
Original Article miR-185 acts as a tumor suppressor by … · miR-185 acts as a tumor suppressor by...
9
Int J Clin Exp Pathol 2015;8(9):11854-11862 www.ijcep.com /ISSN:1936-2625/IJCEP0011167 Original Article miR-185 acts as a tumor suppressor by targeting AKT1 in non-small cell lung cancer cells Shuai Li, Yulian Ma, Xinfang Hou, Ying Liu, Ke Li, Shuning Xu, Jufeng Wang Department of Internal Medicine, Affiliated Cancer Hospital of Zhengzhou University, Henan Cancer Hospital, Zhengzhou, Henan, China Received June 7, 2015; Accepted July 21, 2015; Epub September 1, 2015; Published September 15, 2015 Abstract: Increasing evidence has shown that microRNAs play critical roles in the initiation and progression of non- small cell lung cancer (NSCLC). miR-185 is deregulated in various cancers, whereas its functional mechanism in NSCLC is still unclear. Here, we confirmed that the expression of miR-185 was significantly down-regulated in NSCLC tissues and cell lines. miR-185 over-expression caused significant suppression of in vitro cell proliferation, migration and invasion, and in vivo tumor growth. We subsequently identified that AKT1 was a target gene of miR-185. Re- expression of AKT1 could partially rescue the inhibitory effects of miR-185 on the capacity of NSCLC cell prolifera- tion and motility. Collectively, we conclude that miR-185 has a critical function by blocking AKT1 in NSCLC cells, and it may be a novel therapeutic agent for miRNA based NSCLC therapy. Keywords: miR-185, AKT1, non-small cell lung cancer Introduction Lung cancer is the most common malignant tumors in humans and the leading cause of cancer-related deaths in the world [1]. Non- small cell lung cancer (NSCLC) accounts for at least 80% of all lung cancers [2]. Despite diag- nosis and clinical treatment strategies have been significantly improved, poor prognosis still exists in many NSCLC patients with a low 5-year overall survival rate and a high recurrence rate [3, 4]. Therefore, it is utterly important to under- stand the precise molecular mechanisms of the tumorigenesis of NSCLC. Recently, a large number of studies have shown that microRNAs (miRNAs) play an essential role in NSCLC patho- genesis, which provides new insights into the therapeutic strategy against this disease [5, 6]. MiRNAs are a family of small (approximately 22 nucleotides in length), single-stranded, endog- enous non-coding RNAs which regulate gene expression by causing mRNA degradation or repressing translation though binding to com- plementary sites in 3’-untranslated regions (3’- UTRs) [7-9]. miRNAs are implicated in a wide range of important physiological processes [10-12]. Emerging evidence demonstrates that miRNAs act as novel oncogenes or tumor sup- pressors, and aberrant miRNA expression has been observed in various types of human can- cers including NSCLC [13-15]. Recently, miR- 185 has been reported to be down-regulated in gastric cancer, breast cancer, glioma, clear cell renal cell carcinoma and hepatocellular carci- noma [16-20]. However, the functional mecha- nisms of miR-185 in NSCLC are largely unknown. In the current study, we confirmed that miR-185 was significantly down-regulated in NSCLC tis- sues and cell lines. Over-expression of miR-185 suppressed NSCLC cell growth, migration and invasion in vitro, and tumor growth in nude mice. AKT1 was recognized as a direct and functional target of miR-185. Over-expression of AKT1 could partially rescue the inhibitory effects of miR-185. Our results suggest that miR-185 and its downstream target gene AKT1 may be useful for treatment of NSCLC in the future. Materials and methods Tissue samples Twelve paired NSCLC tissues and adjacent non- tumor tissues used in this study were collected
Transcript of Original Article miR-185 acts as a tumor suppressor by … · miR-185 acts as a tumor suppressor by...

Int J Clin Exp Pathol 2015;8(9):11854-11862www.ijcep.com /ISSN:1936-2625/IJCEP0011167
Original Article miR-185 acts as a tumor suppressor by targeting AKT1 in non-small cell lung cancer cells
Shuai Li, Yulian Ma, Xinfang Hou, Ying Liu, Ke Li, Shuning Xu, Jufeng Wang
Department of Internal Medicine, Affiliated Cancer Hospital of Zhengzhou University, Henan Cancer Hospital, Zhengzhou, Henan, China
Received June 7, 2015; Accepted July 21, 2015; Epub September 1, 2015; Published September 15, 2015
Abstract: Increasing evidence has shown that microRNAs play critical roles in the initiation and progression of non-small cell lung cancer (NSCLC). miR-185 is deregulated in various cancers, whereas its functional mechanism in NSCLC is still unclear. Here, we confirmed that the expression of miR-185 was significantly down-regulated in NSCLC tissues and cell lines. miR-185 over-expression caused significant suppression of in vitro cell proliferation, migration and invasion, and in vivo tumor growth. We subsequently identified that AKT1 was a target gene of miR-185. Re-expression of AKT1 could partially rescue the inhibitory effects of miR-185 on the capacity of NSCLC cell prolifera-tion and motility. Collectively, we conclude that miR-185 has a critical function by blocking AKT1 in NSCLC cells, and it may be a novel therapeutic agent for miRNA based NSCLC therapy.
Keywords: miR-185, AKT1, non-small cell lung cancer
Introduction
Lung cancer is the most common malignant tumors in humans and the leading cause of cancer-related deaths in the world [1]. Non-small cell lung cancer (NSCLC) accounts for at least 80% of all lung cancers [2]. Despite diag-nosis and clinical treatment strategies have been significantly improved, poor prognosis still exists in many NSCLC patients with a low 5-year overall survival rate and a high recurrence rate [3, 4]. Therefore, it is utterly important to under-stand the precise molecular mechanisms of the tumorigenesis of NSCLC. Recently, a large number of studies have shown that microRNAs (miRNAs) play an essential role in NSCLC patho-genesis, which provides new insights into the therapeutic strategy against this disease [5, 6].
MiRNAs are a family of small (approximately 22 nucleotides in length), single-stranded, endog-enous non-coding RNAs which regulate gene expression by causing mRNA degradation or repressing translation though binding to com-plementary sites in 3’-untranslated regions (3’-UTRs) [7-9]. miRNAs are implicated in a wide range of important physiological processes [10-12]. Emerging evidence demonstrates that
miRNAs act as novel oncogenes or tumor sup-pressors, and aberrant miRNA expression has been observed in various types of human can-cers including NSCLC [13-15]. Recently, miR-185 has been reported to be down-regulated in gastric cancer, breast cancer, glioma, clear cell renal cell carcinoma and hepatocellular carci-noma [16-20]. However, the functional mecha-nisms of miR-185 in NSCLC are largely unknown.
In the current study, we confirmed that miR-185 was significantly down-regulated in NSCLC tis-sues and cell lines. Over-expression of miR-185 suppressed NSCLC cell growth, migration and invasion in vitro, and tumor growth in nude mice. AKT1 was recognized as a direct and functional target of miR-185. Over-expression of AKT1 could partially rescue the inhibitory effects of miR-185. Our results suggest that miR-185 and its downstream target gene AKT1 may be useful for treatment of NSCLC in the future.
Materials and methods
Tissue samples
Twelve paired NSCLC tissues and adjacent non-tumor tissues used in this study were collected

miR-185 acts as a tumor suppressor by targeting AKT1 in NSCLC
11855 Int J Clin Exp Pathol 2015;8(9):11854-11862
from patients who underwent surgical resec-tion at Affiliated Cancer Hospital of Zhengzhou University. Tissue samples were immediately snap-frozen and stored in liquid nitrogen until total RNAs or proteins were extracted. Informed consent was obtained from all patients, and the research was approved by the Ethics Committee of Zhengzhou University.
Cell lines and cell culture
All cell lines (A549, H460, H1299 and 16HBE) were obtained from the Cell Resource Center, Shanghai Institute of Biochemistry and Cell Biology at the Chinese Academy of Sciences. All cells were maintained in RPMI-1640 (GIBCO BRL, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS), and incubated in a humidified atmosphere at 37°C under 5% CO2.
RNA extraction and quantitative real-time PCR (qRT-PCR)
Total RNA from tissues or cells was extracted using TRIzol (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocol. Total RNA was transcribed into cDNA using Prime Script RT reagent Kit (Takara, Dalian, China). Total miRNA was transcribed into cDNA using One Step Prime Script miRNA cDNA Synthesis Kit (Takara). qRT-PCR was carried out using a SYBR Premix Ex Taq II (TAKARA, Dalian, China) on ABI 7500 HT system (Applied Biosystems, Foster City, CA, USA). The relative genes expres-sion was calculated the 2-ΔΔCt method. β-actin or U6 was used as endogenous control [21].
Construction of plasmids
miR-185 mimic and negative control were pur-chased from RiboBio (Guangzhou, China). The 3’UTR segment of the human AKT1 (containing the predicted miR-185 binding site) was PCR-amplified from genomic DNA and inserted into the pGL3 vectors (Promega, Madison, WI, USA). The corresponding sequences carrying the miR-185 binding site were mutated by the QuickChange XL Site-Directed Mutagenesis Kit (Stratagene, La Jolla, CA, USA) and inserted into the pGL3 vectors. For AKT1 over-expr- ession, open reading frame cDNA for AKT1 was inserted into pcDNA3.1 vector (Invitrogen). Transfection was done using the Lipofectamine 2000 (Invitrogen) according to the manufactur-er’s protocol.
MTT assay
Cell viability was detected by the MTT assay. After being seeded onto the 96-well culture plates for 24 h, H1299 and A549 cells were transfected with miR-185 mimic or negative control. 1, 2, 3 and 4 days after transient trans-fection, cells were incubated with 20 μl MTT (5 mg/ml) at 37°C for another 4 h. The media was discarded and 150 μl DMSO was added to the cells. Optical density values were examined at 490 nm wavelength with a microplate reader (Bio-Rad, Richmond, CA, USA).
Tumor xenografts
All animal experiments were performed accord-ing to the protocols approved by The Animal Care and Use Committee of Fudan University, China. BALB/c nu/nu female mice (4-week old) were purchased from Experimental Animal Center of Fudan University. 2.5 × 106 H1299 cells treated with miR-185 mimic or negative control were injected subcutaneously into the mice. Tumor growth was monitored by measur-ing maximum (a) and minimum (b) length of the tumor every 5 days. Tumor volume (V) was calculated as follows: V = ½ ab2. After 30 days, all mice were sacrificed, and tumors were collected and weighed.
Wound-healing and transwell invasion assays
For wound-healing assay, 1 × 106 cells were seeded in a six well plate, cultured for 12 h and transfected with miR-185 mimic or negative control. Upon reaching the appropriate conflu-ence, the confluent monolayer was scratched with a 200 μl pipette tip, washed with PBS twice and cultured again for up to 24 h with serum-free medium. Images of the wound clo-sure were taken at 0 and 24 h under a micro-scope and the width of the wound gaps were validated with Image J software and normalized to 0 h wounds [22]. For the invasion assay, 5 × 104 cells in serum-free RPMI-1640 were placed into the upper chamber of each insert with Matrigel (BD Biosciences, SanJose, CA, USA). RPMI-1640 with 10% FBS was added in the lower chamber to act as the nutritional attrac-tant. After 48 h, the cells that had invaded through the membrane were fixed with 20% methanol and stained with 0.1% crystal violet for 30 min, imaged, and counted using a microscope.

miR-185 acts as a tumor suppressor by targeting AKT1 in NSCLC
11856 Int J Clin Exp Pathol 2015;8(9):11854-11862
Luciferase assay
293 T cells were cultured in 24-well plates. After 24 h, the wide-type or mutated AKT1 3’UTR constructs was co-transfected with miR-185 mimic or negative control and pRL-TK vector using Lipofectamine 2000 (Invitrogen). Firefly and renilla luciferase activities of the cell lysates were detected at 48 h post-transfection using the Dual-Luciferase Reporter Assay System (Promega). Renilla luciferase served as an internal control. The relative luciferase activ-ity was a ratio of firefly and renilla luciferase activity for each sample.
Western blotting
Cells were collected and lysed with cell lysis buffer containing protease inhibitors. Protein concentrations were measured using the BCA method (Thermo Scientific, Rockford, IL, USA). Equal amounts of proteins was resolved on SDS-PAGE gels and transferred onto polyvinyli-dene difluoride membranes (Millipore, Bedford, MA, USA). The membranes were blocked with 5% non-fat milk, incubated overnight at 4°C with AKT1 antibody (1:500, Abcam, Cambridge, UK) and GAPDH (1:1000, Santa Cruz Bio- technology, Santa Cruz, CA, USA), and then with the secondary antibody. Signal was visualized with enhanced chemiluminescence (Millipore) [23].
Statistical analysis
All statistical analyses were analyzed using SPSS 16.0 software. Quantitative data are expressed as mean ± standard deviation. Differences between two groups were made using the Student’s t-test. P values less than 0.05 were considered statistically significant.
Results
The expression levels of miR-185 in NSCLC tissues and cell lines
To understand the relationship between miR-185 and NSCLC, we first examined miR-185 expression in NSCLC tissues and their matched non-tumor tissues using qRT-PCR. The results showed that miR-185 was down-regulated in NSCLC samples compared with the non-tumor counterpart (Figure 1A). We further confirmed the miR-185 level in three human NSCLC cell lines including H460, A549 and H1299. As shown in Figure 1B, all three human NSCLC cell lines expressed much lower levels of miR-185 compared to those in the normal lung bronchus epithelial cell line 16HBE. The low miR-185 expression in NSCLC strongly suggests that miR-185 might be involved in the pathogenesis and development of NSCLC.
Figure 1. The expression levels of miR-185 in NSCLC tissues and cell lines. A. qRT-PCR was performed to measure relative miR-185 expression in 12 paired NSCLC tissues and adjacent non-tumor tissues. U6 was used as an inter-nal control. B. Relative miR-185 expression in 3 NSCLC cell lines and the normal lung bronchus epithelial cell line 16HBE. Each sample was analyzed in triplicate. *P < 0.05, **P < 0.01.

miR-185 acts as a tumor suppressor by targeting AKT1 in NSCLC
11857 Int J Clin Exp Pathol 2015;8(9):11854-11862
miR-185 inhibits NSCLC cell proliferation in vitro and in vivo
To investigate the role of miR-185 in NSCLC cells, we first evaluated the effects of miR-185 on NSCLC proliferation using a gain-of-function approach. H1299 and A549 cells were trans-fected with miR-185 mimic or negative control, and over-expression of miR-185 was confirmed by qRT-PCR (Figure 2A). miR-185 over-expres-sion in both cell lines resulted in decreased cell viability at 96 h post-transfection, as detected by MTT assays (Figure 2B). To further evaluate
the effect of miR-185 on tumorigenicity, miR-185-transfected H1299 cells or controls were subcutaneously injected into the nude mice. After 30 days, xenografted tumor volume in miR-185 over-expressing cells was significantly smaller than that in the controls (Figure 2C). Similarly, the tumors in the control group grew much faster and had much heavier tumor weights than those in miR-185-transfected group (Figure 2D). Collectively, these data dem-onstrate that miR-185 over-expression sup-presses NSCLC cell proliferation in vitro and in vivo.
Figure 2. miR-185 inhibited NSCLC cell proliferation in vitro and in vivo. A. miR-185 over-expression in H1299 and A549 cells was confirmed by qRT-PCR. B. Effects of over-expression of miR-185 on the proliferation of H1299 and A549 cells were performed using MTT assay. C. Growth curves for tumor volumes in the nude mice with H1299 cells treated with miR-185 mimic or negative control. D. Representative images and tumor weights of xenograft tumors. Each sample was analyzed in triplicate. *P < 0.05, **P < 0.01.

miR-185 acts as a tumor suppressor by targeting AKT1 in NSCLC
11858 Int J Clin Exp Pathol 2015;8(9):11854-11862
Ectopic expression of miR-185 represses NSCLC cell migration and invasion
Next, we assessed the effects of miR-185 on the migratory and invasive ability of NSCLC cells. The wound healing assay showed that the migratory ability of miR-185-transfected H1299 and A549 cells was much lower than that of the control cells (Figure 3A). Consistent with the migration result, transwell invasion assay dem-onstrated that cell invasive ability was marked-ly inhibited in miR-185 mimic treated cells as compared with control cells (Figure 3B). These results suggest that miR-185 remarkably reduces the migratory and invasive ability of NSCLC cells.
miR-185 directly targets the AKT1 3’UTR
Bioinformatics analyses of AKT1 3’UTR revealed one putative binding site for miR-185. To study the potential interaction, AKT1 3’UTR and its corresponding mutant counterparts were cloned into pGL3 luciferase reporter vec-tors, and co-transfected into 293 T cells with miR-185 mimic or negative control (Figure 4A). As indicated in Figure 4B, miR-185 decreased the luciferase activity of the Luc-AKT1 3’UTR compared to the control vector, whereas the mutant counterpart blocked this decrease. To further investigate whether AKT1 is targeted by miR-185, miR-185 mimic or negative control was transfected into H1299 and A549 cells,
Figure 3. Ectopic expression of miR-185 repressed NSCLC cell migration and invasion. A. The scratch wound healing assay was performed in H1299 and A549 cells treated with miR-185 mimic or negative control oligonucleotides. B. Transwell matrigel invasion assay analysis of cell invasion. Each sample was analyzed in triplicate. *P < 0.05, **P < 0.01.

miR-185 acts as a tumor suppressor by targeting AKT1 in NSCLC
11859 Int J Clin Exp Pathol 2015;8(9):11854-11862
and AKT1 expression levels were analyzed by western blotting and qRT-PCR. The results indi-cated that AKT1 protein and mRNA expression levels were significantly lower in miR-185-trans-fected cells than those in the controls (Figure 4C and 4D). These data suggest that miR-185 negatively regulates AKT1 expression in NSCLC cells.
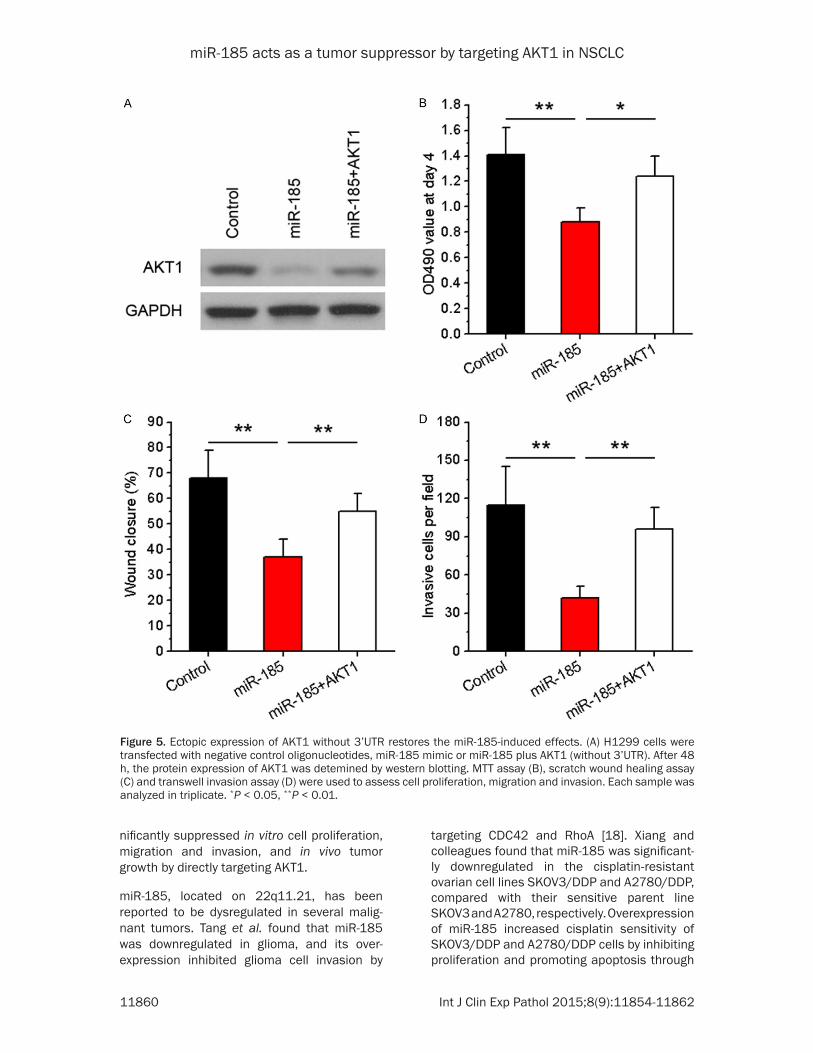
Ectopic expression of AKT1 without 3’UTR restores the miR-185-induced effects
Since miR-185 suppressed NSCLC cell prolifer-ation, migration and invasion, and inhibited AKT1 expression, we predicted that miR-185-mediated proliferative and metastatic inhibition was ascribable to AKT1 gene. Thus, we constructed AKT1 over-expressing vector (lacking its 3’UTR), and co-transfected into
H1299 cells along with miR-185 mimic. AKT1 protein expression was confirmed by western blotting (Figure 5A). In addition, we found that decreased cell viability, migration and invasion via miR-185 mimic were restored by the AKT1 over-expression (Figure 5B-D). Taken together, the cancer suppressor role of miR-185 in NSCLC is at least partially by inhibiting its target gene AKT1.
Discussion
In the current study, we studied the possible role of miR-185 in NSCLC. We found that miR-185 was commonly downregulated in NSCLC specimens and cell lines, compared with adja-cent non-cancerous tissues and the normal lung bronchus epithelial cell line, respectively. Furthermore, over-expression of miR-185 sig-
Figure 4. miR-185 directly targets the AKT1 3’UTR. A. The predictive miR-185 binding site of AKT1 3’UTR and the corresponding mutant binding site were shown. B. Relative activities of luciferase reporters encoding AKT1 3’UTR variants co-transfected with miR-185 mimic or negative control oligonucleotides in 293T cells. C. Relative AKT1 expression levels were examined by western blotting and qRT-PCR. Each sample was analyzed in triplicate. *P < 0.05, **P < 0.01.
miR-185 acts as a tumor suppressor by targeting AKT1 in NSCLC
11860 Int J Clin Exp Pathol 2015;8(9):11854-11862
nificantly suppressed in vitro cell proliferation, migration and invasion, and in vivo tumor growth by directly targeting AKT1.
miR-185, located on 22q11.21, has been reported to be dysregulated in several malig-nant tumors. Tang et al. found that miR-185 was downregulated in glioma, and its over- expression inhibited glioma cell invasion by
targeting CDC42 and RhoA [18]. Xiang and colleagues found that miR-185 was significant-ly downregulated in the cisplatin-resistant ovarian cell lines SKOV3/DDP and A2780/DDP, compared with their sensitive parent line SKOV3 and A2780, respectively. Overexpression of miR-185 increased cisplatin sensitivity of SKOV3/DDP and A2780/DDP cells by inhibiting proliferation and promoting apoptosis through
Figure 5. Ectopic expression of AKT1 without 3’UTR restores the miR-185-induced effects. (A) H1299 cells were transfected with negative control oligonucleotides, miR-185 mimic or miR-185 plus AKT1 (without 3’UTR). After 48 h, the protein expression of AKT1 was detemined by western blotting. MTT assay (B), scratch wound healing assay (C) and transwell invasion assay (D) were used to assess cell proliferation, migration and invasion. Each sample was analyzed in triplicate. *P < 0.05, **P < 0.01.

miR-185 acts as a tumor suppressor by targeting AKT1 in NSCLC
11861 Int J Clin Exp Pathol 2015;8(9):11854-11862
suppressing DNMT1 directly [24]. In triple-neg-ative breast cancer, miR-185 was found to be strongly downregulated in cancer tissues and cell lines and that its expression levels were associated with lymph node metastasis, clini-cal stage, overall survival, and relapse-free sur-vival. Ectopic expression of miR-185 inhibited TNBC cell proliferation in vitro and in vivo by directly targeting DNMT1 and E2F6 [25]. miR-185 was also reported to inhibit HCC cell prolif-eration and invasion in vitro and prevented tumor growth in SCID mice [20]. In NSCLC, previous studies have found that miR-185 sup-pressed cell growth and induce a G1 cell cycle arrest in H1299 cells [26], however, the func-tional mechanism of miR-185 in NSCLC cells are not fully understood. In the present study, we confirmed that miR-185 is downregulated in NSCLC tissues and cell lines, and functions as a tumor suppressor role in the development of cancer cells.
Aberrant PI3K/AKT pathway activation is found in a variety of cancers, including lung cancer, resulting in the development and progression of these malignancies [27, 28]. AKT1/protein kinase B α is the most extensively investigated member of the serine/threonine protein kinase subfamily and is usually referred as AKT [29]. Sun and colleagues showed that AKT1 kinase activity is elevated in prostate, breast cancers and ovarian carcinomas, and its constitutive activation is required for oncogenic transforma-tion in mouse NIH3T3 cells [30]. Linnerth-Petrik et al. found that Akt1 ablation significantly delays initiation of lung tumor growth in a mou- se model [27]. Thus, AKT1 may be a valuable therapeutic target for suppressing oncogene-sis. In this study, we showed that miR-185 sup-presses AKT1 expression by directly targeting its 3’UTR in NSCLC cells, and re-introduction of AKT1 remarkably attenuated the tumor sup-pressive effect of miR-185 on NSCLC cells.
In conclusion, we found that miR-185 functions as a tumor suppressor in NSCLC cells through repressing AKT1 expression. This is the first study to demonstrate that the miR-185/AKT1 axis regulates the proliferation, migration and invasion of NSCLC cells. The current findings facilitate our understanding of the molecular pathogenesis of NSCLC and support the poten-tial of miR-185 as an effective novel therapeu-tic target for treatment of the disease.
Disclosure of conflict of interest
None.
Address correspondence to: Dr. Jufeng Wang, De- partment of Internal Medicine, Affiliated Cancer Hospital of Zhengzhou University, Henan Cancer Hospital, Zhengzhou, Henan, China. E-mail: benben- [email protected]
References
[1] Jemal A, Siegel R, Xu J and Ward E. Cancer statistics, 2010. CA Cancer J Clin 2010; 60: 277-300.
[2] Anglim PP, Alonzo TA and Laird-Offringa IA. DNA methylation-based biomarkers for early detection of non-small cell lung cancer: an update. Mol Cancer 2008; 7: 81.
[3] Verdecchia A, Francisci S, Brenner H, Gatta G, Micheli A, Mangone L and Kunkler I. Recent cancer survival in Europe: a 2000-02 period analysis of EUROCARE-4 data. Lancet Oncol 2007; 8: 784-796.
[4] Lan D, Zhang X, He R, Tang R, Li P, He Q and Chen G. MiR-133a is downregulated in non-small cell lung cancer: a study of clinical significance. Eur J Med Res 2015; 20: 50.
[5] Pan B, Chen Y, Song H, Xu Y, Wang R and Chen L. Mir-24-3p downregulation contributes to VP16-DDP resistance in small-cell lung cancer by targeting ATG4A. Oncotarget 2015; 6: 317-331.
[6] Huang H, Jiang Y, Wang Y, Chen T, Yang L, He H, Lin Z, Liu T, Yang T, Kamp DW, Wu B and Liu G. miR-5100 promotes tumor growth in lung can-cer by targeting Rab6. Cancer Lett 2015; 362: 15-24.
[7] Wang LJ, He CC, Sui X, Cai MJ, Zhou CY, Ma JL, Wu L, Wang H, Han SX and Zhu Q. MiR-21 promotes intrahepatic cholangiocarcinoma proliferation and growth in vitro and in vivo by targeting PTPN14 and PTEN. Oncotarget 2015; 6: 5932-5946.
[8] Liang HL, Hu AP, Li SL, Xie JP, Ma QZ and Liu JY. MiR-454 Prompts Cell Proliferation of Human Colorectal Cancer Cells by Repressing CYLD Expression. Asian Pac J Cancer Prev 2015; 16: 2397-2402.
[9] Jiang J, Zhang Y, Guo Y, Yu C, Chen M, Li Z, Tian S and Sun C. MicroRNA-3127 promotes cell proliferation and tumorigenicity in hepato- cellular carcinoma by disrupting of PI3K/ AKT negative regulation. Oncotarget 2015; 6: 6359-6372.
[10] Yao Q, Cao Z, Tu C, Zhao Y, Liu H and Zhang S. MicroRNA-146a acts as a metastasis suppre- ssor in gastric cancer by targeting WASF2. Cancer Lett 2013; 335: 219-224.

miR-185 acts as a tumor suppressor by targeting AKT1 in NSCLC
11862 Int J Clin Exp Pathol 2015;8(9):11854-11862
[11] Chen P, Wang BL, Pan BS and Guo W. MiR-1297 regulates the growth, migration and inva-sion of colorectal cancer cells by targeting cyc-lo-oxygenase-2. Asian Pac J Cancer Prev 2014; 15: 9185-9190.
[12] Yang T, Thakur A, Chen T, Yang L, Lei G, Liang Y, Zhang S, Ren H and Chen M. MicroRNA-15a induces cell apoptosis and inhibits metastasis by targeting BCL2L2 in non-small cell lung can-cer. Tumour Biol 2015; 36: 4357-4365.
[13] Esquela-Kerscher A and Slack FJ. Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer 2006; 6: 259-269.
[14] Zhang XF, Li KK, Gao L, Li SZ, Chen K, Zhang JB, Wang D, Tu RF, Zhang JX, Tao KX, Wang G and Zhang XD. miR-191 promotes tumorigen-esis of human colorectal cancer through tar-geting C/EBPbeta. Oncotarget 2015; 6: 4144-4158.
[15] He D, Wang J, Zhang C, Shan B, Deng X, Li B, Zhou Y, Chen W, Hong J, Gao Y, Chen Z and Duan C. Down-regulation of miR-675-5p con-tributes to tumor progression and develop-ment by targeting pro-tumorigenic GPR55 in non-small cell lung cancer. Mol Cancer 2015; 14: 73.
[16] Li Q, Wang JX, He YQ, Feng C, Zhang XJ, Sheng JQ and Li PF. MicroRNA-185 regulates chemo-therapeutic sensitivity in gastric cancer by targeting apoptosis repressor with caspase recruitment domain. Cell Death Dis 2014; 5: e1197.
[17] Wang R, Tian S, Wang HB, Chu DP, Cao JL, Xia HF and Ma X. MiR-185 is involved in human breast carcinogenesis by targeting Vegfa. FEBS Lett 2014; 588: 4438-4447.
[18] Tang H, Wang Z, Liu X, Liu Q, Xu G, Li G and Wu M. LRRC4 inhibits glioma cell growth and inva-sion through a miR-185-dependent pathway. Curr Cancer Drug Targets 2012; 12: 1032-1042.
[19] Ma X, Shen D, Li H, Zhang Y, Lv X, Huang Q, Gao Y, Li X, Gu L, Xiu S, Bao X, Duan J and Zhang X. MicroRNA-185 inhibits cell prolifera-tion and induces cell apoptosis by targeting VEGFA directly in von Hippel-Lindau-inactivated clear cell renal cell carcinoma. Urol Oncol 2015; 33: 169. e1-11.
[20] Qadir XV, Han C, Lu D, Zhang J and Wu T. miR-185 inhibits hepatocellular carcinoma growth by targeting the DNMT1/PTEN/Akt pathway. Am J Pathol 2014; 184: 2355-2364.
[21] Zhang N, Wang X, Huo Q, Sun M, Cai C, Liu Z, Hu G and Yang Q. MicroRNA-30a suppresses breast tumor growth and metastasis by target-ing metadherin. Oncogene 2014; 33: 3119-3128.
[22] Zhu S, Deng S, Ma Q, Zhang T, Jia C, Zhuo D, Yang F, Wei J, Wang L, Dykxhoorn DM, Hare JM, Goldschmidt-Clermont PJ and Dong C. MicroRNA-10A* and MicroRNA-21 modulate endothelial progenitor cell senescence via suppressing high-mobility group A2. Circ Res 2013; 112: 152-164.
[23] Dong R, Liu X, Zhang Q, Jiang Z, Li Y, Wei Y, Yang Q, Liu J, Wei JJ, Shao C, Liu Z and Kong B. miR-145 inhibits tumor growth and metas- tasis by targeting metadherin in high-grade serous ovarian carcinoma. Oncotarget 2014; 5: 10816-10829.
[24] Xiang Y, Ma N, Wang D, Zhang Y, Zhou J, Wu G, Zhao R, Huang H, Wang X, Qiao Y, Li F, Han D, Wang L, Zhang G and Gao X. MiR-152 and miR-185 co-contribute to ovarian cancer cells cis-platin sensitivity by targeting DNMT1 directly: a novel epigenetic therapy independent of decitabine. Oncogene 2014; 33: 378-386.
[25] Tang H, Liu P, Yang L, Xie X, Ye F, Wu M, Liu X, Chen B and Zhang L. miR-185 suppresses tu-mor proliferation by directly targeting E2F6 and DNMT1 and indirectly upregulating BRCA1 in triple-negative breast cancer. Mol Cancer Ther 2014; 13: 3185-3197.
[26] Takahashi Y, Forrest AR, Maeno E, Hashimoto T, Daub CO and Yasuda J. MiR-107 and MiR-185 can induce cell cycle arrest in human non small cell lung cancer cell lines. PLoS One 2009; 4: e6677.
[27] Linnerth-Petrik NM, Santry LA, Petrik JJ and Wootton SK. Opposing functions of Akt iso-forms in lung tumor initiation and progression. PLoS One 2014; 9: e94595.
[28] Testa JR and Tsichlis PN. AKT signaling in nor-mal and malignant cells. Oncogene 2005; 24: 7391-7393.
[29] Shen G, Rong X, Zhao J, Yang X, Li H, Jiang H, Zhou Q, Ji T, Huang S, Zhang J and Jia H. MicroRNA-105 suppresses cell proliferation and inhibits PI3K/AKT signaling in human he-patocellular carcinoma. Carcinogenesis 2014; 35: 2748-2755.
[30] Sun M, Wang G, Paciga JE, Feldman RI, Yuan ZQ, Ma XL, Shelley SA, Jove R, Tsichlis PN, Nicosia SV and Cheng JQ. AKT1/PKBalpha ki-nase is frequently elevated in human cancers and its constitutive activation is required for oncogenic transformation in NIH3T3 cells. Am J Pathol 2001; 159: 431-437.









![Research Paper MiR-152-5p as a microRNA passenger strand ... · was first identified in mouse colon in 2002[9], more and more evidence suggests that miR-152 is a tumor suppressor,](https://static.fdocuments.in/doc/165x107/5bcd7b9109d3f2f14c8ddcd1/research-paper-mir-152-5p-as-a-microrna-passenger-strand-was-first-identified.jpg)








