
Opportunistic Immunodeficiency: LP-BM5 fileMiceinfected with LP-BM5murineleukemia viruses develop a...
8
Vol. 60, No. 10 INFECTION AND IMMUNITy, Oct. 1992, p. 4394 4401 0019-9567/92/104394-08$02.00/0 Copyright © 1992, American Society for Microbiology Opportunistic Infections and Retrovirus-Induced Immunodeficiency: Studies of Acute and Chronic Infections with Toxoplasma gondii in Mice Infected with LP-BM5 Murine Leukemia Viruses RICARDO T. GAZZINELLI,l* JANET W. HARTLEY,2 TORGNY N. FREDRICKSON,2'3 SISIR K. CHATTOPADHYAY,2 ALAN SHER,1 AND HERBERT C. MORSE III2 Immunology and Cell Biology Section, Laboratory of Parasitic Diseases,1 and Laboratory of Immunopathology, 2 National Institute ofAllergy and Infectious Diseases, and Registry of Excperimental Cancers, National Cancer Institute,3 Bethesda, Maryland 20892 Received 2 March 1992/Accepted 4 August 1992 Mice infected with LP-BM5 murine leukemia viruses develop a syndrome, termed mouse AIDS (MAIDS), characterized by increasingly severe immunodeficiency and progressive lymphoproliferation. Virus-infected mice were examined for the ability to resist acute infection and to control chronic infection with the protozoan Toxoplasma gondii, a major opportunistic pathogen of individuals infected with human immunodeficiency virus. Mice infected with the retroviruses for 2 or 4 weeks responded normally to challenge with the parasite, but mice inoculated with the protozoan 8 or 12 weeks after viral infection died with acute disease due to T. gondii. Increased sensitivity to acute infection was associated with a reduced ability to produce gamma interferon (IFN-y) and with established changes in CD4+ T-cell function. Mice latently infected with T. gondii and then inoculated with the retrovirus mixture were found to reactivate the parasite infection, with 30 to 40% of dually infected animals dying between 5 and 16 weeks after viral infection. Reactivation was associated with reduced proliferation and impaired production of IFN-y in response to stimulation with soluble T. gondii antigens or to concanavalin A. Continuing resistance to lethal reactivation in the remaining mice was shown to require CD8+ T cells and expression of IFN-y. In addition, it was found that chronic infection with T. gondii altered the course of MAIDS by inhibiting the progression of splenomegaly and immunodeficiency and reducing the expression of both the helper and etiologic defective viruses. These results support previous studies which indicate that infection with T. gondii is controlled by synergistic interactions between CD4+ and CD8+ T cells, the functions of which are progressively impaired during the course of MAIDS. Toxoplasma gondii is a protozoan parasite that is normally controlled by the host immune system and results in an asymptomatic, chronic infection maintained by dormant tissue cysts. In AIDS and other immunocompromised states, however, T. gondii produces an infection which can lead to severe morbidity, as well as mortality. In AIDS patients, clinical toxoplasmosis is thought to result primarily from reactivation of chronic infection, resulting in cyst rupture and release of the rapidly multiplying tachyzoite form of the parasite. However, it is possible that some disease is due to lowered resistance to newly acquired primary infection. Although AIDS-related toxoplasmosis is most often encoun- tered as encephalitis or mass lesions of the central nervous system, extraneural involvement has been demonstrated at autopsy in some patients (6, 19, 22, 27, 28). While the human immunodeficiency virus-dependent CD4+ lymphocyte deficiency is thought to underlie clinically severe toxoplasmosis in AIDS, the precise immune defect has not been characterized. Studies with murine experimen- tal models have implicated gamma interferon (IFN--y) pro- duction as the major host factor that prevents reactivation of chronic infection and determines acquired resistance to challenge infection (13, 35-37). While both CD4+ and CD8+ T cells are important for expression of protective immunity, their role in controlling reactivation is unresolved. * Corresponding author. Infection of C57BV6 (B6) mice with the mixture of C-type murine leukemia viruses (MuLV) termed LP-BM5 MuLV results in a syndrome, designated murine acquired immuno- deficiency syndrome (MAIDS), that has many of the immune abnormalities found in human immunodeficiency virus infec- tion (2, 5, 7, 8, 12, 14, 16, 20, 21, 24, 25, 29, 40). These include (i) polyclonal B-cell activation associated with hy- pergammaglobulinemia marked partly by high levels of im- munoglobulin E (14, 20, 21, 25, 29, 40); (ii) an early, selective defect in CD4+ Th-cell function manifested by impaired production of interleukin 2 and IFN--y, a deficient prolifera- tive response to concanavalin A (ConA) or to soluble antigen without concomitant reduction in CD4+ T-cell numbers, and increased production of interleukins 4 and 10 in response to ConA (7, 14, 24, 25); (iii) a defect in the generation of CD8+ T-cell activity late in the course of disease (24); and (iv) enhanced susceptibility to B-cell lymphomas and ectromelia virus and Cryptosporidium parvum infections (5, 12, 20). In the present study, we attempted to define the effect of immunodeficiency caused by LP-BM5 MuLV on the re- sponse to infection with T. gondii. Specifically, we deter- mined whether prior infection with the viruses alters the resistance of mice to challenge with the protozoan and whether LP-BM5 MuLV can induce reactivation of T. gondii in chronically parasitized animals. Our results indicate that progressively impaired CD4+ T-cell function in the course of MAIDS is associated with the inability of virus-infected mice to respond to acute toxoplasmosis. In contrast, reactivation 4394 on April 25, 2019 by guest http://iai.asm.org/ Downloaded from
Transcript of Opportunistic Immunodeficiency: LP-BM5 fileMiceinfected with LP-BM5murineleukemia viruses develop a...

Vol. 60, No. 10INFECTION AND IMMUNITy, Oct. 1992, p. 4394 44010019-9567/92/104394-08$02.00/0Copyright © 1992, American Society for Microbiology
Opportunistic Infections and Retrovirus-InducedImmunodeficiency: Studies of Acute and Chronic Infectionswith Toxoplasma gondii in Mice Infected with LP-BM5
Murine Leukemia VirusesRICARDO T. GAZZINELLI,l* JANET W. HARTLEY,2 TORGNY N. FREDRICKSON,2'3
SISIR K. CHATTOPADHYAY,2 ALAN SHER,1 AND HERBERT C. MORSE III2Immunology and Cell Biology Section, Laboratory of Parasitic Diseases,1 and Laboratory ofImmunopathology, 2 National Institute ofAllergy and Infectious Diseases, and Registry of
Excperimental Cancers, National Cancer Institute,3 Bethesda, Maryland 20892
Received 2 March 1992/Accepted 4 August 1992
Mice infected with LP-BM5 murine leukemia viruses develop a syndrome, termed mouse AIDS (MAIDS),characterized by increasingly severe immunodeficiency and progressive lymphoproliferation. Virus-infectedmice were examined for the ability to resist acute infection and to control chronic infection with the protozoanToxoplasma gondii, a major opportunistic pathogen of individuals infected with human immunodeficiencyvirus. Mice infected with the retroviruses for 2 or 4 weeks responded normally to challenge with the parasite,but mice inoculated with the protozoan 8 or 12 weeks after viral infection died with acute disease due to T.gondii. Increased sensitivity to acute infection was associated with a reduced ability to produce gammainterferon (IFN-y) and with established changes in CD4+ T-cell function. Mice latently infected with T. gondiiand then inoculated with the retrovirus mixture were found to reactivate the parasite infection, with 30 to 40%of dually infected animals dying between 5 and 16 weeks after viral infection. Reactivation was associated withreduced proliferation and impaired production of IFN-y in response to stimulation with soluble T. gondiiantigens or to concanavalin A. Continuing resistance to lethal reactivation in the remaining mice was shown torequire CD8+ T cells and expression of IFN-y. In addition, it was found that chronic infection with T. gondiialtered the course ofMAIDS by inhibiting the progression of splenomegaly and immunodeficiency and reducingthe expression of both the helper and etiologic defective viruses. These results support previous studies whichindicate that infection with T. gondii is controlled by synergistic interactions between CD4+ and CD8+ T cells,the functions of which are progressively impaired during the course of MAIDS.
Toxoplasma gondii is a protozoan parasite that is normallycontrolled by the host immune system and results in anasymptomatic, chronic infection maintained by dormanttissue cysts. In AIDS and other immunocompromised states,however, T. gondii produces an infection which can lead tosevere morbidity, as well as mortality. In AIDS patients,clinical toxoplasmosis is thought to result primarily fromreactivation of chronic infection, resulting in cyst ruptureand release of the rapidly multiplying tachyzoite form of theparasite. However, it is possible that some disease is due tolowered resistance to newly acquired primary infection.Although AIDS-related toxoplasmosis is most often encoun-tered as encephalitis or mass lesions of the central nervoussystem, extraneural involvement has been demonstrated atautopsy in some patients (6, 19, 22, 27, 28).While the human immunodeficiency virus-dependent
CD4+ lymphocyte deficiency is thought to underlie clinicallysevere toxoplasmosis in AIDS, the precise immune defecthas not been characterized. Studies with murine experimen-tal models have implicated gamma interferon (IFN--y) pro-duction as the major host factor that prevents reactivation ofchronic infection and determines acquired resistance tochallenge infection (13, 35-37). While both CD4+ and CD8+T cells are important for expression of protective immunity,their role in controlling reactivation is unresolved.
* Corresponding author.
Infection of C57BV6 (B6) mice with the mixture of C-typemurine leukemia viruses (MuLV) termed LP-BM5 MuLVresults in a syndrome, designated murine acquired immuno-deficiency syndrome (MAIDS), that has many of the immuneabnormalities found in human immunodeficiency virus infec-tion (2, 5, 7, 8, 12, 14, 16, 20, 21, 24, 25, 29, 40). Theseinclude (i) polyclonal B-cell activation associated with hy-pergammaglobulinemia marked partly by high levels of im-munoglobulin E (14, 20, 21, 25, 29, 40); (ii) an early, selectivedefect in CD4+ Th-cell function manifested by impairedproduction of interleukin 2 and IFN--y, a deficient prolifera-tive response to concanavalinA (ConA) or to soluble antigenwithout concomitant reduction in CD4+ T-cell numbers, andincreased production of interleukins 4 and 10 in response toConA (7, 14, 24, 25); (iii) a defect in the generation of CD8+T-cell activity late in the course of disease (24); and (iv)enhanced susceptibility to B-cell lymphomas and ectromeliavirus and Cryptosporidium parvum infections (5, 12, 20).
In the present study, we attempted to define the effect ofimmunodeficiency caused by LP-BM5 MuLV on the re-sponse to infection with T. gondii. Specifically, we deter-mined whether prior infection with the viruses alters theresistance of mice to challenge with the protozoan andwhether LP-BM5 MuLV can induce reactivation of T. gondiiin chronically parasitized animals. Our results indicate thatprogressively impaired CD4+ T-cell function in the course ofMAIDS is associated with the inability of virus-infected miceto respond to acute toxoplasmosis. In contrast, reactivation
4394
on April 25, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

IMPAIRED IMMUNITY TO TOXOPLASMOSIS IN MAIDS 4395
of chronic infection with the protozoan in virus-infectedmice appears to result primarily from CD8+ T-cell dysfunc-tion.
MATERIALS AND METHODS
Experimental animals. Female B6 and outbred Swiss Web-ster mice 4 to 5 weeks old were obtained from the Divisionof Cancer Treatment, National Cancer Institute (Frederick,Md.).
Viruses and,virus assays. The disease-causing mixture ofLP-BM5 MuLV includes replication-competent B-tropicecotropic and mink cell focus-inducing MuLVs (8, 16, 29)and an etiologic replication-defective virus (2, 8). Viruspools derived from chronically infected SC-1 cultures wereprepared and assayed for ecotropic and mink cell focus-inducing MuLV as previously described (11, 17, 32) and forthe ability to produce disease in B6 mice. Virus pools used inthis study contained 104.1 to 104-3 PFU of ecotropic MuLVper ml. Ecotropic virus recovery from mice was determinedby infectious-center assays of mitomycin C-treated spleen orlymph node cells (10, 11) or XC plaque tests using 10%(wt/vol) extracts of frozen tissues.The presence of integrated copies of the defective virus
genome in tissues of infected mice was evaluated by South-ern blot hybridization of high-molecular-weight DNA fromspleen or lymph nodes. The defective genome contains KpnIsites in the long terminal repeats (8), and copies integrated atvarious chromosomal sites were recruited into a unique4.4-kb fragment on digestion of infected B6 cellular DNAwith this enzyme. The presence or absence of this fragmentwas determined after separation of digested DNA in agarose,transfer to nitrocellulose, and hybridization with a 32p_labelled 100-bp probe homologous to the p12 region of thegag gene of the defective virus as previously described (9).
Parasites and antigen preparation. Three strains of T.gondii were used. The ME-49 strain was used to studyresistance to newly acquired infection, since it is not lethalduring acute infection. To study reactivation of chronicinfection, mice were infected with the C56 strain, whichcauses a mild chronic disease after treatment with sulfadi-azine. The highly virulent RH strain was used for antigenpreparation. Cysts of ME49 (kindly provided by F. Araujoand J. S. Remington, Stanford University) were obtainedfrom brains of B6 mice inoculated with cysts at least 1 monthearlier. Tachyzoites of the C56 strain were obtained from B6mice injected with 20 cysts intraperitoneally (i.p.) 1 weekearlier. For soluble tachyzoite antigen (STAg) preparation,tachyzoites of the RH strain were harvested from peritonealcavities of Swiss Webster mice injected with 5 x 106organisms 3 days earlier. The tachyzoites were sonicatedand centrifuged, and the supernatant was collected as pre-viously described (13).
Experimental infections. In one experimental protocol, B6mice were infected at 4 to 5 weeks of age by i.p. injectionwith 0.1 ml of LP-BM5 virus stock and then challenged atdifferent times postinfection by i.p. injection of 20 cysts of T.gondii ME-49. In the second protocol, mice were infected atthe same age by i.p. injection with 105 C56 strain tachy-zoites, treated for 14 days with sulfadiazine (400 mg/liter) inthe drinking water, and then infected with LP-BM5 MuLV 6weeks later. Mice not infected with T. gondii but treatedwith sulfadiazine did not differ from untreated mice in theirresponses to infection with LP-BM5 MuLV, as evaluated byfluorescence-activated cell sorter (FACS) analyses of spleen
cells, histopathologic studies of spleen and lymph nodes, andvirus recovery from spleen (data not shown).
Treatment of mice with antibodies. Mice were depleted ofCD8+ T cells by i.p. injection with 0.5 mg of a rat monoclo-nal antibody (MAb) (2.43) against mouse CD8 (33) for 3successive days and with additional doses (0.5 mg) everythird day throughout the experiment. For IFN-y neutraliza-tion, 6 mg of a rat anti-mouse IFN--y MAb (XMG 1.2) (26)was injected weekly. Control mice were inoculated with thesame dose of a rat MAb (GL113) to 3-galactosidase. AllMAbs were purified from ascites by ammonium sulfateprecipitation. Depletion of CD8+ cells was monitored byFACS analysis by using a fluorescein isothiocyanate (FITC)-labeled anti-CD8 MAb.Measurement of anti-T. gondii antibody responses. Serum
antibodies to STAg were measured in polystyrene microwellplates (Dynatech Laboratories, Inc., Alexandria, Va.) pre-coated with STAg (20 ,ug/ml) by using mouse serum dilutionsof 1:20 to 1:20,480 in phosphate-buffered saline-Tween 20.Bound immunoglobulin G antibodies were detected follow-ing reaction with peroxidase-conjugated goat anti-mouseimmunoglobulin G (Kirkegaard & Perry Laboratories, Inc.,Gaithersburg, Md.) at a dilution of 1:1,000. Color develop-ment was carried out with ABTS substrate (Kirkegaard &Perry Laboratories).Measurement of cellular immune responses. Spleen or
lymph node single-cell suspensions from infected and con-trol mice were prepared as previously described (13) andused for proliferation and IFN--y assays. Lymphocytes werecultured in Dulbecco modified Eagle medium that contained10% fetal calf serum, 2mM glutamine, 100 U of penicillin perml, 100 mg of streptomycin per ml, 25 mM HEPES (N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid), and 50mM 2-mercaptoethanol in the presence or absence of eitherSTAg (10 p,g/ml) or ConA (5 pg/ml). The proliferativeresponses were assayed in triplicate cultures of 200 ml thatcontained 4 x 105 cells in 96-well flat-bottom tissue cultureplates maintained at 37°C in 5% CO2 in air for 4 days. Wellswere pulsed with 1 pICi of [3H]thymidine for 18 h andprocessed for scintillation counting. IFN--y was measured inculture supernatants of lymph node or spleen cells (107 cellsper ml per well) stimulated with either STAg (10 p,g/ml) orConA (5 p,g/ml) by using a two-site enzyme-linked immuno-sorbent assay employing rabbit polyvalent anti-IFN-y serumand a rat IFN--y-specific MAb (HB170) as previously de-scribed (13).
Statistics. Statistical evaluation of differences betweenmeans of experimental groups was done by analysis ofvariance and the Scheffe F test (STATVIEW 512+, aninteractive statistics and graphics package from AbacurConcepts, Inc.).
RESULTS
Effect of MAIDS on resistance to acute T. gondii infection.In normal B6 mice, infection with T. gondii ME-49 results inaccumulation of numerous cysts in the brain, followed bydeath beginning at 42 days postinfection (Fig. 1A). Todetermine whether infection with LP-BM5 MuLV alters theability of mice to control disease development, animals werechallenged with ME-49 at various times after inoculationwith the retrovirus mixture. As shown in Fig. 1A, micechallenged with ME-49 at 2 or 4 weeks post-virus infectionwere as resistant as normal B6 mice. The deaths thatoccurred in T. gondii-challenged normal mice, as well as inmice infected with virus for 2 or 4 weeks prior to ME-49
VOL. 60, 1992
on April 25, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

4396 GAZZINELLI ET AL.
100 -
80 -
60 -
40 -
20 -
0 10 20 30 40 50
Days post-infection with ME-49
40
E 30
20
10
0
0 4 8 12
Weeks after LP-BM5 MuLV infectionFIG. 1. Resistance of LP-BM5 MuLV-infected mice to challenge with T. gondii correlates with IFN-y response. (A) Normal mice ( )
or mice infected with LP-BM5 MuLV for 2 (0), 4 (@), 8 ([), or 12 (U) weeks were challenged i.p., in groups of at least five animals each,with 20 cysts of strain ME-49. Survival was monitored daily, and cumulative mortality was calculated. (B) IFN-y response of spleen cells frommice during the course of LP-BM5 MuLV infection. Supernatants of spleen cell cultures were collected 48 h after stimulation with ConA (5,ug/ml). The data shown are means and standard errors of IFN--y determinations on three individual mice tested at the indicated times aftervirus infection. The limit of detection of the IFN--y enzyme-linked immunosorbent assay was 50 pg. The asterisks indicate the stages of virusinfection at which mice were challenged with ME-49 in panel A. Responses at 4 through 12 weeks post-virus infection were significantly lowerthan those observed at 0 or 2 weeks (P < 0.05).
inoculation, were associated with encephalitis. The lesionsincluded meningeal infiltration with lymphoid and plasmacells which also formed moderately large cuffs around cere-bral vessels (data not shown). Moderate-size glial noduleswere also scattered throughout the brain. High cyst counts(> 1,000 per brain) accompanied the encephalitis.
In contrast, when challenged at 8 or 12 weeks post-virusinfection, the mice were highly susceptible to ME-49, dyingwithin 15 days of inoculation. Deaths of mice challengedwith ME-49 at 8 or 12 weeks after viral infection were due tolesions of an acute form of toxoplasmosis associated withdiffuse interstitial pneumonitis, myocarditis, hepatic necro-sis, thrombosis, and peritonitis, with occasional hemor-rhagic and necrotic areas in the brain (data not shown).Tachyzoites were evident in some of the ascites, lung, andbrain sections examined, and their presence was confirmedby immunocytochemistry using rabbit antiserum to toxo-plasma antigens (data not shown). Death of these mice wasnot due to MAIDS, as all of the mice infected with the virusalone survived through week 16 postinoculation (Fig. 1 anddata not shown).
Since IFN-y has been implicated as the major cytokinethat determines susceptibility to toxoplasma infection, pro-duction of this cytokine by spleen cells stimulated withConA was monitored at various times after virus inocula-tion. Spleen cells from mice infected for 2 weeks producedIFN--y at levels equivalent to those obtained from uninfectedmice, but the response declined progressively such that at 12weeks of infection the levels were less than 30% of thoseexpressed by normal spleen cells (Fig. 1B). These datademonstrate that enhanced susceptibility to acute infectionwith T. gondii following infection with LP-BM5 MuLV wasassociated with impaired production of IFN-y in response toConA.
Effect of LP-BM5 MuLV infection on mice chronicallyinfected with T. gondii. To determine whether the immuno-deficiency that occurs with MAIDS leads to reactivation ofchronic toxoplasmosis, mice were infected with the lethalstrain T. gondii C56, treated with sulfadiazine for 2 weeks,
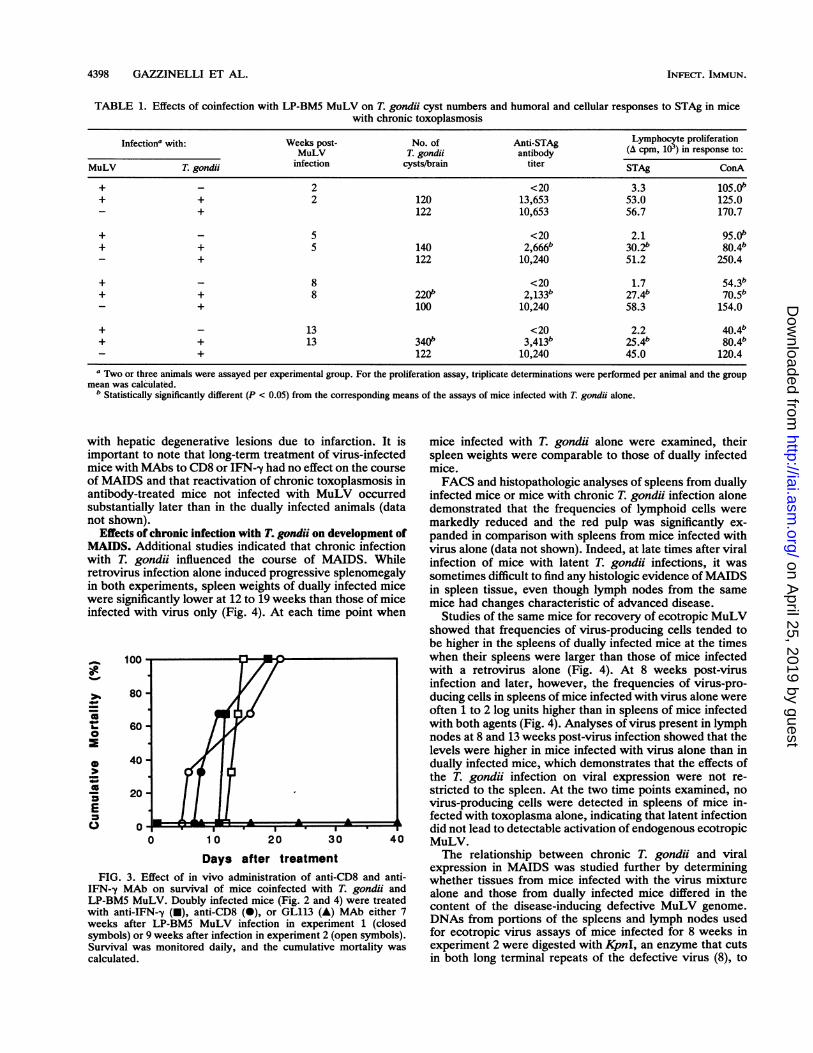
and infected with LP-BM5 MuLV 6 weeks later. No deathswere observed among mice infected with MuLV or T. gondiialone. In two separate studies of mice infected with bothagents, 30 to 40% mortality was observed by 80 daysfollowing viral infection (Fig. 2). Analysis of homogenatesfrom brains of mice killed at various times after viralinfection gave evidence for partial reactivation of T. gondii(Table 1). In mice infected with T. gondii alone, approxi-mately 100 cysts were detected throughout the course ofdisease. In contrast, the numbers of cysts in mice infectedwith both the parasite and the retrovirus were increasedslightly at 5 weeks post-virus inoculation and were signifi-cantly higher at both 8 and 13 weeks (P < 0.05).
Histopathologic studies of the same brains showed thatMAIDS promoted the pathogenic potential of T. gondii. Thispathology was characterized by classic encephalitic lesions,which were significantly more severe in dually infected miceat all times after infection.Humoral and cellular responses to T. gondii in dually
infected mice. To evaluate the mechanisms that might con-tribute to reactivation of latent toxoplasmosis during thecourse of MAIDS, sera from control and singly and duallyinfected mice were tested for specific antibodies againsttoxoplasma antigens, and spleen cells from the same micewere evaluated for the ability to proliferate in response to invitro stimulation with either STAg or ConA (Table 1). Thesestudies showed that the humoral response to T. gondii wassimilar in both groups of mice at the time of MuLV infection(data not shown) and 2 weeks later but was significantlydecreased at 5 weeks post-virus infection and thereafter(Table 1). Studies of cellular immunity to T. gondii antigensshowed that only mice with chronic toxoplasmosis devel-oped vigorous cellular proliferative responses to STAg andthat these responses persisted throughout the period ofobservation. In contrast, the responses of mice with bothinfections were significantly suppressed by 5 weeks afterviral infection and thereafter. Consistent with earlier studies(24, 25), infection with LP-BM5 MuLV alone resulted inmarked reductions in proliferative responses to ConA. It
010-
S
o-
Eco._o
INFECT. IMMUN.
on April 25, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
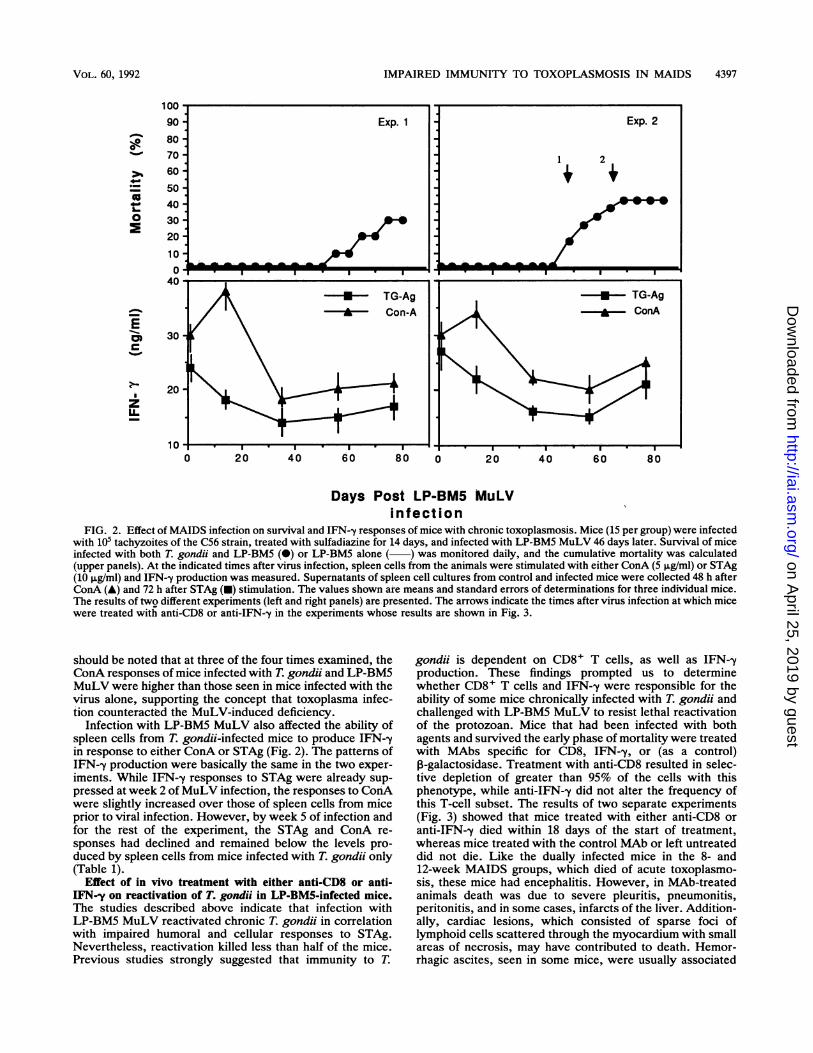
IMPAIRED IMMUNITY TO TOXOPLASMOSIS IN MAIDS 4397
10090
"
800-
70
>, 60
=_ 50- 40
O 30
20
100
40
c 30
20zU-
10
Days Post LP-BM5 MuLVinfection
FIG. 2. Effect of MAIDS infection on survival and IFN--y responses of mice with chronic toxoplasmosis. Mice (15 per group) were infectedwith 105 tachyzoites of the C56 strain, treated with sulfadiazine for 14 days, and infected with LP-BM5 MuLV 46 days later. Survival of miceinfected with both T. gondii and LP-BM5 (@) or LP-BM5 alone ( ) was monitored daily, and the cumulative mortality was calculated(upper panels). At the indicated times after virus infection, spleen cells from the animals were stimulated with either ConA (5 ,ug/ml) or STAg(10 ,ug/ml) and IFN--y production was measured. Supernatants of spleen cell cultures from control and infected mice were collected 48 h afterConA (A) and 72 h after STAg (-) stimulation. The values shown are means and standard errors of determinations for three individual mice.The results of twQ different experiments (left and right panels) are presented. The arrows indicate the times after virus infection at which micewere treated with anti-CD8 or anti-IFN-y in the experiments whose results are shown in Fig. 3.
should be noted that at three of the four times examined, theConA responses of mice infected with T. gondii and LP-BM5MuLV were higher than those seen in mice infected with thevirus alone, supporting the concept that toxoplasma infec-tion counteracted the MuLV-induced deficiency.
Infection with LP-BM5 MuLV also affected the ability ofspleen cells from T. gondii-infected mice to produce IFN--yin response to either ConA or STAg (Fig. 2). The patterns ofIFN--y production were basically the same in the two exper-iments. While IFN--y responses to STAg were already sup-pressed at week 2 of MuLV infection, the responses to ConAwere slightly increased over those of spleen cells from miceprior to viral infection. However, by week 5 of infection andfor the rest of the experiment, the STAg and ConA re-sponses had declined and remained below the levels pro-duced by spleen cells from mice infected with T. gondii only(Table 1).
Effect of in vivo treatment with either anti-CD8 or anti-IFN-y on reactivation of T. gondii in LP-BM5-infected mice.The studies described above indicate that infection withLP-BM5 MuLV reactivated chronic T. gondii in correlationwith impaired humoral and cellular responses to STAg.Nevertheless, reactivation killed less than half of the mice.Previous studies strongly suggested that immunity to T.
gondii is dependent on CD8+ T cells, as well as IFN--yproduction. These findings prompted us to determinewhether CD8+ T cells and IFN--y were responsible for theability of some mice chronically infected with T. gondii andchallenged with LP-BM5 MuLV to resist lethal reactivationof the protozoan. Mice that had been infected with bothagents and survived the early phase of mortality were treatedwith MAbs specific for CD8, IFN-y, or (as a control),-galactosidase. Treatment with anti-CD8 resulted in selec-tive depletion of greater than 95% of the cells with thisphenotype, while anti-IFN-y did not alter the frequency ofthis T-cell subset. The results of two separate experiments(Fig. 3) showed that mice treated with either anti-CD8 oranti-IFN--y died within 18 days of the start of treatment,whereas mice treated with the control MAb or left untreateddid not die. Like the dually infected mice in the 8- and12-week MAIDS groups, which died of acute toxoplasmo-sis, these mice had encephalitis. However, in MAb-treatedanimals death was due to severe pleuritis, pneumonitis,peritonitis, and in some cases, infarcts of the liver. Addition-ally, cardiac lesions, which consisted of sparse foci oflymphoid cells scattered through the myocardium with smallareas of necrosis, may have contributed to death. Hemor-rhagic ascites, seen in some mice, were usually associated
Exp. 1 Exp. 2
M 1. 1 2
i-
;^t - - - ,I ,MEI=I I I,
VOL. 60, 1992
on April 25, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
4398 GAZZINELLI ET AL.
TABLE 1. Effects of coinfection with LP-BM5 MuLV on T. gondii cyst numbers and humoral and cellular responses to STAg in micewith chronic toxoplasmosis
Infection' with: Weeks post- No. of Anti-STAg Lymphocyte proliferationMuLV T. gondii antibody (A cpm, 103) in response to:
MuLV T. gondii infection cysts/brain titer STAg ConA
+ - 2 <20 3.3 105.0"+ + 2 120 13,653 53.0 125.0
+ 122 10,653 56.7 170.7
+ - 5 <20 2.1 95.0"+ + 5 140 2,666b 30.2b 80.4"
+ 122 10,240 51.2 250.4
+ - 8 <20 1.7 54.3"+ + 8 220b 2,133b 27.4" 70.5"
+ 100 10,240 58.3 154.0
+ - 13 <20 2.2 40.4+ + 13 340b 3,413b 25.4" 80.4"
+ 122 10,240 45.0 120.4a Two or three animals were assayed per experimental group. For the proliferation assay, triplicate determinations were performed per animal and the group
mean was calculated.bStatistically significantly different (P < 0.05) from the corresponding means of the assays of mice infected with T. gondii alone.
with hepalimportant Imice with Iof MAIDSantibody-tisubstantialnot shown
Effects oiMAIDS. Awith T. gretrovirus iin both exjwere signifinfected w
-
0
0-
0
2
0
E
FIG. 3. 1IFN-y MAtLP-BM5 Miwith anti-IFweeks aftersymbols) orSurvival wacalculated.
tic degenerative lesions due to infarction. It is mice infected with T. gondii alone were examined, theirto note that long-term treatment of virus-infected spleen weights were comparable to those of dually infectedMAbs to CD8 or IFN--y had no effect on the course mice.and that reactivation of chronic toxoplasmosis in FACS and histopathologic analyses of spleens from duallyreated mice not infected with MuLV occurred infected mice or mice with chronic T. gondii infection alonelly later than in the dually infected animals (data demonstrated that the frequencies of lymphoid cells were
markedly reduced and the red pulp was significantly ex-f chronic infection with T. gondii on development of panded in comparison with spleens from mice infected withLdditional studies indicated that chronic infection virus alone (data not shown). Indeed, at late times after viralondii influenced the course of MAIDS. While infection of mice with latent T. gondii infections, it wasinfection alone induced progressive splenomegaly sometimes difficult to find any histologic evidence of MAIDSperiments, spleen weights of dually infected mice in spleen tissue, even though lymph nodes from the sameScantly lower at 12 to 19 weeks than those of mice mice had changes characteristic of advanced disease.ith virus only (Fig. 4). At each time point when Studies of the same mice for recovery of ecotropic MuLV
showed that frequencies of virus-producing cells tended tobe higher in the spleens of dually infected mice at the timeswhen their spleens were larger than those of mice infectedwith a retrovirus alone (Fig. 4). At 8 weeks post-virusinfection and later, however, the frequencies of virus-pro-ducing cells in spleens of mice infected with virus alone wereoften 1 to 2 log units higher than in spleens of mice infectedwith both agents (Fig. 4). Analyses of virus present in lymphnodes at 8 and 13 weeks post-virus infection showed that thelevels were higher in mice infected with virus alone than indually infected mice, which demonstrates that the effects ofthe T. gondii infection on viral expression were not re-stricted to the spleen. At the two time points examined, novirus-producing cells were detected in spleens of mice in-fected with toxoplasma alone, indicating that latent infectiondid not lead to detectable activation of endogenous ecotropic
0 10 20 30 40 MuLV.Days after treatment The relationship between chronic T. gondii and viralanti-tnexpression in MAIDS was studied further by determiningEffect of in vivo administration of anti-CD8 and anti- whether tissues from mice infected with the virus mixture
b on survival of mice coinfected with T. gondii and alone and those from dually infected mice differed in theuLV. Doubly infected mice (Fig. 2 and 4) were treated4N-,y (U), anti-CD8 (0), or GL113 (A) MAb either 7 content of the disease-iducig defective MuLV genome.r LP-BM5 MuLV infection in experiment 1 (closed DNAs from portions of the spleens and lymph nodes used9 weeks after infection in experiment 2 (open symbols). for ecotropic virus assays of mice infected for 8 weeks inas monitored daily, and the cumulative mortality was experiment 2 were digested with KjpnI, an enzyme that cuts
in both long terminal repeats of the defective virus (8), to
INFECT. IMMUN.
on April 25, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

IMPAIRED IMMUNITY TO TOXOPLASMOSIS IN MAIDS 4399
1000
800E* 600S.C0 4200c0
0Q& 200
.4
= r-O 3
0 >1. 0
0
LI.I.0
0 4 8 12 16 0 4 8 1 2
weeks post virus infectionFIG. 4. Changes in spleen weight (top panels) and expression of infectious ecotropic MuLV (lower panels) in relation to infections with
LP-BM5 MuLV and T. gondu C56. Spleen weights of mice infected with LP-BM5 MuLV alone, LP-BM5 MuLV and T. gondii, or T. gondiialone were determined at the indicated times after MuLV infection in two separate experiments. At some time points, spleens were assayedfor ecotropic MuLV by infectious center tests. Symbols: *, mice infected with LP-BM5 MuLV only; 0, mice infected with virus and T.gondii; A, mice infected with T. gondii only. In experiment 2, pooled peripheral lymph nodes of the same mice examined in infectious centertests of spleen tissue at 8 weeks were used to produce cell extracts that were assayed for infectious ecotropic MuLV or analyzed at 13 weeksby infectious center assays. Symbols: A, mice infected with virus only; El, mice infected with virus and T. gondii. Values indicate PFU (log1o)per milliliter of 10% (wt/vol) extracts at 8 weeks and PFU (log1o) per 107 cells at 13 weeks.
recruit integrated proviral sequences (Fig. 5). A diagnostic4.4-kb fragment that hybridized with a probe for p12 of thedefective virus was detected in DNAs from the spleens andnodes of both mice infected with virus only, indicating thatthe tissues contained multiple copies of the defective ge-nome. In contrast, this band was detected at much reducedintensity in two of the three spleen DNAs from doublyinfected mice and not at all in the third. DNAs from lymphnodes from the same set of mice revealed a 4.4-kb hybrid-izing band of comparable intensity in all samples, except forthat from the third doubly infected animal.
DISCUSSIONToxoplasmosis is a major problem in immunocompro-
mised patients, including those with AIDS. In our studieswith a mouse model of retrovirus-induced immunodefi-ciency, we found that prior infection with LP-BM5 MuLVresulted in enhanced susceptibility to acute infection with T.gondii and that viral infection in mice with latent toxoplas-mosis led to reactivation of the parasite-induced disease.Importantly, chronic infection with T. gondii had prominenteffects on the course of MAIDS, significantly reducing theextent of lymphoproliferation and viral expression associ-ated with this disorder.
Previous studies on the mechanisms that contribute to
A.
1 2 3 4 5 6 7kb
B.
4IISt * : .*
.... ... .. ... .
FIG. 5. Southern blot hybridization analyses of LP-BM5 defectivevirus genomes in DNAs from spleen (A) and lymph node (B) cells ofinfected mice. DNA prepared from the spleen of a normal B6 mouse(lane 1), a B6 cell line chronically infected with LP-BM5 MuLV (lane2), individual mice infected for 8 weeks with virus alone (lanes 3 and4), or mice chronically infected with T. gondii and infected with virusfor 8 weeks (lanes 5 to 7) was digested with KpnI and hybridized witha probe for the defective virus. Size markers include the 4.4-kbfragment containing the recruited BM5d sequences.
TrExperiment 1 Experiment 2
O EE °MENb 0 *AA
00 5 o 0
0 00 .- 00 000 0 0 0
0 A O N AO000 A
4
VOL. 60, 1992
I
4.4---)-0 99 tug
.. A,.: 1."?f.
'. r low. --.! :.*,' vw
2.2- 10
on April 25, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

4400 GAZZINELLI ET AL.
resistance to infection with T. gondii following immunizationshowed that both CD4+ and CD8+ T cells are required forinduction of resistance (13, 37) and that resistance, onceestablished, is in large part dependent on CD8+ T cells aloneand IFN--y production (3, 13, 35, 37). Similar studies of micechronically infected with T. gondii suggest a major role ofCD8+ lymphocyte, as well as IFN--y, in resistance to thisparasite (15, 36). In contrast, although CD4+ lymphocytesare also required for induction of natural resistance to T.gondii (1), their role in controlling reactivation of chronicinfection is more controversial (15, 18, 39). These dataprovide a foundation for an understanding of the interactionsof T. gondii with the immune systems of mice infected withLP-BM5 MuLV.
Earlier studies of mice inoculated with these virusesshowed early and progressive loss of CD4+ T-cell effectorfunctions, including help for responses of normal B cells toantigen (25), help for CD8+ cytotoxic T lymphocyte re-sponses to modified self antigens (24), help for CD8+ Teffectors for rejection of skin grafts across minor histocom-patibility antigen barriers (31), and proliferative responses tosoluble antigens (7). These defects were often evident within2 to 3 weeks of infection and were profound by 8 to 12weeks. As would be expected from the requirement forCD4+ T cells to induce CD8-mediated resistance to T.gondii, there was a striking correlation between the timecourse of virus-induced CD4+ T-cell dysfunction and theability of mice to resist acute challenge with the protozoan atvarious times after viral infection. Mice infected with LP-BM5 MuLV were as resistant as normal mice when chal-lenged with toxoplasma at 2 weeks postinfection, showedsome acceleration in mortality when challenged at 4 weeks,and died quickly when challenged at 8 or 12 weeks (Fig. 1A).Given the requirement for IFN-y in controlling immunity toT. gondii (35), it is not surprising that the decline in resis-tance to this parasite was associated with decreased abilityof cells from virus-infected mice to produce this cytokinefollowing stimulation with ConA (Fig. 1B). However, whilethe IFN--y response was largely suppressed by 4 weeks ofinfection, increased susceptibility to a new T. gondii infec-tion was observed only after 8 weeks of infection. It isnoteworthy that the activity and number of natural killercells are increased during early stages of MAIDS (22a),which may explain the resistance of the mice during weeks 2and 4 after LP-BM5 MuLV infection (Fig. 1).
Unexpectedly, however, mortality in the doubly infectedmice was limited to 30 to 40% of the animals and some micesurvived for as long as 19 weeks after viral infection (Fig. 4).Continuing control of the chronic infection in these mice wasconclusively shown to be mediated by CD8+ T cells andIFN-y, as their elimination resulted in development of fulmi-nant disease (Fig. 4). The predominant pathologic lesionsoutside the central nervous system in dually infected micewith lethal reactivation of chronic T. gonduii were pneumoni-tis, areas of lymphoid infiltration and necrosis in the liver,and, more variably, lymphoid infiltrates and areas of necrosisin the heart. It is noteworthy that pneumonia was also theprimary abnormality observed in mice with reactivation ofchronic T. gondii infection induced by infection with cyto-megalovirus (30).The failure to observe lethal reactivation of the chronic
protozoal infection later than 12 weeks post-virus inoculationsuggests that the decline in CD8+ T-cell function did notprogress as in mice infected with virus alone. Indeed, thefindings that spleen cells from mice infected with both agentsproliferated in response to STAg or ConA at late times after
viral infection (Table 1 and Fig. 3) indicate that CD4+ T-cellfunction was also somewhat preserved relative to that in miceinfected with virus alone. These responses cannot be ex-plained by residual CD8 activity, as purified CD8+ cells fromeither normal or virus-infected mice contribute minimally toproliferative responses to soluble antigen or ConA (7).A basis for understanding the partial preservation of both
CD4+ and CD8+ T-cell functions in mice infected with bothagents in comparison with mice infected with virus alone wasprovided initially by the finding that lymphoproliferationdeveloping as a consequence of viral infection was inhibitedin mice with chronic T. gondii infections. Reductions in thesizes of spleens and lymph nodes were associated withreduced frequencies of cells in these tissues producing infec-tious ecotropic MuLV (Fig. 4). More importantly, the dis-ease-inducing defective virus genome was present at muchreduced levels in DNAs from spleens of each of the threemice with chronic T. gondii infected with MuLV for 8 weeks(Fig. 5). The finding that the genome could be detected readilyin DNAs prepared from lymph nodes of two of these threemice may explain the differences between spleen tissue andlymph nodes in their exhibition of histologic and FACSchanges characteristics of MAIDS. The mechanism by whichchronic infection with T. gondii effects these changes inMuLV expression is not understood but may relate to alter-ations in the cellular composition of the spleen and lymphnodes that develop as a consequence of the infection. It is alsopossible that the Thl type of response to chronic infectionwith T. gondii results in down regulation of the Th2-likeresponse to viral infection, which may be central to thedevelopment of lymphoproliferation and immunodeficiency inMAIDS (14). Conversely, induction of a Th2 cytokine profileby infection with LP-BM5 MuLV may result in down regula-tion of the Thl and CD8 responses to chronic T. gondiiessential for maintenance of the infection in a latent state.
In summary, the data presented in this study reinforce theconcept that CD8+ T lymphocytes play a central role incontrolling infection with T. gondii (3, 13, 35, 37). Assuggested elsewhere (38), efforts to establish and maintainthe responses of CD8+ T cells to various agents may be ofgreat importance to the prevention of acute viral infections(4, 23, 34), as well as the control of reactivation of chronicopportunistic infections, such as T. gondii infection, inretrovirus-induced immunodeficiency. Further analysis ofthe MAIDS-toxoplasma murine model described here shouldbe useful for studying the efficacy of vaccination, chemo-therapeutic agents, or immune modulators in controlling thecourse of this lethal infection in a setting of immunodefi-ciency with many similarities to AIDS. In addition, it mayprovide a system for developing an understanding of themechanism by which infectious agents influence each otherin the natural course of disease.
ACKNOWLEDGMENTS
We are grateful to Sara Hieny, Nancy Wolford, Eva Rudikoff, andFran Hausman for expert technical assistance and thank AllenCheever and Yuhui Xu for help and advice with histopathology andStephanie James for critical reading of the manuscript.
REFERENCES1. Araujo, F. G. 1991. Depletion of L3T4+ (CD4+) T lymphocytes
prevents development of resistance to Toxoplasma gondii inmice. Infect. Immun. 59:1614-1619.
2. Aziz, D. C., Z. Hanna, and P. Jolicoeur. 1989. Severe immuno-deficiency disease induced by a defective murine leukemiavirus. Nature (London) 338:505-508.
INFEcr. IMMUN.
on April 25, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

IMPAIRED IMMUNITY TO TOXOPLASMOSIS IN MAIDS 4401
3. Brown, C. R., and R Mcleod. 1990. Class I MHC genes andCD8+ T cells determine cyst number in T. gondii infection. J.Immunol. 145:3438-3441.
4. Bulier, R. M. L., K. L. Holmes, A. Hugin, T. N. Fredrickson,and H. C. Morse HI. 1987. Induction of cytotoxic T cellresponses in vivo in the absence of CD4 helper cells. Nature(London) 328:77-79.
5. Bulier, R. M. L., R. A. Yetter, T. N. Fredrickson, and H. C.Morsem. 1987. Abrogation of resistance to severe mousepox inC57BL/6 mice infected with LP-BM5 murine leukemia viruses.J. Virol. 61:383-387.
6. Catteral, J. R., J. M. Hofflin, and J. S. Remington. 1986.Pulmonary toxoplasmosis. Annu. Rev. Respir. Dis. 133:704-705.
7. Cerny, A., A. W. Hugin, K. L Holmes, and H. C. Morse m.1990. CD4' T cells in murine acquired immunodeficiency syn-drome: evidence for an intrinsic defect in the proliferativeresponse to soluble antigen. Eur. J. Immunol. 20:1577-1581.
8. Chattopadhyay, S. K., H. C. Morse mII, M. Makdno, S. K.Ruscetfl, and J. W. Hartley. 1989. A defective virus is associatedwith induction of a murine retrovirus-induced immunodefi-ciency syndrome, MAIDS. Proc. Natl. Acad. Sci. USA 86:3862-3866.
9. Chattopadhyay, S. K., D. N. SenGupta, T. N. Fredrickson, H. C.Morse HI, and J. W. Hartley. 1991. Characteristics and contri-butions of defective, ecotropic, and mink cell focus-inducingviruses involved in a retrovirus-induced immunodeficiency syn-drome of mice. J. Virol. 65:4232-4241.
10. Cloyd, M. W., J. W. Hartley, and W. P. Rowe. 1980. Lympho-magenicity of recombinant mink cell focus-inducing murineleukemia viruses. J. Exp. Med. 151:542-552.
11. Cloyd, M. W., J. W. Hartley, and W. P. Rowe. 1981. Geneticstudy of lymphoma induction by AKR mink cell focus-inducingmurine leukemia viruses in AKR x NFS crosses. J. Exp. Med.154:450458.
12. Darban, H., J. Enriquez, C. R. Sterling, M. C. Lopez, G. Chen,M. Abbaszadejan, and R. R. Watgan. 1991. Cryptosporidiosisfacilitated by murine retroviral infection with LP-BM5. J. In-fect. Dis. 164:741-745.
13. Gazzinelli, R. T., F. Hakim, S. Hieny, G. Shearer, and A. Sher.1991. Synergistic role of CD4+ and CD8+ T lymphocytes inIFN-y production and protective immunity induced by an atten-uated Tawoplasma gondii vaccine. J. Immunol. 146:286-292.
14. Gazzinelli, R. T., M. Makino, S. K. Chattopadhyay, C. M.Snapper, A. Sher, A. W. Hugin, and H. C. Morse m. 1992.CD4+ subset regulation in viral infection: preferential activationof Th2 cells during progression of retrovirus-induced immuno-deficiency in mice. J. Immunol. 148:182-188.
15. Gazzinelli, R. T., Y. Xu, S. Hieny, A. Cheever, and A. Sher.1992. Simultaneous depletion of CD4+ and CD8+ T lympho-cytes is required to reactivate chronic infection with Toxo-plasma gondii. J. Immunol. 149:175-180.
16. Hartley, J. W., T. N. Fredrickson, R. A. Yetter, M. Makino, andH. C. Morse II. 1989. Retrovirus-induced murine acquiredimmunodeficiency syndrome: natural history of infection anddiffering susceptibility of inbred mouse strains. J. Virol. 63:1223-1231.
17. Hartley, J. W., N. K. Wolford, L. J. Old, and W. P. Rowe. 1977.A new class of murine leukemia viruses associated with devel-opment of spontaneous lymphomas. Proc. Natl. Acad. Sci.USA 74:789-792.
18. Israelski, D. M., F. G. Araujo, F. K. Conley, Y. Suzuld, S.Sharma, and J. S. Remington. 1989. Treatment with anti-L3T4(CD4) monoclonal antibody reduces the inflammatory responsein toxoplasmic encephalitis. J. Immunol. 142:954-958.
19. Israelski, D. M., G. Showron, J. P. Levental, I. Long, C. F.Blankenship, G. W. Barrio, J. B. Prince, F. G. Araujo, and J. S.Remington. 1988. Toaoplasma peritonitis in a patient withacquired immunodeficiency syndrome. Arch. Intern. Med. 148:1655-1657.
20. Klinken, S. P., T. N. Fredrickson, J. W. Hartley, R. A. Yetter,and H. C. Morse HI. 1988. Evolution of B cell lineage lympho-mas in mice with a retrovirus-induced immunodeficiency syn-drome, MAIDS. J. Immunol. 140:1123-1131.
21. Klinman, D. M., and H. C. Morse Im. 1989. Characteristics of Bcell proliferation and activation in murine AIDS. J. Immunol.142:1144-1149.
22. Loft, B. J., R. G. Brooks, F. K. Conley, R E. McCabe, and J. S.Remington. 1984. Toxoplasmic encephalitis in patients withacquired immune response deficiency syndrome. JAMA 252:913-917.
22a.Makino, M., and H. C. Morse HI. Personal communication.23. Mizuochi, T., A. W. Hugin, H. C. Morse IH, A. Singer, and
R. M. Bulier. 1989. Role of cytokine-secreting CD8+ T cells incytotoxic T lymphocyte responses against vaccinia virus. J.Immunol. 142:270-273.
24. Morse, H. C., Im, R. A. Yetter, C. S. Via, R. R. Hardy, A.Cerny, K. Hayakawa, A. W. Hugin, M. W. Miller, K. L. Holmes,and G. M. Shearer. 1989. Functional and phenotypic alterationsin T cell subsets during the course of MAIDS, a murineretrovirus-induced immunodeficiency syndrome. J. Immunol.143:844-850.
25. Mosier, D. E., R. A. Yetter, and H. C. Morse III. 1985.Retroviral induction of acute lymphoproliferative disease andprofound immunosuppression in adult C57BV6 mice. J. Exp.Med. 161:766-784.
26. Mosmann, T. R., and T. A. T. Fong. 1989. Specific assays forcytokine production by T cells. J. Immunol. Methods 116:151-158.
27. Navia, B. A., C. K. Petito, J. W. M. Gold, E. S. Cho, B. D. Jordan,and R W. Price. 1986. Cerebral toxoplasmosis complicating theacquired immune deficiency syndrome: clinical and neuropath-ological findings in 27 patients. Ann. Neurol. 19.224-228.
28. Parke, D. W., and R L. Font. 1986. Diffuse toxoplasmicretinochoroiditis in patients with AIDS. Am. J. Ophthalmol.104:571-575.
29. Pattengale, P. K., C. R. Taylor, P. Twomey, S. Hill, J. Jonasson,T. Beardsley, and M. Haas. 1982. Immunopathology of B-celllymphomas induced in C57BV/6 mice by dualtropic murineleukemia virus (MuLV). Am. J. Pathol. 107:362-368.
30. Pomeroy, C., S. Kline, M. Colin Jordan, and G. A. Filice. 1989.Reactivation of Toxoplasma gondii by cytomegalovirus diseasein mice: antimicrobial activities of macrophages. J. Infect. Dis.160:305-311.
31. Rosenberg, A. S., T. G. Maniero, and H. C. Morse HI. 1991. Invivo immunologic deficits in mice with MAIDS and effects ofLP-BM5 infection on rejection of skin from infected mice.Transplant. Proc. 23:167-169.
32. Rowe, W. P., W. E. Pugh, and J. W. Hartley. 1970. Plaque assaytechniques for murine leukemia viruses. Virology 42:1136-1139.
33. Sarimiento, M., A. L. Glasebrook, and F. W. Fitch. 1980. IgG orIgM monoclonal antibodies reactive with different determinantson the molecular complex bearing Lyt 2 antigen block T cellmediated cytolysis in the absence of complement. J. Immunol.125:2665-2669.
34. Shearer, G. M., A. Singer, T. Mizuochi, M. Buller, A. W. Hugin,and H. C. Morse HI. 1988. Importance of CD8+ T helper cellfunction in AIDS. J. Infect. Dis. 158:893.
35. Suzuki, Y., F. K. Conley, and J. S. Remington. 1989. Importanceof endogenous IFN--y for prevention of toxoplasmic encephalitisin mice. J. Immunol. 143:2045-2050.
36. Suzuki, Y., M. A. Oreliana, R. D. Schreiber, and J. S. Reming-ton. 1988. Interferon-g: the major mediator of resistance againstT. gondii. Science 240:516-518.
37. Suzuki, Y., and J. S. Remington. 1990. The effect of anti-IFN--yantibody on the protective effect of Lyt-2 immune T cellsagainst toxoplasmosis in mice. J. Immunol. 144:1954-1956.
38. Via, C. S., H. C. Morse m, and G. M. Shearer. 1990. Autoim-mune aspects of human immunodeficiency virus infection andrelevant murine models. Immunol. Today 11:250-255.
39. Vollmer, T. L., M. K. Walder, L. Steinman, and F. K. Conley.1987. Depletion of CD4+ lymphocytes with monoclonal anti-body reactivates toxoplasmosis in central nervous system: amodel of superinfection in AIDS. J. Immunol. 138:3737-3741.
40. Yetter, R. A., R. M. L. Buller, J. S. Lee, K. L. Elkins, D. E. Mosier,T. N. Fredrickson, and H. C. Morse Im. 1988. CD4+ T cells arerequired for development of a murine retrovirus-induced immuno-deficiency syndrome (MAIDS). J. Exp. Med. 168:623-635.
VOL. 60, 1992
on April 25, 2019 by guest
http://iai.asm.org/
Dow
nloaded from


















