On distinguishing the female of Psychodopygus...
14
Reprinted from Annals of Tropical Medicine and Parasitology, Vol. 76, No. 2, 201-214 (1982) On distinguishing the female of Psychodopygus weZZcomei, a vector of mucocutaneous leishmaniasis, from other squamiventris series females I. Characterization of' Ps.* squ'zmiventris squamiventris and Ps. s. mar$aeizsis stat. nov. (Diptera: Psychodidae) BY P. D. READY?, H. FRAIHA The Wellcome Parasitolopy Unitt, Instituto Evandro Chagas, Fundagão Seruifos de Saúde Pública, 4 Belém, Pará, Brazil R. P. LANE British Museum (Natural History) , London, England J. R. ARIAS Instituto flacional de Pesquisas da Amazônia, Manaus, Brazil I AND F.-X. PAJOT Centre O.R.S. T.O,M., Cayenne, French Guiana Received 20 February 1981 The present inability to distinguish accurately Psychodopygus wellcomai females from other females of the series squamiventris, is a serious hindrance to epidemiological studies on mucocutaneous leishmaniasis in Amazônia, where only Ps. wellcomei is a proven vector. The first step in re- appraising the taxonomy of the series has been the characterization of two of its many non- dimensional taxa. These are now recognized as Ps. squamiventrissquamiventris and Ps. s. maripaensis stat. nov., following a morphological study of seven geographically separate adult populations from northeastern Amazônia, including wild and laboratory-reared material from the type locality ofPs. squamiventris (Lutz and Neiva, 1912). The specific status ofPs. complexusis confirmed and the geographical ranges of some of the taxa of the series are amended. The genus Psychodopygus Mangabeira, 1941, sometimes treated as a subgenus of Lutzomyia man-biting catches of phlebotomine sandflies made at ground-level in the non-flooded ('terra firme') primary rain forests of the northern neotropics (Ready et al., 1980). In these forests, however, the vectors of cutaneous leishmaniasis are usually species of .Njssomyia Barretto, 1962, a distinctive subgenus of Lutzomyia (Ready and Fraiha, 1981) ; an exception is Ps. wellcomei Fraiha, Shaw and Lainson, 1971, an important vector of Leishmania braziliensis braziliensis, causative agent of mucocutaneous leishmaniasis in Pará State, Brazil (Lainson et al., 1973). França, 1924, has attracted much attention, partly because its members predominate in I 1 c y >> [ 'F-pQ& *The abbreviation Ps. is used, throughout, for Pyhodopygus, to avoid confusion with P. foPPli-leb?dtolnus? -- i71-, r! p1 I - ! 0003-4983/82/020201+ 14 $02.00/0 Q 1982 Liverpool Sdhool -J.-' No :! 2?.i8 Lx4
Transcript of On distinguishing the female of Psychodopygus...

Reprinted from Annals of Tropical Medicine and Parasitology, Vol. 76, No. 2, 201-214 (1982)
On distinguishing the female of Psychodopygus weZZcomei, a vector of mucocutaneous leishmaniasis, from other
squamiventris series females
I. Characterization of' Ps.* squ'zmiventris squamiventris and Ps. s. mar$aeizsis stat. nov. (Diptera: Psychodidae)
BY P. D. READY?, H. FRAIHA
The Wellcome Parasitolopy Unitt, Instituto Evandro Chagas, Fundagão Seruifos de Saúde Pública, 4 Belém, Pará, Brazil
R. P. LANE
British Museum (Natural History) , London, England
J. R. ARIAS
Instituto flacional de Pesquisas da Amazônia, Manaus, Brazil I
AND
F.-X. PAJOT
Centre O.R.S. T.O,M., Cayenne, French Guiana
Received 20 February 1981
The present inability to distinguish accurately Psychodopygus wellcomai females from other females of the series squamiventris, is a serious hindrance to epidemiological studies on mucocutaneous leishmaniasis in Amazônia, where only Ps. wellcomei is a proven vector. The first step in re- appraising the taxonomy of the series has been the characterization of two of its many non- dimensional taxa. These are now recognized as Ps. squamiventris squamiventris and Ps. s. maripaensis stat. nov., following a morphological study of seven geographically separate adult populations from northeastern Amazônia, including wild and laboratory-reared material from the type locality ofPs. squamiventris (Lutz and Neiva, 1912). The specific status ofPs. complexus is confirmed and the geographical ranges of some of the taxa of the series are amended.
The genus Psychodopygus Mangabeira, 1941, sometimes treated as a subgenus of Lutzomyia
man-biting catches of phlebotomine sandflies made at ground-level in the non-flooded ('terra firme') primary rain forests of the northern neotropics (Ready et al., 1980). In these forests, however, the vectors of cutaneous leishmaniasis are usually species of .Njssomyia Barretto, 1962, a distinctive subgenus of Lutzomyia (Ready and Fraiha, 1981) ; an exception is Ps. wellcomei Fraiha, Shaw and Lainson, 1971, an important vector of Leishmania braziliensis braziliensis, causative agent of mucocutaneous leishmaniasis in Pará State, Brazil (Lainson et al., 1973).
França, 1924, has attracted much attention, partly because its members predominate in I 1 c y >>
[ ' F - p Q & *The abbreviation Ps. i s used, throughout, for Pyhodopygus, to avoid confusion with P. foPPli-leb?dtolnus? -- i71-, r! p1
I - ! 0003-4983/82/020201+ 14 $02.00/0 Q 1982 Liverpool Sdhool -J.-' No :! 2? . i8 Lx4
202 DISTINGUISHING THE FEMALE PSYCHODOPYGUS WELLCOMEZ
r Y ,
A i
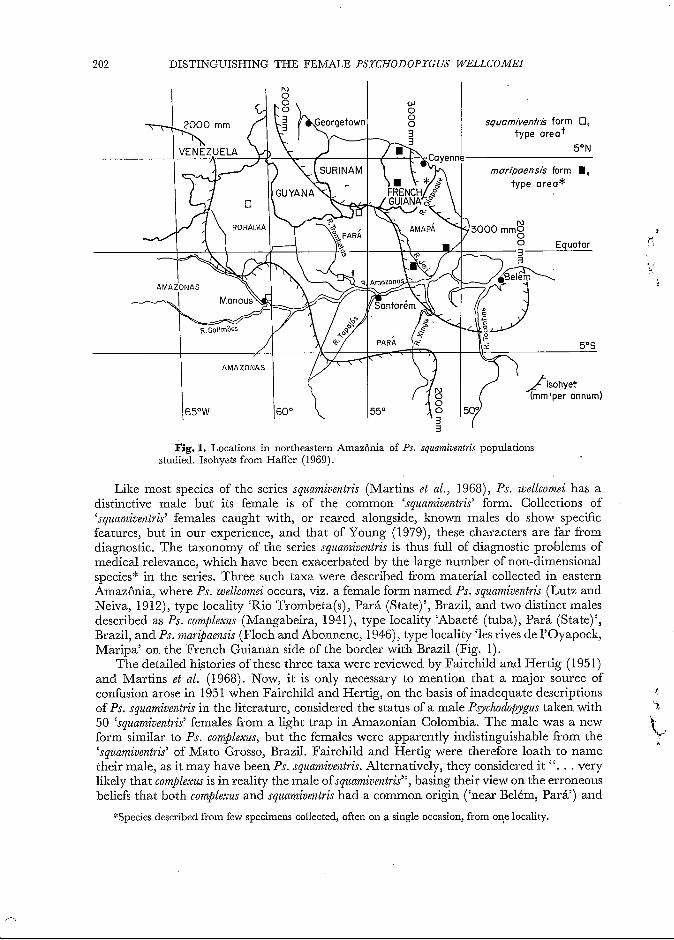
Fig. 1. Locations in northeastern Amazônia of Ps. squamioetztris populations studied. Isohyets from Haffer (1969).
Like most species of the series squamiventris (Martins et al., 1968), Ps. wellcomei has a distinctive male but its female is of the common ‘squamiventris’ form. Collections of ‘squamiventris’ females caught with, or reared alongside, known males do show specific features, but in our experience, and that of Young (1979), these characters are far from diagnostic. The taxonomy of the series squamiventris is thus full of diagnostic problems of medical relevance, which have been exacerbated by the large number of non-dimensional species“ in the series. Three such taxa were described from material collected in eastern Amazônia, where Ps. wellcomei occurs, viz. a female form named Ps. sqziamiventris (Lutz and Neiva, 1912), type locality ‘Rio Trombeta(s), Pará (State)’, Brazil, and two distinct males described as Ps. complexus (Mangabeira, 1941), type locality ‘Abaeté (tuba), Pará (State) ’, Brazil, and Ps. mar$aensis (Floch and Abonnenc, 1946), type locality ‘les rives de l’Oyapock, Maripa’ on the French Guianan side of the border with Brazil (Fig. 1).
The detailed histories of these three taxa were reviewed by Fairchild and Hertig (1951) and Martins et al. (1968). Now, it is only necessary to mention that a major source of confusion arose in 1951 when Fairchild and Hertig, on the basis of inadequate descriptions of Ps. squamiventris in the literature, considered the status of a male Psychodopygus taken with 50 ‘sqziamiventris’ females from a light trap in Amazonian Colombia. The male was a new form similar to Ps. complexus, but the females were apparently indistinguishable from the ‘squamiventris’ of Mato Grosso, Brazil. Fairchild and Hertig were therefore loath to name their male, as it may have been Ps. squamiventris. Alternatively, they considered it “. . . very likely that complexus is in reality the male of squamioe~itris~’, basing their view on the erroneous beliefs that both complexus and squamiventris had a common origin (‘near Belém, Pará’) and
‘“Species described from few specimens collected, often on a single occasion, from one locality.
F 9
k h..

READY E T AL. 203
that Ps. mariljaensis was not another candidate because its palpi, general colouring and wing venation were dissimilar to those of 'squamiventris'.
Later, Martins et al. (1968) recognized the Colombian male as a new species, Ps. fairtigi (Martins, 1970), and designated and fully described the lectotype of Ps. squamiventris. Thereafter, most authors accepted the alternative of Fairchild and Hertig (1951), treating comjlexus as a synonym of squamiventris (Fraiha et al., 1971; Young and Fairchild, 1974; Martins et al., 1978). In contrast, Forattini (1960, 1971, 1973) preferred to treat Ps. maripaensis as the male of Ps. squamiventris, noting the sympatry of these forms both in French Guiana and the adjoining Federal Territory (F.T.) of Amapá, Brazil.
Recently, Ps. com$dexus has been reared in the laboratory (Ward and Ready, unpublished observations), and Fraiha et al. (1978) have recognized it as a distinct species, remarking only that the female was morphologically closer to laboratory-reared females of Ps. wellcomei (Ward, 1971) than to the lectotype of Ps. squamiventris described by Martins et al. (1968). The specific status of Ps. maripaensis was, however, upheld by Fraiha et al. (1978), based on a morphometric difference (the mean length of antennal segment III) between the lectotype female of Ps. squamiveiatris (and seemingly identical material from the F.T. of Roraima) and females from the F.T. of Amapá (Martins et al., 1968).
In this paper, we resolve the status of these three forms by comparing the adult morphology of several geographical populations of 'squainiventris' from northeastern Amazônia, including, for the first time, males bred from females caught in the type locality of Ps. squamiventris (Lutz and Neiva),
7
I Y
MATERIALS AND METHODS
Populations Studied The following adults were collected in 'terra firme' forests north of the river Amazon (Fig. 1).
FRENCH GUIANA (Le Pont and Pajot, col.) Piste de St. Elie (5"26'N, 53'2'W) : 388 (S666-8), 1399 (S678-90) human bait, 1978 Florida (3"36'N, 53'32'W) : 799 (S671-7) human bait, 1978
F.T. OF AMAPÁ, BRAZIL (Fraiha, col.) Curiau, near Macapá (0"8'N, 51'6'W) : 18 (S669), 899 (S554-61) human bait, June 1966
PARÁ STATE, BRAZIL (Ready, Ward, Lane and Shaw, col.) Jari, hill-top forest near Monte Dourado (0"47'S, 52'40'W) : 1588 (S651-65), 1599 (S500-14) light-trap and human bait, February 1976; 1599 (S515-29) human bait, August 1979 Tirios, gallery forest near airstrip (2"20'N, 55'20'W) : 399 (S551-3) human bait, July 1974 Porto Trombetas, hill-top forest near town (1"27'S, 56'24'W) : 3099 (S565-94) human bait,
.
1 April 1980 f
AMAZONAS STATE, BRAZIL (Arias and Freitas, col.) Manaus, CEPLAC forest (2"50'S, 59'55'W) : 2083 (S63 1-50), 3099 (S600-30) horse bait, May-December 1975
F.T. OF RORAIMA, BRAZIL (Fraiha, col.) Mucajai-Caracarai highway, km 13 (2"24'N, 61"O'W) : 1699 (S535-550) human bait, July 1973

204 DISTINGUISHING THE FEMALE PSYCHODOPYGUS WELLCOME1
From these collections, the FI laboratory-reared adults listed below (with slide reference number) were studied; all females came from broods producing at least one male. Jari (as above) 588 (S18-22), 1799 (Sl-17) emerging October 1979 from brood 1 (code 1-79)
” 20 (code 20-79) 488 (S42-5), 299 (S40-1) ” August 1980 ” ” 1 (code 1-80)
” 8 (code 8-80) 388 (S56-8), 499 (S52-5) ” 9 , , l 1 , ” 20 (code 20-80) 18 (S63), 499 (S59-62) ” 2, 11 JY ’ ” 22 (code 22-80) 7$6 (S64-70)
Pt. Trombetas (as above) 488 (S200-3), 299 (S204-5)
a 1 2 1 Y 1 788 (S33-9), 1099 (S23-32) ”
288 (S50-1), 499 (S46-9) ” a , , Y 2 1
emerging August 1980 from other broods
emerging May 1980 from brood 21 (code 21-80) ” 33 (code 33-80)
1, 11 11 ” 43 (code 43-80) ” 56 (code 56-80) ” 63 (code 63-80) ” 53 (code 53-80)
1, Y , 1 3 388 (S206-8), 599 (S209-13) ”
388 (S214-6), 69$! (S217-22) ”
586 (S223-7), 799 (S228-34) ”
888 (S23542), 699 (S243-8 388 (S253-5)
1, 11 , Y
2 , a 1 1 1 11
9 1 11 Y 1 2 1
388 (S249, 251, 256), 299 (S250, 252) emerging May 1980 from other broods.
Rearing Methods Wild, engorged females were caught and maintained in oviposition tubes of the type described by Ward (1977), whose petri dish method of rearing larvae was also followed. Larval food, however, was a mixture of desiccated liver powder and dry, powdered Daphnia. All stages were reared at 23-27°C and approximately 90% RH, in ambient photoperiods in Belém, Pará State, Brazil. Broods were maintained separately at all times.
Adult Morphological Characters Examined All material was slide mounted in Euparal (French Guiana) or Berlese fluid (the remainder). The difference in mounting medium was not thought to affect significantly the measurements made. All qualitative characters of known taxonomic value for neo- tropical phlebotomines were examined using light microscopy (cf. Young, 1979). Following the recognition by Martins et al. (1968) of morphometric differences between some species of the squamiventris series and their importance in accurate identification, a study of morpho- metric characters was made. A preliminary examination of randomly chosen specimens was made to select characters to be recorded. They were (for males and females unless stated otherwise) : length of head, labrum, palp segments 3 and 5, antennal segments III and IV, thorax, wing, and (males only) genital pump and filaments; wing indices a, ß, y, 6 (Young, 1979); the number of upper and lower mesanepisternal setae; and the number of vertical cibarial teeth (females only). The thorax was measured from the anterior edge of the mesoscutum to the posterior tip of the mesoscutellum.
Analysis Quantitative characters were compared using Dice-Leraas type diagrams, after calculating the 95% confidence limits for population means with d or Student’s t statistics, depending on sample size (Bailey, 1968). For any given character, differences between means for two different populations are often significant at the 5% level when these statistics are employed and, partly for this reason, such differences are not always considered to have taxonomic significance; rather, the degree of overlap between a character’s normal frequency distribution for two populations is taken as a more accurate guide to whether or not the observed variation between the two populations is discontinuous (cf. Mayr, 1969).

READY ET AL. 205
A useful means of estimating overlap was given by Mettrick (1963), who adapted the formula for calculating Mayr's coefficient of difference so that, for any given character,
1 /2 (mean, population 1 -mean, population 2) common standard deviation, as for t test
C = '
The percentage overlap is then (1 -P) 100, where P is the integral of the normal distribution from minus infinity to C; tables for obtaining P from C are commonly available, e.g. 'Documenta Geigy' (Diem, 1964).
Mayr et al. (1953) suggested that two populations should be considered different subspecies if there was less than 10% overlap in any one character, once discrete geo- graphical distribution had been ascertained. Amadon (1 949) had been more conservative, electing the value of 3y0, while more recently Mayr (1969) reported that many taxonomists have been treating 5% overlap as critical. A 5% rule was applied in the present study. The shortcomings of such an approach are discussed later.
G
'"4
,I 4
RESULTS
The laboratory-reared, F, females from Pt. Trombetas broods were indistinguishable, on both qualitative and quantitative characters, from the lectotype of Ps. squamiventris described by Martins et al. (1968) (Figs. 2, 3,4) ; the lectotype was examined by one of us (H.F.). F, males from the same broods were morphologically similar both to the male described as Ps. maripaensis by Floch and Abonnenc (1946) and to the laboratory-reared F, males from Jari broods (Figs. 4, 5). However, the observed variation over all the popula- tions studied, both laboratory and wild, was greater than that expected for a single species. Two morphologically distinct groups could be recognized with associated geographical distributions, the east-west boundary between them being near longitude 55"W (Fig. 1).
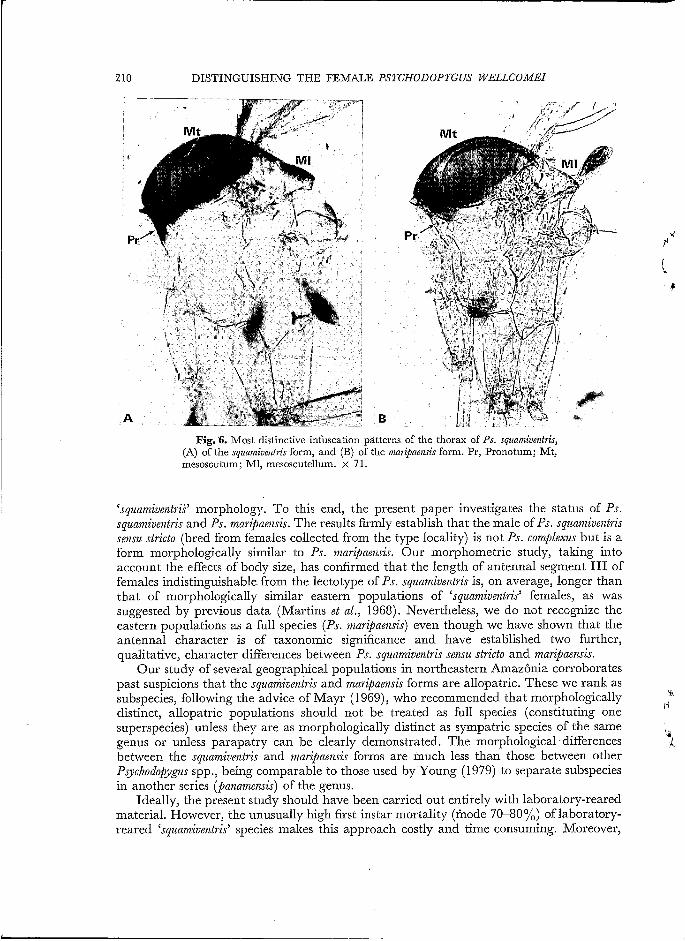
For both sexes, the third and fourth antennal segments of the western populations were significantly longer (overlap <5%) than those of the eastern populations; further, the pronotum was heavily infuscated in all members of the western populations but, with the exception of a few specimens from French Guiana (2/23), more lightly infuscated in members of the eastern populations (Fig. 6) .This difference is diagnostic for populations, not for individuals. The female thoraces figured show further differences which were common to the two forms : the infuscation of the mesoscutum and mesoscutellum was often more intense in the squamiventris (western) form, but this was far from exclusive. Like most sandflies, the male thorax was less intensely infuscated than the female, but the difference between the two geographical forms was still apparent.
For the males, there was, between eastern and western populations, a consistent difference in the structure of the terminal enlargement of the elongate, upturned apical process of the paramere (Fig. 7). The original drawing of the male holotype of Ps. maripaemis is, in this respect, misleading, for the apical process of the paramere was drawn inverted and the terminal enlargement was only sketched.
Concerning the female quantitative characters investigated, the following showed few statistically significant differences between populations and no taxonomically reliable differences (overlaps were 20+%) : head height, length of palp segment 3 and segment 5, number of vertical cibarial teeth, length of thorax, number of lower mesanepisternal setae, wing length, a, ß, y, 6, alß and alS. Moreover, these data suggested no distinctive ratios. Wing length and thorax length were highly correlated and, therefore, the latter was used as a measure of size, being the less easily broken of the two structures. Some results are given in Fig. 3 to illustrate the agreement between the female lectotype of Ps. squamiventris and the western populations studied.
* r,
/ y

206 DISTINGUISHING THE FEMALE PSYCHODOPYGUS WELLCOMEI
Chorocter, Antenna 73 ( p m l LobrumMeod height No. upper mesonepisfernol setae
250300350400450 bo0 1.05 1.10 1.15 1.20 IO 15 20 25 30 Population + Trombelos: FI 99 of brood 21-80 + 2 t
Trombetos; FI 99 of brood 33-80 5
+ 6
$ 6
Trombetas FI pp of brood 43-80
Trombetas FI O? of brood 56-80 7
Trombetos: FI ??of brood 63-80
Trombetas o11 FI ?? -e Trombetos: wild-cought?? $ 29
Monous: wild-cought ?? -$-3O ,+
Roroimo: wild-cough1 99 $- 16 +I5
Tirios: wild-cough1 ??
$- Jori: 6 ?y of brood 1-79
Jori: Fi ?? of brood 20-79
+ + 4-
+
Jori: FI 99 of brood 8-80
Jori: FI 99 of brood 20-80
Jori: FI 99 of brood 22-80
Jori: o11 Fi 99
Jori: wild-caught ?? -$-
AmopÓ: wild -caught ??
Fr. Guiono: wild-caught?? +/+
t,Number In sample
$ 3 3 t
IO.+ IO
4 +
4 4 3
4 I - t - l
17 -$- 17
41 .4, 41
30 30
8 + 8
20 .+ 20
XS,Lectotype I?) of Ps. squumiventris ofter Martins et u/. (1968)
-Et?-
+
2
5
6 '
7
6
28
30
30
15
3
Fig. 2. Morphometric variation within and between female populations of Ps. squamiventris; antenna III, alone, has taxonomic significance. (Vertical bar = estimated population meari; horizontal bar = observed range; rectangle = 95% confidence limits for estimated population mean.)

READY E T AL. 207
Chorocter+ No. Vertical ciborio1 Labrum ( p n 1 Lobrum/Antenna E Antenna lU/Antenno E Population J, teeth
IO 20 30 40 440 480520 560600 1.2 1.4 1 6 1.8 2.4 2.6 2.8 3.0 ---rr”
XS XS X S X S
Trombetas: a11 FI Ç‘P -E+- 28t -!+I- 28 + 28 + 28
Roraimo: wild-caught O? -$ 16 .+I6 16 -E& 16
Jori: a11 FI 99 41 __$_ 41 +- 41
Fr. Guiona: wild-caught 99 + 19 + 20 -+ 20 -+ 20
Trombetas: a11 F1 O? -+ ~-E+- 28--$--- 28 .+. 28
a/ß Wing length /Thorax Charocter+ Antenna E? ( p m ) No. lower Population $ mesonepisternol setae
I20 130 140 150 6 8 IO I2 2.0 2.4 2.8 3.2 36 3.4 3.6 3.8 4.0 ---- X S XS X S
Roraima: wild-cought 99 16 15+ 15 IO
Fr. Guiano:wild-caught -$-- 20 20 20 fB 15
t , Number in sample Xs Lectotype ( 0 ) of Ps. squamiventris after Martins et al. (1968)
Fig. 3. Morphometric variation within and between female populations of Ps. squamiventris; antenna IV, alone, has taxonomic significance.
Two female characters-length of labrum (corrected for body size) and number of upper mesanepisternal setae-showed the greatest variation between individual populations. However, when these characters were analysed population by population, no general geographic trend emerged, i.e. the differences or similarities between individual populations did not correspond to the basic division into ‘eastern populations’ and ‘western populations’ and the variation was found to be as great within these two pools as between them. For example, although some eastern populations showed no significant overlap with some western populations, two widely separated populations such as Manaus and French Guiana showed a complete overlap for both characters (Fig. 2). The results for these two characters are given in full so that their probable lack of diagnostic value is clear to anyone investigating other populations of Ps. squamiventris.
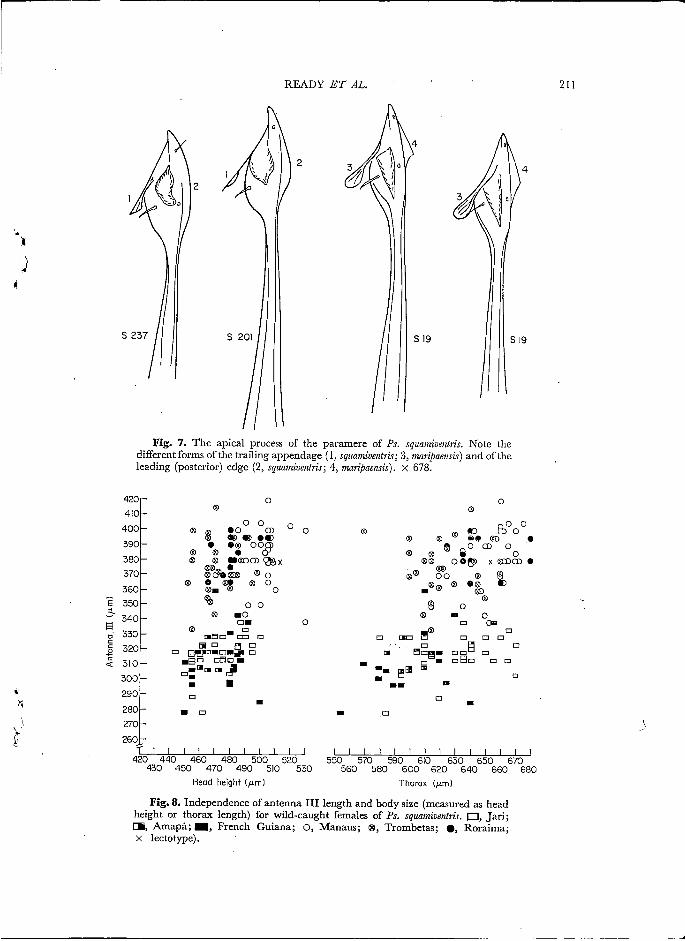
Of the female quantitative characters studied, only the lengths of antennal segment III and antennal segment IV had taxonomic significance; length was independent of body size, as illustrated for antennal segment III in Figs. 4 and 8. Indeed, within the eastern group or the western group of populations the mean length of antennal segment III was remarkably constant (Fig. 2) ; the overlap between pooled eastern wild populations and pooled western wild populations was only 3*4%, while between the laboratory-reared populations of Jari and Pt. Trombetas the overlap was but 1.1%. The length of antennal segment I V was highly correlated with that of antennal segment III and, as for the latter, there was no significant overlap in absolute length between eastern and western populations (Fig. 3).
t P
f ‘P A

208 DISTINGUISHING THE FEMALE PSYCHODOPYGUS WELLCOMEI
Character + Antenna IU / Head ht. Population 4
Antenna IU /Thorax
0.6 0.7 0.8 O9 1.0 111 1.2 1.3 1.4 050 O60 070 0.80 090 I I I I I I r l I I [ I \ ( J I I I )
Fr.Guiana Eh AmopÓ: 28t .+- 23 wild
Trombetas: a11 F, 28 28
Monous wild ?? a Roraima: 45 .-& 40
4 4 Fr. Gui0n.a 81 AmopÓ: wild do"
Jori: a11 FI $d 29 -4- 29
Trombetas: o11 FI d d
Manaus: wild dcr'
t, Number in sample Xs, Lectotype ( 9 ) of Ps. squumiventris after Martins el' u/. (19681 after Floch and Abonnenc (1946)
XM, Holotype (0") of Ps. muripuensis
Fig. 4. Morphometric variation within and between populations of Ps. squamiventris; discontinuity between squamiventris (western) and maripaensis (eastern) forms is greater for males than for females.
The same conclusions hold for the male quantitative characters investigated (Figs.' 4, 5). Again, only the antennae showed differences of taxonomic significance; for antennal segment III, the overlap between eastern and western population groups was less than that for the females, being 0*05°/0 for wild males and 0.13% for laboratory-reared males. This more marked discontinuity (Fig. 4) meant that the ratio labrum length/antenna III length (often quoted for neotropical sandflies) had diagnostic value for the males (Fig. 5) which it had not for the females (Fig. 3).
The ratio length of genital filaments/length of sperm pump (often useful in sandfly taxonomy) had no diagnostic value for the males studied. Further, observations indicated no marked differences between populations with respect to the relative lengths of style, coxite, paramere and lateral lobe of the male external genitalia.
DISCUSSION
Based on male genitalia alone, the series squamiventris could be considered to contain up to ten described species. These are (with the sexes originally described), in chronological order: Ps. squamiventris ( 8 ) ; Ps. chugmi (Costa Lima, 1941) ( 8 ) ; Ps. comtlexus (8); Ps. geniculatus (Mangabeira, 1941) (8) ; Ps. maripaensis (8) ; Ps. bernalei (Osorno-Mesa, Morales- Alarcon & de Osorno, 1966) ($) ; Ps. fairtigi (8) ; Ps. wellcomei ($, $!) ; Ps. lainsoni Fraiha h Ward, 1974 (8, 9 ) ; and Ps. corossoniensis (Le Pont & Pajot, 1978) (8). Laboratory

READY E T AL. . 209
Character LabrumIHead ht. Labrum IAntennoIII Antenna III (pm) Na. upper mesanepisternal setae Papuiatian $
Manaus: wild-caught d d 20
054 0.62 O7 0.47 059 0.71 300 350400 450 500 6 8 IO 12 1416 18202224 mm-m
Trombetas: a11 FI 0” d ++ 29
Jari: a11 F, d d ++ 29
_-
4 3 - 1 5 -4- 15 ‘ .*I5
X M X M --+ l5
Jari: wild -caught a‘ a*
X M
$ 3 +3 t 3 $ 3 Fr. Guiana: wild- caught dd
i
Character, Antenna III /Antenna JZ Antenna E (pm) Wing length/Thorax Genital f i laments/ Population + Sperm pump
2.3 2.5 27 2.9 120 140 160 180 3 4 3.6 3.8 4.0 1.8 2.0 2.2 2.4 ---- Trombetas: a11 FI o”d ,+29 -$- 29 ft3- 29
29 -$- 28 .+ 29
X M
Jari: a11 FI do”
Fr. Guiana: wild-caught$ d + 3 t 3 - i - 3 t 3
t, Number in sample XM, holotype (d) of Ps. maripaensis after Fioch and Abonnenc (1946)
Fig. 5. Morphometric variation within and between male populations of Ps. squamiuenntuis; only antenna III, antenna IV and labrum/antenna III have taxonomic significance.
rearing has associated the sexes of both Ps. wellcomei and Ps. lainsoni, as noted in the original descriptions, of Ps. cliagasi (Fraiha et al., 1974), of Ps. complexus (Ward and Ready, un- published observations) and of both Ps. geiziculatus and Ps. corossoiiieizsis, whose females are similar to that of Ps. gzyatzensis (Ready and Ward, unpublished observations).
The females of Ps. corossoniensis, Ps. geleliiculatus and Ps. lainsoni differ from others in the series squamiventris by having a long common spermatheca1 duct. All other females that have been associated with males of the series have internal genitalia of the common ‘squamiventris’ kind : the common spermathecal duct is short and barely noticeable, while the individual spermathecal ducts are pronounced, being longer than the body of the spermatheca. As stated in the introduction, there are female characters which appear to be typical of certain species, but they are far from being diagnostic at the individual level. For example, the base of the individual spermathecal ducts are usually neither inflated nor sculptured in Ps. chagasi (Martins et al., 1968), while the same structures ase often markedly inflated but only faintly sculptured in Ps. squamiventris (Martins et al., 1968) and slightly inflated and markedly sculptured in Ps. wellcomei (Fraiha st al., 1971) and in Ps. complexus (Ward and Ready, unpublished observations) ; further, females of some populations of Ps. squamiventris and Ps. chagasi are separable on at least one morphometric character (Martins et al., 1968).
The immediate taxonomic goal for the series squamiventris is, therefore, the diagnostic characterization of the females (described or not) which appear to have the common
210 DISTINGUISHING THE FEMALE PSYCHODOPYGUS WELLCOMEI
B
Fig. 6. Most distinctive infuscation patterns of the thorax of Ps. squamive?ztris, (A) of the squamiventris form, and (B) of the mari@ensis form. Pr, Pronotum; Mt, mesoscutum; M1, mesoscutellum. x 71.
‘squamiventris’ morphology. To this end, the present paper investigates the status of Ps. squamiventris and Ps. maripaensis. The results firmly establish that the male of Ps. squamiventris sensu stricto (bred fi-om females collected from the type locality) is not Ps. complexus but is a form morphologically similar to Ps. maripaensis. Our morphometric study, taking into account the effects of body size, has confirmed that the length of antennal segment III of females indistinguishable from the lectotype of Ps. squamiuentris is, on average, longer than that of morphologically similar eastern populations of ‘squamiventris’ females, as was suggested by previous data (Martins et al., 1968). Nevertheless, we do not recognize the eastern populations as a full species (Ps. maripaensis) even though we have shown that the antennal character is of taxonomic significance and have estaljlished two further, qualitative, character differences between Ps. squamioentris sensu stricto and maripaensis.
Our study of several geographical populations in northeastern Amazônia corroborates past suspicions that the squamiventris and maripaensis forms are allopatric. These we rank as subspecies, following the advice of Mayr (1969) , who recommended that morphologically distinct, allopatric populations should not be treated as full species (constituting one superspecies) unless they are as morphologically distinct as sympatric species of the same
between the squamiventris and maripaensis forms are much less than those between other Psychodopygris spp., being comparable to those used by Young (1979) to separate subspecies in another series (panamensis) of the genus.
Ideally, the present study should have been carried out entirely with laboratory-reared material. However, the unusually high first instar mortality (mode 70-80%) of laboratory- seared ‘squamiuentris’ species makes this approach costly and time consuming. Moreover,
l i
ri
genus or unless parapatry can be clearly demonstrated. The morphological differences ‘2
2
S 237
400 390 380 370 360 -
E 350- i - 340-
W 0. 330- 3 320- 2 310-
C
300 290 280
READY ET AL.
- - - - -
- - -
P l 2
4 \
s 19
211
i.
5 19
Fig. 7. The apical process of the paramere of Ps. squanziventris. Note the different forms of the trailing appendage (1, squamiventris; 3, maripaerzsis) and of the leading (posterior) edge (2, squamivetztris; 4, maripaensis). x 678.
420 O 410L o
260 270L
O 0
O
+ 1 1 1 1 1 1 1 1 l 1 1
430 450 470 490 510 530 420 440 460 480 500 520
Heod height ( p m )
fa
O CD
8 A0O0 o o o m m e
oc4 00@ xmCDe BQ O 0
O C D o Q " 8 O
8 0 " 0
o c m o o o m o a o 0 0
I o
I O
1 1 1 1 1 1 l I l I I I l I 550 570 590 610 630 650 670
Thorax (pm) 560 580 600 620 640 660 680
Fig. 8. Independence of antenna III length and body size (measured as head height or thorax length) for wild-caught fema!es of Ps. squamiventris. O, Jari; IE, Amapá ;æ , French Guiana; O, Manaus; O, Trombetas; 0, Roraima; x lectotype).

2 12 DISTINGUISHING THE .FEMALE PSYCHODOPYGUS WELLCOMEI
we feel that i t is most unlikely that our wild material contained other species, for the following reasons : (1) Although females were not reared from the type locality of Ps. maripaensis, laboratory- reared males and females from Jari were indistinguishable from wild flies collected in French Guiana. (2) The majority of characters, both qualitative and morphometric, did not mismatch within either the eastern or the western populations, as defined. (3) Other species whose females could be confused individually with squamiventris and maripaensis forms are extremely rare, or have never been recorded, in the area studied. It is for the last reason that our investigation of the taxonomy of the series squamiventris has been started with populations collected exclusively from! northeastern Amazônia.
The approximate boundaries of the present study area are the river Amazon, the Atlantic Ocean, latitude 6"N and longitude 62"W (Fig. 1). Within this area, the males of Ps. bernalei, Ps. complexus and Ps. fairtigi have never been recorded, while the male of Ps. wellcomei and the male and female of Ps. chagasi have only rarely been captured (and then only to the west of Pará State); in contrast, males similar to Ps. maripaensis have been collected, sometimes in large numbers, from most of the locations providing material for the present study. Previous distributional records come from literature reviews (Forattini, 1973; Martins et al., 1978) and from a large number of captures (from man baits, rodent baits, tree trunks, animal burrows and light traps) made in the F.T. of Roraima (Arias and Freitas, unpublished observations), in Amazonas State (Arias and Freitas, 1977; Arias and Young, unpublished observations), in Pará State (Lainson et al., 1979; Ready and Ward, unpublished observations), in the F.T. of Amapá (Forattini, 1960; Fraiha, un- published observations) and in French Guiana (Léger et al., 1977, 1980; Pajot and Le Pont, unpublished observations).
Our findings mean that some former distribution records need revising. Records of Ps. squamiventris in Colombia, Venezuela, Guyana and Surinam (Martins et al., 1978) should be treated as squamiventris sensu lato, because the females to which they refer may belong to different subspecies or, as in Colombia, to different species. Females of Ps. squamiventris reported from French Guiana are almost certainly Ps. s. maripaensis (Floch and Abonnenc, 1952; Léger et al., 1977) and Arias and Freitas (1977) were clearly dealing with Ps. s. squamiventris, not Ps. s. maripaensis, in Manaus. Martins et al. (1978) would be expected to have reported Ps. complexus, not Ps. squamiventris, from Abaetetuba and Belém, both in Pará State, Brazil. Further, the presence of Ps. squamiventris in the dry states to the east of Pará (Forattini, 1973) is unsubstantiated. Indeed, there are no valid reports of Ps. squamiventris sensu stricto being found south of the river Amazon.
Forattini (1973) lists Ps. squamiuentris as a possible vector of cutaneous leishmaniasis in northeastern Amazônia, but this is a supposition based only on the species anthropophily. In fact, neither squamiventris sensu stricto nor maripaensis is a proven vector of Leishmania, despite numerous dissections in search of flagellates (Arias and Freitas, 1978; Lainson et al., 1979). In this respect, the degree of biological separation between populations of Ps. quamiventris does not have any direct epidemiological significance to date, and the treat- ment of maripaensis as a subspecies can be supported on the grounds that the trinomial conveys two important pieces of information, namely allopatry and closest morphological relationship within the genus (6 Mayr, 1969). Should Ps. squamiventris be incriminated as a vector of leishmaniasis, then its taxonomy will have to be studied further: classification as subspecies is of limited use to the epidemiologist, for the subspecies is not a biological category; its limits are set arbitrarily, with statistical evaluation of morphological dis- continuity serving only to standardize, somewhat, intuitive judgements.
Nevertheless, it is still interesting to speculate on the factors responsible, in the past or present, for isolating the defined subspecies of squamiventris. One possible explanation is

READY E T AL. 213
provided by the ‘refuge theory’ advanced, primarily, by Haffer (1969) and Vanzolini (1973). This postulates that during the dry climatic periods of the Pleistocene, when much water was trapped as ice nearer the Poles, the Amazonian forest was divided into a number of smaller forests, separated by drier savannas, and that these ‘forest refuges’, by encouraging separate evolution by isolation, help to explain the faunal and floral diversity of the Basin. Based on recent climatic patterns and on the observed distributions of various vertebrates, lepidopterans and plants (Prance, 1973), the eastern end of the Guianan Shield and an area north of Manaus are believed to have been the two principal refuges in northeastern Amazônia. Psychodo#ygus s. squamiveiitris and Ps. s. maripaensis could have evolved within these refuges, but their allopatry can be explained equally well in terms of present-day climate. Within much of the known range of Ps. s. squamiveiatris in northeastern Amazônia, the annual rainfall is not more than 1500 mm with a marked dry season lasting three to four months; in contrast, the annual rainfall in the range of Ps. s. maripueilsis is 2000-3000 mm and the dry season is usually shorter and less intense (Fig. 1).
ACKNOWLEDGEMENTS. These studies were funded by the Wellcome Trust (London) , Fundaçã0 Serviços de Saúde Pública (Rio d e Janeiro), WHO Special Programme for Research and Training in Tropical Diseases (Geneva), C.N.Pq. (Brasilia) and O.R.S.T.O.M.’ (Paris). We are much indebted to the companies JAR1 Floresta1 e Agropecuária Ltda and Mineraçã0 Rio Norte for facilities provided in Monte Dourado and Porto Trombetas, respectively. Also, we acknowledge with thanks the technical assistance of Sras. Suêd de N. F. Silva and Graça M. Soares and Srs. Iorlando da R. Barata, José I. de Almeida, Manoel C. M. de Souza and Rui A. Freitas.
i
. I
REFERENCES
AMADON, D. (1949). The seventy-five per cent rule for sub-species. Condor, 51, 250-258. ARIAS, J. R. & FREITAS, R. A. (1977). Flebótomos da Amazônia central do Brasil. 1. Resultados obtidos das
ARIAS, J. R. & FREITAS, R. A. (1978). Sôbre os vetores de leishmaniose cutânea na Amazônia central do
BAILEY, N. T. J. (1968). Statistical Methods in Biology. London: English Universities Press. DIEM, K. (1964). Scientijc Tables (Documenta Geigy). Manchester: Geigy Pharmaceutical Co. Ltd. FAIRCHILD, G. B. & HERTIG, M. (1951). Notes on the Phlebotomus of Panama (Diptera, Psychodidae). VII.
The subgenus Shannonomjina Pratt. Annals of the Entomological Society of America, 44, 39942 1. FLOCH, H. & ABONNENC, E. (1946). Phltbotomes de la Guyane française (XVII). P. maripaensis n. sp. et P. sp.
de Maripa. Publication de l’Institut Pasteur de la Guyane francaise, 140, 1-6. FLOCH, H. & ABONNENC, E. (1952). Diptères Phlébotomes de la Guyane et des Antilles frangaises. Faune de l’Union
Française. XIV. Paris : O.R.S.T.O.M. FORATTINI, O. P. (1960). Notas sôbre Phlebotomus do Território Amapá, Brasil (Diptera, Psychodidae). Studia
Entomologica, 3, 467-480. FORATTINI, O. P. (1971). Nota sôbre Psychodojygus sguamiventris (Lutz & Neiva, 1912) e especies afins (Diptera,
Psychodidae). Revista de Saúde Pública, São Paulo, 5, 151-154. FORATTINI, O. P. (1973). Entomologia Médica. 4. Psychodidae. Phlebotominae. Leishmanioses. Bartonelose. São
Paulo: Blücher. FRAIHA, H., SHAW, J. J. & LAINSON, R. (1971). Phlebotominae brasileiros-II. Psychodojygus wellcomei, nova
especie anthropófila de flebótomo do grupo sguamiventris, do su1 do estado do Pará, Brasil (Diptera, Psycho- didae). Memorias do Instituto Oswaldo Cruz, 69, 489-500.
FRAIHA, H. & WARD, R. D. (1974). A new species of Psychodojygus Mangabeira (Diptera, Phlebotomidae) from the Transamazônica highway, Pará State, Brazil. Bulletin of Entomological Research, 64, 209-2 12.
FRAIHA, H., WARD, R. D., LOUREIRO, C. A. & SOARES, G. M. (1974). Flebotomideos brasileiros-IV. Nota sôbre Psychodojygus chagasi (Costa Lima, 19-11) (Diptera: Phlebotomidae). Revista Brasileira de Biologia, 34,
FRAIHA, H., WARD, R. D., SHAW, J. J. & LAINSON, R. (1978). Fauna antropóiìla de Flebótomus da Rodovia Transamazônica, Brasil (Diptera, Psychodidae). Bulletin of the Pan-American Health Organization, 84, 134-1 39.
capturas feitas com iscas humana e eqüina (Diptera, Psychodidae). Acta Amazonica, 7, 507-527.
Brasil. 2. Incidência de flagelados em flebotómos selváticos. Acta Amaronica, 8, 387-396.
89-9 1.

_ . g" 214
HAFFER, J. (1969). Speciation in Amazonian forest birds. Science, 165, 131-137. LAINSON, R., SHAW, J. J., WARD, R. D. & FRAIHA, H:(1973). Leishmaniasis in Brazil. IX. Considerations on
the Leishmania braziliensis complex: importance of sandflies of ihe genus Psychodopygus Mangabeira in the transmission of L. braziliensis in north Brazil. 'Transdctions of the Royal Socie[v of Tropical Medicine and HTgiene, 67, 184-196.
LAINSON, R., SHAW, J. J., WARD, R. D., READY, P. D. & NAIFF, R. D. (1979). Leishmaniasis in Brazil. XIII. , Isolation of Leishmania from armadillos (Dasypus novemcinctzrs), and observations on the epidemiology of cutaneous leishmaniasis in north Pará State. Transactions of the Royal Society of Tropical Medicine and Hygiene,
LOGER, N., ABONNENC, E. & CLAUSTRE, J. (1980). L'anthropophilie des PhlCbotomes de la Guyane française. Bulletin de la Société de Pathologie Exotique, 73, 112-123.
L~GER, N., ABONNENC, E., PAJOT, F. X., KRAMER, R. & CLAUSTRE, J. (1977). Liste commentCe des Phlébotomes de la Guyane française. Cahiers O.R.S. T.O.M., séric Entomologie Médicale et Parasitologie, 15, 2 17-232.
LE PONT, F. & PAJOT, F. X. (1978). Lutzomyia (Psychadopygus) corossoniemis n. sp. (Diptera: Psychodidae): nouveau Phlébotome découvert en Guyane française. Cahiers O.R.S. T. O.M., série Entomologie Médicale et Parasitologie, 16, 223-226.
MANGABEIRA, O. (1941). 4" contribuição ao estudo dos Flebótomos. Psychodop?tgus n. subg. (Diptera: Psycho- didae). Memorias do Instituto Oswaldo Cruz, 36, 237-250.
MARTINS, A. V. (1970). Lutzomyia (Psychodopygzrs) fairtigi n. sp. from Colombia (Diptera: Psychodidae: Phlebotominae). Proceedings of the Entomological SorieQ Df Washington, 72, 279.
MARTINS, A. V., MARCIEL, C. S. & DA SILVA, J. E. (1968). Notas sôbre os flebótomos do grupo squamiventris do subgenero Psychodopygus Mangabeira, 191.1 (Diptera, Psychodidae). Boletim do Museu Historia Natural da Universidade Federal de Minas Gerais, zoologia, 1, 1-33.
MARTINS, A. V., WILLIAMS, P. & FALCÃO, A. L. (1978). American SandFlies (Diptera: Ptvchodidae: Phlebotominae). Rio de Janeiro: Agademia Brasileira de Ciencias.
MAYR, E, (1969). Princ+les o f Systematic zoology. New York: McGraw-Hill. MAYR, E., LINSLEY, E. G. & USINGER, R. L. (1953). Methods and Principles of Systematic zoology. New York:
McGraw-Hill. METTRICK, D. F. (1963). A statistical analysis of the morphological variation observed between populations of
zouorchis petiolatum (Railliet, 1900) (Trematoda: Dicrocoeliidae) from different hosts and localities. Journal of Parasitolopy, 49, 745-75 1.
PRANCE, G. T. (1973): Phytogeographic support for the theory of Pleistocene forest refuges in the Amazon Basin. Acta Amazonica, 3, 5-28.
READY, P. D. & FRAIHA, H. (1981). Brazilian phlebotomines-VI. Lutzomyia richardwardi sp. n., a new species of Nyssomyia suhg. from Amazônia, with keys for the identification of the males and females of this subgenus (Diptera, Psychodidae): Revista Brasileira de Biologia, 41.
READY, P. D., FRAIHA, H., LAINSON, R. & SHAW, J. J. (1980). Psychodopygus as a genus: reasons for a flexible classification of the phlebotomine sand flies (Diptera: Psychodidae). Journal of Medical Entomology, 17,75-88.
VANZOLINI, P. E. (f973). Paleoclimates, relief and species multiplication in equatorial forests. In Tropical Forest Ecosystems in Afr ia and South America: A Comparative Review, eds B. J. Meggers et al. Washington, D.C.: Smithsonian Institution Press.
DISTINGUISHING THE FEMALE PSYCHODOPYGUS WELLCOMEI
73,239-242.
WARD, R. D. (1971). (Cited'in Fraiha et al. (1971): 9.a) WARD, R. D. (1977).'The colonization of Lutromyia~auiscutellata (Diptera: Psychodidae), a vector of Leishmania
YOUNG, D. G. (1 979). A rmiew of the bloodsucking Psychodidjies ofColombia (Diptera: Phlebotoniinae and Sycoracinae).
YOUNG, D. G. & FAIRCHILD, G. B. (1974). Studies of Phlebotornine San&ies. Gainesville, Florida: Annual Report
mexicana amazonensis in Brazil. Journal of Medical Entomology, 14,469-476.
Gainesville, Florida: Agricultural Experiment Stations Institute of Food and Agricultural Sciences.
of the Department of Entomology and Nematology, University of Florida. 9
I
Printed in Great Britain by Henry Ling Ltd., at the Dorset Press, Dorchester, Dorset I