
Oxygen Transport (II) 1. Special features of myoglobin 1.Isolated haem can bind oxygen but in doing...
19
Oxygen Transport (II)
-
Upload
jane-jacobs -
Category
Documents
-
view
218 -
download
2
Transcript of Oxygen Transport (II) 1. Special features of myoglobin 1.Isolated haem can bind oxygen but in doing...

Oxygen Transport (II)
1
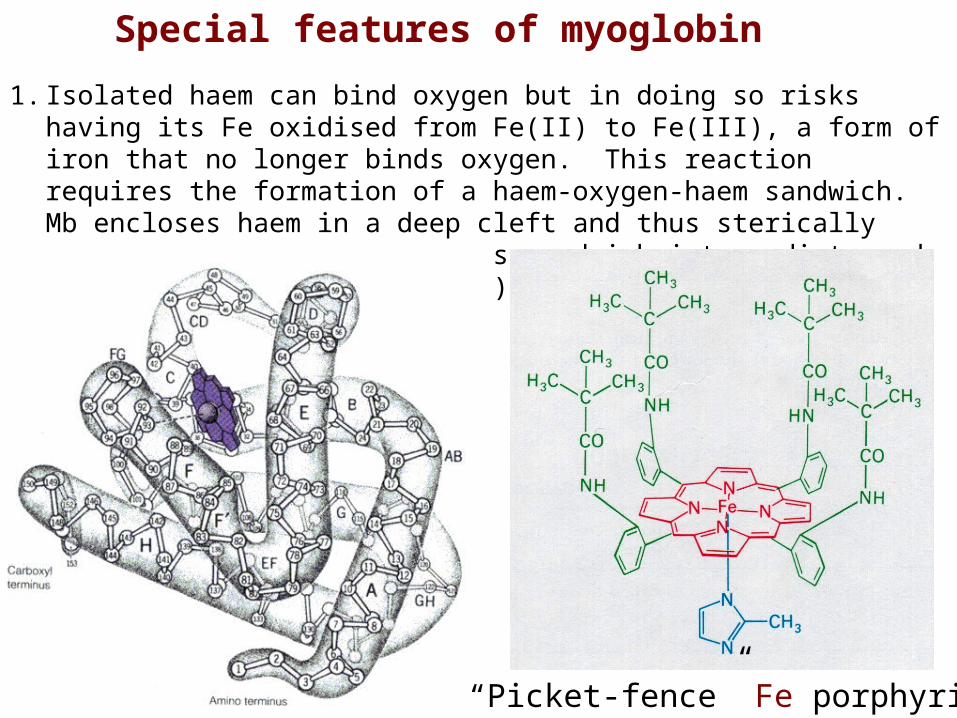
Special features of myoglobin
1. Isolated haem can bind oxygen but in doing so risks having its Fe oxidised from Fe(II) to Fe(III), a form of iron that no longer binds oxygen. This reaction requires the formation of a haem-oxygen-haem sandwich. Mb encloses haem in a deep cleft and thus sterically hinders the formation of this sandwich intermediate and prevents oxidation to Fe(III).
“Picket-fence” Fe porphyrin
2

ProximalHistidine
Distal Histidine
2. The binding environment of the haem reduces its affinity for carbon monoxide (CO) from 25,000 to 200 times greater than its affinity for oxygen. Under normal conditions about 1% of Mb is occupied by CO rather than O2.
Special features of myoglobin3
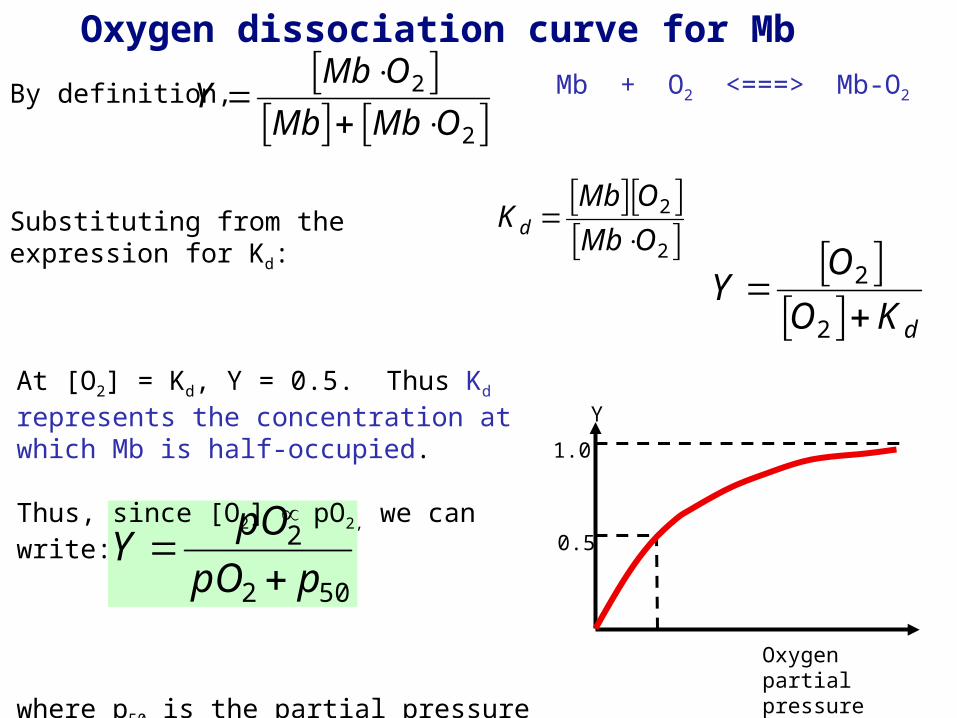
Oxygen dissociation curve for Mb
By definition,
Substituting from the expression for Kd:
Y MbO2
Mb MbO2
Y O2
O2 Kd
4
Mb + O2 <===> Mb-O2
Kd Mb O2 MbO2
Y
Oxygen partial pressure (atm)
1.0
0.5
Y pO2
pO2 p50
At [O2] = Kd, Y = 0.5. Thus Kd represents the concentration at which Mb is half-occupied.
Thus, since [O2] pO2, we can write:
where p50 is the partial pressure of oxygen at which Mb is half-occupied.

Oxygen dissociation curve for Mb
For Mb, p50 = 0.0013 atm (1 torr). In humans, pO2(lungs) = 0.13 atm and pO2(tissue) = 0.026 atm
Mb would be an inefficient O2 transporter (less than 4% of the oxygen picked up in the lungs is released in the tissue) because it binds O2 too tightly.
It stores a reservoir of oxygen in cells that are likely to need rapid access to it under certain conditions; thus Mb will release O2 under conditions of strenuous exercise when the tissue concentration drops to very low levels. Ylungs = 0.990
Ytissue = 0.952
Y = 0.038
Y
Oxygen partial pressure (atm)
1.0
0.5
0.0013 0.026 0.13
lungstissue
Y pO2
pO2 p50
5

Haemoglobin (Hb) - oxygen transporter
Haemoglobin is similar in some ways to myoglobin but has a number of important differences.
Hb is an 22 tetramer (4 polypeptide chains):
2 chains (141 amino acids)
2 chains (146 amino acids)
Max Perutz (1914-2002)
6
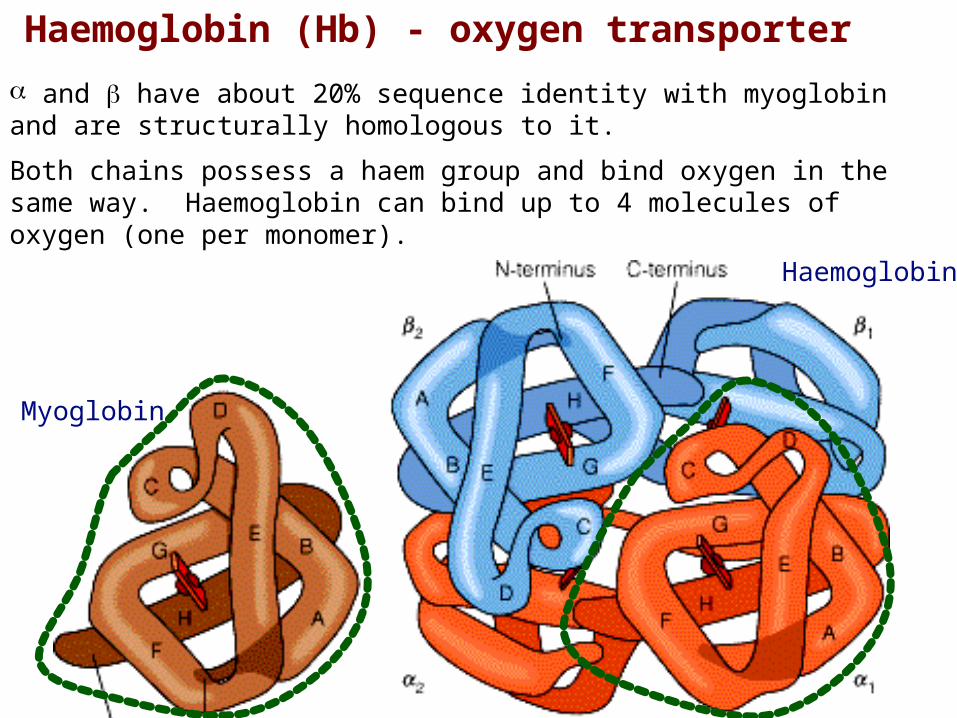
Haemoglobin (Hb) - oxygen transporter
and have about 20% sequence identity with myoglobin and are structurally homologous to it.
Both chains possess a haem group and bind oxygen in the same way. Haemoglobin can bind up to 4 molecules of oxygen (one per monomer).
Myoglobin
Haemoglobin
7

Haemoglobin (Hb) - oxygen transporter
Hb binds a co-factor, 2,3-diphosphoglycerate (DPG) at the interface.
DPG
DPG
8

Haemoglobin (Hb) - oxygen transporter
The oxygen binding curves for Hb and Mb are significantly different.
In contrast to Mb, Hb
(i) binds O2 less tightly
(ii) displays co-operativity (i.e. binding of one molecule of O2 increases the affinity for subsequent O2 binding).
How do we account for this behaviour?
Y
Oxygen partial pressure (atm)
1.0
0.5
0.0013 0.026 0.13
LungsTissue
Hb
Mb
9

Oxygen dissociation curve for Hb
Monod-Wyman-Changeux model
The model makes a number of simple assumptions:
• The protein is an oligomer (>1 polypeptide chain)
• The protein can exist in 2 states: Tense (T) and Relaxed (R)
• T-state has low affinity for oxygen (KT large)
• R-state has high affinity for oxygen (KR small)
• All the subunits of any one molecule are either in the T-state or the R-state (concerted model)
10

Oxygen dissociation curve for Hb
S +
S +
S + + S
+ S
+ S
S S
SS S
S
SS
SSS
S
SS S
S
S
S + + S
S
S
S
L
L1
L2
L3
L4
T0
T1
T2
T3
T4
R0
R1
R2
R3
R4
KT
KT
KT
KT
KR
KR
KR
KR
T to Rconversion
Monod-Changeux-Wyman Model for Hb Allostery
11

Y number of occupied sites
total number of sites
Y R1 2 R2 3 R3 4 R4 T1 2 T2 3 T3 4 T4
4 R0 R1 R2 R3 R4 T0 T1 T2 T3 T4
S +
S +
S + + S
+ S
+ S
S S
SS S
S
SS
SSS
S
SS S
S
S
S + + S
S
S
S
L
L1
L2
L3
L4
T0
T1
T2
T3
T4
R0
R1
R2
R3
R4
KT
KT
KT
KT
KR
KR
KR
KR
T to Rconversion
Monod-Changeux-Wyman Model for Hb Allostery
KT 4 T0 S T1
3 T1 S
2 T2
2 T2 S 3 T3
T3 S 4 T4
Y is defined as the fraction of occupied sites, so:
Oxygen dissociation curve for Hb12
4
3
3
2
2
1
1
0
4
3
2
2
3
4
R
SR
R
SR
R
SR
R
SRKR

To simplify this expression, we can substitute using our expressions for KT and KR and make the following definitions:
[S]
KR ( is proportional to [S])
LT0 R0
(allosteric constant)
c KRKT
(ratio of dissociation constants; c << 1)
S +
S +
S + + S
+ S
+ S
S S
SS S
S
SS
SSS
S
SS S
S
S
S + + S
S
S
S
L
L1
L2
L3
L4
T0
T1
T2
T3
T4
R0
R1
R2
R3
R4
KT
KT
KT
KT
KR
KR
KR
KR
T to Rconversion
Monod-Changeux-Wyman Model for Hb Allostery
Oxygen dissociation curve for Hb13

This gives us:
Y 1 3 Lc 1 c 3
1 4 L 1 c 4
Plotting Y against ,we get:
Y
1.0
S +
S +
S + + S
+ S
+ S
S S
SS S
S
SS
SSS
S
SS S
S
S
S + + S
S
S
S
L
L1
L2
L3
L4
T0
T1
T2
T3
T4
R0
R1
R2
R3
R4
KT
KT
KT
KT
KR
KR
KR
KR
T to Rconversion
Monod-Changeux-Wyman Model for Hb Allostery
Oxygen dissociation curve for Hb14
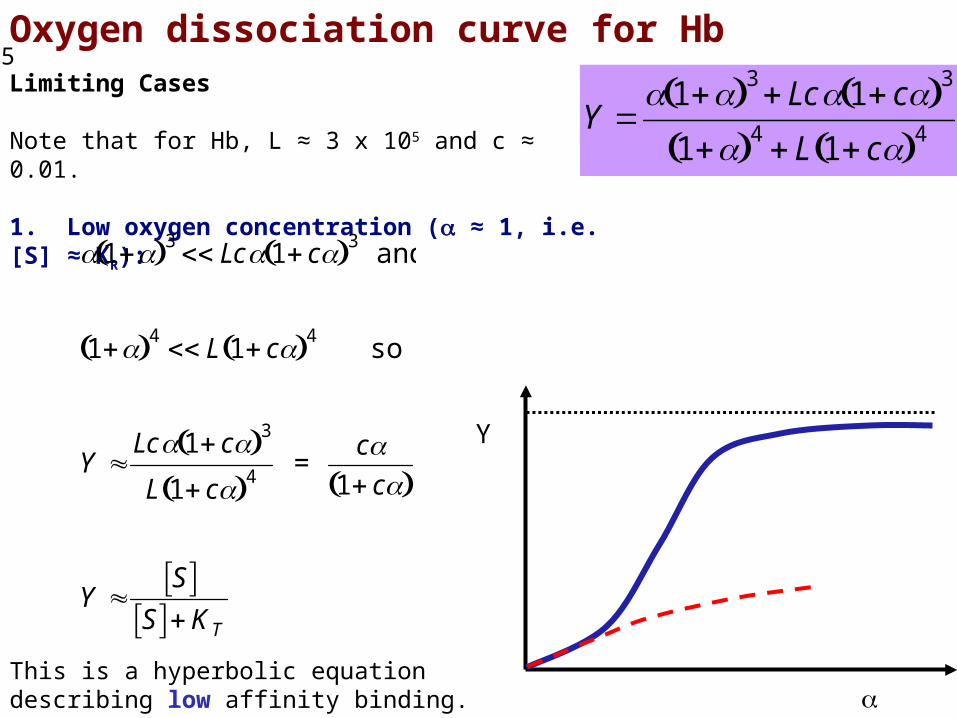
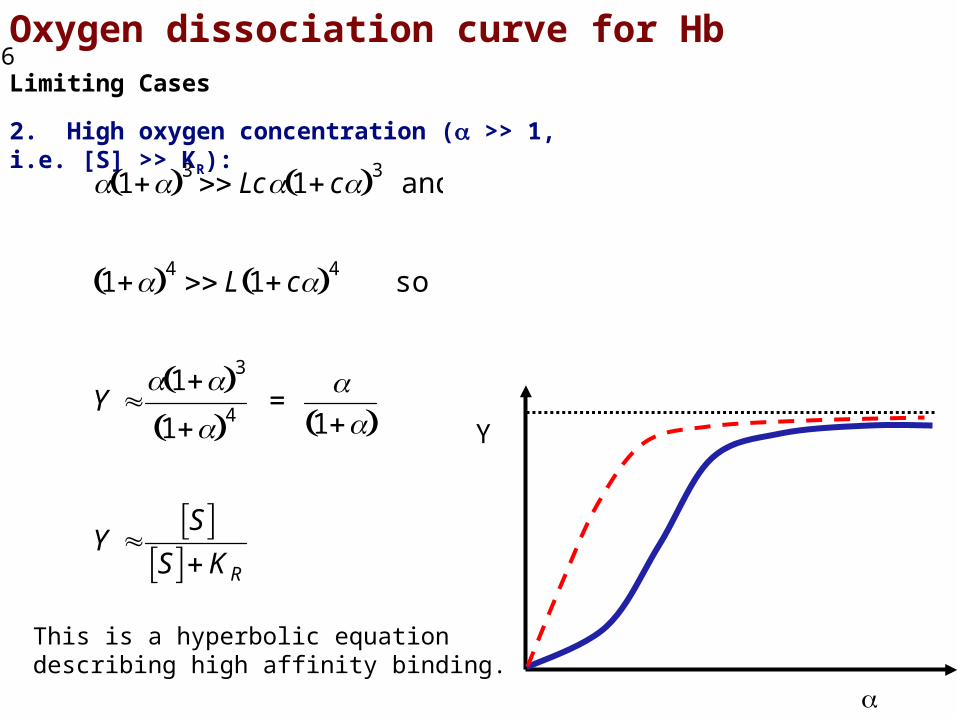
Oxygen dissociation curve for HbLimiting Cases
Note that for Hb, L ≈ 3 x 105 and c ≈ 0.01.
1. Low oxygen concentration ( ≈ 1, i.e. [S] ≈ KR):
1 3 Lc 1 c 3 and
1 4 L 1 c 4 so
Y Lc 1 c 3
L 1 c 4 =
c1 c
Y S
S KTThis is a hyperbolic equation describing low affinity binding.
Y
Y 1 3 Lc 1 c 3
1 4 L 1 c 4
15
Oxygen dissociation curve for HbLimiting Cases
2. High oxygen concentration ( >> 1, i.e. [S] >> KR):
This is a hyperbolic equation describing high affinity binding.
1 3 Lc 1 c 3 and
1 4 L 1 c 4 so
Y 1 3
1 4 =
1
Y S
S KR
Y
16
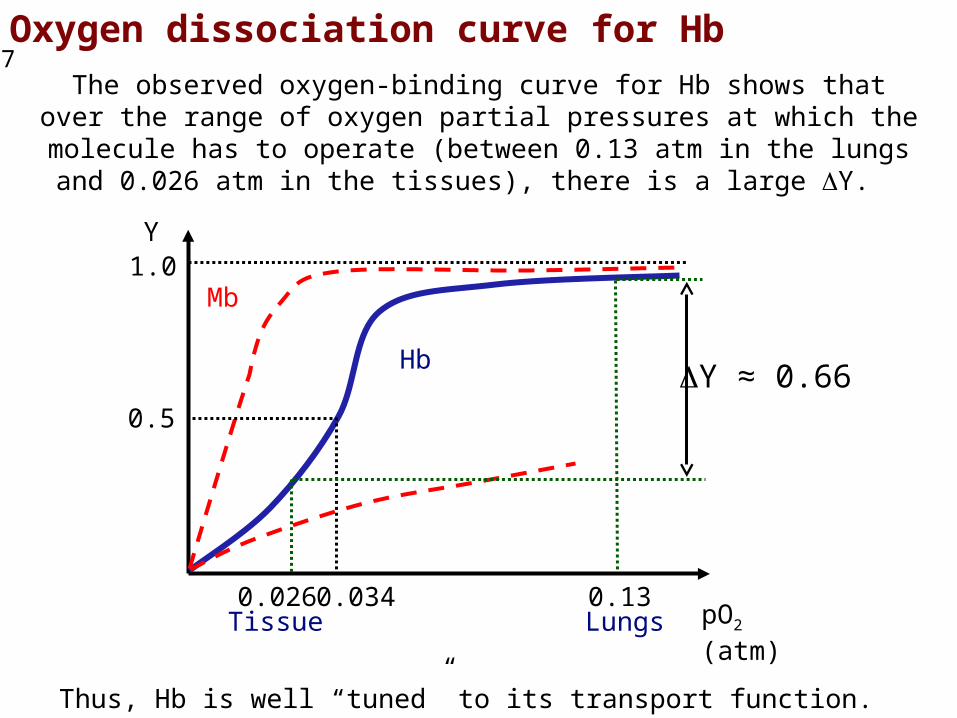
Oxygen dissociation curve for HbThe observed oxygen-binding curve for Hb shows that over the range of oxygen partial pressures at which the molecule has to operate (between 0.13 atm in the lungs and 0.026 atm in the tissues), there is a large Y.
Thus, Hb is well “tuned” to its transport function.
Y
pO2 (atm)
1.0
0.5
0.026 0.034 0.13
Hb
Mb
Y ≈ 0.66
LungsTissue
17
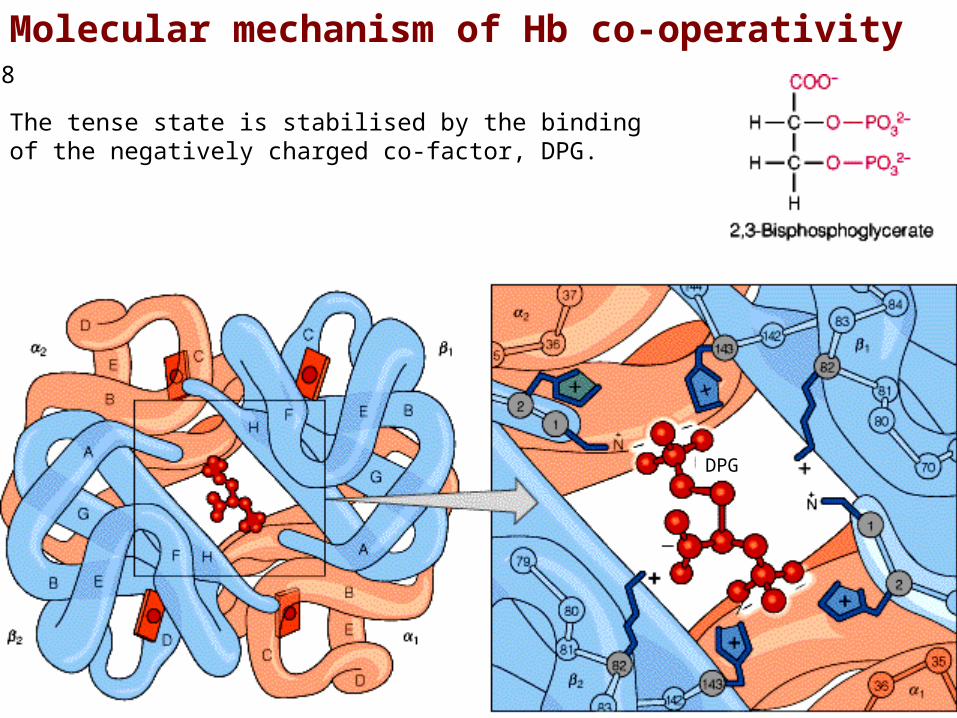
Molecular mechanism of Hb co-operativity
The tense state is stabilised by the binding of the negatively charged co-factor, DPG.
DPG
18

Molecular mechanism of Hb co-operativity
DPG plays a major role in T-state stabilisation; in the absence of DPG, Hb switches to the R-state (high affinity) and is not capable of displaying co-operativity.
Y
Hb without DPG
Hb
19


















