
NT3-chitosan elicits robust endogenous neurogenesis … · NT3-chitosan elicits robust endogenous...
6
NT3-chitosan elicits robust endogenous neurogenesis to enable functional recovery after spinal cord injury Zhaoyang Yang a,b,1 , Aifeng Zhang c , Hongmei Duan b , Sa Zhang d , Peng Hao a , Keqiang Ye e , Yi E. Sun f,g,1 , and Xiaoguang Li a,b,1 a Department of Neurobiology, School of Basic Medical Sciences, Capital Medical University, Beijing 100069, China; b Department of Biomedical Engineering, School of Biological Science and Medical Engineering, Beihang University, Beijing 100191, China; c Beijing Friendship Hospital, Capital Medical University, Beijing 100068, China; d National Center of Biomedical Analysis, Beijing 100850, China; e Department of Pathology and Laboratory Medicine, Center for Neurodegenerative Diseases, Emory University School of Medicine, Atlanta, GA 30322; f Stem Cell Translational Research Center, Tongji Hospital and Department of Regenerative Medicine, Tongji University School of Medicine, Shanghai 200065, China; and g Department of Psychiatry and Biobehavioral Sciences, David Geffen School of Medicine at University of California Los Angeles, Los Angeles, CA 90095 Edited by Thomas C. Südhof, Stanford University School of Medicine, Stanford, CA, and approved August 26, 2015 (received for review May 24, 2015) Neural stem cells (NSCs) in the adult mammalian central nervous system (CNS) hold the key to neural regeneration through proper activation, differentiation, and maturation, to establish nascent neural networks, which can be integrated into damaged neural circuits to repair function. However, the CNS injury microenvironment is often inhibitory and inflammatory, limiting the ability of activated NSCs to differentiate into neurons and form nascent circuits. Here we report that neurotrophin-3 (NT3)-coupled chitosan biomaterial, when inserted into a 5-mm gap of completely transected and excised rat thoracic spinal cord, elicited robust activation of endogenous NSCs in the injured spinal cord. Through slow release of NT3, the biomaterial attracted NSCs to migrate into the lesion area, differentiate into neurons, and form functional neural networks, which interconnected severed ascending and descending axons, resulting in sensory and motor behavioral recovery. Our study suggests that enhancing endogenous neurogenesis could be a novel strategy for treatment of spinal cord injury. spinal cord injury | NT3 | chitosan | functional recovery | endogenous neurogenesis R egeneration of axons in the central nervous system (CNS) is dampened by a nonpermissive microenvironment after injury (1–6). Following spinal cord injury (SCI), a cascade of patho- logical processes occurs, including the breaking down of the vas- culature system, edema, infiltration of immune cells, inflammation, initiation of wound healing processes, seizing of ongoing neuro- transmission, gliosis/glial scar formation, cell death, demyelination, and remyelination (7), along with activation of neural stem/ progenitor cells (NPCs) attempting to participate in neural repair (8). Often, prolonged activation of immune cells, including micro- glia, lymphocytes, and macrophages, leads to secondary lesions of the nervous system, creating a very harsh environment for already compromised CNS neurons to regenerate axons (9). In the past, much of the attention in SCI research focused on how to promote the regrowth of severed corticospinal tract (CST) for fairly long distances to reconnect with their originally innervated alpha motor neurons in the ventral horn and restore function. In recent years, several lines of evidence have suggested that perhaps a more feasible alternative proregeneration strategy is to generate nascent local spinal interneurons to provide a relay neuronal net- work across the injury site, serving as a “bridge” for the severed axons to establish connections with their targeted motor neurons without long-distance axonal growth (10, 11). It has been widely accepted that neural stem cells (NSCs) reside in many regions of the adult CNS (8, 12, 13). CNS injury frequently causes neuronal loss. Although new neurogenesis may occur to some extent spontaneously (14), the inflammatory injury environ- ment does not allow for sufficient neurogenesis. Instead, it is pro- gliosis, which is involved in glial scar formation. Researchers have explored various methods to promote effective neurogenesis (15, 16); however, all of those studies concerned only brain injury repair. Few if any studies have addressed the promotion of regeneration after SCI via activation of endogenous new neurogenesis in the spinal cord. We have previously shown that neurotrophin-3 (NT3) facilitates proliferation of NSCs in vitro through activation of the TrkC re- ceptor and promotes neurogenesis in the spinal cord (17, 18). NT3 has also been shown to facilitate axonal growth, including in the CST, in vivo (19). Using a biodegradable chitosan carrier in com- bination with NT3, we achieved prolonged slow release of NT3 over a period of 14 wk (18). In the present study, we aimed to determine whether slow release of NT3 using chitosan (Fig. S1) to replace a 5-mm thoracic segment of the spinal cord after complete transection and removal of the segment could create an optimized microenvironment to promote new endogenous neurogenesis. We further examined whether the subsequent formation of a nascent synaptic network within the NT3-chitosan tube could functionally bridge the two ends of the severed cord and achieve regeneration in severe cases of SCI. Results NT3-Chitosan Enables Nerve Regeneration and Functional Recovery. Three major experimental groups were included in this study: the lesion control (LC) group, in which a 5-mm thoracic spinal cord segment at T7-8 was surgically removed after complete transec- tion of the cord; the empty tube (ET) group, in which an empty (i.e., non–NT3-loaded) chitosan tube was inserted into the 5-mm gap following surgical removal of the thoracic segment; and the Significance Spinal cord injury (SCI) is a debilitating medical condition with no cure at present time. In this study we have discovered that a bio- degradable material, chitosan, when loaded with, Neurotrophin-3 (NT3), allowed for slow release of this neural trophic factor, pro- viding an optimal microenvironment for regeneration. NT3-chito- san, when inserted into a 5 mm gap of completely transected and excised rat thoracic spinal cord, elicited robust activation of en- dogenous neural stem cells forming functional neural networks, which interconnected the severed ascending and descending axons, resulting in sensory and motor behavioral recovery. Our study suggests that enhancing endogenous neurogenesis by NT3- chitosan could be a novel strategy for treatment of SCI. Author contributions: Z.Y. and X.L. designed research; Z.Y., A.Z., H.D., S.Z., P.H., and X.L. performed research; Z.Y., K.Y., and X.L. analyzed data; and Z.Y. and Y.E.S. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. 1 To whom correspondence may be addressed. Email: [email protected], yi.eve.sun@ gmail.com, or [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1510194112/-/DCSupplemental. 13354–13359 | PNAS | October 27, 2015 | vol. 112 | no. 43 www.pnas.org/cgi/doi/10.1073/pnas.1510194112
Transcript of NT3-chitosan elicits robust endogenous neurogenesis … · NT3-chitosan elicits robust endogenous...

NT3-chitosan elicits robust endogenous neurogenesisto enable functional recovery after spinal cord injuryZhaoyang Yanga,b,1, Aifeng Zhangc, Hongmei Duanb, Sa Zhangd, Peng Haoa, Keqiang Yee, Yi E. Sunf,g,1,and Xiaoguang Lia,b,1
aDepartment of Neurobiology, School of Basic Medical Sciences, Capital Medical University, Beijing 100069, China; bDepartment of Biomedical Engineering,School of Biological Science and Medical Engineering, Beihang University, Beijing 100191, China; cBeijing Friendship Hospital, Capital Medical University,Beijing 100068, China; dNational Center of Biomedical Analysis, Beijing 100850, China; eDepartment of Pathology and Laboratory Medicine, Center forNeurodegenerative Diseases, Emory University School of Medicine, Atlanta, GA 30322; fStem Cell Translational Research Center, Tongji Hospital andDepartment of Regenerative Medicine, Tongji University School of Medicine, Shanghai 200065, China; and gDepartment of Psychiatry and BiobehavioralSciences, David Geffen School of Medicine at University of California Los Angeles, Los Angeles, CA 90095
Edited by Thomas C. Südhof, Stanford University School of Medicine, Stanford, CA, and approved August 26, 2015 (received for review May 24, 2015)
Neural stem cells (NSCs) in the adult mammalian central nervoussystem (CNS) hold the key to neural regeneration through properactivation, differentiation, and maturation, to establish nascent neuralnetworks, which can be integrated into damaged neural circuits torepair function. However, the CNS injury microenvironment is ofteninhibitory and inflammatory, limiting the ability of activated NSCs todifferentiate into neurons and form nascent circuits. Here we reportthat neurotrophin-3 (NT3)-coupled chitosan biomaterial, when insertedinto a 5-mm gap of completely transected and excised rat thoracicspinal cord, elicited robust activation of endogenous NSCs in theinjured spinal cord. Through slow release of NT3, the biomaterialattracted NSCs to migrate into the lesion area, differentiate intoneurons, and form functional neural networks, which interconnectedsevered ascending and descending axons, resulting in sensory andmotor behavioral recovery. Our study suggests that enhancingendogenous neurogenesis could be a novel strategy for treatmentof spinal cord injury.
spinal cord injury | NT3 | chitosan | functional recovery |endogenous neurogenesis
Regeneration of axons in the central nervous system (CNS) isdampened by a nonpermissive microenvironment after injury
(1–6). Following spinal cord injury (SCI), a cascade of patho-logical processes occurs, including the breaking down of the vas-culature system, edema, infiltration of immune cells, inflammation,initiation of wound healing processes, seizing of ongoing neuro-transmission, gliosis/glial scar formation, cell death, demyelination,and remyelination (7), along with activation of neural stem/progenitor cells (NPCs) attempting to participate in neural repair(8). Often, prolonged activation of immune cells, including micro-glia, lymphocytes, and macrophages, leads to secondary lesions ofthe nervous system, creating a very harsh environment for alreadycompromised CNS neurons to regenerate axons (9).In the past, much of the attention in SCI research focused on how
to promote the regrowth of severed corticospinal tract (CST) forfairly long distances to reconnect with their originally innervatedalpha motor neurons in the ventral horn and restore function. Inrecent years, several lines of evidence have suggested that perhaps amore feasible alternative proregeneration strategy is to generatenascent local spinal interneurons to provide a relay neuronal net-work across the injury site, serving as a “bridge” for the severedaxons to establish connections with their targeted motor neuronswithout long-distance axonal growth (10, 11).It has been widely accepted that neural stem cells (NSCs) reside
in many regions of the adult CNS (8, 12, 13). CNS injury frequentlycauses neuronal loss. Although new neurogenesis may occur tosome extent spontaneously (14), the inflammatory injury environ-ment does not allow for sufficient neurogenesis. Instead, it is pro-gliosis, which is involved in glial scar formation. Researchers haveexplored various methods to promote effective neurogenesis (15,16); however, all of those studies concerned only brain injury repair.
Few if any studies have addressed the promotion of regenerationafter SCI via activation of endogenous new neurogenesis in thespinal cord.We have previously shown that neurotrophin-3 (NT3) facilitates
proliferation of NSCs in vitro through activation of the TrkC re-ceptor and promotes neurogenesis in the spinal cord (17, 18). NT3has also been shown to facilitate axonal growth, including in theCST, in vivo (19). Using a biodegradable chitosan carrier in com-bination with NT3, we achieved prolonged slow release of NT3over a period of 14 wk (18). In the present study, we aimed todetermine whether slow release of NT3 using chitosan (Fig. S1) toreplace a 5-mm thoracic segment of the spinal cord after completetransection and removal of the segment could create an optimizedmicroenvironment to promote new endogenous neurogenesis. Wefurther examined whether the subsequent formation of a nascentsynaptic network within the NT3-chitosan tube could functionallybridge the two ends of the severed cord and achieve regenerationin severe cases of SCI.
ResultsNT3-Chitosan Enables Nerve Regeneration and Functional Recovery.Three major experimental groups were included in this study: thelesion control (LC) group, in which a 5-mm thoracic spinal cordsegment at T7-8 was surgically removed after complete transec-tion of the cord; the empty tube (ET) group, in which an empty(i.e., non–NT3-loaded) chitosan tube was inserted into the 5-mmgap following surgical removal of the thoracic segment; and the
Significance
Spinal cord injury (SCI) is a debilitating medical condition with nocure at present time. In this study we have discovered that a bio-degradable material, chitosan, when loaded with, Neurotrophin-3(NT3), allowed for slow release of this neural trophic factor, pro-viding an optimal microenvironment for regeneration. NT3-chito-san, when inserted into a 5 mm gap of completely transected andexcised rat thoracic spinal cord, elicited robust activation of en-dogenous neural stem cells forming functional neural networks,which interconnected the severed ascending and descendingaxons, resulting in sensory and motor behavioral recovery. Ourstudy suggests that enhancing endogenous neurogenesis by NT3-chitosan could be a novel strategy for treatment of SCI.
Author contributions: Z.Y. and X.L. designed research; Z.Y., A.Z., H.D., S.Z., P.H., and X.L.performed research; Z.Y., K.Y., and X.L. analyzed data; and Z.Y. and Y.E.S. wrotethe paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence may be addressed. Email: [email protected], [email protected], or [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1510194112/-/DCSupplemental.
13354–13359 | PNAS | October 27, 2015 | vol. 112 | no. 43 www.pnas.org/cgi/doi/10.1073/pnas.1510194112

NT3-chitosan tube (NT3) group. The loading of NT3 onto thechitosan tube is described in detail in Fig. S1.In all three groups, gross anatomic changes within areas of le-
sions were examined at various time points from 3 d to 1 y afterinjury. At the light dissecting microscopic level, at 3 d after theoperation, a small amount of tissue was observed at the rostral andcaudal ends of the lesion area in all groups (Fig. S2A). At 10 d afterthe operation, the lesion area was occupied with fibrous and cystictissues in the LC group. This process continued and eventuallyformed scar-like tissues, anatomically “gluing” the two ends ofsevered cord together (Fig. S2A, Middle and Fig. 1A, Right). Forthe ET group, at day 10, tissue infiltration was no longer present,and the tube remained empty from day 10 and onward to 1 y afterinjury (Fig. S2A, Bottom, and Fig. 1A, Middle). In the NT3 group,tissues continued to grow into the tube from two ends, graduallynarrowing the tissue gap, and by day 30, tissues from both therostral and caudal ends of the cord fused (Fig. S2A, Top). The di-ameters of newly formed tissues were initially irregular and notuniform. New tissues continued to grow and remodel, and by day90 formed a smooth nerve bundle-like structure that was thick atthe two ends and thin in the middle (Fig. 1A, Left). This anatomicstructure was stabilized from day 90 and onward, up to at least 1 yafter injury (Fig. 1A and Fig. S2B).Using neural filament (NF), Nissl, and neuron-specific enolase
(NSE) staining, we confirmed that regenerated tissues in the NT3group were neural tissues composed of nerve fibers as well asneuronal cells (Fig. 1B and Fig. S3 A and B). NF-positive nervefibers observed in the NT3-chitosan tube could be composed ofregenerated ascending and descending fibers, as well as new fibersoriginating from newly regenerated neuronal cells. Quantitativeanalyses of the exact contribution from each of these three sourcesawait future studies.To determine whether regenerated nerve tissues could elicit be-
havioral functional recovery, we used the Basso–Beattie–Bresnahan(BBB) open-field walking scale to measure hindlimb locomotoractivity of rats after SCI. As expected, hindlimb locomotion waszero immediately after the operation for all of the experimental rats.Over the course of 1 y, the right and left hindlimb BBB scores wereno higher than 1.5 in the LC and ET groups (Fig. 1C). In contrast,
in the NT3 group, the mean BBB scores for right and left hindlimbsincreased significantly at week 4, and continued to rise steadily up to1 y, at which time scores were 10-fold higher than those at week 1(11.4–12.2; Fig. 1C). To evaluate whether this result was related to acompensatory effect rather that spinal cord regeneration, at 52 wkafter the first operation, regenerated nerve bundles were retrans-ected and a plastic diaphragm was placed in the relesion site. As aresult, the mean hindlimb BBB scores dropped from 11.4–12.2 to0–1 (not significantly different from those in the ET and LCgroups), and remained at this low level for the next 5 wk (Fig.1C). The retransection results indicate that the restoration oflocomotor function observed in the NT3 group was most likelyrelated to reestablishment of synaptic transmission from motoraxons crossing the lesion area to elicit motor behavior, but notdue to a compensatory response below the lesion area.We studied five additional control groups to confirm that NT3-
chitosan enabled the nerve-like tissue growth and functional re-covery through activation of the NT3/TrkC pathway. We infusedTrkC tyrosine kinase (NT3 receptor) inhibitors, k-252a, k-252b,and an NT3-neutralizing antibody into NT3-chitosan tubes for12 wk using osmotic pumps and found that, just like in the ETgroup, tissue growth was no longer present inside the NT3-chi-tosan tube with a blockade of NT3/Trk signaling and BBB scoreswere all <1.5 (Fig. 1C). Because the k-252a and k-252b weredissolved in dimethyl sulfoxide (DMSO), we also infused DMSOinto both the NT3-chitosan and empty chitosan tubes to serve ascontrols, and found that DMSO did not influence the experimentaloutcome (Fig. 1C).On EM analysis, the ultrastructure of the regenerated tissue in
the NT3 group showed numerous myelinated axons with typicallayered myelin sheets confined to endoneurium- or perineurium-like structures (Fig. S4A). Over time, the number of myelinatedfibers at the caudal end of the NT3-chitosan tube increased andmore or less stabilized from around 26 wk up to 1 y (Fig. S4B).Based on myelin morphologies and the number of myelinatedaxons associated with each myelinating cell, we determined thatboth Schwann cells (SCs) and oligodendrocytes (OLs) partici-pated in the remyelination process (20) (Fig. S4A). In contrast,no regenerated myelinated axons were found in the middle or
Fig. 1. The NT3-chitosan tube facilitated regeneration and functional recovery after complete SCI. (A) Dorsal view of representative spinal cords from threetreatment groups—the NT3 group, ET group, and LC group—at 12 mo after the operation. Note the presence of nerve tissue-like regenerated cable (blackarrows) in the NT3 group. No tissue growth was seen in the ET group. Arrowheads mark the location of undegraded chitosan tubes. The white arrow indicatesapparent scar tissues. (B) NF staining of regenerated nerve fibers in the NT3 group at 12 mo after the operation. Note presence of the NF-positive regen-erating fibers growing inside the NT3-chitosan tube, traversing the lesion site, and reaching the caudal end of the spinal cord. Almost no NF staining wasfound across the lesion site in the LC group, indicative of no regeneration. (C) BBB open-field walking scores of bilateral hindlimbs for all eight groups overtime, up to 52 wk. At 4 wk after the operation and thereafter, the NT3 groups with or without DMSO had significantly higher BBB scores in both the right andleft hindlimbs compared with the other groups (mean ± SE; n = 8; P < 0.05, one-way ANOVA with Bonferroni post hoc test). Retransection was performed at52 wk after the initial lesion. In the middle of the lesion area (n = 4), a plastic diaphragm was inserted to prevent regeneration, with an additional 5 wk of BBBscoring (mean ± SE; n = 4; P < 0.05, paired Student t test comparing before and after recutting). Retransection dropped BBB scores back to complete SCI levels.
Yang et al. PNAS | October 27, 2015 | vol. 112 | no. 43 | 13355
NEU
ROSC
IENCE
caudal ends of the lesion areas in the LC or ET group (Fig. S4).In the LC group, the bulk of the tissue in the rostral ends wascomposed of nonaxonal scar tissues and some degenerated axons(Fig. S4A). Moreover, blood vessels wrapped by endothelial cellswere frequently found in the regenerating tissue under EM, in-dicative of good vascularization (Fig. S4A).To further confirm sensory and motor functional restorations,
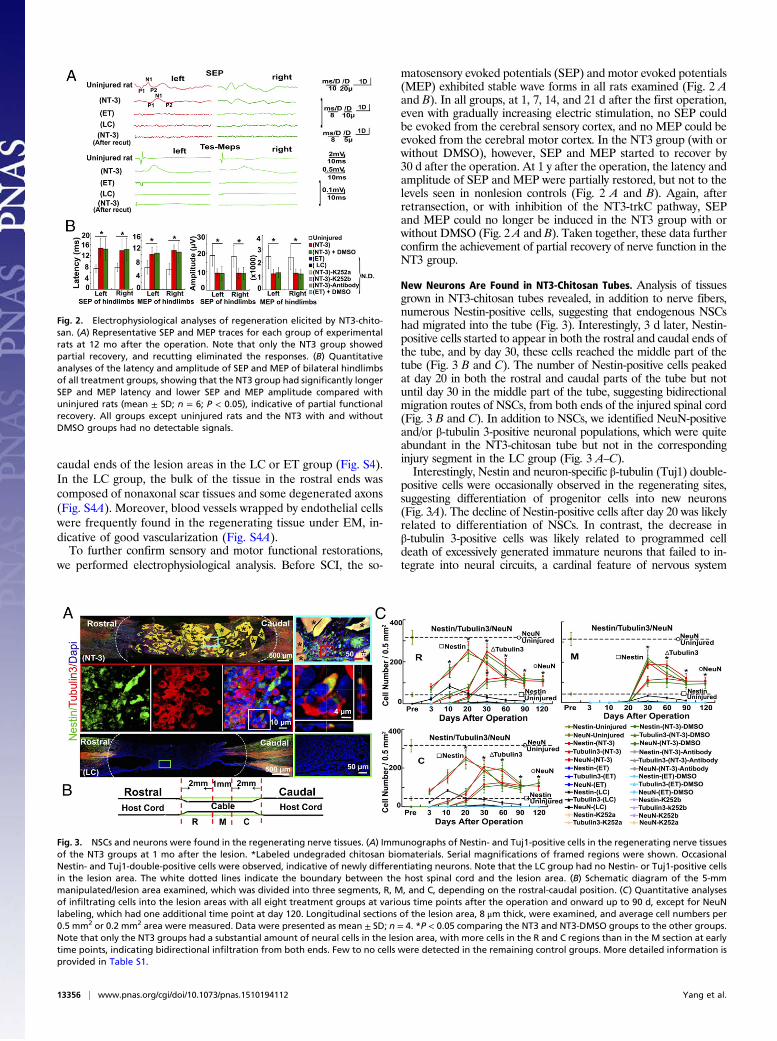
we performed electrophysiological analysis. Before SCI, the so-
matosensory evoked potentials (SEP) and motor evoked potentials(MEP) exhibited stable wave forms in all rats examined (Fig. 2 Aand B). In all groups, at 1, 7, 14, and 21 d after the first operation,even with gradually increasing electric stimulation, no SEP couldbe evoked from the cerebral sensory cortex, and no MEP could beevoked from the cerebral motor cortex. In the NT3 group (with orwithout DMSO), however, SEP and MEP started to recover by30 d after the operation. At 1 y after the operation, the latency andamplitude of SEP and MEP were partially restored, but not to thelevels seen in nonlesion controls (Fig. 2 A and B). Again, afterretransection, or with inhibition of the NT3-trkC pathway, SEPand MEP could no longer be induced in the NT3 group with orwithout DMSO (Fig. 2 A and B). Taken together, these data furtherconfirm the achievement of partial recovery of nerve function in theNT3 group.
New Neurons Are Found in NT3-Chitosan Tubes. Analysis of tissuesgrown in NT3-chitosan tubes revealed, in addition to nerve fibers,numerous Nestin-positive cells, suggesting that endogenous NSCshad migrated into the tube (Fig. 3). Interestingly, 3 d later, Nestin-positive cells started to appear in both the rostral and caudal ends ofthe tube, and by day 30, these cells reached the middle part of thetube (Fig. 3 B and C). The number of Nestin-positive cells peakedat day 20 in both the rostral and caudal parts of the tube but notuntil day 30 in the middle part of the tube, suggesting bidirectionalmigration routes of NSCs, from both ends of the injured spinal cord(Fig. 3 B and C). In addition to NSCs, we identified NeuN-positiveand/or β-tubulin 3-positive neuronal populations, which were quiteabundant in the NT3-chitosan tube but not in the correspondinginjury segment in the LC group (Fig. 3 A–C).Interestingly, Nestin and neuron-specific β-tubulin (Tuj1) double-
positive cells were occasionally observed in the regenerating sites,suggesting differentiation of progenitor cells into new neurons(Fig. 3A). The decline of Nestin-positive cells after day 20 was likelyrelated to differentiation of NSCs. In contrast, the decrease inβ-tubulin 3-positive cells was likely related to programmed celldeath of excessively generated immature neurons that failed to in-tegrate into neural circuits, a cardinal feature of nervous system
Fig. 2. Electrophysiological analyses of regeneration elicited by NT3-chito-san. (A) Representative SEP and MEP traces for each group of experimentalrats at 12 mo after the operation. Note that only the NT3 group showedpartial recovery, and recutting eliminated the responses. (B) Quantitativeanalyses of the latency and amplitude of SEP and MEP of bilateral hindlimbsof all treatment groups, showing that the NT3 group had significantly longerSEP and MEP latency and lower SEP and MEP amplitude compared withuninjured rats (mean ± SD; n = 6; P < 0.05), indicative of partial functionalrecovery. All groups except uninjured rats and the NT3 with and withoutDMSO groups had no detectable signals.
Fig. 3. NSCs and neurons were found in the regenerating nerve tissues. (A) Immunographs of Nestin- and Tuj1-positive cells in the regenerating nerve tissuesof the NT3 groups at 1 mo after the lesion. *Labeled undegraded chitosan biomaterials. Serial magnifications of framed regions were shown. OccasionalNestin- and Tuj1-double-positive cells were observed, indicative of newly differentiating neurons. Note that the LC group had no Nestin- or Tuj1-positive cellsin the lesion area. The white dotted lines indicate the boundary between the host spinal cord and the lesion area. (B) Schematic diagram of the 5-mmmanipulated/lesion area examined, which was divided into three segments, R, M, and C, depending on the rostral-caudal position. (C) Quantitative analysesof infiltrating cells into the lesion areas with all eight treatment groups at various time points after the operation and onward up to 90 d, except for NeuNlabeling, which had one additional time point at day 120. Longitudinal sections of the lesion area, 8 μm thick, were examined, and average cell numbers per0.5 mm2 or 0.2 mm2 area were measured. Data were presented as mean ± SD; n = 4. *P < 0.05 comparing the NT3 and NT3-DMSO groups to the other groups.Note that only the NT3 groups had a substantial amount of neural cells in the lesion area, with more cells in the R and C regions than in the M section at earlytime points, indicating bidirectional infiltration from both ends. Few to no cells were detected in the remaining control groups. More detailed information isprovided in Table S1.
13356 | www.pnas.org/cgi/doi/10.1073/pnas.1510194112 Yang et al.

development (21). NeuN labeled more mature neurons, which didnot decline even at 120 d, long after NT3 had been exhausted (i.e.,at 14 wk after the operation) (Fig. 3C).To confirm that neurons detected within the NT3-chitosan tubes
were newly born neurons from endogenous NSCs rather than mi-grated preexisting neurons, we administered daily i.p. injections of5-bromo-2 -deoxyuridine (BrdU) to rats from day 1 to day 7 afterthe operation. This labeled proliferative cells and newly born neu-rons in vivo during this period. At 4 wk postsurgery, approximately57% of the Nestin-positive cells were labeled with BrdU, and 64%of the Tuj1-positive and 67% of the NeuN-positive neurons wereBrdU-positive, indicating that these neurons were generated afterthe operation (Fig. 4 A and B, Fig. S5, and Fig. S6 A–C).The BrdU-labeling method detected many cell types in addition
to neural progenitors, particularly after SCI. In fact, in the LCgroup, but not in the ET or the NT3 group, some BrdU-positivecells appeared earlier in the middle part of the lesion area (Fig.S6E). Since these BrdU cells in the LC group were not labeled withNestin or Tuj1, they were most likely infiltrating inflammatory im-mune cells (22). Moreover, the fact that those cells did not infiltrateinto the middle part of the chitosan tubes at such early time pointsin the ET and NT3 groups suggests that the chitosan tubes with orwithout NT3 likely created an anti-inflammatory environment earlyafter injury and, when coupled with NT3, supported new neuro-genesis and axonal regeneration (22).
Newly Born Neurons Form a Nascent Functional Neural Network. Toexplore the underlying mechanisms by which newly generatedneurons may participate in locomotor functional recovery, wehypothesized that newly born neurons may form nascent synapticnetworks that serve as information relay stations, bridging as-cending and descending neural transmission signals across the
lesion. Using BDA (dextran, fluorescein, and biotin) to label cor-tical motor axons and BrdU/MAP2 to label newly born neurons inthe NT3-chitosan tube, we found evidence that CST axons makecontact (likely synaptic) with newly born neurons (Fig. S6F).We used immunoelectron microscopy technology to investigate
whether morphological synaptic connections were also establishedamong newly generated neurons. At 4 wk after the operation,immunogold-labeled Tuj1-positive axons and dendrites formedmature symmetrical or nonsymmetrical synapses in the middle ofthe NT3-chitosan tube (Fig. 5A).To determine whether the morphological synapses detected
were electrophysiologically active, we used a planar multielectrodedish system (MED64; Alpha MED Scientific) to reveal features ofthe neuronal network formed in the NT3-chitosan tubes. In ad-dition, we used pharmacologic compounds to determine whetherthe field potentials recorded by the MED64 system in the lesionarea were transmitted by synapses formed among newly bornneurons. Stimulation of the caudal part of the regenerated spinalcord tissue in the lesion area produced multisite neural responseswithin the lesion area, as well as in the host spinal cord caudal tothe lesion (Fig. 5B). The amplitude of field excitatory postsynapticpotentials (fEPSP) recorded at multiple sites in the lesion area aswell as in the host spinal cord caudal to the lesion could besignificantly suppressed by 6-cyano-7-nitroquinoxaline-2, 3-dione(CNQX; 10 μM), suggesting that major excitatory neurotrans-missions were glutamatergic. All of the effects of the pharmaco-logic agents on fEPSP were reversible on reperfusion (i.e., washoutof CNQX), as expected. Taken together, these data indicate thatfunctional synaptic connections can be established among newlygenerated neurons in the lesion area as well as with the host spinalcord, and that these new synaptic connections likely participate inthe locomotor functional recovery.
Fig. 4. NT3-chitosan promoted new endogenous neurogenesis in the lesionarea after SCI. BrdU was injected i.p. at 24 h postsurgery and then daily forthe first 7 d after the operation. Stars indicated undegraded chitosan bio-materials. The white dotted lines indicated the boundary between the hostspinal cord and the lesion area. (A) Immunographs showing BrdU-labeledNestin- and Tuj1-expressing neural lineage cells in the lesion area at 1 mo afterlesion. Serially amplified images of highlighted regions, as well as single opticsections of confocal images with Z-stack demonstrated colabeling. (B) Quan-titative analyses of immunohistochemistry shown in A. Data were presented asmean ± SD; n = 4. *P < 0.05, comparing the NT3 group to the LC group.
Fig. 5. Formation of functional synaptic connections among newly bornneurons in the NT3-chitosan tube and the host spinal cord caudal to thelesion. (A) Immunogold-labeled β-tubulin 3–positive axons and dendritesforming symmetrical (Upper) or asymmetrical (Lower) mature synapses (ar-rows). At, axonal terminal; Den, dendritic side of the synapse as shown bypostsynaptic density. *Undegraded chitosan materials. (B) Photograph oftypical MED64 with 8 × 8 arrays (interelectrode distance, 150 μm) of proberecordings on the spinal cord slice, including the lesion/regenerated areafrom the NT3-chitosan group at 4 wk after operation. The narrow “cable”represented regenerated tissues at the top. While stimulating one site(yellow star) in the caudal part of the regenerated spinal cord, the fieldexcitatory postsynaptic potential (fEPSP) was recorded at multisite in thelesion area and in the caudal of host spinal cord, which could be suppressedby CNQX (10 μM), suggesting that the nature of the synaptic transmissionwas glutamatergic. The red arrowhead pointed to an fEPSP recorded atelectrode 58. The red dotted line indicated the boundary between the re-sidual host caudal spinal cord and the lesion/regenerated area.
Yang et al. PNAS | October 27, 2015 | vol. 112 | no. 43 | 13357
NEU
ROSC
IENCE

NPC Activity and Functional Recovery. To address whether mitoticactivation of endogenous NSCs and subsequent neurogenesis wereimportant for NT3-chitosan–induced functional recovery afterSCI, we used an antimitotic drug, cytosine β-D-arabinofuranoside(Ara-C) (23) to ablate proliferative progenitors. In the NT-3 group,Ara-C was infused into the rat lateral ventricle via a micropump for14 d. As shown in Fig. S7A, Ara-C treatment significantly de-creased the number of Nestin-positive neural progenitors andNeuN-positive neurons within the NT3-chitosan tubes at 4 wkafter the operation. BBB scores in the bilateral hindlimbs at 1 ydecreased by 30–40% after Ara-C treatment compared with noAra-C treatment (Fig. 6A).Interestingly, there was no significant decrease in the latency
of SEP and MEP, but an approximate 30% decrease in ampli-tude was apparent (Fig. 6B). A partial CST tracking experimentwith BDA labeling (with eight 0.4-uL injections of 10% BDA)indicated that Ara-C did not change the number of regeneratingaxons, which likely contributed to the maintenance of SEP andMEP latencies (Figs. S7B and S8). Changes in the amplitude, onthe other hand, suggested that in the NT3 group, large amountof regenerated neurons in the lesion area likely participated inlocomotor functional recovery.
DiscussionAn Excellent Microenvironment Ensured Regeneration and FunctionalRecovery. Through this series of studies, we have demonstrated thatendogenous NSCs have significant potential to regenerate and re-pair CNS injuries. A major obstacle limiting the ability of endoge-nous NSCs to achieve neural repair is the hostile microenvironmentin the injured CNS resulting from myelin-associated axonal growthinhibitory agents and the inflammatory immune environment, whichprohibit neurogenesis from NSCs and promote scar-forming gliosis.Therefore, the establishment of an excellent growth-promoting
microenvironment is key to enable endogenous and, when appli-cable, exogenous NSCs to generate new neurons and form nascentneural circuits that can be used for neural repair.In the present study, prolonged (14 wk) slow release of NT3 was
achieved by coupling it with chitosan (18), a biodegradable ma-terial, to create a regeneration-promoting microenvironment (Fig.S1). With NT3-chitosan, endogenous NSCs and their progenieswere effectively activated and attracted to migrate into the NT3-chitosan tubes and subsequently differentiate into neurons, whichmatured to form a nascent functional synaptic network. We haveshown that such nascent local neural networks can be used byregenerating descending and ascending axons through synapticconnections without the need for long-distance growth, providing aunique regeneration strategy using endogenous NSCs (Fig. 7).Previously, we reported that chitosan loaded with collagen alsopromoted axonal regeneration in semisectioned spinal cord (24);however, there were very few neurons in the regenerated tissues,suggesting that NT3 is much more neurogenic than collagen.
Three Mechanisms Underlying the Regenerative Power of NT3-Chitosan.Previous SCI research has largely focused on ways to overcome theinhibitory CNS environment as well as enhance CNS neuronalintrinsic regenerative potentials for nerve fibers to reextend fromthe CST. In recent years, the Sofroniew and Egerton groups haveshown that spinal proprial interneurons are crucial for motorfunction recovery after partial SCI (25). Moreover the Nakashimagroup has demonstrated that functional recovery from SCI can befacilitated by transplantation of NPCs and that the subsequentneuronal differentiation from NPCs is crucial (11). Our presentfindings show that after complete SCI, the use of the NT3-chitosantube, through activation of NT3-TrkC signaling, elicits two bene-ficial events: enabling of long-distance nerve fiber growth, andenhancement of new neurogenesis and subsequent formation ofa functional new neural synaptic network. Through inhibition ofnew endogenous neurogenesis, behavioral recovery was partiallyimpaired, as were MEP and SEP amplitudes, demonstrating thecontribution of nascent relay neuronal networks created via acti-vation of endogenous NSCs and subsequent neurogenesis. More-over, we also found that the NT3-chitosan tube not only acted onneural cells, but also elicited inhibition of the inflammatory im-mune process (22), both of which could contribute to better nerveregeneration and adult neurogenesis.We do acknowledge that the initial application of Ara-C for 2 wk
not only suppressed NSC activation, but also inhibited proliferationof other cells, including infiltrating immune cells and endothe-lial cells. The fact that neurogenesis was impaired even long afterAra-C was no longer present suggests that the initial phase of NSCactivation remained important for proper production of new neu-rons at the injury site (Fig. S7). Whether the initial blockade of
Fig. 6. Blockade of stem cell activity and new neurogenesis led to worsenedfunctional recovery. (A) BBB scores of animals belonging to the NT3 group,untreated or treated with Ara-C. *P < 0.05 (n ≥ 5), comparing Ara-C–treatedand untreated groups. (B) SEP/MEP latencies and amplitudes in uninjured,NT3-chitosan, Ara-C–treated, and untreated groups. *P < 0.05 (n ≥ 5),comparing uninjured and NT3-chitosan groups and Ara-C–treated and un-treated groups based on labeling.
Fig. 7. Working model. The diagram showed that NT3-chitosan, by providingan excellent microenvironment, enhanced local neurogenesis from endoge-nous NSCs and facilitated the formation of nascent neural synaptic networks,serving as a relay station to transmit nerve impulses from severed nerves.
13358 | www.pnas.org/cgi/doi/10.1073/pnas.1510194112 Yang et al.

proliferation of cells of the other systems, including immune andvasculature systems, might also elicit long-lasting effects remains tobe determined.Clearly there are advantages to engaging endogenous stem cells
for neural repair as opposed to transplantation of exogenous stemcells, where immune rejection as well as safety issues related totumor formation from transplanted stem cells must be considered.In summary, our findings suggest that CNS regeneration can bepossible with new strategies targeting three areas: reduction ofinflammation, promotion of axonal regrowth, and enhancement ofnew local neurogenesis and subsequent synaptic network forma-tion. It is likely that improving these three areas will potentiallylead to novel therapeutic approaches to treat SCI.
Materials and MethodsAll experimental procedures were approved by and performed in accordancewith the standards of the Experimental Animal Center of Capital MedicalUniversity and the Beijing Experimental Animal Association.
NT3-Chitosan Tube Preparation. The chitosan tubes and NT3-chitosan carrierswere prepared as described previously (17, 18, 24), with modifications. De-tailed information is provided in SI Materials and Methods.
Surgical Procedure and Animal Care. Female Wistar rats weighing 200–220 gwere anesthetized by i.p. injection of 6% chloral hydrate (0.6 mL/kg bodyweight), to prepare rat models of thoracic spinal cord transection (SI Ma-terials and Methods). The amounts of k-252a and k-252b used in these ex-periments were based on previously published studies (26).
Immunohistochemistry. Morphological analysis and quantification wereperformed as described previously (24), with modifications, as detailed in SIMaterials and Methods.
Nerve Tract Tracing and Quadruple Immune Fluorescent Labeling. Biotinylateddextran amine conjugated with fluorescein [BDA-fluorescein, 10% (wt/vol)solution; Molecular Probes] was injected into eight different sites within themotor cortex to label the CST (24). Only fibers longer than 40 μm werecounted (27). Details are provided in SI Materials and Methods.
Electrophysiological Studies. Indices measured included SEP and MEP. Elec-trophysiological assays were performed as described previously (28, 29), withmodifications (SI Materials and Methods).
Behavioral Assessment. Observers blinded to the treatment methods andgroupings performed BBB scoring in an open field (24, 30). More details areprovided in SI Materials and Methods.
Labeling of Endogenous Stem Cells. BrdU (50 mg/kg body weight; Sigma-Aldrich) was injected i.p. every 24 h for 1 wk after the operation (SI Materialsand Methods).
Immunoelectron Microscopy Data Analysis. We identified mature synapses onserial sections and defined them as described previously (31). Details areprovided in SI Materials and Methods.
Neural Circuit Test with the Multielectrode Dish. Preparation of the multi-electrode dish system (Alpha MED Scientific) was essentially as describedpreviously (32) (SI Materials and Methods).
Statistical Analysis. Unless stated otherwise, all values are presented as mean ±SD. The Shapiro–Wilk method was used for data normality analysis, and theLevene test was used to test for homogeneity of variance. One-way ANOVAand Bonferroni analysis (multiple comparison for three groups), or theStudent t test or Mann–Whitney U test, were used to determine statisticaldifferences between two groups. P < 0.05 was taken to indicate a statis-tically significant difference.
ACKNOWLEDGMENTS. We express heartfelt thanks to Hui Qiao (NingxiaMedical University) for her kind help and constructive comments. This workwas supported by the State Key Program of National Natural Science Foun-dation of China (Grants 31130022, 313101021, 31320103903, 31271037,81330030, 91319309, and 31271371), the National Science and TechnologyPillar Program of China (Grant 2012BAI17B04), the International Cooperationin Science and Technology Projects of the Ministry of Science Technology ofChina (Grant 2014DFA30640), the National 863 Project (Grant 2012AA020506),the National Ministry of Education Special Fund for Excellent DoctoralDissertation (Grant 201356), the Special Funds for Excellent Doctoral Disserta-tion of Beijing, China (Grant 20111000601), the Key Project of the Departmentof Science and Technology of Beijing (Grant D090800046609004), and the 973Project (Grant 2012CB966303).
1. Wang KC, et al. (2002) Oligodendrocyte-myelin glycoprotein is a Nogo receptor li-gand that inhibits neurite outgrowth. Nature 417(6892):941–944.
2. Mukhopadhyay G, Doherty P, Walsh FS, Crocker PR, Filbin MT (1994) A novel role formyelin-associated glycoprotein as an inhibitor of axonal regeneration. Neuron 13(3):757–767.
3. GrandPré T, Nakamura F, Vartanian T, Strittmatter SM (2000) Identification of the Nogoinhibitor of axon regeneration as a Reticulon protein. Nature 403(6768):439–444.
4. Sivasankaran R, et al. (2004) PKC mediates inhibitory effects of myelin and chon-droitin sulfate proteoglycans on axonal regeneration. Nat Neurosci 7(3):261–268.
5. Andrews EM, Richards RJ, Yin FQ, Viapiano MS, Jakeman LB (2012) Alterations inchondroitin sulfate proteoglycan expression occur both at and far from the site ofspinal contusion injury. Exp Neurol 235(1):174–187.
6. Lang BT, et al. (2015) Modulation of the proteoglycan receptor PTPσ promotes re-covery after spinal cord injury. Nature 518(7539):404–408.
7. Hagg T, Oudega M (2006) Degenerative and spontaneous regenerative processesafter spinal cord injury. J Neurotrauma 23(3-4):264–280.
8. Barnabé-Heider F, et al. (2010) Origin of new glial cells in intact and injured adultspinal cord. Cell Stem Cell 7(4):470–482.
9. Gonzalez R, Glaser J, Liu MT, Lane TE, Keirstead HS (2003) Reducing inflammationdecreases secondary degeneration and functional deficit after spinal cord injury. ExpNeurol 184(1):456–463.
10. Lu P, et al. (2012) Long-distance growth and connectivity of neural stem cells aftersevere spinal cord injury. Cell 150(6):1264–1273.
11. Abematsu M, et al. (2010) Neurons derived from transplanted neural stem cells re-store disrupted neuronal circuitry in a mouse model of spinal cord injury. J Clin Invest120(9):3255–3266.
12. Gage FH (2000) Mammalian neural stem cells. Science 287(5457):1433–1438.13. Luo Y, et al. (2015) Single-cell transcriptome analyses reveal signals to activate dor-
mant neural stem cells. Cell 161(5):1175–1186.14. Cameron HA, Woolley CS, McEwen BS, Gould E (1993) Differentiation of newly born
neurons and glia in the dentate gyrus of the adult rat. Neuroscience 56(2):337–344.15. Zigova T, Pencea V, Wiegand SJ, Luskin MB (1998) Intraventricular administration of
BDNF increases the number of newly generated neurons in the adult olfactory bulb.Mol Cell Neurosci 11(4):234–245.
16. van Praag H, Kempermann G, Gage FH (1999) Running increases cell proliferation andneurogenesis in the adult mouse dentate gyrus. Nat Neurosci 2(3):266–270.
17. Li X, Yang Z, Zhang A (2009) The effect of neurotrophin-3/chitosan carriers on theproliferation and differentiation of neural stem cells. Biomaterials 30(28):4978–4985.
18. Yang Z, Duan H, Mo L, Qiao H, Li X (2010) The effect of the dosage of NT3/chitosancarriers on the proliferation and differentiation of neural stem cells. Biomaterials31(18):4846–4854.
19. Schnell L, Schneider R, Kolbeck R, Barde YA, Schwab ME (1994) Neurotrophin-3 en-hances sprouting of corticospinal tract during development and after adult spinalcord lesion. Nature 367(6459):170–173.
20. Ward RE, et al. (2014) A characterization of white matter pathology following spinalcord compression injury in the rat. Neuroscience 260:227–239.
21. Bera MF, Connors BW, Paradiso MA (2001) Neuroscience: Exploring the Brain (Lip-pincott Williams & Wilkins, Philadelphia), 2nd Ed, pp 675–708.
22. Duan H, et al. (2015) Transcriptome analyses reveal molecular mechanisms underlying func-tional recovery after spinal cord injury. Proc Natl Acad Sci USA 112:13360–13365.
23. Doetsch F, Caillé I, Lim DA, García-Verdugo JM, Alvarez-Buylla A (1999) Subventricular zoneastrocytes are neural stem cells in the adult mammalian brain. Cell 97(6):703–716.
24. Li X, Yang Z, Zhang A, Wang T, Chen W (2009) Repair of thoracic spinal cord injury bychitosan tube implantation in adult rats. Biomaterials 30(6):1121–1132.
25. Courtine G, et al. (2008) Recovery of supraspinal control of stepping via indirectpropriospinal relay connections after spinal cord injury. Nat Med 14(1):69–74.
26. Makino K, et al. (2012) Inhibition of uterine sarcoma cell growth through suppressionof endogenous tyrosine kinase B signaling. PLoS One 7(7):e41049.
27. Piantino J, Burdick JA, Goldberg D, Langer R, Benowitz LI (2006) An injectable, bio-degradable hydrogel for trophic factor delivery enhances axonal rewiring and im-proves performance after spinal cord injury. Exp Neurol 201(2):359–367.
28. Arunkumar MJ, Srinivasa Babu K, Chandy MJ (2001) Motor and somatosensory evokedpotentials in a primate model of experimental spinal cord injury. Neurol India 49(3):219–224.
29. Schlag MG, Hopf R, Redl H (2001) Serial recording of sensory, corticomotor, and brain-stem-derived motor evoked potentials in the rat. Somatosens Mot Res 18(2):106–116.
30. Basso DM, Beattie MS, Bresnahan JC (1995) A sensitive and reliable locomotor ratingscale for open field testing in rats. J Neurotrauma 12(1):1–21.
31. Lessard A, Coleman CG, Pickel VM (2010) Chronic intermittent hypoxia reduces neu-rokinin-1 (NK(1)) receptor density in small dendrites of non-catecholaminergic neu-rons in mouse nucleus tractus solitarius. Exp Neurol 223(2):634–644.
32. Oka H, Shimono K, Ogawa R, Sugihara H, Taketani M (1999) A new planar multi-electrode array for extracellular recording: Application to hippocampal acute slice.J Neurosci Methods 93(1):61–67.
Yang et al. PNAS | October 27, 2015 | vol. 112 | no. 43 | 13359
NEU
ROSC
IENCE


















