
Oxygen binding properties of the coelomic heamoglobin of the polychaete heteromastus filiformis...
16
Netherlands Journal of Sea Research 13 (314) : 581-592 (1979) OXYGEN BINDING PROPERTIES OF THE COELOMIC HEAMOGLOBIN OF THE POLYCHAETE HETEROMASTUS FILIFORMIS RELATED WITH SOME ENVIRONMENTAL FACTORS h) G. PALS* and E. PAUPTIT (fletherlands Institute for Sea Research, Texel, The Netherlands) CONTENTS I. Introduction. . . . . II. Material and Methods III. Results . . . . . . . IV. Discussion . . . V. Summary . . . . . . VI. References. . . . . . . . . . . . . . . . . . . . . . . . 581 . 582 583 . 587 . 591 . 591 I. INTRODUCTION Heteromastusjiliformis (Capitellidae, Polychaeta sedentaria) was chosen for a study of the molecular and oxygen binding properties of its haemoglobin, as this species lives in highly organic sediments where low oxygen tensions can be expected. Heteromastus jilifrmis, the only species of the genus, possesses, like all Capitellidae, haemoglobin in coelomocytes. The species can be found in places where the substratum consists of muddy, fine sands. High densities may occur (up to 8000 m-s) and the species is an important sediment reworker (CADCE, 1979). It is found from the upper littoral zone to a depth of 3000 metres, and can tolerate decreases in salinity down to 8 or 5x0 (HARTMANN-SCHR~DER, 1971). Heteromastus inhabits a vertical burrow that, at a depth of 10 to 20 cm, subdivides in several feeding burrows. The walls of the burrow show no oxidation zone (like those of Arenicola marina) nor are they lined by a mucilagenous layer (like in Notomastus). It is therefore unIikeIy that these burrows are ventilated. Acknowledgements.-The authors wish to thank Prof. Dr R. E. Weber, University of Odense, and Prof. Dr J. H. F. M. Zandee, State Univer- sity of Utrecht, for their helpful criticism on the manuscript. * Present address : Institute of Human Genetics, Free University, Amsterdam.
Transcript of Oxygen binding properties of the coelomic heamoglobin of the polychaete heteromastus filiformis...

Netherlands Journal of Sea Research 13 (314) : 581-592 (1979)
OXYGEN BINDING PROPERTIES OF THE COELOMIC HEAMOGLOBIN OF THE POLYCHAETE
HETEROMASTUS FILIFORMIS RELATED WITH SOME ENVIRONMENTAL FACTORS
h)
G. PALS* and E. PAUPTIT
(fletherlands Institute for Sea Research, Texel, The Netherlands)
CONTENTS
I. Introduction. . . . . II. Material and Methods
III. Results . . . . . . . IV. Discussion . . .
V. Summary . . . . . . VI. References. . . . . .
. .
.
. .
. .
.
. .
. .
. .
. .
.
. 581
. 582 583
. 587
. 591
. 591
I. INTRODUCTION
Heteromastusjiliformis (Capitellidae, Polychaeta sedentaria) was chosen for a study of the molecular and oxygen binding properties of its haemoglobin, as this species lives in highly organic sediments where low oxygen tensions can be expected.
Heteromastus jilifrmis, the only species of the genus, possesses, like all Capitellidae, haemoglobin in coelomocytes. The species can be found in places where the substratum consists of muddy, fine sands. High densities may occur (up to 8000 m-s) and the species is an important sediment reworker (CADCE, 1979). It is found from the upper littoral zone to a depth of 3000 metres, and can tolerate decreases in salinity down to 8 or 5x0 (HARTMANN-SCHR~DER, 1971).
Heteromastus inhabits a vertical burrow that, at a depth of 10 to 20 cm, subdivides in several feeding burrows. The walls of the burrow show no oxidation zone (like those of Arenicola marina) nor are they lined by a mucilagenous layer (like in Notomastus). It is therefore unIikeIy that these burrows are ventilated.
Acknowledgements.-The authors wish to thank Prof. Dr R. E. Weber, University of Odense, and Prof. Dr J. H. F. M. Zandee, State Univer- sity of Utrecht, for their helpful criticism on the manuscript.
* Present address : Institute of Human Genetics, Free University, Amsterdam.

582 G. PALS & E. PAUPTIT
II. MATEKIAL AND METHODS
Specimens of Heteromastusjl~fbrmis were collected from a mud flat near the Institute at Texel.
Coelomic haemoglobin samples were collected by cutting the pro- boscis and sucking up the coelomic fluid with a glass capillary. Fluid of about 3 specimens had to be pooled for measurement of one oxygen equilibrium curve.
Greater amounts of haemoglobin were obtained by homogenizing 150 to 200 sea water rinsed worms in Tris-HCl buffer (pH 7.1; ionic strength 0.1). After centrifugation, the clear solution obtained was chromatographed on Sephadex G 75 (elution with Tris-HCl buffer; pH 7.5; ionic strength 0.1). Column dimensions were 1.6 x 95 cm, elution velocity 27 ml-h- l, fraction volume 2.4 ml. Elution profiles were monitored with an absorptiometer at 280 nm. All preparation was done at a temperature of 4” C.
Oxygen equilibrium curves were determined using the diffusion chamber technique of NIESEL 8r THEWS (1961). The diffusion and calibration gases were adapted to the PSO of the Heteromastus haemo- globin according to SICK & GERSONDE (1969). Oxygen curves were made at a temperature of 15” C except for measurements on the temperature dependence. All solutions measured were buffered using 0.1 N Tris-HCl and Isis-Tris-HCl. To prevent lysis of the coelomocytes, the ionic strength of the buffers was increased to 0.7, by means of sodium chloride.
The Root effect was estimated in a series of 0.1 M Tris-HCl and bis-Tris-HCI buffers, and calculated from the change in optical densities at 540 and 560 nm, after adding a few crystals of sodium dithionite. The change in optical densities at various pH values were then expressed as a percentage of the change of values at pH 8 (ROOT 8: IRVING, 1943).
Iso-electric focusing was done on thin-layer polyacrylamide gels with the aid of an LKB 2117 Multiphor apparatus. Samples of purified haemoglobins of Heteromastus were applied on the middle of the gel-slabs by using small pieces of filter paper. Gels contained ampholines with a pH range of 3.5 to 9.5. After separation the pH ranges of the gels were measured and the gels were treated with coomassie brilliant blue (KARLSSON, DAVIES, OHMAN & ANDERSSON, 1973).
An estimate of molecular weights was made by SDS-disc-gel electrophoresis. Experiments were performed in 5% acrylamide gel rods. Centrifuged samples, after lysing the coelomocytes, were incubated in buffered ls:i, sodium dodecylsulphate (SDS) solution, with and

THE HAEMOGLOBIN OF HETEROMASTUS 583
without P-mercaptoethanol, during 2 hours at 37” C. The reduced samples were mixed with 10 parts of diIution buffer, containing bro- mophenol blue as a tracking dye, and applied on top of the gels. A calibration mixture containing cytochrome C (moIecular weight 12 000), chymotrypsinogen (h1.W. 25 000), ovalbumin (h/I.Fv. 45 000) and human haemoglobin (&l.W. 67 500) was treated simultaneously. A constant current of 8 mA per gel was applied during 3 hours. Then the gels were stained with coomassie brilliant blue-R 250 (SHAPIRO, VIRUELA & PVIAIZEL, 1967; FISH, REYNOLDS 8r TANFORD, 1970).
Oxygen concentration and pH were measured in the interstitial water of the natural habitat of Heteromastus. Small amounts of water were sucked through sintered glass fiIters which had been buried at different depths in the sediment some days before. The water thus obtained was pumped directly into the measuring chamber of an Instrumentation Laboratories 213 blood gas analyzer fitted with a Clark electrode.
The pH in head and tail of Heteromastus was measured with an antimony electrode at 20” C as described by BEELEY, STEVENSON & BEELEY (1972).
III. KESUL1’S
On the tidal flat Heteromastus was found at depth, of 10 to 40 cm, with low oxygen tensions decreasing to zero at about 30 cm depth (Table I). The pH in the body fluid, measured immediately after collection of an animal, was 6.3 to 6.6 in the head and 6.7 to 7.0 in the tail. There was an almost constant pH difference, averaging 0.45 units, between head and tail of an animal.
TAB1.E I
Measurements in the interstitial water in the sediment near burrows of Hetrromastus jliformis, at low tide. Air temperature 20.2” C.
Depth (cm) ~02 (N.m-2)
-10 16.3 15 13.6 20 8.2 25 6.8 30 0.0 35 0.0
PH
7.94 7.82 7.76 7.77 7.70 7.65
Temperature (“C)
22.3 20.5 19.4 18.3 16.5
Haemoglobin concentration could not be measured in the samples of coelomic fluid used to determine the oxygen dissociation curves, because of the small size of the animals. Diluting of the samples with

584 G. PALS & E. PAUPTIT
Tris-HCI buffer in the range of 1 to 5 times had no effect on the Pss, and only slightly decreased the Hill constant.
The oxygen affinity of Heteromastus haemoglobin is high. Whole coelomic fluid and haemolysates had the same I’s0 measured at 15” C and pH 7.4 (tris-HCl buffer), viz. 9 N-m-2 (0.68 mm Hg) while PRO for haemoglobin purified by column chromatography was 3.3 N *m-s (0.25 mm Hg). The oxygen dissociation curves are slightly sigmoidal (Fig. la).
/ I- / ,/ s- i / ,” _ I
f/ ;*/” ; ,,’ i 1 / ,’
- <’
. /’
/
1
/ 01 1 i l”’
0, ( , , 0 10 20 30 ‘lo I 10
PO*lN m-t PO, (N m‘?
Fig. 1. Oxygen dissociation curve of Heteromastus jilijbrmis haemoglobin (a.) and corresponding Hill plot (b.). Coelomic fluid (not haemolysed) with Tris-HCl buffer
(pH 7.82 ; ionic strength 0.7; 15” C).
The Hill plots show an inflection near half-saturation, reflecting two main phases of cooperativity (Fig. lb). The Hill coefficient, n, below 50% saturation (referred to as nr) varied from 1.1 to 2.2, indicating low cooperativity. Above 50% saturation (referred to as ns) the Hill constant varied from 0.83 to 1.5, indicating no or slight cooperativity. Both phases of n varied with pH and haemoglobin con- centration, nr was additionally dependent on temperature. These effects are considered below (page 585).
According to SICK & GERSONDE (1969) application of thick layers of sample in the diffusion chamber and the use of diffusion gas with too high oxygen tension affect the dissociation curves and Hill coeffi- cients. Applying extremely thin layers (<0.02 mm) and using low oxygen concentrations in the diffusion gas, however, still produced Hill plots consisting of two straight lines, with the phase change near

THE HAEMOGLOBIN OF HETEROMASTUS 585
50% saturation, while thick layers and high diffusion gas concentra- tions resulted in multiphasic Hill plots. Hence, we conclude, that the observed phase shift is not an artefact, but represents a real property of the pigment.
The influence of pH on oxygen dissociation was tested between pH 5.86 and pH 8.67 (Fig. 2). The Bohr factor, Q, (A log Pss/ApH), is -0.34 in the entire pH range for whole and haemolysed coelomic fluid as well as for haemolysed washed cells and solutions of stripped haemoglobin. The Root effect (influence of pH on the maximum quantity of oxygen that can be bound) was only important at the lowest pH tested; at pH 5.86, 45% of the haemoglobin does not bind oxygen, while at pH 6.21 this is only 1%.
I lb I60 ~0, (N m-‘1
Fig. 2. Hill plots of HeteromastusJiliformis coelomic fluid at different pH (Tris-HCl buffers; ionic strength 0.7 ; 15” C).
Both Hill constants (nr and nz) vary with pH, but there is no definite trend (Fig. 2). At low pH the Hill plot becomes monophasic.
Temperature dependence of oxygen binding was measured in small portions of a pooled sample to exclude influence of haemoglobin con- centration. Changes in temperature have no effect on a.~, but the nr values decrease with increasing temperature (Fig. 3). The temperature effect which increases PSO with increasing temperature, can be used

586 G. PALS & E. PAUPTIT
to calculate the enthalpy change AH using the integrated van ‘t Hoff equation. The values of AH are: -37.4 (- 11.9) kJ *mol-1 for whole coeIomic fluid; -40.5 (-12.5) kJ* mol-r for haemolysed washed cells and -50.5 (-20.6) kJ* mol-r for solutions of stripped haemoglobin. Figures in parentheses are values corrected for the heat of oxygen dissolution and for the influence of the in vitro Bohr effect caused by temperature induced pH changes. Correction for the Bohr effect was as follows: After measuring the temperature induced pH changes, all P50 values were corrected by abstracting @ - A pH. With the cor- rected Pso values, AH values were recalculated.
, / /‘
/’ I’
, I I
0.1 I IO PO, (N m-‘1
Fig. 3. Hill plots of purified haemoglobin of Heteromastus,Fliformis at different tem- peratures (Tris-HCl buffer; pH 7.50 at 15” C; ionic strength 0.7).
The organic phosphate 2-3 diphosphoglycerate (DPG) has no or only slight effect on the oxygen affinity. ATP has no effect on I’s0 but in the presence of this phosphate, nr increases (1.3 to 1.9). The differ- ence in optical density between oxygenated and deoxygenated form decreases after addition of ATP, reflecting a decrease of about SOqG in the amount of oxygen that can be bound. Addition of DPG does not change the difference in optical density. The exact concentrations of the phosphates and the haemoglobin were not known, but this is not important because the concentration of the phosphates was higher



3 2 3 4 5 6 . .
Above: SDS disc-gel electrophoresis of stripped haemoglobin of Heteromastus firmis: 2, 4 and 6 haemoglobin; 1. 3 and 5 calibration proteins, viz. from the downward human haemoglobin (M.W. 67500), ovalbumin (M.W. 45000), thy trypsinogen (M.W. 25000) and cytochrome C (M.W. 12000). Below: Clustc coelomocytes of HeteromastusJilz~ormir, 400 x enlarged, without staining, the diarr
of the coelomocytes is 13 to 18 Frn (photographed by A. D. G. Dral).
,jil i-
top ‘mo- :r of teter


THE HAEMOGLOBIN OF HETEROMASTUS 587
than that of the pigment and it is improbable that a molecule of pigment with low molecular weight can bind more than one molecule of phosphate (BENESCH, BENESCH & RENTHAL, 1971).
Absorption spectra of oxygenated haemoglobin of Heteromastus jili- formis (Fig. 4) show maxima at 575 and 540 nm, the former being greater, but this may be caused by partial oxidation ( PROSSER, 1973). Spectra of deoxygenated haemoglobin have a maximum at 560 nm.
OD OD I
I I I I I I
500 600 500 h (ml
600
Fig. 4. Absorption spectra of Heteromastus jiliformis haemoglobin in oxygenated (broken line) and deoxygenated form (solid line). a. At pH 6.21. b. At pH 8.10.
Thin-layer iso-electric focusing of Heteromastus haemoglobin in oxy- genated form shows 4 major components at pH 5.86, 6.13, 6.95 and 7.15.
SDS-disc-gel electrophoresis (Plate I) and Sephadex G75 gel filtra- tion (Fig. 5) were used to estimate the molecular weight of the haemo- globin. The molecular weight appeared to be below the range of the calibration proteins used (smaller proteins were not available) ; an accurate determination was therefore not possible. The experiments indicate a value lower than that of cytochrome C (12 000).
Observation of living specimens of Heteromastus under the microscope shows that the coelomocytes move in clusters (Plate I) through the body. Two separate streams are seen, from head to tail and vice versa. This circulation is caused by separating septa in the coelom.The tail has surface-enlarging pleats through which the coelomocytes are circulated.
IV. DISCUSSION
The oxygen binding properties of respiratory pigments of annelids have been investigated in about 40 species (reviews: WELLS & DALES,
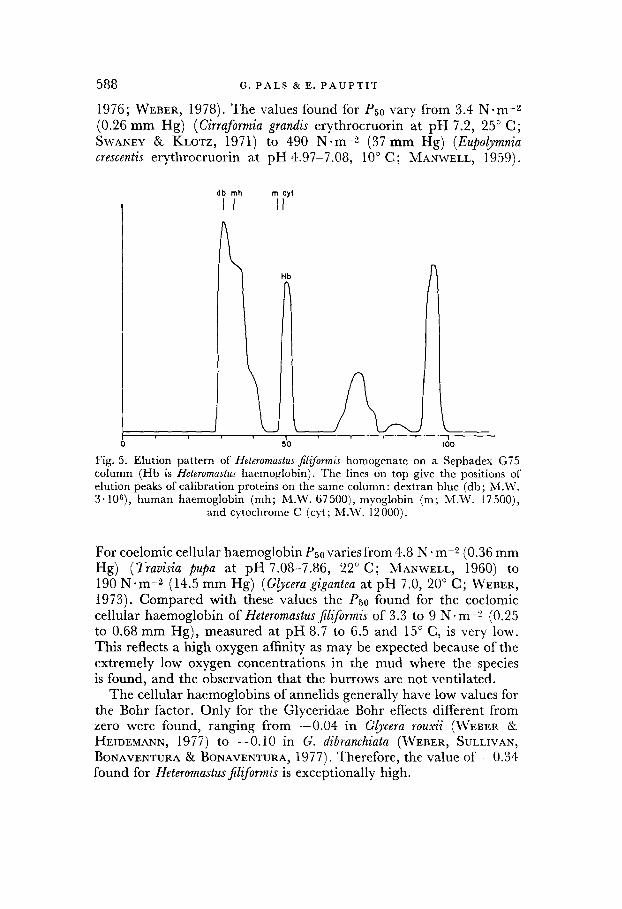
588 G. PALS & E. PAUPTIT
1976; WEBER, 1978). The values found for PSO vary from 3.4 N*m-s (0.26 mm Hg) (Cirraformia grandis erythrocruorin at pH 7.2, 25” C; SWANEY & KLOTZ, 1971) to 490 N-m-2 (37 mm Hg) (Eupolymnia crescentis erythrocruorin at pH 4.97-7.08, 10” C; MANWELL, 1959).
o-
db mh m cyt
II II
II: > -8
HI
3 50
Fig. 5. Elution pattern of Heteromastus filiformis homogenate on a Sephadex G75 column (Hb is Heteromastus haemoglobin). The lines on top give the positions of elution peaks of calibration proteins on the same column: dextran blue (db; M.W. 3. lOs), human haemoglobin (mh; M.W. 67500), myoglobin (m; M.W. 17500),
and cytochrome C (cyt ; M.W. 12 000).
For coelomic cellular haemoglobin PSO varies from 4.8 N * m-2 (0.36 mm Hg) (Travisia pupa at pH 7.08-7.86, 22” C; MANWELL, 1960) to 190 N-m-2 (14.5 mm Hg) (G&era gigantea at pH 7.0, 20” C; WEBER, 1973). Compared with these values the PSO found for the coelomic cellular haemoglobin of HeteromastusJilzjbmis of 3.3 to 9 Nmm-2 (0.25 to 0.68 mm Hg), measured at pH 8.7 to 6.5 and 15” C, is very low. This reflects a high oxygen affinity as may be expected because of the extremely low oxygen concentrations in the mud where the species is found, and the observation that the burrows are not ventilated.
The cellular haemoglobins of annelids generally have low values for the Bohr factor. Only for the Glyceridae Bohr effects different from zero were found, ranging from -0.04 in Gbcera rouxii (WEBER &
HEIDEMANN, 1977) to -0.10 in G. dibranchiata (WEBER, SULLIVAN, BONAVENTURA & BONAVENTURA, 1977). Therefore, the value of -0.34 found for Heteromastusjfjhmis is exceptionally high.

THE HAEMOGLOBIN OF HETEROMASTUS 589
The Hill constants for haemerithrins and cellular haemoglobins are about 1 to 1.8, and for some erythrocruorins values up to 6 were found. The values found for Heteromastus jil@rmis haemoglobin above 50% saturation are normal for cellular haemoglobins, but below 500/b saturation they are comparatively high, up to 2.2 (HILL, 1910).
The Hill plots of Heteromastus jliformis haemoglobin are striking, since a phase shift resulting in a decreased cooperativity at high saturation is not found in other haem respiratory pigments. For some species the reverse was found, viz. for the cellular haemoglobin of the annelid Amphitrite ornata (MANGUM et al., 1975 ; WEBER, MANGUM, STEINMAN, BONAVENTURA, SULLIVAN & BONAVENTURA, 1977) and the haemoglobin of the lamprey Entosphenus japonicus (DOHI, SUGITA &
YONEYAMA, 1973). DOHI, SUGITA 8r YONEYAMA suggest that the increased cooperativity in Entosphenus haemoglobin at high p0s is caused by a monomer-dimer equilibrium of the haemoglobin with increased aggregation in the oxygenated form. The values of the Hill constants for Heteromastus jilifrmis haemoglobin, however, indicate a loss of cooperativity, and consequently decreased aggregation in the oxygenated form. The high values of n2 that were occasionally found (2.0 to 2.2) even suggest a tetrameric form. This need not be in contra- diction with the extremely low estimated molecular weight ( < 12 000)) because this value was determined in the presence of oxygen, which may favour the monomeric form.
There is further evidence for an association equilibrium. The values of n increase with increasing haemoglobin concentration, and below
% oxy Hb 100-l
Fig. 6. Comparison of the measured sigmoid dissociation curve of Heteromastus filiformis haemoglobin (solid line), with the theoretical hyperbolic curve calculated with the same Pss (broken line). The arrows represent the oxygen delivery as $02 decreases from 20 to 5 N.m-2 according to the actual equilibrium (a) and the hyperbolic curve (b) (the measured curve has a 1.6x higher oxygen delivery than
the theoretical one).

590 G. PALS & E. PAUPTIT
50% saturation nr decreases with increasing temperature reflecting increased dissociation with rising temperature.
The phase shift in the Hill plot is connected with a stronger sigmoi- da1 character of the oxygen dissociation curve at low ~0,. The possible significance for oxygen delivery is shown in Fig. 6.
The significant heterotrophic and homotrophic interactions observed (Bohr effect and cooperativity) are remarkable considering the low molecular weight of this haemoglobin.
The fact that Heteromastus maintains an inverted posture (head downwards) in vertical, unventilated burrows in oxygen deficient sediments makes oxygen uptake with the posterior end likely. The circulation of coelomocytes observed will then transport the oxygen to the head. To release oxygen in the head, the extremely high affinity of the haemoglobin must be lowered there. A contribution to this is given by the strong Bohr effect, together with the observed pH difference between head and tail. Considering the affinity of stripped haemoglobin, which is very high compared with that of whole coelomic fluid and haemolysed coelomocytes at the same pH, we conclude that there probably is a “modificating factor” present which lowers the affinity of the haemoglobin for oxygen. The effect of ATP and DPG on the affinity is too small to be involved, so the nature of the suggested factor remains unknown.
% oxy Hb 100
1
Fig. 7. Oxygen uptake and release in Heteromastusjlij3rmir, assuming uptake in the tail and release in the head. The broken line represents the curve of coelomic fluid from the head at pH 6.55, the solid line that of purified haemoglobin at pH 6.94. At the ~0s that was found at a depth of 10 cm in the sediment (16.3 N.m-2) purified haemoglobin would be 70% oxygenated (a), and 52% oxygenated at half this ~0s (c). Therefore, comparison between the blood situation in (a) and that in (b) tentatively reflects the in vivo difference between head and tail. The decrease in oxygenation from 70y0 in (a) to 127; in (b) means an oxygen release of 837;. The
curves, however, show the extremes within which the in vivo curves must fall.

THE HAEMOGLOBIN OF HETEROMASTUS 591
ATP, however, does have a rather strong effect on the total amount of oxygen that can be bound, comparable with the Root effect of Hf ions. According to WEBER & DE WILDE (1975), ATP may increase the Root effect of fish haemoglobin. Since oxygen binding takes place in the tail, a possible “modificating factor” should have its lowest con- centration in the tail. The two curves at the pH values that were found in head and tail (Fig. 7) show the two extreme situations within which the in uivo curves must fall. The curves demonstrate that even at the low oxygen concentrations that were measured (Table I) oxygen can be transported. The extremely high oxygen affinities of the haemo- globin appear to be in agreement with the extremely adverse respira- tory qualities of the habitat.
The intracellular coelomic haemoglobin of Heteromastus jiliformis was studied. The haemoglobin is heterogeneous on the basis of isoelectric point, the 4 main components having in the oxygenated form isoelectric points of 5.86, 6.13, 6.95 and 7.15. SDS electrophoresis in the presence of detergent indicates a molecular weight below 12 000. The oxygen affinity is high; PRO is 9 N-m-2 (0.68 mm Hg) for whole and haemo- lysed coelomic fluid, and 3.3 N.rnp2 (0.25 mm Hg) for purified haemo- globin (15” C, pH 7.4). The Bohr factor is -0.34. There is a distinct Root effect at pH (6. Hill plots show an abrupt decrease of coopera- tivity above 50% saturation. An association equilibrium of the haemo- globin is suggested. There is evidence for an influence of unknown factors on the oxygen affinity. The results are discussed in relation to the environmental parameters.
VI. REFERENCES
BEELEY, J. A., S. M. STEVENSON Pr J. G. BEELEY, 1972. Polyacrylamide gel iso- electric focusing of proteins: Determination of iso-electric points using an antimony electrode.-Biochim. Biophys. Acta 285: 293-300.
BENESCH, R., R. E. BENESCH & R. RENTHAL, 1971. Cofactor binding and oxygen equilibria in haemoglobin.-Nature new Biol., N.Y. 234: 174-l 75.
CADRE, G. Cl., 1979. Sediment reworking by the polychaete Heteromastus jil~formis on a tidal flat in the Dutch Wadden Sea.-Neth. J. Sea Res. 13 (3/4): 441-456.
DOIII, Y., Y. SUCITA & ‘I*. YONEYAM.~, 1973. Th e self-association and oxygen equilibrium of haemoglobin from the lamprey, Entosphenus japonkyJ. biol. Chem. 248: 2354-2363.
FISII, W. W., J. A. REYNOLDS & C. T.~NFORD, 1970. Gel chromatography of proteins in denaturing solvents.-J. biol. Chem. 245: 5166-5168.
HARTMANN-SCHRBDER, G., 1971. Die T’ rerwelt Deutschlands, Teil : Annelida: Borstenwiirmer, Polychaeta. Fischer Verlag. Jena. 58: 3922402.

592 G. PALS & E. PAUPTIT
HII.L, A. V.: 1910. The possible effects of aggregation of haemoglobin on its disso- ciation curves-J. Physio!., Lond. 40: 4-7.
KARLSSON, C., H. DAVIES, J. OHMAN & U. ANDERSSON, 1973. Analytical thin layer gel electrofocusing in polyacrylamide gel. LKB Appl. note 75.
MANGUM, C. P., B. R. WOODIN, C. BONAVENTURA, B. SULLIVAN RT J. BONAVENTUKA, 1975. The role of coelomic and vascular haemoglobin in the annelid family Terebellidae.-Comp. Biochem. Physiol. 51A: 28 l-294.
MANWELL, C., 1959. Alkaline denaturation and oxygen equilibrium of annrlid haemoglobins.-J. cell. camp. Physiol. 53: 61-74.
--, 1960. Histological specificity of respiratory pigments. I. Comparisons of the coelome and muscle hemoglobins of the polychaete worm Travisia Pupa and the cchiurid worm Archynchite pugettensk-Comp. Biochem. Physiol. 1: 267-276.
NIESEL. W. & G. THEWS, 1961. Ein neues Verfahren zur schnellen und genauen Aufnahme der Sauerstoffbindungskurve des Blutes und konzentrierter HBmo- proteidlosungen.-Pfliigers Arch. gea. Physiol. 273: 380-395.
PKOSSER, C. L.. 1973. Comparative animal physiology. Saunders, Philadelphia. London, Toronto : 3 17-361.
Roar, R. W. 8: L. IRVIKG, 1943. The effect of carbon dioxide and lactic acid on the oxygen combining power of whole and hemolysed blood of the marine fish Tautoga onitis (Linn.) .-Biol. Bull. mar. biol. Lab., LYoods Hole 84: 209-2 12.
SH,\PIRO, A. L., E. VI%JELA & .J. V. MAIZEL, 1967. Molecular weight estimation ofpolypeptide chains by electrophoresis in SDS-polyacrylamide gels.---Biochem. biophys. Res. Commun. 28: 815-820.
SICK, H. & K. GERSONDE, 1969. blethod for continuous registration of Os-binding curves of hemoproteins by means of a diffusion chamber.-Analyt. Biochem. 32: 3622376.
SWANE~., ,J. B. Er I. M. KLOTZ, 197 I. Porperties of erythrocruorine from CirruJor/ormia grtlndis.-Archs Biochem. Biophys. 147: 4755486.
WEBER, R. E., 1973. Functional and molecular properties of corpuscular harmo- globin from the bloodworm Glycera gigantea.-Neth. J. Sea Res. 7: 316-327.
--. 1978. Respiratory pigments. In: P. .J. MILL. Physiology of annelids. Academic Press, New York.
WEBI?R, R. E. 8r W. HEIDEMANN, 1977. The coelomic haemoglobin from the blood- worm Gbcern rou.Gi. Molecular and oxygenational properties.- Comp. Biochem. Physiol. 57A: 151-155.
WEBER, R. E.. C. P. MANGUM, H. STEINMAN. C. BONAVENTUR.X, B. SUL.LIVAN & J. BONAVENTURA, 1977. Hemoglobins of two terebellid polychaetes, Enoplo- branchus sanguineus and Amphitrite ornata.--Camp. Biochem. Physiol. 56A: 179-187.
WEBER, R. E., B. SULLIVAN, .J. BONAVENTURA & C. BONAVENTURA, 1977. The haemoglobin systems of the bloodworms Glycern dibranchiata and G. americana. Oxygen binding properties of haemolysates and component haemoglobins.--- Comp. Biochem. Physiol. 58B: 1833187.
WEBER, R. E. &J. A. M. DE WILDE, 1975. Oxygenation properties of haemoglobins from the flatfish plaice (Pleuronectes platessa) and flounder (Platichthys Jesus) .- J. camp. Physiol. BlOl: 99-l 10.
WELL+ R. M. G. Pr R. P. DALES, 1976. Review: Subunit organization in the res- piratory proteins of the Polychaeta.-Camp. Biochem. Physiol. 54A: 3877393.


















