33-53.
Espartel, L. 1977. Curso de Topografia. Porto Alegre, Editora Globo.655p.
Gradwohl, J. & Greenberg, R. 1980. The formation of antwrenflocks on Barro Colorado Island, Panamá. Auk, 97:385-395.
Gram, W.K. 1998. Winter participation by Neotropical migrant andresident birds in mixed-species flocks in northeasthern Mexico.Condor, 100:44-53.
Hill, M.O. & Gauch, H.G. 1980. Detrend correspondence analysis,an improved ordination technique. Vegetatio, 42:47-58.
Hino, T. 1998. Mutualistic and commensal organization of avianmixed-species foraging flocks in a forest of western Madagascar.Journal of Avian Biology, 29:7-24.
Hutto, R.L. 1994. The composition and social organization ofmixed-species flocks in a tropical decidous forest in WesternMexico. Condor, 96:105-118.
Jullien, M. & Thiollay, J.M. 1998. Multi-species territoriality anddynamic of neotropical forest understorey bird flocks. Journalof Animal Ecology, 67:227-252.
Kovac, W.L. 1986. A multivariate statistcs package for the IBM PC andcompatibles.
Latta, S.C. & Wunderle Jr., J.M. 1996. The composition and foragingecology of mixed-species flock in pine forests of Hispaniola.Condor, 98:595-607.
Leck, C.F. 1979. Avian extinctions in an isolated tropical wet-forestpreserve, Ecuador. Auk, 96:343-352.
Lynch, J.F. & Whigham, D.F. 1984. Effects of forest fragmentationon breeding bird communities in Maryland, USA. BiologicalConservation, 28:287-324.
Lovejoy, T.E.; Bierregaard Jr., R.O.; Rylands, A.B.; Malcolm, J.B.;Quintela, C.; Harper, L.H.; Brown Jr., K.S.; Powell, A.H.; Powell,G.V.N.; Schubart, H.O.R. & Hays, M.B. 1986. Edge and othereffects of isolation on Amazon forest fragments. In: Soulé,M.E. (ed.). Conservation biology. The science of scarcity and diversity.Sunderland, MA, Sinauer. p.257-285.
Ludwig, J.A. & Reynolds, J.F. 1988. Statistical Ecology: a primer onmethodsand computing. New York, John Wiley and Sons. 337p.
Machado, C.G. 1997. Vireo olivaceus (Vireonidae): uma espéciemigratória nos bandos mistos de aves na Mata Atlântica dosudeste brasileiro. Ararajuba, 5:60-62.
Machado, C.G. 1999. A composição dos bandos mistos de aves naMata Atlântica da Serra da Paranapiacaba, no sudeste brasileiro.Revista Brasileira de Biologia 59:75-85.
Maldonado-Coelho, M. 2000. Efeitos da fragmentação florestal em bandosmistos de aves de Mata Atlântica, no sudeste de Minas Gerais. BeloHorizonte, Universidade Federal de Minas Gerais. (Tese deMestrado)
Maldonado-Coelho, M. & Marini, M.A. 2000. Effects of forestfragment size and successional stage on mixed-species birdflocks in southeastern Brazil. Condor, 102:585-594.
Marini, M.A. 2001. Effects of forest fragmentation on birds of theCerrado region, Brazil. Bird Conservation International, 11:13-25.
Morrison, M.L.; With, K.A.; Timossi, I.C. & Milne, K.A. 1987.Composition and temporal variation of flocks in the SierraNevada. Condor, 89:739-745.
Morse, D.H. 1977. Feeding behaviour and predator avoidance inheterospecific groups. Bioscience, 27:332-334.
Moynihan, M. 1962. The organization and probable evolution ofsome mixed species flocks of Neotropical birds. SmithsonianMiscellaneous Collection, 143:1-140.
Munn, C.A. 1985. Permanent canopy and understory flocks inAmazonia: species composition and population density. In:Buckley, P.A.; Foster, M.S.; Morton, E.S.; Ridgely, R.S. &Buckley, F.G.(eds.). Neotropical Ornithology. Washington, AmericanOrnithologist’s Union. p.683-712. (Ornithological Monographs,36)
Munn, C.A. & Terborgh, J.W. 1979. Multi-species territoriality inNeotropical foraging flocks. Condor, 81:338-344.
Newmark, W.D. 1991. Tropical forest fragmentation and the localextinction of understory birds in the Eastern UsambaraMountains, Tanzania. Conservation Biology, 5:67-78.
Pereira, L.E.; Suzuki, A.; Souza, R.P.; Souza, M.F.C. & Flauto, G.1998. Sazonalidade das populações de Vireo olivaceus (Linnaeus,1766) (Aves, Vireonidae) em regiões da Mata Atlântica doEstado de São Paulo, Brasil. Ararajuba, 2:117-122.
Poulin, B.; Lefebvre, G. & McNeil, R. 1992. Tropical avian phenologyin relation to abundance and explotation of food resources.Ecology, 73:2295-2309.
Powell, G.V.N. 1979. Structure and dynamics of interspecific flocksin a Neotropical mid-elevation forest. Auk, 96:375-390.
Powell, G.V.N. 1985. Sociobiology and adaptative significance ofinterespecific foraging flocks in the Neotropics. In: Buckley, P.A.;Foster, M.S.; Morton, E.S.; Ridgely, R.S. and Buckley, F.G.(eds.).Neotropical Ornithology. Washington, American Ornithologist’sUnion. p.713-732 (Ornithological Monographs 36)
Rappole, J.H. & E. Morton. 1985. Effects of habitat alteration on atropical avian forest community. In: Buckley, P.A.; Foster, M.S.;Morton, E.S.; Ridgely, R.S. and Buckley, F.G.(eds.). NeotropicalOrnithology. Washington, American Ornithologist’s Union.p.1013-1021. (Ornithological Monographs 36)
Ribon, R. 1998. Fatores que influenciam a distribuição da avifauna emfragmentos de Mata Atlântica nas montanhas de Minas Gerais. BeloHorizonte, Universidade Federal de Minas Gerais. (Tese deMestrado)
Rice, W.R. 1989. Analyzing tables of statistical tests. Evolution,43:223-225.
Rodrigues, M. 1995. Spatial distribution and food utilization amongtanagers in southeastern Brazil (Passeriformes: Emberezidae).Ararajuba, 3:27-32.

PAP. AVULS ZOOL. 43(3), 2003 47
Rodrigues, M.; Álvares, S.M.R. & Machado, C.G. 1994. Foragingbehaviour of the White-collared Foliage-gleaner (Anabazenopsfuscus), a bamboo specialist. Ornitologia Neotropical, 5:65-67.
Sick, H. 1997. Ornitologia Brasileira. Rio de Janeiro, Nova Fronteira.862p.
Sieving, K.E. & Karr, J.R. 1997. Avian extinction and persistencemechanisms in lowland Panama. In: Laurance, W.F. andBierregaard Jr., R.O. (eds.). Tropical forest remnants: ecology,management and conservation of fragmented communities. Chicago, TheUniversity of Chicago Press. p.138-155.
Silva, J.M.C. 1995. Birds of the Cerrado region, South America.Steenstrupia, 21:69-92.
Sokal, R.P. & Rohlf, F.J. 1998. Biometry New York, W.H. Freemanand Company. 887p.
SPSS. 1998. Version 9 ed. SPSS, Inc. Chicago, IL.Stotz, D.F. 1993. Geographic variation in species composition of
mixed species flocks in lowland humid forests in Brazil. PapéisAvulsos de Zoologia, 38:61-75.
Stouffer, P.C. & Bierregaard Jr., R.O. 1995. Use of Amazonian forestfragments by understory insectivorous birds. Ecology,76:2429-2445.
Sullivan, K.A. 1984. The advantages of social foraging in DownyWoodpeckers. Animal Behavior, 32:16-22.
Valverde, O. 1958. Estudo regional da Zona da Mata, de MinasGerais. Revista Brasileira de Geografia, 20:3-82.
Veloso, H.P.; Rangel Filho, A.L.R. &. Lima, J.C.A. 1991. Classificaçãoda vegetação brasileira, adaptada a um sistema universal. Rio de Janeiro,
IBGE, Departamento de Recursos Naturais e EstudosAmbientais. 124p.
Warburton, N.H. 1997. Structure and conservation of forest avifaunain isolated rainforest remnants in tropical Australia. In: Laurance,W.F. & Bierregaard Jr., R.O. (eds.). Tropical forest remnants: ecology,management and conservation of fragmented communities. Chicago, TheUniversity of Chicago Press. p.138-155.
Willis, E.O. 1960. A study of the foraging behaviour of two speciesof Ant-Tanagers. Auk, 77:150-170.
Willis, E.O. 1974. Populations and local extinctions of birds onBarro Colorado Island, Panamá. Ecological Monographs,44:153-169.
Willis, E.O. 1979. The composition of avian communities inremanescent woodlots in southern Brazil. Papéis Avulsos deZoologia, 33:1-25.
Willis, E.O. 1989. Mimicry in bird flocks of cloud forests insoutheastern Brazil. Revista Brasileira de Biologia, 49:615-619.
Willis, E.O. 1991. Expansão geográfica de Netta erythrophthalma,Fluvicola nengeta e outras aves de zonas abertas com a“desertificação” antrópica em São Paulo. Ararajuba, 2:101-102.
Willson, M.F.; De Santo, T.L.; Sabag, C. & Armesto, J.J. 1994. Aviancommunities of fragmented south-temperate rainforests inChile. Conservation Biology, 8:508-520.
Young, B.E. 1994. The effects of food, nest predation and weatheron the timing of breeding in tropical House Wrens. Condor,96:341-353.
Recebido em 09.01.2001Aceito em 26.11.2002
Credenciamento e apoio financeiro doPrograma de Apoio às PublicaçõesCientíficas Periódicas da USPComissão de Credenciamento Se
ção
de P
ublic
açõe
s do
Mus
eu d
e Zo
olog
ia d
a U
nive
rsid
ade
de S
ão P
aulo

48 MALDONADO-COELHO & MARINI: BANDOS MISTOS DE AVESAN
EXO
1
Freq
üênc
ias d
e As
soci
ação
das
Esp
écie
s nos
Ban
dos M
isto
s
ANE
XO 1.
Dist
ribui
ção
de f
reqü
ênci
a da
s es
péci
es r
egist
rada
s no
s ba
ndos
mist
os n
os n
ove
frag
men
tos,
dura
nte
as e
staç
ões
chuv
osa
e se
ca. A
s es
péci
es e
stão
org
aniz
adas
em
ord
em d
ecre
scen
te d
efr
eqüê
ncia.
As l
egen
das p
ara
os b
ando
s são
: H =
ban
dos h
eter
ogên
eos;
S =
ban
dos d
e su
b-bo
sque
e C
= b
ando
s de
copa
.
Frag
men
to (h
a)
Espé
cieBa
ndo
384.5
181.2
120
7545
.138
.89.4
7.63.6
Total
Total
Ger
alN
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
total
Basile
uteru
s culi
civoru
sH
80,0
84,2
100
82,3
84,6
100
92,3
88,8
93,3
100
100
78,5
100
93,7
58,3
73,3
5081
,285
,586
,877
,272
,211
915
127
0FD
, M, I
GS
68,7
55,5
00
00
84,6
41,1
00
53,8
53,3
00
00
00
42,8
50,0
C0
00
00
00
00
00
00
00
00
00
0Tr
ichoth
raupis
mela
nops
H10
0,010
010
076
,492
,366
,676
,977
,780
,073
,385
,771
,438
,475
,016
,656
,225
,031
,278
,862
,070
,852
,110
910
921
8FD
, M, O
NS
62,5
27,7
00
00
69,2
52,4
00
30,7
53,3
00
00
00
54,7
44,0
C0
00
00
00
00
00
00
00
00
00
0Le
ptopo
gon a
maur
oceph
alus
H60
,063
,185
,770
,553
,873
,353
,883
,386
,680
,057
,164
,28,3
18,7
16,6
60,0
043
,750
,062
,039
,647
,861
100
161
FD, M
, IF
S12
,516
,60
00
046
,135
,20
015
,40
00
00
00
23,8
18,0
C0
00
00
00
00
00
00
00
00
00
0To
lmom
yias s
ulphu
rescen
sH
40,0
57,8
28,5
76,4
46,1
86,6
30,7
83,3
26,6
86,6
42,8
92,8
8,331
,225
,060
,00
43,7
27,8
68,2
20,8
48,3
3210
113
3FD
, M, I
FS
6,05,5
00
00
15,4
00
00
6,60
00
00
07,1
4,0C
00
00
00
00
00
00
00
00
00
014
,2Th
amno
phillu
s caer
ulesce
nsH
30,0
63,1
50,0
57,8
30,7
60,0
38,4
38,8
26,6
60,0
14,1
78,5
16,6
68,7
16,6
53,3
50,0
43,7
30,7
58,6
26,7
43,5
4191
132
FD, S
, IF
S18
,716
,60
00
023
,011
,70
07,7
6,60
00
00
016
,612
,0C
00
00
00
00
00
00
00
00
00
00
Ana
bazen
ops f
uscus
H20
,047
,371
,429
,423
,020
,00
5,566
,660
,014
,07,1
00
8,30
87,5
31,2
32,6
22,0
40,2
29,6
6262
124
FD, S
, IF
S75
,077
,70
00
046
,15,8
00
76,9
100
00
00
00
66,6
60,0
C0
00
00
00
00
00
00
00
00
00
0H
yloph
ilus a
maur
oceph
alus
H25
,036
,871
,447
,030
,893
,315
,416
,653
,373
,30
57,1
41,7
43,7
58,3
60,0
12,5
56,2
39,4
52,4
26,1
36,8
4077
117
FD, M
, IF
S0
00
00
00
5,80
00
00
00
00
00
2,0C
00
00
033
,30
00
00
00
00
00
00
7,1X
enops
rutila
nsH
40,0
26,3
50,0
35,2
76,9
60,0
38,4
66,6
26,6
60,0
28,4
50,0
45,7
43,7
8,320
,00
12,5
36,5
40,6
30,5
32,5
4768
115
FD, M
, IT
S6,2
16,6
00
00
38,4
5,80
07,6
6,60
00
00
014
,210
,0C
33,3
010
0,050
,00
00
40,0
00
010
00
00
00
028
,528
,5D
ysitha
mnus
menta
lisH
30,0
63,1
28,6
58,8
38,5
93,3
23,0
44,4
26,6
46,6
14,2
64,2
00
020
,00
019
,243
,421
,538
,733
8111
4FD
, S, I
FS
37,5
38,8
00
00
46,0
41,1
00
7,626
,60
00
00
030
,936
,0C
00
00
00
00
00
00
00
00
00
00

PAP. AVULS ZOOL. 43(3), 2003 49Fr
agm
ento
(ha)
Espé
cieBa
ndo
384.5
181.2
120
7545
.138
.89.4
7.63.6
Total
Total
Ger
alN
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
total
Picum
nus c
irratu
sH
70,0
52,6
50,0
41,1
69,2
53,3
38,4
61,1
33,3
40,0
14,2
50,0
16,6
6,241
,653
,312
,546
,640
,364
,830
,032
,046
6711
3FS
, M, I
TS
00
00
00
15,3
5,80
00
6,60
00
00
04,7
2,0C
00
00
00
00
00
00
00
00
00
00
Syna
llaxi
s rufi
capilla
H20
,052
,628
,552
,946
,180
,07,6
50,0
60,0
60,0
14,2
28,5
012
,516
,626
,60
56,2
25,0
46,8
20,2
36,8
3177
108
FD, S
, IF
S12
,516
,60
00
030
,711
,70
00
26,6
00
00
00
14,2
18,0
C0
00
00
00
00
00
00
00
00
00
0Le
pidoco
laptes
squa
matus
H50
,031
,535
,741
,130
,820
,053
,872
,253
,313
,328
,628
,525
,056
,28,3
025
,050
,033
,942
,725
,431
,539
6610
5FD
, M, I
TS
00
00
00
00
00
06,6
00
00
00
2,37,1
C6,2
50
00
00
060
,00
050
,00
00
00
00
14,2
21,4
Lepid
ocolap
tes fu
scus
H10
,05,2
023
,57,6
00
06,6
00
7,10
00
00
02,8
2,826
,725
,841
5495
FD, S
, IT
S93
,710
00
00
076
,988
,20
010
010
00
00
00
090
,496
,0C
00
00
00
00
00
00
00
00
00
00
Hab
ia ru
bica
H0
00
00
00
00
00
00
00
00
00
027
,423
,942
5092
FD, S
, ON
S10
010
00
00
010
010
00
010
010
00
00
00
010
010
0C
00
00
00
00
00
00
00
00
00
00
Cycla
rhis g
ujanen
sisH
3047
,314
,241
,115
,453
,323
,027
,740
,046
,628
,642
,825
,056
,20
26,6
050
,020
,143
,413
,733
,021
6990
FS, M
, IF
S0
16,6
00
00
00
00
013
,30
00
00
00
10,0
C0
00
00
00
00
00
00
100
00
00
07,1
Camp
ylorha
mphu
s falc
ulariu
sH
05,2
017
,638
,553
,37,6
11,1
33,3
20,0
14,2
7,18,3
00
00
18,7
15,3
14,4
15,6
21,5
2445
69FD
, M, I
TS
2544
,40
00
023
,064
,70
030
,733
,30
00
00
026
,148
,0C
00
00
00
00
00
00
00
00
00
00
Conir
ostru
m spe
ciosum
H30
,026
,314
,223
,57,7
26,6
7,633
,30
6,60
42,8
053
,30
40,0
037
,56,7
31,7
7,125
,811
5465
FD, C
, IF
S0
00
00
00
00
00
00
00
00
00
0C
100
500
100
033
,30
80,0
00
50,0
00
00
00
057
,157
,1Py
rilegn
a leu
copter
aH
026
,342
,835
,215
,413
,315
,35,5
6,640
,014
,214
,20
18,7
8,353
,30
012
,522
,713
,720
,021
4263
FD, S
, IF
S31
,227
,70
00
015
,35,8
00
020
,00
00
00
016
,618
,0C
00
00
00
00
00
00
00
00
00
00
Hem
ithrau
pis ru
ficapil
laH
052
,60
29,4
00
38,4
70,5
00
28,5
21,4
00
00
06,2
6,721
,37,1
20,5
1143
54FD
, C, I
FS
00
00
00
00
00
00
00
00
00
00
C10
010
00
100
066
,60
100
00
100
100
00
00
00
57,1
85,7
Camp
tostom
a ob
soletu
mH
20,0
42,1
011
,70
33,3
7,638
,80
13,3
028
,50
25,0
00
050
,02,8
27,7
1,921
,53
4548
FI, C
, IG
S0
00
00
00
00
00
00
00
00
00
0C
00
00
033
,30
40,0
00
010
00
100
00
00
035
,7V
enillio
rnis
macul
ifron
sH
30,0
26,3
7,135
,27,6
33,3
15,4
33,3
00
14,2
28,5
8,36,2
020
,00
08,6
20,6
6,517
,210
3646
FD, M
, IT
S6,2
33,3
00
00
00
00
00
00
00
00
2,312
,0C
00
00
00
00
00
00
00
00
00
00

50 MALDONADO-COELHO & MARINI: BANDOS MISTOS DE AVESFr
agm
ento
(ha)
Espé
cieBa
ndo
384.5
181.2
120
7545
.138
.89.4
7.63.6
Total
Total
Ger
alN
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
total
Vire
o oliv
aceus
H40
,05,2
50,0
061
,50
23,0
040
,00
014
,225
,018
,78,3
6,60
12,5
30,7
0,621
,55,2
3311
44FD
, C, I
FS
00
00
00
00
00
00
00
00
00
00
C0
00
50,0
00
020
,00
00
00
00
00
00
14,2
Tang
ara cy
anove
ntris
H10
,026
,37,1
47,0
033
,330
,760
,013
,36,6
00
00
06,6
012
,57,6
15,8
8,414
,813
3144
FD, C
, ON
S0
00
00
00
00
00
00
00
00
00
0C
66,6
100
00
020
,00
100
00
500
00
00
00
42,8
57,1
Dacn
is cay
ana
H0
21,0
023
,50
13,3
038
,80
20,0
014
,20
31,2
00
031
,20
22,0
0,620
,01
4243
FS, C
, ON
S0
00
00
66,6
00
00
00
00
00
00
00
C33
,30
010
00
00
100
00
00
00
00
00
14,2
71,4
Piaya
caya
naH
10,0
42,1
029
,40
13,3
016
,60
20,0
014
,20
31,2
013
,30
37,5
0,924
,80,6
17,2
136
37FS
, M, I
FS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0A
utomo
lus le
ucoph
thalm
usH
05,2
21,4
29,4
30,7
33,3
00
00
00
00
16,6
33,3
00
6,711
,09,1
11,0
1423
37FD
, S, I
FS
25,0
38,8
00
00
00
00
7,70
00
00
00
11,9
14,0
C0
00
00
00
00
00
00
00
00
00
0Si
ttasom
us gri
seicap
illus
H10
,00
21,3
29,4
30,7
00
5,50
00
14,2
25,0
56,2
00
00
9,611
,78,4
9,013
1932
FD, M
, IT
S0
00
00
07,6
11,7
00
00
00
00
00
2,34,0
C0
00
00
00
00
00
00
00
00
00
0H
erpsilo
chmus
rufim
argin
atus
H0
26,3
00
15,4
46,6
00
20,0
40,0
042
,80
00
00
04,8
16,5
3,911
,46
2430
FD, M
, IF
S0
00
00
00
00
00
00
00
00
02,3
0C
00
00
00
00
00
00
00
00
00
00
Hem
itricc
us nid
ipend
ulus
H0
00
17,6
026
,60
00
20,0
014
,28,3
12,5
16,6
53,3
031
,22,8
18,6
1,912
,93
2730
FD, S
, IF
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
00
00
00
00
00
Pachy
ramph
us po
lycho
pteru
sH
25,0
5,214
,20
46,1
6,623
,05,5
20,0
014
,20
33,3
00
6,60
022
,12,7
16,3
1,925
429
FS, C
, IF
S0
00
00
00
00
00
00
00
00
00
0C
00
10,0
00
00
010
00
00
00
00
00
28,5
0To
dirost
rum
polio
cepha
lumH
10,0
31,5
011
,70
53,3
00
013
,30
35,7
00
00
025
,00,9
18,6
0,612
,91
2728
FS, M
, IF
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
00
00
00
00
00
Tang
ara ca
yana
H0
21,0
05,8
06,6
022
,20
13,3
00
8,331
,20
20,0
018
,70,9
15,8
0,611
,91
2526
FI, C
, ON
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
040
,00
00
00
00
00
00
14,7
Todir
ostru
m plu
mbeic
epsH
05,2
035
,27,6
33,3
011
,120
,020
,00
14,2
00
06,6
00
3,813
,77,6
9,54
2024
FD, S
, IF
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
00
00
00
00
00

PAP. AVULS ZOOL. 43(3), 2003 51Fr
agm
ento
(ha)
Espé
cieBa
ndo
384.5
181.2
120
7545
.138
.89.4
7.63.6
Total
Total
Ger
alN
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
total
Platy
rinchu
s myst
aceus
H0
15,7
011
,70
26,6
011
,10
6,60
21,4
00
06,6
06,2
011
,70
9,50
2020
FD, S
, IF
S0
11,1
00
00
05,8
00
00
00
00
00
06,0
C0
00
00
00
00
00
00
00
00
00
0Pa
chyram
phus
valid
usH
20,0
07,1
5,80
015
,30
00
57,1
00
00
00
08,6
0,65,8
0,49
1019
FD, C
, IF
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
00
00
00
00
00
Thrau
pis sa
yaca
H0
21,0
05,8
026
,60
11,1
00
00
012
,50
13,3
025
,00
13,1
09,9
019
19FS
, C, O
NS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0M
yiarch
us fer
oxH
05,2
30,8
023
,06,6
05,5
06,6
07,1
018
,70
26,6
06,2
6,76,8
5,24,7
810
18FS
, M, I
GS
00
00
00
00
00
00
00
00
00
00
C33
,30
00
00
00
00
00
00
00
00
28,5
0Pa
chyram
phus
viridi
sH
00
7,15,8
026
,67,6
00
6,60
14,2
00
013
,312
,518
,71,9
8,91,9
6,63
1417
FD, C
, IF
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
010
00
00
00
014
,27,1
Tachy
phon
us cor
onatu
sH
20,0
5,214
,211
,70
6,60
27,7
00
00
8,30
06,6
00
4,86,8
3,25,2
511
16FD
, S, O
NS
6,25,5
00
00
00
00
00
00
00
00
2,32,0
C0
00
00
00
00
00
00
00
00
00
0Ph
yllom
yias f
asciat
usH
010
,50
17,6
033
,30
00
00
00
00
20,0
00
08,9
06,6
014
14FS
, C, I
GS
00
00
00
00
00
00
00
00
00
00
C0
00
00
33,3
00
00
00
00
00
00
07,1
Myio
pagis
virid
icata
H30
,00
35,7
07,6
015
,40
00
00
8,30
8,30
00
12,5
09,1
014
014
FD, M
, IG
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
50,0
00
00
00
014
,20
Myio
pagis
canic
epsH
015
,714
,311
,77,6
6,60
6,60
00
00
00
13,3
00
2,87,5
1,95,2
311
14FD
, C, I
FS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0N
emosi
a pilea
taH
10,0
00
00
00
00
00
7,10
18,7
06,6
025
,00,9
6,20,6
5,21
1112
FD, C
, ON
S0
00
00
00
00
00
00
00
00
00
0C
00
010
00
00
00
00
00
00
00
00
14,2
Xiph
ocolap
tes a
lbicol
lisH
00
00
00
7,65,5
00
00
00
00
00
0,90,6
1,93,8
38
11FD
, M, I
TS
12,5
16,6
00
00
023
,50
00
00
00
00
04,7
14,0
C0
00
00
00
00
00
00
00
00
00
0Fo
rmiciv
ora se
rrana
H0
00
5,80
00
00
00
14,2
8,318
,70
20,0
06,2
0,96,6
0,64,7
111
12FD
, S, I
FS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0

52 MALDONADO-COELHO & MARINI: BANDOS MISTOS DE AVESFr
agm
ento
(ha)
Espé
cieBa
ndo
384.5
181.2
120
7545
.138
.89.4
7.63.6
Total
Total
Ger
alN
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
total
Mack
enziae
na se
vera
H0
10,5
14,2
11,7
06,6
00
6,60
00
00
06,6
012
,52,8
4,81,9
3,33
710
FD, S
, IF
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
00
00
00
00
00
Syna
llaxi
s cine
rascen
sH
010
,57,1
11,7
06,6
7,60
00
00
00
06,6
00
1,94,1
1,33,3
27
9FD
, S, I
FS
00
00
00
00
00
00
00
00
00
02,0
C0
00
00
00
00
00
00
00
00
00
0M
anacu
s man
acus
H10
,015
,70
23,5
00
00
00
00
00
06,6
00
0,95,5
0,63,8
18
9FD
, S, F
RS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0Co
ntopu
s cine
reus
H0
00
00
00
00
00
00
46,6
00
00
04,8
0,63,3
17
8FD
, C, I
AS
00
00
00
00
00
00
00
00
00
00
C33
,30
00
00
00
00
00
00
00
00
14,2
0Le
pidoco
laptes
ang
ustiro
stris
H0
00
00
00
00
00
08,3
31,2
00
00
0,93,4
0,62,3
15
6FI
, M, I
TS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0D
rymop
hyla
ferru
ginea
H0
15,7
00
00
00
00
7,614
,20
00
00
00
3,40,6
2,31
56
FD, S
, IF
S0
00
00
00
00
00
00
00
00
02,3
0C
00
00
00
00
00
00
00
00
00
00
Euph
onia
chloro
ticaH
00
00
06,6
00
00
14,2
7,10
00
00
6,20,9
2,00,6
2,31
56
FS, C
, FR
S0
00
00
00
00
00
00
00
00
00
0C
00
00
033
,30
00
00
100
00
00
00
014
,2Ph
acello
domu
s rufi
frons
H0
00
00
00
00
00
016
,60
020
,00
01,9
2,01,3
1,42
35
FS, S
, IF
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
00
00
00
00
00
Mega
rynchu
s pita
ngua
H0
5,20
00
00
5,50
00
00
00
00
12,5
02,7
02,3
05
5FS
, C, I
GS
00
00
00
00
00
00
00
00
00
00
C0
00
00
33,3
00
00
00
00
00
00
07,1
Pipra
edea
melan
onota
H0
5,20
11,7
00
05,5
00
00
00
06,6
00
03,4
02,3
05
5FD
, M, O
NS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0M
yiorn
is au
ricula
risH
00
14,3
5,87,6
00
00
00
00
00
00
02,8
0,61,9
0,43
14
FD, M
, IF
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
00
00
00
00
00
Coere
ba fla
veola
H0
5,20
00
6,60
5,50
00
00
00
6,60
00
2,70
1,90
44
FS, M
, ON
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
00
00
00
00
00

PAP. AVULS ZOOL. 43(3), 2003 53Fr
agm
ento
(ha)
Espé
cieBa
ndo
384.5
181.2
120
7545
.138
.89.4
7.63.6
Total
Total
Ger
alN
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
total
Colon
ia col
onus
H0
00
00
6,60
5,50
6,60
00
00
00
00
2,00
1,40
33
FD, C
, IA
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
00
00
00
00
00
Dryo
copus
lineat
usH
05,2
00
06,6
00
00
07,1
00
00
00
02,0
00,9
03
3FS
, M, I
TS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0Co
nopo
phaga
linea
taH
05,2
00
06,6
00
00
00
00
00
00
01,3
01,4
03
3FD
, S, I
FS
00
00
00
05,8
00
00
00
00
00
02,0
C0
00
00
00
00
00
00
00
00
00
0Th
raupis
palm
arum
H0
00
00
00
11,1
00
00
00
00
00
01,3
00,9
02
2FS
, C, O
NS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0Ca
mpeph
ilus r
obust
usH
00
00
00
05,5
06,6
00
00
00
00
01,3
00,9
02
2FD
, M, I
TS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0Ce
leus f
lavesc
ensH
00
05,8
00
05,5
00
00
00
00
00
01,3
00,9
02
2FD
, M, I
TS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0M
ionect
es ru
fivent
risH
10,0
07,1
00
13,3
7,60
00
00
00
07,2
6,60
2,82,0
3,20,9
02
2FD
, M, F
RS
12,5
00
00
00
00
00
00
00
00
04,7
0C
00
00
00
00
00
00
00
00
00
00
Myio
zetet
es sim
ilisH
00
00
00
05,5
00
00
00
00
012
,50
1,30
0,90
22
FI, C
, IG
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
00
00
00
00
00
Paru
la pit
yaium
iH
05,2
00
00
00
00
00
00
00
00
00,6
00,9
02
2FD
, C, I
FS
00
00
00
00
00
00
00
00
00
00
C0
00
50,0
00
00
00
00
00
00
00
07,1
Sirys
tes si
bilato
rH
010
,50
00
00
00
00
00
00
00
00
1,30
0,90
22
FD, C
, IG
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
00
00
00
00
00
Baryp
htheng
us ru
ficapil
lusH
00
00
00
00
00
00
00
00
00
00
0,60
10
1FD
, M, O
NS
6,25
00
00
00
00
00
00
00
00
02,3
0C
00
00
00
00
00
00
00
00
00
00
Cnem
otricc
us fus
catus
H0
00
00
00
00
00
00
08,3
00
00,9
00,6
01
01
FD, S
, IG
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
00
00
00
00
00

54 MALDONADO-COELHO & MARINI: BANDOS MISTOS DE AVES
Coccy
zus m
elacor
yphu
sH
00
00
00
7,60
00
00
00
00
00
0,90
0,60
10
1FS
, M, I
FS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0D
rymop
hila o
chrop
ygaH
00
00
00
00
6,60
00
00
00
00
0,90
0,60
10
1FD
, S, I
FS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0M
yiarch
us sw
ainson
iH
00
00
00
00
00
00
00
00
00
00
0,60
10
1FD
, M, I
GS
00
00
00
00
00
00
00
00
00
00
C0
00
00
07,6
00
00
00
00
00
014
,20
Phily
dor r
ufus
H0
00
00
00
00
00
00
00
00
00
00,6
01
01
FD, C
, IF
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
100
00
00
00
00
014
,20
Chiro
xiph
ia cau
data
H0
00
00
00
5,50
00
00
00
00
00
0,60
0,40
11
FD, M
, FR
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
00
00
00
00
00
Latho
triccu
s eule
riH
00
00
00
00
00
00
06,2
00
00
00,6
00,4
01
1FD
, M, I
GS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0M
yioph
obus
fascia
tusH
00
00
00
00
00
00
00
06,6
00
00,6
00,4
01
1FS
, M, I
GS
00
00
00
00
00
00
00
00
00
00
C0
00
00
00
00
00
00
00
00
00
0Tr
oglod
ytes a
edon
H0
00
00
00
00
00
00
6,20
00
00
0,60
0,40
11
FI, S
, IF
S0
00
00
00
00
00
00
00
00
00
0C
00
00
00
00
00
00
00
00
00
00
Atill
a rufu
sH
00
00
00
00
00
00
00
00
00
00
00,4
01
1FD
, M, I
GS
05,5
00
00
00
00
00
00
00
00
02,0
C0
00
00
00
00
00
00
00
00
00
0D
epen
dênc
ia flo
resta
l: FD
= d
epen
dent
e de f
lore
sta; F
S =
sem
idep
ende
nte d
e flo
resta
; FI =
inde
pend
ente
de fl
ores
ta.Es
trato
de f
orra
geam
ento
: C =
copa
; M =
méd
io-e
strato
; S =
sub-
bosq
ue.
Cara
cterís
ticas
de f
orra
geam
ento
: IG
= in
setív
oro
gene
ralis
ta; IF
= in
setív
oro
folh
a; IT
= in
setív
oro
tronc
o; IA
= in
setív
oro
ar; O
N =
oní
voro
; FR
= fru
gívor
o.
Frag
men
to (h
a)
Espé
cieBa
ndo
384.5
181.2
120
7545
.138
.89.4
7.63.6
Total
Total
Ger
alN
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
chuv
ase
cach
uva
seca
total

General Information: Papéis Avulsos de Zoologia covers primarily the fields of Zoology,publishing original contributions in systematics, paleontology, evolutionary biology, ecology,taxonomy, anatomy, behavior, functional morphology, molecular biology, ontogeny, faunisticstudies, and biogeography. Papéis Avulsos de Zoologia also encourages submission of theoreticaland empirical studies that explore principles and methods of systematics.
All contributions must follow the International Code of Zoological Nomenclature. Relevantspecimens should be properly curated and deposited in a recognized public or private,non-profit institution. Tissue samples should be referred to their voucher specimens andall nucleotide sequence data (aligned as well as unaligned) should be submitted to GenBank(http://www.ncbi.nih.gov/Genbank/) or EMBL (http://www.ebi.ac.uk/).
Peer Review: All submissions to Papéis Avulsos de Zoologia are subject to review by at leasttwo referees and the Editor-in-Chief. Three legible copies (including photocopies of originalillustrations) and original illustrations must be submitted; all authors will be notified ofsubmission date. Authors may suggest potential reviewers. Communications regardingacceptance or rejection of manuscripts are made through correspondence with the firstor corresponding author only. Once a manuscript is accepted providing changes suggestedby the referees, the author is requested to return a revised version incorporating thosechanges (or a detailed explanation of why reviewer’s suggestions were not followed) withinfour weeks upon receiving the communication by the editor. Revised manuscripts must besubmitted as both hard copy and electronic file (3.5" disk, Zip Drive, or CD Rom withtext in Microsoft Word format). Figures and graphics should be sent separately (“.jpg”,“.tif ”, “.xls”, “.cdr”).
Proofs: Page-proofs with the revised version will be sent to the first or correspondingauthor. Page-proofs must be returned to the editor in two weeks, preferentially within 48 hours.Failure to return the proof promptly may be interpreted as approval with no changesand/or may delay publication. Only necessary corrections in proof will be permitted.Once page proof is sent to the author, further alterations and/or significant additions oftext are permitted only at the author’s expense or in the form of a brief appendix (“noteadded in proof ”).
Submission of Manuscripts: Manuscripts should be sent to the Editor-in-Chief(H. Zaher, Museu de Zoologia da USP, Caixa Postal 42.494, CEP 04218-970, São Paulo,SP, Brasil). Manuscripts are considered on the understanding that they have not beenpublished or will not appear elsewhere in substantially the same or abbreviated form. Thecriteria for acceptance of articles are: quality and relevance of research, clarity of text, andcompliance with the guidelines for manuscript preparation.
Manuscripts should be written preferentially in English, but texts in Portuguese or Spanishwill also be considered. Studies with a broad coverage are encouraged to be submitted inEnglish. All manuscripts should include an abstract in Portuguese and English regardlessof the original language.
Authors are requested to pay attention to the instructions concerning the preparation ofthe manuscripts. Close adherence to the guidelines will expedite processing of themanuscript, whereas manuscripts deviating from the required form will be returned forrevision prior to review.
Manuscript Form: Manuscripts should not exceed 100 pages of double-spaced typescripton 21 by 29.7 cm (A4 format) or 21.5 by 28 cm (letter format) paper, with wide margins.The pages of the manuscript should be numbered consecutively.
The text of articles should be arranged in the following order: Title Page, Abstracts, Bodyof Text, Literature Cited, Tables, Appendices, and Figure Captions. Each of these sectionsshould begin on a new page. All typescript pages must be double-spaced.
(1) Title Page: This should include the title, author(s) name(s), institutions, and keywordsin English as well as in the language of the manuscript, and a short running title in
Editor-in-Chief: Hussam Zaher, Serviço de Vertebrados, Museu de Zoologia, Universidadede São Paulo, Caixa Postal 42.494, CEP 04218-970, São Paulo, SP, Brasil. E mail:[email protected].
Associate Editors: Antonio C. Marques (Universidade de São Paulo, Brasil), Mario C.C.de Pinna (Universidade de São Paulo, Brasil), Sergio A. Vanin (Universidade de São Paulo,Brasil).
Editorial Board: Aziz N. Ab’Saber (Universidade de São Paulo, Brasil), Rudiger Bieler(Field Museum of Natural History, U.S.A.), Walter A.P. Boeger (Universidade Federal doParaná, Brasil), Carlos Roberto F. Brandão, (Universidade de São Paulo, Brasil), JamesCarpenter (American Museum of Natural History, U.S.A.), Ricardo Macedo Correa eCastro (Universidade de São Paulo, Brasil), Darrel Frost (American Museum of Natural
EDITORIAL COMMITTEEHistory, U.S.A.), W.R. Heyer (National Museum of Natural History, U.S.A.), Ralf Holzentahl(University of Minesotta, U.S.A.), Adriano Kury (Museu Nacional do Rio de Janeiro, Brasil),Gerardo Lamas (Museu Javier Prado de Lima, Peru), John Maisey (American Museum ofNatural History, U.S.A.), Ubirajara Martins (Universidade de São Paulo, Brasil), NaércioMenezes (Universidade de São Paulo, Brasil), Christian de Muizon (Museum Nationald’Histoire Naturelle, France), Nelson Papavero (Universidade de São Paulo, Brasil), JamesPatton (University of Berkeley, U.S.A.), Richard Prum (University of Kansas, U.S.A.), MarcosRaposo (Museu Nacional do Rio de Janeiro, Brasil), Olivier Rieppel (Field Museum ofNatural History, U.S.A.), Miguel T.U. Rodrigues (Universidade de São Paulo, Brasil), RandahlSchuh (American Museum of Natural History, U.S.A.), Marcos Tavares (Universidade deSão Paulo, Brasil), Paulo E. Vanzolini (Universidade de São Paulo, Brasil), Richard Vari(National Museum of Natural History, U.S.A.), Mario de Vivo (Universidade de São Paulo,Brasil) and Paulo Young (Museu Nacional do Rio de Janeiro, Brasil).
INSTRUCTIONS TO AUTHORS(MAY 2002)
the language of the manuscript. The title should be concise and, where appropriate,should include mention of families and/or higher taxa. Names of new taxa shouldnot be included in titles.
(2) Abstract: All papers should have an abstract in English and another in Portuguese,regardless of the original language. The abstract is of great importance as it may bereproduced elsewhere. It should be in a form intelligible if published alone andshould summarize the main facts, ideas, and conclusions of the article. Telegraphicabstracts are strongly discouraged. Include all new taxonomic names for referencingpurposes. Abbreviations should be avoided. It should not include references. Abstractsshould not exceed 350 words.
(3) Body of Text: The main body of the text should include the following sections:Introduction, Materials and Methods, Results, Discussion, and Acknowledgmentsat end. Primary headings in the text should be in capital letters and centered; thefollowing text should begin on the next line, indented. Secondary headings shouldbe in capital and lowercase letters and flush left; the following text should begin onthe next line, indented. Tertiary headings should be in capital and lower case letters,in italics and indented; the following text should be on the same line and separatedfrom the heading by a hyphen.
(4) Literature Cited: Citations in the text should be given as: Silva (1998)..., Silva(1998:14-20)..., Silva (1998: figs. 1, 2)..., Silva (1998a, b)..., Silva & Oliveira (1998)...,(Silva, 1998)..., (Rangel, 1890; Silva & Oliveira, 1998a, b; Adams, 2000)..., (Silva, pers.comm.)..., (Silva et al., 1998), the latter when the paper has three or more authors.The reference need not be cited when author and date are given only as authorityfor a taxonomic name. The literature section should be arranged strictly alphabeticallyand given in the following format:
Journal Article – Silva, H.R.; Oliveira, H. & Rangel, S. Year. Article title. Journal name,00:000-000. Names of journals must be spelled out in full.
Books – Silva, H.R. Year. Book title. Publisher, Place, 000p.
Articles in Books – Silva, H.R. Year. Article title. In: Oliveira, H. & Rangel, S. (Eds.),Book title. Publisher, Place. p.000-000.
Articles in Larger Works – Silva, H.R. Year. Article title. In: H. Oliveira & S. Rangel(Eds.), Title of Larger Work. Serial Publication. Publisher, Place. pp.000-000.
Dissertations and Theses – Silva, H.R. Year. Dissertation title. Ph.D. dissertation,University, Place, 000p.
Tables: All tables must be numbered in the same sequence in which they appear in thetext. Authors are encouraged to indicate where the tables should be placed in the text.They should be comprehensible without reference to the text. Tables should be formattedwith horizontal, not vertical, rules. In the text, tables should be referred as Table 1, Tables2 and 3, Tables 2-6. Use “TABLE” in the table heading.
Illustrations: Figures should be numbered consecutively, in the same sequence theyappear in the text. Separate illustrations of a composite figure should be identified bycapital letters and referred in the text as so (fig. 1A). Where possible, letters should beplaced in the lower right corner of each illustration of a composite figure. Hand-writtenlettering on illustrations is unacceptable. Illustrations should be mounted on stout, whitecardboard. Figures should be mounted in order to minimize blank areas between separateillustrations. High quality color or black and white photographs, and computer generatedfigures are preferable. Authors are encouraged to indicate where the figures should beplaced in the text. Use “(Fig(s).)” and “Figure(s)” for referring to figures in the text, but“FIGURE(S)” in the figure captions and “(fig(s).)” when referring to figures in anotherpaper.
For other details of manuscript preparation of format, consult the CBE Style Manual, available from the Council of Science Editors(http://www.councilscienceeditors.org/publications/style.cfm).
Papéis Avulsos de Zoologia and Arquivos de Zoologia are publications of the Museu de Zoologia da Universidade de São Paulo (www.mz.usp.br).


















