
outcrossing dynamic management genetic predominantly selfing … · PAO and PBO were also studied....
15
Original article The relevance of outcrossing for the dynamic management of genetic resources in predominantly selfing Triticum aestivum L. (bread wheat) Jérome Enjalbert Isabelle Goldringer Jacques David b Philippe Brabant a Station de génétique végétale, Institut national de recherche agronomique, Inapg, Ups, Ferme du Moulon, 91190 Gif-sur-Yvette, France b Station de génétique et amélioration des plantes, Institut national de recherche agronomique, Domaine de Melgueil, 34130 Mauguio, France c Institut national agronomique Paris-Grignon, 16, rue Claude Bernard, 75005 Paris, France Abstract - The mating system is of major concern for the dynamic management of genetic resources. We investigated the outcrossing rate of six experimental wheat populations, derived from two genetic pools (PA and PB) which have been cultivated at four French sites. The replicates of the two segregating populations have undergone 10 years of multiplication in isolation, with no artificial selection. For each population, 78 individuals were randomly sampled and genotyped for 25 codominant loci revealed by 15 restriction fragment length polymorphism (RFLP) probes. Outcrossing rates were estimated on the basis of the multilocus heterozygote deficiency. Estimates ranged from 2.4 to 10.1 %, depending on the population. The outcrossing rate appeared to be affected by the cultivation site, and also by the interaction between the genetic pool and the cultivation site. No significant pool effect was found over sites. The use of natural or modified outcrossing rates is suggested for the management of gene flows between the subpopulations of a metapopulation. Consequences of recombination events within and between populations are discussed in the context of short-term adaptive response of populations and long-term elimination of weakly deleterious mutations. © Inra/Elsevier, Paris outcrossing rate / gene flow / dynamic management / genetic resources / Triticum aestivum L. * Correspondence and reprints Résumé - Taux naturels d’allofécondation dans des populations expérimentales de blé, et conséquences pour une gestion dynamique des ressources génétiques. Le régime de reproduction est un paramètre essentiel pour le pilotage d’un programme de gestion dynamique des ressources génétiques. Cet article s’attache à l’estimation
Transcript of outcrossing dynamic management genetic predominantly selfing … · PAO and PBO were also studied....

Original article
The relevance of outcrossing for thedynamic management of geneticresources in predominantly selfing
Triticum aestivum L. (bread wheat)
Jérome Enjalbert Isabelle GoldringerJacques Davidb Philippe Brabant
a Station de génétique végétale, Institut national de recherche agronomique,Inapg, Ups, Ferme du Moulon, 91190 Gif-sur-Yvette, France
b Station de génétique et amélioration des plantes, Institut national de rechercheagronomique, Domaine de Melgueil, 34130 Mauguio, France
c Institut national agronomique Paris-Grignon, 16, rue Claude Bernard,75005 Paris, France
Abstract - The mating system is of major concern for the dynamic managementof genetic resources. We investigated the outcrossing rate of six experimental wheatpopulations, derived from two genetic pools (PA and PB) which have been cultivatedat four French sites. The replicates of the two segregating populations have undergone10 years of multiplication in isolation, with no artificial selection. For each population,78 individuals were randomly sampled and genotyped for 25 codominant loci revealedby 15 restriction fragment length polymorphism (RFLP) probes. Outcrossing rateswere estimated on the basis of the multilocus heterozygote deficiency. Estimatesranged from 2.4 to 10.1 %, depending on the population. The outcrossing rateappeared to be affected by the cultivation site, and also by the interaction between thegenetic pool and the cultivation site. No significant pool effect was found over sites.The use of natural or modified outcrossing rates is suggested for the managementof gene flows between the subpopulations of a metapopulation. Consequences ofrecombination events within and between populations are discussed in the contextof short-term adaptive response of populations and long-term elimination of weaklydeleterious mutations. © Inra/Elsevier, Parisoutcrossing rate / gene flow / dynamic management / genetic resources /Triticum aestivum L.
*
Correspondence and reprints
Résumé - Taux naturels d’allofécondation dans des populations expérimentalesde blé, et conséquences pour une gestion dynamique des ressources génétiques.Le régime de reproduction est un paramètre essentiel pour le pilotage d’un programmede gestion dynamique des ressources génétiques. Cet article s’attache à l’estimation

du taux d’allogamie dans six populations expérimentales de blé tendre, issues de deuxpools génétiques différents (PA et PB) cultivés dans quatre sites distincts. Chaquepopulation a subi dix années de multiplication en isolement, sans sélection consciente.Dans chacune d’elles, 78 individus ont été aléatoirement échantillonnés et génotypéspour vingt-cinq locus codominants, grâce à quinze sondes RFLP. Les taux d’allogamieont été estimés à partir de l’hétérozygotie résiduelle multilocus. Un niveau d’allogamieélevé pour cette espèce cléistogame a été observé. Les estimations varient de 2,4à 10,1 % suivant les populations. Ces taux d’allofécondation dépendent d’une partdu site de culture, et d’autre part d’une interaction entre composition génétiqueinitiale et site de culture. L’intérêt de l’allogamie tant pour la génération de nouvellescombinaisons génétiques que pour la purge des mutations faiblement délétères estdiscuté. L’utilisation des taux naturels d’allogamie pour instaurer des flux de gènesentre les populations est proposée dans l’optique d’une gestion en métapopulation.© Inra/Elsevier, Paristaux d’allogamie / flux de gènes / gestion dynamique / ressources génétiques /Triticum aestivum L.
1. INTRODUCTION
During the past 30 years, national and international efforts have led to theconstruction of large collections of genetic resources. For the major cereal crops(rice, corn, wheat), these collections mainly comprise sampled seeds, stored incold rooms of central gene banks (Frankel et al., 1995). Though gene banksare a safe and efficient way to preserve genetic resources from extinction, thelong-term conservation of resources in isolation from the environment leads toa decrease in their usefulness. Whereas genetic resources are protected in coldrooms, pathogens are evolving and the climate may change. Resources are nolonger exposed to selective pressures, and eventually will prove ill-adapted to anew environment (Simmonds, 1962; Henry et al., 1991). Hence, complementarystrategies, qualified as ’in situ or ex situ dynamic managements’ (DM) arepresently developed to circumvent this evolutionary freeze. Their aim is tomaintain or mimic natural processes responsible for the diversification andconservation of genetic diversity.On the basis of the metapopulation concept (Levins, 1970; Olivieri et al.,
1990), a pilot programme of dynamic management of genetic resources of breadwheat (TW!CMm aestivum L.) started in France in 1984. Its general rules are tolet genetically heterogeneous populations evolve in different environments andto manage gene flows between populations. This ex situ management developsthe concept of ’mass reservoir’ described by Simmonds (1962). In a first 10-yearperiod, no gene flows were realised in order to allow the study of differentiationand local adaptation with no interference from migration.
After a few generations of cultivation in a French multilocal network,the populations showed significant differentiation for agromorphological andbiochemical traits. These genetic changes are mostly due to the three followingselective pressures:
i) environmental factors, such as climate, which has induced a north-southgradient of flowering time in both PA and PB pools (David et al., 1992);

ii) between-plant competition, which led to the undesirable elimination ofdwarfing genes (rhtl and rht2) segregating in the initial populations (Pontis,1992);
iii) pathogens, as shown by the differentiation of populations for their
composition in resistance genes to powdery mildew (Le Boulc’h, 1994; LeBoulc’h et al., 1994).
Besides these selection pressures, the mating system is of great importancefor the adaptive response of each population, as it plays a major role in thestructuring of within- and between-population diversity, when gene flows areset up between populations.On a within-population scale, a high level of selfing allows the rapid
fixation of the best genotypes and thus a short-term adaptation, whereasoutcrossing maintains genetic diversity and the potential for future adaptation,by producing new recombinant genotypes (Allard, 1979). David et al. (1993)showed that, in a finite population under selection, the optimal adaptiveresponse (accumulation of positive alleles) is obtained with an intermediate
outcrossing rate, whose value depends on the size and fitness of the population.Outcrossing, recombination and natural selection are also critical for purging
populations of low effective sizes (< 1 000) from the genetic load. Recent studiesof natural populations and mutation accumulation experiments have shownthat slightly deleterious mutations (inducing a loss of total fitness of 1 to 5 %)occur at a rate equivalent to 0.5-1 new mutation per diploid genome and pergeneration (Mukai et al., 1972; Lopez and Lopez-Fanjul, 1993; Keightley, 1996).Such empirical data combined with theoretical considerations (Charlesworthet al., 1993) shed new light on what should be the long-term goals of dynamicmanagement of genetic diversity. Mutational load is expected to build uprapidly in a purely selfing population of a few hundred individuals and shouldlead such a population to extinction due to the reduced fertility/viability ofindividuals (phenomenon commonly known as Muller’s ratchet [Muller, 1964!).In this context, a mixed mating system is optimal because it allows bothefficient purging of the genome and recombination (David et al., 1993; Ronfortand Couvet, 1995).On a metapopulation scale, while selection over the metapopulation con-
tributes to the conservation of genetic diversity by the fixation of differentcombinations of adaptive alleles in each site, it also increases the genetic drift,leading to the random loss of alleles with a slightly positive effect on fitness.Genetic drift also leads to the differential fixation of weakly deleterious alle-les randomly appearing through mutation in the different subpopulations. Theoccurrence of interpopulation recombination enables the reintroduction of lostpositive alleles, and to a lesser degree, incorporation of new positive allelesappearing through mutation in other populations. Knowledge of natural out-crossing rate is thus essential to optimise the management of gene flows betweenpopulations and could facilitate its practical realisation.
It is therefore strategic to study the breeding system of the populationscultivated in the wheat dynamic management network. Triticnm aestivnm L.is usually considered as a selfing species because of its cleistogamous flowers.However, outcrossing has been reported, like for other self-fertilising species ofthe Triticeae genus (2-3 % for T. aestivnm L. in Hayes, 1918 and Martin, 1990;1-3 % for T. turgidum L. in Tsegaye, 1996).

This paper gives estimates of outcrossing rates of six genetically heteroge-neous wheat populations of the DM programme submitted to natural selec-tion for 10 years. We used molecular markers to derive outcrossing rates fromthe estimated fixation parameters f (Wright’s statistics). As these populationsoriginate from two segregating populations (PA and PB) cultivated at threedifferent sites, our second aim was to detect if original genetic compositionand cultivation sites influence the outcrossing rate of populations. Lastly, wediscuss the consequences of outcrossing rates for the adaptive response of thewheat populations and consider the use of these natural crossing events for thesetting of gene flows between populations.
2. MATERIALS AND METHODS
2.1. History of populations
Two segregating populations (PA and PB) were created each by a manualpyramidal cross of two distinct sets of 16 inbred lines (for PA, see Thomaset al., 1991). In 1984, after 3 years of bulk multiplication, the populationswere split between, respectively, seven and eight sites of a French multilocalnetwork (either Inra stations or agricultural and agronomic high schools) tosettle local subpopulations. The demographic size of each founding populationwas about 10 000 individuals. At each site, 10 000 to 15 000 plants have beengrown each year in isolated plots, separately harvested and sampled to generatethe next population. As previously mentioned, wheat is usually cleistogamous,with a relatively low pollen/ovule ratio (2 000 to 4 000; Cruden, 1977). Whenoutcrossing occurs, flowers are wind-pollinated.
2.2. Plant material
Six populations were studied after 10 years of multiplication at the samesite: three PA populations cultivated in Le Moulon (48° 9’N 2°E), Rennes (48°1’N 2°W) and Toulouse (43° 6’N 1°E), and three PB populations cultivatedin Le Moulon, Rennes and Venours (45° 8’N 1°W). From the bulk harvest ofthe 1994 season, 78 individuals were randomly sampled and analysed. Addingthe 3 years of multiplication prior to the initial distribution of seeds over thenetwork to the 10-year multiplication period, these populations resulted from13 cycles of natural mating after the last manual crosses. To estimate the initialfrequencies of markers in the founding populations, two random samples ofinbred lines derived by single seed descent (SSD) from the initial populationsPAO and PBO were also studied.
2.3. Molecular study
Because of the low level of polymorphism at the protein level (Gale andSharp, 1988), we used restriction fragment length polymorphism (RFLP)markers, which are codominant and expected to show sufficient polymorphismto reveal heterozygosity.

Total deoxyribonucleic acid (DNA) was extracted from lyophilised youngleaves following a rapid procedure adapted from Dellaporta et al. (1983).Enzyme restriction, electrophoresis, blotting onto Hybond N+ membranes
(Amersham) and nonradioactive hybridisation (DIG@Boehringer-Mannheim)were performed as described by Lu et al. (1994).
P. Leroy provided us with 35 genomic DNA probes, obtained and mappedby Inra-Génoblé of Clermont-Ferrand, France (Nelson et al., 1995a,b). Theseprobes were chosen for their high polymorphism information content (Nelsonet al., 1995a,b) estimated over 14 lines. K. Devos provided us with fiveadditional probes obtained and mapped by the John Innes Centre, Norwich,UK (Gale et al., 1997).
The RFLP diversity of this set of probes was screened on the 32 genitors ofthe PA and PB pools, and the 12 most polymorphic enzyme/probe combina-tions were retained: EcoRl with fba69, fba881 and fbb09; HindIII with fba65,fbal27, fba152, fba!4! and fbbl!; Dra I with fba!04 and fba251 for the Inra-Génoblé probes; EcoRI with PSR 144 and PSR 6!! for the John Innes Centre.Information on the probes can be obtained from the GrainGenes database(http://wheat.pw.usda.gov; French mirror site: http://grain.jouy.inra.fr).
As some probes exhibited polymorphism for nonmapped bands, the relationsof allelism between new segregating bands were tested over the two PAO andPBO SSD and the DM populations.
2.4. Genetic analysis
To estimate the outcrossing rates, we chose measurement of mean heterozy-gosity for two main reasons. First, after 13 seasons of natural mating we ex-pected the neutral markers to be very close to the inbreeding equilibrium, sothat we could use the relationship between t and f. Second, whereas progenyanalysis (Ritland and Jain, 1981) allows unambiguous detection of outcrossingevents, the estimated outcrossing rate is only relevant for one breeding season.Moreover, we needed, for other purposes not developed herein, accurate esti-mates of allelic frequencies more easily achieved in predominantly selfer plantsby random sampling than by progeny analysis.
Outcrossing rates of the populations were estimated from the observedlevel of heterozygosity deficiency f of the sampled genotypes. In a populationof infinite size without over- or underdominant selection, with a constantoutcrossing rate t, the heterozygosity reaches an equilibrium value after a fewgenerations (from one generation for complete outcrossing to ten generations fora 99 % selfing population). As the census size of populations is large (> 10 000)and because of the 13 years of free pollination, we assumed that f was close tothe equilibrium value.
With these assumptions, the estimation of outcrossing rate is given by
(Nei and Syakudo, 1958), where f is the estimated inbreeding coefficient:

Ho is the observed proportion of heterozygotes in the sample,
the gene diversity as defined by Nei (1958), n, the number of alleles for a givenlocus and pi the estimated frequency of the allele i.
f was estimated by the maximum likelihood method proposed by Li andHorovitz (1953) and Curie-Cohen (1982) for monolociis data. Assuming thatthere is no gametic disequilibrium, multilocus f is the root of
(Rousset and Raymond, 1995), where NHet is the number of heterozygotesat locus I, n,zi the number of homozygotes for allele i at locus and pi thefrequency for allele i at the locus I observed in the sample.
Estimate f was numerically computed after estimation of the allelic fre-
quencies, and used to calculate the t value. A bootstrap resampling procedureover individuals was performed to determine the confidence interval of t (Weir,1990). For each population, 500 bootstrap samples were used to calculate adistribution of values (b = 1, ..., 500).
To test the significance of differences in outcrossing rates (ti < tj) betweenpopulations i and j, pairwise subtraction between bootstrapped values ofeach population, 0,b - 4,b allowed us to perform a one-tailed test (Efronand Tibshirani, 1982). The percentage of negative differences provided theprobability associated with the test Ho: ti = tj against H1: ti < t!.
To test the effects of the genetic composition (PA versus PB) and of thesite (Le Moulon versus Rennes), the bootstrap distributions of the followingcontrasts were calculated:
3. RESULTS
3.1. Polymorphism of RFLP markers
Most of the RFLP genomic probes used for this study revealed multibandprofiles, due to the hexaploid constitution of wheat. Over the 12 assayed probes,23 codominant loci were examined, some probe/enzyme combinations showingup to five polymorphic loci. Thus, a single hybridisation led to a mean of1.9 data points per individual. For many probes, new segregating bands were

revealed in comparison with the mapping populations (Nelson et al., 1995a,b).Consequently, the mapping of these 23 loci on the genetic maps was not alwayspossible. The Mendelian inheritance of the new polymorphic bands was verifiedusing the segregation data of the SSD populations extracted from the initialpopulations in which a high level of fixation (96 %) allowed unambiguousdetermination of allelism relationships (results not shown). Over the 23 loci,the mean allelic diversity was 2.48, ranging from two to five alleles per locus(table 1). The mean gene diversity was He = 0.35, ranging from 0.09 to 0.59.
3.2. Outcrossing rates
The heterozygosity observed after 13 generations was much higher thanexpected: 1 to 5 % of mean heterozygosity was measured (data not shown),instead of 0.006 % (2!3 ) expected for strict self-pollination. Values of single213locus estimates of outcrossing rates ranged from 0 to 30 %, most of them beinglower than 10 % (figure 1). The highest outcrossing rates were observed for lociwith a poor gene diversity, and thus a high standard error.
Analysis of variance (ANOVA) over the single-locus estimates (data notshown) did not reveal a significant RFLP effect: no locus exhibited consistentlyhigh or low levels of heterozygosity across the four populations. This indicatedthat no RFLP locus was in linkage disequilibrium with polymorphic genescontrolling the outcrossing rate or submitted to overdominant selection in allpopulations.
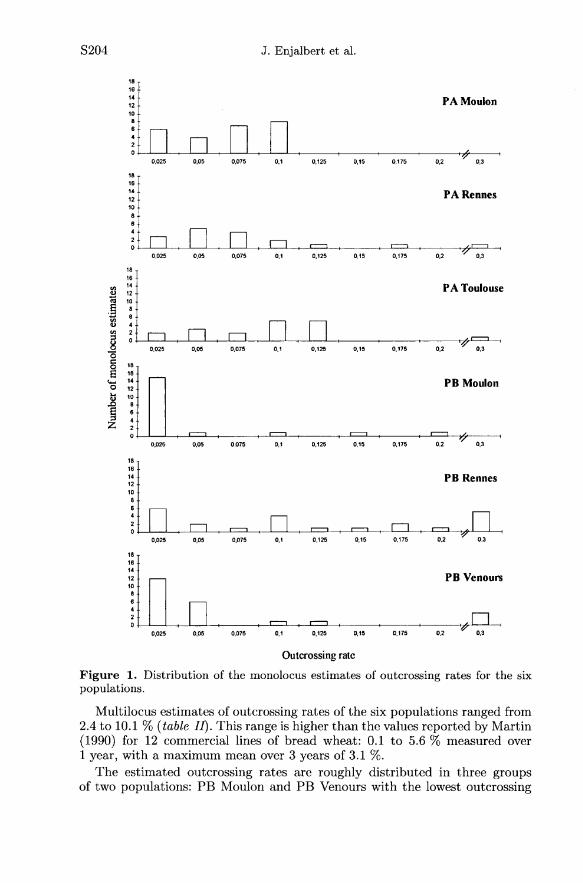
Multilocus estimates of outcrossing rates of the six populations ranged from2.4 to 10.1 % (table 77). This range is higher than the values reported by Martin(1990) for 12 commercial lines of bread wheat: 0.1 to 5.6 % measured over1 year, with a maximum mean over 3 years of 3.1 %.
The estimated outcrossing rates are roughly distributed in three groupsof two populations: PB Moulon and PB Venours with the lowest outcrossing

rates (around 3 %), PA Moulon and PA Rennes with intermediate values (near6 %) and PB Rennes and PA Toulouse with the highest rates (around 10 %).According to comparison tests, PB Moulon differed significantly from all theothers with the exception of PB Venours, a population which, in turn, differssignificantly from all the others except PA Moulon, all other comparisons beingnot significant.A significant site effect was found, with Pr[6site < 0] = 0.004, whereas no
pool effect was detected (Pr[bpool < 0] = 0.4). Nevertheless, site-pool interac-tion was present, as the PA and PB populations had inverse classifications inRennes and Le Moulon.
4. DISCUSSION
Our results showed that the wheat populations of the DM network cultivatedunder natural conditions, though cleistogamous, can exhibit a relatively highoutcrossing rate (ranging from 2.4 to 10.1 %). These estimations rely upon theassumption of selective neutrality and consequently could be biased upwarddue to the selective advantage of heterozygotes in populations in which strongcompetition takes place between individuals. In winter wheat, low heterosis(! 10 %) has been reported for yield surveyed on plots of homogeneous geneticcomposition (Oury et al., 1990), but we might expect heterosis for individ-ual fitness in heterogeneous populations to be stronger, since hybrids are, onaverage, taller and bloom earlier than their inbred parents, both traits con-tributing to a better competitive aptitude. Even with 20 % heterosis on fitnessexplained by a single-locus model, however, 13 cycles of complete selfing onlyraised the heterozygote frequency from 0.006 to 0.08 % (data not shown), avalue significantly lower than the rates observed in this study. This theoreticalconsideration, associated with the observation that no locus exhibited signifi-cant excess of heterozygotes across all studied populations, leads us to arguethat the outcrossing rate of our populations is often as high as several percent,ranging from 2 to 10.

Such levels of outcrossing are commonly reported in other species classicallydescribed as selfing (table I in Schoen and Brown, 1991). In the few highlyselfing species (outcrossing rate < 1 %), such as Arabidopis thaLiana or someAvena species, outcrossing events might be due to accidents in the developmentof some anthers or to the exceptional infiltration of exogenous pollen and mighthave low evolutionary impact. On the other hand, outcrossing rate, even as lowas 5 %, should be explained by different mechanisms.
4.1. Origin and stability of outcrossing ratesin dynamically managed populations
In some grasses, environment has a strong effect on fertility and outcross-ing propensity. Low luminosity and low temperature during meiosis have beenshown to cause pollen sterility in rice (Li et al., 1996) and winter wheat(Demotes-Mainard et al., 1995). Demotes-Mainard et al. (1995) showed thatthis sensitivity to low luminosity has a genetic basis, the two studied lines Per-nel and Moulin exhibiting very contrasted responses to the stress. This phys-iological response should lead to strong year-to-year and site-to-site variationin the rate of outcrossing. Contrasted climatic conditions of the DM networkare most likely to affect the amount of outcrossing realised in the populations,either through partial male sterilities or by shifting of male and female phe-nologies. Thus, one explanation for the high outcrossing rate of populationsgrown in Rennes could be the cloudy weather in spring and in the beginningof the summer in the west of France. On the other hand, Henry et al. (1991),by studying a locus coding for seed storage protein, have previously shownthat populations grown in the south of France displayed significant excesses ofheterozygotes as compared with populations grown in the north. This was ex-plained by a protogynous flowering at high temperatures. Our results, in accordwith this previous study, underline the importance of environmental factors forthe mating system.
Although we did not detect an effect of the genetic background (PA or PB)on the outcrossing ability, the PA population of Le Moulon showed a signifi-cantly higher rate of outcrossing than PB Le Moulon, and the two PB popu-lations of Le Moulon and Venours were the most selfing populations. A ratherhigh level of outcrossing has also been observed in a recurrent selection pro-gramme based on the same genetic background as the PA populations. Spikesor spikelets have often been found with open flowers receptive to exogenouspollen, indicating that no viable pollen was available for a cleistogamous polli-nation, a phenomenon well known in wheat. This trend to partial male sterilitymay be due to the presence in the genitors of PA of the Courtot cultivar, a lineknown for its high degree of pollen sterility. Savy (1991) showed that within apopulation the individuals presented different rates of selfing, with the greatestproportion of plants being mainly selfing, with some individuals showing com-plete outbreeding. Thus, genetic and environmental factors probably interactto determine individual levels of outcrossing.
If site-to-site variation of outcrossing rates could be estimated, the year-to-year or plant-to-plant variations need to be evaluated because they willaffect the efficiency of gene flow in schemes based on the population’s meanoutcrossing rate.

However, which forces maintain the outcrossing ability of these populations ?Simple theoretical models for the evolution of selfing predict that a selfingvariant should spread due to its segregational advantage as soon as inbreedingdepression (defined as one minus the relative fitness of selfed versus outcrossedprogeny) does not exceed one half (Fisher, 1958). Other theoretical models(assuming population of finite size or spatial structuration) show that evenwith low heterosis for fitness, recombination is advantageous and maintainsevolutionary stable rates of outcrossing at a few percent (David et al., 1993;Ronfort and Couvet, 1995). If the optimal multilocus genotype is not initiallypresent in a population, the selection for favourable recombinant types slowsdown the counterselection of outcrossing abilities within the population. In thepresent study, the low heterosis of wheat combined with significant outcrossingrates led us to the conclusion either that a large part of the population mustexhibit a few percent outcrossing, or that the heritability for high outcrossingrates must be low. Otherwise outcrossing plants should have been quicklyeliminated and the outcrossing rates lower than reported here.
Beyond the exact nature of the selective forces acting on the mating system,outcrossing is expected to play a major role in the local adaptation of thepopulations of the DM network. By allowing occasional recombination eventsbetween individuals, such levels of outcrossing enable the production of betteradapted genotypes (Allard, 1979) and simultaneously prevent rapid loss offavourable alleles trapped in inferior lines that would otherwise be wiped out ofthe population. Hence, comparing the level of diversity maintained on polygenictraits and the structure of genetic correlations between traits, a population witha mixed mating system involving a few percent outcrossing is qualitativelydifferent from a pure selfing one.
4.2. Consequences for managing the DM network
The diversity of selective pressures present at the various sites of the net-work is a warrant for the maintenance of polymorphism on loci involved inadaptation (Henry et al., 1991). However, we argue that gene flows betweenpopulations are necessary to compensate for the variability lost within popu-lations through drift and linkage drag due to selection. A rule of thumb couldbe the preservation of a certain level of within-population polymorphism forneutral traits, expecting that in case of change in selection gradients thesepreviously neutral traits become adaptive. Several examples can be given toillustrate this possible feature. First, dramatic changes in environmental con-ditions can induce inversions in the hierarchy of the traits determining localfitness. Second, heterogeneous populations can experiment different successivephases of selection. When the adaptation for an important trait is finally re-alised by the exhaustion of the variability for that trait, natural selection willact more strongly on another trait which was of minor importance in the firststage of adaptation. In these cases of change in selective gradients, gene flowswould allow local populations to enrich their variability for secondary traits,as they restart selection on primary traits when their variability has been ex-hausted by linkage drag or genetic drift. On the other hand, it is importantnot to impede local adaptation of populations to their site because of too highmigration rates (Harrison and Hasting, 1996). By using the neutral theory of

evolution as a guideline, the intensity of gene flow could be determined in orderto limit the level of between-population differentiation on neutral genes, i.e. toensure a minimum rate of migration m, so that Ne*m = 1, where Ne is thelocal effective size of a population (Crow and Kimura, 1970). However, a highermigration could be necessary to preserve diversity of chromosomal segments inthe neighbourhood of genes under strong selection.
To favour recombination between individuals, pollen migration should befavoured instead of seed migration. In outcrossing species both modes of
migration are expected to produce similar effects (except for cytoplasmicgenomes), but markedly different consequences are expected in the case ofselfers. Seed migration could potentially split the recipient population into twocompartments. If migrant genotypes are poorly adapted to local conditions,hybrid/outbreeding depression is expected and the efficiency of gene transfervia such schemes will be low. To circumvent such problems, Hauser et al. (1994)suggested to realise back crosses instead of first-generation hybrids to enhancegenetic diversity in endangered populations. On the other hand, when somemigrants display more competitive ability, they could potentially outcompeteresident genotypes and eliminate the initial variability present in the population(especially if migrant and resident genotypes hardly recombine). Migration viapollen should avoid these phenomena by enforcing recombinatioa of residentand migrant genotypes.On the basis of the data obtained, natural outcrossing rates are sufficient
to allow the incorporation of foreign migrants by the local population. Toachieve a level of gene flow of one effective migrant per generation (neutralistguideline), two outcrossings per migrant pollen per population are required.Assuming a 5 % rate of outcrossing in a population composed of 10 000 plants,500 individuals will potentially outcross. To ensure two crosses, migrant pollenshould thus be provisioned as to represent 1/250 of the total pollen pool. Forwheat, this should be easily realised by surrounding populations with a belt ofmigrant plants.
Estimation of rates of natural outcrossing in populations combined withthe knowledge of local effective sizes will make it possible to rationalise themanagement of the network. Such optimisation should be achieved by realexperiments, analytical approaches and through modelling based on realisticparameters.
ACKNOWLEDGEMENTS
The authors are grateful to T. Bataillon and D. Schoen for scientific commentsand for help in the translation of the paper, and they thank the anonymous reviewersfor very helpful comments on the manuscript.
The wheat dynamic network was initiated by the ’Direction générale de 1’enseigne-ment et de la recherche’ (DGER) of the French agriculture ministry and receivedscientific and financial support from the DGER, ’Institut national de la rechercheagronomique’ and ’Bureau des Ressources Génétiques’ (BRG). This work is supportedby a grant from the BRG. J. Enjalbert is the recipient of a doctoral fellowship fromthe French research and higher education ministry.

REFERENCES
Allard R.W., The mating system and microevolution, Genetics 79 (1979) 115-126.Charlesworth D., Morgan M.T., Charlesworth B., Mutation accumulation in finite
outbreeding and inbreeding populations, Genet. Res. 61 (1993) 39-56.Crow J.F., Kimura M., An Introduction to Population Genetics Theory, Harper
International Edition, New York, 1970.Cruden R.W., Pollen-ovule ratios: a conservative indicator of breeding systems in
flowering plants, Evolution 31 (1977) 32-46.Curie-Cohen M., Estimate of inbreeding in natural populations: a comparison of
sampling properties, Genetics 100 (1982) 339-358.David J.L., Savy J., Trottet M., Pichon M., Méthode de gestion dynamique de la
variabilité génétique en milieu naturel. Exemple de populations composites de ble, in:Colloque international en hommage à Jean Pernes, 8-10 janvier 1992, Paris, France,1992, pp. 337-350.
David J.L., Savy Y., Brabant P., Outcrossing and selfing evolution in populationsunder directional selection, Heredity 71 (1993) 642-651.
Dellaporta S., Wood J., Hicks J., A plant DNA minipreparation: version II, PlantMol. Biol. Rep. 1 (1983) 19-21.
Demotes-Mainard S., Doussinault G., Meynard J.M., Effect of low radiation andlow temperature at meiosis on pollen viability and grain set in wheat, Agronomie 15(1995) 357-365.
Devos K., Gale M.D., The genetic maps of wheat and their potential in plantbreeding, Outlook Agric. 22 (1993) 93-99.
Efron B., Tibshirani R.J., An Introduction to the Bootstraps, Chapman and Hall,New York, 1982.
Fisher R.A., The Genetical Theory of Natural Selection, Oliver and Boyd, London,1958.
Frankel O.H., Brown A.H.D., Burdon J.J., The Conservation of Plant Biodiversity,Cambridge University Press, Cambridge, 1995.
Gale M.D., Sharp P.J., Genetic markers in wheat-developments and prospects,in: Seventh International Wheat Genetic Symposium, Cambridge, 13-19 July 1988,Institute of Plant Science Research, 1988, pp. 469-473.
Gale M.D., Atkinson M.D., Chinoy C.N., Harcourt R.L., Jia J., Li Q.Y., DevosK.M., Genetic maps of hexaploid wheat, in: Eighth International Wheat GeneticsSymposium, Beijing, China, 1997, pp. 29-40.
Harrison S., Hasting A., Genetic and evolutionary consequences of metapopulationstructure, Tree 11 (1996) 180-183.
Hauser T.P., Damgaard C., Loeschcke V., Effects of Inbreeding in Small PlantPopulations: Expectations and Implications for Conservation, Birkhauser Verlag,Basel, 1994.
Hayes H.K., Natural crossing in wheat. A cause of impurities in breeding plots-belief of some agronomists that hybrids frequently revert to the parental type, J.Hered. 9 (1918) 326-330.
Henry J.P., Pontis C., David J.L., Gouyon P.H., An experiment on dynamicconservation of genetic resources with metapopulations, in: Seitz A., Loeschcke V.(Eds.), Species Conservation: A Population-Biological Approach, Birkhaiiser Verlag,Basel, 1991.
Keightley P.D., Nature of mutation load in Drosophila, Genetics 144 (1996) 1993-1999.

Le Boulc’h V., Evolution de la resistance à 1’6idium (Erysiphe graminis) dans despopulations de ble tendre ( Z’riticum aestivum L.) menées en gestion dynamique, these,Institut national agronomique Paris-Grignon, Paris, 1994.
Le Boulc’h V., David J.L., Brabant P., Vallavieille-Pope C., Dynamic conservationof variability: response of wheat populations to different selective forces includingpowdery mildew, Genet. Sel. Evol. 26 (1994) 221-240.
Levins R., Extinction, in: Gerstenhaber M. (Ed.), Some Mathematical Questionsin Biology, American Mathematical Society, Providence, RI, 1970, pp. 77-107.
Li C.C., Horovitz D., Some methods of estimating the inbreeding coefficient, Am.J. Hum. Genet. 5 (1953) 107-117.
Li H.B., Zhang (a., Liu A.M., Zou J.S., Chen Z.M., A genetic analysis of low-temperature-sensitive sterility in indica-japonica rice hybrids, Plant Breeding 115(1996) 305-309.
Lopez M.A., Lopez-Fanjul C., Spontaneous mutation for a quantitative trait inDrosophila melav,ogaster. II. Distribution of mutant effects on trait and fitness, Genet.Res. 61 (1993) 117-126.
Lu Y.H., Merlino M., Isaac P.G., Staccy J., Bernard M., Leroy P., A comparativeanalysis between 32p and digoxigenin-labelled single-copy probes for RFLP detectionin wheat, Agronomic 14 (1994) 33-39.
Martin T.J., Outcrossing in twelve hard red winter wheat cultivars, Crop Sci. 30(1990) 59-62.
Mukai T., Chigusa S.I., Mettler L.E., Crow J.F., Mutation rate and dominance ofgenes affecting viability, Genetics 72 (1972) 335-355.
Muller H.J., The relation of recombination to mutational advance, Mutat. Res. 1(1964) 2-9.
Nei M., Syakudo M., The estimation of outcrossing in natural populations, Jpn.J. Genet. 33 (1958) 46-51.
Nelson J.C., Sorrells M.E., Van Deynze A.E., Lu Y.H., Atkinson M., Bernard M.,Leroy P., Faris J.D., Anderson J.A., Molecular mapping of wheat: major genes andrearrangements in homoeologous group 4, 5 and 6, Genetics 141 (1995a) 721-731.
Nelson J.C., Van Deynze A.E., Autrique E., Sorrells M.E., Lu Y.H., Negre S.,Bernard M., Leroy P., Molecular mapping of wheat. Homoeologous group 3, Genome38 (1995b) 525-533.
Olivieri 1., Couvet D., Gouyon P.-H., The genetic of transient populations: researchat the metapopulation level, Tree 5 (1990) 207-210.
Oury F.-X., Brabant P., Pluchard P., B6rard P., Rousset M., Etude multilocale deblés hybrides : niveaux d’heterosis et elaboration du rendement, Agronomie 10 (1990)735-748.
Pontis C., Utilisation de marqueurs génétiques pour le suivi de la variabilité detrois composites de ble tendre d’hiver (T. aestivam L.) menées en gestion dynamique,these, Institut national agronomique Paris-Grignon, Paris, 1992.
Ritland K., Jain S., A model for the estimation of outcrossing rate and genefrequencies using n independent loci, Heredity 47 (1981) 35-52.
Ronfort J., Couvet D., A stochastic model of selection on selfing rates in structuredpopulations, Genet. Res. 65 (1995) 209-222.
Rousset F., Raymond M., Testing heterozygote excess and deficiency, Genetics 140(1995) 1413-1419.
Savy Y., Evolution de la variabilité génétique en selection naturelle en relation avecle regime de reproduction : étude agronomique de trois pools géniques de ble tendre,étude théorique par simulations informatiques, Rapport de DEA, Institut nationalagronomique Paris-Grignon, Paris, 1991.
Schoen D., Brown A., Intraspecific variation in population gene diversity andeffective population size correlates with the mating system in plants, Proc. Natl.Acad. Sci. USA 88 (1991) 4494-4497.

Simmonds N.W., Variability in crop plants, its use and conservation, Biol. Rev. 37(1962) 442-465.
Thomas G., Rousset M., Pichon M., Trottet M., Doussinault G., Picard E.,Méthodologie de I’amelioration du ble tendre (T. aestivum L.). I. Creation parcroisements et analyse d’une population artificielle à 16 parents, base de cette étudeméthodologique, Agronomie 11 (1991) 359-368.
Tsegaye S., Estimation of outcrossing rate in landraces of tetraploid wheat(Triticum turgidum L.), Plant Breeding 115 (1996) 195-197.
Weir B.S., Genetic Data Analysis, Sinauer Associates, Inc., Sunderland, MA, 1990.







![(PME-PBO (PME-PBO$BSEMFTTvmobile.topica.ne.jp/ebook/pdf/goldloan.pdf(PME-PBO (PME-PBO$BSEMFTT ~ª E Å çÅé ï yyyyyÍ ~ª E Å çÅé ï Åèµ] b ;¨ Å] b ;w ²t Ù c± ï j Øw]](https://static.fdocuments.in/doc/165x107/5cdb825888c99386458cc987/pme-pbo-pme-pbo-pme-pbo-pme-pbobsemftt-a-e-a-cae-i-yyyyyi-a-e-a.jpg)






![Chapter 03.ppt [Mode de compatibilité] - udifmv.ulg.ac.be · Exppgerimental design Pea cultivation Exppgerimental design Pure lines Exppgerimental design Selfing vs outcrossing Mendel’s](https://static.fdocuments.in/doc/165x107/5d651f3b88c993fb748ba81e/chapter-03ppt-mode-de-compatibilite-exppgerimental-design-pea-cultivation.jpg)



